Embed Size (px)
Citation preview

Estudios de seguimiento de la reserva
marina de Cabo de Palos – Islas Hormigas 2013
GARCÍA CHARTON, J.A., LORENZI, M.R., CALÒ, A., TREVIÑO OTÓN, J., IRIGOYEN, A., HERNÁNDEZ ANDREU, R., MUÑOZ
GABALDÓN, I., MARCOS, C., PÉREZ RUZAFA, Á.
Departamento de Ecología e Hidrología
Universidad de Murcia
Informe producido en el marco del Convenio de Colaboración entre la Consejería de Agricultura y Agua – Comunidad Autónoma de la
Región de Murcia y la Universidad de Murcia
MURCIA, DICIEMBRE DE 2013

- 2 -

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 3 -
Tabla de contenidos
11 IInnttrroodduucccciióónn .................................................................... -- 55 --
1.1 La reserva marina de Cabo de Palos – Islas Hormigas - 6 -
1.2 Objetivos ..........................................- 8 -
22 DDiinnáámmiiccaa pplluurriiaannuuaall ddeell ppoobbllaammiieennttoo ddee ppeecceess ddee ffoonnddooss rrooccoossooss eenn llaa rreesseerrvvaa mmaarriinnaa ddee CC.. PPaallooss –– II.. HHoorrmmiiggaass ((11999966--22001133)) .......................................................... -- 99 --
2.1 Introducción .......................................- 9 -
2.2 Objetivos .........................................- 11 -
2.3 Material y métodos ................................- 12 -
2.3.1 Censos visuales de peces.......................- 12 - 2.3.2 Diseño de muestreo.............................- 13 - 2.3.3 Análisis de datos..............................- 14 -
2.4 Resultados ........................................- 15 -
2.4.1 Censos del poblamiento de peces en la reserva marina de Cabo de Palos – Islas Hormigas en 2013 .............- 15 - 2.4.2 Dinámica espacio-temporal de los peces en la reserva marina (1996-2013) ....................................- 15 - 2.4.3 Evolución temporal de la abundancia de las poblaciones de meros en la reserva marina de Cabo de Palos – Islas Hormigas (2009-2013) ............................- 25 -
2.5 Discusión .........................................- 27 -
33 UUssoo ddeell bbrriioozzoooo MMyyrriiaappoorraa ttrruunnccaattaa ccoommoo iinnddiiccaaddoorr ddeell iimmppaaccttoo ddeell ssuubbmmaarriinniissmmoo rreeccrreeaattiivvoo eenn llaa rreesseerrvvaa mmaarriinnaa ddee CC.. PPaallooss –– II.. HHoorrmmiiggaass ................................ -- 3311 --
3.1 Introducción ......................................- 31 -
3.2 Objetivos .........................................- 33 -
3.3 Material y métodos ................................- 34 -
3.3.1 Especie de estudio: Myriapora truncata.........- 34 - 3.3.2 Diseño y técnica de muestreo...................- 36 - 3.3.3 Análisis de datos..............................- 36 -
3.4 Resultados ........................................- 37 -
3.5 Discusión .........................................- 42 -

- 4 -
44 LLaa ppeessccaa aarrtteessaannaall eenn llaa rreesseerrvvaa mmaarriinnaa ddee CC.. PPaallooss –– II.. HHoorrmmiiggaass ...................................................................... -- 5511 --
4.1 Introducción ......................................- 51 -
4.2 Objetivos .........................................- 53 -
4.3 Material y métodos ................................- 53 -
4.3.1 Área de estudio................................- 53 - 4.3.2 Recogida y análisis de datos...................- 55 -
4.4 Resultados ........................................- 58 -
4.4.1 Distribución espacial del esfuerzo pesquero en 2013.- 58 - 4.4.2 Evolución temporal del CPUE y €PUE entre 1993 y 2012 - 64 -
4.5 Discusión .........................................- 73 -
55 CCoonncclluussiioonneess .................................................................. -- 7799 --
5.1 Poblamiento de peces de fondos rocosos ............- 79 -
5.2 Impacto del submarinismo recreativo ...............- 80 -
5.3 Pesca artesanal ...................................- 83 -
66 BBiibblliiooggrraaffííaa .................................................................. -- 8877 --

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 5 -
11 Introducción
Las áreas marinas protegidas (AMPs) han sido propuestas como una
herramienta de gestión para la conservación de los recursos
marinos (García-Charton et al. 2008; Fenberg et al. 2012). Sus
múltiples beneficios para la protección de los ecosistemas y los
procesos ecológicos que los mantienen, a la vez que mejoran las
pesquerías mediante la exportación denso-dependiente de biomasa
de especies de interés comercial a áreas adyacentes (Goñi et al.
2008; Harmelin-Vivien et al. 2008) hacen de las AMPs la medida
más poderosa de gestión espacial del medio ambiente marino.
Un AMP ha de ser adecuadamente monitoreada con el fin de
establecer científicamente si se están produciendo (o no) los
beneficios esperados en términos de conservación (p. ej.
recuperación de poblaciones o ecosistemas) y de objetivos de
gestión (p. ej., mejorar las pesquerías locales). Para ello, se
ha de cumplir la condición previa de que el AMP está bien
vigilada y apropiadamente gestionada, pues de otro modo no es
esperable efecto positivo alguno de la protección (Guidetti et
al. 2008). En años recientes, se ha realizado un enorme esfuerzo
para el establecimiento de nuevas AMPs en el Mediterráneo
(Gabrié et al. 2012). Sin embargo, como en otras regiones del
globo (Mora et al. 2006), muchas AMPs mediterráneas no han
mostrado los efectos esperados. El correcto diseño de redes de
AMPs no puede hacerse sin un adecuado conocimiento de las pautas
y procesos que caracterizan y determinan su éxito (Halpern &
Warner 2003), así como eventuales cambios en los resultados
(biológicos, ecológicos, socioeconómicos) de la protección. Por
ello, resulta esencial contar con datos a largo plazo de los
efectos ecológicos de aquéllas AMPs, como la de Cabo de Palos –
Islas Hormigas, que muestran unos signos inequívocos de éxito de
las medidas de gestión, con el fin de que dichos resultados sean
aplicables a la gestión adaptativa del espacio, así como a otros
casos en el ámbito mediterráneo y mundial.

- 6 -
1.1 La reserva marina de Cabo de Palos – Islas
Hormigas
La reserva marina de Cabo de Palos – Islas Hormigas se creó en
1995, merced a la figura legal de Reserva Marina de Interés
Pesquero, por la entonces Consejería de Agricultura, Ganadería y
Pesca (Decreto 15/1995, de 31 de marzo, por el que se declara
reserva marina de interés pesquero la zona de Cabo de Palos –
Islas Hormigas), junto con el Ministerio de Agricultura, Pesca y
Alimentación (Orden de 22 de junio de 1995, por la que se
establece una reserva marina en el entorno de Cabo de Palos -
Islas Hormigas), con el objeto de proteger a las comunidades
marinas y a las poblaciones de organismos de interés pesquero
que en ellas se desarrollan. La reserva marina de Cabo de Palos
– Islas Hormigas ocupa un área de 1.898 ha, y consta de una zona
de Reserva Integral, alrededor del archipiélago de las Hormigas,
de 270 ha de superficie, en la cual se prohíbe cualquier tipo de
actividad extractiva o recreativa, estando el resto del área
(punta del cabo de Palos, y bajos de La Testa, Piles, Dentro y
Fuera) sometida a uso controlado (pesca artesanal, pesca
deportiva desde tierra, submarinismo deportivo y actividades
científicas) (Fig. 1.1). La pesca recreativa está explícitamente
prohibida en la reserva marina.
Figura 1.1 Área de estudio, con indicación de los límites de la reserva marina de Cabo de Palos –Islas Hormigas, y de las principales biocenosis bentónicas presentes.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 7 -
Esta área marina se encuentra igualmente incluida en sendas
figuras de protección ambiental, al formar parte de un Lugar de
Interés Comunitario (LIC de la red Natura 2000, por la Directiva
“Hábitats” de la UE) y de una Zona de Especial Interés para el
Mediterráneo (ZEPIM, al amparo del Convenio de Barcelona), y
formar parte de una Zona de Especial Protección para las aves
(ZEPA, bajo la Directiva “Aves” de la UE).
El grupo de investigación “Ecología y ordenación de
ecosistemas marinos costeros” de la Universidad de Murcia ha
estado llevando a cabo el seguimiento científico de la reserva
marina de Cabo de Palos – Islas Hormigas desde su creación
(1995), merced a un convenio de colaboración entre la
Universidad de Murcia y la Consejería de Agricultura y Agua de
la Comunidad Autónoma de la Región de Murcia.
Figura 1.2 El mero (Epinephelus marginatus) es quizás la especie piscícola más emblemática en la reserva marina de Cabo de Palos –Islas Hormigas.

- 8 -
1.2 Objetivos
El objeto del presente informe es dar cuenta de los trabajos
realizados entre noviembre de 2012 y diciembre de 2013 para el
seguimiento de los efectos de la reserva marina de Cabo de Palos
– Islas Hormigas, especificándose los antecedentes, objetivos,
métodos llevados a cabo, y los principales resultados. Durante
este período se han llevado a cabo las siguientes tareas:
Estudio del poblamiento de peces de fondos rocosos en la
reserva marina de Cabo de Palos – Islas Hormigas:
- Efectos de la protección pesquera a largo plazo (1995-
2013) sobre la comunidad de peces arrecifales
- Dinámica temporal de las poblaciones de meros
(Serranidae: Epinephilinae)
Estudio de la pesca artesanal en y alrededor de la reserva
marina:
- Caracterización de la flota pesquera artesanal
- Distribución espacial del esfuerzo pesquero
- Evolución temporal de las capturas y rendimientos
económicos de la pesca
Seguimiento de los efectos del submarinismo recreativo sobre
los fondos de la reserva marina: respuesta de la especie
indicadora Myriapora truncata.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 9 -
22 DDiinnáámmiiccaa pplluurriiaannuuaall ddeell ppoobbllaammiieennttoo ddee
ppeecceess ddee ffoonnddooss rrooccoossooss eenn llaa rreesseerrvvaa
mmaarriinnaa ddee CC.. PPaallooss –– II.. HHoorrmmiiggaass ((11999966--
22001133))
2.1 Introducción
Las áreas marinas protegidas (AMPs) con zonas de reserva
integral son herramientas que han demostrado su eficacia para
recuperar y mantener la biodiversidad (Claudet et al. 2008,
2010; García-Charton et al. 2008a; Lester et al. 2009; Fenberg
et al. 2012). Constituyen una solución a la pérdida de
biodiversidad marina, la alteración de las estructuras tróficas
y la sobrepesca (Russ & Alcala 2004), ya que se ha visto que
aumentan la biomasa y diversidad de grandes peces piscívoros y
peces que se alimentan de invertebrados, sobre todo serránidos,
espáridos y lábridos (Prato et al. 2013).
Resulta imprescindible, sin embargo, mantener una
protección continua y duradera para alcanzar la capacidad de
carga del sistema, sobre todo en especies longevas y de lento
crecimiento como los meros (Guidetti et al. 2008; García-Rubies
et al. 2013). En general el tiempo de recuperación de las
poblaciones de peces es muy elevado, especialmente cuando sigue
habiendo una presión pesquera en el borde de la reserva
(McClanahan et al. 2007). En algunos casos se ha observado que
la biomasa de peces depredadores todavía aumenta
exponencialmente después de 14 y 18 años (Russ & Alcala 2004).
Una monitorización bien diseñada y a largo plazo en zonas
con buena protección frente a áreas control son la mejor forma
de documentar el tiempo que necesitan las especies para alcanzar
su capacidad de carga en AMPs (García-Rubies et al. 2013).
Predecir el tiempo necesario para la recuperación de especies
permite determinar el momento en el que se producirá una
exportación de biomasa o ‘spillover’, ya sea porque las especies
alcanzan su capacidad de carga o porque al ser individuos

- 10 -
mayores aumentan su espacio vital (Coll et al. 2012). Además
supone una recuperación funcional del ecosistema, con una
inversión de la pirámide trófica y una mayor importancia del
compartimento de los depredadores (García-Rubies et al. 2013).
En el Mediterráneo, los meros (Epinephilinae) son
depredadores apicales con un efecto crucial sobre las redes
tróficas marinas, e indicadores del estado de conservación de
los ecosistemas (Russ & Alcala 2004; Hackradt 2012; García-
Rubies et al. 2013), y por ello su interés ecológico y comercial
es indudable (Cornish & Harmelin-Vivien 2004). Los meros han
sido declarados especies amenazadas por la IUCN (Cornish &
Harmelin-Vivien 2004; Sadovy de Mitcheson et al. 2012) debido a
un descenso de sus poblaciones por muchos años de
sobreexplotación, su elevada fidelidad a zonas concretas del
territorio y su comportamiento y características reproductivas
(Shunter et al. 2011). Además del interés ecológico, los meros
son especies emblemáticas marinas que tienen un impacto en el
turismo y la economía de determinadas regiones (Shunter et al.
2011).
Evidencias recientes basadas en datos obtenidos entre 2009
y 2011 (García-Charton et al. 2013) apuntaban a que pudo
producirse una disminución importante (próxima al 70%) de la
abundancia de meros en la reserva marina de Cabo de Palos –
Islas Hormigas. En el citado informe, se concluyó que dicha
disminución fue más drástica durante el último año estudiado
(2011), sobre todo en la reserva integral. De confirmarse, la
explicación más plausible para dicha disminución sería el
aumento de la presión por pescadores furtivos. Se imponía pues
la necesidad de comprobar si dicha tendencia a la disminución de
efectivos se mantenía en el tiempo, o por el contrario se había
conseguido atajar con el cambio de estrategia en la vigilancia
acaecido a partir del otoño de 2011 (es decir, después de
obtenidos los datos conducentes a las conclusiones anteriores).

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 11 -
2.2 Objetivos
Continuar el seguimiento a largo plazo de los efectos de la
protección marina sobre el poblamiento de peces de fondos
rocosos en la reserva marina de Cabo de Palos – Islas
Hormigas.
Evaluar la abundancia y estimar la densidad de meros (Fig.
2.1) en la reserva marina de Cabo de Palos – Islas Hormigas,
en comparación con años anteriores, en los que se detectó una
disminución notable de sus efectivos.
Figura 2.1 Las tres especies de meros objeto del presente informe.

- 12 -
2.3 Material y métodos
2.3.1 Censos visuales de peces
El poblamiento de peces fue estimado entre agosto y octubre de
2013, mediante la realización de censos visuales con escafandra
autónoma a lo largo de transectos de 50 metros de largo por 5 m
de ancho. A partir de un punto dado -seleccionado en el momento
de fondear la embarcación en una zona rocosa cualquiera de la
zona correspondiente- el observador descendía hasta la cota de ~
-15 m, y, tras depositar sobre el fondo el extremo de una cinta
métrica atada a un plomo, avanzaba lentamente siguiendo la
isobata y censando peces a la vez que iba desenrollando la cinta
métrica.
El protocolo de censo ha consistido en que el observador,
provisto de equipo de buceo autónomo (equipo ligero de buceo,
botella de aire comprimido, regulador monotráquea, octopus,
consola con manómetro, chaleco estabilizador y ordenador de
inmersión) recorría cada transecto lentamente (2-8 m min-1) y
con natación constante, manteniéndose cerca del fondo, y
anotando en una hoja de poliéster sujeta a una tablilla
submarina de PVC las observaciones realizadas. Un primer vistazo
hacia delante y en la columna de agua (considerando 4 m por
encima del observador) permitía avistar aquéllas especies
mejores nadadoras y/o huidizas. A continuación eran
cuidadosamente inspeccionadas las grietas y oquedades de la
roca, así como el estrato foliar de las manchas de Posidonia
oceanica, a la búsqueda de individuos allí refugiados. Se ha
contado el número de individuos observados adscribiendo cada
observación a una de las siguientes clases de abundancia, cuyos
límites (a, b) de clase coinciden con los términos de una serie
geométrica de base aproximadamente 2 (1, 2-5, 6-10, 11-30, 31-
50, 51-100, 101-200, 201-500, >500). De este modo, se anotaba en
una cada una de las observaciones de individuos (aislados o en
grupo) en el interior de cada unidad muestral, asignándola a una
u otra de las 9 clases de abundancia preestablecidas. Para los
cálculos y análisis de datos se utilizaron como marcas de clase
los números enteros más próximos a las respectivas medias
geométricas (√ab) de estas clases de abundancia (1 / 3 / 8 / 18
/ 39 / 71 / 142 / 317 / 500), por coherencia con la métrica
geométrica utilizada. El protocolo de trabajo incluía la estima

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 13 -
de las tallas individuales, por comparación de los individuos
observados con una regleta graduada portada por el observador.
La talla de los individuos se estimó, tras un periodo de
entrenamiento, aproximándola a clases de 2 cm. Para evitar la
variabilidad debida al comportamiento nictímero del poblamiento
y a las condiciones de iluminación, los censos se llevaron a
cabo entre las 10 y las 15 h y siempre cuando las condiciones
del mar fueran favorables. Las especies de pequeño tamaño que se
caracterizan por su cripsis (Gobiidae, Callyonimidae, Bleniidae,
Gobioesocidae y Tripterygidae), se ignoraron debido a que
requieren diferentes tamaños de la unidad muestral para su
muestreo.
Tras la realización del censo, el observador recorría el
transecto en sentido inverso al de los censos a lo largo de la
cinta métrica, tomando nota de las variables descriptoras de la
complejidad del sustrato y midiendo la heterogeneidad del
hábitat, mientras el buceador acompañante recogía la cinta
métrica. Todo el procedimiento, incluido el censo de peces,
suponía unos 50-60 minutos de inmersión con escafandra autónoma
por zona. Como descriptores de la complejidad del hábitat se han
considerado la verticalidad y el número de bloques rocosos
clasificados por tamaños (piedras, bloques y grandes bloques),
mientras que se ha estimado la heterogeneidad del hábitat como
porcentaje de cobertura por arena, roca y Posidonia, midiéndose
además las profundidades media, mínima y máxima en el interior
del transecto.
2.3.2 Diseño de muestreo
El muestreo trazado siguió, como continuación del seguimiento
realizado desde 1996, un diseño jerarquizado en el espacio. Este
planteamiento permite detectar la variación existente a cada
escala espacial, identificando aquellas escalas espaciales que
más influyen en la distribución de las especies. Consideramos la
variación entre AÑOS, y tres SECTORES en la reserva marina,
separados miles de metros entre sí (SECTOR 1: costa y bajo de la
Testa; SECTOR 2: bajos de Piles y de Dentro; SECTOR 3: reserva
integral y bajo de Fuera). En cada uno de estos tres sectores se

- 14 -
establecieron, de forma aleatoria, tres ZONAS, dentro de las
cuales se realizaron tres réplicas (transectos).
El protocolo ha permitido contar con los siguientes datos
para cada unidad muestral: composición específica, número de
especies (riqueza), y abundancia (total y por especies),
distribución de tallas por especies y tamaño de los cardúmenes.
En junio y julio de 2013 se llevó a cabo un censo de meros
(Serranidae: Epinephelinae) independiente del seguimiento del
poblamiento en su conjunto, con el fin de comparar dichos datos
con los tomados en años anteriores (2009-2011) en el marco de la
tesis doctoral de Carlos W. Hackradt, en los que se observó una
preocupante caída de las abundancias de estas especies.
2.3.3 Análisis de datos
Las diferencias observadas de esas variables biológicas entre
AÑOS, teniendo en cuenta la estructura espacial jerarquizada
(SECTORES, ZONAS y TRANSECTOS) fueron testadas mediante un
análisis de la varianza. Los análisis univariantes se llevaron a
cabo con cada una de las categorías, tanto espaciales como
tróficas, así como con las especies más representativas,
utilizándose para ello el programa GAD del paquete R (Sandrini-
Neto & Camargo 2011). Los datos de abundancia fueron
transformados [log (x + 1)] por coherencia con la escala
logarítmica de adquisición de datos (clases de abundancia), para
homogeneizar las varianzas, y también con el fin de reducir el
peso de las especies más abundantes y aumentar el de las
especies más raras.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 15 -
2.4 Resultados
2.4.1 Censos del poblamiento de peces en la reserva marina de Cabo de Palos – Islas Hormigas en 2013
En los censos realizados durante el verano de 2013 en la reserva
marina han sido observadas 35 especies de peces, pertenecientes
a 11 familias (Tabla 2.1); entre éstas últimas destacan los
espáridos (12 especies), lábridos (9 especies) y serránidos (6
especies). Durante los censos de 2013 se han observado, por
término medio, 13 especies y 285 individuos por transecto de
50×5 m2, cifra ésta última que baja a 105 individuos si se
eliminan del cómputo las especies formadoras de bancos
(abundancia reducida, al restar las especies plantonófagas como
las bogas – Boops boops, más las castañuelas – Chromis chromis y
los trescolas – Anthias anthias) (Tabla 2.1).
2.4.2 Dinámica espacio-temporal de los peces en la
reserva marina (1996-2013)
Con los datos anteriores, el cómputo total de especies ícticas
observadas mediante censos visuales con escafandra autónoma en
la reserva marina es de 56 especies (21 familias) (Tabla 2.1).
En la Tabla 2.2 se muestran los resultados de los análisis de la
varianza mixtos aplicados a los parámetros del poblamiento de
peces (abundancia total – reducida y riqueza) y a los datos de
abundancia de las especies más frecuentes, en los que se explora
el efecto de la variación anual y las diferencias espaciales
(entre sectores, y entre zonas dentro de cada sector). La
abundancia total, así como la abundancia de 15 especies
(Epinephelus marginatus, Mycteroperca rubra, Serranus cabrilla,
Dentex dentex, Diplodus puntazzo, Diplodus vulgaris, Oblada
melanura, Sarpa salpa, Spondyliosoma cantharus, Labrus merula,
Symphodus mediterraneus, Symphodus rostratus, Symphodus tinca,
Thalassoma pavo y Sphyraena viridensis) muestran un efecto
significativo de la interacción entre AÑO y SECTOR, lo cual
significa que las diferencias interanuales no han sido de la
misma naturaleza entre los distintos sectores estudiados (Tabla
2.2). En general, las diferencias interanuales han consistido en
que las abundancias medias máximas han correspondido a uno otro
sector, dependiendo del año, aunque en muchos casos la

- 16 -
abundancia es mayor en los bajos y/o la reserva integral que en
el sector costero (Fig. 2.2). En muchos de esos casos la
tendencia interanual general es a un aumento de las abundancias
medias, mientras que en otras especies no hay una tendencia
clara (p. ej., Serranus cabrilla o Diplodus vulgaris).
Otras 4 especies (Mullus surmuletus, Diplodus annularis,
Pagrus pagrus y Symphodus doderleini) presentan una variabilidad
significativa entre sectores, con independencia del año
prospectado, y sin mostrar diferencias significativas entre años
(Tabla 2.2, Fig. 2.3). En todos los casos, la abundancia sigue
una progresión negativa desde la costa a la reserva integral,
como consecuencia, con toda probabilidad, de las diferencias de
hábitat entre los alrededores del cabo de Palos y los bajos e
islas más alejadas del continente.
Otro grupo de especies presentan simultáneamente efectos
significativos del año y del sector, aunque ambos efectos son
independientes entre sí; tal es el caso de Epinephelus costae,
Serranus scriba, Apogon imberbis, Sciaena umbra, Diplodus
cervinus, D. sargus, Chromis chromis, Coris julis, Labrus
viridis, Symphodus ocellatus y Scorpaena scrofa (Tabla 2.2). En
el caso de las especies con interés comercial (Fig. 2.4), en
general han presentado un aumento temporal claro, además de que
se distribuyen entre los bajos o las islas y no tanto en el
sector costero. Finalmente, únicamente una especie (Symphodus
melanocercus) ha mostrado una variabilidad interanual
significativa (Tabla 2.2).

