Embed Size (px)
Citation preview

Wilhelm Roux' Archly 164, 341--358 (1970)
Hemmung und Induktion yon Proteinsynthesen durch Actinomycin in den wachsenden Oocyten
yon M u s c a domes t ica *
C~RIST~A~ PETZ~LT und KAlCLHEINZ BIER t
Zoologisches Institut der Universit~t Miinster
Eingegangen am 16. Oktober 1969
Inhibition and Induction o/Protein Syntheses by Actinomycin in the Growing Oocytes o/Musca domestica
Summary. The protein synthesis of growing oocytes of Musca domestica L. is investigated by radioautographic and electrophoretic methods on an intermediate developmental stage (stage 3), and after the end of growth (stage 6) in the normal development and after treatment with Aetinomyein. The very high RNA synthesis of the nurse cells and of the follicle epithelium of growing oocytes is completely blocked by injection of 16 ~g/g body weight Actinomycin. The protein synthesis in the euplasm of ~ growing oocyte is relatively high, the stage 6 shows a high protein synthesis in spite of the absence of RNA synthesis. The proteins of the ovary are separated electrophoreticaUy on Cellogel-strips. The ratio of the newly-synthetized proteins is determined after incorporation (30 min) of sAC-labelled amino acids. The protein synthesis is stimulated after 4--6 hours treatment with Actinomycin. After a longer influence of the antibiotic, synthesis of the egg-nurse cell unit decreases to below the control value. The ripe oocyte does not react autoradiographieally to Actinomycin. Only some fractions of the newly-synthetized euplasmatic proteins are sensitive to treatment with Aetinomyein. After intermediate incubation times with Ac~inomyein new proteins appear in the ovary and their synthesis is demonstrable in part even after treatment for 48 hours. There is no newly-synthetized t~NA in the ripe oocyte, which, nevertheless, reacts with Actinomycin to inhibit some and to stimulate some new protein synthesis. In this case the action of the antibiotic is unknown.
Zusammen]assung. Die Proteinsynthese der wachsenden Oocyten von Musca domestica L. wurde autoradiographisch und elektrophoretisch auf einer mittleren Entwicklungsstufe (Sta- dium 3) und nach dem AbsehhiB des Wachstums {Stadium 6) in der Normalentwickhing und nach Applikation yon Actinomycin untersucht. Die sehr hohe RNS-Synthese der iN~hrzel]en und des FoIlikelepithels yon wachsenden Oocyten wurde durch Injektion yon 16 ~g/g KSrper- gewicht Aetinomycin vSllig blockiert. Die Proteinsynthese im Euplasma der waehsenden Oocyte ist relativ hoch, auch das Stadium 6 zeigt trotz fehlender RNS-Synthese eine kr/~ftige Proteinsynthese. Die Proteine des Ovars wurden elektrophoretisch auf Celhiloseacetat- Streifen getrennt. Der Anteil neusynthetisierter Proteine wurde nach vorangegangener Inkorporation (30 min) yon 14C-Aminos~uren bestimmt. 1Nach 4--6 Std Inkubation mit Aetinomyein wlrd die Proteinsynthese stimu]iert. 1Nach l~ngerer Einwh.kung sinkt die Syntheseleistung im Ein~hrverband allm~hlich unter den Ausgangswert. Die reife Oocyte zeigt, autoradiographisch gesehen, keine Reaktion auf das Antibiotikum. Die Synthese der euplasmatischen Proteine im Eini~hrverband des Stadiums 3 sinkt nach Actinomycin-Behandhmg nicht in toto ab, nut einige Fraktionen erweisen sich als empfindlich. Nach mittleren Inkubationszeiten mit Actinomycin treten dagegen neue Proteine im 0vat auf, deren Synthese in begrenztem Urn- fang auch noch nach 48 Std nachweisbar ist. Der reifen Oocyte steht kcine neusyntheti- sierte R1NS zur Vefffigung. Sie reagiert aber ebenfalls auf Actinomycin mit partiellen ttem- mungen und Neusynthesen yon Proteinen. Der Wirkungsmechanismus des Antibiotikums bleibt in diesem Falle offen.
* Die Untersuchungen wurden mit Mitteln unterstiitzt, die yon der Deutschen Forschungs- gemeinschaft Herrn Prof. Dr. K. Bier zur Verfiigung gestellt wurden.

342 C. Petzelt und K. Bier:
Einleitung Die I t e m m u n g der RNS-Syn these bietet die M6gliehkeit, den Prozeg der
EiweiBbildung bereits auf der Ebene der Transkript ion zu bloekieren. Dabei finder Aet inomycin Verwendung (Reich und Goldberg, 1964), ein Antibiot ikum, das in v ivo-Komplexe mit der DNS bildet, wobei sieh besonders Desoxyguanosin durch gutes Komplexbindungsverm6gen auszeichnet (Kersten und Kersten, 1962). Die t)roteinsynthese ist abet nicht immer mit Act inomycin zu bloekieren (Papa- eonstantinou, 1967). Sogar Akt ivierungen wurden unter dem EinfiuB des Anti- biot ikums beobachtet (Eliasson, 1967). Es mfissen demnach stabile Messenger vorhanden sein, die naeh Blockierung der g N S - S y n t h e s e aktiviert werden. Zn gleichen Ergebnissen, ni~mlieh einer Steigerung oder zumindest einem Gleich- bleiben yon Enzymaktivit /~ten naeh Behandlung mit Aetinomycin, kommen noeh andere Autoren (Moog, 1964; Garren et al., 1964; Rosen et al., 1964; Maruzov und Oreklovich, 1966; Baker, 1967; Bose, 1967).
Eine gegenfiber Act inomycin unempfindliche Proteinsynthese wurde aueh wghrend der Frf ihembryogenese yon Seeigeln (Gross und Cousineau, 1964) und Amphibien (Braehet et al., 1964) gefunden. Hier werden w~hrend der Oogenese synthetisierte RNS-Matr izen gespeiehert und wiihrend der frfihen Furehungs- periode aktiviert (vgl. Braehet, 1967). Ziel dieser Arbeit war es, die Periode der Synthese stabiler Matr izen-RNS w/~hrend des Wachs tums der Eier zu ermitteln. Zu diesem Zweek wurde die Proteinsynthese w/thrend der Oogenese yon Musca domestica auf ihre Aet inomycin-Empfindl iehkei t untersucht . Als Versuehsobjekt diente die Stubenfliege, weft ihre Ooeyten rasch und synehron heranwaehsen. Aueh wm~den an diesem Tier bereits stimulierende Effekte des Ant ibiot ikums bei der Synthese yon Reservestoffen beobaehtet (Engels und Bier, 1967).
Material und ~Iethode
Die Zucht und Haltung yon M~sca erfolgte bei 21 ~ C und 18 Std-Tag (vgl. Trepte, 1967). Der Reifegrad der Ovarien wurde nach der Stadieneinteilung yon Bier (1963) bestimmt.
Zur Blockierung dcr RNS-Synthese der wachsenden Follikel wurde Actinomyein C 1 = D appliziert. Das zun~ehst verwendete ,,Sanamyein" Bayer (Gemisch aus Aetinomyein C~, C2, Ca, Verhgltnis Saeeharose:Actinomyein = 1000:1) zeigte auf Grund seines starken Zucker- gehaltes unerwiinsehte Nebenwirkungen. Die Dosis des Actinomyein D betrug pro Versuchs- tier 0,4 tzg (--16 ~g/g K6rpergewicht). Die Behandlungszeiten differierten yon 15 rain bis 48 Std.
Autoradiographie. Als Tracer ffir die Proteinsynthese diente ein Gemiseh von gleiehen Teilen aH-Itistidin, att-Lysin, 3H-Leucin und aII-Valin. Diese Aminos~uren sind in den Pro~ teincn der Musca-Ovarien reichlich vorhanden (Willebrand, 1965). Pro Versuchstier wurden 5 fzC (~ 5 ~1) mit einer graduierten Glaskapillare dureh die Intersegmentalh~ute in die Leibes- hShle injiziert. N~ch entspreehender Inkorporationszeit von 20--30 rain wurden die Tiere mit CO 2 bet~ubt und die Ovarien herauspr~pariert. Sodann wurden sic in modifiziertem Car- noyschen Gemiseh (Alkohol: Chloroform : Eisessig = 8:1 : 1) fixiert und nach den iibliehen histologischen VerfMlren in Paraffin (Schmelzpunkt 58 ~ C) eingebetteL Die 10 ~z dieken Schnitte wurden naeh dem Entparaffinieren bei 4 ~ C in 5 % Trichloressigs~ure zur Entfernung der freien Aminosi~uren 15 min gewasehen und ansehliegend mit Kodak-Stripping-Film AR 10 iiberzogen. Nach der Exposition yon 12 Tagen bei 4 ~ C wurden sie mit RodinM 1 : 100 ent- wickelt und mit Euparal eingedeek~.
Zur Auswertu_ng wurden die Silberk6rner fiber den jewefligen Zellpartien im Phasenkon- trast bei 800facher Vergr6fierung ausgez~hlt, wobei der unspezifische ,,Background" sub- trahiert wurde. Es wurde jeweils die Korndichte an drei verschiedenen Stellen tier zu unter-

