Embed Size (px)
Citation preview

1
HƯỚNG NGHIÊN CỨU VÀ NHỮNG THÍ NGHIỆM
TRONG NGHIÊN CỨU CHĂN NUÔI
NHÀ XUẤT BẢN NÔNG NGHIỆP Hà Nội 2011

2
Dịch từ nguyên tác
Trails and Trials in Livestock Research
Của tác giả E.R Orskov
OBE, BSc, PhD, DSc, FRSE, FPAS International Feed Resource Unit
Macaulay Land Use Research Institute Craigiebuckler, Aberdeen AB15 8QH
Foarmerly of Rowett Research Institute
Bucksburn, Aberdeen AB21 9SB
Người dịch TS. Vũ Chí Cương
Nguyễn Thành Trung
Hiệu đính TS. Vũ Chí Cương

3
MỤC LỤC

4
Lời nói đầu
Có vẻ là hơi lạ khi viết thể loại lý lịch khoa học tự thuật như thế này nhưng
tôi cảm thấy nên làm như vậy! Bởi vì có rất nhiều điều tôi muốn nói với những nhà khoa học trẻ trong cuốn sách này bằng cách cho họ những ví dụ về các con đường nghiên cứu khoa học đã đi như thế nào. Những kỹ thuật mới có thể mở ra những chân trời mới đầy hấp dẫn đôi khi khác hoàn toàn với các mục tiêu ban đầu ra sao. Bằng cách nào từ vẻ ngoài của những thứ tẻ nhạt và buồn chán cuộc sống có thể bất chợt hiện và đầy hấp dẫn. Làm thế nào để nông dân có lợi từ chúng ta trong việc hiểu vấn đề tốt hơn và từ phát triển các kỹ thuật mà họ có thể áp dụng. Càng làm việc lâu trong ngành khoa học nông nghiệp, tôi càng cảm thấy rằng các khách hàng của tôi là người nông dân và tôi là đầy tớ của họ.
Khi tôi đọc danh sách khá dài các tài liệu của mình và đọc lại một số báo cáo quan trọng nhất tôi luôn cảm thấy có một số vấn đề hấp dẫn nhưng đã bị lãng quên. Trong khi không nghi ngờ gì nữa một bài báo khoa học có thể được tô điểm cho mầu mè một chút theo cách của tác giả, tuy nhiên không có chỗ cho các khía cạnh cá nhân ở đây. Điều gì dẫn đến một ý tưởng tốt, liệu có những khó khăn hay không? Thêm nữa, một loạt các bài báo khoa học về một chủ đề tương tự có thể được xuất bản ở các nơi khác nhau và không có đủ chỗ để làm nên một câu truyện hoàn chỉnh. Tôi đã nghiên cứu khoa học nông nghiệp hơn 30 năm. Tôi đã may mắn một cách lạ thường. Tôi được làm việc với cơ sở vật chất tốt và không phải mất nhiều thời gian lo lắng về kinh phí. Nếu như tôi không có đóng góp nào, riêng tôi đáng bị khiển trách. Cùng với các cộng sự, tôi nghĩ chúng tôi cũng đã đóng góp ít nhiều. Tuy nhiên trong cuốn sách nhỏ này, tôi muốn không chỉ mô tả những đóng góp cho các lĩnh vực khác nhau trong chăn nuôi gia súc nhai lại mà còn thêm vào đó những cảm nhận cá nhân của những người tham gia vào nghiên cứu kể cả bản thân tôi, các lãnh đạo của tôi, các đồng nghiệp và những người khác, nhằm làm cho câu chuyện thú vị hơn.
Tôi đã có may mắn lớn trở thành người lãnh đạo của một đơn vị nghiên cứu nơi mà tôi đã tận dụng được rất nhiều kinh nghiệm, sự động viên khuyến khích của các đồng nghiệp và sinh viên từ khắp nơi trên thế giới, nơi mà tôi có được sự hỗ trợ về cơ sở vật chất và danh tiếng của một Viện nghiên cứu nổi tiếng thế giới. Như vậy tôi gặp rất ít trở ngại trong chương trình nghiên cứu của chúng tôi, thay vào đó là các điều kiện thuận lợi cho việc nghiên cứu khoa học, và hy vọng điều này có giá trị thực tiễn.
Lời cám ơn
Rất nhiều người đã giúp đỡ và khuyến khích tôi trong công việc này. Tôi
muốn đặc biệt bày tỏ sự kính trọng sâu sắc tới tiến sĩ Kay về sự giúp đỡ trong việc

5
biên soạn, Tiến sĩ Mike Wilkinson ở Chalcombe Press và vợ tôi, Joan trong việc thiết kế trang bìa.
Một chút về bản thân
Tôi không có ý định làm nhàm chán người đọc với việc viết quá nhiều về
bản thân mình nhưng bởi vì mỗi dự án nghiên cứu tôi làm đều có câu chuyện riêng của nó, và ở một mức độ nào đó các dự án bị ảnh hưởng bởi câu chuyện của người kể do vậy tôi sẽ viết một vài dòng về bản thân mình.
Tôi sinh ra ở một nông trang nhỏ Miền Tây Jutland, Đan Mạch là con thứ 3 trong gia đình có 5 anh em trai và 7 chị em gái. Trang trại của gia đình tôi có 6 bò, 3 cừu và 2 ngựa. Khi tôi lên 9 tuổi, gia đình tôi mua một trang trại lớn hơn một chút với 12 bò, 4 cừu và 3 ngựa. Từ năm lên 7 đến năm lên 10, tôi đi học ở ngôi trường kiểu cũ, 2 ngày một tuần vào mùa đông và 4 ngày một tuần vào mùa hè và từ lúc lên 10 đến năm 14 tuổi là 2 ngày một tuần vào mùa hè và 4 ngày một tuần vào mùa đông. Lịch học sắp xếp như vậy để những đứa trẻ lớn hơn có thể tham gia nhiều hơn vào các công việc ở trang trại và ai cũng có việc làm kể cả tôi. Tôi dự định chỉ làm nông dân. Tôi rời trường học năm 14 tuổi và làm thuê kiếm sống ở một nông trang gần nhà trong 2 năm sau đó. Từ năm 16 đến 18 tuổi, tôi quản lý trang trại gia đình khi cha tôi ốm. Sau đó tôi làm người chăn nuôi bò sữa ở một trang trại xa nhà và khi 20 tuổi tôi đi nghĩa vụ quân sự ở Copenhagen. Đây là một cú sốc lớn về văn hóa đối với tôi! Tôi là một chú bé nông dân không có học với một chất giọng nhà quê bị quoảng vào giữa một nhóm những người được học hành tốt! Lúc đầu tôi cảm thấy thấp kém cho đến khi tôi nhận thấy tôi có thể học nhanh hoặc chí ít cũng nhanh hơn họ nghĩ, tôi chưa từng làm bài thi nào trong đời. Trong tôi rạng lên một chân trời mới rằng tôi có thể làm những thứ khác hơn là chỉ làm nông dân ở trang trại. Tôi bắt đầu đặt mục tiêu cho bản thân, tôi nuôi hoài bão trở thành một chuyên gia nhưng để thực hiện được điều đó tôi phải vào trường đại học chỗ mà tôi chỉ có kiến thức của một đứa trẻ lên mười.
Khi rời quân đội tôi nhận thấy rằng có rất nhiều cơ hội cho những nông dân trẻ làm việc tại các nông trang ở Anh. Sau khi tham gia một lớp học tiếng Anh trong 10 buổi tối, tôi tới làm việc cho một trang trại ở Lincolnshire trong 7 tháng. Đến đây lại là một cú sốc về văn hóa lớn nữa, nhưng may mắn là tiếng Anh của tôi được cải thiện, tiếng Anh đang trở nên cần thiết cho việc học cao hơn. Để kiếm tiền cho việc học hành cao hơn, tôi đã làm việc một thời gian ngắn trong mỏ than và sau đó theo học một khoá 9 tháng ở một trường nông nghiệp (Ladelund). Vào thời điểm đó, do thiếu các chuyên gia trang trại, nên có cửa sau vào các trường đại học. Bạn có thể thi vào đại học mà không cần phải nói kiến thức của bạn có được từ đâu. Năm 1958 tôi vào học trường Cao đẳng Nông nghiệp và Thú y Hoàng gia ở Copenhagen mà không qua chương trình giáo dục trung học. Tôi tốt nghiệp với thành tích cao nhất năm 1961và kết quả là tôi được nhận học bổng học ở nước ngoài, tôi tới trường Đại học Reading bắt đầu nghiên cứu, nghiên cứu này được

6
mô tả trong phần các axit béo bay hơi (thí nghiệm 8). Trong khi bản chất nông dân của tôi không xuất hiện nhiều trong sơ yếu lý
lịch. Tôi mãi mãi tự hào về điều đó và tôi vẫn là một nông dân! Tôi mua một trang trại nhỏ gần Aberdeen năm 1972 và tôi vẫn làm các công việc trang trại. Xuất thân của tôi đã cho tôi rất nhiều kiến thức về những người nông dân, nhất là những người nông nghèo ở các nước đang phát triển, những người đã làm tôi quan tâm rất nhiều trong 15 năm qua.
Trong cuốn sách này mục tiêu đầu tiên của tôi là miêu tả, ở phần đầu của mỗi thí nghiệm, điều gì khiến chúng tôi làm việc đó. Mỗi thí nghiệm có một nền tảng khác nhau. Đôi khi một sinh viên bật lên những tài năng cụ thể dẫn chúng tôi đi vào hướng mà chúng tôi chưa từng dự định trước. Đôi khi một kỹ thuật mới cho chúng tôi một nền tảng mà từ đó chúng tôi có thể phát triển thêm một vài hướng mới trong nghiên cứu những lĩnh vực chúng tôi chưa có kế hoạch trước đó. Có thể giải pháp cho một vấn đề ngay lập tức lại đặt ra cho chúng tôi một vấn đề mới cần giải quyết. Tôi đã tự hỏi tại sao tôi đi tới ý nghĩ dùng hạt xử lý sô đa (NaOH) làm thức ăn cho bò. Nguyên nhân là do tôi đã gặp phải vấn đề với các axit béo mạch nhánh trong mỡ cừu. Sự kết nối nằm ở đâu? Tôi hy vọng khi đọc sách này sự kết nối sẽ lộ rõ!
Tôi đã sắp xếp các chương theo cách mà một hướng nghiên cứu được tiến hành từ đầu tới cuối. Điều này có nghĩa là đôi khi các hướng nghiên cứu đi vào các chủ đề khác nhau. Có 3 điểm khởi đầu chính. Đầu tiên là thí nghiệm về nhu cầu protein của cừu cai sữa sớm, thí nghiệm này đưa chúng tôi đến 4 hướng nghiên cứu tiếp theo. Điểm khởi đầu khác là các nghiên cứu trong luận văn tiến sĩ của tôi về sử dụng axit béo bay hơi, từ điểm khởi đầu này tôi đã có cơ hội theo đuổi vấn đề này sau 15 năm ở Viện nghiên cứu Rowett và điểm khởi đầu cuối cùng là các nghiên cứu về bò sữa, các nghiên cứu này là phần trách nhiệm bổ xung của tôi ở giai đoạn sau này. Tôi cảm thấy điều thú vị nhất là đi theo các hướng nghiên cứu khoa học, ngay cả khi các hướng nghiên cứu khoa học đi vào các chủ đề khác nhau.
Như vậy mỗi hướng nghiên cứu có số thứ tự riêng của nó. Tôi cũng đã làm một mục lục cho những ai chỉ quan tâm đến một chủ đề.
Mục tiêu thứ hai của tôi là miêu tả từng hướng nghiên cứu ở dạng đơn giản nhưng tổng hợp để đi đến những phát hiện then chốt. Nếu ai muốn chi tiết hơn xin hãy lật lại các báo cáo được tham khảo ở cuối mỗi thí nghiệm. Tài liệu tham khảo sẽ cung cấp một tóm tắt các công trình nghiên cứu có thể đã được xuất bản trong các bài báo khác nhau trong các tạp chí khác nhau, theo cách có thể thuận tiện cho các giáo viên dinh dưỡng gia súc, và giáo viên sinh lý và dinh dưỡng sử dụng.
Mục tiêu thứ ba của tôi là chỉ ra tác động hay tác động tiềm năng cho nông dân và các khách hàng khác, hoặc chỉ ra làm thế nào để các dự án đơn giản là cung cấp sự hiểu biết sẽ dẫn chúng ta theo hướng đúng.
Các hướng nghiên cứu khác nhau thường xuất hiện cùng thời điểm và đôi khi thông tin thu được từ một hướng nghiên cứu lại ảnh hưởng đến các thí nghiệm được thực hiện trên một hướng khác và với tính chủ quan như vậy nên vị trí của

7
một dự án hoặc thí nghiệm như thế nào đôi khi rất cần thiết. Mỗi hướng nghiên cứu thường bắt đầu với kiến thức hiện có. Bởi vậy tôi
phải thường xuyên trao đổi nhiều với những người làm cùng một lĩnh vực. Điều này được thể hiện rất rõ trong phần tài liệu tham khảo. Ở đây tôi đã tập trung vào các hướng nghiên cứu mà chúng tôi theo đuổi. Tôi hy vọng rằng đây là cách chúng ta không quên những người đã giúp tạo ra nền tảng kiến thức ban đầu.

8
Hướng nghiên cứu 1. Phân giải, sử dụng protein và sự phát triển kỹ thuật túi ni lông (Nylon bag) Hướng nghiên cứu 1.1. Nhu cầu năng lượng và protein của cừu cai sữa sớm
Công việc đầu tiên khi tôi đến Viện Rowett năm 1967 là phát triển hệ thống chăn nuôi cừu thâm canh. Nhiệm vụ của việc này là tập trung cao độ vào tăng năng suất. Giá dầu rẻ và Dầu Biển bắc ở ngay chân trời. Ngũ cốc rẻ và Tiến sĩ T.R. Preston tại Viện đã nghiên cứu sử dụng ngũ cốc nuôi bò rất thành công, vậy tại sao lại không thể nuôi cừu tốt bằng ngũ cốc. Cừu cái có thể là máy đẻ nhưng cừu con cần được tách mẹ và vỗ béo để cừu mẹ có thể có chửa trở laị ngay sau khi đẻ càng sớm càng tốt.
Chúng tôi đã quyết định thực hiện thí nghiệm lớn sử dụng phương pháp giết mổ so sánh. Theo cách này tôi có thể theo đuổi sự quan tâm của mình về trao đổi năng lượng và tôi có thể xác định được chi phí năng lượng cho duy trì protein và chi phí năng lượng cho duy trì tích lũy mỡ.
Chúng tôi yêu cầu 100 cừu cai sữa sớm. Tôi quyết định cai sữa chúng từ mẹ ở 4 tuần tuổi. Tôi không chắc tại sao chúng tôi lại chọn 4 tuần, nhưng tôi nhớ lại sau này người quản lý trang trại lúc đó ngài Harry Denerley, cảm thấy điều mới mẻ này có thể ‘treo cổ’ anh ta! Việc này sẽ không xảy ra, một vài cừu sẽ chết! May mắn cho tôi, cừu đã không chết! Chúng tôi đã có một cái máy xay thịt mới và chúng tôi đã giết mổ một vài cừu lúc 15 kg, một vài con lúc 27,5 kg và một vài con khác lúc 40 kg. Chúng tôi có 3 mức ăn khác nhau sử dụng yến mạch xay dạng viên với 5 mức protein bổ xung là khô đậu tương nhằm cung cấp 10, 12,5; 15,0; 17,5 và 20% protein thô trong khẩu phần. Thí nghiệm lớn này trở thành một phần luận án của nghiên cứu sinh tiến sỹ của tôi, R P Andrews. Chúng tôi mất 2 con cừu do sỏi niệu đạo và số cừu còn lại của thí nghiệm đều theo đúng kế hoạch (1). Tăng trọng đáp ứng với lượng protein ăn vào đã tạo nên một loạt các đường cong cổ điển, các đường cong phù hợp với kiến thức chấp nhận được ở thời điểm đó: mức ăn càng cao, đáp ứng của tăng trọng với protein khẩu phần càng cao. Ví dụ chúng tôi tính rằng ở 20 kg thể trọng, các tỷ lệ protein của khẩu phần tốt nhất là 17,5; 15 và 12,5% vật chất khô cho các mức ăn cao, trung bình và thấp. Sự khác nhau ở đây giữa các mức cho ăn là 100, 85 và 70% lượng ăn vào tự do ước tính. Khi khối lượng tăng lên, mức protein tối ưu giảm đi. Tôi cảm thấy hài lòng về điều đó. Các bạn tôi ở Viện nghiên cứu quốc gia ở Dairying, Tiến sĩ Clive và tiến sĩ Bill Broster, vừa hoàn thành tổng quan chứng minh rõ các quan hệ kiểu đường cong giữa mức ăn và đáp ứng về tăng trọng với protein khẩu phần, nhưng hãy đợi!
Phân tích hóa học thân thịt (bỏ nội tạng) của cừu cho ra một vài kết quả không phù hợp với kiến thức trong sách. Ví dụ các nhà khoa học Mỹ nói rằng rất khó thay đổi thành phần cơ thể bằng dinh dưỡng ở cừu ở cùng khối lượng thân thịt. Sai lầm; ở khối lượng 40kg và mức nuôi dưỡng cao, protein thô theo vật chất khô của thân thịt tăng từ 29 đến 39% và mỡ giảm từ 64 xuống 53% khi tăng protein khẩu phần. Sự khác nhau này ít hơn nhiều ở mức ăn thấp. Điều đáng ngạc

9
nhiên nhất và tại thời điểm rất khó để hiểu là cừu đạt tới khối lượng giết thịt ở mức ăn thấp béo hơn và ít protein hơn so với cừu đạt khối lượng giết thịt nuôi ở mức ăn cao (2). Ở mức 17,5% protein, protein thô và mỡ theo vật chất khô của thân thịt là 39 và 53% ở cừu nuôi mức ăn cao, trong khi ở cừu nuôi mức ăn thấp các tỷ lệ này là 34 và 59%. Cái mà chúng tôi mong đợi là một tỷ lệ mỡ thấp hơn nhưng sự khác nhau là chắc chắn và đáng kể. Các đồng nghiệp của chúng tôi ở bộ phận nghiên cứu về lợn của Viện không tin chúng tôi !
Thí nghiệm lớn này gồm các tốc độ tăng trọng khác nhau và các thành phần tăng trưởng khác nhau của 90 cừu cho phép chúng tôi đánh giá được năng lượng trao đổi cần cho duy trì, cho tích luỹ mỡ và protein. Chúng tôi đã tính toán khối lượng trao đổi cho từng con cừu và đã xác định được lượng mỡ và protein tăng lên, chúng tôi cũng biết tỷ lệ tiêu hoá của khẩu phần và từ đó ước tính được năng lượng trao đổi (ME) ăn vào, nhờ đó chúng tôi tìm ra hệ số cho sai số ít nhất (3). Nhu cầu ME cho duy trì là 414 KJ/kg0.75, nhu cầu ME cho tích luỹ mỡ là 1,26 KJ/KJ mỡ hay là 47,4 KJ/g mỡ, và cho tích luỹ protein thô là 3,04 KJ/KJ protein hoặc 67,3 KJ/g protein, các kết quả rất chính xác với R2 = 0,99.
Hiệu quả sử dụng thức ăn được ghi chép từ các con cừu được cho ăn đầy đủ protein là khoảng 3 kg vật chất khô khẩu phần cho 1 kg tăng trọng và đôi khi thấp hơn. Kết quả này chư bao giờ được biết đến vào thời điểm đó. Kết quả của thí nghiệm này dẫn đến một vài câu hỏi, một số câu hỏi trong số đó là các câu hỏi không mong đợi. 1. Trong khi chuyển hoá thức ăn tuyệt vời và đáng ngạc nhiên, chất lượng thân
thịt không thật tốt. Mỡ mềm và loãng ở nhiệt độ mát. Không thể cắt một miếng thịt đẹp mắt do đó người bán thịt không thích. Chúng tôi cần theo đuổi vấn đề này.
2. Vách dạ cỏ của gần như tất cả cừu dầy và hơi viêm. Viêm dạ cỏ đã được dự báo trước trên cơ sở những kết quả nuôi dưỡng bò bằng thức ăn hạt, nhưng cừu không bị áp xe gan, trong khi áp xe gan luôn là vấn đề với bò nuôi bằng thức ăn hạt, thỉng thoảng phần lớn gan bò phải loại bỏ ở các lò mổ. Mặc dù có viêm dạ cỏ, năng suất của cừu vẫn tốt; dĩ nhiên chúng tôi muốn tránh viêm dạ cỏ.
3. Như đã đề cập ở trên hàm lượng mỡ thân thịt cao một cách ngạc nhiên ở cừu
nuôi với mức ăn thấp không thể giải thích được và cần nghiên cứu tiếp về vị trí tiêu hoá các chất dinh dưỡng.
4. Từ kết quả trực tiếp của giết mổ so sánh, 2 câu hỏi nảy sinh. Nếu như cừu được cho ăn tự do, sự khác nhau tương tự về thành phần thân thịt có xuất hiện không khi các mức protein khác nhau được sử dụng trong khẩu phần? Sự khác nhau về thành phần thân thịt có giảm đi khi khối lượng cơ thể tăng?
Các câu hỏi 1, 2 và 3 sẽ được giải quyết một cách riêng biệt, ở đây chúng tôi sẽ tiếp tục với vấn câu hỏi 4.
Thí nghiệm tiếp theo là một thí nghiệm giết mổ khác trong đó chúng tôi sử dụng 60 cừu. Chỉ có 3 mức protein được sử dụng là 11,7; 15,7 và 19,4%, sử dụng bột cá và lúa mạch xay ép viên (4). Protein bột cá đã được biết trong các thí

