Embed Size (px)
Citation preview

CICLAGEM DE NUTRIENTES POR PLANTAS DE COBERTURA E
COMPARTIMENTOS DE CARBONO EM SISTEMA DE ROTAÇÃO SOJA
MILHO EM ÁREA DE TRANSIÇÃO CERRADO
Pesquisador Proponente (Coordenador):Fabiano André Petter CPF: 906.242.971-87 Eng. Agr., Dr. Agronomia, ProfFone: (66) 9976-2325 [email protected] Instituição executora: Universidade Federal de Mato Grosso Instituto de Ciências Agrárias e Ambientais CNPJ: 33.004.54010001-00Av. Alexandre Ferronato, 1200, Setor IndustrialCEP: 78557-267 – Sinop-MTFone: (66) 3531-1663 Local da Pesquisa: Sinop- Valor financiado pela Fundação Agrisus Período de execução: outubro
Relatório Parcial – 1617/15
CICLAGEM DE NUTRIENTES POR PLANTAS DE COBERTURA E
COMPARTIMENTOS DE CARBONO EM SISTEMA DE ROTAÇÃO SOJA
MILHO EM ÁREA DE TRANSIÇÃO CERRADO-AMAZÔNIA NO MATO
GROSSO.
(Coordenador):
, Prof. Adjunto da Universidade Federal de Mato Grosso
Federal de Mato Grosso – UFMT Instituto de Ciências Agrárias e Ambientais - ICAA
00 Av. Alexandre Ferronato, 1200, Setor Industrial
MT – Brasil
-MT
ado pela Fundação Agrisus: R$ 20.000,00
outubro de 2015 a outubro de 2017.
CICLAGEM DE NUTRIENTES POR PLANTAS DE COBERTURA E
COMPARTIMENTOS DE CARBONO EM SISTEMA DE ROTAÇÃO SOJA-
AMAZÔNIA NO MATO
Mato Grosso

1. Introdução
A matéria orgânica do solo (MOS) apresenta um papel importante no
condicionamento de propriedades físicas, químicas e biológicas em solos tropicais, por
exemplo, no incremento da capacidade de troca catiônica (CTC), na estruturação do solo e
retenção de umidade, além de constituir o principal compartimento para o carbono orgânico
sequestrado no solo (Bowmann & Germon, 1998). A MOS possui potencial para ser
utilizada como atributo chave da qualidade do solo (Doran & Parkin, 1994), pois além de
satisfazer o requisito básico de ser sensível a modificações pelo manejo do solo, é ainda
fonte primária de nutrientes às plantas.
Para regiões como o Cerrado e no ecótono Cerrado-Amazônia a atividade
microbiana é acelerada pelas condições edafoclimáticas, além de queimadas que ocorrem
naturalmente ou são praticadas, resultando em baixa qualidade e a quantidade de MOS, o
que compromete o sucesso dos sistemas agrícolas. Diante disso, o cultivo de culturas
demandantes como a soja e o milho vem ao longo dos anos passando por alterações nas
técnicas de manejo. Dentre as inovações, está a introdução do sistema plantio direto (SPD),
que preconiza a rotação de culturas, não revolvimento do solo e o uso de plantas de
cobertura. A adoção do sistema de plantio direto (SPD) tem proporcionado a manutenção
dos estoques de carbono no solo (Prior et al., 2004) e consequentemente melhorando os
atributos químicos (Henrichs et al., 2005), físicos e biológicos do solo (Silva et al., 2007).
Nesse sentido, é evidente a importância e a contribuição da matéria orgânica para a
disponibilidade de nutrientes para as plantas. Importante ainda ressaltar, que a baixa
eficiência no uso de nutrientes, implica diretamente no maior uso de recursos ambientais,
pois, as fontes para a fabricação de fertilizantes químicos, constituem-se de um recurso não
renovável.
Como o SPD tem como requisito básico o não revolvimento do solo, rotação de
cultura e o uso de plantas de cobertura visando a formação de palhada sobre o solo, estas
podem afetar a dinâmica da MOS e a disponibilidade de nutrientes para culturas
subsequentes, seja pela alteração dos compartimentos de carbono e/ou pela ciclagem de
nutrientes. Assim, o uso de plantas de cobertura pode por disponibilizar maiores
quantidades de amônio ao solo, via processo de mineralização do N, redução da lixiviação

de potássio e nitrato, maior disponibilidade de fósforo, cálcio, magnésio e outros nutrientes,
e, alterações nos compartimentos de carbono pode viabilizar de forma sustentável, a
produção do milho em rotação com soja. Importante ressaltar, que para a região do Centro
Norte de MT pesquisas dessa natureza ainda são escassas, e tal fato requer atenção, pois
trata-se de uma região destaque no cenário mundial de produção de grãos.
2. Material e Métodos
2.1 Experimento
O experimento está sendo conduzido a campo em Sinop, MT, e em Biomas que
envolvem o ecótono Cerrado-Amazônia, sobre Latossolo Vermelho Amarelo distrófico
(LVAd).
O delineamento experimental é o de blocos ao acaso, sendo a fonte de variação
constituída por oito plantas de cobertura (Crotalaria espectabilis, Cajanus cajan, Mucuna
aterrima, Fagopyrum esculentum, Urochloa ruziziensis, Urochloa brizantha, Pennisetum
glaucum, Elusine coracana) em nove épocas de avaliação (0, 15, 30, 45, 60, 90, 120, 150 e
180 dias após a dessecação de manejo), com quatro repetições. Nas parcelas principais
estarão alocadas as plantas de cobertura e nas subparcelas as épocas de avaliação.
Para carbono orgânico dissolvido, lixiviação de potássio e nitrato será coletada
amostras em quatro profundidades de análises (0-20; 20-40; 40-60 e 60-80 cm), com quatro
repetições a ser realizado aos 90 dias após a dessecação de manejo.
2.2 Implantação, condução do experimento e características avaliadas:
2.2.1 Implantação e condução das culturas e das plantas de cobertura
As culturas da soja (safra 2015/2016) e do milho (2016/2017) foram e serão
cultivadas no verão sobre a palhada de plantas de cobertura que serão dessecadas antes do
plantio destas (dessecação de manejo), e, ao longo de 0; 15; 30; 45; 60; 90, 120; 150 e 180
dias após a dessecação (durante o ciclo da cultura da soja e do milho) serão avaliadas as
características descritas adiante. Assim será possível verificar a dinâmica de liberação de

nutrientes pelas plantas de cobertura ao longo do ciclo das culturas da soja e do milho,
podendo correlacionar com os períodos de maior demanda dessas culturas.
2.2.2 Características a serem avaliadas
Plantas de cobertura – em cada época de avaliação (0; 15; 30; 45; 60; 90, 120, 150
e 180 dias após a dessecação) será determinado: (a) fitomassa seca dos restos culturais; (b)
percentagem de cobertura do solo das plantas de cobertura; (c) dinâmica de decomposição
dos restos culturais das plantas de cobertura; (d) acúmulo e tempo de liberação dos
nutrientes.
Culturas da soja e milho - em pleno florescimento da cultura da soja e do milho
será determinado: (e) fitomassa seca de plantas de milho; (f) altura de plantas; (g) área
foliar; (h) teor de clorofila; (i) acúmulo de nutrientes.
Lixiviação - nas profundidades de 0-20; 20-40; 40-60 e 60-80 cm serão
determinados: (j) lixiviação de potássio; (k) lixiviação de nitrato e (l) e carbono orgânico
dissolvido nas parcelas correspondentes a cada planta de cobertura aos 90 dias após a
dessecação das plantas de cobertura.
Matéria orgânica do solo – em pleno florescimento da soja e do milho (2016 e
2017) será realizado: (m) fracionamento químico da matéria orgânica do solo; (n) carbono
lábil e (o) índice de manejo do carbono e humificação. Na colheita serão avaliados: (p) os
componentes de produção e (q) produtividade da soja e do milho.
Características físicas do solo - em pleno florescimento da soja e do milho (2016 e
2017) será realizado: (r) a resistência à penetração, (s) porosidade total, (t) densidade e (u)
retenção de umidade do solo;
O organograma de cultivo das plantas de cobertura (PC) e das culturas está
representado no esquema abaixo:

2.3 Análises estatísticas
Estão sendo efetuadas análises de variância sendo as médias significativas
agrupadas pelo critério de Tukey. Também serão realizadas análises de regressão,
aplicando o teste “t” de Student (p<0,05), bem como do coeficiente de determinação (R2)
associado a cada modelo de regressão para as épocas de avaliação.
PC entressafra 2015
Dessecação PC out/2015
Plantio Soja out-nov/2015
safra 2015/2016
colheita Soja fev-mar/2016
safra 2015/2016
PC entressafra
safra 2015/2016
Dessecação PC out/2016
Plantio Milho out-nov/2016
safra 2016/2017
colheita Milho fev-mar/2017
safra 2016/2017
PC entressafra
safra 2016/2017
PC entressafra 2014
Dessecação PC out/2014
Plantio Milho out-nov/2014
safra 2014/2015
colheita Milho fev-mar/2015
safra 2014/2015
Safra 2014/2015 – já realizado
Continua safra 2017/2018

