Embed Size (px)
Citation preview

www.elsevier.com/locate/jbiosc
Journal of Bioscience and BioengineeringVOL. 117 No. 3, 343e350, 2014
Aerobic utilization of crude glycerol by recombinant Escherichia coli forsimultaneous production of poly 3-hydroxybutyrate and bioethanol
Pramod Shah,1 Feng-Shen Chiu,1 and John Chi-Wei Lan1,2,*
Graduate School of Biotechnology and Bioengineering, Yuan Ze University, 135 Yuan-Tung Road, Chung-Li, Taoyuan 320, Taiwan1 and Bio-refinery and Bioprocess EngineeringLaboratory, Department of Chemical Engineering and Materials Science, Yuan Ze University, 135 Yuan-Tung Road, Chung-Li, Taoyuan 320, Taiwan2
Received 15 March 2013; accepted 30 August 2013Available online 17 October 2013
* CorrespondDepartment ofYuan-Tung Rofax: þ886 345
E-mail add
1389-1723/$http://dx.doi
Crude glycerol, an inevitable byproduct during biodiesel production, is emerging as a potential feedstock forfermentation, due to its availability and a reasonable price. Biological utilization of abundant crude glycerol to severalvalue added products is contemporary research area with beneficial features. Solving the problem of proper disposal andraising economic viability of biodiesel industries. Several researches have been directed toward the production ofnumerous products by using Escherichia coli, an ideal organism for heterologous expression of various foreign proteins.In this fashion, recombinant E. coli strains were constructed for the simultaneous production of poly 3-hydroxybutyrate(P3HB) and bioethanol from crude glycerol. The incorporation of aldehyde reductase (Alrd) and aldehyde dehydrogenase(AldH) in recombinant strain showed 2-fold increment in crude glycerol utilization under aerobic condition. Moreover,these two enzymes introduced an alternative pathway leading toward the potential production of bioethanol which wasmore than redox-balancing steps. Acetate was accumulated as an intermediate product. Subsequently, acetate wasutilized as substrate in the second pathway, which directly converted acetyl-CoA to P3HB. This strategy demonstrated apotential production manner of bioethanol as an extracellular product and P3HB as water insoluble inclusion bodiesinside E. coli. The maximum production of bioethanol and P3HB in the recombinant strain was 0.8 g LL1 (17.4 mmol LL1)and 30.2% (w/w dry cell weight), respectively, which were higher than the parental strain.
� 2013, The Society for Biotechnology, Japan. All rights reserved.
[Key words: Escherichia coli; Aerobic fermentation; Crude glycerol; Bioethanol; Poly 3-hydroxybutyrate; Aldehyde reductase; Aldehydedehydrogenase]
The alarming rate of consumption and combustion of fossil fuelsto support global energy demand is continuously causing depletionof its limited stocks and severe impact on health and environment(1). Therefore, increasing enthusiasm is focused toward the gen-eration of alternative energy sources. Utilization of biomass forbiofuel production has created unexceptional opportunities indealing with increasing demand and escalating prices of fossil fuel.Biofuel are renewable energy with rather high efficiency and itssustainability and biodegradability contribute to the reduction ofcarbon dioxide emission and other harmful gases during combus-tion (2,3).
The production capacity and market values of biodiesel andbioethanol are gaining lots of attention than other categories ofbiofuels (4). The appropriate method applied for the production ofbiodiesel is a homogeneous base-catalyze trans-esterification oftriglycerides existing in vegetable oils, animal fats, or waste cookingoils with methanol (5). This process separates fatty acids moietyfrom triglycerides resulting in the generation of an inevitablebyproduct, glycerol. One mole of glycerol is produce with the pro-duction of every 3 mol of methyl esters (i.e., biodiesel), resulting in
ing author at: Bio-refinery and Bioprocess Engineering Laboratory,Chemical Engineering and Materials Science, Yuan Ze University, 135ad, Chung-Li, Taoyuan 320, Taiwan. Tel.: þ886 34638800x3550;59373.ress: [email protected] (J.C.-W. Lan).
e see front matter � 2013, The Society for Biotechnology, Japan..org/10.1016/j.jbiosc.2013.08.018
10% by weight (wt %) of the total crude glycerol produce (6). Inrecent years, the mandatory demand of biodiesel has flooded themarket with excessive crude glycerol. As a result, the price of crudeglycerol has drastically plummeted enforcing burden on economicviability in the biodiesel industry. Crude glycerol alone account for13e14 % of the total production costs (7). As a consequence of it,several biodiesel companies need governmental aids for supportingits operation (4).
Biodiesel companies are seeking new alternative for this crudeglycerol to higher-value added products. Biological utilization ofcrude glycerol serves as a feedstock in various fermentation pro-cesses with the production of several value-added products such as1,3-propanediol (PDO), dihydroxy-acetone (DHA), bioethanol,butanol, propionic acid, succinic acid, and so on (8). Utilization ofcrude glycerol in fermentation processes account in reduction ofmanufacturing costs and higher yields of products than other sugarsubstrates. Among several alternative of value-added productsduring the fermentation process, the production of bioethanol is aredox-balance pathway (9,10).
Utilization of crude glycerol by Escherichia coli, a model organ-ism in bioindustries has long been proved under aerobic conditionwith minimal ethanol production. In recent years, anaerobic andmicro-aerobic fermentation of crude glycerol has been well estab-lished for the higher yield of bioethanol by the construction ofdifferent recombinants E. coli (10e17). A high production of bio-ethanol metabolically requires a significant re-engineering of
All rights reserved.

TABLE 1. Microbial strains, plasmids and vectors used in this study.
Strain/Plasmid/Primer Description Source
StrainR. eutropha H16 Wild-type BCRCa-13036E. coli BL21 Wild-type BCRCa-51878E. coli BL21_pARD33-BHB2
pARD33 and pBHB2 This study
PlasmidspGEM-T easy Cloning vector PromegapBAD33 Low level expression
vectorProf. Su Boqi (Tzu ChiUniversity)
pGEM-T easy_alrd pGEM-T easy :: alrdRe This studypGEM-T easy_aldH pGEM-T easy :: aldHRe This studypARD33 pBAD33 :: alrd/aldH This studypBHB2 pBluescript II KS ::
phaCABRE
The Yuan Ze PHA team
PrimersAlrd primerForward GAGGTACCAAGGAGGA
AATGAAGCAAGTCACThis study
Reverse GAGTCGACAGCTCAAAGCATTTCCAG
This study
AldH primerForward AGTCGACAAGGAGAAT
ATGCGAGAAGTCCCCGA
This study
Reverse GAAAGCTTTCAACGCAGAAGCAGGCGCAA
This study
a BCRC: Bioresource Collection and Research Center (Taiwan).
344 SHAH ET AL. J. BIOSCI. BIOENG.,
metabolic pathway to derive bioethanol exclusively from acetyl-CoA with ATP as the energy source and NADPH as the sources ofreducing equivalents (14,18).
