Embed Size (px)
Citation preview

ANKARA ÜNİVERSİTESİ FEN BİLİMLERİ ENSTİTÜSÜ
DOKTORA TEZİ
TÜRKİYE’DEKİ Lathyrus L. (FABACEAE) CİNSİ Cicercula SEKSİYONUNUN RAPD VE SDS-PAGE ANALİZİ
İsmail Ömür ACAR
BİYOLOJİ ANABİLİM DALI
ANKARA 2011
Her hakkı saklıdır

i
ÖZET
Doktora Tezi
TÜRKİYE’DEKİ Lathyrus L. (FABACEAE) CİNSİ Cicercula SEKSİYONUNUN RAPD VE SDS-PAGE ANALİZİ
İsmail Ömür ACAR
Ankara Üniversitesi
Fen Bilimleri Enstitüsü Biyoloji Anabilim Dalı
Danışman: Yrd. Doç. Dr. H. Nurhan BÜYÜKKARTAL Eş Danışman: Prof. Dr. Reyhan ÇOLAK
Lathyrus L. cinsi üyeleri, çoğunlukla hayvan yemi olmakla beraber, toprak için yeşil gübre bitkisi, süs bitkisi ve kısmen de insan besini olarak kullanılan ekonomik önem taşıyan bitkilerdir. Cinsin taksonomik çalışmalarında araştırmacılar arasında görüş ayrılıkları bulunmaktadır. Bu çalışmada Türkiye’deki Lathyrus L. cinsi Cicercula seksiyonunun bazı türlerinin DNA ve tohum depo proteinleri varyasyonları kullanılarak, aralarındaki evrimsel ilişkiler incelenmiştir. Böylece cinsin taksonomik problemlerinin çözümüne katkı sağlanmasına çalışılmıştır. DNA varyasyon analizi için RAPD, tohum depo proteinleri varyasyon analizi için SDS-PAGE teknikleri kullanılmıştır. Elde edilen veriler, POPGENE v1.31 ve MEGA v3.0 bilgisayar programları aracılığıyla istatistiksel olarak değerlendirilmiş ve filogenetik dendogramlar çıkarılmıştır. Sonuç olarak; SDS-PAGE ve RAPD analizi sonuçlarının birebir örtüşmediği görülmüştür. RAPD analizi ve kısmi olarak da SDS-PAGE analizi sonuçlarına göre, daha önceki çalışmalarla uyumlu olarak, L. sativus ve L. cicera’nın birbirlerine genetik açıdan çok yakın türler olduğu belirlenmiştir. Çalışılan Cicercula seksiyonunun diğer türlerinin ise daha önceki çalışmalarla uyumlu olmayan filogenetik ilişkiler sergilediği belirlenmiştir. Aralık 2011, 125 sayfa Anahtar Kelimeler: Lathyrus, Cicercula, Fabaceae, Türkiye, RAPD, SDS-PAGE

ii
ABSTRACT
Ph.D. Thesis
RAPD AND SDS-PAGE ANALYSIS OF Cicercula SECTION OF Lathyrus L. (FABACEAE) GENUS IN TURKEY
İsmail Ömür ACAR
Ankara University
Graduate School of Natural and Applied Sciences Department of Biology
Supervisor: Asst. Prof. Dr. H. Nurhan BÜYÜKKARTAL Co-Supervisor: Prof. Dr. Reyhan ÇOLAK
Individuals of Lathyrus L. genus are economically important plants which are mostly used as animal feed and also as green manure crops, ornamental plant and occasionally for human nutrition. There is dissensus among researchers regarding taxonomic studies of the genus. This study investigates the evolutionary relations among some species of Lathyrus L. genus Cicercula section in Turkey by using variations of DNA and seed storage proteins to contribute to the solution of taxonomic problems of the genus. The study employed RAPD technique for DNA variations analysis and SDS-PAGE technique for seed storage protein variations analyses. POPGENE v1.31 and MEGA v3.0 softwares were used for statistical assessment of the obtained data and phylogenetic dendrograms were produced. As a result of the study, the analysis results obtained with SDS-PAGE and RAPD techniques were identified to be incompatible with each other. Based on RAPD analysis results and partially on SDS-PAGE analysis results, L. sativus and L. cicera were identified to be genetically very similar species, in agreement with former research findings. On the other hand, other species of the Cicercula section were found to display phylogenetic relations which are incompatible with previous literature. December 2011, 125 pages Key Words: Lathyrus, Cicercula, Fabaceae, Turkey, RAPD, SDS-PAGE

iii
TEŞEKKÜR
Çalışmalarım sırasında büyük yardımlarını gördüğüm, daima beni motive eden değerli
danışmanlarım sayın Yrd. Doç. Dr. Nurhan BÜYÜKKARTAL ve Prof. Dr. Reyhan
ÇOLAK (Ankara Üniversitesi, Biyoloji Anabilim Dalı) başta olmak üzere,
araştırmalarıma tecrübeleriyle katkıda bulunan, önerileriyle beni yönlendiren sayın
hocalarım Doç. Dr. E. Sümer ARAS’a (Ankara Üniversitesi, Biyoloji Anabilim Dalı) ve
Prof. Dr. Leyla AÇIK’a (Gazi Üniversitesi, Biyoloji Anabilim Dalı), çalışma
materyalimin büyük çoğunluğunu temin etmeme yardımcı olan ve materyalim
konusundaki engin bilgisini benimle paylaşan sayın hocam Prof. Dr. A. Selçuk
ERTEKİN’e (Dicle Üniversitesi, Biyoloji Anabilim Dalı), değerli fikirlerini benimle
paylaşan Dr. Ömer Faruk KAYA’ya (Harran Üniversitesi, Biyoloji Anabilim Dalı),
Ankara Üniversitesi, Biyoteknoloji Enstitüsü Merkez Laboratuvarı’nda, DNA kalite ve
miktar tespitlerinde yardımcı olan sayın hocam Prof. Dr. Ali ERGÜL’e,
laboratuvarlarındaki görüntüleme sistemleri ile jellerin görüntülenmesine yardımcı olan
sayın hocalarım Prof.Dr. Mustafa AKÇELİK’e (Ankara Üniversitesi, Biyoloji Anabilim
Dalı) ve Doç.Dr. Özlem OSMANAĞAOĞLU’na (Ankara Üniversitesi, Biyoloji
Anabilim Dalı), laboratuvar çalışmalarım boyunca yardımlarını eksik etmeyen sevgili
arkadaşlarım Arş. Gör. Gül OLGUN, Arş. Gör. Canan YAĞCI ve Arş. Gör. Dr. Şenol
ALAN’a, materyal temininde yardımcı olan Ege Tarımsal Araştırmalar Enstitüsü’ne,
daima desteğini arkamda hissettiğim eşim Doç. Dr. Banu Çelikel ACAR’a, onlarla
birlikte geçireceğim zamanlarda, onlarla birlikte olamadığımda büyük olgunluk
gösteren küçük kızlarım Melis ve Selen ACAR’a sonsuz teşekkürlerimi sunarım.
Bu çalışma Ankara Üniversitesi Bilimsel Araştırma Projeleri Müdürlüğü (BAP)
tarafından 09B4240014 nolu proje ile desteklenmiştir.
İsmail Ömür ACAR
Ankara, Aralık 2011

iv
İÇİNDEKİLER
ÖZET.................................................................................................................................i
ABSTRACT.....................................................................................................................ii
TEŞEKKÜR ...................................................................................................................iii
SİMGELER ve KISALTMALAR DİZİNİ .................................................................vii
ŞEKİLLER DİZİNİ ........................................................................................................x
ÇİZELGELER DİZİNİ ...............................................................................................xiii
1. GİRİŞ ...........................................................................................................................1
2. KURAMSAL TEMELLER........................................................................................6
2.1 Lathyrus L. sistematiği ..............................................................................................6
2.1.1 Familya: Leguminosae [Fabaceae (Baklagiller)].................................................7
2.1.1.1 Altfamilya: Papilionoideae .................................................................................8
2.2 Moleküler Sistematik ..............................................................................................11
2.3 Moleküler Belirteçler ..............................................................................................13
2.3.1 Proteinler ..............................................................................................................13
2.3.1.1 Tohum depo proteinleri....................................................................................13
2.3.1.2 Enzimler.............................................................................................................15
2.3.1.3 Protein izolasyonu .............................................................................................16
2.3.2 DNA .......................................................................................................................17
2.3.2.1 Nokta mutasyonlar............................................................................................19
2.3.2.2 İnsersiyonlar ve delesyonlar.............................................................................19
2.3.2.3 Yeniden düzenlenmeler ....................................................................................20
2.3.2.4 DNA izolasyonu .................................................................................................21
2.4 Moleküler Belirteç Analizleri.................................................................................22
2.4.1 Protein elektroforezi ............................................................................................22
2.4.2 DNA varyasyon analizleri ...................................................................................24
2.4.2.1 Restriksiyon enzim analizi................................................................................25
2.4.2.2 DNA dizi analizi ................................................................................................26
2.4.2.3 PZR ve PZR bazlı parmak izi teknikleri ........................................................27
2.4.2.4 Polimeraz Zincir Reaksiyonu (PZR) ...............................................................28

v
2.4.2.5 RAPD (Random Amplified Polymorphic DNA) ............................................ 29
3. MATERYAL ve METOT.........................................................................................32
3.1 Materyal ...................................................................................................................32
3.2 Metot ........................................................................................................................37
3.2.1 RAPD.....................................................................................................................37
3.2.1.1 DNA izolasyonu .................................................................................................37
3.2.1.1.1 CTAB (Cetyl Trimethyl Ammonium Bromide) DNA izolasyonu (Doyle
ve Doyle 1991).................................................................................................. 37
3.2.1.1.2 Modifiye CTAB DNA izolasyonu (Aras vd. 2003) ......................................38
3.2.1.1.3 Hazır ekstraksiyon kiti (Favorprep Plant Genomic DNA extraction
mini kit)............................................................................................................ 39
3.2.1.1.4 DNA saflık ve miktar tayini ..........................................................................41
3.2.1.1.5 Total DNA elektroforezi ................................................................................42
3.2.1.2 Primerler............................................................................................................42
3.2.1.3 PZR optimizasyonu...........................................................................................43
3.2.1.4 Agaroz jel elektroforezi ve görüntüleme.........................................................47
3.2.1.5 Jellerin değerlendirilmesi .................................................................................48
3.2.1.6 RAPD verilerinin analizi (istatistik analizi)...................................................48
3.2.2 SDS-PAGE............................................................................................................48
3.2.2.1 Protein ekstraksiyonu .......................................................................................48
3.2.2.1.1 Protein miktar tayini .....................................................................................49
3.2.2.2 Protein elektroforezi .........................................................................................49
3.2.2.2.1 Poliakrilamid jellerin hazırlanması..............................................................49
3.2.2.2.2 Örneklerin yüklenmesi ..................................................................................51
3.2.2.2.3 Poliakrilamid jel boyanması .........................................................................52
3.2.2.2.4 Bant desenlerinin görüntülenmesi ................................................................53
3.2.2.3 SDS-PAGE verilerinin analizi (istatistik analizi) ...........................................53
4. ARAŞTIRMA BULGULARI...................................................................................54
4.1 RAPD........................................................................................................................54
4.1.1 DNA izolasyonu ....................................................................................................54
4.1.1.1 DNA miktar tayini ............................................................................................54
4.1.1.2 Total DNA elektroforezi ...................................................................................54

vi
4.1.2 PZR optimizasyonu..............................................................................................58
4.1.3 RAPD polimorfizmi ve genetik mesafelerin belirlenmesi.................................59
4.1.3.1 Türler arası genetik mesafeler ........................................................................59
4.1.3.2 Tür içi genetik mesafeler ..................................................................................91
4.2 SDS-PAGE...............................................................................................................97
4.2.1 Protein ekstraksiyonu ..........................................................................................97
4.2.1.1 Protein miktar tayini ........................................................................................97
4.2.2 Depo proteinleri polimorfizmi ve genetik mesafelerin belirlenmesi................97
5. TARTIŞMA ve SONUÇ .........................................................................................102
KAYNAKLAR ............................................................................................................109
EKLER.........................................................................................................................116
EK 1 RAPD bant verileri matrisi ..............................................................................117
EK 2 SDS-PAGE bant verileri matrisi......................................................................124
ÖZGEÇMİŞ.................................................................................................................125

vii
SİMGELER ve KISALTMALAR DİZİNİ
%P Polimorfizm Yüzdesi
(NH4)SO4 Amonyum Sülfat
°C Santigrad Derece
A Adenin
Abs Absorbans
AFLP Çogaltılmıs Parça Uzunluk Polimorfizmi
AP-PCR Rastgele başlatıcılı-Polimeraz Zincir Reaksiyonu
APS Amonyum Per Sülfat
BAPN β-aminopropionitril
BME Beta Merkapto Etanol
bp Baz Çifti
C Sitozin
C:IAA Kloroform:izoamil alkol
CBB Coomassie parlak mavisi
Cm Santimetre
cpDNA Kloroplast DNA’sı
CTAB Setil Trimetil Amonyum Bromid
D Nei’nin genetik mesafe değeri
DABA L-2,4 diaminobutirik asid
DAF DNA Çoğaltım Parmak İzi
ddNTP dideoksinükleotid trifosfat
Dg4 Dış Grup 4. Birey
DNA Deoksiribo Nükleik Asit
dNTP Deoksinükleotid trifosfat
DUF Dicle Üniversitesi Herbaryumu
EDTA Etilen Diamin Tetraasetik Asit
ETAE Ege Tarımsal Araştırma Enstitüsü
ETOH Etanol
G Guanin

viii
GST Populasyonlar arasındaki genetik farklılaşmanın nisbi
büyüklüğü
H Nei’nin genetik çeşitliliği
HCl Hidroklorik asit
I Shannon information indeksi
ISSR Ara Basit Sıra Tekrarlar
ITS Internal Transcribed Spacer
K Negatif Kontrol
kDA Kilodalton
M Marker, Belirteç
MEGA Moleküler Evrimsel Genetik Analizi
Mg Miligram
MgCL2 Magnezyum Klorür
mL Mililitre
mM Milimol
mRNA Mesajcı RNA
mtDNA Mitokondri DNA’sı
Na Gözlenen Allel Sayısı
NaCl Sodyum Klorür
Ne Etkili allel sayısı
ng Nanogram
Nm Gen akışı
nm Nanometre
OD Optik Dansite
ODAP β-N-oxalyl-L, α-βdiamino propionik asid
P Fosfor
PCR Polimeraz Zincir reaksiyonu
pH Hidrojen konsantrasyonunun eksi logaritması
pMol Pikomol
RAPD Rastgele Çogaltılmıs Polimorfik DNA
RE Restriksiyon Enzimleri
RFLP Restriksiyon Parça Uzunluk Polimorfizmi

ix
RNA Ribonükleik asit
rpm Bir Dakikadaki Dönüş Sayısı
S Svedberg Ünitesi
SDS-PAGE Sodyum Dodesil Sülfat–Poliakrilamid Jel Elektroforezi
SEF Sarı Elçi Fiği
T Timin
TAE Tris Asetik asit EDTA
Taq Thermus aquaticus
TE Tris-EDTA
TEMED Tetrametiletilendiamin
Tm Çözünme Sıcaklığı
tRNA Taşıyıcı RNA
TUBİVES Türkiye Bitkileri Veri Sistemi
UPGMA Unweighted Pair Group Method with Arithmetic Mean
UV Ultraviyole
V Hacim
μL Mikrolitre

x
ŞEKİLLER DİZİNİ
Şekil 2.1 L. sativus: a. Çiçek, b. Meyve (
http://de.wikipedia.org/wiki/Datei:Lathyrus_sativus_002.JPG , 2011) ....... 10
Şekil 2.2 L. cicera: a. Çiçek, b. Meyve
(http://www.menudanatura.com/2011/05/lathyrus-cicera-l.html, 2011)......10
Şekil 2.3 DNA Molekülü (http://whyfiles.org/034clone/images/dna_molecule.gif,
2011) ............................................................................................................ 17
Şekil 2.4 DNA’daki baz dizilimi (http://www.webhatti.com/saglik/60365-dna-
nedir-nerede-bulunur.html, 2011) ................................................................ 18
Şekil 2.5 Nokta mutasyon (de Vicente ve Fulton 2003) .............................................19
Şekil 2.6 İnsersiyon (de Vicente ve Fulton 2003) .......................................................20
Şekil 2.7 Delesyon (de Vicente ve Fulton 2003).........................................................20
Şekil 2.8 Yeniden düzenlenme (de Vicente ve Fulton 2003)......................................20
Şekil 2.9 Protein elektroforezi (http://en.wikipedia.org/wiki/SDS-PAGE, 2011) .....23
Şekil 2.10. a. Varyasyon tespiti, b. İlişkilerin hesaplanması, c. Dendrogram (de
Vicente vd. 2004) ......................................................................................... 25
Şekil 2.11 RAPD’in şematik gösterimi (Özcan vd. 2004) ............................................ 31
Şekil 3.1 TUBİVES (Türkiye Bitkileri Veri Sistemi) toplanma bölgeleri, kareler
(http://wwweski.tubitak.gov.tr/tubives/ ,2011) ............................................ 35
Şekil 3.2 Agilent 2100 Bioanalyser NanoDrop ND-1000 spektrofotometre
(http://cgb.indiana.edu/genomics/equipment/15,2011)................................ 41
Şekil 3.3 KODAK Gel Logic 2200 görüntüleme sistemi
(http://blestbj.com/product/dianyong/ningjiao/2009/0605/726.html ,2011).42
Şekil 3.4 Çalışmada kullanılan markerler: a. GeneRuler™ 100 bp Plus DNA
Ladder, 100-3000 bp, b. GeneRuler™ 100 bp DNA Ladder 100-1000 bp. 47
Şekil 3.5 Çalışmada kullanılan tohumlardan protein ektraksiyonu için kullanılan
elektrikli homojenizatör ............................................................................... 49
Şekil 3.6 BIORAD mini protean vertical elektroforez sistemi (http://www.uklabs-
direct.co.uk/html/used_electrophoresis_equipment.html, 2011) ................. 50

xi
Şekil 3.7 SDS-PAGE yönteminde kullanılan protein standartı: Prestained Protein
Molecular Weight Marker 15 kDA- 140 kDA (Fermentas)......................... 52
Şekil 3.8 SDS-PAGE jellerini görüntülemek için kullanılan “Bio-Rad Molecular
imager Gel doc XR” görüntüleme sistemi (http://www.bio-
rad.com/webroot/web/images/lsr/products/imaging_bioinformatics/produ
ct_detail/global/lsr_chemidoc_xrs_system.jpg, 2011)................................. 53
Şekil 4.1 Üç farklı DNA Ekstraksiyon yöntemi kullanılarak L. boissieri’nin
OPA07 primeri ile amplifikasyonu sonucu elde edilen RAPD bant
desenleri. ...................................................................................................... 55
Şekil 4.2 Total DNA elektroforezi (birey no ve isimleri Çizelge 3.4 te verilmiştir).. 57
Şekil 4.3 PCR 1 programı uygulanarak iki Lathyrus türünün iki farklı primer, 5
farklı magnezyum klorür (MgCl2) konsantrasyonu ile gerçekleştirilen
RAPD sonuçları. .......................................................................................... 58
Şekil 4.4 PCR 2 programı uygulanarak iki Lathyrus türünün iki farklı primer, 5
farklı magnezyum klorür (MgCl2) konsantrasyonu ile gerçekleştirilen
RAPD sonuçları ........................................................................................... 59
Şekil 4.5 OPA-01 primeriyle elde edilen bant desenleri (M: DNA Marker) .............. 61
Şekil 4.6 OPA-02 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 62
Şekil 4.7 OPA-03 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 63
Şekil 4.8 OPA-04 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 64
Şekil 4.9 OPA-05 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 65
Şekil 4.10 OPA-06 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 66
Şekil 4.11 OPA-07 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 67
Şekil 4.12 OPA-08 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 68
Şekil 4.13 OPA-09 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 69
Şekil 4.14 OPA-10 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 70
Şekil 4.15 OPA-11 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 71
Şekil 4.16 OPA-15 primeriyle elde edilen bant desenleri(M: DNA Marker) ............... 72
Şekil 4.17 OPB-05 primeriyle elde edilen bant desenleri(M: DNA Marker)................ 73
Şekil 4.18 OPB-10 primeriyle elde edilen bant desenleri (M: DNA Marker)............... 74
Şekil 4.19 OPB-13 primeriyle elde edilen bant desenleri (M: DNA Marker)............... 75
Şekil 4.20 OPB-17 primeriyle elde edilen bant desenleri (M: DNA Marker)............... 76

xii
Şekil 4.21 OPD-03 primeriyle elde edilen bant desenleri (M: DNA Marker) .............. 77
Şekil 4.22 OPD-05 primeriyle elde edilen bant desenleri (M: DNA Marker) .............. 78
Şekil 4.23 OPD-10 primeriyle elde edilen bant desenleri (M: DNA Marker) .............. 79
Şekil 4.24 OPD-11 primeriyle elde edilen bant desenleri (M: DNA Marker) .............. 80
Şekil 4.25 OPD-20 primeriyle elde edilen bant desenleri (M: DNA Marker) .............. 81
Şekil 4.26 OPJ-09 primeriyle elde edilen bant desenleri (M: DNA Marker)................ 82
Şekil 4.27 OPJ-12 primeriyle elde edilen bant desenleri (M: DNA Marker)................ 83
Şekil 4.28 OPJ-14 primeriyle elde edilen bant desenleri (M: DNA Marker)................ 84
Şekil 4.29 OPJ-19 primeriyle elde edilen bant desenleri (M: DNA Marker)................ 85
Şekil 4.31 RAPD verileriyle elde edilen genetik mesafe değerleriyle oluşturulmuş
UPGMA dendrogram (Nei 1978)................................................................. 89
Şekil 4.32 L. annuus popülasyonlarının genetik mesafe verilerine göre (Nei, 1978)
çıkarılan dendrogramı .................................................................................. 92
Şekil 4.33 L. sativus popülasyonlarının genetik mesafe verilerine göre (Nei, 1978)
çıkarılan dendrogramı .................................................................................. 95
Şekil 4.34 Spot test: 1. L. cassius, 2.3.4. L. annuus, 5.6.7. L. cicera, 8. L.
trachycarpus, 9. L. pseudo cicera, 10. L. gorgoni var gorgoni, 11. L.
chrysanthus, 12. 13. L. sativus, 14. L. chloranthus, 15. L. hirsitus, 16. L.
boissieri, 17. Vicia sativa ............................................................................ 97
Şekil 4.35 SDS-PAGE sonucu elde edilen L. cassius, L.annuus, L. cicera, L.
trachycarpus, L. pseudo cicera, tohum depo proteinlerine ait bant
desenleri ....................................................................................................... 99
Şekil 4.36 SDS-PAGE sonucu elde edilen L. gorgoni var gorgoni, L. chrysanthus,
L. sativus, L. chrysanthus, L. hirsitus, L. boissieri, Vicia sativa tohum
depo proteinlerine ait bant desenleri ............................................................ 99
Şekil 4.37 SDS-PAGE verileriyle elde edilen genetik mesafe değerleriyle
oluşturulmuş UPGMA dendrogram (Nei 1978)......................................... 101

xiii
ÇİZELGELER DİZİNİ
Çizelge 1.1 Lathyrus L. cinsinin tarihsel olarak değişik araştırmacılar tarafından
yapılan seksiyonel sınıflandırmaları (Kenicer vd. 2005) ........................ 2
Çizelge 3.1 RAPD yöntemi için kullanılan materyaller, popülasyon no, birey
sayısı, lökaliteler ................................................................................... 33
Çizelge 3.2 SDS-PAGE yöntemi için kullanılan materyal (Tohumlar) ................... 34
Çizelge 3.3 RAPD ve SDS-PAGE için kullanılan dış grup materyali ..................... 34
Çizelge 3.4 RAPD çalışılan materyallerin birey sıra no’ları ve adları (gri kutular,
analizlerden çıkarılan tür ve popülasyonları temsil etmektedir ............ 36
Çizelge 3.5 Çalışmada kullanılan primerler, dizileri, Tm (Temperature melting-
Çözünme sıcaklığı), GC içerikleri ........................................................ 43
Çizelge 3.6 Kaya ve Neale (1995) tarafından uygulanan PZR döngüsü.................. 44
Çizelge 3.7 Optimizasyonda denenen PZR2 döngüsü (Croft vd. 1999) .................. 44
Çizelge 3.8 Çalışmada kullanılan PZR1 döngüsü .................................................... 44
Çizelge 3.9 Optimizasyon: İki farklı PZR döngüsünde, iki farklı türün, iki farklı
primer ile 5 farklı MgCl2 konsantrasyonu denemesi, (K: Kontrol) ...... 46
Çizelge 4.1 Çalışmada kullanılan örneklerden elde edilen DNA’ların
konsantrasyonu ve saflığını veren nanodrop spektrofotometre
sonuçları ................................................................................................ 56
Çizelge 4.2 Çalışmada kullanılan primerler ile yapılan RAPD-PZR sonucunda
elde edilen DNA parçalarının (bantlarının) özellikleri ......................... 60
Çizelge 4.3 Çalışmada kullanılan türlerin RAPD verilerine göre genetik mesafe
değerleri (Nei 1978) .............................................................................. 89
Çizelge 4.4 L. annuus popülasyonları arası genetik varyasyon analizleri (Nei
1987) ..................................................................................................... 92
Çizelge 4.5 L. annuus’un popülasyonları için genetik farklılaşma analizi (Nei
1987) ..................................................................................................... 92
Çizelge 4.6 L. annuus popülasyonlarının genetik mesafe değerleri (Nei 1978) ...... 92
Çizelge 4.7 L. cicera popülasyonlarının genetik varyasyon analizleri (Nei 1987) . 93
Çizelge 4.8 L. cicera’nın populasyonları için genetik farklılaşma analizi (Nei
1987....................................................................................................... 93

xiv
Çizelge 4.9 L. cicera popülasyonlarının genetik mesafe değerleri (Nei 1978)........ 94
Çizelge 4.10 L. sativus popülasyonlarının genetik varyasyon analizleri (Nei 1987) . 94
Çizelge 4.11 L. sativus’un populasyonları için genetik farklılaşma analizi
(Nei 1987) ............................................................................................. 95
Çizelge 4.12 L. sativus popülasyonlarının genetik mesafe değerleri (Nei, 1978)...... 95
Çizelge 4.13 L. cassius popülasyonlarının genetik varyasyon analizleri (Nei 1987). 96
Çizelge 4.14 L. cassius’un populasyonları için genetik farklılaşma analizi (Nei
1987) ..................................................................................................... 96
Çizelge 4.15 L. cassius popülasyonlarının genetik mesafe değerleri (Nei, 1978) ..... 96
Çizelge 4.16 SDS PAGE verilerine göre elde edilen genetik mesafe değerleri (Nei
1978) ................................................................................................... 100

1
1. GİRİŞ Halk arasında mürdümük, burçak, külür gibi adlarla bilinen Lathyrus L. cinsi, Fabaceae
(Leguminosae, Baklagiller) familyasından ekonomik önem taşıyan bitkilerdir. Dünya
genelinde, yeşil ot, kuru ot veya taze yem olarak hayvan beslenmesinde, yeşil gübre
bitkisi olarak toprak yapısının iyileştirilmesinde, yemeklik tane baklagil veya sebze
olarak insan beslenmesinde ya da süs bitkisi olarak bahçelerde kullanılır Dünyada en
fazla tarımı yapılan Lathyrus türleri; L. sativus, L.cicera ve L. ochrus’dur. Bunların
arasında da L. sativus öne çıkar (Kenicer vd. 2005, Başaran vd. 2007).
Tarımsal üretimin dışında doğal olarak da yeryüzünde yayılım gösterir. Fabaceae
(Leguminosae, Baklagiller) familyasının Fabeae (syn. Viceae) oymağının en geniş
cinsidir. Bu oymağın diğer üyeleri gibi hemen hemen tamamen ılıman iklimde yetişen
bir cinstir. Lathyrus üyeleri Kuzey Yarımküre boyunca yayılım göstermekle birlikte bir
grup da Güney Amerika’da görülür. Cinsin asıl yayılım gösterdiği bölge, özellikle
türlerin üçte birini barındıran Türkiye’nin Doğu Akdeniz bölgesi başta olmak üzere,
Batı Avrasya’dır. İkinci bir merkez Kuzey ve Güney Amerika ile Doğu Asya’da
bulunur (Davis 1970, Kenicer 2008). Dünyada yaklaşık 187 türle temsil edilen
Lathyrus cinsinin, Türkiye’de 22 tanesi endemik olmak üzere 68 taksonu bulunmaktadır
(Davis 1970, Allkin vd. 1985, TUBİVES http://wwweski.tubitak.gov.tr/tubives/ 2011).
Lathyrus cinsinin sistematiğiyle ilgili bugüne kadar pek çok çalışma yapılmıştır
(Çizelge 1.1). İlk olarak bitki sistematiğinin ilk yazılı eseri olan “De Historia
Plantarum” adlı eserinde Theophratus (M.Ö. 370-285) habitus özelliklerine göre
sınıflandırdığı 480 bitki arasında, şu anda Lathyrus olarak bilinen cinsi Lathyros olarak
adlandırmıştır. Daha sonra Linneaus (1753) binominal adlandırmada cins için
Lathyros’tan türettiği Lathyrus ismini kullanmıştır (Kenicer 2008). Sonra Godron
(1848) Lathyrus ve Orobus olarak bilinen cinsleri, stiluslarının sırt kısmındaki tüylülük
ile karakterize ederek Lathyrus adı altında toplamıştır. Orobus’u Lathyrus cinsinin bir
seksiyonu olarak düşünmüştür. Boisser (1872) Orobus seksiyonunun sülüksüz üyelerini
Orobus cinsinde birleştirmiş, sülüklü olanları da, Lathyrus içinde Orobastrum
seksiyonunda toplamıştır. Bässler (1966) ise Orobus seksiyonunun çok yıllık türlerini

2
Lathyrus cinsi içinde Orobus altcinsinde toplamış, Lathyrus cinsinin diğer gruplarını
çalışmamıştır. Aynı araştırmacı Orobus ve Lathyrostylis seksiyonlarının Avrasya
türlerinin revizyonunu yapmıştır (Bässler 1973, 1981). Davis (1970), altcins şeklinde
ayrımı kabul etmemiş ve cinsi 10 seksiyona ayırmıştır. Czefronova (1971), Lathyrus
cinsini 6 altcinse ayırmıştır. Kupicha (1983) ise cinsin seksiyonal düzeyde iyi
yapılandığını ifade ederek, cinsi 13 seksiyona ayırmıştır. Genellikle cinsin seksiyonlara
ayrımında son dört yazar benzer bir sınıflamayı kabul etmiştir. Ama Kupicha (1983),
Orobastrum seksiyonunu 3 küçük seksiyona ayırmış, fakat Lathyrus ve Cicercula
seksiyonlarını birleştirmiştir. Amerikan türlerini ise Notolathyrus seksiyonunda
toplamıştır.
Çizelge 1.1 Lathyrus L. cinsinin tarihsel olarak değişik araştırmacılar tarafından yapılan seksiyonel sınıflandırmaları (Asmussen ve Liston 1998, Kenicer vd. 2005)
Taranmış kutular araştırmacılar tarafından ele alınmamış tür gruplarını temsil etmektedir. Boissier (1872) Orobus Kutusunu cins olarak kabul etmektedir. Kesik çizgiler ise şüpheli ayrımları göstermektedir

3
Doğan vd. (1992) Türkiye Lathyrus türlerini nümerik taksonomik yöntemlerle
sınıflandırmışlardır. 54 tür üzerinde yapılan çalışmadan elde edilen fenogramda, cinsin
iki alt cins ve 9 seksiyon şeklinde ayrılabileceğini önermişlerdir. Bu sonuçlar Davis
(1970) ve Kupicha’nın (1983) yaptığı, sınıflamadan oldukça farklıdır. Kupicha’nın
birleştirdiği Lathyrus seksiyonunun Doğan vd. (1992) tarafından 5 ayrı gruba ayrılması
önerilmiştir. Ertekin (1991) tarafından, Güneydoğu Anadolu Bölgesinin Lathyrus türlerinin morfolojik ve anatomik özellikleri üzerine yapılan çalışmada, Lathyrus ve
Cicercula seksiyonlarının bazı üyelerinin ayrı bir seksiyona ayrılabileceği belirtilmiştir.
Yapılan bu çalışmalarda görüldüğü gibi, Lathyrus cinsinin cins altı, tür ve tür altı
kategorilere ayrılmasında bir takım taksonomik problemler vardır. Bu problemlerin
temelinde ise morfolojik homoplazi (ortak atadan gelmeyip, farklı türlerde bağımsız
evrim sonucu ortaya çıkan benzer özellikler) olduğundan sık sık bahsedilmektedir
(Barneby ve Reveal 1971).
Sınıflandırma yapılırken araştırmacıların ortak atadan gelen (homolog) özellikleri analiz
etmeleri ve bu ortak türemiş özellikleri (sinapomorfi) tanımlamaları gerekir. Ancak
böyle benzer özelliklerin farklı grup türlerde tamamen bağımsız olarak evrimleşmesi ve
araştırmacıların bunları homolog özellikler olarak yanlış yorumlaması gibi sebepler
sınıflandırma sırasında hatalar yapılmasına yol açmaktadır. Bu gibi hatalardan
kaçınmanın en etkili yolu evrimsel akrabalıkları oluştururken sadece bir veya birkaç
özellik yerine, çok sayıda özelliği analiz etmektir (Freeman ve Herron 2009).
Bireyler veya populasyonlar arasındaki varyasyon analizi morfolojik özelliklere dayalı
olarak yapılabildiği gibi, moleküler özelliklere dayandırılarak da yapılabilmektedir (Jain
vd. 1983). Bu sebepten 1960’lı yıllara kadar morfolojik veya davranış özelliklerinin incelenmesiyle veri toplanırken 1960’lı yıllardan itibaren moleküler özellikler evrimsel
ve sistematik çalışmalarda giderek önem kazanmışlardır. Bahsedilen moleküler
özelliklerin tespiti için kullanılan kaynak ise; proteinler ve nükleer genom, nükleer
ribozomal DNA, mitokondri genomu, kloroplast genomu gibi makro moleküllerdir.
Sistematik çalışmalarda proteinlerden veri sağlamak için genellikle elektroforetik
teknikler kullanılır. Bu tekniklerde kullanılan protein formları ise çoğunlukla,

4
izoenzimler, allozimler ve tohum depo proteinleridir. Sistematik çalışmalarda veri
toplamak için DNA kullanıldığında ise yararlanılan teknikler; DNA-DNA
hibridizasyonu, restriksiyon enzim analizi, klonlama, dizi analizi ve PZR (polimeraz
zincir reaksiyonu) tabanlı tekniklerdir (Onarıcı ve Sümer 2003). Bu şekilde moleküler analizlere dayalı olarak yapılan sınıflandırmalar moleküler sistematik olarak
adlandırılmaktadır.
Bu güne kadar Lathyrus cinsiyle ilgili pek çok moleküler filogenetik çalışma
yayınlanmıştır. Asmussen ve Liston (1998), Croft vd. (1999), Chtourou-Ghorbel vd.
(2001), Badr vd. (2002), Ben Brahim vd. (2002), Kenicer vd. (2005) gibi. Bu çalışmalar
içinde en kapsamlı olanı Asmussen and Liston’un (1998) çalışması ile Kenicer vd.
(2005)’nin çalışmasıdır. Asmussen ve Liston (1998) RFLP yöntemini kullanarak
Lathyrus cinsinin, kloroplast DNA’larının analizi ile yeni bir sınıflandırma ortaya
koymuşlardır. Yine Kenicer vd. (2005) nükleer DNA’daki bazı internal transcribed
spacer (ITS) ların ve kloroplast DNA’sının trnL-F ve trnS-G bölgelerinin dizi analizi ile
tekrar bir sınıflandırma yapmışlardır. Diğer çalışmalar çoğunlukla ekonomik öneme
sahip olan Lathyrus türleriyle yapılan sınırlı biocoğrafyadaki taksonomik
incelemelerdir. Bu moleküler çalışmalarda kullanılan yöntemler ise DNA dizi analizi,
RFLP (Restriction Fragment Length Polymorphism), RAPD (Random Amplified
Polymorfic DNA), AFLP (amplified fragment length polymorfism) olmuştur. Ghorbel
vd. (2002), RAPD yöntemiyle, Lathyrus cinsinin Tunus, Etiyopya, Portekiz, Fransa,
Macaristan’da yayılış gösteren 5 türü arasındaki, türlerin populasyonları arasındaki ve
popülasyon içindeki (bireyler arasındaki) genetik mesafelere göre akrabalıkları
incelemişlerdir. Araştırmacılar, RAPD çalışmalarının genellikle çok sayıda polimorfik
lokus ortaya çıkardığını ve kendi çalışmalarının da bu bulgularla uyumlu olduğunu
belirtmişlerdir. Ayrıca RAPD çalışmalarının farklı bölgelerdeki populasyonlar
arasındaki genetik çeşitliliğin incelenmesinde ve genetik ilişkilerin belirlenmesinde
oldukça elverişli olduğunu bildirmişlerdir.
Türkiye’de de konuyla ilgili moleküler çalışmalar yapılmıştır: Emre vd.’nin (2006), Cicercula dışındaki 4 farklı seksiyona ait, bazı Lathyrus türleri arasında filogenetik
ilişkilerin ortaya konmasında tohum proteinleriyle yapılmış SDS-PAGE (Sodyum

5
Dodesil Sülfat- PolyAcrylamid Gel Electrophoresis) çalışmasında, araştırıcılar RAPD
çalışmalarının daha ileri düzeyde çözümler sunacağını bildirmişlerdir. Yine aynı
araştırmacılar (Emre vd. 2007, Emre vd. 2010) SDS-PAGE yöntemiyle, Cicercula
seksiyonunun bazı türlerinin, tohum proteinlerinin analizini yapmışlardır. Türkiye’de
Lathyrus cinsine ait DNA seviyesinde bir çalışma bulunmamaktadır .
Moleküler sistematik çalışmalarında DNA’dan veri toplamak için kullanılan pek çok
yöntemden biri de PZR tabanlı bir teknik olan RAPD (random amplified polymorphic
DNA=Rastgele çoğaltılmış polimorfik DNA) tekniğidir. RAPD hemen hemen sınırsız
sayıda DNA belirteci (marker) ortaya koyar (Williams vd. 1990). Aslında izoenzim
elektroforezi ve RFLP tekniklerine alternatif olarak geliştirilmiştir (Waugh ve Powell
1992). Teknik, çok sayıda polimorfik lokus ortaya koyar. Oldukça hızlıdır. Basit bir
tekniktir. DNA hibridizasyonu ve daha yüksek teknik aktiviteler konusunda uzmanlık
gerektirmez. Maliyeti de daha karmaşık tekniklere oranla daha düşüktür. Ayrıca,
radyoaktif madde kullanılmaz.
Proteinlerden veri sağlanarak yapılan moleküler sistematik çalışmalarında teknik olarak
elektroforetik teknikler kullanılmaktadır. Oldukça basit ve etkili olan SDS-PAGE bu
tekniklerden biridir. SDS-PAGE tekniği ile tohum depo proteinlerinin analizi
sonucunda elde edilen veriler, türler arası evrimsel ilişkilerin belirlenmesinde oldukça
kullanışlıdır (Smartt 1990, Przybylska vd. 1999).
Bu çalışmanın amacı Türkiye’deki Lathyrus cinsi Cicercula seksiyonunun bazı
türlerinin DNA ve tohum depo proteinleri varyasyonlarını kullanarak, aralarındaki
evrimsel ilişkileri incelemek ve cinsin taksonomik problemlerinin çözümüne katkı
sağlamaktır. Bu amaçla DNA varyasyon analizi için RAPD, tohum depo proteinleri
varyasyon analizi için SDS-PAGE teknikleri kullanılmıştır. Elde edilen veriler,
POPGENE Microsoft Window-based Freeware for Population Genetics Analysis,
Version 1.31 (Yeh vd. 1997) ve MEGA (Molecular Evolutionary Genetics Analysis)
Versiyon 3.0 (Kumar vd. 2004) bilgisayar programları aracılığıyla istatistiksel olarak
değerlendirilmiş ve filogenetik dendrogramlar çıkarılmıştır.

6
2. KURAMSAL TEMELLER 2.1 Lathyrus L. sistematiği
Alem: Phytae
Bölüm: Spermatophyta
Altbölüm: Angiospermae
Sınıf: Magnoliopsida
Altsınıf: Rosidae
Takım: Fabales
Familya: Leguminosae (Fabaceae)
Altfamilya: Papilionoideae
Cins: Lathyrus L.
Seksiyon: Cicercula (Davis 1970)
Türler:
1. Lathyrus annuus L.
2. Lathyrus hierosolymitanus Boiss. .
3. Lathyrus cassius Boiss.
4. Lathyrus gorgoni Parl. var. gorgoni Parl.
5. Lathyrus gorgoni Parl. var. pilosus C. C. Townsend
6. Lathyrus pseudo-cicera Pamp.
7. Lathyrus cicera L.
8. Lathyrus sativus L.
9. Lathyrus blepharicarpus Boiss.
10. Lathyrus marmoratus Boiss. et Bl.
11. Lathyrus stenophyllus Boiss. et Heldr.
12. Lathyrus lycicus Boiss.
13.Lathyrus phasellitanus Hub.-Mor. et Davis
14. Lathyrus hirsutus L.
15. Lathyrus chrysanthus Boiss.
16. Lathyrus chloranthus Boiss.
17. Lathyrus trachycarpus (Boiss.) Boiss

7
2.1.1 Familya: Leguminosae [Fabaceae (Baklagiller)]
Leguminosae isminin, Latincede toplamak anlamına gelen “legere”den türetildiği
düşünülmektedir. Çünkü çoğunlukla bu familya üyelerinin meyveleri el ile toplanır.
Çoğu Leguminosae üyesi, köklerinde atmosferik azotu bağlama yeteneğine sahip
bakteriler (Rhizobium sp.) içeren nodüllere (yumrucuk) sahiptir. Bakterilerin
atmosferden tespit ettikleri bu azotu bitkiler kullanabilmektedir. Böylece toprak azotuna
ihtiyaç duymamaktadırlar. Bu faaliyet konak bitkinin bütün gelişme evresi boyunca
bakteriler tarafından sürdürülür. Tespit edilen azot uzun süre bitkide kalır. Bu şekildeki
bitkiden, sürülüp toprağa gömülerek, otlatılarak ya da toplanarak yararlanılabilir. Bu
özellikleri baklagillere, “organik tarım”ın en önemli dayanaklarından biri olma yeteneği
vermektedir (Elçi 1988).
Otsu, çalı ya da ağaç formundadırlar. Genellikle dik gövdelidirler. Gövde veya
yapraklarıyla tırmanıcı ya da diğer bitkilere sarılıcı olabilirler. Stipullar (kulakçık)
Acacia’da olduğu gibi geniş veya dikenli olabilir. Genellikle alternat (sarmal) diziliş
gösteren yapraklar, morfolojik olarak çoğunlukla bileşik tiptedir. Daha çok rasemoz
(salkım) tipte olan çiçek durumları (infloresans) oldukça değişkendir. Genellikle
hermafrodit (erkek ve dişi organları bir arada) olan çiçeklerin simetrisi düzenli
(Mimosoideae’de) ya da düzensiz (Papilionoideae’de ve Caesalpinioideae’de) olabilir.
Genelde 5’ten az ya da çok olan kaliks’i (çanak) birleşik sepaller oluşturur. Genellikle 5
serbest ya da birleşik petal korollayı (taç) oluşturur. 10 ya da daha çok sayıdaki stamen
(erkek organ) bir tüp içinde birleşik (monodelphus), veya üst stamen serbest
(diadelphus), ya da bütün hepsi serbest durumda bulunur. 2 veya daha çok sayıdaki üst
durumlu (superior) ovaryumun içerdiği ovül (tohum taslağı), anatrop veya
kampilotropdur. Bakla (legüm) şeklindeki meyve çoğunlukla düz ve uzundur, ancak
taneler arasında boğumlanmış veya bölünmüş de olabilir. Medicago gibi, bazı türlerde
kangal şeklinde görülen meyve spiral şekilde bükülmüştür. Tohumlar bazen
endospermsiz bazen de az endospermlidir (Hickey ve King 1997).

8
Pisum (bezelye), Cicer (nohut), Vicia (bakla), Phaseolus (fasülye), Arachis (yerfıstığı),
Lens (mercimek), Vigna (börülce) gibi yenilebilir tohumları olan birçok cinse sahip olan
familya, insan beslenmesinde, dolayısıyla da ekonomik yönden oldukça önemlidir.
Bununla birlikte kereste, lif, boya, reçine, bitkisel yağ eldesinde ve hayvan yemi
(Trifolium, Medicago, Vicia) olarak da kullanılırlar. Acacia (akasya), Cytisus
(katırtırnağı), Cercis (erguvan) gibi cinsler ise süs bitkisi olarak bahçelerde yer alırlar.
Diğer yandan, azot yönünden toprağı zenginleştirirler ve bu sebeple yeşil gübre olarak
da kullanılırlar.
Dünya çapında yayılım gösteren baklagiller bitkiler aleminin en kalabalık
familyalarından biridir. 657 cinse ait 16400 tür içerir ve Mimosoideae,
Caesalpinioideae ve Papilionoideae olarak 3 alt familyaya ayrılır. En çok tür
Papilionoideae alt familyasında bulunur (Hickey ve King 1997).
2.1.1.1 Altfamilya: Papilionoideae
Genellikle düzensiz olan çiçeklerde en üstteki büyük petal, korollanın diğer 4 petalini
saracak şekilde katlanır. 2 yan petal kanatçıkları temsil ederken, son 2 küçük petal
birbirine yapışık olarak çiçeğin altındaki “kayıkcık (carina)”ı oluşturur. Kayıkcığı
meydana getiren petallerin kenarları çoğunlukla birleşiktir. Bu petaller bazen, dişicik
(pistil) etrafında, bir oluk halinde, 10 stameni içine alır. Yaprakları trifoliolat, dijitat,
pennat ya da basit yapraktır (Elçi 1988).
Cins: Lathyrus L. (Mürdümük)
Lathyrus L. cinsi dünyada 187 civarında tür ve alttürle temsil edilirken, Türkiye’de 68
takson (22 tanesi endemik) yetişmektedir (Davis 1970, Allkin vd. 1985, TUBİVES
http://wwweski.tubitak.gov.tr/tubives/ 2011). Cins, özellikle Akdeniz ve Doğu Anadolu bölgelerinde çok sayıda endemik türe sahiptir. Bu bölgeler dışında diğer yörelerimizde
de birkaç endemik tür dağınık bir şekilde bulunur. Türkiye Florası kayıtlarına göre
Lathyrus cinsine ait 33 taksonun tip örneği Türkiye’den toplanmıştır (Davis 1970).

9
Lathyrus cinsi, Fabaceae üyelerinin gen merkezi olarak kabul edilen ülkemizin Doğu ve
Güneydoğu Anadolu Bölgesi’nde geniş bir yayılış göstermekle birlikte yurdumuzun
hemen hemen tüm bölgelerinde yetişmektedir. Lathyrus cinsi ülkemizde tür, alt tür ve
varyete seviyesinde 68 takson içermekte olup, 22 tanesi endemik olan bu 68 takson, 10
seksiyonda toplanmıştır (Davis 1970). L. odoratus L. süs bitkisi olarak yetiştirildiği için
Flora of Turkey (Davis 1970)’de gösterilen seksiyonlara dahil edilmemiştir. Son
yıllarda yapılan çalışmalarla bu sayıya 4 yeni takson daha eklenmiştir (Davis 1988;
Maxted ve Goyder 1988; Ertekin ve Saya 1996; Liston ve Shmida 1987). Avrupa
florasında ise 54 tür kayıtlıdır (Tutin vd. 2001).
Bütün Lathyrus türleri otsu bitkilerdir. Çok yıllık veya tek yıllık olabilirler. Tek yıllık
türlerin büyük çoğunluğu narin bitkilerdir. Çok yıllık türlerin kökleri rizom veya yumru
şeklindedir. Yanlarda kanatlı ya da kanatsız olabilen gövde, dik tırmanıcı veya sürünücü
olabilir. Ekseni bir arista (kılçık) veya bir sülükle sonlanan yapraklar, paripinnat veya
subdijitattır. Genellikle 1 çift olan yaprakçık sayısı, 8 çifte kadar çıkabildiği gibi bazen
de hiç olmayabilir. Yaprakçıksız olma sebebi, yaprak ekseninin gelişerek fillodik yaprak
şeklini alması veya yaprakların basit sülüklere indirgenmiş olmasıdır. Çoğunlukla
petiyol bulunan yapraklar hipostomatik ya da amfistomatiktir. Kenarları nadiren dişli,
çoğunlukla düz olan stipullar (kulakçık) ok başı, yarı ok başı, yapraksı, yumurta veya
kıl şeklinde olabilir. Çiçekler aksillar (yaprak koltuğundan çıkan), rasemoz (salkım) ya
da soliter (tek çiçek) olabilir. Kaliks düzenli ya da düzensiz, korolla ise değişik
renklerde olabilir. Meyveler şerit halinde yanlardan basık, tipik baklagil (legümen) dir.
2 veya daha çok tohum içerir. Tohumlar silindirik küre biçimli ya da basık ve köşelidir.
Hilum ovaldir. Tohum yeşil, kahverengi, siyah gibi değişik renklerde olabilir (Davis
1970, Ertekin 1991)
Dünyada en fazla tarımı yapılan Lathyrus türü L. sativus’tur (Şekil 2.1). L. sativus
erken dönemde su altında kalmaya geç dönemde de kuraklığa dayanıklıdır. Geniş bir
toprak ve iklim koşulları aralığında rahatça yetişebilir. L. cicera da (Şekil 2.2)
soğuklara karşı dirençlidir . Bu özellikleri sebebiyle baklagil ıslahı açısından önemli
genetik kaynaklardır (Tekele-Haimanot vd. 1990, Clulow vd. 1991, Kumar 1997).

