Embed Size (px)
Citation preview

of February 14, 2018.This information is current as
Interaction with Opsonized Target CellsProfessional Antigen Presentation by
T Lymphocytes Are Licensed forδγHuman
Cohen, Kenth Gustafsson and John AndersonYan, Bertrand Vernay, Luisa Saraiva, Yin Wu, Cyrille J. Nourredine Himoudi, Daniel A. Morgenstern, Mengyong
http://www.jimmunol.org/content/188/4/1708doi: 10.4049/jimmunol.1102654January 2012;
2012; 188:1708-1716; Prepublished online 16J Immunol
MaterialSupplementary
4.DC1http://www.jimmunol.org/content/suppl/2012/01/17/jimmunol.110265
average*
4 weeks from acceptance to publicationSpeedy Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
?The JIWhy
Referenceshttp://www.jimmunol.org/content/188/4/1708.full#ref-list-1
, 16 of which you can access for free at: cites 32 articlesThis article
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2012 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Human gd T Lymphocytes Are Licensed for ProfessionalAntigen Presentation by Interaction with Opsonized TargetCells
Nourredine Himoudi,* Daniel A. Morgenstern,* Mengyong Yan,* Bertrand Vernay,†
Luisa Saraiva,‡ Yin Wu,‡ Cyrille J. Cohen,x Kenth Gustafsson,‡,1 and John Anderson*,1
Activated human blood gd T cells have also been previously demonstrated to behave as professional APCs, although the processes
that control APC function have not been characterized. n this study, we show that the acquisition of potent APC function by
human blood gd T cells is achieved after physical interaction with an Ab-coated target cell, a process that we refer to as licensing.
In cancer models, licensing of gd T cells by tumor-reactive mAbs promotes the uptake of tumor Ags and professional presentation
to tumor-reactive ab T cells. We propose that licensing by Ab is a mechanism whereby the adaptive properties of gd T cells are
induced by their innate functions in a spatially and temporally controlled manner. The Journal of Immunology, 2012, 188: 1708–
1716.
Professional APCs have the hallmark properties of takingup soluble Ag, or of sampling Ags from target cells byphagocytosis, and then presenting processed Ags in the
context of MHC class II, or cross-presenting in the context of MHCclass I, to induce primary immune responses in naive Ag-specificab T cells. Dendritic cells (DCs), the prototypic professionalAPC, take up Ags at sites of infection, injury, or cancer and un-dergo a maturation step involving upregulation of costimulatorymolecules prior to migrating to sentinel lymph nodes. In thissetting, an interaction with CD40L on the surface of CD4-positiveT cells results in CD40 signaling in the DC, which is required foracquisition of full Ag presentation function, a process referred toas licensing (1–3).gd T cells typically represent 1–5% of T lymphocytes in human
blood. In contrast to conventional ab T cells, gd T cells expressa limited repertoire of TCR V-region genes; the most commonsubtype in human blood is Vg9Vd2, which recognizes a groupof nonpeptide phosphoantigens, for example isoprenyl pyrophos-phate (IPP), associated with infection or cancer (4, 5). Stimulatedgd T cells undergo an activation, which results in a plethora of
poorly defined changes including proliferation, proinflammatorycytokine and chemokine secretion, and altered cell surface phe-notype (6). Activated human peripheral blood gd T cells also haveproperties of innate immunity, including cytotoxicity mediatedby NK receptors (7–11), Ab-dependent cell-mediated cytotoxicity(ADCC) (12, 13), and phagocytosis (14). Notably, we and othershave shown FcgRIII (CD16) expressed on at least some human gdT cells to be a potentially important molecule mediating innateeffector functions (15, 16). These changes are likely to be subtypespecific along classical T cell divisions; that is, T naive, T centralmemory, T effector memory, and T effector memory RA+ (16).At least one subtype of human gd T cells has been shown to be
able to express MHC class II, costimulatory molecules, and lymphnode-homing chemokine receptors (e.g., CCL7) and has beencharacterized as a professional APC (17, 18). Previous studieshave thus demonstrated the ability of gd T APC to present andcross-present Ags from soluble protein (17–19), and we and othershave shown that human blood gd T cells can take up large frag-ments from intact cells by the processes of phagocytosis (14) ortrogocytosis (20). Also, we found that Ags associated with theselarge (.1 mm) structures can be processed and presented, boundto MHC class II, by gd T cells to other T cells (14). We thereforehypothesized that when activated gd T cells are cocultured withtransformed cells, the tumor cells would serve not only as targetsfor gd T-mediated killing but also as a source of tumor-associatedAgs (TAAs) for processing and professional presentation by thegd T cells to tumor-reactive ab T cells. In this study, we show thathuman gd T cells do indeed present TAAs from tumor cells.Moreover, we have found that the process of APC function istightly regulated, requiring interaction with cell surface-bound Ab,a requirement we refer to as licensing of gd T cells for profes-sional APC function.
Materials and MethodsCell isolation
Fresh peripheral blood was centrifuged over Ficoll-Hypaque to obtainPBMCs according to the local ethical guidelines on experimentation withhuman samples. Vg9Vd2 T cells were isolated by two rounds of purifi-cation using anti-gd TCR (MicroBead Kit, which consists of a hapten-conjugated anti-TCRg/d Ab and FITC-conjugated anti-hapten microbe-
*Molecular Haematology and Cancer Biology, University College London Instituteof Child Health, London WCN1 1EH, United Kingdom; †Confocal Microscopy CoreFacility, University College London Institute of Child Health, London WCN1 1EH,United Kingdom; ‡Molecular Immunology, University College London Institute ofChild Health, London WCN1 1EH, United Kingdom; and xLaboratory of TumorImmunology and Immunotherapy, Bar-Ilan University, Ramat-Gan 52900, Israel
1K.G. and J.A. contributed equally to this work.