In this work, the knowledge of aerobic fermentation has beenused to engineer E. coli for the effective conversion of crude glycerolto bioethanol with aldehyde reductase and aldehyde dehydroge-nase. Aldehyde reductase (Alrd, EC 1.1.1.21) is an NADPH-dependentoxidoreductase enzyme which catalytically reduces various alde-hydes and carbonyls, including monosaccharide to correspondingpolyalcohol (19). Aldehyde dehydrogenase (AldH, EC 1.2.1.3), on theother hand, catalyzes the NADH-dependent oxidoreductase activ-ity, oxidizing aldehydes to corresponding carboxylic acids (20).These two enzymes provided alternative pathways for the utiliza-tion of glycerol through glycolysis and further, established theconversion of acetate, the end product of glycolysis, to ethanol. Ithas been observed that under aerobic condition, acetic acid is majorproduct which exits as acetate ion (21). Accumulation of acetate istoxic to organism thus a specific approach was considered for theconversion of acetyl-CoA to poly 3-hydroxybutyrate (P3HB), asecond value-added product. These pathways eliminated theaccumulation of acetate.
Poly-hydroxyalkanoates (PHAs) are another value-added prod-uct gaining lots of attention as an ingredient for thermoplastics.With its biodegradable nature and a variety of other materialproperties, they are suitable for several applications (22,23). PHAsare carbon and energy reserve accumulated in many bacteria asintracellular inclusions under excess carbon reserve or unfavorablecondition. These intracellular inclusions enhance the fitness ofbacteria and also contribute to redox-balance (24). P3HB is a wellknown member of PHAs family which has been widely studied.However, the production of P3HB in large scale is limited to sub-strates cost (25). Therefore, the use of crude glycerol for this pur-pose can be an interesting opportunity. Synthesis of P3HB fromglycerol as carbon source has been evaluated in several microbialstrains along with different fermentation strategy (26,27). Ralstoniaeutropha, a well known organism for the production of PHAs con-tains phaCAB operon for the conversion of acetyl-CoA to P3HB. Ithas been established that R. eutropha can accumulate more than80% of P3HB content (28). The recombinant E. coli constructed byharboring phaCAB operon from R. eutropha (phaCABRE) has beenshown to accumulate 67% of P3HB content, using 2e4% glucose ascarbon source (29). In the meantime, bacterial hemoglobin gene(vgb gene) from Vitreoscilla hemoglobin has been noted to enhancecell density, protein production and bioremediation by increasingoxygen availability in the culture (30). The co-expression of pha-CABRE and vgb genes in E. coli was revealed to enhance P3HB pro-duction (31).
The aim of the current study was to generate desirable productsfrom crude glycerol through alternate pathways provided by twoplasmids. Our attempt resulted in simultaneous production of anextracellular and an intracellular product, bioethanol and P3HB,respectively.
MATERIALS AND METHODS
Bacteria strains and plasmid used The wild-type strains, R. eutropha H16and E. coli BL21(lDE3), were kindly provided by Bioresource Collection and ResearchCenter (BCRC), Taiwan. E. coli BL21(lDE3) was used as a host for high level expres-sion of cloned genes. It contained T7 RNA-polymerase gene in a lysogenic andinducible form. This strain is characterized by two protein decomposition enzymegenes lon and ompT that has been artificially damage, making it suitable for the useof enzymes as a foreign gene expression (32). All strains used in this study alongwith plasmids and primers required for the construction of recombinant strainsare listed in Table 1. The molecular biology methods applied here are according tothe manufacturer’s protocols.
Construction of single and double plasmid Plasmid pBAD33 was kindlyprovided by Prof. Soo Po-Chi (Tzu Chi University, Taiwan). This plasmid containedPBAD promoter of araBAD (arabinose) operon and its regulatory gene, araC, followed
by multiple cloning site (MCS2) and M13 intragenic region with chloramphenicol-resistance (cmR) gene, resulting in a high level expression system (33). PolymeraseChain Reaction (PCR) was performed for the amplification of Alrd and AldH fromgenomic DNA of R. eutropha H16 on Select BioProducts (Edison, NJ, USA). Thereagents used were purchased from New England Biolabs (Ipswich, MA, USA) andPromega Corporation (Fitchburg, WI, USA). Expression vector pARD33 wasconstructed as shown in Fig. 2A, by cloning Alrd and AldH genes in cloning vectorpBAD33 with restriction sites of kpnI/SalI and SalI/HindIII, respectively.
The plasmid, pBHB2, was kindly provided by the Yuan Ze PHA team at Yuan ZeUniversity (Taiwan) as mentioned in Table 1. This plasmid contains P3HB synthesisgenes phaCABRE, from R. eutropha H16 and a bacterial hemoglobin gene (vgb gene)from V. hemoglobin with ampicillin-resistance gene (AmpR). The three main en-zymes that has beenwell established for the conversion of acetyl-CoA to the P3HB inR. eutropha are: phaA (b-ketothiolase; EC 2.3.1.9), phaB (NADPH-dependent ace-toacetyl-CoA reductase; EC 1.1.1.36) and phaC (PHA polymerase; EC 2.3.1) (28). Forthis plasmid, Isopropyl b-D-1-thiogalactopyranoside (IPTG) acted as an inducer forprotein regulation and was purchased from SigmaeAldrich Corporation (St. Louis,MO, USA). The expression vectors pARD33 and pBHB2 were transformed into E. coliBL21(lDE3) individually for the construction of recombinant strains with singleplasmid: E. coli BL21_pARD33 and E. coli BL21_pBHB2, respectively (as shown inFig. 2B and C). The size of amplified Alrd and AldH genes, plasmids pBAD33, pARD33and pBHB2 were confirmed by running 0.8% agarose gel, stained with ethidiumbromide and visualized under UV light in Dolphin-Doc Plus (Wealtec Corp., NV,USA). The E. coli BL21_pARD33-BHB2 was constructed with transformation of twoplasmids (pBHB2 and pARD33) one after another in single host, E. coli BL21(lDE3) asshown in Fig. 2D. The constructed recombinant strains were subsequently culturedand applied for further study.
Culture medium The LuriaeBertani (LB) medium was used to prepare LBbroth and LB plates. LB broth contained: 10.0 g L�1 of tryptone, 5.0 g L�1 of yeastextract and 10.0 g L�1 of NaCl, whereas LB plates contained ingredient of LB brothwith addition of 15.0 g L�1 of agar. Appropriate antibiotics were used at followingconcentrations: 50 mg ml�1 chloramphenicol (Cm) and 100 mg ml�1 Ampicillin(Amp) obtained from the SigmaeAldrich.
The crude glycerol used in this study was provided by Greatec Green EnergyCompany, a local biodiesel plant in Taiwan. Greatec Green Energy produces biodieselfrom refined waste cooking oil using sodium methoxide as catalyst. The crudeglycerol was composed of 80% crude glycerol, 11% water, 9% metal ions and otherimpurities. All trials were conducted using the same batch of crude glycerol. Theminimal define medium used in this study contained 30 g L�1 crude glycerol,6.70 g L�1 of Na2HPO4-7H2O, 2.50 g L�1 of (NH4)2SO4,1.50 g L�1 of KH2PO4, 0.20 g L�1
of MgSO4-7H2O, 10.00 mg L�1 of CaCl2 and 5.0 ml of trace metal solution (6.84 g L�1
of H3BO3, 6.00 g L�1 of Na2EDTA, 0.86 g L�1 of MnCl2-4H2O, 0.29 g L�1 of FeCl3-6H2O,0.06 g L�1 of ZnCl2, 0.026 g L�1 of CoCl2-6H2O and 0.002 g L�1 of CuSO4-5H2O) and2 g L�1 of yeast extract. Most of the chemicals were obtained from SigmaeAldrich. Toprevent reaction between chemicals; trace metal solution, crude glycerol, yeastextract and salts were autoclaved separately for 15 min at 121 �C while antibioticswere sterile filtered.