3. Resultados e Discussão
3.1. Dinâmica da fitomassa e de nutrientes das plantas de cobertura
Houve diferenças significativas na produção de fitomassa seca (FS) das plantas de
cobertura no ponto de rolagem (no momento da dessecação de manejo - 0 DAD), com
destaque para U. brizantha (24.172 kg ha-1) e P. glaucum (17.084 kg ha-1) (Tabela 1). Os
valores de FS das plantas de cobertura foram superiores aos obtidos por Pacheco et al.
(2013), Boer et al. (2007), Crusciol e Soratto (2007) e Torres, Pereira e Fabian (2008).
Estas diferenças podem estar relacionadas as condições climáticas, solo e espécies
utilizadas, já que ao longo do ciclo destas espécies houve boa precipitação (Tabela 1). A E.
coracana também se destacou na produção de FS (12.628 kg ha-1), com resultados
superiores ao alcançados por Boer et al., (2007) e Francisco (2002). Embora todas as
espécies avaliadas tenham apresentado significativas reduções na fitomassa seca aos 180
DAD, apenas nos tratamentos com E. coracana não mais se detectou FS na superfície do
solo.
Tabela 1. Fitomassa seca, taxa de cobertura do solo e coeficientes da equação de regressão PL = Poexp(-kt), e meia-vida (T½) da fitomassa seca remanescente das plantas de cobertura semeadas em pré-safra e avaliadas em nove épocas até 180 dias após a dessecação, safra 2014/2015 em Sinop, MT.
Época (DAD)(1)
C. spectabilis M. pruriens U. brizantha E. coracana P. glaucum
Fitomassa seca (kg ha-1)
0 9.524d 6.248e 24.172a 12.628c 17.084b 15(2) 6.992d 4.826d 18.640a 10.548c 14.140b 30 3.718c 3.534c 10.908a 9.738a 6.726b 45 3.500cd 3.436cd 8.642a 5.336bc 6.558ab 60 3.262bc 3.088bc 7.632a 3.872bc 5.820ab 90 2.948bc 2.988bc 5.822a 3.880abc 4.746ab 120 2.876abc 2.692bc 5.566a 3.116abc 3.838abc 150 2.290ab 2.540ab 4.416a 2.406ab 3.204ab 180 2.048ab 2.156ab 4.014a 0c 2.798a CV (%) 22,37
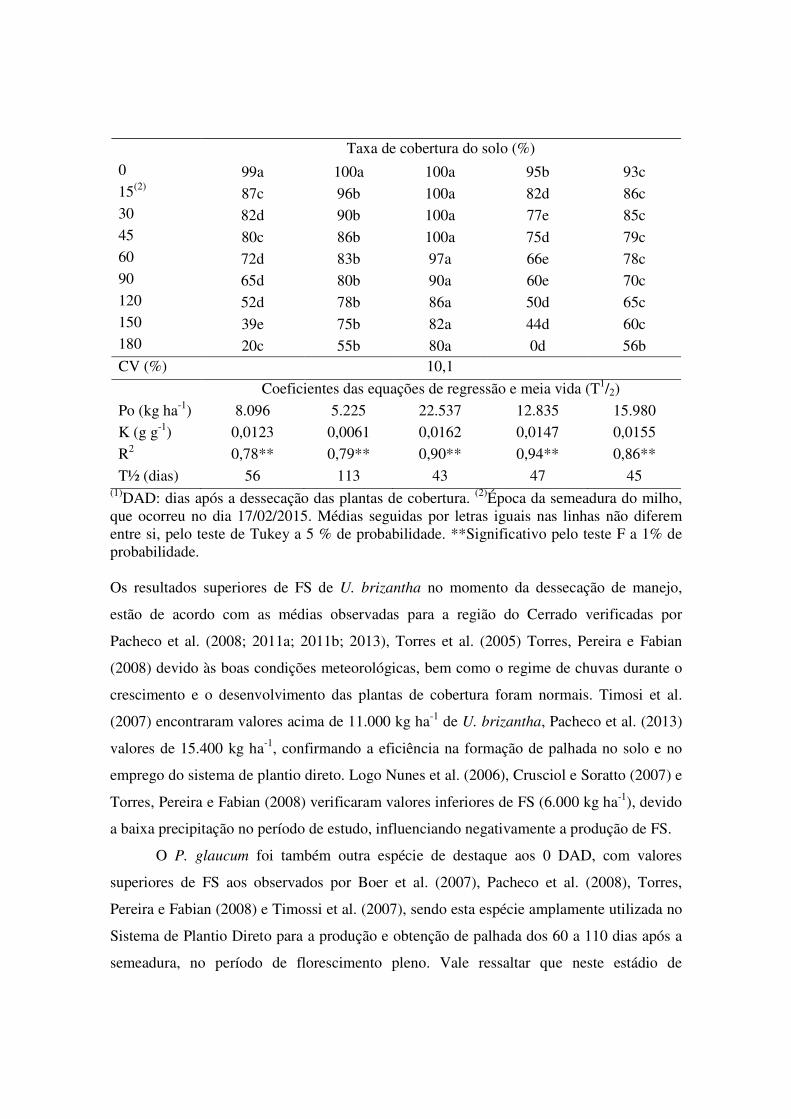
Taxa de cobertura do solo (%)
0 99a 100a 100a 95b 93c 15(2) 87c 96b 100a 82d 86c 30 82d 90b 100a 77e 85c 45 80c 86b 100a 75d 79c 60 72d 83b 97a 66e 78c 90 65d 80b 90a 60e 70c 120 52d 78b 86a 50d 65c 150 39e 75b 82a 44d 60c 180 20c 55b 80a 0d 56b CV (%) 10,1
Coeficientes das equações de regressão e meia vida (T1/2)
Po (kg ha-1) 8.096 5.225 22.537 12.835 15.980
K (g g-1) 0,0123 0,0061 0,0162 0,0147 0,0155
R2 0,78** 0,79** 0,90** 0,94** 0,86**
T½ (dias) 56 113 43 47 45 (1)DAD: dias após a dessecação das plantas de cobertura. (2)Época da semeadura do milho, que ocorreu no dia 17/02/2015. Médias seguidas por letras iguais nas linhas não diferem entre si, pelo teste de Tukey a 5 % de probabilidade. **Significativo pelo teste F a 1% de probabilidade. Os resultados superiores de FS de U. brizantha no momento da dessecação de manejo,
estão de acordo com as médias observadas para a região do Cerrado verificadas por
Pacheco et al. (2008; 2011a; 2011b; 2013), Torres et al. (2005) Torres, Pereira e Fabian
(2008) devido às boas condições meteorológicas, bem como o regime de chuvas durante o
crescimento e o desenvolvimento das plantas de cobertura foram normais. Timosi et al.
(2007) encontraram valores acima de 11.000 kg ha-1 de U. brizantha, Pacheco et al. (2013)
valores de 15.400 kg ha-1, confirmando a eficiência na formação de palhada no solo e no
emprego do sistema de plantio direto. Logo Nunes et al. (2006), Crusciol e Soratto (2007) e
Torres, Pereira e Fabian (2008) verificaram valores inferiores de FS (6.000 kg ha-1), devido
a baixa precipitação no período de estudo, influenciando negativamente a produção de FS.
O P. glaucum foi também outra espécie de destaque aos 0 DAD, com valores
superiores de FS aos observados por Boer et al. (2007), Pacheco et al. (2008), Torres,
Pereira e Fabian (2008) e Timossi et al. (2007), sendo esta espécie amplamente utilizada no
Sistema de Plantio Direto para a produção e obtenção de palhada dos 60 a 110 dias após a
semeadura, no período de florescimento pleno. Vale ressaltar que neste estádio de

desenvolvimento, a planta deve ser manejada quimicamente com o uso de herbicida a fim
de que rapidamente se inicie a decomposição e na sequência, o processo de liberação de
nutrientes (BOER et al., 2007; PACHECO et al., 2008).
De acordo com Petter et al. (2013) o milheto pode ser mais eficiente na
permanência dos resíduos no solo. No milheto cerca de 40% da fitomassa acumulada, se
localiza no caule, que constituem tecidos mais lignificados e com maior relação C/N
comparados às folhas, por isso favorece a menor taxa de decomposição, contribuindo em
uma maior duração desses resíduos no período de entressafra.
Quanto a porcentagem de cobertura no solo (Tabela 1), destacaram-se as espécies
U. brizantha, P. glaucum e M. pruriens, as quais apresentaram uma melhor cobertura do
solo em razão da elevada capacidade de produção de fitomassa com uma degradação lenta
em relação as outras espécies no final do período de avaliação. Timossi et al. (2007)
descreveram que a espécie U. brizantha com plantio no início da entressafra apresentou
maior potencial na produção de fitomassa seca, com cerca de 11.000 kg ha-1 e porcentagem
de cobertura do solo em torno de 97% no momento da dessecação de manejo para
semeadura das culturas anuais.
Já para E. coracana, a quantidade de fitomassa seca (kg ha-1) e a porcentagem de
taxa de cobertura no final dos 180 dias foram nulas, uma vez que esta é uma planta anual
de dia curto que produziu uma menor quantidade de fitomassa e uma maior taxa de
decomposição em comparação as outras espécies observadas (PACHECO et al., 2009).
Pode-se observar que no período entre o 150° e o 180° dia, E. coracana apresentou maior
taxa de decomposição de fitomassa seca, devido uma maior porcentagem de folhas na
fitomassa seca em proporção ao volume de caule na planta, o que acelerou o processo da
decomposição.
Avaliando a fração de nutrientes potencialmente liberados (Po) e a taxa de liberação
de nutrientes (K) na Tabela 1, verifica-se que U. brizantha foi a espécie de cobertura que
mais liberou nutrientes no solo em relação as demais espécies, devido a sua maior
quantidade de fitomassa seca remanescente e menor tempo de meia-vida (T½). Torres et al.
(2005) e Torres e Pereira (2008) também atestaram que a U. brizantha, foi a planta de
cobertura que apresentou o menor (T½) dentre as espécies estudadas. Com relação ao
tempo de meia-vida (T½), M. pruriens se destacou em relação a outras espécies, devido a