Received for publication September 14, 2011. Accepted for publication December 7,2011.
This work was supported by grants from Research in Childhood Cancer, the NationalInstitutes of Health, the Wellcome Trust, and Children with Leukaemia.
Address correspondence and reprint requests to Dr. John Anderson or Kenth Gus-tafsson, Molecular Haematology and Cancer Biology, University College LondonInstitute of Child Health, 30 Guilford Street, London WCN1 1EH, United Kingdom(J.A.) or Molecular Immunology, University College London Institute of ChildHealth, 30 Guilford Street, London WCN1 1EH, United Kingdom (K.G.). E-mailaddresses: [email protected] (J.A.) and [email protected] (K.G.)
The online version of this article contains supplemental material.
Abbreviations used in this article: ADCC, Ab-dependent cell-mediated cytotoxicity;DC, dendritic cell; IPP, isoprenyl pyrophosphate; TAA, tumor-associated Ag.
Copyright� 2012 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/12/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1102654
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

ads). gd T APCs were generated from purified gd T cells via stimulationfor 24 h with 50 mM IPP (Sigma) in the presence of 200 U/ml IL-2.
Cell lines and humanized Abs
Daudi and Raji (B lymphoma) and K562 (erythroleukemia) cell lineswere maintained in RPMI 1640. Neuroblastoma cell line Lan-1 cells weremaintained in culture with DMEM (BioWhittaker, Walkersville, MD).SK-Mel-23 cells were grown as monolayer cultures in F10–Ham medium,and SK-Mel-28 melanoma cell lines were grown in RPMI 1640 medium.All media were supplemented with 10% FBS and 2 mmol/l L-glutamine, inthe absence of antibiotics, at 37˚C and 5% CO2 humidified atmosphere.All cell lines were mycoplasma-free. Humanized mAbs, anti-GD2, IgG1(Hu14.18K322A), anti-CD45RB, IgG1 (Thermo Scientific, Loughborough,U.K.), and anti-CD20 IgG1 (rituximab; Roche Pharma) were used at10 mg/ml to opsonize the cell lines.
Ag cross-presentation assays
Purified gd T cells were IPP activated for 24 h and cocultured for 5 d in thepresence of 200 IU/ml IL-2, with opsonized tumor cells and CFSE-labeledab T cells at a ratio of 10:10:1 (gd T cells/ab T cells/tumor cells) in thepresence of 70-Gy–irradiated HLA-A22 EBV–transformed B cells at a gdT cell/feeder cell ratio of 5:1, or in presence of conditioned media preparedfrom B-lymphoblastoid cell lines. The APCs were washed extensively be-fore culture with responder cells. When required, tumor Ag was added in thecoculture either as Daudi lysate (50 mg/ml), SK-Mel 23 lysate (50 mg/ml),recombinant PAX5 protein at 20 mg/ml (Caltag Medsystem, Milton Keynes,U.K.), or peptides PAX5 (TLPGYPPHV), PRAME (VLDGLDVLL), orMart-1 (ELAGIGILTV) (ProImmune, Oxford, U.K.). All peptides were usedat 5 mg/ml, and IL-2 (200 IU/ml) was added at day 3. In Transwellexperiments, cells were separated by a 1-mm pore-size membrane. Theupper chamber was carefully taken from the well, cells were harvested,washed, and FACS analyzed for CFSE dilution. For the intracellular cy-tokine staining, surface staining was performed by incubating cells withPE-conjugated pentamer raised against HLA-A*0201–restricted epitopeTLPGYPPHV derived from PAX5 (ProImmune). Cells were then per-meabilized in 100 ml Cytofix/Cytoperm solution (BD Biosciences) at 4˚Cfor 20 min, washed with Perm/Wash buffer (BD Biosciences), and fol-lowed by intracellular staining with allophycocyanin-conjugated anti-human IFN-g Ab (BD Biosciences) at 4˚C for 30 min. Intracellular se-cretion of IFN-g was assessed, with gating on the PAX5 ab T clonepentamer-positive cells.
In some experiments (see Fig. 3E), gd T cells were IPP-activated for 24h, cocultured with opsonized Lan-1 tumor cells, bead sorted, then rested ornot in 50 IU/ml IL-2 for 24 h before reincubation with opsonized Lan-1 andresponder ab T cells in the presence of Daudi lysate as described earlier.