FIG. 1. Metabolic pathways involved in glycerol metabolism in Escherichia coli. Abbreviations: Alrd, aldehyde reductase (EC 1.1.1.21); AldH, aldehyde dehydrogenase (EC 1.2.1.3);phaA, b-ketothiolase (EC 2.3.1.9), phaB, NADPH-dependent acetoacetyl-CoA reductase (EC 1.1.1.36) and phaC, PHA polymerase (EC 2.3.1).
VOL. 117, 2014 SIMULTANEOUS REFINERY OF CHEMICALS FROM GLYCEROL 345
Culture condition Each recombinant strain was grown from one loop-fullstock culture, streaking on LB plates with appropriate selection marker and incu-bated for overnight at 37 �C. Single colony from plate was used to inoculate 10 mL LBbroth with specified selection marker and was incubated in a rotary shaker(200 rpm) at 37 �C for 24 h. Batch cultures were conducted in 500 mL shake flaskwith 100 mL working volume. A pre-culture (3% v/v) were used to inoculate 100 mlof minimal define medium and incubated in rotary shaker with a speed of 200 rpmat 37 �C for 48e72 h. Samples were drawn periodically from the post inoculation atregular intervals of time.
Assay of expression enzymes The expression of Alrd and AldH proteinswere analyzed on a 12% sodium dodecyl sulfate poly-acrylamide gel electrophoresis(SDS-PAGE) in a vertical slab gel apparatus, followed by Coomassie Brilliant Bluestaining. The protein concentrations in the supernatant and cell pellet extract aftersonication of wet cell-pallets were determined by using the Bradford method (34)with bovine serum albumin (BSA) as a standard.
The enzyme activities were calculated from the linear slope of increasingabsorption of NADþ/NADPþ reduced to NADH/NADPH (mM mL�1) at 340 nm,respectively. The Alrd activity was performed in a 1 mL mixture at 37 �C by using4.67 mM D,L-glyceraldehyde as substrate in a buffering solution comprised of0.25 M sodium phosphate, 0.38 M ammonium sulfate, 0.11 mM NADPH and0.1 mM EDTA at pH 6.8 (35). The activity of AldH was performed in a 1 mLmixture containing 50 mM sodium phosphate buffering solution at pH 7.5,50e100 mL of crude cell extract or 10e15 mg of purified enzyme, 0.5 mM NADþ
and 1 mM D,L-glyceraldehyde as substrate (36). One unit of enzyme activity isdefined as the amount of protein that produces 1 pmol NADH or NADPH perminute.
Analytical methods Biomass was measured by measuring dry cell weightwhich in turn was correlated with optical density. Cell growth was monitored by
measuring the optical density of culture at 600 nm, on UltraSpec 2100 proUVeVisible spectrophotometer (GE Healthcare, Wauwatosa, WI, USA). In this case,original mediumwas used as the blank. Three consecutive samples were drawn andanalyzed each time for the accuracy of work.
The concentration of bioethanol and residual crude glycerol along with acetatewas analyzed on a LC-2000 plus series high performance liquid chromatography(HPLC) systems (Jasco-Analytical Instruments, Japan) equipped with an ionexclusion column ICEeIONe300 (Transgenomic Labs, Omaha, NE, USA) operatedat 60 �C. Firstly, the samples were filtered through 0.45 mm of HEPA filters(Billerica, MA, USA) and 10 ml was auto-injected to the HPLC. The refractive index(RI-2031 plus) detector at 35 �C was employed to analyze these samples. Thesamples were eluted at 0.4 mL min�1 with 5 mM sulfuric acid (0.0085 N H2SO4)prepared in HPLC grade water. Data were processed and analyzed using theChromNAV software.
P3HB was recovered from the dry cell mass through solvent extraction andthe P3HB content (wt %) was expressed as percentage of P3HB mass in dry cellmass. Briefly, 20 mg of P3HB-containing cell dry weight was mix with 2 mL ofchloroform and 2 mL of methanol solution containing sulfuric acid (1 mL ofH2SO4 and 99 mL of methanol). The solution was incubated at 100 �C for 15 h,and then 1 mL of 1 M NaCl was added to facilitate phase separation. After re-covery of organic (chloroform) phase, 20 mL of gas chromatograph internalstandard (GCIS) was added, and 300 mL of bottom organic layer was collected assample. For analysis, 0.5 mL of sample was injected into a Shimadzu gas chro-matograph GC-2014 system (Shimadzu Scientific Instruments, Columbia, MD,USA) equipped with a flame ionization detector (GC-FID) and a PhenomenexZebron ZB-5 capillary column (30 m � 0.25 mm � 0.25 mm). Oven temperaturewas held at 100 �C. Both injector and detector temperatures were set at 220 �C.Nitrogen was used as the carrier gas and diphenyl ether was used as internalstandard. Data were processed and analyzed by using Peak-ABC software. The
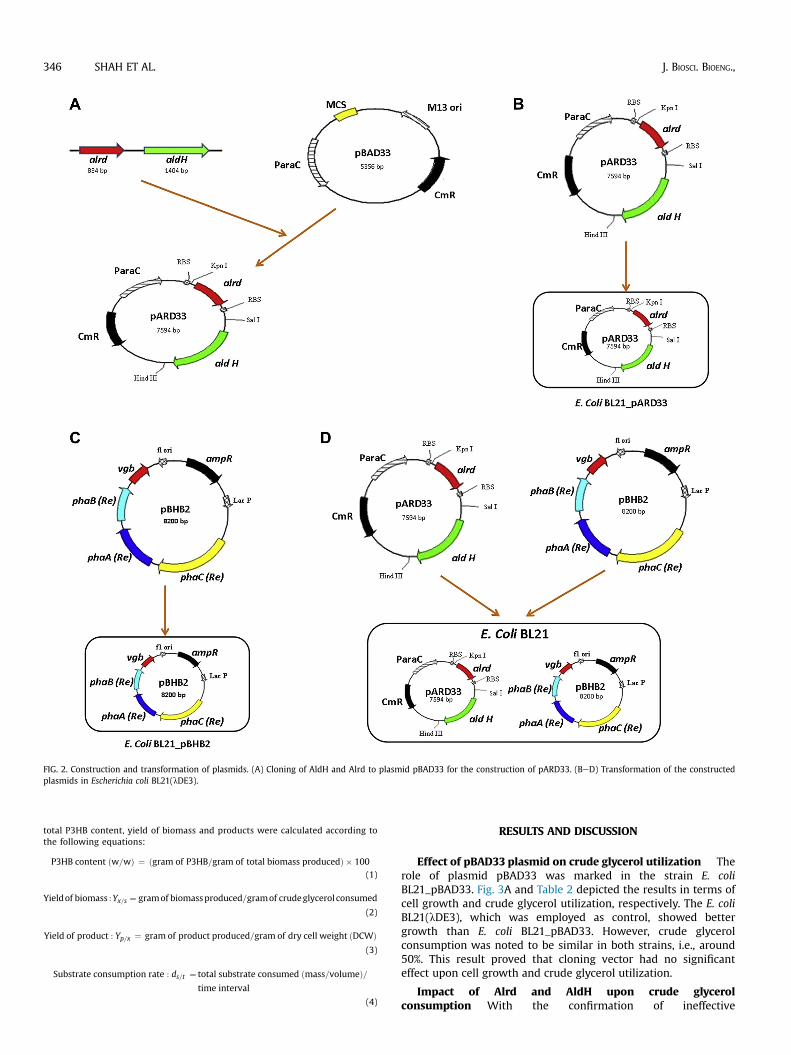
FIG. 2. Construction and transformation of plasmids. (A) Cloning of AldH and Alrd to plasmid pBAD33 for the construction of pARD33. (BeD) Transformation of the constructedplasmids in Escherichia coli BL21(lDE3).