maior proporção de caule em relação as folhas e maior teor de lignina e celulose,
observado também por Petter et al. (2013) em outra espécie do gênero Mucuna, os quais
resultam em uma maior relação C/N, permanência no solo por um maior período, menor
taxa de decomposição e liberação de nutrientes da fitomassa seca (GIACOMINI et al.,
2003). Embora com maior número de folhas, maior altura de plantas e crescimento do
caule, o acúmulo de FS em M. pruriens foi significativamente menor (no ponto de rolagem)
comparado às demais espécies de plantas (Tabela 1). Resíduos com alta relação C/N se
decompõem mais lentamente no solo devido um maior tempo de imobilização até atingir na
mineralização, mas por permanecer mais tempo no solo, com altas temperaturas na região
Norte de Mato Grosso propiciam vantagens como proteção da microbiota na superfície do
solo contra incidência direta solar e desequilíbrio térmico. Porém a curto prazo, o que mais
interfere no rendimento da cultura sucessora é a ciclagem de nutrientes, em especial a
intensidade da liberação de nutrientes pela palhada proveniente da planta de cobertura,
sendo que neste contexto é recomendável os produtores utilizarem culturas com a U.
brizantha, pois com altas temperaturas e altos índices pluviométricos, há uma maior
decomposição e liberação dos nutrientes mais rapidamente no solo.
Houve diferença significativa no acúmulo de nutrientes remanescentes na fitomassa
das plantas de cobertura e na dinâmica de liberação desses nutrientes (Tabela 2). Os
nutrientes que proporcionaram maiores percentuais de acúmulo na matéria seca das
espécies de plantas de cobertura foram o nitrogênio (N) e o potássio (K), corroborando com
os resultados observados, por Torres et al. (2005), Torres, Pereira e Fabian (2008) e Boer et
al. (2007) para N na região do Cerrado. Valores de K superiores foram obtidos por Boer et
al. (2007); Carpim et al. (2008) e Torres, Pereira e Fabian (2008) encontraram valores
semelhantes. Mendonça et al. (2015) encontraram valores semelhantes de N e K na espécie
U. brizantha cv. Marandu, em que destacaram o potencial do uso de gramíneas como
palhada para o sistema de plantio direto em regiões de Cerrado.
Tabela 2. Macronutrientes remanescentes nas fitomassas das plantas de cobertura, semeadas em pré-safra e avaliadas em nove épocas após a dessecação para semeadura do milho em plantio direto em Sinop, MT, safra 2014/2015.
Época (DAD)(1)
C. spectabilis M. pruriens U.
brizantha E. coracana P. glaucum
Nitrogênio (kg ha-1)
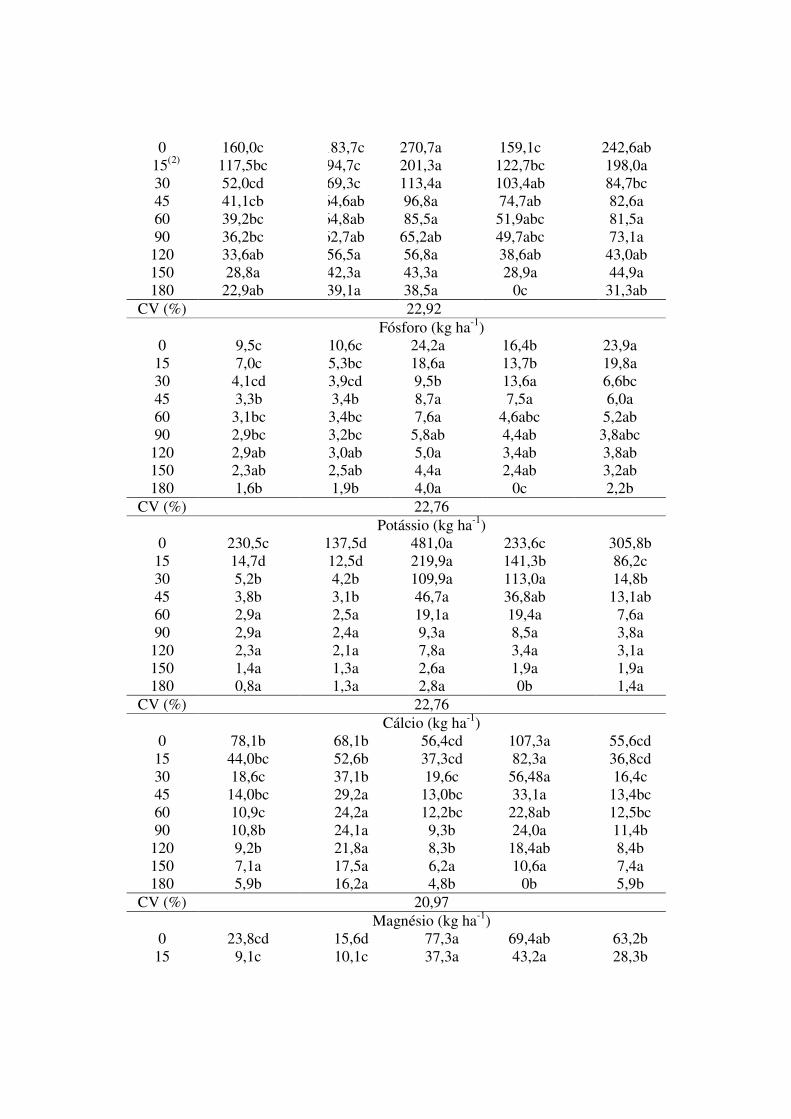
0 160,0c 183,7c 270,7a 159,1c 242,6ab 15(2) 117,5bc 94,7c 201,3a 122,7bc 198,0a
30 52,0cd 69,3c 113,4a 103,4ab 84,7bc 45 41,1cb 64,6ab 96,8a 74,7ab 82,6a 60 39,2bc 64,8ab 85,5a 51,9abc 81,5a 90 36,2bc 62,7ab 65,2ab 49,7abc 73,1a
120 33,6ab 56,5a 56,8a 38,6ab 43,0ab 150 28,8a 42,3a 43,3a 28,9a 44,9a 180 22,9ab 39,1a 38,5a 0c 31,3ab
CV (%) 22,92 Fósforo (kg ha-1) 0 9,5c 10,6c 24,2a 16,4b 23,9a 15 7,0c 5,3bc 18,6a 13,7b 19,8a 30 4,1cd 3,9cd 9,5b 13,6a 6,6bc 45 3,3b 3,4b 8,7a 7,5a 6,0a 60 3,1bc 3,4bc 7,6a 4,6abc 5,2ab 90 2,9bc 3,2bc 5,8ab 4,4ab 3,8abc
120 2,9ab 3,0ab 5,0a 3,4ab 3,8ab 150 2,3ab 2,5ab 4,4a 2,4ab 3,2ab 180 1,6b 1,9b 4,0a 0c 2,2b
CV (%) 22,76 Potássio (kg ha-1) 0 230,5c 137,5d 481,0a 233,6c 305,8b 15 14,7d 12,5d 219,9a 141,3b 86,2c 30 5,2b 4,2b 109,9a 113,0a 14,8b 45 3,8b 3,1b 46,7a 36,8ab 13,1ab 60 2,9a 2,5a 19,1a 19,4a 7,6a 90 2,9a 2,4a 9,3a 8,5a 3,8a
120 2,3a 2,1a 7,8a 3,4a 3,1a 150 1,4a 1,3a 2,6a 1,9a 1,9a 180 0,8a 1,3a 2,8a 0b 1,4a
CV (%) 22,76 Cálcio (kg ha-1) 0 78,1b 68,1b 56,4cd 107,3a 55,6cd 15 44,0bc 52,6b 37,3cd 82,3a 36,8cd 30 18,6c 37,1b 19,6c 56,48a 16,4c 45 14,0bc 29,2a 13,0bc 33,1a 13,4bc 60 10,9c 24,2a 12,2bc 22,8ab 12,5bc 90 10,8b 24,1a 9,3b 24,0a 11,4b
120 9,2b 21,8a 8,3b 18,4ab 8,4b 150 7,1a 17,5a 6,2a 10,6a 7,4a 180 5,9b 16,2a 4,8b 0b 5,9b
CV (%) 20,97 Magnésio (kg ha-1) 0 23,8cd 15,6d 77,3a 69,4ab 63,2b 15 9,1c 10,1c 37,3a 43,2a 28,3b

Médias seguidas por letras iguais nas linhas, não diferem entre si pelo teste Tukey, a 5% de probabilidade. (1)DAD: Dias após a dessecação das plantas de cobertura.
(2)Semeadura do milho em sistema de plantio direto, que ocorreu no dia 17/02/2015.
As espécies U. brizantha e P. glaucum acumularam as maiores quantidades (kg ha-
1) de N, P, K e Mg do início ao fim da avaliação. Tal fato se deve a elevada capacidade de
produção de fitomassa, visto que os teores desses nutrientes nas folhas diferiram entre as
espécies. Esses resultados indicam a possibilidade de diminuição da adubação mineral nas
culturas anuais sucessoras, assemelhando com o reportado por Pacheco et al. (2011b).
Todavia, atenção especial deve ser dada ao N, é um elemento muito móvel no sistema
segundo Mendonça et al. (2015), uma vez que há expressivo potencial de perdas desse
nutriente por volatilização, perlocação, escorrimento laminar e imobilização pela
microbiota do solo durante a decomposição da matéria seca, o que pode reduzir a
disponibilidade imediata para cultivos sucessores (AITA; GIACOMINI, 2003).
Os elevados valores de N acumulados pelas fitomassas de P. glaucum e U.
brizantha podem estar associados à capacidade e exploração do sistema radicular e a
eficiência na absorção desse nutriente. Adicionalmente, esses elevados acúmulos de N
podem também estar associado à atividade simbiótoca de bactérias fixadoras de nitrogênio,
com predominância de Herbaspirillume da bactéria Azospirillum. Conforme Reis et al.
(2001), esse processo pode contribuir com mais de 40 e 20% do nitrogênio acumulado nas
fitomassas de P. glaucum e das espécies de U. brizantha, respectivamente.
Já para os valores elevados de K acumulados pelas fitomassas de U. brizantha e P.
glaucum (Tabela 2), foram encontrados valores superiores da braquiária, aos encontrados
por Crusciol e Soratto (2007) e Pacheco et al. (2013). No milheto foram encontrados
valores inferiores (416 kg ha-1) no ponto de rolagem (0 DAD) por Boer et al. (2007), e
superiores aos encontrados por Teixeira et al. (2005) e Oliveira et al. (2002), em
milheto(comum), cujos obtiveram, respectivamente, 50,9 e 20,8 kg ha-1 de K, com
30 3,0e 5,3de 19,6bc 30,2a 12,1cd 45 2,8c 4,5bc 16,4a 14,4a 11,8ab 60 2,6b 3,7b 11,4a 10,9ab 9,9ab 90 2,3c 3,6bc 9,3a 10,0a 5,7b
120 1,7c 3,0b 6,7a 7,2a 5,0ab 150 1,6b 2,8ab 4,8a 4,1a 3,8ab 180 1,2b 2,1b 4,0a 0c 2,5b
CV (%) 30,37

produção de 2.907 e 14.180 kg ha-1 de FS. O acúmulo de K e até mesmo o de N no milheto,
é devido a cultivar que foi utilizada ser melhorada geneticamente (milheto ADR 300) e ter
boa capacidade para produção de FS, ciclagem e acúmulo de nutrientes. Logo os demais
autores, com exceção de Boer et al. (2007), as quais utilizaram o milheto ADR 500,
realizaram estudos com o milheto(comum), com uma capacidade genética menor. Carpim
et al. (2008) também obervaram grande quantidade de K acumulados na FS do milheto,
pois o fornecimento para a cultura sucessora tem relação com o cultivar utilizado, estádio
fenológico e as condições climáticas.
O aumento inicial no acúmulo de P das espécies U. brizantha (24,2 kg ha-1) e P.
glaucum (23,9 kg ha-1) (Tabela 2), é também explicado pela elevada capacidade de
produção de fitomassa, corroborando com os valores encontrados por Torres et al. (2005),
visto que os teores de P nas folhas não diferiram entre as espécies (Tabela 2). Outro aspecto
que pode ter contribuído para o aumento na absorção de P foi a frequência de índices
pluviométricos regulares, visto que o P (ânion PO4-) se movimenta principalmente por
difusão e esta é dependente de umidade no solo.
Para Cálcio (Ca) (Tabela 2), no manejo de dessecação E. coracana foi a espécie de
maior acúmulo desse nutriente (107,3 kg ha-1), com valores maiores do que os 78,32 kg ha-1
observados por Boer et al. (2007). Entretanto, aos 180 DAD, a espécie de maior destaque
foi a M. pruriens. Os elevados valores de Ca acumulados pelas fitomassas de E. coracana,
C. spectabilis, M. pruriens podem ter sido influenciados pelas estruturas das plantas, bem
como altura e enraizamento, uma vez que o cálcio tem grande importância no
desenvolvimento das raízes, na formação da estrutura da planta e no metabolismo de
nitrogênio (Werner; Haag, 1994).
A E. coracana também é conhecida pelo seu elevado potencial de
desenvolvimento do sistema radicular, sendo considerada uma planta descompactadora do
solo e, esse aspecto, pode aumentar a área de contato íon-raiz e consequentemente a
absorção de Ca. Comportamento semelhante foi verificado para Magnésio (Mg), em que a
E. coracana (69,4 kg ha-1 de Mg) apresentou valores maiores do que os observados por
Boer et al. (2007), 50,94 kg ha-1 de Mg. Entretanto, aos 180 DAD, U. brizantha apresentou
o maior valor dentre as espécies ao passo que E. coracana não mais apresentou nutrientes
remanescente, visto a sua total decomposição da palhada.

