Embed Size (px)
Citation preview
FondamentaliFondamentali
di genetica di genetica
delle popolazionidelle popolazioni
Genetica delle popolazioni
Si occupa:
- delle frequenze dei geni e dei genotipi nelle popolazioni
- del loro cambiamento lungo le generazioni
… e quindi di:
- evoluzione
- selezione naturale e artificiale
Genetica delle popolazioni

Genetica delle popolazioni
Una tale popolazione (di piante allogame, ad esempio) è costituita da individui altamente eterozigoti e tutti geneticamente diversi tra loro
Gruppo di individui sessuati a fecondazione incrociata che si dividono un pool genico comune
Def. Frequenza genotipica: frequenza di un genotipo in una popolazione
Una popolazione è costituita così:
AA 250 (D) x = 250/1000 = 0,25 = D/N
Aa 600 (H) y = 600/1000 = 0,60 = H/N
aa 150 (R) z = 150/1000 = 0,15 = R/N
Tot. 1000 (N) 1,00
Consideriamo il solito locus A con due alleli, A e a
AA Aa aa x y z
Genotipi Frequenze genotipiche x + y + z = 1
Popolazione
Def. Frequenza genica: frequenza di un allele nei loci disponibili per esso in una popolazione
AA Aa aa x y z
Genotipi Frequenze genotipiche
Nel nostro esempio:
x = 0,25
y = 0,60
z = 0,15
p(A) = x + 1/2y = 0,55
q(a) = z + 1/2y = 0,45
p + q = x + 1/2y + z + 1/2y = x + y+ z = 1
Genetica delle popolazioni
Frequenze geniche p(A) = x + 1/2y
q(a) = z + 1/2y
Calcoliamo le frequenze degli alleli A e a
Gli alleli A sono: (2D+H)/2N; quelli a sono (2R+H)/2N, ma più semplicemente:

La legge di Hardy-Weinberg stabilisce che in una generazione di unioni a caso le frequenze genotipiche raggiungono l’equilibrio alle frequenze:
p2(AA) + 2pq(Aa) + q2(aa)
Se tutti gli individui hanno uguali possibilità di riprodursi, le frequenze dei gameti corrispondono alle frequenze geniche. Quindi la generazione successiva, con incroci casuali (panmissia), sarà costituita così:
Gametip(A) q(a)
p(A) p2(AA) pq(Aa)
q(a) pq(Aa) q2(aa)
+
Genetica delle popolazioni
1. Una popolazione è composta da 750 individui AA e 250 individui Aa.
a. Quali sono le frequenze geniche?b. Quali saranno le frequenze genotipiche e geniche nella generazione successiva?
2. Una popolazione è composta da 500 individui AA, 250 individui Aa e 50 individui aa.a. La popolazione è in equilibrio?b. Quali saranno le frequenze genotipiche alla generazione successiva, con unioni casuali?
3. In una popolazione in equilibrio gli individui con genotipo aa sono il 16%. Determinare le frequenze geniche e genotipiche
4. Costruisci un foglio excel per il calcolo e la verifica dell'equilibrio HW date le frequenze assolute dei tre genotipi
Esercizi
Genetica delle popolazioni

Le condizioni dell’equilibrio H-W
* Popolazione numerosa
* Costanza delle frequenze geniche:
* Unioni casuali (panmissia)
Assenza di mutazione differenziale
Assenza di migrazione
Le unioni non sono casuali quando avvengono:
* tra individui fenotipicamente o geneticamente simili: inbreeding
* tra individui fenotipicamente o geneticamente dissimili: outbreeding
Genetica delle popolazioni
Assenza di selezione
Miglioramento genetico = Aumentare le frequenze degli alleli favorevoli mediante selezione
Il breeder sceglie, seleziona, gli individui che hanno le caratteristiche desiderate con l'obiettivo di trasmetterle alle progenie
Fitness (W) o valore di adattamento o valore selettivo
è il parametro della selezione ed esprime contributo di ciascun genotipo alla generazione successiva, in termini di numero di figli
Selezione
Si ha selezione, naturale o artificiale, quando alcuni genotipi hanno
fertilità maggiore di altri e quindi contribuiscono alla generazione successiva in misura diversa
Posto uguale a 1 il numero di figli dell’individuo che fa più figli, la fitness degli altri individui viene definita in relazione a questo valore massimo
0 < W < 1
Genetica delle popolazioni