Tabla 2.1 Valores medios de abundancia de las especies observadas y de los parámetros descriptores del poblamiento de peces (± error típico) en cada año de seguimiento de la reserva marina de Cabo de Palos – Islas Hormigas. Familia Especie 1996 1998 2001 2002 2003 2004 2005
Myliobatidae Myliobatis aquila 0,2 ± 0.2 0.04 ± 0.04 0.4 ± 0.2 0.4 ± 0.2
Engraulidae Engraulis encrasicolus 11,7 ± 11,7 37.0 ± 25.7
Muraenidae Muraena helena 0,8 ± 0,3 0,3 ± 0,1 0,4 ± 0.1 0.5 ± 0.2 2.1 ± 0.5 0.5 ± 0.2 0.1 ± 0.1
Belonidae Belone belone 0,1 ± 0,1
Phycidae Phycis phycis 0,0 ± 0,0 0,1 ± 0.1
Serranidae Anthias anthias 65,1 ± 19,5 39,2 ± 15,1 22,1 ± 7.0 11.3 ± 3.8 44.7 ± 15.6 20.2 ± 7.9 25.3 ± 9.0
Epinephelus costae 0,1 ± 0,1 0,7 ± 0,3 0,4 ± 0.2 1.0 ± 0.6 0.9 ± 0.3 0.7 ± 0.2 1.6 ± 0.8
Epinephelus marginatus 0,2 ± 0,1 0,9 ± 0,2 3,4 ± 0.7 4.0 ± 1.0 3.9 ± 0.8 2.6 ± 0.8 5.6 ± 0.9
Epinephelus caninus 0.1 ± 0.1
Mycteroperca rubra 0,1 ± 0.1 0.3 ± 0.1 0.6 ± 0.3 1.9 ± 0.8
Serranus atricauda 0,04 ± 0,04 0,1 ± 0,1 0,04 ± 0.04 0.04 ± 0.04
Serranus cabrilla 2,3 ± 0,3 1,3 ± 0,3 1,9 ± 0.4 1.4 ± 0.3 1.6 ± 0.4 0.2 ± 0.1 0.5 ± 0.2
Serranus scriba 1,2 ± 0,2 1,4 ± 0,2 2,1 ± 0.4 2.5 ± 0.5 2.2 ± 0.3 1.4 ± 0.3 1.4 ± 0.3
Apogonidae Apogon imberbis 8,2 ± 2,3 3,4 ± 0,9 15,1 ± 4.3 7.9 ± 1.4 4.9 ± 1.3 2.9 ± 0.9 1.6 ± 0.7
Carangidae Seriola dumerili 1,4 ± 1,4 7.1 ± 5.4 0.2 ± 0.1
Trachurus mediterraneus 0,3 ± 0,3 0,8 ± 0,7
Pseudocaranx dentex 0.2 ± 0.1
Scombridae Sarda sarda
Coryphaenidae Coryphaena hippurus 0,04 ± 0,04
Haemulidae Pomadasys incisus 0,04 ± 0,04
Parapristipoma octolineatum
Sciaenidae Sciaena umbra 0,9 ± 0,7 1,8 ± 0,9 5,2 ± 1.7 4.2 ± 1.7 5.5 ± 1.7 8.4 ± 3.3 12.8 ± 4.5
Mullidae Mullus surmuletus 0,5 ± 0,2 0,6 ± 0,2 0,6 ± 0.2 0.6 ± 0.2 0.2 ± 0.1 0.2 ± 0.1 0.3 ± 0.1
Sparidae Boops boops 13,1 ± 13,1 38,5 ± 37,0 190.6 ± 82.4 101.9 ± 46.7
Dentex dentex 2,9 ± 1,7 0,4 ± 0,2 5,8 ± 4.4 2.0 ± 0.7 1.7 ± 0.6 5.3 ± 2.6 2.3 ± 0.5
Diplodus annularis 1,4 ± 0,5 0,4 ± 0,2 0,8 ± 0.2 1.1 ± 0.4 0.7 ± 0.2 0.5 ± 0.2 0.9 ± 0.3
Diplodus cervinus 0,3 ± 0,1 0,2 ± 0,2 0,3 ± 0.1 0.5 ± 0.2 2.0 ± 0.5 0.8 ± 0.2 1.3 ± 0.3
Diplodus puntazzo 3,5 ± 0,8 4,3 ± 1,3 5,4 ± 1.9 4.5 ± 0.8 2.7 ± 0.7 2.6 ± 0.7 2.5 ± 0.8
Diplodus sargus 9,5 ± 3,6 11,6 ± 3,3 12,3 ± 3.7 12.4 ± 1.7 8.2 ± 1.3 7.6 ± 1.7 5.0 ± 1.0

- 18 -
Familia Especie 1996 1998 2001 2002 2003 2004 2005
Diplodus vulgaris 25,0 ± 4,7 32,0 ± 7,0 32,0 ± 9.5 15.1 ± 3.3 12.2 ± 3.0 18.4 ± 4.6 38.4 ± 13.8
Oblada melanura 33,3 ± 13,5 10,5 ± 3,5 5,7 ± 2.7 4.7 ± 2.0 0.9 ± 0.8 11.4 ± 7.5 4.1 ± 2.2
Pagrus pagrus 0,0 ± 0,0 0,1 ± 0,1 0.04 ± 0.04
Pagrus auriga 0.1 ± 0.1
Sarpa salpa 37,7 ± 11,1 46,0 ± 7,9 37,6 ± 7.8 43.4 ± 10.5 11.3 ± 3.8 14.9 ± 5.5 8.3 ± 3.4
Sparus aurata 0,04 ± 0,04 0,1 ± 0,1 0,3 ± 0.2 0.1 ± 0.1 0.3 ± 0.1 0.4 ± 0.1 0.7 ± 0.4
Spondyliosoma cantharus 0,1 ± 0,1 17.9 ± 7.7 3.1 ± 1.7
Centracanthidae Spicara smaris 37,0 ± 37,0 18.4 ± 17.0
Pomacentridae Chromis chromis 527,7 ± 89,2 635,4 ± 84,2 748,0 ± 108.6 571.8 ± 73.4 470.4 ± 105.2 443.7 ± 62.2 441.1 ± 70.3
Labridae Coris julis 32,7 ± 7,7 13,0 ± 1,9 46,1 ± 10.3 9.9 ± 2.2 57.0 ± 13.3 152.4 ± 45.0 88.6 ± 31.1
Labrus merula 0,04 ± 0,04 0,1 ± 0.1 0.1 ± 0.1 0.1 ± 0.1 0.1 ± 0.1 0.1 ± 0.1
Labrus viridis 0.04 ± 0.04 0.1 ± 0.1
Symphodus dordeleini 0,04 ± 0,04 0,5 ± 0,2 0,2 ± 0.1 0.1 ± 0.1 0.1 ± 0.1 0.04 ± 0.04 0.1 ± 0.1
Symphodus mediterraneus 0,5 ± 0,2 1,9 ± 0,4 1,2 ± 0.3 1.2 ± 0.2 2.0 ± 0.4 0.9 ± 0.2 1.4 ± 0.3
Symphodus melanocercus 0,04 ± 0,04 0,7 ± 0.3 0.1 ± 0.1 0.1 ± 0.1 0.7 ± 0.4
Symphodus ocellatus 2,6 ± 0,9 2,0 ± 0,6 13,7 ± 7.5 2.4 ± 0.7 4.8 ± 1.1 6.0 ± 2.4 4.0 ± 1.0
Symphodus roissali 0,4 ± 0,1 0,6 ± 0,2 1,1 ± 0.3 0.4 ± 0.1 0.5 ± 0.2 0.6 ± 0.2 0.2 ± 0.1
Symphodus rostratus 0,1 ± 0,1 0,1 ± 0,1 0,4 ± 0.1 0.3 ± 0.1 0.1 ± 0.1 0.2 ± 0.1 0.3 ± 0.1
Symphodus tinca 2,6 ± 0,5 4,1 ± 0,7 10,0 ± 1.6 3.3 ± 0.7 2.1 ± 0.6 4.4 ± 0.9 3.6 ± 1.0
Thalassoma pavo 99,1 ± 25,9 31,6 ± 6,2 28,9 ± 5.9 58.2 ± 17.2 47.9 ± 9.8 32.4 ± 9.0 16.5 ± 4.3
Sphyraenidae Sphyraena viridensis 40,3 ± 21,8 26,4 ± 14,5 0,3 ± 0.3 15.9 ± 8.6 21.0 ± 6.6 14.8 ± 7.3 21.3 ± 12.7
Mugilidae Mugilidae spp. 0,9 ± 0,7 0,5 ± 0,3 0,4 ± 0.4 0.1 ± 0.1
Scorpaenidae Scorpaena maderensis 0.1 ± 0.04
Scorpaena scrofa 0.3 ± 0.1 0.1 ± 0.1 0.1 ± 0.1
Scorpaena porcus 0,7 ± 0,7 0.04 ± 0.04
Scorpaena notata 0,04 ± 0,04 0.1 ± 0.1 0.1 ± 0.1
Atherinidae Atherina hepsetus 37,0 ± 25,7
Abundancia 951,4 ± 109,3 960,9 ± 139,4 1002,9 ± 119.9 1034.4 ± 135.8 713.1 ± 129.8 773.7 ± 102.7 798.5 ± 91.0
Abundancia reducida 233,6 ± 36,8 159,4 ± 15,0 226,2 ± 20.7 177.4 ± 21.3 175.8 ± 18.5 283.3 ± 52.3 204.3 ± 33.0
Riqueza media 14,2 ± 0,4 15,2 ± 0,6 16,8 ± 0.5 16.4 ± 0.5 16.8 ± 0.6 14.6 ± 0.5 15.4 ± 0.7
Riqueza total 40 39 35 36 39 35 38

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 19 -
Tabla 2.1 Cont. Familia Especie 2006 2007 2008 2009 2010 2013
Myliobatidae Myliobatis aquila 0,2 ± 0,1 0,04 ± 0,04 0.04 ± 0.04 0.1 ± 0.11 0.2 ± 0.12 0.5 ± 0.20
Engraulidae Engraulis encrasicolus 18.5 ± 18.5 18.5 ± 18.5
Muraenidae Muraena helena 0,4 ± 0,1 0,4 ± 0,1 0.4 ± 0.2 0.5 ± 0.2 0.4 ± 0.2 0.5 ± 0.16
Belonidae Belone belone
Phycidae Phycis phycis 0,04 ± 0,04
Serranidae Anthias anthias 1,7 ± 0,8 6,8 ± 2,9 14.3 ± 7.8 8.7 ± 3.3 19.5 ± 11.8 7.8 ± 2.51
Epinephelus costae 1,8 ± 0,8 0,9 ± 0,2 1.7 ± 0.5 2.4 ± 1.2 0.3 ± 0.2 0.9 ± 0.34
Epinephelus marginatus 8,2 ± 1,4 4,7 ± 0,8 6.7 ± 1.3 11.3 ± 1.8 7.7 ± 1.8 8.9 ± 1.80
Epinephelus caninus
Mycteroperca rubra 1,8 ± 0,5 0,6 ± 0,3 2.0 ± 0.6 3.1 ± 1.2 0.9 ± 0.4 1.7 ± 0.53
Serranus atricauda
Serranus cabrilla 1,0 ± 0,2 0,9 ± 0,2 1.4 ± 0.3 1.5 ± 0.3 1.1 ± 0.4 0.6 ± 0.19
Serranus scriba 1,9 ± 0,3 3,1 ± 0,4 7.3 ± 4.7 2.7 ± 0.4 2.5 ± 0.7 0.9 ± 0.23
Apogonidae Apogon imberbis 1,0 ± 0,5 1,6 ± 0,5 6.2 ± 1.7 6.3 ± 1.7 4.2 ± 1.4 0.9 ± 0.54
Carangidae Seriola dumerili 0.1 ± 0.1 0.04 ± 0.0
Trachurus mediterraneus
Pseudocaranx dentex 0,8 ± 0,5 0.04 ± 0.04 0.3 ± 0.26
Scombridae Sarda sarda 0.8 ± 0.8 4.8 ± 3.0
Coryphaenidae Coryphaena hippurus
Haemulidae Pomadasys incisus
Parapristipoma octolineatum 0,04 ± 0,04
Sciaenidae Sciaena umbra 5,9 ± 3,2 7,6 ± 3,0 8.5 ± 4.0 6.4 ± 2.6 10.9 ± 5.8 4.1 ± 2.03
Mullidae Mullus surmuletus 0,4 ± 0,2 0,5 ± 0,2 0.7 ± 0.3 0.7 ± 0.23 0.4 ± 0.25 0.1 ± 0.05
Sparidae Boops boops 443,8 ± 78,0 139,0 ± 44,0 101.8 ± 43.3 18.6 ± 18.5 96.8 ± 57.2 20.0 ± 18.52
Dentex dentex 7,4 ± 2,8 5,3 ± 1,7 11.1 ± 3.6 7.8 ± 2.8 7.8 ± 6.1 7.7 ± 2.76
Diplodus annularis 0,9 ± 0,7 0,7 ± 0,3 0.2 ± 0.1 0.4 ± 0.2 0.2 ± 0.1 0.4 ± 0.14
Diplodus cervinus 1,7 ± 0,3 1,0 ± 0,2 1.2 ± 0.5 1.5 ± 0.3 0.4 ± 0.2 1.2 ± 0.29
Diplodus puntazzo 3,6 ± 0,6 2,9 ± 0,5 4.6 ± 1.1 2.0 ± 0.6 1.3 ± 0.6 2.0 ± 0.67
Diplodus sargus 10,3 ± 2,1 7,0 ± 1,2 13.0 ± 5.1 4.4 ± 1.1 3.7 ± 1.2 4.6 ± 1.85

- 20 -
Familia Especie 2006 2007 2008 2009 2010 2013
Diplodus vulgaris 24,7 ± 9,0 24,3 ± 8,3 21.5 ± 8.1 16.7 ± 5.4 14.4 ± 5.4 11.5 ± 2.73
Oblada melanura 2,1 ± 0,8 11,8 ± 5,8 11.6 ± 9.1 3.2 ± 2.5 16.2 ± 10.8 1.8 ± 0.98
Pagrus pagrus 0,04 ± 0,04 0.04 ± 0.04 0.1 ± 0.05
Pagrus auriga
Sarpa salpa 5,9 ± 2,6 2,2 ± 1,0 9.1 ± 5.6 8.7 ± 3.2 6.0 ± 3.84
Sparus aurata 0,1 ± 0,1 0,1 ± 0,1 0.8 ± 0.4 0.3 ± 0.18 0.1 ± 0.08 0.3 ± 0.11
Spondyliosoma cantharus 3,6 ± 3,3 3,5 ± 1,9 0.5 ± 0.3 0.5 ± 0.3 12.8 ± 6.0 10.2 ± 4.60
Centracanthidae Spicara smaris 0,9 ± 0,4 0.04 ± 0.04 17.6 ± 17.6
Pomacentridae Chromis chromis 323,0 ± 61,6 290,8 ± 55,3 508.7 ± 95.5 227.4 ± 39.8 331.7 ± 75.5 140.2 ± 28.21
Labridae Coris julis 19,5 ± 4,8 28,2 ± 9,6 34.4 ± 10.8 16.5 ± 3.4 39.3 ± 10.9 35.6 ± 8.37
Labrus merula 0,4 ± 0,3 0,1 ± 0,1 0.1 ± 0.1 0.1 ± 0.05
Labrus viridis 0,04 ± 0,04 0,1 ± 0,1 0.1 ± 0.1
Symphodus dordeleini 0,2 ± 0,1 0,04 ± 0,04 0.3 ± 0.1 0.1 ± 0.07 0.2 ± 0.09 0.1 ± 0.05
Symphodus mediterraneus 1,4 ± 0,2 1,9 ± 0,4 1.3 ± 0.3 0.8 ± 0.2 1.5 ± 0.5 0.6 ± 0.14
Symphodus melanocercus 0,1 ± 0,1 0,1 ± 0,1 0.1 ± 0.1 0.2 ± 0.09
Symphodus ocellatus 6,2 ± 2,3 10,6 ± 5,2 10.3 ± 5.0 1.2 ± 0.5 2.3 ± 0.7 0.7 ± 0.18
Symphodus roissali 0,2 ± 0,1 0,3 ± 0,1 0.3 ± 0.2 0.4 ± 0.15 0.4 ± 0.17 0.2 ± 0.09
Symphodus rostratus 0,2 ± 0,1 1,0 ± 0,6 0.4 ± 0.1 0.3 ± 0.13 0.3 ± 0.23 0.04 ± 0.04
Symphodus tinca 3,0 ± 0,7 2,5 ± 0,6 1.4 ± 0.4 1.5 ± 0.5 2.4 ± 0.8 1.6 ± 0.56
Thalassoma pavo 12,4 ± 4,1 4,8 ± 1,3 7.8 ± 2.7 12.1 ± 3.6 5.2 ± 2.0 3.4 ± 1.27
Sphyraenidae Sphyraena viridensis 30,2 ± 15,0 9,8 ± 5,0 14.0 ± 4.3 2.7 ± 1.6 16.7 ± 7.4 9.1 ± 4.77
Mugilidae Mugilidae spp. 0.3 ± 0.30
Scorpaenidae Scorpaena maderensis
Scorpaena scrofa 0,04 ± 0,04 0,04 ± 0,04 0.1 ± 0.1 0.1 ± 0.07 0.1 ± 0.08
Scorpaena porcus
Scorpaena notata
Atherinidae Atherina hepsetus
Abundancia 926,3 ± 99,8 576,4 ± 75,6 822.4 ± 129.5 389.8 ± 40.5 564.6 ± 80.7 284.5 ± 44.7
Abundancia reducida 124,4 ± 15,0 124,9 ± 14,6 153.4 ± 17.6 110.4 ± 9.2 126.5 ± 14.3 105.3 ± 12.8
Riqueza media 16,4 ± 0,4 16,7 ± 0,5 16.2 ± 0.5 14.4 ± 0.6 14.3 ± 0.5 12.7 ± 0.7
Riqueza total 37 40 41 37 35 35

Tabla 2.2 Síntesis de los resultados de los análisis de la varianza, con indicación de la significación estadística del efecto de los factores AÑO, SECTOR y ZONA en la abundancia y riqueza de especies, y la abundancia de las especies más frecuentes en los censos de peces en la reserva marina de Cabo de Palos – Islas Hormigas, y del test C de Cochran de homocedasticidad [***: p < 0,001; **: p < 0,01; *: p < 0,05; (*): p < 0,1].
Variable / Especie C AÑO SECTOR AÑO x SECTOR ZONA
Abundancia reducida *** *** *
Riqueza media *** *** ***
Epinephelus costae * ** *** *
Epinephelus marginatus *** *** *** ***
Mycteroperca rubra ** *** *** **
Serranus cabrilla *** *** * ***
Serranus scriba *** *** *** (*) *** Apogon imberbis *** *** (*)
Sciaena umbra ** ***
Mullus surmuletus * *** * Dentex dentex * *** *** * *
Diplodus annularis *** *** ***
Diplodus cervinus ** *** *** ***
Diplodus puntazzo * (*) *** ** ***
Diplodus sargus * * ** *** Diplodus vulgaris ** *** **
Oblada melanura ** *** *** *
Pagrus pagrus *** * Sarpa salpa *** *** **
Sparus aurata *** (*) *
Spondyliosoma cantharus *** *** *** *** (*)
Chromis chromis ** *** *** **
Coris julis * *** *** *** Labrus merula *** *** *
Labrus viridis *** ** **
Symphodus dordeleini *** * * Symphodus mediterraneus *** ** **
Symphodus melanocercus *** * **
Symphodus ocellatus * *** *** (*) *
Symphodus roissali * (*) *** ***
Symphodus rostratus *** * *** *** Symphodus tinca *** *** ** **
Thalassoma pavo *** ** *** **
Sphyraena viridensis (*) *** * * Scorpaena scrofa *** * *** (*)

- 22 -
Figura 2.2 Valores medios de abundancia por transecto a lo largo del tiempo en la reserva marina de Cabo de Palos – Islas Hormigas, para las tres especies en los que la variación interanual y entre sectores no es independiente entre sí, distinguiéndose entre los tres sectores en los que se ha dividido [ : costa; : bajos; : islas Hormigas y bajo de Fuera].
0
2
4
6
8
10
12
14
16
18
20
Nº
ind
iv.
25
0 m
-2
Año
Epinephelus marginatus
0
1
2
3
4
5
6
7
Nº
ind
iv.
25
0 m
-2
Mycteroperca rubra
96 97 98 99 00 01 02 03 04 05 06 07 08 09 10 11 12 130
5
10
15
20
25
30
Nº
ind
iv.
25
0 m
-2
Año
Dentex dentex
0
2
4
6
8
10
12
14
16
18
20
Nº
ind
iv.
25
0 m
-2
Año
Epinephelus marginatus
0
1
2
3
4
5
6
7
Nº
ind
iv.
25
0 m
-2
Mycteroperca rubra
96 97 98 99 00 01 02 03 04 05 06 07 08 09 10 11 12 130
5
10
15
20
25
30
Nº
ind
iv.
25
0 m
-2
Año
Dentex dentex
96 97 98 99 00 01 02 03 04 05 06 07 08 09 10 11 12 130
5
10
15
20
25
30
Nº
ind
iv.
25
0 m
-2
Año
Dentex dentex