346 SHAH ET AL. J. BIOSCI. BIOENG.,
total P3HB content, yield of biomass and products were calculated according tothe following equations:
P3HB content ðw=wÞ ¼ ðgram of P3HB=gram of total biomass producedÞ � 100(1)
Yieldof biomass :Yx=s ¼ gramof biomassproduced=gramof crudeglycerol consumed
(2)
Yield of product : Yp=x ¼ gram of product produced=gram of dry cell weight ðDCWÞ(3)
Substrate consumption rate : ds=t ¼ total substrate consumed ðmass=volumeÞ=time interval
(4)
RESULTS AND DISCUSSION
Effect of pBAD33 plasmid on crude glycerol utilization Therole of plasmid pBAD33 was marked in the strain E. coliBL21_pBAD33. Fig. 3A and Table 2 depicted the results in terms ofcell growth and crude glycerol utilization, respectively. The E. coliBL21(lDE3), which was employed as control, showed bettergrowth than E. coli BL21_pBAD33. However, crude glycerolconsumption was noted to be similar in both strains, i.e., around50%. This result proved that cloning vector had no significanteffect upon cell growth and crude glycerol utilization.
Impact of Alrd and AldH upon crude glycerolconsumption With the confirmation of ineffective
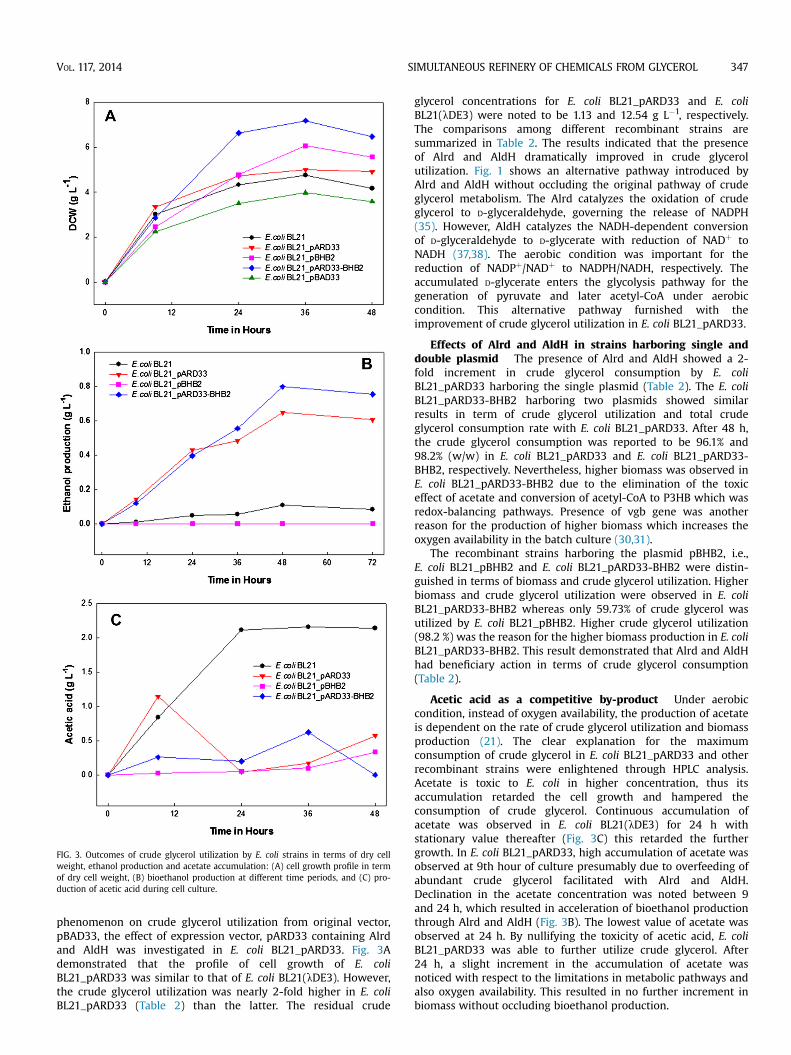
FIG. 3. Outcomes of crude glycerol utilization by E. coli strains in terms of dry cellweight, ethanol production and acetate accumulation: (A) cell growth profile in termof dry cell weight, (B) bioethanol production at different time periods, and (C) pro-duction of acetic acid during cell culture.
VOL. 117, 2014 SIMULTANEOUS REFINERY OF CHEMICALS FROM GLYCEROL 347
phenomenon on crude glycerol utilization from original vector,pBAD33, the effect of expression vector, pARD33 containing Alrdand AldH was investigated in E. coli BL21_pARD33. Fig. 3Ademonstrated that the profile of cell growth of E. coliBL21_pARD33 was similar to that of E. coli BL21(lDE3). However,the crude glycerol utilization was nearly 2-fold higher in E. coliBL21_pARD33 (Table 2) than the latter. The residual crude
glycerol concentrations for E. coli BL21_pARD33 and E. coliBL21(lDE3) were noted to be 1.13 and 12.54 g L�1, respectively.The comparisons among different recombinant strains aresummarized in Table 2. The results indicated that the presenceof Alrd and AldH dramatically improved in crude glycerolutilization. Fig. 1 shows an alternative pathway introduced byAlrd and AldH without occluding the original pathway of crudeglycerol metabolism. The Alrd catalyzes the oxidation of crudeglycerol to D-glyceraldehyde, governing the release of NADPH(35). However, AldH catalyzes the NADH-dependent conversionof D-glyceraldehyde to D-glycerate with reduction of NADþ toNADH (37,38). The aerobic condition was important for thereduction of NADPþ/NADþ to NADPH/NADH, respectively. Theaccumulated D-glycerate enters the glycolysis pathway for thegeneration of pyruvate and later acetyl-CoA under aerobiccondition. This alternative pathway furnished with theimprovement of crude glycerol utilization in E. coli BL21_pARD33.