Coefficiente di selezione S = 1-W
Esprime il minor contributo di un genotipo alla generazione successiva rispetto al genotipo con fitness pari a 1
Esempio: Se la selezione sfavorisce l’omozigote recessivo e in presenza di dominanza completa al locus A si ha:
AA W=1Aa W=1aa W=1-s
Nel caso estremo di un recessivo letale:
AA W=1Aa W=1aa W=0
Se la fitness di un individuo è legata ai geni che porta, la frequenza dei geni degli individui con fitness più elevata aumenterà
da una generazione all’altra, mentre le frequenza dei geni che determinano una bassa capacità riproduttiva diminuirà
La selezione modifica le frequenze geniche
Genetica delle popolazioni
p2 + 2pq + q2
AA Aa aaGenotipi
Frequenze
Fitness 1 1 1-s
Frequenze dopo che ha agito la selezione (un ciclo):
p2 + 2pq + q2(1-s) = p2 + 2pq + q2 - sq2 = 1- sq2
p2 + 2pq + q2p2 + 2pq + q2p2 + 2pq + q2
Per far sì che la somma delle frequenze sia =1:
AA Aa aaGenotipi
Frequenzep2 2pq q2(1-s)
1- sq2 1- sq2 1- sq2+ +
Esempio: la selezione sfavorisce l'omozigote recessivo
Genetica delle popolazioni

Dopo la selezione, quindi, la frequenza di a (q1) sarà (secondo la
formula q= z + 1/2y):
q2(1-s)
1- sq2 1- sq2+
pqq
1 = = …. =
q - sq2
1- sq2
Per convenzione, l'effetto della selezione si misura con la variazione di q:
Hq = q1 – q =
q - sq2
1- sq2– q = … =
- sq2(1-q)
1- sq2
La selezione contro il recessivo è tanto più efficace quanto maggiori sono q e s
Genetica delle popolazioni
Poniamo che una popolazione abbia le frequenze genotipiche e geniche seguenti:
1/3AA + 2/3Aa ; p=2/3; q=1/3
Le frequenze genotipiche e geniche della generazione successiva saranno quelle volute dall’equilibrio H-W:
p2 (AA) + 2pq (Aa) + q2 (aa) 4/9 AA + 4/9 Aa + 1/9 aaCioè:
Si ha quindi: 1/2AA + 1/2Aa e p=3/4; q=1/4
Formula di Liq1 = q
1 + q
Esempio: il destino di un allele letale recessivo che determina albinismo nelle piante
Alla generazione successiva analogamente, si avrà: p=4/5; q=1/5 … e così via
1- sq2q1 = =
1- q2 (1+q)(1- q)=
q - sq2 q(1- q) q(1- q)Infatti:
Genetica delle popolazioni

Con questa formula possimo calcolare la diminuzione della frequenza di un allele letale recessivo nel corso di successive generazioni di interincrocio casuale, e calcolare il numero di generazioni (t) necessario per ridurre la frequenza di una quantità voluta
q1 = q/(1 + q) qt = q/(1 + tq), da cui:
t = 1/qt + 1/q
Genetica delle popolazioni
Quante generazioni ci vogliono per dimezzare la frequenza di un recessivo letale a partire dalla frequenza iniziale q = 0,4 ?
t = 1/qt + 1/q = 1/0,2 + 1/0,4 = 5 + 2,5 = 7,5
E quante generazioni ci vogliono per dimezzare ancora la frequenza a 0,1?
t = 1/0,1 + 1/0,2 = 10 +5 = 15
E per dimezzare ancora la frequenza a 0,05?
t = 1/0,05 + 1/0,1 = 20 + 10 = 30
Quando la frequenza è piccola la selezione è poco efficace anche contro alleli letali
Esercizio
Genetica delle popolazioni

Modello generale della selezione
p2 + 2pq + q2
AA Aa aaGenotipi
Frequenze
Fitness W0 W
1 W
2
p2 + 2pq + q2p2 + 2pq + q2p2 + 2pq + q2
p2W0 + 2pqW
1 + q2W
2 = W Fitness media
o valore adattativo medio
Dopo la selezione le frequenze genotipiche saranno:
p2W0
W W W+ + = 1
La frequenza genica di a dopo la selezione, sarà:
q2W2
2pqW1
W+
q2W2
W=
pqW1 q(qW
2+ pW
1)
Wq
1 =
Genetica delle popolazioni
Hq = q1 – q =
q(qW2+ pW
1)
– q = …....p2W
0 + 2pqW
1 + q2W
2
Hq =pq[q(W
2- W
1) + p(W
1- W
0)]
W
Il modello generale della selezione ci dice che la selezione è più efficace con frequenze geniche di 0,5 e quando la fitness media è bassa
Genetica delle popolazioni

