Embed Size (px)
Citation preview

Aus der Klinik für Kleintiere
der Veterinärmedizinischen Fakultät der Universität Leipzig und der Abteilung für Neuropathologie
der Charité der Freien Universität Berlin
Histochemische und elektronenmikroskopische Untersuchungen der paravertebralen Muskulatur bei
Hunden mit degenerativen Erkrankungen der Wirbelsäule
Inaugural-Dissertation
zur Erlangung des Grades eines Doctor medicinae veterinariae (Dr. med. vet.)
durch die Veterinärmedizinische Fakultät der Universität Leipzig
eingereicht von Romy Herrschelmann
aus Wolgast
Leipzig 2006

Mit Genehmigung der Veterinärmedizinischen Fakultät der Universität Leipzig Dekan: Prof. Dr. Karsten Fehlhaber Betreuer: Prof. Dr. Vera Grevel Prof. Dr. Gisela Stoltenburg-Didinger Gutachter: Prof. Dr. Vera Grevel, Klinik für Kleintiere, Universität Leipzig
Prof. Dr. Gisela Stoltenburg-Didinger, Institut für Neuropathologie, Charité, Freie Universität Berlin Prof. Dr. Heinz-Adolf Schoon, Institut für Veterinär-Pathologie, Universität Leipzig Prof. Dr. Thomas Bilzer, Institut für Neuropathologie, Heinrich-Heine-Universität Düsseldorf
Tag der Verteidigung: 19.01.2006

Für meine Familie

Inhaltsverzeichnis 1 Einleitung 1 2 Literaturübersicht 2 2.1 Die Skelettmuskulatur des adulten Hundes 2 2.1.1 Muskelfasertypen in der Skelettmuskulatur des Hundes 3 2.1.2 Verteilung der Fasertypen in der Skelettmuskulatur des Hundes 5 2.1.3 Unterschiede der Muskelfasertypen hinsichtlich Rasse, Körpermasse,
Alter, Geschlecht und Training 6 2.1.4 Messung des Faserkalibers 8 2.1.5 Bestimmung des Variationskoeffizienten der Muskelfasergrößen 9 2.2 Gewebssyndrome des Skelettmuskels 9 2.2.1 Das myopathische Gewebssyndrom 11 2.2.2 Das entzündliche Gewebssyndrom 11 2.2.3 Das neurogene Gewebssyndrom 12 2.2.4 Metabolische Myopathien 13 2.2.5 Strukturmyopathien in der Skelettmuskulatur 14 2.2.6 Unterscheidungskriterien der Gewebssyndrome 15 2.3 Einfluss neurologischer Störungen auf die Muskulatur 16 2.4 Morphologie und Funktion der Mitochondrien 18 2.4.1 Aufbau der Mitochondrien 18 2.4.2 Struktur der mitochondralen DNS 19 2.4.3 Vermehrung der Mitochondrien 20

2.4.4 Funktion der Mitochondrien 20 2.4.5 Mitochondriopathien 21 2.4.5.1 Mitochondrale Myopathie 21 2.4.5.2 Hypoxidosen 22 2.5 Der Aufbau der Wirbelsäule 23 2.6 Aufgabe und Funktion der Rückenmuskulatur 25 2.7 Rückenmark und begleitende Strukturen 26 2.8 Rückenmarkskompressionen 27 2.8.1 Arten von Rückenmarkskompressionen 27 2.8.1.1 Zervikale Diskopathie 30 2.8.1.2 Thorakolumbale Diskopathie 30 2.8.1.3 Atlantoaxiale Instabilität 31 2.8.1.4 Lumbosakrale Stenose 31 2.8.1.5 Kaudale Zervikale Spondylomyelopathie 32 2.8.2 Diagnose von Rückenmarkskompressionen 33 2.8.3 Operations-Methoden 34 3 Tiere, Material und Methoden 37 3.1 Tiere 37 3.1.1 Klinische Daten der Patienten 37 3.1.2 Art der Rückenmarkskompression 39 3.1.3 Lokalisation der Rückenmarkskompression 40 3.1.4 Auswahl und Entnahme der Muskelbiopsien 44

3.2 Methoden 45 3.2.1 Gefriertechnik der Proben für die Lichtmikroskopie 45 3.2.2 Färbemethoden der Proben für die Lichtmikroskopie 46 3.2.3 Elektronenmikroskopie 51 3.2.4 Morphometrie 53 4 Ergebnisse 55 4.1 Morphologische Veränderungen 55 4.1.1 Neurogenes Gewebssyndrom 57 4.1.2 Myopathisches Gewebssyndrom 64 4.1.3 Lipofuszin in der Skelettmuskulatur 75 4.2 Morphometrische Veränderungen 76 5 Diskussion 78 6 Zusammenfassung 87 7 Summary 89 8 Literaturverzeichnis 91

Abkürzungen BSH Berner Sennenhund Cx Wirbel der Halswirbelsäule (x = Nummer des Wirbels) GM Gliedmaßen HGM Hintergliedmaßen HWS Halswirbelsäule kDa Kilo Dalton KHT Kurzhaarteckel LHT Langhaarteckel Lx Wirbel der Lendenwirbelsäule (x = Nummer des Wirbels) RHT Rauhaarteckel Sx Wirbel der Schwanzwirbelsäule (x = Nummer des Wirbels) Thx Wirbel der Brustwirbelsäule (x = Nummer des Wirbels) VGM Vordergliedmaße

1 Einleitung
Beim Menschen führen Diskopathien zu schmerzbedingten Schon- und Fehlhaltungen der
Wirbelsäule, die in besonderem Maße die Rückenmuskulatur belasten. Diese Überbean-
spruchung der Muskeln kann zu morphologischen und pathophysiologischen Veränderungen
führen, die in unterschiedlichem Maße histologisch, immunhistologisch und/oder elektronen-
mikroskopisch nachweisbar sind. Bei einigen dieser Patienten ließen die gefundenen Verän-
derungen eine primäre Muskelerkrankung vermuten. Eine solche, in ihrer Funktion insuffi-
ziente Muskulatur, könnte eine Wirbelinstabilität nach sich ziehen und Ursache für eine nach-
folgende Diskopathie sein.
Da der Bandscheibenvorfall beim Hund eine häufige Erkrankung darstellt, sollte der Frage
nachgegangen werden, inwiefern auch der Hund bei einer solchen Erkrankung morphologi-
sche, immunhistologische und/oder elektronenmikroskopische Muskelfaserveränderungen der
Rückenmuskulatur aufweist. Lassen sich Veränderungen diagnostizieren, die als Überbean-
spruchung der Muskulatur interpretiert werden können? Sind muskuläre Reaktionen zu fin-
den, die eine Funktionseinschränkung zur Folge haben? Ergibt sich daraus eine Instabilität,
die eine Bandscheibenerkrankung nach sich zieht?
Um diese Fragestellungen beantworten zu können, wurden während des operativen Eingriffes
bei Hunden mit Diskusprolaps, Diskushernien oder einem Menigiom Rückenmuskelbiopsien
im Seitenvergleich sowie ober- und unterhalb der Kompression des Rückenmarks entnommen
und untersucht. Dazu wurden die Biopsien geteilt und in flüssigem Stickstoff bzw. Glutar-
aldehyd gelagert. Später erfolgten eine lichtmikroskopische und eine elektronenmikros-
kopische Untersuchung. Verschiedene Parameter wurden qualitativ und quantitativ erfasst.
Bei ausgewählten Hunden mit Kompressionen im Bereich der thorakalen und/oder lumbalen
Wirbelsäule wurden die Fasergrößen morphometrisch bestimmt.
Zum jetzigen Zeitpunkt liegen nur wenige systematische Untersuchungen beim Hund vor, die
sich mit morphologischen Veränderungen der Skelettmuskulatur befassen. Meist gehen sie
nicht über eine Betrachtung der Fasergrößen hinaus oder beziehen sich auf bestimmte, meist
selten auftretende, Muskelerkrankungen des Hundes. Feingewebliche und morphometrische
Untersuchungen der Rückenmuskulatur fehlen völlig. Somit bildet die Arbeit zudem eine
Basis für Untersuchungen der Rückenmuskulatur, des wichtigsten Halteapparates der Wirbel-
säule des Hundes.

2 Literaturübersicht
2.1 Skelettmuskulatur des adulten Hundes
Die Muskelfasern des normalen Skelettmuskels weisen histologisch im Querschnitt eine
typische polygonale Konfiguration mit geringen Faserkalibervariationen auf und zeigen
hauptsächlich subsarkolemmal (4-8 pro Faserquerschnitt) gelegene, selten binnenständige
Muskelkerne. Binnenständige Kerne kommen physiologisch v.a. in der Nähe von Muskel-
faszien vor und dürfen dann nicht als myopathische Gewebsveränderung fehlinterpretiert
werden.
Die Muskelfaser setzt sich aus mehreren hundert Myofibrillen zusammen, die u.a. die
kontraktilen Proteine Aktin und Myosin enthalten. Diese Aktin- und Myosinfilamente sind
regelmäßig ineinander verzahnt und bilden auf diese Weise das charakteristische Muster der
quergestreiften Muskulatur.
Der Muskelaufbau stellt sich im Elektronenmikroskop deutlich dar (Abb. 1). Dabei sind die
Filamente so angeordnet, dass die dickeren Myosinfilamente nebeneinander liegend ins-
gesamt die A-Bande der Myofibrille ergeben. In der I-Bande liegen die dünneren Aktin-
filamente. Diese ragen zwischen die Myosinfilamente, erreichen jedoch nicht die gegenüber-
liegenden Aktinfilamente, so dass innerhalb der A-Bande ein helleres Areal, die H-Zone
(Hensens-Zone), bleibt. In dieser H-Zone befinden sich die verdickten Anfangsteile der
Myosinfilamente, diese bilden die sogenannte Mittelscheibe (M-Linie). Die Aktinfilamente
sind an der Zwischenscheibe (Z-Linie), die in der Mitte der I-Bande lokalisiert ist, befestigt
(LEONHARDT 1990).
Der Bereich zwischen zwei angrenzenden I-Banden wird Sarkomer genannt. Viele dieser
wiederkehrenden Sarkomere bilden eine Myofibrille. Die Myofibrillen sind untereinander
durch den intermyofibrillären Raum getrennt (DUBOVITZ und BROOK 1973).
Der Aufbau ist in der folgenden Abbildung 1 schematisch dargestellt.

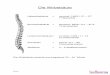
Abb. 1: Muskelaufbau (nach SAJONSKI und SMOLLICH 1990)
Z=Zwischenscheibe, I=I-Bande, A=Bande, Af=Aktinfilamente, Mf=Myosin-
filamente, H=Hensens-Zone, M=Mittelscheibe
Gruppen von 50-100 Muskelzellen werden durch lichtmikroskopisch nicht immer sichtbare
Bindegewebssepten (Endomysium) zu Fasern zusammengefasst. Die im Endomysium liegen-
den Nervenendigungen und Kapillaren sorgen für die Innervation und Energieversorgung der
Muskelzellen. Durch das Perimysium sind die Fasern zu mit bloßem Auge erkennbaren Bün-
deln zusammengefasst. In diesen liegen zu- und abführende Gefäße und Nervenäste. Auch
diese Faserbündel sind wieder von Bindegewebe, dem Epimysium (Faszie) umschlossen,
welches mit den „untergeordneten“ Bindegewebssepten die Verbindung zu Sehnen, Periost
und Aponeurosen herstellt (DUNN et al. 1984, JERUSALEM 1991).
2.1.1 Muskelfasertypen in der Skelettmuskulatur des Hundes
Alle bisher untersuchten Säugetiere (mit Ausnahme des Hundes), weisen in ihrer Skelett-
muskulatur die Fasertypen-I, -IIA, -IIB und -IIC auf. Beim Hund fehlen Typ-IIB-Fasern
(BRAUND et al. 1978). Die Differenzierung der verschiedenen Muskelfasertypen erfolgt mit
Hilfe histochemischer Färbungen (ATPase Reaktionen), womit das charakteristische Schach-
brettmuster dargestellt werden kann (BRAUND et al. 1978).
Der Fasertyp wird durch das die Faser innervierende Motoneuron bestimmt. Da die Muskel-
fasern einer motorischen Einheit nicht als Gruppe zusammenliegen, sondern mit Muskelfa-

sern anderer motorischer Einheiten vermischt sind, kommt es zu dem für die Skelett-
muskulatur typischen Bild des Schachbrettmusters.
Die Muskelfasern-Typ-I zeichnen sich durch einen hohen Myoglobingehalt und einen Mito-
chondrienreichtum aus. Damit sind sie zur Energiegewinnung durch Oxidation in der Lage.
Sie haben eine relativ langsame Kontraktionsgeschwindigkeit, sind aber zur Dauerleistung
befähigt. Deshalb kommen Typ-I-Muskelfasern hauptsächlich in der Stütz- und Haltemus-
kulatur vor.
Die Muskelfasern-Typ-II sind gekennzeichnet durch einen hohen Myofibrillengehalt, weisen
aber eine deutlich geringere Anzahl an Mitochondrien und Myoglobin auf. Sie besitzen eine
hohe glykolytische und eine nur geringe oxidative Aktivität. Die Kontraktionsgeschwindig-
keit dieser Muskelfasern ist sehr schnell, sie sind also zu kurzfristigen Höchstleistungen
befähigt (SAJONSKI und SMOLLICH 1990).
Die Typ-II-Fasern unterteilen sich beim Hund lediglich in die Subtypen-IIA und -IIC
(BRAUND et al. 1978).
CARDINET et al. (1982) unterscheiden bei Hunden zusätzlich zwischen Typ-IA- und Typ-
IB-Fasern. Sie ordnen dabei die Typ-IB-Fasern als Übergangsform von Typ-IIC- in Typ-IA-
Fasern ein.
Die Typ-IIC-Fasern sind intermediäre, unreife Muskelfasern, welche die Vorstufe der
Subtypen-IIA und -IIB (BROOKE et al. 1971) bzw. Typ-IA-Fasern (CARDINET et al. 1982)
darstellen und in weit geringerer Anzahl, weniger als 10% (BRAUND et al. 1978),
vorkommen.
Zusätzlich konnten von SNOW et al. (1982) in der Muskulatur des Hundes durch Differen-
zierung von Peptiden der schweren Myosinkette, eine weitere Klasse von Typ-II-Fasern
gefunden werden, die sie aufgrund ihrer enzymatischen und immunhistochemischen
Eigenschaften als „nichtklassische Typ-IIB-Fasern“ bezeichnen. LATORRE et al. (1993)
sprechen diese Typ-II-ähnlichen Fasern als „ Typ-II Dog-Fasern“ an.
Bei den meisten Carnivoren und Primaten kann man außerdem Typ-IIM-Fasern nachweisen.
Diese finden sich allerdings nur in Muskeln, die vom dorsalen, mesodermalen Primordium
abstammen (M. temporalis, M. masseter, M. pterygoideus medialis und lateralis, M. tensor
tympani, M. tensor veli palatini) (BUBB und SIMS 1986). Erstmals wurden diese Fasern von

ROWLERSON et al. (1981) beschrieben, die sie in den Kieferschließmuskeln der Katze nach-
wiesen.
ORVIS und CARDINET (1981) ordneten sie aufgrund ihrer histochemischen Eigenschaften
zunächst den Typ-IIC-Fasern zu. MASCARELLO et al. (1982) konnten durch immunhisto-
chemische Untersuchungen zeigen, dass diese in der Kaumuskulatur des Hundes vorkommen-
den Fasern, den Typ-IIM-Fasern der Katze entsprechen.
2.1.2 Verteilung der Fasertypen in der Skelettmuskulatur des Hundes
Die Muskelfasern der Skelettmuskulatur setzen sich beim Hundewelpen zum Zeitpunkt der
Geburt zu 90-95 % aus Typ-IIC-Fasern und 5-10 % Typ-I-Fasern zusammen. Dabei stellen
übergroße Typ-I-Fasern, die einen Anteil von 2-4 % besitzen, eine Besonderheit dar. Diese
Fasern sind größer als alle anderen Fasertypen, weisen binnenständige Kerne auf und schei-
nen mit den von WOHLFART (1937) beschriebenen B-Fasern identisch zu sein. Im Alter von
4-5 Wochen sind diese Fasern nicht mehr nachweisbar (BRAUND und LINCOLN 1981).
Ab der 2. Lebenswoche nimmt der Anteil normalgroßer Typ-I und Typ-II-Fasern zu, und im
Alter von etwa zwölf Wochen entspricht die prozentuale Fasertypenverteilung derjenigen
adulter Tiere (BRAUND und LINCOLN 1981, CARDINET et al. 1982).
Die Verteilung der Fasertypen variiert stark zwischen den verschiedenen Muskeln eines
Hundes, und auch bei homologen Muskeln unterschiedlicher Individuen wurden deutliche
Differenzen gefunden. Es konnten keine signifikanten Abweichungen der Fasertypen-
verteilung zwischen rechter und linker Gliedmaße festgestellt werden (ROSENBLATT et al.
1988, ARMSTRONG et al. 1982, NEWSHOLME et al. 1988).
Der Anteil des Fasertyps-I in der Skelettmuskulatur des Hundes reicht von 14% beim M.
extensor carpi radialis bis zu 100 % im M. anconaeus und M. quadriceps femoris. Reine Typ-
II-Muskeln gibt es nicht (ARMSTRONG et al. 1982).
Entsprechend den Verhältnissen beim Mensch weisen die tiefer liegenden Extensoren von
Hunden im Bereich von Oberarm und Oberschenkel die höheren Anteile an Fasertyp-I (96 %
Caput mediale des M. triceps brachii, 88 % M. vastus intermedius) auf. An Unterarm und
Unterschenkel ist das Verhältnis umgekehrt.

Die eher oberflächlich liegenden, antigravitationswirksamen Muskeln haben höhere Typ-I-
Anteile. So lassen sich beispielsweise im M. flexor digitalis superficialis 89 % und im M.
flexor digitalis profundus nur 30 % Typ-I-Fasern nachweisen (ARMSTRONG et al. 1982).
Zu ähnlichen Ergebnissen sind auch MAXWELL et al. (1977), BRAUND und LINCOLN
(1981), RODRIQUEZ-BARBUDO et al. (1984), NEWSHOLME et al. (1988) und AGUERA
et al. (1990) gekommen.
Innerhalb eines Muskels kann abhängig vom Ort der Biopsieentnahme die Fasertypen-
verteilung variieren (ROSENBLATT et al. 1988). ROSENBLATT et al. (1988) stellten am
Beispiel des M. semitendinosus des Hundes, ähnlich dem Menschen, einen signifikant höhe-
ren Fasertyp-I-Anteil in den tiefer gelegenen Muskelbereichen fest.
NEWSHOLME et al. (1988) konnten ein derartiges Verhältnis für den M. tibialis cranialis
nachweisen.
Auch ARMSTRONG et al. (1982) fanden in der Muskulatur von Ober- und Unterarm sowie
Ober- und Unterschenkel die höchsten Typ-I-Anteile in Knochennähe, mit Ausnahme des
Caput laterale des M. triceps brachii. Den ausgeprägtesten Unterschied konnten sie am M.
vastus lateralis mit 15 % Typ-I-Fasern im oberflächlichen Bereich und 85 % Typ-II-Fasern in
der Tiefe aufzeigen.
Der M. masseter besteht zu 85 % aus Typ-IIM-Fasern (BUBB und SIMS 1986).
2.1.3 Unterschiede der Muskelfasertypen hinsichtlich Rasse, Körpermasse, Alter,
Geschlecht und Training
ROSENBLATT et al. (1988) stellten keinen signifikanten, rassespezifischen Unterschied zwi-
schen den Fasertypenanteilen des M. semitendinosus bei Mischlingen, Beagle und Jagd-
hunden fest.
Gegensätzliche Ergebnisse weisen die Untersuchungen von GUNN (1975, 1978b, 1979a, b)
sowie GUY und SNOW (1981) auf. Sie fanden einen deutlich höheren zahlen- und flächen-
mäßigen Anteil von Typ-II-Fasern (z.T. bis 100 %) am Querschnitt des M. semitendinosus
beim Greyhound im Vergleich zu anderen Rassen. Dieses Ergebnis ist trainingsunabhängig.

RODRIQUEZ-BARBUDO et al. (1984) untersuchten den Querschnitt des M. tibialis cranialis
von Greyhounds, Deutschen Schäferhunden und Foxterriern und fanden ebenfalls erhöhte
Fasertyp-II-Anteile beim Greyhound. Der geringste Anteil Typ-II-Fasern tritt beim Deutschen
Schäferhund auf. Der Foxterrier nimmt eine Mittelstellung ein.
Auch AGUERA et al. (1990) stellten bei Vertretern lauffreudiger Hunderassen (Galgo
Espanol, Podenco Iberico) im M. tibialis cranialis und M. flexor digitalis profundus einen sig-
nifikant höheren Anteil an Typ-II-Fasern fest als bei nicht so lauffreudigen Hunderassen
(Deutscher Schäferhund, Mastin Espanol). Der Anteil der Typ-II-Fasern im M. pectineus, der
eine eher statische als dynamische Funktion hat, ist bei den vier untersuchten Rassen etwa
gleich groß.
ROSENBLATT et al. (1988) und AGUERA et al. (1990) geben für größere und schwerere
Rassen größere Faserdurchmesser an. Insbesondere bei Muskeln, die nicht der Fortbewegung
dienen, korreliert die Fasergröße positiv mit der Größe und Masse der Hunde (AGUERA et
al. 1990).
JULIAN und CARDINET (1961) fanden am M. biceps brachii ebenfalls größere Faserquer-
schnitte bei schwereren Hunden. Diese waren jeweils von einer stärkeren Variation der Faser-
größen bei Hunden mit höherem Gewicht begleitet.
Die Untersuchungen von MAXWELL et al. (1977) ergaben eine ebensolche Korrelation zwi-
schen Körpermasse und Faserfläche.
Auch bei dem Verhältnis der Muskelmasse zur Körpermasse bestehen bei adulten Tieren
deutliche Unterschiede (GUNN 1978a).
Bei Greyhounds läßt sich ein Muskelanteil von durchschnittlich 57,1 % nachweisen. Bei
anderen Hunderassen beträgt er lediglich 43,5 %. Welpen zeigen diese Muskelmassedifferen-
zen nicht (GUNN 1978c).
Dahingegen haben GÄRTNER et al. (1987) bei allen von ihnen untersuchten Säugerspezies
einen etwa gleichen Anteil der Muskelmasse an der Gesamtkörpermasse von etwa 40 % ge-
funden.
CASTLE und REYMAN (1984) fanden im M. fibularis longus von ein- bis zweijährigen
Hunden mittlere Faserdurchmesser, die etwa 5 % größer waren als die Durchschnittswerte der
älteren untersuchten Hunde. Bei über 7 Jahre alten Hunden war der mittlere Faserdurchmesser
5-10 % kleiner als der Durchschnitt aller untersuchten Tiere.

Unterschiede zwischen den Geschlechtern in Bezug auf die Muskulatur scheinen nicht zu
bestehen (GUNN 1975, 1978a, b, c 1979a).
2.1.4 Messung des Faserkalibers
Erste Fasermessungen an Hundemuskeln nahmen SCHWALBE und MAYEDA (1890) und
MORPUGO (1897) vor.
Arbeiten aus jüngerer Zeit zur Faserkalibermessung liegen von FALUDI et al. (1964),
BRAUND et al. (1980a,b,c, 1982, 1985a,b, 1987), BRAUND und LINCOLN (1981),
ARMSTRONG (1982), CASTLE und REYMAN (1984), RODRIQUEZ-BARBUDO et al.
(1984), KORNEGAY et al. (1988), ROSENBLATT et al. (1988) und AGUERA et al. (1990)
vor.
Allerdings wurden bei diesen Untersuchungen unterschiedliche Hunderassen und Methoden
verwendet, so dass die Werte nur eingeschränkt vergleichbar sind.
Seit Beginn der 80er Jahre werden vor allem enzymhistochemisch gefärbte Gefrierschnitte
von Muskelbiopsien zur Messung genutzt.
Aufgrund des polygonalen Erscheinungsbildes der quergeschnittenen Muskelfaser wird der
kleinste mittlere Durchmesser ermittelt, um exakte Ergebnisse zu erzielen (DUBOWITZ und
BROOKE 1973, JERUSALEM und ZIERZ 1991). Diese sogenannte Zwei-Punkt-Methode
hat sich in der Praxis bewährt, da die Muskelfasern in den meisten Fällen leicht schräg und
nicht exakt quer geschnitten sind.
Unter Anwendung der Messung des kleinsten mittleren Faserdurchmessers hat HEINZE
(1994) an Muskelbiopsien von verschiedenen Hunderassen Fasergrößen ermittelt. Dabei han-
delte es sich aber um ein post mortem Untersuchungsgut. Außerdem wurden die Proben in
Formalin fixiert und in Paraffin eingebettet, so dass ein Vergleich nur bedingt möglich ist.
In Paraffinschnitten kommt es, im Gegensatz zum Gefrierschnitt, durch Schrumpfung zu
deutlich reduzierten Faserdurchmessern (BANKER und ENGEL 1994). Dagegen besteht die
Möglichkeit bei den Gefrierschnitten, dass sich die Fasern durch den Gefrierprozess etwas
ausdehnen (MOORE et al. 1971).

2.1.5 Bestimmung des Variationskoeffizienten der Muskelfasergrößen
Der Variationskoeffizient (VK) dient in der Myopathologie als Maß für die Faser-
größenstreuung.
Zur Berechnung des Variationskoeffizienten wird der Mittelwert (Median) der Faser-
durchmesser des jeweiligen Muskelfasertyps und seine Standardabweichung herangezogen.
Diese Standardabweichung beträgt normalerweise weniger als ein Viertel des Durchmessers
der Muskelfaser und liegt gewöhnlich unter 10. Dieser Wert wird mit 1000 multipliziert und
durch den Mittelwert (Median) des Muskelfaserdurchmessers dividiert (JERUSALEM und
ZIERZ 1991).
VK= Standartabweichung x 1000 / Mittelwert
Variationskoeffizienten von normalen Skelettmuskeln des Hundes sind nur bei BRAUND et
al. (1982) für den M. biceps femoris, M. gastrocnemius, M. triceps brachii, M. flexor digitalis
superficialis und bei KORNEGAY et al. (1988) für den M. biceps femoris, M. vastus lateralis
und M. triceps brachii angegeben. Beide Autoren nutzten Gefrierschnitte mit der Darstellung
der myofibrillären ATPase für ihre Untersuchungen.
2.2 Gewebssyndrome des Skelettmuskels
Der Skelettmuskel zeigt unter pathologischen Bedingungen nur eine geringe Anzahl morpho-
logischer Reaktionsweisen. Ein und dasselbe morphologisch veränderte Erscheinungsbild
kann unterschiedliche Ursachen haben. Häufiger sind es mehrere Veränderungen, die ein
spezielles morphologisches Syndrom erkennen lassen und seltener krankheitsspezifische Ein-
zelbefunde, die für die Diagnostik entscheidende Hinweise geben. Nach SEITZ (1965) gibt es
das Gewebssyndrom der neurogenen Atrophie, der Dystrophie und Polymyositis. Etwas allge-
meiner, damit aber weitere Krankheitsbilder einbeziehend, spricht JERUSALEM (1991) vom
myopathischen, vom neurogenen und vom myositischen Gewebssyndrom.
Richtungsweisend für die Interpretation einer Muskelläsion können Veränderungen der
schachbrettartigen Verteilung der Muskelfasertypen sein.
Eine Zusammenfassung der möglichen Reaktionen der Skelettmuskulatur ist nach
SCHRÖDER et al. (1989) in der nachfolgenden Tabelle dargestellt.

Tab. 1: Allgemeine Reaktionen der Skelettmuskulatur (nach SCHRÖDER 1989)
1. Kaliberänderungen
a) Atrophie (DD: Hypoplasie)
b) Hypertrophie (DD: Pseudohypertrophie)
2. Charakteristische Strukturveränderungen
a) Ringbinden und sarkoplasmatische Massen
b) Central-Core- und Target-Fasern
c) Nemalinkörper und Z-Band-Streaming
d) Kernvermehrung, zentrale Kernreihen
e) Mitochondriale Veränderungen
f) Glykogen- und Fettspeicherung
g) Proliferationen des sarkoplasmatischen Retikulums
h) Schwellungen des T-Systems;
(DD: Mitochondrienschwellungen, Artefakte)
i) Sarkolemminvaginationen
j) Aufsplitterungen
3. Formen der Muskelfasernekrose
a) Hyaline Degeneration
b) Fokale Degeneration, Lipophanerose
c) Myophagie
4. Muskelfaserregeneration
a) Satellitenzellproliferation
b) Differenzierung zu Myoblasten, Myozyten, Myotuben, Muskelfasern
5. Entzündliche Erkrankungen des Muskels
a) Autoimmunprozesse
b) Infektion
6. Fibrose, Fettvakatwucherung
7. Tumoren
8. Muskeldefekte
9. Veränderungen der motorischen Endplatte
10. Veränderungen der Muskelspindeln

2.2.1 Das myopathische Gewebssyndrom
Dieses Erkrankungsbild ist hauptsächlich durch Faserkalibervariationen, binnenständige Ker-
ne, Degeneration einzelner disseminierter Muskelfasern oder kleiner Gruppen von Muskelfa-
sern und den Ersatz untergegangener Muskelfasern durch Bindegewebe bzw. bei stärkerer
Ausprägung durch Fettgewebe gekennzeichnet.
In verschiedenen Fällen finden sich kleine entzündliche Infiltrate, die eine Abgrenzung zur
Myositis erschweren können (JERUSALEM und ZIERZ 1991).
Auch das Auftreten von regenerierenden Muskelfasern ist möglich. Spaltbildungen von Fa-
sern finden sich zum einen bei chronischen Myopathien, zum anderen aber auch in hyper-
trophen Fasern bei chronisch neurogenen Prozessen.
Manchmal reichen die morphologischen Befundkriterien nicht aus, um ein chronisch myo-
pathisches von einem chronisch neurogenen Schädigungsmuster zu unterscheiden, so dass
eine neurophysiologische Untersuchung notwendig wird.
2.2.2 Das entzündliche Gewebssyndrom
Wichtigstes Kriterium des entzündlichen Gewebssyndroms ist das entzündliche Infiltrat. Die-
ses kann an verschiedenen Lokalisationen schwerpunktmäßig auftreten. Danach wäre ein vor-
wiegend interstitielles Vorkommen, von einem Vorkommen im Parenchym zu unterscheiden.
Infiltrate im Interstitium führen zu keiner wesentlichen Parenchymalteration, wogegen paren-
chymatöse Infiltrationen schwere Muskelfaserveränderungen hervorrufen können.
Auch eine Bildung von Granulomen (granulomatöse Myositis) ist möglich (JERUSALEM
und ZIERZ 1991).
Mitunter werden kleinherdige Infiltrate auch bei nichtentzündlichen myopathischen oder
neurogenen Muskelschäden gefunden, andererseits sind bei Myositiden häufig nicht in allen
untersuchten Ebenen des Bioptates entzündliche Infiltrationen nachweisbar.

