Embed Size (px)
Citation preview
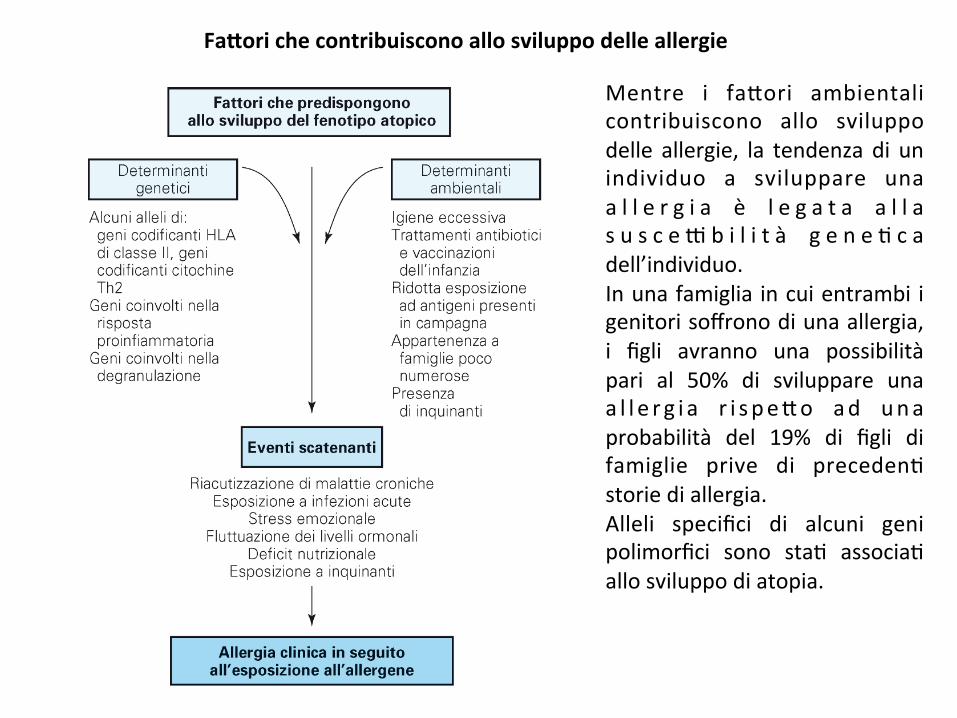
Fa#orichecontribuisconoallosviluppodelleallergie
Mentre i fa*ori ambientalicontribuiscono allo sviluppodelle allergie, la tendenza di unindividuo a sviluppare unaa l l e r g i a è l e g a t a a l l as u s c e 9 b i l i t à g e n e ; c adell’individuo.Inunafamiglia incuientrambi igenitorisoffronodiunaallergia,i figli avranno una possibilitàpari al 50% di sviluppare unaa l l e r g i a r i s pe*o ad unaprobabilità del 19% di figli difamiglie prive di preceden;storiediallergia.Alleli specifici di alcuni genipolimorfici sono sta; associa;allosviluppodiatopia.
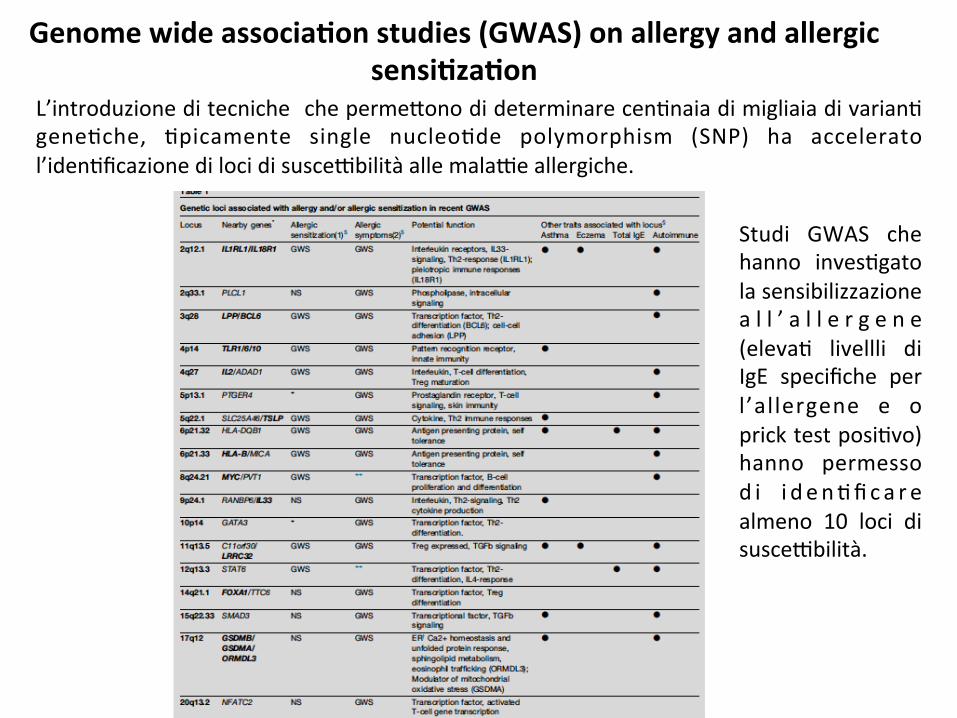
Genomewideassocia8onstudies(GWAS)onallergyandallergicsensi8za8on
L’introduzioneditecniche cheperme*onodideterminarecen;naiadimigliaiadivarian;gene;che, ;picamente single nucleo;de polymorphism (SNP) ha acceleratol’iden;ficazionedilocidisusce9bilitàallemala9eallergiche.
Studi GWAS chehanno inves;gatolasensibilizzazionea l l ’ a l l e r g e n e(eleva; livellli diIgE specifiche perl’allergene e opricktestposi;vo)hanno permessod i i d en;fica r ealmeno 10 loci disusce9bilità.
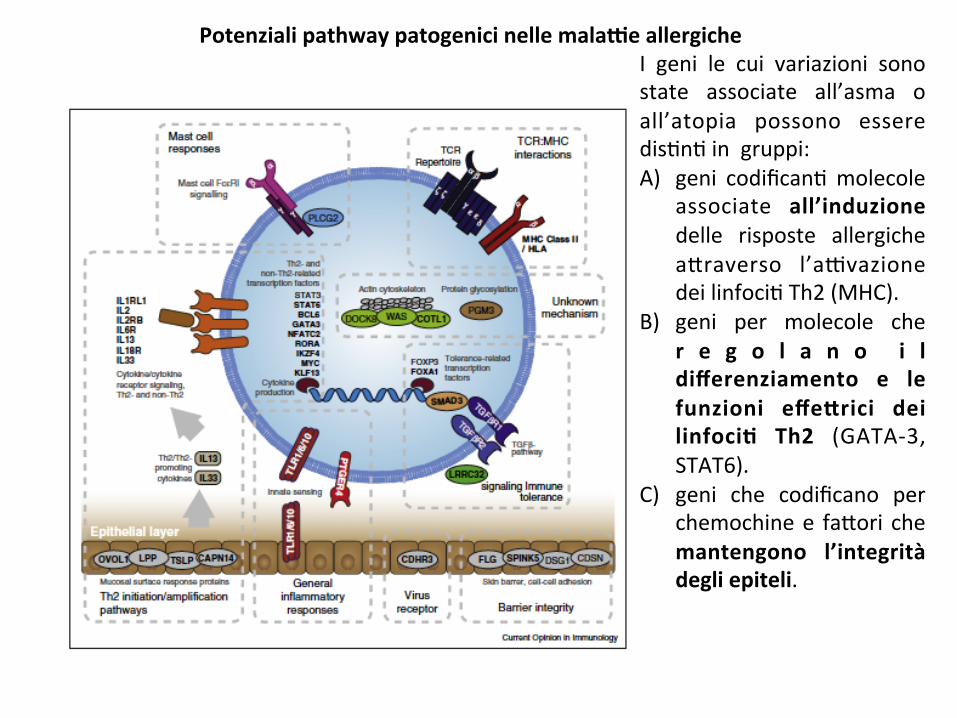
I geni le cui variazioni sonostate associate all’asma oall’atopia possono esseredis;n;ingruppi:A) geni codifican;molecole
associate all’induzionedelle risposte allergichea*raverso l’a9vazionedeilinfoci;Th2(MHC).
B) geni per molecole cher e g o l a n o i ldifferenziamento e lefunzioni effe#rici deilinfoci8 Th2 (GATA-3,STAT6).
C) geni che codificano perchemochinee fa*ori chemantengono l’integritàdegliepiteli.
PotenzialipathwaypatogenicinellemalaGeallergiche

Gene8cadellemalaGeallergicheedellamarciaatopica
Studi gene;ci hanno dimostrato l’importanza dei dife9 della barrieraepiteliale nello sviluppo della derma;te atopica e di altre mala9eallergiche.In par;colare mutazioni in geni che codificano per importan8componen8 della barriera epiteliale (filaggrin, serin pep;dasi inhibitorKazal-type 5 e corneodesmosin) sono state associate allo sviluppo diderma8teatopica.Mutazionidellafilaggrinacausanol’iGosivolgarecheèundisordinedellachera;nizzazione della pelle; mutazioni di SPINK5 e CDSN causanosindromidaesfoliazionedellapelle.