Esercizi
1. Calcolare Hq nel caso di dominanza completa e selezione contro l'omozigote recessivo (W
0=W
1=1; W
2=1-s)
2. Calcolare Hq nel caso di dominanza completa e selezione contro il dominante (W
0=W
1=1-s; W
2=1). Cosa avviene quando s=1?
3. Calcolare Hq nel caso di selezione contro entrambi i dominanti (W
0=1-s
1; W
1=1; W
2=1-s
2). Calcolare il valore di q all'equilibrio
Genetica delle popolazioni
Inbreeding E’ un sistema di unioni in cui le unioni tra individui imparentati è più frequente di quanto voluto dal caso
Può prodursi nel caso di popolazioni costituite da un numero piccolo di individui o molto stanziali, o per intervento dell’uomo
L’omozigosi per alleli identici è molto improbabile in un sistema di unioni casuale, mentre è tanto più probabile quanto più stretta è la parentela tra gli individui che si uniscono
La conseguenza è l’aumento della percentuale di omozigoti per alleli identici
alleli identici
Aa
AA Aa aa
Alleli identici sono alleli che hanno la stessa origine. Sono cioè copie di un unico allele presente inizialmente in un individuo e originatisi per replicazione del DNA
AA Aa
x AA
alleli simili
alleli diversi
Qual è la forma di inbreeding più “stretto” ?
In zootecnia si utilizza il termine Consanguineità
Genetica delle popolazioni

La probabilità che un individuo estratto a caso da una popolazione possieda alleli identici è stata assunta come misura del grado di inbreeding della popolazione. Tale probabilità è espressa dal
0 < F < 1F = 0 individuo non omozigote per alleli identici (per tutti i suoi geni)
F = 1 individuo omozigote per alleli identici (per tutti i suoi geni)
Quindi F esprime allo stesso tempo:
� la percentuale di individui omozigoti per alleli identici in una popolazione� la percentuale dei geni omozigoti per alleli identici di ciascun individuo
Coefficiente di inbreeding: F
(Sewall-Wright, 1922)
Genetica delle popolazioni
AH1H2
B H3H4
C D
E
Unione fratello-sorellaQual è la probabilità che:
E sia omozigote H1H1:
E sia omozigote H2H2:
E sia omozigote H1H1 o H2H2:
E sia omozigote H3H3 o H4H4:
E sia omozigote H1H1 o H2H2 o H3H3 o H4H4: 4(1/2)4
FE= 4(1/2)4 = 4/16 = 0,25
probabilità che E sia omozigote
per alleli identici
In generale:
FX = Σ(1/2)n+1(1 + Fy)
Dove:
Σ fa riferimento a tutti gli antenati comuni ai genitori
dell’individuo consideraton è il numero di passi generazionali che vanno da un genitore all’altro dell'individuo considerato attraverso l’antenato comune
Fy è il coefficiente di inbreeding dell’antenato comune
(1/2)(1/2) (1/2)(1/2)=(1/2)4
(1/2)(1/2) (1/2)(1/2)=(1/2)4
2(1/2)4
2(1/2)4
Calcolo del Coefficiente di inbreeding F
Genetica delle popolazioni

Coefficiente di parentela
Probabilità che due individui (A e B) possiedano alleli identici.
PAB = Σ(1/2)n(1 + Fy)
E' il doppio del coefficiente di inbreeding della loro progenie.
E' un parametro molto utilizzato in campo animale per decidere quali individui vengono fatti accoppiare
Genetica delle popolazioni
Nota
Calcola il coefficiente di inbreeding del toro Comet, uno dei capostipiti della razza Shorthorn, il cui pedigree è qui rappresentato
Esercizio
Lady Maynard Foljambe
Phoenix
YoungStrawberry
Bolingbroke
Favourite
Young Favourite
Comet
?
Genetica delle popolazioni