De maneira geral verifica-se que as espécies U. brizantha e P. glaucum
apresentam elevado potencial de ciclar os macronutrintes N, P e K. Estes resultados
confirmam os alcançados por Torres, Pereira e Fabian (2008) e Torres e Pereira (2008) que
indicaram respectivamente, um maior teor de ciclagem de N, P e K pelas espécies de
cobertura (braquiária e milheto). Diante dos altos custos dos ferlizantes, é provável que
com o contínua utilização de plantas de cobertura na produção de soja e milho, melhore a
eficência na utilização dos nutrientes disponíveis no solo, com readequação nos aspectos
relativos ao custo de produção. Somado a esse fato a espécie E. coracana apresenta elevado
potencial para ciclar Ca e Mg, sendo que tais nutrientes possuem maior simplicidade de
serem manejados, haja vista que são fornecidos via calagem, prática corretiva de acidez
imprescindível e regular realizada nas lavouras do estado de Mato Grosso.
As espécies U. brizantha e P. glaucum apresentaram os maiores acúmulos de
cobre (Cu) (120,86 g ha-1), manganês (Mn) (797,68 g ha-1) e zinco (Zn) (580,85 g ha-1), do
início ao fim da avaliação (Tabela 3). Assim como para os macronutrientes, sendo esses
três micronutrientes associados a processos fotossintéticos (mais intensos e diferenciados
em plantas C4, como braquiária e milheto) e cofatores enzimáticos em diversos
metabolismos. Já as menores quantidades acumuladas na fitomassa seca foram verificados
para M. pruriens e C. spectabilis. Silva et al. (2008) e Cavalcante et al. (2012) observaram
teores de micronutrientes semelhantes ao do presente trabalho. Relataram que a diferença
das espécies em relação a concentração dos nutrientes, sofre influência da diferença da
fertilidade do solo, e com isso a eficiência da ciclagem de nutrientes das espécies de
cobertura dependem de um nível de fertilidade antecedente. Para E. coracana, a quantidade
de micronutrientes remanescente na fitomassa no final dos 180 DAD foi nula, devido total
decomposição da fitomassa.
Tabela 3. Micronutrientes remanescentes nas fitomassas das plantas de cobertura, semeadas em pré-safra e avaliadas em nove épocas após a dessecação para semeadura do milho em plantio direto em Sinop, MT, safra 2014/2015.
Época (DAD)(1)
C. spectabilis M. pruriens U.
brizantha E. coracana P. glaucum
Cobre (g ha-1) 0 76,19b 49,98cd 120,86a 63,1 bc 119,59a
15(2) 42,19c 28,95cd 111,84ab 41,95c 113,12a 30 29,74b 28,27b 54,54ª 38,95ab 40,36ab
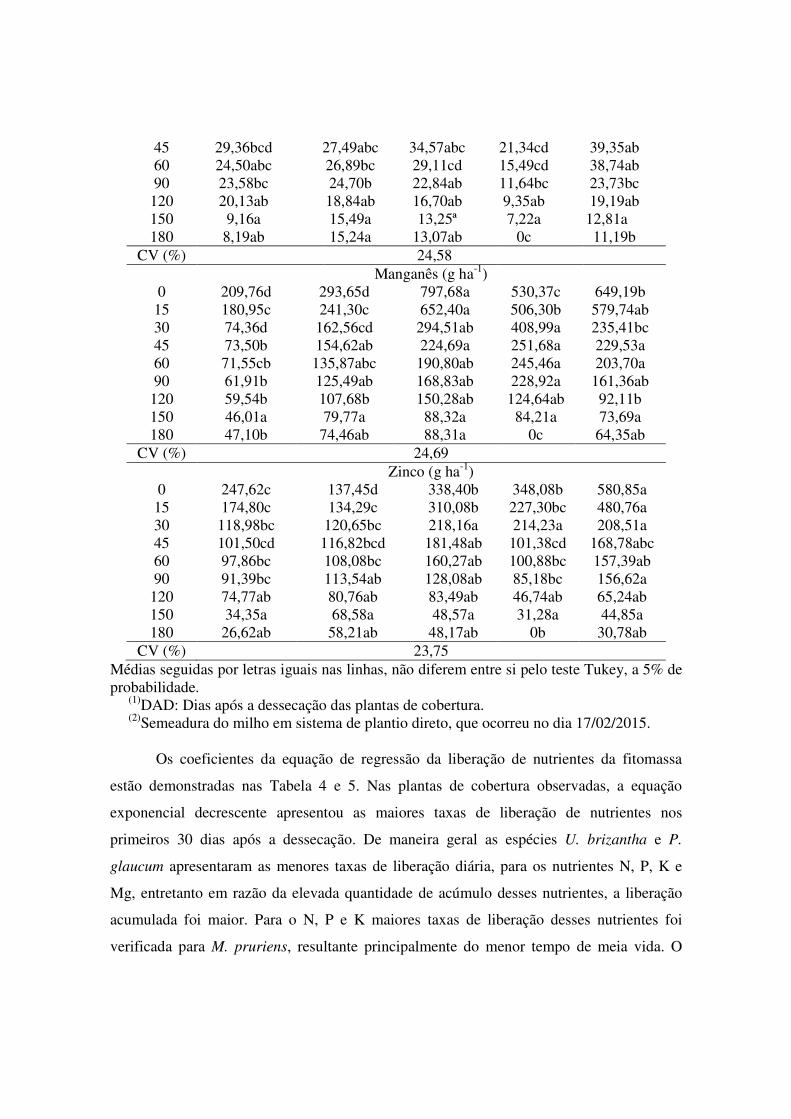
Médias seguidas por letras iguais nas linhas, não diferem entre si pelo teste Tukey, a 5% de probabilidade.
(1)DAD: Dias após a dessecação das plantas de cobertura. (2)Semeadura do milho em sistema de plantio direto, que ocorreu no dia 17/02/2015.
Os coeficientes da equação de regressão da liberação de nutrientes da fitomassa
estão demonstradas nas Tabela 4 e 5. Nas plantas de cobertura observadas, a equação
exponencial decrescente apresentou as maiores taxas de liberação de nutrientes nos
primeiros 30 dias após a dessecação. De maneira geral as espécies U. brizantha e P.
glaucum apresentaram as menores taxas de liberação diária, para os nutrientes N, P, K e
Mg, entretanto em razão da elevada quantidade de acúmulo desses nutrientes, a liberação
acumulada foi maior. Para o N, P e K maiores taxas de liberação desses nutrientes foi
verificada para M. pruriens, resultante principalmente do menor tempo de meia vida. O
45 29,36bcd 27,49abc 34,57abc 21,34cd 39,35ab 60 24,50abc 26,89bc 29,11cd 15,49cd 38,74ab 90 23,58bc 24,70b 22,84ab 11,64bc 23,73bc
120 20,13ab 18,84ab 16,70ab 9,35ab 19,19ab 150 9,16a 15,49a 13,25ª 7,22a 12,81a 180 8,19ab 15,24a 13,07ab 0c 11,19b
CV (%) 24,58 Manganês (g ha-1) 0 209,76d 293,65d 797,68a 530,37c 649,19b 15 180,95c 241,30c 652,40a 506,30b 579,74ab 30 74,36d 162,56cd 294,51ab 408,99a 235,41bc 45 73,50b 154,62ab 224,69a 251,68a 229,53a 60 71,55cb 135,87abc 190,80ab 245,46a 203,70a 90 61,91b 125,49ab 168,83ab 228,92a 161,36ab
120 59,54b 107,68b 150,28ab 124,64ab 92,11b 150 46,01a 79,77a 88,32a 84,21a 73,69a 180 47,10b 74,46ab 88,31a 0c 64,35ab
CV (%) 24,69 Zinco (g ha-1) 0 247,62c 137,45d 338,40b 348,08b 580,85a 15 174,80c 134,29c 310,08b 227,30bc 480,76a 30 118,98bc 120,65bc 218,16a 214,23a 208,51a 45 101,50cd 116,82bcd 181,48ab 101,38cd 168,78abc 60 97,86bc 108,08bc 160,27ab 100,88bc 157,39ab 90 91,39bc 113,54ab 128,08ab 85,18bc 156,62a
120 74,77ab 80,76ab 83,49ab 46,74ab 65,24ab 150 34,35a 68,58a 48,57a 31,28a 44,85a 180 26,62ab 58,21ab 48,17ab 0b 30,78ab
CV (%) 23,75
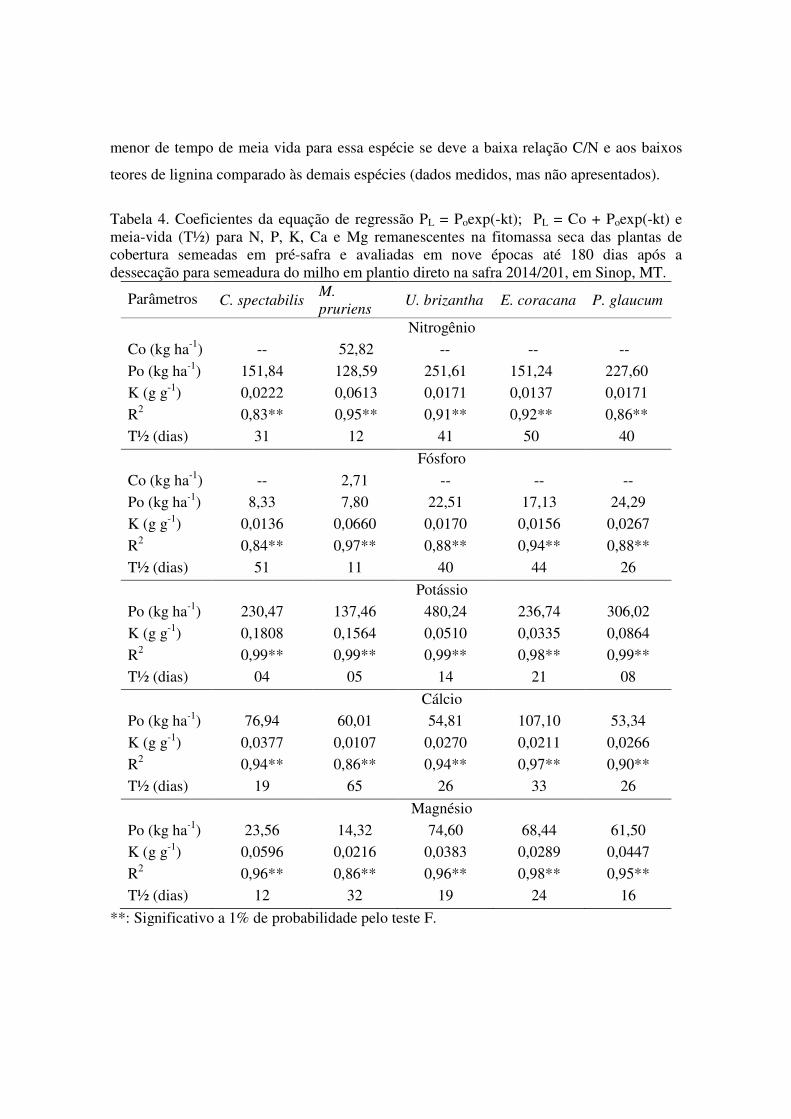
menor de tempo de meia vida para essa espécie se deve a baixa relação C/N e aos baixos
teores de lignina comparado às demais espécies (dados medidos, mas não apresentados).
Tabela 4. Coeficientes da equação de regressão PL = Poexp(-kt); PL = Co + Poexp(-kt) e meia-vida (T½) para N, P, K, Ca e Mg remanescentes na fitomassa seca das plantas de cobertura semeadas em pré-safra e avaliadas em nove épocas até 180 dias após a dessecação para semeadura do milho em plantio direto na safra 2014/201, em Sinop, MT.
Parâmetros C. spectabilis M.
pruriens U. brizantha E. coracana P. glaucum
Nitrogênio
Co (kg ha-1) -- 52,82 -- -- --
Po (kg ha-1) 151,84 128,59 251,61 151,24 227,60
K (g g-1) 0,0222 0,0613 0,0171 0,0137 0,0171
R2 0,83** 0,95** 0,91** 0,92** 0,86**
T½ (dias) 31 12 41 50 40
Fósforo
Co (kg ha-1) -- 2,71 -- -- --
Po (kg ha-1) 8,33 7,80 22,51 17,13 24,29
K (g g-1) 0,0136 0,0660 0,0170 0,0156 0,0267
R2 0,84** 0,97** 0,88** 0,94** 0,88**
T½ (dias) 51 11 40 44 26
Potássio
Po (kg ha-1) 230,47 137,46 480,24 236,74 306,02
K (g g-1) 0,1808 0,1564 0,0510 0,0335 0,0864
R2 0,99** 0,99** 0,99** 0,98** 0,99**
T½ (dias) 04 05 14 21 08
Cálcio
Po (kg ha-1) 76,94 60,01 54,81 107,10 53,34
K (g g-1) 0,0377 0,0107 0,0270 0,0211 0,0266
R2 0,94** 0,86** 0,94** 0,97** 0,90**
T½ (dias) 19 65 26 33 26
Magnésio
Po (kg ha-1) 23,56 14,32 74,60 68,44 61,50
K (g g-1) 0,0596 0,0216 0,0383 0,0289 0,0447
R2 0,96** 0,86** 0,96** 0,98** 0,95**
T½ (dias) 12 32 19 24 16
**: Significativo a 1% de probabilidade pelo teste F.