In blocking experiments, gd T cells were blocked with a mousemonoclonal IgG1 anti-CD16 blocking Ab (clone LNK16; Abcam) orHLA-A2 neutralizing mAb (clone BB7.2, purified from hybridoma;American Type Culture Collection) or nonspecific mouse IgG1 at 10 mg/ml for 30 min at 37˚C/5% CO2 before pulsing with Daudi lysate. In theexperiment comparing DCs and gd T APCs, monocyte-derived DCs werederived from monocytes purified with CD14 microbeads (Miltenyi) andcultured for 6–7 d with 50 ng/ml GM-CSF and 10 ng/ml IL-4 (Peprotech).Mature DCs were pulsed with either Mart-1 peptide, irrelevant PRAMEpeptide, or SK-Mel-23 tumor cell lysate for 2 h, then washed and added tothe coculture with CFSE-labeled ab T cells. In any individual experiment,the numbers of APCs, tumor targets, and T cells were kept constant.
ab CTL clones and ab T cells transduction with a MART-1TCR-expressing retrovirus
High-avidity CTL clones raised against HLA-A2–restricted epitopes,TLPGYPPHV derived from PAX5 and VLDGLDVLL derived fromPRAME, have been previously described (21, 22). To generate Ag-specificbut Ag-inexperienced T cells, we transduced healthy donor PBMCs witha retrovirus expressing MART-1 TCR (containing a Vb12 chain) specificto an HLA-A*0201–restricted epitope, M26-35 (ELAGIGILTV). Briefly,TCR a- and b-chains were isolated from a highly lytic HLA-A2–restrictedCTL clone recognizing the melanoma-associated Melan-A/MART-1 Agand inserted into a retrovirus vector capable of robust and coordinatedexpression of the transgene. The retroviral vector and packaging constructpCL-Ampho (Imagenex) were cotransfected into 293 T cells using Lipo-fectamine 2000 transfection reagent (Invitrogen) according to the manu-facturer’s instructions. Culture supernatants containing retroviruses wereharvested twice at 48 and 72 h after transfection and were concentratedprior to transduction. PBMCs from HLA-A*0201 healthy donors werestimulated in vitro at 1 3 106 cells/well with Dynabeads coated with anti-
CD3, anti-CD28, and anti-CD137 stimulating Abs in the presence of 300IU/ml IL-2 for 3 d. T cell cultures were then transduced as follows: 24-wellplates were coated with RetroNectin (TaKaRa, Otsu, Japan), then pre-loaded with retrovirus according to the manufacturer’s instructions. T cells(2.6 3 106) were added to each well in 1.3 ml Mart-1 TCR retroviralsupernatant, supplemented with 300 IU/ml recombinant human IL-2, andthe plates were centrifuged for 90 min at 1000 3 g. The next day, themedium was replaced with fresh retroviral supernatant, and the centri-fugation was repeated. Transduction efficiency and TCR expression onT cells were confirmed by flow cytometry using 10 mg/ml anti-TCR Vb12(Immunotech). The cells were rested for 24 h, after which transduced cellswere CFSE labeled and assayed for Ag reactivity by 5-d coculture withDCs or with activated gd T cells in the presence of opsonized Lan-1 tumorcells and Ag (SK-Mel-23 tumor lysate, Mart-1 peptide [ELAGIGILTV], orirrelevant peptide). IL-2 was added at day 3.
Confocal microscopy
Opsonized Daudi cells were labeled either with Vybrant DiI (Invitrogen,U.K.) or CFSE and incubated with IPP-activated gd T cells stained withmouse anti-human TCRgd (BD Biosciences, Heidelberg, Germany) andsecondary Ab goat anti-mouse Cy5 (Caltag, U.K.), or Alexa Fluor 488phalloidin (Invitrogen, Grand Island, NY) before being immediately ana-lyzed. Images were captured using LSM 510 confocal microscopy (Zeiss)with Leica confocal software.
Cytotoxicity assays
Freshly isolated or activated gd T cells were used as effector cells. Targetcells were coated or not with the opsonizing mAb and labeled with 100mCi Na2
51CrO4 in cell culture and tested in a standard chromium releaseassay as previously described.
Activation phenotype of gd T after coculture
gd T cells were activated by coculture with lymphoblastoid cell lineconditioned media in the presence of 100 IU/ml recombinant human IL-2and 50 mm IPP and cultured with opsonized or non-opsonized Daudi orRaji cell line for 24 h. gd T cell phenotype was analyzed by FACS usingthe following mAbs: monoclonal mouse IgG2 PE-conjugated anti–HLA-DR, clone L243 (BD Biosciences); monoclonal mouse IgG1 PE-conjugated anti-CD86, clone FUN-1; anti-CD11c–allophycocyanin, cloneB-ly6; anti-CD40–PE–Cy5, clone 5C3; and CD80–FITC, clone L307.4(BD Biosciences). To characterize the gd TCR phenotype, we used anti-Vg9–PE–Cy5 (Beckman-Coulter) to stain activated Vg9Vd2 T cells anda pan anti-gd T–biotin clone B1 (BD Pharmingen).
