Embed Size (px)
Citation preview

EX AFRICA SEMPER ALIQUID NOVI – DIE VERWEEFDE GENEALOGIEË VAN
MIKOBAKTERIEË EN DIE MENS
Prof Nicolaas Claudius Gey van Pittius VI September 2015

EX AFRICA SEMPER ALIQUID NOVI – DIE VERWEEFDE GENEALOGIEË VAN MIKOBAKTERIEË EN DIE MENS
Intreerede gelewer op 1 September 2015
Prof Nicolaas C Gey van Pittius VIAfdeling Molekulêre Biologie en MensgenetikaDepartement Biomediese WetenskappeFakulteit Geneeskunde en GesondheidswetenskappeUniversiteit Stellenbosch
Geredigeer deur US TaalsentrumGedruk deur SUN MeDIAISBN: 978-0-7972-1589-4Kopiereg © 2015 Nicolaas C Gey van Pittius

1
BIOGRAFIe
Professor Nicolaas Claudius (Nico) Gey van Pittius VI is in Pretoria gebore, maar het in Durban
grootgeword. Hy is tans Visedekaan: Navorsing in die Fakulteit Geneeskunde en Gesondheidswetenskappe en ’n volle professor in Molekulêre Biologie en Mensgenetika in die Departement Biomediese Weten-skappe aan die Universiteit Stellenbosch. Hy is ook ’n kernlid van die Departement van Wetenskap en Tegnologie/Nasionale Navorsingstigting se Sentrum van Uitnemendheid in Biomediese Tuberkulosenavorsing, sowel as van die Mediese Navorsingsraad se Sentrum vir Tuberkulosenavorsing by die Universiteit Stellenbosch.
Nadat hy die grade BSc (cum laude), BScHons (cum laude) en MSc van die Universiteit van Potchefstroom en ’n PhD-graad van die Universiteit Stellenbosch verwerf het, werk hy ’n ruk lank as nadoktorale navorsingsgenoot, voordat hy eers as senior navorser, later medeprofessor en nou volle professor in die Departement Biomediese Wetenskappe aangestel word.
Hy het deeltyds aanhou studeer en ’n sertifikaat in Intellektuele Goederereg (cum laude) en die graad LLB (cum laude) van die Universiteit van Suid-Afrika, sowel as die grade LLM en MBA van die Universiteit Stellenbosch verwerf. Hy is tans vir die graad MPhil in die Studie van Wetenskap en Tegnologie by Stellenbosch ingeskryf. As navorser beskik hy oor ’n B-gradering van die Nasionale Navorsingstigting. Hy is in 2006 (op die vroeë ouderdom van 33) tot lid van die Suid-Afrikaanse Akademie vir Wetenskap en Kuns, en in 2010 (op 37-jarige ouderdom) tot lid van ASSAf (oftewel die Wetenskapakademie van
Suid-Afrika) verkies. Hy dien ook in verskeie ander nasionale en internasionale wetenskaplike verenigings en komitees, waaronder die Amerikaanse Vereniging vir Mikrobiologie (ASM) en die Internasionale Unie teen Tuberkulose en Longsiekte (IUATLD).
As ’n molekulêre mikobakterioloog, bestudeer prof Gey van Pittius die afgelope 18 jaar reeds tuberkulose, met ’n bepaalde klem op die evolusiemeganismes van die mikobakterieë en die ontwikkeling van mikobakteriële patogenisiteit en middelweerstandigheid. Hy is mede-outeur van meer as 80 artikels en boekhoofstukke oor verskeie aspekte van tuberkulose, met meer as 3 300 sitasies (en ’n huidige h-indeks van 31), en is mede-uitvinder van twee toegestane en drie voorlopige patente op die gebied. Hy het oor die jare vir menige nagraadse student as studieleier opgetree en nagraadse modules in intellektuele goederereg, bio-informatika, genomika en genoomvariasie aangebied. Sy werk is al by meer as 40 internasionale en nagenoeg 100 nasionale konferensies en byeenkomste in sowel mondelinge as plakkaatvorm aangebied, en hy tree gereeld as gasspreker by verskeie instellings wêreldwyd op.
Vir ’n lys van sy publikasies, skandeer asseblief:

2
DANKBeTUIGING
Ek het soveel wonderlike invloede in my lewe gehad dat ek bang is dat iemand per abuis uit ’n lys
dankbetuigings weggelaat kan word. Tog verleen ek graag erkenning aan veral die volgende persone, wat oor baie jare ’n enorme bydrae tot my persoonlike en professionele ontwikkeling gelewer het:
My vrou, Méshelle. Baie dankie vir jou liefde en ondersteuning. Ek weet dit was vir jou by tye swaar met ’n man wat aanhou studeer en hom in die akademie verdiep. Jy het baie opgeoffer – finansieel én wat tyd en aandag betref. Ek is baie dankbaar daarvoor en is baie lief vir jou.
My pa, Nic, en ma, Kiki. Baie dankie vir die goeie opvoeding wat Pa en Ma vir my gegee het; dat julle my gehelp het om verder te kon studeer; dat julle my belangstelling in boeke, in studie en in die akademie en wetenskap altyd ondersteun het en dit nooit van my weerhou het nie. Dankie dat julle my van kleins af toegelaat het om te wees wie ek is en my nie in ’n rigting gedwing het waarin ek nie wou wees nie. Ek is baie lief vir julle.
My ouma, Maaike. Wat ’n ongelooflike standvastige en liefdevolle mens is my ouma nie – iemand wat my geleer het om altyd vriendelik, toegeeflik, vrygewig en selfstandig te wees en altyd positief te bly, al gaan dit hoe swaar. Baie dankie vir Ouma se wysheid en liefde. Dit is so ’n groot voorreg om hierdie geleentheid met Ouma te kan deel. Ek is baie lief vir Ouma.
My skoonma, Lida, my suster, Marissa, en my broer, Hugo. Julle beteken vir my soveel in my lewe. Ek is baie lief vir julle en baie dankbaar vir julle ondersteuning en liefde oor al die jare.
My close colleague and dear friend Rob Warren, one of the greatest scientists I have ever known. We have been working side by side for most of my academic career. I would like to extend my deepest gratitude to you for all you have done for me over many years. I have really learnt so much from you and am indebted to you for showing me how to be a true scientist. You have been an excellent mentor, a stimulating colleague and a great friend.
Paul van Helden, I am indebted to you for helping to shape my career and for continued advice and support, whether in the realm of the intellectual or the personal. It has been a pleasure to be associated with you and your Centre over the past 18 years.
Albert Beyers, ek het nooit die kans gekry om vir jou dankie te sê dat jy soveel jare gelede vir my die geleentheid gegee het om vir jou te kom werk nie. Ek wonder baie keer waar ek nou sou gewees het indien jy my nie destyds as navorsingsassistent aangestel het en my aan die wonderlike wêreld van tuberkulosenavorsing bekend gestel het nie. Ek mis jou baie en is net jammer dat jy nie kon sien hoe ons werk gegroei en gedy het nie.
Die lede van die Mikobaktomika en Molekulêre Epidemiologie groepe, studente, nadoktorale genote en navorsers, in die verlede en die hede – te veel om te noem. Dankie aan almal vir jul vriendskap en bydraes tot my lewe oor ‘n tydperk van 18 jaar. Dit was wonderlik om soveel ongelooflike ontdekkings met jul te deel.
Al my kollegas in die Afdeling Molekulêre Biologie en Mensgenetika (voorheen die Departement Mediese Biochemie) aan die Universiteit Stellenbosch, my medewerkers regoor die wêreld, Jimmy, Marietjie, Eben, Therese en Sonja, ander kollegas en vriende in die Dekaansafdeling en in die Fakulteit, en my familie, mentors, vriende, dosente en onderwysers, wat almal so baie tot my persoonlike en akademiese lewe bygedra het.
Ten slotte wil ek graag hierdie intreerede opdra aan my twee oupas, Nico C Gey van Pittius IV en Hugo van der Bijl. Ek het so baie eienskappe van julle albei geërf, en ek wens julle kon vandag hier wees om hierdie geleentheid saam met my te vier. Ek mis julle baie en hoop ek het julle darem trots gemaak …
Deus vobiscum

3
eX AFRICA SeMPeR ALIQUID NOVI –DIe VeRWeeFDe GeNeALOGIeË VAN
MIKOBAKTeRIeË eN DIe MeNS
There is a dread disease which so prepares the victim, as it were, for death; which so refines it of its grosser aspect, and throws around familiar looks, unearthly indications of the coming change – a dread disease, in which the struggle between soul and body is so gradual, quiet, and solemn, and the result so sure, that day by day, and grain by grain, the mortal part wastes and withers away, so that the spirit grows light and sanguine with its lightening load, and, feeling immortality at hand, deems it but a new term of mortal life; a disease in which death takes the glow and hue of life, and life the gaunt and grisly form of death; a disease which medicine never cured, wealth warded off, or poverty could boast exemption from; which sometimes moves in giant strides, and sometimes at a tardy pace; but, slow or quick, is ever sure and certain.
’n Beskrywing van tuberkulose uit Nicholas Nickleby – Charles Dickens (1870)
Figuur 1 – Relatiewe wêreldwye sterftesyfers weens infeksiesiektes oor die afgelope 200 jaar (uit Paulson, 2013)
INLeIDING
Mycobacterium tuberculosis is die oorsaaklike agens van tuberkulose (TB), ’n siekte wat vir miljoene
mense wêreldwyd ’n groot gesondheidsgevaar inhou en steeds een van die dodelikste infeksiesiektes ter wêreld is. Tuberkulose word as die noodlottigste infeksiesiekte in menseheugenis beskou en het slegs oor die afgelope 200 jaar na raming meer as ’n miljard mense se lewens geëis (Paulson, 2013) (sien Figuur 1). Daar word voorts geraam dat ongeveer ’n derde van die wêreldbevolking (sowat twee miljard mense) tans met M. tuberculosis geïnfekteer is (WGO, 2014a). In 1993 was kommer oor die skynbaar onkeerbare toename in die tuberkulose-epidemie in die moderne tyd so groot dat die Wêreldgesondheidsorganisasie (WGO) die ongekende stap gedoen het om tuberkulose tot ’n internasionale noodtoestand te verklaar (WGO, 1993). Meer as twintig jaar later is ons nie veel beter daaraan toe nie: In 2013 alleen het ’n geraamde nege miljoen mense wêreldwyd TB opgedoen en het 1.5 miljoen (van wie 360 000 MIV-positief was) daaraan gesterf (WGO, 2014b). Die BRICS-lande (Brasilië, die Russiese Federasie, Indië, China en Suid-Afrika) is gesamentlik verantwoordelik vir bykans 50% van alle aangemelde TB-gevalle wêreldwyd, en 90% van TB-gevalle kom in die ontwikkelende wêreld voor (WGO, 2014b). Die gunstigste ramings dui daarop dat ongeveer een uit elke 100 mense (1 000 per 100 000 lede van die bevolking) in Lesotho, Suid-Afrika en Swaziland jaarliks aktiewe TB opdoen, wat dit die streek met die hoogste voorkoms van tuberkulose ter wêreld maak (sien Figuur 2). Hierdie syfer is soortgelyk aan dié wat in die vroeë 1800’s op die kruin van die tuberkulose-epidemie in industriële Europa, voor die aanbreek van die era van antibiotika, aangemeld is (Daniel, 2006).
Destyds was sterftesyfers in Londen, Stockholm en Hamburg nagenoeg 800 tot 1 000 per 100 000 lede van die bevolking per jaar (Daniel, 2006).
Daar ís ’n paar positiewe ontwikkelings – volskaalse diagnose en behandeling van tuberkulose het die wêreldwye TB-sterftesyfer tussen 1990 en 2013 met 45% laat daal, en doeltreffende diagnose en behandeling het tussen 2000 en 2013 na raming sowat 37 miljoen lewens gered (WGO, 2014b). Dit is ook goeie nuus dat TB-syfers klaarblyklik wêreldwyd stelselmatig afneem. Tog is hierdie afname uiters stadig en wissel dit tussen verskillende wêrelddele. Boonop dreig die onlangse verskyning van multimiddelweerstandige M. tuberculosis-stamme om hierdie tendens om te keer. Suid-Afrika bly die land met die hoogste TB-insidensie en -prevalensie ter wêreld, wat ’n reuse-las op die gesondheid van die bevolking plaas (sien Tabel 1).
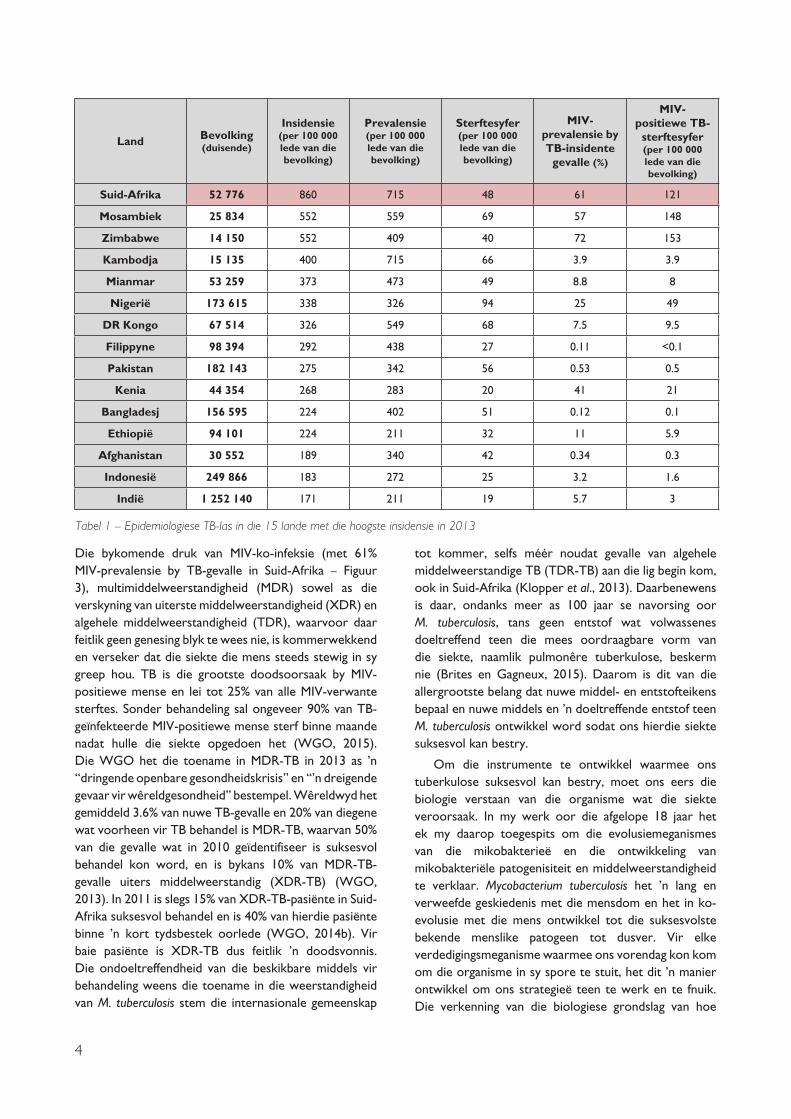
4
Die bykomende druk van MIV-ko-infeksie (met 61% MIV-prevalensie by TB-gevalle in Suid-Afrika – Figuur 3), multimiddelweerstandigheid (MDR) sowel as die verskyning van uiterste middelweerstandigheid (XDR) en algehele middelweerstandigheid (TDR), waarvoor daar feitlik geen genesing blyk te wees nie, is kommerwekkend en verseker dat die siekte die mens steeds stewig in sy greep hou. TB is die grootste doodsoorsaak by MIV-positiewe mense en lei tot 25% van alle MIV-verwante sterftes. Sonder behandeling sal ongeveer 90% van TB-geïnfekteerde MIV-positiewe mense sterf binne maande nadat hulle die siekte opgedoen het (WGO, 2015). Die WGO het die toename in MDR-TB in 2013 as ’n “dringende openbare gesondheidskrisis” en “’n dreigende gevaar vir wêreldgesondheid” bestempel. Wêreldwyd het gemiddeld 3.6% van nuwe TB-gevalle en 20% van diegene wat voorheen vir TB behandel is MDR-TB, waarvan 50% van die gevalle wat in 2010 geïdentifiseer is suksesvol behandel kon word, en is bykans 10% van MDR-TB-gevalle uiters middelweerstandig (XDR-TB) (WGO, 2013). In 2011 is slegs 15% van XDR-TB-pasiënte in Suid-Afrika suksesvol behandel en is 40% van hierdie pasiënte binne ’n kort tydsbestek oorlede (WGO, 2014b). Vir baie pasiënte is XDR-TB dus feitlik ’n doodsvonnis. Die ondoeltreffendheid van die beskikbare middels vir behandeling weens die toename in die weerstandigheid van M. tuberculosis stem die internasionale gemeenskap
tot kommer, selfs méér noudat gevalle van algehele middelweerstandige TB (TDR-TB) aan die lig begin kom, ook in Suid-Afrika (Klopper et al., 2013). Daarbenewens is daar, ondanks meer as 100 jaar se navorsing oor M. tuberculosis, tans geen entstof wat volwassenes doeltreffend teen die mees oordraagbare vorm van die siekte, naamlik pulmonêre tuberkulose, beskerm nie (Brites en Gagneux, 2015). Daarom is dit van die allergrootste belang dat nuwe middel- en entstofteikens bepaal en nuwe middels en ’n doeltreffende entstof teen M. tuberculosis ontwikkel word sodat ons hierdie siekte suksesvol kan bestry.
Om die instrumente te ontwikkel waarmee ons tuberkulose suksesvol kan bestry, moet ons eers die biologie verstaan van die organisme wat die siekte veroorsaak. In my werk oor die afgelope 18 jaar het ek my daarop toegespits om die evolusiemeganismes van die mikobakterieë en die ontwikkeling van mikobakteriële patogenisiteit en middelweerstandigheid te verklaar. Mycobacterium tuberculosis het ’n lang en verweefde geskiedenis met die mensdom en het in ko-evolusie met die mens ontwikkel tot die suksesvolste bekende menslike patogeen tot dusver. Vir elke verdedigingsmeganisme waarmee ons vorendag kon kom om die organisme in sy spore te stuit, het dit ’n manier ontwikkel om ons strategieë teen te werk en te fnuik. Die verkenning van die biologiese grondslag van hoe
Tabel 1 – Epidemiologiese TB-las in die 15 lande met die hoogste insidensie in 2013
Land Bevolking (duisende)
Insidensie(per 100 000 lede van die bevolking)
Prevalensie(per 100 000 lede van die bevolking)
Sterftesyfer(per 100 000 lede van die bevolking)
MIV-prevalensie by TB-insidente
gevalle (%)
MIV-positiewe TB-
sterftesyfer(per 100 000 lede van die bevolking)
Suid-Afrika 52 776 860 715 48 61 121
Mosambiek 25 834 552 559 69 57 148
Zimbabwe 14 150 552 409 40 72 153
Kambodja 15 135 400 715 66 3.9 3.9
Mianmar 53 259 373 473 49 8.8 8
Nigerië 173 615 338 326 94 25 49
DR Kongo 67 514 326 549 68 7.5 9.5
Filippyne 98 394 292 438 27 0.11 <0.1
Pakistan 182 143 275 342 56 0.53 0.5
Kenia 44 354 268 283 20 41 21
Bangladesj 156 595 224 402 51 0.12 0.1
Ethiopië 94 101 224 211 32 11 5.9
Afghanistan 30 552 189 340 42 0.34 0.3
Indonesië 249 866 183 272 25 3.2 1.6
Indië 1 252 140 171 211 19 5.7 3

