Embed Size (px)
Citation preview

UNIVERSIDAD AUTÓNOMA DE SAN LUIS POTOSÍ
FACULTAD DE CIENCIAS QUÍMICAS
PROGRAMA DE POSGRADO EN CIENCIAS EN BIOPROCESOS
IDENTIFICACIÓN Y CUANTIFICACIÓN DE
BIOMOLÉCULAS EN ESCAMOLES
(Liometopum apiculatum Mayr)
TESIS QUE PARA OBTENER EL GRADO DE
MAESTRA EN CIENCIAS EN BIOPROCESOS
PRESENTA:
I.A. MARÍA SARA GARCÍA MUÑOZ
DRA. ALICIA GRAJALES LAGUNES DIRECTORA
DR. MARCO MARTIN GONZÁLEZ CHÁVEZ CODIRECTOR
DRA. ANA PAULINA BARBA DE LA ROSA ASESORA
SAN LUIS POTOSÍ, S.L.P. ABRIL 2016


Proyecto realizado en:
Laboratorio de Ciencia de los Alimentos de la
Facultad de Ciencias Químicas de la
Universidad Autónoma de San Luis Potosí.
Laboratorio de Proteómica y Expresión Génica del
Instituto Potosino de Investigación Científica y Tecnológica
Laboratorio de Química Analítica de la
Facultad de Ciencias Químicas de la
Universidad Autónoma de San Luis Potosí
Con financiamiento de:
Fundación Produce San Luis Potosí
Beca-Tesis del Consejo Nacional de Ciencia y Tecnología
(CONACYT): 289902

Dedicatoria
Dedico esta tesis a mis seres más queridos, mis padres, Pablo García
Tovar y Paula Muñoz Zavala, por ser el pilar fundamental de nuestra familia, por
el apoyo, comprensión, son las personas que siempre están con nosotros en
cada nuevo reto en la vida, a mi hermana Dolores (Loli para mi) quien me da
consejos y comprenderme, cuando nadie más me entiende de lo que hablo, a mi
hermano Pablo por sus consejos, preocuparse y proporcionarme posibles
soluciones, a mi sobrina Paula Alejandra (Paulita para mi) quien es la persona
especial de la familia, le da vida y luz a nuestro hogar, por ser una amiga, a
Ángel por caminar juntos en la vida.

Agradecimientos Académicos
A la directora de tesis de maestría, Dra. Alicia Grajales Lagunes por la
confianza, conocimientos, apoyo, dedicación y entrega a esta investigación y
sobre todo me considero para este proyecto, muchas gracias por su paciencia,
comprensión y amistad, brindada en estos años de trabajo.
Al Dr. Marco Martín González Chávez por todo, la dedicación, apoyo,
confianza, tiempo, conocimientos, y su amistad a lo largo de este proyecto,
muchas gracias por sus enseñanzas tanto profesionales como de vida, por
darme un lugar en su laboratorio e integrarme a su grupo de trabajo, gracias.
A la asesora Dra. Ana Paulina Barba de la Rosa, Dr. José Ángel Huerta
Ocampo y al técnico M. en C. Alberto Barrera Pacheco del IPICYT, por su
confianza, asesoría, paciencia y todo el apoyo brindado para la obtención del
mapa proteómico de los escamoles.
Al Dr. Miguel Ángel Ruiz Cabrera, por sus observaciones, correcciones y
sugerencias en la redacción de esta tesis, gracias.
A las técnicas I.A. Cecilia Rivera Bautista y I.A. María Estela Núñez
Pastrana, por su enseñanza y apoyo técnico en los laboratorios de Ciencia de
los Alimentos y Química Analítica respectivamente, gracias.

Agradecimientos Personales
A mis padres, Pablo y Paula, por todo su apoyo incondicional en todos
mis proyectos de vida y por no dejarme nunca, muchas gracias los amo.
A mi hermana Loli y Pablo los mejores hermanos que puede tener una
persona, porque son solidarios y siempre están incondicionalmente.
A mi sobrina Pau por ser mi acompañante, aun cuando no le gustaba,
pero por convivir lo realizaba, muchas gracias Pau espero que te deje algo de
enseñanza.
A mi novio Ángel David por apoyarme en esta decisión, estar a mi lado,
gracias por esperarme y ser paciente.
A mis amigas de la maestría Fernanda y Lourdes, muchas gracias por
todo el tiempo compartido en esta aventura juntas, fueron una parte muy
importante de enseñanza y amistad, las aprecio mucho por los consejos, y a la
señora Rosa Martha, quien nos cuidó como una madre y amiga, muchas
gracias.
A mis amigos Osiel, Angelitos, Cony, Jessi, Ernesto, Rocío, Evelin, Sonia
y Neyvari, del laboratorio de síntesis orgánica con quienes compartí la mesa de
trabajo, mucho de nuestro tiempo en el laboratorio, el gusto por comer rico,
largas conversaciones que enriquecieron mi vida personal y profesional muchas
gracias.

ÍNDICE
ÍNDICE DE TABLAS ............................................................................................. i
ÍNDICE DE FIGURAS ........................................................................................... ii
RESUMEN ........................................................................................................... iii
ABSTRACT .......................................................................................................... iv
1. INTRODUCCIÓN ............................................................................................. 1
2. ANTECEDENTES ............................................................................................ 3
2.1 Insectos ....................................................................................................... 3
2.1.1 Entomofagia (consumo de insectos) .................................................... 5
2.1.2 Escamoles ............................................................................................ 8
2.1.2.1 Estudios realizados sobre la composición química y la
conservación de los escamoles ............................................................... 12
2.1.3 Proteínas ............................................................................................ 15
2.1.3.1 Función de las proteínas ............................................................. 15
2.1.3.2 Proteómica................................................................................... 16
2.1.3.3 Extracción de proteínas ............................................................... 17
2.1.3.4 Electroforesis bidimensional ........................................................ 17
2.1.3.5 Identificación de péptidos mediante LC-MS/MS .......................... 18
2.1.4 Lípidos y colesterol ............................................................................. 19
2.1.4.1 Función de los lípidos y colesterol ............................................... 19
3. JUSTIFICACIÓN ............................................................................................ 21
4. HIPÓTESIS .................................................................................................... 22
5. OBJETIVOS ................................................................................................... 22
5.1 General ..................................................................................................... 22
5.2 Particulares ............................................................................................... 22

6. METODOLOGÍA ............................................................................................. 23
6.1 Recolección de la muestra ........................................................................ 23
6.2 Análisis del proteoma por electroforesis en 2-D acoplado a espectrometría
de masas ........................................................................................................ 23
6.2.1 Extracción y solubilización de las proteínas ....................................... 23
6.2.2 Electroforesis bidimensional ............................................................... 24
6.2.3 Digestión en gel y análisis por espectrometría de masas tándem (LC-
MS/MS) ....................................................................................................... 25
6.2.4 Búsqueda en base de datos e identificación de las proteínas ............ 25
6.3 Análisis de ácidos grasos por cromatogafía de gases-masas (GC-MS) ... 26
6.3.1 Extracción de lípidos totales ............................................................... 26
6.3.2 Transesterificación de glicéridos ........................................................ 26
6.3.3 Análisis de ácidos grasos por GC-MS ................................................ 26
6.3.4 Cuantificación de ácidos grasos ......................................................... 27
6.4 Análisis de colesterol ................................................................................ 27
6.4.1 Extracción de colesterol ..................................................................... 27
6.4.2 Derivatización del colesterol ............................................................... 27
6.4.3 Análisis de colesterol por GC-MS....................................................... 28
6.4.4 Cuantificación de colesterol ............................................................... 28
7. RESULTADOS Y DISCUSION ....................................................................... 29
7.1 Mapa proteómico de la larva de Liometopum apiculatum Mayr ................ 29
7.2 Identificación de proteínas de Liometopum apiculatum Mayr ................... 30
7.3 Clasificación de proteínas de Liometopum apiculatum Mayr .................... 41
7.3.1 Componentes celulares ...................................................................... 41
7.3.1.1 Estructura celular ......................................................................... 41

7.3.1.2 Regiones extracelulares .............................................................. 42
7.3.2 Funciones moleculares ...................................................................... 43
7.3.2.1 Proteínas asociadas con trascripción y traducción ...................... 43
7.3.2.2 Actividad antioxidante .................................................................. 43
7.3.2.3 Actividad de transporte ................................................................ 44
7.3.2.4 Proteínas de enlace ..................................................................... 45
7.3.3 Procesos biológicos ........................................................................... 46
7.3.3.1 Proteínas relacionadas con metabolismo ................................... 46
7.3.3.1.1 Metabolismo de carbohidratos .............................................. 46
7.3.3.1.2 Metabolismo de aminoácidos ................................................ 47
7.3.3.1.3 Metabolismo de lípidos ......................................................... 48
7.3.3.2 Proteínas de plegamiento y degradación .................................... 49
7.3.4 Señalización celular ........................................................................... 51
7.3.5 Procesos de óxido-reducción ............................................................. 51
7.3.6 Otros procesos ................................................................................... 52
7.4 Perfil de ácidos grasos .............................................................................. 53
7.5 COLESTEROL LIBRE ............................................................................... 59
8. CONCLUSIONES ........................................................................................... 61
9. BIBLIOGRAFÍA .............................................................................................. 62
ANEXO 1 ............................................................................................................ 85
ANEXO 2 ............................................................................................................ 88

i
ÍNDICE DE TABLAS
Tabla 1. Contenido de proteínas y grasa de varios alimentos. ............................. 4
Tabla 2. Composición química de los escamoles recolectados en el estado de
Puebla. ............................................................................................................... 12
Tabla 3. Contenido de aminoácidos esenciales en escamoles, Liometopum
apiculatum Mayr., (g/100g de proteína, base seca). .......................................... 13
Tabla 4. Contenido de algunos minerales en escamoles (g/100g). .................... 14
Tabla 5. Composición proteica de los escamoles, spots de proteínas analizadas
por LC-MS/MS e identificadas contra el orden Hymenoptera de la base de datos
de proteínas de NCBI. ........................................................................................ 32
Tabla 6. Perfil de ácidos grasos en escamoles. ................................................ 54
Tabla 7. Cuantificación de ácidos grasos más abundantes en larvas de
Liometopum apiculatum Mayr con un % relativo ≥ 1.0% en mg/g. ..................... 58

ii
ÍNDICE DE FIGURAS
Figura 1. Registro numérico de especies de insectos comestibles por país,
(Jongema, 2012). ................................................................................................. 6
Figura 2. Micrografía de la hormiga Liometopum apiculatum Mayr. (Del Toro et
al., 2009). ............................................................................................................. 9
Figura 3. Distribución de Liometopum apiculatum Mayr. (Del Toro, et al., 2009). 9
Figura 4. Trabécula extraída en un nido de Liometopum apiculatum Mayr en el
ejido Pocitos, Charcas, S.L.P. (Lara Juárez, 2013). ........................................... 10
Figura 5. Ciclo de vida de la hormiga (http://acuariofiliamadrid.org/Thread-
Hormigas). .......................................................................................................... 10
Figura 6. Fotografía del Laboratorio de Ciencia de los Alimentos, escamoles
(pupas de hormiga). ........................................................................................... 11
Figura 7. Mapa 2-DE de la larva Liometopum apiculatum Mayr. Las proteínas
fueron separadas en tiras IPG de 24 cm con pH de 3-10 (A) y 5-8 (B) y SDS-
PAGE al 13%. .................................................................................................... 29
Figura 8. Mapa proteómico de la larva Liometopum apiculatum Mayr. .............. 30
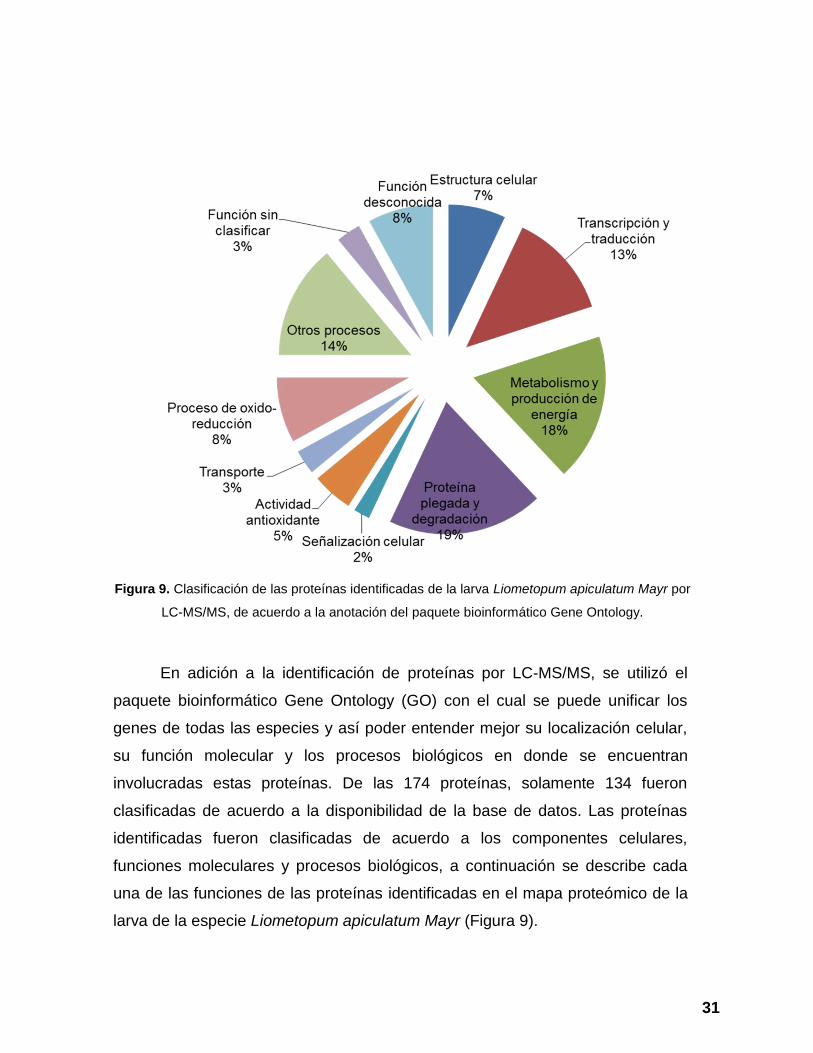
Figura 9. Clasificación de las proteínas identificadas de la larva Liometopum
apiculatum Mayr por LC-MS/MS, de acuerdo a la anotación del paquete
bioinformático Gene Ontology. ........................................................................... 31
Figura 10. Cromatograma de metil esteres de ácidos grasos de larvas
Liometopum apiculatum Mayr (escamoles). 1, Derivatisante (trifluoruro de boro);
2, ácido láurico; 3, ácido tridecil; 4,ácido miristíco; 5, ácido cis-9-tetradecenoico;
6,ácido pentadecílico; 7, ácido palmítico; 8, ácido palmitoleico; 9, ácido
margárico; 10, ácido cis-10-heptadecenoico; 11, ácido esteárico; 12, ácido
oleico; 13, Pentacosano; 14, ácido linoleico; 15, ácido cis-10-nonadecenoico;16,
ácido linolénico; 17, ácido linoleic; 18, ácido araquídico; 19, ácido cis-11-
eicosenoico; 20, Heptacosano; 21, Heptacosanol. ............................................. 55
Figura 11. Cromatograma de ácido cis-10-heptadecenoico, ácido cis-10-
nonadecenoico y ácido cis-11-eicocenoico. ....................................................... 55
Figura 12.Cromatograma comparativo de colesterol estándar y escamoles. ..... 60

iii
RESUMEN
Los escamoles (Liometopum apiculatum Mayr), son los insectos de mayor
consumo en México por su valor nutricional y sus aspectos culinarios. En la
actualidad, se conoce su composición química general, pero no hay informes
específicos respecto a la identificación de proteínas y ácidos grasos. Por lo
tanto, el objetivo de este estudio fue identificar el perfil de proteínas (proteoma) y
ácidos grasos presentes en escamoles.
Las proteínas fueron analizadas empleando herramientas de proteómica y
espectrometría de masas en tándem (LC-MS/MS). El análisis de datos de LC-
MS/MS se realizó empleando el motor de búsqueda MASCOT contra la base de
datos de proteínas de NCBI para organismos del orden Hymenoptera. 175 spots
de proteínas fueron exitosamente identificadas y correspondieron a 134
proteínas diferentes que fueron clasificadas en 11 grupos en relación a su
función conocida: estructura celular (7%), trascripción y traducción (13%),
metabolismo y producción de energía (18%), plegamiento y degradación de
proteínas(19%), señalización celular (2%), actividad antioxidante (5%),
transporte (3%), proceso de óxido-reducción (8%), otros procesos (14%), de
función no clasificada (3%) y función desconocida (8%).
El perfil de ácidos grasos fue analizado como ésteres metílicos de ácidos
grasos, utilizando un cromatógrafo de gases-masas. Los ácidos grasos fueron
identificados por comparación de sus espectros de masas con la biblioteca
NIST11/WILEY09. Del total de ácidos grasos identificados, 66.96%
correspondieron a ácidos grasos insaturados y 31.02% ácidos grasos saturados
de los cuales, los más abundantes fueron el ácido oleico y el ácido palmítico
respectivamente.
En el presente estudio se logró elucidar por primera vez el proteoma de
los escamoles, así como identificar y cuantificar su perfil de ácidos grasos lo cual
sugiere que este sistema alimenticio es una fuente importante de proteínas,
ácidos grasos insaturados esenciales.

iv
ABSTRACT
Escamoles (Liometopum apiculatum Mayr), insect are one of the most
consumed in Mexico, for its nutritional value and culinary aspect. At present,
have been reported overall chemical composition of escamoles, but there are no
specific reports regarding to identification of protein and fatty acids. Therefore,
the objective of this study was to analyze and characterize the profile of proteins
(proteome) and fatty acids presents in escamoles.
Proteins were analyzed using proteomic tools and tandem mass
spectrometry (LC-MS/MS). Data analysis of LC-MS/MS was performed using
MASCOT search engine against NCBI protein database of the organism to the
order Hymenoptera. 175 protein spots were successfully identified were
classified and correspond to 134 different proteins that were classified into 11
groups according to its function: cell structure (7%), transcription and translation
(13%), metabolism and energy production (18%), protein folding and degradation
(19%), cell signaling (2%), antioxidant activity (5%), transport (3%), oxidation-
reduction processes (8%), other processes (14%), unclassified or unknown
function (11%).
The fatty acids profile was analyzed as fatty acids methyl esters were
carried out with a gases-chromatograph mass. The fatty acids were identified by
comparison of their mass spectra with NIST11/WILEY09 library. Unsaturated
(66.96%) and saturated (31.02%) fatty acids were identifies, the most abundant
were oleic acid (56.58%), and palmitic acid (22.3%). In this study was possible to
elucidate the first mapping of escamoles proteome as well as identify and
quantify the fatty acid profile, suggesting foods system an important source of
protein and essentials unsaturated fatty acids.

1
1. INTRODUCCIÓN
La entomofagia se refiere al uso de insectos de diversas especies como
alimentos y es una tradición que se practica en varias regiones del mundo entre
las que destacan Asia, África y América Latina (van Huis et al., 2013).
En algunos países en vías de desarrollo, el consumo de insectos es
utilizado para satisfacer necesidades alimentarias de algunos grupos étnicos
debido a que son altamente nutritivos. Contienen altas concentraciones de
proteínas, grasas, vitaminas A, C y el complejo B, minerales (Ramos-Elorduy et
al., 2006). En los países desarrollados, los platillos elaborados a base de
insectos son considerados exóticos por su aspecto y el sabor característico que
presentan.
En México, las larvas de hormigas (Liometopum apiculatum Mayr),
conocidas como escamoles, se consumen desde épocas pre-hispánicas. Los
escamoles presentan un elevado valor nutricional y son considerados como un
manjar con un sabor cremoso a nuez y con una textura similar al queso cottage.
Sin embargo, a pesar de los estudios realizados para conocer las causas
de deterioro de los escamoles y del desarrollo de tecnologías para incrementar
su vida de anaquel (Salas-García, 2011; Ramírez-García, 2012; Castillo-
Andrade et al., 2015) existe un gran desconocimiento sobre el perfil de proteínas
(proteoma) y lípidos de los escamoles, los cuales son de gran importancia para
la caracterización de la biología, los procesos metabólicos y nutritiva del
escamol.
Por ello, en este trabajo de investigación se propone el uso de proteómica
y la cromatografía de masas GC-MS para elucidar el proteoma y el perfil de
ácidos grasos y así conocer de manera específica las propiedades nutricionales
y funcionales de las proteínas y lípidos presentes en los escamoles.
Este estudio presenta una revisión bibliográfica detallada sobre el tema,
así también se presentan la justificación, hipótesis, objetivo general y de manera
detallada se expresan los objetivos específicos realizados durante el desarrollo

2
de la investigación. Se muestran los materiales y métodos que describen las
técnicas utilizadas para la elucidación del proteoma y el perfil de ácidos grasos.
En la sección de resultados y discusión se aborda la función específica de cada
proteína identificada en base a su función biológica, así como la importancia y
beneficios de los ácidos grasos en la salud del ser humano. Finalmente se
exponen las conclusiones del trabajo y las referencias bibliográficas revisadas
en este estudio.