ResultsCross-presentation of TAA by human blood gd T cells
As a model to study the capacity of human gd T cells to interactwith human cancer cells, we used immunomagnetic positive se-lection from peripheral blood, using a pan-gd TCR Ab. Using thismethod, yields of gd T cells were more than 98% pure (Fig. 1A),and percentage contamination with CD11c-positive cells was be-tween 0.3 and 0.6% (Fig. 1B). As a tumor model, we initially usedthe Burkitt lymphoma cell line Daudi because we (Fig. 1C) andothers (12) showed that opsonization with anti-CD20 IgG1 mAb(rituximab) enhances ADCC of Daudi cells by gd T cells, andbecause we showed that activated human blood gd T cells couldtake up fragments from opsonized Daudi cells (Fig. 1D). In ad-dition, we reasoned that opsonized Daudi cells could, in thismodel, serve as a source of TAA for processing and presentationon gd T MHC to other T cells in a rituximab-dependent manner.To test this, we investigated cross-presentation of PAX5 Ag, anuclear transcription factor expressed in B lineage cancer cellsincluding Daudi (22, 23), by gd T MHC class I. We have previ-ously described a CD8 CTL clone specific for the PAX5 peptidesequence TLPGYPPHV and restricted to HLA-A*0201 (22). Wetested the ability of gd T cells to present PAX5 Ag from intacttumor cells in a 5-d coculture involving 1) IPP-activated gd T cellsfrom an HLA-A*0201–positive donor, 2) rituximab-opsonizedHLA-A*0201–negative Daudi cells, and 3) CFSE-labeled abCTL clone restricted to PAX5/HLA-A*0201 in the presence of
The Journal of Immunology 1709
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

conditioned medium from a B lymphoblastoid cell line (Fig. 2A;see Supplemental Fig. 1 for lymphocytoid cell line detail). Robustproliferation of the ab clone was individually dependent on gdT cells and opsonized Daudi cells and was Ag- and MHC-restricted as demonstrated through use of an ab CTL clone spe-cific for HLA-A0201/PRAME (not expressed on Daudi cells) (Fig.2B) and further demonstrated by lack of proliferation of the PAX5clone when preincubating the gd T cells with an HLA-A0201blocking Ab (Fig. 2C). To exclude the possibility that contami-nating professional APCs in the gd T cell preparations were re-sponsible for the observed stimulation, we titrated cells from thenegative fraction after gd TCR selection back into a cocultureinvolving non-HLA-matched gd T cells. No significant prolifer-ation was observed until the number of negative fraction cellsreached 20% (Supplemental Fig. 2). To confirm the interactionbetween IPP-activated gd T cells and rituximab-opsonized Daudicells, we used live imaging in which the gd TCR was labeled witha fluorescent Ab and was found to form an aggregate at the im-mune synapses with Daudi cells. Moreover, a significant numberof gd T cells (8–13%) were found to be interacting with Daudicells at any time point (Fig. 2D). By the end of the 5-d coculture inthe presence of IPP, the gd T population was found to be exclu-sively of the Vg9 subtype (Fig. 2E). Therefore, we conclude thatthe coculture of activated Vg9Vd2 T cells with intact tumor cellsexpressing an intracellular Ag can result in the MHC class I-restricted Ag-specific proliferation of an ab CTL clone.
Ab-dependent licensing of gd T APC by direct cell–cell contact
Wewanted to compare the cross-presentation function of human gdT cells after culture with either Ag-expressing intact tumor cells or
with soluble tumor Ag. We also were interested in whether acti-
vation of gd T for cross-presentation could cause bystander acti-
vation through release of soluble factors. To this end, we used
Transwells with a 1-mm pore size restricting free diffusion to
soluble proteins or cell fragments. In the presence of opsonized
Daudi cells, gd T cells could, as expected, cross-present PAX5 Ag
to ab CTL in an upper chamber, but the ab CTL clone in the
communicating lower chamber was only modestly stimulated
(Fig. 3A). Similarly, a low level of activation of the CTL clone was
seen if Daudi cells were not opsonized (Fig. 3B, upper panel)
compared with the negligible background activation seen in the
absence of Ag (Fig. 3B, lower panel).Previous studies have demonstrated convincing Ag presenta-
tion to both CD4 and CD8 naive ab T cells by activated human
gd T cells loaded with soluble whole protein Ags (17–19). We
hypothesized that the relatively weak APC function we observed
in the absence of opsonized target cell was being greatly enhanced
as a result of the interaction with Ab-coated cells. To test this
hypothesis, we cultured activated gd T cells and the PAX5 ab
CTL clone with a lysate from Daudi cells in the presence or ab-
sence of a third-party opsonized target cell (Lan-1 neuroblastoma
cells, PAX5 nonexpressing, coated with anti-GD2 IgG1 mAb).
FIGURE 1. Cytotoxic properties of highly purified
activated human blood gd T cells. A, Greater than 98%
purity was obtained after two rounds of positive se-
lection using a pan-gd TCR Ab. B, Purified cells had
insignificant staining for CD11c. In A and B, data are
representative of ∼16 gd TCR preparations from dif-
ferent donors used in this study. C, Overnight chro-
mium release cytotoxicity assay involving gd T cells
activated with IPP or unactivated, and Daudi cells
opsonized with rituximab or non-opsonized. Error bars
are SEM of triplicate readings from a single repre-
sentative experiment. D, Confocal microscopy after
coculture of purified and fluorescently labeled cell
populations. gd T cells were activated with IPP and
stained with a red fluorochrome, and Daudi cells were
pretreated with rituximab and labeled green. Frag-
ments from Daudi cells were seen within interacting
cells. Data is representative of three independent ex-
periments.
1710 gd T CELL LICENSING BY Ab
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Whereas the presence of opsonized target Ag-negative cells in-duced a robust proliferation, we consistently observed only weakCTL response in the absence of opsonized cells (Fig. 3C, leftpanel). A control condition, in which Ab was omitted, resulted inloss of the profound APC function to the level seen in the lowerchamber with Ag alone (Fig. 3C, middle panel). We refer to thisdependency on direct contact with an Ab-coated target cell forinduction of full APC function of gd T cells as “licensing.”Consistent with previous reports (17), proliferation and IFN-gsecretion in the presence of Ag but absence of Ab was consistentlygreater than the background proliferation (Fig. 3C, right panel).We excluded the involvement of soluble factors from Daudi cellsin licensing by substituting recombinant PAX5 protein for theDaudi lysate (Supplemental Fig. 3).To demonstrate this licensing concept in a further cell system,
we substituted K562 erythroleukemia cells coated with an anti-CD45 IgG1 Ab as the licensing cell and found an equivalentand “opsonized cell”-dependent proliferation and IFN-g response(Fig. 3D). Notably, gd T cells that had been licensed by opsonizedLan-1 and then repurified from the licensing conditions and rested
overnight had lost the ability to take up and present soluble PAX5Ag, whereas if these licensed and purified cells were relicensed bycoculture with opsonized Lan-1, they fully regained APC function(Fig. 3E, right panel).