Pacheco et al. (2013, 2011b) na região do Cerrado de Goiás, verificaram-se tempos
de meia-vida (T½) semelhantes para os nutrientes N, P, K Ca e Mg para essas mesmas
espécies. Torres, Pereira e Fabian (2008) observaram em braquiária, menores (T½) para os
nutrientes Ca e Mg, e maiores (T½) para N e P, Gama-Rodrigues et al. (2007) encontraram
em braquiária (T½) superiores para N, P, Ca e Mg e em milheto, Torres, Pereira e Fabian
(2008) encontraram maiores (T½) comparando-os ao presente estudo. Os altos valores de
(T½) observados nestes trabalhos citados, podem ser justificados devido a menor
precipitação e temperatura posteriormente a dessecação (manejo) das plantas de cobertura.
Os menores valores de (T½) encontrados no presente trabalho, comparados aos
obtidos por Pacheco et al. (2013,2011b), são explicados pelas condições de altas
temperaturas e abundantes precipitações do Bioma Amazônia, o que favoreceram a
decomposição dos resíduos das plantas de cobertura, observados também por Espindola et
al. (2006), Carvalho et al. (2008) e Acosta et al. (2013), além do uso da irrigação nos
períodos de veranicos, o que pode ter proporcionado uma rápida e elevada taxa de
decomposição da fitomassa.
Logo, a espécie E. cocarana apresentou maior taxa de liberação para o Ca,
semelhante ao observado por Boer et al. (2007), devido a maior porcentagem de folhas na
fitomassa seca em comparação a proporção de caule da planta. Entretanto, a M. pruriens foi
superior no (T½) deste nutriente, o que pode ter sido influenciado pela estrutura
morfofisiológica da planta, como a formação de ramos que possuem maior concentração e
altos teores de Ca, sendo estruturas mais lignificadas. Segundo Carvalho et al. (2011), altas
concentrações de lignina inibem a decomposição da fitomassa, sendo favorável a cobertura
do solo. Logo, concentrações menores de lignina aceleram a decomposição da fitomassa,
consequentemente resultam em uma ciclagem mais efetiva dos nutrientes.
Para a fração de nutrientes potencialmente liberados (P0) e a taxa de liberação de
nutrientes (K) (Tabela 5), a U. brizantha e a P. glaucum foram as espécies de cobertura que
mais liberaram Cu, Mn e Zn no solo e as leguminosas C. spectabilis e M. pruriens
apresentaram os menores valores entre as espécies justificado pela elevada quantidade de
fitomassa seca remanescente, e pelo metabolismo C3 dessas duas , ou seja, metabolismo
fotossintético menos intenso.

Tabela 5. Coeficientes da equação de regressão PL = Poexp(-kt), e meia-vida (T½) para Cu, Mn e Zn remanescentes na fitomassa seca das plantas de cobertura semeadas em pré-safra e avaliadas em nove épocas até 180 dias após a dessecação para semeadura do milho em plantio direto na safra 2014/201, em Sinop, MT.
Parâmetros C. spectabilis M.
pruriens
U.
brizantha E. coracana P. glaucum
Cobre
Po (kg ha-1) 63,59 39,94 126,47 61,41 122,75
K (g g-1) 0,0148 0,0064 0,0221 0,0199 0,0208
R2 0,84** 0,78* 0,93** 0,97** 0,88**
T½ (dias) 47 108 31 35 33
Manganês
Po (kg ha-1) 190,63 262,69 792,82 559,80 657,23
K (g g-1) 0,0140 0,0088 0,0224 0,0129 0,0197
R2 0,77* 0,90** 0,91** 0,95** 0,90**
T½ (dias) 50 79 31 54 35
Zinco
Po (kg ha-1) 219,30 141,63 341,78 334,42 579,70
K (g g-1) 0,0127 0,0044 0,0122 0,0193 0,0222
R2 0,91** 0,93** 0,98** 0,96** 0,92**
T½ (dias) 55 157 57 36 31
* e **: Significativo a 1 e 5% de probabilidade, respectivamente pelo teste F.
Levando em consideração os tempos de meia-vida obtidos, ou seja, o tempo
necessário para que, cerca de 50 % dos nutrientes remanescentes na fitomassa seca das
plantas de cobertura, fossem liberados, a U. brizantha foi a espécie que mais liberou
rapidamente os nutrientes Cu e Mn, com 31 dias, seguido do P. glaucum com 31 dias do
nutriente Zn.
Já a M. pruriens necessitou um maior tempo para a liberação entre as espécies, com
valores de 157 dias (Zn), 108 dias (Cu) e 79 dias (Mn), devido a maior proporção de caule
(ramos) em relação as folhas, maior teor de lignina e celulose no caule (ramos) do que nas
folhas. Como consequência, uma maior relação C/N, com maior período de permanência no
solo, menor taxa de decomposição e liberação de nutrientes da fitomassa.
3.2. Liberação acumulada de macronutrientes por plantas de cobertura

A liberação acumulada de N, P, K, Ca e Mg (Tabela 6) apresentou comportamento
diferenciado nas cinco plantas de cobertura em estudo. As maiores taxas de liberação
acumulada de nutrientes foram observadas na U. brizantha, seguida da P. glaucum para N,
P, K e Mg, corroborando os resultados observados por Pacheco et al. (2011a). Já para Ca, a
E. coracana foi a espécie com maior taxa de liberação acumulada de nutrientes.
O processo de degradação de matéria seca e a velocidade de liberação de N, P e Mg
se demonstraram semelhantes (Apêndice 1A), ocorrendo uma elevada taxa de
decomposição e liberado do N, P e Mg entre o intervalo de 0 a 45 DAD, e em seguida por
uma taxa mais lenta entre 60 a 180 DAD (modelo exponencial para decomposição
biológica). Já o processo de degradação de matéria seca e a velocidade de liberação de Ca,
também demonstraram semelhança, ocorrendo uma elevada taxa de decomposição e
liberado do Ca entre o intervalo de 0 a 60 DAD, e em seguida por uma taxa mais lenta entre
70 a 180 DAD. Para o K, houve semelhança no processo de degradação de matéria seca e a
velocidade de liberação de K para C. spectabilis e P. glaucum, ocorrendo uma elevada taxa
de decomposição e liberado do K entre o intervalo de 0 a 20 DAD, e para as demais plantas
de 0 a 45 DAD, fato explicado pela elevada mobilidade do K tanto na planta como no solo,
sendo que essas espécies (crotalária e milheto) são portadoras de relação C/N mais reduzida
que as demais, sistema radicular mais profundo (menos superficial) e com maior proporção
de folha do que caule, facilitando assim a liberação de K inicial, com uma fase mais intensa
que as demais, além de possuírem, por questões morfológicas (maior porte e folhas mais
estreitas), maior susceptibilidade a fatores aceleradores da decomposição, como umidade e
temperatura. Em seguida por uma taxa mais lenta entre 30 a 180 DAD para C. spectabilis e
P. glaucum (liberaram de uma vez e estabilizaram a partir do 30 dias) e de 60 a 180 DAD
para as demais plantas, as quais começaram a liberar o K aos poucos e mais controlado.
Entretanto, liberar o K mais rapidamente que as demais plantas, não necessariamente é algo
totalmente desejável, pois a braquiária liberou K em um intervalo maior de dias, em
comparação a crotária e o milheto, e obteve um maior acréscimo na produtividade de
milho.
Tabela 6. Equações de liberação acumulada para os nutrientes N, P, K, Ca e Mg das plantas de cobertura em Sinop, MT, safra 2014/2015. Planta de Equação de liberação R2 Liberação aos 70(1) e 80(1) %
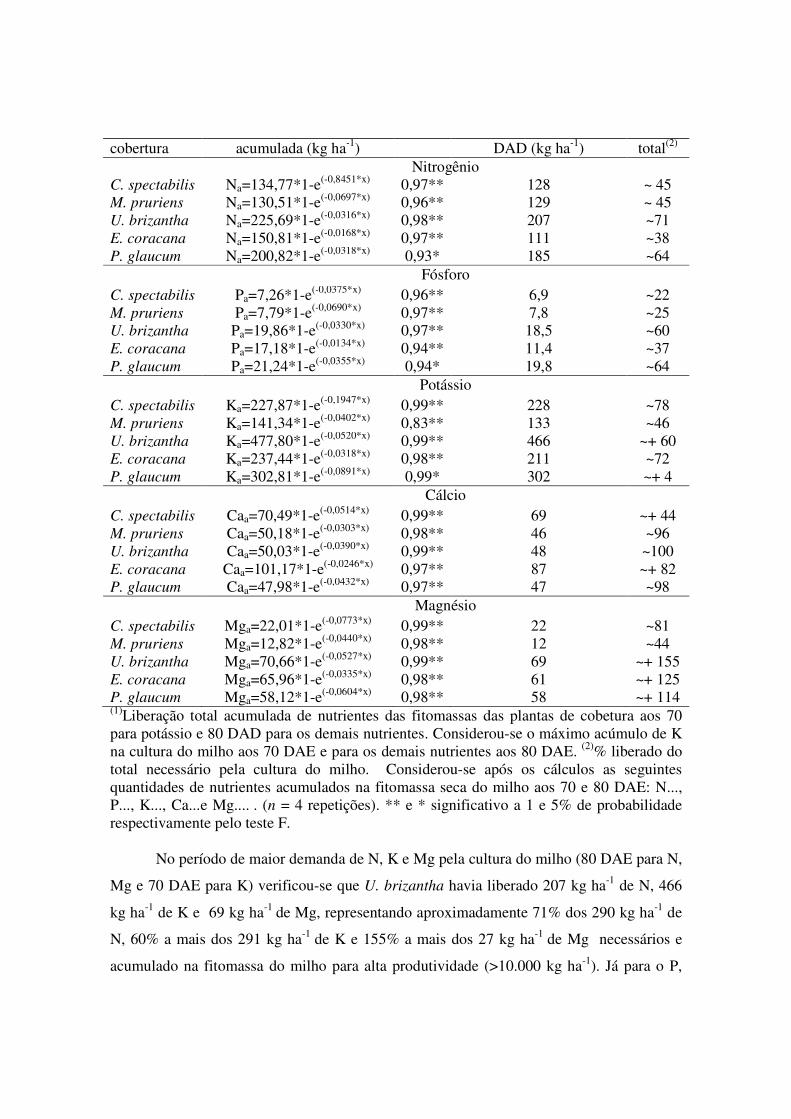
cobertura acumulada (kg ha-1) DAD (kg ha-1) total(2) Nitrogênio C. spectabilis Na=134,77*1-e(-0,8451*x) 0,97** 128 ~ 45 M. pruriens Na=130,51*1-e(-0,0697*x) 0,96** 129 ~ 45 U. brizantha Na=225,69*1-e(-0,0316*x) 0,98** 207 ~71 E. coracana Na=150,81*1-e(-0,0168*x) 0,97** 111 ~38 P. glaucum Na=200,82*1-e(-0,0318*x) 0,93* 185 ~64 Fósforo C. spectabilis Pa=7,26*1-e(-0,0375*x) 0,96** 6,9 ~22 M. pruriens Pa=7,79*1-e(-0,0690*x) 0,97** 7,8 ~25 U. brizantha Pa=19,86*1-e(-0,0330*x) 0,97** 18,5 ~60 E. coracana Pa=17,18*1-e(-0,0134*x) 0,94** 11,4 ~37 P. glaucum Pa=21,24*1-e(-0,0355*x) 0,94* 19,8 ~64 Potássio C. spectabilis Ka=227,87*1-e(-0,1947*x) 0,99** 228 ~78 M. pruriens Ka=141,34*1-e(-0,0402*x) 0,83** 133 ~46 U. brizantha Ka=477,80*1-e(-0,0520*x) 0,99** 466 ~+ 60 E. coracana Ka=237,44*1-e(-0,0318*x) 0,98** 211 ~72 P. glaucum Ka=302,81*1-e(-0,0891*x) 0,99* 302 ~+ 4 Cálcio C. spectabilis Caa=70,49*1-e(-0,0514*x) 0,99** 69 ~+ 44 M. pruriens Caa=50,18*1-e(-0,0303*x) 0,98** 46 ~96 U. brizantha Caa=50,03*1-e(-0,0390*x) 0,99** 48 ~100 E. coracana Caa=101,17*1-e(-0,0246*x) 0,97** 87 ~+ 82 P. glaucum Caa=47,98*1-e(-0,0432*x) 0,97** 47 ~98 Magnésio C. spectabilis Mga=22,01*1-e(-0,0773*x) 0,99** 22 ~81 M. pruriens Mga=12,82*1-e(-0,0440*x) 0,98** 12 ~44 U. brizantha Mga=70,66*1-e(-0,0527*x) 0,99** 69 ~+ 155 E. coracana Mga=65,96*1-e(-0,0335*x) 0,98** 61 ~+ 125 P. glaucum Mga=58,12*1-e(-0,0604*x) 0,98** 58 ~+ 114 (1)Liberação total acumulada de nutrientes das fitomassas das plantas de cobetura aos 70 para potássio e 80 DAD para os demais nutrientes. Considerou-se o máximo acúmulo de K na cultura do milho aos 70 DAE e para os demais nutrientes aos 80 DAE. (2)% liberado do total necessário pela cultura do milho. Considerou-se após os cálculos as seguintes quantidades de nutrientes acumulados na fitomassa seca do milho aos 70 e 80 DAE: N..., P..., K..., Ca...e Mg.... . (n = 4 repetições). ** e * significativo a 1 e 5% de probabilidade respectivamente pelo teste F.
No período de maior demanda de N, K e Mg pela cultura do milho (80 DAE para N,
Mg e 70 DAE para K) verificou-se que U. brizantha havia liberado 207 kg ha-1 de N, 466
kg ha-1 de K e 69 kg ha-1 de Mg, representando aproximadamente 71% dos 290 kg ha-1 de
N, 60% a mais dos 291 kg ha-1 de K e 155% a mais dos 27 kg ha-1 de Mg necessários e
acumulado na fitomassa do milho para alta produtividade (>10.000 kg ha-1). Já para o P,