10
a b
Şekil 2.1 L. sativus: a. Çiçek, b. Meyve (http://de.wikipedia.org/wiki/Datei:Lathyrus_sativus_002.JPG, 2011)
a b
Şekil 2.2 L. cicera: a. Çiçek, b. Meyve (http://www.menudanatura.com/2011/05/Lathyrus-cicera-l.html,2011)
Lathyrus türlerinde beslenme için olumsuz etkili bazı maddeler bulunmaktadır (Urga
vd. 1995). Bunun sebebi yapılarındaki 3 adet protein yapısında olmayan toksik amino asitlerdir. Bunlar:
ODAP (β-N-oxalyl-L, α-βdiamino propionik asid)
DABA (L-2,4 diaminobutirik asid)
BAPN (β-aminopropionitril)’dir.

11
Lathyrus türlerini tüketen insan ve hayvanlarda bu kimyasal maddelere bağlı hastalıklar
görülmekte ve bu hastalıklara genel olarak “lathyrism” adı verilmektedir. Bitki
ıslahçıları cinsdeki bu zararlı maddelerin yok edilmesi veya azaltılmasına yönelik
çalışmalar yapmaktadırlar (Başaran vd. 2007).
Seksiyon: Cicercula
Üst kısımları kanatlı ve sürünücü-tırmanıcı gövdeye sahip üyeler tek yıllıktır. Yapraklar
hipostomatik ya da amfistomatiktir. Alt yapraklar mukronat (L. trachycarpus’un tüm
yaprakları mukronat), üst ve orta yapraklar 3-4 parçalı, sülüklüdür. Çiçekler değişik
renklerde olabilir. Çiçek durumu (infloresans) bir veya birkaç çiçekli olup, stilus
genellikle burulmuştur. Meyve tüylü ya da tüysüz, üst kısımları 2 kanatlı ya da
kanatsızdır. 17 taksonu vardır (Davis 1970, Ertekin 1991).
2.2 Moleküler Sistematik
M.Ö. 3. yüzyılda Aristo tarafından başlatılan organizmaların çeşitliliğini sınıflandırma
çalışmaları sistematik olarak adlandırılır. Klasik sınıflandırma çalışmaları morfolojik
benzerliklere bağlı olarak organizmaların gruplanması şeklindedir. Bu şekildeki
benzerlik analizleriyle oluşturulan sınıflandırma metotları fenetik yaklaşımlar adını
alır. Sınıflandırmadaki diğer yaklaşım şekli ise evrimsel akrabalık temeline dayalıdır ve
filogenetik ya da kladistik yaklaşım adını alır. Darwin’in “Türlerin Kökeni” adlı
kitabının yayınlanmasından sonra bu yaklaşım ortaya çıkmıştır. Basit olarak evrimsel
akrabalık ilişkilerinin ortaya çıkarılmasındaki mantık; “en fazla ortak özelliğe sahip olan
taksonlar en yakın akrabalardır”, yaklaşımıdır (Freeman ve Herron 2009).
Türlerin filogenetik ilişkilerinin belirlenmesinde genetik belirteçlerin benzerliğinden
yararlanılır. Morfolojik belirteçler, protein belirteçleri ve DNA belirteçleri, genetik
belirteçleri oluşturur.
Morfolojik belirteçler çiçek rengi, yaprak tipi, gibi görülebilen özelliklerdir. Morfolojik
belirteçlerin filogenetik çalışmalar için olumlu yönleri arasında, sayıca az olmaları,

12
analizlerinin kolay olması, dominant olmaları, sayılabilir. Olumsuz yönleri ise, çevresel
faktörlerden etkilenebilmeleri, epistatik (bir karakterin ortaya çıkmasından sorumlu olan
farklı genler arasında baskılayıcı etkilerin olması durumu) veya pleiotropik (bir genin
birden fazla fenotipik karakter üzerindeki etkisi) etkilere maruz kalabilmeleri
mutasyonlarla meydana gelmiş olabilmeleridir (Parmaksız 2004, Özcan vd. 2004).
Protein olarak depo proteinleri ya da enzim proteinleri kullanılır. Depo proteinlerinin
olumlu özellikleri analizlerinin çabuk, güvenilir ve tekrarlanabilir olması, olumsuz yönü
ise sayıca çok az olmalarıdır. Enzim proteinlerinin de analizleri çabuk ucuz ve
güvenlidir. Ayrıca çevreden etkilenmezler. Ancak sayıları çok azdır ve bazı enzimler
sadece özel dokularda ve belli bir gelişme döneminde ortaya çıkarlar (Özcan vd. 2004).
DNA molekülleri olarak ise çoğunlukla nükleer genom, nükleer ribozomal DNA,
mitokondri genomu, kloroplast genomu kullanılır. DNA molekülleri çevre şartlarından
etkilenmezler stabildirler. Tüm dokulardan elde edilebilirler, kodominant ya da
dominant özellikte olabilirler, kalıtımı basit ilkelere sahiptir, analizler bitki gelişiminin
herhangi bir evresinde yapılabilir. Tüm genom boyunca sınırsız sayıda varyasyon
gösterebilirler. Ayrıca tüm genom analizi için gereken DNA’yı elde etmek için az
miktarda bitki dokusu yeterlidir. Bitkilerin herhangi bir kısmı DNA ekstraksiyonu için
kullanılabilir (Özcan vd. 2004).
Filogenetik çalışmaları, 1960’lara kadar morfolojik belirteçlere göre yapılmıştır
(Onarıcı ve Sümer 2003). Bugünkü modern sistematikte morfolojik ve anatomik
belirteçlere ek olarak, palinolojik, embriyolojik, sitogenetik, fitokimyasal ve moleküler
belirteçler de kullanılmaktadır. Protein belirteçleri ve DNA belirteçleri, moleküler
belirteçler olarak da adlandırılırlar ve bu belirteçler baz alınarak yapılan filogenetik
çalışmalara moleküler sistematik denir (Özcan vd. 2004).
Moleküler mi yoksa morfolojik özelliklerin mi sistematik çalışmalarında daha iyi veri
kaynağı olduğu tartışmaları günümüzde hala sürmektedir (Freeman ve Herron 2009).
Ancak yapılan çalışmalardan görüldüğü kadarıyla tek birine yönelik yaklaşımdan ziyade
hem moleküler hem de morfolojik ve hatta diğer embriyolojik, palinolojik, sitogenetik

13
ve fitokimyasal özelliklerden elde edilen verilerin bir kaçının bir arada
değerlendirilmesi, biyolojik farklılıkların daha iyi yorumlanmasını ve açıklanmasını
sağlamaktadır (Onarıcı ve Sümer 2003).
2.3 Moleküler Belirteçler
2.3.1 Proteinler
Bir hücrenin kuru ağırlığının yarısından çoğunu oluşturan ve bütün hayati
fonksiyonların gerçekleşmesinde görev alan biyolojik makromoleküller olan proteinler
amino asitlerin belli bir sırada dizilmesi ve bunların daha sonra değişik üç boyutlu
şekillerde katlanmalarıyla ortaya çıkarlar. Bu amino asit dizilişleri proteinin
fonksiyonuyla ilişkilidir (Özcan vd. 2004). Amino asit dizilerinin nasıl olacağı ise
genler tarafından kontrol edilir ve bu kontrol genetik kod ile sağlanır. Dolayısıyla
proteinler genlerin üçüncü kuşak kopyası olduklarından genetik bilgilere çok
yakınlardır. Bu sebeple filogenetik çalışmalarda, başka bir deyişle moleküler
sistematikte genotipler arası farkların gösterilmesinde kullanılırlar (Sammour 1991).
Bitkilere yönelik moleküler sistematik çalışmalarında genellikle kullanılan tohum depo
proteinleri ve enzimler olmak üzere iki tip protein vardır.
2.3.1.1 Tohum depo proteinleri
Önceleri Millerd (1975) tarafından, “metabolik proteinler” ve “depo proteinleri” olarak
iki kategoriye ayrılan tohum proteinleri daha sonraları, Fukushima (1991) tarafından,
“yapısal proteinler ve biyolojik olarak aktif proteinler” diye sınıflandırılmıştır. Biyolojik
olarak aktif olan proteinler, risin, lektinler, enzimler ve enzim inhibitörleridir. Bunların
amino asit kompozisyonları depo proteinlerinden daha dengelidir ve besin değeri olarak
çok önem taşımazlar, tersine hayvanların yememesi için tohumlarda bulunan koruyucu
proteinler olduğu düşünülmektedir (Mandal ve Mandal 2000).
İleride metabolizmanın aktif olduğu bir devrede kullanılmak üzere, gelişimi sırasında
tohumda biriktirilen proteinler ise “depo proteinleri”dir (Derbyshire vd. 1976). Bir zarla
çevrili veziküller veya protein cisimcikleri halinde bulunurlar. Çimlenme esnasında,

14
büyümekte olan bitki tarafından yeni azot bileşikleri sentezlemek amacıyla, azot
kaynağı olarak kullanılırlar. Enzimatik olmayıp, çimlenme ve yeni bitki oluşumu
sırasında azot ve kükürt kaynağı olarak destek sağlarlar.
İnsan ve hayvan beslenmesinin temel elemanlarından olan bitki depo proteinlerinin bazı
esansiyel amino asit içerikleri oldukça düşüktür. Örneğin, baklagillerde “methionin”
eksikliği varken tahıllarda da “treonin, triptofan” ve özellikle “lizin” bakımından
yetersizlik söz konusudur. Oysa bu amino asitler insan metabolizması tarafından
sentezlenemez ve dışarıdan alınmalıdır. Sürekli vejeteryan olarak beslenen insanlar,
özellikle çocuklar, bu gibi amino asit eksikliklerine maruz kalırlar ve bu da onlar da geri
dönüşü olmayan fiziksel ve zihinsel hasarlara neden olabilir. Aynı durum hayvanlar için
de geçerlidir. Bu sebeple bitki genetiği üzerine yapılan çalışmalarda, ürünlerin amino
asit kompozisyonlarının geliştirilmesi de hedeflenmektedir (Heldt 1999).
Derbyshire vd. (1976)’ne göre, depo proteinleri ile ilgili, detaylı ilk çalışma, Osborne ve
Campbell (1898)’ın proteinlerin çözünürlüklerine göre deneysel olarak gerçekleştirdiği
ve Osborne’un 1924’te basılmış olan “The Vegetable Proteins” kitabında yayınladığı
çalışmasıdır. Osborne depo proteinlerini; suda çözünenler (Albuminler), tuz çözeltisinde
çözünenler (Globulinler), su-alkol karışımında çözünenler (Prolaminler) ve seyreltik asit
veya bazda ya da seyreltik SDS çözeltisinde çözünenler (Glutelinler) diye ayırmıştır.
Daha sonraki çalışmalarda, glutelin ve prolaminlerin yapısal olarak birbirleriyle
bağlantılı oldukları belirlenmiş ve glutelinler, prolamin grubunun bir üyesi olarak kabul
edilmiştir (Heldt 1999).
Monokotiledonların (tahıllar gibi) depo proteinleri prolamin ve glutelinlerden
oluşurken, dikotiledonların (baklagiller gibi) başlıca proteinleri albumin ve
globulinlerdir (Derbyshire vd. 1976). Depo proteinleri bazı karakteristik özellikleriyle
diğer proteinlerden ayırt edilebilir. Bu karakteristik özellikler; tohum gelişiminin ara
devresinde fazla miktarlarda biriktirilmeleri ve çimlenme sırasında kullanılmaları,
sadece tohumda sentezlenmeleri, depolanma dışında başka bir fonksiyonel aktiviteye
sahip olmamaları ve protein cisimcikleri veya aleuron tanecikleri şeklinde
saklanmalarıdır (Mandal ve Mandal 2000).

15
“Leguminosae” türlerinin depo proteinleri kotiledonlarda bulunur ve globulinlerdir.
Çökme katsayısına göre globülinler, legumin (11S) ve visilin (7S) olarak ikiye ayrılır.
Her iki globülini de çoklu genler kodlar. “Legüminosa”daki legüminlerde “methionin”
ve “sistein” amino asiti eksikliği vardır (Shewry vd. 1995).
Baklagil tohumlarının asıl depo proteini olan legumindir. Örneğin bakla (Vicia
faba)’nın, toplam depo proteinlerinin %75’ini teşkil eder. Molekül ağırlığı 300-400 kD
olup, bir heksamerdir. İki farklı peptid zincirinden meydana gelir (α, β monomerleri) ve
bunlar disülfid (S-S) köprüsüyle birbirlerine bağlanır. Küçük olan β-zinciri 20 kD,
büyük olan α-zinciri ise 35-40 kD, molekül ağırlığına sahiptir.
Amino asit dizileri bakımından legumine benzerlik gösteren visilin ise genellikle tek
peptid zincirinden meydana gelen monomerlerin oluşturduğu trimerlerdir. Sistein
eksikliği sebebiyle visilin monomerleri disülfid (S-S) köprüleri kuramaz. Moleküler
ağırlığı 150-190 kD olan visilinler leguminlerin tersine çoğu zaman glikozilatlıdır;
yapılarında mannoz, glukoz ve N-asetilglukozamin barındırırlar (Heldt 1999).
2.3.1.2 Enzimler
Enzimler kimyasal reaksiyonların hızını artıran (katalizör) biyolojik makromoleküllerdir
ve hemen hemen tüm enzimler protein yapısındadır (Smith vd. 1997).
Filogenetik çalışmalarda genellikle izoenzim (izozim) veya alloenzim (allozim)’lerden
yararlanılır. İzoenzim farklı genler tarafından üretilen aynı reaksiyonu katalizleyen
enzimlerdir. Alloenzimler ise aynı genin farklı allelleri tarafından meydana getirilirler.
Farklı genler veya alleller tarafından meydana getirildiklerinden dolayı farklı yapıdaki
protein molekülleridir. Bu yapı farklılıkları sebebiyle, sistematik ve filogenetik
çalışmalarda kullanılabilen veriler sunarlar (Carey ve Ganders 1987, Max vd. 1999).

16
2.3.1.3 Protein izolasyonu
Proteinler doğal ortamlarında genellikle başka moleküllerle birlikte ve onlarla daima
etkileşim halinde bulunurlar. Ayrıca birçok farklı işlevden sorumlu, enzimler, membran
proteinleri, reseptörler, antikorlar, aktin, miyozin, flagellin, hormonlar gibi binlerce
farklı tipte protein molekülü vardır. Bunun yanı sıra proteinler hücrenin içinde farklı
yerlerde veya hücrenin dışında bulunabilir. Bu sebeple protein moleküllerini
inceleyebilmek için onları bulundukları yerden çıkarmak ve diğer moleküllerden
ayırarak izole etmek (yalıtmak) gerekir (Arda ve Ertan 2004).
Protein, ekstrasellüler (hücre dışına salınan) ise, ekstraksiyonu için kullanılacak yöntem
kolaydır. Protein salındıktan sonra hücreleri ortamdan uzaklaştırmak (örneğin
santrifüjleme ile) yeterlidir (Arda ve Ertan 2004).
Protein sitoplazmik ya da membrana bağlı ise yöntem uzun ve karmaşıktır. Özellikle
bitki hücrelerinde öncelikle hücre çeperinin parçalanması ve içeriğin serbest kalması
sağlanmalıdır. Bunun için porselen havan, milli homojenizatör veya blender gibi
mekanik olarak parçalamayı sağlayacak aletler kullanılır (Arda ve Ertan 2004).
Protein bir organel içindeyse öncelikle o organelin sitoplazmik proteinlerden
uzaklaştırılması gerekir. Bunun için partiküllerin sedimantasyon (çökelme) derecelerine
göre çöktüğü, giderek artan hızlarda ardışık (diferansiyel) santrifüjleme tekniği
kullanılır (Arda ve Ertan 2004).
Tohum depo proteinlerinin ekstraksiyonu için uygun ekstraksiyon tamponuyla mekanik
parçalama yapılır. Daha sonra yapılacak santrifüj işlemi sonrasında tüpteki berrak
kısımda (supernatan) proteinler çözünmüş halde bulunur (Arda ve Ertan 2004) .

17
2.3.2 DNA
DNA bazı virüsler ve diğer bütün canlıların hayati işlevleri ve biyolojik gelişimleri için
gerekli olan kalıtsal bilgiyi taşıyan nükleik asit molekülüdür. Her hücre içinde bulunan
DNA molekülü kromozomlarda paketlenmiştir. Çift sarmal yapıda organize olmuş olan
nükleotidlerden oluşur. Bir nükleotid ise; bir pentoz şeker (deoksiriboz), bir fosfat
grubu, bir azotlu baz’dan (pürin ya da pirimidin) meydana gelir. Pürin bazlar Adenin
(A) ya da guanin (G), pirimidin bazlar ise sitozin (C) ya da timin (T) olabilir.
DNA molekülünün çift sarmal yapısı, dört temel yapı taşının (A,G,C,T) bir araya
gelmesinden oluşan iki zincir içerir. İki zincirden oluşan sarmal, her biri bir şeker fosfat
omurgalı, adenin-timin (iki hidrojen bağı) ve sitozin-guanin (üç hidrojen bağı) bazları
arasındaki, zayıf hidrojen bağlarıyla bir araya gelmişlerdir (Şekil 2.3)
Şekil 2.3 DNA Molekülü (http://whyfiles.org/034clone/images/dna_molecule.gif, 2011)
A ile T ve C ile G’nin şekli birbirini tamamlayıcıdır ve DNA kendisini bu sayede
kopyalayabilir. Bu bazların zincir boyunca sıralanışı (sekansı) organizmanın
fonksiyonlarının kusursuz bir şekilde yürümesini sağlayan genetik talimatları taşıyan
genetik kodu meydana getirir (Şekil 2.4).

18
Şekil 2.4 DNA’daki baz dizilimi (http://www.webhatti.com/saglik/60365-dna-nedir-nerede-bulunur.html, 2011)
Prokaryotik hücreler genellikle dairesel şekilde çift sarmallı bir DNA molekülünü
sitoplazmalarında içerir. Ökaryotik hücrelerde ise DNA molekülüne bazı proteinlerin
(histon ve histon olmayan proteinler) ve bazı RNA’ların ilavesiyle, DNA’nın kısaltılıp
paketlenmesi sonucu kromozomlar oluşur. Kromozomlar hücre çekirdeğinde bulunur
(Gözükara 1994).
Sitoplazmik DNA: Ökaryotik hücrelerde çok az miktarda DNA çekirdek dışında,
sitoplazmada, kloroplastlarda (cpDNA) ve mitokondride (mtDNA) bulunur.
Kloroplastlar ve mitokondrilerin her biri kendi benzersiz kromozomlarına sahiptir. Bu
genler organellere ait olan tRNA, mRNA ve birkaç proteini kodlar ve organizmanın
fenotipinin ekspresyonunda önemli role sahiptir (Gözükara 1994).
Organel DNA’sı daima olmasa da, genellikle dişi ebeveyn tarafından kalıtılır ve bu
maternal (anneye ait) kalıtım olarak bilinir. cpDNA ve mtDNA dizileri çekirdek
DNA’sından farklıdır. Bitki mtDNA’sı gen düzenlemesi konusunda hızlı bir evrimsel
gelişim ortaya koyarken, nükleotid dizilerinin değişimine dayalı mutasyon birikimi
yavaş kalmaktadır. Bu yavaşlığın sebebi, tam olarak anlaşılamamıştır ancak, oldukça
hatasız DNA replikasyon sistemine veya oldukça etkili DNA hatalarını düzeltme
mekanizmalarına sahip olmalarından kaynaklanıyor olabilir. Buna karşın, cpDNA’sının
evrim hızı, hem genlerin yeniden düzenlenmesi hem de nükleotid dizilerinin değişimine
dayalı mutasyon birikimi bakımından, genellikle yavaştır (de Vicente ve Fulton 2003).
Farklı olaylar, DNA sekansında değişik varyantların ortaya çıkmasına neden olabilir.
Bu varyantlar genellikle moleküler polimorfizm olarak tanımlanır ve genotipteki,

19
uygun bir teknikle araştırıldığında farklı bant desenleri ile gözlemlenebilir. Bu durum
fenotipe yansıyabilir de yansımayabilir de. Moleküler polimorfizmi ortaya çıkaran
birkaç olay aşağıda kısaca açıklanmaktadır:
• Nokta mutasyonlar
• İnsersiyonlar ya da delesyonlar
• Yeni düzenlenmeler
2.3.2.1 Nokta mutasyonlar
Nokta mutasyonlar aynı yerdeki sadece bir baz ya da birkaç bazın yer değiştirmesiyle
ortaya çıkar. Şekil 2.5’de orjinal DNA dizisinin dört bazı (üstteki), farklı dört bazla yer
değiştirmektedir. Bazların orijinal sayısı değişmediği için, dizinin toplam uzunluğu da
değişmez.
Şekil 2.5 Nokta mutasyon (de Vicente ve Fulton 2003)
2.3.2.2 İnsersiyonlar ve delesyonlar
DNA dizisine bir veya birkaç baz ilave olursa bu olaya insersiyon, DNA dizisinden bir
veya birkaç baz çıkarsa bu olaya da delesyon denir. Bu durumda DNA molekülünün
uzunluğu da değişir. Şekil 2.6’da bir DNA dizisine beş adet yeni bazın eklenmesiyle
gerçekleşen bir insersiyon durumu görülmektedir. Burada DNA parçasının boyu diziye
eklenen baz sayısına göre artar.

20
Şekil 2.6 İnsersiyon (de Vicente ve Fulton 2003)
Şekil 2.7’de bir DNA dizisinden dört adet bazın çıkmasıyla gerçekleşen bir delesyon
durumu görülmektedir. Burada DNA parçasının boyu diziden ayrılan baz sayısına göre
azalır.
Şekil 2.7 Delesyon (de Vicente ve Fulton 2003)
2.3.2.3 Yeniden düzenlenmeler
DNA dizisindeki değişiklikler, yeni düzenlenmeler (örneğin; bir parçanın tamamen
kopması) vasıtasıyla da ortaya çıkabilir. Kromozomal yeniden düzenlenmeler genetik
rekombinasyon (Krossing over) ya da yer değiştirebilen genetik parçaların
(transpozonlar) insersiyonu ile gerçekleşir. Molekül uzunluğu değişebilir de
değişmeyebilir de (Şekil 2.8), (de Vicente ve Fulton 2003).
Şekil 2.8 Yeniden düzenlenme (de Vicente ve Fulton 2003).

21
DNA molekülündeki bu varyasyonların analizi ile birbirine çok yakın olan kültür
çeşitleri ayrılabilir ve tanımlanabilir. Türlerin filogenetik akrabalıkları incelenerek
taksonomik tanımlamaları yapılabilir (Lowe vd. 1996). Evrimsel akrabalıklarının
araştırılması, ıslah programları ve bağlantı haritalamalarında yararlanılabilir. DNA
varyasyonları, biyokimyasal veya morfolojik varyasyonlardan fazladır. Bu varyasyonlar
incelenerek, çaprazlaması zor olan türlerin kromozom haritaları ortaya konabilir.
Moleküler belirteçler bunlar gibi birçok avantaja sahiptir.
Doğadaki yabani varyeteleri toplayarak moleküler düzeyde tanımlamak, ekonomik
değeri olan çeşitlerin ıslahı açısından oldukça önemlidir (Bothmer vd. 1991). Bu
varyetelerden gen kaynağı olarak yararlanmak ve moleküler yöntemlere dayanarak
istenilen tipleri belirlemek klasik ıslah metotlarındaki zorlukların aşılmasında kolaylık
sağlar.
2.3.2.4 DNA izolasyonu
Genetik materyal olan DNA hücre içinde paketlenmiş olarak ve diğer bazı moleküllerle
birlikte etkileşim halinde bulunur. DNA ile yapılacak moleküler çalışmalar için
öncelikle DNA’nın hücre dışına çıkarılması ve etkileşim halinde olduğu diğer
moleküllerden ayrılarak izole edilmesi (yalıtılması) gerekir. Hücre duvarının
parçalanması mekanik ya da kimyasal yollarla gerçekleştirilebilir. DNA-protein
kompleksinin çözülmesi ve diğer makromoleküllerden ayrılması işlemleri ise enzimatik
ya da kimyasal yöntemlerle gerçekleştirilir.
DNA izole edildikten sonra saflığı ve konsantrasyonunun tespiti gerekir. Bunun için
spektrofotometre ile optik dansite ölçümü yapılır. Nükleotidlerin heterosiklik halkaları
260 nm dalga boyunda en yüksek ultraviyole (UV) absorbsiyonu (optik dansite) yapar.
1 optik dansite (absorbance) 50 µg/mL çift iplikli DNA molekülünü ifade ederken, 40
µg/mL tek iplikli DNA veya RNA molekülünü ifade eder. Buna göre çift iplikli DNA
konsantrasyon tespitinde aşağıdaki formülden yararlanılır:
DNA (μg/ml)=260 nm’deki OD (Absorbans değeri) x sulandırma oranı x 50

22
DNA 260 nm’de en yüksek UV absorbsiyonu yaparken, proteinler 280 nm’de,
karbonhidratlar da 230 nm’de maksimum absorbsiyonu gösterirler. Bu sebeple 260 ve
280 nm dalga boylarında alınan optik dansite değerlerinin arasındaki oran DNA saflğı
hakkında bilgi verir. 280 nm de proteinlerin maksimum absorbsiyon yapmalarından
dolayı, spektrofotometrede 280 nm’deki okuma sırasındaki değer artışı 260/280
oranında düşüşe sebep olur. 260/280 oranının 1,8-2,0 aralığında olması DNA’nın
yeterince saf olduğunu, protein veya diğer moleküllerle kirlenmemiş olduğunu gösterir
1.8’in altında elde edilen Abs (260 /280) değeri protein kontaminasyonunu, 2’nin
üzerinde elde edilen Abs (260/280) değeri de RNA kontaminasyonunu işaret
etmektedir. PZR yönteminde ne kadar saf ve yüksek molekül ağırlıklı DNA izole
edilebilirse bantların belirginliği ve üretilebilirliği o derece artmaktadır.
DNA analizi için kalitatif olarak da agaroz jel elektroforezi tekniği kullanılabilir. Çok
kolay ve basit bir teknik olmasından dolayı rutin olarak laboratuvarlarda çok sık
kullanılan yöntemlerden biridir. Agaroz konsantrasyonu % 0,5 – 1,5 arasında olacak
şekilde hazırlanan jellere yüklenen ve elektroforeze tabi tutulan DNA molekülleri, daha
sonra jellerin etidyum bromürle boyanmaları sonucu ultraviyole ışık altında görünür
hale getirilerek incelenir (Sarıkaya 2004, Parmaksız 2004,
http://www.drzeydanli.com.tr/index.php?page=icerikgoster&menuID=30#DNA, 2011).
2.4 Moleküler Belirteç Analizleri
2.4.1 Protein elektroforezi
Protein moleküllerindeki organizmanın evrimsel tarihinin anlaşılmasını mümkün
kılacak bilgilere ulaşmak elektriksel alanda bulunan jelatinli bir ortamda, yüklü
moleküllerin, farklı şekilde hareket etmeleri prensibine dayanan elektroforez ile
mümkün olabilmektedir (Şekil 2.9). Protein elektroforezi ile depo proteinleri ya da
enzimlerin yapısal değişiklikleri ortaya konabilir ve bu yapısal değişikliklerin sağladığı
veriler genetik bilgilere çok yakındır. Bunun sebebi ise proteinlerin DNA’nın üçüncü
kuşak kopyası olmalarıdır (Sammour 1991). Proteinlerden sağlanan bu verilerin

23
güvenilirliği mRNA ve DNA moleküllerinden düşük olmakla birlikte, enzim ürünleri
veya bunun gibi moleküllerden üstündür (Zuckerkandl ve Pauling 1965).
Şekil 2.9 Protein elektroforezi (http://en.wikipedia.org/wiki/SDS-PAGE, 2011)
Proteinlerin bu özellikleri, kolay ekstrakte edilmeleri ve jel elektroforezinin proteinler
arasındaki farklılıkları ayırt edebilme gücü fenotipik karakterleri karşılaştırmak için
kullanılabilecek iyi bir kombinasyon oluşturur. Bu sebeple elektroforez genetik
kaynakların belirlenmesinde, genotipler arası farkların ortaya konmasında güçlü ve ucuz
tekniklerdir. Gelişmekte olan ülkelerde DNA veya RNA seviyelerindeki
çalışmalardansa protein elektroforezinin tercih edilmesi daha avantajlıdır. Ayrıca bu
yöntemle; çeşitlerin saflığını belirlemek, çeşitler arası teşhis yapmak, taksonomik
analizlere yardımcı olmak, türlerin filogenisini ortaya koymak, genetik çeşitlilik
hakkındaki bilgileri arttırarak evrime destek sağlamak konularında da yardımcı olur
(Peirce ve Brewbaker 1973, Gorman ve Kiang 1977, Cardy ve Beversdorf 1984,
Hamrick vd. 1991, Sammour 1991). Bunun yanısıra protein veya enzim elektroforezi
çalışmaları, çevresel strese dayanıklılık, bitki veriminin çoğaltılması, çaprazlama
belirlenmesi ve kontrolü, klonlama gibi önemli araştırma konularında da oldukça fazla
kullanılmaktadır (Tanksley ve Orton 1983, van Geyt ve Smed 1984).
Değişik şekillerdeki elektroforez metotlarıyla, bir çok bitkinin tohum ya da yaprak
proteinlerinin veya enzimlerinin analizi yapılmıştır. Bu çalışmalarla yukarıda bahsedilen
çeşitlerin saflığını göstermek, çeşitler arası tanımlama yapmak, taksonomik analizlere
yardımcı olmak, türlerin filogenisini çıkarmak, genetik çeşitlilik bilgilerini arttırarak
evrime destek sağlamak için değerli bilgiler elde edilmiştir (Cooke 1984).

24
2.4.2 DNA varyasyon analizleri
DNA analizleri, çekirdek DNA’sı ve mitokondri ya da kloroplastlarda bulunan organel
DNA’sı kullanılarak yapılabilir. Tüm yüksek yapılı organizmaların (ökaryotlar)
genomlarındaki bir veya daha fazla özelliğin karakterizasyonu elektroforez sonucu
DNA bantlarının ortaya koyulması ile olur. Farklı büyüklükte ve farklı sayıdaki DNA
bantlarının analizi her birey için spesifiktir. Bu bantların ortaya koyulması ile her birey
için DNA parmak izi elde edilmiş olur.
DNA ekstraksiyonu yapıldıktan sonra farklı DNA analiz tekniklerinden biri ile spesifik
DNA parmak izleri elde edilir. Bireylerin DNA parmak izleri birbiriyle karşılaştırılarak
bireyler arasındaki benzerlikler ya da farklılıklar belirlenir. Karşılaştırma için aşağıdaki
işlemler gerçekleştirilir:
1. Varyasyonların tespit edilmesi. Bantların varlığına (1), yokluğuna (0) atanarak
veriler oluşturulur (Şekil 2.10.a).
2. Analizi yapılan üniteler arasındaki ilişkilerin hesaplanması. Bu işlem,
çalışmadaki bütün bireyler ya da populasyonlar arasındaki genetik mesafenin
hesaplanmasını gerektirir. Bunun için, elde edilen veriler matematiksel
formüllerde yerlerine konarak hesaplamalar yapılır (Şekil 2.10.b).
3. Eldeki herhangi bir sınıflandırma yöntemiyle bu ilişkilerin ifade edilmesi.
Günümüzde bu işlemler değişik bilgisayar programları aracılığıyla
gerçekleştirilir. Elde edilen veriler programlara girilerek bireyler veya
populasyonlar arasındaki ilişkiler, kümeleme dendrogramlarıyla görsel hale
getirilir (Şekil 2.10.c), (de Vicente vd. 2004).

25
a b
c
Şekil 2.10.a. Varyasyon tespiti, b. İlişkilerin hesaplanması, c. Dendrogram (de Vicente vd. 2004)
2.4.2.1 Restriksiyon enzim analizi
RFLP (Restriction fragment length polymorphisms) (Kesilmiş parçaların uzunluk
polimorfizmi): Restriksiyon enzimleri (RE), DNA’yı, çok spesifik olarak, belirli
bölgelerinden keserek, genellikle 1.000-20.000 baz çiftlik parçalar haline getiren
enzimlerdir. Sık olarak kullanılan RE arasında EcoRI, ClaI, HindIII, HinfI sayılabilir.
DNA’nın bu enzimlerden bir veya birkaçı ile kesime uğratıldıktan sonra, agaroz jel
elektroforezine tabi tutulması ve sonra etidyum bromür ile boyanan jelde oluşan DNA
bantlarının yeri ve sayısı kıyaslanarak elde edilen çeşitliliğe “restriction fragment length
polymorphisms (RFLP)” adı verilir. RE kesim bölgesindeki tek baz değişikliğini dahi
tanır. Tek bir bazın yer değiştirerek bir enzimin kesim noktasını değiştirmesi, DNA
parçasının uzunluğunu değiştirerek direkt olarak bir bireyin genotipini temsil eden farklı
bir belirteç ortaya çıkarır. Başka bir deyişle, tanıma yerlerinin baz diziliş değişikliği
farklı genotipler arasındaki polimorfizmi meydana getirir. Fakat bu polimorfizm daha
çok RE’nin tanıdığı iki kesim bölgesi arasına parça eklenmesinden (insersiyon) veya
çıkarılmasından (delesyon) kaynaklanır.
RFLP yöntemi kolaylıkla uygulanabilen duyarlı bir yöntemdir. Ancak enzim seçimi
önemlidir. Çok sayıda ya da çok yakın bantların değerlendirilmesi mümkün olmayabilir.

26
Bu zorluğu aşmak için kıyaslanan bantları azaltmak bir yöntem olabilir. Bu amaçla
enzimlerle kesilen DNA parçaları jelde yürütülür. Takiben jelin üzerine aynı boyutta bir
membran konur. Jeldeki DNA parçalarının membrana geçmesi sağlanır (Southern
blotting). Sonraki basamak hibridizasyon basamağıdır. Hibridizasyonda, DNA’daki
zincirlerden birisinin tamamlayıcısı olan tek sarmallı DNA veya RNA probları
kullanılır. Hibridizasyonu görebilmek için probun radyoaktif maddeler (32P) ya da
radyoaktif olmayan maddeler (biotin-digoksigenin) ile işaretlenmesi gerekir. Sonuçta
sadece probla hibridize olan DNA parçaları görüleceğinden özgüllük artırılmış olur.
RFLP’nin avantajları:
• Değişik araştırıcılar tarafından tekrarlandıklarında aynı sonuçlar alınabilir. Bu
sebeple güvenilir bir tekniktir.
• RFLP belirteçleri kodominanttır. Bu sebeple heterozigot tespitinde ve
karakterizasyonunda kullanılabilir.
• Polimorfizm seviyeleri orta düzeydedir.
RFLP’nin dezavantajları:
• Pahalıya mal olur, uzun zaman alır ayrıca gereken iş gücü fazladır.
• Radyoaktif madde kullanılır.
• İzole edilen DNA oldukça kaliteli olmalıdır (Watson vd. 1992, Özcan vd. 2004).
2.4.2.2 DNA dizi analizi
DNA dizi analizi, sistematik için yeni sayılabilecek bir yaklaşım olmasına rağmen
filogenetik ilişkilerin belirlenmesinde en sık kullanılan moleküler analiz yöntemlerinden
biridir (Alverson vd. 1999).
Başlıca iki DNA dizi analizi yöntemi vardır:
1- Maxam ve Gilbert’in kimyasal kırılma yöntemi (Maxam ve Gilbert 1977).
2- Sanger-Coulson’un zincir sonlanma yöntemi (dideoksi ya da enzimatik yöntem de
denir) (Sanger vd. 1977).

27
Bu ikisinden enzimatik olanı, tehlikeli kimyasallardan uzak ve daha hızlı bir yöntemdir.
Bu sebeple daha yaygın olarak kullanılır. Her iki metodun da genel ilkesi DNA’yı sıralı
olarak 4 set halinde, işaretlenmiş fragmentlere ayırmaktır. Enzim metodunda ilk önce
PZR (polimeraz zincir reaksiyonu) uygulanır. Radyoaktif işaretli primerin 3’ hidroksil
grubu deoksinükleozid trifosfat (dNTP) ile yeni fosfodiester bağı yapmak için
reaksiyona girer. DNA sentezini durdurmak için de dideoksinükleozid trifosfat
(ddNTP) kullanılır. Dideoksinükleozid trifosfat (ddNTP), deoksinükleozid trifosfat
(dNTP)’ın yerini alınca DNA sentezi durur çünkü ddNTP’nin 3’ hidroksil grubu yoktur.
Bu şekilde yeni sentezlenen DNA zincirleri daha tamamlanamadan farklı pozisyonlarda
sonlandırılır. Bu prematüre DNA parçaları, denatüre edici poliakrilamid jel
elektroforezi ile ayrıştırılır. Daha sonra otoradyografi ile görüntülenir. Otoradyografiden
sağlanan merdiven şeklindeki bantlar radyoaktif izotop işaretli tek zincirli DNA
moleküllerinin bantlarıdır. En yakın iki bant arasında tek bir nükleotitlik fark bulunur.
Elektroforezde en hızlı hareket eden bant primere en yakındır yani radyoaktif elemente
en yakın bant en küçük DNA fragmentidir. Bu DNA fragmenti hangi ddNTP ile
oluşturulmuşsa işaretten sonra gelen ilk baz odur. İlk banta en yakın ikinci bant da
ikinci bazı verir. Bu şekilde aşağıdan yukarı doğru hareket hızı azalan bantların hangi
ddNTP ile oluşturulduğu tek tek belirlenir. Sonuçta otoradyografideki bu bant
verilerinden yararlanılarak 5’→3’ yönüne doğru olan baz dizilimi belirlenir.
Günümüzde DNA dizi analizlerini otomatik bir şekilde gerçekleştiren cihazlar
geliştirilmiştir. Bu cihazlar sayesinde birkaç saat içinde binlerce nükleotidin dizisi
çıkarılabilmektedir. Ancak filogenetik çalışmalar için teknik belirlerken yüksek maliyet
gerektirdiği göz önüne alınmalıdır (Onarıcı ve Sümer 2003) .
2.4.2.3 PZR ve PZR bazlı parmak izi teknikleri
Polimeraz zincir reaksiyonu (PZR), in vitro şartlarda, enzimatik yollarla, az miktardaki
DNA kalıbından özgül DNA parçaları çoğaltılmasıdır. Bu teknikle istenen DNA
parçalarının replikasyonu hızlandırılmış bir şekilde gerçekleştirilir. Aynen doğal hücre
bölünmesinde olduğu gibi, replikasyon sürecini taklit ederek bir DNA dizisinin yaklaşık
milyar katını kopyalar. Karmaşık DNA kalıplarından özgül DNA parçalarının sentezini

28
ucuz, basit ve çabuk (birkaç saat içinde) gerçekleştirmesinden dolayı moleküler
biyolojide yaygın olarak kullanılan güçlü tekniklerden biridir (Arnheim ve Erlich 1992,
Onarıcı ve Sümer 2003).
Yakın ilişkili türler, bireyler ve popülasyonlar arasındaki genetik çeşitliliğin
belirlenmesi çalışmaları için değişik PZR tabanlı parmak izi teknikleri geliştirilmiştir.
Örnek olarak, rastgele çoğaltılmış polimorfik DNA (RAPD-Random Amplified
Polymorphic DNA), çoğaltılmış parça uzunluğu polimorfizmi (AFLP-(Amplified
Fragment Length Polymorphism), DNA çoğaltılmış parmakizi (DAF- DNA Amplified
DNA), rastgele primerli polimeraz zincir reaksiyonu (Arbitrary Primed-Polymerase
Chain Reaction - AP-PCR), mikrosatellitler, gibi teknikler verilebilir. Bu teknikler,
moleküler sistematik ya da diğer moleküler biyolojik (genetik haritalama, tanı amaçlı
uygulamalar vb.) çalışmalarda kullanılmak üzere pek çok belirteç (marker) sunar
(Onarıcı ve Sümer 2003).
2.4.2.4 Polimeraz Zincir Reaksiyonu (PZR)
PZR’nin kökeni 1980’lerin başında, Kaliforniya’daki “Cetus” şirketinde yapılan önemli
araştırmalara dayanır.1983 baharında Cetus çalışanlarından Karry Mullis arabasıyla San
Fransisko’dan Mendocino’ya giderken aklına gelen fikir bu gün bütün dünyada
kullanılan modern PZR tekniğinin temelini oluşturur. Bu fikri ile Mullis, Cetus
şirketinden 10 000 dolar ikramiye ve 1993 yılı Nobel Kimya Ödülünü kazanmıştır
(Bartlett ve Stirling 2008).
Modern PZR; DNA’nın kendisini kopyalayabilme yeteneğinden yararlanılarak
gerçekleştirilir ve istenen (hedef) bir DNA bölgesinin deney tüpünde çoğaltılması
(amplifikasyonu) işlemidir. Hedef DNA bölgesinin üstel olarak çoğaltılmasını sağlar.
PZR ile karmaşık DNA kalıplarından spesifik DNA parçaları sentezi bir kaç saat içinde
gerçekleştirilebilir. Tekniğin yaygınlaşmasının başlıca sebeplerinden biri budur.
Öncelikle, çalışma yapılan organizmadan DNA eldesi (ekstraksiyon) gerçekleştirilir.
Elde edilen DNA, kalıp DNA olarak deney tüpü içinde, tampon çözeltisiyle birlikte, ısı

29
düzenleyici (thermo cycler) bir cihazda, nükleotidler (dNTP’ler), primerler (başlatıcı
DNA), DNA polimeraz enzimi, magnezyum klorür ile birlikte reaksiyona girer.
Reaksiyon üç basamaktan oluşur:
1. Denatürasyon: Sarmal ikili DNA zincirleri, yüksek sıcaklığın etkisiyle
birbirinden ayrılır.
2. Bağlanma (Annealing): Primerler (başlatıcı DNA’lar), tek iplikçik halindeki
kalıp DNA’ya spesifik olarak bağlanır.
3. Uzama (Extension): Taq DNA polimeraz primerlerin 3’ ucuna nükleotidleri
ekleyerek hedef DNA sekansının bir kopyasını üretir.
Ardı ardına tekrarlanan bu üç adımla DNA parçaları üstel olarak artar. Bu üstel artış,
bir döngüde sentezlenen ürünün sonraki döngülerde kalıp görevi görmesi sebebiyledir
(Welsh ve McClelland 1990, Blair ve Blair Zajdel 1992).
PZR ile hangi dizide DNA çoğaltılacağı iki faktöre bağlıdır. Birincisi ve en önemlisi
primer dizisidir. Yeteri kadar uzunlukta özgül diziler kullanılırsa bütün genomun özgül
bir bölgesine ait DNA çoğaltılır. Kısa ve rastgele primer kullanılırsa rastgele DNA
çoğaltılır. İkinci faktör, bağlanma basamağının gerçekleştiği sıcaklık derecesidir. 30-40
derecelerdeki sıcaklıklarda primerler bir çok yere rahatça bağlanır ve bir çok özgül
olmayan DNA çoğaltılır. Bağlanma sıcaklığı 55-60 dereceler civarında tutulduğu
zamanlarda ise primerler sadece özgül bölgelere bağlanır ve buradan çoğaltım yapılır.
DNA varyasyonlarından dolayı, değişik genotiplerde aynı primer kullanılsa da farklı
DNA dizileri çoğaltılır ve bu farklılıklar DNA belirteçleri oluştururlar (Özcan vd.
2004).
2.4.2.5 RAPD (Random Amplified Polymorphic DNA)
RAPD (Rastgele Çoğaltılmış Polimorfik DNA) nükleotid dizilimi rastgele seçilmiş
primerlerle gerçekleştirilen PZR işlemidir. Williams vd. (1990) tarafından
geliştirilmiştir. Nükleotid dizi bilgisine sahip olmayı gerektirmez. Böylece genom

30
üzerinde rastgele bölgeler çoğaltılılır. Kullanılan primerlerin boyu yaklaşık 6-10
nükleotid uzunluğundadır. Diğer PZR uygulamalarından farklı olarak iki yerine bir
primer kullanılır ve bu primer her iki taraftan DNA üretimini gerçekleştirir (Şekil 2.11).
Bu primerlerin tasarlanması sırasında GC/AT oranının %50 veya daha büyük olması
önemlidir. Ayrıca önemli olan bir diğer faktör de bu primerlerin kalıp DNA’daki
kendilerine uygun dizilere bağlanabilmelerini kolaylaştıracak düşük bağlanma
sıcaklığıdır. Birçok organizmanın genomunda birbirine ters konumlu, 10 nükleotid
uzunluğunda kısa primerlerin bağlanabileceği bölgeler vardır. Bu şekilde primerler,
aralarında birkaç bin baz çifti uzaklıktaki farklı yerlere ters konumlu bağlanabilirler.
Böylece primerlerin bağlantı bölgeleri arasındaki kısımlar çoğaltılabilir.
Bireyler arasındaki DNA dizilerindeki varyasyonlar sebebiyle herhangi bir bireydeki
primer bağlanma yeri kaybolabilir ya da değişebilir. Bu da bireyler arasında PZR
sonucunda elde edilen ürünlerde farklılıklara yol açar. Bu farklılıklar ise PZR
ürünlerinin jel elektroforezi yapılarak bant varlığı ya da yokluğu şeklinde ortaya konur.
Primer Bağlantı bölgelerinin yok olması veya değişmesi bant ya da bantların da yok
olmasıyla sonuçlanır. PZR ürünlerinin elektroforeziyle elde edilen bireysel bant
profillerinin benzerlikleri ya da farklılıkları (polimorfizm) moleküler sistematik
çalışmalarında kullanılacak verileri oluşturur. Bunun yanısıra veriler türlerin teşhis
edilmesi, tohum saflığının belirlenmesi, adli tıp, hastalıkların tanısı gibi alanlarda da
kullanılır (Arı 2004).
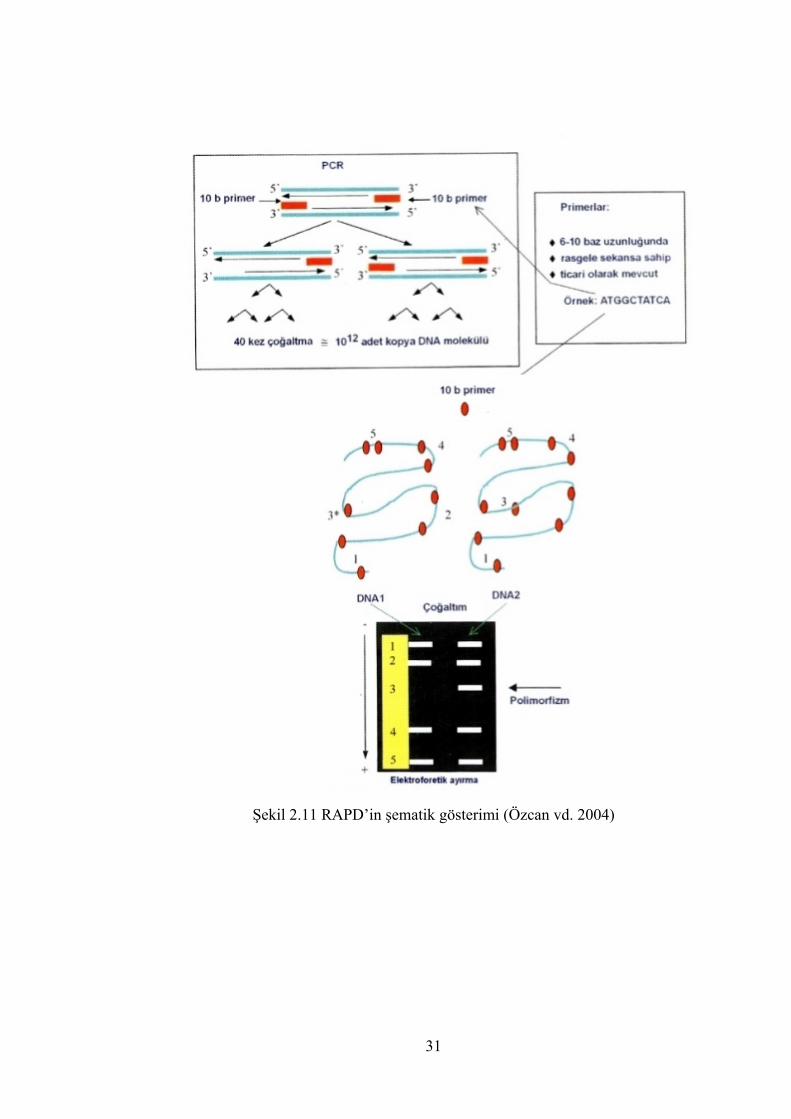
31
Şekil 2.11 RAPD’in şematik gösterimi (Özcan vd. 2004)