3
2. ANTECEDENTES
2.1 Insectos
En la actualidad 1.5 millones de especies de animales que existen en la
tierra, han sido descritos. La mayoría (66%) son artrópodos tales como
crustáceos, arácnidos, miriápodos e insectos (hexápodos) lo cuales representan
el 75% de todos los animales (Chapman, 2013). La palabra insecto
etimológicamente proviene del latín insectum, literalmente, “cortado en medio”,
como copia del griego éntomon (ἒντομον) (Cortés et al., 2014).
Los insectos entre sus características comunes está el tener tres regiones
en su cuerpo: cabeza, tórax y abdomen (Nason, 2000). Se distinguen de los
otros artrópodos porque tienen tres pares de patas, por lo general, dos pares de
alas insertadas en la región media, o torácica, del cuerpo. Además, la cabeza
lleva típicamente un único par de antenas y un par de ojos simples o
compuestos. Hay un sistema traqueal para el intercambio de gases y los
conductos se abren en el extremo posterior del abdomen (Ruppert, Barnes,
1996). Tienen un exoesqueleto que les permite protegerse del medio ambiente,
son de sangre fría, pasan por la metamorfosis, se reproducen rápidamente y
tienen grandes poblaciones (van Huis, 2013).
En algunos casos los insectos se consideran plagas, sin embargo estos
desempeñan muchas funciones benéficas para el medio ambiente por ejemplo:
facilitan la polinización, diseminan las semillas, mejoran la textura del suelo y
descomponen la cubierta vegetal muerta. Proporcionan productos para el
beneficio del ser humano tales como la seda, la miel, la cera de abeja y tintes
(Vantomme, 2010). Además estos juegan un papel clave en el flujo de energía a
través del ecosistema, principalmente como herbívoros, así mismo como
depredadores, parásitos, que a su vez pueden ser consumidos por vertebrados
insectívoros de nivel superior. Alguna especies de insectos son usados como
recursos terapéuticos en los sistemas médicos de muchas culturas alrededor del
mundo (Costa-Neto, 2006). En muchas partes del mundo los insectos continúan

4
teniendo un papel importante como componente normal de la dieta humana
debido a los nutrientes que estos aportan como proteínas, grasas, vitaminas,
minerales (Gillott, 2005).
En la literatura, se ha reportado que el valor nutricional de los insectos
comestibles es muy variado, debido a la amplia variedad de especies que
existen. Incluso en una misma especie se han reportado variaciones que
dependen de la etapa metamórfica del insecto, en particular, para especies con
metamorfosis completa, como hormigas, abejas y escarabajos. Además del
hábitat y la dieta que tenga dicho insecto.
Sin embargo, a pesar de la variabilidad del valor nutricional de los
insectos, este valor se encuentra dentro del rango del valor nutricional de los
alimentos convencionales.
En la Tabla 1 se presenta el contenido de proteínas y grasas de alimentos
convencionales y de insectos. Se puede observar que el contenido de proteínas
en los insectos es significativamente mayor que en cualquier otro tipo de
alimento.
Tabla 1. Contenido de proteínas y grasa de varios alimentos.
Producto Proteína (%) Grasa (%)
Bovino
Cordero
Pollo
Pescado
Leche
Huevo
Insectos
17.4-19.4
14.4-16.8
20.6-23.4
18.2-20.9
3.5-4.5
12.9
20-56.7
15.8-25.1
19.4-27.1
1.9-4.7
1.2-10.0
3.7-3.9
11.5
6.7-9.2
Ghaly (2009).
Numerosos artículos publicados reportan el valor nutricional de diferentes
especie de insectos en las etapas que son consumidos por el humano,

5
presentando: análisis proximales, proteínas, aminoácidos, lípidos, minerales y
vitaminas (Rumpold, Schlüter, 2013). En 1995 Ladrón de Guevara, et al.,
determinaron el contenido de aminoácidos en algunos de los insectos
comestibles en México, gusano blanco de maguey, escamoles entre otros. El
objetivo de este estudio fue determinar los aminoácidos esenciales ya que de
estos dependerá la calidad de las proteínas que permite una mejor digestibilidad
y asimilación del mismo. Los resultados mostraron que los insectos evaluados
de la Orden: Orthoptera, Homoptera, Lipidoptera, Diptera y Hymenoptera,
superan el contenido de proteínas así como de los aminoácidos esenciales
(Lisina, Leucina, Isoleucina, Metionina e Histidina) que la WHO, FAO y ONU
recomienda para la ingesta diaria.
En el oeste de Kenia las termitas son una especie comúnmente
consumida. Estudios revelaron que el contenido de ácidos grasos insaturados
(50.54 a 67.83%) fue superior a los ácidos grasos saturados (32.17 a 49.46%),
esos autores sugieren que los insectos no solamente son una fuente importante
de proteínas, sino que también de ácidos grasos insaturados (Kinyuro et al.,
2013).
Debido a la calidad nutritiva de estas especies, la FAO considera a los
insectos como una fuente alternativa de proteínas y otros nutrientes para
alimentación humana ya que se estima que el crecimiento de la población
mundial alcanzará 8.100 millones en 2025 y 9.600 millones en 2050 (Naciones
Unidas, 2014; van Huis, Dicke, van Loon, 2015).
2.1.1 Entomofagia (consumo de insectos)
El hábito de consumir insectos de diferentes especies por placer o para
satisfacer las necesidades alimenticias se conoce como entomofagia, y ha
existido desde épocas remotas alrededor del mundo (Costa-Neto, 2003; Viesca-
González y Romero-Contreras, 2009).

6
Proporcionar números definitivos de especies de insectos comestibles es
difícil por varias razones, el conocimiento de las especies por una solo persona,
los diferentes nombres coloquiales usados en distintas regiones para referirse a
la misma especie. Jongema (2012), llevó a cabo un inventario a nivel mundial
de los insectos comestibles, estableciendo que existen 1900 especies
comestibles y están distribuidas en diferentes países del mundo: 250 especies
de insectos fueron localizados en África, 180 en China y 549 en México (Figura
1).
El país donde mayormente se consumen insectos es Tailandia, ya que
son más de 200 especies las que se degustan en ese país. Los insectos más
comunes que se encuentran en los mercados de la capital Bangkok o en la
cadena de restaurantes conocida como “Insect King” son grillos, escarabajos,
cucarachas acuáticas, escorpiones y tarántulas. En este país, más de 15,000
familias se dedican al cultivo de granjas de grillos (FAO, 2007). En el sur de
Figura 1. Registro numérico de especies de insectos comestibles por país, (Jongema, 2012).

7
África esta costumbre también juegan un papel muy importante para el
desarrollo económico de algunas comunidades. La ONU afirma que en
Zimbabwe y Botswana cada año se produce una ganancia 9,500 millones de
dólares mediante la cosecha de orugas en los bosques de árboles Mopani.
Estos extravagantes animales son cada vez más apreciados en la región
occidental del mundo, lo que se ha traducido en el aumento de las exportaciones
que llevan a cabo países africanos como Sudán, Nigeria y República de Congo.
Éste último exporta entre tres y cinco toneladas de orugas a Bélgica y Francia.
En Japón no es raro que los restaurantes incluyan en su carta platillos con zaza-
mushi (larva acuática) y hachi-no-ko (larva de avispa); o en China saltamontes o
escarabajos marinados son salsa de soya y jengibre o en Indonesia libélulas
saladas y fritas. En Cerdeña, Italia se elabora un queso llamado casu marzu o
formaggio marcio, cuyo sabor se consigue gracias a que se deja fermentar con
larvas de mosca. En España, Canadá y Colombia se consume las hormigas
culonas de abdomen prominente y alas largas, al cocinarse se les elimina la
cabeza y las alas y únicamente se consume el abdomen asado con un poco de
sal.
Arana (2003) explica que una espontánea evolución cultural ha ocurrido,
lo cual ha conducido a que la entomofagia sea una moda en Estados Unidos y
Europa, debido a que a base de insectos se preparan platillos gourmet y
exóticos a precios elevados.
En México, la entomofagia se practica desde la época prehispánica
(Ramos-Elorduy et al., 1998) y se ha incrementado de manera significativa, ya
que van apareciendo a nuestro alcance alimentos novedosos utilizados de forma
habitual y poco a poco se introducen en la alimentación cotidiana, como es el
caso de los insectos (Matallana-González y Torija-Isasa, 2006).
En la República Mexicana, se han censado a la fecha 540 especies de
insectos comestibles. El mayor grupo son los Coleópteros (443 especies) que
comprenden a los escarabajos y luciérnagas, seguido de los Hymenópteros (307
especies) a la cual pertenecen las abejas, avispas y las hormigas, los Ortópteros

8
(235 especies) comprenden a los grillos y langostas y a los Lepidópteros (228
especies) pertenecen las mariposas y polillas (Costa-Neto, 2003; Ramos-
Elorduy y Pino, 2006). Sin embargo son pocas las especies que se
comercializan, la venta de muchas especies de insectos comestibles se lleva a
cabo de acuerdo a la época de abundancia de cada insecto, pero en general
podemos decir que es durante la época de lluvias. Los insectos que tienen
mayor comercialización son los gusanos blancos (Aegiale hesperiaris), y rojos
(Comadia redtembacheri) del maguey, los chapulines (Sphenarium mexicanum
histrio Gerstaecker), las chicatanas (Atta mexicana Smith, Atta cephalotes), las
abejas sin aguijón (Scaptotrigona mexicana Guérin) y los escamoles
(Liometopum apiculatum Mayr).
El consumo de escamoles se ha ido incrementando significativamente, no
solamente en zonas rurales (cultura indígenas) sino también en mercados y
restaurantes de alto prestigio (tipo gourmet) de zonas urbanas de la parte centro
y sur del país (San Luis Potosí, Estado de México, Hidalgo, Tlaxcala, Oaxaca,
Chiapas, Puebla y Veracruz) (Ramos-Elorduy, 2006).
2.1.2 Escamoles
La palabra escamol proviene del náhuatl, azcatl: hormiga y milli: guiso,
que significa Azcamolli: guisos de hormiga (Tarango-Arámbula, 2012). Los
escamoles son el conjunto en desarrollo de embrión, larva y pupa de la hormiga,
el cual habita en diversos tipos de vegetación como matorrales, magueyes,
nopales, entre otros. La hormiga que produce los escamoles es de la Clase
Insecta, Orden Hymenoptera, Familia Formicidae, Genero Liometopum, Especie
apiculatum Mayr (Juárez Sandoval, 2010) (Figura 2). Su distribución va del
suroeste de los Estados Unidos hacia el norte y suroeste de México (Figura 3)
(Del Toro et al., 2009).

9
Figura 2. Micrografía de la hormiga Liometopum apiculatum Mayr. (Del Toro et al., 2009).
Figura 3. Distribución de Liometopum apiculatum Mayr. (Del Toro, et al., 2009).

10
Estas hormigas se
localizan en nidos que
contienen estructuras
comúnmente llamadas
guacales hechos de tierra,
con una estructura
parecida al de un panal
denominado trabécula,
(Figura 4) en donde en
cada orificio es depositado
un huevecillo (Miranda
Román, 2011).
El ciclo de
desarrollo de las hormigas
es de 6 semanas o 2
meses, lo que depende de
cada especie y de factores
ambientales como la
temperatura, humedad,
disponibilidad de alimento,
disturbios, stress, etc. Son
consideradas especies
sociales, viven en colonias
compuestas de muchos
individuos, se encuentran
organizadas en la reina y
macho, quienes llevan a
cabo la reproducción,
también de obreras,
soldados, nodrizas y los
Figura 4. Trabécula extraída en un nido de Liometopum
apiculatum Mayr en el ejido Pocitos, Charcas, S.L.P. (Lara
Juárez, 2013).
Figura 5. Ciclo de vida de la hormiga
(http://acuariofiliamadrid.org/Thread-Hormigas).

11
estados jóvenes, que son huevos, larvas y pupas (Figura 5).
Se sabe que cada nido puede producir hasta 1.5 Kg durante el año, lo
cual dependerá de distintos factores, como son la edad de la reina, antigüedad
de la colonia, tipo de suelo, producción de nido y condiciones del hábitat. En los
meses de marzo y abril se lleva a cabo la recolección de los escamoles. Los
nidos por lo general se encuentran en una profundidad de 50 cm hasta 1.50 m
(Tarango-Arámbula, 2012).
Los nidos son localizados siguiendo el rastro de cuatro caminos de las
hormigas donde estas se unen, llegando a la entrada del nido, una vez
localizado la tierra se elimina y los huacales son removidos para obtener los
escamoles, sus características son: color blanco opaco, con una longitud de 10
mm y un diámetro de 3 a 5 mm, (Figura 6).
En la literatura pocos son los estudios reportados de esta especie, se han
realizado investigaciones referentes a aspectos biogeográficos y a su valor
nutritivo, los cuales han proporcionado información valiosa acerca de su
distribución geográfica, su estacionalidad y su comercialización, así como su
Figura 6. Fotografía del Laboratorio de Ciencia de los Alimentos,
escamoles (pupas de hormiga).

12
composición química. De esta manera se conoce que los escamoles están
distribuidos principalmente en los estados de México, Hidalgo, Tlaxcala, Puebla,
Zacatecas y San Luis Potosí. Su recolección es muy corta únicamente se
recolectan durante el periodo de febrero-abril y cada nido puede proveer hasta 4
kilos de escamoles. Su comercialización se realiza en los restaurantes de
prestigio del Distrito Federal y de los diferentes estados del país, es considerado
“platillo gourmet” por su sabor especial y su precio. El kilogramo de este
producto en el sector restaurantero llega a valer hasta $3000.00. El platillo típico
que se utiliza para su comercialización es “A la mantequilla” (Castillo-Lara y
Aguirre-Rivera, 2008; Ramos-Elorduy et al. 2007).
2.1.2.1 Estudios realizados sobre la composición química y la
conservación de los escamoles
En la Tabla 2 se indica la composición química de los escamoles, donde
pueden observarse que presentan un alto valor nutritivo que los hace altamente
competitivos con cualquier otro alimento.
Tabla 2. Composición química de los escamoles recolectados en el estado de Puebla.
Componente Porcentaje (%)
Proteínas 39.67
Carbohidratos 19.22
Grasas 36.87
Sales minerales 1.8
Vitamina A 2.91 U.I
Vitamina C 36.14 mg
Tiamina 0.15 mg
Rivoflavina 0.34 mg
Niacina 0.67 mg
Ramos-Elorduy et al. (2001, 2007)

13
Así mismo Ramírez-García (2011) indican que los escamoles
recolectados en el Altiplano Potosino tienen 37.33% de proteínas, 37.54% de
carbohidratos y 22.26% de grasa, los cuales difieren en los valores de
carbohidratos y grasas que se reportan para los escamoles recolectados en el
estado de Puebla (Tabla 2). Estas diferencias pueden ser debida al tipo de
alimentación o a las condiciones ambientales donde se desarrollan los nidos,
porque Ramos Elorduy et al., (1998) reporta valores de 37.33% de proteína,
42.26% de grasa y 17.35% de carbohidratos para insectos recolectados en el
Estado de México.
De igual manera Ramírez-García (2011) indica que los escamoles tienen
una actividad de agua (aw) de 0.99 y un pH de 6.5, así como una elevada
actividad enzimática.
Ladrón de Guevara, et al., (1995) reportaron el contenido de aminoácidos
esenciales de los escamoles en g/100g de proteína, (Tabla 3) y es superior al de
algunos alimentos de origen animal.
Tabla 3. Contenido de aminoácidos esenciales en escamoles, Liometopum apiculatum Mayr.,
(g/100g de proteína, base seca).
Proteína (%) Lys Leu Ile Met Val Phe Tyr Thr Trp His
41.66 6.0 8.9 4.4 1.8 4.8 3.5 6.8 3.5 0.62 2.9
Lys (lisina), Leu (leucina), Ile (Isoleusina), Met (metionina), Val (valina), Phe (fenil alanina), Tyr
(tirosina), Thr (treonina), Trp (triptófano) y His (histidina).
Así mismo en la Tabla 4 se reportan los valores del contenido de
minerales presentes en los escamoles (Ramos-Elorduy et al., 1998).

14
Tabla 4. Contenido de algunos minerales en escamoles (g/100g).
Orden Sales minerales Sodio Potasio Calcio Zinc
Liometopum apiculatum
(Tulancalco, Hidalgo) 5.50 0.075 0.076 0.80 0.032
Liometopum apiculatum
(Puerto México, Hidalgo) 5.05 0.110 0.092 0.083 0.032
La composición química y las propiedades fisicoquímicas (pH y aw) hacen
que los escamoles sean altamente perecederos y que por lo tanto tengan una
corta vida de anaquel.
Por ello el trabajo realizado por Castillo Andrade et al., (2015) fue
enfocado en determinar la vida de anaquel de lo escamoles en parámetros de
textura y actividad enzimáticas de diferentes tipos de proteasas con la finalidad
de obtener información para desarrollar métodos alternativos de conservación.
En dicho estudio se logró elucidar por primera vez que la causa principal
del deterioro de los escamoles, la cual es en primera instancia la actividad
proteolítica de las enzimas endógenas. La evaluación de la activad proteolítica
aunada a los estudios estructurales mostró que durante el almacenamiento, los
escamoles sufren una degradación evidente que afecta directamente su calidad
indicando que su vida de anaquel es de 5 días en refrigeración a 4°C.
Buscando incrementar la vida de anaquel de los escamoles, en los
últimos años se ha trabajado en la estandarización del proceso de enlatado para
la conservación de los mismos. En dicho estudios se logró incrementar la vida
de anaquel de los escamoles procesados en escabeche y salmuera hasta 2
años (Salas García, 2011).
Un segundo estudio relacionado con el incremento de la vida de anaquel
de los escamoles fue realizado por Ramírez-García (2011) utilizando el método
de barrera como método de conservación. La finalidad de este estudio fue
obtener escamoles frescos mínimamente procesados con características
organolépticas aceptadas por el consumidor. Los resultados indicaron que fue

15
posible incrementar la vida de anaquel de los escamoles frescos hasta 35 días
de almacenamiento a 4°C.
En base a la relevancia que ha tomado el consumo de insectos en los
últimos años a nivel mundial y nacional y en que no existen estudios específicos
sobre la composición de proteínas y lípidos en escamoles este estudio fue
enfocado en el estudio de estas biomoléculas ya que tienen diversas funciones
nutritivas y fisiológicas en el organismo humano.
2.1.3 Proteínas
Las proteínas son moléculas complejas constituidas por carbono,
hidrógeno, oxígeno y nitrógeno y, a veces, también otros elementos como
azufre, hierro, cobre, fósforo y zinc. Están formadas por aminoácidos unidos
entre sí por enlaces peptídicos (Cambero Rodríguez, 1998). Son polímeros muy
complejos, constituidos por hasta 20 aminoácidos distintos, unidos por enlaces
peptídicos. (Fennema, 2000).
2.1.3.1 Función de las proteínas
Se encuentran en plantas y animales. En estos últimos ayudan a formar
las estructuras de soporte y protección, tales como cartílago, la piel, las uñas, el
pelo y el músculo. Son los componentes principales de las enzimas, anticuerpos
y de líquidos como la sangre, la leche y la clara del huevo (Potter, 1978).
Los procesos biológicos se encuentran regulados por reacciones
bioquímicas de estas biomoléculas. La totalidad de las proteínas que se
expresan en un organismo es llamado proteoma y el campo de estudio
correspondiente es llamado proteómica (Passarge, 2010). El proteoma es un
elemento altamente dinámico, cuyos componentes varían de un organismo,
tejido, célula o comportamiento subcelular, por consecuencia de cambios en su
entorno, situación de estrés, cambios bioquímicos o fisiológicos. Es por ello que
es importante conocer el perfil proteico de los escamoles, para poder

16
comprender la biología de los escamoles y los tipos de proteínas presentes en
los mismos. La caracterización del proteoma en diversos sistemas biológicos se
ha realizado a través de la proteómica.
2.1.3.2 Proteómica
La proteómica es definida como el análisis simultáneo de mezclas
complejas de proteínas, como son células lisadas y extractos de tejidos, para
observar los cambios cuantitativos de los niveles de expresión. El alcance de su
aplicación se encuentra desde el descubrimiento de fármacos, diagnósticos,
terapia, microbiología, bioquímica e investigación de plantas (Westermeier,
2002).
En la actualidad, la proteómica ha sido la herramienta más utilizada para
identificar el proteoma de diversos sistemas biológicos como plantas,
microorganismos y animales (Uvackova et al., 2013; Stekhoven et al., 2014).
En el campo de alimentos y salud la proteómica ha abierto los temas de
investigación en nutrigenómica o nutriproteómica. Recientemente se ha acuñado
el término “Alimentómica” (Foodomics) al empleo de las herramientas de
proteómica en la investigación de Ciencias de Alimentos y se ha descrito como
el estado-del-arte para el análisis específico y preciso de proteínas en alimentos
(Abu-Reidah et al., 2014; García-Cañas et al., 2012).
En la última década debido a la evolución de la espectrometría de masas
y la disponibilidad de la información genómica, los avances en la proteómica se
han obtenido en paralelo con: la tecnología de electroforesis de dos dimensiones
de alta resolución, con novedosas técnicas de ionización y de nuevos detectores
para espectrometría de masas. De igual forma el crecimiento de las bases de
datos de la secuencia del genoma y la bioinformación ha sido utilizada como
parte de la mejora de las herramientas para búsqueda de información
(Westermeier, 2002). A continuación se muestran las etapas más importantes
para llevar a cabo el estudio proteomico.

17
2.1.3.3 Extracción de proteínas
El estudio proteómico implica un control minucioso de la muestra para la
reproducibilidad de los resultados que se desea obtener. Un aspecto crítico del
estudio es la extracción y solubilización de las proteínas presentes en la
muestra, se debe extraer todas las clases de proteínas de forma reproducible
(Walker, 2005).
Para poder lograrlo es necesario utilizar varios cocteles de solubilización,
los cuales ya existen de forma comercial, pero no funcionan de forma única o
universal, ya que depende del tipo de muestra y los diferentes interferentes que
se puedan presentar, por lo que las proteínas pueden presentar diferentes
solubilidades. Además la presencia de proteínas muy abundantes complica la
extracción, solubilización y el análisis de las especies menos abundantes. Otro
factor importante que se debe visualizar es la compatibilidad de los agentes de
solubilización respecto a los métodos analíticos que se usaran (Walker, 2005).
Los cocteles de solubilización más eficientes que se han encontrado
consisten en una mezcla de agentes caotrópicos, detergentes que contengan
entre 13-15 cadenas largas de carbono hidrófobas y un reductor para
asegurarse de la extracción y solubilización completa de las proteínas presentes
en la muestra (Walker, 2005).
2.1.3.4 Electroforesis bidimensional
La electroforesis bidimensional es un proceso secuencial de dos
electroforesis en distintas condiciones que se aplican sobre una misma muestra.
En la primear dimensión se aplica una separación de muestra según un criterio,
el cual puede ser carga, tamaño o punto isoeléctrico y la segunda de acuerdo a
un parámetro distinto al anterior, carga, tamaño o punto isoeléctrico. Al combinar
dos modos de separación diferentes se consigue el máximo de resolución
posible mediante técnicas electroforéticas. En el caso de la primera dimensión

18
puede ser llevado a cabo en condiciones no desnaturalizantes y la segunda en
condiciones desnaturalizantes (García, 2008).
Lo más habitual para llevar a cabo la separación es que en la primera
dimensión se lleva a cabo por electroenfoque, es un tipo particular de
electroforesis en la que los compuestos anfóteros se separan según su punto
isoeléctrico (pH al cual su carga neta es nula) en un gradiente de pH continuo.
Mientras que en la segunda dimensión se obtiene por electroforesis llevándose a
cabo una separación por tamaño molecular. Una vez obtenido la última
separación se tiñe el gel obtenido y se pueden observar las manchas obtenidas
(García, 2008).
Estas técnicas acopladas se puede considerar como un criterio de pureza
positivo, debido a su gran poder de resolución, llegando a la aceptación de que
la aparición de una sola mancha indica una muestra homogénea (García, 2008)
Permitiendo observar un mapa 2-D de las proteínas presentes en una muestra
analizada, denominado mapa proteómico.
2.1.3.5 Identificación de péptidos mediante LC-MS/MS
Una vez teñido el gel se pueden observar las manchas que son
denominadas “spot”, las cuales pueden ser identificadas por espectrometría de
masas, lo cual se puede realizar con un spot que contenga de 10 a 20 ng de
proteína para una buena señal. El gel obtenido puede ser analizado mediante
software el cual permite asignarle un numero de spot y obtener una imagen para
su posterior análisis de cada spot obtenido (Westermeier, 2002).
El spot es cortado y se lleva por un proceso de digestión dentro del gel
con enzimas proteolíticas, que en su mayoría se usa tripsina. Los péptidos
obtenidos por este proceso son elucidados por espectrometría de masas. La
información de los diferentes péptidos obtenidos es almacenada y comparada
con bibliotecas donde se tienen ya proteínas secuenciadas. Con ello se conoce

19
la secuencia de péptidos que presenta cada proteína dentro de la muestra
analizada.
2.1.4 Lípidos y colesterol
Un lípido se define como un compuesto orgánico de origen natural que es
insoluble en agua y soluble en disolventes orgánicos no polares, tales como un
hidrocarburo o éter dietílico. Las distintas clases de lípidos se relacionan entre sí
por esta propiedad física; pero sus relaciones químicas, estructurales y
funcionales así como sus funciones biológicas, son distintas. La clase de
compuestos usualmente considerado como lípidos: son grasas, aceites,
terpenos, esteroides y otros compuestos (Fessenden, 1983).
2.1.4.1 Función de los lípidos y colesterol
Forman parte de la estructura de la membrana celular y de los sistemas
de transporte de los nutrimentos, son una fuente importante de energía en la
nutrición humana, promueven la síntesis de micelas y bilis, facilitan la absorción
de vitaminas liposolubles, funcionan como aislantes de impulsos eléctricos y
térmicos, al ser malos conductores de la temperatura la mantienen estable
dentro del organismo. En los alimentos son los responsables de las
características sensoriales y de nutrición. Son fuente precursoras de vitaminas
A, D, E, K.
El ácido linoleico componente de las acilglucoceramidas de la piel, el
inositol favorece la traducción de señales y se ha demostrado que los ácidos
grasos insaturados tienen beneficios en la salud del ser humano, ya que
disminuyen enfermedades cardiovasculares, conducen la disminución de
diabetes por la reducción de intolerancia a la glucosa y la prevención de
resistencia a la insulina (Yang et al., 2006).