2.2.3 Das neurogene Gewebssyndrom
Eine frische Denervation (innerhalb weniger Wochen) führt zunächst zu einer disseminiert
auftretenden Einzelfaseratrophie, wodurch die Fasern atrophisch werden sowie eckig und
elongiert erscheinen. Im Anschluss daran kommt es zur Reinnervation durch benachbarte
Motoneurone und damit zum Phänomen der „Fasertypengruppierung“. Dabei prägen die re-
innervierten Fasern entsprechend des innervierenden Motoneurons ihre Fasertypen aus
(kollaterale Reinnervation).
Bei schweren chronischen Verläufen, wenn die kollaterale Reinnervation den Prozess nicht
mehr bewältigen kann, kommt es zur „felderförmigen Atrophie“. Dabei treten stark atrophier-
te Muskelfasergruppen neben Fasergruppen mit zum Teil normalen Muskelfaserdurchmessern
auf. Auch Fasern mit einer kompensatorischen Hypertrophie sind zu finden (JERUSALEM
und ZIERZ 1991).
Gelegentlich kann man bei neurogenen Myopathien sogenannte „rimmed vacuoles“ finden,
die jedoch nicht spezifisch für eine bestimmte Erkrankung sind. Darunter versteht man Vaku-
olen, die von einem granulären Material begrenzt werden, welches sich in der HE-Färbung
basophil und in der Gomori-Trichom-Färbung leuchtend rot (fuchsinophil) darstellt. Sie be-
sitzen eine Größe von 2-25 µm (CARPENTER et al. 1978) und liegen häufiger subsarko-
lemmal als zentral. Da der granuläre Vakuoleninhalt während der Aufarbeitung von Kryostat-
schnitten häufig an den Vakuolenrand gedrängt wird und ihn gewissermaßen „austapeziert“,
entsteht das typische lichtmikroskopische Bild der umrandeten Vakuole.
Häufig finden sich unter den Typ-I-Fasern sogenannte Target-Fasern, die erstmals von SHY
und MAGEE (1956) beschrieben wurden. Dabei handelt es sich um zentrale Ausfälle des
intermyofibrillären Netzwerkes. Diese weisen auf eine neurogen bedingte Störung mit einer
stattfindenden Reinnervation hin, sind aber nicht pathognomonisch.
Binnenständige Kerne sind in der Regel nicht oder kaum zu finden, in seltenen Fällen ist das
Bindegewebe vermehrt, gelegentlich kommt es zu Fettvakatwucherung.
Bei besonders schweren Verläufen kann man allerdings diese sonst eher für myopathische
Prozesse typischen Veränderungen finden, man spricht dann von einer Begleitmyopathie.
Selbst kleine Rundzellinfiltrate können ausnahmsweise vorkommen, und so kann die Ab-
grenzung zum entzündlichen wie auch zum myopathischen Gewebssyndrom schwierig sein.

2.2.4 Metabolische Myopathien
Unter diesem Begriff werden strukturelle und funktionelle Störungen der Skelettmuskulatur
zusammengefasst, die durch Stoffwechselveränderungen hervorgerufen werden.
Derartige Muskelveränderungen sind im Falle von Mitochondriopathien durch das Vorkom-
men vergrößerter, vermehrter und irregulär angeordneter Mitochondrien gekennzeichnet.
Obwohl viele Stoffwechselprozesse in den Mitochondrien ablaufen, wird eine Störung der
Adenosintriphosphat-Produktion während der oxidativen Phosphorilierung als Ursache dieses
Krankheitsbildes angesehen. Es kommt zu morphologischen Umbauten der Mitochondrien
und zu verminderter Funktionalität (MORGAN-HUGES 1994, GHADIALLY 1997).
Lichtmikroskopisch und histochemisch sind Vergröberungen des intermyofibrillären Netz-
werks sowie eine Steigerung der oxidativen Enzymtätigkeit festzustellen. Die erhöhte Enzym-
aktivität kompensiert diese Funktionsstörungen aber nicht, so dass es intrazellulär zum ge-
häuften Auftreten von Stoffwechselprodukten wie z. B. Glykogen und Lipid kommt.
Weiterhin lichtmikroskopisch darstellbar sind „ragged red fibres“ (zerrissene rote Fasern). Bei
„ragged red fibres“ handelt es sich um Muskelfasern, die einen subsarkolemmalen Saum von
Mitochondrien aufweisen oder gänzlich von Mitochondrien ausgefüllt sind. Diese lassen sich
gut mit der Gomori-Trichom-Färbung darstellen, in der sie durch intensive fuchsinophile An-
färbung imponieren. Sie sind Ausdruck einer Vermehrung der Mitochondrien, jedoch nicht
spezifisch für mitochondrale Myopathien. Auch im normalen Muskel älterer Menschen wur-
den vereinzelt „ragged red fibres“ nachgewiesen (RIFAI et al. 1995). Diese interpretieren die
Autoren ebenfalls als Funktionsverlust des Mitochondriums hinsichtlich des oxidativen Stoff-
wechsels, allerdings als normalen altersassoziierten Vorgang.
Auch beim Hund sind mitochondriale Myopathien und deren Ursachen in einigen Fällen
beschrieben (HERRTAGE et al. 1979, OLBY et al. 1997, SHELTON et al. 2000, PACIELLO
et al. 2003). Die Autoren gehen von Mutationen der mitochondrialen DNS aus. Diese
Mutation kann maternal erblich prädisponiert sein. Auch ein spontanes Auftreten ist möglich.
Weitere Autoren vermuten als Ursache der mitochondrialen Dysfunktion des Hundes eine
Laktat-Azidose (BREITSCHWERDT et al. 1992, VIJAYASARATHY et al. 1994). Defekte
im oxidativen Metabolismus des Mitochondriums, die in einer intrazellulären Lipid- oder
Glykogenanreicherung münden, können gelegentlich auch bei anderen Hunderassen gefunden
werden (SHELTON 1991). Deren Ursache konnte bisher nur ungenügend geklärt werden.

2.2.5 Strukturmyopathien in der Skelettmuskulatur
Es existiert eine große Anzahl von strukturellen Anomalien der Muskelfasern, die unspe-
zifisch oder, vorwiegend beim Menschen, im Zusammenhang mit einem speziellen Krank-
heitsbild auftreten können. Im Folgenden sollen nur die für die Arbeit relevanten Anomalien
besprochen werden.
Nemaline-Strukturen oder Rods stellen sich mittels der Gomori-Trichom-Färbung als
1-7 µm lange und 1 µm breite Stäbchen in den Muskelfasern dar. Sie bevorzugen die Typ-I-
Fasern.
Ihr Auftreten kann im Zusammenhang mit einer Nemaline-Krankheit (RYAN et al. 2001)
beobachtet werden. Sie treten aber auch bei anderen Myopathien und Neuromyopathien auf
und sind ebenfalls beim Hund (CARDINET 1984, DELAUCHE et al. 1998) beschrieben.
Bei diesen Prozessen ist die Anzahl der Nemaline-Strukturen enthaltenden Muskelfasern we-
sentlich geringer als bei der echten Nemaline-Krankheit (REWCASTLE und HUMPHREY
1965, DAVON et al. 1980).
Sie lagern in der Z-Streifen-Region und scheinen auch vom Z-Streifen-Material auszugehen.
Verschiedene Extraktionsversuche (ENGEL et al. 1966, ENGEL und GOMEZ 1967) haben
aber bisher keine endgültige Klärung über die Zusammensetzung des Rod-Materials ergeben.
Den Hauptbestandteil der Nemaline-Körper stellt α-actinin dar (YAMAGUGHI et al. 1982).
Die Pathogenese der Nemaline-Myopathie ist ebenfalls noch unklar.
Molekulargenetische Untersuchungen lassen jedoch den Schluß zu, dass es sich bei den
Nemaline-Körpern um sekundäre Veränderungen nach kontraktilen Dysfunktionen der
Muskulatur handelt (ENGEL et al. 1966).
Sphärische Sarkoplasmaeinschlüsse finden sich v.a. in den Typ-I-Fasern und erscheinen in
der Gomori-Trichom-Färbung grün (GOEBEL et al. 1978). Es wird angenommen, dass sie
aus verbreiterten, irregulär verlaufenden Z-Streifen entstehen (NAKASHIMA et al. 1970).
Sphärische Sarkoplasmaeinschlüsse kommen bei Denervation (ENGEL 1962), Polymyositis
(SHAFIQ et al. 1967) und verschiedenen anderen Muskelerkrankungen vor (MACDONALD
und ENGEL 1969).
Ein sogenanntes Streaming des Z-Streifens ist charakterisiert durch eine irreguläre Ausdeh-
nung von Z-Streifen-ähnlichem Material in die I- und A-Bande. In experimentellen

Untersuchungen an der Skelettmuskulatur der Ratte geht dieser Strukturanomalie ein Verlust
der Mitochondrien voraus (TICE und ENGEL 1967). Streaming ist die häufigste patholo-
gische Reaktion der Muskelfasern (BANKER und ENGEL 1994). Weniger ausgeprägte For-
men finden sich gelegentlich auch in gesunder Muskulatur.
2.2.6 Unterscheidungskriterien der Gewebssyndrome
Eine Zusammenfassung der wichtigsten Unterscheidungskriterien der einzelnen Gewebs-
syndrome ist in der nachfolgenden Tabelle 2 dargestellt
Tab. 2: Unterscheidungskriterien der Gewebssyndrome (nach JERUSALEM und ZIERZ 1991)
Myopathisches Myositisches Neurogenes
Gewebssyndrom Gewebssyndrom Gewebssyndrom
disseminierte einzelne oder perivaskuläre, perimysiale Faseratrophien und gruppierte oder endomysiale Fasertypengruppierung Muskelfaserdegenerationen entzündliche Infiltrate (Granulome)
zentrale Kerne (>3 %) zentrale Kerne (>3 %) Kernkonglomerate
pathologische pathologische ---- Kalibervariationen Kalibervariationen mit hypertrophen Fasern
Aufsplitterung ---- ----
Bindegewebsvermehrung Bindegewebsvermehrung Bindegewebsvermehrung
fettige Degeneration fettige Degeneration fettige Degeneration

2.3 Einfluss von neurologischen Störungen auf die Muskulatur
In den letzten Jahren gab es eine Anzahl von histologischen Studien bei Menschen mit Er-
krankungen der Wirbelsäule, um einen Einfluß des Krankheitsgeschehens auf die Rücken-
muskulatur herauszuarbeiten. Dabei wurde in erster Linie auf Veränderungen der Faser-
größen und der Fasertypenverteilungen geachtet.
MATTILA et al. (1986) untersuchten M. multifidus-Biopsien von Patienten mit lumbalen
Diskushernien. Sie arbeiteten Target-Fasern in Typ-I-Fasern, felderförmige Atrophien der
Muskelfasern und Fasertypengruppierungen heraus. Sie interpretierten diese Befunde als
Hinweis für stattgefundene Deinnervation und Reinnervation. Sie konnten in ihrer Arbeit
nicht herausstellen, ob allein der Schmerz (ohne Deinnervation) ausreicht, um derartige Ver-
änderungen hervorzurufen.
Ähnliche Befunde erhielten auch WEBER et al. (1997) bei Untersuchungen von Muskelbiop-
sien entnommen von Patienten mit Beschwerden im Lendenwirbelsäulenbereich. Sie konnten
bei 88 % der erkrankten Menschen pathologische Muskelveränderungen feststellen. Auch in
dieser Arbeit wurden Fasertypengruppierungen, Faseratrophien und einzelne strukturelle Um-
bauten gefunden. Sie konnten einen Zusammenhang zwischen Dauer der Schmerzen und ei-
ner Typ-II-Atrophie statistisch absichern. Keinen Zusammmenhang fanden sie zwischen der
Schmerzintensität und dem Ausmaß der Denervationszeichen.
Die Arbeitsgruppe um ZHU et al. (1989) führte histochemische und morphologische Ana-
lysen des M. erector spinae bei Patienten mit lumbalen Diskushernien durch. In 80 % der
Fälle ließ sich eine Fasertyp-II-Atrophie nachweisen. Lediglich 3 der 22 Patienten zeigten
histologisch keine pathologischen Strukturen in der Muskulatur. Die häufigsten Verände-
rungen waren Target-Fasern und binnenständige Muskelzellkerne. Seltener wurden Faser-
typengruppierungen, Bindegewebszubildungen und Mitochondrienaggregationen gefunden.
ZHU et al. (1989) führten Analysen durch, um einen Einfluß von Geschlecht und Alter der
Patienten, Höhe und Seite der Protrusion sowie der Symptomdauer herauszuarbeiten. Dabei
zeigte sich, dass Höhe und Seite der Diskushernie keinen Einfluß auf die muskulären
Veränderungen hatten. Männer wiesen signifikant größere Faserdurchmesser als Frauen auf.
Ebenso statistisch absichern ließen sich die größeren Faserdurchmesser bei jüngeren Patienten
im Gegensatz zu Älteren. Die Faserdurchmesser bei längerer Symptomdauer waren ebenfalls
nachweislich kleiner und die Bindegewebszubildungen deutlich ausgeprägter.

Auch MANNION et al. (1997, 1999, 2000) haben sich mit der paraspinalen Muskulatur von
an Rückenschmerzen erkrankten Menschen befasst. In der Arbeit von 1997 verglichen sie Bi-
opsien der paraspinalen Muskulatur von gesunden Menschen und Menschen mit Schmerzen
im Lendenwirbelsäulenbereich (mit und ohne neurologische Ausfälle). Auffallend war hier
die signifikant erhöhte Anzahl von Typ-IIB-Fasern (schnell kontrahierend, schnell ermüdend)
in der Patientengruppe. Die weiterhin gefundenen Target-Fasern entsprechen den Befunden
vorangegangener Untersuchungen.
Auch diese Autoren konnten nicht klären, ob die Veränderungen Ursache oder Wirkung der
Erkrankung darstellen. Es wäre möglich, dass die Patienten schon vor Erkrankungsbeginn
diese atypische Faserverteilung aufwiesen und damit prädisponiert waren. Die schneller er-
müdende Rückenmuskulatur kann ihre Funktion als Halte- und Stützapparat der Wirbelsäule
nicht mehr ausreichend aufrechterhalten, wodurch es zu stärkerer Beweglichkeit bei Beugung
und damit zu größerem Stress der Wirbelgelenke kommt (DOLAN und ADAMS 1995,
MANNION et al. 1997).
Im Gegensatz zur vorangegangenen Theorie, kann aber auch das Krankheitsgeschehen zu ei-
ner muskulären Veränderung führen. Das heißt, die Erhöhung des Typ-II-Faseranteils reflek-
tiert mikrostrukturell die schmerzinduzierte Inaktivität bzw. Hemmung der normalen Beweg-
ungsabläufe. Dabei stellten MANNION et al. (1997) auch fest, dass dieser Prozess schon
kurze Zeit nach Beginn der ersten Symptome zu verzeichnen ist und bei längerer Krankheits-
dauer einen Endzustand erreicht.
Die Untersuchung des Einflusses von Alter der Patienten und Dauer der Krankheitssymptome
auf die Fasertypenverteilung und die Fasergrößen durch MANNION et al. (2000) wich etwas
von den Ergebnissen anderer Autoren in früheren Arbeiten ab. Die Untersucher stellten eben-
falls eine Geschlechtsdifferenz der Fasergrößen, aber keinen Einfluß des Alters der Patienten
auf die Größe fest. Es konnte auch kein Zusammenhang zwischen der Dauer der Schmerz-
symptomatik und den Fasergrößen, unabhängig welcher Fasertypenzugehörigkeit, gefunden
werden. Lediglich eine Erhöhung des Fasertyp-II-Anteils konnte bei Patienten mit chroni-
schen Rückenbeschwerden statistisch abgesichert werden.
In einer Arbeit von RAMSBACHER et al. (2001) wurden Biopsien der paravertebralen Mus-
kulatur von Menschen mit vertebralen Instabilitäten untersucht. Der Hauptbefund dieser Un-
tersuchung war das Auftreten von „ragged red fibers“ in den Muskelfasern bei über 70 % der

Patienten. Dabei traten in fast allen dieser Fälle auch morphologische Veränderungen der
Mitochondrien auf.
2.4 Morphologie und Funktion der Mitochondrien
2.4.1 Aufbau der Mitochondrien
Mitochondrien sind Zellorganellen mit einem charakteristischen Aufbau und von sehr variab-
ler Größe. Sie sind durchschnittlich 0,2 µm breit und 2-6 µm lang; Riesenformen mit bis zu
10 µm Länge kommen vor (LEONHARDT 1990).
Die äußere Gestalt der kugel-, ei- oder stabförmigen Mitochondrien ist relativ instabil. Sie un-
terliegt im Zusammenhang mit funktionellen Erfordernissen und damit einhergehenden Be-
wegungen einem leistungsbezogenen Strukturwandel, der in Form von Entfaltung oder Rück-
bildung der mitochondralen Innenarchitektur in Erscheinung tritt (LIEBICH 1990).
Dem Mitochondrienaufbau liegt eine doppelte Zytomembran zugrunde. Durch eine glatt ver-
laufende Außenmembran wird das Mitochondrium gegen das umgebende Zytoplasma abge-
grenzt. Im Abstand von 8-10 nm zum Inneren folgt eine Innenmembran, die durch Ausfal-
tungen eine große Oberfläche herstellt (MITCHELL 1961).
In den meisten Fällen sind die Ausfaltungen kulissenartig leistenförmig und stehen quer zur
Längsachse: als Crista-Typ bezeichnet. Die Anzahl der Cristae innerhalb eines Mitochon-
driums ist funktions- und zellabhängig und Ausdruck der mitochondrialen Stoffwechsel-
aktivität. Daher zeigen besonders die Zellen der Muskulatur, der Leber und der Niere eine
hohe Dichte der Cristae. Nach Hungerzuständen ist ihre Zahl reduziert.
Weniger häufig kommen fingerförmige Ausstülpungen in der Längsachse vor: als Tubulus-
Typ bezeichnet. Vorzugsweise sind diese Mitochondrien mit der Synthese von Steroidhor-
monen verbunden (z.B. in Zellen der Nebennierenrinde oder den Leydig-Zellen).
Eine Sonderform sind Mitochondrien mit Prismen oder Sacculi, die z.B. in einigen Nerven-
zellen vorkommen (LIEBICH 1990).
Das von beiden Mitochondrienmembranen begrenzte Spatium intermembranosum bildet das
äußere Kompartiment, das mit dem intracristalen bzw. intratubulären Raum kommuniziert.

Die innere Mitochondrienmembran schließt das funktionell wesentlich bedeutsamere innere
Kompartiment ein, das mit der Mitochondrienmatrix identisch ist (MITCHELL 1961).
Diese enthält nahezu alle Enzyme des Zitratzyklus, Nukleinsäuren, Ribosomen und 30-50 nm
große Matrixgranula, die reich an Kalzium und anderen Ionen sind und bei der Regulation des
inneren Milieus mitwirken (LIEBICH 1990).
Die Anordnung der membranständigen Enzyme folgt einer Ordnung, weshalb Mitochondrien
auch als „geordnete Multienzymsysteme“ bezeichnet werden (LEONHARDT 1990).
Die Mitochondrienmembranen bestehen zu 50-60 % aus Proteinen und zu rund 40 % aus
Lipiden, allerdings mit chemischen Unterschieden der Fettzusammensetzung, die mit Funkti-
onsunterschieden korrelieren.
In der äußeren Membran befinden sich porenbildende Proteine (Porine), die diese für Mole-
küle bis 10 kDa (und damit für die meisten Metaboliten) passiv permeabel machen.
Charakteristische Leitenzyme der äußeren Mitochondrienmembran sind eine flavinhaltige
Aminooxidase sowie eine NADH-Cytochrom-C-Reduktase.
Im Gegensatz zur äußeren ist die innere Mitochondrienmembran für die meisten Metaboliten
impermeabel, dagegen ist sie für die Atmungsgase Sauerstoff und Kohlendioxid passiv durch-
lässig (MITCHELL 1961,1976, RACKER 1970).
2.4.2 Struktur der mitochondralen DNS
Mitochondrien besitzen ein eigenes Genmaterial, DNS und RNS, das den sogenannten Plas-
magenen (extrachromosomalen Genen) zugeordnet wird.
Das Auftreten dieser extranukleären DNS wird mit der Annahme verbunden, dass Mitochon-
drien aus Prokaryonten entstanden sind (GRIVELL 1983). Ihr Anteil am Gesamt-DNS-Gehalt
der Zelle kann bis zu 2 % betragen.
Die DNS der Mitochondrien ist ringförmig und gelegentlich als verzweigte, filamentöse etwa
5 µm lange Nukleotidkette sichtbar. Sie hat im Gegensatz zur nukleären (chromosomalen)
DNS keine Verbindung mit Histonen (CLAYTON und SMITH 1975).
Die RNS, die für die Übertragung der genetischen Informationen verantwortlich ist, liegt in
Form von etwa 12 nm großen Ribonukleoproteingranula in der Matrix.

2.4.3 Vermehrung der Mitochondrien
Mitochondrien sind aufgrund ihres Genoms semiautonome Zellorganellen mit eigener Repli-
kation, Transkription und Translation. Allerdings kodiert die mitochondriale DNS weniger als
5 % der Mitochondrienproteine. Die Hauptmenge wird zellkodiert an zytoplasmatische Ribo-
somen synthetisiert (CLAYTON und SMITH 1975).
Die Mitochondrien vermehren sich grundsätzlich durch Teilung senkrecht zur Längsachse,
der in der Regel die Replikation mitochondrialer DNS vorausgeht.
Sie haben eine Lebensdauer von ungefähr 20 Tagen und werden durch Autophagolysosomen
intrazellulär abgebaut (LIEBICH 1990).
Die Mitochondrienvermehrung ist prinzipiell während des gesamten Lebenszyklus einer Zelle
möglich. Sie dient der Erneuerung der Mitochondrien und der Adaptation der zellulären Stoff-
wechselleistung an aktuelle Erfordernisse, da die Mitochondrienanzahl in enger Beziehung
besonders zum Energiestoffwechsel der betreffenden Zelle steht.
2.4.4 Funktion der Mitochondrien
Die Lebensfähigkeit einer Zelle hängt von der Leistung eigener stoffwechselaktiver Orga-
nellen und vom intrazellulären Stofftransport fester und gelöster Substanzen ab. Zur Auf-
rechterhaltung dieser Funktionen muss die Zelle über eigene Energiequellen verfügen. Diese
Aufgabe übernehmen die Mitochondrien. Sie setzen Energie aus der Zellatmung durch die
oxidative Phosphorylierung frei und bilden außerdem spezielle Stoffwechselenzyme
(BELITZER und TSIBAKOVA 1937, LIPMAN 1941, MITCHELL 1961).
Durch den Abbau von Zucker und Fettsäuren über mehrere Stufen (Endoxidation) entstehen
Wasser und Kohlendioxid. Die dabei freiwerdende Energie steht der Zelle in Form von
Adenosintriphosphat (ATP) zur Verfügung (LIPMAN 1941, MITCHELL 1961).
Zusätzlich sind weitere Stoffwechselleistungen wie z.B. der Harnstoffzyklus oder die
Glukoneogenese an die Mitochondrien gebunden.

2.4.5 Mitochondriopathien
„Zu den Mitochondriopathien gehören alle diejenigen Krankheitsbilder, bei denen sich der
primäre pathogenetische Prozess in den Mitochondrien abspielt.“ (RIEDE und SCHAEFER
1993)
Dazu gehören: - Mitochondrale Myopathien
- Hypoxidosen
- Autoimmunerkrankungen mit mitochondrialen Autoantikörpern
(RIEDE und SCHAEFER 1993).
2.4.5.1 Mitochondrale Myopathien
Um von einer mitochondralen Myopathie sprechen zu können, müssen verschiedene Kriterien
erfüllt sein.
Es bedarf einer morphologisch nachweisbaren Mitochondrienanomalie, einem spezifischen
biochemischen Defekt und einem klinischen Bild, das durch einen Mitochondrienstoff-
wechseldefekt erklärbar ist (JERUSALEM 1991).
Die Ursachen für die Erkrankung können sehr unterschiedlich sein:
1. Defekte im mitochondralen Substrattransport (z.B. Carnitinmangel).
2. Eine Störung der mitochondralen Substratverwertung.
3. Atmungskettendefekte (Komplex I- bis IV-Defizienz).
4. Defekte der Energiekonservierung und -überführung.
5. Eine m-DNS Mutation.
Die Veränderungen der Mitochondrien betreffen sowohl die Skelettmuskulatur als auch das
Gehirngewebe und werden dort als mitochochondrale Enzephalo-Myopathien bezeichnet
(BRENNER et al. 1992, RIEDE und SCHAEFER 1993).
In der Gomori-Trichom-Färbung fallen auf Muskelquerschnitten irregulär konfigurierte
Muskelfasern mit roten Granula, „ragged red fibres“, auf. Elektronenmikroskopisch sind die
Mitochondrien meist vergrößert, abnorm gebaut und enthalten parakristalline Einschlüsse
(JERUSALEM 1991, RANDAL 1991, GRUBER et al. 2002).

2.4.5.2 Hypoxidosen
Hypoxidosen sind Krankheitsbilder, deren Pathogenese in einer Störung der oxidativen
Energiegewinnung besteht (RIEDE und SCHAEFER 1993).
• Hypoxämische Hypoxidosen
Sie werden durch einen zu geringen Sauerstoffgehalt des arteriellen Blutes aufgrund von
Ventilationsstörungen (Verlegung der Atemwege), pulmonale Perfusionsstörungen (z.B.
Emboli), Diffusionsstörungen (Behinderung des pulmonalen Gasaustausches) oder durch eine
Verminderung des Sauerstoffpartialdruckes in der Außenluft hervorgerufen (RIEDE und
SCHAEFER 1993).
• Ischämische Hypoxidosen
Sie werden durch eine unzureichende Versorgung des Gewebes mit sauerstoff- und nährstoff-
reichem Blut verursacht. Neben diesem Mangel an Substraten ist auch der Abtransport von
Stoffwechselendprodukten und Kohlendioxyd reduziert, so dass eine Gewebsazidose entsteht.
Gründe dafür können Gefäßstenosen und Gefäßverschlüsse durch Thromben oder Emboli sein
(RIEDE und SCHAEFER 1993).
• Hypoglykämische Hypoxidosen
Sie sind die Folge einer unzureichenden Bereitstellung oxydierbarer Substanzen durch
mangelnde Nahrungszufuhr (z.B. Hungern), Malabsorption oder Maldigestion (RIEDE und
SCHAEFER 1993).
• Histotoxische Hypoxidosen
Sie entstehen durch eine Blockierung der intrazellulären Energiegewinnnung, z.B. Ent-
kopplung der oxydativen Phosphorylierung und ATP-Bildung, Störung des Elektronen-
transportes oder Behinderung der Substrataufnahme in die Zelle (RIEDE und SCHAEFER
1993).

2.5 Aufbau der Wirbelsäule
Im Laufe der embryonalen Entwicklung der Wirbelsäule entsteht bei den Vertebraten ein zell-
reicher Achsenstrang, die sogenannte Chorda dorsalis. Diese wird in der frühen embryonalen
Phase durch knorpelige und knöcherne Wirbelsäulenbestandteile ersetzt.
Das Wachstum der Wirbelkörper erfolgt von der Proliferationszone der Knorpelendplatten
her, die beim Hund mit ca. 11 Monaten geschlossen sind (HARE 1961).
Die Wirbelsäule, Columna vertebralis, besteht aus den Wirbeln, den Zwischenwirbelscheiben
und den dazugehörigen Bandstrukturen. Die Wirbel setzen sich aus dem Körper (Corpus
vertebrae), dem Wirbelbogen (Arcus vertebrae), den Fortsätzen zur Kraftübertragung
(Processus transversi, Processus spinosi) sowie den Gelenkfortsätzen (Processus articulares)
zu den benachbarten Wirbeln zusammen. Die aneinandergereihten Wirbelbögen bilden den
dorsalen und lateralen Anteil des Wirbelkanals (Canalis vertebralis), in dem das Rückenmark
eingeschlossen ist.
Zwischen jedem Wirbelpaar befindet sich ein Foramen intervertebrale, durch welches die Spi-
nalnerven und Gefäße hindurchtreten.
Die Wirbelsäule des Hundes weist 7 Hals-, 13 Brust-, 7 Lenden-, 3 Kreuz- und eine unter-
schiedliche Anzahl an Schwanzwirbeln auf (NICKEL et al. 1992).
Zwischen den Wirbelkörpern liegen die Bandscheiben. Sie bestehen aus einem Anulus
fibrosus (bindegewebiger und knorpeliger äußerer Ring) und einem Nucleus pulposus (innerer
Gallertkern).
Der Anulus fibrosus besteht aus bindegewebigen Faserzügen, die lamellenartig zwischen den
Endplatten der Wirbelkörper verlaufen. Zum Zentrum der Bandscheibe hin verliert er seine
typische Struktur und besteht eher aus faserknorpligen Anteilen (NICKEL et al. 1992).
An der ventralen Seite ist der Anulus ca. 1½ bis 3-fach so dick wie an der dorsalen Seite
(EVANS 1993).
Der Nucleus pulposus liegt somit exzentrisch in der Bandscheibe (THATCHER 1989). Er
stellt ein Überbleibsel der Chorda dorsalis dar. Er besteht aus einem Kollagengitter, das in
eine gelatinöse Masse aus Chondrozyten, Hyaluronsäure, Chondroitinsulfat und Keratinsulfat
eingebettet ist (THATCHER 1989).

Funktionell puffern die Zwischenwirbelscheiben die Bewegungen der einzelnen Wirbel
gegeneinander ab, wobei der Druck auf den Nucleus pulposus nach allen Seiten verteilt auf
den Anulus fibrosus weitergegeben und dort in Spannungskräfte umgewandelt wird (NICKEL
et al. 1992).
Lediglich zwischen dem ersten und zweiten Halswirbel (Atlas und Axis) befindet sich eine
gelenkige Verbindung, welche die Drehung des Kopfes ermöglicht.
Im Wesentlichen erfüllt die Wirbelsäule eine Halte- und Ausgleichsfunktion. Kräfte in Form
von Spannungs-, Stoß- und Druckbelastungen werden durch verschiedene Mechanismen
kompensiert und/oder übertragen.
Eine wichtige Rolle spielen dabei auch die Bänder innerhalb und außerhalb des Wirbelkanals.
Es gibt vier bedeutende Bänder zwischen dem 1. und 2. Halswirbel. Das wichtigste ist das
Lig. transversum atlantis, das von der linken zur rechten Atlasseite verläuft und dabei den
Dens axis überquert. Außerdem befinden sich am Wirbelboden von Atlas und Axis noch das
Lig. apicis dentis und die Ligg. alaria. Dorsal verläuft das Lig. atlantoaxiale (NICKEL et al.
1992).
Weiterhin von Bedeutung sind das dorsale und ventrale Lig. longitudinale, welche die Wirbel
von Hals bis Schwanz miteinander verbinden. Die Fasern des Lig. longitudinale dorsale sind
im Bereich des Intervertebralspaltes etwas dünner und verschmelzen mit den Fasern des
Anulus fibrosus. Außerdem wurden in diesem Ligament Schmerzfasern nachgewiesen
(FORSYNTHE und GHOSHAL 1984).
Zwischen den Rippenköpfchen (außer Th1, Th12 und Th13) verläuft das Lig. intercapitale
unter dem Lig. longitudinale dorsale. Mit dem Vorhandensein dieses Ligamentes lässt sich
auch die geringe Inzidenz von Bandscheibenvorfällen im Bereich zwischen dem 2. bis 11.
Thorakalwirbel erklären (THATCHER 1989, TOOMBS und WATERS 2003).
Außerhalb des Wirbelkanales befinden sich noch Bänder zwischen und auf den Processus
spinosi, zwischen den Processus transversus und den Processus articulares (NICKEL et al.
1992).
Von großer Bedeutung für die frühzeitigen degenerativen Wirbelsäulenerkrankungen ist die
besondere Stoffwechsellage des adulten Bandscheibenapparates. Mit Blutgefäßen wird nur
der äußere Anteil des Anulus fibrosus versorgt. Die Ernährung des restlichen Gewebes erfolgt

durch Diffusion aus der Peripherie. Außerdem sind sie fortwährenden Druckbelastungen
ausgesetzt (FREWEIN et al. 1986).
2.6 Aufgabe und Funktion der Rückenmuskulatur
Die Rückenmuskulatur stellt das aktive Element des Wirbelsäulensystems dar. Sie ist den un-
terschiedlich gut beweglichen Abschnitten - Hals-, Brust- und Lendenwirbelsäule - dorsal,
lateral und ventral unmittelbar aufgelagert.
Die Mehrzahl der Muskeln liegt in mehreren Schichten kulissenartig zwischen den Quer- und
Dornfortsätzen der Wirbel und wird von den Dorsalästen der Spinalnerven innerviert. Wenige
Muskeln (M. longus colli, M. scalenus) liegen ventral der Querfortsätze im Innervationsgebiet
der Ventraläste der Rückenmarksnerven (BRAUND et al. 1977, HUTTER 1977, KELLER
1963).
Nach funktionellen Gesichtspunkten lässt sich die Muskulatur der Wirbelsäule in zwei große
Gruppen unterteilen:
1. die Aufrichter, Seitwärtsbieger und Dreher der Hals-, Brust- und Lendenwirbelsäule,
2. die Abwärtsbieger der Halswirbelsäule.
Die erste Gruppe besteht aus den langen Hals- und Rückenmuskeln (M. splenius, M. ilio-
costalis, M. longissimus, M. spinalis, M. semispinalis und die Mm. multifidi) und den kurzen
Hals- und Rückenmuskeln (Mm. intertransversarii, Mm. rotatores).
Zu den Abwärtsbiegern der Halswirbelsäule gehören der M. longus colli und die Mm. scaleni
(KRÜGER 1927, NIEDERER 1985).
Neben der Bewegung der einzelnen Wirbelsäulenabschnitte und Wirbel haben die Hals- und
Rückenmuskeln auch eine statische Funktion im Sinne einer Verstärkung von Zusammenhalt
und Tragfähigkeit der Wirbelsäule.