5
Figuur 2 – Geraamde wêreldwye TB-insidensiesyfers in 2013 (WGO, 2014b)
Figuur 3 – Geraamde wêreldwye MIV-prevalensie by nuwe en relapsgevalle van TB in 2013 (WGO, 2014b)

6
mikobakterieë sulke suksesvolle patogene geword het, bied ons die geleentheid om moontlik hierdie intieme verhouding op ’n meer toegespitste wyse te versteur, wat ons hopelik uiteindelik die band sal help verbreek ten einde dié oersiekte uit te wis.
DIe GeSKIeDeNIS VAN TUBeRKULOSe
The longer you can look back, the further you can look forward.
Winston Churchill (in ’n toespraak by die Koninklike Mediese Kollege – Maart 1944)
Tuberkulose (ook bekend as tering of ftisis) teister die mensdom reeds deur sy hele geskiedenis
én voorgeskiedenis (Daniel, 2006). Die siekte was skynbaar so vroeg as 2 700 vC reeds in Antieke China teenwoordig, waar keiser Shen Nung dit in ’n mediese teks beskryf het (Roberts en Buikstra, 2003; Crawford en Campbell, 2012). Die eerste verwysings na tuberkulose in Indië word in die antieke Sanskrit-tekste genaamd die Vedas aangetref, waarvan die heel oudste (Rigveda, vanaf 1 500 vC) die siekte yaksma (‘geleidelike aftakeling’) noem (Zysk, 1998). Een van die eerste klassieke Mediterreense tekste wat van die siekte melding maak, is The Histories deur Herodotos (450 vC), waarin hy vertel hoe een van Xerxes se generaals die krygstog teen die Spartane moes verlaat omdat hy tering opgedoen het (Herodotos, boek 7 hoofstuk 88). Trouens, in sy werk Of the Epidemics (460 vC) bestempel die vader van moderne geneeskunde, die Griekse geneeskundige Hippokrates, ftisis (wat afgelei is van die Griekse phthiein – ‘om uit te teer, te vergaan’) as die mees wydverspreide siekte van sy tyd en merk hy op dat dit bykans altyd tot die dood lei (Hippokrates, soos aangehaal deur Major, 1945). ’n Aantal ander antieke vakkundiges, onder meer Plinius die jongere, Aristoteles, Galenos, Vitruvius en Aretaeus, het almal verskillende aspekte van die siekte beskryf, party tot in die fynste besonderhede. Selfs in daardie antieke tye was tuberkulose duidelik ’n bekende siekte vir genees- en vakkundiges.
Tog infekteer Mycobacterium tuberculosis die mens al sedert veel vroeër in die geskiedenis van die mensdom: By verskeie argeologiese menslike eksemplare wêreldwyd is patologiese tekens van tuberkulêre verval getipeer en M. tuberculosis- kompleks DNS opgespoor (Roberts en Buikstra, 2003). Tuberkulose kan skeletveranderinge soos die ineenstorting van werwels (oftewel Pott se siekte), periosteale reaktiewe letsels en osteomiëlitis veroorsaak, wat dit moontlik maak om langtermynlyers
aan hulle skeletoorskot uit te ken weens die skade wat die bakterieë gedurende dié individue se leeftyd aan hulle beendere aangerig het (Ortner, 1981). In ’n onlangse studie in Turkye is verslag gedoen oor ’n 500 000 jaar oue fossiel van Homo erectus wat kenmerkende TB-letsels toon (Roberts et al., 2009). Hierdie bevinding is steeds ietwat omstrede, maar indien dit gestaaf word, sou dit beteken dat die evolusie van M. tuberculosis tot voor die oorsprong van die moderne mens teruggevoer kan word (Gagneux, 2012). Die oudste onbestrede bewys van tuberkulose-infeksie by die moderne mens is in menslike oorskot uit die Neolitiese tydperk van 9 000 jaar gelede ontdek in ’n verdronke nedersetting genaamd Atlit-Yam in die oostelike Mediterreense See, digby wat vandag as Israel bekend is (Hershkovitz, et al. 2008). Egiptiese mummies met paleopatologiese veranderinge wat op Pott se siekte dui, dien as ’n ryke bron van antieke tuberkulosebewyse – van ’n 5 400 jaar oue geraamte uit die predinastiese tydperk (Crubézy et al. 1998) tot verskeie ander uit die ou, middel- en nuwe Egiptiese ryke (Nerlich et al. 1997; Zink et al. 2003). Een van die mees onlangse voorbeelde is die sogenaamde Granville-mummie uit die dodestad van Tebe in Antieke Egipte, naamlik dié van die dame Irtyersenu uit die 26ste dinastie, omstreeks 600 vC, wat aan tuberkulose gesterf het (Donoghue et al., 2010). Voorts is tuberkuleuse spondilitis gevind in eksemplare van Italië, waarvan sommige tot die Neolitiese tydperk ongeveer 4 000 vC (Formicola et al., 1987; Canci et al., 1996) en ander tot die Bronstydperk teruggevoer kan word (Canci et al., 2001). M. tuberculosis-DNS is ook opgemerk in Skandinawiese Neolitiese gebeentes in Swede wat uit 3 200–2 300 vC dagteken (Nuorala et al., 2004). Tuberkulose het egter nie net wydverspreid in die Ou Wêreld voorgekom nie, maar is ook in skeletoorskot uit die Nuwe Wêreld ontdek (Roberts en Buikstra, 2003). M. tuberculosis-DNS is byvoorbeeld in die 1 000 jaar oue longweefsel van pre-Columbiaanse mummies uit Peru en noordelike Chile gevind (Allison et al., 1973; Salo et al., 1994, Arriaza et al., 1995). Die manuskrip deur Roberts en Buikstra (2003) bied ’n omvattende uiteensetting van argeologiese tuberkulosebewyse van verskillende plekke oor die ganse Ou en Nuwe Wêreld heen. Uit die bio-argeologiese bewyse van tuberkulose blyk duidelik dat die siekte uit die antieke tyd kom, in verskillende wêrelddele posgevat het, en wydverspreid voorgekom het by menslike populasies wat vir etlike millennia geografies afgesonder van mekaar geleef het, wat te kenne gee dat die siekte moontlik selfs vroeër kon ontstaan het.

10
Gordonia aichiensis
M. gadium
M. cookii
M. fallax M. chitae M. manitobense
M. flavescens M. duvalii M. tuscia
M. vaccae
M. vanbaalenii M. aurum M. novocastrense
M. smegmatis M. goodii
M. moriokaense M. thermoresistibile
M. pulveris M. elephantis
M. confluentis M. phlei
M. brumae M. holsaticum
M. triviale M. heidelbergense
M. simiae M. genavense
M. lentiflavum M. kubicae
M. interjectum M. intermedium
M. scrofulaceum M. lacus
M. celatum M. terrae
M. nonchromogenicum M. hiberniae
M. shimoidei M. heckeshornense
M. botniense M. xenopi
M. ulcerans M. gordonae
M. asiaticum M. szulgai M. malmoense M. gastri
M. kansasii
M. intracellulare
M. komossense M. fortuitum
M. sphagni M. parafortuitum
M. gilvum M. aichiense
M. obuense M. wolinskyi M. peregrinum M. chlorophenolicum
M. chubuense M. septicum
M. alvei M. senegalense
M. farcinogenes
M. frederiksbergense M. diernhoferi
M. neoaurum M. hodleri
M. chelonae
M. microti M. bovis
M. paratuberculosis
RAPID GROWERS
SLOW GROWERS
Long helix 18
Extended helix 18
M. tuberculosis complex
M. avium complex
M. abscessus
M. doricum
M. canettii M. africanum M. tuberculosis
M. marinum M. shottsii
M. avium M. leprae
M. haemophilum
M. palustre
M. sp. JLS M. monacense
M. sp. KMS M. sp. MCS
M. acapulcensis
M. austroafricanum
Outgroup
Figuur 4 – Filogenetiese boom van die lede van die genus Mycobacterium (Gey van Pittius et al., 2006)
7
MYCOBACTERIUM TUBERCULOSIS
Mycobacterium tuberculosis, ’n staafvormige bakterie wat uitsonderlik stadig groei, is uiters goed aangepas
om stresvolle omstandighede in die liggaam te hanteer, met ’n dik, komplekse, lipiedryke selwandstruktuur wat suurtoestande kan weerstaan en ondeurdringbaar is
vir ’n aantal verbindings (Shinnick en Good, 1994). Die bakterie is ’n intrasellulêre patogeen van makrofage wat die maturasie van die fagosoom waarin dit voorkom, kan verander sodat die bakterie die mikrobisidiese effektorfunksies van die gasheersel kan systap (Clemens en Horwitz, 1995). Dit beland gewoonlik in die liggaam deur die inaseming van lugdruppels wat geïnfekteerde