10
nghiệm tiêu hoá song song, là ít bị phân giải trong dạ cỏ so với protein trong thực vật như đậu tương. Cừu được ăn tự do với 3 mức protein và được giết thịt từ 15 đến 50 kg thể trọng và thân thịt của chúng được phân tích. Câu hỏi chủ yếu ở đây là: liệu chúng ta có thể thấy sự khác nhau tương tự trong thành phần thân thịt với chế độ ăn tự do như chúng ta đã thấy trong các thí nghiệm trước đó và liệu thành phần thân thịt có giống nhau ở thể trọng 50 kg?
Thông điệp chính từ các kết quả được thể hiện trong Bảng 1.1, về hàm lượng mỡ ở các mức khối lượng khác nhau khi cừu được nuôi ở 3 mức protein. Bảng 1.1. Mỡ (kg) ở cừu đực và cừu cái với các khối lượng khác nhau được nuôi ở
các mức protein cao (H) trung bình (M) và thấp (L).
Khối lượng (kg)
Cừu đực Cừu cái H M L H M L
20 1,1 1,1 1,5 1,3 1,6 1,9 30 2,9 3,3 4,2 3,7 4,2 5,2 40 5,5 6,2 7,5 6,7 7,4 8,9 50 8,7 9,7 11,2 10,4 11,1 13,0
Khi khối lượng tăng lên, sự khác nhau về mỡ trong thân thịt của cừu giữa
các mức ăn protein khác nhau nhỏ đi. Có sự khác nhau lớn về mỡ trong thân thịt do khác biệt về các mức protein ở cừu khối lượng nhỏ. Cừu đực nuôi ở mức protein thấp thân thịt có nhiều mỡ (11,2 kg) hơn cừu đực nuôi ở mức protein cao hơn (8,7 kg) lúc đạt 50 kg. Thành phần của tăng trọng trở nên gần như nhau khi khối lượng tăng và chúng tôi đã tính toán thấy ở khối lượng 60 kg thành phần thân thịt của chúng gần như nhau. Có sự khác biệt rõ ràng giữa cừu đực và cừu cái về thành phần cơ thể và về đáp ứng với các mức protein của khẩu phần. Cừu cái cần ít protein hơn cừu đực và có nhiều mỡ hơn ở cùng khối lượng cơ thể.
Thí nghiệm này cũng được sử dụng để đánh giá nhu cầu năng lượng cho duy trì và cho tăng trọng mỡ cũng như protein, các kết quả ở đây cũng tương tự như thí nghiệm trước đó.
Câu hỏi tiếp theo chúng tôi muốn giải quyết sử dụng phương pháp giết thịt
so sánh là rất quan trọng với chúng tôi nhưng bị bỏ qua một cách đáng ngạc nhiên (5) có thể bởi vì chúng tôi sử dụng sai tiêu đề. Câu hỏi đó là: điều gì xảy ra nếu như cừu nuôi khẩu phần protein thấp và do đó mỡ ở 28 kg đột nhiên thay đổi khi được cừu được chuyển sang nuôi với mức protein khẩu phần cao? Tương tự như vậy, điều gì xảy ra với lượng thức ăn ăn vào của cừu? Có thể mỡ được sử dụng, như đã bàn, như nguồn năng lượng dùng cho quá trình tích lũy protein. Bởi vì tôi cảm thấy bài báo về thí nghiệm này có một cái gì đó bị lãng quên nên tôi muốn đưa ra những kết quả chi tiết hơn. Cừu được nuôi với khẩu phần protein thấp từ 15 đến 70 kg thể trọng và được giết thịt theo các mức khối lượng khác nhau, và

11
hàm lượng protein, mỡ và khoáng của thân thịt được phân tích. Tương tự, các cừu khác được nuôi khẩu phần protein cao với khối lượng từ 15 đến 70 kg cũng được giết thịt theo kiểu tương tự. Nhóm cừu thứ ba, LPH được nuôi với khẩu phần thấp protein giai đoạn đầu và được đổi sang khẩu phần có tỷ lệ protein cao lúc 28 kg để giám sát những thay đổi thành phần cơ thể. Tất cả cừu đều được ăn tự do ngoại trừ khi khẩu phần thấp protein được đổi thành cao protein, cừu được cho ăn lượng thức ăn bằng với lượng thức ăn của nhóm cừu nuôi với khẩu phần protein cao (HP) đã ăn được ở cùng khối lượng. Kết quả hấp dẫn nhất được trình bày trong Bảng 1.2 và 1.3.
Bảng 1.2. Ảnh hưởng của nuôi khẩu phần thấp (LP), cao protein (HP) hoặc thay đổi từ khẩu phần thấp đến cao protein ở khối lượng 28 kg (LPH) đến tăng trọng và
tiêu tốn thức ăn ở cừu.
Khối lượng (kg)
Tăng trọng (g/ngày)
Tiêu tốn thức ăn (kg DM/ kg tăng trọng)
LP LPH HP LP LPH HP 20-28 135 154 378 5,5 5,0 2,4 28-35 164 457 353 4,4 2,3 3,4 35-45 189 416 397 4,8 2,9 3,5 45-55 180 346 335 6,3 4,0 4,5 55-65 209 279 297 - 5,0 5,7
Cừu nuôi khẩu phần thấp protein được thay đổi khẩu phần lúc 28 kg sang
khẩu phần cao protein có hiệu ứng tăng trưởng bù mạnh mẽ cả về khối lượng và tiêu tốn thức ăn. Ở khoảng 45 kg, tăng trọng và tiêu tốn thức ăn của nhóm cừu này tương tự như ở cừu nuôi với khẩu phần protein cao trong suốt giai đoạn thí nghiệm.
Bảng 1.3. Tỷ lệ tích luỹ protein, mỡ và nước trong thân thịt của cừu nuôi khẩu phần cao (HP), thấp (LP) protein hay khẩu phần thay đổi từ thấp sang cao ở khối
lượng 28 kg (LPH).
Khối lượng (kg)
Tỷ lệ tích luỹ protein (g/ngày)
Tỷ lệ tích luỹ mỡ (g/ngày)
Tỷ lệ tích luỹ nước (g/ngày)
LP LPH HP LP LPH HP LP LPH HP 20-28 19 25 60 29 39 74 65 85 222 28-35 34 62 52 62 108 85 110 276 186 35-45 32 77 59 68 97 116 98 225 199 45-55 27 55 49 65 105 113 80 175 156 55-65 - 43 43 - 98 110 - 133 126

12
Theo quan điểm của tôi các kết quả này rất tuyệt vời. Kết quả cho thấy cừu có khả năng thay đổi đáp ứng rất nhanh khi các trở ngại do thiếu hụt protein trong khẩu phần được loại bỏ. Quá trình tích luỹ nước và protein thay đổi mạnh mẽ, một vài cừu tăng trọng hơn 500g/ngày. Tỷ lệ nước-protein cũng đã thay đổi, mặc dù tỷ lệ này được cho là một hằng số. Nhiều đề xuất của các nhà nghiên cứu trường Đại học Cornell (Reid J.T. và cộng sự (1968) Proc. Cornell Nutr. Conf. P 18-37) cho rằng thành phần cơ thể ở một khối lượng nhất định là khó thay đổi bằng các biện pháp dinh dưỡng! Các số liệu kết quả về thành phần thịt xẻ cũng cho thấy rằng, giống như tăng trọng và chuyển hoá thức ăn, thành phần tăng trọng của cừu nuôi với khẩu phần có sự thay đổi protein từ thấp sang cao (LPH) tương tự như ở cừu nuôi khẩu phần cao protein (LP) ở khối lượng 45 kg.
Ở thời điểm này nghiên cứu giết mổ so sánh đã kết thúc, không phải do tốn kém mà vì chúng tôi cảm thấy ở thời điểm đó không có câu hỏi đặc biệt quan trọng nào cần tìm câu trả lời. Tài liệu tham khảo cho phần 1.1
1. Andrews, R.P. and Orskov, E.R (1970). The nutrition of the early weaned lamb. I. The influence of protein concentration and feeding level on rate of gain in body weight. J. Agric. Sci., Camb. 75, 11-18.
2. Andrews, R.P. and Orskov, E.R (1970). The nutrition of the early weaned lamb. II. The effect of dietary protein concentration, feeding levels and sex on body composition at two live weights. J. Agric. Sci., Camb. 75, 19-26.
3. Orskov, E.R and McDonald, I. (1970). The utilization of dietary energy for maintenance and for protein and for fat deposition in young growing sheep. EAAP Symposium on Early Metabolism, Zurich, pp. 121-124.
4. Orskov, E.R, McDonald, I., Fraser, C and Corse, Elizabeth L. (1971). The nutrition of the early weaned lamb. III. The effect of ad libitum intake of diets varying in protein concentration on performance and body composition at different live weights. J. Agric. Sci., Camb. 77, 351-361.
5. Orskov, E.R, McDonald, I., Grubb, D.A. and Pennie, K. (1976). The nutrition of the early weaned lamb. IV. Effects on growth rate, food utilization and body composition of changing from a low to a high protein diets. J. Agric. Sci., Camb. 86, 411-423.

13
Hướng nghiên cứu 1. Phân giải, sử dụng protein và sự phát triển kỹ thuật túi ni lông
Hướng nghiên cứu 1.2. Phân giải protein và phát triển khái niệm mới về dinh dưỡng protein ở gia súc nhai lại.
Nghiên cứu nhu cầu protein của cừu được trình bày trong hướng nghiên cứu 1.1 cho thấy rõ ràng là có đáp ứng về tăng trọng ở cừu với protein khẩu phần, đặc biệt ở mức protein khẩu phần cao (1,2).
Để nghiên cứu tiếp, chúng tôi đặt canulae ở dạ múi khế và hồi tràng của cừu, nhằm nghiên cứu ở mức độ nào đáp ứng về tăng trọng ở cừu với protein khẩu phần liên quan tới một lượng protein lớn hơn protein đi xuống dạ múi khế, và điều quan trọng hơn là bản chất của protein này là gì? Bao nhiêu phần trăm protein đi xuống dạ múi khế là do tăng protein vi sinh vật đáp ứng với lượng đậu tương tăng lên trong khẩu phần và bao nhiêu phần trăm protein từ đậu tương thoát qua sự phân giải dạ cỏ và đến dạ lá sách để tiêu hoá và hấp thu ở ruột non. Cảm giác riêng của tôi là phần lớn protein đi xuống dạ múi khế là protein của khẩu phần vì các tính toán của tôi về cân bằng lên men khi tính tới các axit béo bay hơi (3), dẫn tôi tới ý nghĩ rằng protein là nguồn năng lượng nghèo cho vi sinh vật trong lên men yếm khí và tổng hợp protein vi sinh vật phải liên quan chủ yếu tới việc lên men carbohydrate.
Thí nghiệm đầu tiên của chúng tôi nhằm kiểm tra khái niệm này sử dụng mức nuôi dưỡng cao với lúa mạch từ thí nghiệm tăng trọng (Thí nghiệm 1.1) và đánh giá lượng protein thô thoát qua dạ lá sách và phần cuối hồi tràng khi tăng mức đậu tương trong khẩu phần (4) (xem Bảng 1.4). Bảng 1.4. Ảnh hưởng tỷ lệ protein thô (CP) và mức đậu tương trong khẩu phần lúa mạch đến hấp thu thực của protein thô (không gồm ammonia) từ ruột non (lượng
CP từ dạ lá sách trừ đi lượng CP ở hồi tràng).
Protein thô theo % vật chất khô của khẩu phần
Đậu tương (%) theo vật chất khô
của khẩu phần
CP ăn vào (g/ngày)
CP (không gồm ammonia) hấp thu
từ ruột non (g/ngày)
10,3 4,5 87 61 13,3 8,8 133 98 16,1 18,4 174 116 19,6 26,0 199 114 SE 5
Rõ ràng có sự tăng đáng kể protein thô (không gồm ammonia) hấp thụ từ
ruột non khi tỷ lệ đậu tương trong khẩu phần tăng lên, đặc biệt ở những khẩu phần có tỷ lệ đậu tương thấp hơn. Tuy nhiên, dường như không có sự thay đổi protein

14
thô (không gồm ammonia) hấp thụ từ ruột non ở mức đậu tương cao hơn, và dự đoán của riêng tôi là mức protein ăn vào cao nhất đã làm giảm sinh tổng hợp của vi sinh vật nhưng làm tăng lượng protein thoát qua không bị phân giải ở dạ cỏ, do đó tổng lượng protein xuống ruột non không thay đổi, nhưng làm sao chúng ta có thể chứng minh rằng protein thô (không gồm ammonia) hấp thụ từ ruột non là protein đậu tương thoát qua sự phân giải ở dạ cỏ? Thành phần amino axit của dưỡng chấp tại dạ lá sách không thay đổi đáng kể, nhưng khó có thể phân biệt ở dạ lá protein nào từ vi sinh vật, protein nào từ đậu tương. Có rất ít sự thay đổi về lượng protein thoát xuống ruột già và bài xuất vào phân, điều này cho thấy phần protein tăng lên thoát xuống ruột non được hấp thu phần lớn. Chúng tôi nghĩ rằng nếu protein thô (không gồm ammonia) hấp thụ từ ruột non là protein vi sinh vật, cừu cũng có thể đáp ứng với urê trong khẩu phần, do đó trong thí nghiệm tiếp theo chúng tôi so sánh ảnh hưởng của mức urê tăng lên trong khẩu phần với các mức khác nhau của bột cá đến lượng protein đi xuống dạ lá sách. Chúng tôi chuyển sang bột cá bởi vì các thí nhiệm sử dụng ống thông thực quản cho thấy việc sử dụng bột cá ưu việt hơn so với đậu nành khi cho ăn khô và ngay cả khi cho ăn ở dạng lỏng. Các kết quả quan trọng nhất được trình bày trong Bảng 1.5 (5). Bảng 1.5. Ảnh hưởng của việc tăng N trong khẩu phần bằng cách bổ sung urê hoặc
bột cá đến protein thô (không kể ammonia) (NACP) thoát qua từ dạ lá sách tới đoạn cuối hồi tràng.
Bổ sung Urê hoặc bột
cá (% theo chất khô của khẩu
phần)
Protein thô ăn vào
(g/ngày)
NACP (g/ngày) Hấp thu từ ruột non
Thoát tới hồi tràng
Trong phân
Không 0,0 97 76 51 37 Urê mức 1 0,9 123 75 54 37 Urê mức 2 2,0 157 66 63 33 Urê mức 3 3,2 193 83 57 34 Urê mức 4 4,3 240 91 50 29 Bột cá mức 1 3,8 116 95 56 29 Bột cá mức 2 8,9 148 89 51 32 Bột cá mức 3 14,1 194 117 57 33 Bột cá mức 4 19,2 220 132 54 31 SE 6 4 6 1
Ở đây, rõ ràng là lượng protein thoát qua từ dạ lá sách tăng lên có nguồn
gốc từ khẩu phần. Có ít hoặc không có quá trình tăng sinh tổng hợp protein vi sinh vật do tăng cung cấp nitơ từ urê nhưng nitơ thoát qua từ dạ lá sách tăng lên đáng kể theo kiểu đường thẳng khi tăng lượng bột cá trong khẩu phần. Các phương trình hồi quy cho thấy khoảng 37% protein bột cá thoát qua sự phân giải dạ cỏ và được

15
hấp thu từ ruột non. Lý do khác để chọn bột cá là thành phần protein bột cá đủ khác với thành
phần protein vi sinh vật, do đó cho phép phân biệt giữa chúng. Ở mức bột cá cao nhất, tỷ lệ lysine và methionine ở dạ lá sách là 7,5 và 3,2 g/16g nitơ. Tỷ lệ tương ứng cho khẩu phần bổ sung urê là 5,9 và 2,1 g/16g nitơ. Đây là bằng chứng tốt nhất cho đến lúc ấy cho thấy protein khẩu phần thoát qua sự phân giải ở dạ cỏ và sinh tổng hợp protein vi sinh vật chỉ do lên men carbohydrate qui định. Chúng ta bây giờ đã có phương pháp cho phép chúng ta phân biệt giữa nhu cầu dinh dưỡng của vi sinh vật và nhu cầu dinh dưỡng của vật chủ (gia súc). Chúng tôi đã bắt đầu nghĩ tới một hệ thống mô tả nhu cầu dinh dưỡng của vi sinh vật và nhu cầu dinh dưỡng của vật chủ (6).
Thí nghiệm tiếp theo (7) nhằm đánh giá nhu cầu nitơ vi sinh vật ở cừu nuôi với khẩu phần cơ sở là lúa mạch. Hai phương pháp được sử dụng ở đây. Ở phương pháp thứ nhất, khẩu phần có tỷ lệ protein thấp được bổ sung urê với các tỷ lệ tăng dần. Chúng tôi đo đặc số lượng tinh bột và protein thoát qua dạ lá sách, vì hình như do nitơ không đủ, tiêu hóa vi sinh vật sẽ chậm lại và nhiều tinh bột hơn có thể thoát qua sự phân giải trong dạ cỏ. Các kết quả được trình bày ở Bảng 1.6. Bảng 1.6. Ảnh hưởng của việc bổ sung urê vào khẩu phần cơ sở lúa mạch đến tinh
bột, protein thoát qua dạ lá sách và tăng trọng của cừu.
Protein thô của khẩu phần (%)
Urê khẩu phần (%)
Tinh bột ăn vào
(g/ngày)
Tinh bột thoát qua dạ lá sách (g/ngày)
Protein thoát qua dạ lá sách (g/ngày)
Tăng trọng
(g/ngày)
10,0 0,0 470 67 100 166 12,4 0,7 464 32 107 217 14,6 1,4 469 16 106 223 16,4 2,1 457 16 104 228 SE 8 31 Ở đây chúng tôi nhận thấy tiêu hoá tinh bột ở dạ cỏ tăng lên cùng với
mức tăng urê trong khẩu phần. Ước lượng có khoảng 1% urê cần được bổ sung vào khẩu phần lúa mạch nhằm đáp ứng nhu cầu của vi sinh vật. Tăng trọng cũng thấy có đáp ứng với urê.
Phương pháp thứ hai được sử dụng là một thí nghiệm về tăng trọng trong đó các nhóm cừu được nuôi với các khẩu phần có mức urê khác nhau. Thiết kế thí nghiệm như vậy là hợp lý vì nitơ vi sinh vật không đáp ứng đủ nhu cầu protein của cừu, cừu đáp ứng rất nhạy cảm với bất kỳ sự tăng sinh tổng hợp protein vi sinh vật nào. Sinh tổng hợp protein vi sinh vật được thể hiện ở việc tăng tỷ lệ tiêu hoá và tăng trọng. Một khẩu phần có bột cá (phần lớn protein trong bột cá đi xuống dạ múi khế mà không bị phân giải) cũng được đưa vào trong thí nghiệm này để chắc chắn rằng tiềm năng tăng trọng sẽ không đạt được nếu chỉ có duy nhất nguồn protein vi sinh vật (xem Bảng 1.7).

16
Bảng 1.7. Ảnh hưởng của việc bổ sung urê trong khẩu phần lúa mạch đến tiêu
hoá, tăng trọng và chuyển hoá thức ăn ở cừu cai sữa sớm.
Mức urê (% DM khẩu phần)
Tỷ lệ protein thô (% DM)
Tỷ lệ tiêu hoá (%)
Tăng trọng (gam/ngày)
0,0 9,0 74,9 158 0,55 10,5 81,2 164 1,10 12,3 80,6 215 1,65 14,1 82,3 196 2,20 15,6 80,7 206
Bột cá 14,7 80,0 261 SE 12
Kết quả đạt được của thí nghiệm này hoàn toàn phù hợp với các kết quả của
các nghiên cứu khác về vị trí tiêu hóa các chất dinh dưỡng trong đường tiêu hóa. Nhu cầu nitơ của vi sinh vật dạ cỏ ở cừu ăn khẩu phần lúa mạch có thể đáp ứng đủ khi bổ sung khoảng 1% urê vào khẩu phần, trên mức này sẽ không có đáp ứng thêm nữa. Kết quả một lần nữa cho thấy tiềm năng tăng trọng của cừu không thể thoả mãn chỉ với protein vi sinh vật và protein trong lúa mạch như nguồn protein thoát qua dạ cỏ, kết quả là cừu đáp ứng với protein không phân giải dạ cỏ để điều chỉnh sự thiếu hụt.
Thí nghiệm cuối cùng trong một loạt các thí nghiệm nhằm tìm hiểu xem urê có thể tiết kiệm protein khẩu phần từ quá trình phân giải trong dạ cỏ không và urê có ưu thế hơn bột cá trong việc đáp ứng nhu cầu nitơ của vi sinh vật của cừu ăn khẩu phần lúa mạch không vì bột cá chỉ bị phân giải một phần ở dạ cỏ.
Câu hỏi đầu tiên được đặt ra trong một thí nghiệm về vị trí tiêu hóa các chất dinh dưỡng trong đường tiêu hóa. Hai mức bột cá được bổ sung cho cừu cùng với 10g urê/ ngày hoặc không có urê (khoảng 1% khẩu phần) vào khẩu phần lúa mạch (Bảng 1.8)(8).
Bảng 1.8. Ảnh hưởng của việc bổ sung urê vào bột cá đến protein thô (không kể ammonia) (NACP) thoát qua dạ múi khế.
Khẩu phần Protein thô ăn
vào (g/ngày) Protein thô (không kể ammonia) (NACP) thoát qua dạ múi khế (g/ngày)
Lúa mạch 97 131 Lúa mạch + bột cá 1 124 144 Lúa mạch +bột cá 1 + 10 g urê 163 142 Lúa mạch + bột cá 2 160 154 Lúa mạch +bột cá 2 + 10 g urê 211 140

17
SE 7
Urê rõ ràng không có ảnh hưởng đến phân giải của bột cá bởi vì không thấy protein thô (không kể ammonia) thoát qua dạ lá sách tăng lên khi urê được bổ sung vào khẩu phần cùng với bột cá.
u hỏi thứ hai được trả lời trong thí nghiệm tăng trọng, thí nghiệm này đã cho thấy urê hơn hẳn bột cá để thoả mãn nhu cầu nitơ của vi sinh vật. Ở mức urê 0,7% tỷ lệ tiêu hoá chất hữu cơ là 78,5% trong khi đó ở mức nitơ ăn vào tương tự từ bột cá tỷ lệ tiêu hoá chất hữu cơ thấp hơn đáng kể: 76,7%. Tuy nhiên, tăng trọng và chuyển hoá thức ăn ở khẩu phần bột cá vượt trội hơn, đúng như mong đợi.
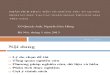
Một thí nghiệm khác được tiến hành cho thấy rõ ràng hơn là nhu cầu nitơ của vi sinh vật tương ứng với khả năng lên men của khẩu phần. Chúng tôi chọn rơm lúa mạch không sử lý và rơm lúa mạch sử lý với NaOH như khẩu phần cơ bản và phun các tỷ lệ urê khác nhau lên rơm. Kết quả được trình bày ở hình 1.1.(9).
Hình 1.1. Ảnh hưởng của bổ sung urê đến lượng thức ăn ăn vào, tỷ lệ tiêu hóa của rơm có xử lý và không xử lý NaOH làm thức ăn cho cứu