Esercizi
1. Calcola l’omozigosi teorica nella F4 di un triibrido, nell’ipotesi che i 3 geni siano
indipendenti.
2. Calcola il coefficiente di inbreeding del cane Crack che ha il pedigree rappresentato in figura.
FUFI DODI
BILLI LILLI
BOBI
TERRI
JACK
PIPPI
BIRBA
CRACK
L’effetto dell’inbreeding sull’equilibrio Hardy-Weinberg
Se una popolazione è inbred essa avrà una percentuale di omozigoti per alleli identici pari a F e la percentuale totale di omozigoti sarà maggiore di quanto voluto dalla legge di H-W
Gli F omozigoti per alleli identici, per un gene A, si ripartiranno tra le due forme alleliche: pF(AA) + qF(aa) = FLa parte restante della popolazione (1-F) avrà le frequenze genotipiche previste dalla legge di H-W:
p2(1-F)(AA) + 2pq(1-F)(Aa) + q2(1-F)(aa)
Sommando: f(AA) = p2(1-F) + pF = p2 – p2F + pF = p2 + pF(1-p) = p2 + pqF f(Aa) = 2pq(1-F) = 2pq- 2pqF
f(aa) = q2(1-F) + qF = q2 – q2F + qF = q2 + qF(1-q) = q2 + pqF
E quindi: (p2 + pqF)(AA) + (2pq- 2pqF)(Aa) + (q2 + pqF)(aa)
Formula dell’equilibrio di Sewall-Wright
Genetica delle popolazioni

L’effetto dell’inbreeding sul fenotipo
Depressione da inbreeding
Diminuzione del vigore e della fertilità (e quindi della produttività) in conseguenza di unioni inbred
Eterosi (Shull, 1905)
Esplosione di vigore e fertilità (e quindi di produttività) quando si uniscono particolari genitori (linee) inbred
per ottenere un “ibrido” F1)
D.F. Jones, Genetics 9:405-418, 1922
Inbred
parentali
F1
Generazioni di autofecondazione1 2 3 4 5 6 7
N.B.L'entità della depressione da inbreeding e dell'eterosi varia in funzione della specie e del genotipo
Genetica delle popolazioni
Plant Cell, Vol. 15, 2236–2239, 2003
Eterosi in mais
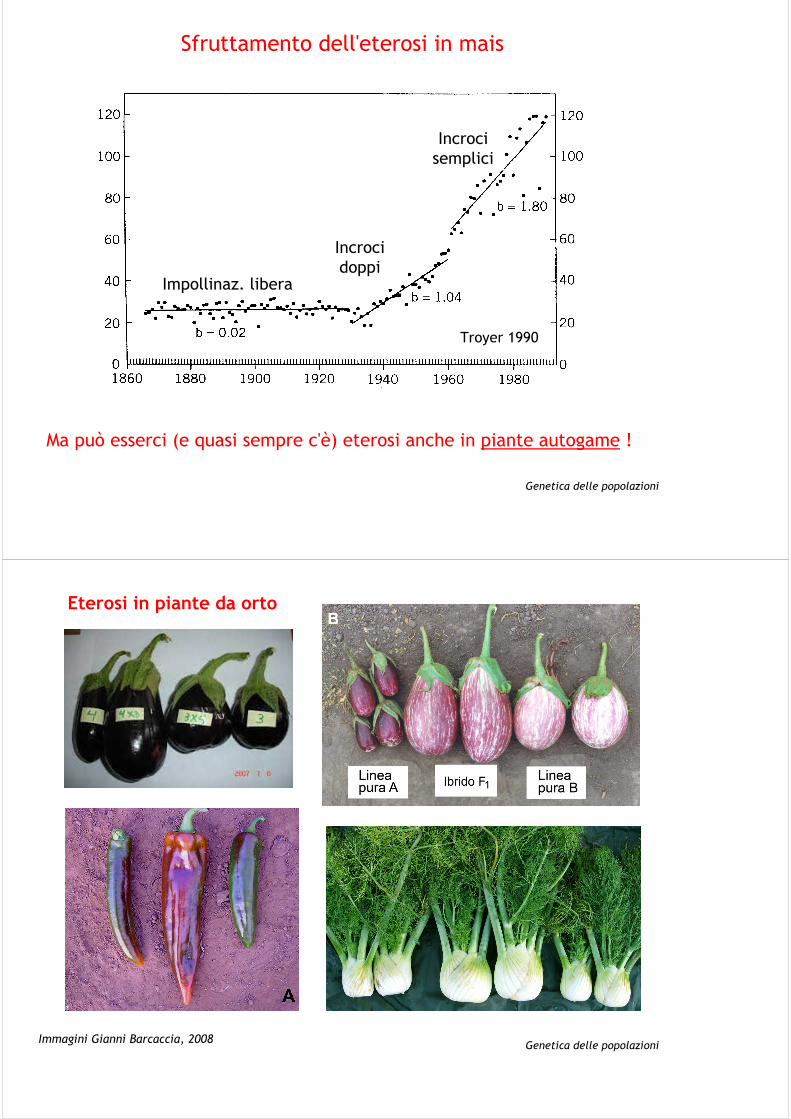
Genetica delle popolazioni
Sfruttamento dell'eterosi in mais
Incroci semplici
Incroci doppi
Impollinaz. libera
Troyer 1990
Ma può esserci (e quasi sempre c'è) eterosi anche in piante autogame !
Genetica delle popolazioniImmagini Gianni Barcaccia, 2008
Eterosi in piante da orto