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 23 -
Figura 2.3 Variación de los valores medios de abundancia por transecto de raspallones (D. annularis) y salmonetes (M. surmuletus) entre sectores la reserva marina de Cabo de Palos – Islas Hormigas.
Es importante notar que las principales especies pesqueras
han visto aumentar sus efectivos en la reserva marina,
considerando todas los sectores juntos (Fig. 2.5). En todos los
casos, sin embargo, son notables valores de abundancia en 2013
más bajos que en años anteriores.
El caso de la gallineta (Scorpaena scrofa) no es tan claro
(Fig. 2.4), debido a que sus hábitos crípticos la hacen
difícilmente censable visualmente.
Finalmente, es destacable que la gran mayoría de las
especies presentan una gran variabilidad espacial a escala fina
(entre zonas dentro de cada sector) (Tabla 2.2).
Diplodus annularis
0
1
2
3N
ºin
div
. 2
50 m
-2
Mullus surmuletus
0
1
2
Costa Bajos Reserva integral
Diplodus annularis
0
1
2
3N
ºin
div
. 2
50 m
-2
Mullus surmuletus
0
1
2
Costa Bajos Reserva integral

- 24 -
Figura 2.4 Valores medios de abundancia por transecto a lo largo del tiempo en la reserva marina de Cabo de Palos – Islas Hormigas, para tres especies comerciales en los que la variación interanual y entre sectores es independiente entre sí, distinguiéndose entre los tres sectores en los que se ha dividido [ : costa; : bajos; : islas Hormigas y bajo de Fuera].
0
1
2
3
4
5
6
7
Nº
ind
iv.
25
0 m
-2
Epinephelus costae
0
5
10
15
20
25
Nº
ind
iv.
25
0 m
-2
Sciaena umbra
96 97 98 99 00 01 02 03 04 05 06 07 08 09 10 11 12 130
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Nº
ind
iv.
25
0 m
-2
Año
Scorpaena scrofa
0
1
2
3
4
5
6
7
Nº
ind
iv.
25
0 m
-2
Epinephelus costae
0
5
10
15
20
25
Nº
ind
iv.
25
0 m
-2
Sciaena umbra
96 97 98 99 00 01 02 03 04 05 06 07 08 09 10 11 12 130
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Nº
ind
iv.
25
0 m
-2
Año
Scorpaena scrofa

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 25 -
Figura 2.5 Variación de los valores medios de abundancia por transecto observada a lo largo del tiempo de las principales especies objetivo de la pesca artesanal en la reserva marina de Cabo de Palos – Islas Hormigas.
2.4.3 Evolución temporal de la abundancia de las
poblaciones de meros en la reserva marina de
Cabo de Palos – Islas Hormigas (2009-2013)
En el censo de meros realizado en junio y julio de 2013 se
observaron en promedio 0,9 ± 0,3 (error típico) meros (E.
marginatus), 8,9 ± 1,8 falsos abadejos (E. costae) y 1,7 ± 0,5
chernas (M. rubra) por transecto de 250 m2 en la reserva marina.
Si se comparan estos datos con lo observado entre 2009 y 2011
por Hackradt (2012), se observa una recuperación de los
efectivos numéricos de estas especies (Fig. 2.5). Por otra
parte, en el caso de meros y chernas estos valores han resultado
menores que los obtenidos a partir de los censos de peces
realizados entre agosto y octubre en la reserva marina cuyo
objetivo era describir todo el poblamiento íctico (incluidas las
tres especies de meros), observándose en algunos casos que se
recuperan los valores anteriores de densidad de meros (Fig.2.6,
véase asimismo las Fig. 2.2, 2.4 y 2.5).
Epinephelus costae
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Sciaena umbra
0
2
4
6
8
10
12
14
Epinephelus marginatus
0
2
4
6
8
10
12
nº
ind
iv.
250
m-2
Dentex dentex
0
2
4
6
8
10
12
Mycteroperca rubra
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
2012
2013
Diplodus cervinus
0.0
0.5
1.0
1.5
2.0
2.5
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
2010
2011
2012
2013

- 26 -
Figura 2.6 Cambio en la abundancia media de los meros, falsos abadejos y chernas en los tres sectores de la reserva marina de Cabo de Palos – Islas Hormigas entre 2009 y 2011 [barras grises, datos de Hackradt (2012)], y en 2013, distinguiendo entre los datos de los censos específicos de meros realizados en junio-julio (barras rojas) y los datos de los censos del poblamiento completo de peces realizados en agosto-octubre (barras rayadas).
Cherna
(Mycteroperca rubra)
0
1
2
3
4
5
Costa Bajos I. Hormigas
2009
2010
2011
2013
2009
2010
2011
2013
2009
2010
2011
2013
Mero
(Epinephelus marginatus)
0
2
4
6
8
10
12
14
16
18
20
Falso abadejo
(Epinephelus costae)
0
1
2
3
4
5
6
7
Nº
ind
iv.
25
0 m
-2
Cherna
(Mycteroperca rubra)
0
1
2
3
4
5
0
1
2
3
4
5
Costa Bajos I. Hormigas
2009
2010
2011
2013
2009
2010
2011
2013
2009
2010
2011
2013
2009
2010
2011
2013
2009
2010
2011
2013
2009
2010
2011
2013
Mero
(Epinephelus marginatus)
0
2
4
6
8
10
12
14
16
18
20
0
2
4
6
8
10
12
14
16
18
20
Falso abadejo
(Epinephelus costae)
0
1
2
3
4
5
6
7
Nº
ind
iv.
25
0 m
-2

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 27 -
2.5 Discusión
En síntesis, se confirma una recuperación de la comunidad de
peces, que había disminuido sus efectivos (al menos en lo que se
refiere a los meros) durante los últimos años, como
consecuencia, posiblemente, de un aumento de la presión por
furtivos, sobre todo en la reserva integral y el bajo de Fuera
(García-Charton et al. 2013).
La dinámica temporal de las poblaciones de peces, por
tanto, parece haber retomado la senda del incremento de las
abundancias, tal y como venía siendo la tónica durante los
últimos años en la reserva marina de Cabo de Palos – Islas
Hormigas (García-Charton et al. 2010). No obstante, no en todos
los casos se han recuperado los niveles de abundancia anteriores
a la drástica disminución observada en 2011, y no puede
descartarse que se mantenga cierta presión por furtivismo en la
reserva marina (a tenor asimismo de diversos testimonios
recogidos en la reserva marina) (García-Charton et al. 2013).
Únicamente con niveles muy elevados de vigilancia (en los que se
verifican a la vez una gran intensidad de salidas de control y
una muy baja frecuencia de episodios de furtivismo, Guidetti et
al. 2008) se pueden hacer cumplir plenamente las normativas
establecidas para la protección de un área marina. Una inversión
suficiente y una óptima organización de las tareas de vigilancia
y monitoreo son factores clave para el éxito de una reserva
marina (Maliao et al. 2004; Samoilys et al. 2007; Guidetti et
al. 2008; Pita et al. 2011; De Santo 2013; Gleason et al. 2013;
Pérez de Oliveira 2013), aunque resulta obvio que entran en
juego otros factores (Warner & Pomeroy 2012; Chuenpagdee et al.
2013). Esta necesidad es aún mayor, si cabe, en el caso de
especies que, como los meros, tienen un alto atractivo para los
pescadores furtivos por su gran tamaño y alto precio en el
mercado (Sadovy de Mitcheson et al. 2012), lo cual exige medidas
de vigilancia especialmente adaptadas a estas características
(Mangubhai et al. 2011).
Se confirma asimismo la gran importancia relativa de la
influencia de la estructura del hábitat sobre el poblamiento de
peces (García-Charton et al. 2001, 2004; Claudet et al. 2011),
representado en este caso por las diferencias entre sectores
(costa vs. bajos o islas), así como por las diferencias

- 28 -
significativas de abundancia a escala espacial fina (entre zonas
separadas por cientos de metros dentro de cada sector),
achacables a diferencias de hábitat a esa escala espacial.
Las especies más favorecidas por las medidas de protección
pesquera son meros (Fig. 2.7), dentones, corvas, sargos reales y
espetones, todas ellas de gran valor comercial, además de ser
especies carnívoras o específicamente piscívoras (es decir,
depredadores apicales en los ecosistemas marinos costeros). Con
ello, con la protección pesquera ocurre una “naturalización” de
dicho ecosistema hacia estadios más prístinos (es decir,
anteriores a la enorme presión antrópica ejercida sobre estos
ecosistemas) (Sandin et al. 2008). Tales cambios han de
acarrear, forzosamente, efectos indirectos o “en cascada”
(Pinnegar et al. 2000; Micheli et al. 2005; Baum & Word 2009),
tales como reducciones en la abundancia de mesodepredadores y
presas potenciales, así como de invertebrados bentónicos.
Es de destacar que los valores de abundancia media de las
tres especies de meros obtenidas en el verano de 2013 no han
coincidido en las dos ocasiones en los que se han muestreado,
una en junio-julio (censos específicos de meros) y otra entre
agosto y octubre (censos de peces incluyendo todo el poblamiento
observable), a pesar de haberse utilizado la misma técnica de
censo visual con escafandra autónoma (transectos de 50×5 m2).
Dos razones son las más plausibles para explicar esta
observación:
1) Las abundancias han sido efectivamente menores a principios
que a finales del verano: esto es quizás explicable por el
atípico comportamiento de las temperaturas del agua de mar,
ya que a mediados de mayo la temperatura superficial aún no
había subido de 14º C (J.A. García-Charton, observ. pers.);
la existencia de anomalías negativas, es decir, un retraso en
el calentamiento estival del agua de mar en un determinado
año, podría explicar que los meros, que usualmente son menos
visibles durante el invierno, bien sea por bajar a zonas más
profundas o por mostrar una movilidad reducida (Hackradt
2012), hubieran retrasado su comportamiento de aumento de
actividad en zonas someras hasta bien avanzado el verano en
el calendario. No obstante, esta hipótesis ha de ser
explorada en detalle, analizando las tendencias temporales de

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 29 -
la temperatura del agua superficial a partir de los datos de
las boyas oceanográficas más próximas, o bien con los
análisis y proyecciones de los organismos internacionales
encargados de tal labor (p. ej. el ‘Mediterranean ocean
Forecasting System – MFS’-http://gnoo.bo.ingv.it/mfs/myocean,
integrado en el EuroGOOS-http://www.eurogoos.org).
2) Las diferencias son achacables a una variabilidad temporal
natural a escala media (entre días o semanas), o bien a
diferencias instantáneas más dependientes de las condiciones
concretas de la mar en cada día de muestreo (oleaje,
corrientes, etc.) (Irigoyen et al. 2013). Esto podría ser
debido a la inadecuación de la forma y tamaño de la unidad
muestral utilizada (pasillos de 50 m de longitud por 5 m de
anchura) para capturar de un modo representativo las
abundancias y biomasas de especies de gran tamaño y/o cuya
movilidad sea mayor que el tamaño del transecto. Para
afrontar tal posibilidad, se impone combinar los seguimientos
tradicionales con otras técnicas de censo visual más
adaptadas a estas circunstancias.
En este último sentido, se ha estado probando una nueva
técnica de censo, con la que se realizan “censos completos”
mediante el método de distancia (Kulbicki & Sarramégne, 1999;
Oedekoven et al 2013), registrándose los ‘tracks’ de estos
recorridos completos mediante un GPS marino y el registro de la
posición de individuos y grupos de meros, así como de otras
observaciones, y de hitos estructurales del hábitat mediante una
cámara submarina (GoProTM) sincronizada con el GPS.
Simultáneamente, una sonda hidroacústica monohaz se encarga de
recoger la información topográfica del fondo, junto con la
localización GPS. La información derivada de la batimetría junto
con variables energéticas procedentes de los registros de la
sonda, permitirán realizar una primera aproximación a una
clasificación de hábitats bentónicos. Esta metodología ha sido
probada satisfactoriamente por Irigoyen, Sánchez-Carnero &
García-Charton en arrecifes rocosos de la Patagonia (Península
Valdés, Chubut, Argentina). Se propone pues desarrollar dicha
metodología en la reserva marina de Cabo de Palos – Islas
Hormigas, y compararla con la aproximación “clásica” basada en
transectos de 50×5 m2.

- 30 -
Figura 2.7 Meros en la reserva marina de Cabo de Palos – Islas Hormigas. FOTO: Carlos W. Hackradt

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 31 -
33 Uso del briozoo Myriapora truncata como
indicador del impacto del submarinismo
recreativo en la reserva marina de C.
Palos – I. Hormigas
3.1 Introducción
El buceo recreativo es la actividad que más se ha desarrollado
en la reserva marina de Cabo de Palos – Islas Hormigas. Según
datos oficiales, entre 1998 y 2010 (último año del que se tenga
un registro completo) el número de inmersiones se había
incrementado en un 225%, pasando de 8.142 inmersiones
registradas a más de 26.000 (Fig. 3.1). Es de suponer que estas
cifras han aumentado en los últimos años, habida cuenta de la
creación de nuevos centros de buceo en Cabo de Palos y del
desarrollo de los ya existentes; de continuar la progresión
observada en los últimos años, el número estimado actual
rondaría las 36.000 inmersiones (Fig. 3.1).
El buceo es una actividad que no se distribuye
uniformemente a lo largo del tiempo en la reserva marina, pues
es esencialmente estival (el 68% de las inmersiones anuales se
realizan en verano, y el 40% tienen lugar en los meses de julio
y agosto) (Fig. 3.2), y el resto del año se concentra en los
fines de semana (García-Charton et al. 2007).
Además, no se distribuye por igual entre los distintos
puntos de buceo, estando concentrado el 43% de las inmersiones
en el bajo de Dentro, y el 49% a partes iguales entre los bajos
de Piles I y Piles II, quedando el resto repartido entre un 5%
en el bajo de Fuera y el 3% para el bajo de la Testa (datos de
2010).

- 32 -
Figura 3.1 Número de inmersiones realizadas entre 1998 y 2010, según los datos oficiales, en la reserva marina de Cabo de Palos – Islas Hormigas (barras sombreadas), y proyección para los últimos años para los que no hay datos disponibles (barras punteadas) ajustada a una progresión exponencial (línea continua)) (Fuente: Servicio de Pesca y Acuicultura, CARM).
Figura 3.2 Número de inmersiones realizadas en cada mes durante el año 2010 en la reserva marina de Cabo de Palos – Islas Hormigas (Fuente: Servicio de Pesca y Acuicultura, CARM).
0
1 000
2 000
3 000
4 000
5 000
6 000
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Mes
Nº
de in
me
rsio
nes
0
1 000
2 000
3 000
4 000
5 000
6 000
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Mes
Nº
de in
me
rsio
nes
81428943 9103
10327
14918
1264511521
15066
17402
19434
21705
2618326388
29094
32170
35571
0
5000
10000
15000
20000
25000
30000
35000
40000
1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012 2013
Años
Nº
inm
ers
ion
es

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 33 -
El buceo recreativo es una de las formas más importantes de
uso comercial de las AMPs en el mundo, llevando aparejado, en
general, niveles variables de impacto sobre la biodiversidad –
especies y comunidades (Davis & Tisdell 1996; Dearden et al.
2007; Uyarra & Côté 2007). Una gran cantidad de estudios se han
aproximado al problema de medir el impacto del submarinismo
recreativo (p. ej., Hawkins & Roberts 1997; Rouphael & Inglis
2001; Garrabou et al. 1998; Hawkins et al. 2005; Hasler & Ott
2008). Los efectos biológicos locales pueden llegar a ser
dramáticos (Rouphael & Inglis 2001; Walters & Samways 2001; Rudd
& Tupper 2002; Zakai & Chadwick-Furman 2002; Barker & Roberts
2004; Coma et al. 2004).
Los buceadores con escafandra autónoma pueden afectar a los
organismos de varios modos, sea intencionadamente o no (Milazzo
et al. 2002; Uyarra & Côté 2007). Los daños son debidos
principalmente al contacto de aletas, equipos y el propio cuerpo
con organismos, por el efecto de las burbujas en cuevas y
extraplomos, o la resuspensión de sedimentos (Milazzo et al.
2002), pudiendo asimismo darse cambios en el comportamiento de
los peces (Kulbicki 1998; Milazzo et al. 2006) [véanse asimismo
los informes anteriores de seguimiento de la reserva marina de
Cabo de Palos – Islas Hormigas].
En el Mediterráneo se ha utilizado en algunas ocasiones
especies frágiles para cuantificar el impacto del buceo, tales
como briozoos (Sala et al. 1996; Garrabou et al. 1998) y
cnidarios (gorgonias: Coma et al. 2004; coral rojo: Linares et
al. 2011).
3.2 Objetivos
En el presente estudio se propone la utilización de la densidad
y características morfométricas del briozoo Myriapora truncata
(conocido como “falso coral”) como un indicador del posible
efecto de la frecuentación por buceadores en la reserva marina
de Cabo de Palos – Islas Hormigas.

- 34 -
3.3 Material y métodos
3.3.1 Especie de estudio: Myriapora truncata
Myriapora truncata (Pallas, 1766) es un briozoo (Gymnolaemata,
Cheilostomatida, Ascophorina, Myriaporidae) comúnmente conocido
como “falso coral”, de amplia distribución, que vive en una
variedad de hábitats rocosos, desde fondos someros submareales
de ambientes resguardados, a ambientes rocosos circalitorales,
tanto en el Mediterráneo como a lo largo de las costas
atlánticas del sur de España y Marruecos (Canu & Bassler 1925;
Gautier 1962; Ferreti et al. 2007). Crece en colonias erectas
arborescentes (zoarios) dotadas de un robusto esqueleto
(zoecio) formando ramas (Fig. 3.3) compuestas por zooides
indiferenciados (unidades alimentarias), dispuestos alrededor de
un amasijo de cenozoides (Berning 2007) con función estructural.
La pared frontal, o armadura, de M. truncata es secretada bajo
una cubierta de celoma y una envoltura externa de cutícula
orgánica. El cuerpo del animal, la polípida, se localiza en el
interior de la pared esquelética con forma de caja del zooide, y
se proyecta a través del orificio primario para alimentarse. La
polípida expandida tiene una corona de tentáculos (lofóforo)
dotados de cilios, los cuales crean una corriente de agua hacia
la boca, situada en la base del lofóforo. Cada tentáculo acaba
en un penacho de cilios rígidos, también presentes en los lados
abfrontal y latero-frontal del tentáculo, a los cuales se les ha
asignado una función sensorial en varias especies de briozoos
(Lombardi et al. 2011). Aunque existe alguna información sobre
la morfología de la larva, el primer zooide formado (ancéstrula;
Ferreti et al. 2007) y la tasa neta de calcificación (Rodolfo-
Metalpa et al. 2010), el ciclo de vida, la tasa de crecimiento
del zooide y la dieta de M. truncata permanecen pobremente
conocidos (Lombardi et al. 2011).

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 35 -
Figura 3.3 Zoario de Myriapora truncata en un fondo rocoso infralitoral [© Nano Sánchez / Wikimedia Commons]
Esta especie ha sido utilizada recientemente como
indicadora de cambios (paleo)ambientales (Berning 2007) y del
efecto de la acidificación marina (como consecuencia del cambio
climático) sobre los organismos bentónicos marinos (Lombardi et
al. 2011). El uso de Myriapora truncata como indicador del
impacto de submarinismo recreativo tiene su antecedente en el
trabajo de Calvisi et al. (2003) para el caso de las cuevas
submarinas, y, más recientemente, Guarnieri et al. (2012) citan
a esta especie como especialmente sensible y frágil al paso de
buceadores en cuevas submarinas. Ya en 1991, Pérez-Ruzafa et al.
destacaban la gran abundancia de esta especie en el área de Cabo
de Palos, lo que, junto a su carácter quebradizo y frágil ante
la presencia de buceadores la hace especialmente adecuada para
su uso como indicador del impacto del buceo recreativo en la
reserva marina.

- 36 -
3.3.2 Diseño y técnica de muestreo
El estudio se ha llevado a cabo en octubre de 2013, es decir
justo después de la temporada alta de buceo en la reserva marina
de Cabo de Palos – Islas Hormigas. Se consideraron dos niveles
de protección: la reserva integral (islas Hormiga y Hormigón, y
bajo del Mosquito), en la cual el submarinismo está prohibido, y
la reserva parcial (bajos de Dentro, Piles I y Piles II), en la
cual dicha actividad está permitida. En cada uno de esos niveles
de protección, se consideraron 2 franjas batimétricas (profunda
– 21-23 m, y somera – 12-14 m), y para cada combinación de nivel
de protección y franja batimétrica se buscaron 3 localidades
(separadas por cientos a miles de metros), en cada una de los
cuales se situaron aleatoriamente 3 sitios (separados por
decenas a cientos de metros entre sí); por último, en cada sitio
se lanzaron al azar 5 ‘quadrats’ de 1x1 m, en los cuales los
observadores, provistos de escafandras autónomas, contabilizaron
el número de zoarios, distinguiéndose entre los que estaban
vivos (coloreados y sujetos al sustrato) y los muertos (por
encontrarse despegados del sustrato y/o con una coloración
blanquecina). Con ello, en total se muestrearon 180 ‘quadrats’,
para cada uno de los cuales se registró la inclinación del
sustrato (en grados). Además, se seleccionaron las 6 colonias
(zoarios) más próximas a la esquina superior derecha de cada
‘quadrat’, para cada una de las cuales se midieron su altura y
anchura máximas, calculándose con estas medidas su área
proyectada (altura x anchura) y su relación de aspecto
(anchura:altura; un valor < 1 indica una forma más alta que
ancha, mientras que una relación de aspecto > 1 indica una
colonia más bien achatada, tanto más cuanto mayor sea ese
índice).
3.3.3 Análisis de datos
Las diferencias de densidad se han testado entre niveles de
protección [factor Reserva, R, fijo con 2 niveles (reserva
integral vs. reserva parcial)], Profundidad (P, con dos niveles
- somero vs. profundo, también fijo) y entre localidades (factor
L, aleatorio con 3 niveles, anidado en la interacción entre los
factores Reserva y Profundidad) y sitios (factor S aleatorio con
3 niveles, anidado en la localidad) mediante ANOVA mixto,
utilizándose para ello el programa GAD del paquete R (Sandrini-

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 37 -
Neto & Camargo 2011). La homogeneidad de las varianzas se ha
comprobado en todos los análisis utilizando el test de Cochran,
transformándose los datos en caso de no resultar homogéneas
dichas varianzas; en los casos en los que no se ha conseguido la
heterocedasticidad, por precaución, aquéllos factores con
valores de p > 0,01 habrán de ser interpretados con cautela
(Underwood 1997). Las diferencias significativas se testaron a
posteriori mediante un test de Student-Newman-Keuls (SNK). Para
el caso de la altura, anchura, área proyectada y relación de
aspecto de las colonias, a ese diseño se le ha añadido el factor
‘quadrat’ (aleatorio y anidado en Sitio, con 5 niveles),
realizándose el análisis mediante una PERMANOVA (análisis de la
varianza por permutaciones) debido a que, al faltar réplicas de
zoarios en algunos ‘quadrats’ (en los casos en los que la
densidad era < 6) la base de datos estaba desequilibrada
(situación no manejable fácilmente con un ANOVA paramétrico).
Las PERMANOVAs se realizaron sobre matrices de distancia
euclidea, testándose mediante 999 permutaciones, y aplicándose
además un test de Monte Carlo (Anderson et al. 2008).
3.4 Resultados
La densidad total de zoarios (colonias) del briozoo M. truncata
ha resultado mayor en la reserva integral que en la reserva
parcial, en la que el buceo recreativo está permitido, lo que da
lugar a la significación estadística del factor Reserva (Tabla
3.1). En esta y en las demás variables dependientes, el test de
Cochran ha resultado significativo (lo cual indica que la
ausencia de homocedasticidad), y las distintas transformaciones
intentadas no han homogeneizado las varianzas. Con independencia
de la profundidad por término medio se han encontrado 27,7
zoarios m-2 en los fondos que rodean las islas Hormigas, frente a
8,1 zoarios m-2 en los bajos de la reserva (Fig. 3.4).