2.7 Rückenmark und begleitende Strukturen
Das Rückenmark bildet mit dem Gehirn das Zentrale Nervensystem. Es liegt im Wirbelkanal
und ist dort von seinen Hüllen (Dura mater und Leptomeninx) umschlossen und wird von
weiteren Strukturen (Gefäße, Fettgewebe usw.) begleitet.
Das Rückenmark geht in Höhe des ersten Halswirbels aus der Medulla oblongata hervor
(BUDRAS u. FRICKE 1991). Es wird durch seine Lage im Wirbelkanal und den segmental
entspringenden Spinalnerven in das Halsmark (Pars cervicalis), das Brustmark (Pars tho-
racica), das Lendenmark (Pars lumbalis), das Kreuzmark (Pars sacralis) und das Schwanz-
mark (Pars coccygea) unterteilt (NICKEL et al. 1992). Sowohl die Länge der einzelnen
Rückenmarkssegmente als auch deren Lage zu den gleichnamigen Wirbeln variiert
(FLETCHER 1993).
Am Hals-Brust-Übergang und im hinteren Bereich des Lendenmarkes weist das Rückenmark
jeweils spindelförmige Anschwellungen auf, die Intumescentia cervicalis bzw. lumbalis. Hier
entspringen die Spinalnerven, die die Vorder- bzw. Hinterextremitäten versorgen und den
Plexus brachialis bzw. lumbosacralis bilden. Hinter der Intumescentia lumbalis verjüngt sich
das Rückenmark in Richtung Schwanz (Conus medullaris) und läuft schließlich als dünner
Endfaden (Filum terminale) aus (FLETCHER 1993).
Durch ein verzögertes Wachstum des Rückenmarkes gegenüber der Wirbelsäule (Ascensus
medullae spinalis) verlaufen die Spinalnerven der letzten Lumbal- und Sakralnerven noch
innerhalb des Wirbelkanals längs des Conus medullaris bis zum Austritt durch die entsprech-
enden Foramina intervertebralia und bilden die Cauda equina (BUDRAS u. FRICKE 1991).
Im Querschnitt lassen sich makroskopisch die graue und die weiße Substanz unterscheiden.
Die zentral gelegene graue Substanz (Substantia grisea) weist einen schmetterlingsförmigen
Querschnitt auf. Sie besteht aus Neuronen, Neuroglia und Blutgefäßen. Median in der grauen
Substanz verläuft der Zentralkanal (Canalis centralis), der mit Liquor cerebrospinals gefüllt
ist.
Umgeben wird die graue Substanz von der weißen Substanz (Substantia alba). Sie besteht aus
myelinisierten und nichtmyelinisierten Axonen, die funktionell und anatomisch (durch längs
verlaufende Rillen und Furchen) in verschiedene Stränge unterteilt werden können (NICKEL
et al. 1992).

Umgeben wird das Rückenmark von seinen drei Häuten (Meningen). Sie gehen kaudal des
Foramen magnum aus den Hirnhäuten hervor. Innen, mit dem Rückenmark direkt verbunden
und sich seinen Formen anschmiegend, befindet sich die Pia mater. Sie stellt eine dünne, ge-
fäßreiche Schicht aus Kollagenfasern und abgeflachten Fibroblasten dar.
Die Spinnwebenhaut (Arachnoidea) ist die mittlere Schicht. Sie ist von der Pia mater durch
den Subarachnoidalraum, in dem sich Liquor cerebrospinalis befindet, getrennt. Pia mater und
Arachnoidea bilden gemeinsam die Endomeninx. Außen liegt die Exomeninx. Sie besteht aus
zwei Blättern, dem Periost, das den Wirbelkanal auskleidet und der Dura mater (Pachyme-
ninx), der die Arachnoidea von innen anliegt. Die Dura mater ist eine derbe, gefäßarme, fibrö-
se Haut, die einen Schlauch um das Rückenmark und die abgehenden Spinalnerven bildet.
Zwischen Periost und Dura mater befindet sich das Cavum epidurale, das mit Fettgewebe,
Lymphgefäßen und einem ventral gelegenen Venenplexus gefüllt ist (THATCHER 1989,
NICKEL et al. 1992, FLETCHER 1993).
2.8 Rückenmarkskompressionen
Im Folgenden soll kurz auf Erkrankungen eingegangen werden, die bei den Hunden vorlagen,
die für eine Muskelbiopsie ausgewählt wurden.
2.8.1 Arten von Rückenmarkskompressionen
Die häufigsten Ursachen für eine Kompression des Rückenmarks beim Hund stellen Band-
scheibenvorfälle dar.
Seltener sind vertebrale Instabilitäten, Neoplasien oder traumatische Einwirkungen von außen
Ursache für eine Schädigung des Rückenmarks.
Degenerative Bandscheibenveränderungen können im Wesentlichen nach zwei unterschied-
lichen Mustern ablaufen. Sie werden aufgrund ihrer Ätiologie nach HANSEN (1952) unter-
teilt. Zum einen spricht man von einer chondroiden Metaplasie. Dabei kommt es zu einer
Wasserabnahme, Degeneration, dystrophischen Verkalkung und Nekrose des Nucleus pulpo-
sus, sowie im weiteren Verlauf des Anulus fibrosus. Der damit verbundene Elastizitätsverlust
des Diskus führt zu verminderter Pufferkapazität und zur Auffaserung des Anulus fibrosus. Es

kommt zur vollständigen Ruptur des Anulus fibrosus und zum Austritt von Nucleus-pulposus-
Material in den Wirbelkanal. Es handelt sich somit um eine Diskusextrusion, einen Diskus-
prolaps oder einen Hansen-Typ-I Vorfall.
Diese Art der Degeneration findet sich in erster Linie bei chondrodystrophen Rassen (z.B.
Dackel, Pekingese, Zwergpudel). Sie beginnt schon im jungen bis mittleren Alter
(SCHOLTYSIK 1962).
Eine zweite Form der Diskusdegeneration ist die fibroide Metaplasie. Es kommt ebenfalls zu
einer Dehydratation und Degeneration des Nucleus pulposus. Dies findet aber erst im fortge-
schrittenen Alter statt.
Bei der fibroiden Metaplasie ist der Anulus fibrosus nicht vollständig, sondern nur partiell
rupturiert, und es kommt zur Vorwölbung in den Wirbelkanal. Man spricht dann von einer
Diskusprotrusion, einer Diskushernie oder einem Hansen-Typ-II Vorfall.
Nach Untersuchungen von GOSH et al. (1975) und GYSLING (1984) ist allerdings die starre
Trennung nach Hansen in chondrodystrophe Hunde mit Prolaps-Typ-I bzw. nichtchondro-
dystrophe Hunde mit Prolaps-Typ-II nicht vollständig aufrechtzuerhalten. Sie fanden bei ein-
zelnen Hunden nichtchondrodystropher Rassen bereits frühzeitige Degenerationen der Zwi-
schenwirbelscheiben. Ebenso konnte RIST (1982) bei Sektionen an 38 Dackeln in immerhin
17 Fällen Vorfälle vom Hansen-Typ-II nachweisen.
Auch GREVEL und COP (1992) fanden bei Hunden großer Rassen (also nichtchondro-
dystropher Rassen) Diskusprolapsus mit vollständiger Ruptur des Anulus fibrosus.
Deutlich seltener als degenerative Bandscheibenveränderungen sind vertebrale Instabilitäten
die Ursache für eine Rückenmarkskompression beim Hund. Instabilitäten sind meistens auf
bestimmte Wirbelsäulenabschnitte beschränkt, die von kranial nach kaudal entsprechenden
Krankheitsbildern zugeordnet werden:
1.) Die Atlantoaxiale Instabilität kann verschiedene Ursachen haben. Der häufigste Grund
ist eine Aplasie oder Hypoplasie des Dens axis. Es können aber auch eine Fraktur des Dens
oder Mißbildung bzw. Ruptur des atlantoaxialen Bandapparates Instabilitäten hervorrufen
(WATSON und DELAHUNTA 1992, WHEELER 1992).
2.) Die kaudale zervikale Spondylomyelopathie (KZS; Wobbler Syndrom) kann ebenfalls
verschiedene Ursachen aufweisen. Die häufigste Lokalisation bei Deutschen Doggen ist zwi-

schen dem 4. und 5. bzw. 5. und 6. Halswirbel; beim Dobermann zwischen dem 5. und 6.
bzw. dem 6. und 7. Halswirbel (LEWIS 1989, JURINA 1996, 2002a). Die KZS kann durch
verschiedene Veränderungen hervorgerufen sein. Es können dynamische und nichtdynami-
sche Diskusprotrusionen, spondylotische Zubildungen, Stenosierung des Wirbelkanals durch
Malformation der Wirbel, Gelenkkapselproliferationen oder Hypertrophie des Lig. flavum die
Ursache für eine Kompression des Rückenmarks sein (DENNY et al. 1977, LEWIS 1992,
SEIM und WITHROW 1982).
3.) Die lumbosakrale Stenose oder das Cauda Equina Syndrom entstehen in Folge von
Veränderungen am Übergang von Lendenwirbelsäule zum Kreuzbein. Das Rückenmark endet
bei den großen Hunden meist auf der Höhe des 5. bis 6. Lendenwirbels, so dass sich lumbo-
sakral nur noch vom Rückenmark abgehende Nervenwurzeln (Cauda equina) befinden. Die
Kompression der Cauda equina oder einzelner Nervenwurzeln kann durch Instabilität im
Lumbosakralgelenk, Stenosierung des Wirbelkanals durch Malformation der Wirbel, Diskus-
protrusion, Weichteilgewebsproliferation oder einer Osteochondrosis des Os sacrum verur-
sacht werden (JAGGY et al. 1987, LANG et al. 1992, WHEELER 1992).
Neoplasien im Bereich der Wirbelsäule treten vergleichsweise selten auf, präsentieren sich
aber häufig mit einer ebenso ausgeprägten Symptomatik wie Diskopathien oder Traumata.
Die erkrankten Tiere sind in der Mehrzahl mittleren bis hohen Alters, doch auch beim jungen
Hund kann eine Neoplasie die Ursache sein (SUMMERS et al. 1988).
Es sind viele verschiedene Tumorarten beschrieben, aber nur mit wenigen Fällen histologisch
belegt (BAGLEY et al. 1993).
Spinale Neoplasien werden, wie auch alle anderen Rückenmarkskompressionen, nach ihrer
Lokalisation zum Rückenmark unterteilt. Extradural gelegene Tumoren kommen mit etwa
50 % am zahlreichsten vor. Intradurale-extramedulläre Veränderungen sind bei etwa 35 % der
Hunde zu finden, seltener sind intramedulläre Neoplasien (PRATA 1977).
Nicht selten sind neben den primären Tumoren auch sekundäre neoplastische Veränderungen
im Sinne einer Metastasierung Grund für eine Rückenmarkskompression.

2.8.1.1 Zervikale Diskopathie
Es sind häufig kleinwüchsige Hunde, vorwiegend chondrodystropher Rassen (Dackel, Beagle,
Shi-Tzu, Chihuahua) betroffen. Die Tiere sind meist älter als 2 Jahre. Man rechnet mit einem
gehäuften Auftreten im Alter von 6 Jahren. Es liegt keine Geschlechtsprädisposition vor
(DENNY 1978, DALLMANN et al. 1992).
Bei über der Hälfte der in der Klinik vorgestellten Patienten mit zervikaler Diskopathie lassen
sich die klinischen Symptome anamnestisch länger als 14 Tage zurückverfolgen (JURINA
1996, JURINA und GREVEL 2002a).
Das Hauptsymptom der Patienten mit einem Diskusprolaps im Bereich der Halswirbelsäule
sind Schmerzäußerungen bei Bewegung des Halses bzw. bei Palpation der Muskulatur und
der Wirbel (SEIM und PRATA 1982). Sie zeigen häufig eine gesenkte Kopf-Hals-Haltung
(TOOMBS und WATERS 2003).
Aufgrund des größeren Durchmessers des Wirbelkanals im Bezug zum Rückenmark treten im
Bereich der Halswirbelsäule, im Gegensatz zum Thorakalbereich, neurologische Ausfälle sel-
tener auf (BERG 1989).
Lahmheit einer Vordergliedmaße kann Hinweis auf eine Irritation der Nervenwurzel sein.
Monoparesen bis hin zu Tetraparesen sind seltener feststellbar.
Ein Diskusprolaps tritt bei den genannten Rassen am häufigsten zwischen dem 2. und 3.
Halswirbel auf. Die Inzidenz nimmt nach kaudal ab (DALLMANN et al. 1992, JURINA
1996, JURINA und GREVEL 2002a).
2.8.1.2 Thorakolumbale Diskopathie
In dieser Lokalisation tritt ein Diskusprolaps hauptsächlich bei chondrodystrophen Rassen im
Alter von 4 bis 8 Jahren, mit einem Maximum im Alter von 5 Jahren, in Erscheinung. Bei
allen anderen Rassen kommt der thorakolumbale Diskusprolaps deutlich seltener vor, auch
sind die Tiere meist schon im mittleren bis hohen Alter (GREVEL und SCHWARTAU 1997).
Es handelt sich bei den chondrodystrophen Tieren vorwiegend um Hansen-Typ-I Läsionen,
im Gegensatz zu den Hunden anderer Rassen, die in der Mehrzahl Hansen-Typ-II Läsionen
aufweisen.
Über 50 % aller thorakolumbalen Bandscheibenvorfälle finden sich zwischen dem 12. und

13. Brustwirbel bzw. zwischen dem 13. Brust- und dem ersten Lendenwirbel (FUNKQUIST
1962, GREVEL und SCHWARTAU 1997).
Die klinischen Symptome sind je nach Ausmaß der Kompression unterschiedlich. Es gibt
Tiere, die eine Schmerzsymptomatik aber keine bzw. kaum neurologische Defizite zeigen. Sie
sind unwillig zu laufen und fallen durch eine Kyphose und Dolenz bei Palpation der
Wirbelsäule auf.
Neurologische Ausfälle reichen von einer Ataxie der Hintergliedmaßen, einer gehfähigen oder
nichtgehfähigen Paraparese bis hin zur Paraplegie mit oder ohne Erhalt der Tiefensensibilität
(LEVINE und CAYWOOD 1984, GRIFFITHS 1992).
2.8.1.3 Atlantoaxiale Instabilität
Die atlantoaxiale Instabilität wird vor allem bei Zwergrassen (Yorkshire Terrier, Zwergpudel,
Zwergspitz, Pekingese) gesehen. Die häufigste Ursache ist eine kongenitale Hypoplasie oder
eine Aplasie des Dens axis, die zu klinischen Symptomen schon bei sehr jungen Hunden
führt. Bei älteren Tieren sind meist traumatisch bedingte Läsionen der Grund für das Krank-
heitsbild (DENNY 1983, WATSON und DELAHUNTA 1992).
Einzelne Fälle von atlantoaxialer Instabilität sind auch bei größeren Hunderassen beschrieben
(WHEELER 1992).
Die Instabilität verursacht Symptome, die ähnlich denen der zervikalen Diskopathie sind. Das
häufigste Symptom ist eine Schmerzhaftigkeit.
Die neurologischen Defizite reichen von einer Ataxie mit Propriozeptionsstörungen bis zu ei-
ner Tetraparese oder Tetraplegie (GEARY et al. 1967, COOK und OLIVER 1981).
2.8.1.4 Lumbosakrale Stenose
Eine lumbosakrale Stenose wird vorwiegend bei Hunden großwüchsiger Rassen, z.B.
Deutscher Schäferhund, diagnostiziert. Gelegentlich können aber auch kleinere Rassen
Probleme aufweisen. Es erkranken Hunde aller Altersklassen, aber junge Hunde, die zum
Leistungssport eingesetzt werden, scheinen prädisponiert zu sein (CHAMBERS 1989,
WHEELER 1990, 1992).
Die Symptome sind vom Grad der Einengung des Wirbelkanals abhängig. Vorherrschende
Symptome sind Bewegungsunlust und Aufjaulen beim Sprung, intermittierende Lahmheiten

sowie Schmerzhaftigkeit bei Druck auf die Wirbelsäule am lumbosakralen Übergang und
beim Strecken einer oder beider Hintergliedmaßen. Die Tiere zeigen typischerweise einen
aufgekrümmten Rücken mit stark abfallender Kruppe und leicht untergestellten
Hintergliedmaßen.
Lahmheiten der Hintergliedmaße können Ausdruck einer Wurzelreizung sein. Die neuro-
logischen Symptome sind meist nicht ausgeprägt, jedoch sind Mono- und Paraparesen
möglich. Kompressionen mit Beeinträchtigung des Nervus pudendus können Harn- und
Kotinkontinenzen hervorrufen (WHEELER 1990, 1992).
2.8.1.5 Kaudale Zervikale Spondylomyelopathie Es sind hauptsächlich Hunde großer und Riesenrassen betroffen, davon über 80 % Dober-
mann und Deutsche Dogge (BRUECKNER et al. 1989, LEWIS 1989).
Die Mehrzahl der Hunde ist zur ersten klinischen Vorstellung mittelalt bis alt, mit Ausnahme
der Doggen. Bei dieser Rasse sind schon junge Tiere (<1 Jahr) klinisch auffällig.
Es handelt sich um ein multifaktorielles Erkrankungsbild, das eine exakte Aussage über die
Wertigkeit der verschiedenen Ursachen und deren klinische Bedeutung kaum zulässt (SEIM
und WITHROW 1982).
Die Symptome richten sich nach der von der Kompression betroffenen Struktur. Bei einer
Affektion der Spinalnerven (Radikulopathie) fallen vor allem eine Dolenz, Lahmheit einer
oder beider Vordergliedmaßen und, bei entsprechend langer Erkrankungsdauer, eine Atrophie
der Schultermuskulatur auf.
Bei einer Kompression des Rückenmarks (Myelopathie) zeigen die Tiere hauptsächlich eine
Ataxie, die an den Hintergliedmaßen ausgeprägter ist. In besonders schwerwiegenden Fällen
können sich die Hunde auch mit einer Tetraparese präsentieren (LEWIS 1989).
Ein breitbeiniger Stand der Vorder- oder Hintergliedmaßen und eine gesenkte Kopf-Hals-
Haltung stellen das typische Bild eines an kaudaler zervikaler Spondylomyelopathie erkrank-
ten Hundes dar.

2.8.2 Diagnose von Rückenmarkskompressionen
In der Mehrzahl der Fälle sind der Vorbericht und die klinischen Symptome der Tiere schon
ausreichend, um den Verdacht auf eine Rückenmarkskompression äußern zu können. Gele-
gentlich können aber unspezifische Schmerzen und unklare Lahmheiten, die zunächst ein
orthopädisches Problem vermuten lassen, die Diagnosefindung erschweren.
Im Anschluss an die Anamnese, die häufig richtungsweisend ist, folgen die allgemeine und
neurologische Untersuchung. Letztere soll herausstellen:
• handelt es sich tatsächlich um ein neurologisches Problem
• welche neurologischen Defizite liegen vor. Ist das obere oder untere motorische Neuron
betroffen: - Ataxie, gehfähige Para- oder Tetraparese
- nichtgehfähige Para- oder Tetraparese
- Para-/Tetraplegie mit fehlendem Harnabsatz und erhaltenem Tiefenschmerz
- Para-/Tetraplegie mit fehlendem Harnabsatz und fehlendem Tiefenschmerz
(GRIFFITHS 1992)
• in welchem Wirbelsäulenabschnitt und auf welcher Seite ist die Kompression zu
vermuten: - C1-C5
- C6-Th2
- Th3-L3
- L4-S3 (BRAUND 1985)
Gegebenenfalls müssen nichtkompressive und entzündliche Wirbelsäulenerkrankungen aus-
geschlossen werden. Dafür empfiehlt sich neben Blut- und Harnuntersuchungen eine Ana-
lyse des Liquors (CHRISMAN 1992).
Der wichtigste Untersuchungsgang bei Patienten mit Wirbelsäulenerkrankungen ist die radio-
logische Diagnostik. Vorraussetzung für die Anfertigung diagnostisch verwertbarer Röntgen-
aufnahmen sind gute technische Bedingungen und vor allem eine gute Lagerung des Patienten
(FLÜCKINGER 1985, DENNIS 1987). Schon eine geringgradig asymmetrische Lagerung
kann bewirken, dass normale Strukturen röntgenologisch verändert erscheinen, bzw. dass
subtile Veränderungen übersehen werden (BRAWNER und HATHCOCK 2003). Dabei wer-
den zunächst Nativröntgenaufnahmen angefertigt. In einigen Fällen sind diese Aufnahmen

bereits ausreichend für eine Diagnose, denn jede Wirbelsäulenerkrankung ruft eine Anzahl
typischer und indirekter Veränderungen hervor, die im Röntgenbild sichtbar sind.
In der Mehrzahl der Erkrankungsfälle erweist sich aber eine Myelographie als notwendig.
Befindet sich der erkrankte Abschnitt im Bereich der Halswirbelsäule, ist eine Kontrast-
mittelinjektion von zisternal angezeigt. Bei Läsionen, die weiter kaudal zu vermuten sind, er-
folgt die Kontrastmittelgabe lumbal (GREVEL 1994).
Die Myelographie ermöglicht eine Differenzierung der Lokalisation von Veränderungen im
Bezug auf das Rückenmark. So können extradurale von intradural-extramedullären und intra-
medullären Läsionen unterschieden werden (GREVEL 1994).
Bei besonderen Fragestellungen wie z.B. der Verdacht auf eine lumbosakrale Schädigung
kann eine Epidurographie (FEENEY und WISE 1981, GREVEL et al. 1993, ROBERT und
SELCER 1993, GREVEL 1994) und/oder eine Diskographie (SISSON et al. 1992, GREVEL
et al. 1993, GREVEL 1994) notwendig werden.
In vielen Fällen können weitere bildgebende Verfahren wie die Magnetresonanztomographie
(DEHAAN et al. 1993) und Computertomographie die Diagnostik erleichtern.
2.8.3 Operations-Methoden
Aufgrund der Vielzahl von Operationstechniken an der Wirbelsäule bei den unterschied-
lichsten Indikationen sollen im Folgenden nur die Verfahren kurz angesprochen werden, bei
denen im Rahmen der vorliegenden Arbeit Biopsien entnommen worden sind.
Partielle Spondylektomie, Ventrale Dekompression, „Ventral Slot“
Darunter versteht man die Entlastung des Rückenmarks bei Diskusprolapsus oder Diskus-
hernien der Halswirbelsäule durch Herstellen einer Öffnung in den Wirbelkanal von ventral
mit Entfernung von Diskusmaterial (WHEELER und SHARP 1994).
Nach Durchtrennung der oberflächlichen Faszie, der Mm. sternohyoidei und Mm. sterno-
mastoidei wird die Trachea freigelegt. Rechts und unterhalb der Trachea stellt man sich, unter
Schonung des N. laryngeus recurrens, der A. carotis communis und des Truncus vagosym-
paticus den M. longus colli dar. Orientierungspunkte stellen das Tuberculum ventrale des

Atlas, die unterschiedlich ausgeprägten Tuberkel jedes Wirbels, sowie die prominenten
Processus transversus des 6. Halswirbels dar (SWAIM 1974, SEIM und PRATA 1982,
JURINA 1996, JURINA und GREVEL 2002b).
Die Anteile des M. longus colli werden von ihrem Ansatz am Tuberculum ventrale gelöst und
von dem ventralen Wirbelkörper verlagert. Die Zwischenwirbelräume liegen direkt kaudal der
Tubercula ventralia der Wirbel.
Nach Darstellung der entsprechenden Wirbelkörper und des Diskus fräst man eine ovale Rin-
ne bis eine Öffnung in den Wirbelkanal geschaffen ist. Die Öffnung darf maximal 40 % der
Wirbelkörperbreite und ein Drittel der Wirbelkörperlänge auf jeder Seite des Interver-
tebralspaltes betragen (SEIM und PRATA 1982, JURINA 1996).
Hemilaminektomie
Im thorakolumbalen Wirbelsäulenabschnitt wird bei Rückenmarkskompressionen die Hemi-
laminektomie durchgeführt.
In den vorwiegenden Fällen werden nach Durchtrennung des Lig. supraspinale und der Faszie
des M. longissimus dorsi die Processus spinosi mit den inserierenden Mm. multifidi darge-
stellt. Die Muskulatur wird dicht am Knochen abpräpariert und das Wirbelgelenk freigelegt.
Danach wird eine ovale Öffnung in die Lamina quer über den Intervertebralspalt bis in den
Wirbelkanal gefräst (HOLMBERG et al. 1990). Das prolabierte oder sequestrierte Bandschei-
benmaterial wird entfernt.
Dorsale Laminektomie
Die dorsale Laminektomie wird hauptsächlich zur chirurgischen Therapie der lumbosakralen
Stenose eingesetzt (CHAMBERS et al. 1988).
Nach Durchtrennung der Faszie des M. longissimus dorsi erreicht man die Processus spinosi
des 7. Lendenwirbels sowie des Os sacrum und die inserierenden Mm. multifidi. Die Pro-
cessus spinosi werden bis zum Wirbelkörper freipräpariert und eine annähernd rechteckige
Öffnung in den Wirbelkanal geschaffen. Diese nimmt etwa 2/3 der dorsalen Begrenzung des
7. Lendenwirbels ein und reicht über den Intervertebralspalt bis zum kranialen Anteil des 1.
Kreuzwirbels (CHAMBERS et al. 1988, WHEELER 1992).

Wirbeldistraktion und –fusion
Eine Indikation für diese Operation stellen dynamische Kompressionen des Rückenmarks im
kaudalen Halswirbelsäulenbereich dar.
Der Zugang zu den Wirbelkörpern entspricht dem Zugang der ventralen Dekompression. Der
Diskus wird, bis auf einen schmalen Rand der den Wirbelkanal verschlossen hält, zusammen
mit einem Teil der Wirbelkörper ausgefräst.
Die Distraktion kann mit Hilfe verschiedener Techniken erfolgen. Es können Steinmann-Pins
eingebracht werden, die in die Wirbel gedrillt und durch Knochenzement fixiert werden
(BRUECKNER et al. 1989) oder Knochenzement kann zwischen die distrahierten Wirbel-
körper nach der Methode von DIXON et al. (1996) eingebracht werden.
Eine weitere Möglichkeit stellt das Einlegen eines zuvor am Beckenkamm oder dem proxi-
malen Humerus entnommenen Knochenspans in die ausgefräste Rinne zwischen den beiden
Wirbeln nach Distraktion dar. Die anschließende Fixation der Wirbel erfolgt mit einer
Kaspar-Platte (VERA GREVEL, LEIPZIG, 1999, persönliche Mitteilung).

3 Tiere, Material und Methoden 3.1 Tiere 3.1.1 Klinische Daten der Patienten Insgesamt wurden 200 Biopsien von 50 Hunden, die zwischen Juni 1999 und April 2001 ope-
riert wurden, ausgewertet. Dabei handelt es sich um 35 männliche und 15 weibliche Tiere.
Die vertretenen Rassen wurden in chondrodystrophe (n = 33) und nichtchondrodystrophe
Tiere (n = 17) unterteilt. Sie sind in den nachfolgenden Abbildungen 2 und 3 graphisch dar-
gestellt.
0 1 2 3
DSH
Rottweiler
Dobermann
DSH-Mix
Großpudel
Wolfsspitz-Mix
Dalmatiner
BSH
Hannov. Schweißhd.
Ras
se
Anzahl
Abb.2: Rasse und Anzahl der nichtchondrodystrophen Hunde im Untersuchungsgut von 50 Hunden mit Rückenmarkskompressionen
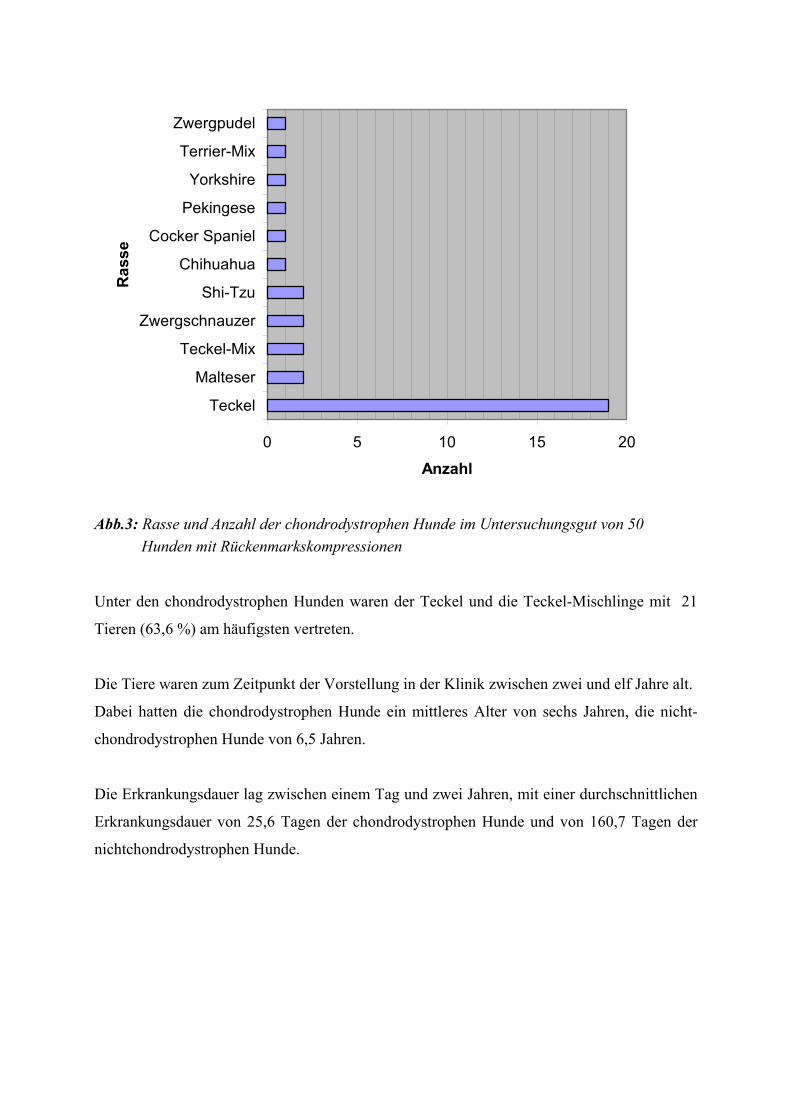
0 5 10 15 20
Teckel
Malteser
Teckel-Mix
Zwergschnauzer
Shi-Tzu
Chihuahua
Cocker Spaniel
Pekingese
Yorkshire
Terrier-Mix
ZwergpudelR
asse
Anzahl
Abb.3: Rasse und Anzahl der chondrodystrophen Hunde im Untersuchungsgut von 50 Hunden mit Rückenmarkskompressionen Unter den chondrodystrophen Hunden waren der Teckel und die Teckel-Mischlinge mit 21
Tieren (63,6 %) am häufigsten vertreten.
Die Tiere waren zum Zeitpunkt der Vorstellung in der Klinik zwischen zwei und elf Jahre alt.
Dabei hatten die chondrodystrophen Hunde ein mittleres Alter von sechs Jahren, die nicht-
chondrodystrophen Hunde von 6,5 Jahren.
Die Erkrankungsdauer lag zwischen einem Tag und zwei Jahren, mit einer durchschnittlichen
Erkrankungsdauer von 25,6 Tagen der chondrodystrophen Hunde und von 160,7 Tagen der
nichtchondrodystrophen Hunde.