Mid-parent heterosis
F1P1 P2 F1P1 P2
High-parent heterosis
Ha rilevanza pratica
Basi genetiche dell'eterosi
• Dominanza direzionale, e/o• Superdominanza/pseudosuperdominanza, e/o• Epistasi
EterosiGeorge H. Shull (1911) “the superiority of heterozygous genotypes over the corresponding homozygotes”
HF1 = F1 – PM
HF1 (%) = 100(F1 – PM)/PM
HF1 = F1 – P
HF1 (%) = 100(F1 – P)/P
–
– –
Un modello utile per misurare l'effetto dell'inbreeding su un carattere quantitativoUn gene che contribuisce, insieme ad altri, al controllo di un carattere
quantitativo può essere descritto così:
Dominanza incompleta:h a-a
I I I I0
AAAaaa
Dominanza completa:
ha-a
I I I0
AA
Aa
aa
Il modello assume pari a 0 il valore della media tra i due omozigoti aa e AA, e che la sostituzione dell'allele a con A incrementa il valore del carattere quantitativo di una quantità a
ha-a
I I I0
AAAaaaAssenza di dominanza:

Effetto dell'inbreeding sui caratteri quantitativi
Genotipi Valore genotipico Frequenza Prodotto
AA a p2 p2a
Aa h 2pq 2pqh
aa -a q2 q2(-a)
p2a + 2pqh + q2(-a) = (p2 – q2)a + 2pqh = (p – q)a +2pqh
Il valore genotipico medio della popolazione è:
Genotipi Valore genotipico Frequenza Prodotto
AA a p2 +pqF (p2 +pqF)a
Aa h 2pq - 2pqF (2pq-2pqF)h
aa -a q2 + pqF (q2-pqF)(-a)
Popolazione inbred:
p2a + pqFa + 2pqh -2pqFh + q2(-a) – pqFa = …. = (p – q)a +2pqh - 2pqFh
Il valore genotipico medio della popolazione è:
2pqFh misura le depressione da inbreeding: è proporzionale ad h e da F
Genetica delle popolazioni
Ipotesi della dominanza (Jones)
Come abbiamo visto, la media di una popolazione inbred diminuisce di 2pqhF rispetto alla popolazione panmittica a causa dell'omozigosi per alleli (identici) recessivi sfavorevoli
AA bb CC DD ee FF GG hh II ll aa BB cc dd EE ff gg HH ii LL
Aa Bb Cc Dd Ee Ff Gg Hh Ii Ll
Inbred 1 Inbred 2
F1
x
HF1 = Σhh a-a
I I I I0
In questo ibrido tutti i geni sono in condizione eterozigote e quindi nessuno dei recessivi sfavorevoli si manifesta