destaque para o período de maior demanda de P pela cultura do milho (80 DAE). A qual
verificou-se que P. glaucum havia liberado 19,8 kg ha-1 de P, representando
aproximadamente 64% dos 31 kg ha-1 de P necessários e acumulados na fitomassa do milho
para alta produtividade (>10.000 kg ha-1).
Para a U. brizantha, há ênfase na liberação total de N, K e Mg durante todo o ciclo
da cultura do milho foi de, respectivamente, 225,69; 477,8 e 70,66 kg ha-1. O rápido
processo de decomposição e liberação de N da fitomassa está relacionado teores reduzidos
de celulose e lignina na biomassa das espécies Urochloa (CARVALHO et al., 2011). Tal
comportamento para K durante todo o ciclo da cultura do milho, é explicado pela produção
de fitomassa inicial U. brizantha (24.172 kg ha-1), devido a U. brizantha apresentou teores
mais elevados de K na biomassa vegetal, demonstrando-se mais eficiente no acúmulo e
liberação de K dentre as plantas estudadas. Na liberação total de Mg durante todo o ciclo da
cultura do milho encontrou-se valor semelhante ao observado por Pacheco et al. (2011a).
Tal fato confirma-se, uma vez que a U. brizantha apresenta uma maior fração de nutrientes
potencialmente liberados (P0) 74,60 kg ha-1.
Agora, já na liberação total de P durante todo o ciclo da cultura do milho foi de 21
kg ha-1. Este aumento no acúmulo de liberação de P pela P. glaucum é explicado pela
ocorrência de índices pluviométricos regulares durante o período estudado, o que auxiliou
no crescimento das plantas e absorção desse nutriente, atestado pela maior produção de
fitomassa dessas espécies.
C. spectabilis e M. pruriens apresentaram semelhança entre as espécies na curva
de resposta, com destaque para N e P. Com liberação de ~129 kg ha-1 de N aos 80 DAD, o
que equivale a 45% do necessário e acumulado na fitomassa do milho para alta
produtividade. Comparando a produção de fitomassa inicial (Tabela 1), pode-se concluir
que a M. pruriens apresentou teores mais elevados de N na biomassa vegetal,
demonstrando-se mais eficiente no acúmulo e liberação de N quando comparado à
quantidade de fitomassa produzida pela C. spectabilis. Já para P, M. pruriens e C.
spectabilis, apresentaram semelhança na curva de resposta, com liberação de ~7 kg ha-1 de
P aos 80 DAD, o que equivale a 25% dos 31 kg ha-1 de P necessários e acumulados na
fitomassa do milho para alta produtividade. Durante todo o ciclo do milho essas espécies
liberaram acumuladamente 7,79 kg ha-1 e 7,26 kg ha-1 de P respectivamente para o sistema.

Comparando-se a produção de fitomassa inicial, M. pruriens (6.248 kg ha-1) e C. spectabilis
(9.524 kg ha-1 ), pode-se concluir que a M. pruriens apresentou teores mais elevados de P
na biomassa vegetal, demonstrando-se mais eficiente no acúmulo e liberação de P quando
comparada à quantidade de fitomassa produzida pela C. spectabilis.
Para Mg, C. spectabilis e M. pruriens apresentaram respectivamente, liberação de
69 e 46 kg ha-1 de Mg aos 80 DAD, o que equivale a 44% a mais do necessário e 96% do
necessário e durante todo o ciclo do milho essas espécies liberaram acumuladamente 70,49
e 50,18 kg ha-1 de Mg respectivamente para o sistema. Comparando-se a produção de
fitomassa inicial, C. spectabilis (9.524 kg ha-1 ) e M. pruriens (6.248 kg ha-1), pode-se
concluir que a M. pruriens apresentou teores mais elevados de Mg na biomassa vegetal,
demonstrando-se mais eficiente no acúmulo e liberação de Mg quando comparados à
quantidade de fitomassa produzida pela C. spectabilis.
Na liberação acumulada de Ca, destaque para o período de maior demanda de Ca
pela cultura do milho (80 DAE), na qual verificou-se que E. coracana havia liberado 87 kg
ha-1 de Ca, representando aproximadamente 82% a mais dos 48 kg ha-1 de Ca necessários e
acumulados na fitomassa do milho para alta produtividade. A liberação total de Ca durante
todo o ciclo da cultura do milho foi de 101,17 kg ha-1. A espécie E. cocarana apresentou
maior taxa de liberação para o Ca, devido à maior porcentagem de folhas na fitomassa seca
em proporção ao caule da planta, logo no final dos 180 DAD, apresentando um teor de Ca
“nulo” na biomassa vegetal, demonstrando-se menos eficiente no acúmulo e liberação de
Ca quando comparado à quantidade de fitomassa produzida pelas demais espécies de
plantas. M. pruriens, U. brizantha e P, glaucum apresentaram semelhança na curva de
resposta, com liberação de ~48 kg ha-1 de Ca aos 80 DAD, o que equivale a 100% do
necessário. Durante todo o ciclo do milho essas espécies liberaram acumuladamente 50,18
kg ha-1, 50,03 kg ha-1 e 47,98 kg ha-1 de Ca respectivamente para o sistema. Comparando a
produção de fitomassa inicial, U. brizantha (24.172 kg ha-1), P. glaucum (17.084 kg ha-1 ) e
M. pruriens (6.248 kg ha-1), pode-se concluir que a C. spectabilis e P. glaucum
apresentaram na fase final teores mais elevados de Ca na biomassa vegetal, demonstrando-
se mais eficientes no acúmulo e liberação de Ca quando comparados à quantidade de Ca
ciclado na biomassa produzida pela E. coracana.