32
3. MATERYAL ve METOT Çalışmalar “Ankara Üniversitesi Fen Fakültesi Biyoloji Bölümü Moleküler Sistematik
Laboratuarı”nda gerçekleştirilmiştir.
3.1 Materyal
Çalışma materyali olarak Türkiye’de doğal ortamda yetişen Lathyrus L. cinsi Cicercula
seksiyonuna ait 17 taksondan elde edilebilen 10 taksona ait örnekler kullanılmıştır.
RAPD çalışmalarında bitki örneklerinin kuru veya taze yaprakları, SDS-PAGE
çalışmalarında ise tohumları kullanılmıştır. Kullanılan materyaller çizelge 3.1-3.3’de
verilmiştir. Materyal temini için dört farklı kaynaktan yararlanılmıştır. Toplanma bölge
kareleri şekil 3.1’de, örnek (birey) numaraları da çizelge 3.4’de belirtilmiştir.
1.Prof. Dr. Alaattin Selçuk ERTEKİN’in (Dicle Üniversitesi Fen Fakültesi, Biyoloji
bölümü) koleksiyonu: Materyalin büyük çoğunluğunun sağlandığı kaynaktır. Sağlanan
bireyler kurutulmuş olduğundan bu bireylerin kuru yaprakları çalışmaya alınmıştır.
2.Ege Tarımsal Araştırma Enstitüsü (ETAE) Gen Bankası: Sağlanan tohum materyali
çimlendirilip 15 günlük fidelerin taze yaprakları RAPD çalışmalarında kullanılmak
üzere toplanmıştır.
3.Yrd. Doç. Dr. Nurhan BÜYÜKKARTAL’ın (Ankara Üniversitesi Fen Fakültesi,
Biyoloji bölümü) koleksiyonu: Cins dışı grup olarak kullanılmak üzere temin edilen,
Vicia sativa tohumları: Sağlanan materyal yine tohum olduğundan yine çimlendirme
yoluyla 15 günlük fidelerin taze yaprakları kullanılmıştır.
4.Yrd. Doç. Dr. Neslihan ERDOĞAN’ın (Mehmet Akif Ersoy Üniversitesi Fen Edebiyat
Fakültesi, Biyoloji bölümü) koleksiyonu: Seksiyon dışı grup olarak kullanılmak üzere
temin edilen Lathyrus cilicicus Hayek et Siehe örnekleri. Temin edilen materyal yine
kuru olduğundan bireylerin kuru yaprakları ile çalışılmıştır.
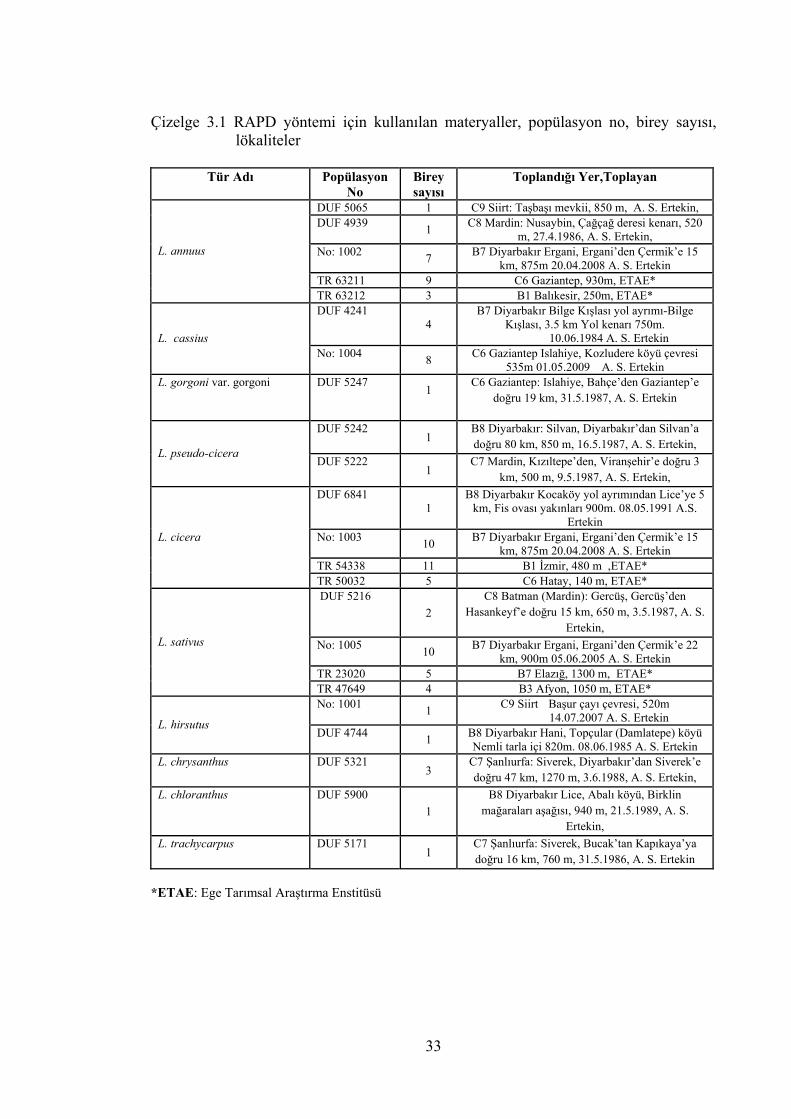
33
Çizelge 3.1 RAPD yöntemi için kullanılan materyaller, popülasyon no, birey sayısı, lökaliteler
Tür Adı Popülasyon
No Birey sayısı
Toplandığı Yer,Toplayan
DUF 5065 1 C9 Siirt: Taşbaşı mevkii, 850 m, A. S. Ertekin, DUF 4939 1 C8 Mardin: Nusaybin, Çağçağ deresi kenarı, 520
m, 27.4.1986, A. S. Ertekin, No: 1002 7 B7 Diyarbakır Ergani, Ergani’den Çermik’e 15
km, 875m 20.04.2008 A. S. Ertekin TR 63211 9 C6 Gaziantep, 930m, ETAE*
L. annuus
TR 63212 3 B1 Balıkesir, 250m, ETAE* DUF 4241
4 B7 Diyarbakır Bilge Kışlası yol ayrımı-Bilge
Kışlası, 3.5 km Yol kenarı 750m. 10.06.1984 A. S. Ertekin L. cassius
No: 1004 8 C6 Gaziantep Islahiye, Kozludere köyü çevresi 535m 01.05.2009 A. S. Ertekin
L. gorgoni var. gorgoni
DUF 5247 1
C6 Gaziantep: Islahiye, Bahçe’den Gaziantep’e doğru 19 km, 31.5.1987, A. S. Ertekin
DUF 5242 1
B8 Diyarbakır: Silvan, Diyarbakır’dan Silvan’a doğru 80 km, 850 m, 16.5.1987, A. S. Ertekin,
L. pseudo-cicera DUF 5222
1 C7 Mardin, Kızıltepe’den, Viranşehir’e doğru 3
km, 500 m, 9.5.1987, A. S. Ertekin, DUF 6841
1 B8 Diyarbakır Kocaköy yol ayrımından Lice’ye 5
km, Fis ovası yakınları 900m. 08.05.1991 A.S. Ertekin
No: 1003 10 B7 Diyarbakır Ergani, Ergani’den Çermik’e 15 km, 875m 20.04.2008 A. S. Ertekin
TR 54338 11 B1 İzmir, 480 m ,ETAE*
L. cicera
TR 50032 5 C6 Hatay, 140 m, ETAE* DUF 5216
2 C8 Batman (Mardin): Gercüş, Gercüş’den
Hasankeyf’e doğru 15 km, 650 m, 3.5.1987, A. S. Ertekin,
No: 1005 10 B7 Diyarbakır Ergani, Ergani’den Çermik’e 22 km, 900m 05.06.2005 A. S. Ertekin
TR 23020 5 B7 Elazığ, 1300 m, ETAE*
L. sativus
TR 47649 4 B3 Afyon, 1050 m, ETAE* No: 1001 1 C9 Siirt Başur çayı çevresi, 520m
14.07.2007 A. S. Ertekin L. hirsutus DUF 4744 1 B8 Diyarbakır Hani, Topçular (Damlatepe) köyü
Nemli tarla içi 820m. 08.06.1985 A. S. Ertekin L. chrysanthus DUF 5321
3 C7 Şanlıurfa: Siverek, Diyarbakır’dan Siverek’e doğru 47 km, 1270 m, 3.6.1988, A. S. Ertekin,
L. chloranthus DUF 5900 1
B8 Diyarbakır Lice, Abalı köyü, Birklin mağaraları aşağısı, 940 m, 21.5.1989, A. S.
Ertekin, L. trachycarpus DUF 5171
1 C7 Şanlıurfa: Siverek, Bucak’tan Kapıkaya’ya doğru 16 km, 760 m, 31.5.1986, A. S. Ertekin
*ETAE: Ege Tarımsal Araştırma Enstitüsü
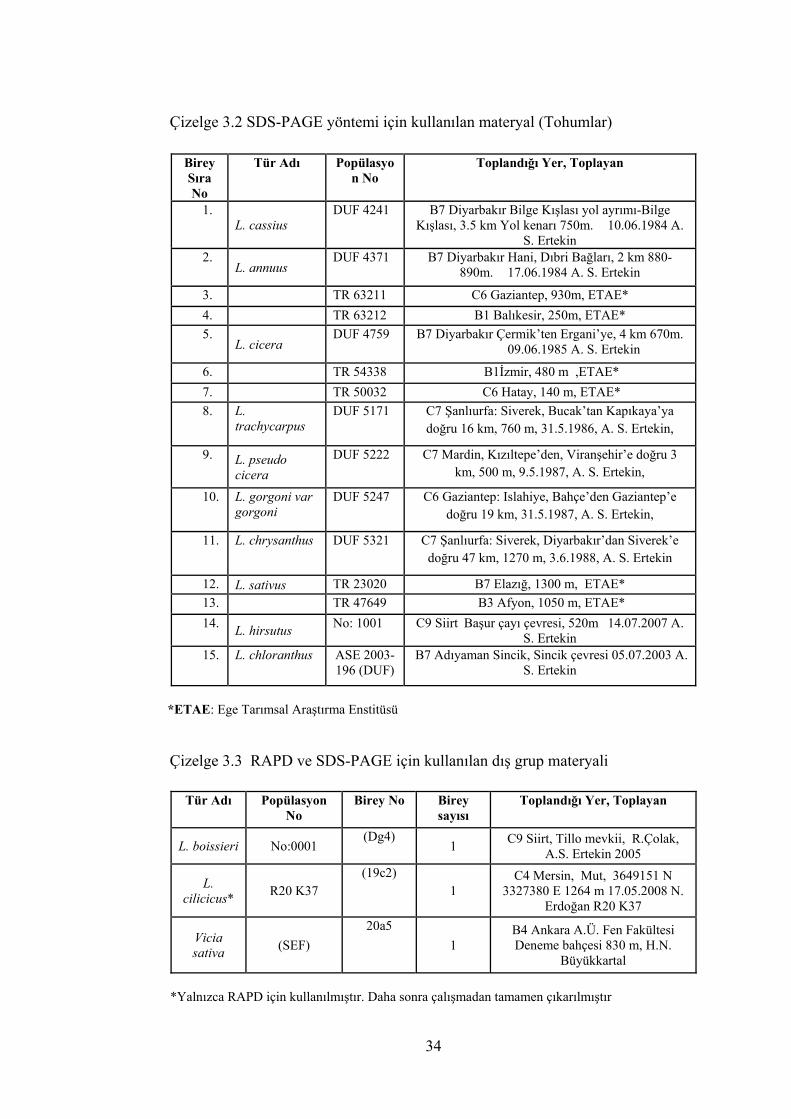
34
Çizelge 3.2 SDS-PAGE yöntemi için kullanılan materyal (Tohumlar)
Birey Sıra No
Tür Adı Popülasyon No
Toplandığı Yer, Toplayan
1. L. cassius
DUF 4241 B7 Diyarbakır Bilge Kışlası yol ayrımı-Bilge Kışlası, 3.5 km Yol kenarı 750m. 10.06.1984 A.
S. Ertekin 2.
L. annuus DUF 4371 B7 Diyarbakır Hani, Dıbri Bağları, 2 km 880-
890m. 17.06.1984 A. S. Ertekin
3. TR 63211 C6 Gaziantep, 930m, ETAE* 4. TR 63212 B1 Balıkesir, 250m, ETAE* 5.
L. cicera DUF 4759 B7 Diyarbakır Çermik’ten Ergani’ye, 4 km 670m.
09.06.1985 A. S. Ertekin
6. TR 54338 B1İzmir, 480 m ,ETAE* 7. TR 50032 C6 Hatay, 140 m, ETAE* 8. L.
trachycarpus DUF 5171 C7 Şanlıurfa: Siverek, Bucak’tan Kapıkaya’ya
doğru 16 km, 760 m, 31.5.1986, A. S. Ertekin,
9. L. pseudo cicera
DUF 5222 C7 Mardin, Kızıltepe’den, Viranşehir’e doğru 3 km, 500 m, 9.5.1987, A. S. Ertekin,
10. L. gorgoni var gorgoni
DUF 5247 C6 Gaziantep: Islahiye, Bahçe’den Gaziantep’e doğru 19 km, 31.5.1987, A. S. Ertekin,
11. L. chrysanthus DUF 5321 C7 Şanlıurfa: Siverek, Diyarbakır’dan Siverek’e doğru 47 km, 1270 m, 3.6.1988, A. S. Ertekin
12. L. sativus TR 23020 B7 Elazığ, 1300 m, ETAE* 13. TR 47649 B3 Afyon, 1050 m, ETAE* 14. L. hirsutus No: 1001 C9 Siirt Başur çayı çevresi, 520m 14.07.2007 A.
S. Ertekin 15. L. chloranthus ASE 2003-
196 (DUF) B7 Adıyaman Sincik, Sincik çevresi 05.07.2003 A.
S. Ertekin
*ETAE: Ege Tarımsal Araştırma Enstitüsü
Çizelge 3.3 RAPD ve SDS-PAGE için kullanılan dış grup materyali
Tür Adı Popülasyon No
Birey No Birey sayısı
Toplandığı Yer, Toplayan
L. boissieri No:0001 (Dg4)
1 C9 Siirt, Tillo mevkii, R.Çolak, A.S. Ertekin 2005
L. cilicicus* R20 K37
(19c2) 1
C4 Mersin, Mut, 3649151 N 3327380 E 1264 m 17.05.2008 N.
Erdoğan R20 K37
Vicia sativa (SEF)
20a5 1
B4 Ankara A.Ü. Fen Fakültesi Deneme bahçesi 830 m, H.N.
Büyükkartal *Yalnızca RAPD için kullanılmıştır. Daha sonra çalışmadan tamamen çıkarılmıştır
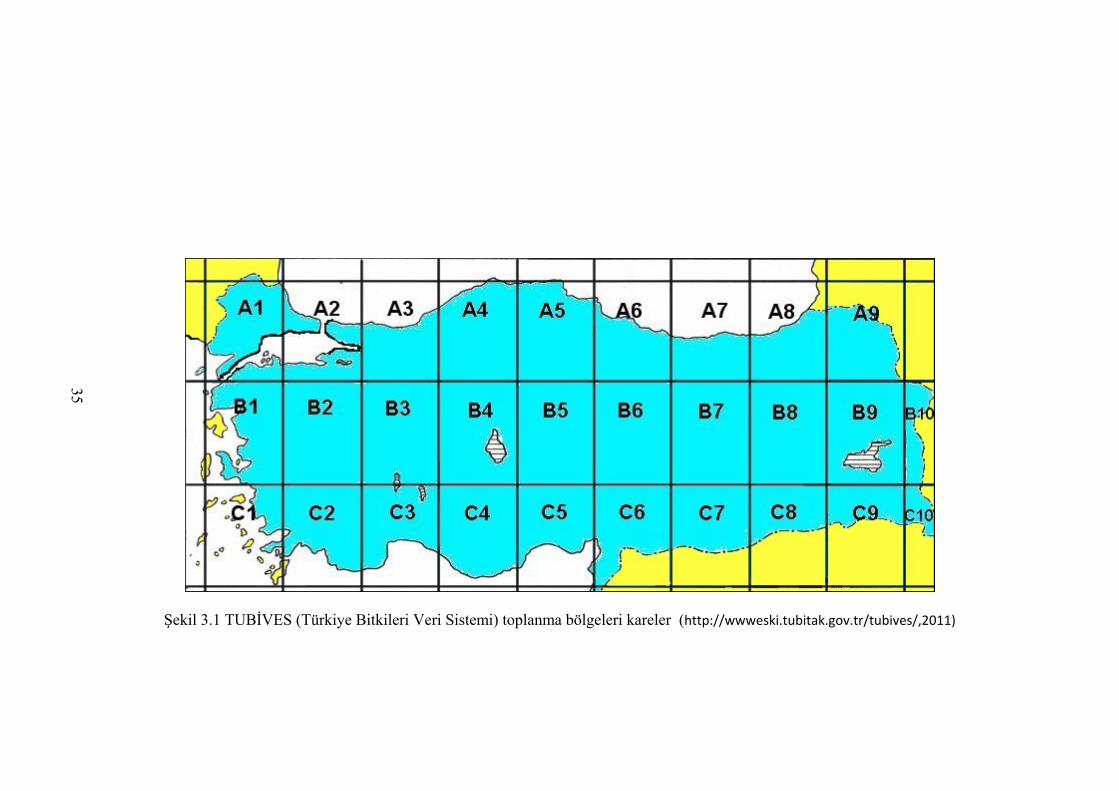
35
Şekil 3.1 TUBİVES (Türkiye Bitkileri Veri Sistemi) toplanma bölgeleri kareler (http://wwweski.tubitak.gov.tr/tubives/,2011)
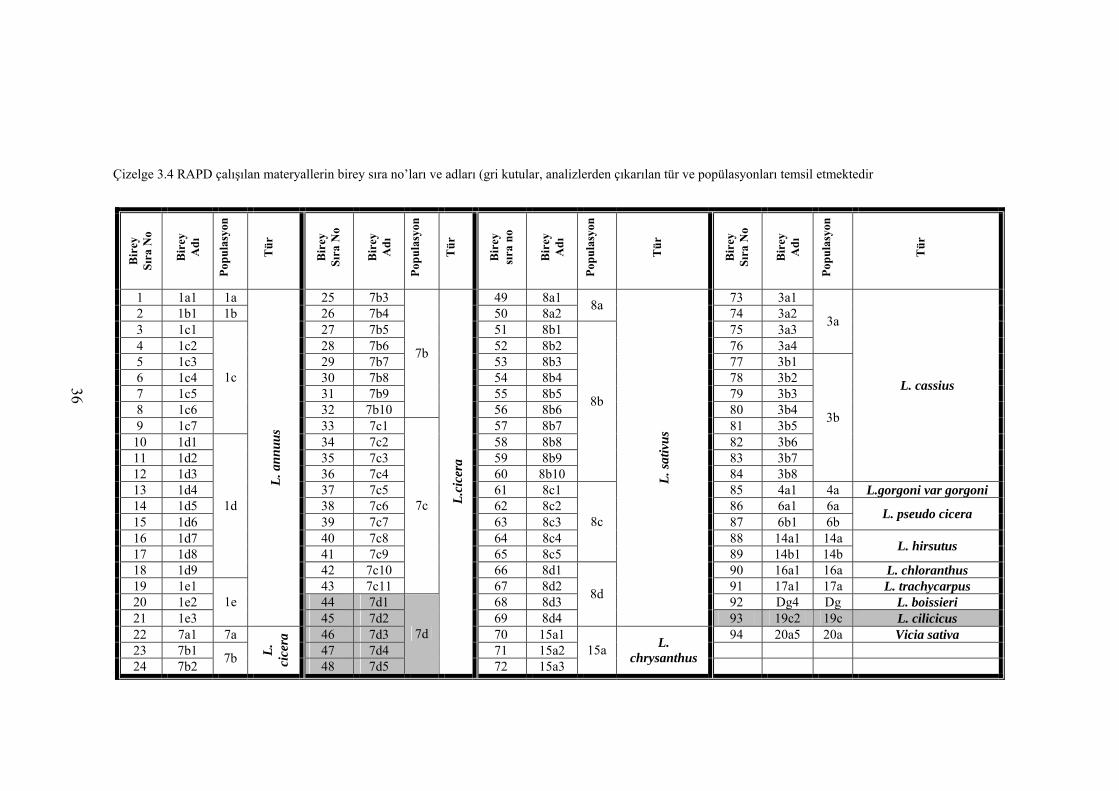
36
Çizelge 3.4 RAPD çalışılan materyallerin birey sıra no’ları ve adları (gri kutular, analizlerden çıkarılan tür ve popülasyonları temsil etmektedir
Bir
ey
Sıra
No
Bir
ey
Adı
Popu
lasy
on
Tür
Bir
ey
Sıra
No
Bir
ey
Adı
Popu
lasy
on
Tür
Bir
ey
sıra
no
Bir
ey
Adı
Popu
lasy
on
Tür
Bir
ey
Sıra
No
Bir
ey
Adı
Popu
lasy
on
Tür
1 1a1 1a 25 7b3 49 8a1 73 3a1 2 1b1 1b 26 7b4 50 8a2 8a 74 3a2 3 1c1 27 7b5 51 8b1 75 3a3 4 1c2 28 7b6 52 8b2 76 3a4
3a
5 1c3 29 7b7 53 8b3 77 3b1 6 1c4 30 7b8 54 8b4 78 3b2 7 1c5 31 7b9 55 8b5 79 3b3 8 1c6 32 7b10
7b
56 8b6 80 3b4 9 1c7
1c
33 7c1 57 8b7 81 3b5 10 1d1 34 7c2 58 8b8 82 3b6 11 1d2 35 7c3 59 8b9 83 3b7 12 1d3 36 7c4 60 8b10
8b
84 3b8
3b
L. cassius
13 1d4 37 7c5 61 8c1 85 4a1 4a L.gorgoni var gorgoni 14 1d5 38 7c6 62 8c2 86 6a1 6a 15 1d6 39 7c7 63 8c3 87 6b1 6b L. pseudo cicera
16 1d7 40 7c8 64 8c4 88 14a1 14a 17 1d8 41 7c9 65 8c5
8c
89 14b1 14b L. hirsutus
18 1d9
1d
42 7c10 66 8d1 90 16a1 16a L. chloranthus 19 1e1 43 7c11
7c
67 8d2 91 17a1 17a L. trachycarpus 20 1e2 44 7d1 68 8d3 92 Dg4 Dg L. boissieri 21 1e3
1e
L. a
nnuu
s
45 7d2 69 8d4
8d
L. sa
tivus
93 19c2 19c L. cilicicus 22 7a1 7a 46 7d3 70 15a1 94 20a5 20a Vicia sativa 23 7b1 47 7d4 71 15a2 24 7b2 7b L.
ci
cera
48 7d5
7d
L.ci
cera
72 15a3 15a L.
chrysanthus

37
3.2 Metot
DNA varyasyon analizi için RAPD (Random amplified polymorphic DNA) tekniği,
depo proteinleri varyasyon analizi için SDS-PAGE (sodyum dodesil sülfat poliakrilamid
jel elektroforezi) tekniği kullanılmıştır.
3.2.1 RAPD
3.2.1.1 DNA izolasyonu
Çalışmalara başlamadan önce hangi DNA ekstraksiyon metodunun kullanılacağına
karar vermek için üç farklı metot denenmiştir. Bu metotlar denenirken materyal olarak
Lathyrus boissieri L. kullanılmıştır. Aşağıda her üç metodun da protokolleri verilmiştir.
3.2.1.1.1 CTAB (Cetyl Trimethyl Ammonium Bromide) DNA İzolasyonu (Doyle ve Doyle 1991)
200 mg kuru yaprak dokusu tartılarak havanda sıvı azot içinde öğütülmüştür.
Üzerine 300 µL CTAB tamponu (2% Cetyl Trimethyl Ammonium Bromide, 1.4 M
NaCl, 20 mM EDTA (Etilen Diamin Tetra Asetik Asit) pH 8.0, 100 mM Tris-HCl, pH
8.0) eklenmiş ve tekrar öğütülmüştür.
Ependorf tüpe aktarılan bu karışımın üzerine 300 µL daha CTAB tamponu eklenmiş ve
tek kullanımlık havaneli ile mümkün olduğunca homojen bir sıvı elde edilinceye kadar
ezilmiş ve karıştırılmıştır.
Daha sonra ependorf tüpler kar içine yerleştirilmiş 50 µL, %2’lik beta merkapto etanol
(BME) eklenip nazikçe karıştırılmıştır.
Karışım 65 °C’deki su banyosunda 1 saat inkübe edilmiştir.
500 µL kloroform:izoamilalkol [(C:IAA) (24:1), (v:v)] ilave edilmiş ve karışım süte
benzer bir kıvama gelene kadar nazikçe karıştırılmıştır.

38
13000 rpm’de +4 °C’de 15 dakika santrifüj edildikten sonra tüpler kar içine
yerleştirilmiştir.
Üst faz pipetle çekilmiş ve yeni bir ependorf tüpe aktarılmıştır.
500 µL -20 °C’de soğutulmuş isopropanol tüplere eklenmiş ve nazikçe karıştırılmıştır.
-80 °C’de 30 dakika bekletilmiştir.
İnkübasyondan sonra, eritilmeden 13000 rpm’de +4 °C’de 10 dakika santrifüj
edilmiştir.
Üst faz dökülmüş ve çökelti (pellet) 2-3 defa %70 ve %96’lık etanol ile yıkanmıştır.
Yıkamadan sonra ependorf tüpler, etanol tamamen uzaklaşana kadar, biyogüvenlik
kabininde en az 1 saat kurutulmuştur.
Ependorf tüpün dibinde çökelti halinde bulunan DNA, üzerine 50 µL TE tamponu (10
mM Tris-HCl pH 8.0, 1 mM EDTA pH 8.0) eklenerek çözülmüş ve kullanılana kadar
bu şekilde -20 °C’de saklanmıştır.
3.2.1.1.2 Modifiye CTAB DNA İzolasyonu (Aras vd. 2003)
Kuru yapraklardan 200 mg tartılmış ve havan içinde sıvı azot ile iyice öğütülmüştür.
Üzerine önceden ısıtılmış 800 µL CTAB tamponu (2% Cetyl Trimethyl Ammonium
Bromide [CTAB], 100 mM Tris-HCl (pH 8.0), 50 mM EDTA, 1.4 M NaCl,) , ilave
edilmiş ve yeniden öğütülmüştür.
Havandan alınan örnekler 1,5 mL’lik ependorf tüpe aktarıldıktan sonra üzerine 2 µL
proteinaz K (20 mg/mL) eklenmiştir.
Örnekler önce 37 °C’de 20 dakika, sonra 65 °C’de 20 dakika inkübe edilmiştir.

39
Örneklerin oda ısısında soğuması beklenmiş ve üzerlerine 250 µL 5 M potasyum asetat
eklenmiştir. Sonra buz üzerinde 30 dakika inkübe edilmiştir.
Örnekler 15 dakika 14000 rpm’de santrifüj edilmiş ve üst faz (süpernatan) yeni bir
ependorf tüpe aktarılmıştır.
Üzerlerine 500 µL fenol-kloroform-isoamilalkol (25:24:1[v:v:v]) karışımından eklenmiş
ve 40–50 defa nazikçe alt-üst edilerek karıştırıldıktan sonra 15 dakika buzda inkübe
edilmiştir.
14000 rpm’de 5 dakika sanrtifüjden sonra üst faz yeni bir 1,5 mL’lik ependorf tüpe
aktarılmış ve üzerine eşit hacimde isopropanol eklenmiştir. Daha sonra yine 15 dakika
buzda bekletilmiştir.
Tekrar 14000 rpm’de 15 dakikalık santrifüjden sonra üst faz atılmış ve çökelti (pellet)
üzerine tüp dolacak şekilde %76’lık etanol eklenmiş ve 14000 rpm’de 10 dakika bir
daha santrifüj edilmiştir.
Çökelti bir kez daha %70’lik etanol ile yıkanmış ve takiben bütün etanol uzaklaşana
kadar biyogüvenlik kabininde kurutulmuştur.
Tüp dibinde kalan DNA çökeltisi üzerine 50 µL TE tamponu (10 mM Tris-HCl (pH 8),
0.1 mM EDTA) eklenerek tekrar çözünmesi sağlanmıştır.
Üzerine 5 µL RNase eklenip vorteks ile iyice karıştırıldıktan sonra -20 °C’de
kullanılacağı zamana kadar saklanmıştır.
3.2.1.1.3 Hazır ekstraksiyon kiti (Favorprep Plant Genomic DNA extraction mini kit)
100 mg kuru yaprak sıvı azot ile havan içinde öğütülerek un haline getirilmiştir.
Öğütülmüş materyal 1,5 mL lik ependorf tüpe aktarılmıştır.

40
Üzerine 400 µL kit içinden çıkan GP1 Tampon ve 5 µL RNase A (10 mg/mL)
eklenmiş ve vortekslenmiştir.
65 °C’de 10 dakika inkübe edilmiş, inkübasyon sırasında 5 dakikada bir alt üst edilerek
karıştırılmıştır. Bu sırada 65 °C’de elüsyon tamponda ısıtılmıştır.
Üzerine 100 µL GP2 tampon eklenmiş ve vortekslenmiştir.
Buzda 3 dakika inkübasyona bırakılmıştır.
Karışım Filter Kolon tüplerine aktarılmıştır
3 dakika 13000 rpm’de santrifüj edilmiştir.
Üst faz dikkatlice yeni ependorf tüpe aktarılmıştır.
GP3 tampon üzerine 3 mL isopropanol eklenmiştir
Yıkama tamponu üzerine 4 mL EtOH (etanol) eklenmiştir.
Yeni tüpe aktarılmış üst fazın üzerine, (her 500 µL’si için 750 µL GP3 tampon ,
(isopropanol eklenmiş) olacak şekilde) GP3 tampon eklenmiş ve 5 saniye
vortekslenmiştir.
GD kolon tüplerine 700 µL karışım eklenmiştir.
13000 rpm’de 2 dakika santrifüj edilmiştir.
Filtreden geçen alttaki kısım atılmış ve ependorfda kalan karışım tekrar eklenmiştir.
13000 rpm’de tekrar 2 dakika santrifüj edilmiş ve filtreden geçen alttaki kısım tekrar
atılmıştır.

41
500 µL Yıkama tamponu (etanol eklenmiş) kolona eklenmiştir. 13000 rpm’de 30
saniye santrifüj edilmiştir. Süzüntü atılmış kolon tüpe tekrar konmuştur.
Yıkama (bir önceki adım) tekrarlanmıştır. Süzüntü atılmış kolon tüpe tekrar konmuştur.
Kolon matriksini kurutmak için 3 dakika 13000 rpm’de santrifüj edilmiştir.
Kurutulmuş GD kolonu temiz bir tüpe aktarılmıştır
GD kolon matriksinin ortasına 100 µL önısıtılmış elüsyon tampon eklenmiştir.
Elüsyon tamponunun kolon tarafından emilmesi için 3-5 dakika beklenmiş sonra 13000
rpm’de 30 saniye santrifüj edilmiştir.
3.2.1.1.4 DNA saflık ve miktar tayini
-20 °C’de saklanan çözünmüş DNA’dan 1 µL alınıp, Ankara Üniversitesi Biyoteknoloji
Enstitüsü Merkez Laboratuvarı’nda, Agilent 2100 Bioanalyser NanoDrop ND-1000
spektrofotometre ile (Şekil 3.2) miktarı ve saflığı (260/280nm) ölçülmüştür.
Şekil 3.2 Agilent 2100 Bioanalyser NanoDrop ND-1000 spektrofotometre (http://cgb.indiana.edu/genomics/equipment/15, 2011)
Elde edilen sonuçlara göre (Çizelge 4.1) DNA konsantrasyonu 1 µL de 50 ng DNA
olacak şekilde distile suyla sulandırılmıştır.

42
3.2.1.1.5 Total DNA Elektroforezi
DNA’nın kalitatif olarak analizi amacıyla modifiye CTAB metodu (Aras vd. 2003) ile
hazırlanan DNA çözeltileri % 0,8 oranında hazırlanan agaroz jellere yüklenmiş, 70
voltta, 2 saat elektroforeze tabi tutulmuştur. Elektroforez sonrasında 5µg/ml’lik etidyum
bromür boyasında 1/2 saat boyanmış, UV ışık altında incelenmiş ve KODAK Gel Logic
2200 görüntüleme sisteminde fotoğraflanmıştır (Şekil 3.3).
Şekil 3.3 KODAK Gel Logic 2200 görüntüleme sistemi (http://blestbj.com/product/dianyong/ningjiao/2009/0605/726.html,2011)
3.2.1.2 Primerler
Primer seçiminde gelişigüzel davranılmış rastgele seçilmiş 10 bazlık primerler
kullanılmıştır. Toplam 25 primer (Çizelge 3.5) ile çalışılmış ve tamamından da iyi bant
desenleri elde edilmiştir. Bu primerler Thermo Electron firması tarafından üretilen
OPA, OPB, OPD ve OPJ serisindendir. Tekrarlanabilirliği test etmek amacıyla OPA 9
primeriyle tekrarlama yapılmış ve aynı sonuçlar alınmıştır. PZR’de denatürasyonu
takiben primerlerin bağlanması aşamasındaki sıcaklığın tespit edilmesi gerekir. Bunun
için değişik formüller vardır. Boyu 20 baza kadar olan primerler için genellikle
aşağıdaki formül kullanılır. Bu formülle elde edilen değer primerlerin Tm (melting
temperature/çözünme sıcaklığı) değeridir. Çoğunlukla bu değerin 3-5 °C altındaki
sıcaklıklar, bağlanma sıcaklığı olarak seçilir.
[(A+T lerin sayısı) x 2 °C + (G+C lerin sayısı) x 4°C]
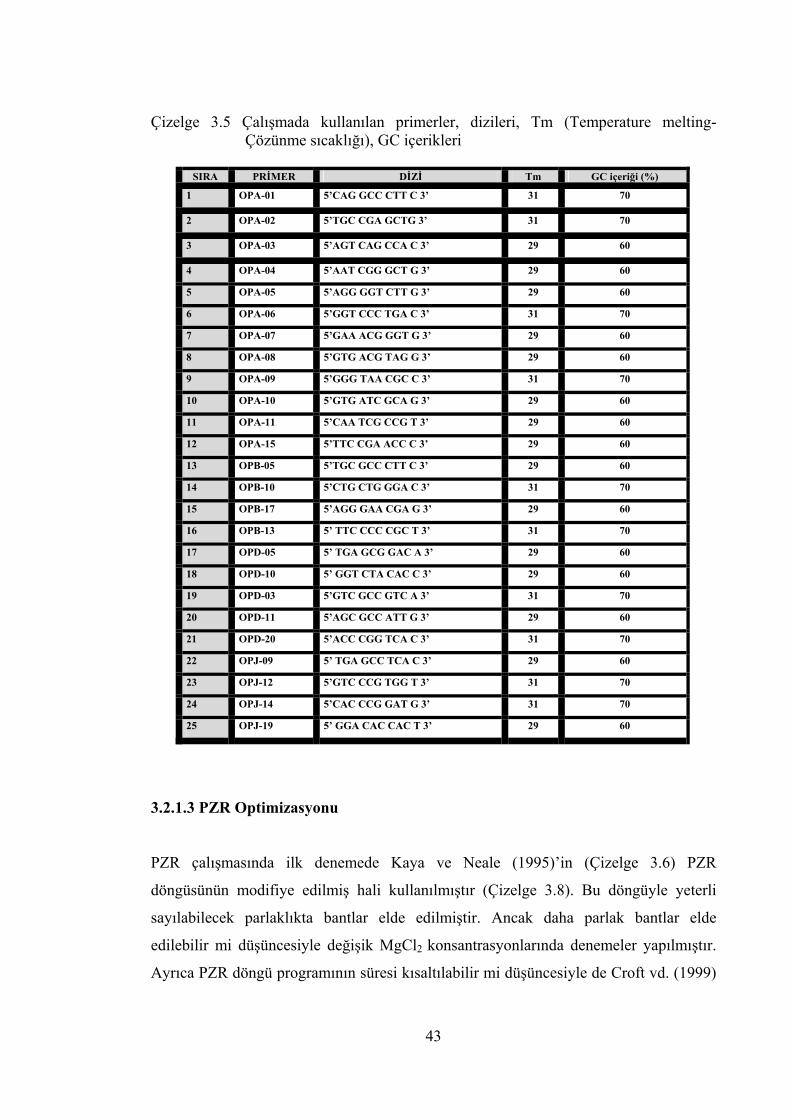
43
Çizelge 3.5 Çalışmada kullanılan primerler, dizileri, Tm (Temperature melting-Çözünme sıcaklığı), GC içerikleri
SIRA PRİMER DİZİ Tm GC içeriği (%)
1 OPA-01 5’CAG GCC CTT C 3’ 31 70
2 OPA-02 5’TGC CGA GCTG 3’ 31 70
3 OPA-03 5’AGT CAG CCA C 3’ 29 60
4 OPA-04 5’AAT CGG GCT G 3’ 29 60
5 OPA-05 5’AGG GGT CTT G 3’ 29 60
6 OPA-06 5’GGT CCC TGA C 3’ 31 70
7 OPA-07 5’GAA ACG GGT G 3’ 29 60
8 OPA-08 5’GTG ACG TAG G 3’ 29 60
9 OPA-09 5’GGG TAA CGC C 3’ 31 70
10 OPA-10 5’GTG ATC GCA G 3’ 29 60
11 OPA-11 5’CAA TCG CCG T 3’ 29 60
12 OPA-15 5’TTC CGA ACC C 3’ 29 60
13 OPB-05 5’TGC GCC CTT C 3’ 29 60
14 OPB-10 5’CTG CTG GGA C 3’ 31 70
15 OPB-17 5’AGG GAA CGA G 3’ 29 60
16 OPB-13 5’ TTC CCC CGC T 3’ 31 70
17 OPD-05 5’ TGA GCG GAC A 3’ 29 60
18 OPD-10 5’ GGT CTA CAC C 3’ 29 60
19 OPD-03 5’GTC GCC GTC A 3’ 31 70
20 OPD-11 5’AGC GCC ATT G 3’ 29 60
21 OPD-20 5’ACC CGG TCA C 3’ 31 70
22 OPJ-09 5’ TGA GCC TCA C 3’ 29 60
23 OPJ-12 5’GTC CCG TGG T 3’ 31 70
24 OPJ-14 5’CAC CCG GAT G 3’ 31 70
25 OPJ-19 5’ GGA CAC CAC T 3’ 29 60
3.2.1.3 PZR Optimizasyonu
PZR çalışmasında ilk denemede Kaya ve Neale (1995)’in (Çizelge 3.6) PZR
döngüsünün modifiye edilmiş hali kullanılmıştır (Çizelge 3.8). Bu döngüyle yeterli
sayılabilecek parlaklıkta bantlar elde edilmiştir. Ancak daha parlak bantlar elde
edilebilir mi düşüncesiyle değişik MgCl2 konsantrasyonlarında denemeler yapılmıştır.
Ayrıca PZR döngü programının süresi kısaltılabilir mi düşüncesiyle de Croft vd. (1999)

44
tarafından kullanılan farklı bir döngü daha denenmiştir (Çizelge 3.7). Bunun için, iki
farklı türün, iki farklı primer ile 5 farklı (2.0 µL, 2.5 µL, 3.0 µL, 4.0 µL, 5.0 µL) MgCl2
konsantrasyonu, iki farklı PZR döngüsü ile denenmiştir (Çizelge 3.9).
Çizelge 3.6 Kaya ve Neale (1995) tarafından uygulanan PZR döngüsü
Basamak Sıcaklık Zaman Döngü Sayısı Ön Denaturasyon 95°C 1 dakika 1
Denaturasyon 94°C 1 dakika Bağlanma
(Annealing) 37°C 1 dakika
Uzama (Extension) 72°C 2 dakika
40
Son Uzama 72°C 15 dakika 1
Çizelge 3.7 Optimizasyonda denenen PZR2 döngüsü (Croft vd. 1999).
Basamak Sıcaklık Zaman Döngü Sayısı Ön Denaturasyon 94°C 1 dakika 1
Denaturasyon 94°C 10 saniye Bağlanma
(Annealing) 40°C 30 saniye
Uzama (Extension) 72°C 1 dakika
35
Son Uzama 72°C 5 dakika 1
Çizelge 3.8 Çalışmada kullanılan PZR1 döngüsü
Basamak Sıcaklık Zaman Döngü Sayısı Ön Denaturasyon 95°C 1 dakika 1
Denaturasyon 94°C 1 dakika Bağlanma
(Annealing) 36°C 2 dakika
Uzama (Extension) 72°C 2 dakika
45
Son Uzama 72°C 15 dakika 1

45
İlk çalışmada kullanılan reaksiyon karışımı;
• 1 μl DNA örneği (50ng/μl),
• 2.5 μl tampon (750mM Tris-HCl ph: 8.8, 200mM (NH4)2 SO4; Fermentas,
Ukrayna),
• 0.3 μl (100 ünite) Taq DNA Polimeraz (Fermentas, Ukrayna),
• 4 μl deoksinükleotit trifosfat karışımı (her bir nükleotitten 200μM) (Fermentas,
Ukrayna),
• 2 μl 2mM MgCl2, (Fermentas, Ukrayna),
• 1 μl (1pmol) primer (Thermo Electron) den oluşmaktadır.
• Toplam hacim 14.2 μl steril distile H20 ile 25 μl’ye tamamlanmıştır.
•
Optimizasyon sonrası kullanılan reaksiyon karışımı;
• 1 μl DNA örneği (50ng/μl),
• 2.5 μl tampon (750mM Tris-HCl ph: 8.8, 200mM (NH4)2 SO4; Fermentas,
Ukrayna),
• 0.3 μl (100 ünite) Taq DNA Polimeraz (Fermentas, Ukrayna),
• 4 μl deoksinükleotit trifosfat karışımı (her bir nükleotitten 200μM), (Fermentas,
Ukrayna),
• 3 μl 2mM MgCl2, (Fermentas, Ukrayna),
• 1 μl (1pmol) primer (Thermo Electron) den oluşmaktadır.
• Toplam hacim 13.2 μl steril distile H20 ile 25 μl’ye tamamlanmıştır.
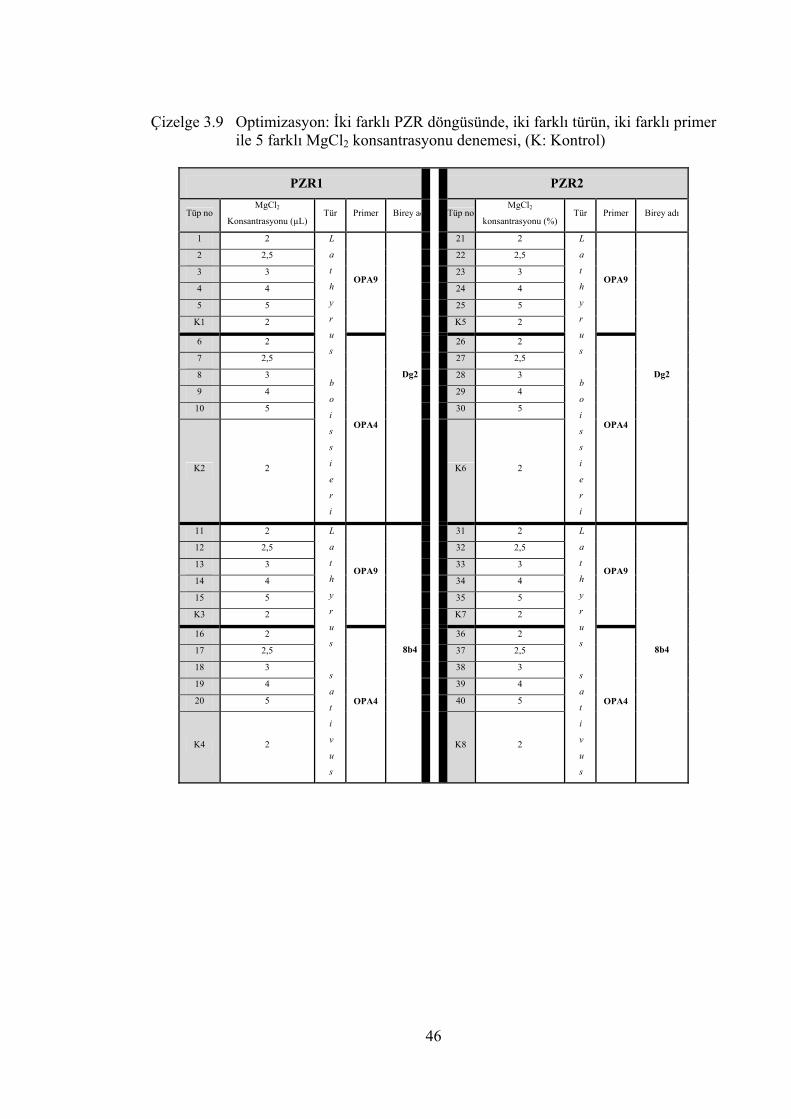
46
Çizelge 3.9 Optimizasyon: İki farklı PZR döngüsünde, iki farklı türün, iki farklı primer ile 5 farklı MgCl2 konsantrasyonu denemesi, (K: Kontrol)
PZR1 PZR2
Tüp no MgCl2
Konsantrasyonu (µL) Tür Primer Birey ad Tüp no
MgCl2
konsantrasyonu (%) Tür Primer Birey adı
1 2 21 2
2 2,5 22 2,5
3 3 23 3
4 4 24 4
5 5 25 5
K1 2
OPA9
K5 2
OPA9
6 2 26 2
7 2,5 27 2,5
8 3 28 3
9 4 29 4
10 5 30 5
K2 2
L
a
t
h
y
r
u
s
b
o
i
s
s
i
e
r
i
OPA4
Dg2
K6 2
L
a
t
h
y
r
u
s
b
o
i
s
s
i
e
r
i
OPA4
Dg2
11 2 31 2
12 2,5 32 2,5
13 3 33 3
14 4 34 4
15 5 35 5
K3 2
OPA9
K7 2
OPA9
16 2 36 2
17 2,5 37 2,5
18 3 38 3
19 4 39 4
20 5 40 5
K4 2
L
a
t
h
y
r
u
s
s
a
t
i
v
u
s
OPA4
8b4
K8 2
L
a
t
h
y
r
u
s
s
a
t
i
v
u
s
OPA4
8b4

47
3.2.1.4 Agaroz jel elektroforezi ve görüntüleme
8,5 gram agaroz tartılıp, 500 mL 1X TAE pH 7.5 [(Tris-Asetik Asit-EDTA) 40 mM
Tris, 20 mM asetik asit, 1 mM EDTA] çözeltisiyle kaynatılarak karıştırılmıştır. Elde
edilen, %1.7 lik agaroz 50-55 ° C ye soğuduktan sonra jel tepsisine dökülmüştür. Bu
sırada 4 adet 26 dişli tarak, kuyu oluşturacak şekilde jel tepsisine yerleştirilmiş, agaroz
donup jeli oluşturduktan sonra da çıkarılmıştır. Jel tepsisi elektroforez tankına
yerleştirilmiş ve üzeri kapanacak şekilde, 1X TAE pH 7.5 çözeltisi dökülmüştür. Daha
sonra tarakların oluşturduğu kuyulara, 5 µL yükleme tamponu [10 mM Tris-HCl (pH
7.6), 0.03% bromophenol blue, 0.03% xylene cyanol FF, 60% glycerol, 60 mM EDTA]
ile karıştırılmış, 25 µL PZR ürünü aktarılmıştır. PZR ürünlerinin elektroforezi
sonrasında büyüklüklerini anlayabilmek için DNA standartı olarak “GeneRuler™ 100
bp DNA Ladder, 100-1000 bp (Fermentas)” ve “GeneRuler™ 100 bp Plus DNA
Ladder, 100-3000 bp (Fermentas)” kullanılmıştır (Şekil 3.4). Marker’dan 4 sıralık 26
kuyunun her iki başına da 5 µL konmuştur. Daha sonra jel 90 Voltta 3-4 saat
elektroforeze tabi tutulmuştur. Elektroforez sonrasında jel 5µg/ml’lik etidyum bromür
çözeltisinde 30 dakika tutularak boyanmış, UV ışığı altında KODAK Gel Logic 2200
görüntüleme sistemi ile fotoğrafları alınmış, jeller atılmıştır.
a b Şekil 3.4 Çalışmada kullanılan markerler: a. GeneRuler™ 100 bp Plus DNA Ladder,
100-3000 bp, b. GeneRuler™ 100 bp DNA Ladder 100-1000 bp

48
3.2.1.5 Jellerin Değerlendirilmesi
Bant varlığına “1” yokluğuna ise “0” değeri verilerek skorlama işlemi yapılmıştır.
Gözlenen tüm bantlar değerlendirmeye alınmıştır.
3.2.1.6 RAPD Verilerinin Analizi (İstatistik Analizi)
Jellerin değerlendirilmesinden elde edilen skorlama verileri POPGENE Microsoft
Window-based Freeware for Population Genetics Analysis, Version 1.31 (Yeh vd.1997)
bilgisayar programına girilerek genetik mesafeler hesaplanmış ve genetik mesafeler ile
MEGA (Molecular Evolutionary Genetics Analysis) versiyon 3.0 (Kumar vd. 2004)
programından filogenetik ilişkileri gösteren dendrogram alınmıştır.
3.2.2 SDS-PAGE
3.2.2.1 Protein Ekstraksiyonu
Tohum depo proteinlerinin ekstraksiyonu için Laemmli (1970)’nin önerdiği protein
ekstraksiyon tamponu (pH 6.8) [0.0625 M Trisma base , %2 Sodyum Dodesil Sülfat-
SDS), %10 Gliserol, %5 2-merkaptoetanol, %0.1 brom fenol mavisi] kullanılmıştır.
İzolasyon işlemine başlamadan önce, türlere ait tohumların kabukları (testa) çıkarılmış
ve havanda dövülerek un haline getirilmiştir. 0.5 g un alınarak bir ependorf tüpte 300
µL protein ekstraksiyon tamponu ile karıştırılmış ve elektrikli homojenizatörde (Şekil
3.5) 1 dakika homojenize edilmiştir. Daha sonra üzerine 1200 µL daha protein
ekstraksiyon tamponu eklenmiş ve tek kullanımlık havan eli ile tüpler içinde iyice ezilip
homojenize edildikten sonra, 14 000 rpm’de 10 dakika santrifüj edilmiştir. Santrifüj
sonrasında tüpler üzerinde kalan berrak kısımlar (supernatan) yeni ependorf tüplere
aktarılmıştır. Bu tüpler elektroforez işlemi gerçekleştirilene kadar -20 °C’de
saklanmıştır. Jellere yüklenmeden hemen önce, denatürasyon için, 100 °C’deki
benmaride 5 dakika bekletilmiştir.