18

19
Bổ sung urê vào rơm không xử lý có ít ảnh hưởng đến lượng thức ăn ăn vào và tỷ lệ tiêu hóa vật chất khô, nhưng ảnh hưởng này không thấy ở rơm đã xử lý NaOH. Nếu như sự thiếu hụt nitơ không được điều chỉnh, sử lý NaOH không có ảnh hưởng đến lượng ăn vào và tỷ lệ tiêu hoá; nhưng bổ sung urê đã làm tăng lượng ăn vào và tỷ lệ tiêu hoá rất nhiều vì vậy thực tế vật chất khô ăn vào tiêu hoá được tăng hơn hai lần. Một quan sát thú vị khác là ảnh hưởng của urê đến tỷ lệ tiêu hoá nitơ. Ở đây kết quả của quá trình lên men tiêu hoá manh tràng cho phép thấy được lượng vật chất có tiềm năng lên men của rơm được sử lý NaOH đi xuống ruột già, được lên men và đóng góp cho quá trình sinh tổng hợp vi sinh vật tại đây, và lượng protein vi sinh vật này được bài xuất dưới dạng nitơ trong phân. Hiển nhiên những thí nghiệm như vậy không thể kéo dài khi nhiều nitơ được bài tiết trong phân hơn nitơ có trong thức ăn. Tuy vậy, thí nghiệm cho thấy tỷ lệ nitơ: năng lượng phụ thuộc lẫn nhau quan trọng như thế nào, thí nghiệm cho thấy tỷ lệ tiêu hóa nitơ biểu kiến không phù hợp như thế nào.
Suy ngẫm lại, ở thời điểm đó năm 1975 chúng ta đã có thể xác định vài nguyên tắc mới liên quan đến dinh dưỡng protein ở gia súc nhai lại.
1. Đáp ứng với protein ở cừu là đáp ứng với protein thoát qua của khẩu phần, không phải là đáp ứng với tăng sinh tổng hợp protein vi sinh vật (Bảng 1.7).
2. Dinh dưỡng nitơ vi sinh vật về khái niệm khác với dinh dưỡng vật chủ vì vi vi sinh vật có thể sử dụng nitơ phi protein (Bảng 1.5) từ urê và nếu không đáp ứng đủ nhu cầu nitơ cho vi sinh vật sẽ dẫn đến tiêu hoá kém và lượng thức ăn ăn vào thấp.
3. Bổ sung protein khác nhau về mức độ phân giải trong dạ cỏ, như đã được chứng minh trong thí nghiệm sử dụng phản xạ đóng, mở rãnh thực quản, cũng sẽ dẫn đến hiệu quả sử dụng sau dạ cỏ, có thể do thành phần axit amin của các protein.
4. Thoả mãn nhu cầu nitơ của vi sinh vật dạ cỏ không có ảnh hưởng thặng dư nào đến phân giải dạ cỏ của thức ăn bổ sung protein (Bảng 1.8).
5. Nhu cầu nitơ của vi sinh vật phụ thuộc vào tính sẵn có của năng lượng tạo ra từ lên men thức ăn trong dạ cỏ (Bảng 1.6).
6. Tỷ lệ tiêu hoá biểu kiến của nitơ không có ý nghĩa gì khi chúng ta quan tâm đến cung cấp protein cho gia súc nhai lại, tỷ lệ tiêu hoá biểu kiến của nitơ có thể hoàn toàn dẫn đến những sai lầm, vì tiêu hóa hồi tràng có ảnh hưởng đến nitơ bài tiết trong phân và do đó ảnh hưởng đến tỷ lệ tiêu hoá biểu kiến của nitơ (Sơ đồ 1.1). Từ những nguyên tắc đó, rõ ràng là việc cung cấp protein cho gia súc nhai
lại không thể được đánh giá khi không có kiến thức về cung cấp protein vi sinh vật và tỷ lệ phân giải của protein khẩu phần. Nếu không biết về cung cấp protein vi sinh vật, không thể đánh giá nhu cầu dinh dưỡng cần có trong khẩu phần và chúng ta buộc phải kết luận rằng tất cả thông tin trong các sách về nhu cầu protein của gia súc nhai lại rất khó tìm thấy các hiểu biết này và các thông tin trong sách là không đáng tin cậy! Sách về các hệ thống dựa trên protein thô tiêu hoá cần phải bỏ riêng ra vào nhóm sách lịch sử !

20
Vào khoảng thời điểm này, ngài Kenneth Blaxter tuyển dụng tiến sĩ E L Miller từ trường Đại học Cambridge và tôi vào nhóm nghiên cứu protein của Uỷ ban ARC nhằm đánh giá nhu cầu dinh dưỡng của gia súc nhai lại. Các thành viên khác của nhóm là tiến sĩ C C Balch, tiến sĩ J H B Roy và tiến sĩ R S Smith, tất cả họ từ Viện Nghiên cứu Bò sữa Quốc gia. Chúng tôi cùng làm việc với nhau rất ăn ý và vào năm 1977 (10, 11) trong một cuộc hội thảo ở Hà Lan, chúng tôi đưa ra hệ thống đánh giá nhu cầu protein, trong đó các nguyên tắc mới trong dinh dưỡng protein ở gia súc nhai đã được đưa vào sử dụng. Tài liệu tham khảo cho phần 1.2
1. Andrews, R.P. and Orskov, E.R (1970). The nutrition of the early weaned lamb. I. The influence of protein concentration and feeding level on rate of gain in body weight. J. Agric. Sci., Camb. 75, 11-18.
2. Andrews, R.P. and Orskov, E.R (1970). The nutrition of the early weaned lamb. II. The effect of dietary protein concentration, feeding levels and sex on body composition at two live weights. J. Agric. Sci., Camb. 75, 19-26.
3. Orskov, E.R, Flatt, W.P. and Moe, P.W. (1968). A fermentation balance approach to estimate extent of fermentation and efficiency of volatile fatty axít formation in ruminants. J. Dairy Sci. 51, 14-29.
4. Orskov, E.R, Fraser, C. and McDonald, I. (1971). Digestion of concentration in sheep. I. The effect of increasing the concentration of soyabean meal in a barley diet on apparent disappearance of food constituents along the digestive tract. Br. J. Nutr. 25, 225-233.
5. Orskov, E.R, Fraser, C. and McDonald, I. (1971). Digestion of concentration in sheep. I. The influence of urea or fish meal in barley on apparent digestion of protein, fat, starch and ash in the rumen, the small intestine and the large intestine. Br. J. Nutr. 25, 243-252.
6. Orskov, E.R, (1970). Nitrogen utilization in the young ruminant. Proc. 4th Nutr. Conf., Nottingham. J. & A. Churchill, London.
7. Orskov, E.R, Fraser, C and McDonald, I. (1972). Digestion of concentration in sheep. IV. The effect of urea on digestion, nitrogen retention and growth in young lambs. Br. J. Nutr. 27, 491-501.

21
Hướng nghiên cứu 1. Phân giải, sử dụng protein và sự phát triển kỹ thuật túi nilông Hướng nghiên cứu 1.3. Nhu cầu nitơ của vi sinh vật ở các thức ăn cơ sở và những ứng dụng để phát triển của kỹ thuật túi ni lông.
Vào năm 1973, Ahmad Z Mehrez - sinh viên tiến sĩ của tôi tham gia nghiên cứu với chúng tôi. Tôi muốn anh ấy tiếp tục nghiên cứu về về nhu cầu nitơ của vi sinh vật trong các thức ăn cơ sở – trong trường hợp này là lúa mạch và ngô. Sự hợp lý ở đây là chúng tôi sẽ để cho phần protein cần thiết bổ sung vào nhu cầu nitơ của vi sinh vật thoát qua dạ cỏ bằng cách ví dụ như sử dụng phản xạ rãnh thực quản. Vào thời điểm đó chúng tôi đã phát triển hệ thống ngũ cốc nguyên hạt trong đó các vitamin, muối khoáng và urê được hấp phụ một cách đơn giản vào ngũ cốc. Chúng tôi chọn 2 loại lúa mạch và tôi muốn chỉ tập trung vào lúa mạch thôi. Chúng tôi đã làm thí nghiêm này tương tự như ở hướng nghiên cứu 1.2 (1,2,3). Cho toàn bộ thí nghiệm, khoảng 1% urê bổ sung vào lúa mạch có 10% protein thô tạo ra khẩu phần có khoảng 12 đến 13% protein thô. Chúng tôi chọn lúa mạch có tỷ lệ protein thô thấp (10,4%) và lúa mạch có tỷ lệ protein thô cao (13,4%). Chúng tôi trông đợi rằng có sự đáp ứng khác nhau với urê, hoặc không có sự đáp ứng với urê ở khẩu phần protein cao. Tuy nhiên, chúng tôi đã rất ngạc nhiên (4) xem Bảng 1.9.
Bảng 1.9. Ảnh hưởng của việc bổ sung urê vào khẩu phần lúa mạch có tỷ lệ protein cao (HP) hoặc thấp (LP) đến lượng thức ăn ăn vào, tăng trọng và chuyển
hoá thức ăn ở cừu cai sữa sớm.
CP (% khẩu phần) Lượng ăn vào (g/ngày) Tăng trọng (g/ngày) HP LP HP LP HP LP 13,4 10,4 982 811 171 87 15,3 13,2 1062 1065 195 200 17,1 15,4 1186 1138 259 216 19,6 17,9 1203 1147 256 225 SE 44 45 16 17
Chúng tôi thấy rằng cừu đáp ứng với urê gần như nhau ở khẩu phần protein
cao và thấp (khoảng 1,1 đến 1,4% urê ở hai khẩu phần). Kết quả này làm vấn đề phức tạp thêm. Tại sao lại như vậy? Chúng tôi nghi nghờ rằng có thể protein trong 2 loại lúa mạch có tỷ lệ phân giải khác nhau và do đó cung cấp những lượng nitơ khác nhau, nhưng chúng ta đo đạc nó như thế nào? Vào thời điểm này phương pháp nylon bag đã được áp dụng. Tôi đã xem phương pháp nylon bag được Tiến sĩ Schoeman ở Stellenbosch tại Nam Phi sử dụng và tôi nhận thấy rằng phương pháp

22
này có thể giúp chúng tôi nhưng lấy nylon ở đâu? Schoeman đã sử dụng vải may dù, vì thế tôi gọi điện cho Trung đoàn nhảy dù tình nguyện ở Aberdeen và hỏi họ nếu họ có cái dù bỏ đi nào không để chúng tôi xin hay mua. Họ tặng chúng tôi một chiếc dù và Ahmad Mehrez, trong luận văn của anh ta đã cảm ơn việc tặng dù của thiếu tá Trung đoàn nhảy dù tình nguyện ở Aberdeen! Ahmad sau khi ổn định cuộc sống đã nhờ vợ-cô Amal và mẹ vợ may một số túi nylon. Công việc đã tiến triển không được tốt, biến động về kết quả là quá lớn. Vào thời điếm đó tôi tới làm việc tại INRA (Pháp) trong 3 tháng và tại đây tôi gặp tiến sĩ De Wet cũng tới từ Nam Phi, người cùng làm việc với tiến sĩ Schoeman. Tôi nói với ông về vấn đề của chúng tôi và háo ra là chúng tôi đã sử dụng quá nhiều mẫu so với kích thước túi. Tôi gửi thư cho Ahmad nói cần phải giảm lượng mẫu hoặc tăng kích thước túi. Khi tôi quay trở về, điều đầu tiên Ahmad nói với tôi là: “Tiến sĩ Orskov, mọi thứ bây giờ ổn rồi’’.
Tôi chỉ muốn nhấn mạnh rằng phương pháp nylon bag không phải do chúng tôi phát triển, đóng góp của chúng tôi là sử dụng phương pháp này như một công cụ năng động để xác định phân giải của thức ăn và sau đó là nghiên cứu về môi trường dạ cỏ. Phương pháp lúc đầu xem ra thô sơ và chúng tôi nhận góp ý rằng phương pháp này thực sự quá sơ sài để có thể đảm bảo đăng bài trên các tạp chí uy tín!
Song song với công việc này chúng tôi đã phát triển khái niệm ngũ cốc nguyên hạt, ở đây này urê được hấp phụ vào ngũ cốc nguyên hạt nhằm giúp cho việc ammonia được giải phóng từ từ trong dạ cỏ. Nhằm làm cho việc giải phóng ammonia đều đặn hơn, chúng tôi tiếp tục sử dụng máng cấp thức ăn liên tục và cho cừu lắp canula dạ cỏ ăn lúa mạch xử lý với 0, 2, 4, 6, 8 và 10g urê cho 1 kg lúa mạch. Điều này giúp chúng tôi có các nồng độ ammonia biến động đều đặn từ 50 đến 450 mg/lít dịch dạ cỏ. Bây giờ chúng tôi có thể xác định nồng độ ammonia tối ưu cho quá trình phân giải lúa mạch (5).
Kết quả thực sự đáng ngạc nhiên. Chúng ta cần khoảng 225 mg ammonia /lít, cao hơn mức đó không có sự tăng lên trong phân giải. Số liệu này cao hơn đáng kể giá trị 50 mg ammonia của Satter và Slyter (1974) Br. J. Nutr. 32, 199.
Nồng độ NH3 tối ưu trong dịch dạ cỏ vẫn còn được tranh luận cho tới ngày nay. Chúng tôi thấy rằng vì vi sinh vật sử dụng nitơ dưới dạng ammonia chứ không phải dưới dạng ammonium, nên pH dịch dạ cỏ rất quan trọng do mối quan hệ pH/pK. Nếu quả là như vậy, thì nồng độ ammonia tối ưu có thể là thấp hơn 10 lần ở pH dạ cỏ 7,0; và cao hơn nhiều mức 235 mg/lít nếu pH dạ cỏ thấp hơn 6,0 nhiều. Trong nghiên cứu của chúng tôi pH dạ cỏ khoảng 6,0. Hơn nữa độ dầy bề mặt, hay mật độ của mẫu có thể rất quan trọng vì vi sinh vật phát triển trong các nhóm, các nhóm này khác nhau về độ sâu xâm nhập và vì thế ammonia trong dịch dạ cỏ có thể không giống nhau vì diện tích bề mặt tiếp xúc của vi sinh vật với mẫu khác nhau.
Như vậy khi nuôi với lúa mạch nguyên hạt, giá trị 235 mg ammonia /lít là phù hợp, tuy vậy mức này có thể không đúng với loại thức ăn khác và điều kiện nuôi dưỡng khác. Lạy chúa, chúng tôi chọn cách không tập trung nhiều hơn nữa

23
vào nồng độ ammonia tối ưu mà tập trung vào lượng nitơ phân giải tối ưu trong mối quan hệ với năng lượng sẵn có trong dạ cỏ từ lên men.
Chúng tôi không bao giờ nghĩ tới việc sử dụng kỹ thuật nylon bag theo hướng tĩnh, nghĩa là với một thời gian ủ mẫu, bởi vì để giải quyết vấn đề lúa mạch của chúng tôi, chúng tôi muốn xác định thời điểm mà ở đó 93% tinh bột đã bị phân giải và sử dụng giá trị protein tại thời điểm đó để xác định tỷ lệ phân giải bởi vì khoảng 7% tinh bột từ lúa mạch thường thoát qua không bị phân giải. Sử dụng phương pháp này, phân giải dạ cỏ của khẩu phần protein cao và của cả hai loại lúa tương ứng là 0,69 và 0,82. Điều này thực sự giúp giải thích tại sao chúng tôi nhận được đáp ứng với urê ở khẩu phần protein cao. Tài liệu tham khảo cho phần 1.3
1. Ørskov, E.R, Fraser, C. and McDonald, I. (1971). Digestion of concentration in sheep. I. The effect of increasing the concentration of soyabean meal in a barley diet on apparent disappearance of food constituents along the digestive tract. Br. J. Nutr. 25, 225-233.
2. Ørskov, E.R, Fraser, C and McDonald, I. (1972). Digestion of concentration in sheep. IV. The effect of urea on digestion, nitrogen retention and growth in young lambs. Br. J. Nutr. 27, 491-501.
3. Ørskov, E.R, Fraser, C and McDonald, I. and Smart, R.. (1972). Digestion of concentrate in sheep. V. The effect of adding fish meal and urea together to cereals diets on protein digestion and utilization by young sheep. Br. J. Nutr. 31, 89-98.
4. Mehrez, A.Z., Ørskov, E.R, and McDonald, I. (1977). Rates of rumen fermentation in relation to ammmonia concentration. Br. J. Nutr. 38, 437-443.
5. Mehrez, A.Z., and Ørskov, E.R. (1978). Protein degradation and optimum urea concentration in cereal-based diets foe sheep. Br. J. Nutr. 40, 337-345.

24
Hướng nghiên cứu 1. Phân giải và sử dụng protein và sự phát triển kỹ thuật túi ni lông Hướng nghiên cứu 1. 4. Kỹ thuật túi ni lông - một công cụ năng động để
nghiên cứu phân giải các chất dinh dưỡng trong dạ cỏ.
Chúng tôi sớm tự tin vào kỹ thuật nylon bag sau khi sử dụng cho các
nghiên cứu phân giải lúa mạch (1.3). Nhằm tăng tính sắc bén của kỹ thuật, chúng tôi nghiên cứu các đặc điểm khác nhau của kỹ thuật như biến động trong một gia súc, giữa các túi và giữa các gia súc, và dĩ nhiên chúng tôi thấy rằng biến động giữa các gia súc là nguồn biến động lớn nhất (1). Đầu tiên chúng tôi sử dụng vải dù và nhận thấy vải nylon cũng có lỗ hổng trên túi tương tự. Chúng tôi đã sử dụng kỹ thuật này cho các nghiên cứu về cỏ và nghiên cứu về các khẩu phần protein (1) và chúng tôi áp dụng một loạt các thời điểm ủ mẫu khác nhau nhưng vấn đề là thời gian ủ mẫu nào cho tỷ lệ phân giải thực sự? Đối với hầu hết thức ăn bổ sung protein, thức ăn thô, phân giải dạ cỏ là một quá trình động.
Trong hướng nghiên cúu 3, các kỹ thuật chúng tôi phát triển nhằm xác định tỷ lệ thoát qua của các mảnh thức ăn nhỏ khỏi dạ cỏ. Chúng tôi thấy rằng bản chất của thức ăn và mức ăn vào có thể dẫn đến tỷ lệ thoát qua thay đổi từ 2 đến 10%/1 giờ. Chúng ta đã biết rằng tỷ lệ phân giải biến động giữa các nguồn protein. Chúng ta cũng biết rằng tốc độ thoát qua biến động với các điều kiện nuôi dưỡng.
Tôi đã đưa vấn đề chúng tôi gặp phải ra thảo luận với ông Ian McDonald, chuyên gia toán thống kê. Ông là người sử lý số liệu tốt nhất mà tôi đã từng làm việc. Ông là nhà toán học và phân tích có năng lực. Ông có kiến thức và cảm nhận tốt về sinh vật học. Ian McDonald luôn lắng nghe các vấn đề của các nhà nghiên cứu hơn là nói với họ vấn đề của họ là cái gì! Ông ta là người viết rất xuất sắc và tôi có lợi từ điều đó. Ian McDonald là đồng tác giả với tôi trong 25 báo cáo. Tôi viết bản phác thảo sau đó ông ta viết gọn lại và phân tích thống kê! Bây giờ vấn đề của tôi ở đây là: có hai hiện tượng xảy ra cùng một thời điểm trong thí nghiệm của tôi: tỷ lệ phân giải phụ thuộc vào nguồn protein, tỷ lệ thoát qua phụ thuộc vào hệ thống nuôi dưỡng và có thể các đặc điểm vật lý khác của khẩu phần, và một vài thức ăn bổ xung protein có phần hòa tan có thể bị phân giải trong thời gian rất ngắn trong dạ cỏ.
Ian đã tìm ra giải pháp cho vấn đề của tôi, đó là sử dụng phương trình:
p = a + b (1-e-ct)
Trong đó p là phần bị phân giải ở thời điểm t, a là phần bị phân giải tức thì), b là phần không hào tan nhưng có thể phân giải, e là logarit tự nhiên, c là hệ số tốc độ phân giải của b và t là thời gian. Nếu bây giờ chúng ta biểu diễn phân giải giải hiệu quả (P) – số lượng bị phân giải trong thời gian ở dạ cỏ, chúng ta có

25
P = a + (bc/c+k) Trong đó a, b và c đã nói ở trên và k là tốc độ thoát qua (2). Bìa báo viết về
vấn đề này là bài báo đuợc trích dẫn nhiều nhất của tôi, chỉ có 4 trang! Phương trình có được các lợi ích từ thực tế rằng tất cả các chữ của phương trình có ý nghĩa riêng, và tất cả mọi người đều hiểu được nó ngay cả những người không phải là nhà toán học hay nhà mô hình hóa! Chúng ta bây giờ có thể đưa ra các bảng về phan giải có hiệu quả và sử dụng chúng để tăng hiệu quả của các hệ thống nuôi dưỡng khác nhau. Chi tiết hơn đã được trình bày trong sách của tôi về chủ đề này (3). Phương pháp nylon bag đã trở thành những điểm xuất phát cho một vài hướng nghiên cứu mới.
Một vài ví dụ phân giải protein ở các tốc độ thoát qua khác nhau được trình bày trong Bảng 1.10. Ở đây có thể thấy rằng việc thức ăn bổ sung protein như bột cá chất lượng cao không bị ảnh hưởng nhiều bởi tốc độ thoát qua bởi vì phần lớn bột cá không bị phân giải trong khi các loại protein thực vật khác bị ảnh hưởng rất nhiều bởi tỷ lệ thoát qua tùy thuộc vào c - hệ số tốc độ phân giải của b. Nói cách khác phân giải protein dạ cỏ là không cố định, không tĩnh, mà rất động và phụ thuộc vào các điều kiện nuôi dưỡng đặc biệt là mức nuôi dưỡng vì mức nuôi dưỡng quyết định tỷ lệ thoát qua của các mảnh thức ăn kích thước nhỏ. Bảng 1.10. Tỷ lệ phân giải của các thức ăn bổ sung protein khác nhau ở các tốc độ
thoát qua khác nhau (3). Tỷ lệ thoát qua (k) 0,02 0,05 0,08 Bột cá 22,7 22,0 21,5 Hạt bông 80,6 69,6 62,7 Hạt lanh 78,1 58,9 46,0 Bột đậu tơng 80,8 62,5 50,4 Lạc 87,4 74,1 64,3
Tài liệu tham khảo cho phần 1.4
1. Mehrez, A.Z., and ỉrskov, E.R. 1977. The use of a Dacron bag technique to determine rate of degradation of protein and energy in the rumen. J. Agri. Sci., Camb. 88, 645-650.
2. Ørskov, E.R. and McDonald, I. 1979. the estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci., Camb. 92, 499-503.
3. Ørskov, E.R. 1982. Protein Nutrition in Ruminants. Academic Press, London, New York.