3.1.2 Art der Rückenmarkskompression
Es handelte sich bei 60 % der Patienten (n = 30) um eine Rückenmarkskompression aufgrund
eines Diskusprolaps.
Bei sieben Tieren war eine Diskushernie Erkrankungsursache. In sechs Fällen konnte eine
Kaudale Zervikale Spondylomyelopathie mit Diskushernie diagnostiziert werden. Bei vier
Hunden lag eine Lumbosakrale Stenose, bei zwei Hunden eine Kompression der Spi-
nalnerven durch knöcherne Zubildungen am 6. bzw. 7. Halswirbel vor. Ein Hund war an ei-
nem intraduralen, extramedullären Tumor zwischen dem ersten und zweiten Halswirbel er-
krankt.
Eine Zusammenfassung ist in der folgenden Abbildung 4 dargestellt.
307
6
42 1
Diskusprolaps
Diskushernie
Kaudale ZervikaleSpondylomyelopathie
Lumbosakrale Stenose
Spinalnervenkompression
Tumor
Abb.4: Art der Rückenmarkskompression im Untersuchungsgut von 50 neurologisch
erkrankten Hunden
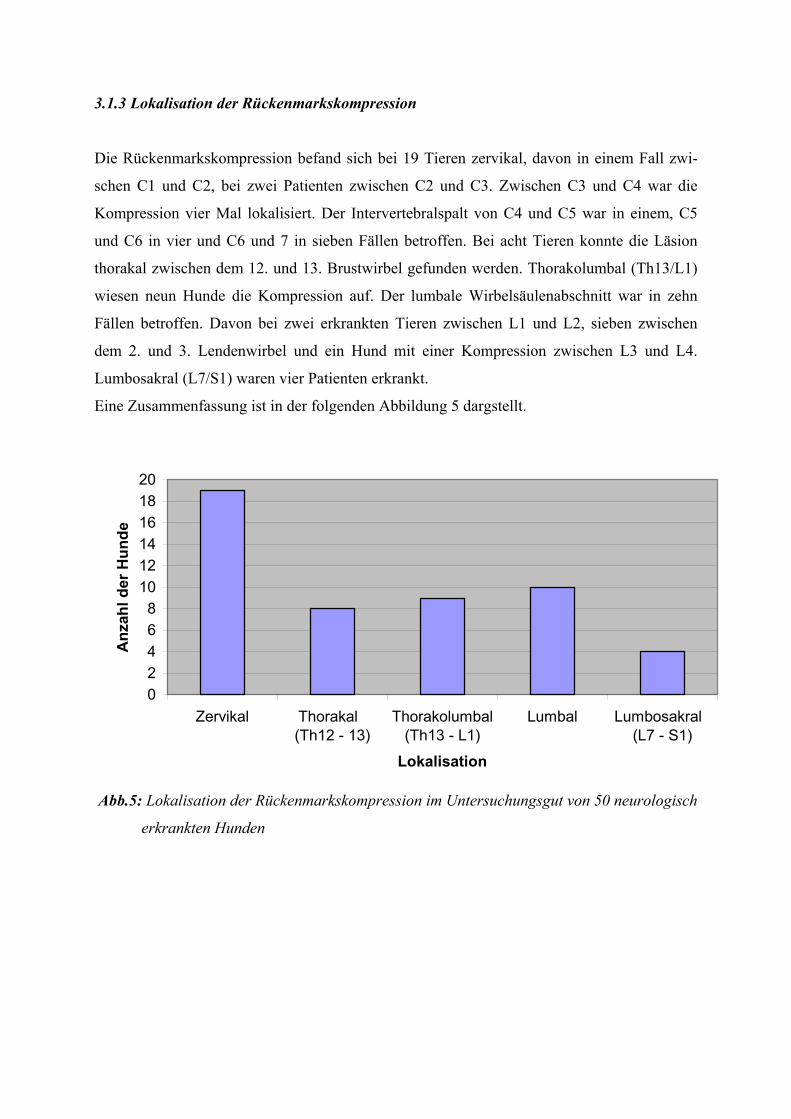
3.1.3 Lokalisation der Rückenmarkskompression
Die Rückenmarkskompression befand sich bei 19 Tieren zervikal, davon in einem Fall zwi-
schen C1 und C2, bei zwei Patienten zwischen C2 und C3. Zwischen C3 und C4 war die
Kompression vier Mal lokalisiert. Der Intervertebralspalt von C4 und C5 war in einem, C5
und C6 in vier und C6 und 7 in sieben Fällen betroffen. Bei acht Tieren konnte die Läsion
thorakal zwischen dem 12. und 13. Brustwirbel gefunden werden. Thorakolumbal (Th13/L1)
wiesen neun Hunde die Kompression auf. Der lumbale Wirbelsäulenabschnitt war in zehn
Fällen betroffen. Davon bei zwei erkrankten Tieren zwischen L1 und L2, sieben zwischen
dem 2. und 3. Lendenwirbel und ein Hund mit einer Kompression zwischen L3 und L4.
Lumbosakral (L7/S1) waren vier Patienten erkrankt.
Eine Zusammenfassung ist in der folgenden Abbildung 5 dargstellt.
02468
101214161820
Zervikal Thorakal (Th12 - 13)
Thorakolumbal(Th13 - L1)
Lumbal Lumbosakral (L7 - S1)
Lokalisation
Anz
ahl d
er H
unde
Abb.5: Lokalisation der Rückenmarkskompression im Untersuchungsgut von 50 neurologisch
erkrankten Hunden
Die Anamnese (A), die klinischen Symptome (S) sowie Diagnose und Lokalisation der
Kompression sind in Tabelle 3 zusammengefasst. Die Gliederung erfolgte nach der Dauer der
Symptome und der Diagnose.
Tab.3: Anamnese, klinische Symptome, Diagnose, Lokalisation von 50 untersuchten Hunden mit Rückenmarkskompression
Nr. Rasse m/w Alter Anamnese/Symptome Diagnose/ Lokalisation
1 RHT m 3J A: seit 1d Bewegungsstörung der HGM S: nichtgehfähige Paraparese
Diskusprolaps TH13/L1 ventral und rechts
2 LHT w 8J A: seit 1d Bewegungsstörung der HGM S: nichtgehfähige Paraparese
Diskusprolaps TH12/13 ventral und rechts
3 Malteser m 3J A: seit 1d Bewegungsstörung der HGM S: Paraplegie ohne Tiefenschmerz
Diskusprolaps Th13/L1 ventral
4 LHT w 5J A: seit 2d Bewegungsstörung der HGM S: nichtgehfähige Paraparese
Diskusprolaps TH12/13 ventral und rechts
5 LHT m 9J A: seit 2d Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps L2/3 ventral und links
6 RHT m 9J A: seit 2d Bewegungsstörung der HGM S: nichtgehfähige Paraparese
Diskusprolaps L2/3 ventral und rechts
7 LHT w 6J A: seit 2d Bewegungsstörung aller 4 Gliedmaßen S: gehfähige Tetraparese
Diskusprolaps C2/3
8 KHT m 7J A: seit 3d Dolenz Rücken, seit 1d Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps L3/4 ventral und links
9 RHT w 4J A: seit 3d Bewegungsstörung der HGM S: Paraplegie mit Tiefenschmerz
Diskusprolaps TH12/13 links
10 RHT w 6J A: seit 3d Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps Th12/13 ventral und rechts
11 KHT w 4J A: seit 3d Bewegungsstörung der HGM S: Paraplegie mit Tiefenschmerz
Diskusprolaps Th13/L1 ventral und links
12 LHT m 5J A: seit 4d Bewegungsstörung der li HGM S: Paraplegie mit Tiefenschmerz
Diskusprolaps TH13/ L1 ventral
13 RHT m 4J A: seit 4d Dolenz im Halsbereich S: HWS-Syndrom
Diskusprolaps C3/4
14 Pekingese m 6J A: seit 5d Dolenz Rücken S: nichtgehfähige Paraparese
Diskusprolaps L2/3 ventral
15 Shih - Tzu m 4J A: seit 1 Woche Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps L1/2 ventral
16 KHT w 4J A: seit 1 Woche Dolenz Rücken S: gehfähige Paraparese
Diskusprolaps Th12/13 ventral und links
17 KHT m 6J A: seit 2 Wochen Dolenz Hals S: gehfähige Paraparese
Diskusprolaps C6/7
18 RHT w 3J A: seit 2 Wochen Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps L2/3 links
19 Chihuahua m 4J A: seit 2 Wochen Dolenz Hals S: HWS-Syndrom
Diskusprolaps C3/4
20 RHT w 5J A: vor 3 Wochen Sturz, seit 2d Bewegungsstörung der HGM S: Paraplegie mit Tiefenschmerz
Diskusprolaps TH13/L1 ventral und rechts
21 Malteser m 3J A: seit 3 Wochen Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps L1/2 ventral und links
22 Wolfsspitz- Mix
m 11J A: seit 3 Wochen Dolenz Hals, Bewegungsstörung linke HGM S: HWS-Syndrom, Hemiparese links
Diskusprolaps C4/5
23 Yorkshire m 4J A: seit 6 Wochen Bewegungsstörung der HGM S: nichtgehfähige Paraparese
Diskusprolaps TH12/13 ventral
24 Teckel-Mix m 6J A: seit 8 Wochen Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps TH12/13 ventral
25 Shih - Tzu m 6J A: seit 8 Wochen dolent im Halsbereich S: HWS-Syndrom
Diskusprolaps C3/4
26 Zwerg-schnauzer
mk 10J A: seit 8 Wochen dolent im Rücken- und Halsbereich S: HWS-Syndrom
Diskusprolaps C3/4
27 LHT m 8J A: seit 8 Wochen Dolenz Rücken S: gehfähige Paraparese
Diskusprolaps TH12/13 ventral
28 KHT m 3J A: seit 3 Monaten Dolenz Rücken S: gehfähige Paraparese
Diskusprolaps TH 13/ L1 ventral
29 Teckel-Mix m 7J A: seit 6 Monaten Bewegungsstörung der HGM S: gehfähige Paraparese
Diskusprolaps L2/3 ventral und rechts
30 Hannov. Schweißhd.
m 10J A: seit 8 Monaten Bewegungsstörung aller 4 GM S: nichtgehfähige Tetraparese
Diskusprolaps C6/7
31 Cocker
Spaniel m 5J A: seit 2d nach Trauma Bewegungs-
störung der HGM S: gehfähige Paraparese
Diskushernie TH13/ L1
32 DSH-Mix m 6J A: seit 3d Dolenz bei Belastung S: gehfähige Paraparese
Diskushernie L2/3 rechts
33 DSH m 8J A: seit 5d Bewegungsstörung der HGM S: gehfähige Paraparese
Diskushernie TH13/ L1 links
34 LHT m 8J A: seit 1 Woche Bewegungsstörung der HGM S: nichtgehfähige Paraparese
Diskushernie L2/3
35 Zwergpudel mk 11J A: seit 3 Wochen Dolenz Hals S: HWS-Syndrom
Diskushernie C2/3
36 Dobermann w 7J A: seit 3 Wochen Bewegungsstörung aller 4 Gliedmaßen S: Tetraplegie, HWS-Syndrom
Diskushernie C5/6
37 DSH m 6J A: seit 6 Monaten Bewegungsstörung der HGM S: Ataxie HGM rechts>links
Diskushernie TH13/L1 rechts
38 Zwerg-
schnauzer m 7J A: seit 14d Dolenz Hals und steifer Gang
S: HWS-Syndrom zervikale Instabilität C5/6
39 Dobermann m 4J A: seit 1 Monat Bewegungsstörung der HGM, Dolenz im Halsbereich S: Ataxie HGM, HWS-Syndrom
zervikale Instabilität C6/7
40 Rottweiler m 4J A: seit 2 Monaten Bewegungsstörung der HGM S: Ataxie der HGM
zervikale Instabilität C6/7
41 Rottweiler m 7J A: seit 3 Monaten Bewegungsstörung der HGM S: gehfähige Paraparese
zervikale Instabilität C6/7
42 Dobermann w 5J A: seit 4 Monaten Bewegungsstörung der HGM S: Ataxie der HGM
zervikale Instabilität C6/7
43 BSH wk 2J A: seit 1 Jahr Bewegungsstörung der HGM S: Ataxie der HGM
zervikale Instabilität C5/6
44 Großpudel wk 6J A: seit 3 Monaten nach Trauma Dolenz bei
Belastung S: Dolenz lumbosakral
Lumbosakrale Stenose L7/ S1
45 Mix m 9J A: seit 6 Monaten Bewegungsstörung der HGM S: gehfähige Paraparese
Lumbosakrale Stenose L7/ S1
46 DSH m 9J A: seit 7 Monaten Schwanken in der Hinterhand, Dolenz Rücken S: Ataxie HGM, Dolenz lumbosakral
Lumbosakrale Stenose L7/ S1
47 Rottweiler w 7J A: seit 2Jahren rezidivierende LH linke HGM S: Dolenz lumbosakral
Lumbosakrale Stenose L7/ S1
48 Dalmatiner m 9J A: seit 6 Monaten progressive Lahmheit
rechte VGM S: HWS-Syndrom
Spinalnerven-kompression C6/7
49 Großpudel m 8J A: vor 1 Jahr Dolenz Hals, seit 6 Monaten angespannte Muskulatur und Dolenz Rücken S: HWS-Syndrom
Spinalnerven- kompression C5/6
50 Terrier-Mix m 9J A: vor 4 Monaten Sturz, seit 2 Monaten Bewegungsstörung der HGM S: nichtgehfähige Paraparese
extraduraler RM-Tumor C1/2
3.1.4 Auswahl und Entnahme der Muskelbiopsien Bei 50 Hunden wurden während einer Wirbelsäulenoperation Muskelbiopsien entnommen. Es
handelt sich dabei entweder um Proben des M. multifidus (n = 31), des M. longus colli (n =
16) oder des M. splenius (n = 3). Dafür wurden je ein Bioptat kaudal links und rechts des er-
krankten Wirbelsäulensegmentes sowie je ein Bioptat kranial sowohl links als auch rechts
ausgewählt, insgesamt vier Bioptate pro Hund.
Die Entnahme erfolgte zu Beginn der Operation, um größere Artefakte durch Instrumente und
Anästhesie zu vermeiden. Außerdem wurde versucht, möglichst entfernt von der Muskelfas-
zie zu bioptieren, um fasziennahe, myopathieähnliche Veränderungen bei der lichtmikros-
kopischen Untersuchung ausschließen zu können.
Die Verarbeitung wurde unmittelbar nach der Probenentnahme durchgeführt. Der Transport
der Bioptate erfolgte in einer mit isotoner Kochsalzlösung angefeuchteten Kompresse.
Die Muskelbiopsien wurden geteilt und der für die Elektronenmikroskopie vorgesehene An-
teil in Glutaraldehyd fixiert.

3.2 Methoden
3.2.1 Gefriertechnik der Proben für die Lichtmikroskopie
Der für die Lichtmikroskopie genutzte Biopsieanteil wurde mittels Gewebekleber (Kryotec®,
Boehringer Mannheim) auf Korkplättchen aufgebracht und in flüssigem Propan für etwa 20
Sekunden eingefroren. Für diesen Vorgang ist ein Stahlbehälter, mit einlegbarem Sieb für die
Biopsien, hergestellt worden. Dieser Behälter wurde in flüssigen Stickstoff verbracht. Über
eine Kupferspule, ebenfalls von Stickstoff umgeben, wurde Propangas in den Biopsiebehälter
eingeleitet. In das durch diesen Vorgang verflüssigte Propan wurden die Muskelbiopsien für
kurze Zeit eingelegt. Danach konnten die Proben bei ca. -196°C in flüssigem Stickstoff
gelagert werden (GISELA STOLTENBURG-DIDINGER, BERLIN, 1999, persönliche
Mitteilung).
Das kurze „Vorfrieren“ in Propan verhindert das Entstehen von Kristallen im Gewebe, die bei
sofortiger Lagerung im Stickstoff auftreten können und zu Artefakten bei der Verarbeitung
und Auswertung führen würden.
Die nachfolgend beschriebenen Färbemethoden der zu untersuchenden Muskelproben wurden
durch die Mitarbeiter des Labors im Institut für Neuropathologie des Universitätsklinikums
Benjamin Franklin in Berlin durchgeführt. Die Beschreibung und Anwendung der enzym-
histochemischen Färbemethoden beziehen sich auf die Ausführungen in den entsprechenden
Veröffentlichungen (DUBOWITZ 1967, ROMEIS 1968 und 1989, JERUSALEM und ZIERZ
1991).
Für die einzelnen Färbungen wurden an einem Kryostaten (Jung Frigocut 2800 E; Cambridge
Instruments GmbH, Nußloch, Deutschland) Gefrierschnitte mit einer Dicke von 8-20 µm an-
gefertigt. Die Präparate wurden auf unbeschichtete Objektträger aufgebracht.

3.2.2 Färbemethoden für die Lichtmikroskopie
Hämatoxylin-Eosin-Färbung
Zur Darstellung von myopathischen Veränderungen (zentrale Kerne, pathologische Kaliber-
variationen, Zunahme des Bindegewebes, Muskelfasernekrosen) eignet sich die Hämatoxylin-
Eosin-Färbung (HE-Färbung).
Das Hämatoxylin ist ein Farbstoff im Holz des in Südamerika vorkommenden Campeche-
baumes. Der wirksame Farbstoff ist eigentlich sein Oxidationsprodukt Hämatein, aber der
Name Hämatoxylin hat sich in der Nomenklatur durchgesetzt. Um diesen Farbstoff histo-
logisch zur Kernfärbung zu nutzen, wurden verschiedene Alaunsalze zugesetzt (Färbung nach
Rawitz). Damit sich das übrige Gewebe besser differenzieren lässt, wird anschließend eine
mäßige Überfärbung mit Eosin durchgeführt (BEHRENS 1908).
Mit der HE-Färbung stellen sich Muskelfasern rot, das Bindegewebe rosarot und die Kerne
der Muskelfasern, des interstitiellen Bindegewebes und der Gefäßendothelzellen dunkelblau
dar. In Querschnitten können die intermyofibrillären Räume als netzförmige Felderung er-
kennbar sein. Das Zytoplasma unreifer Zellen, insbesondere von Myoblasten, erscheint
schwach graublau (basophil).
1. Aqua dest.
2. Hämalaun nach Mayer (Fa. Merck 1.09249) für 4-5 Minuten
3. kurz in Aqua dest. Spülen
4. kurz in 1 %igem HCl-Alkohol differenzieren
5. in Leitungswasser für 5 min bläuen
6. kurz in Aqua dest. spülen
7. in Eosin (Fa. Merck 1345) bis 1 Minute eintauchen
8. aufsteigende Alkoholreihe mit 70, 80, 90 und 100 %igem Aethanol
9. in Xylol (Fa. Baker grade 8118) geben
10. mit Kryostalon (Fa. Harleko-Mounting, USA) eindecken

Eosin-Stammlösung: 1 %ig mit absolutem Alkohol
Eosin-Gebrauchslösung: Stammlösung 1:1 mit 70 %igem Alkohol
verdünnt und einige Tropfen
Eisessig (Fa. Merck 100056)
1 % HCl-Alkohol: ein Teil 3 %iger HCl-Alkohol in zwei
Teile absolutem Alkohol
Gomori-Trichom-Färbung
In der Gomori-Trichom-Färbung (ENGEL und CUNNINGHAM 1963) erscheinen die Mus-
kelfasern grünblau, das Bindegewebe hellgrün, die Zellkerne rotviolett und das sarkoplasma-
tische Retikulum sowie die Mitochondrien des intermyofibrillären Netzwerkes rot. Die Mark-
scheiden der Nervenfasern färben sich blassrosa.
Durch die Gomori-Trichom-Färbung werden Mitochondropathien mit „ragged red fibers“
deutlicher als in der HE-Färbung sichtbar.
Die Färbung wird folgendermaßen durchgeführt:
1. unfixierte Kryostatschnitte für 5 Minuten in frisch filtrierte Papanicolaou-
Lösung geben
2. Spülen mit Leitungswasser
3. 10 Minuten in Gomori-Lösung geben
4. Spülen in 0,2 %iger Essigsäure (Fa. Merck 100065)
5. Entwässerung in aufsteigender Alkoholreihe
6. in Xylol (Fa. Baker grade 8118) geben
7. mit Kryostalon (Fa. Harleko-Mounting, USA) eindecken

Zusammensetzung der Gomori-Lösung:
A. 0,6 g Chromotrop (=Acid red) 2 R (Fa. Chroma 1 B 259) in 100 ml Aqua
dest. lösen
B. 0,3 g Fast-Green (Fa. Serva FCF 21295) zusetzen
C. 0,8 g Wolframatphosphorsäurehydrat (Fa. Merck 583)
D. 1 ml 1 %igen Eisessig (Fa. Merck 100056) zugeben
E. Einstellung mit 1 N NaOH (Fa. Merck Titrisol 109956) auf einen pH von 3,4
Oil-Red-O-Sudan-IV-Färbung
Durch die Oil-Red-O-Färbung (LILLIE und ASHBURN 1943) und die Sudan-IV-Färbung
werden Neutralfette dargestellt. In den blassblauen Muskelfasern mit dunkelblauen
Kernen findet sich im Normalfall gar kein Fett oder nur sehr kleine Fettpartikel in den
Typ-I-Muskelfasern.
In der vorliegenden Arbeit ist für die Färbung der nicht-membrangebundenen Neutralfette
eine Lösung, die zu gleichen Anteilen aus Sudan-IV (Fa. Chroma 11565) und Oil-Red-O (Fa.
Fluka 75087) besteht, verwendet worden. Die Lipide sind kräftig orange und die Zellkerne
blau gefärbt.
1. Gefrierschnitte von 15-20 µm Dicke
2. Schnitte in Aqua dest. auffangen
3. kurz in 50 %igen Alkohol tauchen
4. mindestens 10 Minuten in Oil-Red-O-Sudan IV-Lösung färben
5. mit Aqua dest. spülen
6. für 5 Minuten in Hämalaun (Mayer) eintauchen (Kernfärbung)
7. mit Aqua dest. spülen
8. Bläuen in Leitungswasser und auf Objektträger aufziehen
9. Trocknen und mit Glyzeringelatine (Fa. Merck Kaiserglyzeringelatine) eindecken

Sudan-IV-Lösung:
1 g Pulver in 200 ml 70 %igem Alkohol lösen. Über Nacht im Brutschrank bei 50°C belassen
und anschließend filtrieren.
Oil-Red-O-Lösung:
Stammlösung: 0,5 g Oil-Red-O Pulver in 100 ml 99 %igem Isopropylalkohol lösen.
Danach 6 Teile der Stammlösung A mit 4 Teilen Aqua dest. mischen, 24 Stunden ruhen
lassen und anschließend filtrieren. Zum Abschluß beide Lösungen zu gleichen Teilen (1:1)
mischen.
Nicotinamid-Adenin-Dinukleotid-Tetrazolium-Reduktase (NADH-TR)
Die NAD-Hydrogenase gehört zur Gruppe der oxidativen Enzyme (FARBER et al. 1956).
Dieses in den Muskelfasern vorkommende Ferment entzieht dem im Inkubationsmedium be-
findlichen Substrat NADH den Wasserstoff und überträgt ihn auf ein wasserlösliches und
farbloses Tetrazoliumsalz, das durch diese Reduktion in unlösliches, blauschwarz sichtbares
Formazan überführt wird (FARBER et al. 1956).
Diese NADH-Reduktase-Reaktion eignet sich für die Beurteilung der Mitochondrienvertei-
lung und der Unterscheidung der Muskelfasertypen I und II. Muskelfaser-Typ-I erscheint we-
gen des hohen Mitochondriengehaltes dunkelblau, Muskelfaser-Typ-II hellblau. In der vor-
liegenden Arbeit wurde diese Methode zur Darstellung der Target-Fasern benutzt.
Es werden native Kryostatschnitte für 30 Minuten bei 37°C in folgender Lösung inkubiert:
Phosphat-Puffer 10 ml
Nitroblau-Tetrazoliumchlorid (NBT) (Fa. Serva 30515) 10 mg
Nicotinamidadenindinukleotid (NADH) (Fa. Serva 30312) 8 mg
Anschließend werden die Gefrierschnitte mit Aqua dest. gespült und mit Glyzeringelatine (Fa.
Merck Kaiserglyzeringelatine) eindeckt.

Herstellung des Phosphatpuffers:
Stammlösung A: 177,99 Mol Natriumdihydrogenphosphat (NaH2PO4)
(Fa. Merck 6346) entspricht 3,5598 g Na2HPO4 auf
100 ml Aqua dest.
Stammlösung B : 137,99 Mol Dinatriumhydrogenphosphat (Na2HPO4)
(Fa. Merck 6580) entspricht 2,75 g Na2HPO4auf
100 ml Aqua dest.
Von Lösung A werden 81 ml entnommen, dann langsam mit Lösung B auf 100 ml aufgefüllt
bis ein pH von 7,2 – 7,4 erreicht ist. Danach 100 ml Aqua dest. zugeben.
Cytochrom C Oxidase-Reaktion (COX)
DAB-Methode nach Seeligmann
DAB-Lösung (-20°C) 9 ml
Katalaselösung 1 ml
Cytochrom C 10 mg
Succrose 750 mg
DAB-Lösung auftauen, die weiteren Substanzen nacheinander zugeben und lösen.
Durchführung:
1. Inkubationslösung 24 Stunden bei 22°C
2. kurz in Aqua dest. spülen
3. Dehydrieren mit Xylol und Balsam
Myofibrilläre ATPase
Die myofibrilläre ATPase-Reaktion dient in erster Linie der Darstellung und Unterscheidung
verschiedener Muskelfasertypen. Den Schnitten werden ATP und Calcium bei einem pH von
9,4 zugeführt, worauf die myofibrilläre ATPase des Muskelgewebes vom ATP ein Phosphat

abspaltet, das sich mit Calcium zu Calciumphosphat verbindet. Letzteres ist bei alkalischem
pH unlöslich, also am Ort der Enzymaktivität lokalisiert. Mittels Kobaltchlorid und Ammo-
niumsulfid wird Calciumphosphat in Kobaltphosphat und schließlich in das schwarzbraun er-
scheinende, unlösliche Kobaltsulfid umgewandelt. Die myofibrilläre ATPase-Aktivität er-
weist sich bei einem pH von 9,4 im Inkubationsmedium in den Typ-II-Fasern als sehr hoch,
daher erscheinen diese braunschwarz, während die Typ-I-Faser keine oder nur sehr geringe
Enzymaktivität zeigen. Deshalb bleiben die Typ-I-Muskelfasern blass.
Bei einem pH von 4,3 erweisen sich die Reaktionsintensitäten als invers.
Eine feinere Abstufung der pH-Werte ermöglicht eine Differenzierung in weitere Subtypen.
3.2.3 Elektronenmikroskopie
Die elektronenmikroskopische Bearbeitung der untersuchten Muskelbioptate wurde durch die
Mitarbeiterinnen für Elektronenmikroskopie der Abteilung für Neuropathologie, Universitäts-
klinik Benjamin Franklin, durchgeführt.
Für die Untersuchung wird jeweils ein Teil jeder Muskelprobe in Glutaraldehyd fixiert und in
Epoxidharz (Araldit) eingebettet.
Fixation und Einbettung:
1. Fixieren in 2,5 %iger Glutaraldehydlösung für mindestens 12 Stunden
2. Zerlegen der Muskelprobe in 8 längsorientierte, ca. 4 mm lange
Muskelstückchen
3. Spülung in Cacodylatpuffer für eine Stunde
4. Nachfixieren in Osmiumtetroxid 1 % über 12-24 Stunden
5. Spülung in Cacodylatpuffer
6. Entwässerung in Aceton mit aufsteigender Konzentration; bei 70 %igem Aceton,
zusätzlich Beigabe von 0,5%igem Uranylacetat und 1%iger
Phosphorwolframsäure
7. Aralditstammlösung und 2 % Beschleuniger für 4-5 Stunden
8. Einkapselung in Araldit und 2 % Beschleuniger. Bei der Auswahl der Kapseln wird die
Hälfte der Proben für Längsschnitte, die andere Hälfte für Querschnitte orientiert.
9. Brutschrank bei 60°C für 2 Tage

Herstellung von Glutaraldehyd:
2 % Glutaraldehyd wird in Cacodylatpuffer (Konzentration (c) = 0,1 mol/l) angesetzt.
Substanzen:
500 ml Cacodylatpuffer (c = 0,2 mol/l; Cacodylsäure Natrium Trihydrat,
Fa. Roth 5169.2)
420 ml Aqua dest.
80 ml Glutaraldehyd (25 %ige Lösung in Wasser, Fa. Serva 23115)
Durchführung:
Cacodylatpuffer ansetzen (21,403 g Cacodylatsäure ad 500 ml Aqua dest.,
ergibt c = 0,2 mol/l)
420 ml Aqua dest. unter ständigem Rühren zugeben
80 ml Glutaraldehyd unter ständigem Rühren hinzufügen
Normaldruckfiltration durch Aktivkohle (1 Löffel Aktivkohle in
Filterpapier geben und anschließend die Lösung filtrieren)
den pH-Wert auf pH von 7,2 bis 7,4 einstellen
Herstellung des Cacodylatpuffers:
Cacodylatpuffer wird in einer Konzentration von 0,2 mol/l benutzt.
Substanzen:
42,80 g Cacodylsäure (Cacodylsäure Natrium Trihydrat, Fa. Roth 5169.2)
1000 ml Aqua dest.
Durchführung :
Cacodylsäure mit Präzisionswaage einwiegen
Aqua dest. zugeben und unter ständigem Rühren auflösen

Für die Anfertigung der Semidünnschnitte werden die Araldit-Blöcke pyramidenförmig zu-
geschnitten („getrimmt“), anschließend werden am Ultramikrotom mit einem Glasmesser
0,35 mm dicke Schnitte hergestellt, mit Richardson-Lösung (RICHARDSON et al. 1960)
gefärbt, auf Objektträger gegeben und eingedeckt. Anhand dieser Semidünnschnitte werden
die Areale für die Ultradünnschnitte ausgewählt.
Zusammensetzung der Richardson-Lösung:
A. 1 %ige Perjodsäure-Lösung (1 g auf 100 ml Aqua bidest.)
B. 1 %ige Azur II Lösung (1 g auf 100 ml Aqua bidest.)
C. A und B 1:1 mischen = Mallory’s Azur
D. 1 %ige Methylenblau Lösung mit Na-Tetraborat (1 g Methylenblau auf 100 ml
Aqua bidest., 4 g Na-Tetraborat hinzufügen)
E. C und D mischen und filtrieren
Am Ultramikrotom werden Ultradünnschnitte (70 nm) mit einem Diamantmesser angefertigt,
auf mit Folie beschichteten (100 Maschen = mesh) oder unbeschichteten (200 mesh) Netzen
aufgebracht und mit gebrauchsfertiger Bleizitratlösung im Kontrastierungsautomaten (Leica)
bearbeitet.
Die Elektronenmikroskopie wird an einem Zeiss Elektronenmikroskop (Typ EM 10) durch-
geführt und die relevanten Areale bei unterschiedlichen Vergrößerungen von 1: 1700-fach bis
1: 80000-fach fotografiert.
3.2.4 Morphometrie
Die histometrische Untersuchung wurde computergestützt durchgeführt. Durch die computer-
gesteuerte Morphometrie können die unterschiedlichen Muskelfasertypen erfasst und archi-
viert werden (BRUCHER et al. 1986). An je 200 Fasern pro Biopsie sind in der ATPase-
Färbung die Faserdurchmesser, getrennt nach Typ-I- und Typ-II-Fasern, bestimmt worden.
Die Morphometrie-Apparatur besteht aus einem Mikroskop, Videokamera, Monitor und dazu-
gehörigem Computer. Das Mikroskop wird vor jeder Messung identisch eingestellt. Der zu
untersuchende Abschnitt wird auf den Monitor projiziert und dort vermessen.