49
Şekil 3.5 Çalışmada kullanılan tohumlardan protein ekstraksiyonu için kullanılan elektrikli homojenizatör
3.2.2.1.1 Protein miktar tayini
Jele yüklenecek protein miktarı Esen (1978)’in metoduna göre semikantitatif olarak
belirlendi. Buna göre ekstraksiyon işlemi sonrasında elde edilen supernatandan alınan
50 µL’lik örnekler şerit halindeki Whatman No 4 filtre kağıdına emdirilmiştir. Bu filtre
kağıdı, aşağıda konsantrasyonu verilen, Coomassie Brillant Blue boyasına daldırılıp. 30
dakika bekledikten sonra çıkarılıp, suyla yıkanmıştır. Protein miktarına göre,
supernatan emdirilmiş bölgelerin renk şiddetinin durumu değerlendirilmiştir.
3.2.2.2 Protein Elektroforezi
3.2.2.2.1 Poliakrilamid jellerin hazırlanması
Hazırlanan protein ekstraktlarının, elektroforez için jel üzerine yerleştirilmeleri gerekir.
Bu jeller poliakrilamid yapıdadır ve iki ayrı kısımdan oluşur. Birinci kısım yükleme jeli
(stacking gel), ikinci kısım ayırma jelidir (resolving gel). Adından da anlaşılacağı üzere
protein ekstraktları önce yükleme jeline yüklenir ve bu bölgede önce bir düzene girerler
ve ayırma jeline düzene girmiş şekilde girip ilerlemeye devam ederler. Jellerin
hazırlanması Sambrook vd. (1989)’nin önerdiği gibi aşağıda anlatıldığı şekilde

50
gerçekleştirilmiştir. SDS-PAGE işlemini gerçekleştirmek için “BIORAD mini protean
vertical elektroforez sistemi” kullanılmıştır ( Şekil 3.6).
Şekil 3.6 BIORAD mini protean vertical elektroforez sistemi (http://www.uklabs-direct.co.uk/html/used_electrophoresis_equipment.html, 2011)
Ayırma jeli hazırlanması
Elektroforez ekipmanının jel hazırlama kaseti sızdırmaz bir şekilde hazırlandıktan
sonra, içine içeriği aşağıda verilen şekilde hazırlanan çözelti dökülmüş ve üzeri suya
doymuş n-bütanol ile kapatılmıştır. Bunun sebebi yüzey geriliminden dolayı cam
kenarlarına temas eden, havayla irtibat halindeki üst kısmın, konkavlaşarak jelin
kenarlarının yukarı doğru kıvrılmasını engellemektir. Bu şekilde 30 dakika bekletilen
kaset içindeki çözelti (akrilamid) polimerleşerek katı hale gelir. Sonra üst kısmındaki
n-bütanol dökülmüş ve n-bütanolle temas etmiş olan bu kısım bol distile suyla iyice
yıkanmıştır. Üzerine yükleme jeli dökülmüştür.
Ayırma Jeli Çözeltisi (%T=10 ve molar bisakrilamid:akrilamid oranı 1:29)
• %30 Akrilamid Stok ……..10 ml
• 1,5 M Trisma base (pH 8.8)..7.5 ml
• %10 SDS…………..……….300 µl
• %10 APS…………………….300 µl
• TEMED………………………..18 µl
• Distile Su…………………..12.4 ml

51
Yükleme jeli hazırlanması
Ayırma jeli üzerine aşağıda belirtilen şekilde hazırlanan çözelti dökülmüş ve hemen
içine kuyucuk oluşumunu sağlayacak taraklar yerleştirilmiştir. 15 dakika bekledikten
sonra çözelti (akrilamid) polimerleşerek katı hale gelince tarak dikkatli bir şekilde
çıkarılmış ve kuyucuklar distile suyla yıkanmıştır.
Yükleme Jeli çözeltisi (%T= 4 ve molar bisakrilamid:akrilamid oranı 1:29)
• %30 Akrilamid Stok…..……..660 µl
• 1 M Trisma base (pH 6.8)….. 625 µl
• %10 SDS…………………...….50 µl
• %10 APS…………………...….50 µl
• TEMED……………………..…..5 µl
• Distile Su………………........3.57 ml
3.2.2.2.2 Örneklerin yüklenmesi
İçinde poliakrilamid jel bulunan kaset, elektroforez tankına yerleştirildikten sonra tank
aşağıda belirtilen şekilde hazırlanmış olan tank tamponuyla (Laemmli 1970) kasetin
üzerini kapatacak şekilde doldurulmuştur. Kuyucuklara 7,5 µL örnek yüklenmiştir.
Marker olarak “Prestained Protein Molecular Weight Marker 15 kDA- 140 kDA
(Fermentas)” (Şekil 3.7) kullanılmıştır. Daha sonra elektrik akımı verilerek elektroforez
işlemi başlatılmıştır. Yükleme jeline 30 volt sabit elektrik akımı uygulanmıştır. İzleme
boyası takip edilmiştir. İzleme boyasının ayırma jeline geçmesiyle elektrik akımı 200
volta ayarlanmıştır. İzleme boyasının jel bitimine ulaşmasına 0,5 cm kalınca
elektroforez durdurulmuştur.

52
Şekil 3.7 SDS-PAGE yönteminde kullanılan protein standartı: Prestained Protein Molecular Weight Marker 15 kDA- 140 kDA (Fermentas)
Tank tamponu (pH 8.3 ) (Laemmli 1970).
• 25 mM Trisma base
• 192 mM glisin
• % 0.1 SDS
3.2.2.2.3 Poliakrilamid Jel boyanması
Elektroforez işlemi sonucunda jeller kasetten çıkarılmış Coomassie Brillant Blue boyası
ile 2 saat boyanmış, daha sonra jel zemini boyadan arındırılıncaya kadar şeffaflaştırıcıda
bekletilmiştir.
Coomassie Brillant Blue boyası
• % 0.1 Coomassie blue R-250
• % 45 methanol
• % 10 asetik asit
• % 45 distile su

53
Şeffaflaştırıcı (Destaining) çözeltisi
• % 45 methanol
• % 10 asetik asit
• % 45 distile su
3.2.2.2.4 Bant desenlerinin görüntülenmesi
Şeffaflaştırıcıdan çıkarılan jeller. “Bio-Rad Molecular imager Gel doc XR” görüntüleme
sistemine (Şekil 3.8) yerleştirilmiş, bant deseni fotoğrafları alınmıştır.
Şekil 3.8 SDS-PAGE jellerini görüntülemek için kullanılan “Bio-Rad Molecular imager Gel doc XR” görüntüleme sistemi (http://www.bio-rad.com/webroot/web/images/lsr/products/imaging_bioinformatics/product_detail/global/lsr_chemidoc_xrs_system.jpg, 2011)
3.2.2.3 SDS-PAGE Verilerinin Analizi (İstatistik Analizi)
Bant desenleri incelenmiş ve RAPD yönteminde olduğu gibi bant varlığı “1”, yokluğu
“0” olarak değerlendirilerek skorlama yapılmıştır. Bu şekilde oluşturulan veri dosyaları
POPGENE Microsoft Window-based Freeware for Population Genetics Analysis,
Version 1.31 (Yeh vd.1997) ve MEGA (Molecular Evolutionary Genetics Analysis)
versiyon 3.0 (Kumar vd. 2004) bilgisayar programlarına yüklenmiştir. Programlarla,
taksonlar arasındaki benzerlik indeksleri oluşturulmuş ve dendrogramlar elde edilmiştir.

54
4. ARAŞTIRMA BULGULARI 4.1 RAPD 4.1.1 DNA izolasyonu DNA ekstraksiyonu için denenen üç teknik arasında en parlak bantlar hazır ekstraksiyon
kiti (Favorgen Plant genomik DNA Ekstraksiyon Mini Kit) metoduyla elde edilmiştir.
Kuru yaprak dokularından DNA ekstraksiyonu için modifiye edilmiş CTAB metodunun
(Aras vd. 2003), denenmesiyle hazır kit metoduna nispeten daha az başarılı sonuçlar
alınmıştır. CTAB DNA (Doyle ve Doyle 1991) ekstraksiyonu ile elde edilen bantlar
diğerlerine nazaran oldukça silik çıkmıştır. Bu sonuçların değerlendirmesiyle
kullanılacak ekstraksiyon metodunun modifiye CTAB olmasına karar verilmiştir. Bu
tekniğin tercih edilmesindeki ana etken ekonomik açıdan daha uygun olmasıdır. Bunun
yanısıra elde edilen bant parlaklığının yeterli olduğu görülmüştür (Şekil 4.1).
4.1.1.1 DNA miktar tayini Hazırlanmış olan DNA çözeltilerinden 1’er µL alınıp nanodrop spektrofotometrede
(Agilent 2100 Bioanalyser NanoDrop ND-1000 Spektrophotometer) ölçülen, miktar ve
saflık (Absorbance260/Absorbance280) değerleri çizelge 4.1’de verilmiştir. Bu sonuçlara
göre materyallere ait DNA miktarı 50.89 – 2225.05 ng/ µL aralığında olmuştur. DNA
saflığı (A260/A280) ise 1.92 – 1.63 aralığındadır. Numuneler, bu verilere göre, DNA
konsantrasyonu 50 ng/μL olacak şekilde distile suyla sulandırılmıştır.
4.1.1.2 Total DNA elektroforezi Aras vd. (2003)’nin önerdiği modifiye edilmiş CTAB metodu kullanılarak izole edilen
DNA’nın kalitesini anlamak için yapılan agaroz jel elektroforezi sonucu elde edilen
DNA bantları şekil 4.2’de görülmektedir.
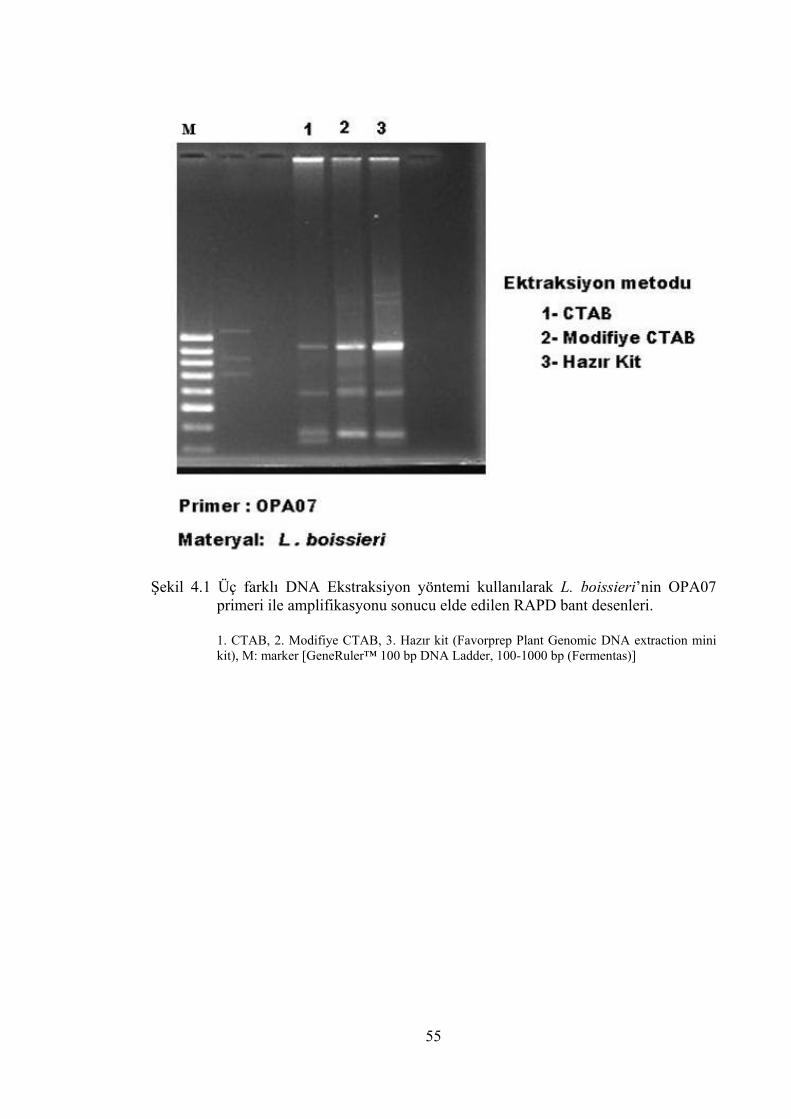
55
Şekil 4.1 Üç farklı DNA Ekstraksiyon yöntemi kullanılarak L. boissieri’nin OPA07
primeri ile amplifikasyonu sonucu elde edilen RAPD bant desenleri.
1. CTAB, 2. Modifiye CTAB, 3. Hazır kit (Favorprep Plant Genomic DNA extraction mini kit), M: marker [GeneRuler™ 100 bp DNA Ladder, 100-1000 bp (Fermentas)]
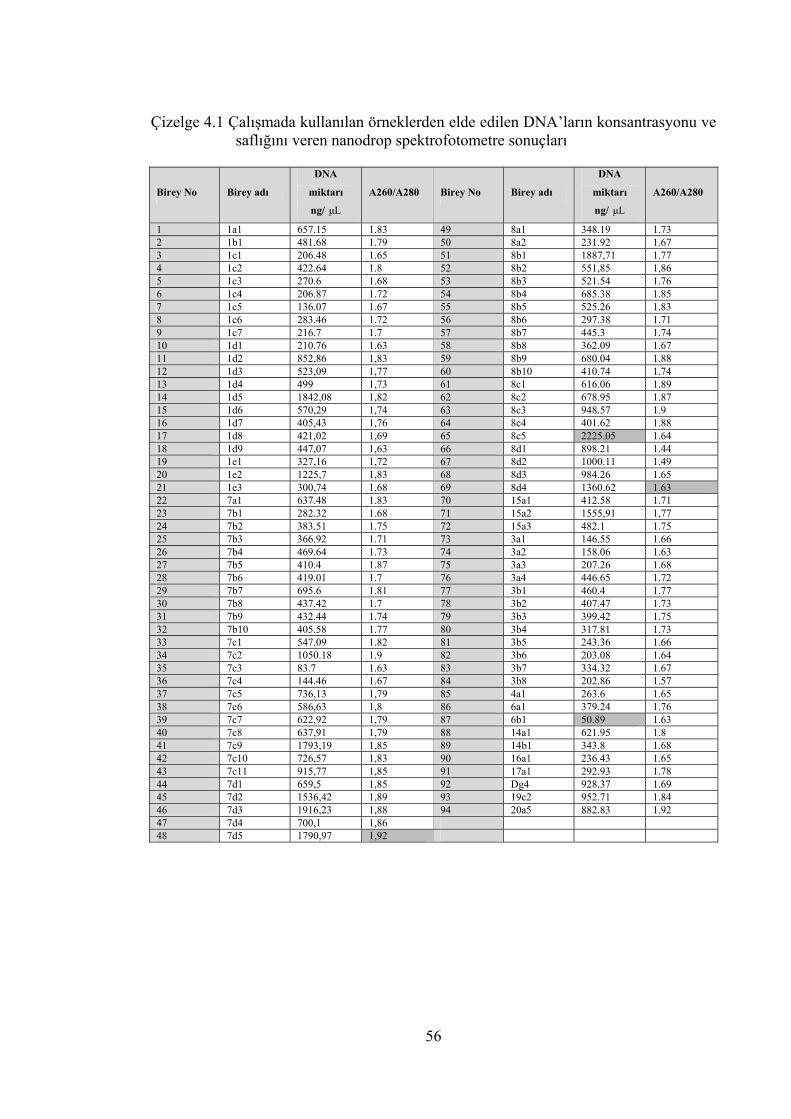
56
Çizelge 4.1 Çalışmada kullanılan örneklerden elde edilen DNA’ların konsantrasyonu ve saflığını veren nanodrop spektrofotometre sonuçları
Birey No Birey adı
DNA
miktarı
ng/ μL
A260/A280 Birey No Birey adı
DNA
miktarı
ng/ μL
A260/A280
1 1a1 657.15 1.83 49 8a1 348.19 1.73 2 1b1 481.68 1.79 50 8a2 231.92 1.67 3 1c1 206.48 1.65 51 8b1 1887,71 1.77 4 1c2 422.64 1.8 52 8b2 551,85 1,86 5 1c3 270.6 1.68 53 8b3 521.54 1.76 6 1c4 206.87 1.72 54 8b4 685.38 1.85 7 1c5 136.07 1.67 55 8b5 525.26 1.83 8 1c6 283.46 1.72 56 8b6 297.38 1.71 9 1c7 216.7 1.7 57 8b7 445.3 1.74 10 1d1 210.76 1.63 58 8b8 362.09 1.67 11 1d2 852,86 1,83 59 8b9 680.04 1.88 12 1d3 523,09 1,77 60 8b10 410.74 1.74 13 1d4 499 1,73 61 8c1 616.06 1.89 14 1d5 1842,08 1,82 62 8c2 678.95 1.87 15 1d6 570,29 1,74 63 8c3 948.57 1.9 16 1d7 405,43 1,76 64 8c4 401.62 1.88 17 1d8 421,02 1,69 65 8c5 2225.05 1.64 18 1d9 447,07 1,63 66 8d1 898.21 1.44 19 1e1 327,16 1,72 67 8d2 1000.11 1.49 20 1e2 1225,7 1,83 68 8d3 984.26 1.65 21 1e3 300,74 1,68 69 8d4 1360.62 1.63 22 7a1 637.48 1.83 70 15a1 412.58 1.71 23 7b1 282.32 1.68 71 15a2 1555,91 1,77 24 7b2 383.51 1.75 72 15a3 482.1 1.75 25 7b3 366.92 1.71 73 3a1 146.55 1.66 26 7b4 469.64 1.73 74 3a2 158.06 1.63 27 7b5 410.4 1.87 75 3a3 207.26 1.68 28 7b6 419.01 1.7 76 3a4 446.65 1.72 29 7b7 695.6 1.81 77 3b1 460.4 1.77 30 7b8 437.42 1.7 78 3b2 407.47 1.73 31 7b9 432.44 1.74 79 3b3 399.42 1.75 32 7b10 405.58 1.77 80 3b4 317.81 1.73 33 7c1 547.09 1.82 81 3b5 243.36 1.66 34 7c2 1050.18 1.9 82 3b6 203.08 1.64 35 7c3 83.7 1.63 83 3b7 334.32 1.67 36 7c4 144.46 1.67 84 3b8 202.86 1.57 37 7c5 736,13 1,79 85 4a1 263.6 1.65 38 7c6 586,63 1,8 86 6a1 379.24 1.76 39 7c7 622,92 1,79 87 6b1 50.89 1.63 40 7c8 637,91 1,79 88 14a1 621.95 1.8 41 7c9 1793,19 1,85 89 14b1 343.8 1.68 42 7c10 726,57 1,83 90 16a1 236.43 1.65 43 7c11 915,77 1,85 91 17a1 292.93 1.78 44 7d1 659,5 1,85 92 Dg4 928.37 1.69 45 7d2 1536,42 1,89 93 19c2 952.71 1.84 46 7d3 1916,23 1,88 94 20a5 882.83 1.92 47 7d4 700,1 1,86 48 7d5 1790,97 1,92
57
a
b
c
d
e
Şekil 4.2 Total DNA elektroforezi (birey no ve isimleri çizelge 3.4 te verilmiştir)

58
4.1.2 PZR Optimizasyonu
Jeller incelendiğinde en iyi bant oluşumunu sağlayan magnezyum klorür (MgCl2)
konsantrasyonunun 3µL’lik miktar olduğu ve PZR1 (Çizelge 3.8) döngüsünde elde
edilen çoğaltmanın iyi olduğu, PZR2 (Çizelge 3.7) döngüsünün ise neredeyse hiç
çoğaltma yapamadığı belirlenmiştir (Şekil 4.3 ve 4.4).
Şekil 4.3 PZR1 döngüsü uygulanarak iki Lathyrus türünün iki farklı primer, 5 farklı magnezyum klorür (MgCl2) konsantrasyonu ile gerçekleştirilen RAPD sonuçları.
M: Marker [(GeneRuler™ 100 bp DNA Ladder, 100-1000 bp (Fermentas)], K: Negatif Kontrol. 1-5 L. boissieri (OPA9), 6-10 L. boissieri (OPA4), 11-15 L. sativus (OPA9), 16-20 L. sativus (OPA4)

59
Şekil 4.4 PZR2 döngüsü uygulanarak iki Lathyrus türünün iki farklı primer, 5 farklı magnezyum klorür (MgCl2) konsantrasyonu ile gerçekleştirilen RAPD sonuçları.
M: Marker [(GeneRuler™ 100 bp DNA Ladder, 100-1000 bp (Fermentas)], K: Negatif Kontrol. 21-25 L. boissieri (OPA9), 26-30 L. boissieri (OPA4), 31-35 L. sativus (OPA9), 16-20 L. sativus (OPA4)
4.1.3 RAPD Polimorfizmi ve Genetik Mesafelerin Belirlenmesi 4.1.3.1 Türler Arası Genetik Mesafeler Çalışmada 25 primer denenmiş ve 25 primerin 94 birey (Çizelge 3.4) ile yapılan RAPD
çalışmasında elde edilen toplam bant sayısı 771 olmuş bu bantlardan 769 (%99,74)’u
polimorfik bulunmuştur. Çalışmada kullanılan primerler ile agaroz jellerde elde edilen,
bant desenlerinin görüntüleri şekil 4.5 – 4.29’da verilmiştir. Çizelge 4.2’de primerlerin
ortaya koyduğu bant sayıları, bu bantların polimorfizm oranları verilmiştir. 25 primer
arasında en fazla bant sayısını 44 olarak OPA-11 (Şekil 4.15) en az bant sayısını ise 19
olarak OPA-3 (Şekil 4.7), ve OPA-9 (Şekil 4.13), primeri ortaya koymuştur. OPB-10
(Şekil 4.18), ve OPJ-19 (Şekil 4.29) primeri dışındaki primerlerin ortaya koyduğu bütün
bantların (%100) polimorfik olduğu belirlenmiştir. OPB-10 ve OPJ-19 primerlerinin
polimorfizm oranı ise sırasıyla, %96,43 ve %95,50 olmuştur.
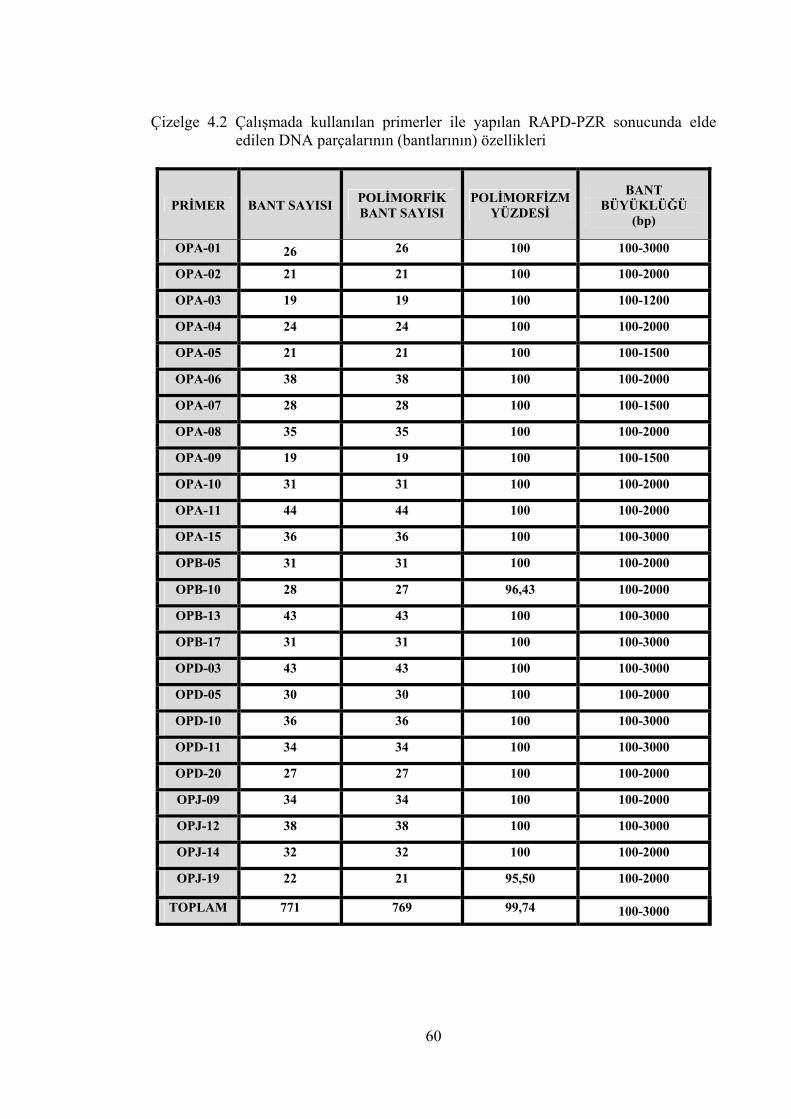
60
Çizelge 4.2 Çalışmada kullanılan primerler ile yapılan RAPD-PZR sonucunda elde edilen DNA parçalarının (bantlarının) özellikleri
PRİMER BANT SAYISI POLİMORFİK BANT SAYISI
POLİMORFİZM YÜZDESİ
BANT BÜYÜKLÜĞÜ
(bp)
OPA-01 26 26 100 100-3000
OPA-02 21 21 100 100-2000
OPA-03 19 19 100 100-1200
OPA-04 24 24 100 100-2000
OPA-05 21 21 100 100-1500
OPA-06 38 38 100 100-2000
OPA-07 28 28 100 100-1500
OPA-08 35 35 100 100-2000
OPA-09 19 19 100 100-1500
OPA-10 31 31 100 100-2000
OPA-11 44 44 100 100-2000
OPA-15 36 36 100 100-3000
OPB-05 31 31 100 100-2000
OPB-10 28 27 96,43 100-2000
OPB-13 43 43 100 100-3000
OPB-17 31 31 100 100-3000
OPD-03 43 43 100 100-3000
OPD-05 30 30 100 100-2000
OPD-10 36 36 100 100-3000
OPD-11 34 34 100 100-3000
OPD-20 27 27 100 100-2000
OPJ-09 34 34 100 100-2000
OPJ-12 38 38 100 100-3000
OPJ-14 32 32 100 100-2000
OPJ-19 22 21 95,50 100-2000
TOPLAM 771 769 99,74 100-3000
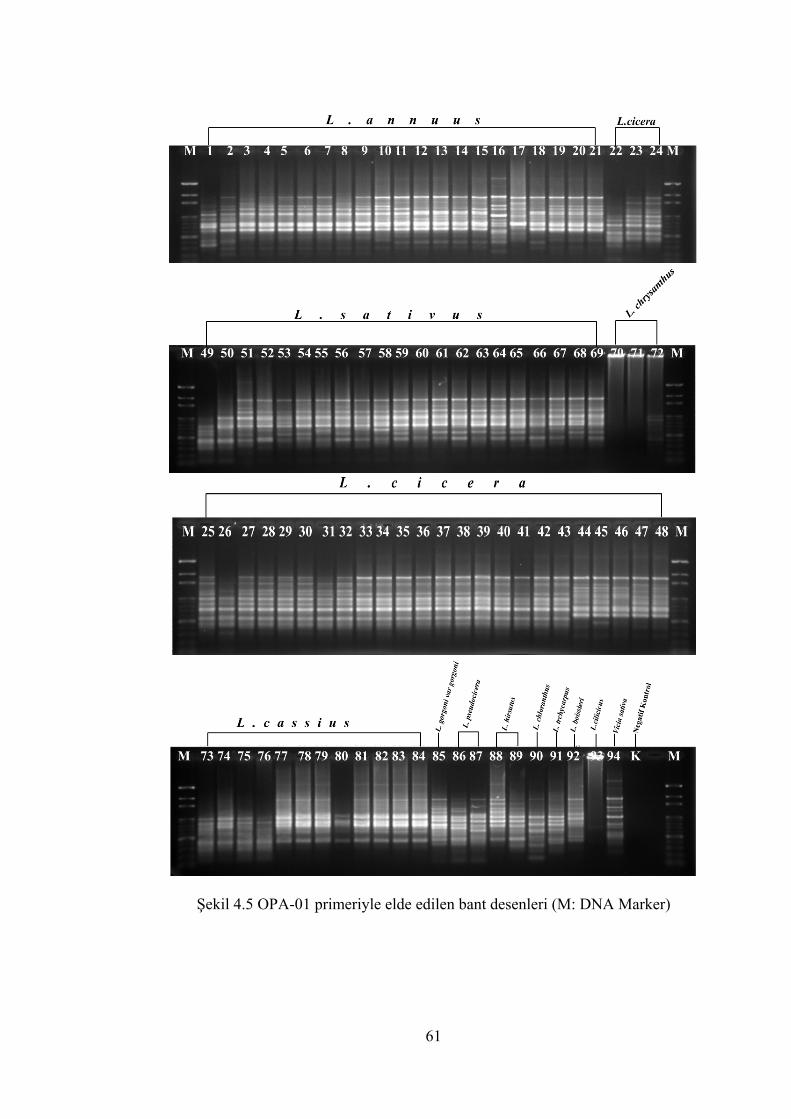
61
Şekil 4.5 OPA-01 primeriyle elde edilen bant desenleri (M: DNA Marker)
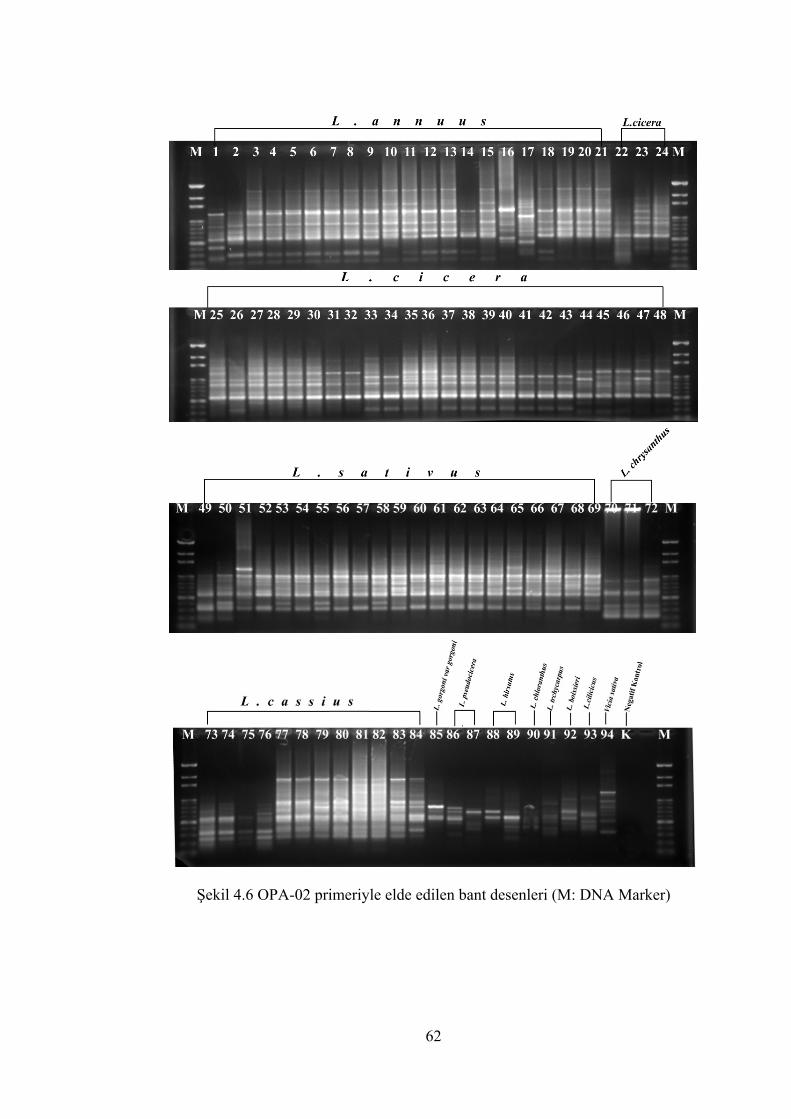
62
Şekil 4.6 OPA-02 primeriyle elde edilen bant desenleri (M: DNA Marker)
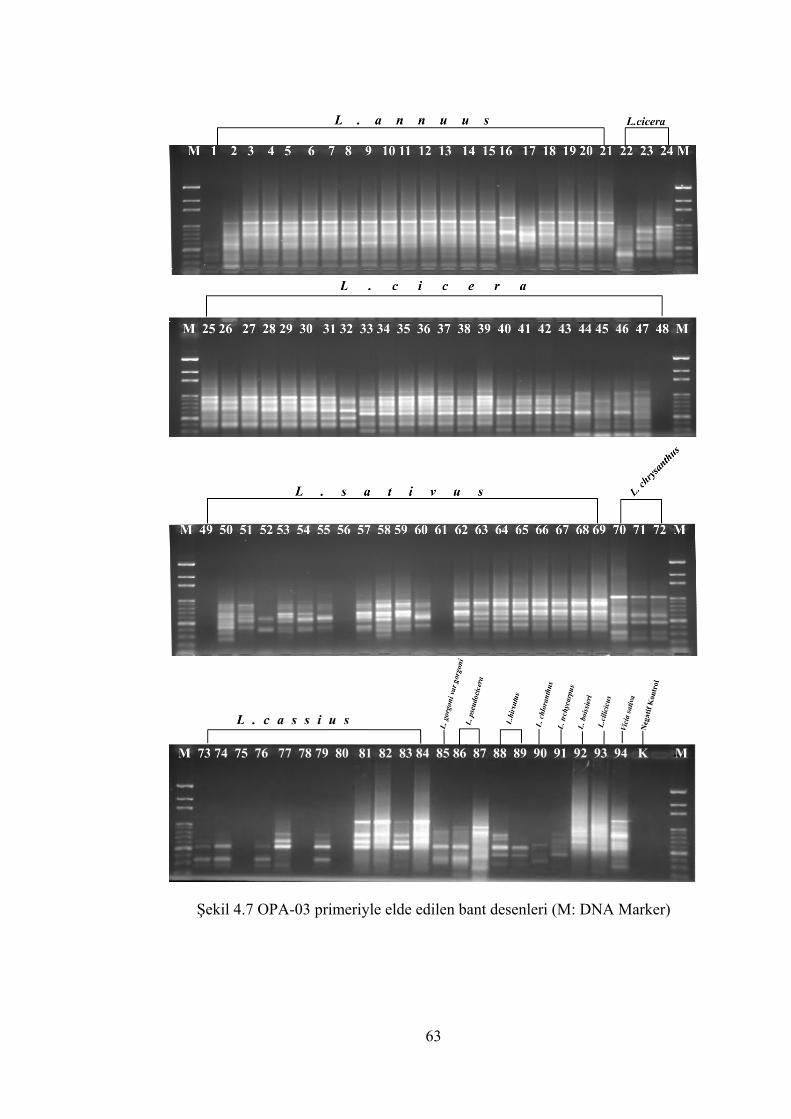
63
Şekil 4.7 OPA-03 primeriyle elde edilen bant desenleri (M: DNA Marker)
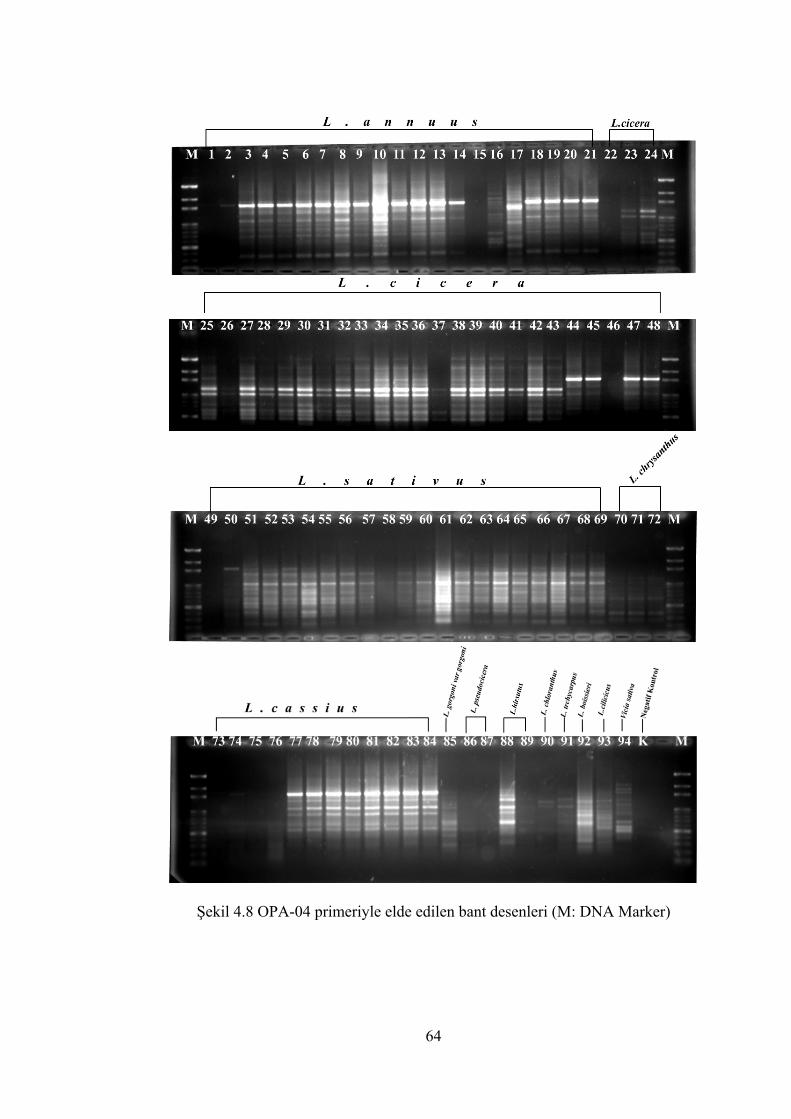
64
Şekil 4.8 OPA-04 primeriyle elde edilen bant desenleri (M: DNA Marker)
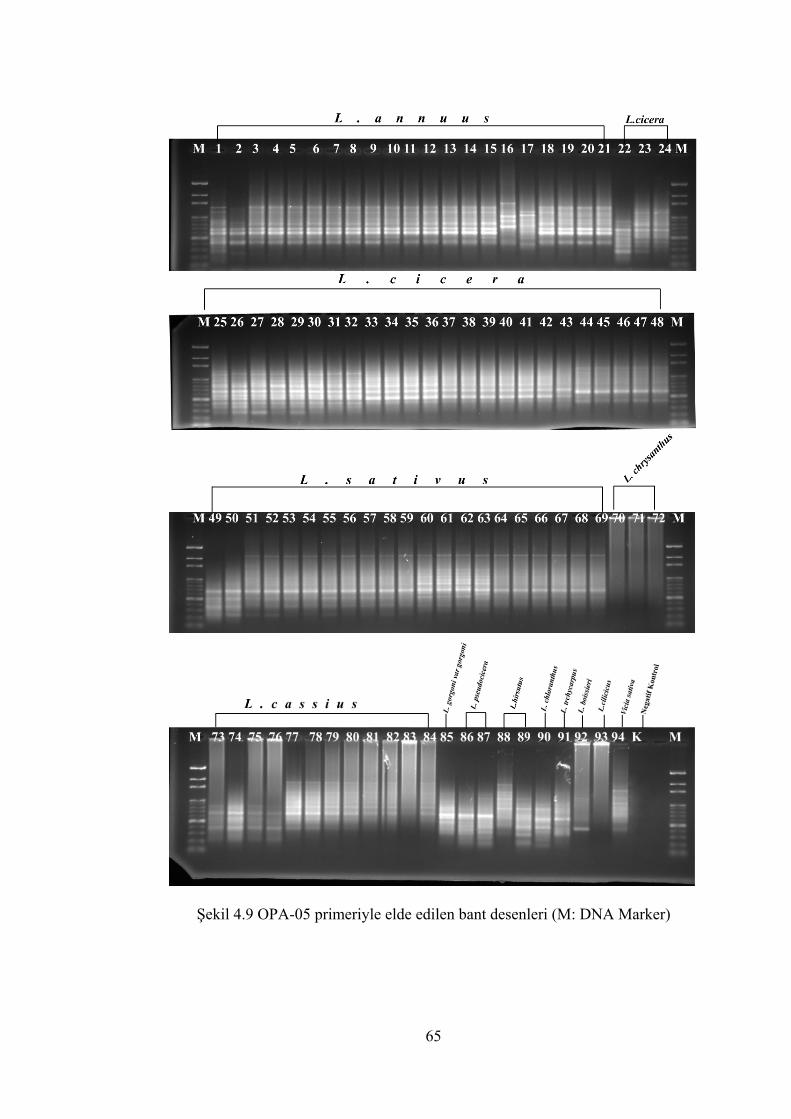
65
Şekil 4.9 OPA-05 primeriyle elde edilen bant desenleri (M: DNA Marker)
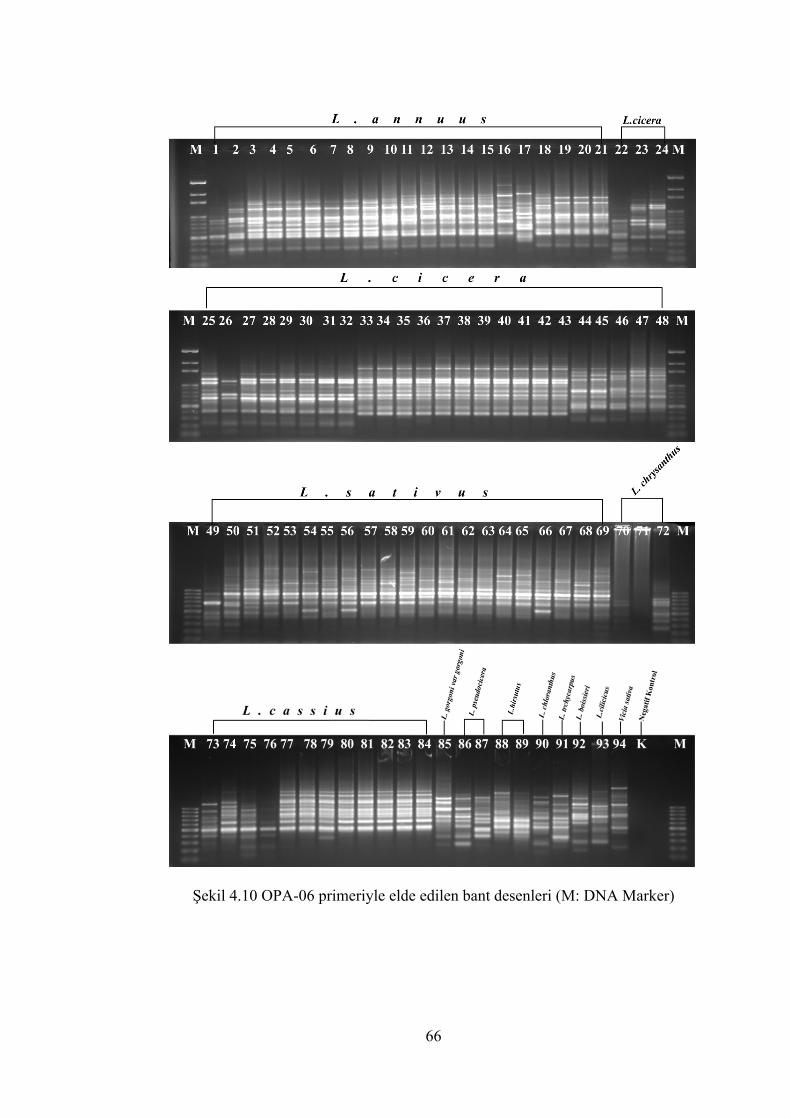
66
Şekil 4.10 OPA-06 primeriyle elde edilen bant desenleri (M: DNA Marker)
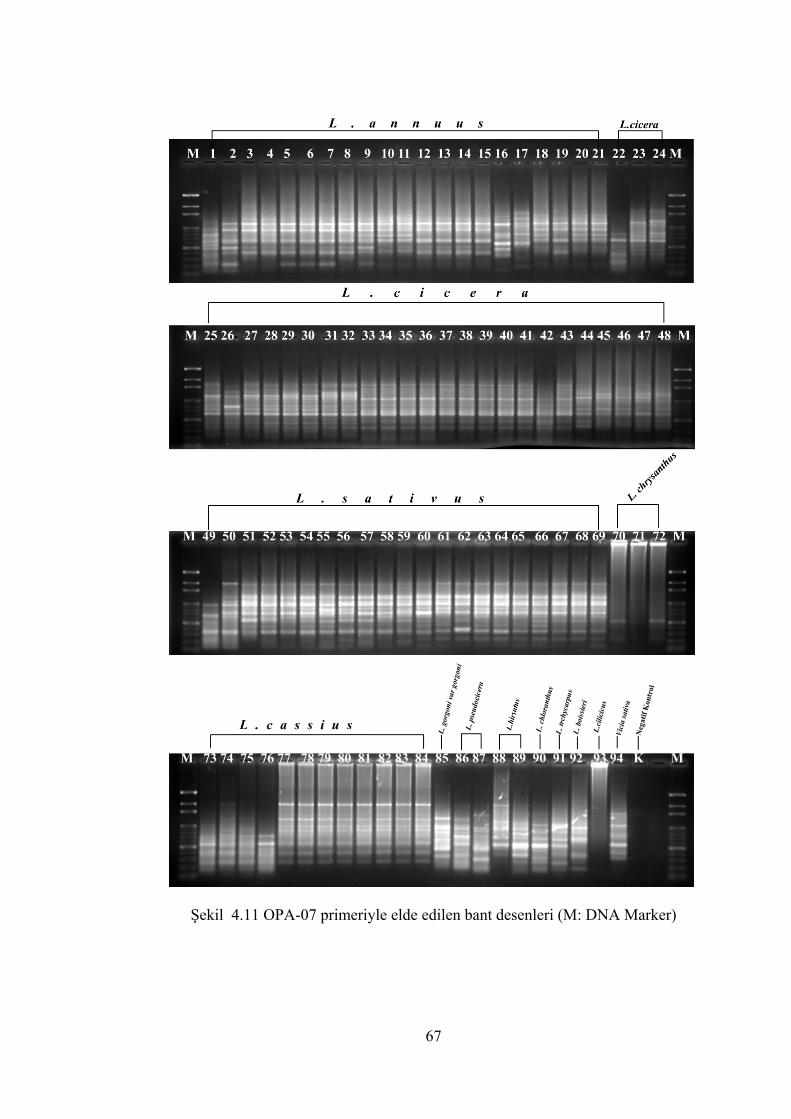
67
Şekil 4.11 OPA-07 primeriyle elde edilen bant desenleri (M: DNA Marker)
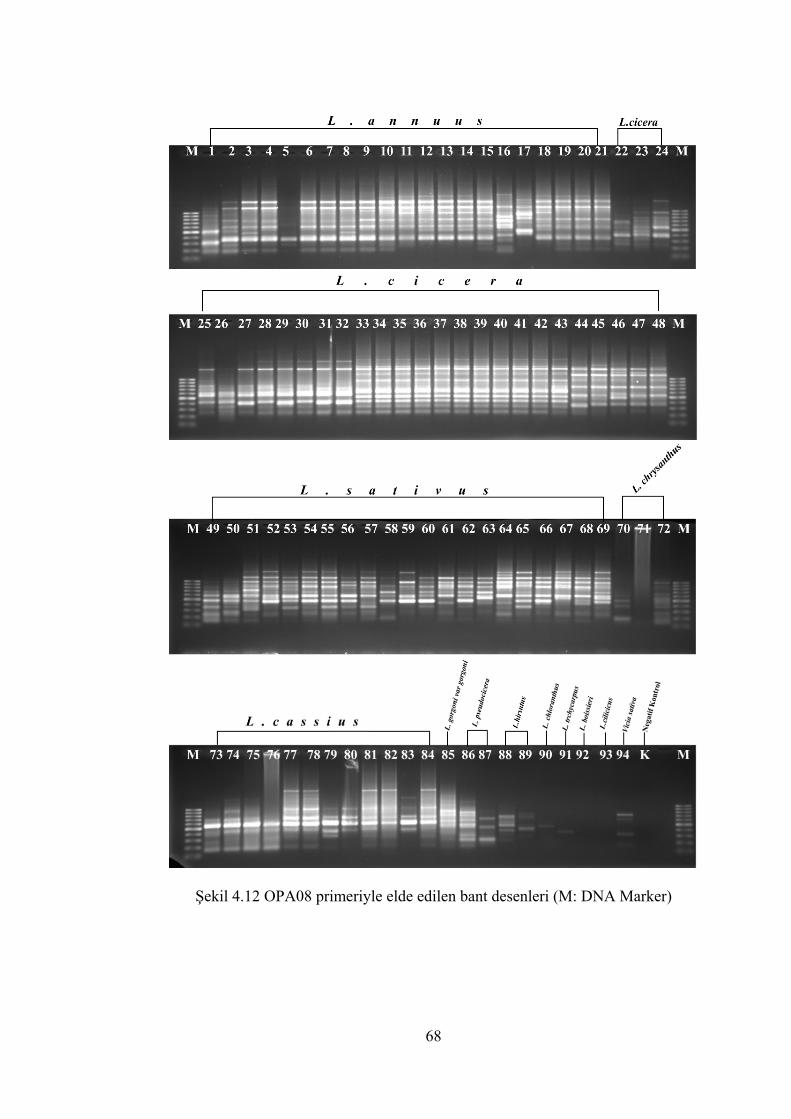
68
Şekil 4.12 OPA08 primeriyle elde edilen bant desenleri (M: DNA Marker)