3.3. Liberação acumulada de micronutrientes por plantas de cobertura
A liberação acumulada de Cu, Mn e Zn apresentaram comportamentos
diferenciados nas cinco plantas de cobertura em estudo (Tabela 7). Já o processo de
degradação de matéria seca e a velocidade de liberação de Cu, Mn e Zn se demonstraram
semelhantes, ocorrendo uma elevada taxa de decomposição e liberado dos nutrientes entre
o intervalo de 0 a 45 DAD, e em seguida por uma taxa mais lenta entre 60 a 180 DAD para
as cinco espécies de plantas (Apêndice 1B).
Já a E. coracana obteve elevada taxa de decomposição e liberação de Mn entre o
intervalo de 0 a 60 DAD e a M. pruriens com elevada taxa de decomposição e liberação de
Zn entre o intervalo de 0 a 60 DAD. Isso é explicado porque a E. coracana e a M. pruriens
são espécies de relação C/N mais alta, com sistema radicular superficial mais distribuído e
consistente, caule e ramos mais resistentes (mais caule do que folhas), portanto mais
lignificados, retardando e estabilizando a liberação gradativa desses micronutrientes, que já
possuem mobilidade reduzida devido à sua própria composição (metais que se deslocam
com mais dificuldade em tecidos lignificados).
Na liberação de micronutrientes pelas plantas de cobertura, destaque para as
espécies U. brizantha e P. glaucum. No período de maior demanda de Cu, Mn e Zn pela
cultura do milho (80 DAE), verificou-se que a U. brizantha e P. glaucum haviam liberado
respectivamente, 96 e 94 g ha-1 de Cu, 645 e 503 g ha-1 de Mn e 208 e 478 g ha-1 de Zn,
representando aproximadamente 47 e 44% a mais das 65 g ha-1 de Cu, 78 e 38% a mais das
363 g ha-1 de Mn e 11% a mais e 155% das 187 g ha-1 de Zn necessários e acumulado na
fitomassa do milho para alta produtividade (>10.000 kg ha-1) (Tabela 7).
Nos maiores valores na liberação acumulada de Cu, Mn e Zn aos 180 DAD,
destaque para U. brizantha e P. glaucum. Essas superioridades em relação as outras
espécies são devido as suas maiores quantidades de fitomassas seca remanescente,
decorrente dos menores tempos de meia-vida (Tabela 5). Considera-se que a elevada taxa
de decomposição e liberação de nutrientes da fitomassa de U. brizantha e P. glaucum
reduziram as concentrações de lignina e celulose devido as menores relação C/N das
gramíneas, em comparação as maiores relação C/N das demais espécies no estudo.
Já a espécie E. coracana, que apresentou maior liberação total de Mn (Tabela 7),
devido à maior porcentagem de folhas na fitomassa seca em comparação à proporção de
caule da planta, haja vista que maior área foliar está associado com maior exportação de
Mn, sendo um micronutriente atuante em processos da fase fotoquímica da fotossíntese.
Logo no final dos 180 DAD (Tabela 3), apresentou um teor de Mn “nulo” na biomassa
vegetal, demonstrando-se menos eficiente no acúmulo e liberação de Mn quando
comparado à quantidade de fitomassa produzida pelas demais espécies de plantas.
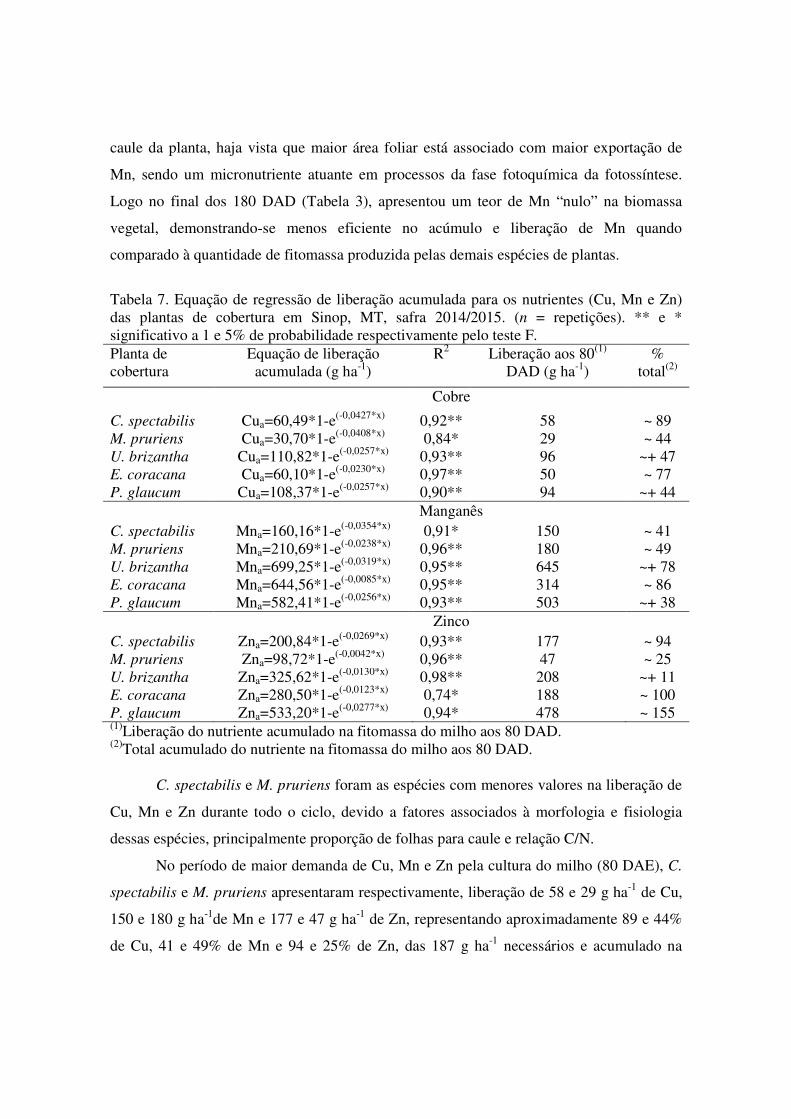
Tabela 7. Equação de regressão de liberação acumulada para os nutrientes (Cu, Mn e Zn) das plantas de cobertura em Sinop, MT, safra 2014/2015. (n = repetições). ** e * significativo a 1 e 5% de probabilidade respectivamente pelo teste F. Planta de cobertura
Equação de liberação acumulada (g ha-1)
R2 Liberação aos 80(1) DAD (g ha-1)
% total(2)
Cobre
C. spectabilis Cua=60,49*1-e(-0,0427*x) 0,92** 58 ~ 89 M. pruriens Cua=30,70*1-e(-0,0408*x) 0,84* 29 ~ 44 U. brizantha Cua=110,82*1-e(-0,0257*x) 0,93** 96 ~+ 47 E. coracana Cua=60,10*1-e(-0,0230*x) 0,97** 50 ~ 77 P. glaucum Cua=108,37*1-e(-0,0257*x) 0,90** 94 ~+ 44 Manganês C. spectabilis Mna=160,16*1-e(-0,0354*x) 0,91* 150 ~ 41 M. pruriens Mna=210,69*1-e(-0,0238*x) 0,96** 180 ~ 49 U. brizantha Mna=699,25*1-e(-0,0319*x) 0,95** 645 ~+ 78 E. coracana Mna=644,56*1-e(-0,0085*x) 0,95** 314 ~ 86 P. glaucum Mna=582,41*1-e(-0,0256*x) 0,93** 503 ~+ 38 Zinco C. spectabilis Zna=200,84*1-e(-0,0269*x) 0,93** 177 ~ 94 M. pruriens Zna=98,72*1-e(-0,0042*x) 0,96** 47 ~ 25 U. brizantha Zna=325,62*1-e(-0,0130*x) 0,98** 208 ~+ 11 E. coracana Zna=280,50*1-e(-0,0123*x) 0,74* 188 ~ 100 P. glaucum Zna=533,20*1-e(-0,0277*x) 0,94* 478 ~ 155 (1)Liberação do nutriente acumulado na fitomassa do milho aos 80 DAD. (2)Total acumulado do nutriente na fitomassa do milho aos 80 DAD.
C. spectabilis e M. pruriens foram as espécies com menores valores na liberação de
Cu, Mn e Zn durante todo o ciclo, devido a fatores associados à morfologia e fisiologia
dessas espécies, principalmente proporção de folhas para caule e relação C/N.
No período de maior demanda de Cu, Mn e Zn pela cultura do milho (80 DAE), C.
spectabilis e M. pruriens apresentaram respectivamente, liberação de 58 e 29 g ha-1 de Cu,
150 e 180 g ha-1de Mn e 177 e 47 g ha-1 de Zn, representando aproximadamente 89 e 44%
de Cu, 41 e 49% de Mn e 94 e 25% de Zn, das 187 g ha-1 necessários e acumulado na

fitomassa do milho para alta produtividade (>10.000 kg ha-1). Comparando os
micronutrientes remanescentes nas fitomassas na fase inicial (Tabela 3), C. spectabilis e M.
pruriens , pode-se concluir que a M. pruriens apresentou teores mais elevados de Cu, Mn e
Zn na biomassa vegetal, demonstrando-se mais eficiente no acúmulo e liberação desses
micronutrientes quando comparados à quantidade de micronutrientes remanescentes nas
fitomassas produzida pela C. spectabilis.
Destaque também para a M. pruriens, com maiores tempos de meia-vida, para o Cu,
Mn e Zn, devido a uma maior proporção de caule em relação à proporção de folhas do que
as outras espécies estudadas, com um maior teor de lignina e celulose no colmo do que nas
folhas, consequentemente uma maior relação C/N, permanecendo no solo por um maior
período, com uma menor taxa de decomposição e liberação de nutrientes da fitomassa.
3.4. Produtividade de Grão de Milho
Houve diferenças entre as plantas de cobertura para a produtividade de grãos de
milho (Tabela 8). As maiores produtividades do milho foram proporcionadas pelas espécies
de cobertura U. brizantha, P. glaucum e M. pruriens. A espécie U. brizantha (Braquiária
brizantha) foi a opção mais viável, por ter possibilitado a maior produtividade de milho,
comprovando a potencialidade dessa espécie para a produção de fitomassa e ciclagem de
nutrientes. A alta produtividade de milho cultivado sob a palhada da U. brizantha (11.034
kg ha-1), foi 17% superior a produtividade com E. coracana.
Tabela 5. Produtividade de grãos de milho cultivado após a palhada de plantas de cobertura em Sinop, MT, safra 2014/15.
Produtividade de grãos (kg ha-1) Diferença (%)
U. brizantha 11.034 a 16,8 P. glaucum 10.450 ab 10,6 M. pruriens 10.134 ab 7,2 C. spectabilis 9.870 b 4,4 E. coracana 9.450 b - CV (%) 12,4
A U. brizantha expressou elevado acúmulo de fitomassa, decomposição mais lenta
da palhada e maior liberação de nutrientes, quando comparada as demais espécies de