Das Vermessen erfolgt mit der „Zwei-Punkt-Technik“. Ermittelt wurde dabei der kleinste
Durchmesser nach DUBOWITZ und BROOKE (1973) bzw. JERUSALEM und ZIERZ
(1991). Dies ergibt auch bei schräg getroffenen Fasern die genauesten Werte. Dennoch wurde
darauf geachtet, dass die Messung in einem Bereich mit möglichst korrekt getroffenen Fasern
erfolgte. Anschließend wurden, durch die im Computer aufgenommenen Messwerte, für beide
Fasertypen Mittelwerte und der Variationskoeffizient berechnet.
Auf die Berechnung von in der Humanpathologie üblichen Atrophie- und Hypertrophie-
faktoren wurde wegen des Fehlens von Vergleichsdaten gesunder Tiere in der Literatur und
zu kleiner eigener Datenmenge verzichtet. Auf die Statistik wurde ebenfalls aus den oben
genannten Gründen verzichtet.

4 Ergebnisse
Insgesamt wurden bei 50 Hunden je vier Muskelbiopsien ausgewertet. Jede dieser Biopsien
wurde in fünf Färbemethoden untersucht. Konnten lichtmikroskopisch abnorme Befunde er-
hoben werden, schloss sich eine elektronenmikroskopische und in ausgewählten Fällen eine
morphometrische Untersuchung an.
4.1 Morphologische Veränderungen
Bei drei Tieren konnten keine pathologischen Veränderungen der Muskulatur festgestellt wer-
den. Sieben Hunde wiesen ein neurogenes Gewebssyndrom auf. Zwölf Patienten zeigten eine
myopathisch veränderte Muskelstruktur. Bei 28 Tieren war ein neurogenes Gewebssyndrom
mit einer Begleitmyopathie zu finden.
Eine Übersicht ist Abbildung 6 dargestellt.
Es wurden zunächst die Biopsien der betroffenen Seite sowohl kranial als auch kaudal der Lä-
sion, und anschließend die Biopsien der gegenüberliegenden Seite, kranial und kaudal, unter-
sucht. Da sich keine lokalen Unterschiede der pathologischen Befunde an den verschiedenen
Muskelabschnitten (kranial, kaudal, links und rechts der Läsion) darstellten, wird im Fol-
genden bei Beschreibung der Befunde nicht extra erwähnt an welcher Lokalisation die Biop-
sie entnommen wurde.
37
1228
normale Muskulatur
neurogenesGewebssyndrom
myopathischesGewebssyndrom
neurogen/ Begleitmyopathie
Abb. 6: histopathologische Befunde in den Paravertebralmuskelbiopsien von 50 Hunden mit
Rückenmarkskompressionen

In Abbildung 7 ist das elektronenmikroskopische Bild der Skelettmuskulatur eines Hundes
dargestellt der histologisch keine pathologischen Veränderungen aufwies.
Es handelt sich um ein längsorientiertes Präparat, d.h. die Muskelfasern (Mf) sind längs
angeschnitten und man kann deutlich die gestreifte Muskulatur erkennen. Dazwischen liegt
der Z-Streifen (Z) welcher sich markant dunkler abhebt.
Zwischen den Muskelbündeln befindet sich Fett- und Bindegewebe (F) das sich im
Elektronenmikroskop weiß darstellt. Darin eingelagert sind die dunkel erscheinenden
Mitochondrien (M).
Abb.7: normale Skelettmuskulatur;EM; x4400,LHT (Nr.5), m, 9J, Diskusprolaps;
Erkrankungsdauer: 2 Tage,Muskelfaser (Mf), Z-Streifen (Z), Fett- und Bindegewebe
(F), Mitochondrien (M)
Z
F
M
Mf
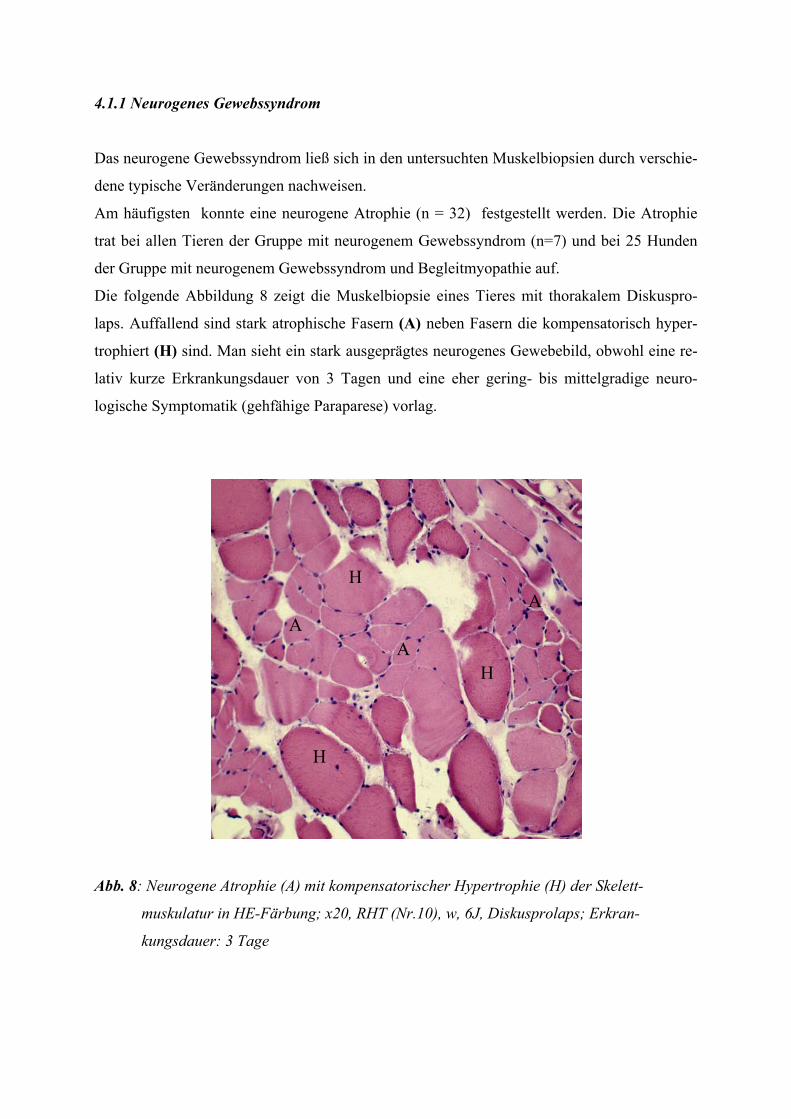
4.1.1 Neurogenes Gewebssyndrom
Das neurogene Gewebssyndrom ließ sich in den untersuchten Muskelbiopsien durch verschie-
dene typische Veränderungen nachweisen.
Am häufigsten konnte eine neurogene Atrophie (n = 32) festgestellt werden. Die Atrophie
trat bei allen Tieren der Gruppe mit neurogenem Gewebssyndrom (n=7) und bei 25 Hunden
der Gruppe mit neurogenem Gewebssyndrom und Begleitmyopathie auf.
Die folgende Abbildung 8 zeigt die Muskelbiopsie eines Tieres mit thorakalem Diskuspro-
laps. Auffallend sind stark atrophische Fasern (A) neben Fasern die kompensatorisch hyper-
trophiert (H) sind. Man sieht ein stark ausgeprägtes neurogenes Gewebebild, obwohl eine re-
lativ kurze Erkrankungsdauer von 3 Tagen und eine eher gering- bis mittelgradige neuro-
logische Symptomatik (gehfähige Paraparese) vorlag.
Abb. 8: Neurogene Atrophie (A) mit kompensatorischer Hypertrophie (H) der Skelett-
muskulatur in HE-Färbung; x20, RHT (Nr.10), w, 6J, Diskusprolaps; Erkran-
kungsdauer: 3 Tage
H
H
H
A A
A

In Abbildung 9 ist eine Fasertypengruppierung dargestellt. Das heißt, es fehlt die regelmäßi-
ge, schachbrettartige Verteilung der beiden Fasertypen. Je nach Motoneuron welches das er-
krankte Gebiet reinnerviert, treten vermehrt Typ-I- oder Typ-II-Fasern im Gesichtsfeld auf.
Hier handelt es sich um eine Gruppierung von Typ-I-Fasern, die braun angefärbt sind, dazwi-
schen einzelne Typ-II-Fasern (blassrosa). Ein typischer Befund nach einer Reinnervation, bei
einem schon länger bestehenden Krankheitsbild.
Abb. 9: Fasertypengruppierung in der Skelettmuskulatur mit ATPase-Reaktion pH 4,2;
x10; Yorkshire (Nr.23), m, 4J, Diskusprolaps; Erkrankungsdauer: 6 Wochen
Weiterhin konnten als Zeichen eines neurogenen Gewebssyndroms Target-Fasern (n = 4) und
„rimmed vacuoles“ (n = 5) festgestellt werden. Die in Abbildung 10 dargestellten Target-Fa-
sern treten hauptsächlich in den Typ-I-Muskelfasern auf.

Die normalerweise vollständig blau angefärbte Faser zeigt im folgenden Bild 10 zentrale Aus-
fälle des intermyofibrillären Netzwerkes. Diese sind als unregelmäßige, weiße Fläche in der
Faser zu erkennen.
Target-Fasern sind ein Hinweis auf stattgefundene De- und Reinnervation. Aus diesem Grund
sind sie erst bei einem längeren Krankheitsgeschehen zu finden. Drei der vier Patienten (Tier
Nr. 42, 47, 48) hatten eine Vorgeschichte ihrer Krankheit, die mindestens 4 Monate zurück-
lag. Bei einem Hund (Nr. 13, RHT, m, 4J, Diskusprolaps C3/4) war vom Besitzer nur eine
neurologische Auffälligkeit seit vier Tagen beschrieben.
Abb. 10: Target-Fasern in der Skelettmuskulatur in NADH-Färbung; x40; Dalmatiner
(Nr.48), m, 9J, Spinalnervenkompression; Erkrankungsdauer: 6 Monate
In Abbildung 11 sind „rimmed vacuoles“ dargestellt. Sie treten, wie auch in der Darstellung,
häufiger subsarkolemmal als zentral auf. Nachgewiesen wurde dieser Befund bei den Hunden
Nr. 2, 14, 22, 23 und 40.
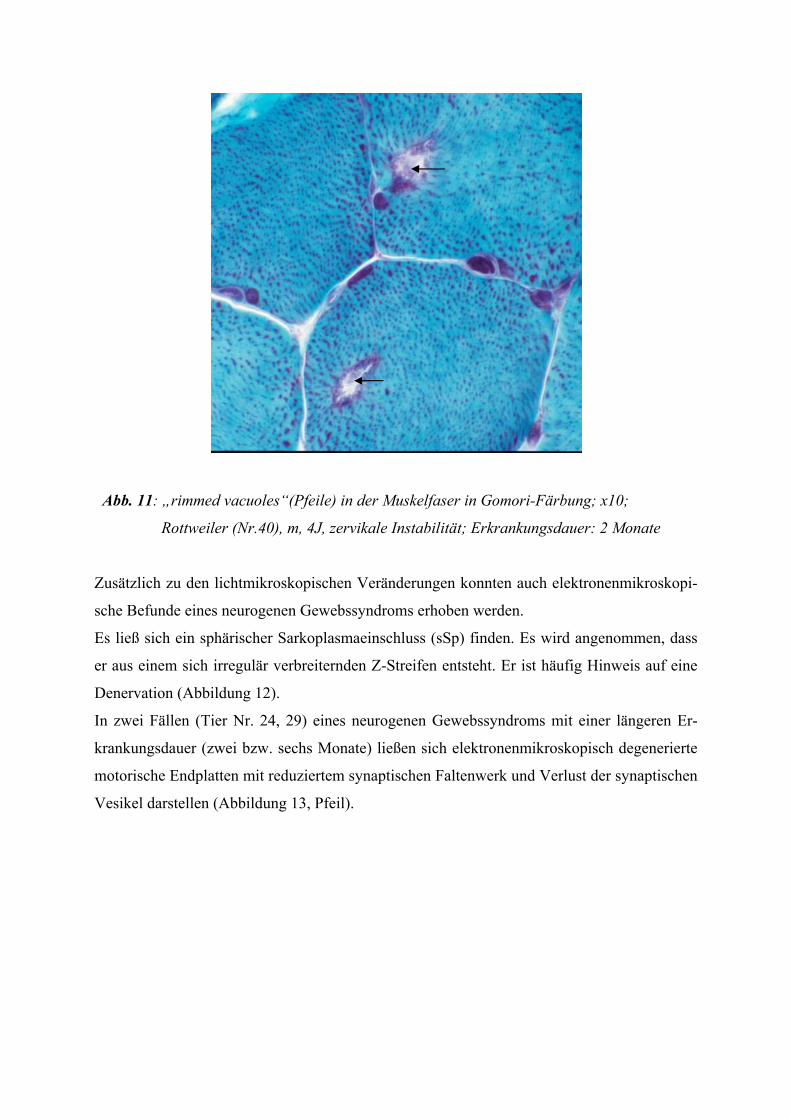
Abb. 11: „rimmed vacuoles“(Pfeile) in der Muskelfaser in Gomori-Färbung; x10;
Rottweiler (Nr.40), m, 4J, zervikale Instabilität; Erkrankungsdauer: 2 Monate
Zusätzlich zu den lichtmikroskopischen Veränderungen konnten auch elektronenmikroskopi-
sche Befunde eines neurogenen Gewebssyndroms erhoben werden.
Es ließ sich ein sphärischer Sarkoplasmaeinschluss (sSp) finden. Es wird angenommen, dass
er aus einem sich irregulär verbreiternden Z-Streifen entsteht. Er ist häufig Hinweis auf eine
Denervation (Abbildung 12).
In zwei Fällen (Tier Nr. 24, 29) eines neurogenen Gewebssyndroms mit einer längeren Er-
krankungsdauer (zwei bzw. sechs Monate) ließen sich elektronenmikroskopisch degenerierte
motorische Endplatten mit reduziertem synaptischen Faltenwerk und Verlust der synaptischen
Vesikel darstellen (Abbildung 13, Pfeil).
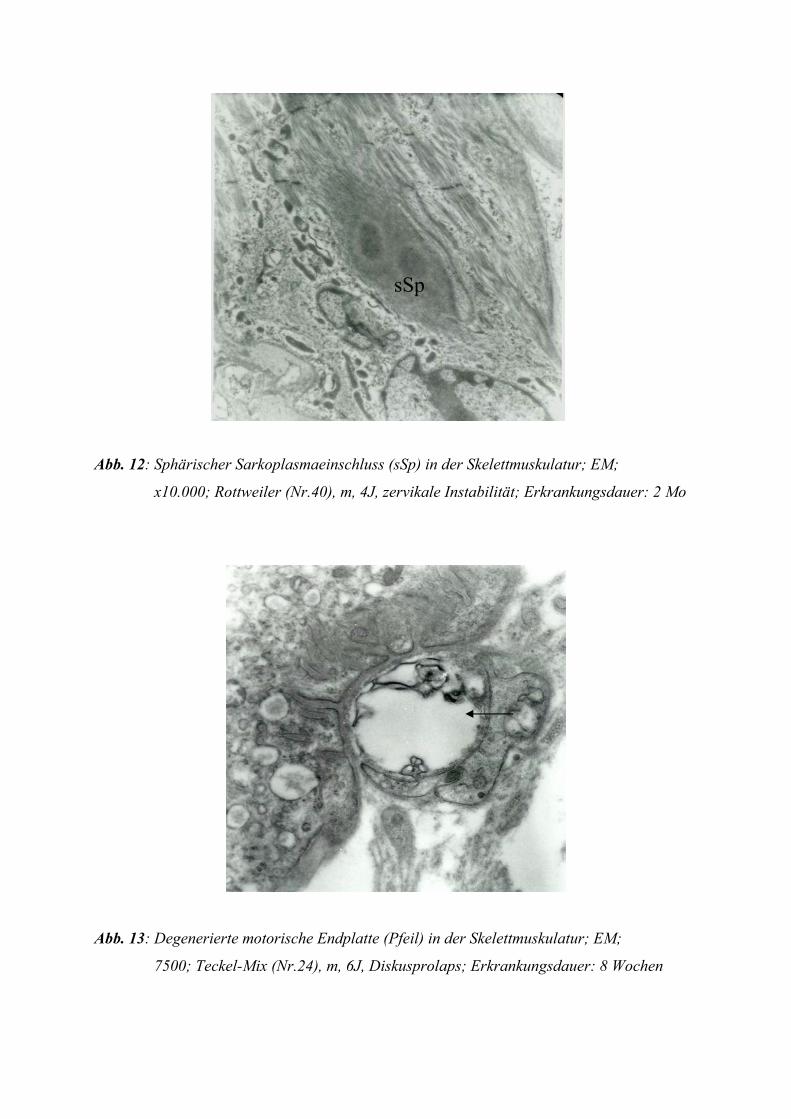
Abb. 12: Sphärischer Sarkoplasmaeinschluss (sSp) in der Skelettmuskulatur; EM;
x10.000; Rottweiler (Nr.40), m, 4J, zervikale Instabilität; Erkrankungsdauer: 2 Mo
Abb. 13: Degenerierte motorische Endplatte (Pfeil) in der Skelettmuskulatur; EM;
7500; Teckel-Mix (Nr.24), m, 6J, Diskusprolaps; Erkrankungsdauer: 8 Wochen
sSp

Zum Vergleich ist in Abbildung 14 die unveränderte motorische Endplatte eines akut erkrank-
ten Teckels dargestellt. Die Endplatte (Pfeil) ist in ihrer Struktur erhalten. Deutlich sind das
synaptische Faltenwerk und die Vesikel zu erkennen.
Abb. 14: Unveränderte motorische Endplatte (Pfeil) in der Skelettmuskulatur; EM; x10.000;
LHT (Nr.2), w, 8J, Diskusprolaps; Erkrankungsdauer: 1Tag
Die lichtmikroskopisch schon nachgewiesenen „rimmed vacuoles“ konnten bei den entspre-
chenden Tieren auch elektronenmikroskopisch aufgezeigt werden. Sie sind in Abbildung 15
gekennzeichnet (rv). Auffallend ist die typische Strukturierung durch den erhaltenen Vakuo-
leninhalt, der beim Gefrierschnitt verloren geht.

Abb. 15: „rimmed vacuoles“(rv) in der Skelettmuskulatur; EM; x20.000; Wolfsspitz-Mix
(Nr.22), m, 11J, Diskusprolaps; Erkrankungsdauer: 3 Wochen
Folgende für ein neurogenes Gewebssyndrom typischen Veränderungen wurden nachgewie-
sen (Tab.4).
Tab.4: Zusammenfassung der Befunde des neurogenen Gewebssyndroms in der Skelettmus-
kulatur von 50 untersuchten Hunden mit Rückenmarkskompressionen
Art der Veränderung Anzahl (n)
Neurogene Atrophie 32
Fasertypengruppierung 5
Target-Fasern 4
„rimmed vacoules“ 5
Sphärischer Sarkoplasmaeinschluß 1
Degenerierte motorische Endplatte 2
rv rv
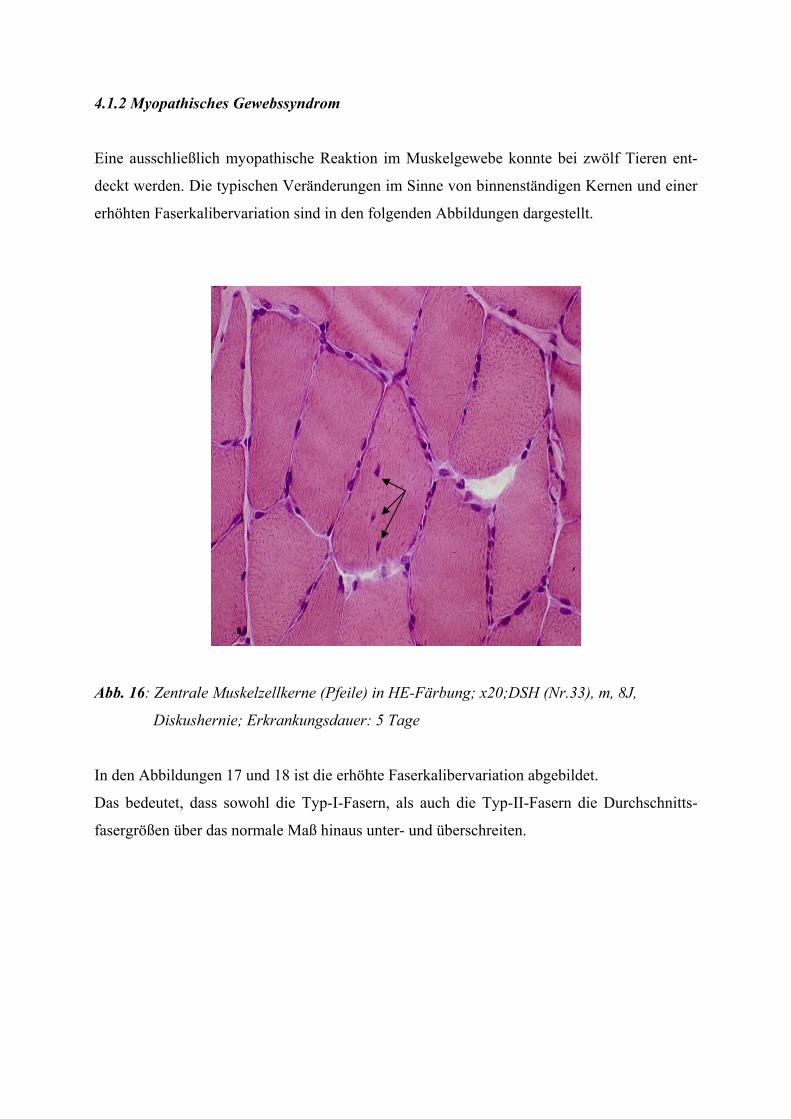
4.1.2 Myopathisches Gewebssyndrom Eine ausschließlich myopathische Reaktion im Muskelgewebe konnte bei zwölf Tieren ent-
deckt werden. Die typischen Veränderungen im Sinne von binnenständigen Kernen und einer
erhöhten Faserkalibervariation sind in den folgenden Abbildungen dargestellt.
Abb. 16: Zentrale Muskelzellkerne (Pfeile) in HE-Färbung; x20;DSH (Nr.33), m, 8J,
Diskushernie; Erkrankungsdauer: 5 Tage
In den Abbildungen 17 und 18 ist die erhöhte Faserkalibervariation abgebildet.
Das bedeutet, dass sowohl die Typ-I-Fasern, als auch die Typ-II-Fasern die Durchschnitts-
fasergrößen über das normale Maß hinaus unter- und überschreiten.
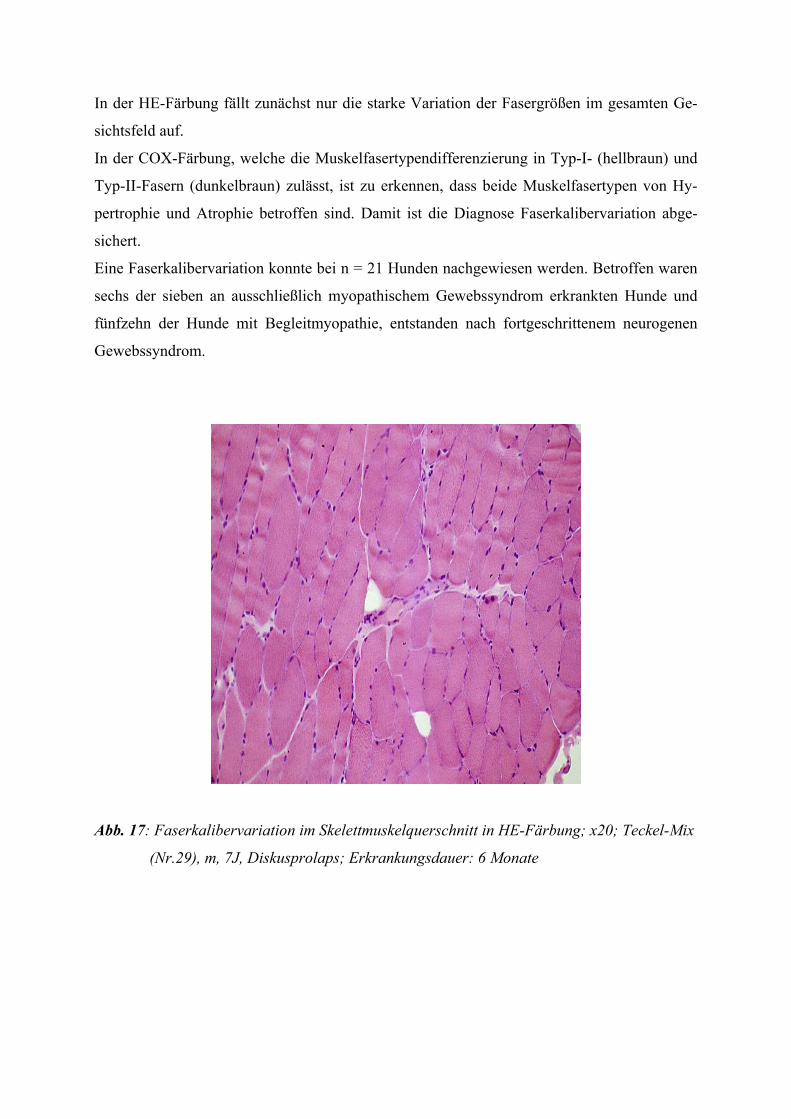
In der HE-Färbung fällt zunächst nur die starke Variation der Fasergrößen im gesamten Ge-
sichtsfeld auf.
In der COX-Färbung, welche die Muskelfasertypendifferenzierung in Typ-I- (hellbraun) und
Typ-II-Fasern (dunkelbraun) zulässt, ist zu erkennen, dass beide Muskelfasertypen von Hy-
pertrophie und Atrophie betroffen sind. Damit ist die Diagnose Faserkalibervariation abge-
sichert.
Eine Faserkalibervariation konnte bei n = 21 Hunden nachgewiesen werden. Betroffen waren
sechs der sieben an ausschließlich myopathischem Gewebssyndrom erkrankten Hunde und
fünfzehn der Hunde mit Begleitmyopathie, entstanden nach fortgeschrittenem neurogenen
Gewebssyndrom.
Abb. 17: Faserkalibervariation im Skelettmuskelquerschnitt in HE-Färbung; x20; Teckel-Mix
(Nr.29), m, 7J, Diskusprolaps; Erkrankungsdauer: 6 Monate
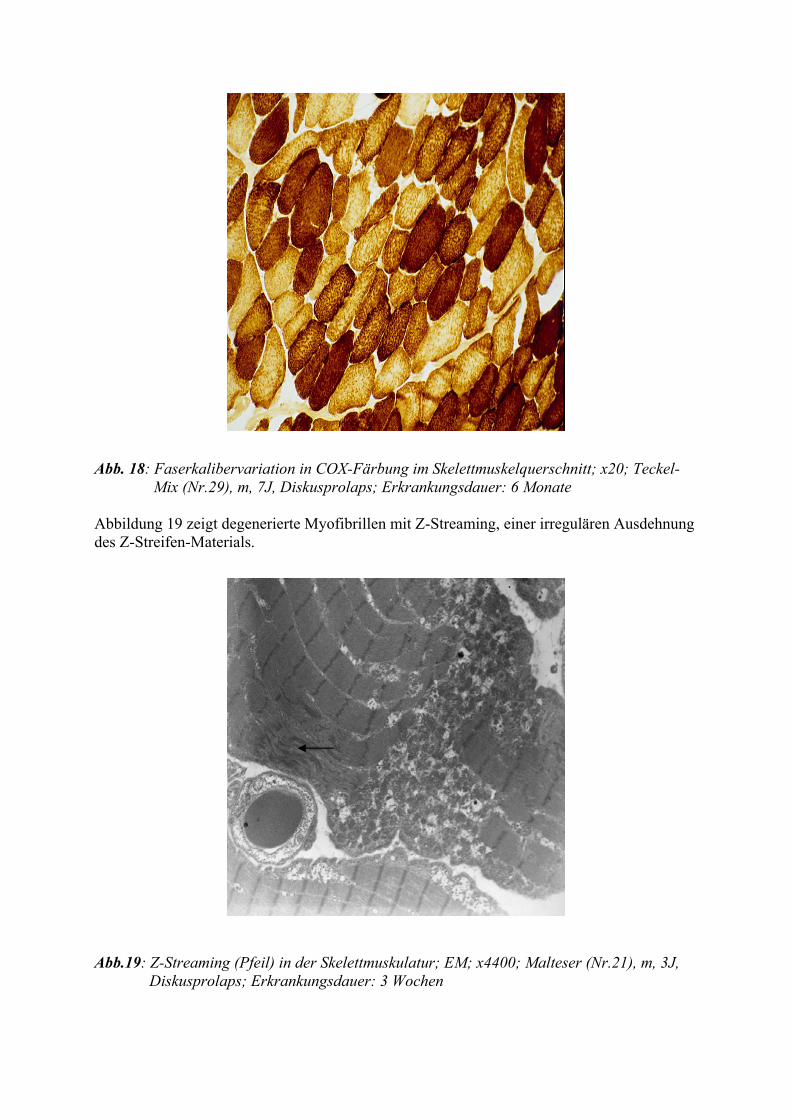
Abb. 18: Faserkalibervariation in COX-Färbung im Skelettmuskelquerschnitt; x20; Teckel- Mix (Nr.29), m, 7J, Diskusprolaps; Erkrankungsdauer: 6 Monate
Abbildung 19 zeigt degenerierte Myofibrillen mit Z-Streaming, einer irregulären Ausdehnung des Z-Streifen-Materials.
Abb.19: Z-Streaming (Pfeil) in der Skelettmuskulatur; EM; x4400; Malteser (Nr.21), m, 3J, Diskusprolaps; Erkrankungsdauer: 3 Wochen

In Abbildung 20 ist eine Zusammenlagerung des Z-Streifen-Materials zu „rods“ (r) darge-
stellt. Diese auch als „nemaline-bodies“ bezeichneten Strukturen waren bei drei Hunden (Tier
41, 49, 50) zu finden.
Abb. 20: „rods“(r) in der Skelettmuskulatur; EM; x7000; Rottweiler (Nr.41), m, 7J, zervikale
Instabilität; Erkrankungsdauer: 3 Monate
r
r r