20
Debido a ello es importante conocer lo ácidos grasos presentes en los
escamoles ya que permitiría conocer si además de ser nutritivos tienen
beneficios en la salud del ser humano.
Con respecto al colesterol, la cantidad de esta biomolécula presente en
los alimentos, es un tema importante dentro de la salud pública, ya que se
relaciona con la incidencia de aterosclerosis. La sociedad americana de la salud
recomienda la ingesta máxima de 300 mg por día para hombres y 225 mg para
mujeres, ya que funciona como precursor de la vitamina D, corticosteroides y
ácidos biliares (Mendoza Martínez, 2010).
En recientes años la micro extracción en fase sólida (SPME) ha sido
utilizada como una alternativa a la extracción con solventes para el estudio de la
composición de lípidos cuticulares de insectos, combinada con GC, GC-MS,
HPLC o LC-MS. Lo anterior debido a que se reduce el tiempo de preparación,
compra de solventes y disposición. La cromatografía de líquidos de alta
resolución es muy usada para resolver problemas analíticos, sin embargo en el
análisis de lípidos no ha ganado terreno debido a la naturaleza química de
estos. A menudo los lípidos cuticulares han sido determinados por cromatografía
en capa fina, ya que separa diferentes clases de lípidos, e isómeros de lípidos
simples. Sin embargo la herramienta principal y más utilizada para el análisis de
lípidos es la Cromatografía de gases y la cromatografía de gases-espectrometría
de masas ya que permite identificar ácidos grasos, alcoholes y aldehídos.
(Golebiowski, et al., 2011).

21
3. JUSTIFICACIÓN
A pesar de que los escamoles están contemplados como una de las
especies de insectos de mayor importancia en nuestro país debido a su
exquisitez culinaria, poca información científica existe acerca de este alimento.
De manera general se conoce que los escamoles poseen un alto
contenido de proteínas, y lípidos pero se desconoce de manera precisa que tipo
de proteínas y lípidos lo constituyen y de qué manera están involucradas en sus
diferentes procesos metabólicos.
Por ello en el presente trabajo de investigación se propone el uso de
tecnologías de la era post-genómica (proteómica) para elucidar el proteoma de
los escamoles y la cromatografía de gases masas para la determinación del
perfil de lípidos.
La identificación del proteoma permitió la generación de conocimientos
acerca de las propiedades nutritivas y/o funcionales de las proteínas presentes
en los escamoles.
De igual forma la identificación de lípidos permitió conocer el tipo de
ácidos grasos saturados e insaturados presentes en los escamoles, y determinar
si son una fuente importante de ácidos grasos esenciales para el cuerpo
humano.
Dichos conocimientos sentaran bases para entender su biología y las
diversas funciones que podría tener cada una de las proteínas y los ácidos
grasos que constituyen a los escamoles.

22
4. HIPÓTESIS
El conocimiento del proteoma y el perfil de ácidos grasos de los
escamoles, permitirán conocer de manera más específica las propiedades
nutricionales y funcionales de las proteínas y los lípidos así como su asociación
con los diferentes procesos biológicos.
5. OBJETIVOS
5.1 General
Identificar y caracterizar el perfil de proteínas (proteoma) y lípidos
presentes en los escamoles (Liometopum apiculatum Mayr).
5.2 Particulares
a) Obtener un mapa proteómico mediante electroforesis bidimensional.
b) Identificar las proteínas más abundantes del mapa proteómico obtenido
mediante LC-MS/MS.
c) Clasificar las proteínas en base a su función biológica.
d) Determinar el perfil de ácidos grasos.
e) Cuantificar los ácidos grasos más abundantes y el colesterol total.

23
6. METODOLOGÍA
6.1 Recolección de la muestra
Los escamoles fueron recolectados en diferentes nidos del ejido de
Pocitos, municipio de Charcas, San Luis Potosí, México. Los escamoles fueron
lavados con agua, mezclados y congelados en nitrógeno líquido; posteriormente
fueron molidos en un molino para granos de café (Braun, Naucalpan, México) el
polvo obtenido fue almacenado a -80°C hasta su análisis.
6.2 Análisis del proteoma por electroforesis en 2-D acoplado a
espectrometría de masas
6.2.1 Extracción y solubilización de las proteínas
5 gramos de escamoles molidos fueron centrifugados con acetona
absoluta fría durante 10 minutos a 13 000 x g a 4°C (Super T21; Sorvall, Kendro
Laboratory Products, Newton, CT, USA) para eliminar la grasa. El pellet
(residuo) se secó a vacío a 30°C por 10 minutos y se suspendió en 14 mL de
buffer de rehidratación (anexo 1), el cual contenía urea 8M, CHAPS 2% (p/v),
azul de bromofenol 0.002% (v/v), DTT (ditiotreitol) 1M, PMSF 200mM y EDTA
0.5M. Una agitación en vortex fue aplicada durante 2 minutos, se dejó reposar
en hielo por 5 minutos y al finalizar se sónico (20KHz a 30% de amplitud) por 3
minutos (GE-505, Ultrasonic Procesor, Sonics & material Inc., Newton, CT,
USA). Nuevamente la muestra fue centrifugada a las mismas condiciones
mencionadas anteriormente, el sobrenadante se filtró con miracloth y se
almacenó a -20°C por 10 minutos, para ser nuevamente centrifugada a 15 000 x
g a 4°C, el sobrenadante (2 mL) fue filtrado y precipitado con 30 mL de acetato
de amonio en metanol al 0.1M y se dejó en reposo durante la noche a -20°C.
Después de 30 minutos de centrifugación a 13 000 x g a 4°C el residuo fue
lavado una vez con metanol y 3 veces con acetona fría, se deshidrató a vacío a
30°C por 10 minutos y se suspendió en buffer de rehidratación.

24
La concentración de proteína fue determinada mediante el método de
Bradford (Kit Protein Assay, Bio-Rad, Hércules, CA, USA), a 595nm.
6.2.2 Electroforesis bidimensional
Para la obtención de la primera dimensión, las proteínas fueron
separadas por isoelectroenfoque, usando tiras de gradiente inmovilizado de pH
de 24 cm de largo (Bio-Rad) con rangos de pH 3-10 y 5-8, las cuales fueron
rehidratadas con 450 µL de solución de rehidratación (anexo 1), que contenía
2.25 mg de proteína y el 1% de anfolitos de 3-10 y 4-7 respectivamente. Este
proceso se llevó a cabo en una charola de rehidratación (Amersham, Pharmacia
Biotech, Uppsala, Suecia) a temperatura ambiente por 16 horas.
Posteriormente a la hidratación, la separación por isoelectroenfoque a 20
°C (Ettan IPGphor system GE healthcare, Piscataway, NJ, USA) a 50 mA
constante por tira con las siguientes condiciones: (I) un gradiente de 150V por 2
h., (II) un gradiente de 300V por 2 h., (III) un gradiente de 1000V por 2 h., (IV) un
gradiente de 3000V por 3 h., (V) un gradiente de 6000V por 3 h. y (VI) mantener
un gradiente de 6000V por 10 h.
La segunda dimensión fue obtenida separando las proteínas por peso
molecular, después de su separación las tiras fueron equilibradas en un buffer
de equilibrio (anexo 1) (50mM Tris-HCl pH 8.8, 8M urea, glicerol 30% v/v, SDS
2% w/v, azul de bromofenol 0.002%w/v) y 1% w/v de DTT. Las tiras fueron
trasferidas a los geles de SDS-gel poliacrilamida verticales al 13% y fueron
selladas con agarosa. La separación se realizó en una cámara EttanTM Daltix
Electrophoresis unit (GE Healthcare) a 12mA por gel. Los geles obtenidos fueron
teñidos con azul de coomassie a 2.2% (PhastGel-BlueR-350). Las imágenes
fueron obtenidas con un equipo Pharos FX Plus Molecular Imager (BioRad) y
analizadas con un software PDQuest Analysis Software v8.0 (Bio-Rad).

25
6.2.3 Digestión en gel y análisis por espectrometría de masas tándem (LC-
MS/MS)
80 spots de proteínas del pH de 3-10 y 300 spots del pH de 5-8 y fueron
seleccionadas. Las cuales fueron cortadas y lavada con agua Milli-Q, y el
desteñido fue realizado 2 veces con acetonitrilo al 50% durante 10 minutos. Un
segundo lavado fue realizado con acetonitrilo al 100% por 10 minutos y
finalmente con bicarbonato de amonio (NH4HCO3) al 25nM (anexo 1) por 15
minutos. Una reducción con DTT 10mM en 25mM de NH4HCO3, fue aplicada,
seguido de una alquilación con iodoacetamida. La digestión con tripsina (anexo
1) (Promega, Madison, WI, USA) fue realizada a 37°C durante toda la noche.
Los fragmentos (péptidos) fueron extraídos con agua Milli-Q y 0.1% de
ácido fórmico hasta obtener una concentración de 10 a 15µL. Posteriormente
fueron separados por nanoACQUITY UPLC System (Waters, Milford, MA, USA)
y analizados por espectrometría de masas (LC-MS/MS) en un SYNAPT HDMS
Q-TOF (Waters). Los datos fueron colectados en modo Data Dependent
Acquisition mode (DDA). En modo de baja energía, los datos fueron
recolectados en energía de colisión constante de 3eV y en modo elevado, la
energía de colisión fue aumentando de 15 a 45 eV durante 8s de integración.
6.2.4 Búsqueda en base de datos e identificación de las proteínas
Con los datos obtenidos de la espectrometría de masas se identificaron
las proteínas mediante el servidor MASCOT (Matrix Sciene,
http://www.matrixscience.com/). La búsqueda se corrió con la base de datos de
NCBI (National Center for Biotechnology Information) , para la orden de las
Hymenopteras.

26
6.3 Análisis de ácidos grasos por cromatogafía de gases-masas (GC-MS)
6.3.1 Extracción de lípidos totales
50 mg de muestra fresca fue utilizada para la extracción de lípidos con
1mL de isooctano (2,2,4–trimetilpentano grado HPLC) usando un equipo de
microondas CEM discover por 10 minutos, bajo las siguientes condiciones:
temperatura de 100°C, una radiación de 150 watts y una presión de 290 psi. La
fase acuosa fue lavada con isooctano, la fase orgánica fue recuperada y secada
con sulfato de sodio anhidro en exceso.
6.3.2 Transesterificación de glicéridos
La fase orgánica recuperada fue derivatizada usando un medio básico
con 0.8mL de una solución de hidróxido de sodio en metanol al 1N (anexo 2), la
mezcla fue calentada a 100°C en un equipo de microondas CEM Discover, bajo
las siguientes condiciones: una radiación de 150 watts, una presión de 290 psi y
un tiempo de 10 minutos, para la derivatización se usó 1.0 mL de trifluoruro de
boro en metanol 14% calentando nuevamente en microondas bajo las mismas
condiciones anteriores, al final se recupera la fase orgánica, inyectando al
cromatógrafo de gases 0.6 µL de esta fase orgánica.
6.3.3 Análisis de ácidos grasos por GC-MS
El análisis de composición de ácidos grasos de los escalomes se llevó a
cabo en un cromatógrafo de gases Agilent Technology modelo 6890N acoplado
a un detector selectivo de masas modelo 5973. Se utilizó una columna capilar
Stabilwax (crossbond carbowax-PEG) de 30m de largo, 0.25 mm de diámetro
interno y 0.25 µm de tamaño de partícula. La temperatura del puerto de
inyección se mantuvo en 250°C, en modo splitless, temperatura del horno de
100°C. El programa usado fue el siguiente: temperatura inicial de 100°C, con un

27
incremento de 15°C/min., hasta 250°C por 5 minutos, con un tiempo de corrida
total de 14 minutos.
Los espectros obtenidos fueron colectados a 71 eV de voltaje de
ionización y el rango de masa analizada fue de 33-800m/z. Los ácidos grasos
fueron identificados como esteres de metilo (FAMEs) usando la librería NIST
11/Wiley 09 y por comparación de los tiempos de retención con ácidos grasos
(de la marca Sigma Aldrich) de referencia previamente esterificados.
6.3.4 Cuantificación de ácidos grasos
La cuantificación se llevó a cabo usando curvas de calibración (anexo 2)
para cada derivado de ácido graso en concentraciones de 10-1000 ppm. Dichas
soluciones fueron tratadas de igual manera que la muestra analizada generando
ácidos grasos de esteres de metilo (FAMEs), en todos los casos se obtuvieron
coeficiente de correlación mayor a 0.99.
6.4 Análisis de colesterol
6.4.1 Extracción de colesterol
50 mg de muestra fresca fueron utilizados para extraer el colesterol 1 mL
de cloroformo como solvente. Un equipo de microondas CEM Discover fue
utilizado bajo las condiciones siguientes: temperatura de 60°C, una radiación de
150 watts, una presión de 290 psi y tiempo de 10 min. Este proceso se realizó
dos veces más para asegurar la extracción de todo el colesterol presente en la
muestra.
6.4.2 Derivatización del colesterol
La fase orgánica recuperada fue derivatizada (silanización) usando 100
µL de N,O-Bis(trimethylsilyl)trifluoroacetamide ≥99%, al 10% de cloruro de
trimetil sililo (anexo 2), se calentó a 60°C en un equipo de microondas CEM

28
Discover, bajo las siguientes condiciones: una radiación de 150 watts, una
presión de 290 psi por un tiempo de 10 minutos. 0.6 µL de la fase orgánica
recuperada fue inyectada al cromatógrafo de gases.
6.4.3 Análisis de colesterol por GC-MS
El análisis de colesterol de los escalomes se llevó a cabo en un
cromatógrafo de gases Agilent Technology modelo 6890N acoplado a un
detector selectivo de masas modelo 5973. Se utilizó una columna capilar DB-
5HT de 15 m de largo, 0.25 mm de diámetro interno y 0.10 µm de tamaño de
partícula. La temperatura del puerto de inyección se mantuvo en 250°C, en
modo splitless y temperatura del horno de 100°C, el programa usado fue el
siguiente: temperatura inicial de 200°C, con un incremento de 20°C/min., hasta
250°C por 2 minutos, con un tiempo de corrida total de 7 minutos.
El espectro obtenido fue colectado a 71 eV de voltaje de ionización y el
rango de masa analizada fue de 33-800m/z. El colesterol fue identificado como
su derivado de éter de trimetilsililo y por comparación con el tiempo de retención
del colesterol de referencia previamente derivatizado de la marca Sigma Aldrich.
6.4.4 Cuantificación de colesterol
La cuantificación de colesterol se llevó a cabo realizando una curva de
calibración (anexo 2) de 1 a 150 ppm de colesterol de referencia al 98%. Cada
concentración fue silanizada de la misma manera que la muestra de escamoles,
el valor del coeficiente de correlación fue de 0.99.

29
7. RESULTADOS Y DISCUSION
7.1 Mapa proteómico de la larva de Liometopum apiculatum Mayr
Un gel 2-DE acoplado con LC-MS/MS fue empleado para generar el mapa
proteómico de los escamoles, las proteínas fueron separadas por IEF (enfoque
isoeléctrico) realizado en tiras de gradiente inmovilizado de pH (IPG) con un
rango de pH de 3-10 (Figura 7A). En dicha Figura se observó que gran parte de
las proteínas fueron localizadas en un rango de pH de 5-8, por lo tanto, con la
finalidad de obtener mayor resolución de las proteínas más abundantes, tiras
IPG con un rango de pH de 5-8 fueron usadas (Figura 7B). El mapa proteómico
compuesto de Liometopum apiculatum Mayr (Figura 8) muestra la cantidad de
las proteínas que fueron analizadas por LC-MS/MS.
Figura 7. Mapa 2-DE de la larva Liometopum apiculatum Mayr. Las proteínas fueron separadas
en tiras IPG de 24 cm con pH de 3-10 (A) y 5-8 (B) y SDS-PAGE al 13%.

30
7.2 Identificación de proteínas de Liometopum apiculatum Mayr
El genoma de Liometopum apiculatum Mayr no está secuenciado,
solamente algunas proteínas (28) y secuencia de nucleótidos (38) del organismo
del genero Liometopum apiculatum Mayr han sido reportados en base de datos.
De 380 proteínas analizadas, únicamente 174 (46%) fueron identificadas
positivamente por búsqueda en base de datos, las proteínas fueron relacionadas
con organismos del orden Hymenoptera. En la Tabla 5, los spots fueron
agrupados de acuerdo a su función: estructura celular, trascripción y traducción,
metabolismo y producción de energía, señalización celular, actividad
antioxidante trasporte y proceso de óxido reducción.
En 23 casos, se identificaron más de 1 proteína por spot. El número de
péptidos similares a las proteínas pertenecientes a la familia Formicidae fue en
general mucho mejor que las proteínas de otros miembros de la orden
Hymenoptera. Sin embargo, en algunos casos la mejor correlación fue la
comparación con el orden Hymenoptera.
Figura 8. Mapa proteómico de la larva Liometopum apiculatum Mayr.
31
En adición a la identificación de proteínas por LC-MS/MS, se utilizó el
paquete bioinformático Gene Ontology (GO) con el cual se puede unificar los
genes de todas las especies y así poder entender mejor su localización celular,
su función molecular y los procesos biológicos en donde se encuentran
involucradas estas proteínas. De las 174 proteínas, solamente 134 fueron
clasificadas de acuerdo a la disponibilidad de la base de datos. Las proteínas
identificadas fueron clasificadas de acuerdo a los componentes celulares,
funciones moleculares y procesos biológicos, a continuación se describe cada
una de las funciones de las proteínas identificadas en el mapa proteómico de la
larva de la especie Liometopum apiculatum Mayr (Figura 9).
Figura 9. Clasificación de las proteínas identificadas de la larva Liometopum apiculatum Mayr por
LC-MS/MS, de acuerdo a la anotación del paquete bioinformático Gene Ontology.

32
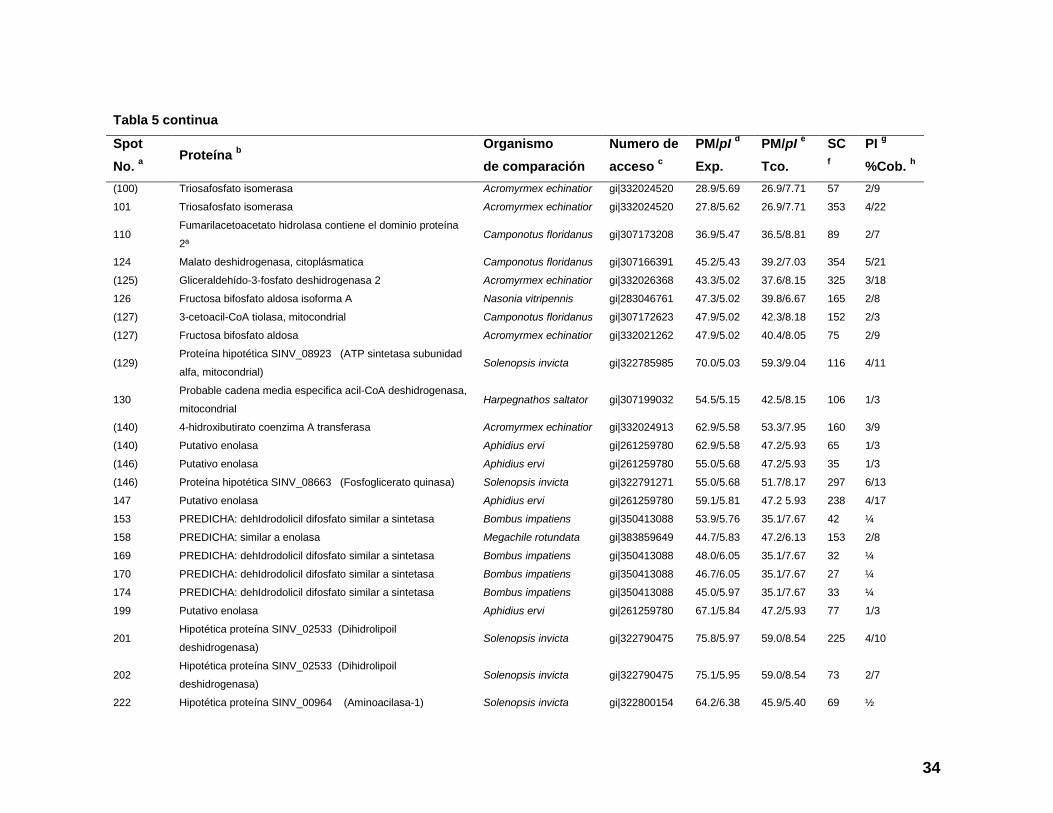
Tabla 5. Composición proteica de los escamoles, spots de proteínas analizadas por LC-MS/MS e identificadas contra el orden
Hymenoptera de la base de datos de proteínas de NCBI.
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
Estructura celular
(24) Miofilina Harpegnathos saltator gi|307212199 25.9/4.24 15.6/9.05 102 1/12
177 Estatmina-4 Acromyrmex echinatior gi|332018661 41.2/5.63 34.9/9.44 277 4/13
198 PREDICHA: kinesina 6ª Apis mellifera gi|328782772 96.6/5.90 97.5/9.17 29 1/1
214 Proteína de interacción con actina 1 Acromyrmex echinatior gi|332023261 97.9/6.10 70.2/6.32 336 4/10
216 PREDICHA: kinesina 6ª Apis mellifera gi|328782772 82.2/6.12 97.5/9.17 31 1/1
234 Timosina beta-4 Camponotus floridanus gi|307189411 22.6/6.43 18.4/5.90 121 3/17
236 Proteína similar al factor despolimerizante cofilina/actina Camponotus floridanus gi|307187751 17.1/6.40 19.6/8.50 136 4/27
254 Timosina beta-4 Camponotus floridanus gi|307189411 15.7/7.21 18.4/5.90 138 4/28
256 Profilina Camponotus floridanus gi|307173635 12.0/7.04 13.9/5.65 140 2/20
257 Profilina Acromyrmex echinatior gi|332028899 11.1/7.02 20.3/9.54 44 1/12
271 Proteína similar al factor despolimerizante cofilina/actina Camponotus floridanus gi|307187751 17.7/6.90 19.6/8.50 291 4/30
305 Tropomiosina 1 isoforma B Nasonia vitripennis gi|229577296 45.6/7.71 32.7/4.73 359 3/10
323 Actina, clon 403 Harpegnathos saltator gi|307195447 59.8/7.15 42.3/5.37 45 ½
324 Actina, clon 403 Acromyrmex echinatior gi|332024486 56.8/7.15 42./5.29 39 2/8
372 Tropomiosina-1 Harpegnathos saltator gi|307196973 45.2/8.00 32.3/4.72 262 4/18
Transcripción y traducción
56 60S Proteína ribosomal ácida P1 Harpegnathos saltator gi|307212258 16.3/9.65 13.3/4.53 96 4/46
58 Proteína hipotética SINV_06628 (Similar a la proteína 60S
ribosomal ácida P2) Solenopsis invicta gi|322795701 14.9/8.29 11.9/4.77 120 3/31
62 Factor de elongación 1-beta' Camponotus floridanus gi|307170891 30.7/8.32 24.3/4.63 65 1/6
63 Factor de elongación 1-beta' Camponotus floridanus gi|307170891 30.1/8.29 24.3/4.63 76 2/9
64 Factor de elongación 1-beta' Harpegnathos saltator gi|307215145 31.4/8.40 24.3/4.72 70 2/7
66 Proteína hipotética SINV_02215 (Proteína que contiene
bromodominio 8) Solenopsis invicta gi|322799152 64.0/8.73 125/4.81 30 1/1
68 Proteína 1 homólogo mitocondrial ribonucleasa P Nasonia vitripenis gi|156551453 66.9/8.24 52.7/9.25 39 ½
237 Factor de empalme, arginina/serina-rica 7 Camponotus floridanus gi|307180295 17.5/6.32 17.8/8.73 94 2/13