Perchè ci sia eterosi la dominanza deve essere “direzionale”: tutti, o la gran parte, dei loci che controllano il carattere devono avere dominanza nella stessa direzione, con l'allele dominante che incrementa il carattere
Dovrebbe allora essere possibile ottenere una inbred vigorosa quanto il migliore ibrido....Si, ma a condizione che avvengano tutti gli eventi di ricombinazione necessari, inclusi i crossing-over tra geni associati!
AA BB CC DD EE FF GG HH II LL
In effetti, nel mais, grazie alla selezione ricorrente che va avanti da 100 anni, le inbred di oggi sono molto migliori di quelle di una volta ...
Single locus overdominance in pomodoro: Single flower
truss (Sft). Krieger et al. Nature Genetics 42, 459–463, 2010
Eterosi
Genetica delle popolazioni
Ipotesi della superdominanza
L'eterozigote supera entrambi gli omozigotiE' stata dimostrata solo per alcuni geni, ad esempio mutazioni clorofilliane in orzo (Gustafsson)
ha-a I I I I
0

Gruppi eterotici
popolazioni o gruppi di popolazioni che “combinano bene” tra loro: Incroci tra piante appartenenti a gruppi eterotici diversi presentano eterosi elevata
Secondo l'ipotesi di East, i gruppi eterotici devono possedere alleli diversi per i geni che controllano i caratteri che manifestano eterosi
East propose che la diversità allelica sia legata al vigore, e che esistano alleli via via “divergenti” che combinandosi insieme nell'ibrido danno effetti di eterosi diversi:
a1 > a2 > a3 > a4 …
a1a4 > a1a3 > a1a2 > a1a1
Alleli multipli
Combinazioni ibride
In effetti esistono molti dati in molte specie diverse che legano l'eterosi alla diversità genetica:Più sono geneticamente diversi i genitori, più aumenta l'eterosi delle loro progenie ibride.
L'effetto del linkage in trans tra un gruppo di geni può fare l'effetto della superdominanza quando in realtà non c'è superdominanza per i singoli geni
A b C D e F G h I l
A b C D e F G h I l
a B c d E f g H i L
A b C D e F G h I l
a B c d E f g H i L
Inbred 1 Inbred 2
F1
a B c d E f g H i L
x
E' rappresentata una porzione di un cromosoma di due inbred “complementari”
Se considerassimo questo tratto cromosomico come un unico locus, concluderemmo che esiste superdominanza
Pseudosuperdominanza
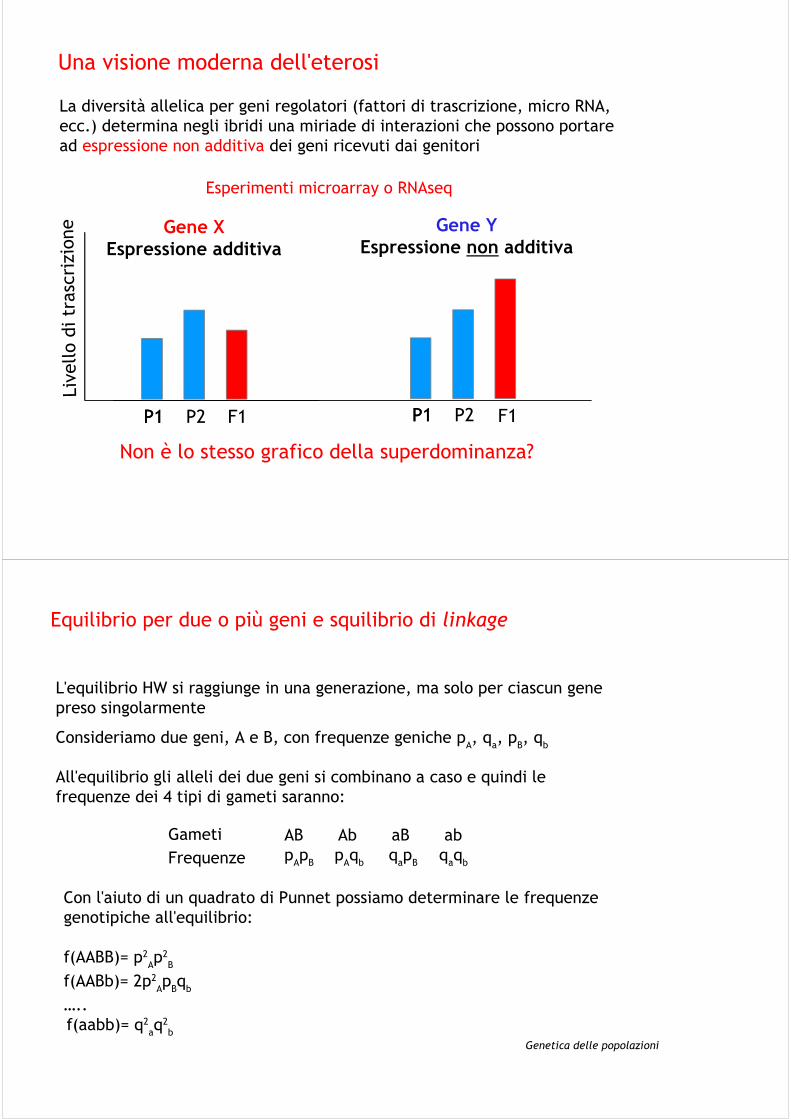
Una visione moderna dell'eterosi
La diversità allelica per geni regolatori (fattori di trascrizione, micro RNA, ecc.) determina negli ibridi una miriade di interazioni che possono portare ad espressione non additiva dei geni ricevuti dai genitori
P1 P2P1
Gene X
Espressione additiva
Liv
ello d
i tr
asc
rizi
one
P1 P2P1
Gene Y
Espressione non additiva
F1 F1
Non è lo stesso grafico della superdominanza?
Esperimenti microarray o RNAseq
Equilibrio per due o più geni e squilibrio di linkage
L'equilibrio HW si raggiunge in una generazione, ma solo per ciascun gene preso singolarmente
Consideriamo due geni, A e B, con frequenze geniche pA, q
a, p
B, q
b
All'equilibrio gli alleli dei due geni si combinano a caso e quindi le frequenze dei 4 tipi di gameti saranno:
AB Ab aB abp
Ap
B p
Aq
b q
ap
B q
aq
b
Gameti
Frequenze
Con l'aiuto di un quadrato di Punnet possiamo determinare le frequenze genotipiche all'equilibrio: f(AABB)= p2
Ap2
B
f(AABb)= 2p2Ap
Bq
b
…..
f(aabb)= q2
aq2
b
Genetica delle popolazioni