Raji cells fail to license gd T cells
Because Daudi cells are well known to be efficiently killed by gdT cells, we sought to investigate whether another very similarcell line, but one that is resistant to gd T cell killing, can alsolicense gd T cells for more potent APC function. To do this, weused Raji cells, which others have described as being more re-sistant to gd T cell cytotoxicity (24). We first confirmed thatactivated HLA-A*0201–positive gd T cells are poor killers ofanti-CD20–opsonized Raji targets. Notably, in contrast to Daudicells, Raji cells failed to induce any APC function of gd T cellsin a 5-d coculture setting (Fig. 4A). We then sought to determinewhether the failure of the induction of APC function of gd T cellswas the consequence of inadequate PAX5 Ag release from Rajicells. To do this, we repeated the 5-d coculture experiment usingopsonized Raji cells in the presence of Daudi lysate as a source
FIGURE 2. Human blood gd T cells cross-present intracellular Ag from an opsonized tumor cell. A, Cartoon schematic of three-cell, 5-d coculture used
in these experiments. B, Ag-specific and MHC-restricted stimulation of an ab CD8 T cell clone disclosed by CFSE dilution measured by flow cytometry.
Data representative of six independent donors. C, Inhibition of ab T cell clone as defined by CFSE dilution and IFN-g secretion measured by flow
cytometry was inhibited by pretreating gd T cells with an HLA-A*0201 blocking Ab. D, Confocal imaging of the three-cell coculture. gd TCR was
visualized on gd T cells with a magenta-labeled anti-pan gd TCR Ab, opsonized Daudi cells labeled in red, and ab T cell clone labeled in green. The
magenta TCR stain was found to localize to a discrete region of the membrane. The lower panel is a magnified portion of the boxed region of the upper. E,
gd T cells after 5-d coculture stained homogeneously positively with a Vd9 specific Ab. Results in panels B and C are representative of three independent
experiments, and panels D and E are representative data of two separate experiments. Percentages shown in the FACS plots are the means of the percentages
of viable positive cells from three experiments yielding similar results.
The Journal of Immunology 1711
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

FIGURE 3. Human blood gd T cells are licensed for Ag cross-presentation by interaction with opsonized cells. A, ab T cell clone activation determined
by CFSE dilution and IFN-g release in top and bottom chambers of a 1-mm-pore Transwell experiment. Daudi targets were opsonized and repeatedly
washed before been added to the top chamber. B, Controls for the three-cell coculture protocol depicted in Fig. 2 in which Daudi cells alone or rituximab
alone were used as the licensing agent. C and D, Five-day coculture 1-mm-pore Transwell experiment set up as depicted in the scheme. E, Human gd T cells
were activated with IPP in the absence of PAX5 Ag and then licensed with Lan-1 cells opsonized with anti-GD2 mAb for 2 d before purifying with two
rounds of positive selection. Licensed gd T cells were then rested for 24 h in the presence of IL-2. Twenty-four-hour rested gd T cells were cocultured with
labeled ab CTL responder clone in the absence (central panel) or presence (right panel) of anti-GD2 opsonized Lan-1 cells. Positive control (left panel)
were non-sorted licensed gd T cells cocultured with responders and Ag. Typical FACS plots are shown from three different experiments performed in
triplicate with two different donor cells each, and percentages shown in the FACS plots are the means of the percentages of viable positive cells from three
experiments yielding similar results.
1712 gd T CELL LICENSING BY Ab
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

of PAX5 Ag. Despite the presence of high amount of solublePAX5 Ag, opsonized Raji cells failed to license completely gd
T cells for APC function (Fig. 4B). Surprisingly, both opsonizedand non-opsonized Daudi and Raji cells induce a strong upreg-ulation of MHC class II (DR), CD40, and costimulatory mole-cules (Fig. 4C) suggesting that the licensing of gd T cells forAg presentation and the upregulation of molecules required forAPC function are distinct events. Thus, Raji cells do not onlyresist gd T cell killing but also fail to license them, independentlyof MHC class II and costimulatory molecule induction on the gdT cells.