cobertura no trabalho. O que pode ter promovido maior tempo de imobilização de N pela
microbiota do solo, encontrado resultados semelhantes por Pacheco et al. (2013) em
produtividade de arroz com o uso da U. brizantha como planta de cobertura.
A P. glaucum é outra espécie de cobertura utilizada para a formação de palhada em
sistema plantio direto pela sua elevada produção de fitomassa e boa ciclagem de nutrientes.
A produtividade de milho cultivado sob a palhada desta espécie foi de 10.450 kg ha-1,
representando um aumento de 10,6 % na produtividade em relação a E. coracana. A
produtividade do milho cultivado na palhada de M. pruriens foi de 10,134 kg ha-1,
representando incremento de 7% na produtividade em relação a E. coracana.
A alta produtividade do milho mediante o uso das espécies de cobertura U.
brizantha, P. glaucum e M. pruriens é explicada pelas boas condições para o maior
desenvolvimento dessas espécies de cobertura, como bom índice pluviométrico mediante
irrigação nos períodos de “veranicos”, elevado acúmulo de fitomassa vegetal,
decomposição lenta e boa liberação de nutrientes nos estádios de maior demanda
nutricional de nutrientes da cultura do milho, com boa potencialidade de uso dessas
espécies para a produção de fitomassa e ciclagem de nutrientes pelo produtor de milho.
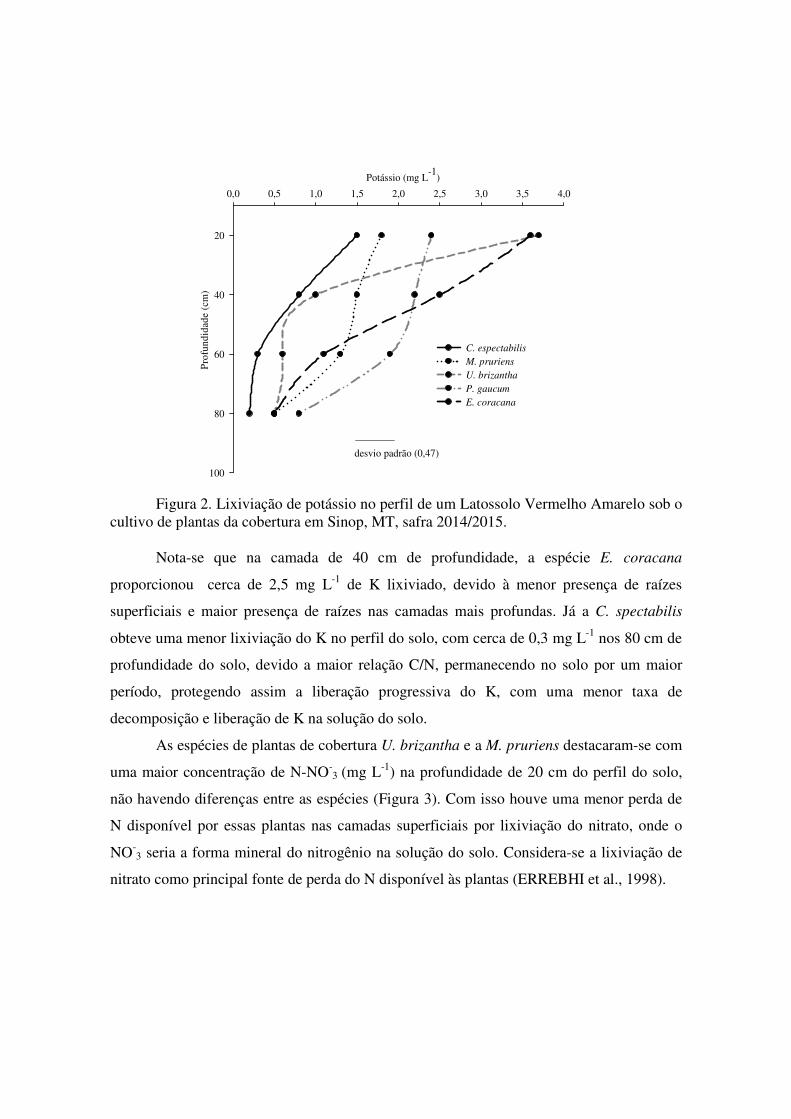
3.5. Lixiviação de potássio e nitrato no solo sob cultivo de plantas de cobertura
As espécies que mais acumularam potássio (K) foram também as que liberaram o
nutriente em maiores quantidades, com destaque para as espécies de cobertura U. brizantha
e E. coracana, com maiores concentração de K trocável disponível na solução do solo na
profundidade de 20 cm com valores superiores a 3.5 mg L-1 de K, onde estatisticamente são
semelhantes com um desvio padrão de 0,47 (Figura 2). Em consideração a lixiviação de
potássio na solução do solo, constatou-se que a espécie P. glaucum sofreu uma maior
lixiviação do K na profundidade de 80 cm no perfil do solo (cerca de 0.9 mg L-1).
Conforme Malavolta (1980), cerca de 80 % do K apresenta-se como substância solúvel na
planta, estando assim, sujeito à lixiviação. Presume-se que esta porcentagem possa ser
menor dependendo da espécie de planta de cobertura utilizada. Com relação ao potencial
de lixiviação da P. glaucum, Rosolem et al. (2003; 2007) relataram estudos comprovando a
maior susceptibilidade do milheto à lixiviação do K.
Potássio (mg L-1
)
0,0 0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0P
rofu
ndid
ade
(cm
)
20
40
60
80
100
C. espectabilis
M. pruriens
U. brizantha
P. gaucum
E. coracana
desvio padrão (0,47)
Figura 2. Lixiviação de potássio no perfil de um Latossolo Vermelho Amarelo sob o
cultivo de plantas da cobertura em Sinop, MT, safra 2014/2015. Nota-se que na camada de 40 cm de profundidade, a espécie E. coracana
proporcionou cerca de 2,5 mg L-1 de K lixiviado, devido à menor presença de raízes
superficiais e maior presença de raízes nas camadas mais profundas. Já a C. spectabilis
obteve uma menor lixiviação do K no perfil do solo, com cerca de 0,3 mg L-1 nos 80 cm de
profundidade do solo, devido a maior relação C/N, permanecendo no solo por um maior
período, protegendo assim a liberação progressiva do K, com uma menor taxa de
decomposição e liberação de K na solução do solo.
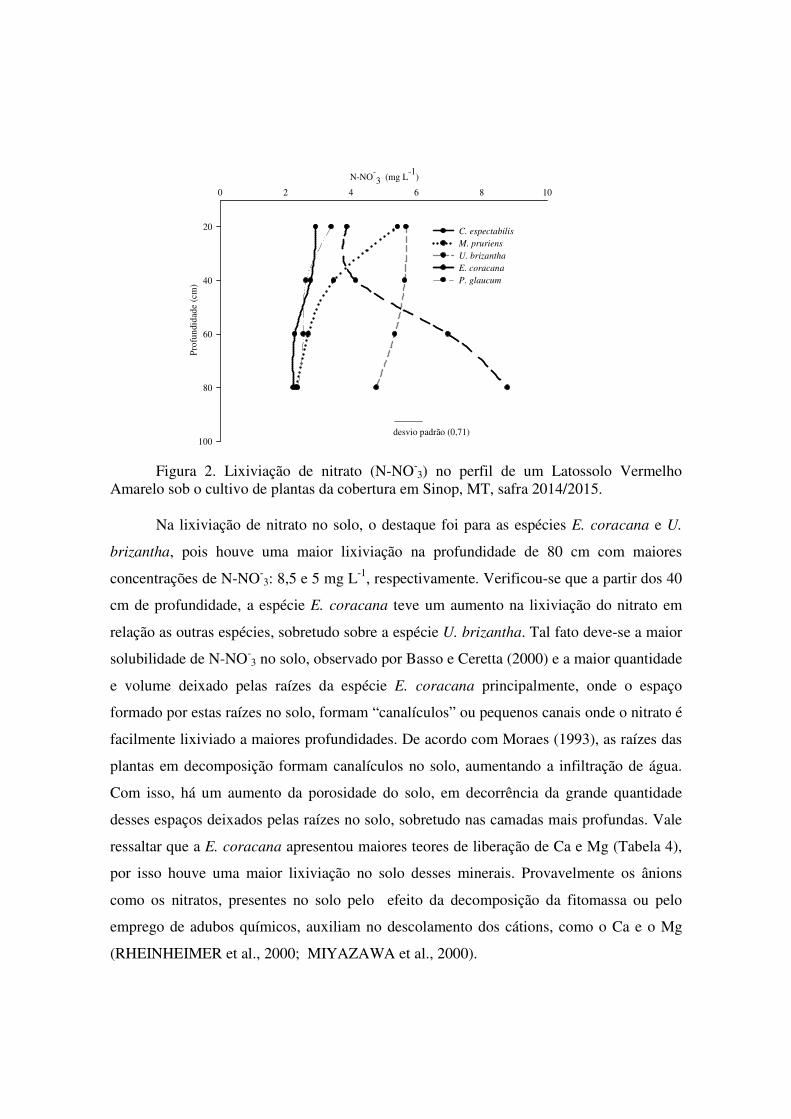
As espécies de plantas de cobertura U. brizantha e a M. pruriens destacaram-se com
uma maior concentração de N-NO-3 (mg L-1) na profundidade de 20 cm do perfil do solo,
não havendo diferenças entre as espécies (Figura 3). Com isso houve uma menor perda de
N disponível por essas plantas nas camadas superficiais por lixiviação do nitrato, onde o
NO-3 seria a forma mineral do nitrogênio na solução do solo. Considera-se a lixiviação de
nitrato como principal fonte de perda do N disponível às plantas (ERREBHI et al., 1998).
N-NO-3 (mg L
-1)
0 2 4 6 8 10P
rofu
ndid
ade
(cm
)
20
40
60
80
100
C. espectabilis
M. pruriens
U. brizantha
E. coracana
P. glaucum
desvio padrão (0,71)
Figura 2. Lixiviação de nitrato (N-NO-
3) no perfil de um Latossolo Vermelho Amarelo sob o cultivo de plantas da cobertura em Sinop, MT, safra 2014/2015.
Na lixiviação de nitrato no solo, o destaque foi para as espécies E. coracana e U.
brizantha, pois houve uma maior lixiviação na profundidade de 80 cm com maiores
concentrações de N-NO-3: 8,5 e 5 mg L-1, respectivamente. Verificou-se que a partir dos 40
cm de profundidade, a espécie E. coracana teve um aumento na lixiviação do nitrato em
relação as outras espécies, sobretudo sobre a espécie U. brizantha. Tal fato deve-se a maior
solubilidade de N-NO-3 no solo, observado por Basso e Ceretta (2000) e a maior quantidade
e volume deixado pelas raízes da espécie E. coracana principalmente, onde o espaço
formado por estas raízes no solo, formam “canalículos” ou pequenos canais onde o nitrato é
facilmente lixiviado a maiores profundidades. De acordo com Moraes (1993), as raízes das
plantas em decomposição formam canalículos no solo, aumentando a infiltração de água.
Com isso, há um aumento da porosidade do solo, em decorrência da grande quantidade
desses espaços deixados pelas raízes no solo, sobretudo nas camadas mais profundas. Vale
ressaltar que a E. coracana apresentou maiores teores de liberação de Ca e Mg (Tabela 4),
por isso houve uma maior lixiviação no solo desses minerais. Provavelmente os ânions
como os nitratos, presentes no solo pelo efeito da decomposição da fitomassa ou pelo
emprego de adubos químicos, auxiliam no descolamento dos cátions, como o Ca e o Mg
(RHEINHEIMER et al., 2000; MIYAZAWA et al., 2000).

4. Conclusões
Do ponto de vista técnico-científico nossas conclusões irão ficar para os relatórios
futuros, visto que as amostras ainda estão em procedimento analítico. Entretanto, do ponto
de vista da coordenação, condução e administração do projeto proposto, todas as etapas
estão rigorosamente em dia e de acordo com o cronograma previsto.
5. Referências
BOUWMANN, A. F.; GERMON, J. C. Introduction: Special issue – soils and climate change. Biology and Fertility Soils, Berlin, v. 27, p. 219. 1998. DORAN, J.W.; PARKIN, T.B. Defining and assessing soil quality. In: DORAN, J.W.; COLEMAN, D.C.; BEZDICEK, D.F.; STEWART, B.A. Defining soil quality for a sustainable environment. Soil Science Society of America, Madison, p. 3-21, 1994. HEINRICHS, R., VITTI, G.C., MOREIRA, A., FIGUEIREDO, P.A.M., FANCELLI, A.L.; CORAZZA, E.J. Características químicas de solo e rendimento de fitomassa de adubos verdes e de grãos de milho, decorrente do cultivo consorciado. Revista Brasileira de Ciência do Solo, v. 29, p. 71-79, 2005.
PRIOR, S.A.; TORBERT, H.A.; RUNION, G.B. & ROGERS, H. Elevated atmospheric CO2 in Agroecosystems: residue decomposition in the field. Environmental Management, New York, v.33, n.3, p.344-354, 2004.
SILVA, M.B.; KLIEMANN, H.J.; SILVEIRA, P. M.; LANNA, A. C. Atributos biológicos do solo sob influência da cobertura vegetal e do sistema de manejo. Pesquisa Agropecuária Brasileira, v.42, p.001-007, 2007.

Fotos: Cultivo das plantas de cobertura e extratores para análise de lixiviação de potássio, nitrato e carbono orgânico dissolvido, em Sinop-MT, safra 2015/2016.
Sinop, MT, 01 de abril de 2017.
____________________________
Prof. Dr. Fabiano André Petter Universidade Federal de Mato Grosso-UFMT





