- 38 -
Tabla 3.1 Resumen de los resultados del análisis de la varianza aplicados a las variaciones de densidad de Myriapora truncata (total, zoarios vivos y muertos) en función de los factores Reserva (R), Profundidad (P), Localidad (L) y Sitio (S) [gl: grados de libertad; MC: media cuadrática; F: F de Snedecor; p: p-valor]. Se indican los resultados del test de Cochran para cada variable dependiente [C: estadístico C de Cochran], en negrita aquéllos valores de p significativos (< 0,05), y en rojo los valores de p < 0,01.
Densidad total
Densidad zoarios vivos
Densidad zoarios muertos
Test de Cochran C = 0,269; p = 2,192e-07 C = 0,305; p = 6,794e-09 C = 0,254; p = 8,021e-07
Fuentes de variación
gl MC F p MC F p MC F p
R 1 17346,0 14,039 0,0057 17091,8 15,207 0,00455 0,8 0,042 0,8427
P 1 3100,0 2,509 0,1519 2599,2 2,313 0,16683 22,8 1,196 0,3060
RxP 1 1110,0 0,898 0,3710 871,2 0,775 0,40431 13,9 0,723 0,4178
L(RxP) 8 1235,6 2,269 0,0577 1124,0 2,879 0,02131 19,0 0,545 0,8109
S(L(RxP)) 24 544,7 2,246 0,0018 390,4 1,829 0,01607 34,9 3,605 8,61e-07
Residual 144 242,5 213,4 9,7
Figura 3.4 Diagrama de cajas de la densidad total de zoarios (colonias) de Myriapora truncata en la reserva integral y la reserva parcial de la reserva marina de Cabo de Palos – Islas Hormigas [la caja representa los cuartiles Q1 (25%) y Q3 (75%), de modo que su altura encierra el Rango Inter-Cuartil (RIC=Q3-Q1) en el que se encuentra el 50% de los datos); la línea gruesa indica la mediana de los datos; y las líneas punteadas delimitan los valores no atípicos – estando los valores atípicos (aquellos inferiores a Q1-1,5*RIC o superiores a Q3+1,5*RIC) representados mediante puntos]
INTEGRAL PARTIAL
02
04
06
08
010
0
Densidad de M. truncata
Nº
zo
ari
os m
-2
INTEGRAL PARCIAL
Nº
zo
ari
os
m-2
INTEGRAL PARTIAL
02
04
06
08
010
0
Densidad de M. truncata
Nº
zo
ari
os m
-2
INTEGRAL PARCIAL
Nº
zo
ari
os
m-2

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 39 -
Por otra parte, no hay un efecto significativo de la
profundidad, ya que, aunque en alguna localidad profunda hay una
densidad mayor que en el resto, el posible efecto de la
profundidad está enmascarado por una variabilidad marginalmente
significativa entre localidades (Fig. 3.5). Por último, se ha
detectado una variabilidad espacial significativa a escala fina
(entre sitios dentro de cada localidad) (Tabla 3.1).
Figura 3.5 Valores medios (± error típico) de densidad total de zoarios de Myriapora truncata en las distintas localidades prospectadas en las franjas somera y profunda de la reserva integral y la reserva parcial.
La variabilidad observada en la densidad total de falso
coral está determinada por la distribución de colonias vivas, ya
que cuando únicamente se consideran los zoarios vivos en los
análisis, se mantiene la influencia significativa del factor
Reserva, mientras que la densidad de zoarios muertos no varía
significativamente con el nivel de protección, aunque sí entre
sitios (Tabla 3.1).
Así, la densidad de zoarios vivos en la reserva integral es
de 24,9 m-2, mientras que en la reserva parcial la densidad
observada es de 5,4 zoarios vivos m-2 (Fig. 3.6 y 3.7). También
en este caso la variabilidad espacial a escala fina (entre
sitios separados decenas de metros) es significativa (Tabla
3.1).
0
10
20
30
40
50
60
Nº
zo
ari
os
m-2
INTEGRAL PARCIAL
PROFUNDO SOMERO PROFUNDO SOMERO

- 40 -
Figura 3.6 Diagrama de cajas de la densidad total de zoarios (colonias) vivos de Myriapora truncata en la reserva integral y la reserva parcial de la reserva marina de Cabo de Palos – Islas Hormigas [ver Fig. 3.2 para una descripción de esta gráfica]
Figura 3.7 Valores medios (± error típico) de densidad total de zoarios vivos de Myriapora truncata en las distintas localidades prospectadas en las franjas somera y profunda de la reserva integral y la reserva parcial.
Nº
zo
ari
os
m-2
INTEGRAL PARCIAL
PROFUNDO SOMERO PROFUNDO SOMERO
0
10
20
30
40
50
60
INTEGRAL PARTIAL
020
40
60
80
100
Densidad de M. truncata
Nº
zo
ari
os m
-2
INTEGRAL PARCIAL
Nº
zo
ari
os
m-2

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 41 -
Tabla 3.2 Resumen de los resultados del análisis PERMANOVA aplicados a las variaciones morfométricas de los zoarios de Myriapora truncata en función de los factores Reserva (R), Profundidad (P), Localidad (L), Sitio (S) y Quadrat (Q) [gl: grados de libertad; MC: media cuadrática; Pseudo-F: pseudo-F de Snédecor; p(perm): p-valor obtenido mediante permutaciones; perms: nº de permutaciones; p(MC): p-valor obtenido mediante el test de Monte Carlo]. Se indican en rojo los valores de p < 0,05.
Fuente de variación
gl MC Pseudo-F p(perm)
perms p(MC)
ANCHO
R 1 2,463 0,11836 0,779 999 0,745
P 1 44,558 2,1413 0,19 998 0,175
RxP 1 3,8428 0,18468 0,705 996 0,676
L(RxP) 8 26,276 1,591 0,166 998 0,183
S(L(RxP)) 24 17,86 3,2217 0,002 999 0,001
Q(S(L(RxP))) 118 687,5 1,6066 0,002 998 0,001
Residual 609 3,6264
ALTO
R 1 3,4858 0,9137 0,354 998 0,350
P 1 0,22395 5,87E-02 0,818 997 0,816
RxP 1 9,87e-02 2,59e-02 0,876 997 0,873
L(RxP) 8 4,596 1,0533 0,428 997 0,450
S(L(RxP)) 24 4,6856 2,3886 0,008 999 0,002
Q(S(L(RxP))) 118 2,0718 1,7064 0,001 999 0,001
Residual 609 1,2141
AREA
R 1 46,995 0,15046 0,686 995 0,682
P 1 191,27 0,61237 0,439 998 0,480
RxP 1 11,092 3,55e-02 0,859 997 0,872
L(RxP) 8 389,36 1,5148 0,217 997 0,188
S(L(RxP)) 24 276,43 3,002 0,001 998 0,001
Q(S(L(RxP))) 118 94,935 1,3051 0,053 998 0,025
Residual 609 72,739
RATIO DE ASPECTO
R 1 0,27527 0,12619 0,744 998 0,740
P 1 12,23 5,6063 0,037 998 0,038
RxP 1 1,65e-04 7,57e-05 0,994 999 0,989
L(RxP) 8 2,6952 1,3802 0,258 998 0,249
S(L(RxP)) 24 2,0983 2,6807 0,006 999 0,001
Q(S(L(RxP))) 118 96,393 1,4822 0,028 998 0,001
Residual 609 0,55113

- 42 -
En lo que respecta a las variaciones morfométricas de los
zoarios, únicamente el ratio de aspecto (anchura:altura)
presenta variaciones significativas entre profundidades (Tabla
3.2), siendo el aspecto de los zoarios más achatado en las zonas
someras (1,51 en promedio) que en las profundas (1,30). Por otra
parte, todas las características morfométricas de las colonias
han mostrado una variabilidad espacial significativa a escala
fina (entre sitios y entre ‘quadrats’) (Tabla 3.2).
3.5 Discusión
El presente estudio pone en evidencia que en la reserva integral
de la reserva marina de Cabo de Palos – Islas Hormigas (es
decir, en los fondos que rodean las islas Hormigas y en el bajo
del Mosquito), en la cual el buceo recreativo no se practica por
estar prohibido, la densidad de zoarios vivos de Myriapora
truncata es 5,6 veces mayor que en la reserva parcial (bajos de
Dentro y Piles), con independencia de la profundidad, así como
una tendencia no significativa a una mayor densidad en la franja
más profunda, con independencia del nivel de protección; por
otra parte, se aprecia una variabilidad espacial significativa a
escala espacial fina (entre sitios alejados cientos de metros
entre sí). Finalmente, no se han observado diferencias
morfométricas de las colonias en función del nivel de protección
o la profundidad, salvo un aspecto ligeramente más achatado (es
decir, una anchura mayor que la altura) de los zoarios que viven
en áreas más someras, en comparación con las que se midieron en
la franja profunda, acaso una respuesta un mayor hidrodinamismo
en las zonas más someras.
Dado que las condiciones ambientales de los lugares de
muestreo, estén éstos protegidos o no de la actividad turística
subacuática, tales como orientación frente a los vientos y
oleajes dominantes, inclinación del sustrato, cobertura algal,
rugosidad del fondo, etc., son comparables en todas las
localidades prospectadas, cabe inferir que es la actividad
humana, y más concretamente la sobrefrecuentación por
buceadores, la causante de las diferencias observadas.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 43 -
Los presentes resultados muestran una probable afección de
la presión por buceo sobre una especie frágil, utilizada aquí
como indicadora de impacto. Se trata del indicio más claro
obtenido hasta la fecha de dicho efecto erosivo. En efecto, el
problema del impacto del submarinismo en la reserva marina de
Cabo de Palos – Islas Hormigas se abordó por vez primera en 2002
(Pérez-Ruzafa et al. 2002), y desde entonces se han ido
realizando diversos estudios, utilizándose diferentes técnicas
de muestreo (muestreo visual con escafandra autónoma,
seguimiento fotográfico, aproximación experimental, seguimiento
del comportamiento erosivo de los buceadores, etc.); y aunque,
como se revisará a continuación, se han obtenido algunos
indicios de efectos erosivos, hasta ahora no había habido un
síntoma tan evidente de posibles efectos de la actividad
subacuática recreativa.
En 2002 se realizó el primer estudio de las diferencias de
cobertura y densidad de algas e invertebrados como indicadores
de erosión debida a la frecuentación por buceadores, y se inició
un estudio del comportamiento de los deportistas recreativos en
la reserva marina. En aquel estudio, en el que se comparaban una
serie de indicadores mediante prospecciones visuales en círculos
de 25 m2 entre antes y después de la temporada alta de buceo
(verano), ya se observó un menor desarrollo algal, con pérdida
de estructura asociada a las algas arbustivas y en un aumento de
los fondos desprovistos de cobertura algal en las zonas
visitadas por los submarinistas; sin embargo, la “señal”
(magnitud del efecto y significación estadística de las
diferencias detectadas) era tenue en el caso de las algas,
debido a la gran variabilidad natural a escalas espaciales finas
(entre sitios separados por cientos a miles de metros, y entre
réplicas separadas por decenas a cientos de metros), y al hecho
de que las comparaciones antes-después coinciden con una
variación estacional. Por su parte, ninguna de las diferencias
de densidad o cobertura de invertebrados bentónicos fue
achacable a la acción erosiva de los buceadores, aunque fue
notable un mayor número de colonias de Eunicella singularis que
aparecieron tumbadas en las zonas de buceo, con un incremento
importante tras la temporada alta de inmersiones.

- 44 -
Por otra parte, el estudio de comportamiento erosivo de los
buceadores en la reserva se culminó al año siguiente (Pérez-
Ruzafa et al. 2003); en este estudio se monitoreó el
comportamiento de los buceadores y su interacción con el fondo,
siguiéndose a 342 personas durante los veranos de 2002 y 2003,
con edades comprendidas entre los 15 y los 67 años. Los
principales resultados de este estudio fueron que el aleteo
constituyó la acción que más daños generó en el fondo,
alcanzando una media de 6,3 impactos por buceador durante los 10
minutos de seguimiento, aunque presentando una gran variabilidad
entre personas, que iba de 1 a 75 interacciones. Los contactos
con el equipo alcanzaron una media de 4,4 por buceador, mientras
que los apoyos con el fondo (que provocara alguna alteración en
el mismo) se produjeron con una frecuencia media de 3,4 veces
por cada 10 minutos para cada buceador. Un factor independiente
del buceador que parece influir en el comportamiento de los
buceadores es la presencia o no de guía durante la inmersión. En
general todas las acciones se ven disminuidas con la presencia
de guía, aunque sólo el aleteo presenta diferencias
significativas (p < 0,05) (Fig. 3.8). Un factor, también
independiente del buceador, que influyó significativamente en el
comportamiento individual bajo el agua es la celebración de un
‘briefing’, considerado éste como el conjunto de normas y
recomendaciones que se deben dar a los buceadores en la
embarcación antes de iniciar la inmersión. En la mayoría de los
casos suele hacer referencia a cuestiones técnicas del buceo
como flotabilidad, consumo de aire, buceo en parejas, itinerario
a recorrer durante la inmersión, profundidad, etc. y en muy
pocos casos incluyó consideraciones de tipo ambiental. A pesar
de ello, se observó una disminución de todas las acciones que
provocan algún tipo de impacto en el fondo cuando previamente ha
habido un ‘briefing’ por parte del instructor en la embarcación
o en el centro justo antes de embarcar, aunque sólo presentaban
diferencias significativas el número de aleteos (p < 0,05) (Fig.
3.8). Asimismo, se observó que el manejo de cámaras aumentaba
significativamente el número de interacciones del buceador con
el fondo.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 45 -
Figura 3.8 Diferencias en el número medio de aleteos realizados por un buceador durante 10 minutos que produzca algún efecto erosivo en el fondo (± error típico) en función de si (a) la inmersión se hace con guía o no y (b) ha habido un ‘briefing’ antes de la inmersión o no lo ha habido (tomado de Pérez-Ruzafa et al. 2003).
En este mismo informe y en años posteriores (Pérez-Ruzafa
et al. 2003, 2004; García-Charton et al. 2005, 2006, 2007, 2008)
se siguieron estudiando visualmente en inmersión con escafandra
autónoma las abundancias de invertebrados bentónicos
(incorporando algunas especies indicadoras adicionales a las
observadas el año anterior) y de coberturas algales, no
llegándose a detectar en ellos efectos claros del submarinismo
recreativo, aunque invertebrados frágiles como la esponja
Dysidea fragilis, el cnidario Cladocora caespitosa y el briozoo
Pentapora fascialis mostraron una tendencia a disminuir sus
abundancias tras la temporada de buceo, sin que estos cambios
resultaran concluyentes. Por su parte, se seguía observando una
mayor proporción de gorgonias tumbadas en los fondos de la
reserva parcial (bajos) (Fig. 3.9). Una observación novedosa fue
la tendencia de los erizos comestibles (Paracentrotus lividus) a
ser más numerosos en las zonas más someras de los bajos en los
cuales el buceo estaba permitido (Fig. 3.10), que en los de la
reserva integral, sin que ello tuviera que ser achacado
necesariamente a la acción erosiva de los buceadores.
(a) (b)(a) (b)

- 46 -
Figura 3.9 Variación espacio-temporal de la relación de colonias de gorgonia blanca (Eunicella singularis) tumbadas respecto al de erectas, comparando lo observado en la isla Hormiga (H) y el Hormigón (HG) con lo que se ha medido en la cueva (CV) y la cara norte (N) del bajo de Dentro (tomado de Pérez-Ruzafa et al. 2004).
Figura 3.10 Valores medios de densidad de erizos en las localidades (BD: Bajo de Dentro; BP: Bajo de Piles; BF: Bajo de Fuera; H1: Hormiga 1; H2: Hormiga 2; HO: Hormigón) y tiempos prospectados en el año 2005 (tomado de García-Charton et al. 2005).

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 47 -
Los valores de densidad de Paracentrotus lividus obtenidos
en 2005 en la reserva integral coinciden con los observados por
Coma et al. (2011) para ese mismo año. Sin embargo, esos mismos
autores dan cuenta de un aumento de la densidad de este erizo en
el archipiélago de las islas Hormigas a partir de 2007,
achacable al éxito del reclutamiento; ello introduciría un
factor de variabilidad temporal que explicaría que no todos los
años se detectaran las diferencias de densidad entre la reserva
integral y la reserva parcial, por lo que tampoco constituye un
efecto claro de la sobrefrecuentación subacuática.
También en 2004 y 2005 se realizaron trabajos
experimentales (Pérez-Ruzafa et al. 2004; García-Charton et al.
2005, 2008) que proporcionan una demostración del mecanismo que
opera con el paso de buceadores en parcelas controladas. Por un
lado, se demostró que el paso de buceadores provoca una pérdida
perceptible de cobertura algal y colonias completas del briozoo
Myriapora truncata (Fig. 3.11). Además, se midió un aumento de
la sedimentación en áreas en las que se bucea, en comparación
con áreas en las que no se bucea (Fig. 3.12).
Figura 3.11 Pérdida media (± error típico) de densidad de zoarios vivos de Myriapora truncata en función del aumento de la intensidad de paso por buceadores sobre parcelas experimentales de 1 m
2 en la reserva marina de Cabo
de Palos – Islas Hormigas en fondos planos (barras blancas) e inclinados (barras grises) (tomado de García-Charton et al. 2008).
Nº
zo
ari
os p
erd
ido
s
0 5 10 25
Nº de pasadas
0
0,5
1,0
1,5
2,0

- 48 -
Figura 3.12 Peso medio del sedimento resuspendido (corregido con el control) para cada intensidad de buceo (nº de pasadas sobre los captadores). Se indica la recta de regresión (línea continua) (R2=0,84, P<0,05) y su extrapolación para una pasada (línea punteada). Las barras de error indican el error típico de la media.
En estos estudios se llevó a cabo asimismo una metodología
basada en la realización de ‘fotoquadrats’ para la medida de
cobertura o densidad de algas e invertebrados. En general, se
han corroborado los resultados obtenidos mediante aproximaciones
visuales, en el sentido de que las algas no indican efecto
erosivo alguno, como consecuencia de su fuerte dinámica
estacional, aunque se detecta una tendencia a una mayor
proporción de fondo desprovisto de vegetación, así como más
erizos y menos falso coral en la reserva parcial (Fig. 3.13).
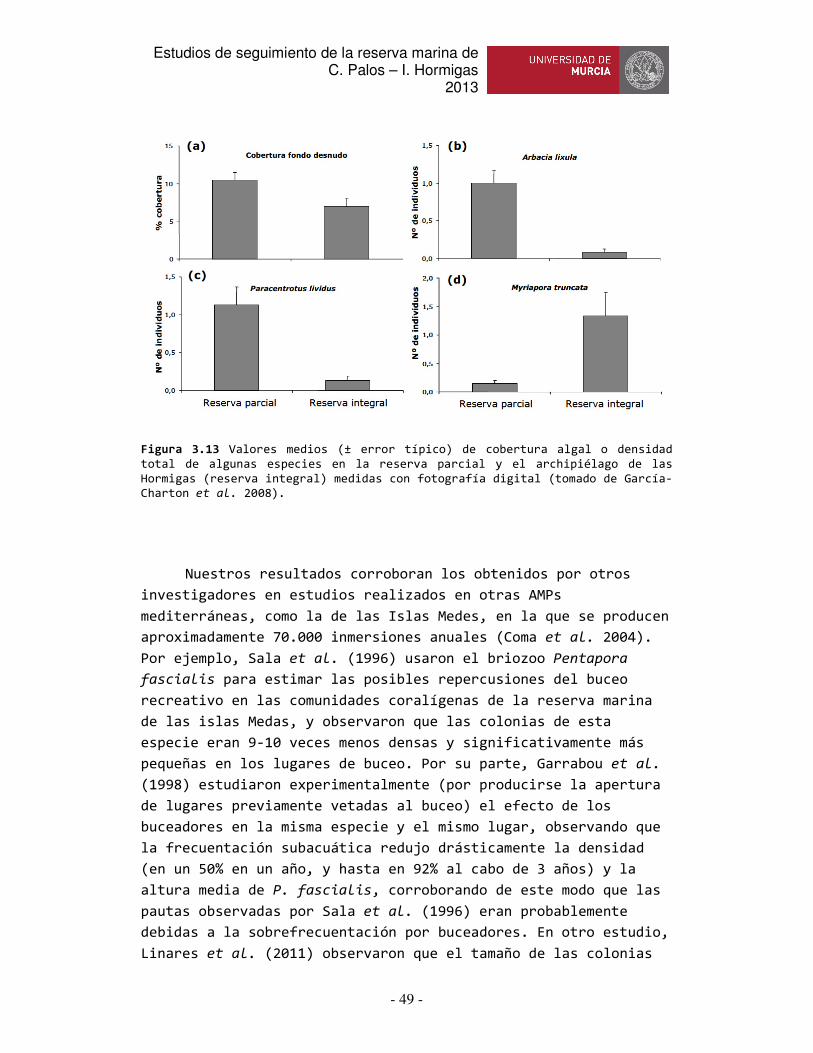
Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 49 -
Figura 3.13 Valores medios (± error típico) de cobertura algal o densidad total de algunas especies en la reserva parcial y el archipiélago de las Hormigas (reserva integral) medidas con fotografía digital (tomado de García-Charton et al. 2008).
Nuestros resultados corroboran los obtenidos por otros
investigadores en estudios realizados en otras AMPs
mediterráneas, como la de las Islas Medes, en la que se producen
aproximadamente 70.000 inmersiones anuales (Coma et al. 2004).
Por ejemplo, Sala et al. (1996) usaron el briozoo Pentapora
fascialis para estimar las posibles repercusiones del buceo
recreativo en las comunidades coralígenas de la reserva marina
de las islas Medas, y observaron que las colonias de esta
especie eran 9-10 veces menos densas y significativamente más
pequeñas en los lugares de buceo. Por su parte, Garrabou et al.
(1998) estudiaron experimentalmente (por producirse la apertura
de lugares previamente vetadas al buceo) el efecto de los
buceadores en la misma especie y el mismo lugar, observando que
la frecuentación subacuática redujo drásticamente la densidad
(en un 50% en un año, y hasta en 92% al cabo de 3 años) y la
altura media de P. fascialis, corroborando de este modo que las
pautas observadas por Sala et al. (1996) eran probablemente
debidas a la sobrefrecuentación por buceadores. En otro estudio,
Linares et al. (2011) observaron que el tamaño de las colonias

- 50 -
de coral rojo (Corallium rubrum) en las islas Medes fue
significativamente menor que el pronosticado por los modelos de
crecimiento y más pequeño que el de las colonias en áreas
marinas protegidas sin buceo en las que esta especie está
presente (Banyus, Carry-le-Rouet, Scandola); además, el número
anual de buceos recreativos y el cambio porcentual en el
diámetro basal de las colonias de coral rojo se correlacionaron
negativamente, lo que sugiere que la abrasión causada por buzos
habría podido incrementar las tasas de mortalidad de la especie.
En el caso de la reserva marina de Cabo de Palos – Islas
Hormigas estaríamos, por tanto, ante una señal de que la gran
cantidad de buceadores podría estar socavando la biodiversidad
local, a través del efecto erosivo de equipos, linternas y
cámaras, y principalmente de las aletas sobre el fondo. El
problema se agrava por el hecho de que en la reserva marina
únicamente se cuenta con 5 puntos de buceo, en los que se
concentra toda la presión de las inmersiones a lo largo del año.
Se impone una limitación precautoria en el uso recreativo
de la reserva marina de Cabo de Palos – Islas Hormigas, en el
sentido de no aumentar la presión de buceo por encima de los
niveles actuales de frecuentación subacuática recreativa
(estimada en alrededor de 36.000 inmersiones anuales). Para
ello, la administración competente, en colaboración con las
entidades interesadas (centros de buceo), y con el acuerdo de
los demás usuarios de la reserva marina (sobre todo pescadores
artesanales) habrá de arbitrar las necesarias medidas de
gestión. Son cada vez más numerosas las iniciativas a nivel
internacional, como el código de conducta ‘Green Fins’ (Hunt et
al. 2013), que pueden mostrar el camino para llevar a cabo con
éxito dicha regulación. Estas medidas tendrán que ser
implementadas mediante un proceso complejo y bien diseñado (Ku &
Chen 2013), basado en los presupuestos de la co-gestión y la
participación pública efectiva, contando con todos los actores y
usuarios de la reserva marina (Hogg et al. 2013).