33
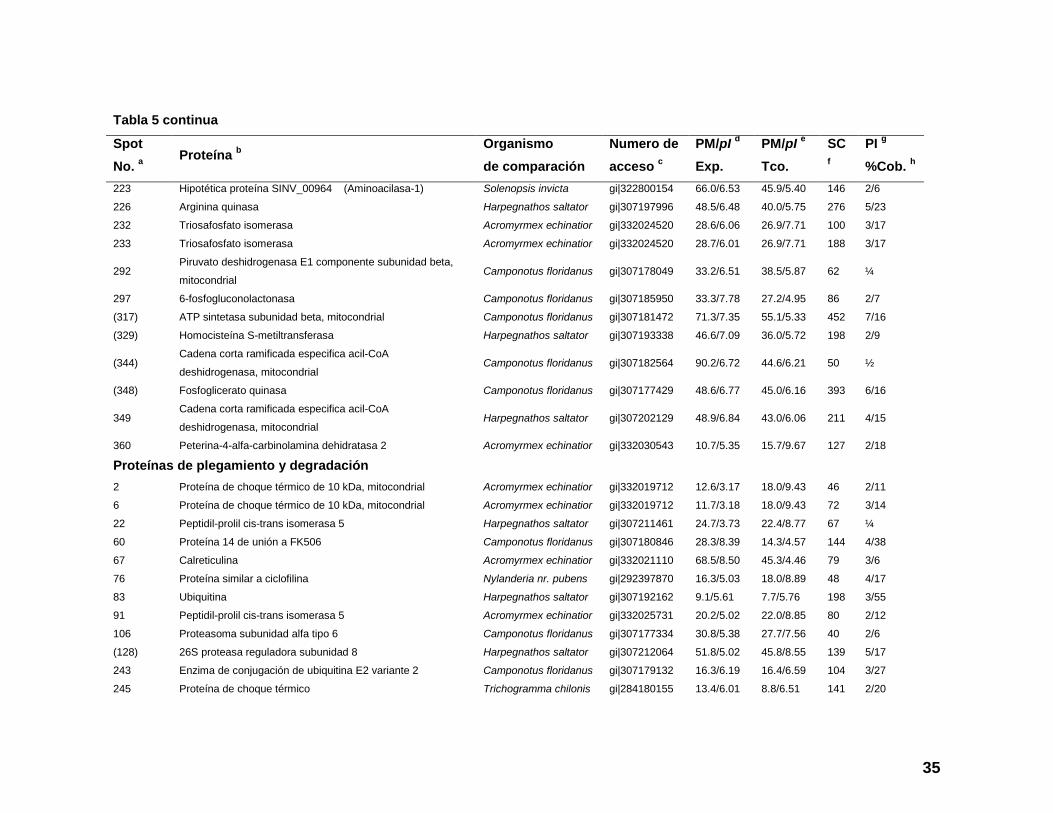
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
255 Proteína S12 ribosomal 40S Camponotus floridanus gi|307187759 15.1/7.27 15.6/5.47 86 2/19
274 Traducción eucariótica factor de iniciación 5ª Camponotus floridanus gi|307180081 18.8/7.47 18.05/07 97 2/14
327 Factor de activación upstream subunidad spp27 Acromyrmex echinatior gi|332018104 45.4/7.28 37.2/8.91 87 2/8
328 Factor de activación upstream subunidad UAF30 Harpegnathos saltator gi|307214555 45.5/7.20 34.3/8.42 43 3/8
(329) Proteínas activadora de la transcripción Pur-alpha Camponotus floridanus gi|307190558 46.6/7.09 33.7/6.86 214 5/21
331 Factor de elongación factor 2 Camponotus floridanus gi|307170298 49.6/6.98 94.3/6.11 164 4/6
345 Factor de elongación 1 alpha Bombus mendax gi|27501884 39.3/6.76 42.5/8.42 58 2/2
346 Factor de elongación 1 alpha Bombus mendax gi|27501884 42.1/6.52 42.5/8.42 41 2/2
Metabolismo y producción energía
17 Nucleósido difosfato quinasa Harpegnathos saltator gi|307193761 18.4/4.88 17.4/7.79 67 1/7
26 3-hidroxiacil-CoA deshidrogenasa tipo 2 Acromyrmex echinatior gi|332024263 35.5/3.05 30.3/8.70 113 6/13
(27) Fosfoglicerato mutasa 1 Harpegnathos saltator gi|307203820 34.5/4.78 28.7/7.77 133 7/27
(33) Gliceraldehído-3-fosfato deshidrogenasa 2 Acromyrmex echinatior gi|332026368 47.6/3.89 37.6/8.15 39 5/10
36 Gliceraldehído-3-fosfato deshidrogenasa 2 Acromyrmex echinatior gi|332026368 48.8/4.33 37.6/8.15 97 8/30
(38) Proteína hipotética SINV_01281 ( Gliceraldehído-3-fosfato
deshidrogenasa ) Solenopsis invicta gi|322794747 48.9/4.21 74.1/6.72 151 6/9
(39) Proteína hipotética SINV_01281 ( Gliceraldehído-3-fosfato
deshidrogenasa ) Solenopsis invicta gi|322794747 48.7/4.08 74.1/6.72 146 6/9
42 Nucleósido difosfato quinasa Camponotus floridanus gi|307173082 18.6/4.76 19.6/8.41 29 ¼
(45) Proteína hipotética SINV_11660 (Proteína de biosíntesis de
purinas bifuncional PURH) Solenopsis invicta gi|322795791 93.9/5.02 65.2/7.58 62 2/4
46 Proteína hipotética SINV_11660 (Proteína de biosíntesis de
purinas bifuncional PURH) Solenopsis invicta gi|322795791 93.0/4.96 65.2/7.58 46 2/4
77 Nucleósido difosfato quinasa Harpegnathos saltator gi|307193761 17.0/5.02 17.4/7.79 45 2/13
79 Pterin-4-alfa-carbinolamina dehidratasa 2 Acromyrmex echinatior gi|332030543 10.6/5.43 15.7/9.67 34 4/17
81 Pterin-4-alfa-carbinolamina dehidratasa 2 Harpegnathos saltator gi|307206440 10.5/5.61 15.5/9.72 100 1/11
94 Fosfoglicerato mutasa 1 Harpegnathos saltator gi|307203820 31.4/5.01 28.7/7.77 150 4/16
(96) 3-hidroxiacil-CoA deshidrogenasa tipo-2 Acromyrmex echinatior gi|332024263 33.9/5.01 30.3/8.70 64 ¼
34
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
(100) Triosafosfato isomerasa Acromyrmex echinatior gi|332024520 28.9/5.69 26.9/7.71 57 2/9
101 Triosafosfato isomerasa Acromyrmex echinatior gi|332024520 27.8/5.62 26.9/7.71 353 4/22
110 Fumarilacetoacetato hidrolasa contiene el dominio proteína
2ª Camponotus floridanus gi|307173208 36.9/5.47 36.5/8.81 89 2/7
124 Malato deshidrogenasa, citoplásmatica Camponotus floridanus gi|307166391 45.2/5.43 39.2/7.03 354 5/21
(125) Gliceraldehído-3-fosfato deshidrogenasa 2 Acromyrmex echinatior gi|332026368 43.3/5.02 37.6/8.15 325 3/18
126 Fructosa bifosfato aldosa isoforma A Nasonia vitripennis gi|283046761 47.3/5.02 39.8/6.67 165 2/8
(127) 3-cetoacil-CoA tiolasa, mitocondrial Camponotus floridanus gi|307172623 47.9/5.02 42.3/8.18 152 2/3
(127) Fructosa bifosfato aldosa Acromyrmex echinatior gi|332021262 47.9/5.02 40.4/8.05 75 2/9
(129) Proteína hipotética SINV_08923 (ATP sintetasa subunidad
alfa, mitocondrial) Solenopsis invicta gi|322785985 70.0/5.03 59.3/9.04 116 4/11
130 Probable cadena media especifica acil-CoA deshidrogenasa,
mitocondrial Harpegnathos saltator gi|307199032 54.5/5.15 42.5/8.15 106 1/3
(140) 4-hidroxibutirato coenzima A transferasa Acromyrmex echinatior gi|332024913 62.9/5.58 53.3/7.95 160 3/9
(140) Putativo enolasa Aphidius ervi gi|261259780 62.9/5.58 47.2/5.93 65 1/3
(146) Putativo enolasa Aphidius ervi gi|261259780 55.0/5.68 47.2/5.93 35 1/3
(146) Proteína hipotética SINV_08663 (Fosfoglicerato quinasa) Solenopsis invicta gi|322791271 55.0/5.68 51.7/8.17 297 6/13
147 Putativo enolasa Aphidius ervi gi|261259780 59.1/5.81 47.2 5.93 238 4/17
153 PREDICHA: dehIdrodolicil difosfato similar a sintetasa Bombus impatiens gi|350413088 53.9/5.76 35.1/7.67 42 ¼
158 PREDICHA: similar a enolasa Megachile rotundata gi|383859649 44.7/5.83 47.2/6.13 153 2/8
169 PREDICHA: dehIdrodolicil difosfato similar a sintetasa Bombus impatiens gi|350413088 48.0/6.05 35.1/7.67 32 ¼ 170 PREDICHA: dehIdrodolicil difosfato similar a sintetasa Bombus impatiens gi|350413088 46.7/6.05 35.1/7.67 27 ¼ 174 PREDICHA: dehIdrodolicil difosfato similar a sintetasa Bombus impatiens gi|350413088 45.0/5.97 35.1/7.67 33 ¼ 199 Putativo enolasa Aphidius ervi gi|261259780 67.1/5.84 47.2/5.93 77 1/3
201 Hipotética proteína SINV_02533 (Dihidrolipoil
deshidrogenasa) Solenopsis invicta gi|322790475 75.8/5.97 59.0/8.54 225 4/10
202 Hipotética proteína SINV_02533 (Dihidrolipoil
deshidrogenasa) Solenopsis invicta gi|322790475 75.1/5.95 59.0/8.54 73 2/7
222 Hipotética proteína SINV_00964 (Aminoacilasa-1) Solenopsis invicta gi|322800154 64.2/6.38 45.9/5.40 69 ½
35
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
223 Hipotética proteína SINV_00964 (Aminoacilasa-1) Solenopsis invicta gi|322800154 66.0/6.53 45.9/5.40 146 2/6
226 Arginina quinasa Harpegnathos saltator gi|307197996 48.5/6.48 40.0/5.75 276 5/23
232 Triosafosfato isomerasa Acromyrmex echinatior gi|332024520 28.6/6.06 26.9/7.71 100 3/17
233 Triosafosfato isomerasa Acromyrmex echinatior gi|332024520 28.7/6.01 26.9/7.71 188 3/17
292 Piruvato deshidrogenasa E1 componente subunidad beta,
mitocondrial Camponotus floridanus gi|307178049 33.2/6.51 38.5/5.87 62 ¼
297 6-fosfogluconolactonasa Camponotus floridanus gi|307185950 33.3/7.78 27.2/4.95 86 2/7
(317) ATP sintetasa subunidad beta, mitocondrial Camponotus floridanus gi|307181472 71.3/7.35 55.1/5.33 452 7/16
(329) Homocisteína S-metiltransferasa Harpegnathos saltator gi|307193338 46.6/7.09 36.0/5.72 198 2/9
(344) Cadena corta ramificada especifica acil-CoA
deshidrogenasa, mitocondrial Camponotus floridanus gi|307182564 90.2/6.72 44.6/6.21 50 ½
(348) Fosfoglicerato quinasa Camponotus floridanus gi|307177429 48.6/6.77 45.0/6.16 393 6/16
349 Cadena corta ramificada especifica acil-CoA
deshidrogenasa, mitocondrial Harpegnathos saltator gi|307202129 48.9/6.84 43.0/6.06 211 4/15
360 Peterina-4-alfa-carbinolamina dehidratasa 2 Acromyrmex echinatior gi|332030543 10.7/5.35 15.7/9.67 127 2/18
Proteínas de plegamiento y degradación
2 Proteína de choque térmico de 10 kDa, mitocondrial Acromyrmex echinatior gi|332019712 12.6/3.17 18.0/9.43 46 2/11
6 Proteína de choque térmico de 10 kDa, mitocondrial Acromyrmex echinatior gi|332019712 11.7/3.18 18.0/9.43 72 3/14
22 Peptidil-prolil cis-trans isomerasa 5 Harpegnathos saltator gi|307211461 24.7/3.73 22.4/8.77 67 ¼
60 Proteína 14 de unión a FK506 Camponotus floridanus gi|307180846 28.3/8.39 14.3/4.57 144 4/38
67 Calreticulina Acromyrmex echinatior gi|332021110 68.5/8.50 45.3/4.46 79 3/6
76 Proteína similar a ciclofilina Nylanderia nr. pubens gi|292397870 16.3/5.03 18.0/8.89 48 4/17
83 Ubiquitina Harpegnathos saltator gi|307192162 9.1/5.61 7.7/5.76 198 3/55
91 Peptidil-prolil cis-trans isomerasa 5 Acromyrmex echinatior gi|332025731 20.2/5.02 22.0/8.85 80 2/12
106 Proteasoma subunidad alfa tipo 6 Camponotus floridanus gi|307177334 30.8/5.38 27.7/7.56 40 2/6
(128) 26S proteasa reguladora subunidad 8 Harpegnathos saltator gi|307212064 51.8/5.02 45.8/8.55 139 5/17
243 Enzima de conjugación de ubiquitina E2 variante 2 Camponotus floridanus gi|307179132 16.3/6.19 16.4/6.59 104 3/27
245 Proteína de choque térmico Trichogramma chilonis gi|284180155 13.4/6.01 8.8/6.51 141 2/20

36
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
282 Proteína hipotética SINV_11930 (cis-trans isomerasa
peptidil-prolil) Solenopsis invicta gi|322798830 22.2/7.91 19.3/4.69 111 3/23
286 Proteína de choque térmico beta-1 Camponotus floridanus gi|307183305 30.9/6.66 29.5/6.37 326 5/25
287 Proteína de choque térmico beta-1 Camponotus floridanus gi|307183305 32.6/6.72 29.5/6.37 42 2/11
311 26S proteasa reguladora subunidad 6ª Camponotus floridanus gi|307190289 69.7/7.55 47.9/4.98 567 10/34
312 Proteína de interacción con Hsc70 Harpegnathos saltator gi|307215258 66.0/7.73 31.3/4.56 206 3/14
313 PREDICHA: similar a la proteína de choque térmico 70 kDa
afín 3 Apis florea gi|380030032 107/7.45 106/5.71 427 7/8
314 PREDICHA: similar a la proteína de choque térmico 70 kDa
afín 3 Megachile rotundata gi|383848513 109/7.49 106/5.52 311 5/7
(317) 26S proteasa reguladora subunidad 6B Camponotus floridanus gi|307172856 71.3/7.35 45.8/5.16 48 ½
318 26S proteasa reguladora subunidad 6B Camponotus floridanus gi|307172856 74.1/7.28 45.8/5.16 263 5/16
319 Proteína hipotética SINV_14312 (60 kDa proteína de
choque térmico, mitocondrial) Solenopsis invicta gi|322787697 89.4/7.31 60.3/5.60 336 7/18
320 PREDICHA: similar a la proteína de choque térmico de 60
kDa, mitocondrial Apis mellifera gi|66547450 91.3/7.28 60.5/5.64 371 6/12
321 Proteína hipotética SINV_14312 (60 kDa proteína de
choque térmico, mitocondrial) Solenopsis invicta gi|322787697 90.0/7.20 60.3/5.60 305 6/16
322 Procesamiento mitocondrial peptidasa subunidad beta Harpegnathos saltator gi|307207091 67.2/7.16 53./5.77 355 6/11
338 Proteína de choque térmico 70 kDa afín 5 Camponotus floridanus gi|307181923 104/6.86 72.3/6.37 547 8/12
(342) Proteína de choque térmico 70 kDa afín 5 Camponotus floridanus gi|307176326 106/6.97 71.7/5.43 541 9/21
347 Proteasoma tipo 1 subunidad alfa Camponotus floridanus gi|307180286 42.0/6.57 31.1/5.91 196 4/15
363 Prefoldina subunidad 3 Camponotus floridanus gi|307172404 29.4/7.25 22.1/5.27 94 3/18
Señalización celular
55 Calmodulina Camponotus floridanus gi|307181039 14.8/9.62 17.7/4.14 94 3/25
(96) Canal de aniones selectivo dependiente de la tensión Harpegnathos saltator gi|307198439 33.9/5.01 30.9/8.21 75 1/3
(100) Proteína hipotética EAG_02089 (lectina tipo C) Camponotus floridanus gi|307166128 28.9/5.69 27.0/6.30 84 1/6
Actividad antioxidante

37
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
78 Proteína hipotética SINV_03933 (Peroxiredoxina 5,
mitocondrial) Solenopsis invicta gi|322796535 16.1/5.41 20.3/9.06 107 1/9
98 Mn superóxido dismutasa Apis mellifera ligustica gi|33089106 23.0/5.37 24.7/9.11 121 2/12
229 PREDICHA: similar peroxirredoxina 6 Bombus terrestris gi|340718481 29.6/6.32 25.4/5.46 56 1/6
247 Probable fosfolípidos hidroperóxido glutatión peroxidasa Harpegnathos saltator gi|307192508 18.8/5.55 19.3/6.96 140 2/15
253 2-cys peroxirredoxina Bombus ignitus gi|227976948 16.7/6.65 21.9/6.90 46 1/5
289 Proteína hipotética SINV_03768 (Peroxiredoxina 6) Solenopsis invicta gi|322797657 31.2/6.57 26.5/5.98 64 ¼
290 PREDICHA: similar peroxirredoxina 6 Bombus terrestris gi|340718481 30.5/6.54 25.4/5.46 81 1/6
364 Fosfolípido hidroperóxido glutatión peroxidasa, mitocondrial Acromyrmex echinatior gi|332027619 24.7/7.80 38.2/6.54 46 2/11
369 Peroxirredoxina 1 Acromyrmex echinatior gi|332021321 28.9/5.06 21.8/5.94 116 1/7
Transporte
34 Similar a PREDICHA: similar a la proteína mielina P2 Megachile rotundata gi|383853580 14.8/5.03 15.2/6.37 136 2/10
(75) Proteína de unión a ácidos grasos, músculo Acromyrmex echinatior gi|332024570 14.4/5.06 15.6/6.43 52 2/16
(75) Proteína mielina P2 Harpegnathos saltator gi|307206040 14.4/5.06 19.8/6.64 138 5/16
105 Trasferencia de electrones flavoproteína subunidad beta Acromyrmex echinatior gi|332025657 31.8/5.48 27.7/6.92 44 ¼
107 Trasferencia de electrones flavoproteína subunidad beta Acromyrmex echinatior gi|332025657 32.0/5.35 27.7/6.92 86 2/10
183 Trasferencia de electrones flavoproteína subunidad alfa,
mitocondrial Camponotus floridanus gi|307181673 37.0/5.15 35.7/8.97 50 1/3
249 Proteína mielina P2 Harpegnathos saltator gi|307206040 14.8/6.66 19.7/6.64 145 3/9
Procesos de óxido-reducción
(128) Proteína hipotética SINV_12515 (proteína similar
hidroxisteroide deshidrogenasa 2) Solenopsis invicta gi|322801306 51.8/5.02 42.9/8.83 95 ¼
315 Proteína disulfuro isomerasa Harpegnathos saltator gi|307207787 80.7/7.84 56.3/4.73 249 3/6
316 Proteína disulfuro isomerasa Harpegnathos saltator gi|307207787 79.7/7.86 56.3/4.73 193 4/6
333 Proteína disulfuro isomerasa A6 Camponotus floridanus gi|307167982 69.3/6.99 48.6/5.14 185 4/8
336 Proteína disulfuro isomerasa A3 Acromyrmex echinatior gi|332023180 92.5/6.79 56.1/5.66 184 5/9
(348) Retinal deshidrogenasa 1 Camponotus floridanus gi|307178537 48.6/6.77 53.5/5.46 64 ½
(350) Retinal deshidrogenasa 1 Camponotus floridanus gi|307178537 51.3/6.88 53.5/5.46 48 2/3