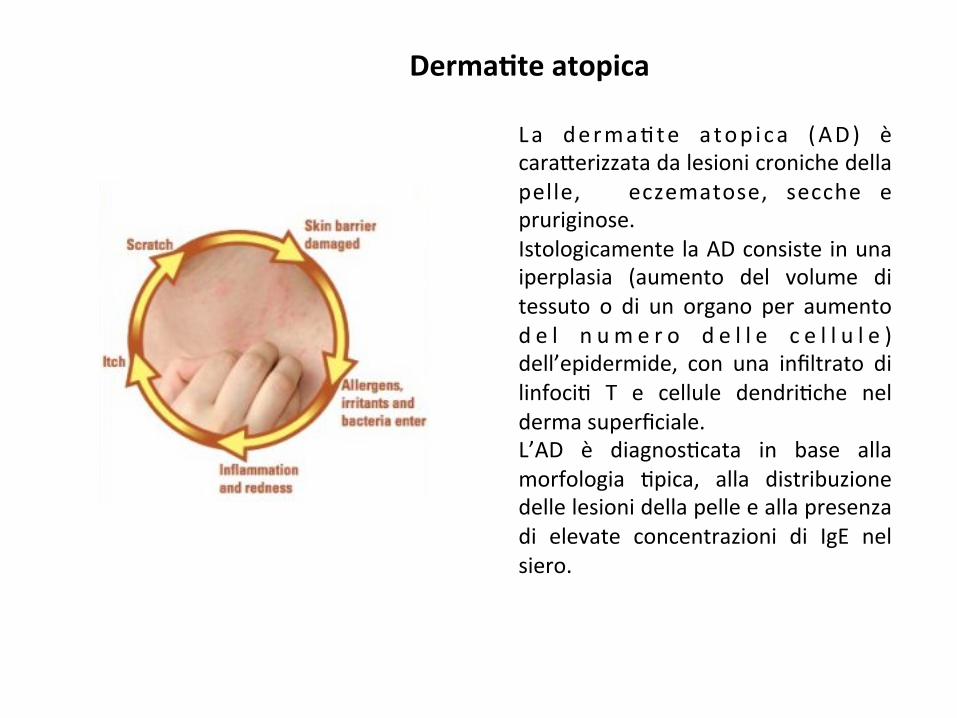
Derma8teatopica
La derma;te a top i ca (AD) ècara*erizzatadalesionicronichedellapelle, eczematose, secche epruriginose.Istologicamente laADconsiste inunaiperplasia (aumento del volume ditessuto o di un organo per aumentod e l n u m e r o d e l l e c e l l u l e )dell’epidermide, con una infiltrato dilinfoci; T e cellule dendri;che neldermasuperficiale.L’AD è diagnos;cata in base allamorfologia ;pica, alla distribuzionedellelesionidellapelleeallapresenzadi elevate concentrazioni di IgE nelsiero.
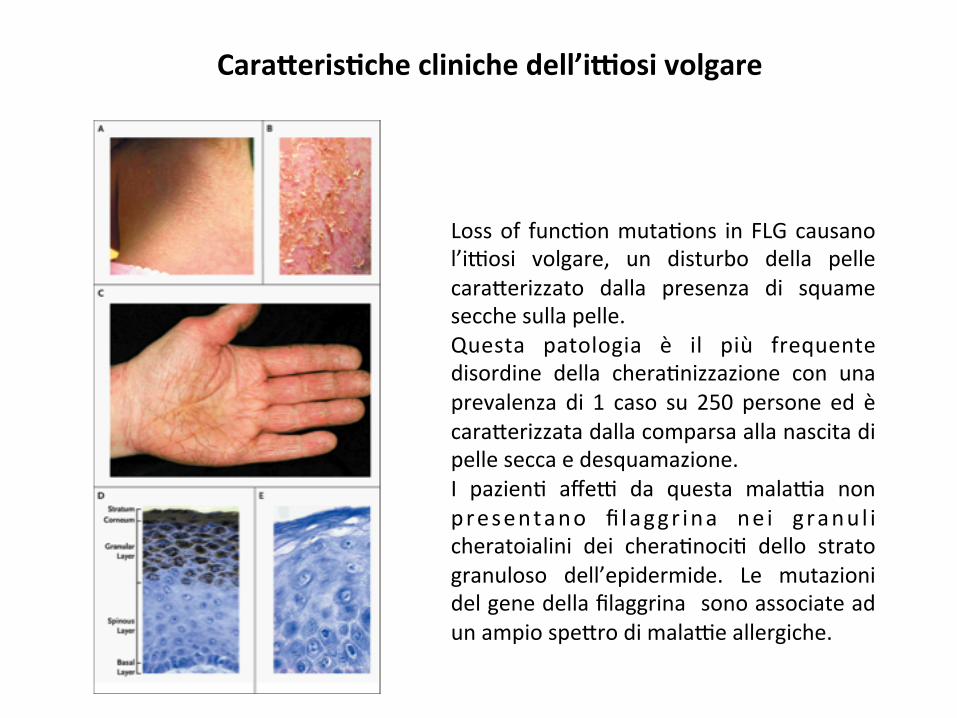
Cara#eris8checlinichedell’iGosivolgare
Lossof func;onmuta;ons inFLGcausanol’i9osi volgare, un disturbo della pellecara*erizzato dalla presenza di squamesecchesullapelle.Questa patologia è il più frequentedisordine della chera;nizzazione con unaprevalenza di 1 caso su 250 persone ed ècara*erizzatadallacomparsaallanascitadipelleseccaedesquamazione.I pazien; affe9 da questa mala9a nonpresentano fi laggr ina ne i g ranu l icheratoialini dei chera;noci; dello stratogranuloso dell’epidermide. Le mutazionidelgenedellafilaggrina sonoassociateadunampiospe*rodimala9eallergiche.
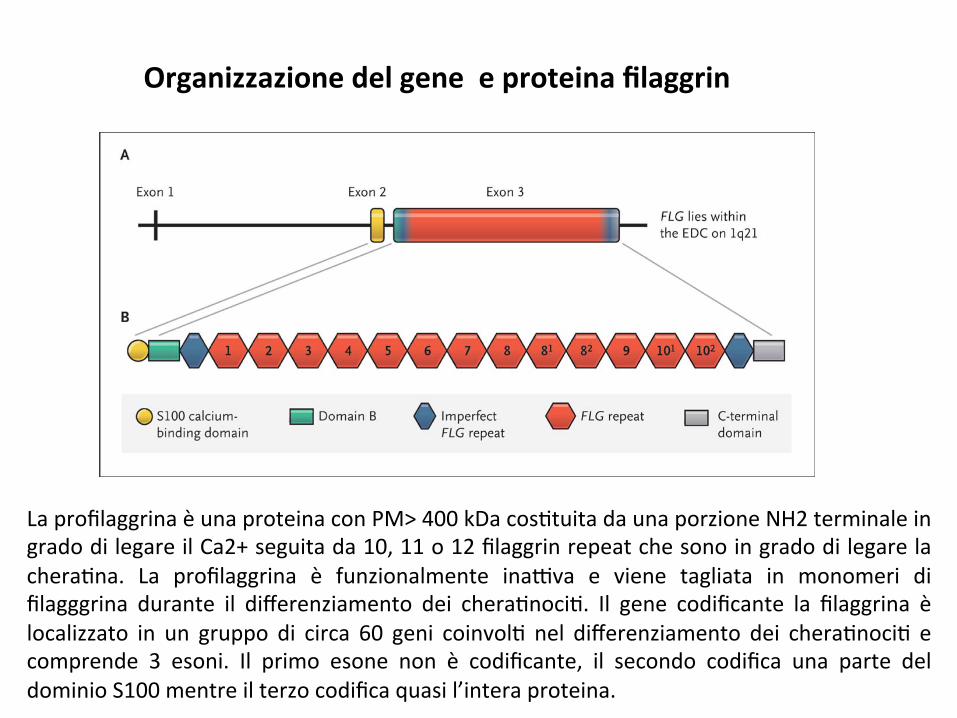
Organizzazionedelgeneeproteinafilaggrin
LaprofilaggrinaèunaproteinaconPM>400kDacos;tuitadaunaporzioneNH2terminaleingradodilegareilCa2+seguitada10,11o12filaggrinrepeatchesonoingradodilegarelachera;na. La profilaggrina è funzionalmente ina9va e viene tagliata in monomeri difilagggrina durante il differenziamento dei chera;noci;. Il gene codificante la filaggrina èlocalizzato in un gruppo di circa 60 geni coinvol; nel differenziamento dei chera;noci; ecomprende 3 esoni. Il primo esone non è codificante, il secondo codifica una parte deldominioS100mentreilterzocodificaquasil’interaproteina.