Effects of Alrd and AldH in strains harboring single anddouble plasmid The presence of Alrd and AldH showed a 2-fold increment in crude glycerol consumption by E. coliBL21_pARD33 harboring the single plasmid (Table 2). The E. coliBL21_pARD33-BHB2 harboring two plasmids showed similarresults in term of crude glycerol utilization and total crudeglycerol consumption rate with E. coli BL21_pARD33. After 48 h,the crude glycerol consumption was reported to be 96.1% and98.2% (w/w) in E. coli BL21_pARD33 and E. coli BL21_pARD33-BHB2, respectively. Nevertheless, higher biomass was observed inE. coli BL21_pARD33-BHB2 due to the elimination of the toxiceffect of acetate and conversion of acetyl-CoA to P3HB which wasredox-balancing pathways. Presence of vgb gene was anotherreason for the production of higher biomass which increases theoxygen availability in the batch culture (30,31).
The recombinant strains harboring the plasmid pBHB2, i.e.,E. coli BL21_pBHB2 and E. coli BL21_pARD33-BHB2 were distin-guished in terms of biomass and crude glycerol utilization. Higherbiomass and crude glycerol utilization were observed in E. coliBL21_pARD33-BHB2 whereas only 59.73% of crude glycerol wasutilized by E. coli BL21_pBHB2. Higher crude glycerol utilization(98.2 %) was the reason for the higher biomass production in E. coliBL21_pARD33-BHB2. This result demonstrated that Alrd and AldHhad beneficiary action in terms of crude glycerol consumption(Table 2).
Acetic acid as a competitive by-product Under aerobiccondition, instead of oxygen availability, the production of acetateis dependent on the rate of crude glycerol utilization and biomassproduction (21). The clear explanation for the maximumconsumption of crude glycerol in E. coli BL21_pARD33 and otherrecombinant strains were enlightened through HPLC analysis.Acetate is toxic to E. coli in higher concentration, thus itsaccumulation retarded the cell growth and hampered theconsumption of crude glycerol. Continuous accumulation ofacetate was observed in E. coli BL21(lDE3) for 24 h withstationary value thereafter (Fig. 3C) this retarded the furthergrowth. In E. coli BL21_pARD33, high accumulation of acetate wasobserved at 9th hour of culture presumably due to overfeeding ofabundant crude glycerol facilitated with Alrd and AldH.Declination in the acetate concentration was noted between 9and 24 h, which resulted in acceleration of bioethanol productionthrough Alrd and AldH (Fig. 3B). The lowest value of acetate wasobserved at 24 h. By nullifying the toxicity of acetic acid, E. coliBL21_pARD33 was able to further utilize crude glycerol. After24 h, a slight increment in the accumulation of acetate wasnoticed with respect to the limitations in metabolic pathways andalso oxygen availability. This resulted in no further increment inbiomass without occluding bioethanol production.
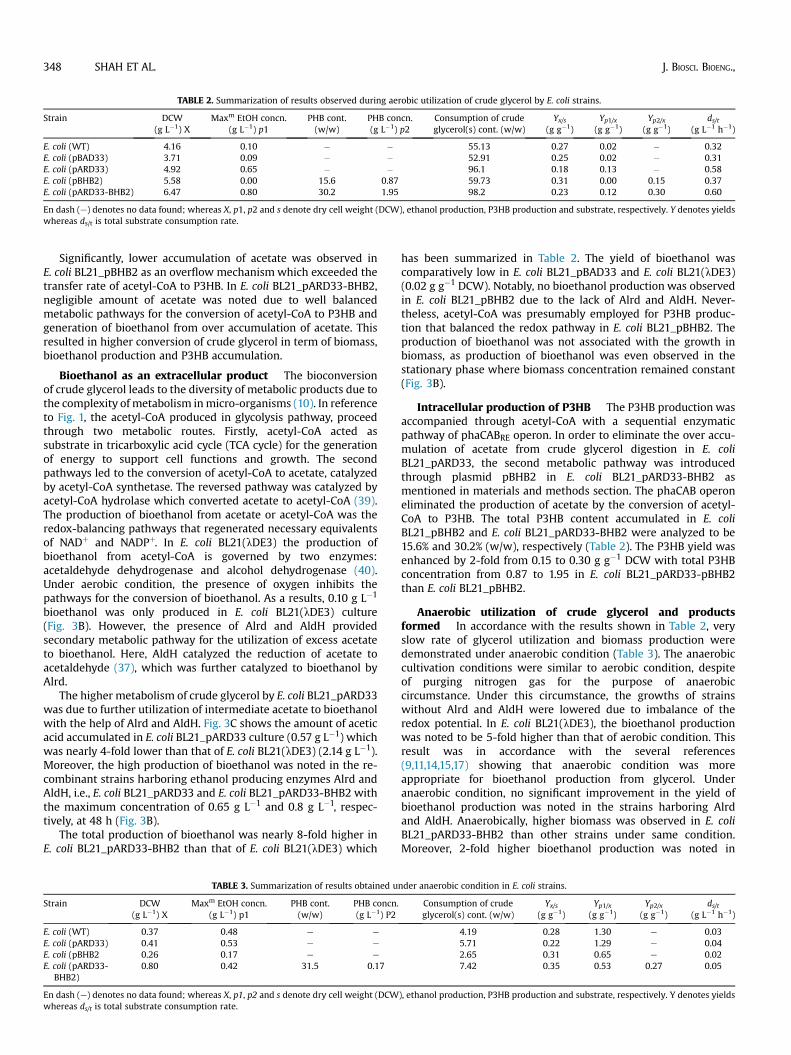
TABLE 2. Summarization of results observed during aerobic utilization of crude glycerol by E. coli strains.
Strain DCW(g L�1) X
Maxm EtOH concn.(g L�1) p1
PHB cont.(w/w)
PHB concn.(g L�1) p2
Consumption of crudeglycerol(s) cont. (w/w)
Yx/s(g g�1)
Yp1/x(g g�1)
Yp2/x(g g�1)
ds/t(g L�1 h�1)
E. coli (WT) 4.16 0.10 � � 55.13 0.27 0.02 � 0.32E. coli (pBAD33) 3.71 0.09 � � 52.91 0.25 0.02 � 0.31E. coli (pARD33) 4.92 0.65 � � 96.1 0.18 0.13 � 0.58E. coli (pBHB2) 5.58 0.00 15.6 0.87 59.73 0.31 0.00 0.15 0.37E. coli (pARD33-BHB2) 6.47 0.80 30.2 1.95 98.2 0.23 0.12 0.30 0.60
En dash (e) denotes no data found; whereas X, p1, p2 and s denote dry cell weight (DCW), ethanol production, P3HB production and substrate, respectively. Y denotes yieldswhereas ds/t is total substrate consumption rate.
348 SHAH ET AL. J. BIOSCI. BIOENG.,
Significantly, lower accumulation of acetate was observed inE. coli BL21_pBHB2 as an overflow mechanism which exceeded thetransfer rate of acetyl-CoA to P3HB. In E. coli BL21_pARD33-BHB2,negligible amount of acetate was noted due to well balancedmetabolic pathways for the conversion of acetyl-CoA to P3HB andgeneration of bioethanol from over accumulation of acetate. Thisresulted in higher conversion of crude glycerol in term of biomass,bioethanol production and P3HB accumulation.