38
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
351 Hemocianin Acromyrmex echinatior gi|332025198 66.5/6.15 85.2/6.85 51 1/1
Otros procesos
23 Homólogo de la proteína de unión a fosfatidiletanolamina Bombus impatiens gi|350403780 24.6/4.02 23.1/8.85 82 1/6
(27) Proteína de la cutícula 21 Camponotus floridanus gi|307175910 34.5/4.78 26.1/6.20 59 2/9
(27) PREDICHA: proteína de la cutícula Nasonia vitripennis gi|156548084 34.5/4.78 35.0/7.29 32 1/3
(32) Poli(A) especifico ribonucleasa PARN Camponotus floridanus gi|307167442 54.9/3.29 65.5/6.35 41 1/1
(45) Proteína no especifica de trasferencia de lípidos Harpegnathos saltator gi|307194938 93.9/5.02 60.0/8.66 38 2/4
51 Proteína de la cutícula flexible 12 Acromyrmex echinatior gi|332023422 11.4/8.51 11.3/4.50 59 1/12
59 Miosina de cadena corta alcalina Harpegnathos saltator gi|307208335 16.9/8.57 17.3/4.57 152 4/37
65 Proteína hipotética SINV_02991 (Calumenin-B) Solenopsis invicta gi|322796781 51.4/8.51 40.0/4.55 79 3/10
70 PREDICHA: similar a hexamerin Bombus terrestris gi|340721852 143/8.4 120/6.19 53 1/1
(99) Proteína de la cutícula 21 Camponotus floridanus gi|307175910 28.7/5.77 26.1/6.20 42 1/3
(129) PREDICHA: proteína similar Idgf4 similar quitinasa Apis mellifera gi|66514614 70.0/5.03 49.0/8.06 116 2/6
135 PREDICHA: proteína similar Idgf4 similar quitinasa Apis mellifera gi|66514614 75.0/5.37 49.0/8.06 94 1/3
152 Sin caracterizar glicosiltransferasa AER61 Acromyrmex echinatior gi|332019577 55.3/5.77 45.8/8.51 28 1/3
230 Proteína de unión a fosfatidiletanolamina 1 Acromyrmex echinatior gi|332028806 26.5/6.17 13.8/9.36 69 3/41
295 Subunidad ferritina Acromyrmex echinatior gi|332027165 35.9/7.08 26.0/5.66 60 3/14
299 Proteína similar a nucleoplasmina Acromyrmex echinatior gi|332017234 29.9/7.91 24.2/4.67 49 ¼
302 Proteoglicanos de condroitín 2 Harpegnathos saltator gi|307212770 41.0/7.90 29.7/5.20 103 2/7
330 Proteína hipotética SINV_15827 (Regucalcina) Solenopsis invicta gi|322800197 47.5/7.06 42.7/5.35 50 1/3
334 Proteína de unión a selenio 1-A Acromyrmex echinatior gi|332021867 88.1/6.82 52.7/6.99 150 2/6
335 Proteína de unión a selenio 1-A Acromyrmex echinatior gi|332021867 89.4/6.92 52.7/6.99 161 3/9
337 Proteína de unión a selenio 1-A Camponotus floridanus gi|307191199 91.0/7.01 52.5/5.49 83 2/5
339 Transferrina Acromyrmex echinatior gi|332029256 120/6.82 79.4/5.47 185 4/6
340 Transferrina Solenopsis invicta gi|62912066 125/6.74 78.6/5.66 174 4/6
341 Transferrina Acromyrmex echinatior gi|332029256 123/6.89 79.4/5.47 172 4/5
(342) Proteína hipotética SINV_08631 (Hexamerina) Solenopsis invicta gi|322781221 106/6.97 103/6.66 36 1/1
343 Transferrina Solenopsis invicta gi|62912066 124/6.96 78.6/5.66 139 3/5
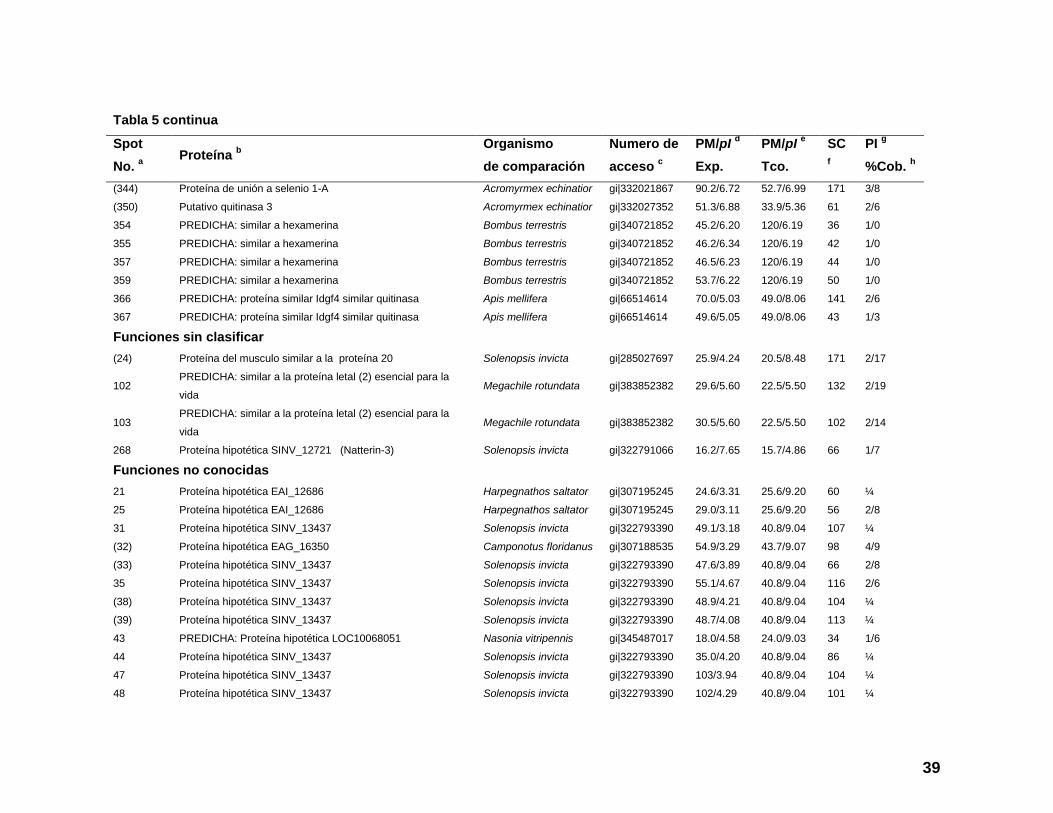
39
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
(344) Proteína de unión a selenio 1-A Acromyrmex echinatior gi|332021867 90.2/6.72 52.7/6.99 171 3/8
(350) Putativo quitinasa 3 Acromyrmex echinatior gi|332027352 51.3/6.88 33.9/5.36 61 2/6
354 PREDICHA: similar a hexamerina Bombus terrestris gi|340721852 45.2/6.20 120/6.19 36 1/0
355 PREDICHA: similar a hexamerina Bombus terrestris gi|340721852 46.2/6.34 120/6.19 42 1/0
357 PREDICHA: similar a hexamerina Bombus terrestris gi|340721852 46.5/6.23 120/6.19 44 1/0
359 PREDICHA: similar a hexamerina Bombus terrestris gi|340721852 53.7/6.22 120/6.19 50 1/0
366 PREDICHA: proteína similar Idgf4 similar quitinasa Apis mellifera gi|66514614 70.0/5.03 49.0/8.06 141 2/6
367 PREDICHA: proteína similar Idgf4 similar quitinasa Apis mellifera gi|66514614 49.6/5.05 49.0/8.06 43 1/3
Funciones sin clasificar
(24) Proteína del musculo similar a la proteína 20 Solenopsis invicta gi|285027697 25.9/4.24 20.5/8.48 171 2/17
102 PREDICHA: similar a la proteína letal (2) esencial para la
vida Megachile rotundata gi|383852382 29.6/5.60 22.5/5.50 132 2/19
103 PREDICHA: similar a la proteína letal (2) esencial para la
vida Megachile rotundata gi|383852382 30.5/5.60 22.5/5.50 102 2/14
268 Proteína hipotética SINV_12721 (Natterin-3) Solenopsis invicta gi|322791066 16.2/7.65 15.7/4.86 66 1/7
Funciones no conocidas
21 Proteína hipotética EAI_12686 Harpegnathos saltator gi|307195245 24.6/3.31 25.6/9.20 60 ¼
25 Proteína hipotética EAI_12686 Harpegnathos saltator gi|307195245 29.0/3.11 25.6/9.20 56 2/8
31 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 49.1/3.18 40.8/9.04 107 ¼
(32) Proteína hipotética EAG_16350 Camponotus floridanus gi|307188535 54.9/3.29 43.7/9.07 98 4/9
(33) Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 47.6/3.89 40.8/9.04 66 2/8
35 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 55.1/4.67 40.8/9.04 116 2/6
(38) Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 48.9/4.21 40.8/9.04 104 ¼
(39) Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 48.7/4.08 40.8/9.04 113 ¼
43 PREDICHA: Proteína hipotética LOC10068051 Nasonia vitripennis gi|345487017 18.0/4.58 24.0/9.03 34 1/6
44 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 35.0/4.20 40.8/9.04 86 ¼
47 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 103/3.94 40.8/9.04 104 ¼
48 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 102/4.29 40.8/9.04 101 ¼
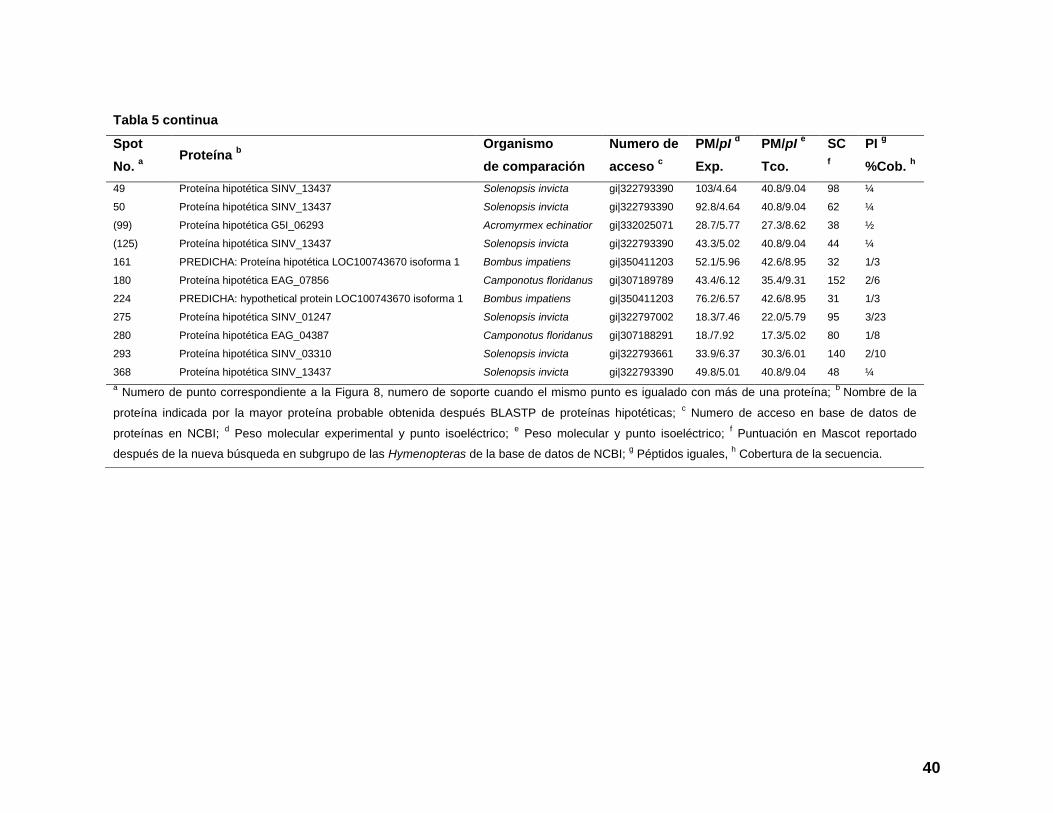
40
Tabla 5 continua
Spot
No. a
Proteína b
Organismo
de comparación
Numero de
acceso c
PM/pI d
Exp.
PM/pI e
Tco.
SC
f
PI g
%Cob. h
49 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 103/4.64 40.8/9.04 98 ¼
50 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 92.8/4.64 40.8/9.04 62 ¼
(99) Proteína hipotética G5I_06293 Acromyrmex echinatior gi|332025071 28.7/5.77 27.3/8.62 38 ½
(125) Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 43.3/5.02 40.8/9.04 44 ¼
161 PREDICHA: Proteína hipotética LOC100743670 isoforma 1 Bombus impatiens gi|350411203 52.1/5.96 42.6/8.95 32 1/3
180 Proteína hipotética EAG_07856 Camponotus floridanus gi|307189789 43.4/6.12 35.4/9.31 152 2/6
224 PREDICHA: hypothetical protein LOC100743670 isoforma 1 Bombus impatiens gi|350411203 76.2/6.57 42.6/8.95 31 1/3
275 Proteína hipotética SINV_01247 Solenopsis invicta gi|322797002 18.3/7.46 22.0/5.79 95 3/23
280 Proteína hipotética EAG_04387 Camponotus floridanus gi|307188291 18./7.92 17.3/5.02 80 1/8
293 Proteína hipotética SINV_03310 Solenopsis invicta gi|322793661 33.9/6.37 30.3/6.01 140 2/10
368 Proteína hipotética SINV_13437 Solenopsis invicta gi|322793390 49.8/5.01 40.8/9.04 48 ¼
a Numero de punto correspondiente a la Figura 8, numero de soporte cuando el mismo punto es igualado con más de una proteína;
b Nombre de la
proteína indicada por la mayor proteína probable obtenida después BLASTP de proteínas hipotéticas; c Numero de acceso en base de datos de
proteínas en NCBI; d Peso molecular experimental y punto isoeléctrico;
e Peso molecular y punto isoeléctrico;
f Puntuación en Mascot reportado
después de la nueva búsqueda en subgrupo de las Hymenopteras de la base de datos de NCBI; g Péptidos iguales,
h Cobertura de la secuencia.

41
7.3 Clasificación de proteínas de Liometopum apiculatum Mayr
7.3.1 Componentes celulares
7.3.1.1 Estructura celular
Miofilina (spot 24) es una proteína que mostró una homología significativa
con calponinas, las cuales son filamentos delgados asociados a proteínas que
están implicadas en la regulación de la contracción del músculo liso y
organización citoesquelética. El dominio N terminal de calponinas tiene
homología con α actina, que es la responsable con la interacción con F-actina,
tropomiosina y calmodulina (Horowitz et al., 1996).
Stathmina (spot 177) es una fosfoproteína reguladora citosólica ubicua
cuya actividad es regular por fosforilación en respuesta a la señalización o a las
fases del ciclo celular, está enlazada a los micro túbulos y promueve la
depolimerización secuestrando los dímeros de tubulina (Yip et al., 2014). En
Drosophila Stathmina es esencial para la migración de células germinales y la
formación del sistema nervioso en el embrión, también es importante en el
mantenimiento de los micro túbulos y la regulación del trasporte axonal (Duncan
et al., 2013)
Timosinas (spots 234 y 254) pertenecen a la familia de hormonas tímicas
que juegan un rol crucial en el desarrollo y mantenimiento de un sistema inmune
competente (Zhang et al., 2013). En humanos se ha demostrado que esta
proteína es un péptido antimicrobiano y posee aplicación farmacéutica en la
rehabilitación y renovación de los tejidos ulcerados (Goldstein et al., 2005).
Colifikin/actina-depolymerización factor-like proteínas (spots 236 y 271) es
una proteína ubicua expresada en la familia de F actina, puede ser encontrada
tanto en el reino animal como en el vegetal. Su actividad es fundamental para la
motilidad de actina inducida, a través de la aceleración de filamentos de actina
(Gurniak, Perlas y Witke, 2005).
Profilina (spot 256 y 267) es una proteína de unión a actina globular que
tiene varias funciones regulatorias en el autoensamble de filamentos de actina.
También vincula las vías de señalización de la actina en el citoesqueleto por

42
enlace a fosfoinosítidos y L-prolina en varias proteínas involucradas en la
cascada de señalización y transducción (Lu y Pollard, 2001).
Tropomiosinas (spot 305 y 372) son un grupo de proteínas con diferentes
isoformas identificadas en el músculo estriado y liso, y en células no musculares
de animales superiores e inferiores (Anyanful et al., 2001).
Actina (spot 323 y 324) es una proteína eucariota ubicua que tiene un rol
importante en diverso procesos fisiológicos como la contracción muscular,
transmisión citoplasmática, fagocitosis, movimientos morfogenéticos,
movimientos ameboides, citocinesis y mitosis.
Kinesina 6 (spot 192 y 216) es un motor molecular que es considerado
como transporte en organelos de membrana y filamento intermedio a lo largo de
los microtúbulos (Shimizu et al., 2000).
7.3.1.2 Regiones extracelulares
Las proteínas de cutícula fueron identificadas en diferentes puntos (spot
27, 51 y 99), estas proteínas y fibras de quitina son los constituyentes de la
matriz extracelular (Charles, 2010). Las principales funciones de la cutícula en
los insectos es darles forma, flexibilidad, fuerza, libre movimiento, defensa,
protección y asistencia en la homeostasis (Souza-Ferreira et al., 2014). Estas
funciones dependen de las proteínas presentes en la cutícula, una cutícula dura
es típica de larvas o adultos.
Los glucosaminoglicanos condoitrina sulfato fueron identificados en el
spot 302, son los principales componentes de los proteoglicanos en la superficie
de la célula y en la matriz extracelular. Esta proteína juega un rol importante en
el control de las cascadas de señalización, algunos trabajos sugieren que la
biosíntesis de condoitrina sulfato podría tener efectos benéficos en la
regeneración de enfermedades cardiovasculares (Prinz et al., 2014).

43
7.3.2 Funciones moleculares
7.3.2.1 Proteínas asociadas con trascripción y traducción
La metamorfosis en insectos es un fenómeno remarcable cuando la larva
sufre una reorganización morfológica para dar lugar al adulto. Tal trasformación
es posible debido a que las vías fisiológicas que controlan la metamorfosis
traducen la expresión del gen que promueve cambios a gran escala (Mitra,
2013). En Drosophila, probablemente el insecto mejor estudiado se ha
encontrado 872 genes que cambian su expresión durante la metamorfosis
(Pavlopoulos y Akam 2011). En L. apiculatum el mapa proteómico revela varias
proteínas relacionadas con la traducción y la trascripción tal como proteínas
ribosomales (spot 56, 58 y 255), factor de elongación (spot 62-64, 331, 345 y
346), factores de iniciación de la traducción (spot 274) así como subunidades
del factor de activación (spots 327 y 328) y la proteína activadora transcripcional
Pur-alpha (spot 329) fueron identificadas.
Proteínas que contienen bromodominios (spot 66) regula la estructura de
la cromatina, que es crucial en la regulación de algunos procesos fundamentales
de la célula como procesos funcionales cromosómicos: como la trascripción,
replicación de ADN, reparación y recombinación (Wang, et al., 2012). Factor de
empalme arginina/serina 7 (spot 237) puede modular el empalme en el sitio de
selección y jugar un rol importante en la regulación de los genes de expresión,
es conocido que este factor tiene implicaciones en neurodegeneración (van Abel
et al., 2011).
7.3.2.2 Actividad antioxidante
Peroxidorredoxinas (Prx) poseen tiorredoxinas o glutatión peroxidasa y
actividad como chaperón, de esta forma protege a las células del estrés
oxidativo (Ishii et al., 2012). Se ha reportado que en humanos peroxirredoxin-5
mitocondrial (PRX-5, spot 78) protege al ADN mitocondrial del ataque oxidativo
(Banmeyer et al., 2005).

44
PRX-6 (spot 229, 289 y 290) es el único miembro de esta familia que
presenta una sola actividad redox en la cisteína y utiliza peroxidasa de glutatión
para catalizar la reducción de H2O2 y puede reducir los fosfolípidos en
hidroperóxidos (Manevich y Fisher, 2005).
2-Cys peroxirredoxina (spot 253) son enzimas ubicuas que actúan como
peroxidasas (desintoxicación de peróxidos) o como holdasas (chaperonas
independientes de moléculas de ATP) (Soito et al., 2011).
PR-1(spot 369) es un miembro de la familia de 2-cys peroxirredoxinas, en
humanos se sabe que la PR-1 favorece la toxicidad de las células naturales
asesinas e inhibe la oncogenes c-myc y c-Abl (Wood et al., 2003).
Manganeso superóxido dismutasa (spot 98) es una reconocida enzima
que está involucrada en los procesos de estrés oxidativo (ROS) así también en
respuestas al estrés por calor, frío, hambre y metales pesados (Kim et al., 2010).
Fosfolípido hidroperóxido glutatión peroxidasa (spot 247 y 364), se
encuentra presente en varios tejidos de mamíferos, es la mayor enzima
antioxidante y puede proteger contra la hidroperoxidación de lípidos en
biomembranas (Cai et al., 2008).
7.3.2.3 Actividad de transporte
Mielina P2 (spot 34, 75 y 249) es una proteína extrínseca básica de la
membrana localizada en los espacios citoplásmicos en las laminillas de mielina,
se han encontrado en el sistema nervioso central y periférico (Hunter et al.,
2005). Esta proteína es un miembro de la familia de proteínas que unen ácidos
grasos, presentan una alta afinidad con el ácido oleico, ácido retinoico y retinol
(Maddalo et al., 2010).
Las proteínas de unión a los ácidos grasos en el musculo, fueron
detectadas en el spot 75, esta proteína es muy importante en el metabolismo y
trasporte de cadenas largas de ácidos grasos y es responsable de la modulación

45
de algunas funciones como es el crecimiento y proliferación celular (Kizilbasch et
al., 2013).
Flavoproteínas de transporte de electrones (ETF) es un heterodímero de
la matriz mitocondrial con α ETFA; 30kDa, (spot 183) y β ETFB; 28 kDa (spot
105 y 107) subunidades. Sirve como aceptor especifico de varias
deshidrogenasas incluyendo 5-acil CoA deshidrogenasa, glutaril CoA, y
sarcosina deshidrogenasa. Transfieren electrones a la principal cadena
respiratoria mitocondrial vía ETF ubiquitina oxidoreductasa. Defectos en EFTA y
EFTB son la causa de la aciduria glutarica tipo 2 A, que es un desorden
hereditario de ácidos grasos, aminoácidos y el metabolismo de colina
(Rosenbohm et al., 2014).
7.3.2.4 Proteínas de enlace
Fosfotiadiletanolamina (PEBP, spot 23 y 230), son un grupo de proteínas
que se encuentran en el citosol y tienen la habilidad de enlazar fosfolípidos
(Zhang et al., 2007). Se ha reportado que en moscas transgénicas que sobre
expresan PEBP 1 están altamente protegidas contra la infección bacteriana
debido a la liberación de proteínas relacionadas contra la inmunidad en su
hemolinfa (Reumer et al., 2009).
Calumenina (spot 65) es un multicomplejo que presentan un dominio
estructural EF-Ca2+, se encuentra en el retículo endo/sarcoplasmico de
corazones de mamíferos. Esta proteína es altamente expresada durante la etapa
inicial de desarrollo de corazón y juega un rol esencial en el alivio del estrés del
retículo endoplásmico en cardiomiocitos de rata neonatal (Lee et al., 2013).
Nucleoplasmina (spot 299) ha sido descrita como una proteína que tiene
un rol activo en el ensamble de nucleosomas de histonas purificadas y ADN in
vitro. El nucleoplamino homólogo de Drosophila funciona como una chaperona
de histonas que puede remodelar la cromatina en los embriones (Padeken et al.,
2013).

46
Hexamerinas (spot 70, 342, 354, 355, 356, y 359) son proteínas derivadas
de hemocianina que han perdido la habilidad de enlazar iones de cobre y
transportar oxígeno y que han sido convertidas en proteínas de reserva (Martins
et al., 2010).
7.3.3 Procesos biológicos
7.3.3.1 Proteínas relacionadas con metabolismo
7.3.3.1.1 Metabolismo de carbohidratos
Enzimas relacionadas con el metabolismo de carbohidratos fueron
identificadas (glicolisis, Krebs y la vía de la pentosa fosfato) tal como:
gliceraldehido 3-phosphate dehidrogenasa, fructosa-bis fosfato aldosa, triosa
fosfato isomerasa, enolasa, fosfoglicerato mutasa, fosfoglicerato kinasa, malato
dehidrogenasa, piruvato dehidrogenasa y 6-fosfogluconalactonasa.
La enolasa es una enzima clave implicada en la vía glicolítica (spots 140,
146, 147, 158, y 199) y cataliza la conversión reversible de 2 fosfoglicerato a
fosfoenol piruvato. La enolasa también se conoce como una proteína
multifuncional involucrada en diferentes procesos fisiológicos y patológicos por
ejemplo, como proteína de unión del plasminogeno y la proteína de choque
térmico (Nguyen et al., 2013). Se ha demostrado que la enolasa de la superficie
celular puede mediar la interacción huésped-patógeno incluyendo las
alteraciones patológicas y la penetración de los tejidos del huésped por
patógenos y células tumorales (Diaz-Ramos et al., 2012).
Fosfoglicerato mutasa 1 (PGAM, spots 27 y 94) cataliza la conversión de
3-fosfoglicerato a 2-fosfoglicerato en la glicolisis, así como la mayoría de los
compuestos intermediarios que son usados como precursores para la biosíntesis
de anabólicos, en muchos tipos de canceres la actividad de PGAM1 se
incrementa en comparación a la de los tejidos normales (Hitosugi et al., 2012).
Fosfoglicerato kinasa (PGK spots 146 y 348) tiene como principal función
catalizar la conversión reversible de 1-3 difosfoglicerato a 3-fosfoglicerato
produciendo una molécula de ATP en la glucolisis. La deficiencia de PGK en

47
humanos está relacionada con disfunciones neurológicas y miopatías (Chiarelli
et al., 2012).
Difosfato kinasa (NDPK spots 17,42 y 77), la actividad de NDPK está
implicada en múltiples aspectos de la fisiología celular que incluye la regulación
de la expresión génica en células de mamíferos, patogénesis bacteriana y
limpieza de parásitos. En los insectos ayuda a alterar la actividad fisiológica del
anfitrión dando como resultado el establecimiento exitoso de las larvas (Sinha et
al., 2012). De Morais Guedes et al. (2005) reportaron que NDPK es una proteína
expresada diferencialmente en la hemolinfa del sistema inmune activado de D.
melanogaster larva.
7.3.3.1.2 Metabolismo de aminoácidos
Diversas enzimas relacionadas con el metabolismo de aminoácidos
fueron identificadas.
Arginina kinasa (spot 226) cataliza la trasferencia reversible de un grupo
fosforilo de alta energía por ATP a L-arginina para formar fosfoarginina, que es
usada como una energía amortiguadora, en insectos, crustáceos y algunos
organismos unicelulares. Debido a los papeles fundamentales en el metabolismo
energético y la ausencia en vertebrados, la arginina kinasa podría ser elegida
como proteína diana para el desarrollo de agentes quimioterapéuticos contra la
enfermedad de Chagas y otras enfermedades parasitarias causados por
organismos relacionados (Bragg et al., 2012).
Pterin-4α-carbinolamina dehidratasa (PCD, spots 79, 81 y 360) está
involucrada en la vía de la hidroxilación de fenilalanina, donde participa en la
producción de tetrahidrobiopterina (BH4). BH4 es el cofactor esencial de
fenilalanina hidroxilasa, la cual convierte de fenilalanina a tirosina; la falta de
BH4 causa desordene neurológico y retraso mental asociado con
fenilalanainemia o fenilketonuria (Eskinazi et al., 1999).
Aminoacialsa 1 (spots 222 y 223) es una enzima de enlace de zinc que
hidroliza al aminoácido N-acetil en aminoácidos libres y ácido acético. En