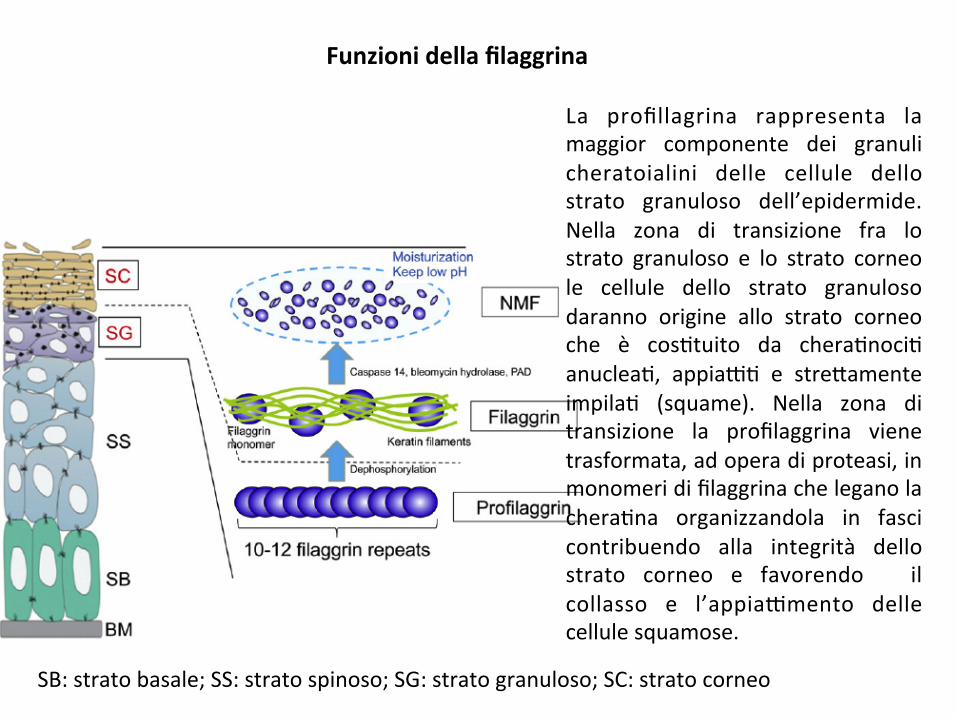
Funzionidellafilaggrina
La profillagrina rappresenta lamaggior componente dei granulicheratoialini delle cellule dellostrato granuloso dell’epidermide.Nella zona di transizione fra lostrato granuloso e lo strato corneole cellule dello strato granulosodaranno origine allo strato corneoche è cos;tuito da chera;noci;anuclea;, appia9; e stre*amenteimpila; (squame). Nella zona ditransizione la profilaggrina vienetrasformata,adoperadiproteasi,inmonomeridifilaggrinacheleganolachera;na organizzandola in fascicontribuendo alla integrità dellostrato corneo e favorendo ilcollasso e l’appia9mento dellecellulesquamose.
SB:stratobasale;SS:stratospinoso;SG:stratogranuloso;SC:stratocorneo
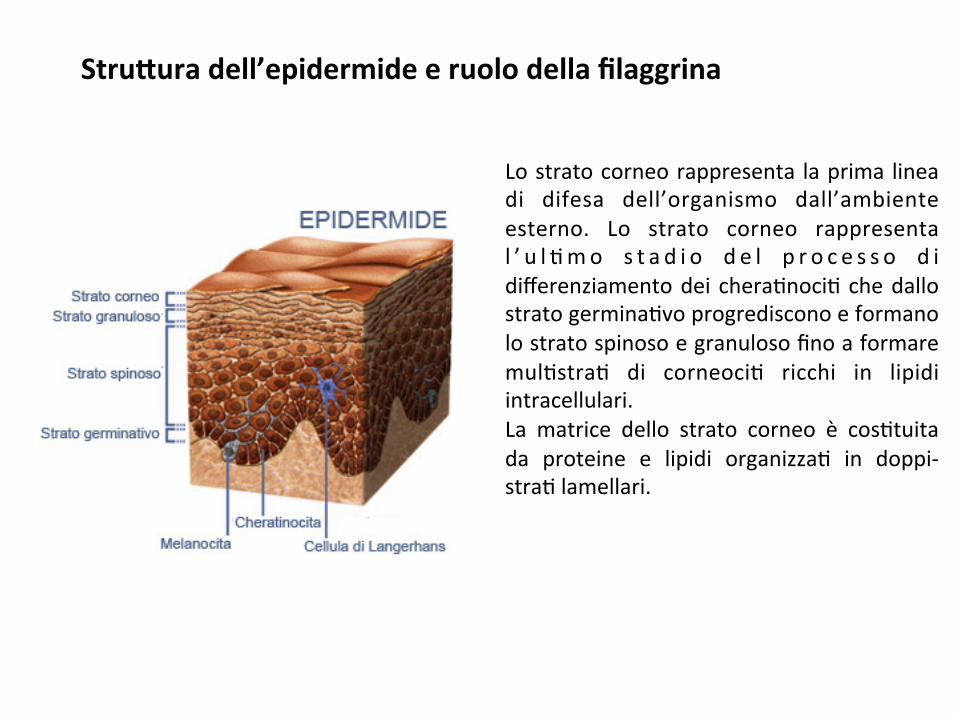
Stru#uradell’epidermideeruolodellafilaggrina
Lostratocorneorappresenta laprima lineadi difesa dell’organismo dall’ambienteesterno. Lo strato corneo rappresental ’ u l ;mo s t a d i o d e l p r o c e s s o d idifferenziamentodei chera;noci;chedallostratogermina;voprogredisconoeformanolostratospinosoegranulosofinoaformaremul;stra; di corneoci; ricchi in lipidiintracellulari.La matrice dello strato corneo è cos;tuitada proteine e lipidi organizza; in doppi-stra;lamellari.
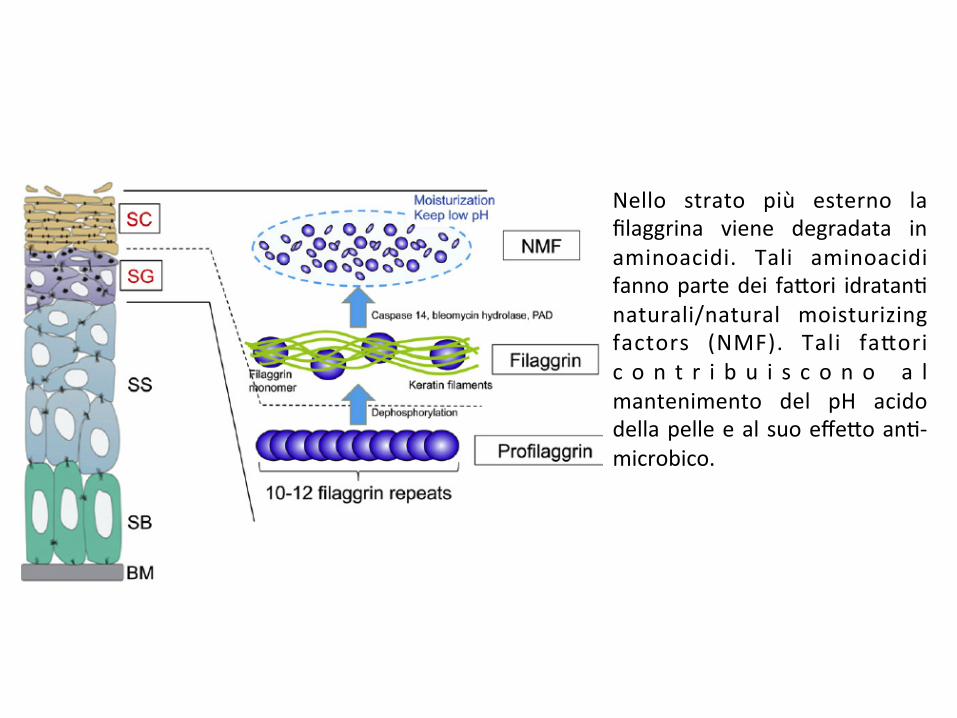
Nello strato più esterno lafilaggrina viene degradata inaminoacidi. Tali aminoacidifannopartedei fa*ori idratan;naturali/natural moisturizingfactors (NMF). Tali fa*oric o n t r i b u i s c o n o a lmantenimento del pH acidodellapelleealsuoeffe*oan;-microbico.