Licensed gd T cells can present TAAs to TCR-transduced naiveprimary T cells
A hallmark of professional APCs is the ability to stimulate naiveCTL. To generate a population of Ag-inexperienced human T cellsof a single antigenic specificity, we transduced T cells from an
HLA-A*0201–positive donor with a retrovirus encoding a TCRwith specificity for an immunodominant HLA-A*0201–restrictedpeptide from the melanoma Ag MART-1 (Fig. 5A). CFSE-labeledMART-1 TCR-transduced T cells were cocultured with either gdT cells (licensed by the presence of anti-GD2–opsonized Lan-1cells) or with conventional DCs, both derived from the sameHLA-A*0201–positive donor. The Ag was either MEL-23 mela-noma cell line lysate or soluble MART-1 peptide. Licensed gdT cells were found to present either Ag to the same degree asmonocyte-derived DCs (Fig. 5B). Opsonization of target cells wasmandatory to license gd T cells for potent APC function (Fig. 5C).We speculated that interaction of gd cells with Fc receptors mightbe critical for the licensing phenomenon and therefore stainedactivated gd T cells with Abs against the Fcg receptors CD16,CD32, and CD64, of which only CD16 was positive (data notshown). We then found that preincubating gd T with a blockingAb directed against the FcgRIII (CD16) abrogated the licensing
FIGURE 4. Raji target cells do not license gd T cells. A, Four-hour chromium release cytotoxicity assay involving gd T cells activated with IPP or
unactivated and cultured with Raji or Daudi target cells opsonized with rituximab or non-opsonized. Data are the mean of triplicate readings from a single
representative experiment. B, ab T cell clone activation determined by CFSE dilution and IFN-g release in 5-d coculture incubation in which rituximab-
coated Raji cells were used as the licensing target cells in the presence (right panel) or absence (left panel) of Daudi lysate (PAX5 Ag source). Repre-
sentative FACS plots are shown. C, Immunophenotype of purified human blood gd T cells after 5-d coculture with opsonized or non-opsonized target cells
(Daudi or Raji) in the presence or absence of IPP. The mean of triplicate readings from a single representative experiment are shown. Each experiment was
performed at least twice yielding similar results.
The Journal of Immunology 1713
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

effect (Fig. 5D). Therefore, the acquisition of full APC function inhuman peripheral blood gd T cells requires a licensing event thatinvolves a physical interaction with an Ab-coated target cell, andour data are consistent with a CD16–Fc interaction being essentialfor licensing.
DiscussionWe have confirmed the finding of previous studies that, givenappropriate stimulation, phosphoantigen-activated human bloodgd T cells, which overwhelmingly express the Vg9Vd2 TCR,can take up and cross-present exogenous Ag to Ag-experienced
FIGURE 5. Human blood gd T cells are licensed by opsonized target cells to prime Ag-inexperienced ab T cells. A, A population of human CTL
expressing a single TCR with specificity for an HLA-A0201–restricted peptide from MART-1 was obtained by retroviral transduction. B, DCs and gd
T cells were obtained from the same HLA-A0201–positive donor and cocultured as depicted for 5 d. gd T cells were licensed in the presence of opsonized
Daudi cells. C, Controls for the experiment in B showing that omission of Ag or licensing results in loss of stimulation. D, Coculture with responders in
licensing conditions in which gd T cells were pretreated with a CD16 blocking Ab or isotype control Ab. Data are representative of two independent
experiments from different donors. In B–D, proliferation of Ag-specific cells was measured after gating on the Vb12 transduced population. Percentages
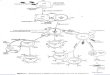
shown in the FACS plots are the means of the percentages of viable positive cells from three experiments yielding similar results. E, Possible physiological
licensing and Ag presentation functions of gd T cells.
1714 gd T CELL LICENSING BY Ab
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

or inexperienced ab T cells. For the first time to our knowledge,we show that this process is subject to regulation, and we haveidentified one mechanism of likely important physiological sig-nificance in the host response to cancer. We have reproduciblyseen, using two different antigenic systems and three different“licensing” opsonized cells, that the degree of Ag-specific stim-ulation of responder ab T cells is massively increased in thepresence of opsonized cells. In the presence of Ab-coated cells,the degree of stimulation is equivalent to that seen with matureAg-loaded DCs, whereas in the absence of licensing, a moremodest degree of stimulation above background is apparent.Physiologically, we propose that the requirement for Ab coating
of target cells provides an important regulatory circuit betweeninnate and adaptive immunity, which will localize functional an-tigenic sampling of gd T cells to their site of activation (Fig. 5D).Hence in the cancer models demonstrated here, activation of cir-culating gd T cells at the tumor site by TCR ligand will activatetheir expansion and innate killing. However, full APC functionwill only be engaged after the binding of Ig to tumor cells. This inturn will both activate gd T-mediated killing (by ADCC) (Fig. 1C)and will license the cells for Ag presentation. It is not yet clear,however, whether gd T cells kill their target in the process ofbeing licensed. In this regard, it is of interest that there is a cor-relation between sensitivity to cytotoxic killing (in Daudi and Rajicells, respectively) and licensing ability. This differential sensi-tivity to killing has been reported previously, but the precisemechanism is not known although is speculated to be related toexpression of an unknown ligand for Vg9Vd2 TCR (25). Thedifferential expression of F1-ATPase related protein (putative li-gand for Vg9Vd2 TCR) on Daudi and Raji cells might alsocontribute to the differential sensitivity (26). In support of thephysiological significance of this concept in cancer are datadocumenting infiltration of gd T cells into a broad array of tumortypes (27–29), which when analyzed have been found to be pre-dominantly of the Vg9Vd2 subtype (7, 9, 30, 31).Licensing of DCs by CD40 ligation is a distinct process from
their maturation, the latter involving upregulation of costimulatorymolecules, MHC class II, and chemokine receptors. Analogously,for IPP-activated gd T cells, we found that upregulation of co-stimulatory molecules and HLA-DR was dependent on interactionwith tumor target cells but independent of interaction with Ab.Moreover, activation via ligation of the TCR was not sufficient forfull upregulation of costimulatory molecules or HLA-DR (Fig.4C). The signaling that results in upregulation of costimulatorymolecules on gd T cells is not known, nor is it known whether itrequires direct interaction with and/or killing of a target cell.Cells with combined innate killing and Ag presentation function
are well known; for example, macrophages. We have previouslydemonstrated the capacity of gd T cells to take up and present Ags,using phagocytosis (14), and gd T cells also take up Ag by tro-gocytosis (20). Hence, at the tumor site, gd T cells have the po-tential to kill by innate cytotoxic mechanisms releasing Ag intosolution and to take up Ags directly by phagocytosis, trogocytosis,or pinocytosis. Ab coating of target cells potentially enhances allthese processes but additionally provides a distinct licensing sig-nal for potent activation of tumor-reactive CTL.It is interesting to speculate how licensing could regulate APC
function spatially and temporally within the setting of response tocancer or infection. Previous data have demonstrated that Vg9Vd2TCR activation induces upregulation of secondary lymphoid tissuechemokines CCR7 and CCR4 and enhanced lymphoid tissue-homing capabilities, as well as clustering within germinal centersof B cell follicles (32). However, regardless of the role of gdT licensing in physiological systems, the availability of clinical
reagents for gd T activation and therapeutic mAbs to effect li-censing in the tumor niche suggests new approaches toward Ag-specific vaccination/treatment for cancer and infection.