Bioethanol as an extracellular product The bioconversionof crude glycerol leads to the diversity of metabolic products due tothe complexity of metabolism inmicro-organisms (10). In referenceto Fig. 1, the acetyl-CoA produced in glycolysis pathway, proceedthrough two metabolic routes. Firstly, acetyl-CoA acted assubstrate in tricarboxylic acid cycle (TCA cycle) for the generationof energy to support cell functions and growth. The secondpathways led to the conversion of acetyl-CoA to acetate, catalyzedby acetyl-CoA synthetase. The reversed pathway was catalyzed byacetyl-CoA hydrolase which converted acetate to acetyl-CoA (39).The production of bioethanol from acetate or acetyl-CoA was theredox-balancing pathways that regenerated necessary equivalentsof NADþ and NADPþ. In E. coli BL21(lDE3) the production ofbioethanol from acetyl-CoA is governed by two enzymes:acetaldehyde dehydrogenase and alcohol dehydrogenase (40).Under aerobic condition, the presence of oxygen inhibits thepathways for the conversion of bioethanol. As a results, 0.10 g L�1
bioethanol was only produced in E. coli BL21(lDE3) culture(Fig. 3B). However, the presence of Alrd and AldH providedsecondary metabolic pathway for the utilization of excess acetateto bioethanol. Here, AldH catalyzed the reduction of acetate toacetaldehyde (37), which was further catalyzed to bioethanol byAlrd.
The higher metabolism of crude glycerol by E. coli BL21_pARD33was due to further utilization of intermediate acetate to bioethanolwith the help of Alrd and AldH. Fig. 3C shows the amount of aceticacid accumulated in E. coli BL21_pARD33 culture (0.57 g L�1) whichwas nearly 4-fold lower than that of E. coli BL21(lDE3) (2.14 g L�1).Moreover, the high production of bioethanol was noted in the re-combinant strains harboring ethanol producing enzymes Alrd andAldH, i.e., E. coli BL21_pARD33 and E. coli BL21_pARD33-BHB2 withthe maximum concentration of 0.65 g L�1 and 0.8 g L�1, respec-tively, at 48 h (Fig. 3B).
The total production of bioethanol was nearly 8-fold higher inE. coli BL21_pARD33-BHB2 than that of E. coli BL21(lDE3) which
TABLE 3. Summarization of results obtained u
Strain DCW(g L�1) X
Maxm EtOH concn.(g L�1) p1
PHB cont.(w/w)
PHB concn.(g L�1) P2
E. coli (WT) 0.37 0.48 e e
E. coli (pARD33) 0.41 0.53 e e
E. coli (pBHB2 0.26 0.17 e e
E. coli (pARD33-BHB2)
0.80 0.42 31.5 0.17
En dash (e) denotes no data found; whereas X, p1, p2 and s denote dry cell weight (DCWwhereas ds/t is total substrate consumption rate.
has been summarized in Table 2. The yield of bioethanol wascomparatively low in E. coli BL21_pBAD33 and E. coli BL21(lDE3)(0.02 g g�1 DCW). Notably, no bioethanol production was observedin E. coli BL21_pBHB2 due to the lack of Alrd and AldH. Never-theless, acetyl-CoA was presumably employed for P3HB produc-tion that balanced the redox pathway in E. coli BL21_pBHB2. Theproduction of bioethanol was not associated with the growth inbiomass, as production of bioethanol was even observed in thestationary phase where biomass concentration remained constant(Fig. 3B).
Intracellular production of P3HB The P3HB production wasaccompanied through acetyl-CoA with a sequential enzymaticpathway of phaCABRE operon. In order to eliminate the over accu-mulation of acetate from crude glycerol digestion in E. coliBL21_pARD33, the second metabolic pathway was introducedthrough plasmid pBHB2 in E. coli BL21_pARD33-BHB2 asmentioned in materials and methods section. The phaCAB operoneliminated the production of acetate by the conversion of acetyl-CoA to P3HB. The total P3HB content accumulated in E. coliBL21_pBHB2 and E. coli BL21_pARD33-BHB2 were analyzed to be15.6% and 30.2% (w/w), respectively (Table 2). The P3HB yield wasenhanced by 2-fold from 0.15 to 0.30 g g�1 DCW with total P3HBconcentration from 0.87 to 1.95 in E. coli BL21_pARD33-pBHB2than E. coli BL21_pBHB2.
Anaerobic utilization of crude glycerol and productsformed In accordance with the results shown in Table 2, veryslow rate of glycerol utilization and biomass production weredemonstrated under anaerobic condition (Table 3). The anaerobiccultivation conditions were similar to aerobic condition, despiteof purging nitrogen gas for the purpose of anaerobiccircumstance. Under this circumstance, the growths of strainswithout Alrd and AldH were lowered due to imbalance of theredox potential. In E. coli BL21(lDE3), the bioethanol productionwas noted to be 5-fold higher than that of aerobic condition. Thisresult was in accordance with the several references(9,11,14,15,17) showing that anaerobic condition was moreappropriate for bioethanol production from glycerol. Underanaerobic condition, no significant improvement in the yield ofbioethanol production was noted in the strains harboring Alrdand AldH. Anaerobically, higher biomass was observed in E. coliBL21_pARD33-BHB2 than other strains under same condition.Moreover, 2-fold higher bioethanol production was noted in
nder anaerobic condition in E. coli strains.
Consumption of crudeglycerol(s) cont. (w/w)
Yx/s(g g�1)
Yp1/x(g g�1)
Yp2/x(g g�1)
ds/t(g L�1 h�1)
4.19 0.28 1.30 e 0.035.71 0.22 1.29 e 0.042.65 0.31 0.65 e 0.027.42 0.35 0.53 0.27 0.05
), ethanol production, P3HB production and substrate, respectively. Y denotes yields

VOL. 117, 2014 SIMULTANEOUS REFINERY OF CHEMICALS FROM GLYCEROL 349
E. coli BL21_pARD33-BHB2 under aerobic condition than anaerobic.In spite of no bioethanol production in E. coli BL21_pBHB2 underaerobic condition, slightly bioethanol production was notedanaerobically. This was for sure to balance the redox-equivalentPathways. However no P3HB was accounted because ofsignificantly low biomass.
The production of bioethanol under anaerobic culturewas lowerthan the aerobic culture in the recombinant strains harboring Alrdand AldH. This showed the beneficiary role of Alrd and AldH for theproduction of bioethanol aerobically. Improvement of crude glyc-erol utilization under aerobic condition yielded in higher produc-tion of bioethanol and P3HB. Total crude glycerol consumption rate(ds/t) was noted to be nearly 10-fold lower in anaerobic than aerobiccondition (Table 2).
Use of crude glycerol over pure glycerol Crude glycerol is acheap and abundantmaterial containing unreactedmethanol, soap,salts, water and solid organic materials acquire during biodieselproduction (40). Presence of these impurities in crude glycerol hasbeen expected to influence the negative effect in bioconversionprocess. Hence, for achieving a higher yield of bioethanol, manyresearchers focused on using pure glycerol under anaerobic ormicro-aerobic conditions due to high glycerol content andnegligible amount of impurities. Several grades of refined glycerolhave been commercialized that differ in glycerol content.Purification of crude glycerol is an expensive process due to nomarket value of crude glycerol. Therefore, adopting the process ofpurification for crude glycerol or using commercially purifiedglycerol for the production of value added products is aninfeasible route in bio-industry (13e18). Table 4 summarizes theproduction of bioethanol by different strains of E. coli on pure andcrude glycerol, under different cultivation condition. Underaerobic condition, the complete utilization of crude glycerol canbe achieved with higher biomass and low bioethanol yield(12,14). The production of only bioethanol was not a feasibleproposition due to high operational cost and no effective use ofbiomass. Hence, the strategy discussed here resulted in theeffectual use of biomass for the accumulation of P3HB.