48
humanos la deficiencia de Aminoacialsa 1 sigue un patrón autosómico recesivo y
es caracterizada por la acumulación de N-acetilaminoácidos en la orina
(Sommer et al., 2011).
Fumarilacetoacetato hidrolasa (FAH, spot 110) cataliza la hidrolisis de 4-
fumarilacetoacetato en fumarato y acetoacetato, se trata de la última enzima en
la vía del catabolismo de la tirosina, la deficiencia de FAH está asociada con
tirosemia hereditaria tipo 1 (Mohamed et al., 2013).
En el Spot 329 fue identificada la homocisteína S-metiltransferasa. La
homocisteína es un aminoácido que contiene azufre y un factor de riesgo
implicado en las enfermedades cerebrovasculares y neurodegenerativas. Uno
de los productos del metabolismo de la homocisteína es la metionina,
aminoácido esencial en los mamíferos (Zang, et al., 2013). En mamíferos la
implementación de vitamina B puede reducir los niveles de homocisteína (Zang,
et al., 2013).
Purina (PURH, spots 45 y 46) es una proteína bifuncional que posee dos
actividades enzimáticas (fosforribosil aminoimidazol carboxamida formilo
trasferasa e IMP ciclohidrolasa) que cataliza de manera secuencial los dos
últimos pasos en la vía para la síntesis de novo de inosina 5-monofosfato. Esta
enzima bifuncional es de particular interés debido a su potencial en procesos
quimioterapéuticos (Bulock, et al., 2002).
7.3.3.1.3 Metabolismo de lípidos
La β oxidación mitocondrial, es el sistema metabólico más importante
para metabolizar ácidos grasos y producir Acetil CoA mediante la degradación
de ácidos grasos a través de una cascada de cuatro reacciones: deshidratación,
hidratación, oxidación y tiolisis.
Las proteínas trifuncionales mitocondriales (MTP) catalizan los tres
últimos pasos en la β oxidación de los ácidos grasos de cadena larga. MTP
consiste de un hetero-octámero de 4 MTPα y 4 subunidades MTPβ. La
subunidad MTPα posee actividades de 3-enoly-CoA hidratasa de cadena larga y

49
3-hidroxiacil-CoA deshidrogenasa la cual fue encontrada en el spot 96. La
subunidad MTPβ presenta actividad de 3-cetoacil CoA tiolasa que cataliza la
cuarta etapa. La deficiencia de MTP en humanos provoca el síndrome de Reye
(Kishita et al., 2012).
En el spot 140 fue encontrada la 4-hidroxibutirato CoA transferasa; en
bacterias intestinales estas enzimas desempeñan un papel clave en la formación
de butirato en el colon humano (Charrier et al., 2006).
Acil Co-A deshidrogenasa de cadena media (MCAD spot 130) es una de
las enzimas involucradas en la β oxidación de ácidos grasos que alimenta la
acetogénesis hepática, es una fuente importante de energía una vez que la
reservas de glucógeno se agotan durante el ayuno y en los periodos
prolongados de alta demanda de energía.
Acil Co-A deshidrogenasa de cadena corta (SBCAD) fue encontrada en el
spot 344 y 349. SBCAD cataliza la deshidrogenasa acil-CoA derivados en el
metabolismo de ácidos grasos. La deficiencia de SBCAD provoca un trastorno
autosómico recesivo y es estimulado por un defecto en el catabolismo de L-
isoleucina, el cual es caracterizado por un incremento de 2-metilbutirilglicina y 2-
metil butirilcarnitina en sangre y orina (Madsen et al., 2006).
Dehidrolipoil deshidrogenasa (spots 201 y 202) es un componente del
sistema de segmentación de la glicina.
7.3.3.2 Proteínas de plegamiento y degradación
Diferentes isoformas de proteínas de choque térmico HSC70 y HSC70-
interación con proteínas fueron identificadas en los spots 312, 313. 314, 338 y
342. HSC70 es una proteína chaperona constitutiva que pertenece a la familia
de proteínas de choque térmico 70 (HSP70). HSC70 realiza funciones de
cooperación con co-chaperonas como la proteína de interacción-HSC70 (Liu, et
al., 2012). La interacción con otras moléculas de proteínas regula varias

50
funciones celulares y podrían ser utilizadas como biomarcadores diagnóstico
para diferentes enfermedades (Liu et al., 2012).
La proteína HSP60 fue identificada como una proteína mitocondrial
(spots 319, 320 y 321). HSP60 forma un anillo complejo heptamérico que, con la
co-chaperona HSP10 (spot 2 y 6), permiten el adecuado plegamiento de las
proteínas mitocondriales en respuesta al estrés oxidativo. HSP10 es crucial para
la sobre vivencia de la células y su deficiencia en el cuerpo conduce a la
apoptosis celular y muerte embrionaria temprana en ratones (Christensen et al.,
2010). En humanos ha sido demostrado que existen diferentes mutaciones en el
gen (HSPD1) que codifica HSP60 y han sido asociadas con enfermedades
neurodegenerativas lo que indica que las neuronas son especialmente
vulnerables al mal funcionamiento de HSP60 (Magen et al., 2008). La
disminución aguda de HSP60 en el hipotálamo también induce la resistencia de
insulina (Kleinridders et al., 2013).
Prefoldina (spot 363) es una molécula chaperona encontrada en el
dominio de eucariotas y arqueobacterias que actúan en conjunto del grupo II
chaperonina para un correcto plegado de las proteínas nacientes (Whitehead, et
al., 2007).
En los spots 22, 91 y 282 las proteínas fueron identificadas como peptidil-
prolil cis-trans isomerasa (PPlasa), y en el spot 60 como FK506-proteina de
enlace ambos contienen el dominio de PPlasa. Esta actividad enzimática induce
la conversión de prolil-peptido de enlaces trans a cis, un frecuente paso limitante
en el plegamiento de la proteína. La actividad de PPIasa también ha sido
reportada como dominio de las inmunofilinas que se expresan en gran parte
durante la fase de larvas (Zhang et al., 2013).
Calreticulina (spot 67) participa en la homeostasis del calcio, funciones
inmunes y desempeña un papel importante en el plegamiento de nuevos
polipéptidos. La ubiquitina-proteosome sistema (UPS) fue identificada en los
spots 83, 106, 128, 243, 311, 318 y 347, es la principal vía proteolítica
responsable de la degradación intracelular contralada de proteínas de corta

51
duración y mal plegadas a través de la degradación de sustrato ubiquitina-
marcada (Lipinszki et al., 2013).
7.3.4 Señalización celular
Las proteínas relacionadas con la señalización celular también fueron
identificadas en el mapa proteómico de Liometopum apiculatum Mayr. El spot 55
fue identificado como calmodulina y el 100 como lectina tipo C. La señalización
de calcio está asociada a la detección de frio en los tejidos de insectos a través
de proteína quinasa II dependiente calcio/calmadulina (Teets, et al., 2013).
Lectina tipo C son una familia de proteínas con actividad de enlace a
carbohidratos, en mamíferos o artrópodos son empleados como receptores o
factores de fijación facilitando el reconocimiento de patógenos. En Drosofila se
han encontrado más de 30 genes que codifican lecitina C (Dodd y Drickmar,
2001).
7.3.5 Procesos de óxido-reducción
Entre las proteínas relacionadas con procesos de óxido-reducción, la
enzima disulfuro isomerasa (PDI) fue encontrada en los spots 315, 316, 333 y
336, pertenece a la familia disulfuro oxido reductasa con actividad tiol isomerasa
y oxido reductasa La función de PDI es la formación de enlaces disulfuro en la
formación de la proteína y en el plegamiento del retículo endoplasmático,
además podría regular la reorganización del citoesqueleto por el intercambio tiol-
disulfuro en β-actina a través de mecanismos redox-dependientes (Soblerajska,
et al., 2014).
En los spots 348 y 350 la enzima retinol hidrogenasa fue identificada. La
importancia biológica de los retinoides (vitamina A o retinol trans) radica en que
tanto el exceso como la deficiencia causan anormalidades embrionarias
importantes (Isken et al., 2007).

52
Hemocianinas identificada en el spot 361, tiene como función trasportar
oxígeno en la hemolinfa de insectos como plecoptera. En la actualidad ninguna
característica fisiológica o morfológica se conoce que pueda explicar la
presencia o perdida de hemocianinas en estos insectos (Burmester y Hankeln,
2007).
7.3.6 Otros procesos
Poly(A) ribonucleasa especifica (PARN, spot 32) es una de las enzimas
bioquímicamente caracterizadas y conservadas en células eucariotas y está
implicada en el control de la longitud de la cadena y descomposición de mRNA
en núcleo y citoplasma (Yoda et al., 2013).
La proteína no específica de transferencia de lípidos (spot 45) es una
familia de proteínas ubicuas, la cual participa en la transferencia de ácidos
grasos y fosfolípidos entre membranas (Hoh, et al., 2005).
Miosina de cadena ligera alcalina (spot 59), es una proteína del
citoesqueleto, regula múltiples procesos que están involucrados en el trasporte
de material, contracción muscular y división celular (Han, et al., 2010).
Calumenina (spot 65) es una proteína de unión a Ca2+ en conjunto con el
dominio estructural EF-Ca2+ localizado en el retículo endo/sarcoplasmico del
corazón de los mamíferos. La nucleoplasmina, (spot 299), se ha descrito como
una proteína que tiene una papel en el ensamblado de nucleosomas para
purificación de histonas y ADN in vitro (Padeken, et al., 2013).
Ferritina (spot 295) en insectos tiene dos funciones: almacenar y
trasportar hierro. En insectos la transferrina (spots 339, 340, 341 y 343) está
presente en la hemolinfa y puede funcionar como proteína de trasporte de hierro
(Phamy y Winzerling, 2010).
La regulcanina fue identificada en el spot 330, sus funciones es regular el
calcio intracelular y podría ser un candidato para ayudar a disminuir la
resistencia del frío a corto plazo (Reis et al., 2011).

53
Hexamerinas (spots 70, 342, 354, 355, 356 y 359) son proteínas
derivadas de hemocianina que han perdido la capacidad de enlazarse con los
iones de cobre y trasportar oxígeno; en cambio se convierten en proteínas de
almacenaje (Martins, et al., 2010).
El selenio es un nutriente esencial que exhibe un potente efecto anti
cancerígeno. La deficiencia de este puede causar enfermedades neurológicas
en humanos, la función antitumor del selenio es mediado a través de la proteína
selenio-enlace proteína 1 (SELENBP1 spots 334, 335, 337, y 344), SELENBP1
se expresa abundantemente en tejidos humanos normales y disminuye
significativamente cuando se detecta cualquier tipo de cáncer (Zhang et al.,
2013).
Quitinasa 3 y Quitinasa-like fueron identificadas en los spots (129, 135,
350, 366 y 367) tienen una extensa cantidad de funciones y están involucradas
en digestión, en la muda de piel de artrópodos, defensa/inmunidad y
patogenicidad (Koch, et al., 2014).
7.4 Perfil de ácidos grasos
Los resultados del contenido de ácidos grasos obtenidos del análisis de
GC-MS son mostrados en la Tabla 6 y el respectivo cromatograma del perfil de
los ácidos grasos en la figura 10. El resultado indica que los escamoles
presentan alto contenido de ácidos grasos insaturados y saturados, 67% y
31.02% respectivamente. Los ácidos grasos insaturados identificados en mayor
concentración fueron el ácido oleico (C18:1) y el ácido linoleico (C18:2) 56% y
4% respectivamente, otros ácidos mono (MUFA) y poliinsaturados (PUFA) como
ácido cis-10 nonadecenoico (C17:1), ácido cis-11-eicosenoico (C20:1), linolénico
(C18:3), palmitoleico (C16:1), miristoleico (C14:1) también fueron identificados
en menor proporción.
La isomería de los ácidos grasos CIS fue validada a través de tiempos de
retención con estándares de la marca de Sigma Aldrich confirmando así la

54
isomería de los ácidos grasos ácido cis-10-Heptadecenoico (7.815 min.), ácido
cis-10-Nonadecenoico (9.283 min.) y ácido cis-11-Eicosenoico (10.086 min.)
(Figura 11).
Tabla 6. Perfil de ácidos grasos en escamoles.
Nombre IUPAC Nombre trivial Carbonos % Relativo
Ácido dodecanoico Ácido láurico C12:0 0.403 ± 0.038
Ácido Tridecanoico Ácido tridecil C13:0 0.187 ± 0127
Ácido Tetradecanoico Ácido mirístico C14:0 2.389 ± 0.122
Ácido cis-9-Tetradecenoico Ácido miristoleico C14:1 cis 0.149 ± 0.033
Ácido Pentadecanoico Ácido pentadecílico C15:0 0.175 ± 0.099
Ácido Hexadecanoico Ácido palmítico C16:0 22.390 ± 0.498
Ácido cis-9-Hexadecenoico Ácido palmitoleico C16:1 cis 3.993 ± 0.180
Ácido Heptadecanoico Ácido margárico C17:0 0.558 ± 0.198
Ácido cis-10-Heptadecenoico Ácido cis-10-Heptadecenoico C17:1 cis 0.741 ± 0.161
Ácido Octadecanoico Ácido esteárico C18:0 4.673 ± 0.238
Ácido cis-9-Octadecenoico (Z) Ácido oleico C18:1n-9 cis 56.301 ± 0.245
Ácido cis-9,12-Octadecadienoico Ácido linoleico C18:2n-6 cis 4.205 ± 0.106
Ácido cis-10-Nonadecenoico Ácido cis-10-Nonadecenoico C19:1 cis 0.393 ± 0.042
Ácido cis-9,12,15-Octadecatrienoico Ácido linolénico C18:3n-3 cis 0.612 ± 0.051
Ácido Eicosanoico Ácido araquídico C20:0 0.621 ± 0.358
Ácido cis-11-Eicosenoico Ácido cis-11-Eicosenoico C20:1 cis 0.730 ± 0.115
Total ácidos graos saturados (SFA) --- --- 31.396 ± 0.328
Total ácidos grasos insaturados --- --- 67.124 ± 0.074
Mono insaturados (MUFA) --- --- 62.307 ± 0.048
Poliinsaturados (PUFA) --- --- 4.817 ± 0.116
n-6/n-3 --- --- 6.870 ± 0.768
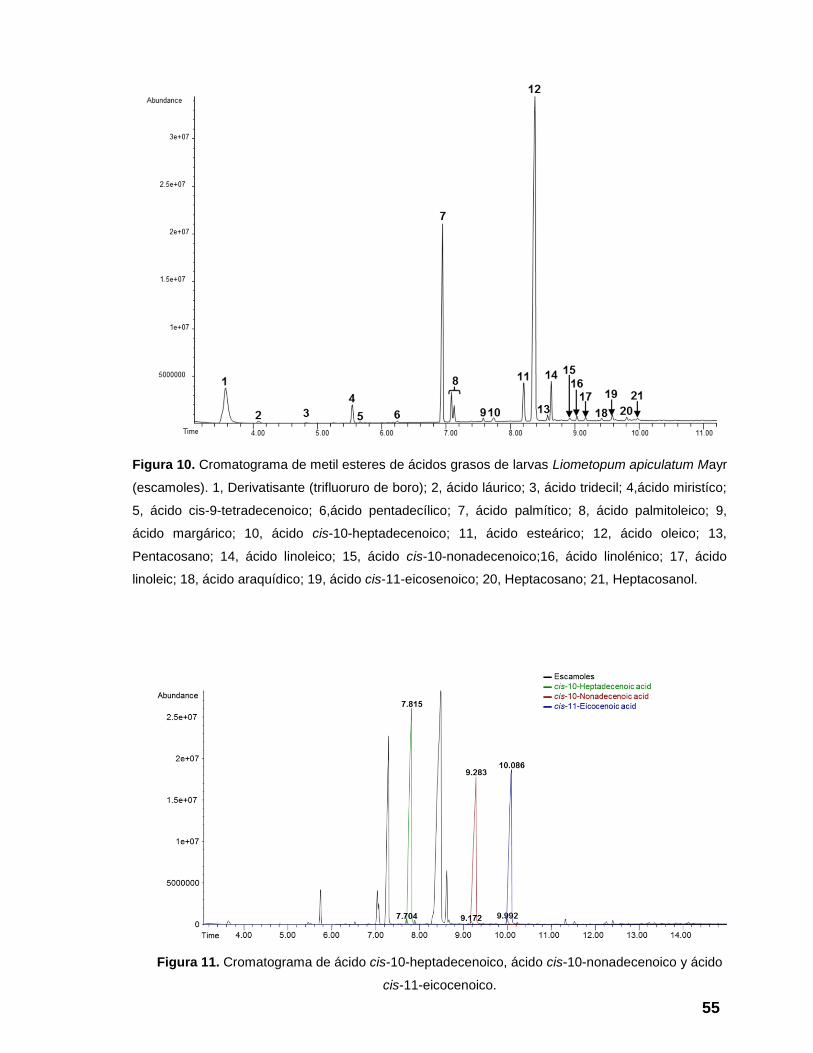
55
Figura 10. Cromatograma de metil esteres de ácidos grasos de larvas Liometopum apiculatum Mayr
(escamoles). 1, Derivatisante (trifluoruro de boro); 2, ácido láurico; 3, ácido tridecil; 4,ácido miristíco;
5, ácido cis-9-tetradecenoico; 6,ácido pentadecílico; 7, ácido palmítico; 8, ácido palmitoleico; 9,
ácido margárico; 10, ácido cis-10-heptadecenoico; 11, ácido esteárico; 12, ácido oleico; 13,
Pentacosano; 14, ácido linoleico; 15, ácido cis-10-nonadecenoico;16, ácido linolénico; 17, ácido
linoleic; 18, ácido araquídico; 19, ácido cis-11-eicosenoico; 20, Heptacosano; 21, Heptacosanol.
Figura 11. Cromatograma de ácido cis-10-heptadecenoico, ácido cis-10-nonadecenoico y ácido
cis-11-eicocenoico.

56
Estos resultados están de acuerdo a lo reportado en la bibliografía para
diversos insectos comestibles, debido a que algunos de estos ácidos grasos
insaturados han sido encontrados en otros insectos como Hymenoptera
(hormigas, abejas), Coleoptera (escarabajos y gusanos), Lepidopteras
(Caterpillars) (Raksakantong et al., 2010; Rumpold y Schlüter, 2013). En termitas
(Isopteras) de la especie Macrotermes bellicosus también se observó un alto
contenido de ácido oleico (41.74%) (Kynyuro et al., 2013).
Sin embargo, estos resultados difieren de lo reportado por Melo-Ruiz et al.
(2013) respecto al contenido de ácidos grasos en escamoles, estos autores
únicamente reportan 4 ácidos grasos e indican que el ácido linoleico (C18:2) es
el de mayor concentración (68%) seguido de linolénico (C18:3) (2.61%). Ácido
araquidónico (C20:4) (0.16%) y ácido eicosapentanoico (C20:5) (0.16%).
Las diferencias que existen entre ambos resultados pueden ser debidas al
método de extracción y/o separación de la muestra, esto debido a que en este
estudio en el cromatograma de la figura 10 se observó que los picos del ácido
oleico y linoleico son muy cercanos, por ello es preciso diferenciar los dos picos
modificando el método de separación. Esto pudiera sugerir que en el trabajo de
dichos autores se tomó un solo pico dándole prioridad al ácido linoleico ya que el
ácido oleico no fue reportado. Otro factor importante puede estar relacionado
con la región donde fueron recolectados los escamoles, debido a que el medio
ambiente, cambios climáticos, alimentación, sexo y edad de la hormiga influye
sobre la composición de los ácidos grasos entre diferentes especies y dentro de
la misma especie (Sihamala et al., 2010).
En la literatura se ha reportado que los insectos que pertenecen al género
Hymenoptera (hormigas y abejas) al cual pertenecen los escamoles presentan
mayor cantidad de ácidos grasos monoinsaturados (MUFA) que poliinsaturados
siendo estos últimos mayormente encontrados en especies marinas (Rumpold y
Schlüter, 2013). De esta manera el porcentaje de MUFA y PUFA reportado para
este género fue de 73.1 y 3.1% respectivamente (Rumpold y Schlüter, 2013) los
cuales son similares a los reportados en este estudio. Dentro de los ácidos