DisclosuresThe authors have no financial conflicts of interest.
References1. Bennett, S. R., F. R. Carbone, F. Karamalis, R. A. Flavell, J. F. Miller, and
W. R. Heath. 1998. Help for cytotoxic-T-cell responses is mediated by CD40signalling. Nature 393: 478–480.
2. Schoenberger, S. P., R. E. Toes, E. I. van der Voort, R. Offringa, and C. J. Melief.1998. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40Linteractions. Nature 393: 480–483.
3. Ridge, J. P., F. Di Rosa, and P. Matzinger. 1998. A conditioned dendritic cell canbe a temporal bridge between a CD4+ T-helper and a T-killer cell. Nature 393:474–478.
4. Morita, C. T., C. Jin, G. Sarikonda, and H. Wang. 2007. Nonpeptide antigens,presentation mechanisms, and immunological memory of human Vgamma2V-delta2 T cells: discriminating friend from foe through the recognition of prenylpyrophosphate antigens. Immunol. Rev. 215: 59–76.
5. Gober, H. J., M. Kistowska, L. Angman, P. Jeno, L. Mori, and G. De Libero.2003. Human T cell receptor gammadelta cells recognize endogenous mevalo-nate metabolites in tumor cells. J. Exp. Med. 197: 163–168.
6. Ribot, J. C., A. debarros, and B. Silva-Santos. 2011. Searching for “signal 2”:costimulation requirements of gd T cells. Cell. Mol. Life Sci. 68: 2345–2355.
7. Viey, E., G. Fromont, B. Escudier, Y. Morel, S. Da Rocha, S. Chouaib, andA. Caignard. 2005. Phosphostim-activated gamma delta T cells kill autologousmetastatic renal cell carcinoma. J. Immunol. 174: 1338–1347.
8. Das, H., V. Groh, C. Kuijl, M. Sugita, C. T. Morita, T. Spies, and J. F. Bukowski.2001. MICA engagement by human Vgamma2Vdelta2 T cells enhances theirantigen-dependent effector function. Immunity 15: 83–93.
9. Corvaisier, M., A. Moreau-Aubry, E. Diez, J. Bennouna, J. F. Mosnier, E. Scotet,M. Bonneville, and F. Jotereau. 2005. V gamma 9V delta 2 T cell response tocolon carcinoma cells. J. Immunol. 175: 5481–5488.
10. Rincon-Orozco, B., V. Kunzmann, P. Wrobel, D. Kabelitz, A. Steinle, andT. Herrmann. 2005. Activation of V gamma 9V delta 2 T cells by NKG2D. J.Immunol. 175: 2144–2151.
11. Maniar, A., X. Zhang, W. Lin, B. R. Gastman, C. D. Pauza, S. E. Strome, andA. I. Chapoval. 2010. Human gammadelta T lymphocytes induce robust NK cell-mediated antitumor cytotoxicity through CD137 engagement. Blood 116: 1726–1733.
12. Gertner-Dardenne, J., C. Bonnafous, C. Bezombes, A. H. Capietto, V. Scaglione,S. Ingoure, D. Cendron, E. Gross, J. F. Lepage, A. Quillet-Mary, et al. 2009.Bromohydrin pyrophosphate enhances antibody-dependent cell-mediated cyto-toxicity induced by therapeutic antibodies. Blood 113: 4875–4884.
13. Tokuyama, H., T. Hagi, S. R. Mattarollo, J. Morley, Q. Wang, H. F. So,F. Moriyasu, M. Nieda, and A. J. Nicol. 2008. V gamma 9 V delta 2 T cellcytotoxicity against tumor cells is enhanced by monoclonal antibody drugs—rituximab and trastuzumab. Int. J. Cancer 122: 2526–2534.
14. Wu, Y., W. Wu, W. M. Wong, E. Ward, A. J. Thrasher, D. Goldblatt, M. Osman,P. Digard, D. H. Canaday, and K. Gustafsson. 2009. Human gamma delta T cells:a lymphoid lineage cell capable of professional phagocytosis. J. Immunol. 183:5622–5629.
15. Wu, H. Y., Q. Wang, Y. J. Chen, F. Xie, Y. X. Mao, G. Q. Liu, and X. G. Zhang.2007. [Preparation and characterization of two novel functional monoclonalantibodies against human OX40L]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 23:757–760.