69
Şekil 4.13 OPA09 primeriyle elde edilen bant desenleri (M: DNA Marker)
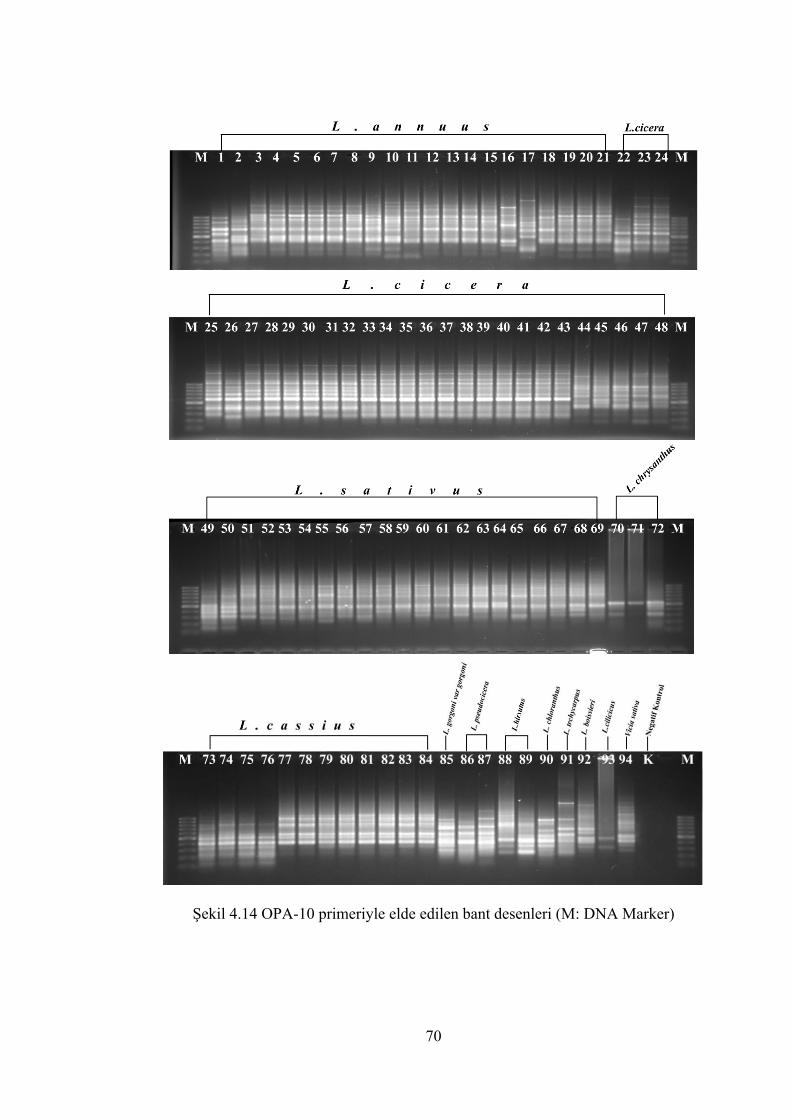
70
Şekil 4.14 OPA-10 primeriyle elde edilen bant desenleri (M: DNA Marker)
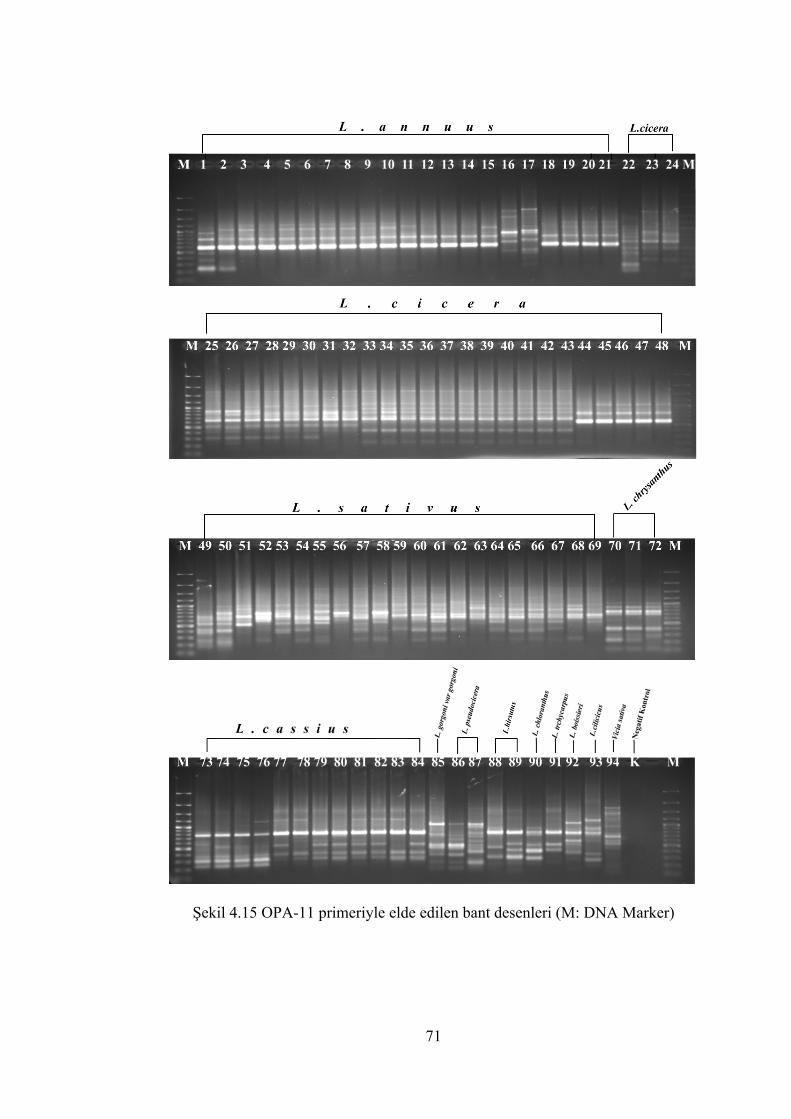
71
Şekil 4.15 OPA-11 primeriyle elde edilen bant desenleri (M: DNA Marker)

72
Şekil 4.16 OPA-15 primeriyle elde edilen bant desenleri (M: DNA Marker)
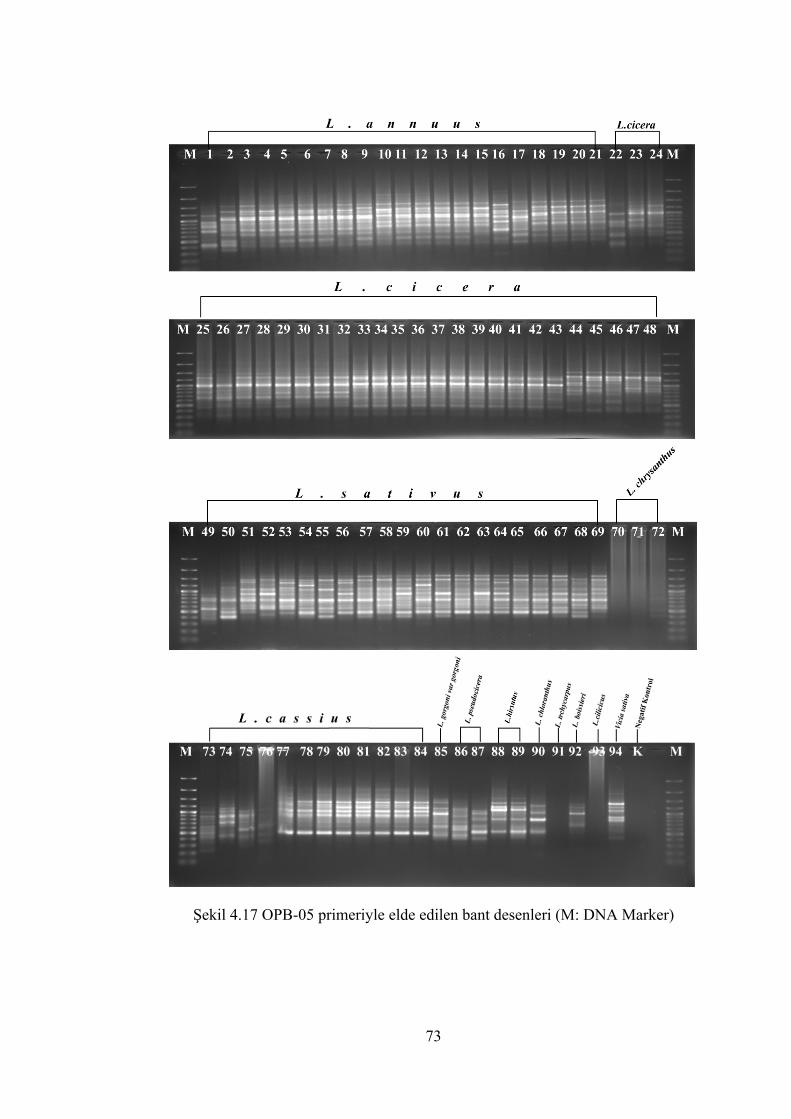
73
Şekil 4.17 OPB-05 primeriyle elde edilen bant desenleri (M: DNA Marker)
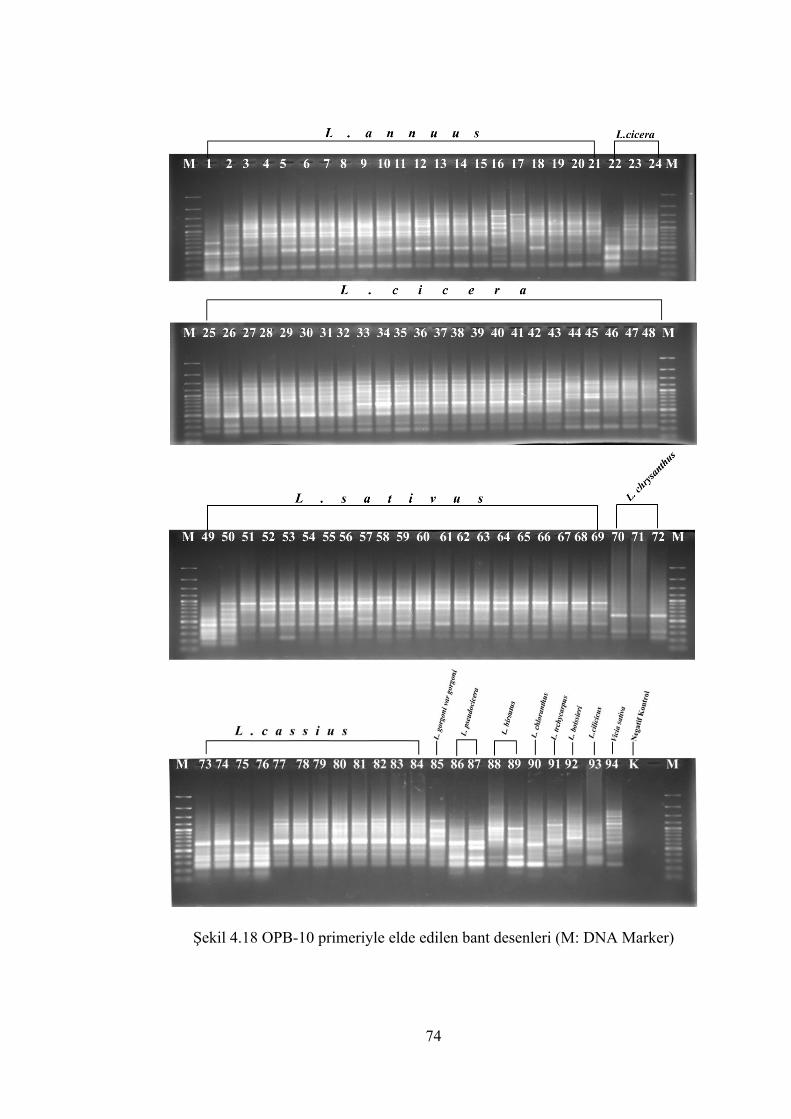
74
Şekil 4.18 OPB-10 primeriyle elde edilen bant desenleri (M: DNA Marker)
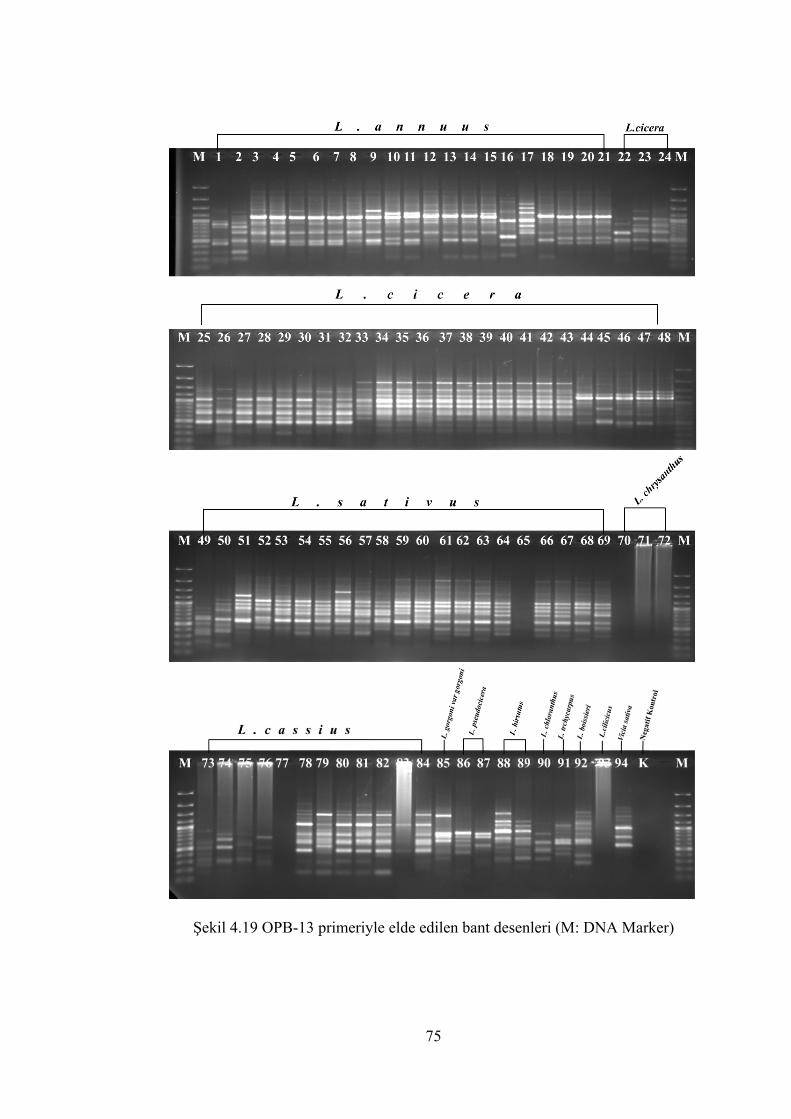
75
Şekil 4.19 OPB-13 primeriyle elde edilen bant desenleri (M: DNA Marker)
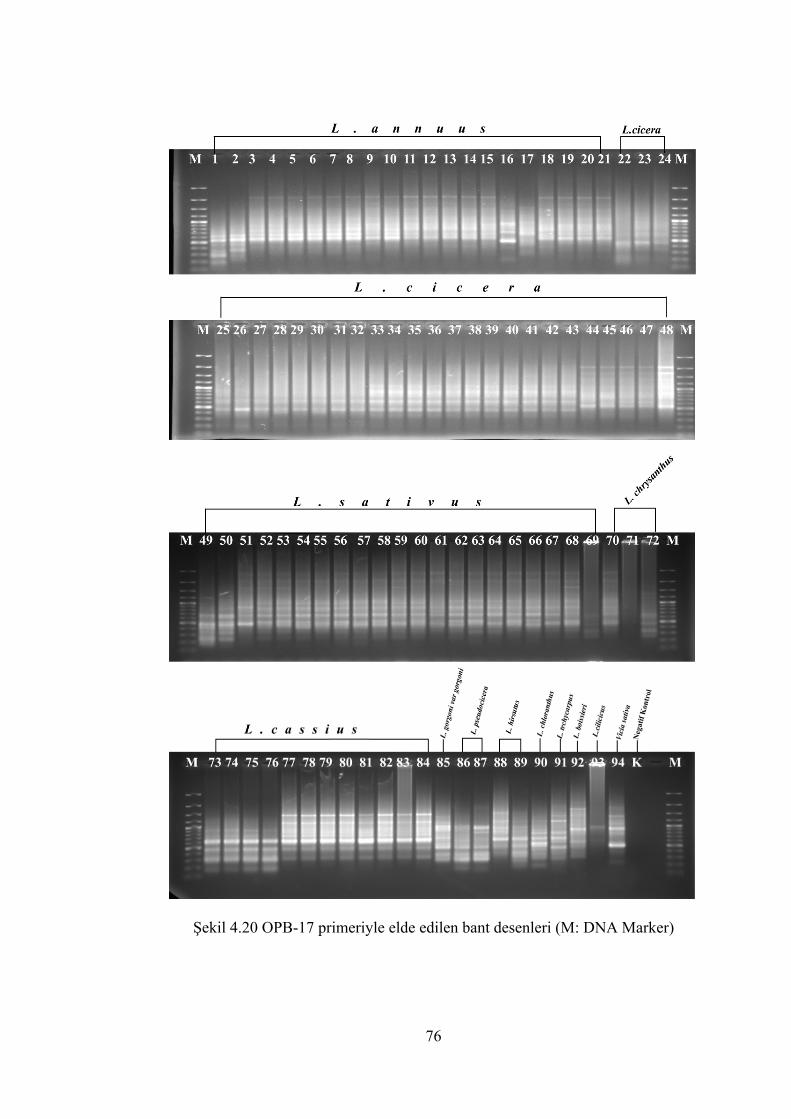
76
Şekil 4.20 OPB-17 primeriyle elde edilen bant desenleri (M: DNA Marker)

77
Şekil 4.21 OPD-03 primeriyle elde edilen bant desenleri (M: DNA Marker)
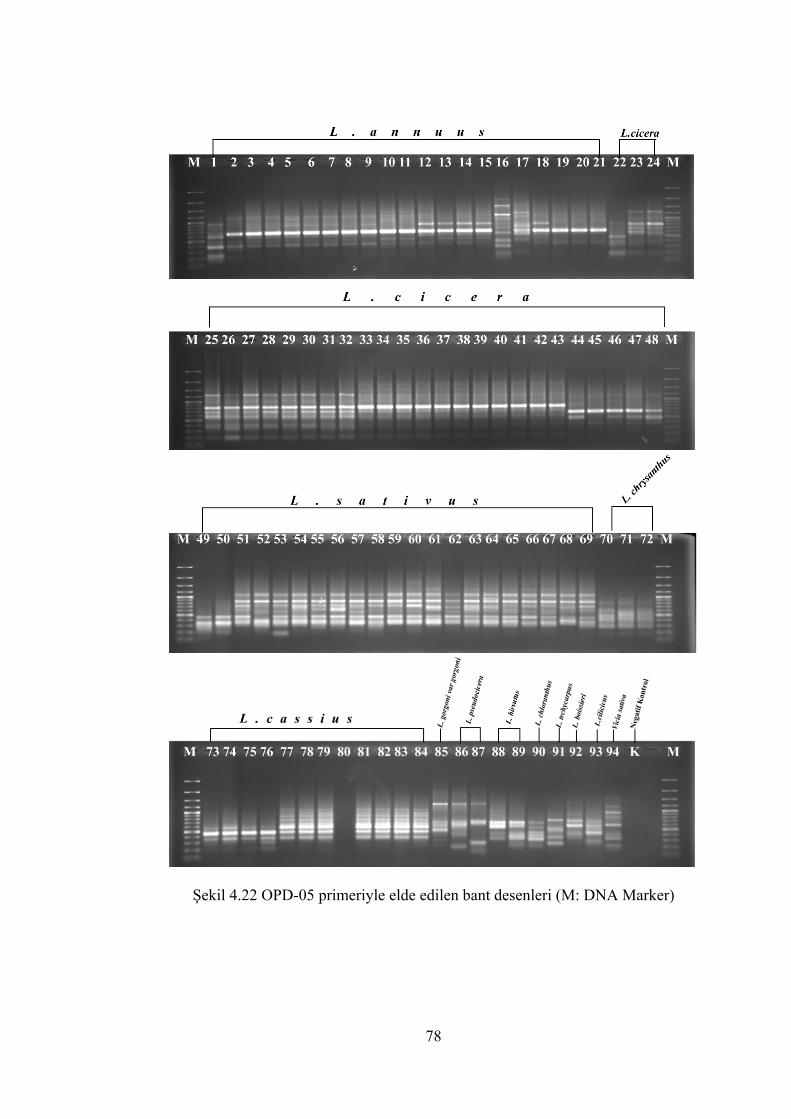
78
Şekil 4.22 OPD-05 primeriyle elde edilen bant desenleri (M: DNA Marker)
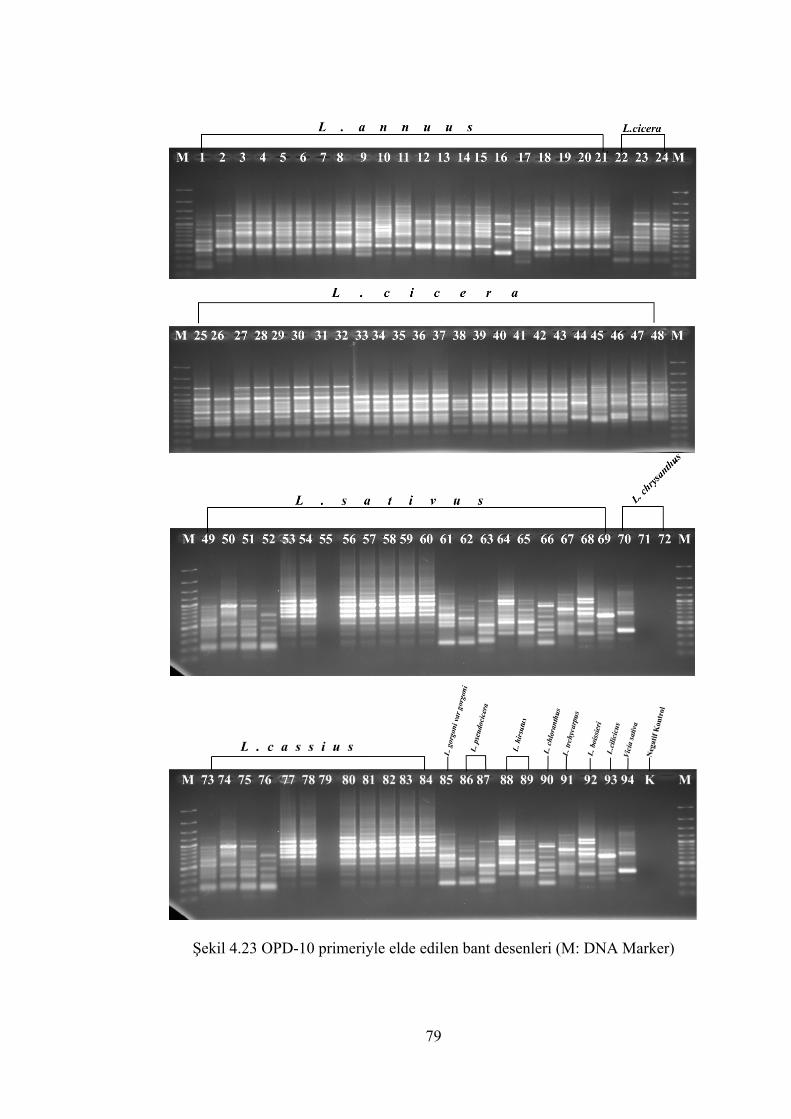
79
Şekil 4.23 OPD-10 primeriyle elde edilen bant desenleri (M: DNA Marker)
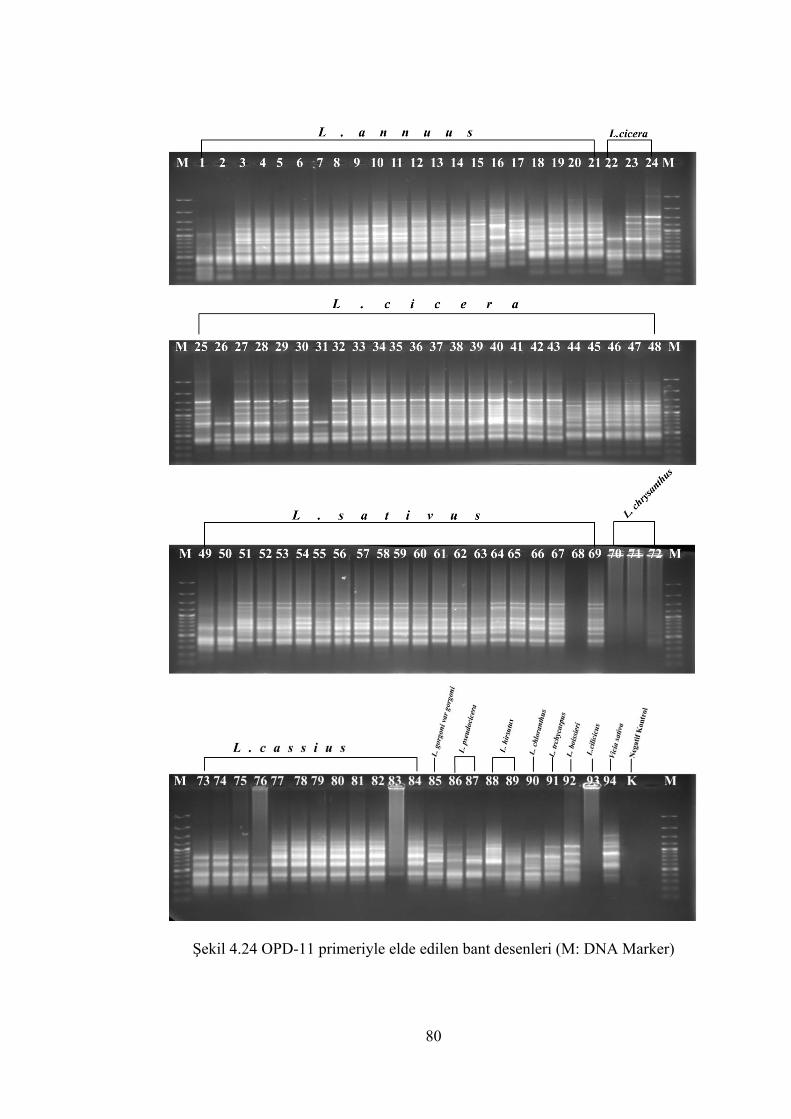
80
Şekil 4.24 OPD-11 primeriyle elde edilen bant desenleri (M: DNA Marker)
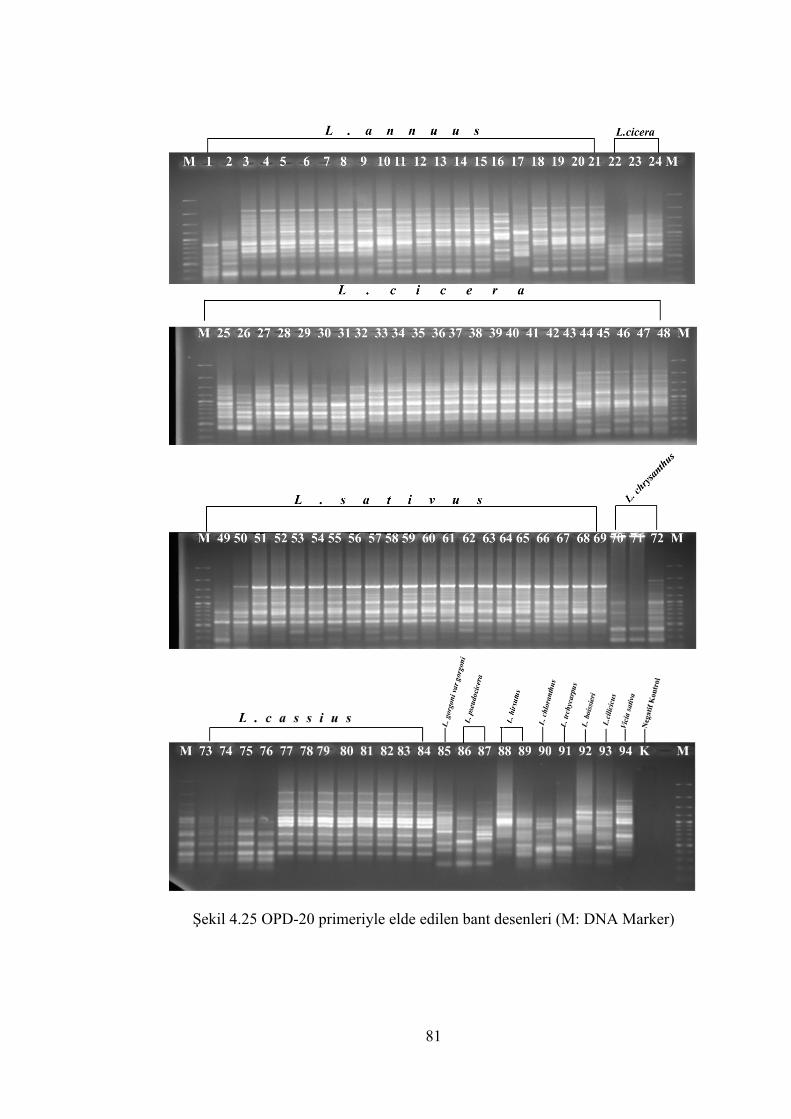
81
Şekil 4.25 OPD-20 primeriyle elde edilen bant desenleri (M: DNA Marker)
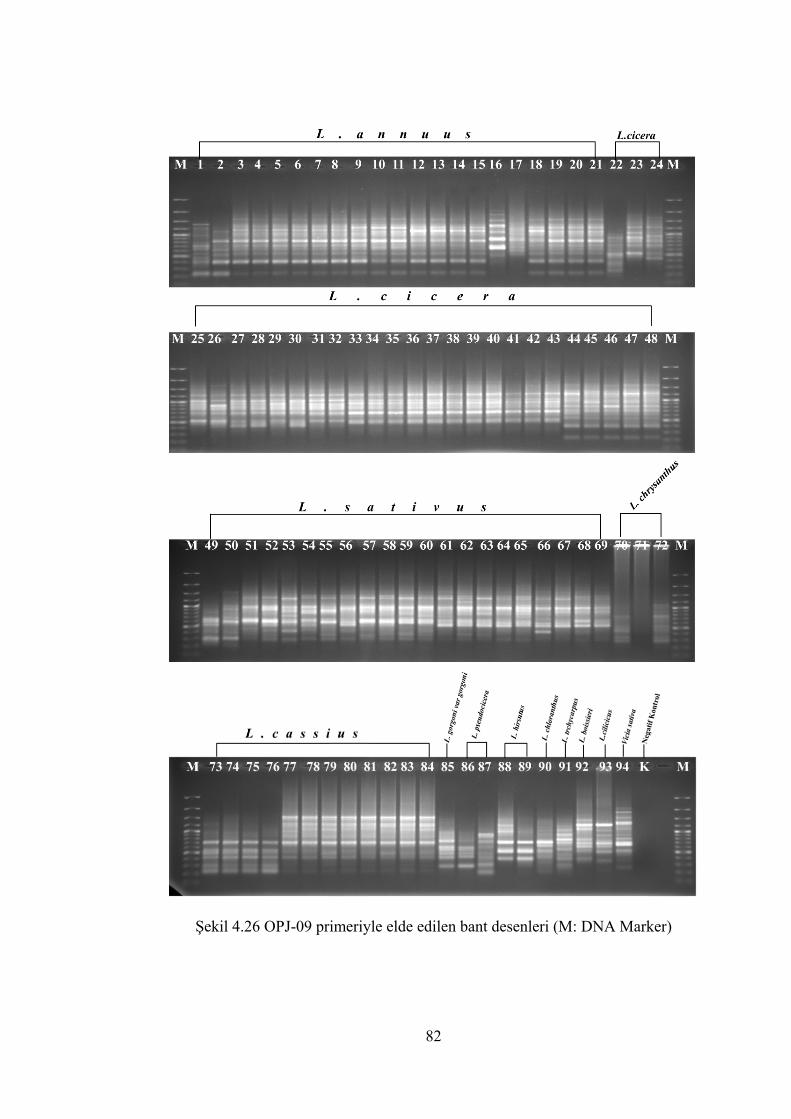
82
Şekil 4.26 OPJ-09 primeriyle elde edilen bant desenleri (M: DNA Marker)
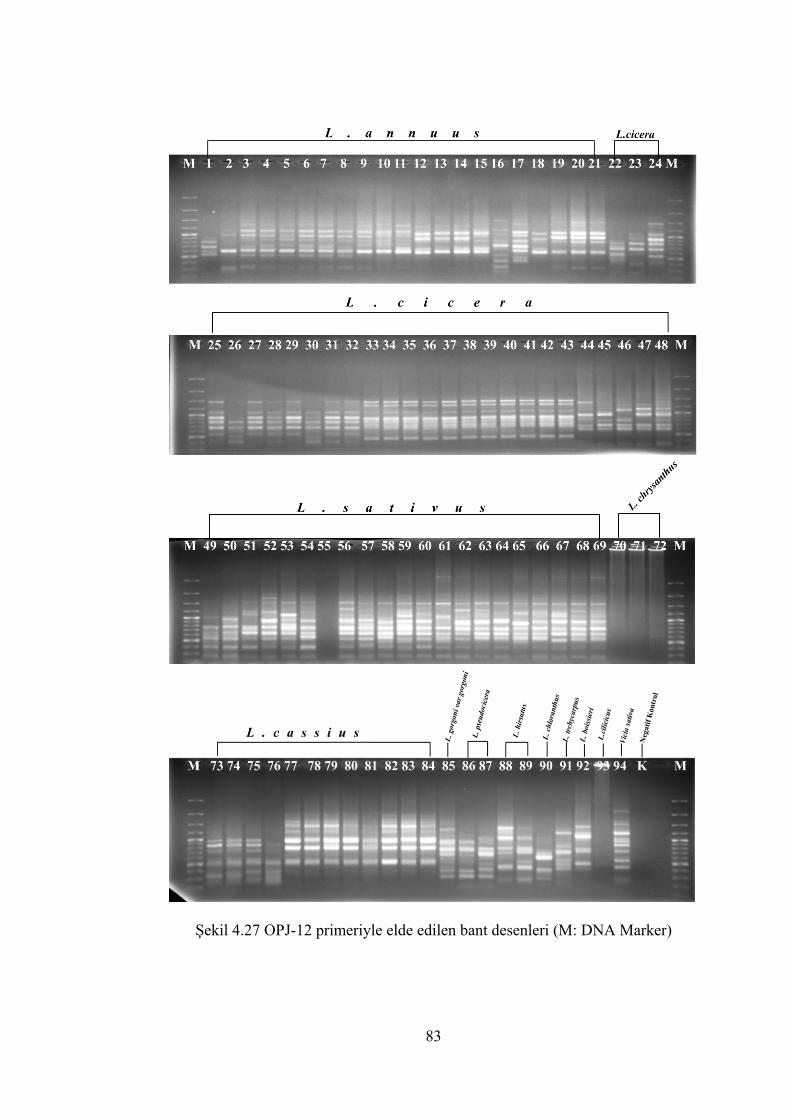
83
Şekil 4.27 OPJ-12 primeriyle elde edilen bant desenleri (M: DNA Marker)
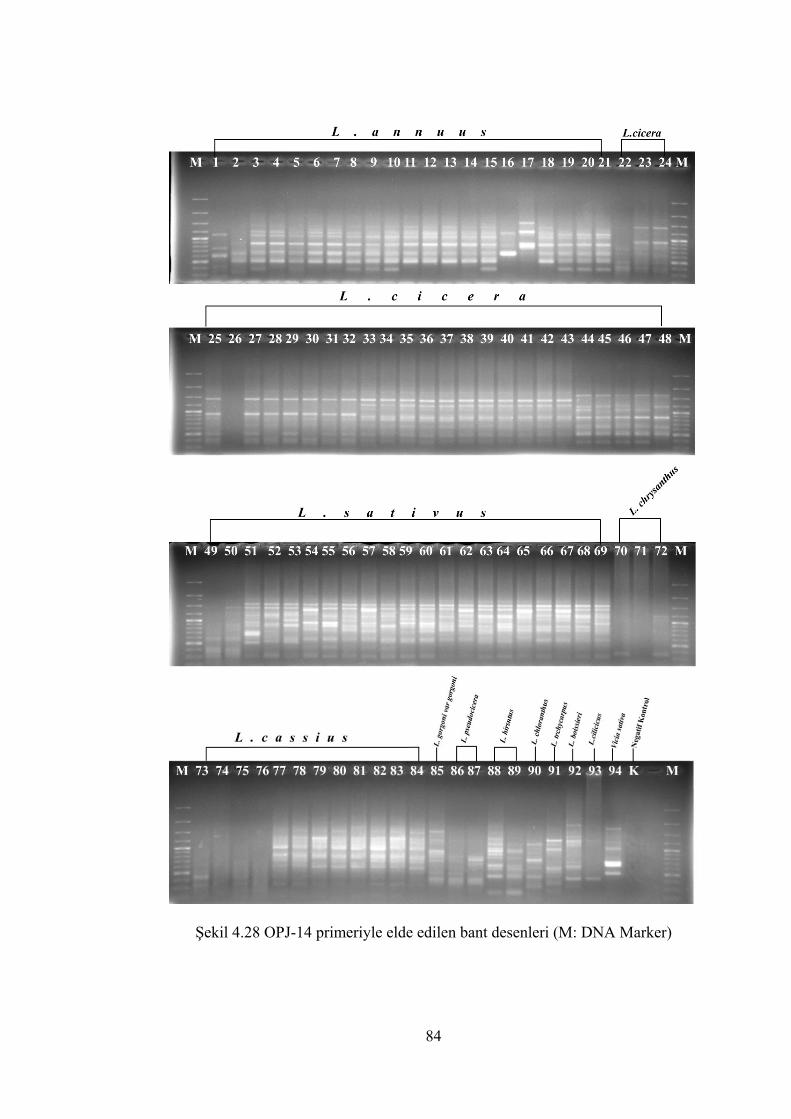
84
Şekil 4.28 OPJ-14 primeriyle elde edilen bant desenleri (M: DNA Marker)
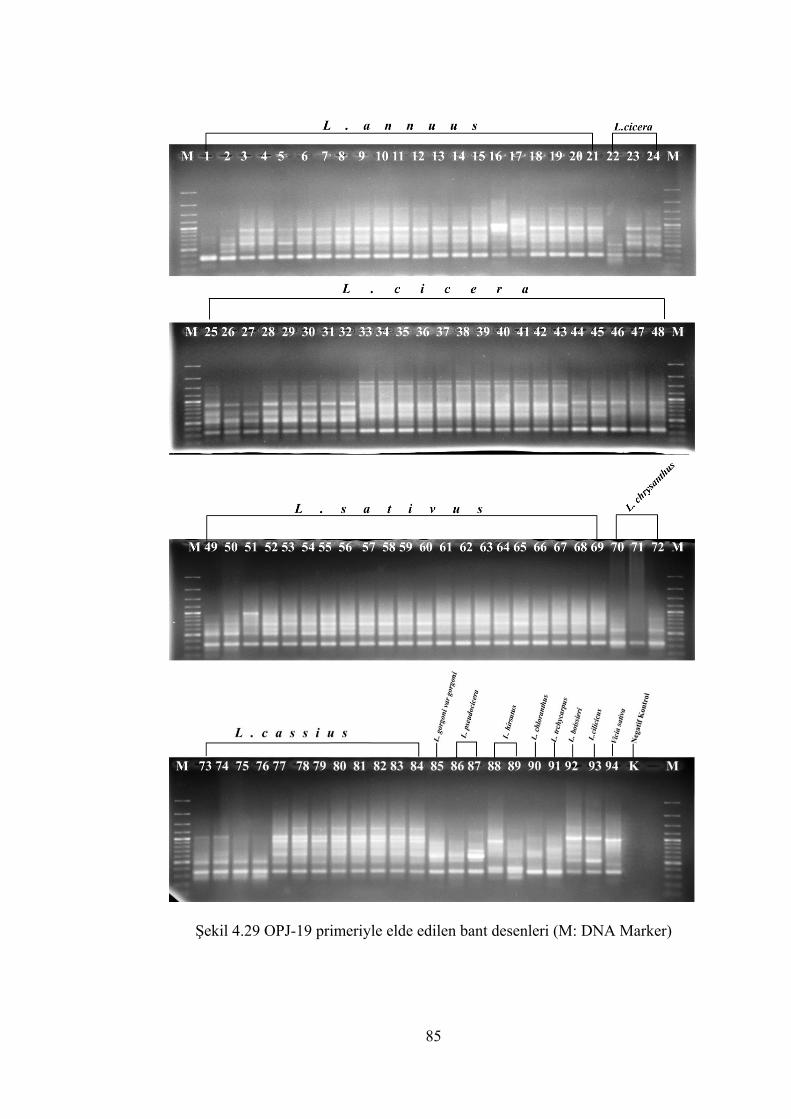
85
Şekil 4.29 OPJ-19 primeriyle elde edilen bant desenleri (M: DNA Marker)

86
Gözlenen tüm bantlar değerlendirmeye alınarak bant varlığına “1” yokluğuna ise “0”
değeri verilerek oluşturulan veri dosyası (Ek1) POPGENE Microsoft Window-based
Freeware for Population Genetics Analysis, Version 1.31 (Yeh vd. 1997) programına
yüklenerek genetik mesafe değerleri (Çizelge 4.3) belirlenmiştir. Bu değerler Nei (1978)
formülüne dayanmaktadır.
MEGA (Molecular Evolutionary Genetics Analysis) Versiyon 3.0 (Kumar vd. 2004)
programı ise genetik mesafe değerleri ile dendrogram [UPGMA (Unweighted Pair-
Group Method Arithmetic Average) ağacı (Şekil 4.30)] oluşturmak için kullanılmıştır.
Yapılan ilk analizlerde elde edilen bireyler arası filogenetik ilişkileri gösteren
dendrogramda (Şekil 4.30) ETAE’den temin edilmiş olan TR 500532 etiket nolu
Lathyrus cicera türünün, 7d nolu popülasyonuna ait kolların (Çizelge 3.4’deki gri
kutularla belirtilen) diğer L. cicera kollarıyla beraber olması beklenirken, L. annuus
kollarıyla beraber çıktığı görülmektedir. Aynı durum SDS-PAGE verilerinin analizinde
de gözlenmiştir. Bu sebeple, 44, 45, 46, 47, 48 birey sıra nolu örneklere (Çizelge 3.4) ait
veriler çalışmadan çıkarılmıştır. Ayrıca ikinci seksiyon dışı grup olması amacıyla
kullanılan L. cilicicus’a ait 93 birey sıra nolu örnek de (Çizelge 3.4), bir adet seksiyon
dışı grubun yeterli olacağı düşünülerek çalışmadan çıkarılmıştır.
Daha sonra bireylerin kendi türleriyle birleştirilerek elde edilen genetik mesafe
değerlerine baktığımızda (Çizelge 4.3) çalışmadaki türler arasındaki en yakın genetik
mesafenin 0.0859 ile L. pseudo-cicera ile L. gorgoni var. gorgoni arasında, en uzak
genetik mesafenin ise 0.3459 ile Vicia sativa ile L. cicera arasında olduğu
görülmektedir. En uzak mesafenin Vicia sativa ile olması beklenen bir durumdur.
Çünkü Vicia sativa çalışmada dış grup olarak kullanılmıştır. Aynı şekilde L. boissieri de
Cicercula seksiyonu dışından dış grup olarak kullanılmıştır ve gerçekten de diğer
türlerden genetik mesafe olarak uzakta çıkmıştır. Çalışmadaki Cicercula seksiyonu
üyelerinin kendi içlerindeki en uzak mesafe ise L.cicera ile L.chloranthus arasındaki
0,2816 değeridir (Çizelge 4.3).

87
Çizelge 4.3’teki genetik mesafe değerlerine göre oluşturulmuş olan dendrogram
incelendiğinde, çalışmaya alınan türler, Vicia sativa ve L. boissieri dış grupları hariç
olmak üzere, iki ana grup altında toplanmaktadır (Şekil 4.31):
1. Ana grup L. cicera ve L. sativus dallarından meydana gelmiştir.
2. Ana grup kendi içinde 2 alt gruba ayrılmıştır. Bu alt gruplardan birisini L. annuus dalı
oluşturmuştur. Diğer alt grup yine iki alt gruba ayrılmıştır. Bu alt gruplardan bir kol
Lathyrus gorgoni var. gorgoni ve Lathyrus pseudo- cicera dallarına ayrılırken, diğer kol
yine iki gruba ayrılmıştır. Bu gruplardan birini Lathyrus cassius ve Lathyrus hirsutus
dalları oluştururken diğer grubu Lathyrus chrysanthus, Lathyrus chloranthus ve
Lathyrus trachycarpus dalları meydana getirmiştir.
Çalışmada cins dışı grup olarak kullanılan Vicia sativa beklendiği gibi tüm bu grupların
dışında yer almıştır. Daha önceki çalışmalarda (Davis 1970) Lathyrostylis (Platystylis)
seksiyonuna dahil edilmiş olan ve çalışmada seksiyon dışı grup olarak kullanılan
Lathyrus boissieri de beklendiği gibi Vicia sativa ile diğer türler arasında yer almıştır.