8
individue uithoes. Hierdie druppels dra die bakterie tot in ’n individu se longe (Riley et al., 1959). Hierna behels die progressie van die siekte fagositose van die organismes deur middel van alveolêre makrofage, daaropvolgende oorlewing en replisering binne hierdie selle, gevolg deur die aftakeling van die longweefsel deur die gasheer se eie immuunreaksie, wat op sy beurt verspreiding na nuwe gashere tot gevolg het (McKinney et al., 1998). Meer as 80% van tuberkulosegevalle is in die vorm van pulmonêre tuberkulose, wat ook die mees aansteeklike vorm van die siekte is, en lei tot ’n gehoes, borskaspyn, koors en nagsweet, gewigsverlies (uittering), ’n swakker eetlus, uitputting en die uithoes van bloed (Hopewell, 1994).
Franciscus de le Boë (dr Sylvius) het die term ‘tuberkulose’ die eerste keer in sy Opera Medica van 1679 gebruik (Trail, 1970) met betrekking tot die tuberkels, tuberkuleuse holtes en tuberkuleuse limfnodusse wat met die siekte verbind word. Die eerste groot deurbraak in die stryd teen tuberkulose was toe ’n Engelse geneesheer, Benjamin Marten, besef het dat tuberkulose moontlik veroorsaak word deur ’n organisme wat deur die lug versprei (McKinney et al., 1998). Tot in daardie stadium het vakkundiges en geneeshere hewig debat gevoer oor of tuberkulose ’n infeksie- of oorerflike siekte is. In sy werk A New Theory of Consumption wat in 1722 verskyn het, reken Marten die etiologiese agens “may possibly be some certain Species of Animalculae or wonderfully minute living creatures that, by their peculiar Shape or disagreeable Parts are inimicable to our Nature; but, however, capable of subsisting in our Juices and Vessels …” (Dubos en Dubos, 1952). Hierdie “wonderfully minute creatures” is eers meer as 100 jaar later, teen die einde van die 1800’s, die eerste keer geïsoleer en beskryf deur die Duitse geneeskundige Robert Koch (Koch, 1882). Hy het sy ontdekking van die identifikasie van M. tuberculosis as oorsaaklike agens van tuberkulose op 24 Maart 1882 in ’n geskiedkundige toespraak voor die Berlynse Fisiologiese Vereniging bekend gemaak (McKinney et al., 1998). Tog was ’n neef van M. tuberculosis, naamlik Mycobacterium leprae – die oorsaaklike agens van melaatsheid by mense, die mikobakteriële spesie wat heel eerste geïdentifiseer is. Dié ontdekking deur Armauer Hansen was ’n deurbraak in die mediese geskiedenis, synde die heel eerste oortuigende verbintenis van ’n mikro-organisme met ’n menslike siekte (Hansen, 1874; Hansen, 1880).
DIe GeNUS MYCOBACTERIUM
Die genus Mycobacterium behoort tot die klas Actinobacteria, ’n groep guanien- en sitosien- (G+C-)
ryke Gram-positiewe bakterieë wat algemeen in grond voorkom, waar dit ’n aktiewe rol in die koolstofsiklus en ander ekologiese prosesse speel (Jang et al., 2008). Die genus bestaan tans uit sowat 130 spesies en vyf subspesies (sowel as ’n bykomende 34 spesies waaroor daar nog nie stawend gepubliseer is nie), waarvan 60% maar eers in die afgelope 15 tot 20 jaar geïdentifiseer is (sien Figuur 4). Die genus Mycobacterium bevat heelwat spesies wat voëls, vis, amfibieë en soogdiere (waaronder mense) kan infekteer, en het meganismes ontwikkel waardeur dit gasheerselle kan binnedring en daarin groei. Dus sluit die genus ’n aantal patogenies beduidende spesies in wat menslike en dieresiektes veroorsaak, onder meer tuberkulose (M. tuberculosis en M. bovis) (Michel et al., 2007; Espie et al., 2009); melaatsheid (M. leprae) (Gelber, 1994); Buruli-ulkus (M. ulcerans) (Sizaire et al., 2006); pulmonêre en niepulmonêre infeksies, hoofsaaklik by immuunkompromitteerde individue (M. avium) (Havlir, 1994), en paratuberkulose of Johne se siekte (’n chroniese granulomateuse enteritis by herkouerdiere wat deur M. paratuberculosis veroorsaak word) (Chiodini et al., 1984).
Die genus Mycobacterium het na bewering meer as 150 miljoen jaar gelede ontstaan. Dié hipotese is gegrond op die baie bepaalde habitatvereistes én huidige geografiese verspreiding van M. ulcerans in Wes-Afrika, Australië en Suid-Amerika – endemiese streke waarvan die geografiese grense so ver verwyderd is van mekaar dat dit feitlik onmoontlik is dat dié spesie hierdie gebiede in die onlangse verlede kon bereik het. Hierdie streke was laas aangrensend as deel van die oervastelandsmassa Gondwanaland, wat van ongeveer 510 tot 180 miljoen jaar gelede in die Juratydperk bestaan het (Daniel, 2006). Dít dui daarop dat die spesie moes ontwikkel het voordat die afsonderlike vastelande as gevolg van kontinentskuiwing gevorm het.
Mikobakteriële spesies word in twee groepe verdeel: (1) snelgroeiende spesies wat binne minder as sewe dae kolonies op vaste media kan vorm, en (2) stadig groeiende spesies wat eers ná sewe dae of langer kolonies op vaste media kan vorm (Shinnick en Good, 1994; Springer et al., 1996). Die meeste van die stadig groeiende mikobakterieë is patogeniese spesies wat oënskynlik meer onlangs ontwikkel het, met inbegrip van die spesies wat tuberkulose, melaatsheid, paratuberkulose en ander siektes veroorsaak. Daarteenoor blyk die snelgroeiendes nouer verbind te wees met die progenitor van die mikobakterieë en ander nou verwante genusse, waaronder Corynebacterium, Nocardia, Rhodococcus en ander aktinomisete genusse soos Streptomyces (Wayne en Kubica, 1986), wat hoofsaaklik niepatogenies is en meestal as saprofiete in

9
die omgewing voorkom (Shinnick en Good, 1994). Die stadig groeiende lede het dus duidelik in die een of ander stadium van die evolusie van die genus Mycobacterium die vermoë ontwikkel om in ’n intrasellulêre omgewing te leef en te óórleef, en het tot opportunistiese patogene (soos M. avium) sowel as obligatoriese patogene (soos M. tuberculosis en M. leprae) ontwikkel.
DIe MYCOBACTERIUM TUBERCULOSIS-KOMPLeKS
Mycobacterium tuberculosis behoort tot ’n groter groep nou verwante, stadig groeiende
mikobakteriële spesies wat tuberkulose veroorsaak binne die genus Mycobacterium, oftewel die Mycobacterium tuberculosis-kompleks (MTBK) of tuberkelbasille. Die M. tuberculosis-kompleks bestaan uit genotipies en fenotipies kenmerkende linies wat deur klonale uitbreiding vanuit ’n gemeenskaplike progenitor ontwikkel het (Brites en Gagneux, 2015). Op grond van gasheeraanpassingsprofiele en fenotipes, bestaan die kompleks uit drie verskillende subgroepe, naamlik (1) die gladde tuberkelbasille (waaronder Mycobacterium canetti en ander gladde tuberkelbasille wat slegs in Oos-Afrika voorkom), waarvan die werklike gasheerverspreiding steeds onbekend is (Gutierrez et al., 2005), (2) die mens-aangepaste spesies (waaronder M. tuberculosis en M. africanum) (Brites en Gagneux, 2015), en (3) die dier-aangepaste spesies, wat insluit die sjimpansee-basil; M. mungi (die muishond-basil); die dassie-basil; M. suricattae (die meerkat-basil); M. pinnipedii (die rob-basil); M. microti (die vleirot-basil); M. orygis (die gemsbok-basil); M. caprae (die bok-basil), en M. bovis (die bees-basil) (Alexander et al., 2010; Gey van Pittius et al., 2012a; Gey van Pittius et al., 2012b; Van Ingen et al, 2012; Coscolla et al, 2013; Parsons et al., 2013). Al hierdie spesies kan tuberkelsiekte by enige soogdier, waaronder die mens, veroorsaak, maar is meestal beperk tot spesifieke ekotipes wat tot siekte by ’n bepaalde gasheer lei: Die dassie-basil kom byvoorbeeld hoofsaaklik by die klipdassie (Procavia capensis) voor, M. pinnipedii by robbe, ensovoorts (Parsons et al., 2008).
MIKOBAKTeRIËLe GeNOMIKA
n Groot deurbraak in tuberkulosenavorsing en die begrip van die mikobakterieë was
die volledige volgordebepaling (in 1998) van die eerste heelgenoomvolgorde van die M. tuberculosis-laboratoriumverwysingstam H37Rv, wat algemeen gebruik word (Cole et al., 1998), sowel as van duisende ander stamme en spesies binne die genus Mycobacterium
in die jare daarna, waaronder meer as 600 M. tuberculosis-stamme in ons eie laboratorium aan die Universiteit Stellenbosch. Die volgordedata van M. tuberculosis het getoon dat die genoom sowat 4.4 Mb groot is, oor ’n hoë G+C-inhoud van 67% beskik en uit sowat 4 000 gene bestaan wat eweredig oor albei stringe versprei is. Die funksie van ongeveer 40% van die gene was bekend, dié van 20% was hipoteties, terwyl die oorblywende 40% geheel en al onbekend was. Die volgordebepaling het dus ’n groot aantal voorheen onbekende gene aan die lig gebring, wat alles moontlik by die patogenese van die organisme betrokke kon wees (Cole et al., 1998). Interessant genoeg het ongeveer 52% van die genoom as gevolg van geenduplikasie of domeinskommelinge ontstaan (Tekaia et al., 1999). Nóg ’n verrassende waarneming was dat twee onbekende, nuwe geenfamilies (wat in genoomareas met ’n uitsonderlik hoë G+C-inhoud van meer as 80% voorkom) sowat 10% van die genoom uitmaak (Cole, 1999). Hierdie twee families is gevolglik die PE- (99 lede) en die PPE-geenfamilie (67 lede) gedoop weens die prolien- (P-) en glutamiensuur- (E-)motiewe wat gekonserveerd voorkom in die N-termini van die gekodeerde proteïene. Die volgordebepaling van die genome van duisende mikobakteriële stamme en spesies oor die afgelope 18 jaar het ’n buitengewone geleentheid gebied om belangrike gene deur die nuwe wetenskap van vergelykende genomika te ontdek (Cole, 1998), waardeur die genome en geenvolgordes van mikobakteriële spesies en stamme vergelyk en die verskille uitvoerig ontleed word.
Mikobakteriële genomika het die weg gebaan tot ’n begrip van hoe mikobakteriële spesies en stamme met mekaar verband hou en watter veranderinge gedurende hulle evolusie plaasgevind het wat aanleiding gegee het tot die wisselende fenotipes wat ons deesdae teëkom. Die byvoeging van daaropvolgende tegnologieë, soos vergelykende proteomika, vergelykende transkriptomika en vergelykende metabolomika, het ’n reuse-bydrae gelewer tot die magdom inligting tot ons beskikking, en het tot ’n nuwe vakgebied genaamd vergelykende mikobakteriële ‘omika’ – kortweg ‘mikobaktomika’ – gelei. Die resultate van hierdie ontledings het ’n ongeëwenaarde blik op die genomiese en proteomiese variasie van die mikobakterieë gebied, en insig in die evolusie van fiksheid, virulensie en middelweerstandigheid verleen. Mikobaktomika het ook nuwe instrumente opgelewer om belangrike chromosomale streke, gene, proteïene sowel as metaboliese verbindings te identifiseer wat gebruik kan word in die ontwikkeling en toepassing van metodes om patogeniese en niepatogeniese stamme en spesies op te spoor, met ander woorde vir die ontwikkeling van nuwe mikobakteriële diagnostika.
’