26
Hướng nghiên cứu 2. Sử dụng phương pháp nylon bag cho các nghiên cứu môi trường dạ cỏ và đánh giá thức ăn thô
Hướng nghiên cứu 2.1 Tương tác của thức ăn Những ảnh hưởng kết hợp bất lợi
Khái niệm ảnh hưởng kết hợp, một cách đơn giản là những ảnh hưởng thêm ra khi hai loại thức ăn được cho ăn cùng nhau, chúng tôi phát hiên ra ảnh hưởng này khi thực hiện những nghiên cứu (năm 1975),. Những ảnh hưởng kết hợp luôn luôn bất lợi và thường thấy khi thức ăn thô và thức ăn tinh được cho cùng nhau. Lý do được nghĩ đến là do pH thấp từ quá trình lên men thức ăn tinh đã ảnh hưởng và làm giảm tiêu hoá thức ăn thô. Tôi thực sự không có bất cứ ước muốn định trước nào tham gia vào các nghiên cứu về ảnh hưởng kết hợp vì có vẻ như là đây là chủ đề không mấy hấp dẫn cho đến khi chúng tôi nhận thấy rằng đã có một phương pháp có thể thực hiện các cuộc đột nhập vào vấn đề nêu trên! Đó là phương pháp nylon bag, phương pháp này cho đến lúc đó chúng tôi chỉ mới sử dụng để xác định sự phân giải protein. Để nghiên cứu ảnh hưởng kết hợp chúng tôi giữ môi trường dạ cỏ ổn định với khả năng có thể và thay đổi mẫu thức ăn ủ theo ý muốn, nhưng có thể chúng tôi sẽ sử dụng phương pháp theo cách ngược lại: giữ nguyên mẫu ủ và thay đổi môi trường dạ cỏ. Vào thời điểm đó chúng tôi đang ở nửa đường nghiên cứu nuôi cừu với thức ăn ngũ cốc không qua chế biến và đã thấy rằng pH dạ cỏ vào khoảng 6,2 khi nuôi cừu bằng ngũ cốc nguyên hạt và 5,2 khi nuôi cừu bằng hạt ngũ cốc đã qua chế biến. pH đã ảnh hưởng đến tiêu hoá cellulose như thế nào? Chúng tôi đã không phát triển phương trình mô tả tốc độ phân giải nhưng chúng tôi đã có kết quả đủ rõ ràng khi chúng tôi sử dụng cỏ khô là thức ăn ủ trong túi nylon và nuôi cừu mổ lỗ dò với lúa mạch viên hoặc nguyên hạt. Bảng 2.1. Lượng mất đi của cỏ khô (mg/g) từ các túi nylon được ủ trong dạ cỏ cừu
nuôi với lúa mạch viên hoặc nguyên hạt (1)
Thời gian ủ (h) Lúa mạch nguyên hạt Lúa mạch viên SE 6 285 242 36 12 453 373 11 18 530 406 20 24 625 423 23
Những sự khác nhau trong tiêu hoá thức ăn thô đã dẫn đến những thí
nghiệm nuôi dưỡng cuối cùng sử dụng các khẩu phần cỏ khô được bổ sung lúa mạch nguyên hạt hoặc dạng viên (2) như trình bày trong hướng nghiên cứu 7 về chế biến ngũ cốc. Tuy nhiên, có thể tránh cái gọi là ảnh hưởng kết hợp hay không?
Thí nghiệm nhỏ tiếp theo cụ thể hơn một chút và ở đây như trước đó chúng tôi yêu cầu sinh viên thạc sĩ, trong trường hợp này là Andrew Chimwano từ

27
Zambia, nghiên cứu sâu hơn nữa. Thí nghiệm của Chimwano (2) được thiết lập nhằm xem xét lượng ngũ cốc đã chế biến có thể cho ăn là bao nhiêu trước khi tiêu hoá thức ăn thô bị ức chế (Hình 2.1).
Tăng lượng lúa mạch lên chút ít đã có chút ít hoặc không ảnh hưởng đến phân giải, khi tiếp tục tăng lượng lúa mạch lên phân giải giảm tuyến tính với lượng lúa mạch tăng lên. Khác với cỏ khô, phân giải của sợi bông giảm ở tất cả các mức tăng lúa mạch. Phải nhớ rằng cỏ khô chứa khoảng 40 đến 50% phần hòa tan do đó mức lúa mạch cao nhất tiêu hoá cellulose trong cỏ khô cũng bằng 0 như sợi bông.
Sơ đồ 2.1 Ảnh hưởng của lượng lúa mạch chế biến và cỏ khô đến lượng chất khô mất đi từ cỏ khô và sợi bông được ủ trong các túi nylon trong dạ cỏ 24 giờ. (mg/g
được ủ)
Kết quả đáng khích lệ. Phương pháp có thể sử dụng để xác định cách quản lý thức ăn tối ưuu và xác định phương pháp chế biến nhằm tối ưu hoá tiêu hoá xơ khi thức ăn thô được cho ăn với thức ăn tinh. Rõ ràng chế biến ngũ cốc là phương

28
pháp hiển nhiên. Nghiên cứu này cho chúng ta nhận ra rằng phương pháp nylon bag đơn giản có thể sử dụng cho mục đích dẫn đến nhiều tiến bộ trong sử dụng thức ăn nói chung.
Tiếp theo thí nghiệm trên, chúng tôi nỗ lực hơn nữa và ở đây tôi đã có cơ hội tốt đó có được 3 nghiên cứu sinh tiến sĩ xuất sắc, Fergus Mould từ Scotland, Louis Istasse từ Bỉ và Ayona Silva từ Sri Lanka. Fergus Mould là người đầu tiên xác định pH dạ cỏ in vivo, tại giá trị pH đó quá trình phân giải cellulose bị ức chế (4). Cừu được nuôi bằng cỏ khô cho pH dạ cỏ khoảng 6,6, cừu được nuôi bằng lúa mạch viên cho pH dạ cỏ 5,2. Bằng cách sử dụng axit khoáng Fergus Mould đã từ từ làm giảm pH của cừu ăn cỏ từ 6,6 còn khoảng 5,2. Sau đó Fergus Mould, có thể là lần đầu tiên, xác định giá trị pH dạ cỏ ở đó quá trình phân giải cellulose bắt đầu bị ức chế, cụ thể là pH từ 6,2 đến 6,3; và xác định được giá trị pH mà tại đó quá trình phân giải cellulo giảm xuống gần như bằng 0, đó là khi pH dưới 6,0. Ngược lại pH dạ cỏ tăng dần từ 5,2 đến 6,6 ở khẩu phần lúa mạch viên đã có ảnh hưởng tương đối nhỏ đến phân giải cellulose và thực ra là đến hệ vi sinh vật dạ cỏ. Tinh bột ở dạng lơ lửng có ảnh hưởng lớn đến hệ vi sinh vật dạ cỏ. Tiêu hoá cellulose không thể được phục hồi khi hệ vi sinh vật không thay đổi nhiều và thành phần các axit béo bay hơi ít thay đổi. Điều này khác với việc nuôi gia súc bằng ngũ cốc nguyên hạt với pH tương tự tiêu hoá cellulose tích cực vẫn được duy trì (Bảng 2.1).
Như vậy, cả chất nền trong dịch dạ cỏ và pH là quan trọng. Nuôi gia súc bằng lúa mạch nguyên hạt, cho pH dạ cỏ cao hơn, đã có ảnh hưởng khác nhau đến hệ vi sinh vật hơn là khẩu phần lúa mạch viên và sau đó phục hồi pH đến giá trị cao hơn.
Khi pH giảm ở khẩu phần cỏ, không có sự thay đổi thành phần lớn nào ở hệ vi sinh vật nhưng hệ vi sinh vật giảm đáng kể về số lượng và khả năng sống khi pH nhỏ hơn 6,0. Vật chất khô mất từ cỏ khô trong các túi nylon bag ở thời điểm 24 giờ là khoảng 300mg/g ở pH 6,5 trong dạ cỏ của cừu nuôi khẩu phần cơ sở là lúa mạch và khoảng 600 mg/g ở dạ cỏ của cừu nuôi bằng cỏ ở pH 6,5. Như vậy, pH không phải là vấn đề duy nhất ảnh hưởng đến tiêu hoá cellulose.
Sau đó chúng tôi nghiên cứu xem lượng lúa mạch có thể dùng trong khẩu phần cơ bản là cỏ khô trước khi tiêu hoá xơ bị giảm là bao nhiêu, và lượng lúa mạch này phụ thuộc ở mức độ nào vào chế biến ngũ cốc. Ở đây chúng tôi thấy rằng với lượng lúa mạch 50% trong khẩu phần không thể phát hiện sự khác nhau về phân giải cellulose do chế biến; nhưng ở thời điểm ủ 24 giờ, phân giải cỏ khô trong túi giảm từ 51 xuống 40%. Khi lúa mạch chiếm 75%, sự khác nhau về phân giải do chế biến trở nên rất rõ ràng và giá trị pH dạ cỏ cũng khác nhau. Ở mức 75% lúa mạch trong khẩu phần, giá trị pH dạ cỏ là 6,2 với lúa mạch nguyên hạt và 5,5 với lúa mạch nghiền và ép thành viên. Vật chất khô tương ứng mất đi từ các túi nylon là 36 và 24%. Giá trị 24% phần lớn là phần hoà tan mất đi từ cỏ khô. Nói cách khác gần như không có tiêu hoá xơ (5).
Thí nghiệm tiếp theo nhằm sử dụng các nguồn xơ khác nhau, cỏ khô, cỏ khô chất lượng cao, cỏ khô chất lượng thấp, và rơm bổ sung bằng 65% lúa mạch

29
nguyên hạt, lúa mạch viên, ngô nguyên hạt, ngô viên hoặc rỉ mật. Mẫu thức ăn ủ trong dạ cỏ là thức ăn thô gia súc đang ăn. Sau khi cho ăn khẩu phần 65% ngũ cốc, bicarbonate được đưa vào dạ cỏ để tăng pH dạ cỏ tới 6,6-6,7 để xem liệu tiêu hoá cellulose có được phục hồi bởi khi pH đạt mức tối ưu. Hai kết luận ở đây là:
1. So với khẩu phần chỉ có thức ăn thô, mức thức ăn tinh có thể làm giảm phân giải thức ăn thô phụ thuộc vào mức giảm pH dạ cỏ do tốc độ hoà tan của thức ăn tinh và phụ thuộc vào loại thức ăn thô. Giảm tiêu hóa lớn nhất xẩy ra với thức ăn thô chất lượng kém và khi rỉ mật là nguồn thức ăn tinh trong khẩu phần.
2. Giảm đáng kể tỷ lệ phân giải cellulose có thể đảo ngược bằng cách đưa bicarbonate vào dạ cỏ để tăng pH, nhưng phục hồi pH đến 6,7 không cải thiện được phân giải cellulose khi làm t\có ảnh hởng đến sự suy giảm phân giải khi rỉ mật là nguồn thức ăn tinh trong khẩu phần.
Thí nghiệm cuối cùng là thí nghiệm in vivo (6) nhằm xem xét xem liệu các
nguyên tắc đã được xác định có đúng trong thực tế không. Bảng 2.2 trình bày ảnh hưởng của mức lúa mạch, chế biến thức ăn thô và bổ sung bicarbonate. Tỷ lệ tiêu hoá mong đợi được tính toán với các giả thiết về tương tác từ các thức ăn riêng biệt. Vì bất cứ ảnh hưởng nào đến tỷ lệ tiêu hoá đều do giảm tiêu hóa xơ, vì vậy tỷ lệ tiêu hoá mong đợi cũng được tính toán trên cơ sở giảm tiêu hoá cỏ khô. Bicarbonate đã không có ảnh hưởng với cỏ khô được chặt nhỏ nên không trình bày trong bảng 2.2. Bảng 2.2. Ảnh hưởng của mức lúa mạch ép viên và bicarbonate đến tỷ lệ tiêu hoá
mong đợi và tỷ lệ tiêu hóa đo được in vivo.
Khẩu phần cỏ khô
Lúa mạch (%)
Bicarbonate Tỷ lệ tiêu hoá của
toàn khẩu phần (đo được)
Tỷ lệ tiêu hoá mong đợi của
toàn khẩu phần
Thay đổi tỷ lệ tiêu
hoá (%)
Cỏ khô chặt 0 - 58 58 0 44 - 66 67 -3 62 - 70 71 -4 78 - 73 74 -5
Cỏ khô nghiền
0 - 52 52 0
33 - 56 61 -13 46 - 58 64 -23 57 - 59 67 -37
Cỏ khô nghiền
0 + 47 47 0
37 + 59 59 -1

30
50 + 63 63 + 1 60 + 67 66 -3
Ở đây tầm quan trọng của bài tiết nước bọt là rất rõ ràng. Lúa mạch làm
giảm tỷ lệ tiêu hóa tương đối nhỏ ở khẩu phần cỏ khô chặt, cỏ khô chặt kích thích tiết ra nhiều nước bọt. Cỏ khô nghiền cũng không làm giảm tỷ lệ tiêu hóa nhiều. Có thể thấy rằng nghiền thức ăn thô không chỉ giảm tỷ lệ tiêu hoá của nó khi nó được cho ăn như thức ăn duy nhất mà còn làm giảm tỷ lệ tiêu hoá nhiều hơn khi thức ăn tinh được cho thêm khẩu phần cỏ khô nghiền. Tuy nhiên, giảm tỷ lệ tiêu hóa có thể phục hồi bằng cách bổ sung dung dịch đệm bicarbonate vào dạ cỏ để thay thế cho việc nước bọt giảm do nghiền thức ăn thô.
Cuối cùng chúng tôi nghiên cứu xác định ảnh hưởng tương tác của chế biến ngũ cốc và chế biến thức ăn thô. (Bảng 2.3). Bảng 2.3. Ảnh hưởng của chế biến cỏ khô và chế biến lúa mạch đến việc giảm tỷ
lệ tiêu hoá.
Tỷ lệ ngũ cốc (%)
Tỷ lệ cỏ khô (%)
Chế biến cỏ khô
Chế biến lúa mạch
Giảm tỷ lệ tiêu hóa tổng số
(%)
Giảm tỷ lệ tiêu hóa cỏ
khô (%)
34 66 Dài Nguyên hạt 7,7 12,9 67 33 Dài Nguyên hạt 2,9 10,9 34 66 Dài Viên 4,2 6,9 67 33 Dài Viên 8,1 29,7 34 66 Nghiền Nguyên hạt 3,8 6,9 67 33 Nghiền Nguyên hạt 5,4 22,4 34 66 Nghiền Viên 6,0 10,9 67 33 Nghiền Viên 9,1 38,8
Ở đây chúng tôi đã có thể xác định rõ ràng có 3 nhân tố ảnh hưởng đến
tương tác và tiêu hoá cellulose. 1. Thức ăn tinh càng nhiều, tiêu hoá xơ càng giảm (Hình 2.1). 2. Thức ăn thô được chế biến càng kỹ, tỷ lệ tiêu hoá càng giảm (Bảng 2.3). 3. Thức ăn tinh càng chế biến kỹ, trong trường hợp này là lúa mạch, tỷ lệ tiêu hoá cellulose càng giảm mạnh (Bảng 2.3).
Các bước tiếp theo trong nghiên cứu ảnh hưởng kết hợp bất lợi do Istasse (7) đảm nhiệm, anh ta thêm một yếu tố khác là quản lý nuôi dưỡng vào thí nghiệm. Anh ta cố gắng cho ăn thức ăn tinh 2 hoặc 4 lần mỗi ngày, hoặc trộn thật kỹ thức ăn thô với thức ăn tinh. Sử dụng cừu mổ lỗ dò, pH dạ cỏ được đo 2 giờ/lần và một loại cỏ khô đã tách phần hòa tan được sử dụng để ủ trong các túi nylon (Bảng 2.4).

31
Bảng 2.4. Ảnh hưởng của phương pháp cho ăn thức ăn tinh đến pH dạ cỏ, thời gian giảm pH xuống dưới 6, tổng thời gian pH dưới 6 và phân giải vật chất khô từ
các túi nylon ủ trong 18 và 24 giờ.
Phương pháp cho
ăn
pH trung bình
Biến động của
pH
Thời gian pH dưới 6 (giờ)
Mức độ giảm pH dưới 6 (pH x giờ)
Tỷ lệ phân giải (%) trong
24 giờ 48 giờ
2 lần 6,0 5,4-6,5 14 4,10 18,7 31,8 4 lần 5,9 5,6-6,3 21 4,64 18,2 29,0
Hỗn hợp 5,8 5,9-6,3 19 8,75 10,3 21,9
Sự khám phá đúng là ở đây. Chúng tôi đã nghi nghờ rằng sự ổn định pH dạ cỏ đạt được với một khẩu phần trộn hoàn toàn sẽ cho tỷ lệ tiêu hoá cellulose tốt nhất nhưng với 65% thức ăn tinh sử dụng ở đây pH thấp hơn 6,0 trong phần lớn thời gian; do đó tổng số lần pH giảm, độ dài thời gian pH dưới 6 lớn nhất và mức độ giảm pH lớn nhất với khẩu phần trọn thô tinh.
Khẩu phần trộn hoàn toàn có thể cho một môi trường dạ cỏ ổn định nhưng không nhất thiết là điều kiện tốt nhất cho tỷ lệ tiêu hoá tối ưu. Với kiểu cho ăn thức ăn tinh không liên tục (2 lần/ngày), tiêu hoá xơ được phục hồi trong phần lớn thời gian của ngày so với các kiểu cho ăn khác do đó tỷ lệ tiêu hoá có thể cao nhất khi tỷ lệ thức ăn tinh trong khẩu phần cao. Tương quan cao nhất có được là giữa giảm phân giải và mức độ giảm pH dưới 6 (r = 0,74).
Nhằm xem có sự khác nhau tương tự ở mức thức ăn tinh thấp hơn, một thí nghiệm khác được tiến hành, trong thí nghiệm này tỷ lệ tiêu hoá in vivo cũng được nghiên cứu (Bảng2.5).
Bảng 2.5. Ảnh hưởng của mức thức ăn tinh trong khẩu phần sơ sở lúa mạch và phương pháp cho ăn đến tỷ lệ phân giải vật chất khô ở thời điểm 24 và 48 giờ
trong các túi nylon và tỷ lệ tiêu hoá in vivo.
Mức thức ăn tinh (%)
Phương pháp cho ăn thức ăn tinh
Tỷ lệ phân giải ở Tỷ lệ tiêu hóa chất khô của cỏ khô (%)
24 giờ 48 giờ
40 2 lần/ngày 35,0 49,4 52,6 40 Trộn với thức ăn thô 35,1 50,4 52,0 65 2 lần/ngày 30,3 44,9 48,2 65 Trộn với thức ăn thô 18,0 32,9 42,0 SE 1,7 1,9 1,8
Ở đây một lần nữa cho thấy rõ ràng một lần nữa là ở mức thức ăn tinh thấp

32
hơn, phương pháp cho ăn không quan trọng; pH dạ cỏ rõ ràng không giảm tới mức có thể hạn chế tiêu hoá. Ở mức thức ăn tinh cao hơn, một lần nữa cho thấy khẩu phần hỗn hợp hoàn toàn không cho tỷ lệ tiêu hoá tối ưu mặc dù nó có thể giúp ổn định pH dạ cỏ. Cuối cùng chúng tôi chuyển sự chú ý sang bò sữa (8) để đánh giá ảnh hưởng của tỷ lệ thức ăn tinh – cỏ khô và phương thức cho ăn (Bảng 2.6). Ở đây thật không may chúng tôi không có bò mổ lỗ dò để đo pH dạ cỏ. Bảng 2.6. Ảnh hưởng của tỷ lệ thức ăn tinh và phương pháp cho ăn đến khả năng
suất bò sữa
Tỷ lệ thức ăn tinh trong toàn bộ khẩu
phần DM
Phương pháp cho ăn thức ăn tinh
SE
0,40 0,65 Trộn 2 lần/ngày Năng suất sữa (kg/ngày) 23,8 26,2 26,2 23,8 0,77Mỡ sữa (%) 3,79 3,57 3,58 3,79 0,11Chất khô ăn vào (kg/ngày) 16,2 16,7 17,8 15,2 0,3 Tỷ lệ tiêu hoá DM (%) 58,5 61,4 61,0 58,9 0,6 Tỷ lệ tiêu hoá cỏ khô* 46,8 38,9 43,3 42,4 -
* Được tính toán với giả thiết rằng bất kỳ sự khác nhau nào từ việc bổ xung là do giảm tỷ lệ tiêu hoá cỏ khô.
Ở đây, chúng tôi thấy rằng trong trường hợp này tỷ lệ tiêu hoá không giảm với khẩu phần hỗn hợp hoàn chỉnh (trộn thức ăn tinh và thô) ở mức 65% thức ăn tinh trong vật chất khô khẩu phần. Tỷ lệ tiêu hoá của cỏ khô giảm khi tăng tỷ lệ thức ăn tinh trong khẩu phần và sản lượng sữa cao hơn khi bò ăn khẩu phần hỗn hợp hoàn chỉnh. Sự khác nhau trong cấu trúc thức ăn thô do việc nghiền thức ăn thô để tạo khẩu phần hỗn hợp hoàn chỉnh đã ảnh hưởng đến pH dạ cỏ.
Bản chất của thức ăn tinh cũng rất quan trọng (Hình 2.2), như Fahmy (9) đã chúng minh. Cừu mổ lỗ dò được nuôi bằng khẩu phần với các tỷ lệ lúa mạch hoặc củ cải đường tăng dần, và phân giải chất khô trong túi nylon được đo đạc. Thức ăn thô là rơm sử lý ammoniac. Sơ đồ 2.2. Ảnh hưởng của mức củ cải đường hoặc lúa mạch xay đến phân giải rơm
ủ ammoniac trong túi ny lon trong 24 giờ (mg/g mẫu ủ)