Hemmung und Induktion yon Proteinsynthesen durch Actinomycin 343
suchenden Struktur bei je vier Quadratfeldern yon je 440 ~2 (Okularmikrometer) ermittelt. Um den Z~hlfehler zu verringern, wurde dieser Vorgang an drei verschiedenen Schnitten desselben Objektes wiederholt und das Mi~tel aus diesen insgesamt neun Zghlungen gewertet.
Elelctrophorese au/Celluloseacetat. Als Tracer fiir die Proteinsynthese diente ein 14C-markiertes Proteinhydrolysat der New England Nukleo-Corp. (start in 0,01 n tiC1 in Aqua bidest, gelSst). Pro Versuchstier wurden 0,6 ~C (~ 6 ~l) injiziert. Die Inkorporationszeit betrug 20--30 min. Actinomycin wurde mit der gleichen Dosis und mit den gleiehen Zeiten wie oben angegeben appliziert.
Die Auswertung erfolgte mit Hilfe eines Methan-Durehfluitz~hlers mit 4 ~-Geometrie (Diinnschicht-Scanner der Firma Berthold, Wildbad). Die Blendenbreite betrug 1 mm, die Durchlaufgesehwindigkeit 60 und 120 mm/h.
Naeh Betgubung der Versuehstiere mit CO 2 wurde unter dem Binokular bei 10facher VergrSl~erung das Abdomen dorsal in den Intersegmentalh~uten mit Uhrmacherpinzetten aufgerissen und die H~molymphe mit einer Glaskapillare abgesaugt. Dann wurden die Ovarien in einem vorgekiihlten 1 ml-Homogenisator nach Zugabe yon 0,1 m] Elektrophorese- Puffer bei niedriger Tourenzahl homogenisiert. Die H~molymphe und 20 ~1 des Ovarhomo- genats wurden mit einer Mikropipette auf Celluloseaeetat-Streifen (Serva, Heidelberg) aufge- tragen find zur Trennung in die entsprechende Elektrophorese-Kammer gelegt. Als Puffer diente Aronson-Gronvall-Puffer (60,5 g Trihydroxymethylaminomethan, 6,6 g Athylendi- amintetraessigs~ure, 4,6 g Borsgure, auf 1 Liter Aqua bidest., pH 9,0). Die Spannung betrug 140 Volt, Laufzeit 180 min, Temperatur 4 ~ C. Nach der Trennung wurden die Streifen in einer ges~ttigten Amidoschwarzl5sung yon MethanohEisessig (9:1) ca. 8 rain ge~rbt und in Methanol:Eisessig (9:]) entfgrbt. Die Transparenz und HHa]tbarkeit der Streifen wurde dureh ein 2 min-Bad in Methanol: Eisessig (85:15) und anschliel~endes Trocknen auf einer Glasplatte bei 60 ~ C erreicht.
Ergebnisse A. A utoradiographische Untersuchungen
1. Normalbed ingungen
Die euplasmat i sche Pro te insyn these im Stadium 3 i s t im Vergleich zu den Synthese le i s tungen der T rophocy ten und des Fol l ike lepi the ls rech t niedrig. Doch ist ihre Intensit i~t insofern bemerkenswer t , a]s der Oocytenkern fast vSllig iner t is t und die notwendige l~NS-Versorgung du tch die Ns erfolgt (Bier, ]963).
Naeh 20 rain I n k u b a t i o n mi t e inem 8It-Aminos~uregemisch is t im Ooplasma eine ziemlich kr~ft ige Markierung zu sehen, die nahezu vS1]ig gleiehm~l~ig den ooeyt~ren R a u m umlau t . Ein Gradien t is t n icht festzustel len, al lenfalls is t im p rox ima len Teil des Ooplasmas eine etwas versti~rkte Synthese le i s tung an den Or ten des t~NS-Zuflusses aus den] N~hrfach zu finden. (~ber dem Oocytenkern sind im Vergleieh zum Ooplasma keine Aktivi t i~tsdifferenzen zu unterscheiden. Ls m a n den Tracer 30 rain und ]~nger einwirken, beginnen die ers ten Anzeichen der p inoeyt~ren Dot te re in lagerung aus der H ~ m o l y m p h e ira A u t o r a d i o g r a m m hervorzu t r e t en und verf~lschen das Bi ld der Synthese le is tung der Oocyte. Des- ha lb wurden l~ngere Inkuba t ionsze i t en als 30 rain vermieden. Die Aufnahme und Ein lagerung yon I t g m o l y m p h p r o t e i n e n in das Do~tersys tem wird an anderer Stelle beschr ieben (Petzel t und Bier, 1970).
Die Pro te insyn these der Ni~hrzellen is t deut l ieh hSher als die des Ooplasmas. Die Nghrzel lkerne s ind st i i rker mark ie r t als alas Cytoplasma, besonders die Nukleolen t r e ten hervor . Dieses Gesamtve rha l t en der P ro te insyn these der Ni~hr- zellen s t i m m t gu t mi t den Befunden iiberein, dab die T rophoey t en als t t a u p t - p roduzen ten yon t~NS im Eini~hrverband eine s t a rk erhShte Aktivit i~t zeigen (Bier, 1963).
23 Wilhelm l~oux' Arch., Bd. 164