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 51 -
44 La pesca artesanal en la reserva marina
de C. Palos – I. Hormigas
4.1 Introducción
Las pesquerías marinas globales han tenido un gran desarrollo y
han sufrido grandes cambios desde que la FAO (Organización de
las Naciones Unidas para la Alimentación y la Agricultura),
empezó a recolectar datos pesqueros en 1950, aumentando
enormemente su tamaño, capacidad de almacenamiento y de
autonomía (Pauly et al. 2002). Las capturas mundiales alcanzaron
su pico máximo en 1996 con un total de 86,4 millones de
toneladas de pescado desembarcadas y desde entonces han estado
declinando hasta estabilizarse en aproximadamente 80 millones de
toneladas anuales. El mar Mediterráneo y el mar Negro presentan
una tendencia de disminución en la captura total de pescado
desde mediados de 1980. En 2009 estos mares tenían un 33% de sus
poblaciones de peces plenamente explotadas y un 50%
sobreexplotadas (FAO 2012, Froese et al. 2012).
La disminución de los ‘stocks’ pesqueros, así como los
efectos derivados de los demás impactos de las actividades
humanas (Worm et al. 2009) motivaron cambios en las medidas de
gestión con el fin de mitigar e intentar revertir los efectos
deletéreos del ser humano sobre el medio ambiente marino
(Lubchenco et al. 2003, Fox et al. 2012). Las Áreas Marinas
Protegidas – AMPs – han sido creadas para promover estos cambios
e incrementar la protección de la biodiversidad marina y la
conservación de los recursos marinos (Lubchenco et al. 2003,
Pauly et al. 2005, Claudet et al. 2008) así como las actividades
pesqueras y socioculturales relacionadas con el medio ambiente
marino. Entretanto, actualmente las AMPs protegen alrededor de
1% de la superficie global de los océanos (Cullis-Suzuki et al.
2010). En aguas europeas las reservas marinas, un tipo de AMP
donde existe al menos un área con extracción totalmente
prohibida, muestran evidencias de que están promoviendo la
recuperación de la biodiversidad local, siendo así una

- 52 -
importante herramienta de gestión basada en el ecosistema
(Halpern et al. 2004, García-Charton et al. 2008, Katsanevakis
et al. 2011, Fenberg et al. 2012). Entre los efectos positivos
esperados en las áreas marinas protegidas están el incremento de
la biomasa de las especies objetivo, la exportación de biomasa a
zonas adyacentes, el aumento de la biodiversidad, la protección
de hábitats, la mejora de los rendimientos pesqueros, los
beneficios de las actividades recreativas, la reducción de los
conflictos entre usuarios y la mejora de los mecanismos de
participación y gestión del área marina (Pelletier et al. 2005).
En relación a las pesquerías, las reservas marinas, cuando
están bien establecidas y gestionadas, generan beneficios
directos e indirectos. Goñi et al. (2008), en un estudio en 6
AMPs en el mar Mediterráneo, mostraron que las tasas de captura
de las pesquerías artesanales aumentan a medida que se aproximan
a las reservas marinas. El tamaño de la reserva también influye
en la tasas y en los tipos de captura, ya que reservas
integrales mayores tienden a generar mayores cantidades de
capturas y aumentar las capturas de las especies objetivo
(Vandeperre et al. 2011).
Las pesquerías artesanales se caracterizan por utilizar
caladeros cercanos a su puerto base, con barcos pequeños donde
trabajan pocos pescadores con una gran variedad de artes de
pesca (Guyader et al. 2013), que incluyen redes de trasmallo,
redes de enmalle, palangre, cadufos y trampas (Colloca et al.
2004, FAO 2011). En el Mediterráneo europeo los barcos
artesanales representan más de un 80% de la flota, y en 2010
representaban más de un 85% de la flota mundial (Berthou et al.
2005, Battaglia et al. 2010, FAO 2012). En innumerables países
las pesquerías artesanales tienen una larga tradición cultural y
una gran importancia socio-económica (Stergiou et al. 2006,
Crilly et al. 2013).
La actividad pesquera en la zona del cabo de Palos se lleva
realizando desde la antigüedad con artes similares a los
actuales. La flota artesanal de Cabo de Palos concentra su faena
dentro y en el entorno la Reserva Marina de Cabo de Palos –
Islas Hormigas (Fig. 1.1) y tiene como objetivo una gran
variedad de especies, valiéndose de varias artes de pesca para
capturarlas (García 1997, Esparza-Alaminos 2010). El uso de esas

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 53 -
artes se cambia a lo largo del año en función de la biología de
las especies, del valor y demanda del mercado, de las épocas de
veda y condiciones del mar, entre otros factores (García-Charton
et al. 2010).
4.2 Objetivos
El objetivo del presente capítulo es describir la distribución
del esfuerzo pesquero realizado en el año de 2013 por la flota
artesanal de Cabo de Palos, así como actualizar la
caracterización de la variación temporal a largo plazo en las
capturas por unidad de esfuerzo (CPUE) y en los ingresos por
unidad de esfuerzo (€PUE) a partir de los datos registrados en
lonja de la flota artesanal que opera en la reserva marina de
Cabo de Palos – Islas Hormigas antes y después de su creación,
hasta el año 2012, último para el cual se cuenta con el registro
completo en lonja.
4.3 Material y métodos
4.3.1 Área de estudio
El cabo de Palos (37°38’N; 00°42’O) y el archipiélago de las
Islas Hormigas, situados en el sudeste la península Ibérica
(Mediterráneo sur-occidental), tiene una elevada biodiversidad.
Por ser un área submarina singular en el Mediterráneo occidental
(García-Charton & Marcos 2004), y con el fin de proteger los
recursos marinos que se desarrollan en la zona, en 1995 se creó
la Reserva Marina de Interés Pesquero de Cabo de Palos-Islas
Hormigas, comprendiendo un área total de 1934 hectáreas (Fig.
4.1).
La reserva está dividida en dos áreas con figuras de
protección distintas, una zona con protección integral y otra
con protección parcial. La reserva integral, con 270 ha de
superficie, está localizada al entorno de las islas Hormigas, en
este área está prohibida cualquier actividad profesional o
recreativa tanto extractiva como deportiva, salvo la
investigación científica. A su alrededor está situada la reserva
parcial, donde están regulados la pesca artesanal para la flota
basada en el puerto de Cabo Palos y el buceo recreativo. La

- 54 -
pesca recreativa está explícitamente prohibida en toda el área
de la AMP. La reserva marina de Cabo de Palos tiene su gestión
compartida entre la Secretaria General de Pesca Marítima en la
zona de aguas exteriores y la Consejería de Agricultura y Agua
de la Región de Murcia en la zona de aguas interiores (García-
Charton et al. 2010). El área de estudio está limitada por la
zona de actividades de la flota artesanal que tiene como puerto
base el Puerto de Cabo de Palos y permiso para faenar en la
Reserva Marina de Interés Pesquero de Cabo de Palos.
Figura 4.1 Delimitación del área de estudio incluido en el estudio pesquero, con indicación de los límites de la Reserva Marina de Interés Pesquero de Cabo de Palos y la localización del puerto de Cabo de Palos.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 55 -
4.3.2 Recogida y análisis de datos
Los datos fueran obtenidos a partir de los registros históricos
de capturas declaradas en la lonja de la Cofradía de Pescadores
de Cartagena, de la bibliografía disponible (García 1997,
Esparza-Alaminos 2010) y de los registros oficiales del Servicio
de Pesca Marítima de la Consejería de Agricultura y Agua de la
Región Autónoma de Murcia. Fue posible recoger informaciones
para el período entre los años de 1993 a 2012, excluyendo los
años 1997 y 1998, para los cuales no hay datos disponibles.
Los datos publicados por García (1997) son la única fuente
de información para los años cercanos a la creación de la
Reserva Marina de Cabo de Palos-Islas Hormigas, entre 1993-1996
y los datos de entre 1999 y 2009 fueran facilitados por Esparza-
Alaminos (2010). De forma general, el conjunto de datos
proporcionaron informaciones sobre: (1) el nombre del barco, (2)
el nombre común de la especie capturada, (3) la cantidad de
pescado subastado (en kg) en lonja, (4) el valor medio bruto (en
€) de la venta del pescado en lonja y (5) el valor de ingreso
total (€) por cada venta. La lista de embarcaciones con licencia
para pescar en la Reserva Marina de Cabo de Palos fue obtenida
de la legislación vigente (Resolución de 22 de febrero de 2010,
de la Secretaría General del Mar, por la que se actualiza el
censo de embarcaciones autorizadas a ejercer la pesca marítima
profesional en la reserva marina del entorno de Cabo de Palos-
Islas Hormigas). Las características técnicas de las
embarcaciones se han obtenido a partir del censo de
embarcaciones del Ministerio de Agricultura, Alimentación y
Medio disponibles en su sitio web (www.magrama.gob.es), y se han
validado con cuestionarios a los patrones de las embarcaciones.
La captura por unidad de esfuerzo (CPUE) se ha calculado a
través del cociente entre la suma de la captura anual declarada
de la flota de Cabo de Palos y la suma de la potencia de las
embarcaciones en kilovatios (kW). En el cálculo del importe por
unidad de esfuerzo (€PUE) se ha dividido la suma de los importes
de cada año por el total de embarcaciones de Cabo de Palos en
cada año.
No hay registro histórico de los valores de la venta en
lonja para los años anteriores a 2001, por lo que únicamente ha

- 56 -
sido posible obtener la €PUE para el intervalo 2001-2012. Para
el cálculo del €PUE los valores de venta en lonja del pescado
fueran corregidos para valores constantes de 2001 a través de
ajuste por la inflación interanual en la Región de Murcia, cuyos
valores anuales fueron obtenidos por medio del sitio web del
Instituto Nacional de Estadística (www.ine.es).
Los datos fueron analizados mediante regresiones lineales.
En los cálculos de captura e importe se han excluido los
registros de especies que no son capturadas por ningún arte
artesanal pero que son subastadas en la lonja de Cartagena por
las flotas de arrastre, palangre y cerco de altura basadas en el
Puerto de Cartagena. Estas especies son (1) el atún, Thunnus
thynnus (Linnaeus, 1758); (2) el emperador o pez-espada, Xiphias
gladius (Linnaeus, 1758); (3) la gamba roja, Aristeus antennatus
(Risso, 1816); (4) el langostino, Penaeus kerathurus (Forsskål,
1775); (5) la sardina, Sardina pilchardus (Walbaum, 1792); (6)
la alacha, Sardinella aurita (Valenciennes, 1847); y todos los
moluscos con concha.
Asimismo, se describió el cambio en las capturas totales
para las especies positivamente afectadas por la regulación
pesquera en la reserva marina (ver capítulo 1 del presente
informe) y aquéllas más propias de los fondos rocosos del área
protegida: mero, Epinephelus marginatus (Lowe, 1834); gallineta
o cabracho, Scorpaena scrofa (Linnaeus, 1758); dentón, Dentex
dentex (Linnaeus, 1758); corva, Sciaena umbra (Linnaeus, 1758);
sargos, Diplodus spp.; y espetón o barracuda, Sphyraena
viridensis (Cuvier, 1829).
Para la determinación de la distribución del esfuerzo
pesquero fueran realizados encuestas y embarques en la flota de
artesanal de Cabo de Palos entre los meses de mayo y noviembre
de 2013. Las bases cartográficas utilizadas en la creación de
los mapas fueran obtenidas del archivo del Grupo de Ecología y
Hidrología de la Universidad de Murcia. A partir de los puntos
de pesca obtenidos fue estimada la densidad del esfuerzo
pesquero en el área de estudio con la aplicación del cálculo de
Estimación de Densidad Kernel a través de la extensión de
análisis espaciales del software QGIS 2.0. El estimador de
Kernel crea alrededor de cada punto de muestreo un área
circular, dividida en ‘pixels’, que corresponde al radio de

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 57 -
influencia del punto. El valor del resultante de la estimación
es calculado para cada ‘pixel’ a través de la suma de los
valores Kernel superpuestos y divididos por el área del radio de
influencia. La variación en estos valores está representada en
el mapa por variaciones de color, siendo el verde zonas de menor
densidad, naranjas de densidad media y rojas de alta densidad.
En este estudio se ha considerado como radio de influencia la
longitud media de las artes de pesca de la flota de Cabo de
Palos, estimada por Esparza-Alaminos (2010) en 1000 metros.

- 58 -
4.4 Resultados
4.4.1 Distribución espacial del esfuerzo pesquero
en 2013
Las encuestas y embarques para el estudio de la distribución del
esfuerzo pesquero han sido realizadas entre los meses de mayo y
noviembre de 2013. Se han registrado 7 de las 11 distintas artes
de pesca descritas a lo largo del seguimiento de las pesquerías
en el entorno de la reserva marina: trasmallo claro, trasmallo
fino, red de nailon, cadufo, palangre, chirretera y moruna, de
estos el trasmallo claro (61,7%) y el cadufo (14%) representaran
75,7% del esfuerzo total.
Figura 4.2 Distribución espacial de la densidad del esfuerzo pesquero de la flota artesanal del puerto de Cabo de Palos, considerando todas las artes de pesca juntas.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 59 -
La mayor densidad del esfuerzo está concentrada (Fig. 4.2)
en la zona sudoeste de la reserva marina, un área donde se
encuentran bajos y praderas de Posidonia oceanica. Las zonas con
intensidad media al norte del límite de la reserva se
distribuyen sobre la isobata de los 15 metros en fondo
recubierto por P. oceanica. Al sur de la reserva marina se
localiza otra zona de esfuerzo alto, en las cercanías de
Calbanque, relacionada con la agregación del esfuerzo de la
pesca con cadufos y trasmallo fino (Fig. 4.2).
El trasmallo claro ha sido el arte más empleado por la
flota artesanal, habiendo estado concentrado su uso durante el
período de estudio en las proximidades de los bajos de Dentro,
Piles y la Testa, es decir dentro de la Reserva Marina de Cabo
de Palos – Islas Hormigas(Fig. 4.3).
Figura 4.3 Distribución espacial de la densidad del esfuerzo pesquero realizado con trasmallo claro por la flota artesanal del puerto de Cabo de Palos en 2013.

- 60 -
La pesca con cadufos, que tiene el pulpo (Octopus
vulgaris) como especie objetivo, ha estado predominantemente
localizada al sur de la reserva marina, a una profundidad
promedio de 25 m (Figura 4.4).
Figura 4.4 Distribución espacial de la densidad del esfuerzo pesquero realizado con cadufo por la flota artesanal del puerto de Cabo de Palos en 2013.
Al igual que el cadufo, el esfuerzo pesquero con trasmallo
fino (Fig. 4.5) también es mayor en las cercanías de Calblanque,
y rara vez ha sido empleado dentro de la reserva marina.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 61 -
Figura 4.5 Distribución espacial de la densidad del esfuerzo pesquero realizado con trasmallo fino por la flota artesanal del puerto de Cabo de Palos en 2013.
Por su parte, la red de nailon (Fig. 4.6), el palangre
(Fig. 4.7) y la charretera (Fig. 4.8) han sido poco empleadas
durante este año, de modo que los puntos de muestreo están
dispersos en el área de estudio, con lo que la distribución del
esfuerzo no es muy clara en los correspondientes mapas de
distribución. La moruna es un arte empleada solamente por un
barco, y su localización se quedó fija en todo el periodo de
estudio, por lo que su esfuerzo está limitado a un punto
específico – Cala Túnez (Fig. 4.9).

- 62 -
Figura 4.6 Distribución espacial de la densidad del esfuerzo pesquero realizado con red de nailon por la flota artesanal del puerto de Cabo de Palos en 2013.
Figura 4.7 Distribución espacial de la densidad del esfuerzo pesquero realizado con palangre por la flota artesanal del puerto de Cabo de Palos en 2013.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 63 -
Figura 4.8 Distribución espacial de la densidad del esfuerzo pesquero realizado con chirretera por la flota artesanal del puerto de Cabo de Palos en 2013.
Figura 4.9 Distribución espacial de la densidad del esfuerzo pesquero realizado con moruna por la flota artesanal del puerto de Cabo de Palos en 2013.

- 64 -
4.4.2 Evolución temporal del CPUE y €PUE entre 1993
y 2012
En los últimos veinte años la flota artesanal del Puerto de Cabo
de Palos ha sufrido una drástica disminución en el número de
embarcaciones, pasando de 14 en 1993 a 7 en 2012. Los barcos
artesanales en el año de 2012 tenían, de media, una antigüedad
de 26 años, una eslora total de 8,8 metros y las tripulaciones
compuestas por 2,3 personas (Tabla 4.1). La flota concentra su
esfuerzo casi exclusivamente en las cercanías del puerto de
Cabo de Palos. No obstante, un único barco suele dividir su
esfuerzo entre los caladeros de la zona de Cabo de Palos y la
zona de San Pedro del Pinatar, subastando sus capturas en la
cofradía de la zona en que ha pescado. Esto hace que su
presencia media en los datos de la Cofradía de Cartagena sea
menor que los demás barcos del puerto, sin embargo aun así
mantiene su derecho a explotar los recursos de la reserva
marina.
Tabla 4.1 Resumen de las características técnicas de la flota pesquera artesanal en el año de 2012 del puerto de Cabo de Palos. Se muestran los promedios de cada característica, la desviación estándar (±de) y el número de barcos (N) incluidos en los cálculos.
Características Media ±de N Edad (años) 26 29 7 Eslora total (m) 8,8 1,48 7 Potencia del motor declarada (kW) 39,1 26,09 7 Arquero TRB 4,0 1,66 7 Arqueo GT 4,01 2,30 7 Números de tripulantes por barco 2,3 0,95 7 Días activos de pesca 134 54 7