57
grasos saturados (SFA) los de mayor proporción fueron el ácido palmítico
(C16:0) y el ácido esteárico (C18:0) 22.3 y 4.7% respectivamente. Estos
resultados están de acuerdo a lo reportado en la literatura para otros insectos
como abejas, termitas y orugas (Rumpold y Schlüter, 2013). El porcentaje de
SFA reportado en la literatura para el género Hymenoptera fue de 30.8% y para
Isoptera (termitas) 42%. El porcentaje de SFA reportado en este estudio fue de
31.4% el cual fue similar al reportado para el género Hymenoptera.
Los ácidos grasos de los insectos comestibles han sido comparados con
los ácidos grasos de pollo y pescado de acuerdo a su grado de insaturación
indicando que el pollo y pescado contienen mayor cantidad de PUFA (ácidos
poliinsaturados) (Rumpold y Schlüter, 2013). Por el contrario la carne de bovino
y cerdo contienen menor cantidad de MUFA y PUFA comparado con insectos
comestibles.
Los resultados de este estudio claramente muestran que los ácidos
grasos insaturados fueron predominantes en el perfil de ácidos grasos de los
escamoles. Para poder realizar una comparación con la ingesta necesaria de
ácidos grasos que recomienda la Organización Mundial de la Alimentación
(FAO, 2012), para la prevención de diversas enfermedades, se realizó una
cuantificación de los ácidos grasos que presentaron un porcentaje relativo mayor
al 1.0%. Así los resultados de la concentración en mg/g de cada ácido graso
fueron obtenidos por interpolación del área de cada pico correspondiente a cada
ácido graso evaluado: ácido mirístico, ácido palmítico, ácido palmitoleico, ácido
esteárico, ácido oleico y ácido linoleico, las curvas de calibración de estos ácidos
se muestran en el ANEXO 2.
La Tabla 7 muestra la concentración en mg/g de cada ácido graso
analizado. Los resultados dan una reafirmación de la existencia de un alto
contenido de ácido oleico con un aporte de 26.07 mg/g, siendo mayor que el
resto de los ácidos grasos.
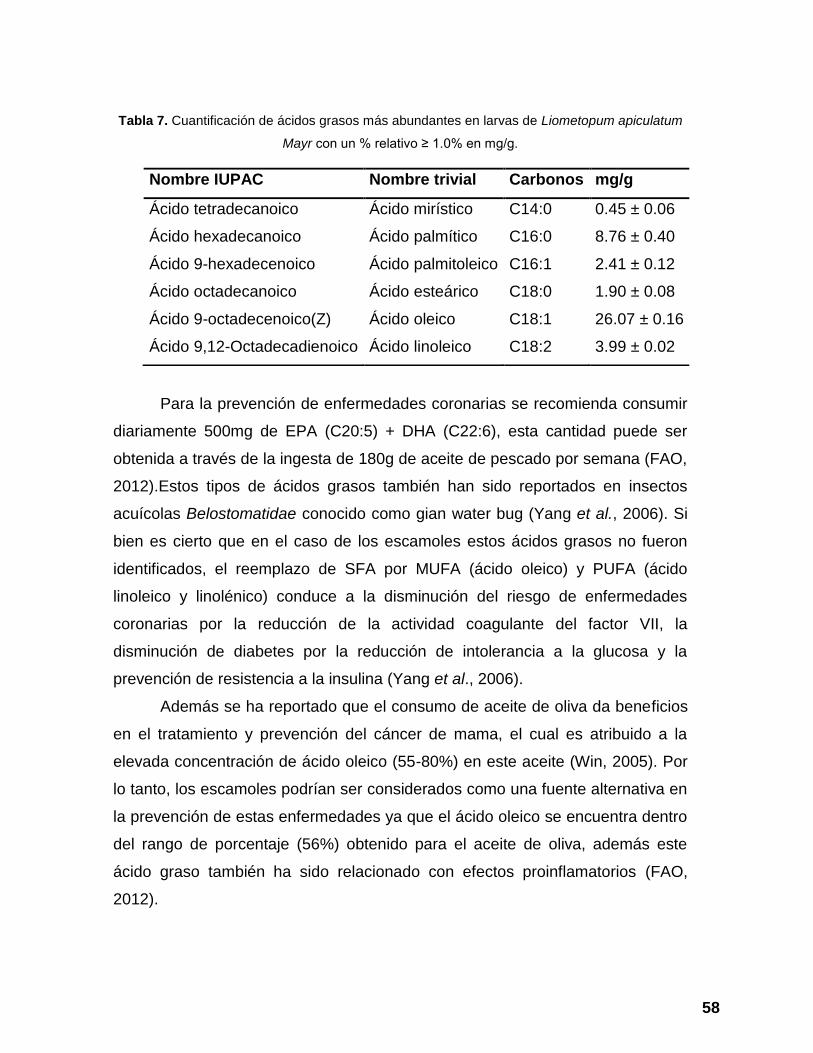
58
Tabla 7. Cuantificación de ácidos grasos más abundantes en larvas de Liometopum apiculatum
Mayr con un % relativo ≥ 1.0% en mg/g.
Para la prevención de enfermedades coronarias se recomienda consumir
diariamente 500mg de EPA (C20:5) + DHA (C22:6), esta cantidad puede ser
obtenida a través de la ingesta de 180g de aceite de pescado por semana (FAO,
2012).Estos tipos de ácidos grasos también han sido reportados en insectos
acuícolas Belostomatidae conocido como gian water bug (Yang et al., 2006). Si
bien es cierto que en el caso de los escamoles estos ácidos grasos no fueron
identificados, el reemplazo de SFA por MUFA (ácido oleico) y PUFA (ácido
linoleico y linolénico) conduce a la disminución del riesgo de enfermedades
coronarias por la reducción de la actividad coagulante del factor VII, la
disminución de diabetes por la reducción de intolerancia a la glucosa y la
prevención de resistencia a la insulina (Yang et al., 2006).
Además se ha reportado que el consumo de aceite de oliva da beneficios
en el tratamiento y prevención del cáncer de mama, el cual es atribuido a la
elevada concentración de ácido oleico (55-80%) en este aceite (Win, 2005). Por
lo tanto, los escamoles podrían ser considerados como una fuente alternativa en
la prevención de estas enfermedades ya que el ácido oleico se encuentra dentro
del rango de porcentaje (56%) obtenido para el aceite de oliva, además este
ácido graso también ha sido relacionado con efectos proinflamatorios (FAO,
2012).
Nombre IUPAC Nombre trivial Carbonos mg/g
Ácido tetradecanoico Ácido mirístico C14:0 0.45 ± 0.06
Ácido hexadecanoico Ácido palmítico C16:0 8.76 ± 0.40
Ácido 9-hexadecenoico Ácido palmitoleico C16:1 2.41 ± 0.12
Ácido octadecanoico Ácido esteárico C18:0 1.90 ± 0.08
Ácido 9-octadecenoico(Z) Ácido oleico C18:1 26.07 ± 0.16
Ácido 9,12-Octadecadienoico Ácido linoleico C18:2 3.99 ± 0.02

59
Otro ácido graso importante que presentan los escamoles es el ácido
linoleico (omega-6), este ácido graso es considerado un ácido graso esencial, ya
que el organismo no es capaz de sintetizarlo, el cual cumple funciones
principales en el mantenimiento del equilibrio metabólico y estructural (Morón
Moreno, 2008). Recientemente se ha demostrado que es un agente
antiinflamatorio de enfermedades intestinales (Reifen et al., 2015). La relación
de ácidos grasos omega 6 (linoleico) y omega 3 (linolenico) debe ser equitativo
ya que un desequilibrio puede inhibir la producción de ácidos ecosapentanoico
(EPA) y de ácido decosahexanoico (DHA), elementos importantes para
mantener la salud cardiovascular y el desarrollo del cerebro, la recomendación
de la relación de omega 6 y omega 3 es de 2:1 a 4:1 (Rodota, 2012). Por ello la
relación que presentan los escamoles es muy cercana a una relación
recomendada.
7.5 COLESTEROL LIBRE
En la Figura 12 se muestra el cromatograma de colesterol libre presente
en los escamoles, el tiempo de retención comparado con un estándar de la
marca Sigma Aldich con un grado de pureza del 98% fue a los 5 minutos. La
interpretación del área bajo curva también fue realizada como se indicó
anteriormente para ácidos grasos, obtenido de esta manera que la
concentración de colesterol en escamoles fue de 0.32±0.05 mg/g. La curva de
calibración se muestra en el ANEXO 2.

60
Diferentes estudios han sido realizados para evaluar el contenido de colesterol
en distintas especies de insectos colectados en Tailandia, por ejemplo los grillos,
langostas y escarabajos tienen un contenido de colesterol de 105 mg/100g, 66
mg/100g, 56 mg/100g respectivamente, (Rumpold y Schlüter, 2013) dichos
valores son superiores a los obtenidos para los escamoles. En huevos crudos
frescos se han reportado 372 mg/100g de colesterol, lo que significa que
contiene 10 veces más cantidad de colesterol que los escamoles y que otras
especies de insectos. De igual forma mayor cantidad de colesterol fue
observada en carne de bovino 2.739 mg/g (Pereira et al., 2003).
El contenido de colesterol en escamoles es muy similar al encontrado en
alimentos de origen marino como lo es el langostino con 0.709 mg/g de
colesterol (Tsape et al., 2010). Es importante considerar el consumo de
alimentos con bajo contenido de colesterol ya que es sintetizado por el
organismo, un consumo elevado es relacionado con un incremento en el riesgo
de enfermedades cardiovasculares (Gil, 2010).
Figura 12.Cromatograma comparativo de colesterol estándar y escamoles.

61
8. CONCLUSIONES
Este trabajo presenta por primera vez el mapa proteómico de los
escamoles de la especie Liometopum apiculatum Mayr, el cual es de gran
importancia para conocer el tipo de proteínas presentes en estos insectos y así
poder comprender la biología de estos.
Los escamoles son una fuente importante de proteínas con diversas
funciones biológicas, aportando un valor agregado a quien los consume.
El estudio de proteómica realizado en esta investigación proporciona una
base científica para identificar péptidos bioactivos que pueden tener un beneficio
a la salud convirtiéndose en un modelo para el estudio de enfermedades
degenerativas en seres humanos.
Los escamoles son una fuente importante de ácidos grasos insaturados
principalmente de ácido oleico y linoleico, los cuales son importantes para la
nutrición humana ya que estos aportan beneficios a la salud, como la prevención
de enfermedades cardiovasculares y cáncer de mama.
Los escamoles, contienen bajo contenido de colesterol (0.32 mg/g) 10
veces menor que alimentos convencionales (huevo y carne).
Por lo tanto el consumo de escamoles proporciona a los consumidores
nutrientes de calidad como proteínas, ácidos grasos insaturados y colesterol
bajo por lo que pueden formar parte de una dieta balanceada proporcionando
beneficios a su salud.

62
9. BIBLIOGRAFÍA
A
Abu-Reidah, I. M., Ali-Shtayeh, M. S., Jamous, R. M., Arráez-Román, D.,
Segura-Carretero, A. (2015). HPLC–DAD–ESI–MS/MS screening of
bioactive components from Rhus coriaria L. (Sumac) fruits. Food
Chemistry, 166, pp. 179–191.
Anyanful, A., Sakube, Y., Takuwa, K., Kagawa, H. (2001). The Third and Fourth
Tropomyosin Isoforms of Caenorhabditis elegans are Expressed in the
Pharynx and Intestines and are Essential for Development and
Morphology. Journal of Molecular Biology, 313, 525-537.
Arana, A. F. (2003). Insectos Comestibles, Entomofobia y Entomofagía. PhD
Thesis (Tesis Doctoral Facultad de Ciencias, Biología), UNAM, México
City.
B
Banmeyer, I., Marchand, C., Clippe, A. & Knoops, B. (2005). Human
mitochondrial peroxiredoxin 5 protects from mitochondrial DNA damages
induced by hydrogen peroxide. FEBS Letters, 579, 2327–2333.
Bragg, J., Rajkovic, A., Anderson, C., Curtis, R., Van Houten, J., Begres, B.,
Naples, C., Snider, M., Fraga, D. & Singer, M. (2012). Identification and
Characterization of a Putative Arginine Kinase Homolog from Myxococcus
xanthus Required for Fruiting Body Formation and Cell Differentiation.
Journal of Bacteriology, 194, p. 2668–2676.

63
Buckner, J. S., Hagen, M. M. & Nelson, D. R. (1999). The composition of the
cuticular lipids from nymphs and exuviae of the Silverleaf Whitefly,
Bemisia argentifolii. Comparative Biochemistry and Physiology Part B 124,
201–207
Bulock, K. G., Beardsley, G. P. & Anderson, K. S. (2002). The Kinetic
Mechanism of the Human Bifunctional Enzyme ATIC (5-Amino-4-
imidazolecarboxamide Ribonucleotide Transformylase/Inosine 5-
Monophosphate Cyclohydrolase). The Journal Of Biological Chemistry,
Vol. 277, No. 25, Pp. 22168–22174.
Burmester, T., Hankeln, T. (2007). The respiratory proteins of insects. Journal of
Insect Physiology, 53, 285–294.
C
Cai, G. B., Bae, Y. A., Kim, S. H., Sohn, W. M., Lee, Y. S., Jiang, M. S., Kim, T.
S. & Kong, Y. (2008). Vitellocyte-specific expression of phospholipid
hydroperoxide glutathione peroxidases in Clonorchis sinensis.
International Journal for Parasitology, 38, 1613–1623.
Cambero Rodriguez, M. I. (1998). Tecnología de los alimentos: Alimento de
origen animal. Ordóñez Pereda, J. A. (Ed.). Ciencias químicas.
Tecnología bioquímica y de los alimentos. Editorial illustrated. Madrid. Pp.
368.
Castillo-Andrade, A., García-Barrientos, R., Ruiz-Cabrera, M. A., Rivera-Bautista,
C., Pérez-Martínez, J. D., González García, R., Fuentes-Ahumada, C.,
Grajales-Lagunes, A. (2015). Structural and physicochemical changes due
to proteolytic deterioration of escamoles (Liometopum apiculatum) a

64
traditional Mexican food. Journal of Insects as Food and Feed. Pages: 271
– 280.
Castillo-Lara, P., Aguirre-Rivera, J. R. (2008). Platillos gourmet del altiplano:
Escamoles. Revista “El Cenzontle” SEDARH, Gobierno del Estado de San
Luis Potosí. 3(2):8.
Chapman, R. F. (2013). The insects: Structure and Function. Simpson, S. J. and
Douglas A. E. (Ed.). Cambridge University Press, United States of
America pp. Xiii.
Charles, J. P. (2010). The regulation of expression of insect cuticle protein
genes. Insect Biochemistry and Molecular Biology, 40, 205-213.
Charrier, C., Duncan, G. J., Reid, M. D., Rucklidge, G. J., Henderson, D., Young,
P., Russell, V. J., Aminov, R. I., Flint, H. J. & Louis, P. (2006). A novel
class of CoA-transferase involved in shortchain fatty acid metabolism in
butyrate-producing human colonic bacteria. Microbiology, 152, 179–185.
Chiarelli, L. R., Morera, S. M., Bianchi, P., Fermo, E., Zanella, A., Galizzi, A.,
Valentini, G. (2012). Molecular Insights on Pathogenic Effects of Mutations
Causing Phosphoglycerate Kinase Deficiency. PLoS One 7(2), e32065.
Christensen, J. H., Nielsen, M. N., Hansen, J., Füchtbauer, A., Füchtbauer, E.
M., West, M., Corydon, T. J., Gregersen, N., Bross, P. (2010). Inactivation
of the hereditary spastic paraplegia-associated Hspd1 gene encoding the
Hsp60 chaperone results in early embryonic lethality in mice. Cell Stress
and Chaperones, 15, 851–863.

65
Cortés Gabaudan, F., Ureña Bracero, J. (2014). Dicciomed.eusal.es. Diccionario
médico-biológico, histórico y etimológico. Recuperado de Dirección URL:
http://dicciomed.eusal.es. Cid Ledesma, E., Cortés Gabaudan, F.,
Marchena, M., Pedrero, P., Ledesma Martín, M. C., Ureña Bracero, J.,
Villalba Álvaréz, J., Cid Ledesma, T. (Ed.). Salamanca.
Costa-Neto, E. M. (2003). Insects as sources of proteins for man: Valorization of
disgusting resources. Interciencia, 28, pp. 136–140.
Costa-Neto, E. M., Ramos-Elorduy, J. (2006). Los insectos comestibles de
Brasil: Etnicidad, diversidad e importancia en la alimentación. Boletín
sociedad entomológica Aragonesa, no. 38: 423-442.
Costa-Neto, E. M., Ramos-Elorduy, J., Pino, J. M. (2006). Los insectos
Medicinales del Brasil: Primeros resultados. Boletín Sociedad
Entomológica Aragonesa, no. 38: 395−414.
D
Del Toro I., Pacheco J. A., Mackay W. P., (2009). Revision of the Ant Genus
Liomeopum Mayr (Hymenoptera:Formicidae). Sociobiology 52:299-369.
DeFoliart, G. R. (1999). INSECTS AS FOOD: Why the Western Attitude Is
Important. Annual Review Entomology. 44:21–50
De Morais Guedes, S., Victorino, R., Domingues, R., Tomer, K., Correia, A. J. F.,
Amado, F. Domingues, P., (2005). Proteomics of immune-challenged
Drosophila melanogaster larvae hemolymph. Biochemical and Biophysical
Research Communications 328, 106–115.

66
Díaz-Ramos, A., Roig-Borrellas, A., García-Melero, A., López-Alemany, R.
(2012). α-Enolase, a Multifunctional Protein: Its Role on
Pathophysiological Situations. Journal of Biomedicine and Biotechnology
Volume 2012, 1-12.
Dodd, R. B., Drickamer, K. (2001). Lectin-like proteins in model organisms:
implications for evolution of carbohydrate-binding activity. Glycobiology,
11, 71R-9R.
Duncan, J. E., Lytle, N. K., Zuniga, A., Goldstein, L. S. B. (2013). The
Microtubule Regulatory Protein Stathmin Is Required to Maintain the
Integrity of Axonal Microtubules in Drosophila. PLoS One, 8, 6, e68324.
Durst, P. B., Shono, K. (2010). Edible forest insects: exploring new horizons and
traditional practices. Durst, P. B., Johnson, D. V., Leslie, R. N., Shono, K.
(Ed.). Pp. 1-4.
E
Eskinazi, R., Thöny, B., Svoboda, M., Robberecht, P., Dassesse, D., Heizmann,
C.W., Van Laethem, J. L. & Resibois, A. (1999). Overexpression of Pterin-
4a-carbinolamine Dehydratase/Dimerization Cofactor of Hepatocyte
Nuclear Factor 1 in Human Colon Cancer. The American Journal of
Pathology, Volume 155, Pages 1105–1113.
Estudio FAO Alimentación y Nutrición, (2012). Grasas y ácidos grasos en
nutrición humana, Consulta de expertos, Granada, España, 15, 24, 104.

67
F
Farb, P. (2011). Los insectos, Colección de la naturaleza de Time-Life. Ediciones
culturales internacionales. México, D.F. pp14.
FAO. (2012). Los insectos comestibles de los bosques, visto en el línea el
18/02/13 http://www.fao.org/forestry/edibleinsects/65425/es/
FAO. (2007). Promises and challenges of the informal food sector in developing
countries. Rome.
Fennema, O. R. (2000). Química de los Alimentos. Acribia. Zaragoza 2ª.Ed. Pp.
1258.
Fessenden R. J. (1983). Química orgánica. 2ª. Edición. Editorial Iberoamérica
S.A. de C.V. México D.F. Pp. 814, 894, 858.
G
García-Ca as, V., Simó,C., Herrero, M., Ibá ez, E., Cifuentes, A. (2012). Present
and Future Challenges in Food Analysis: Foodomics. American Chemical
Society. 84, 10150−10159.
García S. (2008). Técnicas instrumentales de análisis en bioquímica. Editorial
Síntesis, S. A. España. pp. 250, 251, 259.
Ghaly, A. E. (2009). The use of insects as human food in Zambia. Journal of
Biological Sciences. 9 (4): 93-104.

68
Gil Hernández, A., Martínez de Victoria Muñoz, E. & Maldonado Lozano, J.,
(2010). Tratado de nutrición, Tomo III Nutrición Humana en el Estado de
Salud, 2ª Edición, Editorial Medica Panamericana, S. A., Madrid, España,
300-301.
Goldstein, A. L., Hannappel, E., Kleinman, H., K. (2005). Thymosin β4: actin-
sequestering protein moonlights to repair injured tissues. Trends in
Molecular Medicine Vol.11 No.9.
Gołębiowski, M., Boguś, M. I., Paszkiewicz, M. & Stepnowski, P. (2011).
REVIEW: Cuticular lipids of insects as potential biofungicides: methods of
lipid composition analysis. Analytical and Bioanalytical Chemistry.
399:3177–3191.
Guillott, C. (2005). Entomology. 3ª Edition. Springer, Netherlands. Pp. 731.
Gurniak, C. B., Perlas, E. & Witke, W. (2005). The actin depolymerizing factor n-
cofilin is essential for neural tube morphogenesis and neural crest cell
migration. Developmental Biology, 278 231– 241.
H
Han, F., Wang, Z. & Wang, X. (2010). Characterization of myosin light chain in
shrimp hemocytic phagocytosis. Fish & Shellfish Immunology, 29, 875-
883.
Hitosugi, T., Zhou, L., Elf, S., Fan,J., Kang, H. B., Seo, J. H., Shan, C., Dai, Q.,
Zhang, L., Xie, J., Gu, T. L., Jin, P., Alečković, M., LeRoy, G., Kang, Y.,
Sudderth, J. A., DeBerardinis, R. J., Luan, C. H., Chen, G. Z., Muller, S.,
Shin, D. M., Owonikoko, T. K., Lonial, S., Arellano, M. L., Khoury, H. J.,

69
Khuri, F. R., Lee, b. h., Ye, k., Boggon, T. J., Kang, S., He, C., Chen, J.
(2012). Phosphoglycerate mutase 1 coordinates glycolysis and
biosynthesis to promote tumor growth. Cancer Cell, 13, 585-600.
Hoh, F., Pons, J. L., Gautier, M. F., Lamotte, F. & Dumas C. (2005). Structure of
a liganded type 2 non-specific lipid-transfer protein from wheat to the
molecular basis of lipid binding. Acta Crystallogr D: Biological
Crystallography, 61, 397-406.
Horowitz, A., Clément-Chomienne, O., Walsh, M. P.; Tao, T., Katsuyama, H. &
Morgan, K. G. (1996). Effects of calponin on force generation by single
smoth muscle cells. American Journal of Physiology-Heart and Circulatory
Physiology, 270, H1858.1863.
Hunter, D. J. B., Macmaster, R., Roszak, A. W., Riboldi-Tunnicliffe, A., Griffiths, I.
R. & Freer, A. A. (2005). Structure of myelin P2 protein from equine spinal
cord. Acta Crystallographica Section D: Biological Crystallography, 61,
1067–1071.
I
Ishii, T., Warabi, E. & Yanagawa, T. (2012). Novel roles of peroxiredoxins in
inflammation, cancer and innate immunity. Journal Clinical Biochemistry
and Nutrition, vol. 50, pp. 91–105.
Isken, A., Holzschuch, J., Lampert, J. M., Fischer, L., Oberhauser, V.,
Palczewski, K. and von Linting. (2007). Sequestration of Retinyl Esters Is
Essential for Retinoid Signaling in the Zebrafish Embryo. The Journal Of
Biological Chemistry, Vol. 282, No. 2, Pp. 1144–1151.

70
J
Jongema, Y. (2012). List of edible insects of the world (Julio 15, 2013).
http://www.fao.org/forestry/edibleinsects/65425/es/
Juárez Sandoval, J. J., Melo Ruiz, V. E., Pèrez Santiago, D., Calvo Carrillo, C.
(2010). Contenido de proteínas y aminoácidos en escamoles (liometopum
apiculatum M.) capturados en el estado de hidalgo. Congreso
internacional de QFB 2010, Edición especial No. 10-2010.
K
Kim, Y. I., Kim, H. J., Kwon, Y. M., Kang, Y. J., Lee, I. H., Jin, B. R., Han, Y. S.,
Cheon, H. M., Ha, N. G. & Sep, S. J. (2010). Modulation of MnSOD
protein in response to different experimental stimulation in Hyphantria
cunea. Comparative Biochemistry and Physiology, Part B 157, 343–350.
Kishita, Y., Tsuda, M., Aigaki, T. (2012). Impaired fatty acid oxidation in a
Drosophila model of mitochondrial trifunctional protein (MTP) deficiency.
Biochemical and Biophysical Research Communications 419, 344–349.
Kizilbach, N. A., Hai, A., Alruwaili, J. (2013). Model of β-Sheet of Muscle Fatty
Acid Binding Protein of Locusta migratoria Displays Characteristic
Topology. Bioinformation, 9: 1003-1009.
Kleinridders, A., Lauritzen, H. P. M. M., Ussar, S., Christensen, J. H., Mori, M. A.,
Bross, P., Kahn, C. R. (2013). Leptin regulation of Hsp60 impacts
hypothalamic insulin signaling. The Journal of Clinical Investigation, 13,
4667-4680.

71
Kinyuru, J. N., Konyole, S. O., Roos, N., Onyango, C. A., Owino, V. O., Owuor,
B. O., Estambale, B. B., Friis, H., Aagaard-Hansen, J., Kenji, G. M.,
(2013). Nutrient composition of four species of winged termites consumed
in western Kenya. Journal of Food Composition and Analysis 30, 120-124.
Koch, B. E. V., Stougaard, J. & Spaink, H. P. (2014). Spatial and temporal
expression patterns of chitinase genes in developing zebrafish embryos.
Gene Expression Patterns, 14, 69–77.
L
Lara Juárez, P. (2013). Etnobiología de escamoles (liometopum apiculatum) en
el altiplano potosino. (Tesis que para obtener el grado de Maestría en
ciencias ambientales). Universidad Autónoma de San Luis Potosí. México,
S.L.P.
Ladrón de Guevara, O., Padilla, P., García, L., Pino, J. M. & Ramos-Elorduy, J.
(1995). Amino acid determination in some edible Mexican insects. Amino
Acids 9, 161-173.
Lee, J. H., Kwon, E. J., Kim, D. H. (2013). Calumenin has a role in the alleviation
of ER stress in neonatal rat cardiomyocytes. Biochemical and Biophysical
Research Communications 439, 327–332.
Lipinszki, Z., Klement, E., Hunyadi-Guylyas, E., Medzihradszky, K. F., Márkus,
R., Pál, M., Deák, P. & Udvardy, A. (2013). A novel interplay between the
ubiquitin–proteasome system and serine proteases during Drosophila
development. Biochemical Journal, 454, 571–583.