88
L. annuus 1a1 L. cicera 7a1 L. sativus 8a1 L. sativus 8a2 L. cassius 3a1 L. cassius 3a2 L. cassius 3a3 L. cassius 3a4 L. chloranthus 16a1 L. trachycarpus 17a1 L. chrysanthus 15a1 L. chrysanthus 15a2 L. chrysanthus 15a3 L. cilicicus 19c2 L. gorgoni var gorgoni 14a1 L. pseudo cicera 6a1 L. pseudo cicera 6b1 L. hirsutus 14a1 L. hirsutus 14b1 L. boissieri dg4 L. cassius 3b1 L. cassius 3b2 L. cassius 3b5 L. cassius 3b6 L. cassius 3b7 L. cassius 3b8 L. cassius 3b4 L. cassius 3b3 L. sativus 8b1 L. sativus 8b2 L. sativus 8b3 L. sativus 8b4 L. sativus 8b6 L. sativus 8b7 L. sativus 8b8 L. sativus 8b5 L. sativus 8b9 L. sativus 8b10 L. sativus 8c1 L. sativus 8c2 L. sativus 8c3 L. sativus 8c4 L. sativus 8d1 L. sativus 8d2 L. sativus 8d4 L. sativus 8c5 L. sativus 8d3 L. cicera 7b1 L. cicera 7b2 L. cicera 7b3 L. cicera 7b5 L. cicera 7b6 L. cicera 7b8 L. cicera 7b7 L. cicera 7b9 L. cicera 7b10 L. cicera 7b4 L. cicera 7c1 L. cicera 7c2 L. cicera 7c3 L. cicera 7c4 L. cicera 7c5 L. cicera 7c6 L. cicera 7c7 L. cicera 7c8 L. cicera 7c9 L. cicera 7c10 L. cicera 7c11 L. annuus 1b1 L. annuus 1c1 L. annuus 1c2 L. annuus 1c6 L. annuus 1c3 L. annuus 1c4 L. annuus 1c5 L. annuus 1c7 L. annuus 1d1 L. annuus 1d2 L. annuus 1d3 L. annuus 1d4 L. annuus 1d5 L. annuus 1d6 L. annuus 1d9 L. annuus 1e1 L. annuus 1e2 L. annuus 1e3 L. cicera 7d1 L. cicera 7d2 L. cicera 7d3 L. cicera 7d4 L. cicera 7d5 L. annuus 1d7 L. annuus 1d8 Vicia sativa 20 a5
5
Şekil 4.30 Çalışmada kullanılan bireylerin UPGMA dendrogramı (Nei 1978)
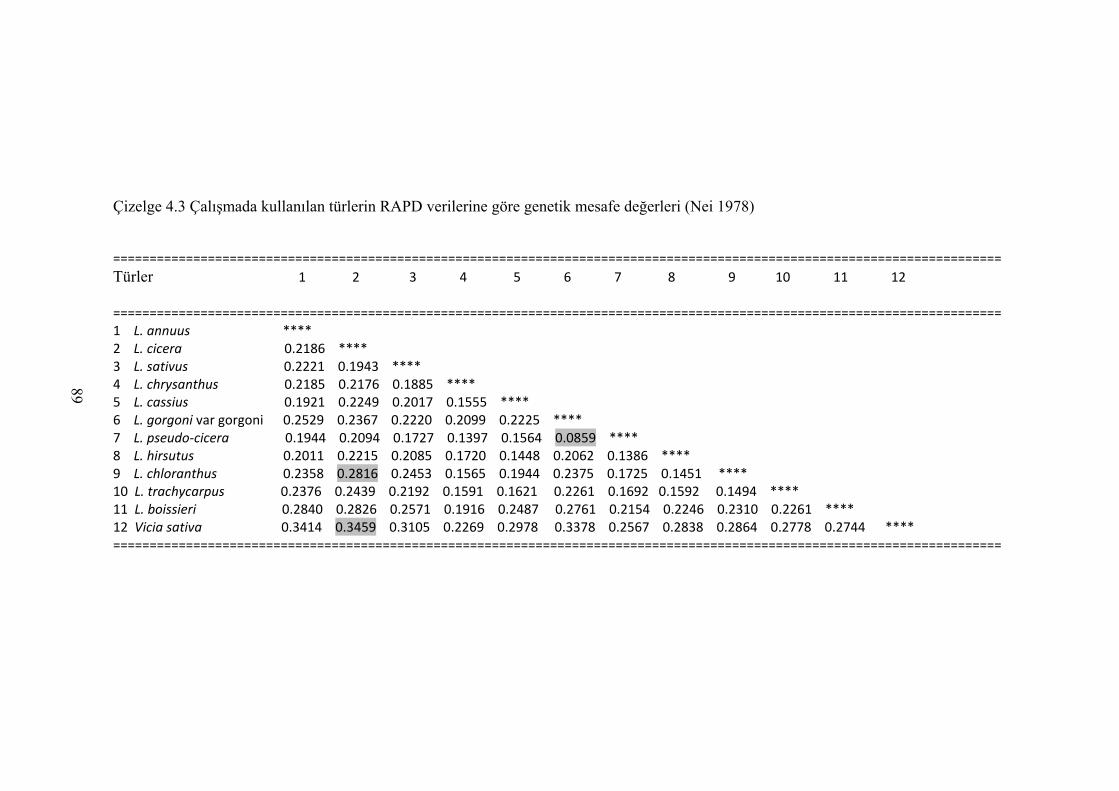
89
Çizelge 4.3 Çalışmada kullanılan türlerin RAPD verilerine göre genetik mesafe değerleri (Nei 1978)
========================================================================================================================== Türler 1 2 3 4 5 6 7 8 9 10 11 12 ========================================================================================================================== 1 L. annuus **** 2 L. cicera 0.2186 **** 3 L. sativus 0.2221 0.1943 **** 4 L. chrysanthus 0.2185 0.2176 0.1885 **** 5 L. cassius 0.1921 0.2249 0.2017 0.1555 **** 6 L. gorgoni var gorgoni 0.2529 0.2367 0.2220 0.2099 0.2225 **** 7 L. pseudo‐cicera 0.1944 0.2094 0.1727 0.1397 0.1564 0.0859 **** 8 L. hirsutus 0.2011 0.2215 0.2085 0.1720 0.1448 0.2062 0.1386 **** 9 L. chloranthus 0.2358 0.2816 0.2453 0.1565 0.1944 0.2375 0.1725 0.1451 **** 10 L. trachycarpus 0.2376 0.2439 0.2192 0.1591 0.1621 0.2261 0.1692 0.1592 0.1494 **** 11 L. boissieri 0.2840 0.2826 0.2571 0.1916 0.2487 0.2761 0.2154 0.2246 0.2310 0.2261 **** 12 Vicia sativa 0.3414 0.3459 0.3105 0.2269 0.2978 0.3378 0.2567 0.2838 0.2864 0.2778 0.2744 **** ==========================================================================================================================

90
L. cicera
L. sativus1. ANA GRUP
1. ALT GRUP L. annuus
L. cassiius
L. hirsutus
L. chrysanthus
L. chloranthus
L. trachycarpus
L. gorgoni var gorgoni
L. pseudo cicera
2. ALT GRUP2. ANA GRUP
SEKSİYON DIŞI GRUP L. boissieri
CİNS DIŞI GRUP Vicia sativa
2 Şekil 4.31 Çalışmada kullanılan türlerin RAPD verileriyle elde edilen genetik mesafe değerleriyle oluşturulmuş UPGMA dendrogram
(Nei 1978)

91
4.1.3.2 Tür içi genetik mesafeler
Çalışmada birey sayısı birden fazla olan popülasyonların tür içi genetik varyasyon
analizleri yapılmıştır. Birey sayısı birden fazla olan popülasyonlar L. annuus türü için
1c, 1d,1e, L. cicera için 7b, 7c, L. sativus için 8a, 8b, 8c, 8d, L. cassius için 3a ve 3b
popülasyonlarıdır. Bu analizlerde yine POPGENE Microsoft Window-based Freeware
for Population Genetics Analysis, Version 1.31 (Yeh vd. 1997) programı kullanılmıştır.
Bu program ile yukarıda belirtilen popülasyonlar için aşağıdaki değerler elde edilmiştir:
Na : Her örnek için gözlenen allel sayısı
Ne : Etkili allel sayısı
Nm : Gen akışı
H : Nei’nin genetik çeşitliliği (1987)
I : Shannon information indeksi
GST : Alt populasyonlar arasındaki genetik farklılaşmanın nisbi büyüklüğü
D : Nei’nin genetik mesafe değeri (1978)
L. annuus popülasyonları
L. annuus popülasyonlarının POPGENE [Microsoft Window-based Freeware for
Population Genetics Analysis, Version 1.31 (Yeh vd. 1997)] programı ile yapılan
genetik varyasyon (Nei 1987) analizleri çizelge 4.4’de, genetik farklılaşma (Nei 1987)
değerleri çizelge 4.5 de, genetik mesafe değerleri çizelge 4.6’da ve genetik mesafe
verileriyle elde edilen dendrogram şekil 4.31’de gösterilmiştir.
L. annuus popülasyonları için hesaplanan genetik çeşitlilik değerlerine bakıldığında, en
düşük değerin H= 0,0042 (polimorfik lokus yüzdesi, %P=0,91) ile 1e (Balıkesir)
popülasyonunda, en yüksek değerin ise H= 0,1534 (%P=51,88) ile 1d (Gaziantep)
popülasyonunda olduğu görülmüştür. Nei’ye (1987) göre L. annuus popülasyonları için
hesaplanan ortalama genetik çeşitlilik değeri H= 0,1468 (%P=55,64)’dir (Çizelge 4.4).
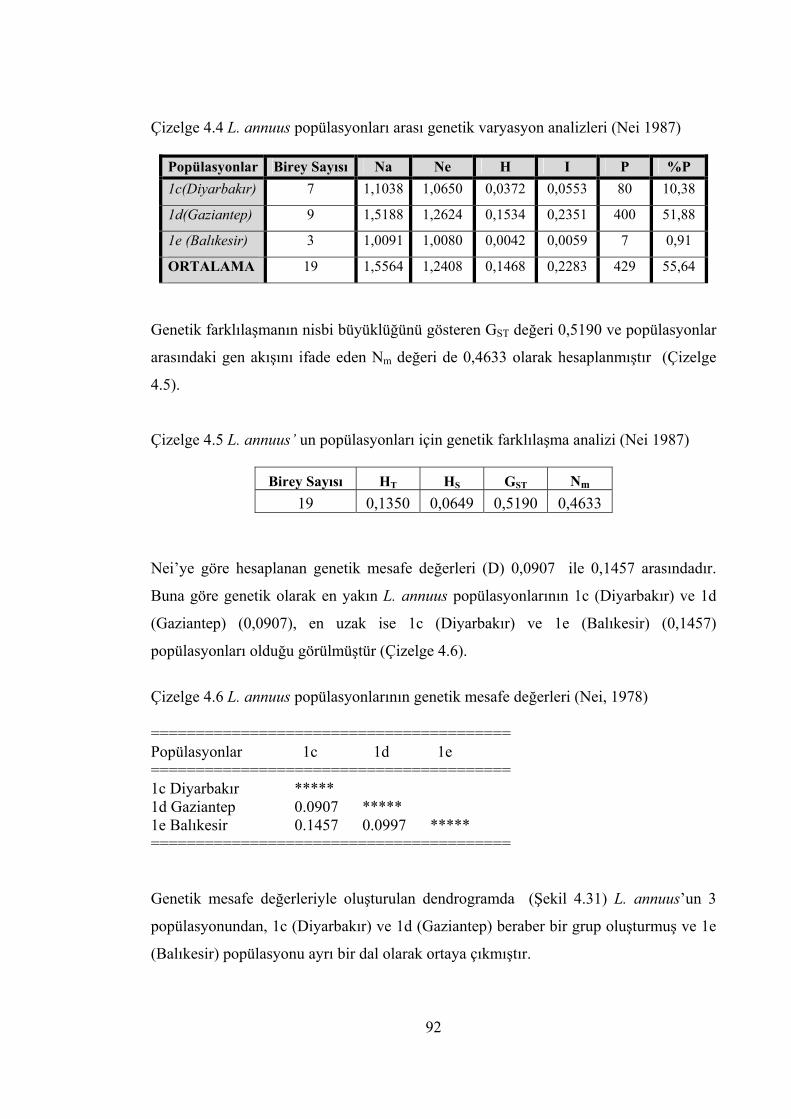
92
Çizelge 4.4 L. annuus popülasyonları arası genetik varyasyon analizleri (Nei 1987)
Popülasyonlar Birey Sayısı Na Ne H I P %P 1c(Diyarbakır) 7 1,1038 1,0650 0,0372 0,0553 80 10,38
1d(Gaziantep) 9 1,5188 1,2624 0,1534 0,2351 400 51,88
1e (Balıkesir) 3 1,0091 1,0080 0,0042 0,0059 7 0,91
ORTALAMA 19 1,5564 1,2408 0,1468 0,2283 429 55,64
Genetik farklılaşmanın nisbi büyüklüğünü gösteren GST değeri 0,5190 ve popülasyonlar
arasındaki gen akışını ifade eden Nm değeri de 0,4633 olarak hesaplanmıştır (Çizelge
4.5).
Çizelge 4.5 L. annuus’ un popülasyonları için genetik farklılaşma analizi (Nei 1987)
Birey Sayısı HT HS GST Nm
19 0,1350 0,0649 0,5190 0,4633
Nei’ye göre hesaplanan genetik mesafe değerleri (D) 0,0907 ile 0,1457 arasındadır.
Buna göre genetik olarak en yakın L. annuus popülasyonlarının 1c (Diyarbakır) ve 1d
(Gaziantep) (0,0907), en uzak ise 1c (Diyarbakır) ve 1e (Balıkesir) (0,1457)
popülasyonları olduğu görülmüştür (Çizelge 4.6).
Çizelge 4.6 L. annuus popülasyonlarının genetik mesafe değerleri (Nei, 1978) ======================================== Popülasyonlar 1c 1d 1e ======================================== 1c Diyarbakır ***** 1d Gaziantep 0.0907 ***** 1e Balıkesir 0.1457 0.0997 ***** ======================================== Genetik mesafe değerleriyle oluşturulan dendrogramda (Şekil 4.31) L. annuus’un 3
popülasyonundan, 1c (Diyarbakır) ve 1d (Gaziantep) beraber bir grup oluşturmuş ve 1e
(Balıkesir) popülasyonu ayrı bir dal olarak ortaya çıkmıştır.

93
1c Diyarbakır 1d Gaziantep 1e Balıkesir
1 Şekil 4.32 L. annuus popülasyonlarının genetik mesafe verilerine göre (Nei, 1978)
çıkarılan dendrogramı
L. cicera popülasyonları
L. cicera popülasyonları için hesaplanan genetik çeşitlilik değerlerine bakıldığında, en
düşük değerin H= 0,0180 ( %P=5,06) ile 7c (İzmir) popülasyonunda , en yüksek değerin
ise H= 0,0682 (%P=20,10) ile 7b (Diyarbakır) popülasyonunda olduğu görülmüştür
(Çizelge 4.7).
Nei’ye (1987) göre L. cicera popülasyonları için hesaplanan ortalama genetik çeşitlilik
değeri H= 0,1161 (%P=32,56)’dir (Çizelge 4.8).
Çizelge 4.7 L. cicera popülasyonlarının genetik varyasyon analizleri (Nei 1987)
Popülasyonlar Birey Sayısı Na Ne H I P %P 7b(Diyarbakır) 10 1,2010 1,1185 0,0682 0,1022 155 20,10
7c (İzmir) 11 1,0506 1,0301 0,0180 0,0268 39 5,06
ORTALAMA 21 1,3256 1,2083 0,1161 0,1712 251 32,56
Genetik farklılaşmanın nisbi büyüklüğünü gösteren GST değeri 0,6330 ve popülasyonlar
arasındaki gen akışını ifade eden Nm değeri 0,2899 olarak hesaplanmıştır (Çizelge 4.8).
Çizelge 4.8 L. cicera’ nın populasyonları için genetik farklılaşma analizi (Nei 1987)
Birey Sayısı HT HS GST Nm 21 0,1174 0,0431 0,6330 0,2899
94
Nei’ye (1978) göre hesaplanan, iki popülasyon arasındaki genetik mesafe değeri (D)
0,1662’dir (Çizelge 4.9).
Çizelge 4.9 L. cicera popülasyonlarının genetik mesafe değerleri (Nei 1978) ============================== Popülasyonlar 7b 7c ============================== 7b Diyarbakır **** 7c İzmir 0.1662 **** ============================== L. sativus popülasyonları
L. sativus popülasyonları için hesaplanan genetik çeşitlilik değerleri incelendiğinde, en
düşük değer H= 0,0301 ( %P=7,26) ile 8a (Batman) popülasyonunda , en yüksek değer
ise H= 0,0840 (%P=27,89) ile 8b (Diyarbakır) popülasyonunda görülmüştür (Çizelge
4.10)
Nei’ye (1987) göre L. sativus popülasyonları için hesaplanan ortalama genetik çeşitlilik
değeri H= 0,1045 (%P = 41,12)’dir (Çizelge 4.10).
Çizelge 4.10 L. sativus popülasyonlarının genetik varyasyon analizleri (Nei 1987)
Popülasyonlar Birey Sayısı Na Ne H I P %P
8a (Batman) 2 1,0726 1,0514 0,0301 0,0439 56 7,26 8b(Diyarbakır) 10 1,2789 1,1411 0,0840 0,1288 215 27,89 8c (Elazığ) 5 1,1556 1,0954 0,0557 0,0833 120 15,56 8d (Afyon) 4 1,1284 1,0853 0,0484 0,0716 99 12,84 ORTALAMA 21 1,4112 1,1675 0,1045 0,1650 317 41,12
Genetik farklılaşmanın nisbi büyüklüğünü gösteren GST değeri 0,5193 ve popülasyonlar
arasındaki gen akışını ifade eden Nm değeri 0,4628 olarak hesaplanmıştır (Çizelge
4.11).

95
Çizelge 4.11 L. sativus’ un populasyonları için genetik farklılaşma analizi (Nei 1987)
Birey Sayısı HT HS GST Nm
21 0,1135 0,0546 0,5193 0,4628
Nei’ye göre hesaplanan genetik mesafe değerleri (D) 0,0236 ile 0,1428 arasındadır
(Çizelge 4.12). Buna göre genetik olarak en yakın L. sativus popülasyonlarının 8d
(Afyon) ve 8c (Elazığ) (0,0236), en uzak ise 8d (Afyon) ve 8a (Batman) (0,1428)
popülasyonları olduğu görülmüştür.
Çizelge 4.12 L. sativus popülasyonlarının genetik mesafe değerleri (Nei 1978) ============================================================== Popülasyonlar 8a 8b 8c 8d ================================================== 8a Batman ****** 8b Diyarbakır 0.0954 ****** 8c Elazığ 0.1358 0.0353 ****** 8d Afyon 0.1428 0.0478 0.0236 ***** ==================================================
Genetik mesafe değerleriyle oluşturulan dendrogramda (Şekil 4.32) L. sativus
popülasyonlarının 2 gruba ayrıldığı görülmüştür. 1. Grupta 8a (Batman) popülasyonu
tek başına yer alırken, 2. Grupta 2 alt grup oluşmuş bu alt gruplardan birincisini 8b
(Diyarbakır) popülasyonu tek başına oluştururken ikincisini ise 8c (Elazığ) ve 8d
(Afyon) popülasyonlarının dalları meydana getirmiştir.
8a Batman 8b Diyarbakır 8c Elazığ 8d Afyon
1 Şekil 4.33 L. sativus popülasyonlarının genetik mesafe verilerine göre (Nei 1978)
çıkarılan dendrogramı
96
L. cassius popülasyonları
L. cassius popülasyonları için hesaplanan genetik çeşitlilik değerlerine bakıldığında, en
düşük değerin H= 0,0286 ( %P=8,04) ile 3a (Diyarbakır) popülasyonunda, en yüksek
değerin ise H= 0,0500 (%P=13,36) ile 3b (Gaziantep) popülasyonunda olduğu
görülmüştür. Nei’ye (1987) göre L. cassius popülasyonları için hesaplanan ortalama
genetik çeşitlilik değeri H= 0,1092 (%P=30,74)’dir (Çizelge 4.13).
Çizelge 4.13 L. cassius popülasyonlarının genetik varyasyon analizleri (Nei 1987)
Popülasyon Birey Sayısı Na Ne H I P %P 3a(Diyarbakır) 4 1,0804 1,0486 0,0286 0,0429 62 8,04
3b(Gaziantep) 8 1,1336 1,0888 0,0500 0,0736 103 13,36
ORTALAMA 12 1,3074 1,1886 0,1092 0,1623 237 30,74
Genetik farklılaşmanın nisbi büyüklüğünü gösteren GST değeri 0,6549 ve popülasyonlar
arasındaki gen akışını ifade eden Nm değeri 0,2635 olarak hesaplanmıştır (Çizelge 4.14).
Çizelge 4.14 L. cassius’ un populasyonları için genetik farklılaşma analizi (Nei 1987)
Birey Sayısı HT HS GST Nm
12 0,1139 0,0393 0,6549 0,2635
Nei’ye göre hesaplanan, iki popülasyon arasındaki genetik mesafe değeri (D)
0,1648’dir (Çizelge 4.15).
Çizelge 4.15 L. cassius popülasyonlarının genetik mesafe değerleri (Nei, 1978) ============================== Popülasyonlar 3a 3b ============================== 3a (Diyarbakır) **** 3b (Gaziantep) 0.1648 **** ==============================

97
4.2 SDS-PAGE 4.2.1 Protein ekstraksiyonu 4.2.1.1 Protein miktar tayini Esen (1978)’in metoduna göre Whatman No 4 filtre kağıdına ekstraksiyon işlemi
sonrasında elde edilen supernatan emdirilmiş bölgelerin Coomassie Brillant Blue
boyasıyla boyandıktan sonraki renk şiddetinin durumu değerlendirildiğinde bütün
örneklerin renk şiddetinin aynı olduğu sonucuna varılmıştır (Şekil 4.33).
Şekil 4.34 Spot test: 1. L. cassius, 2.3.4. L. annuus, 5.6.7. L. cicera, 8. L. trachycarpus, 9. L. pseudo cicera, 10. L. gorgoni var. gorgoni, 11. L. chrysanthus, 12. 13. L. sativus, 14. L. chloranthus, 15. L. hirsutus, 16. L. boissieri, 17. Vicia sativa
4.2.2 Depo proteinleri polimorfizmi ve genetik mesafelerin belirlenmesi
Elektroforez sonrasında boyanan jellerdeki bant desenleri şekil 4.34-4.35’de
görülmektedir.
10 adet Cicercula seksiyonuna ait türün ve bir seksiyon dışı bir de cins dışı türün
tohumlarından elde edilen SDS-PAGE bant desenleri incelendiğinde toplam 52 adet
bant gözlenmiştir. Bütün bantların molekül ağırlığı 15 kD ile 140 kD arasında
değişmektedir. Marker olarak “Prestained Protein Molecular Weight Marker
(Fermentas)” kullanılmıştır. Toplamda gözlenen 52 bantın tamamının polimorfik olduğu
tespit edilmiştir.

98
Bant varlığına “1”, yokluğuna “0” değeri verilerek oluşturulan veri dosyaları (Ek 2)
POPGENE Microsoft Window-based Freeware for Population Genetics Analysis,
Version 1.31 (Yeh vd. 1997) programına girilerek Nei (1978)’ye göre genetik mesafe
değerleri elde edilmiştir (Çizelge 4.16). Buna göre en yakın genetik mesafe değeri
0.1710’dir ve L. cicera ile L. hirsutus arasındadır. En uzak genetik mesafe değeri ise
0,5500 olup, Vicia sativa’nın hem L. gorgoni var. gorgoni, hem de L. chloranthus ile
arasındaki genetik mesafedir.
Seksiyona ait türler arasındaki en uzak mesafe ise yine 0,5500 ile hem Lathyrus
hirsutus ve Lathyrus pseudo-cicera, hem de L. chloranthus ve Lathyrus trachycarpus
arasındaki genetik mesafedir.
Genetik mesafe değerleri ile elde edilen dendrogram (Şekil 4.36) incelendiğinde türlerin
iki ana gruba ayrıldığı görülmüştür.
1.Ana grup kendi içinde de 2 alt gruba ayrılmıştır. Bu alt gruplardan birincisini
Lathyrus cassius ve Lathyrus annuus’a ait iki dal oluşturmuştur. İkinci alt grubu 2
grup meydana getirmiştir. Bu gruplardan birincisi yine iki kola ayrılmış ve birinci kolu
Lathyrus cicera ile Lathyrus hirsutus dalları, ikinci kolu Lathyrus sativus dalı meydana
getirmiştir. İkinci grup ise yalnızca L. chloranthus dalından oluşmuştur.
2.Ana grup da kendi içinde yine 2 ayrı alt gruba ayrılmıştır. 1. alt grupta, iki kol
bulunur. Birinci kolda Lathyrus trachycarpus, ve Lathyrus pseudo-cicera’ya ait alt
dallar varken, 2. kolda Lathyrus chrysanthus ve Lathyrus boissieri’ye ait dallar
bulunur. 2. alt grupta Lathyrus gorgoni var. gorgoni’ye ait tek bir dal mevcuttur.
Vicia sativa dış grup olarak bu grupların dışında yer almıştır. Ancak Lathyrostylis
(Platystylis) seksiyonunda olduğu bilinen (Davis 1970) ve çalışmada seksiyon dışı grup
olarak kullanımı amaçlanan Lathyrus boissieri 2. ana grup içinde yer almıştır.
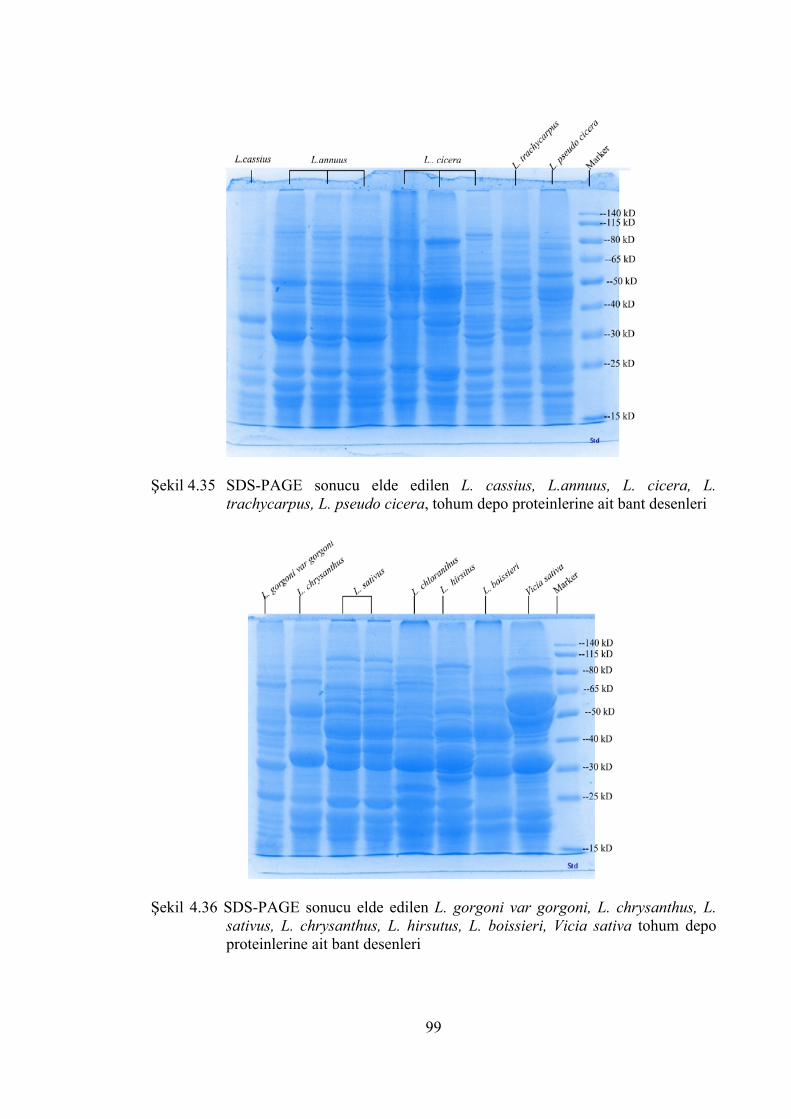
99
Şekil 4.35 SDS-PAGE sonucu elde edilen L. cassius, L.annuus, L. cicera, L.
trachycarpus, L. pseudo cicera, tohum depo proteinlerine ait bant desenleri
Şekil 4.36 SDS-PAGE sonucu elde edilen L. gorgoni var gorgoni, L. chrysanthus, L. sativus, L. chrysanthus, L. hirsutus, L. boissieri, Vicia sativa tohum depo proteinlerine ait bant desenleri
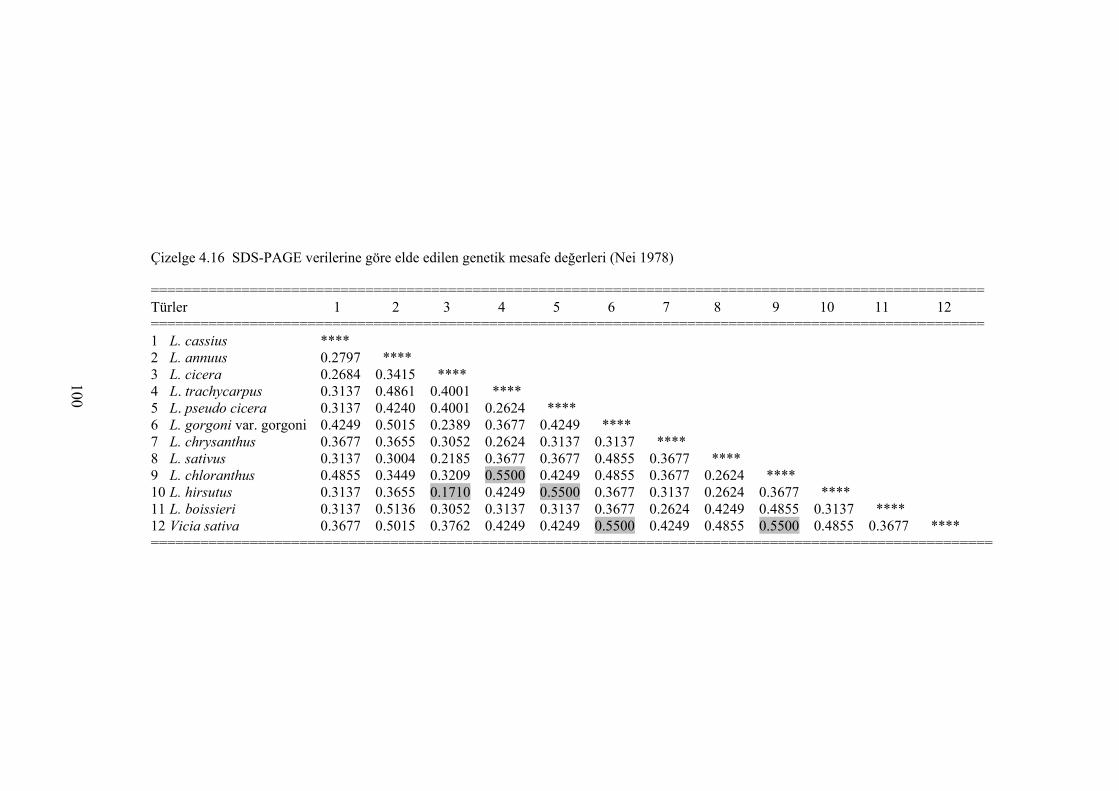
100
Çizelge 4.16 SDS-PAGE verilerine göre elde edilen genetik mesafe değerleri (Nei 1978)
===================================================================================================== Türler 1 2 3 4 5 6 7 8 9 10 11 12 ===================================================================================================== 1 L. cassius **** 2 L. annuus 0.2797 **** 3 L. cicera 0.2684 0.3415 **** 4 L. trachycarpus 0.3137 0.4861 0.4001 **** 5 L. pseudo cicera 0.3137 0.4240 0.4001 0.2624 **** 6 L. gorgoni var. gorgoni 0.4249 0.5015 0.2389 0.3677 0.4249 **** 7 L. chrysanthus 0.3677 0.3655 0.3052 0.2624 0.3137 0.3137 **** 8 L. sativus 0.3137 0.3004 0.2185 0.3677 0.3677 0.4855 0.3677 **** 9 L. chloranthus 0.4855 0.3449 0.3209 0.5500 0.4249 0.4855 0.3677 0.2624 **** 10 L. hirsutus 0.3137 0.3655 0.1710 0.4249 0.5500 0.3677 0.3137 0.2624 0.3677 **** 11 L. boissieri 0.3137 0.5136 0.3052 0.3137 0.3137 0.3677 0.2624 0.4249 0.4855 0.3137 **** 12 Vicia sativa 0.3677 0.5015 0.3762 0.4249 0.4249 0.5500 0.4249 0.4855 0.5500 0.4855 0.3677 **** ======================================================================================================

101
L.cassius
L.annuus1. Alt Grup
L. cicera
L. hirsitus
L. sativus
L. chloranthus
2 Alt Grup
1. Ana Grup
L. trachycarpus
L. pseudo-cicera
L.chrysanthus
L. boissieri
1. Alt grup
2. Alt grup L. gorgoni var. gorgoni
2. Ana Grup
Cins Dışı Grup Vicia sativa
5 Şekil 4.37 Çalışmada kullanılan türlerin SDS-PAGE verileriyle elde edilen genetik mesafe değerleriyle oluşturulmuş UPGMA dendrogram
(Nei 1978)

102
5. TARTIŞMA ve SONUÇ Lathyrus cinsinin Cicercula seksiyonunun Türkiye’de dağılım gösteren bazı türlerinin
RAPD ve SDS-PAGE yöntemi ile genetik varyasyonları incelenmiş, filogenetik
ilişkileri değerlendirilmiştir. Değerlendirmeye alınan türlerin, tamamının kullanıldığı
başka bir çalışma bulunmamaktadır. Bu bakımdan çalışmanın ortaya koyduğu veriler
ilerideki daha kapsamlı çalışmalar için iyi bir referans olacaktır.
Bu çalışmada denenen üç DNA ekstraksiyonu tekniği arasında en iyi bantları hazır kit
yöntemi vermiştir. Ancak Aras vd. (2003)’nin önerdiği modifiye edilmiş CTAB metodu
da polimorfizmin belirlenmesinde yeterli olacağı düşünülen bantlar ortaya koymuştur.
Bu sebeple ekonomik yönden avantajlı olan bu teknik çalışmada tercih edilen yöntem
olmuştur. Ayrıca çalışmalara bakıldığında da laboratuvarlarda en çok kullanılan
metodun Doyle ve Doyle (1991)’un önerdiği CTAB yöntemi veya bunun değişik
modifikasyonları olduğu görülmüştür (Laucou vd. 1998, Chen ve Ronald 1999, Bay
2009).
Sarıkaya (2004) tarafından agaroz jel üzerindeki genomik DNA’nın keskin bantlar ve
bunların aşağı ve yukarısına doğru yayılan (smear) bir görüntü verdiği bildirilmektedir.
Bu çalışmada da elde edilen genomik DNA’nın kalitatif açıdan incelenmesi için yapılan
agaroz jel elektroforezi sonrasında bu bildirimle uyumlu sonuçlar alınmıştır.
Çalışmada PZR döngüsü için iki farklı metot denenmiş ancak bunlardan PZR2 (Şekil
4.4) döngüsünden yeterli derecede PZR ürünü temin edilememiştir. Oysa Croft vd.
(1999) bu tekniği yine Lathyrus türleri üzerinde kullanmışlardır. Kullandıkları
primerlerden sadece OPA4, üzerinde çalıştıkları türlerden de sadece L. sativus bu
çalışmada da optimizasyon sırasında kullanılmıştır. Croft vd. (1999) kullandıkları bu
PZR2 (Şekil 4.4) döngüsüyle gerçekleştirdikleri çoğaltma işleminde OPA4 primerinin
diğer türlerle 18 bant, L. sativus türü ile 9 bant ürettiğini bildirmişlerdir. Bu çalışmada
da OPA4 primeri L. sativus türü ile 9 bant vermiştir, ancak bu durum Croft vd.
(1999)’un önerdiği PZR2 döngüsüyle değil, Kaya ve Neale (1995)’in önerdiği PZR
döngüsünden modifiye edilmiş PZR1 (Şekil 4.3) döngüsüyle gerçekleşmiştir. PZR2’nin

103
ürünleri jel üzerinde çok çok silik bantlar vermiş, bu sebeple de çalışmada tercih
edilmemiştir.
Bu çalışmada SDS-PAGE ve RAPD sonuçlarının birebir örtüşmediği görülmüştür.
Yüzbaşıoğlu’nun (2005) Lens culinaris Medik. (mercimek) türüne ait 14 çeşit ve 36 hat
kullanarak yaptığı çalışmada ve Babaoğlu vd. (2004)’nin Alyssum L. türleri ile
yaptıkları çalışmada da protein ve DNA düzeyindeki analiz sonuçlarının örtüşmediği
görülmektedir. SDS-PAGE yönteminde, kullanılan örnek sayısının artırılması ve RAPD
yönteminde kullanılan bireylere ait tohumların kullanılması durumunda analiz sonuçları
birbiriyle örtüşebilir.
RAPD sonuçlarından elde edilen dendrogramda, Cicercula seksiyonu iki ana gruba
ayrılmıştır. L. cicera ile L. sativus, L.cassius ile L. hirsutus, L. chloranthus ile L.
trachycarpus, L. gorgoni var. gorgoni ile L.pseudo-cicera kardeş türler olarak ortaya
çıkmıştır.
Asmussen ve Liston (1998), Lathyrus cinsinin kloroplast DNA’larından yaptıkları
RFLP yöntemi ile ortaya koydukları sınıflandırmada, 41 farklı Lathyrus türü
incelemişlerdir. Bu türler içinde, bu çalışmadaki türlerden L. cicera, L. sativus, L.
hirsutus ve L. annuus bulunmaktadır. Araştırmacılar Cicercula seksiyonu
tanımlamamışlar, bu türleri Lathyrus seksiyonunda kabul etmişlerdir. Ortaya
çıkardıkları dendogamda L. cicera ile L. gorgoni, L. sativus ile L. amphicarpos kardeş
tür olarak belirlenmiştir. Ancak bu dört tür aynı grup içinde yer almıştır. Bu sebeple bu
çalışmayla uyumludur.
Croft vd. (1999) sekiz Lathyrus türünün RAPD yöntemiyle elde ettikleri dendrogramda
L. sativus ile L. gorgoni’ yi, L. cicera ile L. odoratus’u kardeş tür olarak
belirlemişlerdir. Bu dört tür yine aynı grup içinde yer almıştır. Yine L. cicera ve L.
sativus’un aynı grupta bulunması sebebiyle bu çalışmayla uyumludur.
Ghorbel vd. (2002) beş Lathyrus türünün (L. sativus, L. cicera, L. ochrus, L. sylvestris,
L. latifolius) RAPD yöntemiyle elde ettikleri dendrogramında, Lathyrus seksiyonunda

104
kabul ettikleri L. sativus ile L. cicera’yı bu çalışmayla uyumlu olarak kardeş türler
olarak tespit etmişlerdir.
Badr vd. (2002) AFLP yöntemiyle Lathyrus seksiyonuna ait türlerin analizlerini
gerçekleştirmişlerdir. Araştırmacılar bu çalışmada kullanılan L. annuus, L. cicera, L.
hirsutus, L. pseudo-cicera, L. sativus türlerini çalışmalarında kullanmışlar ancak bunları
Lathyrus seksiyonunda kabul etmişlerdir. AFLP yöntemiyle elde ettikleri dendrogramda
L. sativus ile L. gorgoni, L. cicera ile L. hirsutus aynı grup içinde kardeş tür olarak
ortaya çıkmıştır. Bunun yanısıra L. annuus ile L. hierosolymitanus, L. pseudo-cicera ile
L. blepharicarpus aynı grup içinde kardeş tür olarak belirlenmiştir. L. cicera ve L.
sativus’un yine aynı grup içinde yer alması bu çalışmayla uyumludur.
Kenicer vd. (2005) 53 Lathyrus türünün nükleer DNA’daki bazı internal transcribed
spacer (ITS)’ların ve cpDNA’sının trnL-F ve trnS-G bölgelerinin dizi analizi ile
yaptıkları çalışmada Cicercula seksiyonu tanımlamamışlardır. Çalışmalarında 53 tür
içinde bu çalışmadaki L. cicera, L. sativus, L. annuus türleri bulunmaktadır. Bu türleri
Lathyrus seksiyonu içinde kabul etmişlerdir. Ortaya koydukları dendrogramda, bu
çalışmadaki bulgularla uyumlu olarak, L. sativus ve L. cicera kardeş türler olarak tespit
edilmiştir. L. annuus ise bu çalışmada yer almayan başka türlerle kardeş tür olarak
ortaya çıkmıştır.
Yukarıdaki çalışmalar değerlendirildiğinde genellikle L. sativus ile L. cicera’ nın aynı
grupta yer aldığı, görülmektedir. Bu çalışmada ve Ghorbel vd. (2002)’ in çalışmasında
görüldüğü gibi çalışmaya sadece L. sativus ve L. cicera alındığında kardeş türler olarak
ortaya çıkmaktadırlar.
L. hirsutus, L. pseudo-cicera, L. annuus türlerinin bu çalışmada kardeş çıkan türleri
bahsedilen çalışmalarda kullanılmadığından yorum yapmak zordur. Ayrıca L. cassius L.
chloranthus, L. trachycarpus L. gorgoni var. gorgoni türleri de yukarıdaki çalışmalarda
kullanılmadığından bunlar için de yorum yapmak zordur. Ancak morfolojik verilere
dayalı geleneksel sınıflandırmalarla bir karşılaştırma yapmak mümkündür.

105
Ertekin (1991) çalışmasında incelenen morfolojik özelliklerin genellikle Davis (1970)
ve Kupicha (1983) ile uyumluluk gösterdiğini, ancak Kupicha’nın (1983) Cicercula
seksiyonunu Lathyrus seksiyonuyla birleştirmesini hatalı bulduğunu, çünkü Cicercula
seksiyonu üyelerinin Lathyrus üyelerine çok benzer olmalarına rağmen, belirgin olarak
tek yıllık ve genellikle tek çiçekli ya da Lathyrus seksiyonuna göre daha seyrek çiçekli
olmalarıyla bu seksiyondan ayrılmaları gerektiğini bildirmiştir. Ayrıca Cicercula
seksiyonundan, L. hirsutus, L. chrysanthus, L. chloranthus ve L. trachycarpus’un
seksiyonunun diğer üyelerinden 2-8 çiçekli ve belirgin tuberkulat tüylü baklalarıyla
farklı olduklarını ve morfolojik olarak birbirlerine daha yakın olduklarını bildirmiştir.
Yine seksiyonun diğer üyelerinden L. cicera, L. sativus L. gorgoni var. gorgoni, ve L.
pseudo-cicera’nın yaprak damarlanması, tek çiçekli olması, tohum şekilleri gibi
özellikleriyle birbirlerine daha yakın olduğunu bildirmiştir. L. annuus ve L. cassius’un
da birbirine çok yakın türler olup morfolojik olarak seksiyon içinde her iki gurup
arasında yer aldığını belirtmiştir (Ertekin 1991).
Bu çalışmada RAPD yöntemiyle elde edilen dendrogramda L. cicera ile L. sativus’un ve
L. gorgoni var. gorgoni ile L. pseudo-cicera’nın kardeş tür çıkmasıyla L. hirsutus, L.
chloranthus, L. chrysanthus ve L trachycarpus’un aynı grup içinde yer almaları
Ertekin’in (1991) çalışmasıyla uyumludur. Ancak L. sativus’un ve L. gorgoni. var.
gorgoni ile L. pseudo-cicera’nın kardeş tür çıkmalarına karşın aynı grup içinde yer
almamaları Ertekin’in (1991) morfolojik belirteçlere göre yaptığı gruplandırmaya
uymamaktadır. SDS-PAGE yönteminden elde edilen dendrogram sonuçlarına
bakıldığında L. annuus ve L. cassius Ertekin’in (1991) çalışmasıyla uyumlu bir şekilde
kardeş türler olarak ortaya çıkmış ancak diğer gruplanmalar farklı bulunmuştur. Buna
göre bu çalışmadan sağlanan moleküler belirteçlere göre yapılan gruplandırma,
morfolojik belirteçlere göre yapılan gruplandırmayla kısmen uyumlu olsa da büyük
oranda uyumlu değildir.
Emre vd. (2007) yaptıkları çalışmada, Türkiye’nin 14 farklı coğrafik bölgesinden
topladıkları Lathyrus cinsinin Cicercula seksiyonuna ait 8 türün ( L. stenophyllus, L.
hirsutus, L. chloranthus, L. cicera, L. sativus, L. annuus, L. cassius ve L. phaselitanus)
filogenetik ilişkilerini, tohumlarının SDS-PAGE bant desenlerine dayanarak

106
incelemişlerdir. Hem bu çalışmada hem de Emre vd. (2007)’nin çalışmasında incelenen
türlerden altısı (L.annuus, L. cassius, L. hirsutus, L. chloranthus, L. cicera, L. sativus)
aynı türlerdir. Ancak toplanma yerleri farklıdır. Araştırmacılar elde ettikleri bulgulara
göre oluşturdukları dendrogramda iki grup ortaya çıktığını bildirmişlerdir. Bu
gruplardan birincisini L. stenophyllus, L. hirsutus, L. chloranthus, L. cicera, L. sativus
türleri, ikincisini ise L. annuus, L. cassius ve L. phaselitanus türleri oluşturmaktadır.
Bu çalışmada olup Emre vd. (2007)’nin çalışmasında olmayan türleri (L. gorgoni var
gorgoni, L. pseudo-cicera, L. chrysanthus, L. trachycarpus) ve Emre vd. (2007)’nin
çalışmasında olup bu çalışmada olmayan türleri (L. stenophyllus ve L. phaselitanus)
dikkate almadığımızda, her iki SDS-PAGE yönteminden elde edilen dendrogramlardaki
gruplaşmanın aynı olduğu görülmektedir. Ancak, SDS-PAGE sonuçlarına göre bu
çalışmada genetik mesafesi en yakın türler L. cicera ve L. hirsutus olarak
belirlenmişken, diğer çalışmada en yakın genetik mesafe L. hirsutus ile L. stenopyllus
arasında görülmüştür. Bu çalışmada L. stenophyllus türünün bulunmayışı bu konuda
yorum yapmayı zorlaştırmaktadır. Ancak genel olarak bakıldığında her iki çalışma
birbiriyle uyumludur.
Emre (2009) tarafından gerçekleştirilen başka bir çalışmada L. cassius dışında yine aynı
türlerin ( L. stenophyllus, L. hirsutus, L. chloranthus, L. cicera, L. sativus, L. annuus ve
L. phaselitanus) tohum depo proteinlerinin alt ünitelerinin (albüminler, globülin A,
globülin B, glutelinler) SDS-PAGE yöntemi ile filogenetik ilişkileri değerlendirilmiştir.
Emre (2009) çalışmasında L. sativus ve L. cicera arasındaki benzerliğin çok yüksek
olduğunu ve bunun da yapılmış olan pek çok çalışmayla uyumlu olduğunu bildirmiştir.
Bu çalışmada da SDS-PAGE sonucunda L. sativus ve L. cicera aynı grup içinde yer
almıştır.
El-Shanshoury (1997) çalışmasında SDS-PAGE yöntemiyle Lathyrus cinsinin 4
seksiyonunun, 14 türünün, 18 farklı bölgeye ait tohum örneklerinin depo proteinleri
analizini yapmış ve Cicercula seksiyonunun 3 gruba ayrıldığını ve elde ettiği sonuçların
önceden yapılmış olan morfolojik ve karyolojik çalışmalarla uyumlu olduğunu
bildirmiştir. Bu çalışmada ve El-Shanshoury (1997)’nin çalışmasında ortak olan

107
türlerden L. hirsutus, L. chloranthus L. cicera, L. sativus her iki çalışmada da aynı
grupta yer almıştır.
Bu çalışmada RAPD yöntemiyle elde edilen veriler ışığında tür içi popülasyonlar arası
genetik ilişkiler de değerlendirilmiştir. Buna göre, değerlendirilebilen türlerden (L.
sativus, L. cicera, L. cassius, L. annuus) tür içi popülasyonlar arası en yüksek
polimorfizm oranı (%P=51,88), L. annuus’un Gaziantep (1d) populasyonunda
bulunmuştur. En düşük polimorfizm oranı ise (%P=0,91) yine L. annuus’un Balıkesir
(1e) popülasyonunda görülmüştür. Balıkesir (1e) popülasyonuna ait olan 3 birey ETAE
tarafından gönderilen tohumlardan çimlendirilerek elde edilmiş bireylerdir. Bu düşük
polimorfizmin sebebi çimlendirilen bu tohumların aynı bireye ait olma olasılığı olabilir.
Belaid vd. (2006) yaptıkları çalışmada 3 Lathyrus türünün (L. cicera, L. sativus ve L.
ochrus) 5 populasyonunda ISSR (inter simple sequence repeats) metoduyla yaptıkları
analizde popülasyonlar içinde ve arasında yüksek genetik varyasyon tespit etmişlerdir.
Bununla beraber L. cicera ve L. sativus arasında çok yakın ilişki olduğunu ve bu türlerin
büyük olasılıkla ortak bir atadan ayrıldıklarını belirtmişlerdir. Bu çalışmada da yine
RAPD sonuçlarına göre elde edilen dendrogramda L. sativus ve L. cicera kardeş türler
olarak çıkmıştır.
Bu çalışmanın ve daha önce yapılmış morfolojik ve moleküler çalışmaların verilerinin
benzerliklerine göre bir değerlendirme yapacak olursak, çalışılan bu 10 taksonun kendi
arasında 3 gruba ayrılabileceğini söyleyebiliriz. Bunlar:
1.Grup: L. cicera, L.sativus, L. gorgoni var. gorgoni ve L. pseudo-cicera
2.Grup: L. chloranthus, L. trachycarpus, L. chrysanthus ve L. hirsutus,
3.Grup: L. annuus ve L. cassius
Yukarıda bahsedilen bugüne kadar yapılan çalışmaların çoğunda ve bu çalışmanın
RAPD ve kısmi olarak da SDS-PAGE sonuçlarında L. sativus ve L. cicera’nın çok
yakın türler olduğu görülmektedir. Cicercula seksiyonuna ait diğer türler ise
çalışmalarda her zaman aynı grup içinde yer almamaktadır. Bu da türlerin gerçekten

108
hem morfolojik hem de moleküler belirteçler aracılığıyla taksonomik sınıflandırmasının
gerçekten de güç olduğunun göstergesidir. Daha fazla sayıda materyalle, daha ileri
düzeydeki teknikler kullanılarak gerçekleştirilecek çalışmalar türler arasındaki
filogenetik ilişkilerin belirlenmesindeki bu güçlüklerle başa çıkmada önemli avantajlar
sunabilir.

109
KAYNAKLAR
Allkin, R., Macfarlane, T.D. White, R.J. Bisby, F.A. and Adey, M.E.1985. The geographical distribution of Lathyrus, Southampton University, Vicieae Database Project 6, pp. 1–35.1. Southampton, UK.
Alverson, W.S., Whitlock, B.A., Nyffeler, R., Bayer, C. and Baum, D.A. 1999. Phylogeny of the core Malvales: evidence from ndhF sequence data. American Journal of Botany Vol. 86(10), pp. 1474-86.
Anonim. 2011. Web Sitesi: http://wwweski.tubitak.gov.tr/tubives/, Erişim Tarihi: 05.08.2011.
Anonim. 2011. Web Sitesi: http://de.wikipedia.org/wiki/Datei:Lathyrus_sativus_002. JPG, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://www.menudanatura.com/2011/05/lathyrus-cicera-l.html, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://whyfiles.org/034clone/images/dna_molecule.gif, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://www.webhatti.com/saglik/60365-dna-nedir-nerede-bulunur.html, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://www.drzeydanli.com.tr/index. php?page= icerikgoster &menuID=30#DNA, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://en.wikipedia.org/wiki/SDS-PAGE, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://cgb.indiana.edu/genomics/equipment/15, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://blestbj.com/product/dianyong/ningjiao/2009/0605 /726.html, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://www.uklabs-direct.co.uk/html /used_electrophoresis _ equipment.html, Erişim Tarihi: 04.09.2011
Anonim. 2011. Web Sitesi: http://www.bio-rad.com/webroot/web/images /lsr/products/ imaging_ bioinformatics/product_detail/global/lsr_chemidoc_xrs_system.jpg, Erişim Tarihi: 04.09.2011
Aras, S., Duran, A. and Yenilmez, G. 2003. Isolation of DNA for RAPD analysis from dry leaf material of some Hesperis L. specimens. Plant Molecular Biology Reporter., Vol. 21, pp. 461a-461f.
Arda, N. ve Ertan, H. 2004, Proteinlerin izolasyonu, analizi ve saflaştırılması. In: Moleküler Biyolojide Kullanılan Yöntemler. Temizkan, G. ve Arda, N. (eds), Nobel Tıp Kitabevleri, s. 161-274, İstanbul.
Arı, Ş. 2004. DNA’nın polimeraz zincir reaksiyonu (PCR) ile çoğaltılması, In: Moleküler Biyolojide Kullanılan Yöntemler. Temizkan, G. ve Arda, N. (eds), Nobel Tıp Kitabevleri, s. 101-120, İstanbul.

110
Arnheim, N. and Erlich, H. 1992. Polymerase Chain Reaction Strategy. Annual Review of Biochemistry. Vol. 61, pp. 131–56.
Asmussen, C. and Liston, A. 1998. Chloroplast DNA characters, phylogeny and classification of Lathyrus (Fabaceae ). American Journal of Botany, Vol. 85, pp. 387–401.
Babaoglu, S. Acik, L. Celebi, A. And Adiguzel, N. 2004. Molecular analysis of Turkish Alyssum L. (Brasiıcaceae) species by RAPD-PCR and SDS-PAGE methods. Gazi Üniversitesi Fen Bilimleri Dergisi, Vol. 17(3), pp. 25-33.
Badr, A., El Shazly, H., El Rabey, H. and Watson, L. E. 2002. Systematic relationships in Lathyrus sect.Lathyrus (Fabaceae) based on amplified fragment length polymorphism (AFLP) data. Canadian Journal of Botany, Vol. 80, pp.962–969.
Barneby, R. C., and Reveal, J. L. 1971. A new species of Lathyrus (Fabaceae ) from the Death Valley region of California and Nevada. Aliso, Vol. 7, pp. 361–364.
Bartlett, J. M. S.and Stirling, D. 2008. Methods in Molecular Biology, Vol. 226: PCR Protocols, Second Edition © Humana Press Inc. 568 p, Totowa, NJ.
Bässler, M. 1966. Die Stellung des Subgenus Orobus (L.) Baker inder Gattung Lathyrus L. und seine systematische Gliederung. Feddes Repertorium, Vol. 72, pp. 69–97.
Bässler, M. 1973. Revision der eurasiatischen Arten von Lathyrus L. Sect. Orobus (L.) Gren. et Godr. Feddes Repertorium, Vol. 84, pp. 329–447.
Bässler, M.1981. Revision von Lathyrus L. sect. Lathyrostylis (Griseb.) Bässler (Fabaceae ). Feddes Repertorium, Vol. 92, pp. 179–254.
Başaran, U., Acar, Z., Önal Aşcı, Ö., Mut, H. ve Ayan, İ. 2007. Mürdümük (Lathyrus sp.) türlerinin önemi, tarımda kullanım olanakları ve zararlı madde içerikleri. Ondokuz Mayıs Üniversitesi Ziraat Fakültesi Dergisi, Vol. 22 (1), pp. 139-148.
Bay, S. 2009. Türk çeltik çeşitlerinin (Oryza satıva L.) tohum depo proteinleri ve random amplified polymorphic DNA (RAPD) belirleyicileri ile genetik analizi. Yüksek Lisans Tezi, Kahramanmaraş Sütçü İmam Üniversitesi, 43 s., Kahramanmaraş.
Belaıd, Y., Chtourou-Ghorbel, N., Marrakchi, M. and Trifi-Farah, N. 2006. Genetic diversity within and between populations of Lathyrus genus (Fabaceae) revealed by ISSR markers. Genetic Resources and Crop Evolution, Vol. 53, pp. 1413–1418.
Ben Brahim, N., Salhi, A. Chtourou, N. Combes, D. and Marrakchi, M. 2002. Isozymic polymporphism and phylogeny of 10 Lathyrus species. Genetic Resources and Crop Evolution, Vol. 49, pp. 427–436.
Blair, G.E. and Blair Zajdel, M.E. 1992 The polymerase chain reaction already an established technique in biochemistry. Biochemical Education, Vol. 20(2), pp. 87-91.
Boissier, E. 1872. Lathyrus and Orobus. In: Flora Orientalis 2. Boissier, E. (ed), pp. 599–622. Geneva, Basel.