Mitochondrienveränderungen sind bei 22 Patienten aufgetreten. Dabei waren fünf Tiere der
Gruppe mit rein myopathischen Veränderungen betroffen und außerdem 17 Tiere der Gruppe
die an ausgeprägten neurogenen Veränderungen mit dabei entstehender Begleitmyopathie
litten.
Alle diese Tiere zeigten eine Mitochondrienvermehrung. Mitochondrienvermehrungen fanden
sich vor allem subsarkolemmal. Bei zwei Patienten war die Vermehrung so stark ausgeprägt,
dass „ragged red fibres“, also eine Mitochondrienanhäufung in der gesamten Muskelfaser,
gefunden wurden.
Die folgenden Abbildungen 21-23 zeigen verschiedene Formen von Mitochondrienvermeh-
rungen in der Gomori-Färbung.
Abb. 21: Subsarkolemmale Mitochondrienvermehrung (Pfeile)in den Muskelfasern in
Gomori-Färbung; x20; Malteser (Nr.21), m, 3J, Diskusprolaps; Erkrankungsdauer:
3 Wochen
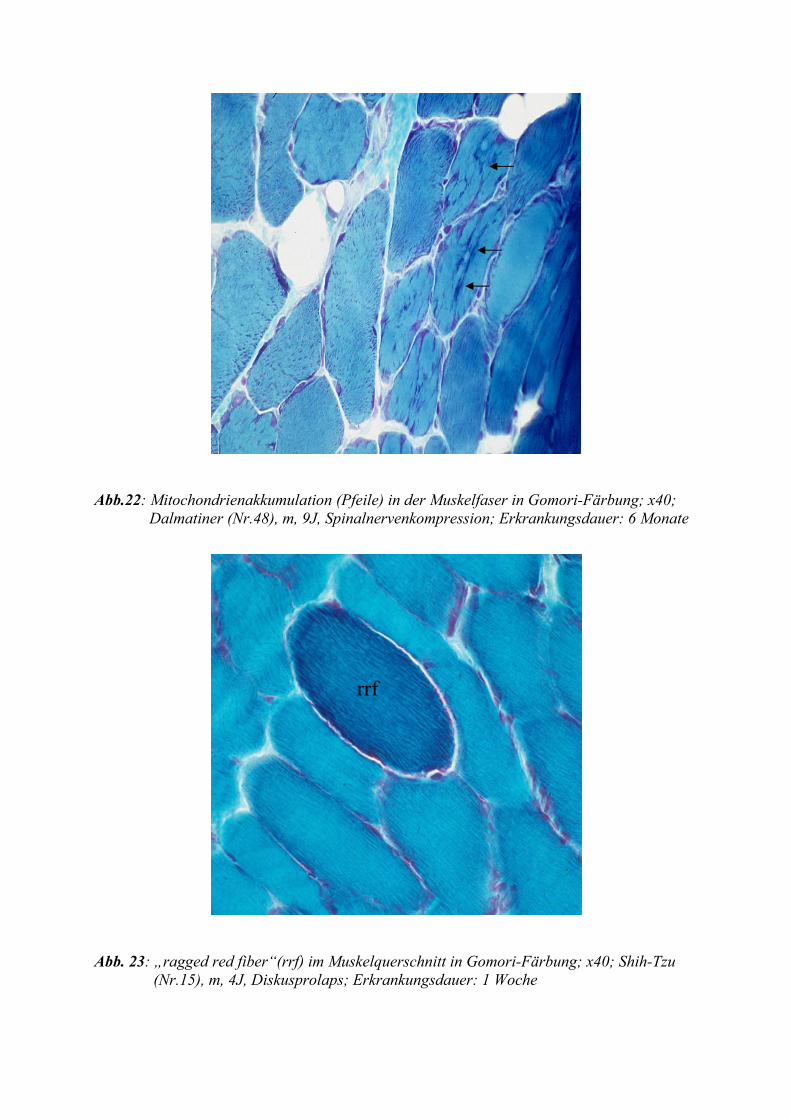
Abb.22: Mitochondrienakkumulation (Pfeile) in der Muskelfaser in Gomori-Färbung; x40; Dalmatiner (Nr.48), m, 9J, Spinalnervenkompression; Erkrankungsdauer: 6 Monate
Abb. 23: „ragged red fiber“(rrf) im Muskelquerschnitt in Gomori-Färbung; x40; Shih-Tzu (Nr.15), m, 4J, Diskusprolaps; Erkrankungsdauer: 1 Woche
rrf

Die Mitochondriopathien wurden weiterhin besonders deutlich durch die Darstellung mittels
Elektronenmikroskopie.
Die Mitochondrien stellten sich hauptsächlich mit allgemeiner Vermehrung (Abbildung 24),
in zwei Fällen auch mit Formvariationen und Cristae-Anomalien dar.
Bemerkenswert ist, dass diese beiden Tiere (Abbildung 25 und 26) auch die längste Erkran-
kungsdauer (6 Monate) in der Gruppe der Hunde mit metabolischen Myopathien aufwiesen.
Abb. 24: Mitochondrienvermehrung (M) in der Muskelfaser; x4400; Malteser (Nr.21), m, 3J,
Diskusprolaps; Erkrankungsdauer: 3 Wochen
M
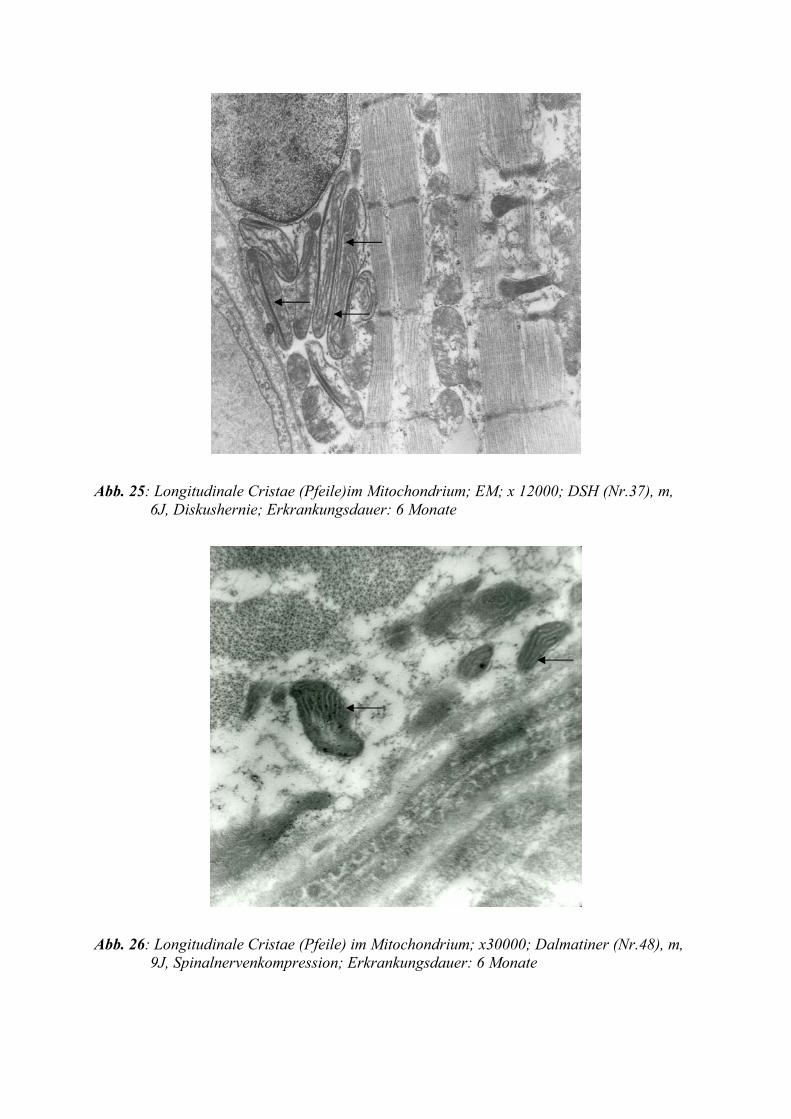
Abb. 25: Longitudinale Cristae (Pfeile)im Mitochondrium; EM; x 12000; DSH (Nr.37), m, 6J, Diskushernie; Erkrankungsdauer: 6 Monate
Abb. 26: Longitudinale Cristae (Pfeile) im Mitochondrium; x30000; Dalmatiner (Nr.48), m, 9J, Spinalnervenkompression; Erkrankungsdauer: 6 Monate
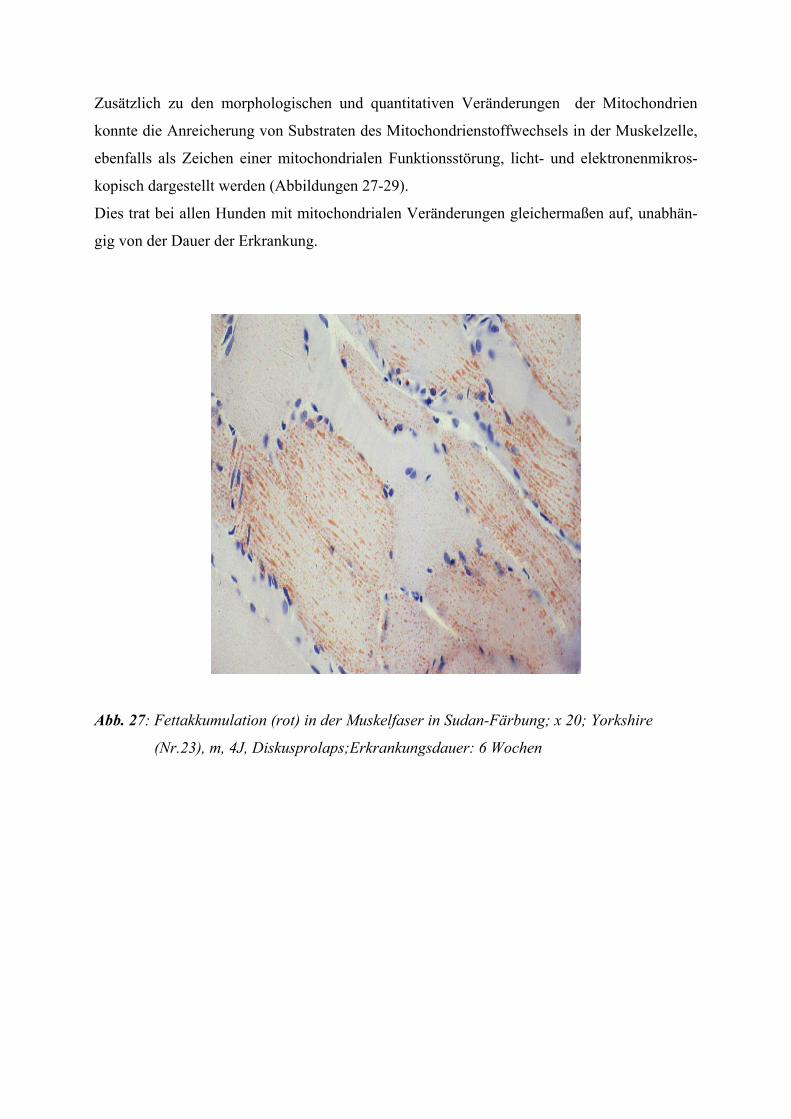
Zusätzlich zu den morphologischen und quantitativen Veränderungen der Mitochondrien
konnte die Anreicherung von Substraten des Mitochondrienstoffwechsels in der Muskelzelle,
ebenfalls als Zeichen einer mitochondrialen Funktionsstörung, licht- und elektronenmikros-
kopisch dargestellt werden (Abbildungen 27-29).
Dies trat bei allen Hunden mit mitochondrialen Veränderungen gleichermaßen auf, unabhän-
gig von der Dauer der Erkrankung.
Abb. 27: Fettakkumulation (rot) in der Muskelfaser in Sudan-Färbung; x 20; Yorkshire
(Nr.23), m, 4J, Diskusprolaps;Erkrankungsdauer: 6 Wochen
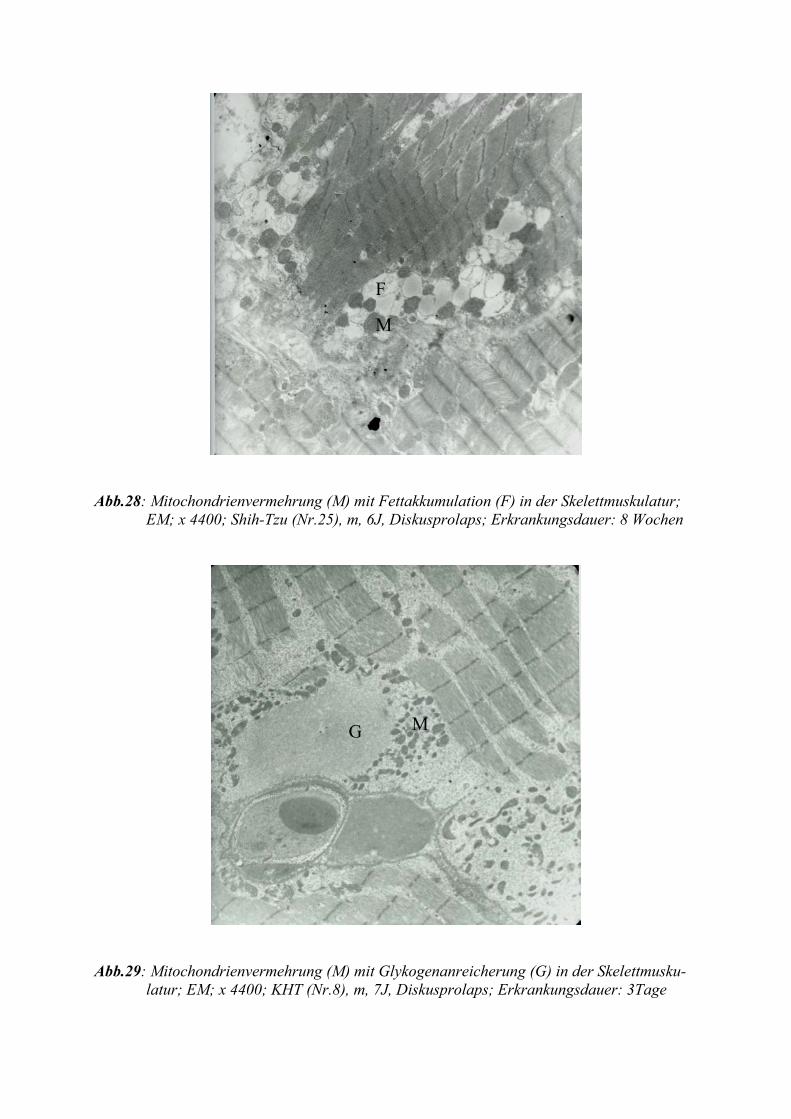
Abb.28: Mitochondrienvermehrung (M) mit Fettakkumulation (F) in der Skelettmuskulatur; EM; x 4400; Shih-Tzu (Nr.25), m, 6J, Diskusprolaps; Erkrankungsdauer: 8 Wochen
Abb.29: Mitochondrienvermehrung (M) mit Glykogenanreicherung (G) in der Skelettmusku- latur; EM; x 4400; KHT (Nr.8), m, 7J, Diskusprolaps; Erkrankungsdauer: 3Tage
G M
F
M
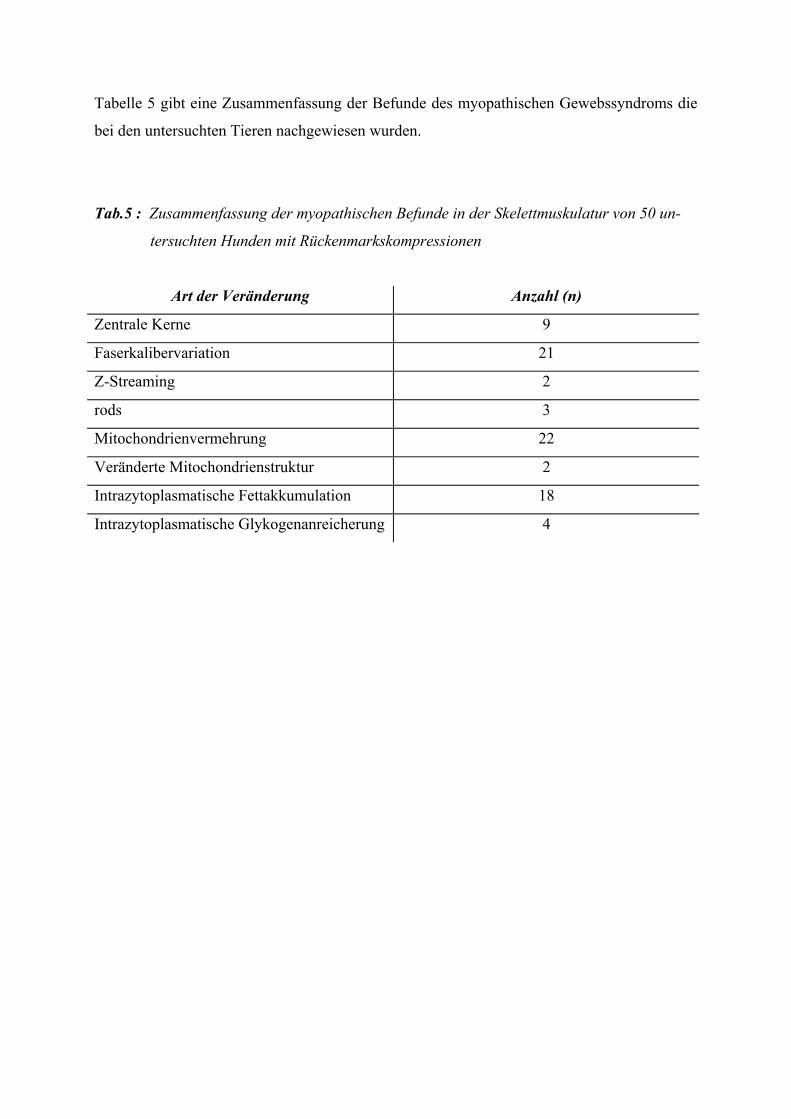
Tabelle 5 gibt eine Zusammenfassung der Befunde des myopathischen Gewebssyndroms die
bei den untersuchten Tieren nachgewiesen wurden.
Tab.5 : Zusammenfassung der myopathischen Befunde in der Skelettmuskulatur von 50 un-
tersuchten Hunden mit Rückenmarkskompressionen
Art der Veränderung Anzahl (n)
Zentrale Kerne 9
Faserkalibervariation 21
Z-Streaming 2
rods 3
Mitochondrienvermehrung 22
Veränderte Mitochondrienstruktur 2
Intrazytoplasmatische Fettakkumulation 18
Intrazytoplasmatische Glykogenanreicherung 4

4.1.3 Lipofuszin in der Skelettmuskulatur
Einen weiteren interessanten Befund stellte das bei vier jungen Tieren (Hunde Nr.1, 13, 16,
18) nachgewiesene, eisenfreie Alterspigment Lipofuszin dar.
Es reichert sich in allen postmitotischen Zellen an, erscheint elektronenmikroskopisch
schwarz, in unregelmäßiger Form und Gestalt und variiert mit seinem Durchmesser von ei-
nem bis zu mehreren Mikrometern. Lipofuszin liegt als Restlysosom vor.
Alle vier Hunde hatten nur eine relativ kurze Krankheitsanamnese von einem, vier, sieben
bzw. vierzehn Tagen.
In Abbildung 30 ist der Nachweis von Lipofuszin in der Muskelbiopsie eines drei Jahre alten
Rauhhaarteckels dargestellt.
Abb. 30: Lipofuszinanreicherung (Pfeil) in der Skelettmuskulatur; EM; x 12000; RHT (Nr.1),
m, 3J, Diskusprolaps; Erkrankungsdauer: 1 Tag
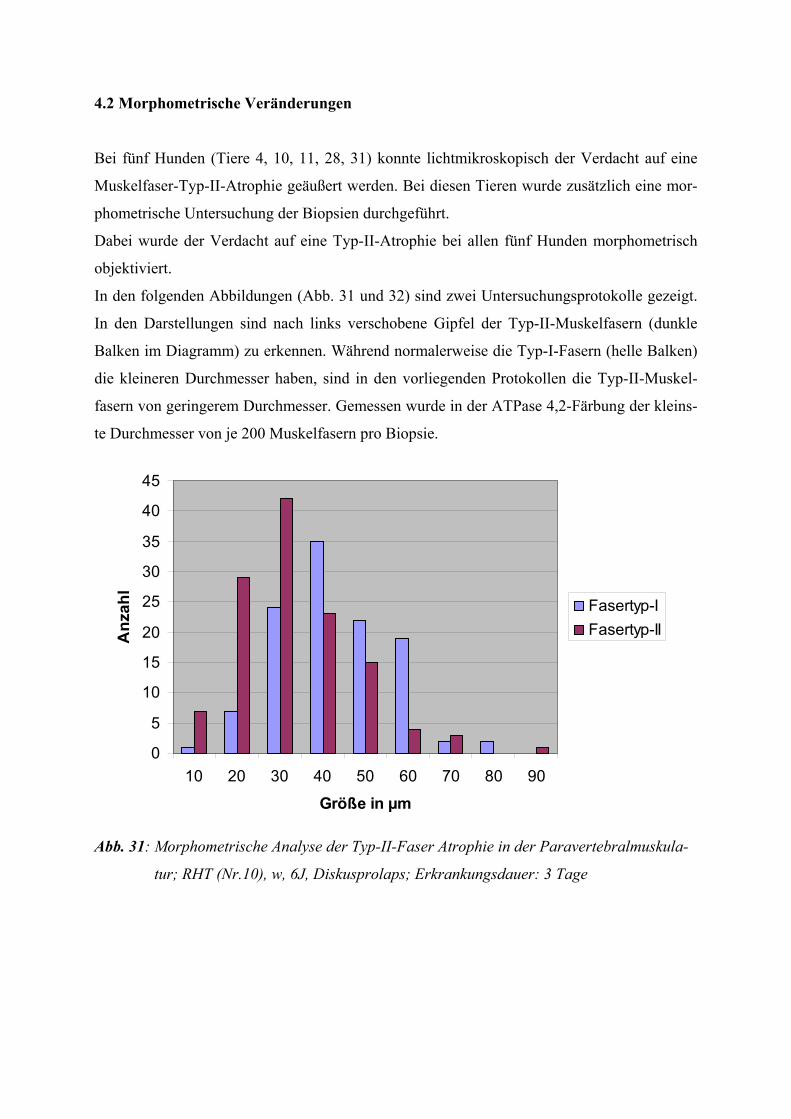
4.2 Morphometrische Veränderungen
Bei fünf Hunden (Tiere 4, 10, 11, 28, 31) konnte lichtmikroskopisch der Verdacht auf eine
Muskelfaser-Typ-II-Atrophie geäußert werden. Bei diesen Tieren wurde zusätzlich eine mor-
phometrische Untersuchung der Biopsien durchgeführt.
Dabei wurde der Verdacht auf eine Typ-II-Atrophie bei allen fünf Hunden morphometrisch
objektiviert.
In den folgenden Abbildungen (Abb. 31 und 32) sind zwei Untersuchungsprotokolle gezeigt.
In den Darstellungen sind nach links verschobene Gipfel der Typ-II-Muskelfasern (dunkle
Balken im Diagramm) zu erkennen. Während normalerweise die Typ-I-Fasern (helle Balken)
die kleineren Durchmesser haben, sind in den vorliegenden Protokollen die Typ-II-Muskel-
fasern von geringerem Durchmesser. Gemessen wurde in der ATPase 4,2-Färbung der kleins-
te Durchmesser von je 200 Muskelfasern pro Biopsie.
Abb. 31: Morphometrische Analyse der Typ-II-Faser Atrophie in der Paravertebralmuskula-
tur; RHT (Nr.10), w, 6J, Diskusprolaps; Erkrankungsdauer: 3 Tage
0
5
10
15
20
25
30
35
40
45
10 20 30 40 50 60 70 80 90
Größe in µm
Anz
ahl
Fasertyp-IFasertyp-II
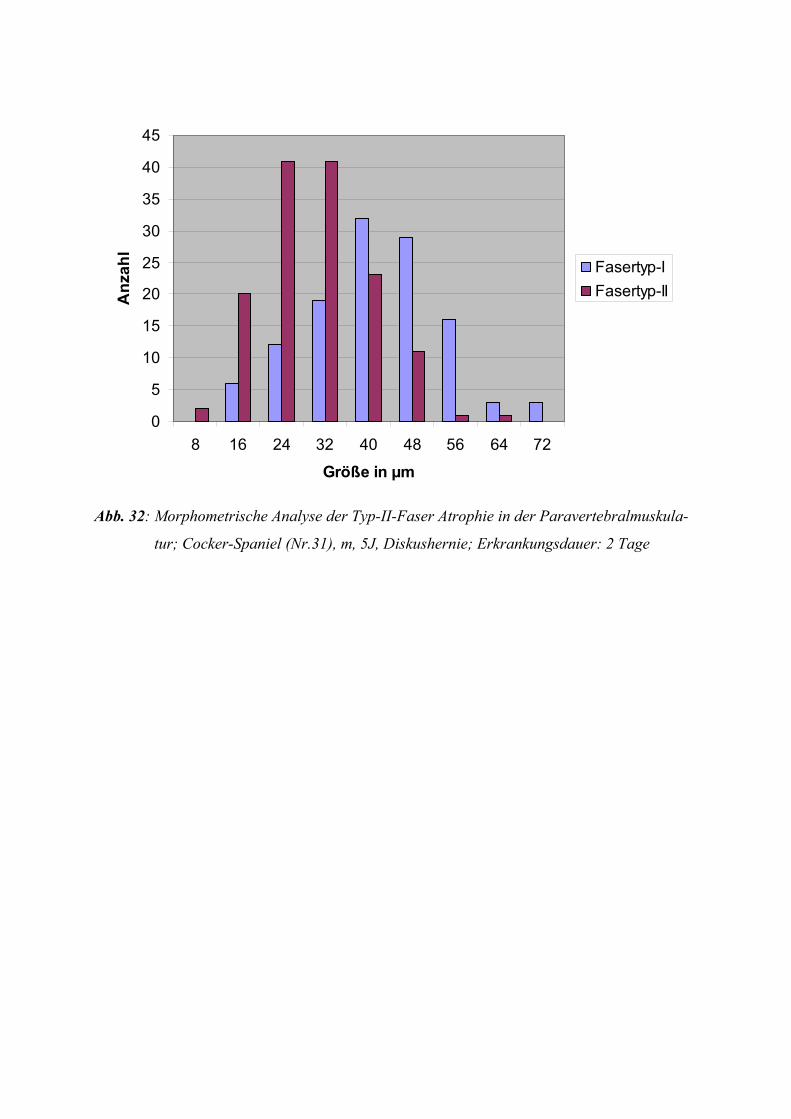
Abb. 32: Morphometrische Analyse der Typ-II-Faser Atrophie in der Paravertebralmuskula-
tur; Cocker-Spaniel (Nr.31), m, 5J, Diskushernie; Erkrankungsdauer: 2 Tage
0
5
10
15
20
25
30
35
40
45
8 16 24 32 40 48 56 64 72
Größe in µm
Anz
ahl
Fasertyp-IFasertyp-II

5 Diskussion
Es sollte untersucht werden, ob durch Bandscheibenvorfälle und dadurch verursachte Rücken-
markskompressionen bzw. Nervenwurzelirritationen eine Schädigung der Muskulatur hervor-
gerufen wird, die sich histologisch nachweisen lässt.
Die Fragestellung ist neu und auch beim Menschen bisher nur unvollständig bearbeitet.
Morphologische Untersuchungen eines größeren Kollektivs fehlen.
Die bisher in der Literatur dargestellten Ergebnisse von Muskeluntersuchungen bei Hunden
beziehen sich fast ausschließlich auf morphometrische Analysen von gesunder und trainierter
Skelettmuskulatur. Dabei wurden zum größten Teil andere Methoden der Muskelkonser-
vierung genutzt. Die Ergebnisse dieser Arbeit sind deshalb nur eingeschränkt mit den bisheri-
gen Resultaten anderer Autoren zu vergleichen.
Da man häufig bei den Patienten mit thorakolumbalen Veränderungen durch die klinische Un-
tersuchung und bildgebende Diagnostik eine Seitenbetonung der neurologischen Störungen
feststellen kann, wurde jeweils der Muskelabschnitt links und rechts der Wirbelsäule sowie
oberhalb und unterhalb der Läsion untersucht. Dies sollte Vergleiche und gezieltere Aussagen
ermöglichen.
Das Erstellen einer Kontrollgruppe erwies sich als nicht durchführbar, da die Entnahme von
Muskelbiopsien an gesunden Hunden nicht zu rechtfertigen ist. Die Auswertung von Muskel-
biopsien von Hunden, die aufgrund einer anderen Erkrankung euthanasiert werden mussten,
war nicht sinnvoll. Es handelte sich meist um onkologische oder endokrinologische Erkrank-
ungen, bei denen man von einer Beeinträchtigung der Skelettmuskulatur aufgrund der Krank-
heit ausgehen muss (DUNCAN et al. 1977, BRAUND et al. 1980, CARDINET 1984,
SHELTON und CARDINET 1987). Auch sehr alte Hunde, die euthanasiert wurden, stellten
keine Datengrundlage für eine Kontrollgruppe dar, da auch hier mit muskulären Umbaureak-
tionen zu rechnen ist (BLOOM 1968, TOMLINSON et al. 1969, REBEIZ et al. 1972).
Diese Arbeit soll eine Grundlage zur weiteren Forschung auf dem Gebiet der Myopathologie
des Hundes liefern. Das Anlegen einer Datenbank, die auch biochemische Untersuchungen
und molekulare Genetik einschließt, wäre nützlich und sinnvoll, um die Ursachenforschung
und Rehabilitation bei Hunden mit degenerativen Wirbelsäulenveränderungen weiter zu
führen.

Patientengut
In der Studie waren insgesamt 20 verschiedene Rassen vertreten. Der größte Teil waren
Teckel und Teckel-Mischlinge (63,6 %). Die Gruppe der chondrodystrophen Rassen war mit
33 Tieren deutlich zahlreicher vertreten als die nichtchondrodystrophen Rassen mit 17 Tieren.
Prinzipiell kann jede Hunderasse von einer degenerativen Bandscheibenveränderung be-
troffen sein. Die Beobachtungen anderer Autoren decken sich allerdings in Hinblick auf das
gehäufte Vorkommen bei chondrodystrophen Rassen bzw. Rassen mit chondrodystrophen
Merkmalen. So fand HANSEN (1952) bei seinen Untersuchungen öfter Hunde chondrodys-
tropher Rassen, namentlich Teckel, Pekinesen und Französische Bulldoggen, von einem
Bandscheibenvorfall betroffen. Auch VAUGHAN (1958) hatte in seinem Patientengut 52 %
Teckel. Für Teckel fanden GOGGIN et al. (1970) ein 12,6-fach höheres Risiko für einen
Bandscheibenvorfall. GAGE (1975) stellte ein vermehrtes Vorkommen von Teckeln mit
70,1 % fest.
In dieser Untersuchung überwiegen die männlichen Patienten mit 70 % gegenüber den weib-
lichen mit 30 %. Im Vergleich mit anderen Arbeiten scheint dies zufällig begründet zu sein.
VAUGHAN (1958), GAGE (1975), HOERLEIN (1978), FRY et al. (1991) und TOOMBS
und BAUER (1993) fanden bei ihren Untersuchungen ein annähernd ausgeglichenes Verhält-
nis zwischen männlichen und weiblichen Tieren. Eine Erklärung für eine etwaige Ge-
schlechtsdisposition kann aufgrund der Untersuchungen dieser Arbeit nicht gegeben werden.
Eine Beeinflussung des Patientengutes durch regionale Gegebenheiten, Gewohnheiten der
Hundebesitzer oder Haltungsbedingungen spielen hier eine Rolle.
Die Altersverteilung der in dieser Arbeit untersuchten Hunde deckt sich mit den Ergebnissen
anderer Autoren. Das jüngste Tier war zwei Jahre, das älteste elf Jahre alt. Dabei hatten die
chondrodystrophen Hunde ein mittleres Alter von sechs Jahren, die nicht-chondrodystrophen
Hunde von 6,5 Jahren.
RUSSEL und GRIFFITHS (1968) berichten bei ihren Patienten einen Altersdurchschnitt von
5,9 Jahren. Bei der Studie von GAGE (1975) war die Altersgruppe der fünfjährigen Hunde
besonders stark vertreten. Auch HOERLEIN (1978) berichtet von 73,1 % von 2395 Hunden
mit Diskopathie, die zwischen drei und sechs Jahren alt sind.
Insgesamt handelt es sich aber um ein sehr heterogenes Patientengut, welches nur bedingt
endgültige Aussagen über bestimmte Krankheitserscheinungen zulässt. Eine Unterteilung der

Tiere in Gruppen entsprechend des Gewichtes erschien aufgrund der geringen Anzahl nicht
sinnvoll.
Es wäre erstrebenswert, in weiteren Untersuchungen Hunde mit etwa gleichem Gewicht und
von der gleichen Rasse zu beurteilen, um aussagekräftige Daten vor allem in Bezug auf Faser-
größen der Rückenmuskulatur zu erhalten.
Strukturelle Veränderungen
Bei 47 der 50 untersuchten Tiere konnten pathologische Veränderungen an der Rückenmus-
kulatur nachgewiesen werden. Davon bei sieben Hunden ein neurogenes, bei 12 Hunden ein
myopathisches und bei weiteren 28 Hunden ein neurogenes Gewebssyndrom mit Begleit-
myopathie.
Bei vier Hunden (Tier 1, 13, 16, 18) wurde Lipofuszin beobachtet. Lipofuszin ist als Zellalte-
rungspigment bzw. Abnutzungspigment bekannt. Es akkumuliert bei Alterungsprozessen im
normalen Muskel (PRINCE 1963). Da es sich bei den betroffenen Hunden um drei- und vier-
jährige Teckel handelt, ist es nicht wahrscheinlich, dass als alleinige Ursache das Alter der
Tiere in Frage kommt.
Man kann vermuten, dass es durch die übermäßige Beanspruchung der Rückenmuskulatur auf
Grund chronischer Fehlbelastung zu verfrühten Alterungsprozessen in der Zelle gekommen
ist. Diese frühzeitige Alterung kann durch eine erhöhte Stoffwechselbelastung der Muskel-
zellen infolge andauernder Arbeitsvorgänge verursacht werden (TRITSCHLER et al. 1994).
Allerdings wiesen alle vier Hunde nur Krankheitsanamnesen von längstens 14 Tagen auf.
Möglicherweise lag schon ein längeres Schmerzgeschehen ohne neurologische Symptomatik
bei den Tieren vor, dass für die Besitzer nicht sichtbar war. Dies ist vor allem bei Hund 13,
der an einem HWS-Syndrom litt, vorstellbar, da hier die Symptome zu Beginn der Erkran-
kung häufig nicht so gravierend sind.
Da es sich ausschließlich um Teckel handelt, liegt möglicherweise bei dieser Rasse eine
Diskopathie-Prädisposition auch aufgrund einer frühzeitig veränderten Rückenmuskulatur
vor.