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 65 -
Entre 1993 y 2012 (excluyendo los años 1997 y 1998, para
los cuales no se tienen datos), la captura total declarada de la
flota artesanal de Cabo de Palos ha sido de 449.972 kg de
pescado, habiéndose observado un incremento de 376% en el total
anual de capturas (Fig. 4.10). Después de la creación de la
reserva marina de Cabo de Palos – Islas Hormigas, en 1995, hubo
un aumento significativo (R2 = 0,80; p < 0,001) en las capturas,
que pasaron de 6.000 kg en 1995 a 39.951 kg en 2012. En este
período destaca el incremento ocurrido en 2002, año en el cual
por primera vez se alcanzaron los 30.000 kg de pescado, un 118%
más que el año anterior y aproximadamente 22.000 kg más que en
1993.
Figura 4.10 Evolución de las capturas totales declaradas (en kg) por las embarcaciones artesanales de Cabo de Palos entre 1993 y 2012.
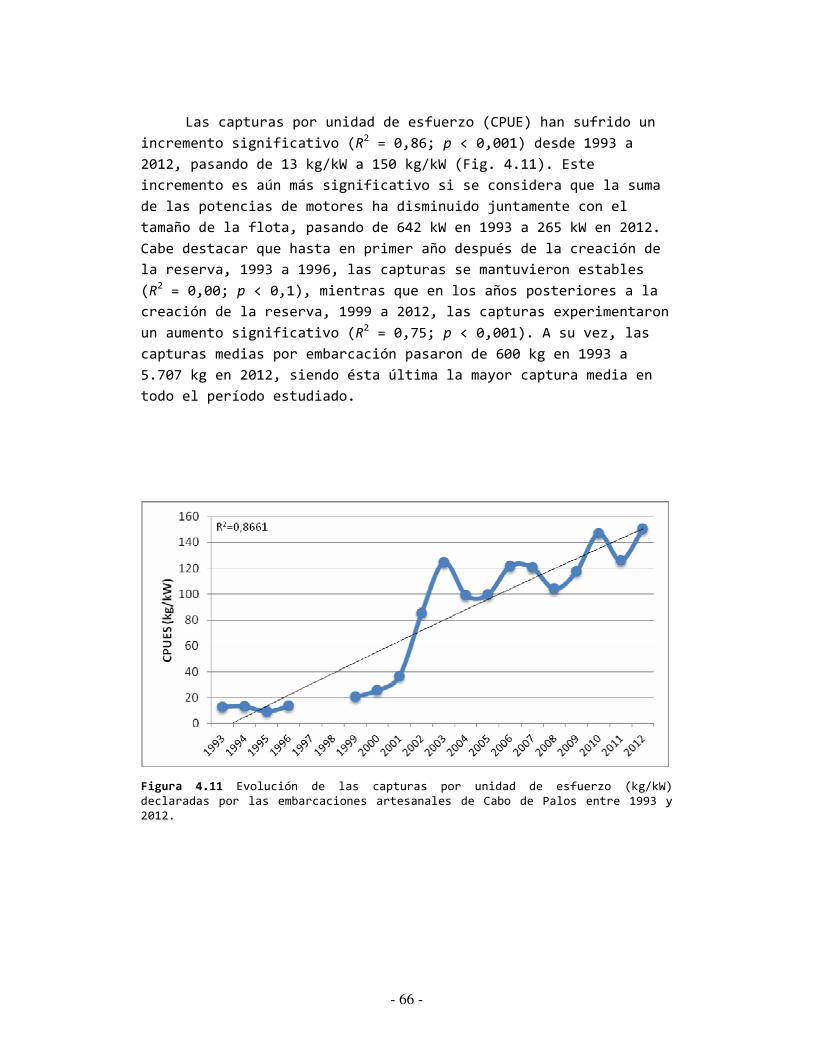
- 66 -
Las capturas por unidad de esfuerzo (CPUE) han sufrido un
incremento significativo (R2 = 0,86; p < 0,001) desde 1993 a
2012, pasando de 13 kg/kW a 150 kg/kW (Fig. 4.11). Este
incremento es aún más significativo si se considera que la suma
de las potencias de motores ha disminuido juntamente con el
tamaño de la flota, pasando de 642 kW en 1993 a 265 kW en 2012.
Cabe destacar que hasta en primer año después de la creación de
la reserva, 1993 a 1996, las capturas se mantuvieron estables
(R2 = 0,00; p < 0,1), mientras que en los años posteriores a la
creación de la reserva, 1999 a 2012, las capturas experimentaron
un aumento significativo (R2 = 0,75; p < 0,001). A su vez, las
capturas medias por embarcación pasaron de 600 kg en 1993 a
5.707 kg en 2012, siendo ésta última la mayor captura media en
todo el período estudiado.
Figura 4.11 Evolución de las capturas por unidad de esfuerzo (kg/kW) declaradas por las embarcaciones artesanales de Cabo de Palos entre 1993 y 2012.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 67 -
Los ingresos económicos obtenidos por la flota artesanal de
Cabo de Palos entre los años 2001 y 2012 suman más 2,8 millones
de euros, que en valores constantes de 2001 ajustados teniendo
en cuenta la inflación interanual en la Región de Murcia, son
2,1 millones de euros (Tabla 4.2), y muestran una tendencia,
aunque estadísticamente no significativa (R2 = 0,15; p = 0,20),
a aumentar con el paso del tiempo. Los ingresos por unidad de
esfuerzo (€PUE) presentan la misma tendencia no significativa de
crecimiento a lo largo de los años (R2 = 0,24; p = 0,10) (Fig.
4.12). Sin embargo, en tres años a partir de 2001, el sexto
después de la creación de la reserva, se registró un aumento por
valor de seis veces la media del primer año, para después
quedarse los ingresos medios anuales por barco fluctuando
alrededor de 25.000 €, a pesar del aumento de las CPUE.
Año Importe ajustado N €PUE
2001 74.173,93 € 10 7.417,39 € 2002 171.959,07 € 9 19.106,56 € 2003 213.056,94 € 7 30.436,71 € 2004 175.017,33 € 7 25.002,48 € 2005 145.947,58 € 7 20.849,65 € 2006 211.470,31 € 8 26.433,79 € 2007 227.309,60 € 8 28.413,70 € 2008 200.024,52 € 8 25.003,06 € 2009 187.484,29 € 8 23.435,54 € 2010 176.658,04 € 7 25.236,86 € 2011 158.181,65 € 7 22.597,38 € 2012 201.930,01 € 7 28.847,14 € Total 2.143.213,26 €
Tabla 4.2 Variación del importe total generado por la flota artesanal de Cabo de Palos entre los años 2001 y 2012, en valores constantes de 2001 ajustados por la inflación interanual de la Región de Murcia, y de los importes por unidad de esfuerzo (€PUE) (en ingresos medios por barco).

- 68 -
Figura 4.12 Evolución de los importes (en €) por unidad de esfuerzo (ingresos por barco) de los barcos de Cabo de Palos entre 2001 y 2012 en valores constantes de 2001 ajustados por la inflación interanual de la Región de Murcia.
Analizando la variación interanual de biomasa declarada, se
percibe un incremento en las capturas de las especies objetivo y
una disminución del conjunto de especies no objetivo. Las cinco
especies más abundantes sumadas representaban en 1999 un 8% del
total de las capturas, sin embargo en el año 2012 pasan a
representar un 42%. Por su parte, la categoría ‘morralla’ pasó
de representar un 36% de las capturas en 1999, a un 5% en 2012.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 69 -
Figura 4.13 Variación anual de la biomasa (kg) declarada de las seis especies más capturadas por la flota artesanal de Cabo de Palos entre años de 1999 y 2012.
Las especies objetivo tienen aún más importancia en
relación a los ingresos generados por las capturas. Incluso no
siendo las especies más capturadas al inicio de la serie
histórica de datos, las especies objetivo ya representaban la
principal fuente de ingresos para la flota. De los más de 74.000
euros generados por las capturas en el año 2001, un 55% provino
de cinco especies objetivo: chanquete (A. minuta, 29%), lecha
(Seriola dumerili, 11%), gallineta (Scorpaena scrofa, 7%),
dentón (Dentex dentex, 4%) y salmonete (Mullus spp., 4%); en
2012 las mismas especies tuvieron una importancia aún mayor,
representando el 63% de los ingresos totales, correspondiendo el
21%, 11%, 9%, 9% y 12% a cada especie respectivamente. Sin
embargo, los ingresos generados por la captura de morralla
fueron un 13% en 2001 y un 2% en 2012, estos valores se explican
porque este grupo representó el 22% de la biomasa total en 2001
y apenas un 5% en 2012.

- 70 -
Especies TOTAL Especies TOTAL
Anguilla anguilla 783,00 Pagellus spp. 3.732,84
Aphia minuta 42.280,29 Pagrus pagrus 673,40
Atherina spp. 6.714,88 Palinurus spp. 1.023,95
Auxis spp. 2.595,85 Parapristipoma octolineatum 576,80
Boops boops 322,60 Phycis spp. 2.877,70
Campogramma glaycos 666,74 Plectorhinchus mediterraneus 255,90
Chelon labrosus 418,35 Polyprion americanus 457,80
Citharus linguatula 15,80 Pomatomus saltator 191,75
Conger conger 11.577,34 Psetta máxima 91,00
Coryphaena hippurus 535,15 Rajiformes* 8.456,00
Crangon spp. 30,15 Sarda sarda 3.593,65
Dactylopterus volitans 410,70 Sarpa salpa 675,30
Dasyatis pastinaca 1.232,40 Sciaena umbra 18.461,92
Dentex dentex 19.953,55 Scomber japonicus 144,85
Dentex spp. 188,70 Scomber scombrus 559,80
Dicentrarchus labrax 947,90 Scorpaena scrofa 12.661,27
Diplodus spp. 14.510,72 Scorpaena spp. 12.305,56
Engraulis encrasicholus 59,00 Scyliorhinus canicula 174,05
Epinephelus costae 219,14 Scyllarides ocellatus 24,15
Epinephelus marginatus 2.688,64 Sepia officinalis 7.574,49
Euthynnus alletteratus 10.293,64 Seriola dumerili 42.074,82
Helicolenus dactylopterus 49,90 Serranus spp. 1.214,14
Homarus gammarus 194,49 Solea spp. 516,55
Labrus spp. 513,10 Sparus aurata 5.170,82
Lepidorhombus spp. 248,55 Sphyraena spp. 5.891,70
Liocarcinus depurator 16,70 Spicara smaris 1.280,50
Lithognathus mormyrus 6.000,20 Spondyliosoma cantharus 457,85
Liza aurata 1.627,65 Sprattus sprattus 11,00
Loligo vulgaris 700,74 Squaliformes* 4.394,54
Lophius piscatorius 3.860,70 Thunnus alalunga 77,60
Merluccius merluccius 1.458,80 Todarodes sagittatus 110,95
Morralla* 43.359,82 Trachinotus ovatus 5.057,18
Mullus spp. 35.098,98 Trachinus draco 131,89
Muraena spp. 620,70 Trachurus spp. 1.499,44
Murex spp. 77,15 Trichiurus lepturus 4.409,00
Mustelus mustelus 835,05 Trigla spp. 1.089,95
Myliobatis aquila 5.244,53 Trisopterus minutus 1,70
Nephrops norvegicus 146,50 Trisopterus spp. 49,30
Oblada melanura 134,15 Uranoscopus scaber 399,50
Octopus vulgaris 35.727,65 Zeus faber 229,05
Pagellus erythrinus 19.033,85
Tabla 4.3 Listado de especies y biomasa total capturada, en kg, por la flota artesanal de Cabo de Palos entre los años 1999 y 2012, donde (*) representa un grupo de especies.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 71 -
Cuando se examina la evolución de las capturas totales de
las especies directamente afectadas por la protección pesquera
ejercida por la reserva marina, se observa que el dentón (D.
dentex), los sargos (Diplodus spp.) y los espetones (Sphyraena
viridensis), y en menor medida los meros (Epinephelus
marginatus) y las corvas (Sciaena umbra) presentan una tendencia
clara al aumento de capturas a lo largo del tiempo (Fig. 4.14).
Por su parte, las gallinetas (Scorpaena scrofa) presentan un
comportamiento más irregular, sin que haya una tendencia clara
al aumento en sus capturas, aunque sí algunos años especialmente
favorables (Fig. 4.14). Sin embargo, en casi todos los casos es
notable un descenso en las capturas durante los últimos años
examinados, lo cual también es evidente en la suma de capturas
de estas especies emblemáticas (Fig. 4.15).
Figura 4.14 Variación anual de la biomasa (kg) declarada de las seis especies más relacionadas con los efectos de la reserva marina por la flota artesanal de Cabo de Palos entre años de 1999 y 2012.
Dentex dentex
0
500
1000
1500
2000
2500
3000
3500Diplodus spp.
0
500
1000
1500
2000
2500
Sciaena umbra
0
1000
2000
3000
4000
5000
6000
7000
Scorpaena scrofa
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Epinephelus spp.
0
100
200
300
400
500
600
700
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
Sphyraena viridensis
0
100
200
300
400
500
600
700
800
900
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
Ca
ptu
ras
(kg
)

- 72 -
Figura 4.15 Variación anual entre 1999 y 2012 de la suma de la biomasa (kg) declarada de las seis especies más relacionadas con los efectos de la reserva marina por la flota artesanal de Cabo de Palos (ver Fig. 4.14 para la identidad de esa especies).
La evolución temporal de los valores de CPUE para estas
mismas especies es prácticamente la misma que la observada para
las capturas brutas (Fig. 4.16), al igual que ocurre con los
valores de €PUE.
Figura 4.16 Variación anual entre años de 1999 y 2012 de la captura por unidad de esfuerzo (CPUE, en kg kW-1) declarada de las seis especies más relacionadas con los efectos de la reserva marina por la flota artesanal de Cabo de Palos.
Ca
ptu
ras
(kg
)
0
2000
4000
6000
8000
10000
12000
14000
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
Dentex dentex
0
2
4
6
8
10
12
Diplodus spp.
0
2
4
6
8
Sciaena umbra
0
5
10
15
20
25
Scorpaena scrofa
0
2
4
6
8
Epinephelus spp.
0.0
0.5
1.0
1.5
2.0
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
Sphyraena viridensis
0.0
0.5
1.0
1.5
2.0
2.5
3.0
1999200020012002200320042005 2006 2007 2008 2009 2010 2011 2012
Dentex dentex
0
2
4
6
8
10
12
Diplodus spp.
0
2
4
6
8
Sciaena umbra
0
5
10
15
20
25
Scorpaena scrofa
0
2
4
6
8
Epinephelus spp.
0.0
0.5
1.0
1.5
2.0
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
Sphyraena viridensis
0.0
0.5
1.0
1.5
2.0
2.5
3.0
1999200020012002200320042005 2006 2007 2008 2009 2010 2011 2012
CP
UE
(k
g k
W-1
)

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 73 -
4.5 Discusión
El presente estudio tiene como objetivo describir la evolución
temporal y evaluar el efecto de la creación de la reserva marina
de Cabo de Palos-Islas Hormigas en las capturas e ingresos
económicos de la flota artesanal que utiliza el área protegida y
sus aledaños como zona de pesca. Nuestros resultados muestran,
como resultado general más reseñable, que ha habido un aumento
significativo en las tasas de captura y en los ingresos totales
de la flota artesanal local a lo largo de los años, aunque los
ingresos por barco, tras un aumento inicial, se han estabilizado
alrededor de 25.000 euros. Sin embargo, gran parte de las
especies objetivo que han experimentado un mayor aumento, tales
como chanquete, salmonete o pulpo, no pueden relacionarse
directamente con los efectos de la reserva marina. El aumento en
las capturas de otras especies objetivo, como dentones, meros,
corvas, espetones y sargos, en cambio, sí son achacables a un
efecto positivo de la reserva marina, mientras que para otras
las capturas han sido más irregulares (caso de las gallinetas).
Ello ha determinado un aumento de las capturas del conjunto de
especies favorecidas por las medidas pesqueras en la reserva.
Asimismo, es notable una disminución de dichas capturas de
especies objeto de las medidas de protección durante los últimos
años, y especialmente en 2011 y 2012.
Los efectos positivos de las AMPs en las pesquerías locales
ya han sido encontrados en otras reservas marinas en el
Mediterráneo (Goñi et al. 2008, Stelzenmüller et al. 2008,
Forcada et al. 2009, Guidetti & Claudet 2009, Vandeperre et al.
2011). Así, en general las capturas por unidad de esfuerzo son
aproximadamente 12 veces mayores y los ingresos anuales por cada
barco más de cuatro veces mayores que los valores de los años
anteriores a protección. De este modo, las AMPs se han mostrado
como una importante herramienta de gestión (Gell & Roberts 2003,
Fenberg et al. 2012) en el intento de mantener la actividad
pesquera (Roberts et al. 2001), de impedir el colapso de los
‘stocks’ pesqueros (Myers & Worm 2003; Claudet et al. 2010) y de
conservar la biodiversidad (Guidetti 2006, Beddington et al.
2007, McCay et al. 2011).

- 74 -
Sin embargo, para que alcancen sus objetivos las reservas
marinas necesitan ser adecuadamente vigiladas y gestionadas (Di
Franco et al. 2009, Guidetti et al. 2008, 2010). La posible
existencia de furtivismo en Cabo de Palos por pescadores pseudo-
profesionales (es decir, personas que, utilizando métodos
deportivos, como arpones, capturan ilegalmente determinadas
especies en la reserva marina) puede explicar las disminuciones
de capturas durante los últimos años. Otra posible causa de
dicha disminución de capturas podría ser las interferencias con
el buceo recreativo en los mismos lugares utilizados para la
pesca artesanal (bajos de Piles y Dentro), cuestión ésta que
merece ser investigada con mayor detalle.
El mayor incremento en los valores de biomasa capturada ha
ocurrido entre los años 2001 y 2002, con un aumento de 118% en
la tasa de captura, periodo en que el modelo teórico propuesto
por Pérez-Ruzafa et al. (2008) sugería una estabilización en el
efecto de protección (Russ & Alcala 2010), estimado en 3-5 años
tras la implantación de la reserva. Sin embargo, otros factores
pueden influenciar el incremento de las tasas de captura
(Kellner et al. 2007, Merino et al. 2009). La flota artesanal de
Cabo de Palos tiene una heterogeneidad tanto en su dinámica
temporal como en su distribución espacial (Esparza-Alaminos
2010), así como otras flotas artesanales (Berthou et al. 2005,
Tzanatos 2006, Stelzenmüller et al. 2008, Forcada et al. 2010).
Eligiendo el arte y el caladero, el pescador puede hacer frente
las variaciones ambientales y comerciales, así como a periodos
de veda y regulaciones pesqueras (Esparza-Alaminos 2010). El
pescador basa su elección del caladero en sus conocimientos
sobre la biología de los peces, las características del hábitat,
las condiciones climatológicas y por el conocimiento tradicional
(Wilcox & Pomeroy 2003, Merino et al. 2009). No obstante, la
creación de un AMP es un factor importante en la elección del
caladero y de agregación de la CPUE (Halpern et al. 2004), por
limitar el uso de otras áreas o artes, o bien por generar una
expectativa de incremento de las capturas y importes (Murawski
et al. 2004, Goñi et al. 2008, Forcada et al. 2010, Esparza-
Alaminos 2010).
También han tenido lugar diferencias en la composición de
la captura en el periodo estudiado. Las especies objetivo de las

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 75 -
artes pesqueras utilizadas fueron progresivamente aumentando su
abundancia a lo largo del tiempo, convirtiéndose en la principal
fuente de biomasa; sin embargo, únicamente en algunos casos
(espetones, sargos, dentones, meros y corvas) este aumento puede
ser debido a un mecanismo de exportación de biomasa de la
reserva hacia las áreas en las que la pesca está permitida
(Claudet et al. 2008, 2010, Esparza-Alaminos 2010) como se ha
visto en otras reservas marinas (Wilcox & Pomeroy 2003, Willis
et al. 2003, Abesamis et al. 2005, Murawski et al. 2005,
Stelzenmüller et al. 2007, Stobart et al. 2009, Forcada et al.
2009, Amargós et al. 2010, Goñi et al. 2010). Este
“desbordamiento” de biomasa hacia el exterior de la reserva se
ve favorecido en función de la estructura y distribución
espacial de los hábitats de la zona (Stelzenmüller et al. 2007,
Goñi et al. 2008, Forcada et al. 2009). El incremento en la
abundancia y/o biomasa de dichas especies indica que la reserva
marina de Cabo de Palos está contribuyendo a la recuperación de
especies altamente explotadas (Myers & Worm 2003, Unsworth et
al. 2007) y recuperando la estructura de la comunidad de peces
(McClanahan et al. 2007).
Por otra parte, se ha detectado una influencia de la
presencia de la reserva marina en la distribución del esfuerzo
pesquero, tal y como se ejemplifica en los datos
correspondientes al año 2013, en el que estuvo concentrado en el
interior o al borde de la reserva marina y vinculado a fondos de
elevada diversidad, principalmente relacionadas a presencia de
praderas bien desarrolladas de P. oceanica. Otro factor que ha
influido en la distribución espacial es el trasmallo claro, que
ha sido el arte de pesca más utilizado. Las características de
esta modalidad hacen que los pescadores busquen fondos y
profundidades específicas, que son precisamente las encontradas
en el área de la reserva marina (bajos rocosos) que difícilmente
se encuentran fuera de ella. Con ello, habría que estudiar las
motivaciones que llevan, en última instancia, a los pescadores a
seguir determinadas tácticas y estrategias de pesca a la hora de
utilizar dichos caladeros
Estos resultados muestran, para el caso de algunas
especies, el efecto positivo de las reservas marinas en las
pesquerías locales (Vandeperre et al. 2011). La flota artesanal

- 76 -
que faena en la reserva parcial y en el entorno de la Reserva
Marina de Cabo de Palos-Islas Hormigas ha visto un aumento en
sus capturas, mientras las demás pesquerías de la región no
asociadas a ninguna área protegida muestran una tendencia a
disminuir (Esparza-Alaminos 2010). Aunque algunos estudios
afirman que las zonas de reserva parcial pueden ser
perjudiciales para protección de los peces (Claudet et al. 2008)
y que las reservas parciales no son eficaces, debiéndose centrar
los esfuerzos de protección en la creación de reservas
integrales (‘no-take zones’) (Denny & Babcock 2004), nuestro
estudio apunta a que las zonas con protección parcial pueden ser
beneficiosas para las pesquerías. Sin embargo, se podrían
obtener mejores resultados si se implementaran planes de co-
gestión integrando a los pescadores y científicos, además de los
gestores (Guidetti & Claudet 2010; Hogg et al. 2013).
A pesar de los efectos sobre las pesquerías, históricamente
las AMPs son creadas para proteger la biodiversidad, los
hábitats prístinos y para el incremento del turismo (Holland et
al. 1996, Abdulla et al. 2008, Russ & Alcala 2010). De este modo
generan beneficios más allá de los pesqueros, contribuyendo al
desarrollo económico de las comunidades costeras cercanas
(Boncoeur et al. 2002, Roncin et al. 2008). La reserva marina de
Cabo de Palos-Islas Hormigas también cumple este papel,
promoviendo la protección de las praderas de fanerógamas marinas
e incrementando el turismo subacuático, extendiendo así los
beneficios a toda la comunidad (García-Charton et al. 2005,
2006, 2007, 2008, 2009, 2010). Sumado a esto, el sector pesquero
también se beneficia económicamente, una vez que el aumento de
los valores de venta del pescado y del €PUE generados por la
flota artesanal están directamente ligados al efecto de
protección, dado que ésta tiende a aumentar la abundancia, el
tamaño y biomasa de los peces (Barrett et al. 2007) y
consecuentemente la calidad y el valor la pesca (Guyader et al.
2013, Maynou et al. 2013). Sin embargo, nuestra observación de
que los ingresos económicos generados por la pesca declarada en
lonja no han aumentado significativamente tras un fuerte
incremento inicial, pese a que las capturas sí que han seguido
aumentando, merece ser explorada en más detalle. Ello es una
indicación de que la rentabilidad de las pesquerías en un AMP
está vinculada a los planes de gestión y comercialización, los

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 77 -
cuales deberían ser más sensibles a las realidades locales,
promoviendo la participación activa de pescadores, científicos y
demás usuarios de los recursos (Florido 2008), y pudiendo así
fortalecer social y comercialmente a las comunidades costeras a
la vez que se está preservando la herencia cultural asociada a
las pesquerías (Salas & Gaertner 2004). Sin embargo, debido a la
complejidad de las relaciones entre un área marina protegida y
sus diversos usuarios, la biodiversidad y los órganos de
gestión, aún se necesita mejorar el conocimiento sobre factores
como el diseño, el tamaño y los planes de uso para optimizar las
AMPs como herramientas de gestión (García-Charton et al. 2008).