33
Ở đây chúng tôi thấy rằng tỷ lệ lớn hơn nữa củ cải đường có thể thêm vào khẩu phần trước khi giảm tỷ lệ phân giải rơm xảy ra. Điều này được thể hiện rất rõ trong thí nghiệm in vivo khi cừu được nuôi với khẩu phần hỗn hợp hoàn chỉnh có 30% rơm sử lý ammoniac và 70% củ cải đường hoặc lúa mạch xay (9) Bảng 2.7. Bảng 2.7. Ảnh hưởng của tỷ lệ củ cải đường hoặc lúa mạch xay đến tỷ lệ tiêu hoá
của khẩu phần cơ sở là rơm sử lý ammoniac. Khẩu phần Tỷ lệ tiêu hoá
toàn khẩu phần (%)
Tỷ lệ tiêu hoá toàn khẩu phần dự đoán có tính đến ảnh hưởng
kết hợp
Tỷ lệ tiêu hoá rơm theo tính
toán (%)
Chỉ có rơm xử lý ammoniac (AS)
53,3 53,3 53,3
AS + 70% củ cải đường 70,5 74,5 40,3 AS + 70% lúa mạch xay 65,4 74,9 21,8 SE 1,7
Tỷ lệ tiêu hoá rơm sử lý ammoniac tính được giảm từ 53,3 xuống 40,3% do
bổ sung thêm củ cải đường, nhưng tỷ lệ này giảm nhiều hơn, giảm đến 21,8% do bổ sung lúa mạch xay. Có khả năng tỷ lệ tiêu hoá của rơm không xử lý sẽ giảm tới các mức thấp tương tự như vậy, do đó nâng cao chất lượng rơm bằng cách ủ ammoniac là không hợp lý. Ảnh hưởng khác nhau của các loại thức ăn tinh đến

34
tiêu hoá cellulose sẽ phải được tính đến khi thức ăn tinh được sử dụng cho bò sữa. Ảnh hưởng kết hợp bất lợi trở nên phức tạp hơn chúng tôi dự đoán nhưng
phương pháp nylon bag là công cụ hữu hiệu, phương pháp này cho phép nghiên cứu nhiều nhân tố, những nhân tố này có thể gây trở ngại cho tiêu hoá cellulose khi thức ăn tinh có trong khẩu phần.
1. Nhân tố quan trọng nhất là pH dạ cỏ. Giá trị pH dạ cỏ có thể được điều chỉnh trong một phạm vi nhất định, để ít khi pH dưới 6,2 là pH mà tại đó tiêu hoá xơ bắt đầu bị ức chế.
2. pH dạ cỏ có thể điều chỉnh bằng quản lý nuôi dưỡng, tỷ lệ thức ăn tinh, chế biến thức ăn tinh, chế biến thức ăn thô và loại thức ăn tinh.
3. pH dạ cỏ bị ảnh hưởng bởi tốc độ mà tại đó carbonhydrate trong thức ăn tinh trở nên hoà tan (rỉ mật hòa tan nhiều hơn ngũ cốc), và dĩ nhiên cũng bị ảnh hưởng bởi tỷ lệ thực của thức ăn tinh trong khẩu phần và mức độ chế biến thức ăn tinh.
4. Lưu tâm đến các vấn đề nêu trên có thể có những ảnh hưởng lớn đến tiêu hoá các chất dinh dưỡng của thức ăn và nói chung ảnh hưởng đến chi phí để nghiền thức ăn. Trong thực tế, chế biến quá kỹ sự hoặc quá tốn kém thức ăn tinh và thô là những yếu tố góp phần lớn nhất làm cho dụng thức ăn không hiệu quả.
Những ảnh hưởng kết hợp tích cực Ảnh hưởng kết hợp tích cực xảy ra khi tỷ lệ tiêu hoá hoặc lượng thức ăn ăn
vào của một khẩu phần hỗn hợp hoàn chỉnh cao hơn tổng tỷ lệ tiêu hoá hoặc lượng thức ăn ăn vào của các thức ăn thành phần khi cho ăn riêng lẻ. Một lần nữa, lúc đầu tôi không thực sự thích thú với vấn đề này, và tôi đã bắt đầu một nghiên cứu như là một nhiệm vụ được giao hơn là một niềm say mê! Nhưng một cách tình cờ sau đó chúng tôi đã tìm ra một mỏ vàng! Ayona Silva, trong luận văn thạc sĩ của mình tiến hành một thí nghiệm nhỏ trong đó cừu mổ lỗ dò được nuôi với cỏ khô, rơm sử lý ammoniac hoặc rơm không sử lý, nhưng để chắc chắn rằng ammonia dạ cỏ không phải là yếu tố hạn chế, urê 1% được phun vào cỏ khô và rơm không sử lý. Sau đó Ayona Silva ủ các mẫu thức ăn trong túi nylon trong dạ cỏ của cừu được nuôi với 3 khẩu phần trên. Đây là kết quả cô đã đạt được (10). (Bảng 2.8). Bảng 2.8. Ảnh hưởng của khẩu phần đến nồng độ NH3, pH dạ cỏ và phân giải thức
ăn.
Khẩu phần NH3 dạ
cỏ (mg/100
ml)
pH dạ cỏ
Phân giải chất khô sau 48 giờ (mg/g) mẫu ủ
Rơm không xử
lý
Rơm xử lý ammoniac
Cỏ khô
Rơm không xử lý + 1% urê
26,8 6,9 44,3 541 495
Rơm xử lý ammoniac 24,4 6,8 607 648 629

35
(3% NH3) Cỏ khô + 1% urê 21,2 6,5 605 652 638 SE 1,9 0,05 17 19 21
Ở đây chúng tôi có được một quan sát thú vị là rơm không xử lý mất đi một
cách nhanh hơn trong dạ cỏ cừu nuôi bằng rơm xử lý ammoniac so với ở cừu nuôi bằng rơm không xử lý, thậm chí nồng độ ammonia và pH dạ cỏ là thuận lợi tương đương trong cả hai trường hợp. Do vậy, bây giờ chúng tôi có thể theo đuổi một loại nghiên cứu khác, để xem thức ăn nào khi bổ sung vào khẩu phần rơm không xử lý sẽ tạo ra môi trờng dạ cỏ tốt như môi trường dạ cỏ ở cừu nuôi bằng rơm xử lý ammoniac và thực sự cơ chế ở đây là gì?
Để cho đơn giản, tôi đã tóm tắt trong Bảng 2.9, một thí nghiệm với bã củ cải đường, thí nghiệm đã chỉ ra ảnh hưởng của rơm không xử lý và rơm xử lý ammoniac đến tỷ lệ tiêu hoá và lượng thức ăn ăn vào (11). Lượng thức ăn ăn vào của rơm không xử lý tăng lên khi bổ sung bã củ cải đường, có lẽ do môi trường dạ cỏ được cải thiện, nhưng lượng thức ăn ăn vào và tỷ lệ tiêu hoá của rơm xử lý ammoniac không tăng khi bổ sung củ cải đường vì môi trờng dạ cỏ đã được tối ưu hoá nhờ quá trình ammoniac hóa.
Bảng 2.9. Ảnh hưởng của bổ xung bã củ cải đường đến tỷ lệ tiêu hoá và lượng
thức ăn ăn vào của rơm xử lý và không xử lý ở cừu. Khẩu phần Tỷ lệ tiêu hóa
chất khô (%) Lượng thức ăn ăn vào (g/ngày)
Rơm Tổng Rơm không xử lý 49 414 414 Rơm không xử lý + 15% củ cải đường
54 505 589
Rơm xử lý ammonia 57 729 729 Rơm xử lý ammonia + 15% củ cải đờng
56 717 852
SE 1.2 30 29
Cuối cùng, một loạt các thức ăn bổ xung đã được sử dụng nhằm xác định xem thức ăn bổ sung nào ảnh hưởng đến tốc độ phân giải rơm lúa mạch không xử lý trong túi nylon dạ cỏ (11). Chúng tôi đã thử nghiệm nhiều loại thức ăn bổ sung như bã củ cải đường, bã cam quýt, cỏ khô, bột đậu tương và bột cá. Bã củ cải đường và cỏ khô ảnh hưởng đáng kể đến tốc độ phân giải. Bã cam quýt cũng như bột đậu tương và bột cá đã không có ảnh hưởng đến tốc độ phân giải (Bảng 2.10). Kết quả cho thấy rằng nếu chúng ta bổ xung bã củ cải đường hoặc cỏ khô, tốc độ phân giải của rơm không sử lý ủ trong túi nylon đặt trong dạ cỏ cừu được cho ăn rơm không sử lý và thức ăn bổ xung tương tự tốc độ phân giải của rơm không sử lý ủ trong túi nylon đặt trong dạ cỏ cừu ăn rơm xử lý ammoniac. Hình như, vấn đề xẩy ra ở đây là xơ dễ lên men và/hoặc pectin làm đã làm tăng hoạt động của vi

36
sinh vật phân giải cellulose trong dịch dạ cỏ vì vậy thức ăn mới được ăn vào bị vi sinh vật xâm nhập và lên men nhanh chóng hơn. Nếu quả thực là như vậy, thì lượng thức ăn ăn vào của các thức ăn trong khẩu phần cơ sở phải tăng.
Bảng 2.10. Ảnh hưởng của bổ sung đến phân giải vật chất khô của rơm không sử
lý, pH và nồng độ NH3 dạ cỏ. Thức ăn bổ xung g thức ăn bổ
xung/kg khẩu phần
Phân giải tại pH dạ cỏ
Nồng độ NH3 dạ cỏ
(mg/100ml) 24 giờ (%) 48 giờ (%)
Không bổ sung 0 38,6 46,3 6,9 22,6 Bã củ cải đường 150 40,7 50,3 6,8 Không bổ xung 0 41,0 48,8 6,8 21,7 Cỏ khô 150 45,8 56,3 6,9 14,5 Không bổ xung 0 44,3 51,7 6,8 22,5 Cam quýt 150 44,3 53,4 7,0 21,0 Không bổ xung 0 40,8 49,6 6,7 18,6 Bột đậu tương 100 40,2 49,5 6,9 29,1 Không bổ xung 0 50,5 57,2 6,9 19,4 Bột cá 100 52,5 59,5 6,9 30,7 SE 0,7 0,7 0,3 1,6
Trong khi thực hiện những thí nghiệm này, chúng tôi chợt nảy ra ý nghĩ tìm
ra kỹ thuật cải tiến để xác định hoạt động phân giải cellulose. Chúng ta đã biết rằng vi sinh vật bám một cách chắc chắn vào rơm vì hàm lượng nitơ của rơm luôn tăng theo thời gian ủ trong các túi ny lon ủ trong dạ cỏ, thậm trí ngay cả sau khi rửa túi cẩn thận. Liệu chúng ta có thể, thay vì xác định số lượng vi khuẩn ở trên rơm, đơn giản là chúng ta chỉ đo hoạt lực enzyme của chúng? Chúng tôi chọn cách xác định hoạt lực của enzyme glutamate dehydrogenase (GDH), vì enzyme phổ biến này tham gia vào quá trình hấp thụ ammonia của vi sinh vật phân giải cellulose (12). Chúng tôi hy vọng rằng phương pháp này sẽ nhậy hơn kỹ thuật nylon bag. Chúng tôi ủ rơm trong các túi nylon trong dạ cỏ của cừu nuôi bằng lúa mạch hoặc bằng thức ăn thô. Điều làm chúng tôi ngạc nhiên là hoạt lực GDH ở trên rơm đã rửa từ các túi nylon cao hơn khi rơm được ủ trong dạ cỏ cừu cho ăn ngũ cốc, mặc dù có ít hoặc không có sự phân giải cellulose ở cừu cho ăn ngũ cốc, so với cừu chỉ được nuôi với thức ăn thô. Lý do đơn giản là vi khuẩn không phân giải cellulose cũng bám chặt vào rơm mặc dù chúng không lên men xơ nhưng lên men tinh bột trong dạ cỏ. Vì lý do này, chúng tôi chuyển sang sử dụng hoạt lực của enzyme carboxymethyl cellulase (CMC). Đây là enzyme tốt hơn và rất nhạy. Tương quan giữa hoạt lực của CMC và phân giải cellulose dạ cỏ của các thức ăn khác nhau là r = 0,94 ở 24 giờ phân giải và r = 0,85 ở 48 giờ phân giải. Hoạt lực CMC phải đại diện một cách chính xác hoạt động của vi sinh vật phân giải cellulose và dễ xác định hơn việc đếm vi sinh vật nhiều. Có được phương pháp

37
mới này, chúng tôi trở lại với rơm xử lý ammoniac. Trong một vài chuyến đi đến Chân Phi tôi thấy rằng gần như tất cả phụ phẩm trồng trọt được sử dụng cho gia súc ngay tại chỗ và thức ăn bổ xung không sẵn có. Liệu rơm hoặc thân cây ngô xử lý ammoniac có thể sử dụng làm thức ăn bổ sung cho gia súc chăn thả trên cánh đồng sau vụ thu hoạch ăn các phế phụ phẩm trồng trọt (rơm rạ khôngb xử lý)?
Một thí nghiệm nhỏ do Benjamin Manyuchi tiến hành (13) trong đó rơm bổ xung urê cho cừu ăn tự do và rơm sử lý ammoniac được cho cừu ăn như thức ăn bổ xung. (Bảng 2.11).
Bảng 2.11. Ảnh hưởng của cho ăn rơm xử lý ammoniac đến lượng thức ăn ăn vào
của rơm không xử lý, tổng lượng vật chất khô ăn vào và hoạt lực của cellulase carbonxymethyl (CMC).
Lượng rơm
xử lý ammoniac
cho ăn (g/ngày)
Rơm không xử lý ăn vào (g DM/ngày)
Rơm xử lý ammoniac ăn
vào (gDM/ngày)
Tổng vật chất khô ăn vào
(g/ngày)
Hoạt lực CMC
0 641 0 641 114 200 816 187 1003 152 400 667 352 1019 147 Ăn tự do - 1152 1152 146 SE 7,2 73 50
Ở đây chúng tôi thấy một lượng nhỏ (200 g/ngày) rơm xử lý ammoniac đã
làm tăng lượng ăn vào của rơm không xử lý. Hoạt lực CMC cũng tăng, “c” trong phương trình diễn giải sự phân giải (p) (p=a+b (1-e-ct)) cũng tăng, có thể thấy tỷ lệ tiêu hoá đã tăng. Như vậy, có thể sử lý ammoniac một phần nhỏ phụ phẩm trồng trọt và cho gia súc ăn thức ăn đó vào buổi tối sẽ cải thiện sử dụng phụ phẩm trồng trọt ăn vào ban ngày khi chăn thả.
Một thí nghiệm thú vị khác chúng tôi thực hiện ở Sri Lanka (14) trong đó lá cây họ đậu Glyricidia masculata được sử dụng để bổ xung cho rơm lúa không xử lý. Trong thí nghiệm này chúng tôi có hai ảnh hưởng. Glyricidia có nhiều nitơ và có thể cung cấp nitơ cũng như xơ dễ lên men cho rơm không xử lý. Nhằm tách riêng hai ảnh hưởng, một nhóm cừu được nuôi với rơm lúa được phun dung dịch urê (2g urê/100g rơm) và nhóm kia cho ăn rơm không xử lý. (Bảng 2.12).
Khía cạnh thú vị nhất của đáp ứng trong thí nghiệm này là trong khi Glyricidia làm tăng lượng rơm ăn vào đáng kể khi không có urê bổ xung, Glyricidia cũng làm tăng lượng rơm ăn vào khi urê được bổ xung thêm vào. Kết quả này cho thấy Glyricidia có 2 ảnh hưởng, đóng góp cả nitơ và xơ dễ lên men cho khẩu phần không có urê, nhưng chỉ đóng góp xơ dễ lên men cho khẩu phần xử lý urê vì NH3 đã có quá nhiều.

38
Bảng 2.12. Ảnh hưởng của bổ sung lá cây Glyricidia có/hoặc không có urê đến l-ượng thức ăn ăn vào, tỷ lệ tiêu hoá của rơm lúa và nồng độ NH3 dạ cỏ.
Urê (%
DM rơm) Glyricidia (% khẩu
phần)
Rơm ăn vào
(g/ngày)
Tổng ăn vào
(g/ngày)
Tỷ lệ tiêu hoá chất
hữu cơ (%)
NH3 dạ cỏ (mg/100
ml) 0 0 574 574 41,2 43 0 5 673 708 46,7 103 0 15 848 998 50,4 185 0 30 721 1030 49,8 202 2 0 690 690 46,3 162 2 5 727 765 46,3 136 2 15 880 1035 50,7 183 2 30 715 1021 51,0 221
Sử dụng thức ăn bổ xung nhằm tác động tích cực đến tiêu hoá và lượng
thức ăn ăn vào của các thức ăn thô chất lượng thấp trở nên quan trọng hơn việc tránh tương tác bất lợi. Bổ xung có thể áp dụng cho số lượng gia súc lớn hơn vì phần lớn gia súc trên thế giới được nuôi bằng thức ăn thô, phần lớn thức ăn thô này có chất lượng thấp và tình hình này sẽ vẫn còn như vậy trong tương lai thấy trước được. Thức ăn bổ xung không có nhiều và thường đắt ở thế giới thứ ba và một điều quan trọng là nếu sẵn có, chúng nên được sử dụng theo cách mà chúng không làm giảm việc sử dụng thức ăn thô. Đó là điều tốt nhất, dĩ nhiên, nếu thức ăn bổ xung, ngoài việc được sử dụng như là nguồn năng lượng, thức ăn bổ xung cũng có thể cũng nâng cao tỷ lệ tiêu hoá và lượng thức ăn ăn vào của khẩu phần cơ sở là thức ăn thô. Chiến lược bổ xung, do đó, trở nên vô cùng quan trọng nhưng phải nhớ rằng các nghiên cứu môi trường dạ cỏ chỉ có thể được thực hiện ở điều kiện địa phương, sử dụng các nguồn thức ăn địa phương. Kỹ thuật nylon bag, tuy nhiên, là công cụ hữu hiệu, đơn giản, năng động cho các nghiên cứu về môi trường dạ cỏ, kỹ thuật này chỉ cần gia súc mổ lỗ dò và thiết bị đơn giản. Kỹ thuật không phụ thuộc vào việc kiểm soát chính xác nguồn cung cấp điện như trong các nghiên in vitro, và có thể rất quan trọng, đặc biệt ở các nước đang phát triển, nơi thức ăn thô là nguồn thức ăn thô xanh chính cho gia súc. Thức ăn bổ sung có thể đánh giá qua tương tác tích cực của chúng với thức ăn của khẩu phần cơ sở cũng như qua tỷ lệ tiêu hoá của chúng và số năng lượng chúng đóng góp. Tài liệu tham khảo cho phần 2.1
1. Ørskov, E.R. and Mehrez, A.Z. 1975. The effect of feeding whole or pelleted barley on rate of digestion of dried grass in the rumen of sheep. Proc. Nutr. Soc. 34, 64A.
2. Ørskov, E.R and Fraser, C. 1975. The effect of processing of barley based supplements on rumen pH, rate of digestion and voluntary intake in sheep. Br. J. Nutr. 34, 493-500.

39
3. Chimwano, A.M., Ørskov, E.R and Stewart, C.S. 1976. Effect of dietary proportions of roughage and concentrate on rate of digestion of dried grass and cellulose in the rumen of sheep. Proc. Nutr. Soc. 35, 101A.
4. Mould, F.L. and Ørskov, E.R. 1984. Manipulation of rumen fluid pH and its influence on cellulolysis in sacco, dry matter degradation and the rumen microflora of sheep offered either hay or concentrate. Anim. Fd. Sci. Tech. 10, 1-14.
5. Mould, F.L., Ørskov, E.R. and Mann, S.O. 1984. Associative effects of mixed feeds. I. Effects of type and level of supplementation and the influence of the rumen fluid pH on cellulolysis in vivo and dry matter and digestion on various roughages. Anim. Fd. Sci. Tech. 10, 15-30.
6. Mould, F.L., Ørskov, E.R. and Gauld, Shirley A. 1984. Associative effects of mixed feeds. II. The effect of dietary addition of bicarbonate salts on the voluntary intake and digestibility of diets containing various proportions of hay and barley. Anim. Fd. Sci. Tech. 10, 31-47.
7. Istasse, L., Smart, R.I. and Ørskov, E.R. 1986. Comparison between two methods of feeding concentrate to sheep given a diet high or low in concentrate with or without buffering substances. Anim. Fd. Sci. Tech. 16, 37-49.
8. Istasse, L., Reid, G.W., Tait, C.A.G. and Ørskov, E.R. 1986. Concentrates
for dairy cows: effects of feeding method, proportion in diet and type. Anim. Feed Sci. Technol. 15, 167-182.
9. Fahmy, S.T.M., Lee, N.H. and Ørskov, E.R. 1984. Digestion and utilization of straw. II. Effect of different supplements on the digestion of ammonia-heep. Tech. Bull. No. 23, IAFMM, Potters Bar, Herts.
10. Silva, Ayona T. and Ørskov, E.R. 1988. Fibre degradation in the rumen of animals receiving hay, untreated or ammonia-treated straw. Anim. Fd. Sci. Technol. 19, 277-287.
11. Silva, Ayona T., Greenhalgh, J.F.D. and Ørskov, E.R. 1989. Influence of ammonia treatment and supplementation on the intake, digestibility and weight gain of sheep and cattle on barley straw diets. Anim. Prod. 48, 99-108.
12. Silva, Ayona T., Wallace, R.J. and Ørskov, E.R. 1987. Use of particle-bound microbial enzyme activity to predict the rate and extend of fibre degradation in the rumen. Br. J. nutr. 57, 407-415.
13. Manyuchi, B., Ørskov, E.R. and Kay, R.N.B. 1992. Effects of feeding small amounts of ammonia treated straw on degradation rate and intake of untreated straw. In: Anim. Fd. Sci. & Tech. 38, 293-304.
14. Pathirana, K.K. and Ørskov, E.R. 1995. Effect of supplementing rice straw with urea and glyricidia forage on intake and digestibility by sheep. Livestock Research for Rural Development, 7, No. 2, 40-43.