16. Angelini, D. F., G. Borsellino, M. Poupot, A. Diamantini, R. Poupot,G. Bernardi, F. Poccia, J. J. Fournie, and L. Battistini. 2004. FcgammaRIIIdiscriminates between 2 subsets of Vgamma9Vdelta2 effector cells with dif-ferent responses and activation pathways. Blood 104: 1801–1807.
17. Brandes, M., K. Willimann, and B. Moser. 2005. Professional antigen-presentation function by human gammadelta T Cells. Science 309: 264–268.
18. Brandes, M., K. Willimann, G. Bioley, N. Levy, M. Eberl, M. Luo, R. Tampe,F. Levy, P. Romero, and B. Moser. 2009. Cross-presenting human gammadeltaT cells induce robust CD8+ alphabeta T cell responses. Proc. Natl. Acad. Sci.USA 106: 2307–2312.
19. Meuter, S., M. Eberl, and B. Moser. 2010. Prolonged antigen survival and cy-tosolic export in cross-presenting human gammadelta T cells. Proc. Natl. Acad.Sci. USA 107: 8730–8735.
20. Poupot, M., F. Pont, and J. J. Fournie. 2005. Profiling blood lymphocyte inter-actions with cancer cells uncovers the innate reactivity of human gamma deltaT cells to anaplastic large cell lymphoma. J. Immunol. 174: 1717–1722.
21. Yan, M., N. Himoudi, B. P. Basu, R. Wallace, E. Poon, S. Adams, F. Hasan,S. A. Xue, N. Wilson, A. Dalgleish, et al. 2011. Increased PRAME antigen-specific killing of malignant cell lines by low avidity CTL clones, followingtreatment with 5-Aza-2’-Deoxycytidine. Cancer immunology, immunotherapy:CII. 60: 1243–1255.
22. Yan, M., N. Himoudi, M. Pule, N. Sebire, E. Poon, A. Blair, O. Williams, andJ. Anderson. 2008. Development of cellular immune responses against PAX5,a novel target for cancer immunotherapy. Cancer Res. 68: 8058–8065.
The Journal of Immunology 1715
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

23. Torlakovic, E., A. Slipicevic, C. Robinson, J. F. DeCoteau, G. C. Alfsen,M. Vyberg, R. Chibbar, and V. A. Flørenes. 2006. Pax-5 expression in non-hematopoietic tissues. Am. J. Clin. Pathol. 126: 798–804.
24. L’Faqihi, F. E., M. Guiraud, N. Dastugue, P. Brousset, P. Le Bouteiller, F. Halary,M. Bonneville, J. J. Fournie, and E. Champagne. 1999. Acquisition of a stimu-latory activity for Vgamma9/Vdelta2 T cells by a Burkitt’s lymphoma cell linewithout loss of HLA class I expression. Hum. Immunol. 60: 928–938.
25. Thedrez, A., C. Sabourin, J. Gertner, M. C. Devilder, S. Allain-Maillet,J. J. Fournie, E. Scotet, and M. Bonneville. 2007. Self/non-self discrimination byhuman gammadelta T cells: simple solutions for a complex issue? Immunol. Rev.215: 123–135.
26. Scotet, E., L. O. Martinez, E. Grant, R. Barbaras, P. Jeno, M. Guiraud,B. Monsarrat, X. Saulquin, S. Maillet, J. P. Esteve, et al. 2005. Tumor recog-nition following Vgamma9Vdelta2 T cell receptor interactions with a surfaceF1-ATPase-related structure and apolipoprotein A-I. Immunity 22: 71–80.
27. Xu, C., H. Zhang, H. Hu, H. He, Z. Wang, Y. Xu, H. Chen, W. Cao, S. Zhang,L. Cui, et al. 2007. Gammadelta T cells recognize tumor cells via CDR3deltaregion. Mol. Immunol. 44: 302–310.
28. Groh, V., R. Rhinehart, H. Secrist, S. Bauer, K. H. Grabstein, and T. Spies.1999. Broad tumor-associated expression and recognition by tumor-derivedgamma delta T cells of MICA and MICB. Proc. Natl. Acad. Sci. USA 96:6879–6884.
29. Maeurer, M. J., D. Martin, W. Walter, K. Liu, L. Zitvogel, K. Halusczcak,H. Rabinowich, R. Duquesnoy, W. Storkus, and M. T. Lotze. 1996. Human in-testinal Vdelta1+ lymphocytes recognize tumor cells of epithelial origin. J. Exp.Med. 183: 1681–1696.
30. Nanno, M., H. Seki, G. Mathioudakis, R. Suzuki, K. Itoh, C. G. Ioannides,S. Suzuki, P. F. Chen, and C. D. Platsoucas. 1992. Gamma/delta T cell antigenreceptors expressed on tumor-infiltrating lymphocytes from patients with solidtumors. Eur. J. Immunol. 22: 679–687.
31. Bachelez, H., B. Flageul, L. Degos, L. Boumsell, and A. Bensussan. 1992. TCRgamma delta bearing T lymphocytes infiltrating human primary cutaneousmelanomas. J. Invest. Dermatol. 98: 369–374.
32. Brandes, M., K. Willimann, A. B. Lang, K. H. Nam, C. Jin, M. B. Brenner,C. T. Morita, and B. Moser. 2003. Flexible migration program regulates gammadelta T-cell involvement in humoral immunity. Blood 102: 3693–3701.
1716 gd T CELL LICENSING BY Ab
by guest on February 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from