Immaginiamo di formare una popolazione mescolando un ugual numero di individui con genotipi AABB e aabb, lasciati poi alla panmissia
Le frequenze geniche iniziali sono: pA= q
a= p
B= q
b= 0,5
Le frequenze gametiche sono inizialmente 0,5 sia per AB che per ab (diverse da quelle all'equilibrio, pari a 0,25 per tutti i tipi gametici), quindi dopo la prima generazione di unioni a caso si avrà:
Gameti0,5(AB) 0,5(ab)
0,5(AB) 0,25(AABB) 0,25(AaBb)
0,5(ab) 0,25(AaBb) 0,25(aabb)
Alla prima generazione si formano solo 4 dei possibili 9 genotipi.Alla generazione successiva le frequenze gametiche cambieranno per effetto della segregazione degli eterozigoti. Se A e B sono indipendenti:
f(AB) = 0,25 + 2(0,25)2 = 0,25 + 0,125 = 0,375f(Ab) = 2(0,25)2 = 0,125 f(aB) = 2(0,25)2 = 0,125 f(ab) = 0,25 + 2(0,25)2 = 0,25 + 0,125 = 0,375
Equilibrio per due o più geni e squilibrio di linkage
Genetica delle popolazioni
Le frequenze gametiche si avvicinano progressivamente a quelle di equilibrio nel corso delle generazioni di interincrocio casuale.
Abbiamo ipotizzato che A e B siano indipendenti.Se due geni sono associati, allora la formazione di gameti ricombinanti è possibile solo grazie al crossing-over e il raggiungimento dell'equilibrio ed è quindi molto più lento
Squilibrio gametico (gametic phase disequilibrium)
In assenza di linkage viene perduto (dissipato) nel corso di pochissime generazioni.
Si parla comunque (non del tutto propriamente) di
Squilibrio di linkage (Linkage disequilibrium, LD)
Il LD si stima, di solito, con il parametro
D = p(AB) - pAp
B
Dove p(AB) è la frequenza osservata del gamete AB e pa e p
B sono le frequenze
geniche di A e B.Il valore assoluto di D è lo stesso per tutti i gameti ma è di segno positivo per i gameti parentali e negativo per i ricombinanti.
Equilibrio per due o più geni e squilibrio di linkage
Genetica delle popolazioni