In der Annahme, dass durch vermehrte Muskelarbeit eine erhöhte Stoffwechselaktivität ent-
steht, liegt die Vermutung nahe, dass eine Mitochondrienvermehrung als Anpassungsversuch
der Zelle zu verstehen ist.
Bei 22 Tieren fiel eine Mitochondrienvermehrung auf, davon bei zwei Hunden (Nr. 14, 22) in
so ausgeprägter Form, dass „ragged red fibres“ entstanden. Es traten vergrößerte, vermehrte,
irregulär angeordnete und zum Teil strukturell veränderte Mitochondrien auf.
Die Mitochondrienvermehrung war auch bei Hunden mit eher kurzem Krankheitsverlauf
(3-4 Tage) nachweisbar.
Bei zwei Hunden (Dalmatiner, m, 9 J, Spinalnervenkompression; DSH, m, 6J, Diskushernie
Th13/L1) mit einer Krankheitsdauer von ca. 6 Monaten waren zusätzlich strukturelle Mito-
chondrienveränderungen feststellbar.
Morphologisch abnorme Mitochondrien wurden bei unterschiedlichen neuromuskulären Er-
krankungen des Menschen z.B. Hypothyreose mit Myopathie (NORRIS und PANNER 1966),
Polymyositis (CHOU 1967) oder Mitochondriopathie mit Myopathie (SHY et al. 1966) be-
schrieben.
Das klinische Erscheinungsbild der Hunde offenbarte keine Anzeichen einer systemischen
Muskelerkrankung. Man kann vermuten, dass nach einer anfangs noch physiologischen Reak-
tion des Mitochondriums bei fortdauernder Belastung eine Insuffizienz des mitochondrialen
Stoffwechsels durch eine steigende Anzahl freier Radikale entsteht, denn „oxidativer Stress“
führt zu einer eingeschränkten Mitochondrienfunktion (TRITSCHLER et al. 1994). Die Mito-
chondrienvermehrung stellt somit wahrscheinlich den Versuch der Kompensation des mito-
chondrialen Funktionsverlustes dar.
Experimentell konnte bei Ratten durch eine elektrische Muskeldauerreizung eine mitochon-
driale Reaktion nachgewiesen werden (WALTER 1981).
Die Mitochondrienfunktionsstörung war in der vorliegenden Arbeit neben den strukturellen
Veränderungen der Mitochondrien und einer Mitochondrienvermehrung auch durch den
Nachweis der Ansammlung von Fett und Glykogen in der Zelle darstellbar.
Eine abnormale Lipidanreicherung in der Muskelzelle ist bei Hunden beiderlei Geschlechts,
unterschiedlichster Rassen und verschiedenen Alters beschrieben (SHELTON et al. 1998,
PLATT et al. 1999). In diesen beschriebenen Fällen deuteten die biochemischen Unter-
suchungen der Muskelbiopsien auf eine mitochondrale Stoffwechselstörung hin.
Eine intrazytoplasmatische Anreicherung von Lipiden konnte in der vorliegenden Arbeit bei
18 Hunden gefunden werden, eine Glykogenvermehrung wurde bei vier Hunden festgestellt.

Heinze untersuchte 1994 Rücken- und Gliedmaßenmuskelbiopsien von Hunden aus dem Sek-
tionsgut des Pathologischen Instituts der FU Berlin. Sie entdeckte ebenfalls Glykogenanrei-
cherungen in den Myozyten und fand dies in atrophischen aber auch in unveränderten Mus-
kelfasern. Sie schreibt dieses Phänomen ebenso einem gestörten Zellstoffwechsel zu.
Der Rückenmuskel neigt, wie von HEINZE (1994) herausgefunden, zur allimentär bedingten
Verfettung. Die Lipidablagerungen finden sich in diesen Fällen aber endomysial und inter-
faszikulär. Diese in der vorliegenden Arbeit nachgewiesene intrazelluläre Fettanreicherung ist
damit nicht zu erklären. Da die Rückenmuskulatur regelmäßig ein Überwiegen von Typ-I-
Fasern aufweist und Typ-I-Fasern mehr Neutralfett enthalten (JERUSALEM und ZIERZ
1991) als die glykolytischen, schnell zuckenden Typ-II-Fasern, könnte die Fasertypenprä-
dominanz die Verfettung begünstigen.
Weiterhin wurden „rimmed vacuoles“ bei fünf untersuchten Tieren (Nr. 2, 14, 22, 23, 40) ge-
funden. Erstmalig wurden „rimmed vacuoles“ von DUBOWITZ und BROOKE (1973) bei
Menschen mit oculopharyngealer Dystrophie beschrieben.
In den Untersuchungen von KNIRSCH (1997) traten in 1460 ausgewerteten Muskelbiopsien
von menschlichen Patienten mit neuromuskulären Erkrankungen 11,44 % Fälle mit neuro-
gener Gewebsreaktion und „rimmed vacuoles“ auf. Das Untersuchungsmaterial wurde aus un-
terschiedlichen Muskelbioptaten ausgewählt, die zu diagnostischen Zwecken an das Institut
für Neuropathologie der Freien Universität Berlin eingesandt worden waren.
Beim Tier wurden derartige Befunde meines Wissens noch nicht beschrieben.
Alle fünf Tiere bei denen in der vorliegenden Arbeit diese intrazytoplasmatischen Einschlüsse
auftraten, wiesen deutliche neurologische Störungen der Gliedmaßen auf (Ataxie, Para- oder
Hemiparese) und zeigten auch histologisch ausgeprägte neurologische Gewebssyndrome. Da
aber die eigenen Fallzahlen zu klein für eine endgültige Aussage zum Auftreten von „rimmed
vacuoles“ bei Hunden sind, müsste man weitere Untersuchungen mit einem entsprechend
großen Patientenkollektiv anstreben.
Bei zwei Hunden (Nr. 18, 21) waren degenerierte Myofibrillen mit Z-Streaming, einer ir-
regulären Ausdehnung des Z-Streifen-Materials, zu sehen. In fortgeschrittenen Fällen führt
das, wie bei drei weiteren Tieren (Hund 41, 49, 50) zu finden, zur Zusammenlagerung von
Z-Massen, sogenannten „rods“ oder „nemaline-bodies“, den stäbchenförmigen, intrazyto-
plasmatischen Anhäufungen von Z-Streifen-Material. Dies sind die häufigsten patholo-
gischen Reaktionen bei Myopathien des Hundes (CARDINET 1984), aber nicht pathogno-

monisch. Auch in der vorliegenden Arbeit sind sie am wahrscheinlichsten als unspezifische
myopathische Reaktion zu werten.
Es kann angenommen werden, dass es sich bei dem Untersuchungsgut um klinisch muskel-
gesunde Patienten mit einer isolierten Schädigung der Rückenmuskulatur infolge der degene-
rativen Wirbelsäulenerkrankungen handelt.
Ausgehend von unserer Hypothese, dass durch Schonhaltungen und Fehlbelastungen bei Rü-
ckenmarkskompressionen die Muskulatur überbeansprucht wird, konnte dies histologisch be-
stätigt werden. Es traten bei der überwiegenden Anzahl der untersuchten Patienten deutliche
myopathologische Veränderungen unterschiedlichster Form und Ausprägung auf.
Es konnten allerdings keine typischen histologischen Befunde herausgearbeitet werden, die
ausschließlich nach Wurzelreizung, Wurzelkompression oder Rückenmarkskompression auf-
treten.
Fasergrößenveränderungen
Im gesamten Untersuchungsgut trat die neurogene Atrophie n = 32 (zum Teil vergesell-
schaftet mit Begleitmyopathie) am häufigsten auf.
Die neurogene Atrophie ist gekennzeichnet durch Gruppen von atrophischen Fasern und
Target-Fasern. Die Target-Fasern werden seit Anfang der sechziger Jahre als Erkennungs-
merkmal bei Denervierungsprozessen herangezogen (ENGEL 1961). Es wurden aber auch
Target-Fasern bei Reinnervationsprozessen nachgewiesen (DUBOWITZ 1967) und seit eini-
gen Jahren sogar eher mit diesen in Verbindung gebracht (BANKER und ENGEL 1994).
Auch beim Hund konnten Target-Fasern als Biopsiebefunde bei einer Myopathie
(NEWSHOLME und GASKELL 1987, TARGETT et al. 1994) und bei Denervation
(GRIFFITH et al. 1973) nachgewiesen werden.
Da keiner der untersuchten Hunde einen Hinweis auf eine systemische Muskelerkrankung
zeigte und nach der Operation die klinischen Symptome in fast allen Fällen behoben wurden,
handelt es sich am wahrscheinlichsten um eine neurogene Ursache.
In der vorliegenden Untersuchung konnte bei den Tieren, die Target-Fasern aufwiesen, keine
Zuordnung zur Phase der Denervierung oder Reinnervation vorgenommen werden. Es war
lediglich festzustellen, dass es sich jeweils um besonders stark ausgeprägte neurogene Ge-
webssyndrome mit Begleitmyopathie handelte.

Nach den Untersuchungen von HEINZE (1994) ist die Muskelatrophie von der Dauer der spi-
nalen Leitungsunterbrechung und vom Ort der Läsion (je weiter kaudal der Wirbelsäule die
Läsion befindlich ist, umso stärker ausgeprägte Atrophie) abhängig.
Sie stufte die Muskulatur von Tieren mit ein- bis zweiwöchiger Lähmungsdauer durch Dis-
kusprolaps als unverändert ein.
Dies konnte in der vorliegenden Arbeit nicht bestätigt werden. Dabei wurden bei zum Teil nur
wenige Tage erkrankten Hunden (z.B. Fall Nr. 2, 3, 14) ausgeprägte neurogene Gewebs-
syndrome mit deutlicher Atrophie und Target-Fasern festgestellt.
Diese unterschiedlichen Befunde könnten damit erklärt werden, dass vor dem eigentlichen
Insult schon eine längere Krankheitsphase mit gelegentlichen Schmerzen vorlag, die den Be-
sitzern entgangen ist. Es ist aber auch vorstellbar, dass zusätzlich zur Inaktivitätsatrophie eine
Denervationsatrophie vorlag, welche zu einem ausgeprägteren Krankheitsbild in kürzerer Zeit
geführt hat. Als Zeichen für eine Denervationsatrophie können die gruppenweise Faser-
atrophie (HULLAND 1985), Fasertypengruppierung oder die Target-Fasern (SHY und
MAGEE 1956) angenommen werden wie Untersuchungen beim Menschen zeigen.
In normalen menschlichen Muskeln ist die Zahl der Fasern gleichen histochemischen Typs,
die unmittelbar nebeneinander liegen, nicht größer als 50, meistens sogar geringer als 15
(BLACK et al. 1974, MELTZER et al. 1976). JERUSALEM und ZIERZ (1991) sprechen
beim Zusammenliegen von über 40 Muskelfasern eines Typs oder bei mindestens 13 einge-
schlossenen, d.h. vom selben Muskelfasertyp umgebenen Muskelfasern, von Fasertypen-
gruppierung.
Die Fasertypengruppierung wird als Folge einer kollateralen Nervenfasersprossung gedeutet,
die auch im Experiment zu reproduzieren ist (KARPATI und ENGEL 1968).
Fasertypengruppierungen wurden auch bei Menschen mit Erkrankungen der Motoneurone
und bei Polyneuropathien beobachtet (STALBERG et al. 1976).
In den hier erfolgten Untersuchungen ließ sich bei fünf Hunden (Tiere 14, 23, 27, 29, 36) eine
Fasertypengruppierung nachweisen. Keines der Tiere hatte klinische Hinweise auf eine Poly-
neuropathie, so dass man am ehesten von einer kollateralen Nervenfasereinsprossung nach
Denervation ausgehen kann. In zwei Fällen (Tier 23: Yorkshire, m, 4J, Diskusprolaps
Th12/13, Erkrankungsdauer 6 Wochen; Tier 29: Teckel-Mix, m, 7J, Diskusprolaps L2/3,
Erkrankungsdauer 6 Monate) ließen sich im elektronenmikroskopischen Präparat ent-
sprechende degenerierte motorische Endplatten mit reduziertem synaptischen Faltenwerk und
Verlust der synaptischen Vesikel darstellen.

Fällt ein Teil der Muskelfasern in ihrer Funktion aus, können intakte Fasern kompensatorisch
hypertrophieren. Die Verteilungskurven der Fasertypen in den Histogrammen solcher Mus-
keln werden, wie schon HULLAND (1985) bemerkte, durch diese hypertrophierten Fasern
nach rechts verschoben und dadurch deutlich breiter.
Meist sind mehr Fasern atrophiert als hypertrophiert. Das deutet darauf hin, dass eine kom-
pensatorische Hypertrophie einzelner Fasern in der Regel nicht gleichzeitig mit der Atrophie
einsetzt, sondern erst, wenn die Atrophie bereits ein gewisses Ausmaß erreicht hat.
Allein anhand des Variationskoeffizienten ist es beim Hund nicht möglich festzustellen, ob
ein Muskel unverändert, atrophiert oder hypertrophiert ist (HEINZE 1994). Nur sehr niedrige
und sehr hohe Werte lassen eine Aussage über den jeweiligen Muskel zu.
Da ausgeprägte Fasergrößenveränderungen, ungeachtet ob sie nur einen Muskelfasertyp oder
beide betreffen, lichtmikroskopisch leicht erkannt werden können, wurde in der vorliegenden
Arbeit von der Bestimmung des Variationskoeffizienten abgesehen.
Ein Altersunterschied in Bezug auf die Fasergrößen wie er von BRAUND et al. (1992) und
CASTLE und REYMAN (1984) bei adulten Hunden bemerkt wurde, konnte hier bei Hunden
mit einem Altersspektrum von zwei bis elf Jahren nicht festgestellt werden.
Übereinstimmend mit den Untersuchungen von GUNN (1975, 1978 a, b, c, 1979a) wurden
auch in dieser Arbeit keine Größenunterschiede zwischen den Muskelfasern von männlichen
und weiblichen Tieren gefunden.
Über den Einfluss des Trainingszustandes lässt sich bei den hier untersuchten Hunden keine
Aussage machen, da die Haltungsbedingungen der Tiere nicht bekannt sind.
Lediglich bei einer sechsjährigen Großpudelhündin (Tier 44) war aus der Anamnese ein regel-
mäßiges, intensives Training bekannt. Bei diesem Tier fiel lichtmikroskopisch eine Hyper-
trophie beider Muskelfasertypen auf. Dies entspricht den Ergebnissen der Studien von
AGUERA et al. (1990), wurde aber, da es sich um einen Einzelfall handelte, nicht morpho-
metrisch abgesichert.
HEINZE (1994) stellte fest, dass bei einer spinalen Leitungsunterbrechung zunächst der kau-
dal der Läsion befindliche Teil der Rückenmuskulatur atrophiert. Erst wenn die Atrophie ein
gewisses Ausmaß erreicht hat, wird auch der kraniale Anteil mit einbezogen.
In der hier vorliegenden Arbeit wurden ebenfalls kranial und kaudal der Läsion Mus-
kelbiopsien entnommen. Ein vergleichbares Untersuchungsergebnis ließ sich nicht heraus-
arbeiten.

Ein Grund dafür könnte sein, dass die Biopsien sowohl kranial als auch kaudal relativ nah an
der Läsion entnommen wurden, da sich der Operationszugang üblicherweise nur über die an-
grenzenden beiden Wirbel erstreckt. HEINZE (1994) entnahm, unabhängig vom Ort der Lä-
sion, stets auf Höhe des 8. Thorakal- und des 5. Lendenwirbels die Biopsien. Da die häu-
figsten Rückenmarksalterationen zwischen dem 12. Brustwirbel und dem 3. Lendenwirbel la-
gen, ergeben sich deutlich größere Abstände zur Läsion.
Bei fünf Hunden (Tiere 4, 10, 11, 28, 31) konnte in den untersuchten Muskelbiopsien eine
Muskelfasertyp-II-Atrophie festgestellt werden. Derartige Veränderungen sind unter anderem
nach Tenotomie (ENGEL et al. 1966), bei Maldigestion, Kachexie, Kortikosteroidüber-
produktion und Inaktivität (SCHRÖDER 1989, SHIRES et al. 1982) beim Menschen be-
schrieben.
Bei den fünf Hunden ist es am wahrscheinlichsten, dass für die Typ-II-Atrophie eine Inakti-
vität aufgrund von Schmerzen bzw. Lähmungserscheinungen (gehfähige Paraparese bei Tier
10, 28 und 31, nichtgehfähige Paraparese bei Tier 4, Paraplegie bei Hund 11) verantwortlich
war.
Übereinstimmend mit der Arbeit von HEINZE (1994) wurden bei den in der vorliegenden
Arbeit untersuchten Tieren mit Rückenmarkskompressionen beidseits der Wirbelsäule
gleichsinnige neurogene und morphologische Veränderungen gefunden. Dies trifft auch für
die Hunde zu, bei denen klinisch und intraoperativ eine Seitenzuordnung der Kompression
festgestellt wurde.

6 Zusammenfassung
Romy Herrschelmann Histochemische und elektronenmikroskopische Untersuchungen der paravertebralen Muskulatur bei Hunden mit degenerativen Erkrankungen der Wirbelsäule Klinik für Kleintiere, Veterinärmedizinische Fakultät, Universität Leipzig Institut für Neuropathologie, Charité, Freie Universität Berlin Eingereicht im Mai 2005 106 S., 32 Abb., 5 Tab., 204 Lit. Schlüsselworte: Myopathie, Mitochondriopathie, Paravertebralmuskulatur, Elektronenmikroskopie, Rückenmarkskompression
Bandscheibenschäden und Instabilitäten der Wirbelsäule mit Rückenmarkskompression kön-
nen zu Schmerzuständen und/oder unterschiedlichen Graden neurologischer Ausfälle der
Gliedmaßen führen. Dies kann zu schmerzbedingten Schonhaltungen, Fehlbelastungen oder
hypertonen Zuständen der Rückenmuskulatur führen.
Von 50 Hunden verschiedener Rassen (Teckel und Teckelmischlinge n=21, kleinwüchsige
Rassen n=12, großwüchsige Rassen n=17) mit Rückenmarkskompression infolge eines Dis-
kusprolaps, einer Diskushernie, Instabilität der Wirbelsäule oder einem thorakal gelegenen
Menigiom wurden Muskelbiopsien während der Operation entnommen. Die Biospieentnahme
erfolgte aus den paravertebralen Rücken- bzw. Halsmuskeln kranial und kaudal der Rücken-
marksveränderung. Alle Biopsien, die lichtmikroskopisch pathologische Befunde aufwiesen
(n=47), wurden zusätzlich elektronenmikroskopisch und teilweise morphometrisch unter-
sucht. Besondere Beachtung fanden dabei die histochemischen und ultrastrukturellen Verän-
derungen.
Das Alter der Hunde lag zwischen zwei und elf Jahren (im Mittel 6,25 Jahre), die Erkran-
kungsdauer zwischen einem Tag und zwei Jahren (im Mittel 25,6 Tage). Zehn Hunde (20%)
zeigten ausschließlich Schmerzen, 32 Hunde (64%) eine Paraparese, zwei Hunde (4%) eine
Tetraparese, ein Hund (2%) eine Hemiparese, vier Hunde (8%) eine Paraplegie und ein Hund
(2%) eine Tetraplegie. Bei allen Tieren war der Tiefenschmerz erhalten.

Lichtmikroskopisch zeigten zwölf Tiere (24%) ein myopathisches Gewebsmuster. Neurogene
Veränderungen vergesellschaftet mit einem myopathischen Gewebsbild (Begleitmyopathie)
wiesen 28 von 50 (56%) Patienten auf. Eine ausschließlich neurogene Schädigung war bei
sieben Hunden (14%) zu finden, davon in fünf Fällen mit Ausbildung so genannter „rimmed
vacuoles“ (in der Muskelfaser vorkommende Vacuole mit granulärem Inhalt bzw. einer gra-
nulären Umrandung).
Bei fünf Hunden mit myopathischem Gewebssyndrom wurden degenerierte Myofibrillen mit
Z-streaming, in fortgeschrittenen Stadien Z-Massen bzw. „rods“ (Zusammenlagerung von Z-
Streifen Material) nachgewiesen.
Bei vier jungen Tieren konnte intrazytoplasmatisch Lipofuszin, in einem Fall ein sphärischer
Zytoplasmaeinschluß festgestellt werden.
Die myopathischen und neurogenen Veränderungen wie „rimmed vacuoles“, „rods“, Faser-
kalibervariationen und Faseratrophien stellten unspezifische Hinweise auf eine neuromus-
kuläre Erkrankung dar.
Bei 22 Hunden (44%) traten Mitochondrienvermehrungen auf. In zwei dieser Fälle wurden
lichtmikroskopisch „ragged red fibres“ (massive Vermehrung der Mitochondrien in der ge-
samten Muskelfaser) beobachtet, ein charakteristisches Bild für eine Verminderung des Mito-
chondrienstoffwechels. Elektronenmikroskopisch wurde bei zwei Hunden eine strukturelle
Umbaureaktion der Mitochondrien festgestellt.
Bei allen 22 Hunden wurden außerdem vermehrt Neutralfett, Glykogen oder beide Substrate
innerhalb der Muskelfaser gefunden. Diese Veränderungen weisen zusätzlich auf eine Ein-
schränkung des Mitochondrienstoffwechsels hin.
Das Auftreten mitochondrialer Veränderungen korreliert nicht wie üblich mit einem höheren
Lebensalter. Eine vorzeitige Alterung des Muskelgewebes aufgrund einer gestörten Stoff-
wechsellage der Mitochondrien (oxidativer Stress) erscheint wahrscheinlich.
Es kann angenommen werden, dass es sich bei dem Untersuchungsgut um klinisch muskel-
gesunde Patienten mit einer isolierten Schädigung der Rückenmuskulatur infolge der Rücken-
markskompression handelt. Die ausgeprägten myopathischen Veränderungen sind am ehesten
auf eine chronische Fehlbelastung oder Hypertonie der Rückenmuskulatur zurückzuführen.

7 Summary Romy Herrschelmann Histochemic and electron microscopic examinations of the paravertebral muscles of dogs having degenerative spinal column disease. Department of Small Animal Medicine, Faculty of Veterinary Medicine, University of Leipzig; Institute for Neuropathology, Charité, Open University of Berlin Submitted in May 2005 106 pp., 32 figures, 5 tables, 204 references. key words: myopathy, mitochondriopathy, paravertebral musculature, electron microscopy, spinal cord compression Intervertebral disc diseases and instabilities of the vertebral column with spinal cord com-
pression results in painful conditions and/or different neurological deficits. This may lead to
an overuse of paravertebral muscles caused by strenuous postures or hypertonic situations.
Paravertebral muscle biopsies were taken during the operation from 50 dogs (dachshounds
n=21, small breed dogs n=12, large breed dogs n=17) with spinal cord compression as a result
of a prolapsed disc, a disc hernia, instability of the vertebral column or a thoracal menigiom.
From both sides of the vertebral column, as well as cranial and caudal of the compression site
biopsies were examined.
All biopsies showing pathological findings (n=47) during microscopy were additionally
examined using an electron microscope and some were evaluated by a morphometric test.
Special emphasis was laid on histochemical and ultrastructural alterations.
The dogs age at presentation ranged from two to eleven years (mean 6,25 years). The duration
of signs before presentation varied between one day and two years (mean 25,6 days). Ten
dogs (20%) showed severe back pain, 32 dogs (64%) were paraparetic, two dogs (4%)
tetraparetic, one dog (2%) suffered from hemiparesis, four (8%) from paraplegia, and one
(2%) from tetraplegia, all with deep pain.
In the microscopical examinations muscles of twelve dogs (24 %) had changes compatible
with an myopathic tissue pattern. Twenty-eight out of 50 dogs (56 %) showed neurogenic
alterations combined with a myopathic tissue pattern (secondary myopathy).

The muscles of seven dogs (14 %) showed exclusively neurogenic changes; in five cases with
so-called “rimmed vacuoles” (vacuole with granular content or granular border within the
muscle fibre) were detected.
In five dogs with myopathic tissue pattern degenerated myofibrils with Z-streaming were
discovered, as well as Z-masses or “rods” (clumping of Z-stripe material), seen in an ad-
vanced stage.
In four young dogs lipofuszin was detected intracytoplasmatically, in one case a spherical
cytoplasmatic inclusion could be identified.
The myopathic and neurogenic alterations in form of “rimmed vacuoles”, “rods”, fibre caliber
variations and fibre atrophies represent non-specific changes for neuromuscular diseases.
In 22 dogs (44 %) the number of mitochondria was increased whereas in two cases structural
transformations in the mitochondrion itselfs were discovered. Lightmicroscopically two cases
showed “ragged red fibres” (increase of mitochondria in the whole fibre).
In all cases increased levels of neutral lipid, glycogen or both substrates were found within the
muscle fibre. These alterations indicate a limited mitochondrial metabolism. The presence of
mitochondrion abnormalities did not correlate with the usual higher age.
A premature aging of the muscle due to an impaired metabolism of mitochondria (oxidative
stress) seems reasonable.
It can be assumed that the dogs of this study did not suffer from a generalized myopathy be-
fore the spinal cord compression occurred but developed lesions of the vertebral muscles
caused by spinal cord compression.
The advanced myopathic alterations may be explained by inappropriate, chronic stress situ-
ation of paravertebral muscles due to pain and/or hypertonic status.

8 Literaturverzeichnis 1. Aguera E, Diz A, Vazquez-Auton J, Monterde JG. Differences in fibre population in dog muscles of different functional purpose. Anat Histol Embryol. 1990a; 19: 128-34. 2. Aguera E, Diz A, Vazquez-Auton J, Monterde JG. Muscle fibre morphometry in three dog muscles of different functional purpose in different breeds. Anat Histol Embryol. 1990b; 19: 289-93. 3. Armstrong RB, Saubert CW, Seeherman HJ, Tayler CR. Distribution of fiber types in locomotory muscles of dogs. Am J Anat. 1982; 163: 87-98. 4. Bagley RS, Kornegay JN, Page RL, Thrall DE. Central nervous system. in: Slatter DH: Textbook of Small Animal Surgery. Saunders, Philadelphia: 1993; 2137-66. 5. Bancroft JD. An indroduction to histochemical technique. Butterworth, London. 1976. 6. Banker BQ, Engel AG. Basic reactions of muscle. in: Engel AG, Franzini-Armstrong C: Myology, 2.Aufl. McGraw-Hill, New York. 1994; 832-88. 7. Behrens W. Tabellen zum Gebrauch bei mikroskopischen Arbeiten. 4.Aufl. S. Hirzel, Leipzig: 1908. 8. Belitser VA, Tsibakova ET. Oxidative Phosphorylierung. Biokhimiia. 1939; 4: 516-35. 9. Berg J. Problems in neurolocalization. in: Thacher C: Problems in veterinary medicine 1: Intervertebral disc disease. Lippincott, Philadelphia: 1989; 358-65. 10. Black JT, Bhatt GP, Dejesus PV, Schotland DL, Rowland LP. Diagnostic accuracy of clinical data, quantitative electromyography and histochemistry in neuromuscular disease. A study of 105 cases. J Neurol Sci. 1974; 21: 59-70. 11. Bloom F.Geriatrics. The locomotor system. in: Catcott EJ: Canine medicine Veterinary Publications: 1986; 837. 12. Braund KG, Taylor TKF, Ghosh P, Sherwood AA. Spinal mobility in the dog. A study in chondrodystrophoid and nonchondrodystrophoid animals. Res Vet Sci. 1977; 22: 78-82.
13. Braund KG, Hoff EJ, Richardson KEY. Histochemical identification of fiber types in canine skeletal muscle. Am J Vet Res. 1978; 39: 561-5. 14. Braund KG, Vandevelde M. German shepherd dog myelopathy – A morphologic and morphometric study. Am J Vet Res. 1978; 39: 1315. 15. Braund KG, Dillon AR, Mikeal RL. Experimental investigation of glucocorticoid-induced myopathy in the dog. Exp Neurol. 1980a; 68: 50-71.

16. Braund KG, Dillon AR, Mikeal RL, August JR. Subclinical myopathy associated with hyperadrenocorticism in the dog. Vet Pathol. 1980b; 17: 134-48. 17. Braund KG, Shires PK, Mikeal RL. Type I fiber atrophy in the vastus lateralis muscle in dogs with femoral fractures treated by hyperextension. Vet Pathol. 1980c; 17: 164-76.
18. Braund KG, Lincoln CE. Histochemical differentiation of fiber types in neonatal canine muscle. Am J Vet Res. 1981; 42: 407-15. 19. Braund KG, MacGuire JA, Lincoln CE. Observations on normal skeletal muscle of mature dogs: A cytochemical, histochemical and morphometric study. Vet Pathol. 1982; 19: 577-95. 20. Braund KG.Localizing lesions using neurologic syndromes – 2: Spinal cord syndromes. Vet Med. 1985a; 80: 54-63. 21. Braund KG. Skeletal muscle biopsy. in: Slatter DH: Textbook of Small Animal Surgery Saunders, Philadelphia: 1985b; 108-15. 22. Braund KG. Diseases of peripheral nerves, cranial nerves and muscles. in: Oliver JE, Hoerlein BF, Mayhew IG: Veterinary neurology. 1987; 353-92. 23. Brawner WR, Hathcock J T Neuroradiology. in: Slatter DH: Textbook of Small Animal Surgery, 3. Aufl. Saunders, Philadelphia: 2003; 1118-31. 24. Breitschwerdt EB, Kornegay JN, Wheeler SJ, Stenens JB, Baty CJ. Episodic weakness associated with exertional lactic acidosis and myopathy in Old English sheepdog littermates. J Am Vet Med Assoc. 1992; 201: 731-6. 25. Brenner O, de Lahunta A, Cummings JF, Summers BA, Monachelli M. A canine encephalomyelopathy with morphological abnormalities in mitochondria. Acta Neuropathol. 1997; 94: 390-7. 26. Brooke MH, Williamson E, Kaiser KK. The behaviour of four fiber types in developing and reinnervated muscle. Arch Neurol. 1977; 25: 360-6. 27. Brucher JM, Musoglu E, Bangels M. Computer-assisted morphometry of muscle inclu-ding four fiber types. Clin Neuropath. 1986; 5: 122. 28. Brueckner KA, Seim HB, Blass CE. Caudal cervical spondylomyelopathy: Decompres-sion by linear traction and stabilization with Steinmann pins and polymethyl methacrylate. J Am Anim Hosp Assoc. 1989; 25: 677. 29. Brueckner KA, Seim HB, Withrow SJ. Clinical evaluation of three surgical methods for treatment of caudal cervical spondylomyelopathy of dogs. Vet Surg. 1989; 18: 197.
30. Bubb WJ, Sims MH. Fiber type composition of rostral and caudal portions of the digastric muscle in the dog. Am J Vet Res. 1986; 47: 1834-42.