344 C. Petzelt und K. Bier:
Abb. 1. Dunkelfeldaulnahme eines Follikels im Stadium 6 nach 30 rain Inkorporation eines 3H-Aminos~uregemisches. Das Ooplasma ist gleichm~l]ig markiert, lediglich im Periplasma
ist der Einbau scheinbar verst~rkt, da dor~ keine inerten Dotterschollen liegen
Eine noch wenig bekannte Rolle in diesem System spielt das Follikelepithel. Sichergestellt ist bis jetzt nur seine Funktion als Durchlaufstation fiir Dotter- proteine, die yon der t t~molymphe dutch Interzellularen an das Oolemm wan- dern (Teller, 1961).
I m Stadium 6 sind die Ni~hrzellen bereits resorbiert und die Follikelzellen degeneriert. Dieses letzte Entwicklungsstadium der Eizelle ist durch das Fehlen einer aktuellen l%NS-Synthese gekennzeichnet. Kontrollversuche mit aH-Uridin bis zu 30 rain Inkubationszeit bestatigen das Fehlen einer RNS-Synthese. Die zu einer Karyosphi~re kondensierten, inaktiven Chromosomen des Oocytenkerns treten ohne Entspiralisierung in die Metaphase I ein (Chandley, 1966; Bier, Kunz und l%ibbert, 1967). Trotzdem werden aIt-Aminosguren eingebaut. Wie Abb. 1 zeigt, existiert in diesem Stadium eine kr/iftige Proteinsynthese, die nahezu v6]lig gleichm/~gig das gesamte Euplasma umfal~t. Zwar wird die Aktivit/it des 0oplasmas des Stadiums 3 nicht ganz erreieht, doch ist die It6he der Markierung um so bemerkenswerter, als ja die Basis, n~mlich die de novo gebildete RNS, v511ig fehlt. Hier zeigt sich bereits in der 0ogenese das Phgnomen einer Protein- synthese ohne aktuelle RNS-Synthese, wie sie fiir die fr/ihe Embryogenese charak- teristiseh ist.
2. Die Proteinsynthese unter Actinomycin-Einflu~
I m Stadium 6 ist unter diesen Bedingungen yon vornherein keine Inhibition der Proteinsynthese durch Actinomycin zu erwarten. Da jedoch wiederholt auch eine Aktivierung der Proteinsynthese unter Einwirkung yon Actinomycin beschrie- ben wurde (s. Einleitung), mul~ ein Versuch gerade in diesem Stadium ohne I%NS- Synthese yon Interesse sein. Weiterhin bleibt durch Applikation dieses Antibio-

t{emmung und Induktion yon Proteinsynthesen durch Actinomyein 345
Tabelle. ~nderung der Protein. und RNS-Synthese nach verschiedenen Inkubationszeiten mit H20 ( ~ t ) und anschlieflender In/corporation (30 rain) yon aH-Aminosdiuren bzw. 3H-Uridin. In beiden Fdillen ergeben sich nach 2 Std keine Di]]erenzen mehr gegeniiber den KontroUen.
% -Angabe ~ Standardabweichung
Pro~einsynthese RN S- Synthese
t (rain) % (Einbaurate) t (rain) % (Einbaurate)
0 100 --1,8 0 100 • 20 89,4 • 1,4 20 91,5 • 1,1 40 97,1 ::J:: 1,9 40 92,0~ 1,6 60 95,3 ::J:: 2,3 60 97,2 • 2,5
120 100 • 2,0 120 100 • 2,2
tikums im Stadium 3 zu kl/~ren, ob diese Proteinsynthese allein auf dem st/~ndigen Naehschub trophoeyti~rer RNS basiert oder ob sie zumindest teilweise bereits naeh ~hnlichen Prinzipien wie im Stadium 6 vor sieh geht. Im ersten Falle wiirde die Proteinsymthese dutch Actinomycin ganz zum Erliegen kommen, w~h- rend die Aktivit/s unter den letzteren Voraussetzungen nur absinken miil~te.
a) Vorversuche. Aetinomyein ist in der angewandten Dosis nieht toxisch. Die Tiere fiberleben bis zu 48 Std nach der Injektion (und wahrseheinlieh ls die Weiterentwiek]ung der Ovarien ist allerdings stark gehemmt. Um die unspezifi- sche Beeinflussung der Syntheserate durch die experimentellen Eingriffe zu prfifen, wurden Kurzzeitinkubationen mit einer Vorinjektion yon It20 stat t Aetino- mycin und in verschiedenem Zeitabstand injizierten aH-Aminos/~uren bzw. att- Uridin vorgenommen. Dabei ergab sich eine Minderung der RNS- und Protein- synthese. Diese Reaktion, die dann auch bei den Versuchen mit Actinomycin auftrat, ist also wahrseheinlieh zum grSgten Teil nicht auf die Wirkung des Injektionsmittels selbst zuriiekzuffihren, sondern seheint eine unmittelbare geak- tion des allgemeinen Stoffweehsels anI die Injektion zu sein. Im Unterschied zur Actinomycinbehandlung zeigt der VerlauI der Syntheseintensit/~t 2 Std naeh der HsO-Injektion keinen Unterschied mehr zu den Kontrollen (Tabe]le). Die elektrophoretischen Befunde (vgl. S. 348) sichern den spezifischen Einbau der applizierten markierten Aminos~uren in Proteine. Trotzdem wurden noch zusiitz- ]ich Versuehe re_it Puromycin nnternommen, um das Fortbestehen einer Protein- synthese nach Actinomycinbehandlung mit einer anderen Methode zu unter- mauern. Puromyein hemmt in der applizierten Dosis yon 4 ~g/Vt die Inkorpo- ration markierter Aminos~uren in den Kontrollen wie aueh in den Tieren, die bis zu 6 Std mit Actinomyein vorbehandelt worden waren, vSllig.
Weiterhin wurde die Wirkung des Actinomycins auf die RNS-Synthese yon Musca noch einmal gepriift, obwohl sie sehon 1/~nger bekannt is~ (Engels und Bier, 1967). Schon nach einer Gesamtinkubation yon 30 rain (10 rain Vorbehand- lung mit Actinomycin und zus/~tzlich 20 rain aH-Uridin-Inkorporation) erfolgt eine BlocMerung der RNS-Synthese zu 98 %. Die geringe restliche Markierung ist auch mit RNase-Behandlung nicht zu beseitigen. Diese so raseh einsetzende Inhibie- rung h/~lt bis 48 Std Aetinomycin-Einwirkung in gleichem AusmaB an.
b) Resultate. In den drei untersuchten Zellarten des Eifollikels wirkt sich die Applikation yon Aetinomycin untersehiedlieh aus (Abb. 2).
23*
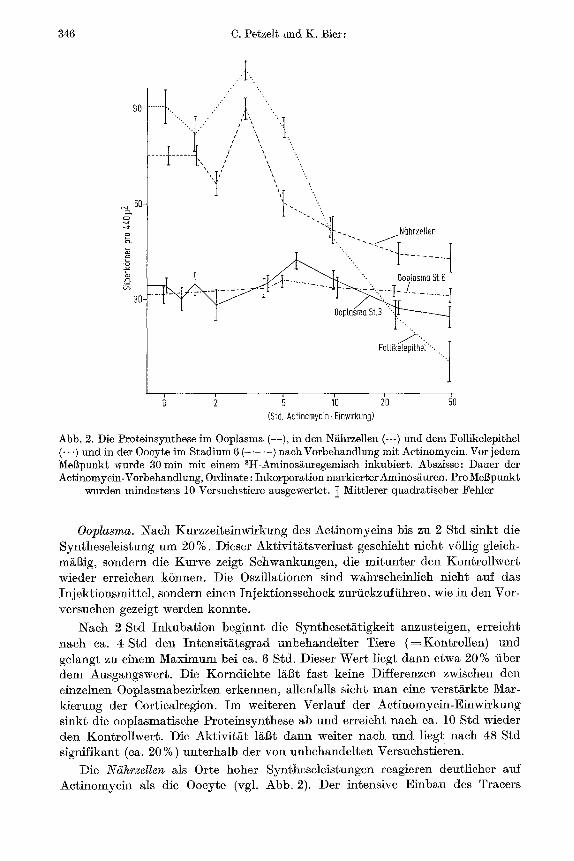
346 C. Petzelt und K. Bier:
�9 /
"" t """ "'"'t i t ~ ..
\,
�9 i
(Std. Actinomycin-Einwirktm 9)
Abb. 2. Die Proteinsynthese ira Ooplasma ( ), in den N~hrzellen (---) und dem Fo]likelepithel (-- -) und in der 0oeyte im Stadium 6 ( - . . . . ) n a c h Vorbehandlung mit Actinomycin. Vor jedem Mel]punkt wurde 30 lnh~ mit einem SH-Aminos~uregemisch inkubiert. Abszisse: Dauer der Actinomycin-Vorbehandlung, Ordinate: Inkorporation markierter Aminos~uren. Pro ]r
wurden mindestens 10 Versuchstiere ausgewertet. ~ Mittlerer quadratischer Fehler
OopIasma. Nach Kurzzeiteinwirkung des Actinomycins bis zu 2 Std sinkt die Syntheseleistung um 20 %. Dieser Aktivit'~tsverlust geschieht nicht vSllig gleich- m&~ig, sondern die Kurve zeigt Schwankungen, die mitunter den Kontrollwert wieder erreichen k6nnen. Die Oszillationen sind wahrscheinlieh nicht auf das Injektionsmittel, sondern einen Iniektionsschock zurfickzuffihren, wie in den Vor- versuchen gezeigt werden konnte.
Nach 2 Std Inkubation beginnt die Syntheset~ttigkeit anzusteigen, erreicht nach ca. 4 Std den Intensit&tsgrad unbehandelter Tiere (=Kontrol len) und gelangt zu einem Maximum bei ca. 6 Std. Dieser Wert ]iegt dann etwa 20% fiber dem Ausgangswert. Die Korndichte li~l~t fast keine Differenzen zwischen den einzelnen Ooplasmabezirken erkennen, alleafalls sieht man eine verstarkte Mar- kierung der Cortica]region. Im weiteren VerIauf der Actinomycin-Einwh~kung sinkt die ooplasmatische Proteinsynthese ab und erreicht nach ca. 10 Std wieder den Kontrollwert. Die Aktivit~tt ]al3t dann weiter n~ch und liegt nach 48 S~d signifikant (ca. 20 %) unterhalb der von unbehandelten Versuchstieren.
Die Niihrzellen als Orte hoher Syntheseleistungen re~gieren deutlicher attf Actinomycin als die Oocyte (vgl. Abb. 2). Der intensive Einbau des Tracers