TABLE 4. Summary table showing utilization of pure and crude glycerol for bio-ethanol production by different strains of E. coli under different conditions.
Strain Condition Initial glycerolconcn. (g L�1)
Ethanol yields(mol mol�1)
Source
Pure glycerolEscherichia coliMG1655 Anaerobic 10 0.10 13MG1655 (ATCC700926)
Anaerobic 10 0.76 16
MG1655 Anaerobic 10 0.92 12SS1 Anaerobic 20 1.00 15BL21 Anaerobic 20 0.99 15BW25113 Anaerobic 20 0.59 15SY04 (pZSKL0MgldA) Anaerobic 10 1.04 9CT1061LER Micro-aerobic 30 1.02 17EH05 [pZSKLMgldA] Micro-aerobic 60 0.84 14BW25113 Micro-aerobic 20 0.66 14MG1655 Micro-aerobic 20 0.52 14TCS099 e50rep1/pLOI297
Micro-aerobic 40 0.98 18
TCS099/pLOI297 Aerobic 40 0.01 18Crude glycerolEscherichia coliMG1655(pZSKLMgldA)
Anaerobic 50 0.80 17
SY04 (pZSKLMgldA) Anaerobic 10 0.97 9SY04 (pZSKLMgldA) Anaerobic 20 1.02 9BL21_WT Aerobic 30 0.006 This studyBL21_pARD33 Aerobic 30 0.04 This studyBL21_pARD33-BHB2 Aerobic 30 0.06 This study
Awareness to environmental and energy issues have led to thedevelopment of biomass-based bioconversion processes. Produc-tion of fuels and chemicals from crude glycerol is cost-effectiveprocess which has been evaluated in recent years. E. coli can utilizecrude glycerol for the production of bioethanol under aerobic aswell as anaerobic condition (Table 2) (13,14). Bioethanol is only thesecondary product of the crude glycerol fermentation under anycondition. As a result, the yield of bioethanol production is rela-tively low than other metabolites. Hence, several modificationshave been made with the help of biotechnology and molecularbiology for the improvement of bioethanol production from crudeglycerol. In this work, the natural pathway for crude glycerol uti-lization was enhanced by the addition of Alrd and AldH, whichfurther provided the alternative and effective route for bioethanolproduction in E. coli BL21_pARD33 (Fig. 1). Under aerobic condition,acetate was the major product derived through acetyl-CoA. ThephaCAB operon used acetyl-CoA as a substrate for the production ofP3HB which reduced the formation of acetate (28). Higher yield ofbioethanol was observed under aerobic condition in the strainsharboring Alrd and AldH. This result was contradictory with thegeneral belief that high bioethanol can be only produced underanaerobic condition. This was probably due to the role of Alrd andAldH in aerobic condition which provided with addition pathwayfor the production of bioethanol and also balanced the redox-equivalents. The production of bioethanol and P3HB at the sametime facilitated with high degree of value-added products fromcrude glycerol under both aerobic and anaerobic condition. How-ever, higher yields of bioethanol and P3HB were noted under aer-obic condition along with complete utilization of crude glycerol inE. coli BL21_pARD33-BHB2 at 48 h of post-culture.
Considering the huge crude glycerol surplus, our findingestablished a newmodel for the conversion of crude glycerol underaerobic condition by using an E. coli-based platform. The totalproduction of bioethanol and P3HB content was noted to be0.8 g L�1 and 30.2% (w/w), respectively, in E. coli BL21_pARD33-BHB2.
ACKNOWLEDGMENTS
This research was supported by National Science Council(Taiwan) with funding number 99-2622-E-155-002-CC2 and 101-2221-E-155-042. We thankfully acknowledge help from Prof.Shaw-Shan Wang (Distinguished Professor, Department of Chemi-cal Engineering and Materials Science, Yuan Ze University) forimproving English grammar, verb usage, sentence structure andgeneral readability of the manuscript revision.
References
1. Nigam, P. S. and Singh, A.: Production of liquid biofuels from renewable re-sources, Prog. Energy Combust. Sci., 37, 52e68 (2011).
2. Janaun, J. and Ellis, N.: Perspectives on biodiesel as a sustainable fuel, Renew.Sust. Energy Rev., 14, 1312e1320 (2010).
3. Girard, P. and Fallot, A.: Review of existing and emerging technologies for theproduction of biofuels in developing countries, Energy Sustain. Dev., 10,92e108 (2006).
4. Deenanath, E. D., Iyuke, S., and Rubold, K.: The bioethanol industry in sub-Saharan Africa: history, challenges, and prospects, J. Biomed. Biotechnol., 2012,4416491 (2012).
5. Kolesarova, N., Hutnan, M., Bodik, I., and Spalkova, V.: Utilization of biodieselby-products for biogas production, J. Biomed. Biotechnol., 2011, 126798 (2011).
6. Melero, J. A., Vicente, G., Paniagua, M., Morales, G., and Munoz, P.: Ether-ification of biodiesel-derived crude glycerol with bioethanol for fuel formula-tion over sulfonic modified catalysts, Bioresour. Technol., 103, 142e151 (2012).
7. Zhang, Y., Dub, M. A., McLean, D. D., and Kates, M.: Biodiesel production fromwaste cooking oil: economic assessment and sensitivity analysis, Bioresour.Technol., 90, 229e240 (2003).

350 SHAH ET AL. J. BIOSCI. BIOENG.,
8. Dobson, R., Gray, V., and Rumbold, K.: Microbial utilization of crude glycerolfor the production of value-added products, J. Ind. Microbiol. Biotechnol., 39,217e226 (2012).
9. Yazdani, S. S. and Gonzalez, R.: Engineering Escherichia coli for the efficientconversion of crude glycerol to ethanol and co-products, Metab. Eng., 10,340e351 (2008).
10. Stephanopoulos, G.: Challenges in engineering microbes for biofuels produc-tion, Science, 315, 801e804 (2007).
11. Gonzalez, R., Murarka, A., Dharmadi, Y., and Yazdani, S. S.: A new modelfor the anaerobic fermentation of crude glycerol in enteric bacteria: trunkand auxiliary pathways in Escherichia coli, Metab. Eng., 10, 234e245(2008).
12. Murarka, A., Dharmadi, Y., Yazdani, S. S., and Gonzalez, R.: Fermentativeutilization of crude glycerol in Escherichia coli and its implication for the pro-duction of fuels and chemicals, Appl. Environ. Microbiol., 74, 1124e1135(2008).
13. Dharmadi, Y., Murarka, A., and Gonzalez, R.: Anaerobic fermentation of crudeglycerol by Escherichia coli: a new platform for metabolic engineering, Bio-technol. Bioeng., 94, 821e829 (2006).