111
Bothmer, R., Van Jacobsen, N., Baden, C., Linde-Laursen, I. and Jorgessen, R.B. 1991. An ekogeographical study of the genus Hordeum. Systematic and Ecogeographic Studies on Crop Genopools. 7. Int. Board for Plant Genetic Resources, Rome.
Cardy, B.J. and Beversdorf, W.P. 1984. Identification of soybean cultivars using isoenzyme electrophoresis. Seed Science and Technology, Vol. 12, pp. 943-954.
Carey, K. and Ganders, F.R. 1987. Patterns of isoenzyme variation in Plectritis (Valerianaceae). Systematic Botany, Vol. 12(1), pp. 125-132.
Chen, D.-H., and Ronald, P.C. 1999. A Rapid DNA minipreparation method suitable for AFLP and other PCR applications. Plant Molecular Biology Reporter, Vol. 17, pp. 53–57.
Chtourou-Ghorbel, N., Lauga, B. Combes, D. and Marrakchi, M. 2001. Comparative genetic diversity studies in the genus Lathyrus using RFLP and RAPD markers. Lathyrus Lathyrism Newsletter, Vol. 2(2), pp. 62–68.
Clulow, S. A., Matthews, P. Lewis, B.G., 1991. Genetical analysis of resistance to Mycosphaerella pinodes in pea seedlings. Euphytica, Vol. 58, pp. 183–189.
Cooke, R.J. 1984. The characterisation and identification of crop cultivars by electrophoresis. Electrophoresis, Vol. 5, pp. 59-72.
Croft, A. M., Pang, E. C. K. and Taylor, P. W. J. 1999. Molecular analysis of Lathyrus sativus L. (grasspea) and related Lathyrus species. Euphytica, Vol. 107, pp. 167–176.
Czefranova, Z. 1971. Conspectus systematis generis Lathyrus L. Novitates Systematicae. Plantarum Vascularum (Leningrad), Vol. 8, pp. 191–201.
Davis, P. H. 1970. Lathyrus L. In: Flora of Turkey P. H. Davis (ed), Edinburgh University Press, Vol. 3, pp. 328–369, Edinburgh.
Davis, P.H. 1988. Flora of Turkey and East Aegean Islands (Supplement), Edinburgh University Press, Vol. 10, pp.125-126, Edinburgh.
de Vicente, M.C. and Fulton, T. 2003. Using molecular marker technology in studies on plant genetic diversity. Illus. Nelly Giraldo. IPGRI, Rome, Italy and Institute for Genetic Diversity, Ithaca, New York, USA. http://www.bioversityinternational.org/training/training_materials/using_molecular_marker_technology_in_studies_on_plant_genetic_diversity_vol_1/molecular_markers_vol_1.html Erişim Tarihi: 30.05.2011.
de Vicente, M.C., Lopez, C. and Fulton, T. 2004. Genetic Diversity Analysis with Molecular Marker Data: Learning Module. International Plant Genetic Resources Institute (IPGRI), Rome, Italy. http://www.bioversityinternational.org/training/training_materials/genetic_diversity_analysis_with_molecular_marker_data_learning_module_volume_2/molecular_markers_vol_2.html Erişim Tarihi: 30.05.2011.
Derbyshire, E., Wright, D. J. and Boulter, D. 1976. Legumin and vicilin, storage proteins of legume seeds. Phytochemistry, Vol. 15 (1), pp. 3-24.

112
Dogan, M., Kence, A. and Tıgın, C. 1992. Numerical taxonomic study on Turkish Lathyrus (Leguminosae). Edinburgh Journal of Botany, Vol. 49, pp. 333–341.
Doyle, J.J. and Doyle, J.L. 1991. Isolation of plant DNA from fresh tissiue. Focus Vol. 12(1), pp. 13-15.
Elçi, Ş. 1988. Ziraatte Baklagiller. Tarım İşletmeleri Genel Müdürlüğü Yayınları, 422 s, Ankara.
El-Shanshoury, A.R. 1997. The use of seed proteins revealed by SDS PAGE in taxonomy and phylogeny of some Lathyrus species. Biologia plantarum, Vol. 39 (4), pp. 553-559.
Emre, İ., Turgut-Balık, D., Genç, H. and Şahin, A. 2006. The Use of Seed Proteins Revealed by SDS-PAGE in Taxonomy of Some Lathyrus L Species Grown in Turkey. Pakistan Journal of Biological Sciences Vol. 9 (12), pp. 2358-2361.
Emre, İ.,Turgut-Balık, D. and Sahin, A. 2007. Electrophoretic Analysis of Total Protein Profiles of Some Lathyrus L (Sect. Cicercula) Grown in Turkey. Pakistan Journal of Biological Sciences, Vol. 10 (17), pp. 2890-2894.
Emre,İ. 2009. Electrophoretic analysis of some Lathyrus L. species based on seed storage proteins. Genetic Resources and Crop Evolution, Vol. 56(1), pp. 31-38.
Emre,İ., Turgut- Balık, D., Genç, H., Şahin, A. 2010. Total seed storage protein patterns of some Lathyrus species growıng in Turkey using SDS-PAGE. Pakistan Journal of Botany, Vol. 42(5), pp. 3157-3163.
Ertekin, S. A. 1991. Güneydoğu Anadolu Bölgesinin Lathyrus L. (Fabaceae) türleri üzerine sistematik, morfolojik ve anatomik araştırmalar. Doktora Tezi (basılmamış), 123 s., Dicle Üniversitesi, Diyarbakır.
Ertekin, S. A., Saya, Ö., 1996. Türkiye Florası İçin Yeni Bir Kayıt. Doğa Turkish Journal of Botany, Vol. 15(1), pp. 75-77.
Esen, A. 1978. A simple method for quantitative, semiquantitative and qualitative assay of protein. Analytical Biochemistry, Vol. 89, pp. 264-273.
Freeman, S. ve Herron, J. C. 2009. Evrimsel Analiz (çeviri: B. Çıplak, H.H. Başıbüyük, S. Karaytuğ, İ. Gündüz), 2. Baskı. Palme Yayıncılık, 838 s., Ankara.
Fukushima, D. 1991. Recent progress of soybean protein foods: Chemistry, technology, and nutrition. Food Reviews International, Vol. 7, pp. 323–351.
Ghorbel, N.C., Lauga, B., Brahim, N.B., Combes, D. and Marrakchi, M. 2002. Genetic variation analysis in the genus Lathyrus using RAPD markers. Genetic Resources and Crop Evolution, Vol. 49, pp. 363–370.
Godron, M. 1848. Lathyrus. In: Flore de France 1. Grenier, M. and Godron, M. (eds), Baillie`re, pp. 478–492, Paris.
Gorman, M.B. and Kiang, Y.T. 1977. Variety-specific electrophoretic variants of four soybean enzymes. Crop Science, Vol. 17, pp. 963-965.
Gözükara, E.M.1994. Biyokimya 2. Baskı. Evin Matbaası,1177 s., Malatya.

113
Hamrick, J.L., Godt, M. J. W., Murawski, D.A., Loveless, M.D. 1991. Correlation between species traits and allozyme diversity: implications for conservation biology, In: Genetics and conservation of rare plants. Falker, D.E. and Holsinger, K.E. (eds), Oxford University Press, pp. 75-86, U.K.
Heldt, H.W. 1999. Plant biochemistry and molecular biology. Oxford University Press Inc, 310 p., New York.
Hickey, M. and King, C. 1997. Common families of flowering plants. Cambridge University press, 197 p., U.K.
Jain, S., Srivastava, J.P. and Guotam, P.L. 1983. Evaluation of genetic resources of durum wheat for salt stress tolerance, In: Proceeding 6th International Wheat Genetics Symposium. pp. 137-141, Kyoto, Japan.
Kaya, Z. and Neale, D.B. 1995. Linkage Map Based on Randomly Amplified Polymorphic DNA (RAPD) Markers in Pinus brutia. Silvae Genetica, Vol. 44, pp. 110-116.
Kenicer, G.J., Kajita, T., Pennington, T.R. and Murata, J. 2005. Systematics and Biogeography of Lathyrus (Leguminosae) Based on Internal Transcribed Spacer and CpDNA Sequence Data. American Journal of Botany, Vol. 92(7), pp. 1199–1209.
Kenicer, G. 2008. An introduction to the genus Lathyrus L. Curtis’s Botanical Magazine, Vol. 25(4), pp. 286-295.
Kumar, 1997. Utilization of Lathyrus. Lathyrus Genetic Resources Network. 8-10 December. pp. 57-59, New Delhi/India.
Kumar, S., Tamura, T. and Nei, M. 2004. MEGA3: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment. Briefings in Bioinformatics, Vol. 5(2), pp. 150-163.
Kupicha, F.K. 1983. The infrageneric structure of Lathyrus. Notes from the Royal Botanic Garden Edinburgh, Vol. 41, pp. 209-244.
Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of Bacteriophage T4. Nature, Vol. 227, pp. 680-685.
Laucou, V., Haurogne, K., Ellis, N., Rameau, C. 1998. Genetic mapping in pea. 1. RAPD-based genetic linkage map of Pisum sativum. Theoretical and Applied Genetics, Vol. 97, pp. 905-915.
Linnaeus, C. (1753). Species Plantarum, Impensis direct Laurentii Salvii, vol. 2, 1684 p., Stockholm.
Liston, A., Shmida, A. 1987. Contributions to the Flora of Mount Hermon (Israel). I. Species with Interesting Disjunct Distributions – Kitaibelia balansae and Lathyrus roseus. Botanische Jahrbücher für Systematik, Vol. 109(1), pp. 59-69.
Lowe, A. J., Hanotte, O. and Guarino, L. 1996. Standardization of Molecular Genetic Techniques for the Characterization of Germplasm Collections: The Caase of Random Amplified Polymorphic DNA (RAPD). Plant Genetic Resources Newsletter. Vol. 107, pp. 50–54.

114
Mandal, S. and Mandal, R.K. 2000. Seed storage proteins and approaches for improvement of their nutritional quality by genetic engineering. Current Science, Vol. 79(5), pp. 576-589.
Max, K.N., Mouchaty, S.K., Schwaegerle, K.E. 1999. Allozyme and morphological variation in two subspecies of Dryas octopetala (Rosaceae) in Alaska. American Journal of Botany, Vol.86(11), pp. 1637-1644.
Maxam, A., Gilbert, W. 1977, A new method of sequencing DNA. Proceedings of the National Academy of Sciences, Vol. 74, pp. 560-4.
Maxted, N., Goyder, D.J. 1988. A New Species of Lathyrus (Leguminosae, Papilionodeae) from Turkey. Kew Bulletin, Vol. 43(4), pp. 711-714.
Millerd, A. 1975. Biochemistry of legume seed proteins. Annual Review of Plant Biology, Vol. 26, pp. 53–72.
Nei, M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, Vol. 89, pp. 583–590.
Nei, M. 1987. Molecular Evolutionary Genetics. Columbia University Press, 512 p., New York.
Onarıcı, S. G. and Sümer, S. 2003. Protein and DNA in systematic biology. Turkish Journal of Biology, Vol. 27, pp. 47-55.
Özcan, S., Gürel, E. ve Babaoğlu, M. 2004. Bitki Biyoteknolojisi II Genetik Mühendisliği ve Uygulamaları 2. Baskı. Selçuk Üniversitesi Vakfı, 456 s, Konya.
Parmaksız, İ. 2004. Papaver Cinsi Oxytona Seksiyonunun Türkiye’de Yetişen Türlerinde Genetik Çeşitliliğin RAPD Markörleri ile Analizi. Doktora Tezi. Ankara Üniversitesi, 174 s., Ankara.
Peirce, L.C. and Brewbaker, J.L. 1973. Application of isozyme analysis in horticultural science. Hortscience, Vol. 8, pp. 17-22.
Przybylska, J., Zimniak-Przybylska, Z., Krajewski, P. 1999. Diversity of seed albumins in some Lathyrus species related to L. sativus L.: an electrophoretic study. Genetic Resources and Crop Evolution, Vol. 46, pp. 261–266.
Sambrook, J., Fritsch, E.F. and Maniatis, T. 1989. Moleculer cloning, a laboratory manual. Second edition. Cold Spring Harbor Laboratory Press, 1659 p., New York.
Sammour, R.H. 1991. Using electrophoretic techniques in varietal identification, biosystematic analysis, phylogenetic relations and genetic resources management. Journal of Islamic Academy of Sciences Vol. 4(3), pp. 221-226.
Sanger, F., Nicklen, S., Coulson, A.R. 1977. DNA sequencing with chain- terminating inhibitors, Proceedings of the National Academy of Sciences, Vol. 74, pp. 5463-5467.
Sarıkaya, A. T. 2004. DNA’nın izolasyonu ve analizi, In: Moleküler Biyolojide Kullanılan Yöntemler. Temizkan, G. ve Arda, N. (eds), Nobel Tıp Kitabevleri, s. 55-80, İstanbul.
Shewry, P.R., Napier, J.A. and Tatham, A.S. 1995. Seed storage proteins: structure and biosynthesis. The Plant Cell, Vol. 7, pp. 945-956.

115
Smartt, J. 1990. Grain Legumes: Evolution and genetic resources. Cambridge University Press, 392 p., Cambridge.
Smith, A. D., Datta, S. P., Howard Smith, G., Campbell, P. N., Bentley, R. and McKenzie, H.A. 1997. Oxford Dictionary of Biochemistry and Molecular Biology. Oxford University Press., 738 p., Oxford.
Tanksley, S.D. and Orton, T.J. 1983. Isozymes in plant genetics and breeding. Elsevier, 516 p., Amsterdam.
Tekele-Haimanot, R., Kidane, Y., Wuhib, E., Kalissa, A., Alemu, T., Zein, Z.A., Spencer, P.S., 1990. Lathyrism in rural Northwestern Ethiopia: a highly prevalent neurotoxic disorder. International Journal of Epidemiology, Vol. 19, pp. 664–672.
Tutin, T.G., Heywood, V.H., Burges, N.A., Valentine, D.H., Walters, S.M. and Webb, D.A. 2001. Flora Europea. Cambridge University Press, Vol. 2, pp.136-143. Cambridge.
Urga, K., Fite, A., Kebede, B. 1995. Nutritional and antinutritional factors of grasspea (Lathyrus sativus) germplasms. Bulletin of the Chemical Society of Ethiopia, Vol. 9, pp. 9-16.
van Geyt, J.P.C.F. and Smed, F. 1984. Polymorphism of some marker enzymes of sugarbeet (Beta vulgaris L.) investigated by polyacrylamide gel electrophoresis and starch gel electrophoresis. Journal of Plant Breeding, Vol. 92, pp. 295-308.
Watson, J.D., Gilman, M., Witkoski, J. and Zoller, M. 1992. Recombinant DNA 2nd Ed. Scientific American Books, 626 p., New York.
Waugh, R. and Powell, W. 1992. Using RAPD markers for crop improvement. Trends in Biotechnology, Vol. 10, pp. 186–191.
Welsh, J. and McClelland, M. 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Research, Vol. 18(24), pp. 7213-7218.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A. and Tingey, S.V. 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research, Vol. 18(22), pp. 6531–6535.
Yeh, C.F., Yang, R. and Boyle, T. 1997. POPGENE Version 1.31. Windows-based Software for Population Genetics Analysis. http://www.ualberta.ca/~fyeh/ popgene_download.html. Erişim Tarihi: 15.06.2008.
Yüzbaşıoğlu, E. 2005 Mercimek hat ve çeşitlerinde genetik varyasyonun tespiti. Doktora tezi, (basılmamış). Gazi Üniversitesi, 85 s., Ankara.
Zuckerkandl, E. and Pauling, L. 1965. Molecules as documents of evolutionary history. Journal of Theoretical Biology, Vol. 8, pp. 357-366.