Hemmung und Induktion yon Proteinsynthesen durch Actinomycin 347
erreicht schon vor dem Maximum der ooplasmatischen Proteinsynthese seinen H6hepunkt. Allerdings bleibt dieser Gipfel im Gegensatz zur Ooeyte nieht lange erhMten, sondern die Syntheset/s sinkt steil ab und erreicht naeh 5 Std einen Punkt 20% unterhMb des Ausgangswertes. Im weiteren Versuchsverlauf nimmt die Aktivit/it nur noch allm~hlich ab, die Kurve verflaeht sich und erreicht naeh 48 Std etwa den Wert der maximalen Einbaurate des Ooplasmas.
Aueh morphologisch maeht sieh die St6rung des Stoffwechsels bemerkbar. Die in den Kontrollen so hervorsteehende Markierung der Nuk]eolen ist v6]lig ver- schwnnden. Es lassen sieh nur noch geringe Markierungsunterschiede zwisehen den Kernen und dem N/~hrzellplasma finden. Die ehemMs kreisrunden Kerne ver]ieren ihre Form, sie werden lappig und zeigen Einbuehtungen. Das Bild hat grol~e Xhnliehkeit mit den Degenerationserseheinungen der N/~hrzellen ira Stadium 4--5 yon unbehandelten Tieren.
Das Follikelepithel ist das Gewebe mit der hSchsten Einbaurate von Amino- s/~uren aller drei Zelltypen des Ovars (Bier, 1963). ttier zeigt sieh die Aetinomyein- wirkung besonders drastiseh. Naeh 48 Std ist die Proteinsynthese nahezu voll- kommen inhibiert. Ebenfalls gehen mit der 1/~ngeren Einwirkung des Antibioti- kums charakteristische Formver/~nderungen einher. Das ursprfingheh hoehprisma- tische Epithel wird kubiseh, die einzelnen Zellen beginnen sieh ab 16 Std auf der yon der Ooeyte abgewandten Seite abzurunden. Im weiteren Verlauf der Actinomyein-Behandlung verflaeht das Epithel immer mehr, bis es nach 48 Std als diehtgepackter Kranz abgerundeter Zel]en die Ooeyte umgibt.
Im Stadium 6 liegt die Proteinsynthese der Ooeyte, gemessen a]s Markierungs- diehte fiber den Z/s bereits bei den Kontrolltieren unterhalb der Aktivit/~t des Stadiums 3. Eine Applikation yon Actinomycin mit der gleiehen Folge der Inkubationszeiten wie im Stadium 3 ffihrt kaum zu einer Ver/inderung der Einbaurate. Es bleibt aber ebenso wie bei den Experimenten mit dem Stadium 3 die Frage often, ob naeh langen Behandlungszeiten mit Aetinomycin noch die gleichen Proteine wie in der unbehandelten Oocyte synthetisiert werden.
B. Elelctrophorese
1. Normalbedingungen
Radioelektrophoretische Untersuchungen geben fiber den Proteinbestand und die aktuel]e Eiweil3synthese qualitativen und quantitativen Aufschlul~. Ffir das Stadium 3 wird die Aussage gegenfiber den autoradiographischen Befunden in- sofern eingeschr~nkt, als Ooplasma, Niihrzellen und Follikelepithel nur als Ganzes errant wurden. Im Stadium 6 werden, da die N/~hr- und Follikelzellen bereits de- generiert sind, nur die Proteine der Oocyte gemessen.
Im Stadium 3 linden sich in Pherogrammen auf Celluloseacetat bis zu aeht Banden nach Anf~rbung mit Amidoschwarz, wobei besonders eine Bande in Startn~he durch starke Bindung des Farbstoffes herausragt (Abb. 3). Die Aus- wertung der Radiopherogramme mit dem Methan-Durchflul3z/~hler nach 20 rain Inkorporation yon laC-Proteinhydrolysat zeigt, dal~ zwar eine gr613ere Anzahl yon Proteinen markiert ist, abet keine Bande sieh dureh besondere Intensit~t aus- zeiehnet. Demnach resultiert die naeh 20 rain doeh schon relativ hohe Einbau- rate (vgl. Abb. 2) nieht aus der bevorzugten Herstellung eines Proteins, sondern