14. Durnin, G., Clomburg, J., Yeates, Z., Alvarez, P. J. J., Zygourakis, K.,Campbell, P., and Gonzalez, R.: Understanding and harnessing the micro-aerobic metabolism of glycerol in Escherichia coli, Biotechnol. Bioeng., 103,148e161 (2009).
15. Suhaimi, S. N., Phang, L. Y., Maeda, T., Abd-Aziz, S., Wakisaka, M., Shirai, Y.,and Hassan, M. A.: Bioconversion of glycerol for bioethanol production usingisolated Escherichia coli SS1, Braz. J. Microbiol., 43, 506e516 (2012).
16. Chaudhary, N., Ngadi, M. O., Simpson, B. K., and Kassama, L. S.: Biosynthesisof ethanol and hydrogen by crude glycerol fermentation using Escherichia coli,Adv. Chem. Eng. Sci., 1, 83e89 (2011).
17. Nikel, P. I., Ramirez, M. C., Pettinari, M. J., Mendez, B. S., and Galvagno, M. A.:Methanol synthesis from crude glycerol by Escherichia coli redox mutantsexpressing adhE from Leuconostoc mesenteroides, J. Appl. Microbiol., 109,492e504 (2010).
18. Trinh, C. T. and Srienc, F.:Metabolic engineering of Escherichia coli for efficientconversion of crude glycerol to ethanol, Appl. Environ. Microbiol., 75,6696e6705 (2009).
19. El-Kabbani, O., Old, S. E., Ginell, S. L., and Carper, D. A.: Aldose and aldehydereductases: structure-function studies on the coenzyme and inhibitor-bindingsites, Mol. Vis., 5, 20e25 (1999).
20. Perozich, J., Nicholas, H., Wang, B. C., Lindahl, R., and Hempel, J.: Relation-ships within the aldehyde dehydrogenase extended family, Protein Sci., 8,137e146 (1999).
21. Eiteman, M. A. and Altman, E.: Overcoming acetate in Escherichia coli re-combinant protein fermentation, Trends Biotechnol., 24, 530e536 (2006).
22. Chen, G. Q.: A microbial polyhydroxyalkanoates (PHA) based bio- and mate-rials industry, Chem. Soc. Rev., 38, 2434e2446 (2009).
23. Andreessen, B., Lange, A. B., Robenek, H., and Steinbuchel, A.: Conversion ofglycerol to poly(3-hydroxypropionate) in recombinant Escherichia coli, Appl.Environ. Microbiol., 76, 622e626 (2010).
24. Lopez, N. I., Floccari, M. E., Steinbuchel, A., Garcia, A. F., and Mendez, B. S.:Effect of poly(3-hydroxybutyrate) (PHB) content on the starvation-survival ofbacteria in natural waters, FEMS Microbiol. Ecol., 16, 95e102 (1995).
25. Andre, A., Chatzifragkou, A., Diamantopoulou, P., Sarris, D., Philippoussis, A.,Galiotou-Panayotou,M., Komaitis, M., and Papanikolaou, S.: Biotechnologicalconversions of bio-diesel-derived crude glycerol by Yarrowia lipolytica strains,Eng. Life Sci., 9, 468e478 (2009).
26. Garcia, I. L., Lopez, J. A., Dorado, M. P., Kopsahelis, N., Alexandri, M.,Papanikolaou, S., Villa, M. A., and Koutinas, A. A.: Evaluation of by-productsfrom the biodiesel industry as fermentation feedstock for poly(3-hydrox-ybutyrate-co-3-hydroxyvalerate) production by Cupriavidus necator, Bioresour.Technol., 130, 16e22 (2013).
27. Ibrahim, M. H. A. and Steinbuchel, A.: Poly(3-hydroxybutyrate) productionfrom glycerol by Zobellella denitrificans MW1 via high-cell-density fed-batchfermentation and simplified solvent extraction, Appl. Environ. Microbiol., 17,6222e6231 (2009).
28. Sudesh, K., Abe, H., and Doi, Y.: Synthesis, structure and properties of poly-hydroxyalkanoates: biological polyesters, Prog. Polym. Sci.,25, 1503e1555 (2000).
29. Chien, C. C., Hong, C. C., Soo, P. C., Wei, Y. H., Chen, S. Y., Cheng, M. L., andSun, Y. M.: Functional expression of phaCAB genes from Cupriavidus taiwa-nensis strain 184 in Escherichia coli for polyhydroxybutyrate production, Appl.Biochem. Biotechnol., 162, 2355e2364 (2010).
30. Urgun-Demirtas, M., Pagilla, K. R., and Stark, B.: Enhanced kinetics ofgenetically engineered Burkholderia cepacia: the role of vgb in the hypoxicmetabolism of 2-CBA, Biotechnol. Bioeng., 87, 110e118 (2004).
31. Horng, Y. T., Chang, K. C., Chien, C. C., Wei, Y. H., Sun, Y. M., and Soo, P. C.:Enhanced polyhydroxybutyrate (PHB) production via the coexpressed phaCABand vgb genes controlled by arabinose P(BAD) promoter in Escherichia coli, Lett.Appl. Microbiol., 50, 158e167 (2010).
32. Jeong, H., Barbe, V., Lee, C. H., Vallenet, D., Yu, D. S., Choi, S. H., Couloux, A.,Lee, S. W., Yoon, S. H., and Cattolico, L.: Genome sequences of Escherichia coliB strains REL606 and BL21(DE3), J. Mol. Biol., 394, 644e652 (2009).
33. Guzman, L. M., Belin, D., Carson, M. J., and Beckwith, J.: Tight regulation,modulation, and high-level expression by vectors containing the arabinosepBAD promoter, J. Bacteriol., 177, 4121e4130 (1995).
34. Bradford, M. M.: A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principles of protein-dye binding, Anal.Biochem., 72, 248e254 (1976).
35. Del Corso, A., Costantino, L., Rastelli, G., Buono, F., and Mura, U.: Aldosereductase does catalyse the reduction of glyceraldehyde through a stoichio-metric oxidation of NADPH, Exp. Eye Res., 71, 515e521 (2000).
36. Ho, K. K. and Weiner, H.: Isolation and characterization of an aldehyde de-hydrogenase encoded by the aldB gene of Escherichia coli, J. Bacteriol., 187,1067e1073 (2005).
37. Eriksson, C. J. P., Saarenmaa, T. P. S., Bykov, I. L., and Heino, P. U.: Acceler-ation of ethanol and acetaldehyde oxidation by D-glycerate in rats, Metab. Clin.Exp., 56, 895e898 (2007).
38. Millo, H. and Werman, M. J.: Hepatic fructose-metabolizing enzymes andrelated metabolites: role of dietary copper and gender, J. Nutr. Biochem., 11,374e381 (2000).
39. Shimazu, T., Hirschey, M. D., Huang, J. Y., Ho, L. T., and Verdin, E.: Acetatemetabolism and aging: an emerging connection, Mech. Ageing Develop., 131,511e516 (2010).
40. Kumar, M. and Gayen, K.: Developments in biobutanol production: new in-sights, Appl. Energy, 88, 1999e2012 (2011).