40
Hướng nghiên cứu 2. Sử dụng phương pháp nylon bag cho các nghiên cứu môi trường dạ cỏ và đánh giá thức ăn thô Hướng nghiên cứu 2. 2. Cải thiện chất lượng phụ phẩm trồng trọt
Vào năm 1980, chúng tôi bắt đầu rất quan tâm đến việc các nước khác nhau sử dụng rơm làm thức ăn cho gia súc nhai lại như thế nào. Ví dụ ở Bangladesh, chúng tôi thấy rơm lúa chiếm 80 đến 90% thức ăn cho bò, trong khi cùng thời điểm đó ở Anh phần lớn rơm bị đốt bỏ, do đó tôi cảm thấy đây là thời gian rất thích hợp cho chúng tôi nghiên cứu rơm một cách nghiêm túc và nói một cách ẩn dụ, đánh thức tiềm năng của rơm. Thực tế tôi đã nghiên cứu và một vài năm sau ngừơi ta gọi đùa tôi là ngài rơm! Một điều khác đã đưa chúng tôi đến các nghiên cứu này là do hợp tác của Anh với các nước đang phát triển tăng lên, chúng tôi có những sinh viên và thực tập sinh quan tâm đến thức ăn thô chất lượng thấp.
Chính là kỹ thuật nylon bag, với kỹ thuật này chúng tôi có thể xác định chất lượng thức ăn một cách nhanh chóng và đáng tin cậy, một lần nữa cho chúng tôi vai trò hàng đầu trong nghiên cứu. Để cải thiện chất lượng rơm, chúng tôi nâng cao chất lượng bằng các biện pháp xử lý, chọn lọc giống và phân chia phụ phẩm riêng ra. Thỉnh thoảng các chủ đề nghiên cứu chồng chéo lên nhau, tôi thấy rằng thảo luận chúng một cách riêng rẽ là thuận tiện nhất.
Vì rất nhiều lý do, chúng tôi thấy việc xử lý rơm với dung dịch NaOH là không thực tế, bởi vì nếu dùng quá nhiều dung dịch NaOH sẽ gây ô nhiễm môi trường, và gây thiếu nitơ trong rơm. Sử dụng ammonia dường như hấp dẫn hơn vì một lượng ammonia đủ nhiều bám vào rơm sẽ khắc phục sự thiếu nitơ trong rơm, sự thiếu nitơ là kết quả của kiềm hóa đã làm cho nhiều carbonhydrate của rơm có thể lên men bởi vi sinh vật dạ cỏ. Cũng đã có kinh nghiệm khích lệ từ Na Uy và Đan Mạch về sử dụng phương pháp ammonia trong thực tế.
Thí nghiệm đầu tiên được thực hiện với bò cái tơ đã cho một vài kết quả thú vị khi sử dụng anhydrous ammonia khan ở mức 30 gNH3/kg rơm (1) (Bảng 2.13). Bảng 2.13. Ảnh hưởng của rơm lúa mạch bổ sung 1% urê hoặc rơm lúa mạch xử
lý ammonia đến tỷ lệ tiêu hoá, lượng thức ăn ăn vào và tăng trọng ở bò cái tơ. Xử lý Tỷ lệ tiêu hoá
vật chất khô (%)
Chất khô ăn vào (kg/ngày)
Chất khô tiêu hoá ăn vào (kg/ngày)
Tăng trọng (g/ngày)
Urê(1%) 49,9 3,87 1,93 - 447 Ammonia 58,8 5,88 3,45 324 SE 1,8 0,31 - Tăng lên (%) 18 52 79

41
Ở đây tóm tắt lại ảnh hưởng quan trọng của việc nâng cao chất lượng bằng
các biện pháp xử lý là: tăng đáng kể lượng rơm ăn vào (52%), chất khô ăn vào tiêu hóa được (79%), và tăng trọng. Kết quả không hoàn toàn đáng ngạc nhiên; chúng tôi một cách đơn giản đã thử nghiệm công nghệ đã biết. Tôi cảm thấy rằng nông dân ở Anh sẽ có lợi nhưng ở đây có một rào cản. Cục khuyến nông Anh cho rằng công nghệ quá nguy hiểm cho nông dân Anh! Tôi cảm thấy thất vọng và thấy cần trình diễn cho nông dân! Tôi mua một cái thùng nhỏ từ Calor gas và đổ đầy anhydrous ammonia, đặt sau xe kéo của tôi và thực hiện một thí nghiệm nhỏ về xử lý ammonia với ba người bạn nông dân của tôi, ông J. Thomson, ông S. Catto và ông C. Esson. Sau khi nuôi bò với rơm xử lý ammonia, BBC được mời đến phỏng vấn họ. Ông Thomson đặc biệt ấn tượng với thí nghiệm. Ông ta chỉ phàn nàn rằng bò ăn rơm xử lý quá nhanh. Thực tế ông ta đã nói về điều này trong các cuộc họp của nông dân. Sau thí nghiệm nhỏ này, các trường nông nghiệp ở Scoland mua các thùng nhỏ cho các thí nghiệm tiếp theo và ICI đã sẵn sàng cho việc áp dụng công nghệ ở quy mô lớn nhưng họ muốn dùng ammonia lỏng hơn là anhydrrous ammonia. Điều này có nghĩa là không cần tới những thùng điều áp nữa. Dùng ammonia lỏng được thử nghiệm và trình bày trong luận văn thạc sĩ của Stella Holland (2). Kết qủa tương tự như kết quả chúng tôi đã đạt được với anhydrous ammonia khô về lượng thức ăn ăn vào và tỷ lệ tiêu hóa.
Sau đó việc áp dụng đã được tiến hành trên diện rộng nhưng vẫn còn một số vấn đề. Tôi đã phát biểu tại cuộc hội thảo của Hội Nông nghiệp Hoàng gia năm 1982 về “Rơm xử lý, một thức ăn kinh tế” (3) và tranh luận rằng rơm xử lý có thể được xem ngang hàng với thức ăn ủ chua hay cỏ khô, và có thể sử dụng rộng rãi. Sau đó người ta phỏng vấn tôi ở Highland Show bởi vì Cục Khuyến nông đã đưa ra các tính toán chi phí, họ tranh luận rằng trừ phi tỷ lệ tiêu hoá tăng 12%, ví dụ từ 48 lên 60%, thấp hơn mức đó xử lý sẽ không mang lại hiệu quả kinh tế; và trong phần lớn trường hợp tỷ lệ tiêu hoá không thể tăng tới 12%. Cùng với ông McDonald, nhà thống kê của chúng tôi; chúng tôi dã tính rằng nếu lượng ăn vào tăng thêm 50%, riêng nó đã tạo nên hiệu quả kinh tế ngay cả khi không có sự thay đổi về nào về tỷ lệ tiêu hoá! Những tranh luận như vậy không tìm thấy trong các tài liệu của Cục Khuyến nông, và chúng tôi đã có những quan điểm bất đồng. Một bản kiến nghị đã được gửi đến giám đốc của Viện nghiên cứu Rowett. Tôi được cho phép có một cuộc nói chuyện nhẹ nhàng, tôi không có nhiệm vụ nói chuyện với nông dân, việc áp dụng công nghệ là công việc của khuyến nông. Tuy nhiên nông dân có vẻ hiểu các luận cứ của tôi và công nghệ tìm thấy vị trí của nó trong nền nông nghiệp Anh!
Phạm vi mà axit có thể sử dụng để tăng tỷ lệ tiêu hoá cũng được thử nghiệm (5). Tỷ lệ phân giải của rơm không xử lý đã tăng từ 42 lên 67% ở 48 giờ và tỷ lệ phân giải của rơm xử lý ammonia đã tăng từ 61 lên 69% ở 48 giờ khi tăng lượng H2SO2 từ 0 đến 80 g/kg rơm.
Năm 1985, tiến sĩ A.K.Tuah từ Ghana và tiến sĩ E.Lufadeja từ Nigeria nhận một nhiệm vụ nặng nề là xử lý ammonia rơm của 19 loại lúa mạch, 14 loại lúa mì,

42
11 loại yến mạch và 1 loại triticale (lai giữa lúa mỳ và lúa mạch đen) để nghiên cứu biến động của các ảnh hưởng có lợi giữa các loài và giữa các giống (287). Về tổng thể, hình như là tỷ lệ phân giải của rơm không xử lý càng thấp thì đáp ứng với xử lý ammonia càng cao; nhưng rơm chất lượng tốt hơn đáp ứng tốt hơn với xử lý ammonia ở mức thấp hơn. Tiến sĩ M.Ramazin từ Italia ở cùng một thời điểm đó cho thấy cọng rơm bị ảnh hưởng bởi xử lý ammonia nhiều hơn lá (7).
Trong khoảng 6 năm, đàn bò sữa ở Viện nghiên cứu Rowett được cho ăn thức ăn mùa đông có 50% rơm lúa mạch xử lý ammonia mà không sụt giảm sản lượng sữa. Chúng tôi cũng thực hiện một thí nghiệm sử dụng 2 loại rơm lúa mạch khác nhau, chất lượng thấp (Gerbil) và chất lượng cao (Corgi), không xử lý (U) và xử lý (A), được trộn vào khẩu phần hỗn hợp hoàn chỉnh (TMR) với tỷ lệ 50% trong khẩu phần và cho ăn tự do (8) (Bảng 2.14). Bảng 2.14. Ảnh hưởng của việc cho ăn TMR có rơm lúa mạch Gerbil hoặc Corgi
xử lý và không xử lý đến năng suất bò sữa. 10 tuần sau khi đẻ.
Loại rơm Xử lý DM ăn vào (kg/ngày)
Tỷ lệ tiêu hoá (%)
Năng suất sữa
(kg/ngày)
Tỷ lệ phân giải của
rơm ở 48 giờ
Gerbil U 13,1 61,5 21,3 52,8 Gerbil A 15,0 65,2 22,3 61,2 Corgi U 14,8 66,3 21,5 66,0 Corgi A 17,1 67,2 27,2 72,1 SE 0,5 0,9 1,2 1,0
Rõ ràng có ảnh hưởng của xử lý ammonia và hai loại rơm đáp ứng khác
nhau. Năng suất sữa 27 kg ở 10 tuần sau khi đẻ gần với tiềm năng sản xuất của bò. Một quan sát thú vị ở đây là nếu so sánh tỷ lệ tiêu hoá của rơm với tỷ lệ phân giải, sai khác lớn hơn xẩy ra ở khẩu phần xử lý ammonia. Kết quả này cho phép nghĩ rằng có thể tỷ lệ phân giải vật chất khô của phân sẽ lớn hơn ở bò ăn rơm sử lý ammonia. Kết quả đúng là như vậy, phân giải vật chất khô của phân vào khoảng 30% ở bò ăn khẩu phần không xử lý và hơn 40% ở bò ăn khẩu phần xử lý.
Rơm xử lý ammonia có thể dùng để vỗ béo bò ở mức độ nào cũng đã được nghiên cứu trong một thí nghiệm lớn. Rơm lúa mạch Corgi xử lý và không xử lý ammonia và rơm lúa mì xử lý ammonia được trộn với các tỷ lệ khác nhau cùng với hạt lúa mạch và được cho ăn tự do trong thí nghiệm này. Kết quả thí nghiệm đuợc trình bày trong Bảng 2.15 (9). Bảng 2.15. Ảnh hưởng của việc cho ăn các loại rơm khác nhau đến lượng thức ăn
ăn vào, tỷ lệ tiêu hoá và tăng trọng của bò thịt trong 80 ngày vỗ béo. Loại rơm Tỷ lệ Chất khô ăn Tỷ lệ tiêu hoá Tăng trọng

43
rơm vào (kg/ngày)
chất khô (g/kg)
(kg/ngày)
Lúa mạch không sử lý 35 9,37 700 1,17 Lúa mạch không sử lý 45 8,48 667 1,02 Lúa mạch không sử lý 55 6,87 643 0,70 Lúa mạch xử lý ammonia 35 9,50 708 1,32 Lúa mạch xử lý ammonia 45 10,08 693 1,27 Lúa mạch xử lý ammonia 55 8,59 669 1,02 Lúa mì xử lý ammonia 35 9,85 703 1,26 Lúa mì xử lý ammonia 45 9,39 679 1,20 Lúa mì xử lý ammonia 55 7,54 670 0,95 SE 0,50 49 0,10
Tiềm năng tăng trọng của bò khoảng 1,3 kg/ngày. Một điều thú vị là 45% rơm lúa mạch không xử lý trong khẩu phần cho
tăng trọng tương đương 55% rơm xử lý trong khẩu phần. Sự khác nhau giữa rơm xử lý và không xử lý tăng khi tỷ lệ rơm trong khẩu phần tăng. Dù thế nào đi nữa, bò có thể được vỗ béo một cách thỏa đáng với khẩu phần có trên 50% rơm xử lý. Sự khác nhau giữa rơm lúa mì và rơm lúa mạch xử lý ammonia một cách đơn giản là do khác nhau về tỷ lệ phân giải. Tỷ lệ phân giải của 3 loại rơm ở 48 giờ là 55,8 (rơm lúa mạch không xử lý); 64,6 (rơm lúa mạch xử lý ammonia) và 60,9% (rơm lúa mì xử lý ammonia).
Vào năm 1983, dường như với chúng tôi, việc tách hạt ngũ cốc từ rơm bằng máy thu hoạch liên hoàn và chỉ làm khô hạt, rồi trộn chúng với nhau một lần nữa khi cho gia súc ăn là không cần thiết và lãng phí. Chúng tôi vì thế đã thử nghiệm xem liệu chúng ta có thể thu hoạch lúa mạch khi vẫn còn tươi không. Lúa mạch cả cây được thu hoạch khi vật chất khô khoảng 45% ở thời điểm 10 ngày trước khi chín (10). Lúa mạch cả cây được xử lý hoặc không với anhydrous ammonia hoặc ammonia lỏng khi thu hoạch ở mức 30 g NH3/kg vật chất khô, hoặc trộn với urê ở mức 60 g/kg chất khô và ủ trong hố nhỏ. Mẫu không xử lý giảm chất lượng một cách nhanh chóng nhưng mẫu được xử lý với urê, anhydrous ammonia hoặc ammonia lỏng được bản quản tốt và được sử dụng để nuôi bò thí nghiệm. (Bảng 2.16).
Bảng 2.16. Ảnh hưởng của các dạng sử lý ammonia đến bảo quản và vật chất khô
ăn vào của rơm lúa mạch cả cây và tỷ lệ tiêu hoá ở bò tơ. Xử lý Chất khô ăn vào
(kgDM/ngày)
Tỷ lệ tiêu hoá DM Tinh bột
Urê 5,01 54,2 78,0 Anhydrous NH3 2,79 58,8 92,8 NH3 lỏng 3,11 56,9 88,0

44
SE 0,21 1,3 1,5
Lượng vật chất khô ăn vào của khẩu phần rơm lúa mạch cả cây xử lý ammonia rất đáng thất vọng do ammonia phân bố phân bố không đều với rơm tươi. Urê hứa hẹn nhất và được phân bố đều khi trộn với lúa mạch cả cây trong máy trộn. Tất cả các mẫu xử lý ammonia đều được bảo quản tốt.
Cuối cùng chúng tôi xem xét phương pháp khác để cải thiện chất lượng rơm. Một phơng pháp hấp dẫn là sử dụng nước ôxy già (H2O2), nước ôxy già sẽ oxy hoá lignin và giải phóng sản phẩm cuối cùng là nước. Tuy nhiên nước oxy già chỉ có thể hoạt động trong môi trường kiềm và thường được trộn với NaOH. Chúng tôi thấy rằng có thể sử dụng phương pháp khác, phun dung dịch H2O2 vào rơm sau đó xử lý ammonia, ammonia sẽ cung cấp kiềm. Thí nghiệm này được tiến sĩ F. Adebowale từ Nigeria thực hiện (11) (Bảng 2.17).
Bảng 2.17. Ảnh hưởng của xử lý ammonia sau khi phun các lượng H2O2 khác
nhau đến phân giải tiềm năng và c của rơm lúa mì, thân ngô và lõi ngô.
Tỷ lệ phân giải ở 48 giờ (%)
(c)
Rơm lúa mì
Thân cây ngô
Lõi ngô
Rơm lúa mì
Thân cây ngô
Lõi ngô
Không xử lý 51 62 40 0,043 0,049 0,024Xử lý NH3 60 65 51 0,045 0,054 0,042Xử lý NH3+10gH2O2/kgDM 64 70 60 0,048 0,054 0,047Xử lý NH3+50gH2O2/kgDM 68 72 65 0,047 0,058 0,048Xử lý NH3+100gH2O2/kgDM 71 77 69 0,056 0,069 0,068
SE 1,1 0,6 0,7 0,002 0,0016 0,002
Có khả năng là sử dụng phương pháp này đã giải phóng nhiều hơn chất dễ lên men từ rơm. Thật không may H2O2 quá đắt nhưng về nguyên tắc phương pháp này có tiềm năng trong tương lai nếu giá giảm xuống.
Có thể cải thiện chất lượng rơm bằng cách cho enzyme tấn công rơm, và khi giáo sư Yoshio Nakashima từ Nhật tới tham gia với chúng tôi trong thời gian nghỉ phép, chúng tôi chuyển sự quan tâm sang việc sử dụng enzyme cellulase và xylanase.
Hỗn hợp enzyme cellulase chắc chắn giải phóng một số loại đường từ rơm và rơm đã xử lý với NaOH và NaOH + H2O2. Một ví dụ được trình bày trong Bảng 2.18 (12). Khi rơm ủ với enzyme trong phòng thí nghiệm, các đường được lên men và giảm pH. Tuy nhiên, hầu như không hoặc có ít ảnh hưởng của enzyme đến tỷ lệ phân giải (12).

45
Bảng 2.18. Ảnh hưởng của xử lý enzyme cellulase đến lá và thân của rơm lúa
mạch đã được xử lý NaOH hoặc NaOH + H2O2 từ trước.
pH Tỷ lệ nước (%)
Tỷ lệ phân giải ở 48 giờ
(%) Lá Thân Lá Thân Lá Thân
Không xử lý 0 5,00 5,60 18,5 13,6 68,0 29,9 Không xử lý 5 3,90 4,70 29,0 17,4 67,7 29,3 NaOH 0 6,75 7,25 18,1 11,7 73,5 28,3 NaOH 5 4,50 5,15 37,6 17,9 73,3 33,2 NaOH+H2O2 0 7,40 7,70 16,5 11,3 73,9 38,0 NaOH+H2O2 5 5,05 5,30 38,2 19,5 77,3 43,0
SE - - - - 1,5 1,0
Ở đây rõ rằng là nhiều đường bị hoà tan trong lá hơn là trong thân và về tổng thể tỷ lệ phân giải không thay đổi nhiều. Tuy nhiên việc này một phần do sự lên men của các chất hoà tan. Một thí nghiệm được thực hiện trong đó axit propionic được thêm vào nhằm hạn chế lên men và trong trường hợp này phân giải tiềm năng được cải thiện đôi chút (12).
Xử lý enzyme hình như hiệu quả hơn với rơm lúa, phân giải tiềm năng của rơm lúa tăng đáng kể. Phần đốt và thân đáp ứng nhiều hơn với xử lý enzyme so với lá nên thực tế ở rơm lúa, thân có tỷ lệ phân giải cao hơn lá. Một số kết quả được trình bày trong Bảng 2.19. Bảng 2.19. Ảnh hưởng của xử lý enzyme cellulase và ammonia lên các đặc điểm
phân giải của thân rơm lúa.
Sử lý Nước hoà tan (%)
Phân giải tiềm năng (%)
(c)
Không xử lý 30,0 63,8 0,0487 Xử lý ammonia 31,7 68,3 0,0424 Enzyme cellulase 36,4 68,1 0,0342
SE 0,0040
Phân giải tiềm năng tăng trong khi c giảm do xử lý bằng enzyme. Tuy nhiên, hỗn hợp enzyme cellulase được sử dụng không nguyên chất, trong thí nghiệm cuối cùng với enzyme, chúng tôi sử dụng một enzyme xylanase đặc biệt và cũng bổ sung thêm xylanase và cellulase vào cùng nhau (13) (Bảng 2.20). Bảng 2.20. Ảnh hưởng của ủ rơm lúa mạch với xylanase, cellulase hoặc hỗn hợp

46
của xylanase và cellulase trong 20 ngày. Xử lý Tỷ lệ nước hoà
tan (%) Phân giải 48 giờ
(%) Phân giải tiềm năng (a+b) (%)
Đối chứng 17,5 52,5 58,1 Xylanase 32,8 63,7 68,2 Cellulase 28,3 57,8 68,0 Xylanase+cellulase 33,6 62,7 74,5 SE - 1,5 1,6
Có thể thấy rằng xử lý enzyme tạo cơ hội thực sự để nâng cao chất lượng
rơm, nhưng giá của enzyme cần phải giảm để xử lý enzyme có hiệu quả kinh tế. Xử lý bằng enzyme có lợi ích nữa là thân thiện với môi trường vì không để lại các chất hoá học không mong muốn. Phương pháp dùng enzyme cho phép ủ chua rơm lúa vì lượng đường được giải phóng và bị lên men, đủ để tạo pH đủ thấp để bảo quản rơm
Chúng tôi cũng đã thử tìm hiểu ảnh hưởng của làm ớt rơm hoặc ủ rơm với cỏ ủ. Tuy nhiên như mong đợi, về tổng thể không có ảnh hưởng nào đến tỷ lệ phân giải của rơm khi rơm được ủ trong hố với cỏ ủ chua ở pH 4 trong 21 ngày (14).
Phương pháp cuối cùng chúng tôi nghiên cứu cải thiện chất lượng rơm là xử lý hơi nước ở mức nhẹ. Với phương pháp này, một phần hemicellulose được thủy phân, đặc biệt khi rơm được trộn với axit loãng trước khi xử lý hơi nước ở mức nhẹ. Xử lý hơi nước nghe có vẻ là cách tốn kém vì nó được thực hiện dưới áp suất ở nhiệt độ cao. Tuy nhiên một phần của rơm có thể dùng làm nhiên. Trong bất cứ trường hợp nào, chúng tôi cũng phải thực hiện nghiên cứu này vì đấy là luận án tiến sỹ của Fernando Castro đến từ Brazil. Kết quả của nghiên cứu này đầy hứa hẹn và vẫn đang tiếp tục, tôi cũng sẽ thảo luận một vài khía cạnh của nghiên cứu này trong phần nghiên cứu từng phần khác nhau của phụ phẩm sử dụng phương pháp mạnh để xử lý rơm. Một số kết quả được trình bày trong bảng 2.21 (15 và 16). Bảng 2.21. Ảnh hưởng của xử lý hơi nước rơm lúa mỳ đã xử lý với axit loãng (18g H2SO4/kgDM) ở các nhiệt độ khác nhau trong 120 phút đến tỷ lệ phân giải ở 24 và
96 giờ ủ.
Nhiệt độ (Oc) Tỷ lệ phân giải (%) 24 giờ 96 giờ
Nhiệt độ phòng 38,7 56,3 98 34,3 56,2 121 42,6 63,4 134 45,0 69,1 SE 2,0 1,8