10
eVOLUSIe VAN VIRULeNSIe IN DIe MIKOBAKTeRIeË
Die meeste mikobakterieë is omgewingsorganismes wat in grond en water voorkom en net sóms siekte
tot gevolg het wanneer hulle ’n vatbare, gewoonlik immuunkompromitteerde, mense- of dieregasheer teëkom. ’n Paar stadig groeiende spesies, soos die lede van die M. tuberculosis-kompleks, het egter die vermoë ontwikkel om in die loop van evolusie in gasheermakrofages te oorleef, en het beduidende patogene van mense én diere geword (Jang et al., 2008). Soos reeds vroeër genoem is, het die beskikbaarheid van heelgenoomvolgordedata vir ’n groot aantal mikobakteriële spesies die weg gebaan tot vergelykende genoomnavorsing met die doel om genetiese veranderlikheid binne en tussen mikobakteriële spesies te bepaal. Een so ’n studie het 48 M. tuberculosis- chromosomale streke met atipiese kenmerke geïdentifiseer, wat op verwerwing deur horisontale geenoordrag dui (Becq et al., 2007). Hierdie bepaalde streke verteenwoordig 4.5% van die genoom (199 kb) en sluit ’n groot aantal gene in wat ortoloog is met dié van ander bakteriële spesies wat in grond voorkom, bepaald lede van die proteobakterieë en aktinobakterieë (Becq et al., 2007). Na aanleiding hiervan het Veyrier en kollegas (2009) later 137 gene in M. tuberculosis geïdentifiseer wat waarskynlik verkry is deur ’n stapsgewyse proses van horisontale geenoordrag gedurende die skeiding tussen die stadig groeiende en snelgroeiende mikobakterieë. Altesaam 60% van hierdie gene het uit ander aktinobakterieë gespruit wat in dieselfde omgewing as die saprofitiese mikobakterieë voorkom. Onder andere was daar ’n spesifieke verkryging van gene wat kodeer vir proteïene gemoeid met die metaboliese funksies wat met aanpassing by anaërobiese omstandighede verband hou (Veyrier et al., 2009), wat die organisme sodoende op ’n intrasellulêre leefstyl voorberei.
Hierdie vroeë episodes van horisontale oordrag van genomiese eilande vanaf omliggende omgewingspesies kon bygedra het tot die ontwikkeling van die virulensie van M. tuberculosis deur spesifiek die basille eerstens te help oorleef in protosoë wat in hulle natuurlike omgewing aangetref word, en gevolglik ook in soogdierfagosiete. Etlike studies het al getoon dat patogeniese mikobakteriële spesies die vermoë ontwikkel het om in protosoë te oorleef. M. fortuitum, M. xenopi, M. avium, M. marinum en M. bovis ontduik almal uitwissing en vermenigvuldig in Acanthamoeba sp. (Taylor et al, 2003), terwyl sommige studies selfs getoon het dat die virulensie van dié spesies toeneem met opeenvolgende verspreidings in hierdie amebe (Cirillo et al. 1997). Die hipotese is dat hierdie
intraprotosoïese leefstyl, en die versterkte vermoë van die bakterie om intrasellulêr te leef en uitwissing deur die gasheersel te ontduik, die progenitor van die M. tuberculosis-kompleks vir ’n intramakrofagiese leefstyl toegerus het.
Nadat die voorouers van die M. tuberculosis-kompleks die vermoë ontwikkel het om intrasellulêr te oorleef, moes hulle met strategieë vorendag kom om die immuunreaksie van die gasheer te weerstaan, doeltreffend tussen gashere te versprei, en te keer dat hulle in klein menslike populasies uitsterf deur hulle eie virulensie te temper (Brites en Gagneux, 2015). In die een of ander stadium van hulle evolusie het die lede van die mens- en dier-aangepaste linies van die M. tuberculosis-kompleks weggebreek van die gladde tuberkelbasille, en klaarblyklik die vermoë verloor om horisontale geenoordrag te ondergaan. Dít was gedurende ’n evolusiebottelnek of selektiewe opruiming sowat 35 000 tot 40 000 jaar gelede, wat waarskynlik verband gehou het met die verandering in leefstylstatus van ’n opportunistiese patogeen tot ’n obligatoriese patogeen (Gutierrez et al., 2005). Dit blyk of hierdie wegbreek van die gladde tuberkelbasille (waaronder M. canetti) onmiddellik voor dié bottelnek of selektiewe opruiming gebeur het, terwyl alle ander lede van die kompleks die gevolg van die klonale uitbreiding van slegs ’n klein getal oorlewende bakterieë is (McEvoy et al., 2012). Vergelykende genomika-ontledings toon dat M. tuberculosis ’n tweefasige evolusionêre proses ondergaan het, wat genoomuitbreiding (deur geenverwerwing en -duplikasie) sowel as reduksie-evolusie (deur geenskrapping) behels het (Veyrier et al., 2011). Veral geenduplikasie het oënskynlik ’n beduidende rol in die wegbreek en aanpassing van die spesie, sowel as in die daaropvolgende toename in die virulensie van M. tuberculosis, gespeel – studies toon dat sowat 52% van die organisme se proteome uit geenduplikasies spruit (Tekaia et al., 1999). Hierdie duplikasiegebeure sluit in die vyf kopieë van die immunopatologies beduidende ESAT-6- (ESX-)geengroepe (Gey van Pittius et al., 2001) sowel as die groot multiproteïenfamilies PE en PPE (Gey van Pittius et al., 2006), wat albei na bewering ’n kernrol in die oorlewing en virulensie van die patogeniese mikobakterieë gespeel het.
Die ESAT-6- (of ESX-)geengroepstreke druk proteïene uit wat ’n toegewyde, bindende, proteïenafhanklike aktiewe transportstelsel met etlike komponente uitmaak en wat ’n nuwe tipe VII-sekresiestelsel kodeer wat verantwoordelik is vir die sec-onafhanklike sekresie van onder andere die familielede van die T-sel-antigene ESAT-6 en CFP-10 (Gey van Pittius et al., 2001, Abdallah et al., 2007, Bitter et al., 2009). Die vyf verskillende ESX-stelsels

11
in die genus Mycobacterium het deur geenduplikasie ontwikkel in die volgorde ESX-4, ESX-1, ESX-3, ESX-2 en, mees onlangs, ESX-5 (Gey van Pittius et al., 2001). Hoewel ál vyf gemoeid is met die transport van klein proteïene oor die selmembraan, het dié immunologies beduidende sekresiestelsels so ontwikkel dat hulle verskillende funksies vervul (’n bekende uitwerking van geenduplikasie en die oortolligheid wat gevolglik ontstaan) en mekaar oënskynlik nié by uitskakeling aanvul nie. ESX-4 blyk die mees voorouerlike streek te wees, synde die kleinste ESX-lokus (met slegs die minimale stel gene wat vir transport vereis word), waarvan ortoloë gebiede ook in bakterieë buite die genus Mycobacterium aangetref word en wat klaarblyklik geen rol in virulensie speel nie (Gey van Pittius et al., 2001; Sassetti et al., 2003). ESX-1 was die eerste ESX-stelsel wat ontdek is en blyk direk by patogenisiteit en die fagosomale ontsnapping van die organisme betrokke te wees, hoewel dit nie vir in vitro-groei vereis word nie (Simeone et al., 2012). Daarteenoor is ESX-3 blykbaar betrokke by yster- en sink-homeostase, wat dit wél noodsaaklik maak vir die in vitro-groei van hierdie organisme (Serafini et al., 2013). Die virulensie van ESX-1-mutante is beduidend laer in makrofagiese sellyne en in vivo-infeksiemodelle omdat hulle nie aan die fagolisosoom van makrofages kan ontsnap nie (Houben et al., 2012, Simeone et al., 2012). Die skrapping van ’n groot gedeelte van die ESX-1- genetiese lokus is ook die hoofrede vir die verlaagde virulensie van die vaksienstam M. bovis BCG (Pym et al., 2002) en ander nievirulente mikobakteriële spesies. ’n Interessante waarneming is dat die laaste duplikasie (van ESX-5) saamgeval het met die verskyning van die stadig groeiende mikobakteriële spesies en nie teenwoordig is in die genome van enige snelgroeiende spesie wat tot dusver bestudeer is nie (Gey van Pittius et al., 2006, Abdallah et al., 2009). Hierdie stelsel is verantwoordelik vir die sekresie van etlike lede van die groot PE- en PPE-proteïenfamilies in M. marinum (Abdallah et al., 2009), meer bepaald die onlangs ontwikkelde glisienryke en herhalende PE_PGRS-proteïene, wat slegs in die stadige groeiers aangetref word en na bewering by virulensie en immuunontduiking betrokke is (Gey van Pittius et al., 2006, Abdallah et al., 2009). Aangesien die meeste patogeniese mikobakterieë stadig groei, blyk die aanwesigheid van ESX-5 en die uitbreiding van die PE- en PPE-proteïenfamilies in slegs die stadige groeiers te dui op ’n verband met die aanpassing by ’n intrasellulêre en virulente leefstyl. ’n Moontlike rol van ESX-5 is dat dit tot ’n voedingsverskaffingstelsel ontwikkel het in die plek van die porienstelsels in die snelgroeiendes (wat as nadelig bewys is vir die intrasellulêre oorlewing van snelgroeiende mikobakterieë, en afwesig is by die
stadige groeiers). Hierdie verskaffingstelsel verseker voldoende voeding en handhaaf terselfdertyd die ondeurdringbaarheid van die buitenste membraan wat vir in vivo-volharding vereis word (Ates et al., 2015). Duplikasie van die oorspronklike geengroep wat vir die ESAT-6-sekresiestelsel kodeer, en die verwante proteïene van die PE- (bepaald die PE_PGRS-), PPE- en ESAT-6-agtige proteïenfamilies, het dus funksionele diversifikasie tot ’n aantal nuwe patogeniese meganismes moontlik gemaak.
OORSPRONKLIKe SeLeKTIeWe KRAGTe – GASHeeRAANPASSING eN KO-eVOLUSIe
Die verweefde genealogieë van mikobakterieë en die mens het ’n aanvang geneem die oomblik toe die
eerste progenitor van die M. tuberculosis-kompleks die vermoë ontwikkel het om ’n lid van Homo sapiens stabiel te infekteer. Sedertdien het ’n wisselwerkingsverhouding tussen gasheer en bakterie ontwikkel, wat tot vandag toe voortduur. Die oorskakeling van ’n opportunistiese patogeniese leefstyl na dié van ’n obligatoriese patogeen het ongelooflike druk op die bakterie geplaas om te verseker dat dit in die gasheer sou oorleef en (deur die meganisme van aktiewe siekte) na nuwe gashere sou kon versprei, maar nie op so ’n manier dat dit alle beskikbare gashere sou uitwis en dus sy eie oorlewingsvermoë sou vernietig nie. Juis die oorlewing van die bakterie het dus op verwikkelde wyse met die bestaan van die gasheer vervleg geraak (en is deur die gasheer se immuunstelsel beïnvloed), en enige veranderinge in die interne en eksterne omgewing van die gasheer het die moontlike bedreiging ingehou dat dit ook die oorlewing van die bakterie ingrypend kon raak. ’n Herhalende proses van gasheeraanpassing en ko-evolusie het derhalwe oor millennia heen plaasgevind (en vind steeds plaas), waardeur evolusionêre veranderinge in die patogeen wat infektiwiteit en oorlewing verhoog, teëgewerk word deur evolusionêre veranderinge in die gasheer wat weerstand teen, en opklaring van, infeksie versterk (Brites en Gagneux, 2015). Hierdie proses van wederkerige, antagonistiese uitwerkings op die fiksheid van sowel die mikobakterie as die mens het aanleiding gegee tot een van die kerneienskappe van tuberkulose soos ons dit ken, naamlik latensie – ’n toestand waarin die bakterie die gasheer latent en aanhoudend kan infekteer sonder om siekte te veroorsaak of te versprei, met die gasheer se immuunstelsel wat dit in toom hou, totdat gunstige omstandighede vir aktiewe siekte en verspreiding na ’n nuwe gasheer ontstaan (soos bejaardheid of immuunonderdrukking) (Brites

12
Figuur 5 – Die stamboom van die lede van die Mycobacterium tuberculosis-kompleks (aangepas uit Alexander et al., 2010)
en Gagneux, 2012). Mycobacterium tuberculosis het so eiesoortig en besonder doeltreffend by hierdie leefstyl aangepas dat dit na raming sowat ’n derde van die wêreld se menslike bevolking latent geïnfekteer het (WGO, 2015). In die millennia nadat die voorouer van M. tuberculosis die eerste mense geïnfekteer het, was die vernaamste twee oorspronklike selektiewe kragte wat meestal op die aanpassing van die organisme ingewerk het, die immuunreaksie van die gasheer en veranderinge in menslike demografie (Brites en Gagneux, 2012).
EX AFRICA SEMPER ALIQUID NOVI – ALTYD IeTS NUUTS UIT AFRIKA
Vergelykende genomika-ontledings het aan die lig gebring dat hoewel die lede van die M. tuberculosis-
kompleks opmerklike verskille toon in fenotipe, patogenisiteit en primêre gashere (wat strek van die sjimpansee tot die mens, vleirot, bok, gemsbok, dassie, bees, meerkat, muishond en rob), hulle geneties nou verwant is en dat hulle DNS-volgordes oor 99.9% gemeenskaplike identiteit beskik (Brosch et al., 2002). Lede van die M. tuberculosis-kompleks het die vermoë verloor om horisontale geenoordrag te ondergaan, en dus is die enigste genetiese veranderinge wat nog in hierdie organismes kan plaasvind skrappings, duplikasies, genetiese herskikkings, die invoeging van transposons, en enkelnukleotied-polimorfismes (Karboul et al., 2006; McEvoy et al., 2007; Karboul et al., 2008; McEvoy et al.,
2009a; McEvoy et al., 2009b; Weiner et al., 2012). M. tuberculosis het van al hierdie meganismes gebruik gemaak (en gebruik dit steeds) om te verseker dat hy enige van die gasheer se aanvalle op sy oorlewing met ’n teenaanval kan beantwoord (sien byvoorbeeld hieronder vir ’n beskrywing van die evolusie van middelweerstandigheid deur enkelnukleotied-polimorfismes). Navorsers glo dat die evolusie van hierdie verskillende spesies en stamme van die M. tuberculosis-kompleks uit ’n gemeenskaplike progenitor spruit wat meer as 70 000 jaar gelede bestaan het, toe die eerste mikobakterie na bewering ’n lid van Homo sapiens in Oos-Afrika geïnfekteer het (Comas et al., 2013). Tot ongeveer ’n dekade gelede is daar aanvaar dat M. tuberculosis uit M. bovis ontwikkel het en eers ná die temming van beeste (wat tans as instandhoudingsgasheer dien) sowat 10 000 jaar gelede na die mens begin versprei het (Stead, 1997). Tog het genomika-ontledings hierdie teorie verkeerd bewys, aangesien mens-aangepaste linies van M. tuberculosis filogeneties meer voorouerlik is as dié van die dier-aangepaste spesies (sien Figuur 5) en M. bovis verskeie gene verloor het wat steeds in M. tuberculosis en ander dier-aangepaste spesies aanwesig is (Brosch et al, 2002). Trouens, etlike studies het getoon dat dieretuberkulose reeds aan die einde van die Pleistoseentydperk (tussen 20 000 en 11 000 jaar gelede) voorgekom het, nog lank voordat beeste getem is (Rothschild en Martin, 2006). Dít word gestaaf deur die aanwesigheid van kenmerkende tuberkulêre letsels by fossiele van die sogenaamde
RD7
M. canettii and otherextant smooth tubercle bacilli
“M. prototuberculosis”TbD1
gyrA95ACCAGC
katG463CTCCGG
“ancestral” M. tuberculosis (gr. 1, TbD1 intact)
gr. 1, e.g. Beijing
gr. 3, e.g. H37Rv
“modern” M. tuberculosis(TbD1 deleted)
M. africanum subtype 1(b) (West-African 1 lineage)
RD9
RD8
RD10
gr. 2, e.g. LAM
sublineage 1
sublineage 2sublineage 3
M. africanum subtype 1(a)(West-African 2 lineage)
Rv15101129
Chimpanzee bacillus
M. mungi (mongoose bacillus)
Dassie bacillus
M. suricattae (meerkat bacillus)
M. orygis (oryx bacillus)
M. microti (vole bacillus)
M. pinnipedii (seal bacillus)
M. caprae (goat bacillus)
M. bovis (‘classical’ bovine bacillus)
M. bovis BCG (vaccine strains)
RD701/2rpoB1163
rpoB1049
N-RD25das
RD1mon
RD1das
RD713
RD5das
RD5sur
mmpL6551AACAAG
gyrBoryx
RD1micoxyR n285 GA
RD2sealRD12bov
RD13
RD4
RD1BCG
Chimpanzee
Mongoose
Dassie
Meerkat
Oryx
Vole
Goat
Seal
Cattle
Humans
animal-adapted species

Figuur 6 – ‘Uit Afrika’- en Neolitiese verspreiding en uitbreiding van die mens-aangepaste spesies van die Mycobacterium tuberculosis-kompleks (M. tuberculosis en M. africanum). Belangrike wegbrekings word aangedui deur die middelwaarde (in duisende jare) van die betrokke nodusdatering (uit Comas et al., 2013).
13
groothoringskaap (Ovis canadensis catclawensis), die uitgestorwe muskusbees (Bootherium bombifrons) en die mastodon (Mammut americanum) in Noord-Amerika (Martin en Rothchild, 1989; Rothschild en Laub, 2006), sowel as letsels en die aanwesigheid van DNS uit die M. tuberculosis-oerkompleks in die metakarpaal van ’n uitgestorwe Noord-Amerikaanse langhoring-bison, waarvan die ouderdom met radiokoolstofdatering op meer as 17 800 jaar geskat is (Rothschild et al., 2001). Mastodons het vanaf ongeveer twee miljoen jaar gelede volop in Noord-Amerika voorgekom totdat hulle sowat 11 000 jaar gelede (om en by die tyd toe die mens op die vasteland aangekom het) saam met baie ander groot Noord-Amerikaanse soogdiere uitgesterf het – die grondslag vir die teorie dat menslike kontak met hierdie groot soogdiere voorouerlike tuberkulose in omloop gebring het, wat moontlik tot die uitsterwing van die diere bygedra het (Rothschild en Laub, 2006).
Ondanks die hoë klonaliteit van die spesies wat tot die M. tuberculosis-kompleks behoort, het verskillende linies, sublinies en stamme (deur bogenoemde meganismes) oor die afgelope 70 000 jaar ontwikkel, onder meer ook die progenitor van die dier-aangepaste spesies, wat ’n streek van die genoom genaamd RD9 verloor het, welke genoomstreek derhalwe afwesig is by alle nie-uitgestorwe dier-aangepaste spesies, dog
steeds in die mens-aangepaste lede van die kompleks voorkom (Brosch et al., 2002). Hoewel daar bewys is dat M. tuberculosis en M. africanum ander soogdierspesies kan infekteer (sien byvoorbeeld Parsons et al., 2012, en ander), kan dit slegs mense stabiel infekteer en in mense versprei. Daarom kan dit wees dat hierdie skrapping van RD9 die weg gebaan het tot die differensiasie en verspreiding van die organisme na ander soogdiergasheerspesies, waarvan die heel eerstes van die Afrikavasteland gekom het (sjimpansee, dassie, meerkat en muishond). Dít staaf die siening dat die kompleks in Afrika ontstaan en dat diversifikasie op hierdie vasteland plaasgevind het voordat dit na die res van die wêreld versprei het. Daar bestaan bewyse dat dieretuberkulose in Eurasië voorgekom het lank voordat dit na Noord-Amerika versprei het, en dat lede van die Bovidae dit na Noord-Amerika gebring het, wat die teorie oor die verspreiding vanaf Afrika deur Eurasië na Amerika ondersteun (Rothschild en Martin, 2006). Al die verskillende dier-aangepaste spesies het ná RD9 hulle eie spesifieke genetiese veranderinge (merendeels skrappings) ondergaan om by hulle nuutgevonde gasheernisse aan te pas, dermate dat hierdie spesifieke skrappings vandag as ’n instrument dien om geneties tussen al die verskillende lede van die M. tuberculosis-kompleks te onderskei (Warren et al., 2006).

14
Volgens vergelykende genomika- en filogeografiese ontledings het M. tuberculosis as ’n menslike patogeen in Afrika ontwikkel en daarná saam met die moderne mens na die res van die wêreld versprei namate dié oor die afgelope 70 000 tot 40 000 jaar uit Afrika gemigreer het (Gutierrez et al., 2005; Hershberg et al., 2008) (sien Figuur 6). Namate menslike populasies gediversifiseer het, het hierdie M. tuberculosis-oerstamme ook op grond van die veranderende menslike demografie en genotipes vroeë diversifikasie ondergaan ten einde die optimale gasheer-patogeen-verhouding te handhaaf (Brites en Gagneux, 2015). Weens gasheerspesifieke aanpassing by geïsoleerde menslike populasies, het verskeie M. tuberculosis-hooflinies met kenmerkende eienskappe oor duisende jare in isolasie ontwikkel. Die spesie M. tuberculosis wat ons deesdae teëkom, bestaan uit ’n groot getal verskillende stamme (daar is tans meer as 1 000 stamme waarvan ons weet) wat tot meer as 40 verskillende groter stamfamilies behoort (waarvan heelwat oor verskillende patogeniese kenmerke, fiksheid en gasheeraanpassings beskik) en onder sewe groot bekende linies val (Gagneux et al., 2006; Coscolla en Gagneux, 2014). Omdat hulle gedifferensieer en by verskillende menslike populasies in verskillende wêrelddele aangepas het, het hierdie sewe linies ’n sterk filogeografiese struktuur. Hulle word in oerlinies en moderne linies verdeel op grond van die skrapping van ’n geengebied genaamd TbD1 (Brosch et al., 2002). Die oerlinies is die nouste verwant aan die progenitor en die dier-aangepaste spesies, en bestaan uit linie 5 (M. africanum Wes-Afrika 1) en linie 6 (M. africanum Wes-Afrika 2), wat albei geografies tot die lande in Wes-Afrika beperk is, sowel as die onlangs beskrewe linie 7 (die Ethiopiese linie wat tot Ethiopië beperk is) en linie 1 (wat ook die Indo-Oseaniese linie genoem word en in Oos-Afrika, op die rand van die Indiese Oseaan, en in die Filippyne voorkom) (Coscolla en Gagneux, 2014). Die teenwoordigheid van die oerlinies in Afrika ondersteun die teorie wat Afrika as die oorsprong- en verspreidingskern van M. tuberculosis voorhou. Die moderne linies het ná die skrapping van TbD1 gevorm, en bestaan uit linie 2 (die Oos-Asiatiese linie wat wydverspreid in Oos-Asiatiese lande voorkom en die baie groot en suksesvolle Beijing-stamfamilie insluit), linie 3 (die Oos-Afrika- en Sentraal-Asiatiese linie) en linie 4 (die Euro-Amerikaanse linie wat wydverspreid in Europa en Amerika voorkom en die baie groot en suksesvolle LAM-stamfamilie insluit). Hoewel die sewe linies geskiedkundig met verskillende menslike gasheerpopulasies van verskillende wêreldstreke verbind word (die CAS-stamme word byvoorbeeld meestal met menslike gashere in Indië, Pakistan en Oos-
Afrika verbind; die Beijing-stamme met gashere in Oos-Asië; die EAI-stamme met gashere in die Indo-Oseaniese streke, en die LAM-, Haarlem- en LCC-stamme met gashere in Europa), kon hierdie stamme die afgelope 400 jaar deur middel van kolonisasie, die ingebruikneming van skeepshandelsroetes tussen die westelike en oostelike halfronde sowel as die ontdekking en verowering van die Noord- en Suid-Amerikas en Australië tussen die 16de en 19de eeu na ander wêrelddele versprei (Gagneux et al., 2006). Met die opheffing van historiese geografiese versperrings tussen die menslike populasies van die wêreld, was die identifikasie van, interaksie met en infeksie van nuwe gashere, wat oor ander genetiese en immunologiese agtergronde beskik het, nou moontlik vir die stamme van M. tuberculosis, wat voorheen geografies ingeperk was. Dít sou ’n splinternuwe hoofstuk in die verweefde geskiedenisse van mikobakterieë en die mens inlui.
UIT AFRIKA … eN WeeR TeRUG …
Suider-Afrika was honderde jare lank geografies sentraal geleë op die historiese handelsroetes tussen
die Ooste en die Weste, en tienduisende reisigers van sowel Oos-Asië as Europa het deur hierdie streek gereis en hulle hier gevestig (die Europeërs merendeels uit vrye wil, en die Oos-Indiërs en Asiate meestal teensinnig as slawe). Dit is ook bekend dat tuberkuloselyers van Europa (veral Brittanje) en Indië na Suid-Afrika gekom het weens die sonnige, matige klimaat, wat as voordelig beskou is en glo genesing sou aanhelp. Advertensies in die Illustrated London News het selfs verkondig hoe voortreflik die Kaapse klimaat vir die genesing van tuberkulose is, wat groot getalle nuwe tuberkulosegevalle in die 1870’s en 1880’s na die streek laat stroom het, van wie die bekendste straks Cecil John Rhodes was (Packard, 1989). Daarom kom ons deesdae plaaslik soveel diverse M. tuberculosis-stamme van Europese en Oos-Asiatiese oorsprong as dominante stamsoorte teë. Soos reeds hierbo genoem is, het verskillende stamme van M. tuberculosis deur evolusie verskillende fenotipiese eienskappe ontwikkel, wat gelei het tot stamme wat byvoorbeeld hiper- of hipovirulent is by sekere populasies (Aguilar et al., 2010). In bepaalde omstandighede blyk sekere stamfamilies ook ander families uit te oorlê (Mardassi et al., 2005, Hanekom et al., 2007). So byvoorbeeld het die voorkoms van gevalle met stamme van die Beijing-genotipe (wat oorspronklik van Oos-Asië kom) mettertyd eksponensieel toegeneem in ons gasheerpopulasie in die Wes-Kaap, Suid-Afrika, teen ’n geraamde verdubbelingstempo van slegs 3.8 jaar

15
(Johnson et al., 2010, Hanekom et al., 2011). Volgens studies groei die Beijing-stamfamilie vinniger as ander stamme, versprei makliker, en is meer geneig om op siekte uit te loop (López et al., 2003). Aan die ander kant is die F11 LAM-genotipe (wat oorspronklik uit Europa kom) ook tans een van die grootste stamfamilies in die Wes-Kaap, en word dit met ongeveer 15% van tuberkulosegevalle wêreldwyd verbind (Gibson et al., 2008). Die twee families Beijing en F11, wat uit gans verskillende wêrelddele kom, verteenwoordig gesamentlik nagenoeg 45% van die tuberkulosegevalle in die Wes-Kaap (Victor et al., 2004). In KwaZulu-Natal, ’n ander Suid-Afrikaanse studieomgewing, vind ons die KZN-stamfamilie (’n lid van die groter LAM-genotipe en familie van die F11 LAM-stamfamilie wat volop in die Wes-Kaap voorkom), ’n hoogs virulente stam wat endemies en oorheersend is in die KwaZulu-Natalse streek, waar daar onlangs ’n uitbreking van uiters middelweerstandige tuberkulose was (Pillay en Sturm, 2007). In Zimbabwe en Zambië blyk die F9 LAM11-stam te oorheers, en is ongeveer die helfte van die pasiënte in hierdie streek daarmee geïnfekteer (Gibson et al., 2008). Buite Afrika kom die RDRio M. tuberculosis-familie, ’n LAM-sublinie, wêreldwyd voor, maar is dit veral die hoofoorsaak van TB in Suid-Amerika, waar dit 38% van alle stamme en 70.4% van alle LAM-stamme uitmaak (Gibson et al., 2008). Soos die Beijing-familie, beskik stamme in die LAM-familie oor ’n fiksheidsvoordeel wanneer hulle groeitempo met dié van nie-LAM-stamme vergelyk word. Die aanvang van globalisasie het die verhouding tussen mikobakterieë en die mens beduidend verander. Wat na ’n betreklik stabiele verhouding gelyk het en oor millennia deur ko-evolusie en wederkerige antagonistiese druk gevorm is, is versteur toe die tradisionele geografiese grense opgehef is. Gevolglik het die mikobakterie nuwe genotipes van die mens teëgekom en het die mens nuwe genotipes van die mikobakterie teëgekom, en so het ’n heel nuwe, wêreldwye konflik tussen die twee oervyande uitgebreek. Hierdie nuwe toedrag van sake word verder bemoeilik deur twee onlangse gebeure in die lang en verweefde geskiedenis van mikobakterieë en die mens, naamlik die bekendstelling van antimikrobiese middels en die aanvang van die MIV/vigs-epidemie. Hierdie nuwe selektiewe kragte het ’n beduidende impak op die verhouding en sal die epidemie nog jare lank beïnvloed.
MODeRNe SeLeKTIeWe KRAGTe – MIV eN ANTIMIKROBIeSe MIDDeLS
Die ontdekking van antimikrobiese middels het ’n revolusie in 20ste-eeuse geneeskunde veroorsaak.
Nog nooit in die geskiedenis van die mensdom was daar soveel verskillende vorme van beskerming teen mikrobiese siektes soos nou nie. In samehang met inenting, kon antimikrobiese behandeling etlike vreesaanjaende siektes bykans uitwis. Tog het oormatige gebruik, ondoeltreffende voorskrifpraktyke en swak behandelingsgetrouheid tot ’n nuwe vlaag van bakteriële weerstandigheid gelei. Weens die toename in bakteriële weerstandigheid teen antibiotika die afgelope paar dekades het die Wêreldgesondheidsorganisasie antimikrobiese weerstandigheid geklassifiseer as ’n “ernstige bedreiging wat nie meer ’n toekomsvoorspelling is nie, maar reeds in elke wêreldstreek gebeur en enigeen van enige ouderdom en in enige land kan raak” (WGO, 2014c).
Selman A Waksman (1964), wat die antibiotikum streptomisien ontdek het, skryf soos volg in sy boek The Conquest of Tuberculosis:
But most important, the ancient foe of man, known as consumption, the great white plague, tuberculosis, or by whatever other name, is on the way to being reduced to a minor ailment of man. The future appears bright indeed, and the complete eradication of the disease is in sight…
Waksman kon dit nie verder mis gehad het nie. Die mensdom het vergeet: Vir elke aksie is daar ’n reaksie – ook vir daardie aksies wat die mens se verweefde verhouding met mikobakterieë raak. Aanvanklik was die gebruik van antibiotika teen M. tuberculosis skynbaar baie suksesvol en tuberkulosesyfers het dermate begin daal dat alle ontwikkeling van nuwe antimikobakteriële middels vir ’n aantal dekades gestaak is nadat die eerste suksesvolle aktiewe verbindings teen M. tuberculosis tussen die 1940’s en 1960’s ontdek en klinies in gebruik geneem is. Maar die onoordeelkundige gebruik van hierdie middels het tot middelweerstandigheid, MDR (multimiddelweerstandigheid), XDR (uiterste middelweerstandigheid) en selfs TDR (algehele middelweerstandigheid) gelei (Müller et al., 2013) – afkortings wat deesdae sonder uitsondering vrees inboesem by deelnemers aan tuberkulosegesprekke wêreldwyd. Die mens het ’n nuwe aanval op die mikobakterie geloods, synde antibiotika, en sodoende die eeue oue gasheer-patogeen-verhouding in sy guns probeer swaai. Oorwinnings is gevier, maar die stryd was nog lank nie verby nie. Die mikobakterieë het op hulle eie manier gereageer om hulleself te beskerm deur meganismes soos weerstandigheidseleksie met behulp van enkelnukleotied-polimorfismes in middelteikengene, sowel as die vermindering van vatbaarheid deur

middel-effluks (Louw et al., 2009; Louw et al., 2011; Müller et al., 2011; Sirgel et al., 2012). Die moderne selektiewe druk van antimikrobiese behandeling was selfs voordelig vir sekere mikobakteriële stamme, wat nou beter kon reageer op die nuwe omgewing wat die skielike teenwoordigheid van antibiotika geskep het (Van Rie et al., 2005). Dus sien ons veel hoër vlakke van middelweerstandigheid in die Beijing-familie in die Wes-Kaap vergeleke met ander stamfamilies soos die LAM-familie. Dít is ’n belangrike waarneming, aangesien die LAM-stamme tans byna ewe veel as die Beijing-stamme in die Kaapse populasie voorkom, en dui daarop dat die Beijing-familie oor stamspesifieke meganismes beskik waardeur middelweerstandigheid ontwikkel en aangepas word. Volgens ons epidemiologiese data is daar ’n eksponensiële toename in Beijing-stamme in hierdie omgewing vergeleke met ander stamme, wat oor die afgelope 13 jaar betreklik konstant gebly of selfs verminder het (Van der Spuy et al., 2009). Ons merk dus veranderinge in die M. tuberculosis-populasiepatrone, wat op klade-spesifieke patogeniese en aanpassingskenmerke dui. In die Oos-Kaap, ’n ander Suid-Afrikaanse studieomgewing, is middelweerstandige atipiese Beijing-stamme uiters belangrik, en dié streek gaan tans gebuk onder ’n groot uitbreking van hierdie stamme (Klopper et al., 2013). Middelweerstandigheid en multimiddelweerstandigheid ontwikkel en ver-sprei op groot skaal (Ioerger et al., 2010) en neem wêreldwyd toe. Die verskyning van uiters en algeheel middelweerstandige mikobakterieë, wat nie op eerste- óf tweedeliniebehandeling reageer nie, is geweldig kommerwekkend (Hoek et al., 2009; Chihota et al., 2012; Streicher et al., 2012). Ons moet dringend soek na nuwe maniere om tradisionele metodes van genotipering te vervang (Hanekom et al., 2008, Warren et al., 2009b), sowel as na nuwe instrumente om die tydsverloop tot en met diagnose te verkort en weerstandigheid vinniger op te spoor (Hoek et al., 2008, Warren et al., 2009a, Parsons et al., 2011, Barnard et al., 2012a, Barnard et al., 2012b). Middelweerstandigheid is ’n selektiewe druk in die mikobakterie se guns, en ’n ontoereikende reaksie van die gasheer sal ’n nadelige uitwerking hê op die mensdom se oorlewing in die stryd tussen mikobakterie en die mens.
Die ander onlangse gebeurtenis wat ’n beduidende impak op die verhouding tussen mikobakterie en die mens gehad het, was die aanvang van die MIV/vigs-epidemie (Brites en Gagneux, 2012). Hoewel MIV-siekte nog maar slegs die afgelope 30 jaar pandemies is, het dit ’n reuse-invloed op menslike gesondheid en op die verweefde verhouding tussen mikobakterieë en die mens. Sowat 35 miljoen mense wêreldwyd leef tans met MIV en ongeveer 2.3 miljoen nuwe gevalle word jaarliks
geïnfekteer (Lucas en Nelson, 2015). Sewentig persent van die wêreld se MIV-positiewe mense woon in Afrika suid van die Sahara. MIV kan sistemiese T-selaftakeling en verswakte selbemiddelde immuniteit veroorsaak, wat op sy beurt tot ’n wye verskeidenheid opportunistiese infeksies en kankers lei (Lucas en Nelson, 2015). Een van die vernaamstes hiervan is infeksie met M. tuberculosis, ’n organisme wat in noue wisselwerking met die gasheer se immuunstelsel funksioneer en dus gedy in omstandighede waar die immuunstelsel gekompromitteer is. Die moderne selektiewe krag van MIV-ko-infeksie verseker nou dat stamme van M. tuberculosis die gunstigste gasheeromstandighede ondervind wat hulle tot nog toe in hulle lang verhouding met die mensdom teëgekom het. Hierdie gunstige, immuunkompromitteerde omstandighede as gevolg van MIV stel M. tuberculosis in staat om nuwe nisse vir infeksie te vind, wat tot meer gereelde ekstrapulmonêre siekte lei en atipiese kliniese profiele tot gevolg het (Wasserman en Meintjes, 2014). TB-MIV-ko-infeksie is die vernaamste oorsaak vir die hoë infeksiesiekte- en -sterftesyfers onder Suid-Afrikaanse volwassenes (Wasserman en Meintjes, 2014). MIV-positiewe pasiënte loop tot 40 keer groter gevaar om TB op te doen as hulle MIV-negatiewe eweknieë (Getahun et al., 2010). TB is ook die grootste doodsoorsaak onder MIV-positiewe mense en lei tot 25% van alle MIV-verwante sterftes. Sonder behandeling sal sowat 90% van alle TB-geïnfekteerde MIV-positiewe mense sterf binne maande nadat hulle die siekte opgedoen het (WGO, 2015). Die risiko dat ’n MIV-negatiewe persoon met latente M. tuberculosis-infeksie TB-siekte sal opdoen, is 10% in sy leeftyd; ’n MIV-positiewe ko-geïnfekteerde persoon se risiko is 10% per jaar (McShane, 2005).
Die moderne selektiewe druk van die ingebruikneming van antimikrobiese middels en die aanvang van die MIV/vigs-epidemie het duidelik ’n beduidende rol gespeel in die onlangse gebeure wat die verweefde genealogieë van mikobakterieë en die mens betref. Hoewel die mens met behulp van antibiotika ’n aantal oorwinnings behaal het, het hy veel meer verloor deur sy onoordeelkundige gebruik daarvan. Die stryd het in die bakterie se guns gedraai – nie net deur die meganismes van multimiddelweerstandigheid nie, maar ook deur die verdere komplikasies van ’n nuwe siekte genaamd MIV/vigs, wat immuniteit kompromitteer en sodoende die gasheer se eerste verdedigingslinie teen die bakterie neutraliseer.
16