348 0. Petzelt and K. Bier:
, St 180
i L.LI
i -4
IV, I I I / " I
.1ZO~ r -
E
-50
-25
Abb. 3. Pherogramm veto Musca-Ovar, Stadium 3. Darstellung der gef~rbten (...) und der markierten (--) Fraktionen. St Start. Inkorporation des 14C-Aminos~uregemisches 20 rain. Extinktionsmessung der gef~rbten Banden mit Zeiss-Photometer, Schlitzbreite 0,2 ram, Wellen]~nge 620 nm. Auswertung der radio~ktiven Banden mit Methan-DurchfluBz~Mer, Blende 1 ram, Durehlaufgeschwindigkeit 60 ram/h, Maxima]~usschlag bei 180 I/rain, D~mpfung 30 sec
hat in der Synthese eines breiten EiweiBspektrums ihren Ursprung. Allerdings lassen sich einige charakteristische Banden starker Intensi ts yon schw~eher markierten unterscheiden, wobei der Vergleieh zwischen Aktivit/it und Anf~rbung der einzelnen Banden interessant ist. Der besonders stark tingierten Bande Nr. I I entsprieht nur eine vergleiehsweise sehwache Markierung, w&hrend anderer- seits an Stellen, an denen keine gefi~rbten Banden erseheinen, eine deutliehe Akti- vit/~t ~estzustellen ist. Diese Diskrepanz yon Markierung und F/irbung 1/~Bt Sehlfisse auf die tIalbwertszeit der OoeyteneiweiBffaktionen und eine gestaffelte Synthese versehiedener Proteine zu. Die stark ge~s abet nut sehwaeh markierten Banden mfigten demnach aus einem Protein bestehen, dessen Synthese sehr lang- sam erfolgt oder dessen Bildungsort auBerhalb der Eizelle liegt. Der zweiten Hypothese entspreehen die Dotterproteine. Ihre Einlagerung kann sowohl auto- r~diographiseh (Bier, 1963) als aueh radioelektrophoretiseh (Petzelt und Bier, 1970) verfolgt werden. In dieser Arbeit ist die exogene N~tur der Proteine des Extink- tionsgipfels bei Bande Nr. I I be]egt.
Das Stadium 6 zeigt fi~rberiseh ein ~hnliehes Bandenmuster wie das Stadium 3 (Abb. 4). Die Differenzen beziehen sich aut den Bereich Nr. I und I I I , we einige Banden zus/~tzlieh auftauchen und andere fehlen. Die Auswertung mit dem

Hemmtmg und Induktion yon Proteinsynthesen durch Actinomycin 349
l..l.J
-, St
It
:- I m /'.../-
o..~ "
t 180
I
1
E aa
25
Abb. 4. Pherogramm vom Muzca-Ovar, Stadium 6. Darstellung der gef~rbt~n (---) und der markierten (--) Fraktionen. Inkorporation 30 rain. Extinktionsmessung und Auswertung der
Radioaktivit~t wie in Abb. 3
Scanner nach 20 rain Inkubationszeit ergibt Abweiehungen vom Pherogramm der jfingeren Oocyte. Die Gesamtaktivit&t ist wesentlich niediiger, die MarMerung liegt bei den Banden Nr. I und I I I bei anderen Proteinfraktionen, die Uberein- stimmung einiger deutlich aktiven Banden ist aber gegeben. Nach 30 rain ver- &ndert sich das Bild nut dalfingehend, dab alle Proteine eine hSherc Einbaurate zeigen. Es fehlt abet die stark aktive , ,Dotterfraktion" des Stadiums 3.
2. Die Beeinflussung des Proteinspektrums durch Actinomycin im Stadium 3
Nach 2 Std Inkubation mit Actinomycin tr i t t keine signifikante Ver~nderung in Lage, Umfang und fiirberischem Verhalten der Banden des Pherogramms ehl. Auch die Verteilung der Radioaktivitiit hat sich im Vergleich zur Kontrollkurve nicht verschoben, nur ihre Intensit~t hat nachgelassen.
Nach 4 Std Aetinomyein-Behand]ung haben bestimmte Proteine, besonders die Dotterproteine, an Aktivit/~t verloren (Abb. 5). Es treten aber z. T. neue Proteine auf, deren Bfldung dureh Actinomyein demnach nicht gehemmt, sondern sogar gesteigert zu werden seheint. Dieser Effekt zeigt sich besonders bei den Banden Nr. 1, 2 kurz vor der ,,Dotterfraktion". Wie auch die autoradiographischen Ergebnisse zeigen, ist an der Gesamtproteinsynthese, also quantitativ, keine Ver- ~inderung eingetreten, qualitativ ergibt sieh aber neben der Verringerung auch

350 C. Petzelt und K. Bier:
St
1 i 180
2 1
c 60-
- 25 �84
-120
5O -E
25
Abb. 5. Vergleich der radioaktiven Proteinfraktionen des Ovars im Stadium 3 zwischen Kon~rollen (obere Kurve), Inkorporation des Aminosguregemisehes 30 rain, und naeh 4 Std Aetinomyein-Behandlung und ansehlieBender Inkorporation des Aminosguregemisehes yon
30 rain (untere Kurve)
eine vermehrte Bildung VOlt Eiweil3en mit unterschiedlichen Ladungseigenschaften. DM3 dieses Ph/~nomen nieht etwa durch Assoziation einzelner Proteine an Aetino- myein vorget~useht werden kann, haben Koberstein et al. (1968) gezeigt.
Naeh 8 Std lassen sieh deutlichere Unterschiede im Vergleich zur unbehan- delten Ooeyte erkennen. Das urspriingliehe Muster ist zwar immer noch erhalten, doeh treten zus~tzlieh neue ]3anden auf. Das Radiogramm zeigt starke Abwei- chungen von dem der Kontrolltiere. Die anfangs am st&rksten markierte ,,Dotter- f rakt ion" ist v611ig versehwunden. Ebenso ist die Synthese ehliger anderer Proteine fast ganz zum Erliegen gekommen. Start dessen hat sieh die Einbaurate neu aufgetretener Banden erh6ht, die Gesamtakt ivi ts liegt fiber dem Kontrollwert, obwohl die Bildung yon Proteinen, die im unbehandelten Ovar auftreten, durch das Antibiotikum gehemmt wird. Die Bildung der in der unbehandelten Oocyte synthetisierten Proteine ist naeh 16 Std Aetinomycin-Einwirkung zum grSl3ten Teil s tark zurfiekgegangen (Abb. 6). Die Einlagerung markierter Dotterproteine hat aufgehSrt, die Gesamtaktivit/~t liegt unter dem Kontrollwert. Dennoeh ist die Synthese der Proteine, die erst nach der Applikation yon Aetinomycin erscheinen, bei weitem nieht in dem Ma6 gehemmt, wie es bei den ursprfinglieh vorhartdenen Fraktionen der Fall ist.

Hemmung und Induktion yon Proteinsynthesen dutch Actinomycin 351
" 5 t . ,-180
60- r -
E
25-
-120
r
E - 5 0 - -
25
Abb. 6. Vergleich der radioaktiven Pro~einfraktionen des Ovars im Stadium 3 zwischen Kontrollen (obere Kurve), Inkorporation des Aminos~uregemisches 30 rain, und nach 16 Std Actinomycin-Behandlung und anschlieBender Inkorporation des Aminos~uregemisches von
30 min (untere Kurve)
Nach 48 Std Vorbehandlung mit Actinomycin ist die Synthese der meisten Proteine praktisch eingestellt, wenn auch das angef&rbte Pherogramm in etwa dem Ausgangsbild gleicht. Die Auswertung mit dem Scanner bringt die Besti~- tigung, da~ nur noch ganz wenige Proteine gebfldet werden (Abb. 7). Sie ]iegen im Bereich der Banden Nr. l, 2, 3, 4, ihre Einbaurate ist im Vergleich zu einer 16stfindigen Einwirkung yon Actinomycin nur geringffigig gesunken. Sie gehSren zu den Banden, die fiberhaupt erst nach der Applikation des Injektionsmittels auftreten. Ihre Inhibierung scheint im Gegensatz zu den Proteinen, die zum Ovar yon Anfang an gehSren, sehr verzSgert einzusetzen. Es l&13t sieh also durch Aetinomycin eine Hemmung des konstitutiven EiweiBspektrums erreiehen, wobei das Ph&nomen einer adaptiven Eiwefl~bildung ab 4 Std Aetinomycin-Behandlung ffir die vor~bergehende Zunahme der Gesamtinkorporation yon Aminosguren ver- antwortlich ist.
3. Die l~eaktion des Stadiums 6 auf Actinomycin
In der autoradiographischen Analyse zeigt Actinomycin im Stadium 6 nur eine sehr geringe Wirkung, die Silberkornzahl fiber dem Ooplasma bleibt ann&hernd gleieh. Das elektrophoretische Bandenmuster unterscheidet sich nach kurzen Inku- bationszeiten bis zu 4 Std nicht yon dem der Kontrollen. Nach 6 Std sind