47
Xử lý hơi nước cho phép tạo ra thức ăn chất lượng cao. Nếu đốt rơm để
cung cấp năng lượng cho quá trình xử lý, sẽ tốn khoảng 10% rơm cho việc này. Phương pháp dùng hơi nước có thể tiến hành ở phạm vi lớn mang tính công nghiệp với nồi hơi cố định như được dùng cho xử lý bã mía hoặc tinh chế phụ phẩm ngũ cốc trong tương lai. Cũng có thể sản xuất các nồi hơi di động, các trang trại nhỏ có thể sử dụng áp dụng được, sử dụng một phần của rơm hoặc rơm kém chất lượng để cung cấp năng lượng cho nồi hơi.
Có một lượng lớn năng lượng cất giữ trong phụ phẩm trồng trọt, năng lượng này có thể được giải phóng để cung cấp dinh dưỡng cho gia súc nhai lại. Phần lớn phụ phẩm trồng trọt có tỷ lệ tiêu hoá khoảng 40%. Về mặt lý thuyết, tỷ lệ này có thể được nâng lên khoảng 70% với những hiệu quả to lớn sau đó. Với việc sử dụng enzyme hợp lý, trong tương lai có thể biến đổi phụ phẩm trồng trọt thành thức ăn cho con người và tăng gần gấp đôi lượng lương thực sản xuất trên thế giới. Về chăn nuôi gia súc, hiện nay chỉ có phương pháp xử lý anhydrous ammonia hoặc ammonia lỏng là đã có ảnh hưởng ở các nước công nghiệp. Sử lý với urê là một phương pháp sản sinh ammonia rất quan trọng ở nhiều nước như Trung Quốc dùng cho xử lý rơm ở qui mô nhỏ. Có nhiều khả năng ứng dụng một vài phương pháp chúng tôi đã thử nghiệm. Chi phí và hoàn cảnh biến động lớn ở mỗi môi trường làm cho đôi khi phương pháp xử lý sẽ rất hiệu quả về kinh tế, nhưng thông thường thì không phải như vậy. Điều quan trọng nhất là phải cân nhắc hậu quả môi trường lâu dài của các phương pháp xử lý nâng cao chất lượng thức ăn. Sử dụng hơi nước, H2O2 và enzyme và ở một vài phạm vi nào đó dùng ammonia hấp dẫn hơn việc sử dụng kiềm khoáng như NaOH. Một điều quan trọng để cân nhắc, ngoài chi phí sử lý là có thừa rơm không, nếu thừa rơm thì phần lớn chi phí bỏ ra để xử lý rơm sẽ được bù lại do nhiều thức ăn thừa được sử dụng cho chăn nuôi. Nhiều nghiên cứu tìm kiếm và khám phá vẫn đang cần để tìm ra phương pháp giúp giải phóng nguồn năng lợng lớn bị giữ trong phụ phẩm trồng trọt và không được sử dụng cho gia súc và con ngời. Về tổng thể, xử lý enzyme cellulase không tăng tỷ lệ tiêu hoá nhưng tăng độ hoà tan vì một số cellulose bị hoà tan. Tăng độ hoà tan chủ yếu từ phần lá trừ rơm lúa tăng độ hoà tan lớn nhất là phần thân. Nếu axit propionic được thêm vào, men của phần hoà tan có thể tránh được và vì thế tỷ lệ phân giải tiềm năng tăng lên. Hiện tại giá enzyme đang giảm. Sự áp dụng hứa hẹn nhất của sử dụng enzyme sẽ có thể là lên men rơm ướt vì rơm lúa tươi có thể được ủ tránh được việc lãng phí một số lượng lớn rơm lúa tươi. Xử lý rơm với ammonia có thể vẫn cần khai thác thêm. Ở những nước công nghiệp sử dụng anhydrous ammonia và ammonia lỏng là hợp lý nhất. Ở các nước đang phát triển, đặc biệt là trong các trang trại nhỏ, nơi nhiệt độ cao, urê là tốt nhất cho xử lý ammonia các loại phụ phẩm nông nghiệp. Tài liệu tham khảo cho phần 2.2
1. Orskov, E.R., Tait, C.A.G. and Reid, G.W. 1981. Utilization of ammonia

48
treated or urea treated barley straws as the only feed for dairy heifers. Anim. Prod. 32, 388.
2. Orskov, E.R., Reid, G.W., Holland, S.M., Tait, C.A.G. and Lee, N.H. 1983. The feeding value for ruminants of straw and whole crop barley and oats treated with anhydrous or aqueous ammonia or urea. Anim. Fd. Sci. Tech. 8, 247-257.
3. Adebowale, E.A., Ørskov, E.R. and Hotten, P.M. 1989. Rumen degradation of straw. 8. Effect of alkaline hydrogen peroxide on degradation of straw using either sodium hydroxide or gaseous ammonia as source of alkali. Anim. Prod. 48, 553-559.
4. Fahmy, S.T.M. and Ørskov, E.R. 1984. Digestion and utilization of straw. I. Effect of different chemical treatments on degradability and digestibility of barley straw by sheep. Anim. Prod. 38, 69-74.
5. Fahmy, S.T.M., Lee, N.H. and Orskov, E.R. 1984. Digestion and utilization of straw. II. Effect of different supplements on the digestion of ammonia fed straw. Anim. Prod. 38, 74-84.
6. Tuah, A.K., Lufadeju, E., Orskov, E.R. and Blackett, G.A. 1986. Rumen degradation of straw. 1. Untreated and ammonia treated barley, oat and wheat straw varieties and triticale straw. Anim. Prod. 43, 261-269.
7. Ramanzin, M., Orskov, E.R. and Tuah, A.K. 1986. Rumen degradation of straw. 2. Botanical fractions of straw from two barley cultivars. Anim. Prod. 43, 271-278.
8. Orskov, E.R., Tait, C.A.G., Reid, G.W. and Flachowsky, G. 1988. Effect of straw quality and ammonia treatment on voluntary intake, milk yield and degradation characteristics of faecal fibre. Anim. Prod. 46, 1, 23-27.
9. Orskov, E.R., Reid, G.W. and Kay, M. 1991. Influence of straw quality and level of concentrate in a completely mixed diet on intake and growth rate in steers. Anim. Prod. 52, 461-464.
10. Orskov, E.R., Reid, G.W., Holland, S.M., Tait, C.A.G. and Lee, N.H. 1983. The feeding value for ruminants of straw and whole crop barley and oats treated with anhydrous or aqueous ammonia or urea. Anim. Fd. Sci. Tech. 8, 247-257.
11. Orskov, E.R., Reid, G.W., G.W., Holland, S.M., Tait, C.A.G. and Lee, N.H. 1983. The feeding value for ruminants of straw and whole crop barley and oats treated with anhydrous or aqueous ammonia or urea. Anim. Fd. Sci. tech. 8, 247-257.
12. Nakashima, Y. and ỉrskov, E.R. 1989. Rumen degradation of straw. 7. Effects of chemical pre-tretament and addition of propionic axít on degradation characteristics of botanical fractions of barley straw treated with a cellulase preparation. Anim. Prod. 48, 543-551.
13. Ørskov, E.R. and Smith, D.C. 1990. Enzyme treatment to improve nutritive value of straws. PIRA 2nd International Conference, Peterborough. PIRA Paper Board Division, Leatherhead, Surrey.

49
14. Bhargava, P.K., Ørskov, E.R. and Walli, T.K. 1989. Effect of soaking, ensilage and hydrogen peroxide treatment of barley straw on rumen degradability. Anim. Fd. Sci. Tech. 22, 295-303.
15. Castro, F.B., Hotten, P.M. and Ørskov, E.R. 1993. Effect of dilute-axít hydrolysis treatment on the physico-treatment features and bio-utilization of wheat straw. Anim. Fd. Sci. Tech. 42, 55-67.
16. Castro, F.B., Hotten, P.M. and Orskov, E.R. 1993. The potential of dilute-axit hydrolysis as a treatment of improving the nutritional quality of industrial lignocellulosic by-products. Anim. Fd. Sci. Tech. 42, 39-53.

50
Hướng nghiên cứu 2. Sử dụng phương pháp túi ni lông (nylon bag) cho các nghiên cứu môi trường dạ cỏ và đánh giá thức ăn thô Hướng nghiên cứu 2.3. Sự khác nhau về chất lượng rơm giữa các giống ngũ cốc
Quyết định đi vào nghiên cứu rơm một cách nghiêm túc được định hướng không chỉ bởi vì chúng tôi cảm thấy sử dụng tốt hơn nguồn thức ăn to lớn này rất quan trọng, mà còn bởi vì chúng tôi đã có phương pháp nylon bag tiến bộ, chúng tôi tin tưởng vào nó, và phương pháp này có thể xác định một cách nhanh chóng giá trị dinh dưỡng của rơm. Có một câu hỏi nhất định phải trả lời là liệu các giống có sự khác nhau? Chúng tôi hợp tác với bộ phận di truyền giống Trường cao đẳng Nông nghiệp Scotland và có, như đã đề cập ở đâu đó, 19 loại rơm lúa mạch khác nhau, 14 loại rơm lúa mỳ, 11 loại kiều mạch và 1 loại triticale (lai giữa lúa mỳ và lúa mạch đen) (1). Kết quả nhiên cứu rất khác thường. Ví dụ tỷ lệ phân giải 48 giờ của rơm lúa mạch mùa xuân biến động từ 38,5 đến 60,7; của rơm lúa mỳ từ 45,2 đến 50,5 và rơm kiều mạch từ 44,1 đến 51,6 %. Cái gì quan trọng tương đương hoặc quan trọng hơn? Không có mối liên hệ giữa năng suất hạt và chất lượng rơm. Biến động đặc biệt lớn ở rơm lúa mạch có nghĩa là sự khác nhau giữa một vài giống lúa mạch lớn hoặc lớn hơn sự khác nhau có được khi xử lý ammonia và việc xử lý là không cần thiết. Cũng không có mối liên hệ giữa chiều dài rơm và hàm lượng nitơ.
Cái gì tạo nên sự khác nhau đó? Có thể sự khác nhau này là do sự khác nhau giữa chúng về phần thực vật học của lá và thân. Ramazin từ Italia (2) đã nghiên cứu rơm của 2 giống rất khác biệt đại diện cho loại rơm chất lượng cao (Corgi) hoặc chất lượng thấp (Gerbel). Rơm được tách thành lá + bẹ lá, gióng, mấu và rơm vụn. Tỷ lệ các phần lá + bẹ lá, gióng, mấu và rơm vụn của rơm Corgi tương ứng là 49,9; 38,0; 0,55 và 6,5% và của rơm Gerbel là 40,4; 51,2; 5,9 và 2,5%. Bởi vì tỷ lệ phân giải của lá cao hơn tỷ lệ phân giải của cuống, sự khác nhau về thành phần thực vật học có thể giải thích một phần cho sự khác nhau nhưng ngay cả như vậy, trong sai khác về tỷ lệ phân giải lúc 48 giờ 13,2%, chỉ 2,5% là do sự khác nhau về thành phần thực vật, phần còn lại 10,7% là do sự khác nhau về tỷ lệ phân giải của các phần, chủ yếu là tỷ lệ phân giải của cuống.
Sự khác nhau tương tự cũng quan sát thấy ở rơm lúa cả cây Ấn Độ (3), Nhật Bản (4) và Indonesia (5). Walli thấy rằng tỷ lệ phân giải tiềm năng (a+b) của rơm của một giống là 62 và của một giống khác là 56. Sự khác nhau này ở rơm các giống Indonesia còn lớn hơn. Sau đó Bill Shand đã thu thập rất nhiều số liệu từ 6 giống kiều mạch và 11 giống rơm lúa mỳ (6). Có 2 thứ xuất hiện ở đây: tỷ lệ phân giải của rơm các giống kiều mạch thay đổi từ 36,7 đến 40,7; biến động ít hơn tỷ lệ phân giải của rơm các giống lúa mỳ 39,0 đến 48,0; và điều làm nông dân ngạc nhiên là giá trị dinh dưỡng của rơm lúa mỳ cao hơn của rơm kiều mạch. Tỷ lệ

51
phân giải trung bình lúc 48 giờ của phần lá rơm lúa mỳ và phần lá rơm kiều mạch tương ứng là 61,5 và 50,1.
Để tìm kết quả trong thực tế của phát hiện trên, một thí nghiệm (cũng đã được thảo luận ở trong hướng nghiên cứu 2.5) so sánh 4 giống rơm lúa mỳ sử dụng cho bò đực thiến (7) (Bảng 2.22).
Bảng 2.22. Ảnh hưởng của giống lúa mỳ đến sử dụng rơm của bò đực thiến cho ăn
1,5 kg thức ăn tinh/ngày. Giống Chất khô
ăn vào (kg/ngày)
Tỷ lệ tiêu hoá (%)
Năng lợng trao đổi (MJ/kg)
Tăng trọng (g/ngày)
Gerbil 3,43 40,9 7,23 106 Igri 3,56 41,2 7,16 126 Golden promise 4,43 45,2 7,35 198 Corgi 5,16 48,4 7,82 400 SE 0,18 1,2 - 40
Có thể thấy sự khác nhau xác định từ các thí nghiệm nylon bag cũng được
phản ánh trong tỷ lệ tiêu hoá in vivo, lượng thức ăn ăn vào và tốc độ tăng trọng từ 106 đến 400 g/ngày chỉ do sự khác nhau về giống rơm sử dụng. Sự khác nhau này có thứ tự như sự khác nhau do xử lý ammonia nhưng không tốn kém chi phí.
Một thí nghiệm được thực hiện trên bò sữa nhằm xem xem bò sữa có đáp ứng tương tự hay không. Rơm Gerbel và Corgi chiếm 50% trong khẩu phần hỗn hợp hoàn chỉnh (8) (Bảng 2.23).
Bảng 2.23. Ảnh hưởng của cho ăn rơm lúa mạch Corgi và Gerbil đến năng suất của bò cái Friesian trong 10 tuần vắt sữa.
Giống Tỷ lệ phân
giải lúc 48 giờ (%)
Tổng lượng ăn vào
(kg/ngày)
Năng suất sữa (kg/ngày)
Giảm khối lượng (kg)
Gerbil 52,2 13,1 21,3 -36 Corgi 66,8 14,8 21,5 -17 SE 0,5 1,1 6
Ở đây sự khác nhau được thể hiện ở lượng thức ăn ăn vào và giảm khối
lượng nhưng năng suất sữa là như nhau. Cuối cùng chúng tôi nghiên cứu tính ổn định của khác biệt giữa các giống
trên 9 giống lúa mạch mùa xuân và 10 giống lúa mỳ mùa đông trong 3 năm, và 12 giống lúa mạch mùa đông, 7 giống kiều mạch trong 2 năm. Biến động lớn nhất giữa các giống quan sát thấy ở lúa mạch mùa xuân. Ở đây tương quan trong các giống từ năm này qua năm khác phần lớn vượt quá r = 0,90; có sự biến động đáng

52
kể giữa các năm (9), nhưng hầu hết xếp hạng thứ tự về chất lượng là tương tự cho từng năm.
Chọn lọc cây trồng để nâng cao chất lượng phụ phẩm trồng trọt cần được các nhà chọn giống cây trồng thực hiện một cách nghiêm túc. Nâng cao chất lượng phụ phẩm trồng trọt có thể thực hiện được với điều kiện không làm ảnh hưởng đến năng suất và chất lượng hạt. Sự khác nhau về chất lượng phụ phẩm có thể lớn như sự khác nhau có được do xử lý ammonia, sự thay đổi này không tốn kém gì cả. Mặc dù có thể chọn lọc cây trồng để nâng cao chất lượng phụ phẩm trồng trọt, cho đến hiện nay các nhà tạo giống cây trồng trên thế giới ít quan tâm đến việc chọn lọc này.
Tài liệu tham khảo cho phần 2.3
1. Tuah, A.K., Lufadeju, E., Ørskov, E.R. and Blackett, G.A. 1986. Rumen degradation of straw. 1. Untreated and ammonia-treated barley, oat and wheat straw varieties and triticale straw. Anim. Prod. 43, 261-269.
2. Ramazin, M., Ørskov, E.R. and Tuah, A.K. 1986. Rumen degradation of straw. 2. Botanical fractions of straw from two barley cultivars. Anim. Prod. 43, 271-278.
3. Walli., T.K., Ørskov, E.R. and Tuah, A.K. 1988. Rumen degradation of straw. 3. Botanical fractions of two straw varieties and effects of ammonia treatment. Anim. Prod. 46, 347-352.
4. Nakashima, Y. and Ørskov, E.R. 1989. Rumen degradation of straw. 9. Effect of cellulase and ammonia treatment on different varieties of rice straw and their botanical fractions. Anim. Prod. 50, 309-317.
5. Soebarinoto, Hermanto, Ørskov, E.R. and van Bruchem, J. 1992. In sacco degradation characteristics as a predictor of digestibility and voluntary intake of rice straw. In: Livestock and Feed Development in the Tropics. (eds. Ibrahim, M.N.M., de Jong, R., van Bruchem, J., and Purnomo, H.,) pp. 233-238. Proc. International Seminar Brawaijaja University, Malang.
6. Shand, W.J., Ørskov, E.R. and Morrice, L.A.F. 1988. Rumen degradation of straw. 5. Botanical fractions and degradability of different varieties of oat and wheat straws. Anim. Prod. 47, 387-392.
7. Reid, G.W., Ørskov, E.R. and Kay, M. 1988. A note on the effect of variety, type of straw and ammonia treatment on digestibility and on growth rate in steers. Anim. Prod. 47, 157-160.
8. Ørskov, E.R., Tait, C.A.G., Reid, G.W. and Flacjowsky, G. 1988. Effect of straw quality and ammonnia treatment on voluntary intake, milk yield and degradation characteristics of faecal fibre. Anim. Prod. 46, 1, 23-27.
9. Ørskov, E.R., Shand, W.J., Tedesco, D. and Morrice, L.A.F. 1990. Rumen degradation of straw. 10. consistency of differences in nutritive value between varieties of cereal straws. Anim. Prod. 51, 155-162.

53
Hướng nghiên cứu 2. Sử dụng phương pháp nylon bag cho các nghiên
cứu môi trường dạ cỏ và đánh giá thức ăn thô Hướng nghiên cứu 2. 4. Tách phụ phẩm trồng trọt thành những phần nhỏ
Triết lý tách phụ phẩm trồng trọt thành những phần nhỏ không mới. Tôi đã được khuyến khích khi đọc cuốn sách do Finn Rexen và Lars Munch viết “Cây ngũ cốc dùng trong công nghiệp” (EEC xuất bản năm, 1984). Trong cuốn sách này, các tác giả đã thảo luận nhiều cách sử dụng hạt ngũ cốc và tách rơm thành những phần nhỏ. Thân tách ra từ rơm, có thành phần tương tự như gỗ còn phần lá tách ra từ rơm dễ tiêu hóa hơn và hữu ích cho gia súc, vì vậy thân có thể sử dụng trong công nghiệp làm giấy và ván cứng. Như đã đề cập từ trước, ở giai đoạn đầu trong nghiên cứu của chúng tôi, chúng tôi đã tách rơm thành các phần lá, thân, mấu và rơm vụn và xác định giá trị dinh dưỡng của chúng.
Một vài giá trị trung bình các phần thực vật của các loại rơm khác nhau được trình bày dưới đây (Bảng 2.24). Tuy nhiên ở đây, cần nhấn mạnh rằng có biến động rất lớn giữa các giống. Nói chung, thực vật cao cây cho tỷ lệ thân nhiều hơn và bởi vì thân và lá là hai phần chính của cây, chắc chắn có mối quan hệ nghịch giữa phần lá và thân. Bảng 2.24. Trung bình phần lá, cuống, mấu và rơm vụn trong các nghiên cứu khác
nhau (1, 2, 3).
Nguồn tham khảo
Loại rơm Cuống (%)
Tỷ lệ phân giải cuống ở 48 giờ
Lá (%)
Tỷ lệ phân giải lá ở 48
giờ
Rơm vụn (%)
Mấu (%)
2 Lúa mạch 44,6 45,8 45,1
73,4 4,5 5,7
1 Lúa mỳ 46,5 33,1 34,0
61,5 13,6 5,7
1 Kiều mạch 56,9 27,0 31,0
50,1 4,8 7,3
3 Lúa 20,2 53,9 65,6
53,3 6,2 8,0
Trừ rơm lúa, giá trị dinh dưỡng của lá nói chung cao hơn cuống, đôi khi
khoảng 70% ở lúa mạch. Mặt khác, lá rơm kiều mạch không dễ tiêu hoá như lá rơm lúa mạch và lúa mỳ.
Nguyên tắc chung đã được phát hiện trong nghiên cứu của chúng tôi (2) là phần cuống đáp ứng với xử lý ammonia nhiều hơn phần lá, ngược lại phần lá đáp ứng với xử lý enzyme tốt hơn nhiều do hoà tan một lượng lớn những phần không