- 78 -

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 79 -
55 Conclusiones
5.1 Poblamiento de peces de fondos rocosos
� El poblamiento de peces muestra una recuperación de sus
efectivos numéricos, que parecía haber disminuido (al menos
en lo que se refiere a los meros) durante los últimos años,
como consecuencia, posiblemente, de un aumento de la presión
por furtivos ocurrido durante los últimos años, sobre todo en
la reserva integral y el bajo de Fuera. Parece que las
medidas de vigilancia se han reajustado y han conseguido
limitar (aunque posiblemente no erradicar) la presión por
furtivos en la reserva marina.
� Las especies de peces más favorecidas por las medidas de
protección son meros, dentones, corvas, sargos reales y
espetones. Estas especies han visto aumentar sus abundancias,
aunque en los últimos años este aumento se ha ido atenuando.
Se hace necesario establecer si esta dinámica temporal
responde a que las poblaciones de peces se están acercando a
la capacidad de carga del hábitat (tras casi 20 años de
protección), o bien se trata de una respuesta a la presión
por furtivismo.
� Estas especies tienen unos hábitos tróficos macrocarnívoros,
favoreciéndose de este modo una recuperación del
compartimento de depredadores apicales, propia de las
comunidades marinas mejor conservadas.
� La estructura del hábitat rocoso determina parte de las
diferencias espaciales y temporales observadas, con especies
como raspallones, salmonetes y pequeños lábridos que son más
abundantes en la parte más costera de la reserva.
� La abundancia media de las tres especies de meros obtenidas
en el verano de 2013 no han coincidido en las dos ocasiones
en los que se han muestreado, una en junio-julio (censos
específicos de meros) y otra entre agosto y octubre (censos
de peces incluyendo todo el poblamiento observable), a pesar

- 80 -
de haberse utilizado la misma técnica de censo visual con
escafandra autónoma (transectos de 50×5 m2). Se proponen dos
posibles causas de este hecho: respuesta a anomalías en el
calentamiento estival del agua, y variabilidad temporal a
escala de días o semanas, que el muestreo aplicado no sería
capaz de absorber. Se hace necesario afinar en las técnicas
de censo visual a aplicar, con el fin de acceder a una
fracción representativa de las poblaciones presentes en la
reserva marina.
5.2 Impacto del submarinismo recreativo
� El presente estudio, unido a resultados de estudios
anteriores, nos indican que, aun no siendo de gran magnitud,
son detectables diversas señales de que la actividad
subacuática recreativa está ejerciendo cierto efecto erosivo
sobre los fondos de la reserva:
- La especie utilizada como indicadora de impacto del
submarinismo en la reserva marina (el briozoo Myriapora
truncata, o falso coral) es 5,6 veces más densa en la
reserva integral que en los bajos en los que se practica
el buceo recreativo.
- Estudios anteriores no han mostrado efectos tan
evidentes, aunque señales más tenues de una posible
influencia (sin poderse establecer claramente una
relación causa-efecto) han sido una mayor proporción de
fondo desprovisto de cobertura algal, menor densidad de
otras especies frágiles – esponjas, cnidarios, etc.,
mayor proporción de gorgonias tumbadas, y mayor densidad
de erizos comestibles – Paracentrotus lividus en los
bajos en los que se bucea en comparación con la reserva
integral.
- Estudios experimentales demostraron que el paso
continuado de buceadores sobre parcelas marcadas ejerce
un efecto erosivo sobre algas y determinados
invertebrados bentónicos, entre los que se cuenta el
falso coral, así como una resuspensión del sedimento
depositado sobre el sustrato rocoso.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 81 -
� Aunque dichos impactos no son de gran magnitud, sí
constituyen una señal temprana (‘early-warning’) de que la
actividad subacuática practicada actualmente es capaz de
ejercer cambios detectables en diversas características de
los fondos de la reserva marina. Ante la posibilidad más que
probable de que la relación entre perturbación y respuesta de
los fondos de la reserva marina no sea lineal, sino abrupta e
incluso discontinua (histéresis) (García-Charton et al.
2008), la mejor estrategia de gestión para mantener y
garantizar el estado de salud del ecosistema en la reserva
marina consiste en mantener los niveles de perturbación (sea
natural – p. ej. densidad de erizos, o antrópica – intensidad
de uso público por el submarinismo recreativo) alejados del
punto crítico de bifurcación. Ante la imposibilidad actual de
conocer dicho punto crítico, y apelando al Principio de
Incertidumbre, creemos imprescindible limitar en lo posible
ulteriores incrementos en el número de inmersiones
permisibles en la reserva marina.
� De lo anterior, se impone, en nuestra opinión, la necesidad
de establecer límite al número de inmersiones en la reserva
marina. Dicha limitación tendría que marcarse, con carácter
precautorio, lo más cerca posible de los niveles actuales de
frecuentación por buceadores, estimados, a falta de datos
oficiales, en unas 36.000 inmersiones anuales.
� El establecimiento de cupos al buceo es un problema de
gestión, cuyo principal objetivo sea promover la
compatibilidad de usos de la reserva, principalmente entre el
buceo recreativo y la pesca artesanal. Para dicha gestión,
las administraciones competentes deberían establecer
mecanismos participativos y de autorregulación del propio
sector, siguiendo esquemas de co-gestión, en los que se
satisfagan las expectativas y condicionantes de los sectores
implicados en la explotación de la reserva marina (centros de
buceo, pero también pescadores artesanales), con base en la
mejor información ecológica disponible. Además, dichos
mecanismos de co-gestión han de ser adaptativos, en un ciclo
de planificación-acción-evaluación-adaptación en el que los
resultados del seguimiento científico jueguen un papel
central (Fig. 5.1).

- 82 -
� El cupo anual, a establecer mediante un mecanismo
participativo, y revisable (con una periodicidad por decidir)
siguiendo esquemas adaptativos, tendrá que ir acompañado de
otras medidas de gestión del buceo recreativo. En pasados
informes se desarrollaban una serie de posibles medidas, que
reiteramos como aconsejables para minimizar los efectos que
el control y limitación del número de inmersiones en la
reserva marina pudieran tener sobre el sector:
Figura 5.1 Ciclo de gestión adaptativa, aplicable al establecimiento de medidas de regulación del buceo recreativo en la reserva marina. La periodicidad del ciclo podría ser anual o bianual.
- reparto espacio-temporal de la presión de buceo: cupos
semanales y/o mensuales, límite al número de
embarcaciones por boya y al número de submarinistas por
embarcación, etc.;
- obligación de realizar ‘briefings’ previos a la
inmersión, en los que se realicen recomendaciones y
orientaciones para minimizar el impacto individual de
los buceadores;
- obligatoriedad de llevar guías de las inmersiones, ya
que se ha demostrado que su presencia reduce el efecto
planificaciónplanificaciónplanificaciónplanificaciónobjetivosobjetivosobjetivosobjetivos
estrategiaestrategiaestrategiaestrategia
medidasmedidasmedidasmedidas
acciónacciónacciónacción
implementaciónimplementaciónimplementaciónimplementación
seguimientoseguimientoseguimientoseguimiento
acciónacciónacciónacción
implementaciónimplementaciónimplementaciónimplementación
seguimientoseguimientoseguimientoseguimiento
adaptaciónadaptaciónadaptaciónadaptación
evaluaciónevaluaciónevaluaciónevaluación
análisisanálisisanálisisanálisis
comunicacióncomunicacióncomunicacióncomunicación
evaluaciónevaluaciónevaluaciónevaluación
análisisanálisisanálisisanálisis
comunicacióncomunicacióncomunicacióncomunicación

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 83 -
erosivo de los buceadores, por su acción de control, o
por simple imitación de un desenvolvimiento adecuado
bajo el agua; para facilitar dicha tarea, se propone la
organización de cursos oficiales de eco-guía y la
edición de materiales divulgativos;
- oferta de puntos alternativos de buceo: itinerarios
submarinos, promoción de puntos de buceo fuera de la
reserva marina, arrecifes artificiales, pecios, etc.;
- organización de actividades de voluntariado científico,
utilizable incluso para complementar las tareas de
muestreo para el seguimiento de los efectos de la
reserva marina;
- edición de un manual de buenas prácticas subacuáticas;
- cobro de eco-tasas a los buceadores (sean éstos clientes
de centros de buceo o no), para contribuir a la
sostenibilidad económica de la reserva marina y/o a la
rentabilidad de la actividad para los centros de buceo;
- desarrollo de una eco-marca (fuente de prestigio y
reconocimiento) ‘Reserva marina de Cabo de Palos – Islas
Hormigas’, cuya adhesión exija la firma de convenios
específicos entre centros de buceo y la administración
de la reserva, sujetos al cumplimiento de una serie de
normas auto-impuestas orientadas a la sostenibilidad de
la reserva marina.
5.3 Pesca artesanal
� Los datos plurianuales indican que tanto las capturas totales
como las capturas por unidad de esfuerzo de las unidades de
pesca artesanal autorizadas a faenar en la reserva marina han
aumentado significativamente desde antes del establecimiento
de las medidas de protección. � Gran parte de esos aumentos, sin embargo, no son achacables
directamente a los efectos de la reserva marina, pues
descansan en el aumento en las capturas de especies objetivo
de las artes utilizadas que no se benefician directamente de

- 84 -
las medidas de protección (caso de chanquete, salmonete y
pulpo). � Otras especies (dentón, sargo, espetón, meros, corvas) que
han visto aumentar sus capturas sí pueden estar relacionadas
directamente con el éxito de las medidas de protección,
consistentes en un aumento de sus abundancias y biomasas (a
tenor de los datos obtenidos con métodos independientes de la
pesca – censos visuales en inmersión con escafandra
autónoma), y la exportación de sus efectivos desde la reserva
integral hacia la reserva parcial, y desde ésta a los fondos
exteriores a la reserva marina. � La consideración conjunta de las especies anteriores
asociadas a la reserva marina, más la gallineta (cuyas
abundancias han podido aumentar en la reserva marina, aunque
los censos visuales no están especialmente adaptados a su
control debido a sus características crípticas y miméticas),
como especies más preciadas por la pesca artesanal en las
zonas rocosas, determina una tendencia al aumento en la
biomasa capturada. Sin embargo, durante los dos últimos años
(2011 y 2012) se ha observado una disminución de dichas
capturas totales. Esta disminución puede tener su origen en
dos causas no excluyentes: aumento del furtivismo en la
reserva marina, e interferencia entre pesca y turismo
subacuático en los pocos lugares en los que ambas actividades
se pueden realizar (bajos rocosos). � Los ingresos obtenidos en lonja por la flota pesquera
artesanal de Cabo de Palos experimentó un aumento inicial,
pero desde hace varios años los ingresos por barco, aunque
fluctuantes, se mantienen constantes, y ello a pesar de los
aumentos registrados en las capturas por unidad de esfuerzo. � Se proponen las siguientes medidas de gestión de la pesca
artesanal en y alrededor de la reserva marina:
- erradicar la pesca furtiva, mejorando los mecanismos de
vigilancia y control del furtivismo en la reserva
marina;
- mejorar las estadísticas y los mecanismos de validación
de los datos pesqueros, para aumentar su utilidad en la
evaluación y gestión del sector;

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 85 -
- fomentar la “marca” del pescado de la reserva marina
(eco-etiqueta, ferias gastronómicas, campañas de
sensibilización, etc.);
- organizar actividades que mejoren el conocimiento del
sector por parte del gran público (artes de pesca,
especies, organización de la pesca, etc.);
- mejorar los circuitos y mecanismos de comercialización,
con el fin de mejorar el rendimiento económico de la
pesca, eliminando intermediarios;
- explorar mecanismos de diversificación del sector, con
fomento de actividades alternativas (p. ej.: pesca-
turismo, visitas guiadas, etc.);
- junto con la continuación de los seguimientos de captura
y esfuerzo, iniciar estudios específicos de los
siguientes aspectos de la pesca artesanal en la reserva:
(1) factores que influyen en las tácticas y estrategias
pesqueras y la distribución del esfuerzo pesquero, (2)
diagnóstico y mejora de los circuitos y mecanismos de
comercialización y puesta en valor de los productos de
la pesca artesanal en la reserva marina, (3) exploración
de alternativas a la pesca por parte del sector
pesquero: pesca-turismo, visitas guiadas, etc.;
- fomentar la compatibilidad entre usos en la reserva
marina (principalmente entre pesca artesanal y turismo
subacuático), arbitrando los mecanismos de participación
pública y co-manejo siguiendo esquemas adaptativos (ver
apartado anterior, referido a los cupos de buceo).

- 86 -

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 87 -
66 Bibliografía
Abdulla A, Gomei M, Maison E, Piante C. (2008) Status of Marine Protected Areas in the Mediterranean Sea. IUCN-Málaga and WWF-France. 152 pp.
Abesamis RA, Russ GR (2005) Density-dependent spillover from a Marine reserve: long term evidence. Ecological Applications, 15: 1798-1812.
Amargós F, Sansón GG, Castillo AJ, Fernández AZ, Blanco FM (2010) An experiment of fish spillover from a marine reserve in Cuba. Environmental Biology of Fishes 87: 363-372.
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for PRIMER: Guide to software and statistical methods. Primer-e, Plymouth, UK.
Barker NHL, Roberts C (2004) Scuba diver behaviour and the management of diving impacts on coral reefs. Biological Conservation 120: 481-489.
Barrett NS, Edgar GE, Buxton CD, Haddon M (2007) Changes in fish assemblages following 10 years of protection in Tasmanian marine protected áreas. Journal of Experimental Marine Biology and Ecology 345:141-157.
Battaglia P, Romeo T, Consoli P, Scotti G, Andaloro F (2010) Characterization of the artisanal fishery and its socio-economic aspects in the central Mediterranean Sea (Aeolian Islands, Italy). Fisheries Research, 112:87–97.
Baum JK, Worm B (2009) Cascading top-down effects of changing oceanic predator abundances. Journal of Animal Ecology 78: 699-714.
Beddington JRB, Agnew J, Clark CW (2007) Current problems in the management of marine fisheries. Science 316: 1713-1716.
Berning B (2007) The Mediterranean bryozoan Myriapora truncata (Pallas, 1766): a potential indicator of (palaeo-) environmental conditions. Lethaia 40: 221-232.
Berthou P, Daurès F, Demaneche S (2005) Some considerations about small-scale coastal fisheries in Europe. Workshop on Small-Scale Fisheries, 12–16th September. Kavala, Greece.
Boncoeur J, Alban F, Guyader O, Thébaund O (2002) Fish, fishers, seals and tourists: economic consequences of creating a marine reserve in a multi-species, multi-activity context. Natural Resources Modelling 15: 387-411.
Calvisi G, Pais M, Floris A (2003) Un metodo di valutazione dell’impacto dell’attività subacquea in un’area marina protteta sarda. Biologia Marina Mediterranea 10: 503-505.

- 88 -
Canu F, Bassler RF (1925) Les bryozoaires du Maroc et de Mauritanie. Mém Soc Sci Nat Maroc 18: 1-85.
Chuenpagdee R, Pascual-Fernández JJ, Szeliánszky E, Alegret JL, Fraga J, Jentoft S (2013) Marine protected areas: Re-thinking their inception. Marine Policy 39: 234-240.
Claudet J, García-Charton JA, Lenfant P (2011) Combined effects of levels of protection and environmental variables at different spatial resolutions on fish assemblages in a marine protected area. Conservation Biology 25: 105-114.
Claudet J, Osenberg CW, Benedetti-Cecchi L, Domenici P, García-Charton JA, Pérez- Ruzafa A, Badalamenti F, Bayle-Sempere J, Brito A, Bulleri F, Culioli JM, Dimech M, Falcón JM, Guala I, Milazzo M, Sánchez-Meca J, Somerfield PJ, Stobart B, Vandeperre F, Valle C, Planes S (2008) Marine reserves: size and age do matter. Ecology Letters 11: 481–489.
Claudet J, Osenberg CW, Domenici P, Badalamenti, M, Falcón, JM, Bertocci, I, Benedetti-Cecchi, L, García-Charton JA, Goñi, R, Borg JA, Forcada A, De Lucia GA, Pérez-Ruzafa A, Afonso P, Brito A, Guala I, Le Diréach L, Sánchez-Jerez P, Somerfield PJ, Planes S (2010) Marine reserves: fish life history and ecological traits matter. Ecological Applications 20: 830-839.
Coll M, Piroddi C, Albouy C, Lasram FBR, Cheung WWL, Christensen V, Karpouzi VS, Guilhaumon F, Mouillot D, Paleczny M, Palomares ML, Steenbeek J, Trujillo P, Watson R, Pauly D (2012) The Mediterranean Sea under siege: spatial overlap between marine biodiversity, cumulative threats and marine reserves. Global Ecology and Biogeography 21: 465-480.
Colloca F, Crespi V, Cerasi S, Coppola S (2004) Structure and evolution of the artisanal fishery in a southern Italian coastal area. Fisheries Research, 69 (3):359–369.
Coma R, Pola E, Ribes M, Zabala M (2004) Long-term assessment of temperate octocoral mortality patterns, protected vs. unprotected areas. Ecological Applications 14: 1466-1478.
Cornish A, Harmelin-Vivien ML (2004) Epinephelus marginatus. IUCN 2010. IUCN Red List of Threatened Species. Version 3.1. www.iucnredlist.org.
Crilly R, Esteban A (2013) Small versus large-scale, multi-fleet fisheries: The case for economic, social and environmental access criteria in European fisheries. Marine Policy 37: 20-27.
Cullis-Suzuki S, Pauly D (2010) Marine protected area costs as “beneficial” fisheries subsidies: a global evaluation. Coastal Management 38: 113-121.
Davis D, Tisdell C (1996) Economic management of recreational scuba diving and the environment. Journal Environmental Management 48: 229-248.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 89 -
De Santo EM (2013) Missing marine protected area (MPA) targets: How the push for quantity over quality undermines sustainability and social justice. Journal of Environmental Management 124: 137–146.
Dearden P, Bennett M, Rollins R (2007) Perceptions of diving impacts and implications for reef conservation. Coastal Management 35: 305-317.
Denny CM, Babcock RC (2004) Do partial marine reserves protect reef fish assemblages? Biological Conservation 116:119-129.
Di Franco A, Bussotti S, Navone A, Panzalis P, Guidetti P (2009) Evaluating effects of total and partial restrictions to fishing on Mediterranean rocky-reef fish assemblages. Marine Ecology Progress Series 387: 275-285.
Esparza-Alaminos O (2010) Estudio de la pesca artesanal en el entorno de la reserva marina de Cabo de Palos – Islas Hormigas. Estrategias de pesca, efecto de la protección y propuestas para la gestión. Tesis doctoral, Universidad de Murcia. 227 pp.
FAO (2011) Review of the state of world marine fishery resources. FAO Fisheries and Aquaculture Technical Paper No. 569. Rome, Italy. 344 pp.
FAO (2012) The state of world fisheries and Aquaculture. 209 pp. Fenberg PB, Caselle JE, Claudet J, Clemence M, Gaines SD,
García-Charton JA, Gonçalves EJ, Grorud-Colvert K, Guidetti P, Jenkins SR, Jones PJS, Lester SE, McAllen R, Moland E, Planes S, Sørensen T (2012) The science of European marine reserves: Status, efficacy, and future needs. Marine Policy 36: 1012–1021.
Ferretti C, Magnino G, Balduzzi A (2007) Morphology of the larva and ancestrula of Myriapora truncata (Bryozoa, Cheilostomatida), Italian Journal of Zoology, 74: 341-350.
Florido D (2008) Las flotas artesanales andaluzas en un contexto de crisis. González Laxe. Lecciones de Economía Pesquera. Instituto de Estudios Marítimos, Netbiblio. A Coruña, España.
Forcada A, Valle C, Bonhomme P, Criquet G, Cadiou G, Lenfant P, Sánchez-Lizaso JL (2009) Effects of habitat on spillover from marine protected areas to artisanal fisheries. Marine Ecology Progress Series, 379: 197–211.
Forcada A, Valle C, Sánchez-Lizaso JL, Bayle-Sempere JT, Corsi F (2010) Structure and spatio-temporal dynamics of artisanal fisheries around a Mediterranean marine protected area.ICES Journal of Marine Science 67: 191-203.
Fox HE, Mascia MB, Basurto X, Costa A, Glew S, Heinemann D, Karrer LB, Lester SE, Lombana AV, Pomeroy RS, Recchia CA, Roberts CM, Sanchirico JN, Pet-Soede L, White AT (2012)

- 90 -
Reexamining the science of marine protected areas: linking knowledge to action. Conservation Letters 5: 1-10.
Froese R, Zeller D, Kleisner K, Pauly D (2012) What catch data can tell us about the status of global fisheries. Marine Biology. DOI 07/s00227-012-1909-6.
Gabrié C, Lagabrielle E, Bissery C, Crochelet E, Meola B, Webster C, Claudet J, Chassanite A, Marinesque S, Robert P, Goutx M (2012) The status of the Marine Protected Areas in the Mediterranean Sea 2012, MedPAN & CAR/ASP, 2012.
García B (1997) Las pesquerías artesanales en los alrededors de la reserva marina de Cabo do Palos-Islas Hormigas (Murcia). Congreso Biología Pesquera. Murcia : Consejería de Medio Ambiente, Agricultura y Agua.
García-Charton JA, Hackradt CW, Félix-Hackradt FC (2013) Evolución temporal de la abundancia de las poblaciones de meros en la reserva marina de Cabo de Palos – Islas Hormigas (1996-2011). Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
García-Charton JA, Hackradt CW, Félix-Hackradt FC, Segovia-Viadero M, Martínez-Garrido J, González-Andrés C, Tubío A, Sanchis-Martínez A, Morcillo-Buendía E, Treviño-Otón J, Marcos-Diego C, Pérez-Ruzafa A (2010) Estudios de seguimiento de la reserva marina de Cabo de Palos – Islas Hormigas 2010. Universidad de Murcia, Comunidad Autónoma de la Región de Murcia.
García-Charton JA, Herrero-Pérez A, Esparza-Alaminos O, Espejo-Cayuela C, Pérez-Ruzafa A, Marcos C (2005) Estudios de seguimiento de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
García-Charton JA, Herrero-Pérez A, Esparza-Alaminos O, Pérez Ruzafa A, Marcos C (2006) Estudios de seguimiento de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
García-Charton JA, Herrero-Pérez A, Esparza-Alaminos O, Pérez-Ruzafa A, Marcos C (2007) Estudios de seguimiento de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
García-Charton JA, Herrero-Pérez A, Esparza-Alaminos O, Pérez-Ruzafa A, Marcos C, Treviño J, Cenci E, Segovia-Viadero M (2008a) Estudios de seguimiento de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 91 -
García-Charton JA, Marcos C (2004) Los fondos marinos de la costa murciana. En: Esteve-Selma MA, Riquelme MLP, Martínez-Gallur C. (Eds.) Los recursos naturales de la Región de Murcia: un análisis interdisciplinar. Servicio de Publicaciones de la Universidad de Murcia: 385- 394.
García-Charton JA, Pérez-Ruzafa Á (2001) Spatial pattern and the habitat structure of a Mediterranean rocky reef fish local assemblage. Marine Biology 138: 917-934.
García-Charton JA, Pérez-Ruzafa Á, Marcos C, Claudet J, Badalamenti F, Benedetti-Cecchi L, Falcón JM, Milazzo M, Schembri PJ, Stobart B, Vandeperre F, Brito A, Chemello R, Dimech M, Domenici P, Guala I, LeDiréach L, Maggi E, Planes S (2008b) Effectiveness of European Atlanto-Mediterranean MPAs: Do they accomplish the expected effects on populations, communities and ecosystems? J Nat Conserv 16: 193-221.
García-Charton JA, Pérez-Ruzafa A, Sánchez-Jerez P, Bayle-Sempere JT, Reñones O, Moreno-Lampreave D (2004) Multi-scale spatial heterogeneity, habitat structure, and the effect of marine reserves on Western Mediterranean rocky reef fish assemblages. Marine Biology 144 : 161–182.
García-Rubies A, Hereu B, Zabala M (2013) Long-term recovery patterns and limited spillover of large predatory fish in a Mediterranean MPA. PLoS ONE 8(9): e73922. doi:10.1371/journal.pone.0073922
Garrabou J, Sala E, Arcas A, Zabala M (1998) The impact of diving on rocky sublittoral communities: A case study of a bryozoan population. Conservation Biology 12: 302-312.
Gautier YV (1962) Recherches écologiques sur les Bryozoaires Chilostomes en Méditerranée Occidentale. Recueil des Travaux de la Station Marine d’Endoume 38: 1-434.
Gell FR, Roberts CM (2003) Benefits beyond boundaries: the fishery effects of marine reserves. Trends in Ecology & Evolution 18: 448-455.
Gleason M, Fox E, Ashcraft S, Vasques J, Whiteman E, Serpa P, Saarman E, Caldwell M, Frimodig A, Millar-Henson M, Kirlin J, Ota B, Pope E, Weber M, Wiseman K (2013) Designing a network of marine protected areas in California: Achievements, costs, lessons learned, and challenges ahead. Ocean & Coastal Management 74: 90-101.
Goñi R, Adlerstein S, Alvarez-Berastegui D, Forcada A, Reñones O, Criquet G, Polti S, Cadiou G, Valle C, Lenfant P, Bonhomme P, Pérez-Ruzafa A, Sánchez-Lizaso JL, García-Charton JA, Bernard G, Stelzenmüller V, Planes S (2008) Spillover from six western Mediterranean marine protected areas: evidence from artisanal fisheries. Marine Ecology Progress Series 366: 159-174.