31. Budras KD, Fricke W. Atlas der Anatomie des Hundes: Lehrbuch für Tierärzte und Studierende. 3. Aufl. Schlütersche, Hannover: 1991. 32. Cardinet GH, Leong CL, Means PS. Myofiber differentiation in normal and hypotrophied canine pectineal muscles. Muscle & Nerve. 1982; 5: 665-73. 33. Cardinet GH. Nemaline rods in neuromuscular disorders of the dog. Anat Histol Embryol. 1984; 13: 87. 34. Carpenter S, Karpati G, Heller I, Eisen A. Inclusion body myositis. A distinct variety of idiopathic inflammatory myopathy. Neurology. 1978; 28: 8-17. 35. Castle ME, Reyman TA. The effect of tenotomy and tendon transfers on muscle fiber types in the dog. Clin Orthop Relat Res. 1984; 186: 302-10. 36. Chambers JN. Degenerative lumbosacral stenosis in dogs. Vet Med Rep. 1986; 2: 166-80. 37. Chou SM. “Megaconial” mitochondria observed in a case of chronic polymyositis. Neuro-logy (Minneap.). 1967; 17: 309-14. 38. Chrisman CL. Cerebrospinal fluid analysis. Vet C N Am: Small Anim Pract. 1992; 22: 781-810. 39. Clayton DA, Smith CA. Complex mitochondrial DNA. Int Rev Exp Pathol. 1975; 14: 1-15. 40. Cook JR, Oliver JE. Atlantoaxial subluxation in the dog. Comp Cont Ed Pract Vet. 1981; 3: 242-50. 41. Dallmann MJ, Pelattas P, Bojrab MJ. Characteristics of dogs admitted for treatment of cervical intervertebral disk disease: 105 cases (1972-1982). J Am Anim Hosp Assoc. 1992; 200: 2009-11. 42. Danon MJ, Sarpel G, Manaligod JR. Nemaline rod myopathy and Charcot-Marie-Tooth disease. Arch Neurol. 1980; 37: 123-7. 43. DeHaan JJ, Shelton SB, Ackerman N. Magnetic resonance imaging in the diagnosis of degenerative lumbosacral stenosis in four dogs. Vet Surg. 1993; 22: 14. 44. Delauche AJ, Cuddon PA, Podell M, Devoe K, Powell HC, Shelton GD. Nemaline rods in canine myopathies: 4 case reports and literature review. J Vet Intern Med. 1998; 12: 424-30. 45. Dennis R. Radiographic examination of the canine spine. Vet Rec. 1987; 121: 31-5. 46. Denny HR. The surgical treatment of cervical disc disease in the dog: a review of 40 cases. J Small Anim Pract. 1978; 19: 251-97.

47. Denny HR, Gibbs C, Gashell CJ. Cervical spondylopathy in the dog – a review of thirty-five cases. J Small Anim Pract. 1977; 18: 117-32. 48. Denny HR. The lateral fenestration of thoracolumbar disc protrusions: a review of 30 cases. J Small Anim Pract. 1978; 19: 259-66. 49. Denny HR. Fractures of the cervical vertebrae in the dog. Vet Ann. 1983; 23: 236-40. 50. Dixon BC, Tomlinson JL, Kraus KH. Modified distraction-stabilization technique using an interbody polymethyl methacrylate plug in dogs with caudal cervical spondylomyelopathy. J Am Vet Med Assoc. 1996; 208: 61-3. 51. Dolan P, Adams MA. Muscle fatigue increases the bending stresses acting on the lumbar spine during manual handling. J Bone Joint Surg (Br). 1995; 77: 326-7. 52. Dubovitz V. Pathology of experimentally reinnervated skeletal muscle. J Neurol Neuro-surg Psychiatr. 1967; 30: 99-105. 53. Dubovitz V, Brooke MH. Muscle biopsy: A modern approach. Saunders, London: 1973; 231-41. 54. Dubovitz V. Muscle biopsy: A practical approach. Baillères Tindall, Eastbourne: 1985; 3-129. 55. Duncan ID, Griffith IR, Nash AS. Myotonia in canine Cushings disease. Vet Rec. 1977; 100: 30-1. 56. Engel AG, Gomez MR. Nemaline (Z disk) myopathy: observations on the origin, structure and solubility properties of the nemaline structures. J Neuropathol exp Neurol. 1967; 26: 601-19. 57. Engel WK. Muscle target fibres, a newly recognized sign of denervation. Nature (Lon-don). 1961; 191: 389-90. 58. Engel WK. The essentiality of histo- and cytochemical studies of skeletal muscle in the investigation of neuromuscular disease. Neurology (Minneap.). 1962; 13: 919-23. 59. Engel WK, Cunningham GG. Rapid examination of muscle tissue. An improved trichome method for freshfrozen biopsy sections. Neurology (Minneap.). 1963; 12: 778-94. 60. Engel WK, Brooke MH, Nelson PG. Histochemical studies of denervated or tenotomized cat muscle: illustrating difficulties in relating experimental animal conditions to human neuromuscular diseases. Ann NY Acad Sci. 1966; 138: 160-85. 61. Evans HE. Miller`s anatomy of the dog. 2.Aufl. Saunders, Philadelphia: 1993.
62. Faludi G, Mills LC, Chayes ZW. Effects of steroids on muscles. Acta Endocrinol. 1964; 45: 68-78.

63. Farber E, Sternberg WH, Dunlap CD. Histochemical localization of specific oxidative enzymes. Tetrazolium stains for diphosphopyridine nucleotide diaphorase and triphos-phopyridine nucleotide diaphorase. J Histochem Cytochem. 1956; 4: 147-66. 64. Feeney DA, Wise M. Epidurography in the normal dog: technique and radiographic findings. Vet Radiol. 1981; 22: 35-9. 65. Flückinger M. Radiologische Untersuchung der Wirbelsäule. Proceedings Jahresver-sammlung der Schweizerischen Vereinigung für Kleintiermedizin 2.-4. Mai Lausanne 29-38: 1985. 66. Fletcher TF. Spinal cord and meninges. in: Evans H.E.: Miller`s anatomy of the dog. 3. Aufl. Saunders, Philadelphia: 1993; 800-28. 67. Forsynthe WB, Ghoshal HG. Innervation of the canine thoracolumbar vertebral column. Anat Rec. 1984; 208: 63-75. 68. Frewein J, Heininger-Meylan M, Brunner K. Zur Blutgefäßversorgung der Zwischen-wirbelscheiben im Lendenbereich beim Hund. Tierärztl Wschr. 1986; 99: 174-6. 69. Fry T, Johnson AL, Hungerford L, Toombs J. Surgical treatment of cervical disc herniations in ambulatory dogs. Ventral decompressions versus fenestration, 111 cases. Progr Vet Neurol. 1991; 2: 165-73.
70. Funkquist B. Thoracolumbar disc protrusion with severe cord compression in the dog. I: Clinical and pathoanatomical observations with special reference to the role of development of the symptoms of motor loss. Acta Vet Scand. 1962; 3: 256-74. 71. Gärtner K, Reulecke W, Hackbarth H, Wollnik F. Zur Abhängigkeit von Muskelmasse und Körpergewicht im Vergleich von Maus, Ratte, Kaninchen, Hund, Mensch und Pferd. Dtsch Tierärztl Wschr. 1987; 94: 52-3. 72. Gage ED. Incidence of clinical disc disease in the dog. J Am Anim Hosp Assoc. 1975; 11: 135-8. 73. Geary JC, Oliver JE, Hoerlein BF. Atlantoaxial subluxation in the canine. J Small Anim Pract. 1967; 8: 577-82. 74. Ghadially NF. Ultrastructural Pathology of the Cell and Matrix. 4. Aufl. Butterworth-Heinemann, Boston. 1997; 195-327. 75. Goebel HH, Müller J, Gillen HW, Merrit AD. Autosomal dominant „spheroid body myopathy“. Muscle & Nerve. 1978; 1: 14-26. 76. Goggin JE, Li A, Franti CE. Canine intervertebral disc disease: characterization by age, sex, breed and anatomic site of involvement. Am J Vet Res. 1970; 31: 1687-92. 77. Gosh P, Taylor KF, Yarroll JM. Genetic factors in the maturation of the canine intervertebral disc. Res Vet Sci. 1975; 19: 304-11.

78. Grevel V. Liquorzytologie und bildgebende Verfahren zur Diagnostik neurologischer Er-krankungen bei Hund und Katze. Habilitationsschrift, Freie Universität Berlin: 1994.
79. Grevel V, Cop Z. Ungewöhnliche Bandscheibenvorfälle beim Hund. Teil I: Vorge-fallenes Bandscheibenmaterial aus klinischer Sicht. Kleintierpraxis. 1992; 37: 657-64. 80. Grevel V, Cop Z, Wagner S. Die Bedeutung der röntgenologischen Untersuchungs-methoden für die Diagnosestellung einer Lumbosakralen Stenose beim Hund. Proceedings XVIII. WSAVA Weltkongress und 39. Jahrestagung der FKDVG: 19936.-9. Oktober, Berlin: 42-6. 81. Grevel V, Schwartau K. Die Hemilaminektomie beim thorakolumbalen Bandscheiben-vorfall des Hundes. 1.Teil: Klinische Befunde und Röntgendiagnostik. Kleintierpraxis. 1997; 42: 5-20. 82. Grevel V, Schwartau K. Die Hemilaminektomie beim thorakolumbalen Bandscheiben-vorfall des Hundes. 2. Teil: Operationsbefunde und Operationsergebnisse. Kleintierpraxis. 1997; 42: 173-96. 83. Griffiths IR, Duncan ID, Quirk C. “The central areas” of denervated canine muscle. J Comp Pathol. 1973; 83: 493-8. 84. Griffiths IR. Spinal disease in the dog. Practice. 1992; 4: 44-52. 85. Grivell LA. Mitochondrial DNA. Sci Am. 1983; 248: 78-89. 86. Gruber AD, Wessmann A, Vandevelde M, Summers BA, Tipold A. Mitochondriopathy with Regional Encephalic Mineralization in a Jack-Russel Terrier. Vet Pathol. 2002; 39:732-6. 87. Gunn HM. Adaptions of the skeletal muscle that favour athletic ability. N Z Vet J. 1975; 23: 249-54. 88. Gunn HM. The mean fibre area of the semitendinosus, diaphragm and pectoralis transversus muscles in differing types of horse and dog. J Anat. 1978a; 127: 403-14. 89. Gunn HM. Differences in the histochemical properties of skeletal muscles of different breeds of horses and dogs. J Anat. 1978b; 127: 615-34. 90. Gunn HM. The proportions of muscle, bone and fat in two different types of dogs. Res Vet Sci. 1978c; 24: 277-82. 91. Gunn HM. Total fibre numbers in cross section of the semitendinosus in athletic and non-athletic horses and dogs. J Anat. 1979a; 128: 821-8. 92. Gunn HM. The growth of the transverse sectional area (TSA) of M. semitendinosus in the dog and horse and its relation to athletic ability in the two species. Anat Histol Embryol. 1979b; 8: 365-8.

93. Guy PS, Snow DH. Skeletal muscle fibre composition in the dog and its relationship to athletic ability. Res Vet Sci. 1981; 31: 244-8. 94. Gysling C. Der Alterungsprozess der Zwischenwirbelscheibe beim Deutschen Schäfer-hund. Diss Vet-med Fakultät Zürich: 1984. 95. Hansen HJ. A pathologic-anatomical study on disc degeneration in dog with special reference to the so called Enchondrosis intervertebralis. Acta Orthop Scand. 1952; 11: 130-42. 96. Hare WCD. Zur Ossifikation und Vereinigung der Wirbelepiphysen beim Hund. Wien Tierärztl Wschr. 1961; 48: 210-5. 97. Heinze C. Histometrische und histologische Untersuchungen zur Diagnostik von Muskelveränderungen bei Hunden mit einer Darstellung der in der Literatur beschriebenen Muskelerkrankungen des Hundes. Diss Freie Universität Berlin: 1994. 98. Herrtage ME, Houlton JEF. Collapsing clumber spaniels. Vet Rec. 1979; 105: 1260-6. 99. Herrtage ME, Houlton JEF. Mitochondrial myopathy in the Sussex spaniel. Vet Rec. 1979; 106: 206-10. 100. Hoerlein BF. Canine neurology. 3. Aufl. Saunders, Philadelphia: 1978. 101. Holmberg DL, Palmer NC, Vanpelt D, Willan AR. A comparison of manual and power assisted thoracolumbar disc fenestration in dogs. Vet Surg. 1990; 19: 323-7. 102. Hulland TJ. Muscles and Tendons. in: Jubb KVF, Kennedy PC, Palmer N: Pathology of domestic animals. Bd. I Academic Press, Orlando. 1985; 139-99. 103. Hutter H. Mm. intertransversarii cervicis, thoracis, lumborum et caudae bei Pferd, Rind, Schwein und Hund. Diss Vet-med Fakultät Wien: 1977. 104. Jaggy A, Lang J, Schawalder P. Cauda equina-Syndrom beim Hund. Schweiz Arch Tierheilkd. 1987; 129: 171-92. 105. Jerusalem F, Zierz S. Muskelerkrankungen. Klinik, Therapie, Pathologie. Thieme, Suttgart: 1991. 106. Julian LM, Cardinet GH. Fiber sizes of the biceps brachii muscle of dogs which differ greatly in body size. Anat Rec. 1961; 139: 243. 107. Jurina K. Zervikale Bandscheibenvorfälle beim Hund - Klinische und röntgenologische Untersuchungsbefunde prae und post operationem. Diss Freie Universität Berlin: 1996. 108. Jurina K, Grevel V. Der zervikale Bandscheibenvorfall beim Hund. Teil 1: Klinische und röntgenologische Befunde. Tierärztl Prax. 2002a; 30: 88-96.

109. Jurina K, Grevel V. Der zervikale Bandscheibenvorfall beim Hund. Teil 2: Operation und Nachuntersuchung. Tierärztl Prax. 2002b; 30: 192-201. 110. Karpati G, Engel WK. „Type grouping“ in skeletal muscles after experimental reinnervation. Neurology (Minneap.). 1968; 18: 447-55. 111. Keller H. Die Topographie der ventralen Seite des Hundehalses unter spezieller Berücksichtigung chirurgischer Eingriffe. Zbl Vet Med A. 1963; 10: 513-35.
112. Knirsch U. Ubiquitin und Amyloid bei Einschlusskörperchenmyositis und anderen Muskelerkrankungen mit „Rimmed Vacuoles“. Diss Freie Universität Berlin: 1997. 113. Kornegay JN, Tuler SM, Miller DM, Levesque DC. Muscular dystrophy in a litter of golden retriever dogs. Muscle & Nerve. 1988; 11: 1056-64. 114. Krüger W. Musculus multifidus und Mm. rotatores der Haussäugetiere und ihre Beziehungen zur Drehfähigkeit der Wirbelsäule. Anat Anz. 1927; 63: 305-27. 115. Lang J, Hani H, Schawalder P. A sacral lesion resembling osteochondrosis in the German shepherd dog. Vet Radiol. 1992; 33: 69-76. 116. Latorre R, Gil F, Vazquez JM, Moreno F, Mascarello F, Ramirez G. Skeletal muscle fibre types in the dog. J Anat. 1993; 182: 329-37. 117. Leonhardt H. Histologie, Zytologie und Mikroanatomie des Menschen. 8. Aufl. Springer, New York. 1990: 45-47, 202-13. 118. Levine SH, Caywood DD. Recurrence of neurological deficits in dogs treated for thoracolumbar disc disease. J Am Vet Med Assoc. 1984; 20: 889-94. 119. Lewis DG. Cervical spondylomyelopathy (wobbler`s syndrome) in the dog: A study based on 224 cases. J Small Anim Pract. 1989; 30: 657-65. 120. Lewis DG. Cervical spondylomyelopathy (wobbler`s syndrome) in the dog. Practice. 1992; 14: 125-30. 121. Liebich HG. Stoffwechselsysteme der Zelle. in: Funktionelle Histologie. Schattauer, Stuttgart: 1990; 12-4. 122. Lillie RD, Ashburn LL. Optimisation of oil red O staining permits combination with Sudan. in: Bancroft JD, Stevens A: Theory and practice of histological technique. Springer, New York: 1943; 113-6. 123. Lipman F. Energiereiche Phosphatbindungen. Enzymologia. 1941; 2: 52-74. 124. Müller H. Der Bandscheibenvorfall beim Hund. Tierärztl Umschau. 1955; 10: 435-40. 125. MacDonald RD, Engel AG. The cytoplasmatic body: another structural anomaly of the Z disc. Acta Neuropathol. 1969; 14: 99-107.

126. Mannion AF, Weber BR, Dvorak J, Grob D, Müntener M. Fibre type characteristics of the lumbar paraspinal muscles in normal healthy subjects and in patients with low back pain. J Orthop Res. 1997; 15: 881-7. 127. Mannion AF. Fibre type characteristics and function of the human paraspinal muscles: normal values and changes in association with low back pain. Electromyogr Kinesiol. 1999; 9: 363-77. 128. Mannion AF, Käser L, Weber E, Rhyner A, Dvorak J, Müntener M. Influence of age and duration of symptoms on fibre type distribution and size of the back muscles in chronic low back pain. Eur Spine J. 2000; 9: 273-81. 129. Mattila M, Hurme M, Alaranta H, Paljärvi L, Kalimo H, Falck B, Lehto M, Einola S, Järvinen M. The multifidus muscle in patients with lumbar disc herniation. A histochemical and morphometric analysis of intraoperative biopsies. Spine. 1986; 11: 732-8. 130. Mitchell P. Coupling of phosphorylation to electron and hydrogen transfer by chemiosmotic type of mechanism. Nature. 1961; 191: 144-8.
131. Mitchell P. Possible molecular mechanisms of the protonmotive function of cytochrome systems. J Theor Biol. 1976; 62: 327-67.
132. Mascarello F, Vegetti, A, Carpene E, Rowlerson A, Jenny E. The tensor timpani muscle of cat and dog contains IIM and slow-tonic fibres: an unusual combination of fibre types. J Muscle Res Cell Motil. 1982; 3: 363-74. 133. Maxwell LC, Barclay JK, Mohrmann DE, Faulkner JA. Physiological characteristics of skeletal muscles of dogs and cats. Am J Physiol. 1977; 233: 14-8. 134. Meltzer HY, Rastogi S, Ellison J. Quantitative histochemical evaluation of normal human skeletal muscle. Neurology (Minneap.). 1976; 26: 849-52. 135. Moore MJ, Rebeiz JJ, Holden M, Adams RD. Biometric analyses of normal skeletal muscle. Acta Neuropathol. 1971; 19: 51-69. 136. Morgan-Huges JA. Mitochondrial Disease. Myology. 2. Aufl. McGraw-Hill, New York. 1994; 1610-60. 137. Morpurgo B. Über Activitäts-Hypertrophie der willkürlichen Muskeln. Arch pathol Anat Phys klin Med. 1897; 150: 522-54. 138. Nakashima N, Tamura Z, Okamoto S, Goto H. Inclusion bodies in human neuromuscular disorders. Arch Neurol. 1970; 22: 270-8. 139. Newsholme SJ, Gaskell CJ. Myopathy with core-like structures in a dog. J Comp Pathol. 1987; 97: 597-600. 140. Newsholme SJ, Lexel J, Downham DY. Distribution of fibre types and fibre sizes in the tibialis cranialis muscle of beagle dogs. J Anat. 1988; 160: 1-8.

141. Nickel R, Schummer A, Seiferle E. Bewegungsapparat. in: Lehrbuch der Anatomie der Haussäugetiere. Bd 1, 6. Aufl. Parey, Stuttgart: 1992; 28-52, 225-30. 142. Niederer M. Der Musculus splenius capitis und der Musculus splenius cervicis bei unseren Haustieren. Diss Vet-med Fakultät Zürich: 1985.
143. Norris FH, Panner BJ. Hypothyreoid myopathy – clinical, electromyographical and ultrastructural observation. Arch Neurol (Chic.). 1966; 14: 574-89. 144. Olby NJ, Chan KK, Targett MP, Houlton JEF. Suspected mitochondrial myopathy in a Jack Russel terrier. J Small Anim Pract. 1997; 38: 213-6. 145. Orvis JS, Cardinet GH. Canine muscle fiber types and susceptibility of masticatory muscles to myositis. Muscle & Nerve. 1981; 4: 354-9. 146. Paciello O, Maiolino P, Fatone G, Papparella S. Mitochondrial Myopathy in a German Sheperd Dog. Vet Pathol. 2003; 40: 507-11. 147. Platt SR, Chrisman CL, Shelton GD. Lipid storage myopathy in a cocker spaniel. J Small Anim Pract. 1999; 40: 31-4. 148. Prata RG. Diagnosis of spinal cord tumors in the dog. Vet C N Am: Small Anim Pract. 1977; 7: 165-85. 149. Price HM. The skeletal muscle fiber in the light of electron microscopic studies. Am J Med. 1963; 35: 589. 150. Racker E. Function and structure of the inner membrane of mitochondria and chloroplasts. in: Membranes of mitochondria and chloroplasts. Van Nostrand, New York: 1970; 127-71. 151. Ramsbacher J, Theallier-Janko A, Stoltenburg-Didinger G, Brock M. Ultrastructural changes in paravertebral muscles associated with degenerative spondylolisthesis. Spine. 2001; 26: 2180-5. 152. Randal T. Mitochondrial DNA. A new frontier in acquired and inborn gene defects. J Am Med Ass. 1991; 266: 1739. 153. Rebeiz JJ, Moore MJ, Holden EM, Adams RD. Variations in muscle status with age and systemic diseases. Acta Neuropath (Berl.). 1972; 22: 127-44. 154. Rewcastle NB, Humphrey JG. Vacuolar myopathy: Clinical, histochemical and microscopic study. Arch Neurol. 1965; 12: 570-82. 155. Richardson KC, Jarett L, Finke EH. Embedding in exposy resins for ultrathin sectioning in electronic microscopy. Stain Technol. 1960; 35: 337-45. 156. Riede UN, Schaefer HE. Störungen der zellulären und extrazellulären Organisation. in: Allgemeine und spezielle Pathologie, 3.Aufl. Thieme, Stuttgart: 1993; 24-5.

157. Rifai Z, Welle S, Kamp C, Thornton CA. Ragged red fibers in normal aging and inflammatory myopathy. Ann Neurol. 1995; 37: 24-9. 158. Rist G. Über das Schicksal prolabierten Bandscheibengewebes beim Hund. Diss Ludwig-Maximilians Universität München: 1982. 159. Roberts RE, Selcer BA. Myelography and epidurography. Vet Clin North Am (Small Anim Pract). 1993; 23: 307-29. 160. Rodriguez-Barbudo MV, Vaamonde R, Aguera E, Carpio M, Moreno F, Fuentes S, Robina A. Estudios histoquimico y morfometrico del muscolo tibial craneal en perros de diferentes aptitudes dinamicas (galgo, pastor aleman y fox terrier). Histochemische und morphometrische Untersuchungen am M. tibialis cranialis von Hunden mit verschiedenen dynamischen Fähigkeiten (Windhund, Deutscher Schäferhund und Foxterrier). Anat Histol Embryol. 1984; 13: 300-12. 161. Romeis B. Mikroskopische Technik. 16. Aufl. WR Oldenbourg, München: 1968. 162. Rosenblatt JD, Kuzon WM, Pynn BR, Plyley MJ, MacKee NH. Fiber type, fiber size and capillary geometric features of the semitendinosus muscle in three types of dogs. Am J Vet Res. 1988; 49: 1573-6. 163. Rowlerson A, Pope B, Murray J, Whalen RB, Weeds AG, Adams RD. A novel myosin present in cat jaw-closing muscles. J Muscle Res Cell Motil. 1981; 2: 417-38. 164. Russel SW, Griffith RC. Recurrence of cervical disc syndromes in surgically and conservatively treated dogs. J Am Vet Med Assoc. 1968; 153: 1412-7. 165. Ryan MM, Schnell C, Strickland CD. Nemaline myopathy: a clinical study of 143 cases. Ann Neurol. 2001; 50: 312-20. 166. Sajonski H, Smollich A. Mitochondrien. in: Zelle und Gewebe. 7. Aufl. S Hirzel, Leipzig: 1990. 167. Scholtysik G. Die normale Zwischenwirbelscheibe des Hundes normaler und chondro-dystopher Rassen. Diss Freie Universität Berlin: 1962. 168. Schröder JM. Pathologie der Muskulatur. in: Spezielle pathologische Anatomie. Doerr, Seifert: 1989; 385-460. 169. Schwalbe G, Mayeda R. Über die Kaliberverhältnisse der quergestreiften Muskelfasern des Menschen. Z Biol. 1890; 27: 482-516. 170. Seim HB, Withrow SJ. Pathophysiology and diagnosis of caudal cervical spondylomyelopathy with emphasis on the Doberman pinscher. J Am Vet Med Assoc. 1982; 18: 241-50. 171. Seim HB, Prata RG. Ventral decompression for the treatment of cervical disc disease in the dog: a review of 54 cases. J Am Anim Hosp Assoc. 1982; 18: 233-40.

172. Seitz D. Die Bedeutung der Muskelbiopsie für die Diagnose und Therapie chronischer neuromuskulärer Prozesse. Dtsch Z Nervenheilk. 1965; 187: 136-65. 173. Shafiq SA, Milhorat AT, Gorycki MA. Fine structure of human muscle in neurogenic atrophy. Neurology (Minneap.). 1967; 17: 934-48. 174. Shelton GD, Cardinet GH. Pathophysiologic basis of canine muscle disorders. J Vet Intern Med. 1987; 1: 36-44. 175. Shelton GD. Differential diagnosis of muscle diseases in companion animals. PVN Review. 1991; 2: 27-32. 176. Shelton GD, Nyhan WL, Kass PH. Analysis of organic acids, amino acids, and carnitine in dogs with lipid storage myopathy. Muscle & Nerve. 1998; 21: 1202-5. 177. Shelton GD, van Ham L, Bhatti S. Pyruvate dehydrogenase deficiency in Clumber and Sussex Spaniels in the United States. J Vet Intern Med. 2000; 14: 342. 178. Shires PK, Braund KG, Milton JL. Effect of localized trauma and temporary splinting on immature skeletal muscle and mobility of the femorotibial joint in the dog. Am J Vet Res. 1982; 43: 454-60.
179. Shy GM, Magee KR. A new congenital non-progressive myopathy. Brain. 1956; 79: 610-21. 180. Shy GM, Gonatas NK, Perez M. Two childhood myopathies with abnormal mitochondria, Part 1 (Megaconial myopathy), Part 2 (Pleoconial myopathy). Brain. 1966; 89: 133-58. 181. Sisson AF, LeCouteur RA, Ingram JT, Park RD, Child G. Diagnosis of cauda equine abnormalities by using electromyography, discography and epidurogaphy in dogs. J Vet Intern Med. 1992; 6: 253-63. 182. Snow DH, Billeter R, Mascarello F, Carpene E, Rowlerson A, Jenny E. No classical type IIB fibres in dog skeletal muscle. Histochemistry. 1982; 75: 53-65. 183. Stalberg E, Schwartz MS, Thiele B, Schiller HH. The normal motor unit in man. A single fibre EMG multielectrode investigation. J Neurol Sci. 1976; 27: 291-301. 184. Summers BA, DeLahunta A, McEntee M, Kuhajda FP. A novel intradural extra-medullary spinal cord tumors in young dogs. Acta Neuropathol. 1988; 75: 402-10. 185. Suter PF. Systemische Interpretation von Röntgenbildern der Wirbelsäule. Proceedings 4th Small Animal Surgical Course for Foreign Practioneers: Neurologic Surgery in the Dog. 1984; 16.-20. Juli, Davis (USA). 186. Swaim SF. Ventral decompression of the cervical spinal cord in the dog. J Am Vet Med Assoc. 1974; 164: 491-5.

187. Targett MP, Franklin RJM, Olby NJ. Central core myopathy in a great dane. J Small Anim Pract. 1994; 35: 100-3. 188. Thatcher C. Neuroanatomic and pathophysiologic aspects of intervertebral disc disease in the dog. in: Problems in veterinary medicine I: Intervertebral disc disease. Lippincott, Philadelphia: 1989; 337-57. 189. Tice LW, Engel AG. The effects of glucocorticoids on red and white muscles of the rat. Am J Pathol. 1967; 50: 311-33. 190. Tomlinson BE, Walton JN, Rebeiz JJ. The effects of ageing and of cachexia upon skele- tal muscle. A histopathological study. J Neurol Sci. 1969; 9: 321-46. 191. Toombs JP, Waters DJ. Intervertebral disc disease. in: Slatter DH: Textbook of small animal surgery. 3.Aufl. Saunders, Philadelphia: 2003; 1193-209. 192. Tritschler HJ, Packer L, Medori MD. Oxidative stress and mitochondrial dysfunction in neurodegeneration. Biochem Mol Biol Int. 1994; 34: 169-81. 193. Vaughan LC. Studies on intervertebral disc protrusion in the dog. 2. Diagnosis of the disease. Brit Vet J. 1958; 114: 203-09. 194. Vijayasarathy C, Giger U, Prociuk U, Patterson DF, Breitschwerdt EB, Avadhani NG. Canine mitochondrial myopathy associated with reduced mitochondrial mRNA and altered cytochrome c oxidase activities in fibriblasts and skeletal muscle. Comp Biochem A Physiol. 1994; 109: 887-94. 195. Walter GF. Neuromuskuläre Mitochondriopathie. Urban & Fischer, München: 1981. 196. Watson AG, DeLahunta A. Atlantoaxial subluxation and absence of transverse ligament of the atlas in a dog. J Am Vet Med Assoc. 1992; 195: 235-7. 197. Weber BR, Grob D, Dvorak J, Muntener M. Posterior surgical approach to the lumbar spine and its effect on the multifidus muscle. Spine. 1997; 22: 1765-72. 198. Wheeler SJ. Lumbosacral disorders in dogs. Vet Ann. 1990; 30: 262-8. 199. Wheeler SJ. Atlantoaxiale subluxation with absence of the dens in a rottweiler. J Small Anim Pract. 1992; 33: 90-3. 200. Wheeler SJ. Lumbosacral disease. Vet C N Am: Small Anim Pract. 1992; 22: 937-50. 201. Wheeler SJ, Sharp NJH. Small animal spinal disorders. Diagnosis and surgery. Mosby-Wolfe, London: 1994. 202. Wohlfart G. Über das Vorkommen verschiedener Arten von Muskelfasern in der Skelettmuskulatur des Menschen und einiger Säugetiere. Acta Psychatr Neurol Scand (Suppl.). 1937; 12: 1-119.

203. Yamaguchi M, Robson RM, Stromer MH. Nemaline myopathy rod bodies. Structure and composition. J Neurol Sci. 1982; 56: 35-56.
204. Zhu XZ, Parnianpour M, Nordin M, Kahanovitz N. Histochemistry and morphology of erector spinae muscle in lumbar disc herniation. Spine. 1989; 14: 391-7.

Danksagung Mein herzlicher Dank gilt Frau Professor Dr. Grevel für die Überlassung des interessanten
Themas, die Probenentnahme, die gewährte Hilfe sowie die kritische und stets rasche Durch-
sicht der Arbeit.
Weiterhin meinen allerherzlichsten Dank an Frau Professor Dr. Stoltenburg-Didinger für ihre
jederzeit gewährte Hilfe, die vielen Stunden gemeinsamer Arbeit am Mikroskop, ihr Engage-
ment und vor allem für die wiederkehrenden Motivationsschübe.
Ich danke den technischen Mitarbeiterinnen des Institutes für Neuropathologie für ihre
schnelle und sehr gute Bearbeitung der Muskelbiopsien.
Außerdem ein Dank an Herrn Dr. Scharvogel für die Muskelprobenentnahme und den An-
sporn, die Arbeit zu vollenden.
Schließlich gilt mein Dank meinem lieben Mann, der mir bei allem zur Seite stand, für seine
technische Assistenz und Geduld und meinen Eltern, die mir die Ausbildung zur Tierärztin
ermöglicht haben.