352 C. Petzelt und K. Bier:
50- r
E
25-
" ~ 1180
-120.~
...... o , - - . w .
Abb. 7. Vergleich der radioaktiven l~roteinfraktionen des Ovars im Stadium 3 zwischen Kontrollen (obere Kurve), Inkorporation des Aminosauregemisches 30 rain, und nach 48 Std Actinomycin-Behandlung und ansch]ieBender Inkorporation des Aminosauregemisches yon
30 rain (untere Kurve)
jedoch die urspriinglich vorhandenen Frsktionen weniger msrkiert, dagegen treten neue Bsnden suf (1, 2, 3), deren Einbsurate eine betr/~chtliche HShe erreicht (Abb. 8). Nach 8 Std hat die Markierung der neu sufgetretenen Fraktionen ihren HShepunkt iiberschritten und yon Bsnde zu Bande verschieden sbgenommen. Die urspriinghch synthetisierten Proteine werden such jetzt noch weitergebildet, allerdings in wesenthch geringerem AusmsB.
I~sch 48 Std wird die dutch Actinomycin sngeregte Bildung neuer Proteine immer noch sufrechterhalten; die snfangs vorhandenen Fraktionen haben zwar in ihrer Markierung sbgenommen, sind abet noch deutlich nachweisbar (Abb. 9). Der sutoradiogrsphische Befund, dab nsch diesen Actinomycinzeiten die Einbsurate der Aminos~turen im Vergleich zu den Kontrollen gleichgeblieben ist, wird dutch das Auftreten neuer Fraktionen hervorgerufen, welche die teilweise Synthesehemmung bet den snfsngs vorhandenen Proteinen ausgleichen.
AufschluBreich fiir die Bedeutung der nach Actinomycinbehandlung in Erschei- nung tretenden Proteine ist der Vergleich zwischen Oocyten des Stadiums 6, die vor der 30 min-Aminos/~ure-Inkorporstion 48 Std mit Actinomycin behandeIt wnrden, und frisch sbgelegten, befruehteten Eiern, denen w~hrend ihrer Weiter- entwicklung ansehlie6end 30 rain dss 14C-Aminos~uregemisch zur Verfagung stand

Hemmung und Induktion von Prot~insyn~hesen durch Actinomycin
" St
=60
--25
353
.180
.120
6O -E
25
Abb. 8. Vergleich der radioaktiven Proteinfraktionen des Ovars im Stadium 6 zwischen Kon~rollen (obere Kurve), Inkorporation des Aminos~uregemisches 30 rain, und nach 6 Std Actinomycin-Behandlung und ansehlieBender Inkorporation des Aminosi~uregemisches yon
30 rain (untere Kurve)
(Methodik s. Pietruschka). Man sieht aus Abb. 10, dab adaptive Proteine, die erst auf Grund der Aetinomycin-Einwh.kung im Stadium 6 gebildet werden, z. T. im Proteinspektrum unbehandelter fftiher Embryonen auftreten (Banden Nr. 2, 3).
Diskussion
A. Die Actinomycin-unemp/indliche Proteinsynthese Die Bildung yon Proteinen ist an das Vorhandensein entspreehender tuRNS
gebunden. Da dureh Actinomycin schon naeh kurzer Behandlungsdauer fast die gesamte RNS-Synthese bloekiert wird, muB fiir die sowohl im Stadium 3 als auch im Stadium 6 weiterlaufende Eiweil?synthese ein Messenger angenommen werden, der schon vor der Applikation des Antibiotikums gebildet worden ist. Diese RNS-Frakt ion besitzt demnach in beiden Stadien eine so lange Halbwerts- zeit, dab sie auch noch naeh 48 Std in der Lage ist, eine relativ hohe Protein- synthese auffeehtzuerhalten. Eine stabile RNS als Grundlage einer Actinomycin- resistenten Proteinsynthese ist yon vielen Autoren zur Erkls ihrer Ergebnisse herangezogen worden und z .T . aueh naehgewiesen. Crippa et al. (1967) ver- glichen bei Xenopus mit ttflfe yon Hybridisierungsversuchen die RNS friiher und

354 C. Petzelt und K. Bier:
9 5t -180
,_50-
- 25-
120
50--
25
Abb. 9. Vergleich der radioaktiven Proteinfraktionen des Owrs im Stadium 6 zwisehen Xontro]len (obere Kurve), Inkorporation des Aminos~uregemisches 30 rain, und nach 48 Std Actinomycin-Behandlung und anschlieBender Inkorporation des Aminos~uregemisches von
30 rain (untere Kurve)
sps Embryogenesestadien. Sie fanden, dal~ die RNS der ersten Stadien sich signifikant besser mit der oocyts t~NS parallelisieren lggt als die neugebildete RNS glterer Stadien. Da. aber in den friihen Stadien nur eine ganz minimale aktuelle RNS-Synthese stattfindet, muB die RNS aus der Oocyte zur Protein- synthese im Anfang der Embryogenese herangezogen werden, sie ist also stabi- lisiert.
Bei Musca reagiert auch schon im Stadium 3, ~lso zu einem Zeitpunkt, in dem die RNS-Versorgung der Oocyte aus den N~hrzellen ihre volle Leistung erreicht hat, die Proteinsynthese nut verhs wenig auf Actinomycin. Die Eiwei~bildung in diesem Stadium setzt sich also einerseits aus der Protein- synthese zusammen, die von kurzlebiger ml%NS codiert wird und durch Actino- mycin gehemmt werden kann, zum anderen aus der Eiweil~synthese, die auf la.ng- lebiger tuRNS beruht und Actinomycin-resistent ist. I m Stadium 3 ist die Reaktion auf das injizierte Mittel mit ttilfe der Autoradtographi e und auf Grand mor- phologischer Ver~nderungen bei den Nghrzellen und dem Follikelepithel festzu- stellen. Die bei beiden gefundene Steigerung der Einbaurate nach mittleren Inku- bationszeiten macht auch bier eine induzierte Proteinsynthese wahrscheinlich. Die N~hrzellen als abortive Keimzellen reagieren s auf das Antibiotikum wie die Oocyte, ihre Proteinsynthese lgBt sich nur in begrenztem Umfang inhibieren. Dagegen erlischt bei den Follikelzellen - - Somazellen mesodermalen Ursprungs - -