54
hòa tan. Xử lý enzyme có thể đủ để hòa tan để phần lá rơm thích hợp cho các da súc dạ dầy đơn.
Kết quả của các nghiên cứu trên dẫn chúng tôi đến một dự án hợp tác về phân đoạn cây trồng trong công nghiệp sản xuất giấy, ván cứng; và chế tạo máy để tách lá và cuống sử dụng kết hợp chà sát và sàng sảy cơ học. Máy hoạt động không tốt trong việc tách lá từ cuống nhưng phần lá tiêu hoá tốt hơn nhiều so với phần cuống. Một khía cạnh quan trọng khác ở đây liên quan đến sự phát triển tương lai của các loại máy tinh chế ngũ cốc thay thế cho máy thu hoạch kết hợp là giá trị dinh dưỡng của lá và thân không tương quan chặt, do đó thức ăn gia súc chất lượng cao có thể được chọn lọc cùng thời gian với phần cây trồng có giá trị công nghiệp cao bởi vì để sử dụng trong công nghiệp, cuống không cần phải có giá trị dinh dưỡng cao (3). Cũng có thể tách một phần của rơm làm thức ăn chăn nuôi và phần còn lại làm chất độn chuồng không bụi vì hiện phần lớn rơm ở một số vùng đang được dùng làm chất độn chuồng.
Cuối cùng chúng tôi quan tâm nhiều hơn đến xử lý hơi nước sau khi chúng tôi thiết lập một thiết bị xử lý hơi nước nhỏ trong phòng thí nghiệm. Công việc này phần lớn do Fernando Castro từ Brazil và Mohammed Zahidifar từ Iran thực hiện bởi và phần lớn các nghiên cứu này sẽ được xuất bản. Về cơ bản, sử lý hơi nước và nén từ từ, có thể tách rơm thành những thành phần cơ bản như lignin, cellulose và hemicellulose. Hemicellulose có thể được tách ra bằng nước và được sử dụng như siro cho gia súc ăn hoặc được sử dụng như chất nền để lên men riệu và các chất khác. Lignin và cellulose có thể tách ra bằng kiềm nhẹ. Cellulose tinh chất sau đó có thể sử dụng cho sản xuất giấy ít hoặc không gây ô nhiễm môi trường. Lignin nguyên chất có thể là giá trị nhất. Lignin nguyên chất có thể sử dụng như nguyên liệu thô, keo dán, nhựa sinh học phân hủy được, sơn hoặc vật liệu làm đường xá về mặt dunh dưỡng lignin nguyên chất giúp urê phân giả chậm, bảo vệ protein khỏi bị phân giải ở dạ cỏ (4).
Có rất nhiều lựa chọn tiềm năng, trong đó chúng ta có thể sử dụng những thứ bỏ đi như phụ phẩm trồng trọt làm nguyên liệu thô cho công nghiệp và thức ăn chất lượng cao cho gia súc, tôi đã phát biểu như vậy năm 1991 trong một cuộc họp về Hoá học, Nông nghiệp và Môi trường do Hội hoá học Hoàng gia tổ chức (5). Tôi đã kết luận, tôi sẽ ủng hộ kết luận này với sự nhấn mạnh hơn vào thời điểm hiện nay.
“Lựa chọn sử dụng phụ phẩm nông nghiệp là sự thúc đẩy kinh tế to lớn và là sự ủng hộ lớn lao cho các nghiên cứu cần thiết và cho sự phát triển. Chúng ta hãy ủng hộ, giáo dục, thể chế hóa và khuyến khích sử dụng đầy đủ nhất các nguồn tài nguyên có thể phục hồi vì lợi ích của môi trường toàn cầu nói chung và vì sự quan tâm lâu dài của mỗi quốc gia thông qua thúc đẩy kinh tế và các biện pháp khác. Làm như vậy chúng ta sẽ biến rác thải hôm nay thành tài nguyên của ngày mai”. Tài liệu tham khảo cho phần 2.4
1. Shand, W.J., Ørskov, E.R. and Morrice, L.A.F. 1988. Rumen degradation of

55
straw. 5. Botanical fractions and degradability of different varieties of oat and wheat straws. Anim. Prod. 47, 387-392.
2. Ramazin, M., Ørskov, E.R. and Tuah, A.K. 1986. Rumen degradation of straw. 2. Botanical fractions of straw from two barley cultivars. Anim. Prod. 43, 271-278.
3. Walli., T.K., Ørskov, E.R. and Tuah, A.K. 1988. Rumen degradation of straw. 3. Botanical fractions of two straw varieties and effects of ammonia treatment. Anim. Prod. 46, 347-352.
4. Zahidifar, M., Castro, F.B. and Ørskov, E.R. (2001). Effect of hydrolytic lignin on formation of protein lignin complexes and protein degradation by rumen microbes. Anim. Fd. Sci. Tech. (in press).
5. Ørskov, E.R. 1991. Potential for using agricultural waste as a valuable resource. In: Chemistry, Agricaulture and the Environment (ed. Richardson, M.L.) pp. 31-45. The Royal Society of Chemistry, Cambridge.

56
Hướng nghiên cứu 2. Sử dụng phương pháp túi ni lông (nylon bag) cho các nghiên cứu môi trường dạ cỏ và đánh giá thức ăn thô Hướng nghiên cứu 2. 5. Ước tính lượng rơm ăn vào sử dụng phương pháp nylon bag
Tôi thực ra không thiết lập thí nghiệm đầu tiên để dự đoán lượng ăn vào. Tuy nhiên, bởi vì chúng tôi đã thực hiện một thí nghiệm với 5 giống lúa có tỷ lệ phân giải khác nhau ở 48 giờ, hoặc xử lý ammonia hoặc không, chúng tôi quyết định thực hiện một thí nghiệm lớn với bò đực thiến nhằm xem xem có thể ước đoán lượng thức ăn ăn vào đến đâu. Chúng tôi cũng đã xác định thành phần hoá học của rơm và năng lượng trao đổi của nó sử dụng buồng hô hấp (Respiration chamber). Khi Charlie Tait đưa cho tôi kết quả về thí nghiệm nylon bag, các kết quả rất hấp dẫn và thực sự chỉ sau đó tôi mới nhận ra rằng 3 thông số chính từ phương trình p = a+b(1-e-ct) phản ánh các đặc điểm quan trọng của rơm nếu chúng ta giả thiết rằng có hạn chế mặt vật lý đến lượng thức ăn ăn vào. Các đặc điểm đó là: phần hoà tan (a), phần không hoà tan được nhưng có thể lên men (b) và tốc độ tiêu hoá của phần không hoà tan (c). Kết quả thích đáng nhất được trình bày trong bảng 2.25 (1).
Tương quan giữa hàm lượng năng lượng trao đổi của thức ăn với vật chất khô ăn vào chỉ là r = 0,70 nhưng hồi qui nhiều chiều của a, b và c mạnh và có độ chính xác cao: với lượng thức ăn ăn vào r = 0,88; với vật chất khô tiêu hóa ăn vào r = 0,95 và với tăng trọng r = 0,95 (2).
Chúng tôi sớm nhận ra rằng do phải dừng xảy ra khi phần xơ không hoà tan của thức ăn thô lên men, giá trị ngoại suy cho phần chất hoà tan thường âm nhiều; điều này dĩ nhiên không nói lên điều gì cả vì tỷ lệ hoà tan không thể âm. Sau đó chúng tôi nhận ra rằng có ít nhất một đặc điểm của thưc ăn thô có thể được xác định trong phòng thí nghiệm: tỷ lệ hoà tan trong nước, đặc điểm này chúng tôi gọi A, giá trì này không giống như (a) trong phương trình trên. Do đó B = (a+b) - A.
A, phần hòa tan trong nước B, (a+b) - a, phần không hoà tan nhưng có thể lên men C, hằng số độ của b
Bảng 2.25. Ảnh hưởng của loại và giống rơm không xử lý (U) và xử lý ammonia
(A) đến năng lượng trao đổi (ME), đặc điểm phân giải của chúng, và đến tăng trọng, lượng vật chất khô ăn vào, tỷ lệ tiêu hoá vật chất khô (DDM) ở bò đực thiến. Loại MJ
(ME/kg DM)
a (%)
b (%)
c (%)
Tăng trọng
(g/ngày)
Ăn vào (kg/ngày)
DDM
(%) Lúa mạch Gerbil U 7,23 6,0 32,9 0,0337 106 3,43 40,9

57
mùa đông
A 8,53 7,9 54,4 0,0258 359 4,70 48,7
Lúa mạch Igri
U 7,16 5,1 38,2 0,0391 126 3,56 41,2 A 7,90 7,9 45,2 0,0351 332 4,82 45,5
Lúa mạch Corgi mùa xuân
A 9,50 3,4 48,7 0,0483 400 5,16 48,4 U 7,35 6,4 60,4 0,0451 608 5,86 59,6
Golden A 8,40 7,5 48,0 0,0303 198 4,43 45,2 Promise U 5,98 9,3 52,1 0,0376 602 4,93 50,4 Norman mùa đông A 8,37 7,7 40,9 0,0345 273 4,57 34,3 Lúa mỳ 5,98 9,0 51,9 0,0364 516 5,81 48,4 SE 40 0,17 1,4
U; không xử lý; A: xử lý ammonia; a: vật chất khô hoà tan; b: vật chất khô không hoà tan có khả năng phân giải; c: hằng số tốc độ.
Các nghiên cứu này được 2 sinh viên của chúng tôi, A. Kinbon ở Nigeria (3) và Martin Shem ở Tazania (4) tiếp tục tiến hành ở nước ngoài. Kinbon thí nghiệm trên cừu trong khi Martin thí nghiệm với bò đực thiến ở vùng núi Kilimanjaro trên 16 loại thức ăn thô khác nhau. Khazal cũng làm việc ngoài phòng thí nghiệm nhưng với các đồng nghiệp từ Bồ Đào Nha sử dụng cừu và chủ yếu thức ăn thô họ đậu (5). Kết quả từ tất cả 4 phòng thí nghiệm được trình bày trong Bảng 2.26 dưới đây. Ở đây có thể thấy rõ rằng tất cả các thông số quan trọng được chẩn đoán một cách chính xác. Tuy nhiên trong thí nghiệm ở Tanzania, lượng ăn vào của thân cây chuối rất thấp và thấp hơn nhiều so với dự đoán do đó số liệu đã không được đưa đây. Thân cây chuối có 95% nước và xơ rất dai. Bảng 2.26. Tương quan giữa chất khô ăn vào (DM), DM tiêu hóa ăn vào, tỷ lệ tiêu hoá DM, tăng trọng và các đặc điểm phân giải A, B và C của rơm ở Scotland (1)
Nigeria (3) Tanzania (4) và Bồ Đào Nha (5).
Tham khảo DM ăn vào DDM ăn vào Tỷ lệ tiêu hoá DM
Tăng trọng
1 0,93 0,96 0,90 0,95 3 0,99 0,92 0,88 0,99 4 0,88 ND 0,95 ND 5 0,90 0,93 0,98 0,93
ND: không xác định
Gần đây, các nghiên cứu ở Bồ Đào Nha, Ethiopia và Thổ Nhĩ Kỳ cho kết quả tương tự như kết quả trên.
Như vậy là từ một quan điểm nghiên cứu, chúng tôi đã có thể phân loại thức ăn thô một cách đơn giản dưới dạng các giá trị A, B và C. Tuy nhiên nông dân chỉ cần một giá trị, giá trị ấy cho biết gia súc sẽ ăn cái gì? Chúng tôi đã nỗ lực để đơn giản hoá yêu cầu ấy. Chúng ta hãy giả thiết là lượng chất khô (DM) ăn vào

58
= X1A + X2B + X3C. Nếu cơ sở dữ liệu đủ lớn, các hệ số sẽ cho một số chỉ dẫn về tầm quan trọng của mỗi giá trị trong chẩn đoán lượng thức ăn ăn vào. Nếu quả thật là như vậy, chúng ta có thể đơn giản hóa bằng cách chia tất cả với X1 do đó:
A = 1, tức là. ( ) A + ( ) B + ( ) C Trong nghiên cứu của mình, chúng tôi sau đó đã có được các giá trị: A=1,
B = 0,4 và C=200. Chúng ta giả sử rằng chúng ta có một loại rơm trong đó A=12, B = 40 và C = 0,03, như vậy giá trị chỉ số là (1 x 12) + (0,4 x 40) + (200 x 0,03) = 12 + 16 + 6 = 34. Vậy sau đó là cái gì? Bây giờ, chúng ta có thể vẽ đồ thị về các giá trị chỉ số với lượng thức ăn ăn vào và xác định các giá trị chỉ số cần thiết để có các mức ăn vào tương đương với nhu cầu duy trì. Trong nghiên cứu của chúng tôi, với giá trị chỉ số khoảng 30, bò đực thiến ăn đủ nhu cầu duy trì. Vậy tương quan giữa giá trị chỉ số và năng suất súc như thế nào? Kết quả từ 3 thí nghiệm được trình bày trong bảng 2.27 dưới đây.
Bảng 2.27. Tương quan giữa chất khô ăn vào (DM), tỷ lệ tiêu hoá chất khô, chất khô tiêu hoá ăn vào và tăng trọng tính được từ các giá trị chỉ số dựa trên các đặc
điểm phân giải. Tham khảo Chất khô ăn
vào Tỷ lệ tiêu hoá
chất khô Chất khô ăn vào tiêu hoá
được
Tăng trọng
1 0,95 0,74 0,94 0,96 4 0,90 0,75 0,88 0,81 5 0,90 0,95 0,92 0,89
Tương quan cao đương nhiên có thể xảy ra vì chỉ số giá trị dựa trên các đặc
điểm phân giải thức ăn trong dạ cỏ. Tuy nhiên, tiềm năng thức ăn của thức ăn thô phụ thuộc vào các giá trị hoặc A, B hoặc C. Cải thiện tốc độ tiêu hoá không cải thiện tỷ lệ tiêu hoá nhưng nâng cao lượng tiêu thụ thức ăn và do đó có thể nâng cao năng suất của gia súc. Cuống hoặc thức ăn thô họ đậu thường được ăn nhiều hơn rơm hòa thảo và thức ăn thô thân cỏ có tỷ lệ tiêu hóa tương đương do C lớn hơn.
Khả năng dự đoán tiềm năng thức ăn của thức ăn thô có thể mang lại kết quả to lớn. Kinh nghiệm chăn nuôi gia súc nhai lại ở Thế giới thứ ba đã rất rõ ràng là phần lớn gia súc nhai lại được nuôi bằng thức ăn thô và sẽ còn tiếp tục như vậy trong một tương lai nhìn thấy được.
Ở khía cạnh này tôi khá thất vọng với hệ thống đánh giá thức ăn hiện nay của chúng ta, dù nó dựa trên năng lượng tiêu hoá, năng lượng trao đổi hay năng lượng thuần, nó cũng không thực sự đúng với thức ăn thô. Một hệ thống đánh giá tốt sẽ cho nông dân biết được một chỉ dẫn về tỷ lệ thay đổi của thức ăn, nhờ vậy
1
1
X
X
1
2
X
X
1
3
X
X

59
khi mua hay bán thức ăn, người nông dân sẽ biết thức ăn nào rẻ nhất. Khi chúng tôi nói rằng năng lượng trao đổi từ rơm là rẻ nhất, nông dân sẽ mua rơm, nhưng gia súc chỉ có thể ăn được một lượng rơm nhất định vì sự hạn chế về vật lý của rơm. Các hệ thống đánh giá thức ăn tĩnh tại hiện nay hoạt động tốt cho thức ăn tinh tiêu hoá nhanh nhưng không hoạt động tốt cho thức ăn thô. Hậu quả là, sự xuất khẩu các hệ thống đánh giá thức ăn phương tây đến các nước nơi mà thức ăn thô là thức ăn duy nhất cho gia súc nhai lại chỉ co chút ít hoặc có giá trị gì, việc xuất khẩu này thường dẫn đến những sai lầm hay có gái trị tiêu cực. Sự thiếu hụt một hệ thống đánh giá thức ăn, cho phép chỉ ra được gia súc sẽ ăn bao nhiêu một loại thức ăn nào đó, đã khuyến khích xuất khẩu các gia súc có tiềm năng năng suất cao nhưng không thích hợp như bò Friesian đến những phần còn lại của thể giới nơi mà chất lượng thức ăn quá thấp không đủ để gia súc phát hyu hết tiềm năng sản xuất của chúng. Kết quả đã rõ. Bò phải cố gắng đáp ứng tiềm năng của chúng bằng cách huy động các mô của cơ thể, chúng ngừng chu ký sinh dục và thậm chí mất khả năng kháng bệnh và chết. Hiện tượng này hiện vẫn còn đang tiếp diễn. Nếu như chúng ta có một phương pháp xác định tiềm năng của thức ăn, thay vì các giá trị thức ăn được thể hiện là ME, DE hoặc NE, chúng ta có thể có một cơ sở quan trọng để làm cho tiềm năng của thức ăn phù hợp với tiềm năng của gia súc. Nói một cách khác, chúng ta có thể quyết định về các hệ thống sản xuất thích hợp nhất ở các vùng khác nhau nhằm làm cho tiềm năng của nguồn thức ăn phù hợp với tiềm năng sản xuất của gia súc một cách bền vững. Ví dụ, chúng ta sẽ quyết định xem liệu tiềm năng nguồn thức ăn có phù hợp với sản xuất sữa cao hơn nhu cầu của bê bú hay không? Hay chúg ta có thể quyết định tiềm năng thức ăn có đủ cho sinh sản, vỗ béo và làm việc hay không, hay chỉ cho duy trì và tồn tại. Yak ở vùng Ngoại Mông hai đến ba năm mới sinh một bê Tỷ lệ sinh sản thấp, có thể là như vậy, nhưng đấy là sự phù hợp hoàn hảo giữa tiềm năng năng suất và tiềm năng nguồn thức ăn của địa phương. Cừu mặt đen ở vùng núi Scotland sinh ít hơn một con mỗi năm. Một lần nữa đây là sự phù hợp hoàn hảo giữa tiềm năng năng suất và tiềm năng nguồn thức ăn của địa phương, nhưng chúng tôi chưa thấy có hệ thống đánh giá thức ăn nào đề xuất điều này. Nếu như chúng ta nỗ lực để tăng tỷ lệ sinh sản trong những tình huống như vậy, tỷ lệ chết tăng lên sẽ giảm số con sinh ra và hệ thống sẽ không còn bền vững nữa.
Cách tiếp cận mới này có thể tạo nên nền tảng của một hệ thống đánh giá thức ăn mới phù hợp với thức ăn thô và thay thế cho hệ thống đã lỗi thời 100 năm mà chúng ta đang có?
Chỉ số giá trị đề xuất cho thức ăn thô có thể được xem như chỉ số tiềm năng thức ăn. Nếu giá trị hơn 30, bò của chúng ta có thể ăn hơn mức duy trì; nếu nhỏ hơn 30 chúng sẽ giảm khối lượng. Chỉ số giá trị đương nhiên biến động giữa các loại và giống gia súc nhai lại. Chỉ số giá trị ở trâu dường như thấp hơn ở bò và chỉ số này ở bò địa phương đã thích nghi thấp bò cải tiến. Các giống với dung tích dạ cỏ so với khối lượng cơ thể lớn yêu cầu chỉ số giá trị thấp hơn giúp chúng ăn đủ cho duy trì.
Phương pháp tính toán ở trên rẻ, chỉ khoảng 1% chi phí xác định năng

60
lượng trao đổi in vivo. Tôi tin rằng phương pháp này tốt hơn nhiều và có thể vẫn rẻ hơn. Blummel (người đã đến làm việc với chúng tôi) sử dụng phơng pháp in vivo gas production để xác định tốc độ sản xuất khí thay vì các giá trị tĩnh tại. Bằng cách sử dụng phương trình ‘nhỏ’ thôi p = a + b (1-e-ct) nhưng áp dụng cho gas production invivo (6) thấy rằng có thể ớc đoán lượng ăn vào (r = 0,88) tăng trọng (r = 0,94) và chất khô tiêu hóa ăn vào (r = 92) với độ chính xác gần như độ chính xác thu được từ phương pháp nylon bag.
Vào thời điểm đó có vô số các hệ thống đánh giá thức ăn có sự biến động lớn. Tôi nghĩ chúng ta có thể đưa ra một hệ thống toàn cầu xác định tiềm năng của thức ăn thô và lý do là hệ thống được đề xuất dựa trên cơ sở thức ăn đáp ứng với vi sinh vật dạ cỏ như thế nào. Vi sinh vật trong dạ cỏ là tương tự nhau ở khắp thế giới. Mặc dù chúng ta cần một cơ sở dữ liệu lớn hơn nữa đêr có các chỉ số tiềm năng thức ăn tốt hơn, điều thú vị là các hệ số cho A, B và C tương tự nhau trong các thí nghiệm tại Scottland và Tanzania.
Không nghi ngờ gì nữa sẽ có những thức ăn ngoại lệ như thân cây chuối, những thức ăn này không thể đưa vào hệ thống. Thức ăn với các nhân tố kháng dinh dưỡng, có thể ảnh hưởng đến hoặc vi sinh vật hoặc vật chủ, sẽ không thể đưa vào hệ thống, nhưng những thức ăn này bản thân chúng lại gợi ra câu hỏi thú vị. Hoàn toàn có thể tiến hành một cuộc điều tra về tiềm năng thức ăn cho các mùa khác nhau ở các hệ sinh thái khác nhau nhằm làm cho năng suất của gia súc phù hợp với tiềm năng của nguồn thức ăn và luôn nhớ rằng nhiều giống gia súc có đặc tính sản xuất theo mùa vụ rất mạnh mẽ. Chúng ta cũng có thể đánh giá xem tiềm năng thức ăn có thể tăng lên chẳng hạn bằng xử lý hóa học có kinh tế không? Hay có cần nhập khẩu các thức ăn khác nữa không?
Tôi thiết nghĩ lĩnh vực nghiên cứu này xứng đáng được đầu tư nhiều hơn vì nó rất quan trọng giúp nông dân biết được tỷ lệ thay đổi thức ăn hợp lý, giúp những người lập kế hoạch chăn nuôi dự đoán được năng suất của gia súc, lập được kế hoạch xuất hoặc nhập khẩu, và nhất định lĩnh vực nghiên cứu này cũng quan trọng cho gia súc vì trong tương lai chúng ta có thể tránh được rất nhiều bất cập giữa tiềm năng nguồn thức ăn và năng suất.
Tài liệu tham khảo cho phần 2.5 1. Ørskov, E.R., Reid, G.W. and Kay, M. 1988. Prediction of intake by cattle
from degradation cgaracteristics of roughages. Anim. Prod. 46, 1, 29-34. 2. Reid, G.W., Ørskov, E.R. and Kay, M. 1988. A note on the effects of
variety, type of straw and ammonia treatment on digestibility and on growth rate in steers. Anim. Prod. 47, 157-160.
3. Kibon, A. and Ørskov, E.R. 1993. The use of degradation characteristics of browse plants to predict intake and digestibility by goats. Anim. Prod. 57, 247-251.
4. Shem, M.N., Ørskov, E.R. and Kimambo, A.E. 1995. Prediction of vonluntary dry matter intake, digestible dry matter intake and growth rate of cattle from the degradation characteristics of tropical foods. Animal

61
Science. 60, 65-74. 5. