Embed Size (px)
DESCRIPTION
hem
Citation preview

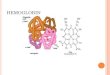
• Hemoglobina este o heteroproteină (proteină conjugată)
• Cromoproteină – HEMUL care reprezintă partea neproteică este colorată
• CARACTERISTICI:
- partea proteică – globina ≈ 95-96% din moleculă
- partea neproteică – hemul ≈ 4%
- fierul ≈ 0,34% fier porfirinic
HEMUL – structură tetrapirolică:
- 4 nuclee pirolice I, II, III, și IV, coplanare, legate prin punți metin notate α, β, ɣ, δ
- substituenții: - CH3 în pozițiile 1,3,5,8
- CH=CH2 (vinil) în pozițiile 2,4
- CH2-CH2-COOH (propionil) în pozițiile 6,7
- Fierul în stare de valență +2
HEXACOORDINAT: 4 legături de azotul pirolic:
-1 legătură directă cu His din globină
- 1 legătură cu His din globină prin intermediul H2O sau O2
STRUCTURA HEMOGLOBINEI

LEGĂTURA HEMULUI CU CATENELE POLIPEPTIDICE ALE GLOBINEI
-Se leagă de catenele de tip α ale globinei - Se realizează prin intermediul histidinelor din poziția 58 și 87 - FIERUL – hexacoordinat - este legat diferit:
CH = CH2
N
CH3
N
CH3
CH = CH2
N
CH3
CH3
N Fe2+
COOH
COOH
IV HEM
I
II
III
1 2
3
4
56
7
8
CH2
CH2
CH2
CH2
Globinã
His (87 sau 93)
His (58 sau 63)Globinã
O2 (H2O)
1. DIRECT cu o histidină (legătură proximală) 1. Indirect cu altă histidină (legătură distală) Cu ajutorul unei legături de hidrogen, prin intremediul unei molecule de apă

GLOBINA – Caracteristici structurale
• Proteină bazică- subclasa HISTONE – predomină aminoacizi cu caracter bazic (histidina, arginina)
• Globina funcțională are o structură cuaternară: 4 lanțuri polipeptidice (TETRAMER)
• 574 aminoacizi : 141 aminoacizi în 2 lanțuri de tip α și 146 de aminoacizi în 2 lanțuri de tip β (2 monomeri α și 2 monomeri β)
• La nivelul tetramerilor, legaturile între monomerii α și β sunt mai puternice decât cele între monomerii de același tip
• Structura spațială a catenelor polipeptidice permite înglobarea în interiorul moleculei a 4 structuri heminice; fiecare catenă formează un “buzunar” hidrofob în care este inclusă o moleculă de hem, prin intermediul fierului divalent
• Fiecare protomer are structură primară, secundară, terțiară (globulară)
• Lanțurile polipeptidice sunt legate între ele prin legături labile (punți de hidrogen, forțe Van der Vaals)

GLOBINA – Caracteristici structurale
• Există mai multe tipuri de protomeri:
• Hemoglobina normală: 2 monomeri identici - 2 câte 2: α1α2β1β2
• 2 legături ale fierului se realizează direct cu anumite histidine din structura globinei
2 tipuri de legături
Legăturile dintre protomeri
Legăturile dintre protomeri și HEM


FUNCȚIILE HEMOGLOBINEI
1. Se combină reversibil cu O2
Hb + O2 HbO2 + O2 Hb(O2)2 Hb(O2)3 Hb(O2)4
+O2 +O2
Fe NN
N
N
H2O
GlobinãHis
Fe NN
N
N
O2
HisGlobinã
GlobinãHis
+ O2 + H2O
HisGlobinã
DEZOXIHEMOGLOBINĂ OXIHEMOGLOBINĂ
Proteină alosterică!

FACTORI : 1) Presiunea parțială a O2
2) pH –ul 3) Prezența 2,3-difosfogliceratului
1) La o pO2 mai mare de 100mm Hg, în alveolele pulmonare, echilibrul este deplasat spre dreapta; la pO2 mai mică de 35 mm Hg în capilare, echilibrul deplasat spre stânga
2) La pH acid (în capilare) Hb are afinitate mai mică pentru oxigen pe care î-l cedează usor; la pH alcalin afinitatea Hb pentru oxigen este mai mare
3) cantităţi crescute de acid 2,3 difosfogliceric deplasează echilibrul spre stânga
Factorii care influențează capacitatea hemoglobinei de a lega oxigenul

Forma R În capilarele pulmonare adiacente
alveolelor
Forma T la nivelul țesuturilor

2. Se combină reversibil cu CO2 Prin intermediul grupării –NH2 libere a globinei => carbhemoglobină
FUNCȚIILE HEMOGLOBINEI continuare
Hb – NH2 + CO2 Hb – NH – COO- + H+
3. Se combină ireversibil cu CO => carboxihemoglobină
Hb + CO HbCO afinitate > 300 ori
4. Oxidarea Fe2+ - e- Fe3+ methemoglobină (ireversibilă)
5. Sulfhemoglobina

• Globinele se deosebesc între ele prin
• Importanță practică în studiile de biologie evolutivă
• În cursul evoluției ontogenetice există mai multe variante de hemoglobine normale:
- Hb embrionară – HbE α2ε2
- Hb fetală – HbF α2δ2 (sintetizată în ficat între lunile 3-8 de sarcină)
- Hb juvenilă
- Hb adultului HbA1 α2β2 (este forma majoritara 97-99%); mai există o formă minoră HbA2 (α2 δ 2 1-2%) – începe să fie sintetizată din viața intrauterină. Înlocuiește treptat HbF
• Globinele au speciospecificitate (cristalizează diferit în funcție de specie):
-oxihemoglobina de cal – cristale monoclinice
- oxihemoglobina de cobai – cristale tetraedrice
- oxihemoglobina de câine – cristale ortorombice
- oxihemoglobina umana – bipiramide ortorombice simetrice - Importanță în laboratoarele de criminalistică
Conținutul Proporția Aranjamentul
aminoacizilor

Biosinteza hemoglobinei

BIOSINTEZA HEMOGLOBINEI
Etapele: I. Biosinteza HEMULUI
II. Biosinteza GLOBINEI
III. Cuplarea hemului cu globina
David SHEMIN, David RITTENBERG – 1945
I. Biosinteza HEMULUI. Faze:
1. Sinteza acidului delta – aminolevulinic
2. Biosinteza PORFOBILINOGENULUI
3. Biosinteza PROTOPORFIRINEI – IX
4. Biosinteza HEMULUI
Precursori: - Succinil ~ SCoA
- Glicocol activat cu PLP
Autoexperiment cu 15N-glicină
•Hemoglobinogeneză •Eritropoieză

N
CH2
CH
OH
CH3
O PO3H
2
N CH2
COOHCOOH
CH2
CH2O
~SCoA
COOH
CH2
CH2
O
NH2
COOH
CO2
COOH
CH2
CH2
O
CH2
NH2
+
C
Glicocol activat cu PLP Succinil~SCoA
H2O PLP(se reface)
- ALAS
( - aminolevulinatsintetaza)
C
Acid - amino, - cetoadipic
HC
- ALAS C
Acid - aminolevulinic
( - ALA)
1) Sinteza acidului delta aminolevulinic (δ – ALA)
HSCoA

1. Biosinteza acidului delta aminolevulinic
• Reprezintă prima etapă din biosinteza hemului
• Presupune reacţia între succinil CoA şi glicocol sub acţiunea δ aminolevulinatsintazei
• Este obligatorie prezenţa PLP care activează glicocolul
• Aceasta etapă se desfasoară intramitocondrial, iar δ ALA trece în citoplasmă
• Enzima δ ALA sintaza este enzima determinantă de viteză a întregului proces; hemul acţionează ca inhibitor al activităţii enzimei
• Plumbul inhibă δ ALA sintaza

COOH
CH2
CH2
O
H2C
NH2
COOH
CH2
CH2
O
CH2
NH2
CH2
COOH
N
CH2
CH2
NH2
CH2
COOH
CH2
COOH
CH2
NH2
CH2
N
CH2
COOH
CH2
C
C+
2 H2O
C C
C
PORFOBILINOGEN(PBG)
izomeraza
( - aminolevulinatdehidratataza
H
Acid - aminolevulinic
( - ALA)
2) Biosinteza PORFOBILINOGENULUI
porfobilinogen sintetaza

• Se realizează prin condensarea a 2 molecule de δ ALA sub acţiunea enzimei δ aminolevulinat dehidrataza (porfobilinogen sintetaza)
• Această etapă se desfăşoară în citoplasmă
• Enzima conţine zinc şi este foarte sensibilă la acţiunea inhibitorie a metalelor grele, mai ales a plumbului
2. Biosinteza porfobilinogenului (PBG)

3) Biosinteza PROTOPORFIRINEI – IX
COOH
CH2
NH
CH2
NH2
CH2
CH2
COOH
NH
AP
NH
A
P
NH
A P
NH
NH
AP
NH
A
P
NH
A P
NH
- CH2 - COOH
- CH2 - CH
2 - COOH
PORFOBILINOGEN(4 PBG)
porfobilinogendezaminaza
A
P
H2C
H2C CH2
CH2
I
II
III
IV
A
P
UROPORFIRINOGEN izomer - I
H2C
H2C CH2
CH2
IV
UROPORFIRINOGEN izomer - III
izomeraza
A
P
A
P
4 NH3
- acetil
- propionil
4 molecule

CH3
NH
P
NH
CH3
P
NH
CH3
P
NH
CH3- CH
2 - COOH
- CH2 - CH
2 - COOH
CH3
NH
P
NH
CH3
CH = CH2
NH
CH3
CH = CH2
NH
CH3
CH3
N
P
NH
CH3
CH = CH2
N
CH3
CH = CH2
NH
CH3
O2
H2C
H2C CH2
CH2
IV
COPROPORFIRINOGEN izomer - III
P
A
P
UROPORFIRINOGEN - III
uroporfirinogendecarboxilaza
4 CO2
I
II
III
coproporfirinogen oxidaza
H2C
H2C CH2
CH2
IV
PROTOPORFIRINOGEN izomer - IX
P
I
II
III
HC
HC CH
CH
IV
PROTOPORFIRINÃizomer- IX
P
I
II
III
protoporfirinogen oxidaza
6 H
ferochelataza
2 H2O
2 CO2

3. Biosinteza protoporfirinei IX
Transformarea porfobilinogenului (PBG) în protoporfirina IX are loc în citoplasmă
PBG este transformat iniţial în uroporfirinogen, sub acţiunea a 2 enzime:
– Porfobilinogen deaminaza care determină formarea unor punţi metilenice între inele pirolice
– Uroporfirinogen III sintaza care are proprietatea de a închide ciclul tetrapirolic
Uroporfirinogenul este transformat în coproporfirinogen sub acţiunea uroporfirinogen decarboxilazei

Fe2+
CH = CH2
N
CH3
N
CH3
CH = CH2
N
CH3
CH3
N Fe2+
COOH
COOH
PROTOPORFIRINÃizomer- IX
ferochelataza
4.) Biosinteza HEMULUIIII. Cuplarea hemului cu globina
IV HEM
I
II
III
1 2
3
4
56
7
8
CH2
CH2
CH2
CH2
Globinã
His (87 sau 93)
His (58 sau 63)Globinã
Globinã
O2 (H2O)
2H

4. Biosinteza hemului
• Chelatarea ferului de către protoporfirina IX se desfășoară
intramitocondrial
• Enzima ferochelataza este sensibilă la acţiunea inhibitoare a metalelor grele (mai ales a plumbului)
În paralel, la nivelul ribozomilor are lor sinteza globinei , care se va lega de hem prin intermediul resturilor de Hys din poziţiile 58 si 87, pentru a forma hemoglobina

REGLAREA BIOSINTEZEI HEMULUI
8 GLICINÃ + 8 SUCCINIL~SCoA
- aminolevulinatsintetaza
8 ACID - AMINOLEVULINIC
porfobilinogensintetaza
4 - PORFOBILINOGEN
uroporfirinogenI sintetaza
UROPORFIRINOGEN III
uroporfirinogendecarboxilaza
COPROPORFIRINOGEN III
coproporfirinogenoxidaza
PROTOPORFIRINOGEN IX
protoporfirinogenoxidaza
PROTOPORFIRINÃ IX
ferochelataza
HEMAPOREPRESOR
8 HS-CoA + 8 CO2
8 H2O
4 NH3
Izomerizarea ciclului IV
4 CO2
2 CO2 + 4 H
6 H
Fe2+
Fe3+
feedback (-)
UROPORFIRINOGEN Iuroporfirinogen
III sintetaza

BIOSINTEZA HEMOGLOBINEI - COMENTARII -
=> PRECURSORI = COMPUȘI FOARTE SIMPLI
- Glicocol
- Succinil ~SCoA
- Piridoxal fosfat (PLP)
=> Legătura cu CICLUL KREBS prin Succinil ~SCoA
(ciclul KREBS – SHEMIN)
=> Prima enzimă = δ – aminolevulinat sintetaza – este enzimă limitantă
de viteză → etapă reglatoare a procesului
=> ERITROPOIETINA = hormon ce modulează procesul de biosinteză
=> Proces foarte complex ce se petrece în reticulocite
=> Are loc la nivelul tuturor organitelor subcelulare:
- formarea protoporfirinei IX → MITOCONDRIE
- încorporarea Fe2+ → CITOPLASMĂ
- biosinteza globinei → RIBOZOMI
- cuplarea globinei cu hem → CITOPLASMĂ

Catabolismul hemoglobinei

Eritrocitele au o durată medie de viaţă de 120 de zile; celulele îmbătrânite
sunt recunoscute datorită modificarilor ce au loc la nivel membranar şi
sunt fagocitate de către ţesutul reticuloenditelial extravascular (splina,
ficat, maduva osoasă). Are loc denaturarea globinei şi eliberarea
hemului în citoplasmă. Globina este degradată cu eliberarea
aminoacizilor ce vor intra în fondul metabolic al organismului.
Hemul este degradat prin intervenţia sistemului enzimatic
microzomal; enzima hem-oxigenaza care desface ciclul tetrapirolic este
dependentă de fier şi de NADPH; această enzimă desface una din
punţile α metinice care leagă două inele pirolice, iar C este convertit în
CO, aceasta fiind singura sursă endogenă de CO. Această oxidare se
realizează în prezenţa oxigenului molecular.
Catabolismul hemoglobinei

În urma acţiunii hemoxigenazei se formează biliverdina care se va
transforma sub acţiunea biliverdinreductazei în bilirubină.
Bilirubina nu rezultă numai din degradarea eritrocitelor
îmbătrânite dar şi din catabolismul celorlalte proteine heminice.
Bilirubina este puţin solubilă în mediu apos la pH-ul
fiziologic; ea este transportată în plasmă legată de albumină
(70mg/dL de plasma). În această formă bilirubina este denumită
bilirubină totală/ neconjugată/ indirectă, şi nu dă reacţia de
diazotare. La concentraţii mai mari de 25mg/dL se manifestă
toxicitatea bilirubinei care va determina apariţia icterului nuclear.
Catabolismul hemoglobinei

V
N
M
N
MP
V
N
M
M
N
P
Fe2+
O2
NADPH2
V
N
M
N
MP
V
N
M
M
N
P
OH
Fe3+
O2 CO
Fe3+
V
NH
M
NH
MP
O
V
ONH
M
M
P
N
HEM
Globinã
Globinã
hem oxigenazã
H2O
Globinã
Globinã
I
II
- HIDROXIHEMINÃ
VERDOGLOBINÃ
Globinã
Cit P450
I
II
BILIVERDINÃ
NADP+
III
IV
CATABOLISMUL HEMOGLOBINEI

V
NH
M
NH
MP
O
V
ONH
M
M
P
N
NADPH2
NH
M V
O NH
CH
2
M P
NH
P M
NH
M V
O
I
II
BILIVERDINÃ
III
IV
biliverdin reductazã
NADP+
IIIIIIIV
BILIRUBINÃ

NH
M V
O NH
CH
2
M
NH
P M
NH
M V
O
COOH
COOH
NH
M V
O NH
CH
2
M
NH
M
NH
M V
O
COOH
IIIIIIIV
BILIRUBINÃ
O
OOC
(CH2)2
O
O CO
(CH2)2
IIIIIIIV
O
O CO
(CH2)2
1 1
Bilirubinã - monoglucuronid Bilirubinã - diglucuronid
UDP-GLUCURONILtransferaza
UDP-GLUCURONAT 2 UDP-GLUCURONAT
La FICAT:
Bilirubina glucuronoconjugată (bilirubina directă)

Bilirubina este transportată la ficat unde este separată de albumine, şi
suferă o reacţie de conjugare cu acizii glucuronici; la această reacţie
bilirubina participă prin grupările carboxilice de la resturile propionice
ce substituie inele pirolice. Conjugarea are loc în prezenţa enzimei
bilirubin: UDP glucuronil transferaza.
Bilirubina glucuronoconjugată (bilirubina directă) este
hidrosolubilă, ceea ce permite excreţia sa la nivel biliar. Bilirubina
glucuronoconjugată nu este absorbită in intestinul subţire; la nivelul
intestinului gros, sub acţiunea hidrolazelor bacteriene, se realizează
hidroliza esterilor cu acizii glucuronici şi eliberarea resturilor propionice.
Bilirubina eliberată astfel este redusă la un compus tetrapirolic liniar,
numit urobilinogen (punţile metinice devin metilenice iar inelele
tetrapirolice au duble legături paralele).

Urobilinogenul poate urma mai multe căi:
- o parte este oxidat la urobilină, sau redus la stercobilinogen,
compuşi care se elimină prin fecale
- o parte este reabsorbit la nivel intestinal, revine la nivel
hepatic şi este din nou excretat prin bilă (circuitul
enterohepatic)
-o parte se elimină ca atare prin urină

CICLUL ENTEROHEPATIC
AL PIGMENȚILOR
SANGVINI
HEMOGLOBINA
VERDOGLOBINA
BILIVERDINA
BILIRUBINA
BILIRUBINA LEGATÃ DE ALBUMINE
INDIRECTÃ
PRIMARÃ (NECONJUGATÃ)
BILIRUBINA DIRECTÃ
CONJUGATÃ (DIGLUCURONID)
BILÃ(colecist)
INTESTIN
Reduceri
BILIRUBINÃ
MEZOBILIRUBINÃ
UROBILINOGEN
STERCOBILINOGEN
Reducere
Oxidare
Oxidare
UROBILINÃ
STERCOBILINÃ
FECALE
PLASMÃ
RINICHI
URINÃurobilinogenstercobilinogen
urobilinogenstercobilinogen
FICAT
MÃDUVÃ OSOASÃ
SPLINÃ
CELULE RETICULO-ENDOTELIALE
EXTRAHEPATICE
ruperea puntii
- metinice
Fe2+
globinã
Reduceri
PLASMÃ
{
FICAT
{
{