GeVOLGTReKKING
So it is said that if you know your enemies and know yourself, you can win a hundred battles without a single loss. If you only know yourself, but not your opponent, you may win or may lose. If you know neither yourself nor your enemy, you will always endanger yourself.
Sun Tzu, The Art of War, Hoofstuk 3 (6de eeu vC)
In aansluiting by Sun Tzu: Om die siekte te oorwin, moet ons nie net onsself verstaan nie, maar ook die organisme
wat die siekte veroorsaak. My werk die afgelope twee dekades het bepaald daarop gekonsentreer om te probeer begryp hoe mikobakterieë tot sulke suksesvolle menslike patogene ontwikkel het. Dít het my op ’n boeiende ontdekkingsreis deur tyd geneem en my ’n blik gebied op die buitengewone, verweefde genealogieë van mikobakterieë en die mens – ’n verhouding wat duisende jare gelede begin het op die Afrikavasteland, waar een van die grootste veldslae in hierdie eeue oue oorlog nou wéér afspeel. Net die tyd sal leer wie as oorwinnaar uit die stryd sal tree ... Met albei kante wat die ander probeer uitoorlê, is die stryd nog lank nie verby nie en is die verweefde geskiedenis van mikobakterieë en die mens nog nie klaar geskryf nie …
“We have made substantial progress toward TB elimination in this country, but TB remains a formidable opponent with thousands of cases still diagnosed each year. TB can be fatal and treatment remains long and difficult. This ancient bacterium has demonstrated its ability to evade our attacks many times before.”
Dr Jonathan Mermin, Direkteur: Nasionale Sentrum vir die Voorkoming van MIV/vigs, Virale Hepatitis, SOS en TB, Amerikaanse Sentrums vir Siektebeheer en -voorkoming (2014)
~~~~
17

BRONNeLYSAbdallah AM, Gey van Pittius NC, Champion PA, Cox J, Luirink J, Vandenbroucke-Grauls CM, Appelmelk BJ, Bitter W., 2007. Type VII secretion –
mycobacteria show the way. Nat. Rev. Microbiol. Nov;5(11):883–891.
Abdallah AM, Verboom T, Weerdenburg EM, Gey van Pittius NC, Mahasha PW, Jiménez C, Parra M, Cadieux N, Brennan MJ, Appelmelk BJ, Bitter W., 2009. PPE and PE_PGRS proteins of Mycobacterium marinum are transported via the type VII secretion system ESX-5. Mol. Microbiol. 73:329–340.
Aguilar D, Hanekom M, Mata D, Gey van Pittius NC, Van Helden PD, Warren RM, Hernandez-Pando R., 2010. Mycobacterium tuberculosis strains with the Beijing genotype demonstrate variability in virulence associated with transmission. Tuberculosis (Edinb.). Sep;90(5):319–325.
Alexander KA, Laver PN, Michel AL, Williams M, Van Helden PD, Warren RM, Gey van Pittius NC., 2010. Novel Mycobacterium tuberculosis complex pathogen, M. mungi. Emerg. Inf. Dis. 16:1296–1299.
Allison MJ, Mendoza D, Pezzia A., 1973. Documentation of a case of tuberculosis in pre-Columbian America. Am. Rev. Respir. Dis. 107:985–991.
Arriaza BT, Salo W, Aufderheide AC, Holcomb TA., 1995. Pre-Columbian tuberculosis in northern Chile: molecular and skeletal evidence. Am. J. Phys. Anthropol. 98:37–45.
Ates LS, Ummels R, Commandeur S, Van der Weerd R, Sparrius M, Weerdenburg E, Alber M, Kalscheuer R, Piersma SR, Abdallah AM, Abd El Ghany M, Abdel-Haleem AM, Pain A, Jiménez CR, Bitter W, Houben EN., 2015. Essential role of the ESX-5 secretion system in outer membrane permeability of pathogenic mycobacteria. PLoS Genet. Mei 4;11(5):e1005190.
Barnard M, Gey van Pittius NC, Van Helden PD, Bosman M, Coetzee G, Warren RM., 2012a. The diagnostic performance of the GenoType MTBDRplus version 2 line probe assay is equivalent to that of the Xpert MTB/RIF assay. J. Clin. Microbiol. Nov;50(11):3712–3716.
Barnard M, Warren R, Gey Van Pittius N, Van Helden P, Bosman M, Streicher E, Coetzee G, O’Brien R., 2012b. Genotype MTBDRsl line probe assay shortens time to diagnosis of extensively drug-resistant tuberculosis in a high-throughput diagnostic laboratory. Am. J. Respir. Crit. Care Med. Des 15;186(12):1298–1305.
Becq J, Gutierrez MC, Rosas-Magallanes V, Rauzier J, Gicquel B, Neyrolles O, Deschavanne P., 2007. Contribution of horizontally acquired genomic islands to the evolution of the tubercle bacilli. Mol. Biol. Evol. Aug;24(8):1861–1871.
Bitter W, Houben EN, Bottai D, Brodin P, Brown EJ, Cox JS, Derbyshire K, Fortune SM, Gao LY, Liu J, Gey van Pittius NC, Pym AS, Rubin EJ, Sherman DR, Cole ST, Brosch R., 2009. Systematic genetic nomenclature for type VII secretion systems. PLoS Pathog. Okt;5(10):e1000507.
Brites D, Gagneux S., 2012. Old and new selective pressures on Mycobacterium tuberculosis. Infect. Genet. Evol. Jun;12(4):678–685.
Brites D, Gagneux S., 2015. Co-evolution of Mycobacterium tuberculosis and Homo sapiens. Immunol. Rev. Mrt;264(1):6–24.
Brosch R, Gordon SV, Marmiesse M, Brodin P, Buchrieser C, Eiglmeier K, Garnier T, Gutierrez C, Hewinson G, Kremer K, Parsons LM, Pym AS, Samper S, Van Soolingen D, Cole ST., 2002. A new evolutionary scenario for the Mycobacterium tuberculosis complex. Proc. Natl. Acad. Sci. U.S.A. Mrt;19;99(6):3684–3689
Canci A, Minozzi S, Borgognini Tarli SM., 1996. New evidence of tuberculous spondylitis from Neolithic Liguria (Italy). Int. J. Osteoarchaeol. 6:498–501.
Canci A, Minozzi S, Borgognini Tarli SM., 2001. Tuberculous spondylitis during the Bronze Age: Two cases from Italy. In Proceedings to the XIIIth European Meeting of the Paleopathology Association, La Verghetta M en Capasso L (reds.). Edigrafital: Teramo. 67–71.
Chihota VN, Müller B, Mlambo CK, Pillay M, Tait M, Streicher EM, Marais E, Van der Spuy GD, Hanekom M, Coetzee G, Trollip A, Hayes C, Bosman ME, Gey van Pittius NC, Victor TC, Van Helden PD, Warren RM., 2012. Population structure of multi- and extensively drug-resistant Mycobacterium tuberculosis strains in South Africa. J. Clin. Microbiol. Mrt;50(3):995–1002.
Chiodini RJ, Van Kruiningen HJ, Merkal RS., 1984. Ruminant paratuberculosis (Johne’s disease): The current status and future prospects. Cornell Vet. 74:218–262.
Cirillo JD, Falkow S, Tompkins LS, Bermudez LE.,1997. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 65:3759–3767
Clemens DL, Horwitz MA.,1995. Characterization of the Mycobacterium tuberculosis phagosome and evidence that phagosomal maturation is inhibited. J.Exp.Med. 181:257–270.
Cole ST., 1999. Learning from the genome sequence of Mycobacterium tuberculosis H37Rv. FEBS Lett. 452:7–10.
Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, Gordon SV, Eiglmeier K, Gas S, Barry CE III, Tekaia F, Badcock K, Basham D, Brown D, Chillingworth T, Connor R, Davies R, Devlin K, Feltwell T, Gentles S, Hamlin N, Holroyd S, Hornsby T, Jagels K, Barrell BG., 1998. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393:537–544.
Comas I, Coscolla M, Luo T, Borrell S, Holt KE, Kato-Maeda M, Parkhill J, Malla B, Berg S, Thwaites G, Yeboah-Manu D, Bothamley G, Mei J, Wei L, Bentley S, Harris SR, Niemann S, Diel R, Aseffa A, Gao Q, Young D, Gagneux S., 2013. Out-of-Africa migration and Neolithic coexpansion of Mycobacterium tuberculosis with modern humans. Nat. Genet. Okt;45(10):1176–1182.
Coscolla M, Lewin A, Metzger S, Maetz-Rennsing K, Calvignac-Spencer S, Nitsche A, Dabrowski PW, Radonic A, Niemann S, Parkhill J, Couacy-Hymann E, Feldman J, Comas I, Boesch C, Gagneux S, Leendertz FH., 2013. Novel Mycobacterium tuberculosis complex isolate from a wild chimpanzee. Emerging Infect. Dis. Jun;19(6):969–976.
Coscolla M, Gagneux S., 2014. Consequences of genomic diversity in Mycobacterium tuberculosis. Semin Immunol. 2014 Des;26(6):431-44
Crawford MH, Campbell BC., 2012. Causes and consequences of human migration: An evolutionary perspective. Cambridge University Press. 317.
Crubézy E, Ludes B, Poveda JD, Clayton J, Crouau-Roy B, Montagnon D., 1998. Identification of Mycobacterium DNA in an Egyptian Pott’s disease of 5400 years old. C.R. Acad. Sci. III-Vie 321:941–951.
18

Daniel TM., 2006. The history of tuberculosis. Resp. Med. 100:1862–1870.
Donoghue HD, Lee OY, Minnikin DE, Besra GS, Taylor JH, Spigelman M., 2010. Tuberculosis in Dr Granville’s mummy: A molecular re-examination of the earliest known Egyptian mummy to be scientifically examined and given a medical diagnosis. Proc. Biol. Sci. Jan;7;277(1678):51–56.
Dubos R, Dubos J., 1952. The White Plague: Tuberculosis, man, and society. Little, Brown, and Company: Boston.
Espie IW, Hlokwe TM, Gey van Pittius NC, Lane E, Tordiffe AS, Michel AL, Müller A, Kotze A, Van Helden PD., 2009. Pulmonary infection due to Mycobacterium bovis in a black rhinoceros (Diceros bicornis minor) in South Africa. J. Wildl. Dis. Okt;45(4):1187–1193.
Formicola V, Milanesi Q, Scarsini C., 1987. Evidence of spinal tuberculosis at the beginning of the fourth millennium BC from Arene Candide (Liguria, Italy). Am. J. Phys. Anthropol. 72:1–6.
Gagneux S, DeRiemer K, Van T, Kato-Maeda M, de Jong BC, Narayanan S, Nicol M, Niemann S, Kremer K, Gutierrez MC, Hilty M, Hopewell PC, Small PM., 2006, Variable host-pathogen compatibility in Mycobacterium tuberculosis. Proc Natl Acad Sci U S A. 2006 Feb 21;103(8):2869-73
Gagneux S., 2012. Host–pathogen coevolution in human tuberculosis. Phil. Trans. R. Soc. B. 367:850–859.
Gelber RH.,1994. Chemotherapy of lepromatous leprosy: Recent developments and prospects for the future. Eur. J. Clin. Microbiol. Infect. Dis. 13:942–952.
Getahun H, Gunneberg C, Granich R, Nunn P., 2010. HIV infection-associated tuberculosis: The epidemiology and the response. Clin. Infect. Dis. 50(3):S201–207.
Gey van Pittius NC, Gamieldien J, Hide W, Brown GD, Siezen RJ, Beyers AD., 2001. The ESAT-6 gene cluster of Mycobacterium tuberculosis and other high G+C Gram-positive bacteria. Genome Biol, 2:0044.
Gey van Pittius NC, Perrett KD, Michel AL, Keet DF, Hlokwe T, Streicher EM, Warren RM, Van Helden PD., 2012a. Infection of African buffalo (Syncerus caffer) by oryx bacillus, a rare member of the antelope clade of the Mycobacterium tuberculosis complex. J. Wildl. Dis. Okt;48(4):849–57.
Gey van Pittius NC, Sampson SL, Lee H, Kim Y, Van Helden PD, Warren RM., 2006. Evolution and expansion of the Mycobacterium tuberculosis PE and PPE multigene families and their association with the duplication of the ESAT-6 (esx) gene cluster regions. BMC. Evol. Biol. Nov;15:6:95.
Gey van Pittius NC, Van Helden PD, Warren RM., 2012b. Characterization of Mycobacterium orygis. Emerg. Infect. Dis. Okt;18(10):1708–1709.
Gibson AL, Huard RC, Gey van Pittius NC, Lazzarini LC, Driscoll J, Kurepina N, Zozio T, Sola C, Spindola SM, Kritski AL, Fitzgerald D, Kremer K, Mardassi H, Chitale P, Brinkworth J, Garcia de Viedma D, Gicquel B, Pape JW, Van Soolingen D, Kreiswirth BN, Warren RM, Van Helden PD, Rastogi N, Suffys PN, Lapa e Silva J, Ho JL., 2008. Application of sensitive and specific molecular methods to uncover global dissemination of the major RDRio Sublineage of the Latin American-Mediterranean Mycobacterium tuberculosis spoligotype family. J. Clin. Microbiol. Apr;46(4):1259–1267.
Granville AB., 1825. An essay on Egyptian mummies; with observations on the art of embalming among the ancient Egyptians. Proc. R. Soc. Lond. 115:269–316.
Gutierrez MC, Brisse S, Brosch R, Fabre M, Omaïs B, Marmiesse M, Supply P, Vincent V., 2005. Ancient origin and gene mosaicism of the progenitor of Mycobacterium tuberculosis. PLoS Pathog.;1:e5.
Hanekom M, Gey van Pittius NC, McEvoy C, Victor TC, Van Helden PD, Warren RM., 2011. Mycobacterium tuberculosis Beijing genotype: A template for success. Tuberculosis (Edinb.). Nov;91(6):510–523.
Hanekom M, Van der Spuy GD, Gey van Pittius NC, McEvoy CR, Hoek KG, Ndabambi SL, Jordaan AM, Victor TC, Van Helden PD, Warren RM., 2008. Discordance between mycobacterial interspersed repetitive-unit-variable-number tandem-repeat typing and IS6110 restriction fragment length polymorphism genotyping for analysis of Mycobacterium tuberculosis Beijing strains in a setting of high incidence of tuberculosis. J. Clin. Microbiol. Okt;46(10):3338–3345.
Hanekom M, Van der Spuy GD, Gey van Pittius NC, McEvoy CR, Ndabambi SL, Victor TC, Hoal EG, Van Helden PD, Warren RM., 2007. Evidence that the spread of Mycobacterium tuberculosis strains with the Beijing genotype is human population dependent. J. Clin. Microbiol. Jul;45(7):2263–2266.
Hansen GHA., 1874. Undersogelser angaende spedalskhedens aasager. Norsk Magazin for Laegervidenskaben 4 (Suppl.), 1–88.
Hansen GHA., 1880. Bacillus leprae. Virchows Archiv. 79:32–42.
Havlir DV., 1994. Mycobacterium avium complex: Advances in therapy. Eur. J. Clin. Microbiol. Infect. Dis. 13:915–924.
Herodotus, The Histories, Boek 7 (Polymnia), Hoofstuk 88, vertaler G. C. Macaulay, derde uitgawe, gedruk in 1914, MacMillan and Co., Limited, St. Martin’s Street, Londen.
Hershberg R, Lipatov M, Small PM, Sheffer H, Niemann S, Homolka S, Roach JC, Kremer K, Petrov DA, Feldman MW, Gagneux S., 2008. High functional diversity in Mycobacterium tuberculosis driven by genetic drift and human demography. PLoS Biol. 6:e311.
Hershkovitz I, Donoghue HD, Minnikin DE, Besra GS, Lee OY, Gernaey AM, Galili E, Eshed V, Greenblatt CL, Lemma E, Bar-Gal GK, Spigelman M., 2008. Detection and molecular characterization of 9000-year-old Mycobacterium tuberculosis from a Neolithic settlement in the Eastern Mediterranean. PLoS ONE 3, e3426.
Hoek KG, Gey van Pittius NC, Moolman-Smook H, Carelse-Tofa K, Jordaan A, Van der Spuy GD, Streicher E, Victor TC, Van Helden PD, Warren RM., 2008. Fluorometric assay for testing rifampin susceptibility of Mycobacterium tuberculosis complex. J. Clin. Microbiol. Apr;46(4):1369–1373.
Hoek KG, Schaaf HS, Gey van Pittius NC, Van Helden PD, Warren RM., 2009. Resistance to pyrazinamide and ethambutol compromises MDR/XDR-TB treatment. S. Afr. Med. J. Nov;99(11):785–787.
Hopewell PC., 1994. The baby and the bath water. The case for retaining categorical services for tuberculosis control in a reformed health care system. Am. J. Respir. Crit Care Med. 150:895.
19

Houben D, Demangel C, Van Ingen J, Perez J, Baldeón L, Abdallah AM, Caleechurn L, Bottai D, Van Zon M, De Punder K, Van der Laan T, Kant A, Bossers-de Vries R, Willemsen P, Bitter W, Van Soolingen D, Brosch R, Van der Wel N, Peters PJ., 2012. ESX-1-mediated translocation to the cytosol controls virulence of mycobacteria. Cell. Microbiol. 14:1287–1298.
Ioerger TR, Feng Y, Chen X, Dobos KM, Victor TC, Streicher EM, Warren RM, Gey van Pittius NC, Van Helden PD, Sacchettini JC., 2010. The non-clonality of drug resistance in Beijing-genotype isolates of Mycobacterium tuberculosis from the Western Cape of South Africa. BMC Genomics. Nov;26(11):670.