Lorenzetti et al. 2010
Dt = D
0(1-c)t
Questa formula ci permette di calcolare la diminuzione di D dopo t generazioni di incrocio casuale; c = frequenza di ricombinazione
Equilibrio per due o più geni e squilibrio di linkage
Genetica delle popolazioni
La presenza di LD è essenziale per la costruzione di mappe genetiche.Per fare una mappa si incrociano due genotipi contrastanti e si utilizza la F
2 o una
popolazione da reincrocio, quindi c'e stata una sola meiosi che che ha potuto creare ricombinazione: il LD è massimo
In laternativa si può ricorrere al cosiddetto
LD mapping, o (impropriamente) Association mapping
Si utilizzano popolazioni preesistenti cercando associazione tra il carattere studiato (ad esempio un malattia genetica nell'uomo, o un carattere quantitativo in una pianta coltivata) con le frequenze alleliche di marcatori molecolari (prevalentemente SNP)
L'ipotesi di base è che il tratto cromosomico dove è ubicato il gene responsabile del fenotipo di interesse presenti LD almeno con i marcatori più vicini
LD e mappe genetiche
Nella forma più avanzata si parla di Genome Wide Association Studies (GWAS)
LD mapping è molto utile negli organismi in cui non è possibile creare popolazioni ad hoc, o anche quando tale creazione sia lunga e complessa, ma viene oggi utilizzato anche nelle piante
Genetica delle popolazioni

Se disponiamo di molti marcatori molto vicini possiamo individuare un aplotipo specifico per il carattere studiato.
Nella genetica umana, con il LD mapping, si cerca di mettere in relazione particolari aplotipi con la suscettibilità o predisposizione a particolari patologie
L’uso pratico del LD mapping nel miglioramento genetico delle piante oggi è ancora limitato, anche se ci sono ormai molti studi pubblicati
SNPSNP11 SNPSNP22 SNPSNP33 SNPSNP44
CC AA CC TT
AA CC AA AAMM
Questo perchè nel momento in cui è comparso, in un individuo della popolazione, l'allele responsabile del fenotipo (ad esempio, l'allele mutato MM), esso si trovava accanto a specifici alleli dei marcatori vicini, e assieme ad essi rappresenta una combinazione di alleli (aplotipo) unica a loci associati . Questa combinazione di alleli viene progressivamente distrutta dalla ricombinazione, ma ciò avviene molto lentamente per loci molto vicini tra loro
Genetica delle popolazioni
Zhu et. al. Plant Genome, 2008, 1:5-20 Status and Prospects of Association Mapping in Plants Genetica delle popolazioni

Selezione genomica (Genomic selection, GS)
Una volta che è stata stabilita l'associazione genome-wide tra marcatori e caratteri, diventa possibile fare GS
Hamblin et al 2011. Population genetics of genomics-based crop improvement methods. Trends in Genetics 27 (3)
E' una forma di MAS in cui si utilizzano marcatori che coprono l'intero genoma, scelti in modo tale che tutti i QTL individuati nell'analisi “Genome Wide” sono in LD con almeno un marcatore.
Si selezionano gli individui della popolazione segregante sulla base degli alleli favorevoli a tutti i marcatori scelti
E' diventata possibile grazie alla disponibilità di molti SNP e dell'automazione della loro genotipizzazione
Termolino Metologie integrate per la selezione genomica di piante ortive (25)Tripòdi Metodi di selezione genomica di peperone (20)Portis Metodi di selezione genomica in melanzana (20)Formisano Metodi di selezione genomica in zucchino (20)Riccardo Metodi di selezione genomica in patata (20)
Genetica delle popolazioni
Sommario
Le popolazioni vengono descritte in termini di frequenze geniche e genotipiche
Le frequenze geniche e genotipiche vengono modificate dalla selezione
Il miglioramento genetico spesso comporta l'incrocio tra individui imparentati (inbreeding)
I caratteri quantitativi sono soggetti alla depressione da inbreeding in misura diversa per diverse specie agrarie. L'eterosi è il lato positivo della medaglia!
L'eterosi varia per I diversi caratteri, specie e genotipi, e viene sfruttata grazie all'incrocio controllato (prossima lezione)
Lo squilibrio di linkage permette di mappare i geni utili ai fini della selezione e dell'isolamento dei geni
La selezione genomica è resa possibile dalla sintesi delle conoscenze di genetica delle popolazioni e genetica quantitativa, insieme alle tecniche più recenti di analisi del genoma (SNP genotyping)
Genetica delle popolazioni