72
Liu, T., Daniels, C. K. & Cao, S. (2012). Comprehensive review on the HSC70
functions, interactions with related molecules and involvement in clinical
diseases and therapeutic potential. Pharmacology & Therapeutics, 136,
354–374.
Lu, J. & Pollard, T. D. (2001). Profilin Binding to Poly-L-Proline and Actin
Monomers along with Ability to Catalyze Actin Nucleotide Exchange Is
Required for Viability of Fission Yeast. Molecular Biology of the Cell, Vol.
12, 1161–1175.
M
Madsen, P. P., Kiæk, M., Roca, X., Sachidanandam, R., Krainer, A. R.,
Christensen, E., Steiner, R. D., Gibson, K. M., Corydon, T. J., Knudsen, I.,
Wanders, R. J. A., Ruiter, J. P. N., Gregersen, N., Andresen, B. S. (2006).
Short /branched-chain acyl-CoA dehydrogenase deficiency due to an
IVS3+3A>G mutation that causes exon skipping. Human Genetics. 118,
680-690.
Magen, D., Georgopoulos, C., Bross, P., Ang, D., Segev, Y., Goldsher, D.,
Nemirovski, A., Shahar, E., Ravid, S., Luder, A., Heno, B., Gershoni-
Baruch, R., Skorecki, K., Mandel, H. (2008). Mitochondrial Hsp60
chaperonopathy causes an autosomal-recessive neurodegenerative
disorder linked to brain hypomyelination and leukodystrophy. The
American Journal of Human Genetics 83, 30–42.
Maddalo, G., Shariatgorji, M., Adams, C. M., Fung, E.,Nilsson, U., Zubarev, R.
A., Sedzik, J., Ilag, L. L. (2010). Porcine P2 myelin protein primary
structure and bound fatty acids determined by mass spectrometry.
Analytical and Bioanalytical Chemistry. 397, 1903–1910.

73
Manevich, Y. & Fisher, A. B. (2005). Peroxiredoxin 6, a 1-Cys peroxiredoxin,
functions in antioxidant defense and lung phospholipid metabolism. Free
Radical Biology & Medicine, 38, 1422 – 1432.
Martins, J. R., Nunes, F. MF., Cristino, A. S., Simões, Z. LP. & Bitondi, M. MG.
(2010). The four hexamerin genes in the honey bee: structure, molecular
evolution and function deduced from expression patterns in queens,
workers and drones. BMC Molecular Biology, 11:23.
Matallana-González, M. C., Torija-Isasa, M. E. (2006) Los insectos como
alimento: ¿están de moda? El consumo de insectos en el mundo.
Facultad de Farmacia. Universidad Complutense de Madrid. Schironia, 5:
41-49.
Melo Ruiz, V., Sánchez Herrera, K., Sandoval Trujillo, H., Quirino Barreda, T.
and Calvo Carrillo, C., (2013). Lipids Data Composition of Edible Ant Eggs
Liometopum apiculatum M. Escamoles. Journal of Life Sciences, ISSN
1934-7391, USA, Vol. 7, No. 5, pp. 547-552.
Mendoza Martínez, E. & Calvo Carrillo, M. C. (2010). Bromatología Composición
y propiedades de los alimentos. McGraw Hill. México. Pp. 67-68.
Miranda Román, G., Quintero Salazar, B., Ramos Rastro, B. (2011). La
recolección de insectos con fines alimenticios en la zona turística de
Otumba y Teotihuacán, Estado de México. Revista de turismo y
patrimonio cultural, vol. 9 No. 1, Estado de México, pp. 81-98
Mitra, A. (2013). Cinderella’s new shoes – how and why insects remodel their
bodies between life stages. Current Science, 104, 1028-1036.

74
Mohamed, S., Kambal, M. A., Jurayyan, N. A. A., Al-Nemri, A., Babiker, A.,
Hasanato, R., Al-Jarallah, A. S. (2013). Tyrosinemia type 1: a rare and
forgotten cause of reversible hypertrophic cardiomyopathy in infacy.
BioMed Central, 6, 362.
Morón Moreno, A., (2008). Las grasas esenciales y mortales. Qué son, Dónde
están y Cómo reconocerlas. Editorial Díaz de Santos, España, 43, 47.
N
Naciones Unidas. (2014). La situación demográfica en el mundo. Departamento
de asuntos económicos y sociales. División de población. Pp. 1-4.
Nason, A. (2000). Biología. Editorial Limusa. México, D.F. pp. 425.
Nguyen, T. T. A., Magnoli, I., Cloutier, C., Michaud, D., Muratori, F., Hance, T.
(2013). Early presence of an enolase in the oviposition injecta of the aphid
parasitoid Aphidius ervi analyzed with chitosan beads as artificial hosts.
Journal of Insect Physiology. 59, 11–18.
O
OMS, serie de Informes Técnicos 916, (2003). Dieta, Nutrición y Prevención de
enfermedades crónicas. Consulta Mixta de Expertos OMS/FAO. Ginebra,
87.

75
P
Padeken, J., Mendiburo, M. J., Chlamydas, S., Schwarz, H. J., Kremmer, E. &
Heun P. (2013). The Nucleoplasmin Homolog NLP Mediates Centromere
Clustering and Anchoring to the Nucleolus. Molecular Cell, 50, 236–249.
Passarge, E. (2010). Genética texto y atlas. 3º. Edición. Editorial medica
Panamericana. España. pp. 2,3.
Pavlopoulos, A., Akam, M. (2011). Hox gene Ultrabithorax regulates distinct sets
of target genes at successive stage of Drosophila haltere morphogenesis.
Proceeding of the National Academy of Sciences. 108, 2855-2860.
Pereira, N. P., Ferrarese-Filho, O., Matsushita, M. & E. de Souza, N., (2003).
Proximate composition and fatty acid profile of Bombyx mori L. chrysalis
toast. Journal of Food Composition and Analysis, 16, 451–457.
Pham, D. Q. D., Winzerling, J. J. (2010). Insect ferritins: Typical or atypical?.
Biochimica et Biophysica Acta, 1800, 824–833.
Potter, N. N., Ph. D. (1978). La ciencia de los alimentos. Editorial Edutex, S.A.
México, D.F. Pp. 41, 47.
Prinz, R. D., Willis, C. M., van Kuppevelt, T. H., Klüppel, M. (2014). Biphasic Role
of Chondroitin Sulfate in Cardiac Differentiation of Embryonic Stem Cells
through Inhibition of Wnt/β-Catenin Signaling. PLoS One, 3, e92381.

76
R
Ramírez García, G. L. (2012). Uso del método de barrera en la conservación de
escamoles (Liometopum apiculatum) frescos (Tesis que para obtener el
grado de Maestría en ciencias en bioprocesos). Universidad Autónoma de
San Luis Potosí. México, S.L.P. pp. 47
Ramos-Elorduy, J., Pino Moreno, J. M., Escamilla Prado, E., Alvarado Pérez, A.,
Lagunez Otero, J. & Ladrón de Guevara, O. (1997). Nutritional Value of
Edible Insects from the State of Oaxaca, Mexico. Journal of food
composition and analysis 10, 142–157.
Ramos-Elorduy, J., Pino M., J. M., Cuevas Correa, S., (1998). Insectos
comestibles del Estado de México y determinación de su valor nutritivo.
Anales Instituto de Biología, Universidad Nacional Autónoma de México,
Serie Zoología, 69(1), 65-104.
Ramos-Elorduy, J., Pino, J. M., Conconi, M. (2006). Ausencia de una
reglamentación y normalización de la explotación y comercialización de
insectos comestibles en México. Folia Entomológica Mexicana, vol. 45,
núm. 3, pp. 291-318.
Ramos-Elorduy J., Pino J. M., Morales de León J., (2002). Análisis químico
proximal, vitaminas nutrimentos inorgánicos de insectos consumidos en el
estado de Hidalgo, México. Folia Entomológica Mexicana 41:15-29.
Ramos-Elorduy, J., Costa N., Pino-Moreno, J., Cuevas, C., García, F., (2007).
Conocimiento de la entomofauna útil en el poblado La Purísima Palmar de
Bravo, Estado de Puebla, México. Biotemas 20: 121-134.

77
Ramos-Elorduy, J., Viejo Montesinos, J. L. (2007). Los insectos como alimento
humano: Breve ensayo sobre la entomofagia, con especial referencia a
México. Boletín de la Real Sociedad Española de la Historia Natural
Sección Biología. 102 (1-4). Pp. 61-84.
Ramos-Elorduy, J. (2009). ¿Los insectos se comen?. Dirección general de
divulgación de la ciencia, UNAM. México. Pp. 7
Ramos Rostro B., Quintero Salazar B., Ramos-Elorduy J., Pino Moreno J. M.,
Ángeles Campos S. C., García Pérez Á., et al. (2012). Análisis químico y
nutricional de tres insectos comestibles de interés comercial en la zona
arqueológica del municipio de San Juan Teotihuacán y en Otumba, en el
estado de México. Interciencia 37:914-20.
Raksakantong, P., Meeso, N., Kubola, J., Siriamornpun, S., (2010). Fatty acids
and proximate composition of eight Thai edible terricolous insects. Food
Research International 43, 350–355.
Reifena, R., Karlinskya, A., Starka, A. H., Berkovicha, Z., Nyskab, A. (2015). α-
Linolenic acid (ALA) is an anti-inflammatory agent in inflammatory bowel
disease. Journal of Nutritional Biochemistry, 26, 1632–1640.
Reis, M., Vieira, C. P., Morales-Hojas, R., Aguiar, B., Rocha, H., Schlötterer, C. &
Vieria, J. (2011). A Comparative Study of the Short Term Cold Resistance
Response in Distantly Related Drosophila Species: The Role of regucalcin
and Frost. PLoS ONE, 6 (10), e25520.
Reumer, A., Bogaerts, A., Van Loy, T., Husson, S. J., Temmerman, L., Choi, C.,
Clynen, E., Hassan, B., Schoofs, L. (2009). Unraveling the protective
effect of a Drosophila phosphatidylethanolamine-binding protein upon

78
bacterial infection by means of proteomics. Developmental and
Comparative Immunology, 33, 1186–1195.
Rodota, L. P. & Castro, M. E., (2012). Nutrición y dietoterapia, 1ª Ed., Médica
Panamericana, Buenos Aires, Argentina, 46.
Romero-Contreras, A. T., Viesca-González, F. C. (2009). La Entomofagia en
México. Algunos aspectos culturales. El Periplo Sustentable. 16, pp. 57-
83.
Rosenbohm, A., Süssmuth, S. D., Kassubek, J., Müller, H. P., Pontes, C., Abicht,
A., Bulst, S., Ludolph, A. C., Pinkhardt, E. (2014). Novel ETFDH mutation
and imaging findings in an adult with glutaric aciduria type II. Muscle &
Nerve, 49, pp. 446-450
Rowe, A., Macebo, F. A. F., Visentainer, J. V., Souza, N. E. & Matsushita, M.
(1999). Muscle composition and fatty acid pro®le in lambs fattened in
drylot or pasture. Meat Science 51, 283-288.
Rumpold, B. A., Schlülter, O. K. (2013). Potential and challenges of insects as an
innovative source for food and feed production. Innovative Food Science
and Emerging Technologies. 17, 1–11.
Ruppert, E. E., Barnes, R. D. (1996). Zoología de los invertebrados. 6ed.
McGraw Hill interamericana. México. Pp. 833.
S
Salas García, S. V. (2011). Determinación del tiempo de proceso experimental y
simulación para el enlatado de escamoles (Liometopum apiculatum.)

79
(Tesis que para obtener el título de Ingeniero en Alimentos). Universidad
Autónoma de San Luis Potosí. México, S.L.P. pp. 11
Senser, F. & Scherz, H., (1991). Tablas de composición de alimentos, El
Pequeño “Souci-Fachmann-Kraut”. Editorial Wissenschaftliche
Verlasgesellscharft mbH, Birkenwaldstr. 44, D-70191 Stuttgart, Grmany,
116.
Shapley, H. (1920). Thermokinetics of Liometopum apiculatum Mayr. Physiology:
H. Shapley. Vol. 6. 206-211.
Shimizu, T., Thorn, K. S., Ruby, A. & Vale, R. D. (2000). ATPase Kinetic
Characterization and Single Molecule Behavior of Mutant Human Kinesin
Motors Defective in Microtubule-Based Motility. Biochemistry, 39, 5265-
5273.
Sihamala, O., Bhulaidok, S., Li-rong, S. & Duo, L., (2010). Lipids and Fatty Acid
Composition of Dried Edible Red and Black Ants. Agricultural Sciences in
China, 9, 1072-1077.
Sinha, D. K., Atray, I., Bentur, J. S., Nair, S. (2012). Expression of Orseolia
oryzae nucleoside diphosphate kinase (OoNDPK) is enhanced in rice gall
midge feeding on susceptible rice hosts and its over-expression leads to
salt tolerance in Escherichia coli. Insect Molecular Biology, 21, 593-603.
Sobierajska, K., Skurzynski, S., Stasiak, M., Kryczka, J., Cierniewski, C. S. &
Swiatkowska. (2014). Protein Disulfide Isomerase Directly Interacts with β-
Actin Cys374 and Regulates Cytoskeleton Reorganization. The Journal Of
Biological Chemistry, Vol. 289, No. 9, Pp. 5758–5773.

80
Soito, L., Williamson, C., Knutson, S. T., Fetrown, J. S., Poole, L. B. & Nelson, K.
J. (2011). PREX: PeroxiRedoxin classification indEX, a database of
subfamily assignments across the diverse peroxiredoxin family. Nucleic
Acids Research, Vol. 39, D332–D337.
Sommer, A., Christensen, E., Schwenger, S., Seul, R., Haas, D., Olbrich, H.,
Omran, H., Sass, J. O. (2011). The molecular basis of aminoacylase 1
deficiency. Biochimica et Biophysica Acta, 1812, 685–690.
Souza-Ferreira, P., Moreira, M. F., Atella, G. C., Oliveira-Carvalho, A. L.,
Eizemberg, R., Majerowicz, D., Melo, C. A., Zingali, R. B., Masuda, H.
(2014). Molecular characterization of Rhodnius prolixus’ embryonic cuticle.
Insect Biochemistry and Molecular Biology, 51, 89-100.
Stanley-Samuelson, D. W., Jurenka, R. A., Cripps, C., Blomquits, G. J. &
Renobales, M. (1988). REVIEW ARTICLE: Fatty Acids in Insects:
Composition, Metabolrsm, and Biological Significance. Archives of Insect
Biochemistry and Physiology 9, l-33.
Stekhoven, D. J., Omasits, U., Quebatte, M., Dehio, C., Ahrens, C. H. (2014).
Proteome-wide identification of predominant subcellular protein
localizations in a bacterial model organism. Journal of proteomics 99,
123–137.
T
Tarango-Aràmbula, L. A. (2012). Los escamoles y su producción en el altiplano
potosino-zacatecano. RESPYN “Revista Salud Pública y Nutrición”
Edición Especial No. 04 2012 pp 139 – 144.

81
Teets, N. M., Yi, S. X., Lee, R. E. & Delinger, D. L. (2013). Calcium signaling
mediates cold sensing in insect tissues. Proceedings of the National
Academy of the Sciences USA, 110, 9154-9159.
Tsape, K., Sinanoglou, V. J. & Miniadis-Meimaroglou, S. (2010). Comparative
analysis of the acid and sterol profiles of widely consumed Mediterranean
crustacean species. Food Chemistry, 122, 292-299.
U
Uvackova, L., Skultety, L., Bekesova, S., McClain, S., Hajducha, M. (2013). The
MSE-proteomic analysis of gliadins and glutenins in wheat grain identifies
and quantifies proteins associated with celiac disease and baker's asthma.
Journal of proteomics, 93, 65–73.
V
Van Abel, D., Hölzel, D. R., Jain, S., Lun, F. M. F., Zheng, Y. W. L., Chen, E. Z.,
Sun, H., Chiu, R. W. K., Lo, Y. M. D., van Dijk, M. & Oudejans, C. B. M.
(2011). SFRS7-Mediated Splicing of Tau Exon 10 Is Directly Regulated by
STOX1A in Glial Cells. PLoS ONE, 6(7), e21994.
van Huis A., van Itterbeeck J., Klunder H., Mertens E., Halloran A., Muir G.,
(2013). Edible insects: Future prospects for food and feed security. In:
FAO Forestry Paper 171. Rome, Italy: Food and Agriculture Organization
of the United Nations.
van Huis, A., Dicke, M. & van Loon, J. A. (2015). Insects to feed the world.
Journal of Insects as Food and Feed. 1(1): 3-5.

82
Vantomme, P. (2010). Los Insectos Forestales Comestibles, Una Fuente De
Proteínas Que Suele Pasar Por Alto. Unasylva 236, Vol.61. pp. 19
W
Walker, J., M. (2005). The proteomics protocols handbook. Editorial Humana
Press. Totowa. New Jersey. pp. 1,2.
Wang, R., Li, Q., Helfer, C. H., Jiao, J. & You, J. (2012). Bromodomain Protein
Brd4 Associated with Acetylated Chromatin Is Important for Maintenance
of Higher-order Chromatin Structure. The Journal Of Biological Chemistry,
Vol. 287, No. 14, Pp. 10738–10752.
Whitehead, T. A., Boonchai, B. B., Höllrigl, V. & Clark, D. S. (2007). A
filamentous molecular chaperone of the prefoldin family from the deep-sea
hyperthermophile Methanocaldococcus jannaschii. Protein Science,
16:626–634.
Win, D. T. (2005). Oleic Acid – The Anti-Breast Cancer Component in Olive Oil.
AU Journal of Technology, 9, 75-78.
Westermeier, R., Naven, T. (2002). Proteomics in practice. 3º. Edición, Editorial
Wiley-Vch. Germany. pp. 4, 5.
Wood, Z. A., Schroder, E., Harris, J. R.,Poole, L. B. (2003). Structure,
mechanism and regulation of peroxiredoxins. TRENDS in Biochemical
Sciences. Vol.28, No.1.

83
Y
Yang, L. F., Siriamornpun, S., Li, D. (2006). Polyunsaturated fatty acid content of
edible insects in Thailand. Journal of Food Lipids, 13, 277–285.
Yen, A. L. (2015). Insects as food and feed in the Asia Pacific region: current
perspectives and future directions. Journal of Insects as Food and Feed.
1(1): 33-55.
Yip, Y. Y., Yeap, Y. Y. C., Bogoyevitch, M. A., & Ng, D. C. H. (2014). cAMP-
dependent Protein Kinase and c-Jun N-terminal Kinase Mediate Stathmin
Phosphorylation for the Maintenance of Interphase Microtubules during
Osmotic Stress. The Journal Of Biological Chemistry Vol. 289, No. 4, Pp.
2157–2169.
Yoda, M., Cifuentes, D., Izumi, N., Sakaguchi, Y., Suzuli, T., Giraldez, A. J. &
Tomari, Y. (2013). Poly(A)-Specific Ribonuclease Mediates 3’-End
Trimming of Argonaute2-Cleaved Precursor MicroRNAs. Cell Reports, 5,
715-726.
Z
Zhang, Y., Zhu, T., Wang, L., Pan, Y. H. & Zhang, S. (2013). Homocysteine
Homeostasis and Betaine-Homocysteine S-Methyltransferase Expression
in the Brain of Hibernating Bats. PLos One, 8(12), e856332.
Zhang, S., Li, F., Younes, M., Liu, H., Chen, C., Yao, Q. (2013). Reduced
Selenium-Binding Protein 1 in Breast Cancer Correlates with Poor Survival
and Resistance to the Anti-Proliferative Effects of Selenium. PLoS One, 8,
e63702.

84
Zhang, Y. L., Xue, R. Y., Cao, G. L., Zhu, J. X., Pan, Z. H., Gong, C. L. (2013).
Shotgun proteomic analysis of wing discs from the domesticated silkworm
(Bombyx mori) during metamorphosis. Amino Acids, 45, 1231–1241.
Zhang, Y., Wang, X., Xiang, Z., Li, H., Qiu, J., Sun, Q., Wan T., Li, N., Cao, X. &
Wang, J. (2007). Promotion of cellular migration and apoptosis resistance
by a mouse eye-specific phosphatidylethanolamine-binding protein.
International Journal Of Molecular Medicine, 19, 55-63.

85
ANEXO 1
Soluciones Usadas En Proteómica
1. Solución de rehidratación sin IPG Buffer.
Urea 8M, CHAPS 2%, Azul de bromofenol 0.02%.
Reactivo Concentración final Cantidad
Urea 8 M 12 g CHAPS 2% (w/v) 0.5 g Azul de bromofenol al 1% 0.002% (w/v) 50 μL Agua Milli-Q c.b.p. 25 mL
Agregar antes de usar: 0.56% (w/v) de DTT y 0.5% (w/v) de IPG Buffer.
2. Buffer de equilibrio SDS.
Reactivo Concentración final Cantidad
Tris 1.5M pH 8.8 50 mM 10 mL Urea 6 M 72.07 g Glicerol 30% (v/v) 60 mL SDS 2% (v/v) 4.0 g Azul de bromofenol al 1% 0.002% (p/v) 400 μL Agua Milli-Q c.b.p. 200 mL
Agregar antes de usar: 1% (p/v) de DTT ó 2.5% (p/v) de Iodacetamida por
cada mililitro de solución de equilibrio.
3. Buffer de electroforesis
Reactivo Concentración final Cantidad
Tris-base (PM 121.1) 25 mM 30.275 g Glicina (PM 75.07) 192 mM 144.134 g SDS (PM 288.38) 0.1% (p/v) 10 g Agua Milli-Q c.b.p. 10 L
4. Gel de poliacrilamida desnaturalizante al 13% para 2-DE de tira IPG
de 24cm (separador).
Reactivo Cantidad
Acrilamida 35.83 mL Agua Milli-Q 23.33 mL

86
Tris 1.5 M pH 8.8 20.83 mL SDS 10% 0.83 mL PSA 10% 66.67 μL TEMED 66.67 μL
5. Gel de poliacrilamida desnaturalizante al 4% para 2-DE de tiras IPG
de 24cm (concentrador).
Reactivo Cantidad
Acrilamida 0.765 mL Agua Milli-Q 3.483 mL Tris 1.5 M pH 8.8 1.45 mL SDS 10% 58.15 μL PSA 10% 29.02 μL TEMED 2.90 μL
6. Solución de sellado.
Reactivo Concentración final Cantidad
Buffer de electroforesis
Tris 25 mM, Glicina 192 mM, SDS 0.1%
100 mL
Agarosa 0.5% 0.5 g Azul de bromofenol 1%
0.002% 200 μL
7. Solución de azul de Coomasie R-250 al 0.025%.
Reactivo Concentración final Cantidad
Azul de Coomasie R-250 0.025% (p/v) 250 mg Metanol 40% (v/v) 400 mL Ácido acético glacial 10% (v/v) 100 mL Agua Milli-Q c.b.p. 1L
8. Solución de fijado y desteñido.
Reactivo Concentración final Cantidad
Metanol 30 % 300 mL Ácido acético glacial 10 % 100 mL Agua Milli-Q c.b.p. 1L
9. Bicarbonato de amonio (NH4HCO3)100nM.
Reactivo Concentración final Cantidad

87
NH4HCO3 100mM 79 mg Agua Milli-Q c.b.p. 10mL
10. Solución de digestión con tripsina.
Reactivo Cantidad
Tripsina (0.1µg/μL) 5 μL NH4HCO3 50 mM 45 μL

88
ANEXO 2
Soluciones Usadas Análisis de Lípidos
1. Solución de hidróxido de sodio 1N.
Reactivo Cantidad
NaOH 0.1g Metanol c.b.p. 5 mL
2. Solución de N,O-Bis(trimethylsilyl) trifluoroacetamide ≥99%,
Reactivo Cantidad
N,O-Bis(trimethylsilyl) trifluoroacetamide 900 μL Chlorotrimethylsilane 100 μL

89

90

91

92





