116
EKLER
EK 1 RAPD bant verileri matrisi
EK 2 SDS-PAGE bant verileri matrisi

117
EK 1 RAPD bant verileri matrisi LATHYRUS 12 species with 25 primer and 771 loci Number of species = 12 Number of loci = 771 locus name = OPA0101 OPA0102 OPA0103 OPA0104 OPA0105 OPA0106 OPA0107 OPA0108 OPA0109 OPA0110 OPA0111 OPA0112 OPA0113 OPA0114 OPA0115 OPA0116 OPA0117 OPA0118 OPA0119
OPA0120 OPA0121 OPA0122 OPA0123 OPA0124 OPA0125 OPA0126
OPA0201 OPA0202 OPA0203 OPA0204 OPA0205 OPA0206 OPA0207 OPA0208 OPA0209 OPA0210 OPA0211 OPA0212 OPA0213 OPA0214 OPA0215 OPA0216 OPA0217 OPA0218 OPA0219 OPA0220 OPA0221
OPA0301 OPA0302 OPA0303 OPA0304 OPA0305 OPA0306 OPA0307 OPA0308 OPA0309 OPA0310 OPA0311 OPA0312 OPA0313 OPA0314 OPA0315 OPA0316 OPA0317 OPA0318 OPA0319
OPA0401 OPA0402 OPA0403 OPA0404 OPA0405 OPA0406 OPA0407 OPA0408 OPA0409 OPA0410 OPA0411 OPA0412 OPA0413 OPA0414 OPA0415 OPA0416 OPA0417 OPA0418 OPA0419 OPA0420 OPA0421 OPA0422 OPA0423 OPA0424
OPA0501 OPA0502 OPA0503 OPA0504 OPA0505 OPA0506 OPA0507 OPA0508 OPA0509 OPA0510 OPA0511 OPA0512 OPA0513 OPA0514 OPA0515 OPA0516 OPA0517 OPA0518 OPA0519 OPA0520 OPA0521
OPA0601 OPA0602 OPA0603 OPA0604 OPA0605 OPA0606 OPA0607 OPA0608 OPA0609 OPA0610 OPA0611 OPA0612 OPA0613 OPA0614 OPA0615 OPA0616 OPA0617 OPA0618 OPA0619 OPA0620 OPA0621 OPA0622 OPA0623 OPA0624 OPA0625 OPA0626 OPA0627 OPA0628 OPA0629 OPA0630 OPA0631 OPA0632 OPA0633 OPA0634 OPA0635 OPA0636 OPA0637 OPA0638
OPA0701 OPA0702 OPA0703 OPA0704 OPA0705 OPA0706 OPA0707 OPA0708 OPA0709 OPA0710 OPA0711 OPA0712 OPA0713 OPA0714 OPA0715 OPA0716 OPA0717 OPA0718 OPA0719 OPA0720 OPA0721 OPA0722 OPA0723 OPA0724 OPA0725 OPA0726 OPA0727 OPA0728
OPA0801 OPA0802 OPA0803 OPA0804 OPA0805 OPA0806 OPA0807 OPA0808 OPA0809 OPA0810 OPA0811 OPA0812 OPA0813 OPA0814 OPA0815 OPA0816 OPA0817 OPA0818 OPA0819 OPA0820 OPA0821 OPA0822 OPA0823 OPA0824 OPA0825 OPA0826 OPA0827 OPA0828 OPA0829 OPA0830 OPA0831 OPA0832 OPA0833 OPA0834 OPA0835
OPA0901 OPA0902 OPA0903 OPA0904 OPA0905 OPA0906 OPA0907 OPA0908 OPA0909 OPA0910 OPA0911 OPA0912 OPA0913 OPA0914 OPA0915 OPA0916 OPA0917 OPA0918 OPA0919
OPA1001 OPA1002 OPA1003 OPA1004 OPA1005 OPA1006 OPA1007 OPA1008 OPA1009 OPA1010 OPA1011 OPA1012 OPA1013 OPA1014 OPA1015 OPA1016 OPA1017 OPA1018 OPA1019 OPA1020 OPA1021 OPA1022 OPA1023 OPA1024 OPA1025 OPA1026 OPA1027 OPA1028 OPA1029 OPA1030 OPA1031
OPA1101 OPA1102 OPA1103 OPA1104 OPA1105 OPA1106 OPA1107 OPA1108 OPA1109 OPA1110 OPA1111 OPA1112 OPA1113 OPA1114 OPA1115 OPA1116 OPA1117 OPA1118 OPA1119 OPA1120 OPA1121 OPA1122 OPA1123 OPA1124 OPA1125 OPA1126 OPA1127 OPA1128 OPA1129 OPA1130 OPA1131 OPA1132 OPA1133 OPA1134 OPA1135 OPA1136 OPA1137 OPA1138 OPA1139 OPA1140 OPA1141 OPA1142 OPA1143 OPA1144
OPA1501 OPA1502 OPA1503 OPA1504 OPA1505 OPA1506 OPA1507 OPA1508 OPA1509 OPA1510 OPA1511 OPA1512 OPA1513 OPA1514 OPA1515 OPA1516 OPA1517 OPA1518 OPA1519 OPA1520 OPA1521 OPA1522 OPA1523 OPA1524 OPA1525 OPA1526 OPA1527 OPA1528 OPA1529 OPA1530 OPA1531 OPA1532 OPA1533 OPA1534 OPA1535 OPA1536
OPB0501 OPB0502 OPB0503 OPB0504 OPB0505 OPB0506 OPB0507 OPB0508 OPB0509 OPB0510 OPB0511 OPB0512 OPB0513 OPB0514 OPB0515 OPB0516 OPB0517 OPB0518 OPB0519 OPB0520 OPB0521 OPB0522 OPB0523 OPB0524 OPB0525 OPB0526 OPB0527 OPB0528 OPB0529 OPB0530 OPB0531
OPB1001 OPB1002 OPB1003 OPB1004 OPB1005 OPB1006 OPB1007 OPB1008 OPB1009 OPB1010 OPB1011 OPB1012 OPB1013 OPB1014 OPB1015 OPB1016 OPB1017 OPB1018 OPB1019 OPB1020 OPB1021 OPB1022 OPB1023 OPB1024 OPB1025 OPB1026 OPB1027 OPB1028
OPB1301 OPB1302 OPB1303 OPB1304 OPB1305 OPB1306 OPB1307 OPB1308 OPB1309 OPB1310 OPB1311 OPB1312 OPB1313 OPB1314 OPB1315 OPB1316 OPB1317 OPB1318 OPB1319 OPB1320 OPB1321 OPB1322 OPB1323 OPB1324 OPB1325 OPB1326 OPB1327 OPB1328 OPB1329 OPB1330 OPB1331 OPB1332 OPB1333 OPB1334 OPB1335 OPB1336 OPB1337 OPB1338 OPB1339 OPB1340 OPB1341 OPB1342 OPB1343
OPB1701 OPB1702 OPB1703 OPB1704 OPB1705 OPB1706 OPB1707 OPB1708 OPB1709 OPB1710 OPB1711 OPB1712 OPB1713 OPB1714 OPB1715 OPB1716 OPB1717 OPB1718 OPB1719 OPB1720 OPB1721 OPB1722 OPB1723 OPB1724 OPB1725 OPB1726 OPB1727 OPB1728 OPB1729 OPB1730 OPB1731
OPD0301 OPD0302 OPD0303 OPD0304 OPD0305 OPD0306 OPD0307 OPD0308 OPD0309 OPD0310 OPD0311 OPD0312 OPD0313 OPD0314 OPD0315 OPD0316 OPD0317 OPD0318 OPD0319 OPD0320 OPD0321 OPD0322 OPD0323 OPD0324 OPD0325 OPD0326 OPD0327 OPD0328 OPD0329 OPD0330 OPD0331 OPD0332 OPD0333 OPD0334 OPD0335 OPD0336 OPD0337 OPD0338 OPD0339 OPD0340 OPD0341 OPD0342 OPD0343
OPD0501 OPD0502 OPD0503 OPD0504 OPD0505 OPD0506 OPD0507 OPD0508 OPD0509 OPD0510 OPD0511 OPD0512 OPD0513 OPD0514 OPD0515 OPD0516 OPD0517 OPD0518 OPD0519 OPD0520 OPD0521 OPD0522 OPD0523 OPD0524 OPD0525 OPD0526 OPD0527 OPD0528 OPD0529 OPD0530
OPD1001 OPD1002 OPD1003 OPD1004 OPD1005 OPD1006 OPD1007 OPD1008 OPD1009 OPD1010 OPD1011 OPD1012 OPD1013 OPD1014 OPD1015 OPD1016 OPD1017 OPD1018 OPD1019 OPD1020 OPD1021 OPD1022 OPD1023 OPD1024 OPD1025 OPD1026 OPD1027 OPD1028 OPD1029 OPD1030 OPD1031 OPD1032 OPD1033 OPD1034 OPD1035 OPD1036
OPD1101 OPD1102 OPD1103 OPD1104 OPD1105 OPD1106 OPD1107 OPD1108 OPD1109 OPD1110 OPD1111 OPD1112 OPD1113 OPD1114 OPD1115 OPD1116 OPD1117 OPD1118 OPD1119 OPD1120 OPD1121 OPD1122 OPD1123 OPD1124 OPD1125 OPD1126 OPD1127 OPD1128 OPD1129 OPD1130 OPD1131 OPD1132 OPD1133 OPD1134
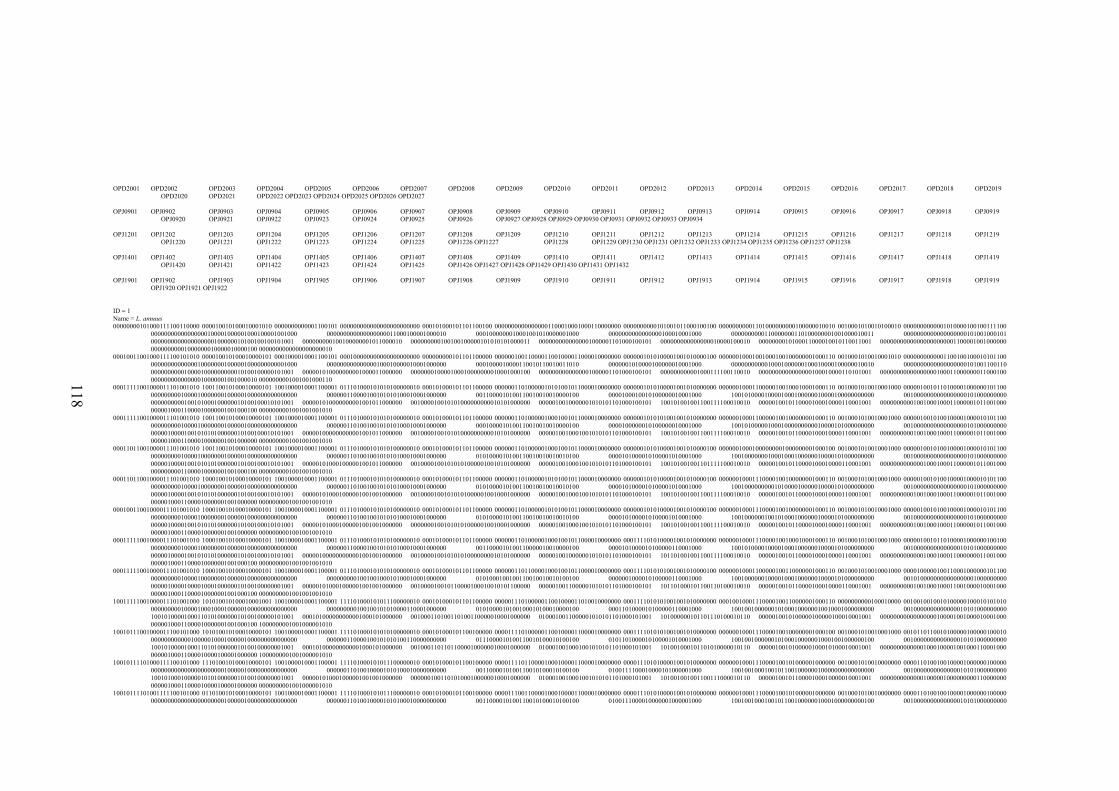
118
OPD2001 OPD2002 OPD2003 OPD2004 OPD2005 OPD2006 OPD2007 OPD2008 OPD2009 OPD2010 OPD2011 OPD2012 OPD2013 OPD2014 OPD2015 OPD2016 OPD2017 OPD2018 OPD2019 OPD2020 OPD2021 OPD2022 OPD2023 OPD2024 OPD2025 OPD2026 OPD2027
OPJ0901 OPJ0902 OPJ0903 OPJ0904 OPJ0905 OPJ0906 OPJ0907 OPJ0908 OPJ0909 OPJ0910 OPJ0911 OPJ0912 OPJ0913 OPJ0914 OPJ0915 OPJ0916 OPJ0917 OPJ0918 OPJ0919 OPJ0920 OPJ0921 OPJ0922 OPJ0923 OPJ0924 OPJ0925 OPJ0926 OPJ0927 OPJ0928 OPJ0929 OPJ0930 OPJ0931 OPJ0932 OPJ0933 OPJ0934
OPJ1201 OPJ1202 OPJ1203 OPJ1204 OPJ1205 OPJ1206 OPJ1207 OPJ1208 OPJ1209 OPJ1210 OPJ1211 OPJ1212 OPJ1213 OPJ1214 OPJ1215 OPJ1216 OPJ1217 OPJ1218 OPJ1219 OPJ1220 OPJ1221 OPJ1222 OPJ1223 OPJ1224 OPJ1225 OPJ1226 OPJ1227 OPJ1228 OPJ1229 OPJ1230 OPJ1231 OPJ1232 OPJ1233 OPJ1234 OPJ1235 OPJ1236 OPJ1237 OPJ1238
OPJ1401 OPJ1402 OPJ1403 OPJ1404 OPJ1405 OPJ1406 OPJ1407 OPJ1408 OPJ1409 OPJ1410 OPJ1411 OPJ1412 OPJ1413 OPJ1414 OPJ1415 OPJ1416 OPJ1417 OPJ1418 OPJ1419 OPJ1420 OPJ1421 OPJ1422 OPJ1423 OPJ1424 OPJ1425 OPJ1426 OPJ1427 OPJ1428 OPJ1429 OPJ1430 OPJ1431 OPJ1432
OPJ1901 OPJ1902 OPJ1903 OPJ1904 OPJ1905 OPJ1906 OPJ1907 OPJ1908 OPJ1909 OPJ1910 OPJ1911 OPJ1912 OPJ1913 OPJ1914 OPJ1915 OPJ1916 OPJ1917 OPJ1918 OPJ1919 OPJ1920 OPJ1921 OPJ1922
ID = 1 Name = L. annuus 00000000101000111100110000 000010010100010001010 0000000000001100101 000000000000000000000000 000101000101101100100 00000000000000001100010001000110000000 0000000000101001011000100100 00000000001101000000000100000010010 0010001010010100010 0000000000001010000100100111100
00000000000000000100001000001000100001001000 000000000000000000011000100001000010 0001000000010001001010000001000 0000000000000000100010001000 0000000000110000000110100000001001000010011 0000000000000000000101001000101 0000000000000000000100000010100100100101001 000000000100100000001011000010 000000000100100100000101010101000011 0000000000000001000001101000100101 000000000000000010000100010 0000000010100011000010010110011001 00000000000000000000001100001001000000 00000000001000000010000010000100 0000000000000000000010
00010011001000111100101010 000010010100010000101 0001000010001100101 000100000000000000000000 000000000101101100000 00000010011000011001000011000010000000 0000001010100001001010000100 00000010001001000100100000001000110 0010001010010001010 0000000000001100100100010101100 00000000000000100000001000001000000000001000 000000000000000100010000010001000000 0001000010000110010110010011010 0000001010000100000010001000 0000000000010001000000100010000100000010010 0000000000000000000101001100110 0000000000100001000000000010100100000101001 000001010000000001000011000000 000000010000100010000000010001000100 0000000000000001000001101000100101 000000000001000111100110010 0000000000000000100010000110101001 00000000000000000100011000000011000100 00000000000000010000001001000010 0000000001001001000110
00011111001000011101001010 100110010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011010000010101001011000010000000 0000001010100001001010000000 00000010001100000100100010001000110 0010001010010001000 0000010010110100001000000101100 00000000010000100000001000001000000000000000 000000110000100101010100010001000000 0011000010100110010010010000100 0000100010010100000010001000 1001010000100001000100000010000100000000000 0010000000000000000101000000000 0000000000100101000100000010100100010101001 000001010000000001001011000000 001000010010101000000000010101000000 0000010010000001010101101000100101 100101001001100111100010010 0000010010110000100010000110001001 00000000001001000100011000001011001000 00000100011000010000001001000100 0000000001001001001010
00011111001000011101001010 100110010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011010000010001001011000010000000 0000001010101001001010000000 00000010001100000100100000001000110 0010001010010001000 0000010010100100001000010101100 00000000010000100000001000001000000000000000 000000110100100101010100010001000000 0001000010100110010010010000100 0000100000010100000010001000 1001010000010001000000000010000101000000000 0010000000000000000101000000000 0000010000100101010100000010100100010101001 000001000000000001001011000000 001000010010101000000000010101000000 0000010010001001010101101000100101 100101001001100111100010010 0000010010110000100010000110001001 00000000001001000100011000001011001000 00000100011000010000001001000000 0000000001001001001010
00011011001000011101001010 100110010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011010000010001001011000010000000 0000001010100001001010000100 00000010001000000001000000001000100 0010001010010001000 0000010010100100001000010101100 00000000010000100000001000001000000000000000 000000110100100101010100010001000000 0101000010100110010010010010100 0000101000010100001010001000 1001000000010001000100000010000101000000000 0010000000000000000101000000000 0000010000100101010100000010100100010101001 000001010001000001001011000000 001000010010101010000010010101000000 0000010010001001010101101000100101 100101001001101111100010010 0000010010110000100010000110001001 00000000000001000100011000001011001000 00000000011000010000001001000100 0000000001001001001010
00011011001000011101001010 100010010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011010000010101001011000010000000 0000001010100001001010000100 00000010001110000100100000001000110 0010001010010001000 0000010010100100001000010101100 00000000010000100000001000001000000000000000 000000110100100101010100010001000000 0101000010100110010010010010100 0000101000010100001010001000 1001000000000101000010000010000101000000000 0010000000000000000101000000000 0000010000100101010100000010100100010101001 000001010001000001001001000000 001000010010101010000010010001000000 0000010010001001010101101000100101 100101001001100111100010010 0000010010110000100010000110001001 00000000001001000100011000001011001000 00000100011000010000001001000000 0000000001001001001010
00010011001000011101001010 100010010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011010000010101001011000010000000 0000001010100001001010000100 00000010001110000100100000001000110 0010001010010001000 0000010010100100001000010101100 00000000010000100000001000001000000000000000 000000110100100101010100010001000000 0101000010100110010010010010100 0000101000010100001010001000 1001000000100101000100000010000101000000000 0010000000000000000101000000000 0000010000100101010100000010100100010101001 000001010001000001001001000000 000000010010101010000010010001000000 0000010010001001010101101000100101 100101001001100111100010010 0000010010110000100010000110001001 00000000001001000100011000001011001000 00000100011000010000001001000000 0000000001001001001010
00011111001000011101001010 100010010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011010000010001001011000010000000 0001111010100001001010000000 00000010001110000100100010001000110 0010001010010001000 0000010010110100001000000100100 00000000010000100000001000001000000000000000 000000110000100101010100010001000000 0011000010100110000010010000100 0000101000010100000110001000 1001010000100001000100000010000101000000000 0010000000000000010101000000000 0000010000100101010100000010100100010101001 000001000000000001001001000000 001000010010101010000000010101000000 0000010010000001010101101000100101 101101001001100111100010010 0000010010110000100010000110001001 00000000000001000100011000000011001000 00000100011000010000001001000100 0000000001001001001010
00011111001000011101001010 100010010100010000101 1001000010001100001 011101000101010100000010 000101000101101100000 00000011011000010001001011000010000000 0001111010101001001010000100 00000010001100000100110000001000110 0010001010010001000 0000100000100110001000000101100 00000000010000100000001000001000000000000000 000000000100100100010100010001000000 0101000100100110010010010100100 0000001000010100000110001000 1001000000100001000100000010000101000000000 0010100000000000000001000000000 0000010000100001000100000010100100000001001 000001010001000001001001000000 001000010010110000100010010101100000 0000010011000001010101101000100101 101101000101100110100010010 0000010010110000100010000110001001 00000000001001000100011001000010001000 00000100011000010000001001000100 0000000001001001001010
10011111001000011101001000 101010010100010001001 1001000010001100001 111101000101011100000010 000101000101101100000 00000111010000011001000011010010000000 0001111010101001001010000000 00010010001110000100110000001000110 0000000000100010000 0010010010010100000100010101010 00000000010000100010001000001000000000000000 000000000100100101010000110001000000 0101000010100100010100010000100 0001101000010100000110001000 1001001000000101000100000010010001000000000 0010000000000000010101000000000 1001010000100011010100000010100100000101001 000101000000000001000101000000 001000110100110100110000010001000000 0100010011000001010101101000101001 101000000101101110100010110 0000010010100000100000100010001001 00000000000000000100001001000010001000 00000100011000010000001001000100 1000000001001000001010
10010111001000011100101000 101010010100010000101 1001000010001100001 111101000101010100000010 000101000101100100000 00001111010000011001000011000010000000 0001111010101001001010000000 00000010001110000100100000001000100 0010001010010001000 0010110110010100000100000100010 00000000000010000010001000001000000000000000 000000110000100101010100110000000000 0111000010100110010100010100100 0101101000010100001010001000 1001001000000101000100000010001001000000100 0010000000000000010101000000000 1001010000100011010100000010100100000001001 000101000000000001000101000000 001000110110110000100000010001000000 0100010010001001010101101000101001 101001000101101010000010110 0000010010100000100010100010001001 00000000000001000100001001000110001000 00000100011000010000100001000000 1000000001001000001010
10010111101000111100101000 111010010100010000101 1001000010001100001 111101000101011100000010 000101000101100100000 00001111011000010001000011000010000000 0000111010100001001010000000 00000010001110000100101000001000000 0010001010010000000 0000110100100100001000000100000 00000000000000000000001000001000000000000000 000000110100100001010100010000000000 0011000010100110010100010100100 0100111100010000101000001000 1001001000100101100100000010000000000000000 0010000000000000010101000000000 1001010001000001010100000010100100000001001 000001010001000001001001000000 000000100110101000100000010001000000 0100010010001001010101101000101001 101001001001100111000010110 0000010010110000100010000010001001 00000000000001000001000000000110000000 00000100011000010000100001000000 0000000001001000001010
10010111101001111100101000 011010010100010000101 1001000010001100001 111101000101011100000010 000101000101100100000 00001110011000010001000011000010000000 0000111010100001001010000000 00000010001110000100101000001000000 0010001010010000000 0000110100100100001000000100000 00000000000000000000001000001000000000000000 000000110100100001010100010000000000 0011000010100110010100010100100 0100111000010000001000001000 1001001000100101100100000010001000000000100 0010000000000000010101000000000
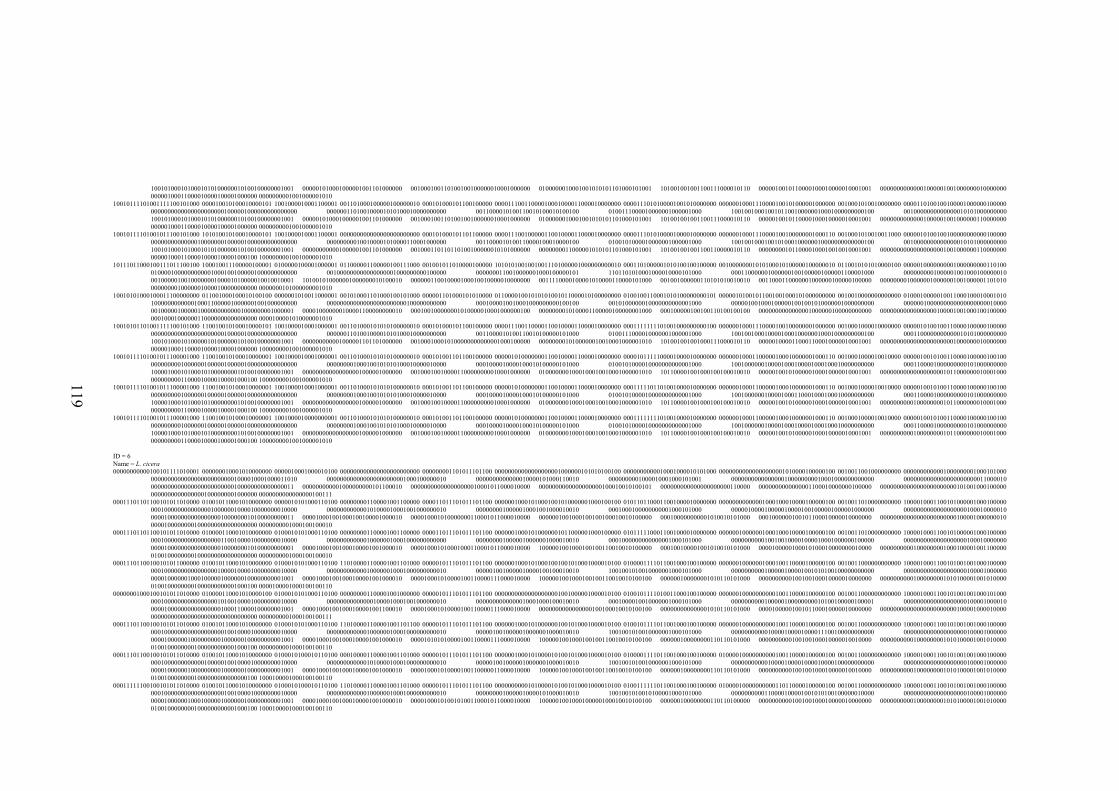
119
1001010001010001010100000010100100000001001 000001010001000001001101000000 001000100110100100100000010001000000 0100000010001001010101101000101001 101001001001100111000010110 0000010010110000100010000010001001 00000000000001000001001000000010000000 00000100011000010000100001000000 0000000001001000001010
10010111101001111100101000 000010010100010000101 1001000010001100001 001101000100000100000010 000101000101100100000 00001110011000010001000011000010000000 0000111010100001001010000000 00000010001110000100101000001000000 0010001010010000000 0000110100100100001000000100000 00000000000000000000001000001000000000000000 000000110100100001010100010000000000 0011000010100110010100010100100 0100111000010000001000001000 1001001000100101100100000010001000000000100 0010000000000000010101000000000 1001010001010001010100000010100100000001001 000001010001000001001101000000 001000100110100100100000010001000000 0100000010001001010101101000101001 101001001001100111000010110 0000010010110000100010000010001001 00000000000001000001001000000110000000 00000100011000010000100001000000 0000000001001000001010
10010111101001011100101000 101010010100010000101 1001000010001100001 000000000000000000000000 000101000101101100000 00001110010000011001000011000010000000 0000111010100001000010000000 00000010001110000100100000001000110 0010001010010011000 0000010100100100000000000100000 00000000000000100000001000001000000000000000 000000000100100001010000110001000000 0011000010100110000100010000100 0100101000010000001000001000 1001001000100101000100000010000000000000100 0010000000000000010101000000000 1001010001010001010100000010100100000001001 000000000001000001001101000000 001000110110110100100000010101000000 0000000011000001010101101000101001 101001001001100110000010110 0000000010110000100010010010001001 00000000000000000001001000000110000000 00000100011000010000100001000100 1000000001001000001010
10111011000100111011100100 100010011100000100001 0100000100001000001 011000001100000100111000 001001011010000100000 10101010010010011101000001000000000010 0001101000001010100100100000 00100000001010100010100000100000010 0110010101010000100 0000010000000001000000000110100 01000010000000000010001001000001000000000000 001000000000000000001000000000100000 0000000110010000001000100000101 1101101010001000010000101000 0001100000010000001001000001000001100001000 0000000001000001001000100000010 0010000010010000000100001010000010010010001 101001010000001000000010100010 000000110010000100010010000010000000 0011110000100001010000110000101000 001001000000110101010010010 0011000110000001000000100000100000 00000000100000010000001001000001101010 00000000100000010000100000000000 0000000101000000001010
10010101000100011100000000 011001000100010100100 0000001010011000001 001010001101000100101000 000001101000101010000 01100001001010101001011000010100000000 0100100110001010100000000101 00000101001011001001000101000000000 0010010000000000000 0100010000010011000100010001010 10000000000001000110000010000001001000000000 000000000000000000000000100000000000 0001000010010001000000000100100 0010100000010000000000001000 0000010010001000001001001010000001000000000 0000001000000000000000000010000 0010000010000010000000000001000000001000001 000010000000100001100000000010 000100100000001010000010001000000100 0000000010100001100000100000001000 000100000100100110100100100 0000000000000001000000100000000000 00000000000000000100001001000100100000 00010001000000110000000000000000 0000100001010000001010
10010101101001111100101000 110010010100010000101 1001000010001000001 001101000101010100000010 000101000101100100000 00001110011000011001000011000010000000 0001111111101001000000000100 00000010001110000100100000001000000 0010001000010000000 0000010100100110000100000100000 00000000000000000000001000001000000000000000 000000110100100001010100010000000000 0011000010100110010100000101000 0100111000010000001000001000 1001001000100001000100000010001000000000100 0001100000000000010101000000000 1001010001010000010100000010100100000001001 000000000001000001101101000000 001000100010100000000000001000100000 0000000010100000010010001000001010 101001001001000111000010110 0000010000110001100010000010001001 00000000000000000000001000000010000000 00000100011000010000100001000000 1000000001001000001010
10010111101001011100001000 110010010100010000001 1001000010001000001 001101000101010100000010 000101001101100100000 00000101000000011001000011000010000000 0000101111100001000010000000 00000010001100000100010000001000110 0010001000010010000 0000010010100110000100000100100 00000000010000001000001000001000000000000000 000000001000100101010100010000010000 0001000010000100010100000101000 0100101000010000000000001000 1001000000100001000100001000100010000000000 0001100001000000000101000000000 1000010001010001010000000010100100000001001 000000000000000001000001000000 001000100100001100000000010001000000 0100000001000100010010001000001010 101100001001000100100010010 0000010010100000100010000010001001 00000000000000000001011000000010001000 00000000011000010000100001000100 1000000001001000001010
10010111101001011100001000 110010010100010000001 1001000010001000001 001101000101010100000010 000101001101100100000 00000101000000011001000011000010000000 0001111101101001000010000000 00000010001100000100010000001000110 0010001000010010000 0000010010100110000100000100100 00000000010000001000001000001000000000000000 000000001000100101010100010000010000 0001000010000100010100000101000 0100101000010000000000001000 1001000000100001000110001000100010000000000 0001100001000000000101000000000 1000010001010001010000000010100100000001001 000000000000000001000001000000 001000100100001100000000010001000000 0100000001000100010010001000001010 101100001001000100100010010 0000010010100000100010000010001001 00000000001000000001011000000010001000 00000000011000010000100001000100 1000000001001000001010
10010111101001011100001000 110010010100010000001 1001000010000000001 001101000101010100000010 000101001101100100000 00000101000000011001000011000010000000 0001111111101001000010000000 00000010001100000100010000001000110 0010001000010010000 0000010010100110000100000100100 00000000010000001000001000001000000000000000 000000001000100101010100010000010000 0001000010000100010100000101000 0100101000010000000000001000 1001000000100001000100001000100010000000000 0001100001000000000101000000000 1000010001010001010000000010100100000001001 000000000000000001000001000000 001000100100001100000000010001000000 0100000001000100010010001000001010 101100001001000100100010010 0000010010100000100010000010001001 00000000001000000001011000000010001000 00000000011000010000100001000100 1000000001001000001010
ID = 6 Name = L. cicera 00000000000100101111010001 000000010001010000000 0000010001000010100 000000000000000000000000 000000001101011101100 00000000000000000001000000101010100100 0000000000010001000010101000 00000000000000000001010000100000100 0010011001000000000 0000000000001000000001000101000
00000000000000000000000001000010001000011010 000000000000000000000001000100000010 0000000000000010000101000110010 0000000001000010001000101001 0000000000000000100000000010001000000000000 0000000000000000000000011000010 0000000000000000000000000100000000000000011 000000000001000000000101100010 000000000000000000010001011000010000 0000000000000000000100010010100101 000000000000000000000110000 0000000000000001100010000000100000 00000000000000000000000101001000100000 00000000000000001000000001000000 0000000000000000100111
00011101101100101011010000 010010110001010000000 0000010101000110100 000000001100001001100000 000011011101011101100 00000010001010001001010000001000100100 0101101100011001000010000000 00000000000001000100010000100000100 0010011010000000000 1000010001100101000001000100000 00010000000000000010000001000010000000010000 000000000000101000010001001000000010 0000000010000010001001000010010 0001000100000000001000101000 0000010000100000100001001000001000001000000 0000000000000000000100010000010 0000100000000000000001000000010100000000011 000010001001000100100001000010 000010001010000000110001011000010000 0000001001000100100100010010100000 000100000000001010010101000 0001000000100101100010000010000000 00000000000000000000000100001000000010 00001000000001000000000000000000 0000000001000100100010
00011101101100101011010000 010000110001010000000 0100010101000110100 000000001100001001100000 000011011101011101100 00000010001010000001011000001000100000 0101111100011001000010000000 00000010000001000100010000100000100 0010011010000000000 1000010001100101000001000100000 00010000000000000000011001000010000000010000 000000000000100000010001000000000000 0000000010000010000001000010010 0001000000000000001000101000 0000000000100100100001000010001000000100000 0000000000000000000100010000000 0000100000000000000001000000010100000000001 000010001001000100001001000010 000010001010001000110001011000010000 1000001001000100100110010010100000 000100100001001010010101000 0000100000100010100010000000010000 00000000001000000001000100001001100000 01001000000001000000000000000000 0000000001000100100010
00011101100100101011000000 010010110001010000000 0100010101000110100 110100001100001001101000 000001011101011101100 00000010001010001001001010001000010100 0100001111011001000100100000 00000010000001000100110000100000100 0010011000000000000 1000010001100101001001000100000 00010000000000000000100001000010000000010000 000000000000100000010001000000000010 0000010010000010000100100010010 1001001010010000001000101000 0000000000100000100001001010100100000000000 0000000000000000000100001000000 0000100000010001000001000000100000000001001 000010001001000100001001000010 000010001010000100110000111000010000 1000001001000100100110010010100100 000000100000001010110101000 0000000000100100100010000010000000 00000000001000000001010100001001010000 01001000000001000000000001000100 0000100001000100100110
00000001000100101011010000 010000110001010000100 0100010101000110100 000000001100001001000000 000001011101011101100 00000000000000000001001000001000010100 0100101111010011000100100000 00000010000000000100110000100000100 0010011000000000000 1000010001100101001001000101000 00010000000000000000101001000010000000010000 000000000000001000010001001000000010 0000000000000010001000100010010 0001000010010000001000101000 0000000000100000100000000010100100000100001 0000000000000000000100001000010 0000100000000000000001000110000100000001001 000010001001000100001001100010 000010001010000100110000111000010000 0000000000000000100100010010100100 000000000000001010110101000 0000100000100101100010000010000000 00000000000000000000000100001000010000 00000000000000000000000000000000 0000000001000100100111
00011101100100101011010000 010010110001010000000 0100010101000110100 110100001100001001101100 000001011101011101100 00000010001010000001001010001000010100 0100101111011001000100100000 00000010000000000100110000100000100 0010011000000000000 1000010001100101001001000100000 00010000000000000000001001000010000000010000 000000000000100000010001000000000010 0000010010000010000001000010010 1001001010010000001000101000 0000000000010000100001000011100100000000000 0000000000000000000100001000000 0000100000010000000001000000100000000001001 000010001001000100001001000010 000010101010000100110000111000010000 1000001001000100100110010010100100 000000100000000110110101000 0000000000100100100010000010010000 00000000001000000001010100001001010000 01001000000001000000000001000100 0000000001000100100110
00011101100100101011010000 010010110001010000000 0100010100010110100 000100001100001001101000 000001011101011101100 00000010001010000101001010001000010100 0100001111011001000100100000 01000010000000000100110000100000100 0010011000000000000 1000010001100101001001000100000 00010000000000001000001001000010000000010000 000000000000101000010001000000000010 0000010010000010000001000010010 1001001010010000001000101000 0000000000010000100001000010000100000000000 0000000000000000000100001000000 0000100000010000000001000000100000000001001 000010001001000100001001000010 000010001010000100110000011000010000 1000001001000100100110010010100100 000000100000000110110101000 0000000000100100100010000010010000 00000000001000000001010100001001010000 01001000000001000000000000000100 1000100001000100100110
00011111100100101011010000 010010110001010000000 0100010100010110100 110100001100001001101000 000001011101011101100 00000000001010000101001010001000010100 0100111111011001000100100000 01000010000000000110110000100000100 0010011000000000000 1000010001100101001001000100000 00010000000000000000001001000010000000010000 000000000000100000010001000000000010 0000000010000010000101000010010 1001001010010100001000101000 0000000000110000100001001010100100000010000 0000000000000000000100001000000 0000100000010001000001000000100000000001001 000010001001000100001001000010 000010001010010100110001011000010000 1000001001000100000100010010100100 000000100000000110110100000 0000000000100100100010000010000000 00000000001000000001010100001001010000 01001000000001000000000001000100 1000100001000100100110
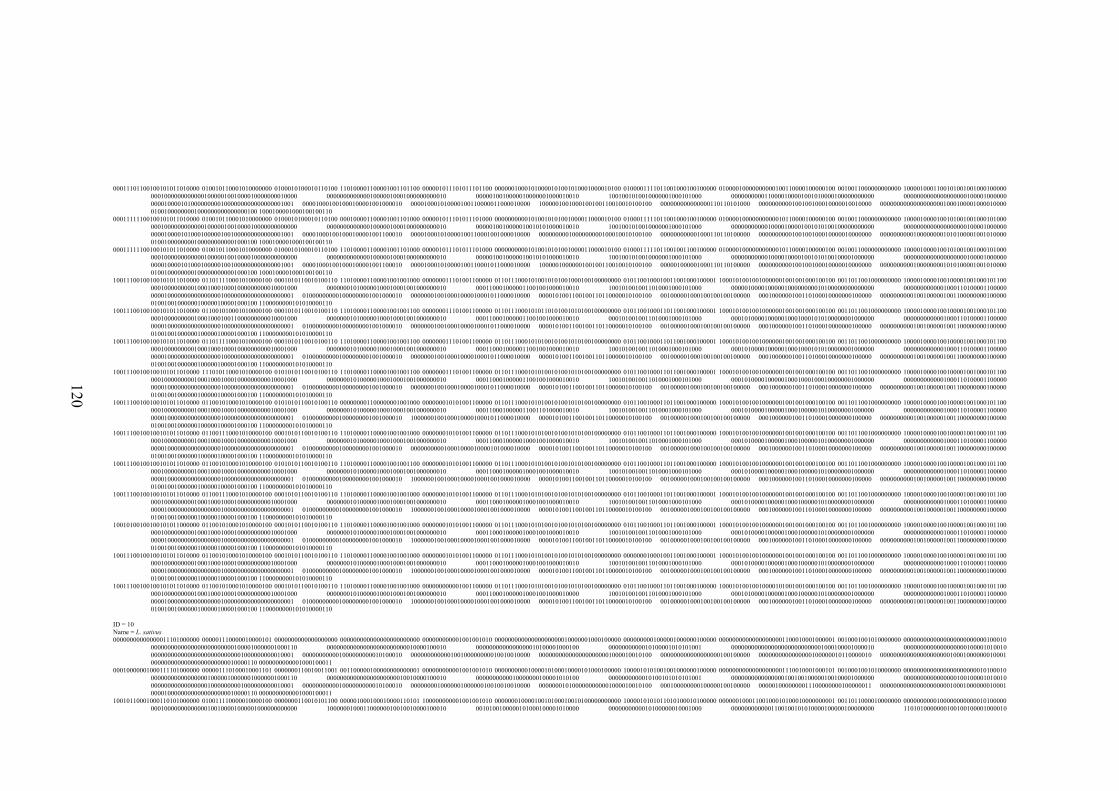
120
00011101100100101011010000 010010110001010000000 0100010100010110100 110100001100001001101100 000001011101011101100 00000010001010000101001010001000010100 0100001111011001000100100000 01000010000000000100110000100000100 0010011000000000000 1000010001100101001001000100000 00010000000000001000001001000010000000010000 000000000000010000010001000000000010 0000010010000010000001000010010 1001001010010000001000101000 0000000000110000100001001010000100000000000 0000000000000000000100001000000 0000100001010000000001000000000000000001001 000010001001000100001001000010 000010001010000100110000011000010000 1000001001000100100110010010100100 000000000000000110110101000 0000000000100100100010000010010000 00000000000000000001000100001000010000 01001000000001000000000000000100 1000100001000100100110
00011111100100101011010000 010010110001010000000 0100010100010110100 000100001100001001101000 000001011101011101000 00000000001010010101001000011000010100 0100011111011001000100100000 01000010000000000010110000100000100 0010011000000000000 1000010000100101001001000101000 00010000000000001000001001000010000000000000 000000000000010000010001000000000010 0000010010000010010101000010010 1001001010010000001000101000 0000000000010000100001001010100100000000000 0000000000000000000100001000000 0000100001010001000001001000000000000001001 000010001001000100001001100010 000010001010000100110001001000010000 0000000001000000000100010010100100 000000000001000110110100000 0000000000100100100010000010000000 00000000001000000001010100001001010000 01001000000001000000000001000100 1000100001000100100110
00011111100100101011010000 010010110001010000000 0100010100010110100 110100001100001001101000 000001011101011101000 00000000001010010101001000011000010100 0100011111011001001100100000 01000010000000000010110000100000100 0010011000000000000 1000010000100101001001000101000 00010000000000001000001001000010000000000000 000000000000010000010001000000000010 0000010010000010010101000010010 1001001010010000001000101000 0000000000010000100001001010100100001000000 0000000000000000000100001000000 0000100001010001000001001000000000000001001 000010001001000100001001100010 000010001010000100110001011000010000 1000001000000100100110010010100100 000001000001000110110100000 0000000000100100100010000010000000 00000000001000000001010100001001010000 01001000000001000000000001000100 1000100001000100100110
10011100100100101011010000 011011110001010000100 0001010110010100110 110100001100001001001000 000000011101001100000 01101110001010010101001000100100000000 0101100100010011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000011001001000010010 1001010010011010001000101000 0000010000100000100000000010100000000000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 000000010010001000010001011000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001010001010000100 0001010110010100110 110100001100001001001100 000000011101001100000 01101110001010110101001010100100000000 0101100100011011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010011000000000010001000 000000010100000100010001001000000010 0001100010000011001001000010010 0001010010011010001000101000 0001010000100000100010001010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 000000010010001000010001011000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011011110001010000100 0001010110010100110 110100001100001001001100 000000011101001100000 01101110001010100101001010100100000000 0101100100011011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000011001001000010010 1001010010011010001000101000 0001010000100000100010001010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 000000010010001000010001011000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 111010110001010000100 0101010110010100110 110100001100001001001100 000000011101001100000 01101110001010100101001010100100000000 0101100100011011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000011001001000010010 1001010010011010001000101000 0001010000100000100010001000100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 000000010010001000010001011000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001010001010000100 0101010110010100110 000000001100000001001000 000000010101001100000 01101110001010100101001010100100000000 0101100100011011001000100000 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000011001101000010010 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 100000010010001000010001011000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001110001010000100 0001010110010100110 110100001100001001001000 000000010101001100000 01101110001010100101001010100100000000 0101100100011011001000100000 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000010001001000010010 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 000000000010001000010000101000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001010001010000100 0101010110010100110 110100001100001001001100 000000010101001100000 01101110001010100101001010100100000000 0101100100011011001000100000 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000010001001000010010 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 100000010010001000010001001000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001110001010000100 0001010110010100110 110100001100001001001000 000000010101001100000 01101110001010100101001010100100000000 0101100100011011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000010001001000010010 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 100000010010001000010001001000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10010100100100101011000000 011001010001010000100 0001010110010100110 110100001100001001001000 000000010101001100000 01101110001010100101001010100100000000 0101100100011011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000010001001000010010 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 100000010010001000010001001000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001010001010000100 0001010110010100110 110100001100001001001000 000000010101001100000 01101110001010100101001010100100000000 0000000100010011001000100001 10001010010010000001001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000010001001000010010 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 100000010010001000010001001000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
10011100100100101011010000 011001010001010000100 0001010110010100110 110100001100001001001000 000000000001001100000 01101110001010100101001010100100000000 0101100100011011001000100000 10001010010010000101001001000100100 0011011001000000000 1000010000100100001001000101100 00010000000001000100010001000000000010001000 000000010100000100010001001000000010 0001100010000010001001000010000 1001010010011010001000101000 0001010000100000100010000010100000001000000 0000000000001000110100001100000 0000100000000000000001000000000000000000001 010000000001000000001001000010 100000010010001000010001001000010000 0000101001100100110110000010100100 001000001000100100100100000 0001000000100110100010000000100000 00000000001001000001001100000000100000 01001001000000100000100001000100 1100000000101010000110
ID = 10 Name = L. sativus 00000000000000011101000000 000001110000010000101 0000000000000000000 000000000000000000000000 000000000001001001010 00000000000000000000010000001000100000 0000000001000001000000100000 00000000000000000001100010001000001 0010001001010000000 0000000000000000000000000100010
00000000000000000000000001000010000001000110 000000000000000000000000010000100010 0000000000000000010100001000100 0000000000010100001010101001 0000000000000000000000000001000100001000010 0000000000000000000100001010010 0000000000000000000000000001000000000010001 000000000001000000000010100010 000000000000100100000000010010010000 0000000000000000000001000010010100 000000000000000000100100000 0000000000000000100000010110000010 00000000000000000000010001000000010001 00000000000000000000000010000110 0000000000001000100011
00010000001000111101000000 000001110100010001101 0000000110010011001 001100000100000000000001 000000000001001001010 00000000001000010100010000101000100000 1000010101001001000000100000 00000000000000000001110010001000101 0010001001010000000 0000000000000000000000010100010 00000000000000000100000100000010000001000110 000000000000000000000010010000100010 0000000000010000000100001010100 0000000000010100101010101001 0000000000000000100100100000100100001000000 0000000000000000100100001010010 0000000000000000010000000001000000000010001 000000000001000000000010100010 000000001000000100000010010010010000 0000000101000000000001000010010100 000100000000100000100100000 0000010000000011100000000100000011 00000000000000000000010001000000010001 00001000000000000000000010000110 0000000000001000100011
10010110001000110101000000 010011110000010000100 0000000110010101100 000001000100010000110101 100000000001001001010 00000001000010010100010010100000000000 1000010101011010100010100000 00000010001100100010100010000000001 0011011000010000000 0000000000100000000000010100000 00010000000000000100100001000001000000000000 100000010001100000010010010000100010 0010100100000101000100001010000 0000000000010100000010001000 0000000000001100100101010000100000100000000 1101010000000100100100001000010
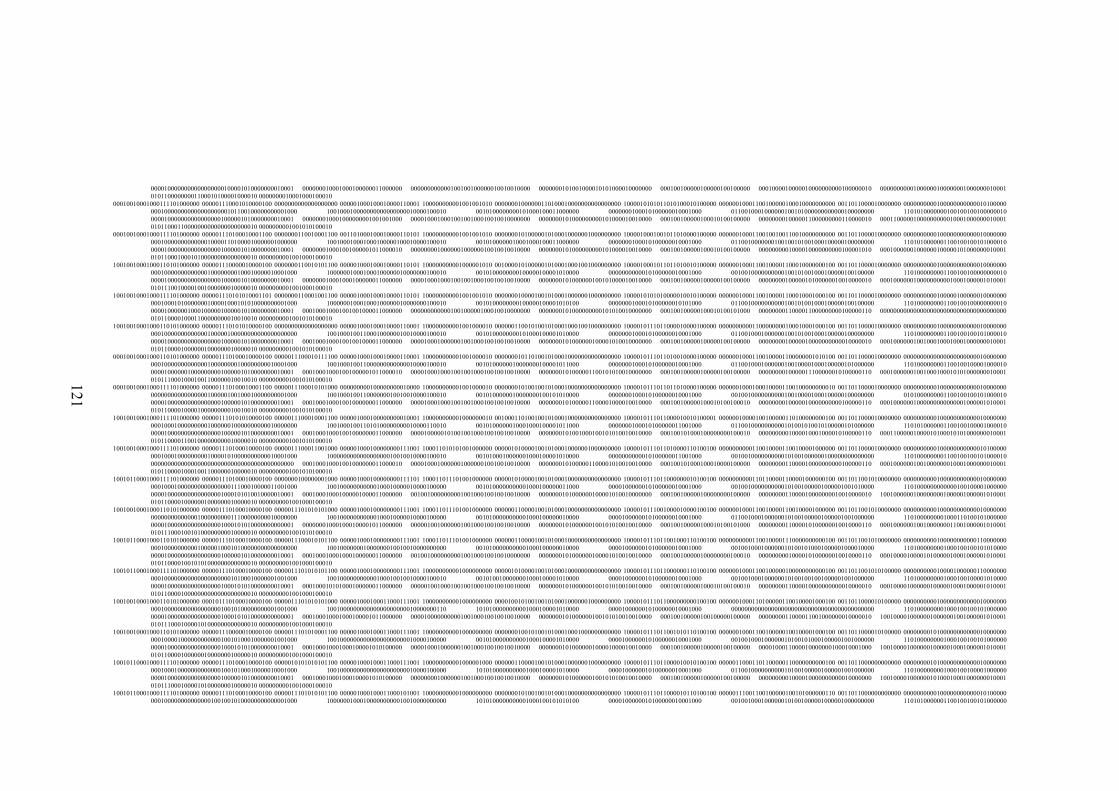
121
0000100000000000000000100001010000000010001 000000010001000100000011000000 000000000000100100100000010010010000 0000000101001000010101000010000000 000100100000100000100100000 0001000010000010000000000100000010 00000000001000000100000001000000010001 01011000000001100010100001000010 0000000010001000100010
00010010001000111101000000 000001110001010000100 0000000000000000000 000001000100010000110001 100000000001001001010 00000001000000110100010000000000000000 1000010101011010100010100000 00000010001100100000100010000000000 0011011000010000000 0000000000100000000000010100000 00010000000000000000000101100100000000001000 100100010000000000000000010000100010 0010100000000101000100011000000 0000000100010100000010001000 0110010001000000100101000000000000100000000 1101010000000100100100100000010 0000100000000000000001000001010000000010001 000000010001000000001001001000 000010001000100100100010010010000000 0000000101000000000101000010010000 000100100000100010100100000 0000000010000011000000000110000010 00011000001000000000010001000000010001 01011000110000000000000000000010 0000000001001010100010
00010010001000111101000000 000001110100010001100 0000000110010001100 001101000100010000110101 100000000001001001010 00000001010000010100010000001000000000 1000010001001011010000100000 00000010001100100100110010000000000 0011011000010000000 0000000000100000000000010000000 00010000000000000100001101000010000001000000 100100010001000100000100010000100010 0010100000010001000100011000000 0000000100010100000010001000 0110010000000100100101001000100000100000000 1101010000001100100100101000010 0000100000000000000001000001010000000010001 000000010001001000001011000010 000000001000000100000010010010010000 0000000101000000000101000010010000 000100100000100010100100000 0000000001000010000000000100001010 00010000001000000100000101000000010001 01011000100010100000000000000010 0000000001001000100010
10010010001000110101000000 000001110000010000100 0000000110010101100 000001000100010000110101 100000000001000001010 00100001010000010100010001001000000000 1000010001011011010010100000 00000010001100100001100010000000100 0011011000010000000 0000000000100000000000010000000 00010000000000000100000000100010000010001000 100000010001000100000010000000100010 0010100000000100000100001010000 0000000000010100000010001000 0010010000000000100101001000100000100100000 1101000000001100100100000000010 0000100000000000000001000001010000000010001 000000010001000100000011000000 000010001000100100100010010010010000 0000000101000000100101000010010000 000100100000100000100100000 0000000010000010100000010010000010 00010000001000000000010001000001010001 01011100100001001000000010000010 0000000001001000100010
10010010001000111101000000 000001110101010001101 0000000110001001100 000001000100010000110101 100000000001001001010 00000001000010010100010000001000000000 1000010101010000010010100000 00000010001100100001100010001000100 0011011000010000000 0000000000100000100000010000000 00010001010000000100001000101010000000001000 100000010001000100000010000000100010 0010100000000100000100001010100 0000000100010100000010101000 0110010000000000100101001000100000100100000 1101000000001100100100000000010 0000100000010001000001000001010000000010001 000100010001001001000011000000 000000000000100100000100010010000000 0000000101000000000101010010000000 000100100000100010100101000 0000000011000011000000000100000110 00000000000000000000000000000000000000 01011000010001100000000010010010 0000000001001010100010
10010010001000110101000000 000001110101010000100 0000000000000000000 000001000100010000110001 100000000001001000010 00000011001010010100010001001000000000 1000010111011000010000100000 00000000001100000000100010001000100 0011011000010000000 0000000000100000000000010000000 00010000000000000100000100000000000000000000 100100010011000100000010010000100010 0010100000000101000100001010000 0000000100010100000010001000 0110010001000000100101001000100000100000000 1101000000001100100100101000010 0000100000000000000001000001010000000010001 000100010001001001000011000000 000010001000000100100010010010010000 0000000101000000100001010010000000 000100100000100000100100000 0000000010000010000000000010000010 00010000001001000100010001000000010001 01011000010000001000000010000010 0000000001001010100010
00010010001000110101000000 000001110100010000100 0000011100010111100 000001000100010000110001 100000000001001000010 00000001011010010100010000000000000000 1000010111011010010000100000 00000010001100100001100000001010100 0011011000010000000 0000000000000000000000010000000 00010000000000000100000000100000000010001000 100100010011000000000000010000100010 0010100000010000000100001011000 0000000100010100000010001000 0110010001000000100100001000100000101000000 1101000000001100100100001000010 0000100000010000000001000001010000000010001 000100010001001000001011000010 000010001000100100100010010010010000 0000000101000001100101010010000000 000100100000100000100100000 0000000010000011100000010100000110 00010000001001000100010101000000010001 01011100010001001100000010010010 0000000001001010100010
00010010001000111101000000 000001110100010001100 0000011100010101000 000000000100000000010000 100000000001001000010 00000001010010010100010000000000000000 1000010111011011010000100000 00000010001000100001100100000000010 0011011000010000000 0000000000100000000000010000000 00000000000000000100000100100010000000001000 100100010011000000010010010000100010 0010100000010000000100101010000 0000000100010100000010001000 0010010000000000100100001000100000100000000 0101000000001100100100101000010 0000100000000000000001000001010000000010001 000100010001001000000011000000 000010001000100100100010010010010000 0000000101000001100001000010010000 000100100000100010100100010 0000000010000010000000000100000110 00010000001000000000000001000001010001 01011000010000100000000010010010 0000000001001010100010
10010010001000111101000000 000001110101010000100 0000011100010001100 000001000100000000010001 100000000001000000010 00100011010010010100010000000000000000 1000010111011000010010100001 00000010000100100000110100000000100 0011011000010000000 0000000000100000000000010000000 00010001000000000100000010000000000010000000 100100010011010100000000010000110010 0010100000010001000100001011000 0000000100010100000011001000 0110010000000000101001010010100000101000000 1101010000001100100100001000010 0000100000000000000001000001010000000010001 000100010001001000000011000000 000010000010100100100010010010010000 0000000101001000100101010010010000 000100101000100000000100010 0000000001000010001000010100000110 00011000001000010100010101000000010001 01011000011001000000000010000010 0000000001001010100010
10010010001000111101000000 000001110100010000100 0000011100011001000 000001000100000000111001 100011010101001000000 00000101000010010100010000001000000000 1000010111011010000110100100 00000000001100100001100100001000000 0011011000010000000 0000000000100000000000010100000 00010001000000000100001010000000000010001000 100000000000000000010010010000100010 0010100010000001000100001010000 0000000000010100000011001000 0010010000000000101001000000100000000000000 1101000000001100100100101000010 0000000000000000000000000000000000000000000 000100010001001000000011000010 000010001000000100000010010010010000 0000000101000001100001010010010000 000100101000100010000100000 0000000011000010000000000100000110 00010000001001000000010001000000010001 01011000010001001100000010000010 0000000001001010100010
10010110001000111101000000 000001110100010000100 0000000100000001000 000001000100000000111101 100011011101001000000 00000101000010010100010000000000000000 1000010111011000000010100100 00000000001101100001100001000000100 0011011001010000000 0000000000100000000000010000000 00010001000000000000000011100010000011001000 100100000000000100010000010000100000 0010100000000001000100000011000 0000100000010100000010001000 0010010000000000101001000001000001001010000 1101000000000000100100001000000 0000100000000000000001000101010010000010001 000100010001000001000011000000 001001000000000100100010010010010000 0000000101000000100001010010000000 000100100000100000000100000 0000000011000010000000010010000010 10010000001000000001000001000001010001 01011000010000001000000010000010 0000000001001000100010
10010010001000110101000000 000001110100010000100 0000011101010101000 000001000100000000111001 100011011101001000000 00000011000010010100010000000000000000 1000010111001000010000100100 00000010001100100001100100001000000 0011011001010000000 0000000000100000000000010000000 00000000000000100000000011100000000010000000 100100000000000100010000010000100000 0010100000000001000100000010000 0000100000010100000010001000 0110010001000000101001000001000001001000000 1101000000001000110100101000000 0000100000000000000001000101010000000000001 000000010001000100001011000000 000001001000000100100010010010010000 0000000101000000100101010010010000 000100100000100010100101000 0000000011000010100000010010000110 00010000001001000000011001000001010001 01011100010010100000000010000010 0000000001001010100010
10010110001000110101000000 000001110100010000100 0000011100010101100 000001000100000000111001 100011011101001000000 00000011000010010100010000000000000000 1000010111011001000110100100 00000000001100100001110000000000100 0011011001010000000 0000000000100000000000110000000 00010000000000100000100010100000000000000000 100100000001000000010010010000000000 0010100000000001000100000010000 0000100000010100000010001000 0010010001000000101001010001000001000010000 1101000000001000100100101010000 0000100000000000000001000001010000000010001 000100010001000100000011000000 001001000000000100100010010010000000 0000000101000000100001010010010000 000100100000100000000100010 0000000001000010100000010010000110 00010000100001010000010001000001010001 01011000010010101000000000000010 0000000001001000100010
10010110001000111101000000 000001110100010000100 0000011101010101100 000001000100000000111001 100000000001000000000 00000101000010010100010000000000000000 1000010111011000000110100100 00000010001100100000100000000000100 0011011001010100000 0000000000100001000000110000000 00010000000000000000000010100010000001001000 100100000000000100010010010000100010 0010100100000001000100001010000 0000100000010100000010001000 0010010001000000101001001001000001001000000 1101000000001000100100001010000 0000100000000000000001000101010000000010001 000100010101000100000011000000 000001001000100100100010010010010000 0000000101000000100101010010010000 000100100000100010100100010 0000000011000010000000000010000010 00010000100000010000010001000001010001 01011000010000000000000000000010 0000000001001000100010
10010010001000110101000000 000101110100010000100 0000011101010101000 000001000100011000111001 100000000001000000000 00001001010010010100010000001000000000 1000010111011000000000100100 00000010001101000001100100001000100 0011011000010100000 0000000000100000000000010000000 00010000000000000000010010100000000001001000 100100000000000000000000010000000110 1010100000000001000100001010000 0000100000010100000010001000 0000000000000000000000000000000000000000000 1101000000001000100100101000000 0000100000000000000001000101010000000000001 000100010001000100001011000000 000010000000000100100010010010010000 0000000101000000100101010010010000 000100100000100100000100000 0000000011000011001000000010000010 10010000100000010000001001000001010001 01011100010000101000000000000010 0000000001001000100010
10010010001000110101000000 000001110000010000100 0000011101010001100 000001000100011000111001 100000000001000000000 00000001001010010100010001000000000000 1000010111011001010110100100 00000010001100100000100100001000100 0011011000010100000 0000000010100000000000010000000 00010000100000000000010010100010000001001000 100100000000000000000000010000100000 0010100000000001000100001010000 0000100000010100000010001000 0010010001000000101001010001000001001000000 1101000000001000100100101000000 0000100000000000000001000101010000000010001 000100010001000100001010100000 000010001000000100100010010010010000 0000000101000000100001000010010000 000100100000100000100100000 0000100011000010000000100010001000 10010000100000010000010001000001010001 01011000010000001000000010000010 0000000001001000100010
10010110001000111101000000 000001110100010000100 0000010101010101100 000001000100011000111001 100000000001000001000 00000011000010010100010000001000000000 1000010111011000010010100100 00000110001101100000110000000000100 0011011000000000000 0000000010100000000000010000000 00010001000000000000010010100010000010001000 100100000000000000000000010000100000 1010100000000001000100001010000 0000100000010100000010001000 0110010000000000101001000001000001001000000 1101000000001000100100001000000 0000100000000000000001000001010000000010001 000100010001000100001010100000 000000001000000100100010010010010000 0000000101000000100101010010010000 000100100000100000100100000 0000000001000010000000000010000000 10010000100000010100010001000000010001 01011100010000101000000010000010 0000000001001000100010
10010110001000111101000000 000001110100010000100 0000011101010101100 000001000100011000101001 100000000001000000000 00000001010010010100010000000000000000 1000010111011000010110100100 00000111001100100000100101000000110 0011011000000000000 0000000000100000000000010100000 00010000000000000010010010100000000000001000 100000010001000000000010010000000000 1010100000000001000100101010100 0000100000010100000010001000 0010010001000000101001000001000001000000000 1101010000001100100100101000000
122
0000100000000000000001000101000000000010001 000100010001000000000011000000 000010000000000100100010010010010000 0000000000000000000000000000000000 000100100000100000100100000 0000100010000010000000010010000000 10010000100001000100010001000001010001 01011000010010101000000000000010 0000000001001000100010
10010110001000110101000000 000001110100010000100 0000011101010101100 000001000100011000101001 100000000001000000000 00000101000010010100010000010000100000 1000010111011001010000100100 00000110001100100001110010001000100 0011011000000100000 0000000000100000000000010000000 00010001000000000000000010100010000000000000 100000000000000000000010010000100000 1010100000000001000100001010000 0000100000010100000010001000 0010010001000000101001010001000100001000000 0000000000000000000100000100100 0000100000010000000001000001010000000010001 000100010001000100001011000000 000010001000000100100010010010010000 0000000101000000100001010010010000 000100100000100000000100000 0000000010000010000000010010000000 10010000100001010100010001000001010001 01011000010001000000000010000010 0000000001001000100010
ID = 14 Name = L. chrysanthus 00000000000000000000000000 000000010000010000101 0000100000011010011 000000000000000001000100 000000000000000000000 00000000000000000001010000000000100000 0000000000000000010000000100 00000000000000000000000000100000010 0000001000010100010 0000000000000000000000100000000
00000000000000000000101010000000000010000001 000000000000000000001000001000010000 0000000000000000000000000000000 0000000000000000010000101000 0000000000000000000000000000000000000000000 1101000000001000100100101000000 0000000000000001010000000001001010001010001 000000000001000000100100001000 000000000000010010000001001010010010 0000000000000000000000000000000000 000000000000100001010001001 0000000000000000001000000000010010 00000000000000000000000000000000000000 00000000000000000000000000000100 0000000000000001000010
00000000000000000000000000 000000010000010000101 0000100000011010011 000000000000000001000100 000000000000000000000 00000000000000000001000000000000000000 0000000000000000000000000000 00000000000000000000000000000000000 0000001000010100010 0000000000000000000000100000000 00000000000000000000101010000000000010000101 000000000000000000001000001000010000 0000000000000000000000000000000 0000000000000000010000101000 0000000000000000000000000000000000000000000 0000000000000000000000000000000 0000000000000000000000000000000010000000001 000000000001000000000000001000 000000000000010010000001001010010010 0100000000000000000000000000000000 000000000000100000010001001 0000000000000000000000000000000000 00000000000000000000000000000000000000 00000000000000000000000000000000 0000000000000000000010
00000000001001000000000000 000000010000010000101 0000100000011010011 000000000000000001000100 000000000000000000000 00000000001000000101000010100000100000 0000000000000000010000000100 00000000000000000001000000100000101 0000001000010100010 0000000000000000000000100100100 00000000000000000000101010000000000010000101 000000000000000000001000001000010000 0000000000000000000000000000000 0000000000000000010000101000 0000000000000000000000000000000000000000000 0000000000000000000100001100100 0000000000000001010000000001001010001010001 000000000001000000000000001000 000000000000000010000100001010000010 0000000000000000000000000000000000 001100000000100101010001001 0000000000000000001000000010000110 00000000000000000000000000000000000000 00000000000000000000000000000100 0000000000000001000010
ID = 15 Name = L. cassius 00000000000100110101000000 000000000100010000111 0000000000001100011 000000000000000000000000 000000000101000001010 00000000001000010001000011000100000000 0000000000001000100010001000 00000000000000000010110000000000010 0010001001000001010 0000000000000010000001010110100
00000000000000000000001000001000001001010000 000000000000000000000010010001000010 0000000000000001010000100100001 0000000000000000010000111000 0000000000000000000000000000000100001000000 0000000000000000010100001001001 0000000000000000010000100010001010001001001 000000000000000000100010100000 000000010000010000100010101000100010 0000000000000000000100100000101001 000000000000100101010101010 0000000000000000000010000010001011 00000000000000000000011001000001001001 00000000000000000000000000000100 0000010000000000001011
00000000000100110101000000 000000000000010000111 0000000000000100011 000000000000000000000000 000000000101000001010 00001000000010010101001001000100000000 1000000000101000100010001000 00000010000010000010110001000000010 0000001001000001010 0000000000000010000001010110100 00000000000000000000001000000010001001010000 000000000000000000001010010001000010 0000000000010101000100000010000 0000000000000000010000111000 0000000000000000000000010010000100000001000 0000000000000000010100001001001 0000000000000000010000000010001010001001001 000000000000000000100010100000 000000000100010000100000101000100010 0000000000000000000100100000101001 000000000000100101010101010 0000000000000000000010000010001011 00000000000000000000011001000001001001 00000000000000000000000000000000 0000010000000000001011
00000000000100110101000000 000000000000010000111 0000000000000000000 000000000000000000000000 000000000101000001010 00000000000010010101001001000100010100 0000000000101000100010001000 00000010000000000000110000000000010 0000000001000001010 0000000000000010000001010110100 00000000000000000000001000000000001001010000 000000000000000000000010010001000010 0000000000000101000000001000000 0000000000000000010000111000 0000000000000000000000000000000000001010000 0000000000000000010100001001001 0000000000000000010000100010001010001001001 000000000000000000100010100000 000000000100010000100010101000100010 0000000000000000000100100000101001 000000000000100101000101010 0000000000000000000010000010001011 00000000000000000000011001000001010001 00000000000000000000000000000000 0000000000000000001011
00000000000100110101000000 000000000100010000111 0000000000000100011 000000000000000000000000 000000000101000001010 00000000000000010000001001001100010000 0000000000001000100010001000 00000000000000000000110000000000010 0010000001000001010 0000000000000010000001010110100 00000001000000000000001000000000001001010000 000000000000000000000010010000000010 0000000000000000000000000010000 0000000000000000010000111000 0000000000000000000000001000000100000000000 0000000000000000000100001001001 0000000000000000000000100010001010001001001 000000000000000000100010100100 000000000000010000100010001000100010 0000000000000000000100100000101001 000000000000100001000101010 0000000000000000000010000010001011 00000000000000000000000000000001010001 00000000000000000000000000000000 0000000000000000001011
01010000001100110101000000 010000010100010000010 0000000010010100001 000101000101000001100000 010000000101000001000 00001010010010100101001001000000000000 1000010010100000100010001000 01000000000100000100100000000000010 1101000100000000100 0000000001011010000001100100000 00100001000000000000100000000100000010100000 001000000010101001000010010000000000 0000000000000101000000000100000 0000011010000000100000101000 0000000000000000000000000000000000000000000 0000000100010000110100001010000 0010000001010000010001000001100010101000001 000000000000101100101010100000 000000101100010010010000100001000000 0000000000100001000100100000101001 100100001100110101010101000 0100000101000000000010000000001001 00011010001000010000010100000001000000 00000010000010000000000010000010 0010010001000010001010
01010000001100110101000000 010000010000010000010 0000000000000000000 000101000101000001100000 010000000101000001000 00001010010010100101001001000000000000 1000010010100000100010001000 01010110000100000100100000000000010 1101000100000000100 0000000001011010000001100100000 00100001000000000000100000000000000010100000 001000000000100001010010000000000000 0000010010000101000000000100000 0000011010000000100000101000 0001001001000000100000000001000001000000000 0000000100010000110100001010000 0010000001010000010001000001001010101000001 000000000000101100101010100000 000100101100010010010000100001000000 0000000000100001000100100000101001 100100001100110101010101000 0100000101000100000010000000001001 00011010001000010000010100000001000000 00000010000010000000000010000010 0010010001000010001010
01010000001100110101000000 010000001000010000010 0000000010010100010 000101000101000001100000 010000000101000001000 00001010010010101101001001100010000000 1000010010100000100010001000 00000000000100000100100001001000000 1101000110000000100 0000000001011010000001100100000 00100000000000000000100001000100000010100000 000000000000000000010010000000000000 0100010010000101000100000100000 0000011010000000100000101000 0000001001001000100010000001001000000000000 0000000100010000110100001010000 0010000001010000010001000001100010101000001 000000001000101100101010100000 000100000000000000000000000000000000 0000000000100001000100100000101001 100100001100110101010101000 0100000101000100000010000000001001 00011010001000010000010100000001000000 00000010000010000000000010000000 0010010001000010001010
00000000001000110101000000 010000001000010000010 0000000000000000000 000101000101000001100000 010000000101000001000 00001010010010101101001001100000000000 1000010010100000100010001000 00000000000100000100100010000000000 1101000110000000100 0000000001011010000001100100000 00100000000000000000100000000100000010100000 001000000000010001010010010000000000 0100010010000101000100000100000 0000011000010000100000101000 0001001001001000100000010001000001000001000 0000000100010000110100001010000 0010000001010000010001000001001010101000001 000000000000000000000000000000 000100101100010010010000100001000000 0000000000100001000100100000101001 100100001100110101010101000 0100000101000100000010000000001001 00011010001000000000010100000001000000 00010010000010000000000010000000 0010010001000010001010
01010000001100110101000000 011000000101010000010 0001000010010100000 000101000101000001100000 010000000101000000000 00001010010010101101001001100000000000 1000010010100000100010001000 10000000000100000100100000000000010 1101000110000000100 0000000001011010000001100100000 00100001000000000000100001001000000010100000 001000000011110001010010000000000000 0101010010000101000100000100000 0000011000000000100000101000 0001001001001000100000010001000001000001000 0000000100010000110100001010000 0010000001010000010001000001001010101000001 000000000100101100101010100000 000100101100010010010000100001000000 0000000000100001000100100000101001 100100001100110101010101000 0100000101000100000010000000001001 00000000001000000000010100000001000000 01000010000010000000000010000000 0010010001000010001010
01010000001100110101000000 011000000101010000010 0001000010010100000 000101000101000001100000 010000000101000000000 00001010010010101101001001100000000000 1000010010100000100010001000 10000000000100000100100000000000010 1101000110000000100 0000000001011010000001100100000 00100001000000000000100000001000000010100000 001000000011110001010010010000000000 0101010010000101000100000100000 0000011010000000100000101000 0000001001001000100000010001000001001000000 0000000100010000110100001010000 0010000001010000010001000001000010101000001 000000000100101100101010100000 000100101100010010010000100001000000 0000000000100001000100100000101001 100100001100110101010101000 0100000101000000000010000000001001 00011010001000010000010100000001000000 00010010000010001000000010000010 0010010001000010001010
01010000001100110101000000 010000011101010000010 0001000010010100000 000101000101000001100000 000000000101000000000 00001010010010101101001001100000000000 1000010010100000100010001000 00000100000100000100100010000000000 1101000010000000100 0000000001011010000001100100000 00100001000000000000100000001000000010100000 001000000011010001010110000000000000 0100010010000101000100000100000 0000011010000000100000101000 0000000000000000100000000000000000000000000 0000000100010000110100001010000 0010000001010000010001000001001010101000001 000000000100101100101010100000 000100101100010110010000100001000000 0000000000000000000000100000101000 100100001100110101010101000 0100000101000100000010000000001001 00011010001000010000010100000001000000 00010010000010001000000010000010 0010010001000010001010
01010000001100110101000000 010000011001010000010 0001000010010000000 000101000101000001100000 010000000101000000000 00001010010010101101001001100000000000 1001010010100000100010001000 10000000000100000100100000000000010 1101000010000000000 0000000001011010000001100100000 00100000000000000000100000001000000010100000 000000000010010001010110010000000000 0100010010000101000100000100000 0000011010000000100000101000 0000001001000000100000010000100001000000000 0000000100010000110100001010000
123
0010000001010000010000000001001010101000001 000000000100101100101010100000 000100101100010010010000100001000000 0000000000100001000100100000101000 100100001100110101010101000 0100000101000100000010000000001001 00011010001000010000010100000001000000 00010001000010001000000010000000 0010010001000010001010
ID = 17 Name = L. gorgoni var gorgoni 00010110101100111001100000 000000011100010000000 0000010000000100100 000000000100000000100001 000000000001100000000 00000001001000100001001000000100100000 0001100011010010001000100000 00000000000000001000101000000000010 0011001000010000000 0000000000001010100001100100000
00100000000001000000010000000110000001000100 000100000000000100000000000000000000 0000010000100010000101000010000 0000010000010000010000011000 0000001001100000000000101000000000000010000 0000000001010100001000010101000 0000100100000001000001000001001001000000001 000010000000000001000000000000 000000100000000000010000110000000010 0000000000001001010001010000100100 000000000001000000000010001 0000000000000010001010000100001010 00000000000001000000010001001000010000 00010001000001000000000000000100 0000000001000010100010
ID = 18 Name = L. pseudo-cicera 00010111000100111001100000 000000000100010100010 0000010000000100100 000000000000000000100000 000000000001100000010 00000010010010100101001010000100100000 0000000011001000100000100000 00000000100000100100111000001001010 0011001000010000000 0000000000000000100001000100000
00000000000001000000000000000110000001000100 000100000000000000000000000100000001 0000000000100001000101010010000 0000010000010000010000001000 0000000000000000000001101000010000000000000 0000000000000100000000010101001 0000000000000000000001000000001001000100001 000010000000000001000000000000 000000000000000100000010100000000010 0000000000001000010001000000100100 000000000001000000010010001 0000000000000000001010000100001010 00000000000000000000010001000000010000 00000000000000000000000000000100 0000000000000010100010
00010100001100110100000000 000000000100000000000 0000010000000000001 000000000000000000000000 000001000001000000010 00000001010000100100011001110100000000 0001000001100000001000010001 00000000000001000010000100000001000 0011001000010000000 0000000100011000100001100100000 01000000000001000000000100000000000010000000 000000000000000000000110000100000001 0000000000000110000001001001000 0000000010010000011100011000 0000000000000000000000101000010000000000000 0000000000000100100000010101001 0000000000000000010100000001001001000010001 000010000000000001000000000001 000000000010000100000010010001000100 0000000000000001000001000000100000 000000000000000001100010001 0000000000000001001010000100001011 00000000000000000000000001001000000000 00000000000000000100000000000100 0000000000001010100010
ID = 20 Name = L. hirsutus 01010111000100110101000000 000000000100010000000 0000000010000100010 000011000101000100100010 010011000101000000000 00000100000000001001001001010000000000 1000010011001000100010000000 00000000000000000110101001000000000 1001001010000000001 0000000011000010000000000011000
00010000000000000000100000000010000000100000 000000000000100001000000000100000000 0101010000010000001000001000000 0000001000010010100000001000 0000000010000000100010000010000000000010000 0000001000100000100100010000000 0000000000000100010000001000000010000010001 000000000000000001101000001000 000000101010000100000110100001000000 0000000000001001000011010000100000 000000000100110000000000000 0000000000100000000010000000101000 00000000000010010001000100001000100000 00000010010011010000001010010001 0000000001000000000010
00010111000100110101000000 000000000000010101010 0000000000000100000 000000000000000000000000 000001000100000100010 00000000010000000000001010010100000000 1000000001011000100000101001 00000000000000001110000010000000000 0001001010000000110 0000000000000011000000000011000 00110000000000000000100000001000100000100010 000000000000100000000000000100000000 0001010000010000001000001000000 0000000000010010100000001000 0000000000000000000010000010000000000010000 0000000000100100000000000000110 0000000000000000000000000000000010000010001 000000000000000001101000001010 000000000010000010000100100001000000 0000000000000001000001000000101000 000000000000100000000100000 0000000000000000000010000000101000 00000000000010000001000100001001100010 00000000010001000000001000010001 0000000001000010000011
ID = 22 Name = L. chloranthus 00010000000100010100000000 000000000000010101000 0000000000001000000 000000010000000000000000 000000000100100100001 00000001001000001001000001000000100100 0000000101000010100000011001 00000000000000000000000100000000000 0000000000000000010 0000000000001010000100000010000
00000000000000000000100000000100101000100000 000000000000000000000000000100010000 0000000010000100001000001000000 0000000010000000000000011000 0000000000000000000000100001000001000010000 0000000000100001001001100000010 0000000000000000010000000000100010000010001 000000000000000000000101001010 000000000000001010000100010010000010 0000000000000001000101010000101000 000000000000101000000000010 0000000000000000000010010000101001 00000000000000000000000000000010000100 00000000010000000100000000010010 0000000000000010000010
ID = 23 Name = L. trachycarpus 00010000000100110101000000 000000010000010000100 0000000000000000000 000001000100000000000000 000000000100100000000 00000000000010000000011001000010000000 0000000100000010100000001001 00000000000000000000000000000000000 0000000010000000010 0010000000001000001001000000001
01110000000000000000100000000100100000000000 000000000000100101000000000000000100 0000000000000000000000000000000 0000000100000000100010111000 0000000000000000100000000001000100001000000 0000000000100000101000010001000 0000000000000001010000000010000010000000001 000000000000000100000100100010 000000000000000100000000010000000000 0000000000000100000101010000101000 000000000000101001010000010 0000000000000101001010000000010110 00000000000000010000001000001001000000 00000001010010000000000010000000 0000000000000000000010
ID = 24 Name = L. boissieri 10010000000100010000000000 000010101000000000100 0100000100000000000 000000000000000100110000 001000100000000000000 00010000000010000010000001000000000000 0000000010000101000001100010 00000000000000000000000000000000000 0010101000000000000 0001000000000000010010011010000
00000000001000000001000010000000010000000000 000000000000000001000000000100001000 0000000000000010000010000000000 0000000000000010000000000000 0000000100000010001000000110000000010001000 0000000000000001000010000000000 0000000001001000000000100100000000000000001 000000000000000010001000000000 000000000001010001001000010000001000 0000000000000010000100010001000000 000000010010011000000000000 0000000000101000010001001000000000 00000000100001000000110000100000000000 00000001000000000000000100001000 0000001000000000000100
ID = 26 Name = Vicia sativa 00010000010100100000000000 000000000010001000000 0000001100100000000 000000010010100010000010 000000000000000100000 00000000100100100100110100000001000000 0010000100000000100000000000 00000000000000010010000000010000000 0010000100000100000 0000000001000000000000101000000
01001000000000000000001000010000000000000000 010010000001000001000000000001000000 0000001001001000101000000000000 0000100100100010010000000010 0000000000000000010000010000100000000000000 0000000010000010000000100000000 0100001000000001001000010000000001000000101 000000100010010000010000010000 000000000000000000010000000001000000 0000010000010010001000000100000000 010110100001100010000000000 0011001011000000100100100010000000 01000101000100101010010010010000000000 01000000000100000001010000100000 0001100010001000010100

124
EK 2 SDS-PAGE bant verileri matrisi LATHYRUS 12 populations with primer and 52 loci Number of populations = 12 Number of loci = 52 locus name = TP1 TP2 TP3 TP4 TP5 TP6 TP7 TP8 TP9 TP10 TP11 TP12 TP13 TP14 TP15 TP16 TP17 TP18 TP19 TP20 TP21 TP22 TP23 TP24 TP25 TP26 TP27 TP28 TP29 TP30 TP31 TP32 TP33 TP34 TP35 TP36 TP37 TP38 TP39 TP40 TP41 TP42 TP43 TP44 TP45 TP46 TP47 TP48 TP49 TP50 TP51 TP52 ID = 1 Name = L.cassius 0000000000000101000000000100000100000000001100100101 ID = 2 Name = L.annuus 0011000001000011001010000100000101000010001001010100 0111000101000011101010000100000101000010001001010100 0000000001000011101010000100000101000010001001010100 ID = 5 Name = L. cicera 0000100000000001000000001000010000000000011001001000 0000100000001001000000001001010000000000001001001000 ID = 8 NAME = L.trachycarpus 0001010000000101001001000001100010000000010010100100 ID = 9 NAME = L. pseudo-cicera 0001011010001001001010000000001000000000010100100100 ID = 10 NAME = L. gorgoni var gorgoni 0000000100000001000101010001001000000000110001001000 ID = 11 NAME = L. chrysanthus 0001001000000011000100000000100000000000010010010000 ID = 12 NAME = L. sativus 0101000010010101001000001000010000000000001001010100 0101000010010101001000001000010000000000001001010100 ID = 14 Name = L. chloranthus 0101010110000011010010000000010000000100001001010010 ID = 15 Name = L. hirsutus 0010100000000101000100000000010010100000001001010000 ID = 16 Name = L.boissieri 0000000010001000000100000010000010000000000010100000 ID = 17 Name = Vicia sativa 1100100000100001000010100000000000011001000010100100

125
ÖZGEÇMİŞ
Adı Soyadı: İsmail Ömür ACAR
Doğum Yeri: Doğubeyazıt
Doğum Tarihi: 09/03/1969
Medeni Hali: Evli
Yabancı Dili: İngilizce
Eğitim Durumu (Kurum ve Yıl)
Lise : İzmir Tepecik Sağlık Meslek Lisesi (1986)
Lisans : Ankara Üniversitesi Fen Fakültesi Biyoloji Bölümü (1998)
Yüksek Lisans: Ankara Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim
Dalı (2005)
Çalıştığı Kurum/Kurumlar ve Yıl
S.B. Ankara Eğitim ve Araştırma Hastanesi 1993-Halen
Yayınları (SCI ve diğer)
Acar, I. O. Colak, R. and Buyukkartal, H. N. 2006. Electrophoretic Investigation of Isoenzymes and Soluble Proteins Between Two Varieties of Vicia sativa L. Seeds. Pakistan Journal of Biological Sciences, Volume 9, Number 13, 2432-2437,
Ulusal Konferans ve Seminerler
Acar, İ. Ö. Çolak, R. ve Büyükkartal, H. N. 2006. Vicia sativa L. nın iki çeşidi (Karae1çi ve Sarıelçi Fiğleri)’nin Tohumları arasındaki İzoenzim ve Çözünür Proteinlerin Elektroforetik İncelenmesi. 8. Ulusal Biyoloji Kongresi, 26-30 Haziran, Kuşadası/Aydın
Acar, İ. Ö. 2006. Bitkilerde Moleküler Polimorfizm. Doktora Semineri, Ankara
Acar, İ. Ö. 2004.Tohum Gelişiminin Genomik Açıdan İncelenmesi. Yüksek Lisans Semineri, Ankara