- 92 -
Goñi R, Hilborn R, Diaz D, Mallol S, Adlerstein S (2010) Net contribution of spillover from a marine reserve to fishery catches. Mar Ecol Prog Ser. 400:233–243.
Guarnieri G, Terlizzi A, Bevilacqua S, Fraschetti S (2012) Increasing heterogeneity of sensitive assemblages as a consequence of human impact in submarine caves. Marine Biology 159: 1155-1164.
Guidetti P (2006) Marine reserve reestablish lost predatory interactions and cause community changes in rocky reefs. Ecological Applications 16: 963-976.
Guidetti P, Bussotti S, Pizzolante F, Ciccolella A (2010) Assessing the potential of an artesanal fishing co-management in the Marine Protected Area of Torre Guaceto (Southern Adriatic Sea, SE Italy). Fisheries Research 101: 180-187.
Guidetti P, Claudet J (2010) Co-management practices enhance fisheries in marine protected areas. Conservation Biology 24: 312-318.
Guidetti P, Milazzo M, Bussotti S, Molinari A, Murenu M, Pais A, Spanò N, Balzano R, Agardy T, Boero F, Carrada G, Cattaneo-Vietti R, Cau A, Chemello R, Greco S, Manganaro A, Notarbartolo di Sciara G, Russo G, Tunesi L (2008) Italian marine reserve effectiveness: does enforcement matter?. Biological Conservation 141: 699-709.
Guyader O, Berthou P, Koutsikopoulos C, Alban F, Demanèche S, Gaspar MB, Eschbaum R, Fahy E, Tully O, Reynal L, Curti O, Frangoudes K, Maynou F (2013) Small scale fisheries in Europe: A comparative analysis based on a selection of case studies. Fisheries Research 140: 1-13.
Hackradt CW (2012) Population ecology and mobility patterns of groupers (Serranidae: Epinephelinae) on temperate rocky reefs on Southwestern Mediterranean Sea: Implications for their conservation. Tesis doctoral, Universidad de Murcia.
Halpern BS, Floeter SR (2008) Functional diversity responses to changing species richness in reef fish communities. Marine Ecology Progress Series 364: 147-156.
Halpern BS, Gaines SD, Warner RR (2004) Confounding effects of the export of production and the displacement of fishing effort from marine reserves. Ecological Applications 14: 1248–1256.
Halpern BS, Lester SE, McLeod KL (2010) Placing marine protected areas onto the ecosystem-based management seascape. Proceedings of the National Academy of Sciences of the United States of America 107: 1-6.
Halpern BS, Warner RR (2002) Marine reserves have rapid and lasting effects. Ecology Letters 5: 361-366.
Harmelin J-G (1987) Structure et variabilité de l’ichtyofaune d’une zone rocheuse protégée en Méditerranée (Parc

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 93 -
national de Port-Cros, France). PSZN I: Marine Ecology 8: 263-284.
Harmelin-Vivien M, Le Diréach L, Bayle-Sempere J, Charbonnel E, García-Charton JA, Goñi R, Ody D, Pérez-Ruzafa A, Reñones O, Sánchez-Jerez P, Valle C (2008) Gradients of abundance and biomass across reserve boundaries in six Mediterranean marine protected areas: evidence of fish spillover? Biological Conservation 141: 1829-1839.
Hasler H, Ott JA (2008) Diving down the reefs? Intensive diving tourism threatens the reefs of the northern Red Sea. Marine Pollution Bulletin 56: 1788-1794.
Hawkins JP, Roberts CM (1997) Estimating the carrying capacity of coral reefs for scuba diving. En: Lessios HA, Macintyre IG (Ed.) Proceedings 8th International Coral Reef Symposium, Vol. 2. Panama: 8th International Coral Reef Symposium Executive Committee, Smithsonian Tropical Research Institute: 1923-1926.
Hawkins JP, Roberts CM, Kooistra D, Buchan K, White S (2005) Sustainability of scuba diving tourism on coral reefs of Saba. Coastal Management 33: 373–387.
Hogg K, Noguera-Méndez P, Semitiel-García M, Giménez-Casalduero M (2013) Marine protected area governance: Prospects for co-management in the European Mediterranean. Advances in Oceanography and Limnology 4: 241-259.
Holland DS, Brazee RJ (1996) Marine reserves for fisheries management. Marine Resource Economics 11: 157-171.
Hunt CV, Harvey JJ, Miller A, Johnson V, Phongsuwan N (2013) The Green Fins approach for monitoring and promoting environmentally sustainable scuba diving operations in South East Asia. Ocean & Coast Manage 78: 35-44.
Irigoyen AJ, Galván DE, Venerus LA, Parma AM (2013) Variability in abundance of temperate reef fishes estimated by visual Census. PLoS ONE 8(4): e61072.
Katsanevakis S, Stelzenmüller V, South A, Sørensen TK, Jones PJS, Kerr S, Badalamenti F, Anagnostou C, Breen P, Chust G, D’Anna G, Duijn M, Filatova T, Fiorentino F, Hulsman H, Johnson K, Karageorgis AP, Kröncke I, Mirto S, Pipitone C, Portelli S, Qiu W, Reiss W, Sakellariou D, Salomidi M, van Hoof L, Vassilopoulou V, Vega Fernández T, Vöge S, Weber A, Zenetos A, ter Hofstede R (2011) Ecosystem-based marine spatial management: review of concepts, policies, tools, and critical issues. Ocean Coastal Management 54: 807-820.
Kellner JB, Tetreault I, Gaines SD, Nisbet RM (2007) Fishing the line near marine reserves in single and multispecies fisheries. Ecological Applications 17: 1039–1054.
Ku KC, Chen T-C (2013) A conceptual process-based reference model for collaboratively managing recreational scuba diving in Kenting National Park. Marine Policy 39: 1-10.

- 94 -
Kulbicki M (1998) How the acquired behaviour of commercial reef fishes may influence the results obtained from visual censuses. Journal of Experimental Marine Biology and Ecology 222: 11–30.
Kulbicki M, Sarramégne S (1999) Comparison of density estimates derived from strip transect and distance sampling for underwater visual censuses: a case study of Chaetodontidae and Pomacanthidae. Aquatic Living Resources 12: 315-325.
Lester SE, Halpern BS, Grorud-Colvert K, Lubchenco J, Ruttenberg BI, Gaines SD, Airamé S, Warner RR (2009) Biological effects within no-take marine reserves: a global synthesis. Marine Ecology Progress Series 384: 33-46.
Lombardi C, Cocito S, Gambi MC, Cisterna B, Flach F, Taylor PD, Keltie K, Freer A, Cusack M (2011) Effects of ocean acidification on growth, organic tissue and protein profile of the Mediterranean bryozoan Myriapora truncata. Aquatic Biology 13: 251-262.
Lubchenco J, Palumbi SR, Gaines SD, Andelman S (2003) Plugging a hole in the ocean: the emerging science of marine reserves. Ecological Applications 13:S3–S7.
Maliao RJ, Webb EL, Jensen KR (2004) A survey of stock of the donkey’s ear abalone, Haliotis asinina L. in the Sagay Marine Reserve, Philippines: evaluating the effectiveness of marine protected area enforcement. Fisheries Research 66: 343-353.
Mangubhai S, Saleh M, Suprayitno, Muljadi A, Purwanto, Rhodes KL, Tjandra K (2011) Do not stop: The importance of seamless monitoring and enforcement in an Indonesian marine protected area. Journal of Marine Biology 2011, art. ID 501465: 1-11 (doi:10.1155/2011/501465).
Maynou F, Morales-Nin B, Cabanellas-Reboredo M, Palmer M, García E, Grau AM (2013) Small-scale fishery in the Balearic Islands (W Mediterranean): A socio-economic approach. Fisheries Research 139: 11-17.
McCay BJ, Jones PJS (2011) Marine Protected Areas and the governance of marine ecosystems and fisheries. Conservation Biology 25: 1130-1133.
McClanahan TR, Nicholas A, Graham J, Jacqulyn MC, MacNeil MA (2007) Toward pristine biomass reef fish recovery in coral reef marine protected areas in Kenya. Ecological Applications 17: 1055–1067.
Merino G, Maynou F, Boncoeur J (2009) Bioeconomic model for three-zone Marine Protected Area: a case study of Medes Islands (northwest Mediterranean). ICES Journal of Marine Science 66: 147-154.
Micheli F, Benedetti-Cecchi L, Gambaccini S, Bertocci I, Borsini C, Osio GC, Romano F (2005) Cascading human impacts,

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 95 -
marine protected areas, and the structure of Mediterranean reef assemblages. Ecological Monographs 75: 81-102.
Milazzo M, Anastasi I, Willis TJ (2006) Recreational fish feeding affects coastal fish behaviour and increases frequency of predation on damselfish (Chromis chromis) nests. Marine Ecology Progress Series 310: 165-172.
Milazzo M, Chemello R, Badalamenti F, Camarda R, Riggio S (2002) The impact of human recreational activities in marine protected areas: what lessons should be learnt in the Mediterranean Sea? Marine Ecology 23: 280–290.
Mora C, Andréfouët S, Costello MJ, Kranenburg C, Rollo A, Veron J, Gaston KJ, Myers RA (2006) Coral reefs and global network of marine protected areas. Science 312: 1750–1751.
Murawski SA, Rago PJ, Fogarty MJ (2004) Spillover effects from temperate marine protected areas. In: Aquatic Protected Areas as Fisheries Management Tools (Proceedings of the American Fisheries Society Symposium 42, Quebec, 11-12 August, 2003). American Fisheries Society, Bethesda, pp. 167– 184.
Murawski SA, Wigley SE, Fogarty MJ, Rago PJ, Mountain DG (2005) Effort distribution and catch patterns adjacente to temperate MPAs. ICES Journal of Marine Science 62: 1150–1167.
Myers RA, Worm B (2003) Rapid worldwide depletion of predatory fish communities. Nature 423: 280-283.
Oedekoven CS, Buckland ST, Mackenzie ML, Evans KO, Burger LW (2013) Improving distance sampling: accounting for covariates and non-independency between sampled sites. Journal of Applied Ecololgy 50: 786-793.
Pauly D, Christensen V, Guénette S, Pitcher TJ, Sumaila UR, Walters CJ, Watson R, Zeller D (2002) Towards sustainability in world fisheries. Nature 418: 689-695.
Pauly D, Watson R, Alder J (2005) Global trends in world fisheries: impacts on marine ecosystems and food security. Philosophical Transactions of the Royal Society B 360:5-12.
Pelletier D, García-Charton JA, Ferraris J, David G, Thébaud O, Letourneur Y, Claudet J, Amand M, Kulbicki M, Galzin R (2005) Designing indicators for evaluating the impact of Marine Protected Areas on coral reefs ecosystems: a multidisciplinary standpoint. Aquatic Living Resources 18: 15-33.
Pérez de Oliveira L (2013) Fishers as advocates of marine protected areas: a case study from Galicia (NW Spain). Marine Policy 41: 95-102.
Pérez-Ruzafa A, Calvín JC, Marcos C, Marín A, Pérez-Ruzafa IM, Terrados J, Ros JD (1991) Contribución al estudio de las comunidades bentónicas del cabo de Palos (Murcia, SE de

- 96 -
España) y su cartografía bionómica. Actas V Simp. Ibér. Estud. Bentos Mar. 1: 385-420.
Pérez-Ruzafa A, García-Charton JA, Espejo-Cayuela C, Herrero-Pérez A, Lawrence KE, Marcos-Diego C, Polti S, Cánovas-García F (2003) Estudios de seguimiento de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
Pérez-Ruzafa A, García-Charton JA, Herrero-Pérez A, Espejo-Cayuela C, González-Wangüemert M, Marcos C (2004) Estudios de seguimiento de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
Pérez-Ruzafa A, Marcos C, García-Charton JA, González-Wangüemert M, Polti S, Vera J, Clemente C (2002) Seguimiento de la Reserva Marina de Cabo de Palos – Islas Hormigas. Valoración del impacto de los buceadores. de la Reserva Marina de Cabo de Palos-Islas Hormigas. Universidad de Murcia y Consejería de Agricultura y Agua de la Comunidad Autónoma de la Región de Murcia.
Pérez-Ruzafa A, Martín E, Marcos C, Zamarro JM, Stobart B, Harmelin-Vivien M, Polti S, Planes S, García-Charton JA, González-Wangüemert M (2008) Modelling spatial and temporal scales for spillover and biomass exportation from MPAs and their potential for fisheries enhancement. Journal of Nature Conservation 16: 234-255.
Pinnegar JK, Polunin NVC, Francour P, Badalamenti F, Chemello R, Harmelin-Vivien ML, Hereu B, Milazzo M, Zabala M, D’Anna G, Pipitone C (2000) Trophic cascades in benthic marine ecosystems: lessons for fisheries and protected-area management. Environmental Conservation 27: 179-200.
Pita C, Pierce GJ, Theodossius I, Macpherson K (2011) An overview of commercial fishers’ attitudes towards marine protected areas. Hydrobiologia 670: 289-306.
Prato G, Guidetti P, Bartolini F, Mangialajo L, Francour P (2013) The importance of high-level predators in marine protected area management: Consequences of their decline and their potential recovery in the Mediterranean context. Advances in Oceanography and Limnology 4: 176-193.
Roberts CM, Bohnsack JA, Gell F, Hawkins JP, Goodridge R (2001) Effects of marine reserves in adjacent fisheries. Science 294: 1920-1923.
Rodolfo-Metalpa R, Lombardi C, Cocito S, Hall-Spencer JM, Gambi MC (2010) Effects of ocean acidification and high temperatures on the bryozoan Myriapora truncata at natural CO2 vents. Marine Ecology, an Evolutionary Perspective 31: 447-456.

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 97 -
Roncin N, Alban F, Charbonnel E, Crec’hriou R, Modino R, Culioli JM, Dimech M, Goñi R, Guala I, Higgins R, Lavisse E, Le Diréach L, Luna B, Marcos C, Maynou F, Pascual J, Person J, Smith P, Stobart B, Szelianszky E, Valle C, Vaselli S, Boncoeur J (2008) Uses of ecosystem services provided by MPA: how much do they impact the local economy? A Southern Europe perspective. Journal of Nature Conservation 16: 256-270.
Rouphael AB, Inglis GJ (2001) ‘Take only photographs and leave only footprints’? An experimental study of the impacts of underwater photographers on coral reef dive sites. Biological Conservation 100: 281–287.
Rudd MA, Tupper MH (2002) The impact of Nassau grouper size and abundance on scuba diver site selection and MPA economics. Coastal Management 30: 133–151.
Russ G, Alcala A (2010) Decadal-scale rebuilding of predator biomass in Philippine marine reserves. Oecologia 163: 1103-1106.
Russ GR, Alcala AC (2004) Marine reserves: long-term protection is required for full recovery of predatory fish populations. Oecologia 138: 622-627.
Sadovy de Mitcheson Y, Craig MT, Bertoncini AA, Carpenter KE, Cheung WWL, Choat JH, Cornish AS, Fennessy ST, Ferreira BP, Heemstra PC, Liu M, Myers RF, Pollard DA, Rhodes KL, Rocha LA, Russell BC, Samoilys MA, Sanciangco J (2012) Fishing groupers towards extinction: a global assessment of threats and extinction risks in a billion dollar fishery. Fish and Fisheries, doi: 10.1111/j.1467-2979.2011.00455.x
Sala E, Garrabou J, Zabala M (1996) Effects of diver frequentation on Mediterranean sublittoral populations of the bryozoan Pentapora fascialis. Marine Biology 126: 451–459.
Salas S & Gaertner D (2004) The behavioural of fishers: management implications. Fish and Fisheries 5: 153-167.
Samoilys MA, Martin-Smith KM, Giles BG, Cabrera B, Anticamara JA, Brunio EO, Vincent ACJ (2007) Effectiveness of five small Philippines’ coral reef reserves for fish populations depends on site-specific factors, particularly enforcement history. Biological Conservation 136: 584-601.
Sandin SA, Smith JE, DeMartini EE, Dinsdale EA, Donner SD, Friedlander AM, Konotchick T, Malay M, Maragos JE, Obura D, Pantos O, Paulay G, Richie M, Rohwer F, Schroeder RE, Walsh S, Jackson JBC, Knowlton N, Sala E (2008). Baselines and degradation of coral reefs in the Northern Line Islands. PLoS ONE 3: 1-11.

- 98 -
Sandrini-Neto L, Camargo MG (2011) GAD (General ANOVA Design), an R package for ANOVA designs from the general principles. Disponible en CRAN.
Schunter C, Carreras-Carbonell J, Planes S, Sala E, Ballesteros E, Zabala M, Harmelin JG, Harmelin-Vivien M, Macpherson E, Pascual M (2011) Genetic connectivity patterns in an endangered species: The dusky grouper (Epinephelus marginatus). Journal of Experimental Marine Biology and Ecology 401: 126-133.
Stelzenmüller V, Maynou F, Bernard G, Cadiou G, Camilleri M, Crec’hriou R, Criquet G, Dimech M, Esparza-Alaminos O, Higgins R, Lenfant P, Pérez-Ruzafa A (2008) Spatial assessment of fishing effort around European marine reserves: implications for sucessful fisheries management. Marine Pollution Bulletin 56: 2018-2016.
Stelzenmüller V, Maynou F, Martin P (2007) Spatial assessment of benefits of a coastal Mediterranean marine protected area. Biological Conservation 136: 571–583.
Stergiou KI, Moutopoulos DK, Soriguer MC, Puente E, Lino PG, Zabala C, Monteiro P, Errazkin LU, Erzini K (2006) Trammel net catch species composition, catch rates and metiers in southern European waters: a multivariate approach. Fisheries Research 79: 170–182.
Stobart B, Warwick R, González C, Mallol S, Díaz D, Reñones O, Goñi R (2009) Long-term and spillover effects of a marine protected area on an exploited fish community. Marine Ecology Progress Series 384: 47-60.
Tzanatos E, Somarakis S, Tserpes G, Koutsikopoulos C (2006) Identifying and classifying small-scale fisheries métiers in the Mediterranean: a case study in the Patraikos Gulf, Greece. Fisheries Research 81: 158-168.
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press.
Unsworth RKF, Powell A, Hukom F, Smith DJ (2007) The ecology of Indo-Pacific grouper (Serranidae) species and the effect of small scale no take area on grouper assemblage, abundance and size frequency distribution. Marine Biology 152: 243-254.
Uyarra MC, Côté IM (2007) The quest for cryptic creatures: impacts of species-focused recreational diving on corals. Biological Conservation 136: 77–84.
Vandeperre F, Higgins RM, Sánchez-Meca J, Maynou F, Goñi R, Martín-Sosa P, Pérez-Ruzafa A, Afonso P, Bertocci I, Crec’hriou R, D’Anna G, Dimech M, Dorta C, Oscar-Esparza O, Falcón JM, Forcada A, Guala I, Le Direach L, Concepción M, Ojeada-Martínez C, Pipinote C, Schembri PJ, Stelzenmüller V, Stobart B, Santos RS (2011) Effects of

Estudios de seguimiento de la reserva marina de C. Palos – I. Hormigas
2013
- 99 -
no-take area size and age of marine protected áreas on fisheries yields: a meta-analytical approach. Fish and Fisheries 12: 412-426.
Walters RDM, Samways MJ (2001) Sustainable dive ecotourism on a South African coral reef. Biodiversity and Conservation 10: 2167–2179.
Warner TE, Pomeroy RS (2012) Creating compliance: A cross-sectional study of the factors associated with marine protected area outcomes. Marine Policy 36: 922-932.
Wilcox C, Pomeroy C (2003) Do commercial fishers aggregate around marine reserves? Evidence from Big Creek Marine Ecological Reserve, central California. North American Journal of Fisheries Management 23: 241–250.
Willis TJ, Millar RB, Babcock RC, Tolimieri N (2003) Burdens of evidence and the benefits of marine reserves: Putting Descartes before Des horse? Environmental Conservation 30: 97–103.
Zakai D, Chadwick-Furman NE (2002) Impacts of intensive recreational diving on reef corals at Eilat, northern Red Sea. Biological Conservation 105: 179-187.

- 100 -





