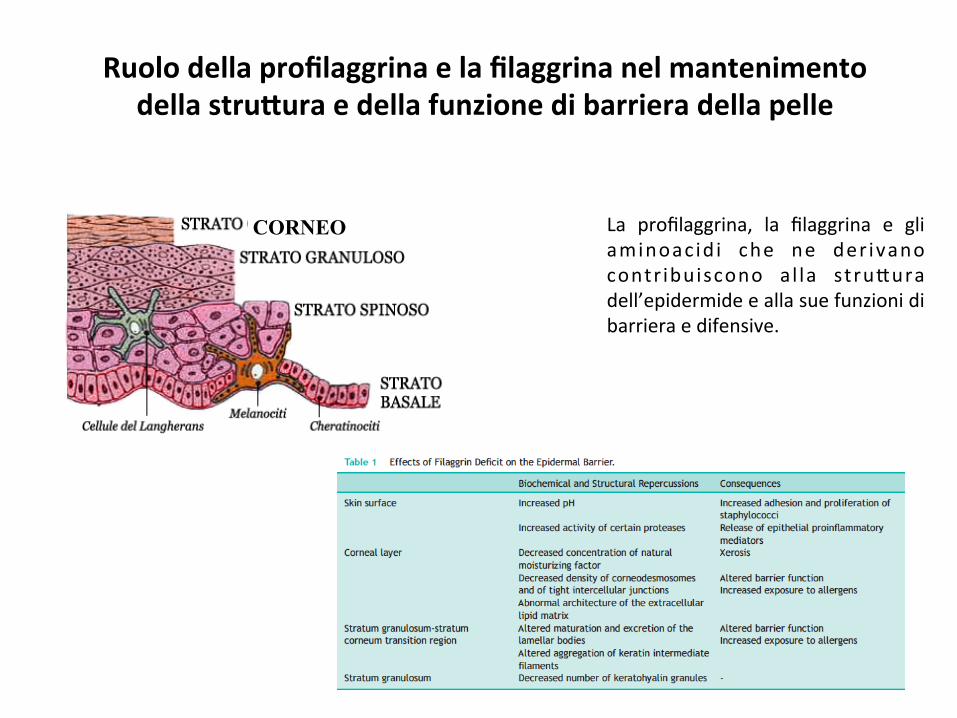
Ruolodellaprofilaggrinaelafilaggrinanelmantenimentodellastru#uraedellafunzionedibarrieradellapelle
La profilaggrina, la filaggrina e gliaminoac id i che ne der ivanocontr ibuiscono al la stru*uradell’epidermideeallasuefunzionidibarrieraedifensive.
CORNEO

Mutazioninellafilaggrinaaumentanoilrischiodisvilupparederma8teatopica
• Nel 2006 iden;ficazionedi mutazioni
loss of func;on nel gene FLG(filaggrina) in individuiaffe9da ic;osivolgare(IV).
• Moltedellefamiglieirlandesiaffe*eda
IV mostravano una alta incidenza dimala9e allergiche come derma;teatopica,febbredafieno,asma.
• Dimostrazione che la presenza dimutazioni loss of func;on della FLGconferiscono un alto rischio disviluppare AD. Questa associazione èstatapoiconfermatainaltri30studi.
• Nel modello murino “flaky tail”deficiente nella filaggrina è statodimostrato che la deficienza infilaggrina perme*e il passaggiodell’allergene a*raverso l’epidermideinducendo una risposta immune Th2.Ques; studi hanno confermatol’ipotesichel’alterazionedellabarrieraepiteliale induce risposte Th2 versol’allergene.

Mutazionidellafilaggrinasonoassociateallosviluppodiderma8teatopica
• mutazioninelgenedellafilaggrinaumanorappresentanoilmaggiorefa*oredirischionellosviluppodiderma;teatopica.
• Laderma;teatopicacolpiscecircal’11%deibambininegliSta;Uni;ecircail25%nelRegnoUnito.
• Laderma;teatopicaècara*erizzatadaalterazionidellabarrieraepidermica,infiammazione cutanea,presenzadi una risposta immunedi;po2 sistemicafrequentecolonizzazionedapartediStaphylococcusaureus.
• Laderma;teatopicaè lapiù frequentemala9a infiammatoriacronica della
prima infanzia e rappresenta spesso lo stadio iniziale di quella che vienedefinita”atopicmarch”chesimanifestacon ilsuccessivosviluppodiallergie,allergieadalimen;,rini;allergicheeasmaallergico.
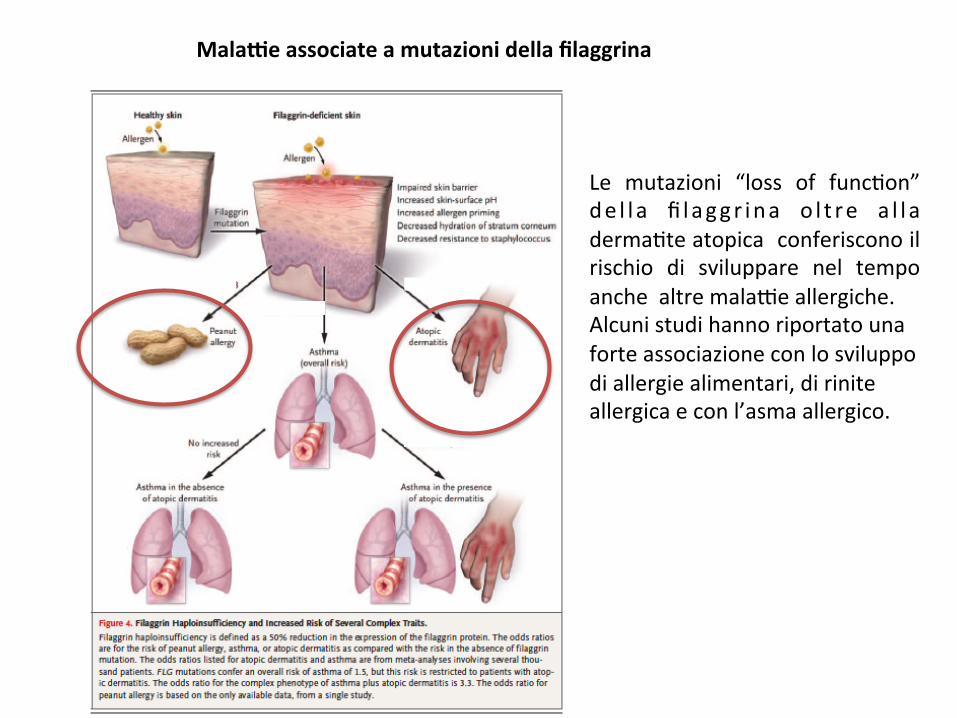
MalaGeassociateamutazionidellafilaggrina
Le mutazioni “loss of func;on”de l l a fi laggr ina o l t re a l l aderma;teatopicaconferisconoilrischio di sviluppare nel tempoanchealtremala9eallergiche.Alcunistudihannoriportatounaforteassociazioneconlosviluppodiallergiealimentari,diriniteallergicaeconl’asmaallergico.
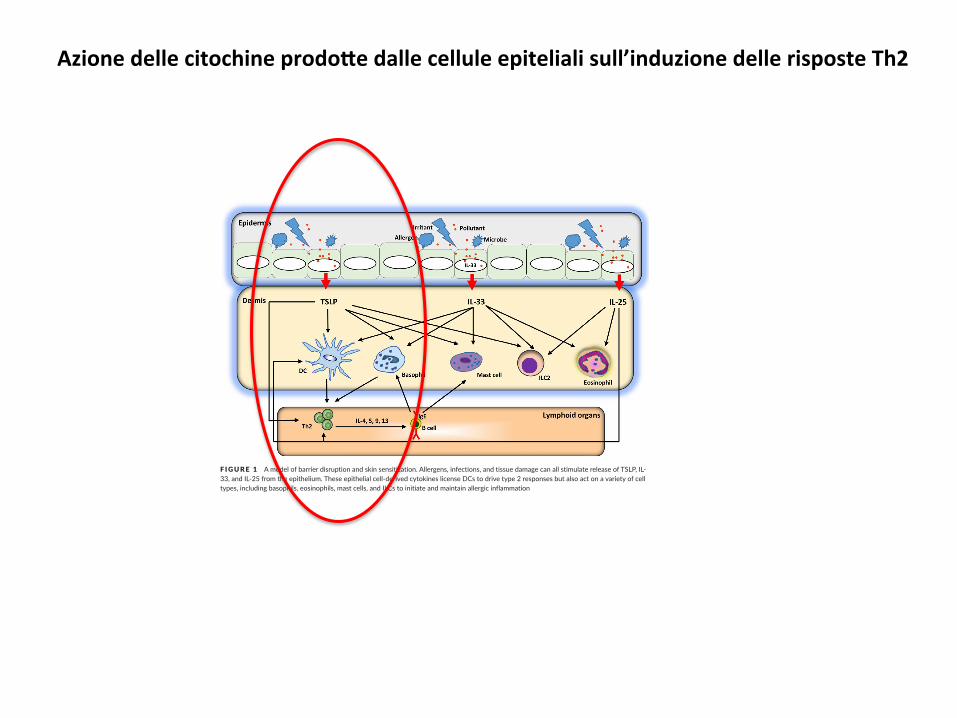
Azionedellecitochineprodo#edallecelluleepitelialisull’induzionedellerisposteTh2
122 | HAN et Al.
pathways interact with one another and the extent to which they are functionally distinct. Some studies indicate that there is some cross- regulation at the level of gene expression. These cytokines also show different expression kinetics, suggesting that they may be involved in driving disease at different stages of inflammation. In a T. muris infec-tion model, the kinetics of TSLP expression lagged that seen for IL- 33, and intraperitoneal injection of recombinant IL- 33 induced cecal ex-pression of TSLP and TSLPR mRNA.189 TSLP mRNA can also be up-regulated following epithelial cell- specific overexpression of IL- 25 in the lung.167 In a model of chronic HDM exposure, IL- 33 mRNA levels were elevated at early and late time points, whereas TSLP mRNA was only upregulated at early time points, and IL- 25 mRNA was upregu-lated only at late time points.190 Reporter mice for TSLP,191 IL- 25,176 and IL- 33 157 may help define the cell- type- specific expression kinet-ics of these cytokines.
Models of allergic inflammation have shown distinct requirements for TSLP, IL- 33, and IL- 25. After mice were exposed to intranasal HDM or intragastric peanut, IL- 33 but not TSLP or IL- 25 was required to drive inflammation.192 In these models, IL- 33 induced a robust induction of OX40L expression on DCs and expansion of ILCs. Consistent with the findings by Chu and colleagues, IL- 33 but not IL- 25 was found to be required for airway hyperreactivity after intranasal ragweed challenge, which robustly increased the expression of IL- 33, but not IL- 25, and expanded IL- 13- producing ILC2 in the lung.162 We have found that the TSLP- TSLPR axis is not required in IL- 33- driven food allergy; in
contrast, mice lacking ST2 or IL- 17RB had attenuated disease in TSLP- driven food allergy 105 (H. Han and SF. Ziegler, unpublished data). IL- 33 may play a critical role in amplifying type 2 responses initiated by TSLP, as TSLP induces the accumulation of many inflammatory cells that ex-press the IL- 33 receptor and respond to IL- 33.123
In some cases, different mouse strains have been shown to have distinct cytokine requirements. Several studies have shown that ILC2s are important in allergic responses in the skin and accumulate in the skin of individuals with atopic dermatitis; however, studies in mouse models have demonstrated striking differences in cytokine depen-dence in C57BL/6 and BALB/c mouse strains. MC903- driven skin in-flammation in C57BL/6 mice was dependent on TSLP but not on IL- 33 or IL- 25,184 whereas inflammation and ILC2 accumulation in the skin in BALB/c mice were only mildly affected by lack of TSLP and were instead dependent on IL- 25.145 BALB/c mice lacking ST2 had an in-termediate phenotype between TSLPR knockouts and IL- 17RB knock-outs in MC903- driven disease.145 We have not detected induction of IL- 25 or IL- 33 mRNA after topical application of MC903 in BALB/c mice (H. Han and SF. Ziegler, unpublished data); additional studies will be required to determine whether IL- 25 and IL- 33 may act down-stream of TSLP or may be important in amplifying TSLP- mediated Th2 responses in this model.
The type of allergen and the allergen dose and duration of expo-sure may also affect the requirements for TSLP, IL- 33, and IL- 25 in al-lergic responses. In a chronic model of HDM lung inflammation, triple
F IGURE 1 A model of barrier disruption and skin sensitization. Allergens, infections, and tissue damage can all stimulate release of TSLP, IL- 33, and IL- 25 from the epithelium. These epithelial cell- derived cytokines license DCs to drive type 2 responses but also act on a variety of cell types, including basophils, eosinophils, mast cells, and ILCs to initiate and maintain allergic inflammation

• Apar;redal‘900l’incidenzadellemala9eatopicheènotevolmenteaumentataneipaesimaggiormenteindustrializza;.
• Per spiegare tale aumento è stata proposta “l’ipotesi dell’igiene”. Tale teoria
suggerisce che l’esposizione precoce ad agen; infe9vi o microbici svolga unruoloprote9voneiconfron;dellosviluppodellemala9eallergiche.
• Nel 1989 uno studio aveva dimostrato una relazione inversa fra il numero difratellieiltassodimala9eallergicheinunindividuo.Proponendocheunrido*onumerodifratellieal;standardigienicideterminasserounariduzionedicross-infezioniconconseguenteaumentodellosviluppodimala9eallergiche.
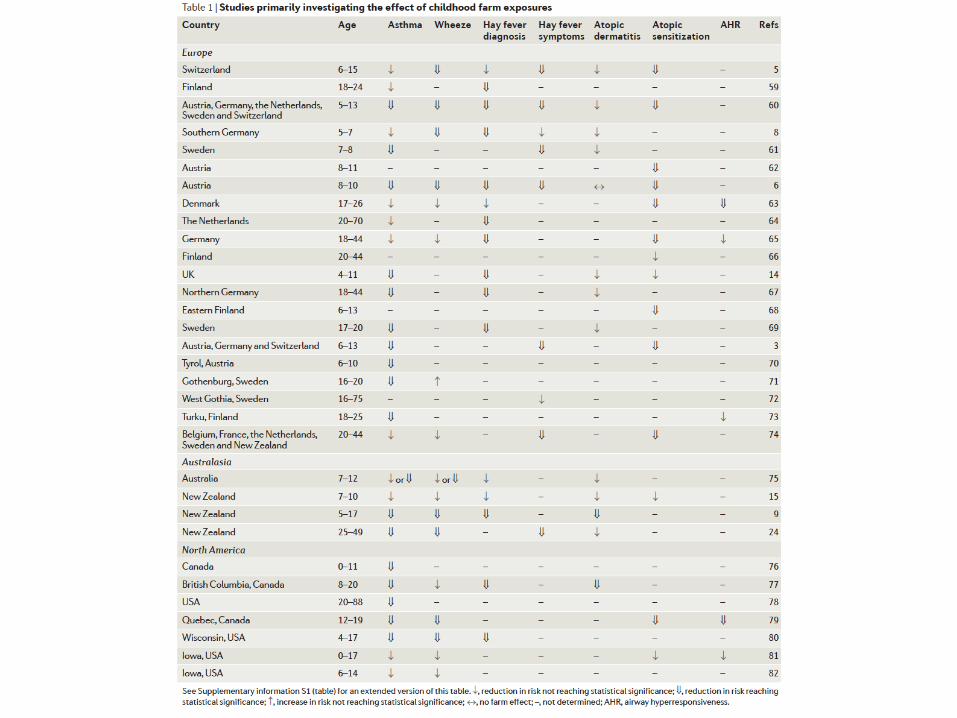
• Ida;amaggiorsostegnodellaTeoriadell’Igienederivanodastudisuibambinicresciu; nelle fa*orie chedimostrano come crescere in tali ambien;, dove èaumentatal’esposizioneaimicrorganismi,proteggedallosviluppodiasma.
Fa#oriambientalinellosviluppodellemalaGeatopiche
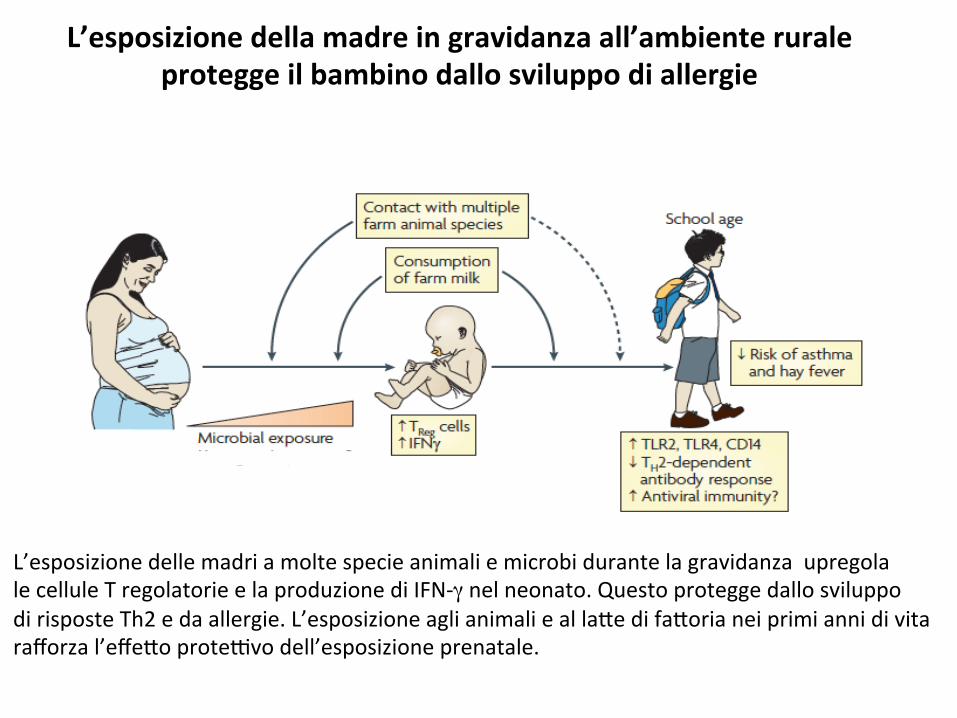
L’esposizionedellamadreingravidanzaall’ambienteruraleproteggeilbambinodallosviluppodiallergie
L’esposizionedellemadriamoltespecieanimaliemicrobidurantelagravidanzaupregolalecelluleTregolatorieelaproduzionediIFN-γnelneonato.QuestoproteggedallosviluppodirisposteTh2edaallergie.L’esposizioneaglianimaliealla*edifa*orianeiprimiannidivitarafforzal’effe*oprote9vodell’esposizioneprenatale.
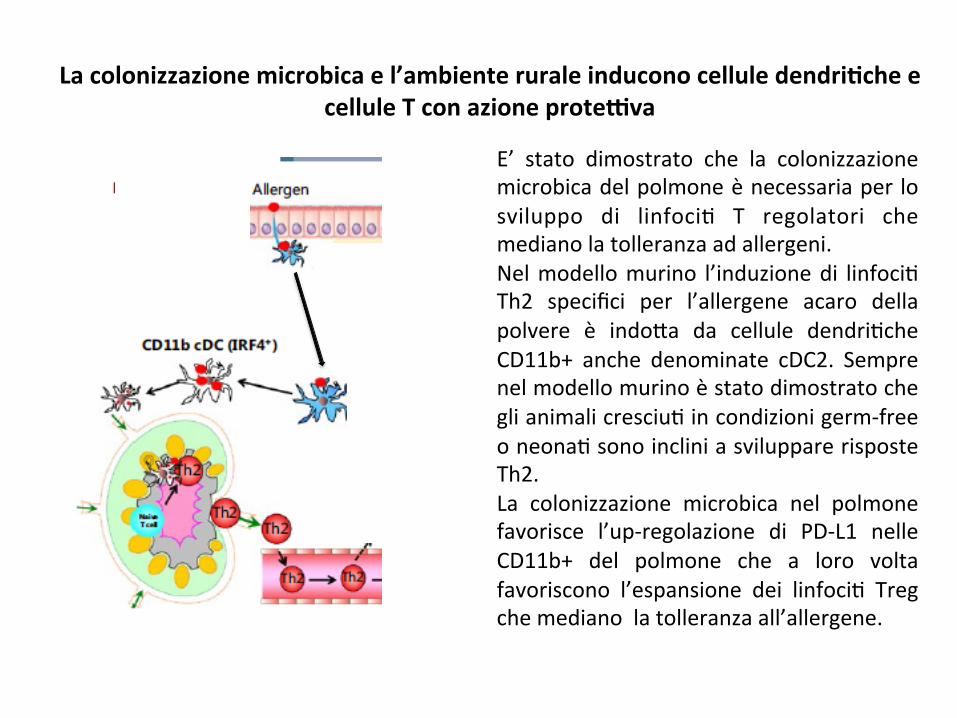
Lacolonizzazionemicrobicael’ambienteruraleinduconocelluledendri8cheecelluleTconazioneproteGva
E’ stato dimostrato che la colonizzazionemicrobicadelpolmoneènecessariaper losviluppo di linfoci; T regolatori chemedianolatolleranzaadallergeni.Nelmodellomurino l’induzionedi linfoci;Th2 specifici per l’allergene acaro dellapolvere è indo*a da cellule dendri;cheCD11b+ anche denominate cDC2. Semprenelmodellomurinoèstatodimostratocheglianimalicresciu;incondizionigerm-freeoneona;sonoincliniasvilupparerisposteTh2.La colonizzazione microbica nel polmonefavorisce l’up-regolazione di PD-L1 nelleCD11b+ del polmone che a loro voltafavoriscono l’espansione dei linfoci; Tregchemedianolatolleranzaall’allergene.
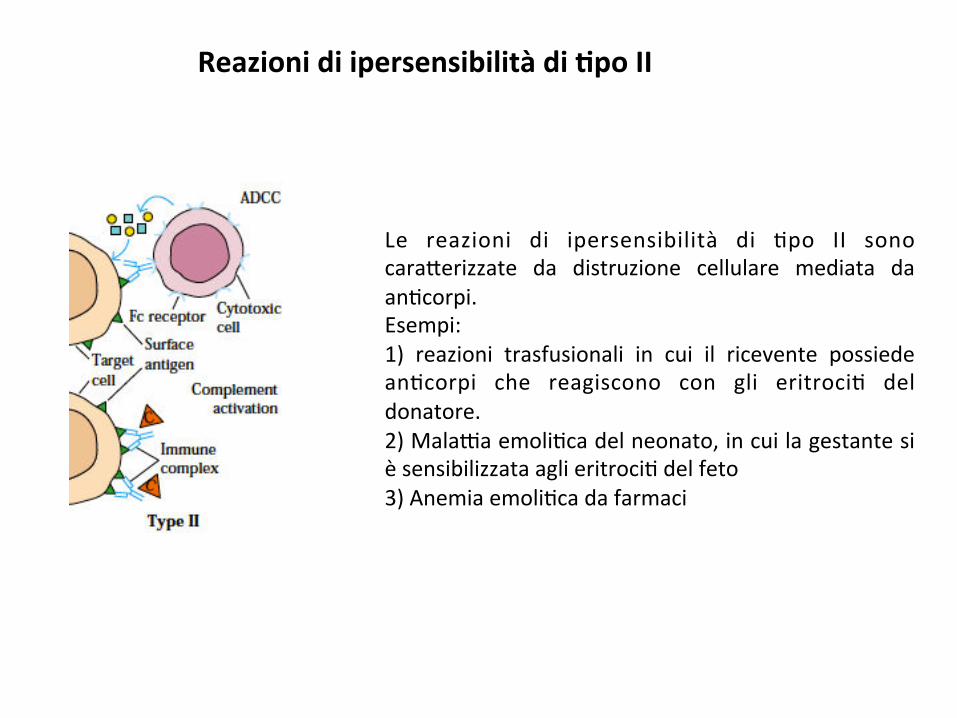
Le reazioni di ipersensibilità di ;po II sonocara*erizzate da distruzione cellulare mediata daan;corpi.Esempi:1) reazioni trasfusionali in cui il ricevente possiedean;corpi che reagiscono con gli eritroci; deldonatore.2)Mala9aemoli;cadelneonato,incuilagestantesièsensibilizzataaglieritroci;delfeto3)Anemiaemoli;cadafarmaci
Reazionidiipersensibilitàdi8poII
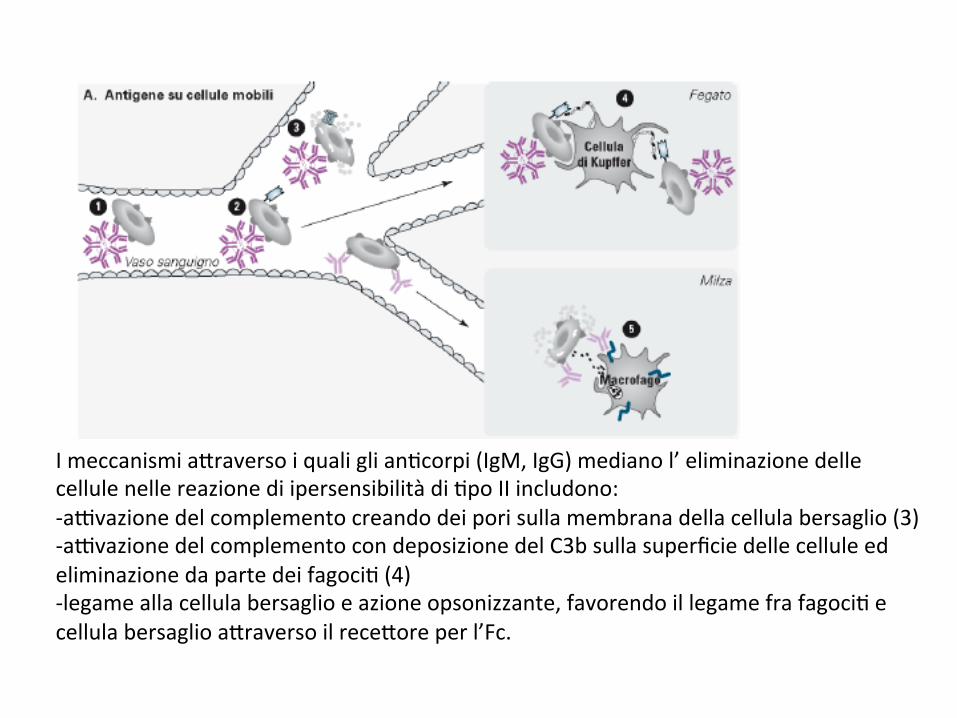
Imeccanismia*raversoiqualiglian;corpi(IgM,IgG)medianol’eliminazionedellecellulenellereazionediipersensibilitàdi;poIIincludono:-a9vazionedelcomplementocreandodeiporisullamembranadellacellulabersaglio(3)-a9vazionedelcomplementocondeposizionedelC3bsullasuperficiedellecelluleedeliminazionedapartedeifagoci;(4)-legameallacellulabersaglioeazioneopsonizzante,favorendoillegamefrafagoci;ecellulabersaglioa*raversoilrece*oreperl’Fc.
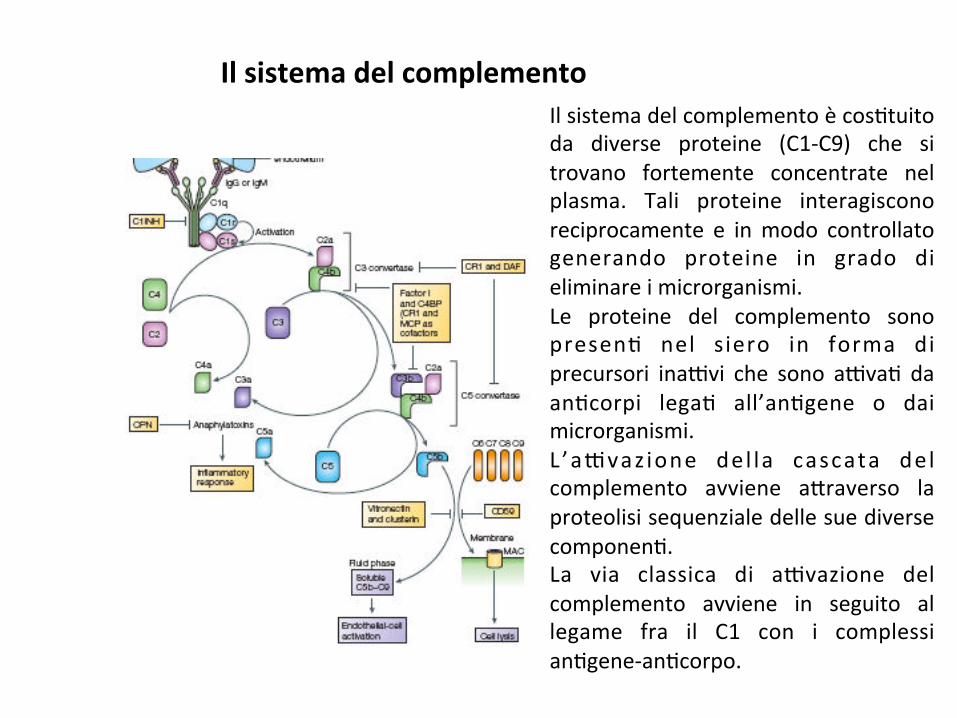
Ilsistemadelcomplementoècos;tuitoda diverse proteine (C1-C9) che sitrovano fortemente concentrate nelplasma. Tali proteine interagisconoreciprocamente e inmodo controllatogenerando proteine in grado dieliminareimicrorganismi.Le proteine del complemento sonopresen; nel siero in forma diprecursori ina9vi che sono a9va; daan;corpi lega; all’an;gene o daimicrorganismi.L ’a9vaz ione del la cascata delcomplemento avviene a*raverso laproteolisisequenzialedellesuediversecomponen;.La via classica di a9vazione delcomplemento avviene in seguito allegame fra il C1 con i complessian;gene-an;corpo.
Ilsistemadelcomplemento
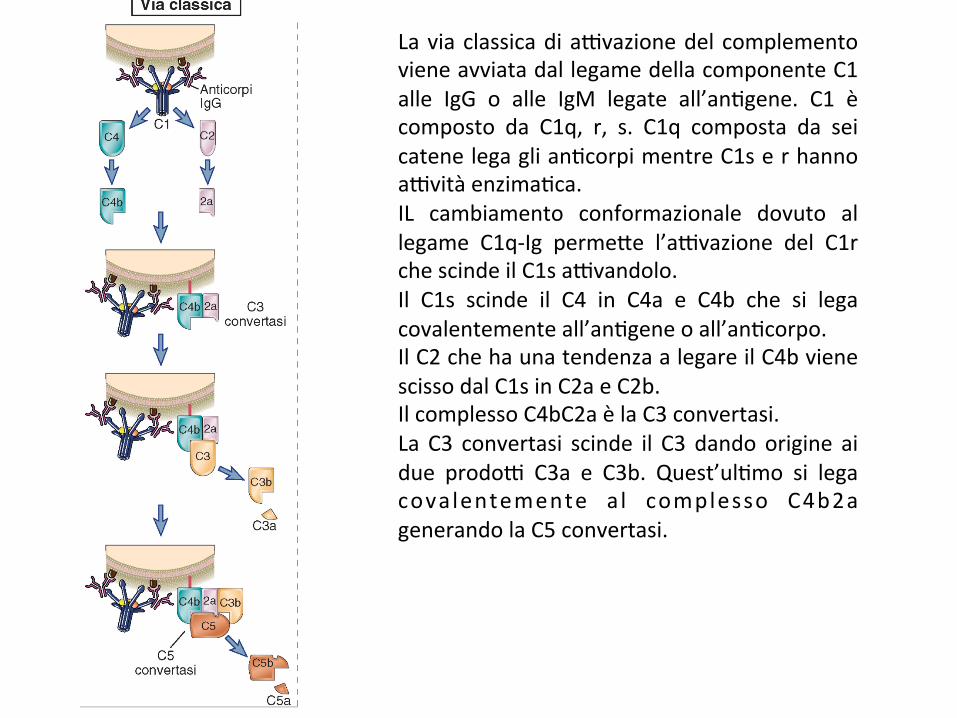
Laviaclassicadia9vazionedelcomplementovieneavviatadallegamedellacomponenteC1alle IgG o alle IgM legate all’an;gene. C1 ècomposto da C1q, r, s. C1q composta da seicatenelegaglian;corpimentreC1serhannoa9vitàenzima;ca.IL cambiamento conformazionale dovuto allegame C1q-Ig perme*e l’a9vazione del C1rchescindeilC1sa9vandolo.Il C1s scinde il C4 in C4a e C4b che si legacovalentementeall’an;geneoall’an;corpo.IlC2chehaunatendenzaalegareilC4bvienescissodalC1sinC2aeC2b.IlcomplessoC4bC2aèlaC3convertasi.La C3 convertasi scinde il C3 dandoorigine aidue prodo9 C3a e C3b. Quest’ul;mo si legacovalentemente a l complesso C4b2agenerandolaC5convertasi.
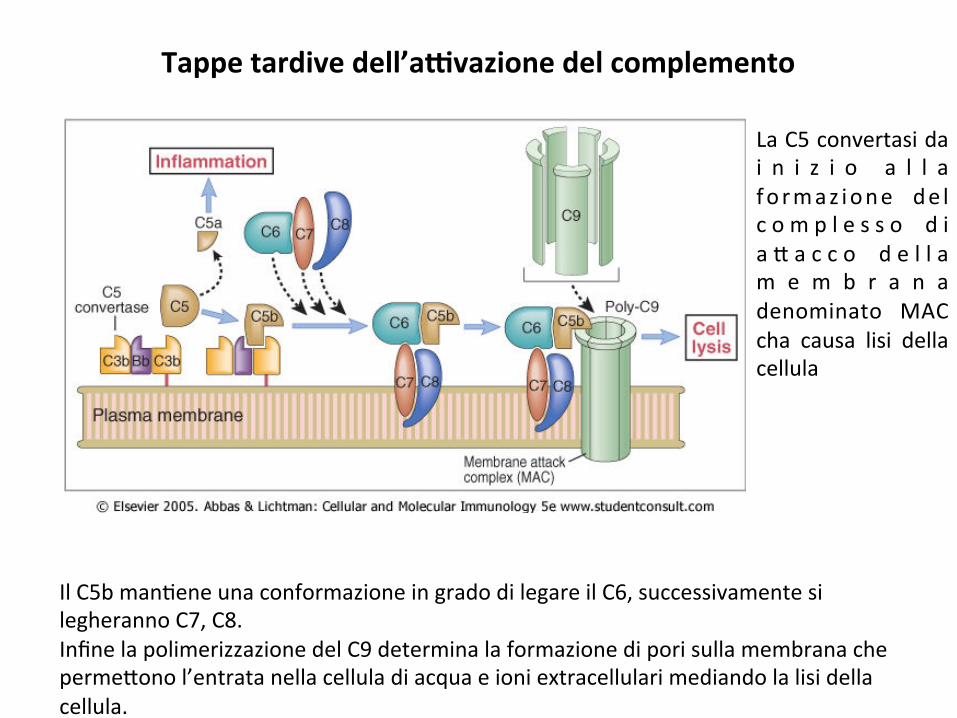
LaC5convertasidai n i z i o a l l afo rmaz ione de lc o m p l e s s o d ia * a c c o d e l l am e m b r a n adenominato MACcha causa lisi dellacellula
Tappetardivedell’aGvazionedelcomplemento
IlC5bman;eneunaconformazioneingradodilegareilC6,successivamentesilegherannoC7,C8.InfinelapolimerizzazionedelC9determinalaformazionediporisullamembranacheperme*onol’entratanellacelluladiacquaeioniextracellularimediandolalisidellacellula.
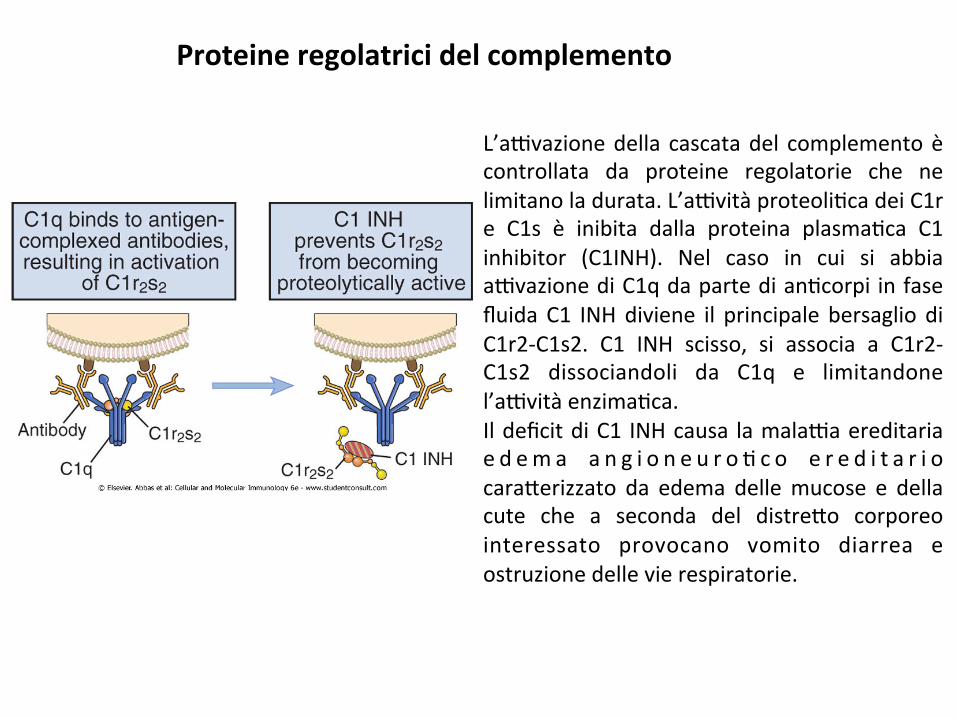
L’a9vazionedella cascatadel complementoècontrollata da proteine regolatorie che nelimitanoladurata.L’a9vitàproteoli;cadeiC1re C1s è inibita dalla proteina plasma;ca C1inhibitor (C1INH). Nel caso in cui si abbiaa9vazionediC1qdapartedian;corpi infasefluidaC1 INHdiviene il principale bersaglio diC1r2-C1s2. C1 INH scisso, si associa a C1r2-C1s2 dissociandoli da C1q e limitandonel’a9vitàenzima;ca.IldeficitdiC1 INHcausa lamala9aereditariae d em a a n g i o n e u r o ; c o e r e d i t a r i ocara*erizzato da edema dellemucose e dellacute che a seconda del distre*o corporeointeressato provocano vomito diarrea eostruzionedellevierespiratorie.
Proteineregolatricidelcomplemento
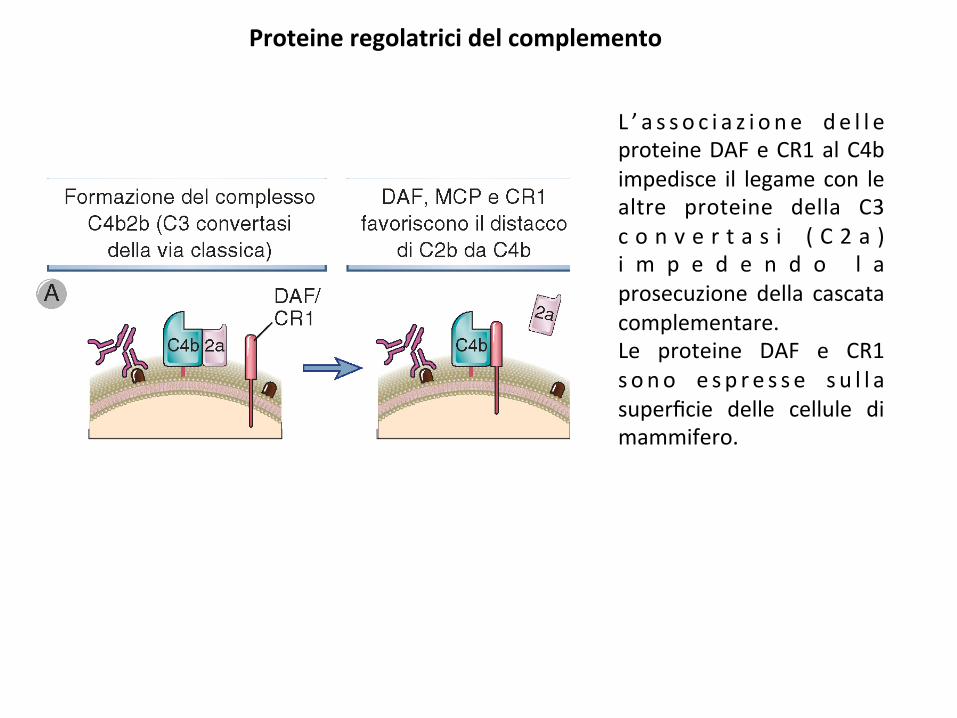
Proteineregolatricidelcomplemento
L ’ a s s o c i a z i o n e d e l l eproteineDAFeCR1alC4bimpedisce il legamecon lealtre proteine della C3c o n v e r t a s i ( C 2 a )i m p e d e n d o l aprosecuzione della cascatacomplementare.Le proteine DAF e CR1s ono e s p r e s s e s u l l asuperficie delle cellule dimammifero.
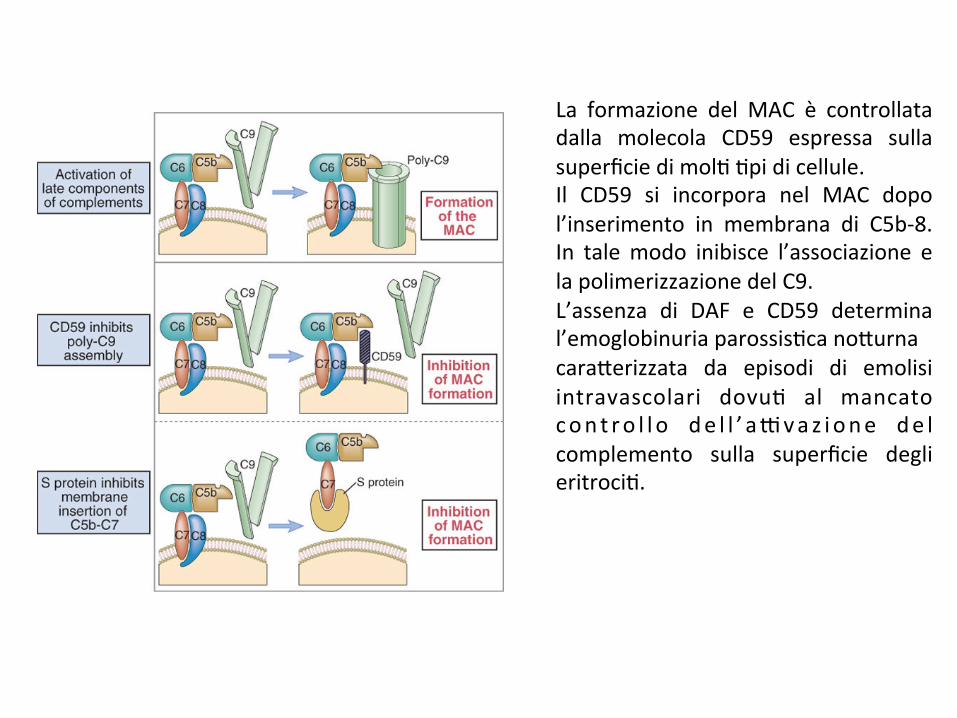
La formazione del MAC è controllatadalla molecola CD59 espressa sullasuperficiedimol;;pidicellule.Il CD59 si incorpora nel MAC dopol’inserimento in membrana di C5b-8.In talemodo inibisce l’associazione elapolimerizzazionedelC9.L’assenza di DAF e CD59 determinal’emoglobinuriaparossis;cano*urnacara*erizzata da episodi di emolisiintravascolari dovu; al mancatocon t ro l l o de l l ’ a9vaz ione de lcomplemento sulla superficie deglieritroci;.





