Jang J, Becq J, Gicquel B, Deschavanne P, Neyrolles O., 2008. Horizontally acquired genomic islands in the tubercle bacilli. Trends Microbiol. Jul;16(7):303–308.
Johnson R, Warren RM, Van der Spuy GD, Gey van Pittius NC, Theron D, Streicher EM, Bosman M, Coetzee GJ, Van Helden PD, Victor TC., 2010. Drug-resistant tuberculosis epidemic in the Western Cape driven by a virulent Beijing genotype strain. Int. J. Tuberc. Lung. Dis. Jan;14(1):119–121.
Karboul A, Gey van Pittius NC, Namouchi A, Vincent V, Sola C, Rastogi N, Suffys P, Fabre M, Cataldi A, Huard RC, Kurepina N, Kreiswirth B, Ho JL, Gutierrez MC, Mardassi H., 2006. Insights into the evolutionary history of tubercle bacilli as disclosed by genetic rearrangements within a PE_PGRS duplicated gene pair. BMC Evol. Biol. Des;12(6):107.
Karboul A, Mazza A, Gey van Pittius NC, Ho JL, Brousseau R, Mardassi H., 2008. Frequent homologous recombination events in Mycobacterium tuberculosis PE/PPE multigene families: Potential role in antigenic variability. J. Bacteriol. Des;190(23):7838–7846.
Klopper M, Warren RM, Hayes C, Gey van Pittius NC, Streicher EM, Müller B, Sirgel FA, Chabula-Nxiweni M, Hoosain E, Coetzee G, van Helden PD, Victor TC, Trollip AP., 2013. Emergence and spread of extensively and totally drug-resistant tuberculosis, South Africa. Emerg. Infect. Dis. Mrt;19(3):449–455.
Koch R., 1882. Die Aetiologie der Tuberkulose. Ber. Klin. Wochenschrift. 15.
López B, Aguilar D, Orozco H, Burger M, Espitia C, Ritacco V, Barrera L, Kremer K, Hernandez-Pando R, Huygen K, van Soolingen D., 2003. A marked difference in pathogenesis and immune response induced by different Mycobacterium tuberculosis genotypes. Clin Exp Immunol. 2003 Jul;133(1):30-7.
Louw GE, Warren RM, Gey van Pittius NC, Leon R, Jimenez A, Hernandez-Pando R, McEvoy CR, Grobbelaar M, Murray M, Van Helden PD, Victor TC., 2011. Rifampicin reduces susceptibility to ofloxacin in rifampicin-resistant Mycobacterium tuberculosis through efflux. Am. J. Respir. Crit. Care Med. Jul 15;184(2):269–276.
Louw GE, Warren RM, Gey van Pittius NC, McEvoy CR, Van Helden PD, Victor TC., 2009. A balancing act: Efflux/influx in mycobacterial drug resistance. Antimicrob. Agents Chemother. Aug;53(8):3181–3189.
Lucas S, Nelson AM., 2015. HIV and the spectrum of human disease. J. Pathol. Jan;235(2):229–241.
Major RH., 1945. Classic descriptions of disease, 3de uitg. Springfield, IL: Charles C. Thomas..
Mardassi H, Namouchi A, Haltiti R, Zarrouk M, Mhenni B, Karboul A, Khabouchi N, Gey van Pittius NC, Streicher EM, Rauzier J, Gicquel B, Dellagi K., 2005. Tuberculosis due to resistant Haarlem strain, Tunisia. Emerg. Infect. Dis. Jun;11(6):957–961.
Martin LD, Rothschild BM., 1989, Frequency of pathology in a large natural sample from Natural Trap Cave (late Pleistocene). J Vert Paleontol. 9:31
McEvoy CR, Falmer AA, Gey van Pittius NC, Victor TC, Van Helden PD, Warren RM., 2007. The role of IS6110 in the evolution of Mycobacterium tuberculosis. Tuberculosis (Edinb.). Sep;87(5):393–404.
McEvoy CR, Van Helden PD, Warren RM, Gey van Pittius NC., 2009a. Evidence for a rapid rate of molecular evolution at the hypervariable and immunogenic Mycobacterium tuberculosis PPE38 gene region. BMC Evol. Biol. Sep 21;9:237.
McEvoy CR, Warren RM, Van Helden PD, Gey van Pittius NC., 2009b. Multiple, independent, identical IS6110 insertions in Mycobacterium tuberculosis PPE genes. Tuberculosis (Edinb.). Nov;89(6):439–442.
McEvoy CRE, Cloete R, Müller B, Schürch AC, Van Helden PD, Gagneux S, Warren RM, Gey van Pittius NC., 2012. Comparative analysis of Mycobacterium tuberculosis PE and PPE genes reveals high sequence variation and an apparent absence of selective constraints. PLoS ONE 7(4):e30593.
McKinney JD, Jacobs WR Jr, Bloom BR., 1998. Persisting problems in tuberculosis. In ‘Emerging Infections’. 51–146. (Academic Press)
McShane H., 2005. Co-infection with HIV and TB: Double trouble. Int. J. STD AIDS.;16:95–101.
Michel AL, De Klerk LM, Gey van Pittius NC, Warren RM, Van Helden PD., 2007. Bovine tuberculosis in African buffaloes: Observations regarding Mycobacterium bovis shedding into water and exposure to environmental mycobacteria. BMC Vet. Res. Sep 27;3:23.
Müller B, Chihota VN, Pillay M, Klopper M, Streicher EM, Coetzee G, Trollip A, Hayes C, Bosman ME, Gey van Pittius NC, Victor TC, Gagneux S, Van Helden PD, Warren RM., 2013. Programmatically selected multidrug-resistant strains drive the emergence of extensively drug-resistant tuberculosis in South Africa, PLoS One. Aug 23;8(8):e70919.
Müller B, Streicher EM, Hoek KG, Tait M, Trollip A, Bosman ME, Coetzee GJ, Chabula-Nxiweni EM, Hoosain E, Gey van Pittius NC, Victor TC, Van Helden PD, Warren RM., 2011. InhA promoter mutations: A gateway to extensively drug-resistant tuberculosis in South Africa? Int. J. Tuberc. Lung Dis. Mrt;15(3):344–351.
Nerlich AG, Haas CJ, Zink A, Szeimies U, Hagedorn HG., 1997. Molecular evidence for tuberculosis in an ancient Egyptian mummy. Lancet 350:1404.
Nuorala, E; Götherström, A; Ahlström, T; Donoghue, HD; Spigelman, M; Lidén, K, 2004. MTB complex DNA in a Scandinavian Neolithic passage grave, Theses and Papers in Scientific Archaeology 6, Argeologiese Navorsingslaboratorium, Universiteit van Stockholm.
20

Ortner DJ, Putschar WG., 1981. Identification of pathological conditions on human skeletal remains. Washington DC: Smithsonian Institution Press.
Packard RM., 1989. White Plague, Black Labor – Tuberculosis and the Political Economy of Health and Disease in South Africa. November, University of California Press.
Parsons S, Smith SGD, Martins Q, Horsnell WGC, Gous TA, Streicher EM, Warren RM, Van Helden PD, Gey van Pittius NC., 2008. Pulmonary infection due to the dassie bacillus (Mycobacterium sp) in a free-living dassie (rock hyrax – Procavia capensis) from South Africa. Tuberculosis. Jan;88(1):80–83.
Parsons SD, Cooper D, McCall AJ, McCall WA, Streicher EM, Le Maitre NC, Müller A, Gey van Pittius NC, Warren RM, Van Helden PD., 2011. Modification of the QuantiFERON-TB Gold (In-Tube) assay for the diagnosis of Mycobacterium bovis infection in African buffaloes (Syncerus caffer). Vet. Immunol. Immunopathol. Jul 15;142(1–2):113–118.
Parsons SD, Drewe JA, Gey van Pittius NC, Warren RM, Van Helden PD., 2013. Novel cause of tuberculosis in meerkats, South Africa. Emerg. Infect. Dis. Des;19(12):2004–2007.
Parsons SD, Warren RM, Ottenhoff TH, Gey van Pittius NC, Van Helden PD., 2012. Detection of Mycobacterium tuberculosis infection in dogs in a high-risk setting. Res. Vet. Sci. Jun;92(3):414–419.
Paulson T, 2013. Outlook Tuberculosis – A mortal foe. Nature. Okt 10;502(7470):S2-3.
Pillay M, Sturm AW., 2007. Evolution of the extensively drug-resistant F15/LAM4/KZN strain of Mycobacterium tuberculosis in KwaZulu-Natal, South Africa. Clin Infect Dis. Des 1;45(11):1409-14
Pym AS, Brodin P, Brosch R, Huerre M, Cole ST., 2002. Loss of RD1 contributed to the attenuation of the live tuberculosis vaccines Mycobacterium bovis BCG and Mycobacterium microti. Mol. Microbiol. Nov;46(3):709–717.
Riley RI, Mills CC, Nyka W, Weinstock N, Storey PB, Sultan LU, Riley MC, Wells WF., 1959. Aerial dissemination of pulmonary tuberculosis: A two-year study of contagion in a tuberculosis ward. Am. J. Hygiene. 70:185–196.
Roberts CA, Buikstra JE. 2003. The Bioarchaeology of Tuberculosis: A Global View on a Reemerging Disease, University Press of Florida.
Roberts CA, Pfister LA, Mays S., 2009. Letter to the editor: Was tuberculosis present in Homo erectus in Turkey? Am. J. Phys. Anthropol. 139:442–444.
Rothschild BM, Martin LD, Lev G, Bercovier H, Bar-Gal GK, Greenblatt C, Donoghue H, Spigelman M, Brittain D., 2001. Mycobacterium tuberculosis complex DNA from an extinct bison dated 17,000 years before the present. Clin Infect Dis. Aug 1;33(3):305-11.
Rothschild BM, Martin LD., 2006. Did ice-age bovids spread tuberculosis? Naturwissenschaften. 93:565–569.
Rothschild BM, Laub R., 2006. Hyperdisease in the late Pleistocene: validation of an early 20th century hypothesis. Naturwissenschaften. Nov;93(11):557-64.
Salo WL, Aufderheide AC, Buikstra J, Holcomb TA., 1994. Identification of Mycobacterium tuberculosis DNA in a pre-Columbian Peruvian mummy. Proc. Natl. Acad. Sci. U.S.A. 91:2091–2094.
Sassetti CM, Boyd DH, Rubin EJ., 2003. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 48:77–84.
Serafini A, Pisu D, Palù G, Rodriguez GM, Manganelli R., 2013. The ESX-3 secretion system is necessary for iron and zinc homeostasis in Mycobacterium tuberculosis, PLoS One. Okt 14;8(10):e78351.
Shinnick TM, Good RC., 1994. Mycobacterial taxonomy. Eur. J. Clin. Microbiol. Infect. Dis. 13:884–901.
Simeone R, Bobard A, Lippmann J, Bitter W, Majlessi L, Brosch R, Enninga J., 2012. Phagosomal rupture by Mycobacterium tuberculosis results in toxicity and host cell death. PLoS Pathog. 8:e1002507 d.
Sirgel FA, Tait M, Warren RM, Streicher EM, Böttger EC, Van Helden PD, Gey van Pittius NC, Coetzee G, Hoosain EY, Chabula-Nxiweni M, Hayes C, Victor TC, Trollip A., 2012. Mutations in the rrs A1401G gene and phenotypic resistance to amikacin and capreomycin in Mycobacterium tuberculosis. Microb. Drug Resist. Apr;18(2):193–197.
Sizaire V, Nackers F, Comte E, Portaels F., 2006. Mycobacterium ulcerans infection: control, diagnosis, and treatment. Lancet Infect. Dis. 6(5): 288–296
Springer B, Stockman L, Teschner K, Roberts GD, Bottger EC., 1996. Two-laboratory collaborative study on identification of mycobacteria: Molecular versus phenotypic methods. J. Clin. Microbiol. 34:296–303.
Stead WW., 1997. The origin and erratic global spread of tuberculosis. How the past explains the present and is the key to the future. Clin. Chest Med. 18:65–77.
Streicher EM, Müller B, Chihota V, Mlambo C, Tait M, Pillay M, Trollip A, Hoek KG, Sirgel FA, Gey van Pittius NC, Van Helden PD, Victor TC, Warren RM., 2012. Emergence and treatment of multidrug resistant (MDR) and extensively drug-resistant (XDR) tuberculosis in South Africa. Infect. Genet. Evol. Jun;12(4):686–694.
Taylor SJ, Ahonen LJ, De Leij FA, Dale JW., 2003. Infection of Acanthamoeba castellanii with Mycobacterium bovis and M. bovis BCG and survival of M. bovis within the amoebae. Appl. Environ. Microbiol. 69: 4316–4319.
Tekaia F, Gordon SV, Garnier T, Brosch R, Barrell BG, Cole ST., 1999. Analysis of the proteome of Mycobacterium tuberculosis in silico. Tuber. Lung Dis.79(6):329–342.
Trail RR., 1970. Richard Morton (1637–1698). Med. Hist. Apr;14(2):166–174.
Van der Spuy GD, Kremer K, Ndabambi SL, Beyers N, Dunbar R, Marais BJ, van Helden PD, Warren RM., 2009. Changing Mycobacterium tuberculosis population highlights clade-specific pathogenic characteristics. Tuberculosis (Edinb). Mrt;89(2):120-5
21

Van Ingen J, Rahim Z, Mulder A, Boeree MJ, Simeone R, Brosch R, Van Soolingen D., 2012. Characterization of Mycobacterium orygis as M. tuberculosis complex subspecies. Emerg. Infect. Dis. Apr;18(4):653–655.
Van Rie A, Victor TC, Richardson M, Johnson R, Van der Spuy GD, Murray EJ, Beyers N, Gey van Pittius NC, Van Helden PD, Warren RM., 2005. Reinfection and mixed infection cause changing Mycobacterium tuberculosis drug-resistance patterns. Am. J. Respir. Crit. Care Med. Sep 1;172(5):636–642.
Veyrier F, Pletzer D, Turenne C, Behr MA., 2009. Phylogenetic detection of horizontal gene transfer during the step-wise genesis of Mycobacterium tuberculosis. BMC Evol. Biol. Aug 10;9:196.
Veyrier FJ, Dufort A, Behr MA., 2011. The rise and fall of the Mycobacterium tuberculosis genome. Trends Microbiol. Apr;19(4):156–161.
Victor TC, de Haas PE, Jordaan AM, van der Spuy GD, Richardson M, van Soolingen D, van Helden PD, Warren R., 2004. Molecular characteristics and global spread of Mycobacterium tuberculosis with a Western Cape F11 genotype. J. Clin. Microbiol. 42:769-772
Waksman SA., 1964. The Conquest of Tuberculosis, University of California Press; Eerste Uitgawe.
Warren RM, Gey van Pittius NC, Barnard M, Hesseling A, Engelke E, De Kock M, Gutierrez MC, Chege GK, Victor TC, Hoal EG, Van Helden PD., 2006. Differentiation of Mycobacterium tuberculosis complex by PCR amplification of genomic regions of difference. Int. J. Tuberc. Lung Dis. Jul;10(7):818–822.
Warren RM, Streicher EM, Gey van Pittius NC, Marais BJ, Van der Spuy GD, Victor TC, Sirgel F, Donald PR, Van Helden PD., 2009a. The clinical relevance of Mycobacterial pharmacogenetics. Tuberculosis (Edinb.). Mei;89(3):199–202.
Warren RM, Van Helden PD, Gey van Pittius NC., 2009b. Insertion element IS6110-based restriction fragment length polymorphism genotyping of Mycobacterium tuberculosis. Methods Mol. Biol. 465:353–370.
Wasserman S, Meintjes G., 2014.The diagnosis, management and prevention of HIV-associated tuberculosis. S. Afr. Med. J. Des;104(12):886–893.
Wayne LG, Kubica GP., 1986. The mycobacteria. In ‘Bergey’s Manual of Systematic Bacteriology’, Reds. PHA Sneath, NS Mair, ME Sharpe, JG Holt., Williams and Wilkins: Baltimore, p1436–1457.
WGO.,1993. WHO declares tuberculosis a global emergency. Soz. Praventivmed. 38:251–252.
WGO., 2013. Global tuberculosis report 2013. Geneva: World Health Organization; http://www.who.int/tb/publications/global_report/en/. (Toegang bekom Jul 30, 2015)
WGO., 2014a. Guidance for National Tuberculosis Programmes on the Management of Tuberculosis in Children. 2de uitgawe. Genève: Wêreldgesondheidsorganisasie; 2014.
WGO., 2014b. Global tuberculosis report 2014. Genève: Wêreldgesondheidsorganisasie; 2014. http://www.who.int/tb/publications/global_report/en/. (Toegang bekom Jul 30, 2015)
WGO., 2014c. “WHO’s first global report on antibiotic resistance reveals serious, worldwide threat to public health” (Persverklaring). Wêreldgesondheidsorganisasie. April 30, 2014.
WGO., 2015. Feiteblad N°104 Hersien Maart 2015 Fact sheet on Tuberculosis (TB).
Weiner B, Gomez J, Victor TC, Warren RM, Sloutsky A, Plikaytis BB, Posey JE, Van Helden PD, Gey van Pittius NC, Koehrsen M, Sisk P, Stolte C, White J, Gagneux S, Birren B, Hung D, Murray M, Galagan J., 2012. Independent large scale duplications in multiple M. tuberculosis lineages overlapping the same genomic region. PLoS One. 7(2):e26038.
Zink AR, Grabner W, Reischl U, Nerlich AG., 2003. Molecular study on human tuberculosis in three geographically distinct and time delineated populations from ancient Egypt. Epidemiol. Infect. 130:239–249.
Zysk KG., 1998. Medicine in the Veda: Religious Healing in the Veda. Motilal Banarsidass Publ. ISBN 8120814002.
22





