ttemmung und Induktion yon Proteinsynthesen durch Actinomych~
- St
355
-180
50. r
~2 C 3 u
E
25
3 2
-120
r.--
2
e ~
E
6O
25
Abb. 10. Vergleich der radioaktiven Proteinfraktionen des Ovars im Stadium 6 nach 48 Std Actinomycin-Behandlung und anschliel3ender Aminos~ure-Inkorporation yon 30 mhl (obere Kurve), und yon unbehandelten, frisch abgelegten Eiern, die 30 rain mit dem Aminos~ure-
gemisch inkubiert wurden (untere Kurve)
die Proteinsynthese fast v511ig; die relativ hobe RNS-Synthese dieses EpJthels bei unbehandelten Tieren ist also anscheinend mit einer sofortigen Translation der produzierten kurzlebigen Matrizen verbunden, die Eiweil~bildung ist daher durch Aetinomyein zu blockieren.
I m Stadium 3 ist in den Trophoeyten eine sehr hohe RNS-Synthese. Nur ein geringer Teil davon wird aber als kurzlebige I~NS gebildet, da v o n d e r Protein- synthese n~tmlieh nur ca. 20 % durch Actinomycin zu inhibieren sind, obwohl die l%NS-Synthese se]bst ja vSllig bloekiert wird. Das wfirde bedeuten, dal~ die I~NS- Synthese der N&hrzellen im Stadium 3 zu einem grol~en Teil dazn dient, die Proteinsynthese sp&terer 0ogenesestadien bzw. in der friihen Embryogenese zu gew&hrleisten, wenn naeh Degeneration der N&hrzellen und der bis zur frfihen Em- bryogenese anhaltenden Inaktivierung des Oocytengenoms und seiner Deszenden- ten das System ohne l~NS-Synthese ist, We~terhin ist daraus ersichtlich, dab die langlebige talONS, die zur Actinomycin-unempfindliehen Proteinsynthese im Stadium 3 herangezogen wh'd, sehon sehr frfih gebildet worden ist.
Die Proteinsynthese des Stadiums 6 ist auch ohne Actinomycin-Einwirkung nicht mit einer gleichzeitigen BNS-Synthese korreliert. Der dennoch stattfin- dende Einbau markierter Aminos~uren mu8 also in jedem Fall an Matrlzen vor

356 C. Petzelt und K. Bier:
sich gehen, die in frfiheren Stadien gebfldet und entweder bis zu ihrer Trans- lation stabilisiert wurden oder eine entspreehend lange IIalbwertszeit aufweisen. Die Erscheinung der yon de novo gebildeten RNS unabh/~ngigen Proteinsyn- these ist auch bei Linsenzellen (Marks et al., 1963; Spector und Kinoshita, 1965) und Reticuloeyten (Scott und Bell, 1965) gefunden worden.
Diesen Zellen ist die Eigensehaft gemeinsam, sehr spezifische P r o t e i n e - Cristallin, H/~moglobin - - zu synthetisieren. Die Synthese geht anseheinend eben- falls an stabflisierten Matrizen vor sieh. Die Stabilisierung ist naeh diesen Unter- suchungen erst ein Ph/inomen der Endstadien der Differenzierung. Als ,,Differen- zierung" wird der ProzeB einer vorfibergehenden oder andauernden Spezialisierung einer Zelle bezeichnet, in der ein Teil des Genoms ffir bestimmte Produkte aktiv ist, die nieht ffir den unmittelbaren Stoffweehsel der Zelle erforderhch sind (Brown und Dawid, 1968). Eine so differenzierte Zelle ist im allgemeinen dutch folgende MerkmMe gekennzeiehnet: Mangel an Kernaktivit/it, Abnahme der Polysomen, stabile tuRNS, Weiterlaufen einer Proteinsynthese ohne RNS-Neusynthese. Eben diese Kennzeichen weist auch die Eizelle nach dem Abschlul3 der Oogenese auf. Man k6rmte also sagen, dab das E i - eigentlich die undifferenzierte Zelle par excellence - - yon der Oogenese her betrachtet, eine gewisse Spezialisierung mit dem Abschlug der Eireifung erfahren hat.
B. Die durch Actinomycin induzierte Proteinsynthese Verschiedene Autoren fanden nach einer Actinomycin-Applikation eine Stimu-
lierung der Proteinsynthese (Lit. vgl. S. 342). Bei diesen Experimenten wurden allerdings selten qualitative Untersehiede gemaeht. Es warde nieht differenziert zwisehen einer Syntheseaktivit/it, die zu einer Verst~rkung bereits vorhandener Proteinmuster ffihrt, und einem zus/~tzlichen Auftreten neuer Fraktionen trotz Hemmung der konstitutiven Proteinbi]dung.
Die autoradiographischen Untersuchungen ergeben im Fliegenovar naeh mitt- leren Inkubationszeiten eine eindeutige Steigerung der Einbaurate in allen unter- suehten Zelltypen des Ein/~hrverbandes. Verfolgt man an Hand der Pherogramme das VerhMten der ursprfinglieh vorhandenen Fraktionen, sieht man ihre im Lauf der Acthlomyein-Einwh'kung immer mehr zunehmende Inhibierung. DaB ihre Bildung nicht" sofort blockiert ist, liegt wohl an den Ha]bwertszeiten der sie codierenden I~NS. Da aber aueh Fraktionen existieren, deren Synthese nicht oder nur wenig dureh das Antibiotikum beeinflugt wird, andererseits aber die Neu- bfldung yon RNS naeh der Applikation yon Aetinomyein praktisch sofort gestoppt ist, lassen sich aus dem Verhalten der einzelnen konstitutiven Proteine Schliisse auf den Metabolismus der fiir sie spezifischen Matrizen ziehen. Neue Proteine, die auf Grund der Aetinomycin-Einwirkung auftreteu, sind ffir eine Erh6hung der Einbaurate verantwortlich. Das autoradiographisch ermittelte Aktivit~tsmaxi- mum ist also der Zeitpunkt, an dem die absinkende, aber noch relativ kr/~ftige Synthese der konstitutiven Proteine mit der anlaufenden Bildung adaptiver Ei- weiBe sieh zu ihrem hSchsten Wert addiert.
Wie 1/~gt sieh nun 4iese Neubildung bestimmter Proteine erkl~ren ? Eine ihr vorausgehende, induzierte prim~re RNS-Synthese kommt nieht in Frage, da die Transkription an der DNS blockiert ist und im unbehandelten Stadium 6 keine aktivierten Genome zur Verfiigung stehen, denn die Chromosomen sind zu einer inaktiven Karyosph/~re kondensiert. Die verwandten Matrizen mfissen bereits vor

Hemmung und Induktion yon Proteinsynthesen durch Actinomycin 357
der Actinomycin-Applikation gebildet sein. Eine Vergnderung der elektrophoreti- schen Eigenschaften durch Bildung eines Protein-Actinomycin-Komplexes, der durch eine Bandenversehiebung neue Proteine im Pherogramm vortguscht, ist nach den Untersuchungen yon Koberstein et al. (1968) ausgeschlossen.
Nach allgemeiner Ansicht wird die Inaktivierung der fiir spi~tere Funktionen best immten RNS durch die Anlagerung spezifischer Proteine an diese Nuklein- s~uremolekfile erreicht. I m unbefruchteten Seeigelei sind nach Maggio et al. (1965) die Ribosomen, nach Monroy (1968) ein Tell der Polysomen in einem dureh Proteinanlagerung reprimierten Zustand. Dadurch wird eine vorzeitige Verwen- dung bereits vorhandener Matrizen verhindert. Wenn man dicse Ergebnisse auf das Verhalten der Musca-Oocyten unter Actinomycin-Einflug iibertrggt, und dabei am klassischen Wirknngsmodus des Actinomycins festzuhalten versucht, liege sieh die Syntheseaktivierung bestimmter Proteine dureh die Blockierung der Produk- tion solcher Proteine erklgren, die diese Polysomen im Normalfa]l inaktivieren. )As Zusatzhypothese mug jedoch eine relativ geringe Halbwertszeit der an der Stabilisation beteiligten Proteine angenommen werden.
I m Stadium 6 zeigt die oocyti~re Proteinsynthese eine unerwartete Reaktion; die hier zu findende adaptive Eiweigsynthese ist auf Grund der natfirlieherweise fehlenden RNS-Bildung schwer verstgndlich. Nach unseren jetzigen Kenntnissen greift Aetinomycin in physiologischer Dosis nur an der DNS an (Reich und Goldberg, 1964). Da in diesem Stadium die RNS-Versorgung dutch die Niihr- zel]en erloschen und der Oocytenkern v6llig inert ist, die Eizelle abet t rotzdem aui das Antibiotikum reagiert, mug es zus~tz]ich noch einen anderen Wirk~ngs- modus ffir Actinomycin geben. Die nur wenig beeinflugte Bildung ursprfinglich vorhandener Proteine erscheint unter der Annahme einer Existenz stabiler Matri- zen plausibel. Die Indukt ion der Synthese neuer Proteine liege sich allenfalls durch eine Sprengung des Protein-RNS-Komplexes inaktivierter RNS durch Aeti- nomycin interpretieren. Es mug auch often b]eiben, ob die gefundene partielle Aktivierung im Stadium 3 auf einer direkten Wirkung des Actinomycins beruht. Pastan und Friedman (1968) geben eine Zusammenstellung yon Wirkungen des Actinomycins, die augerhalb der Hemmung der DNS-abhiingigen RNS-Synthese liegen.
Literatur
Baker, W. : A clonal system of differential gene activity in Drosophila. Develop. Biol. 16, 1-- 17 (1967).
Bier, K. : Autoradiographische Untersuchungen fiber die Leistungen des Fo]likelepithels und der Nahrzellen bei der Dotterbfldung und Eiweigsynthese im Fliegenovar. Wilhelm Roux'Arch. Entwickl.-Mech. 0rg. 154, 552--575 (1963).
- - Kunz, W., l~ibbert, D. : Struktur und Funktion der Oocytenchromosomen und Nukleolen sowie der Extra-DNS wahrend der Oogenese panoistischer und meroistischer Insekten. Chromosoma (Berl.) 2~, 214--254 (1967).
Bose, S. : Differential sensitivity of protein synthesis in human cell lines to actinomycin D. Exp. Cell Res. 46, 599--603 (1967).
Braehet, J.: Biochemical changes during fertilization and early embryonic development. In: Cell differentiation (Reuck and Knight, ed.). London: Churchill Ltd. 1967.
- - Denis, H., Vitry, F. de: The effects of actinomycin D and puromycin on morphogenesis in amphibian eggs and Acetabularia mediterranea. Develop. Biol. 9, 398--412 (1964).
Brown, D., Dawid, I. : Specific gene amplification in oocytes. Science 160, 272--274 (1968). Chandley, A. C. : Studies on oogenesis in Drosophila melanogaster with 3-H-thymidine label.
Exp. Cell Res. 44, 201--215 (1966).

358 Petzelt und Bier: Hemmung und Induktion yon Proteinsynthesen dutch Actinomycin
Crippa, 3s Davidson, E. I-I., 3/[irksy, A. E. : Persistence in early amphibian embryos of infor- mational RNA's from the lampbrush chromosome stage of oogenesis. Prec. nat. Aead. Sei. (Wash.) 57, 885--897 (1967).
Eliasson, E. : Repression of arginase synthesis in Chang liver cells. Exp. Cell Res. 48, 1--17 (1967).
Engels, W., Bier, K. : Zur Glykogenspeicherung w~hrend der Oogenese und ihrer vorzeitigen Aus]Ssung durch Blockierung der RNS-Vcrsorgung (Untersuchungen an Musca domestica L.). Wilhelm Roux' Arch. Entwickl.-Mech. Org. 158, 64--88 (1967).
Garren, L. D., Howell, R. R., Tomkins, G. M.: A paradoxical effect of aetinomycin D: The mechanism of regulation of enzyme synthesis by hydrocortisone. Prec. nat. Acad. Sci. (Wash.) 52, 1121--1129 (1964).
Gross, P. R., Cousineau, G. H. : Macromolecule synthesis and the influence of actinomyein on early development. Exp. Cell Res. 311, 368--395 (1964).
Kersten, W., Kersten, H. : Bindung yon Actinomyein C an Nucleins~uren und Nucleotide. Hoppe-Seylers Z. physiol. Chem. 11110, 21--30 (1962).
Koberstein, R., Weber, B., Jaenicke, R. : Wechselwirkungen yon Proteinen mit Aetinomycin C. Z. Naturforsch. 2]b, 474-483 (1968).
Maggie, R., 3s A., Rinaldi, A. M., Vittorelli, H. : Activation in vitro des ribosomes des oeufs vierges d'oursin. C. R. Aead. Sci. (Paris) 260, 1293---1295 (1965).
Marks, P. A., Rifkind, R.A., Darien, D. : The existence of long-lived RNA templates in embryonic chick. Prec. nat. Acad. Sei. (Wash.) 50, 336~342 (1963).
Maruzov, V. I., OreMovich, N. N. : Unusual effect of aetinomyein on collagen synthesis in perichondral tissue of chicken embryo. Prec. nat. Acad. Sci. (USSR) 166, 742--745 (1966).
Monroy, A. : The ribosomes of the unfertilized sea urchin eggs. Acad. Naz. Lincci 104, 127-- 132 (1968).
Moog, F. : Intestinal phosphatase activity: Acceleration of increase by puromycin and actino- mycin. Science 144, 414~-416 (1964).
Papaconstantinou, J. : A stabilization of RNA templates in lens cell differentiation. Prec. nat. Acad. Sci. (Wash.) 58, 95--101 (1967).
Pastan, I., Friedman, R. : Actinomycin D: Inhibition of phospholipid synthesis in chick embryo cells. Science 160, 316--317 (1968).
Petzelt, Ch., Bier, K. : Synthese der H~molymphproteine and die Aufnahme der Dotterfl'aktion in die Ooeyte unter Aetinomyein-EinfluB. Wilhelm Roux' Arehiv 164, 359--366 (1970).
Pietruschka, F. : In Vorbereitung. Reich, E., Goldberg, I. I-I. : Aetinomyein and nucleic acid function. In: Progress in nucleic
acid research and molecular biology, vol. 3, p~ 183--234. (J. N. Davidson and W. E. Cohn, ed.). New York-London: Academic Press 1964.
Rosen, F., Raina, P.N., Milholland, l~. J., Niehol, C. A. : Induction of several adaptive enzymes by aetinomycin. Science 146, 661--663 (1964).
Scott, R. B., Bell, E. : Protein synthesis during development: Control through mRNA. Science 145, 711--714 (I964).
Specter, A., Kinoshita, J. I-I. : Long-lived RNA template in calf lenses. Bioehim. biophys. Aeta (Amst.) 95, 561--570 (1965).
Teller, W. H.: The route of entry and locMisation of blood proteins in the ooeytes of saturnfid moths. J. biophys, bioohem. Cytol. 9, 747--761 (1961).
Trepte, It.: Uber den EinfluB dcr larvalen and imaginalen Ernghrung auf die Eient- wicklung bei der Stubcnfliege Musca domesticct. Staatsexamensarbeit Zool. Inst. Miinster (1967) (unverSff.).
Willebrand, G. : Vergleiehende and experimentelle Untersuehungen des Aminosgure-Invcntars und Zuekerbestandes yon Insekten- and Wirbeltierovarien mittels Diinnsehicht-Chromato- graphie. Staatscxamensarbeit Zool. Inst. Mtinster (1965) (unverSff.).
Dr. Christian Petzelt Max-Planck-Institut f. Zellbiologie Institut Tiibingen 74 Ttibingen lVielanchthonstr. 36





























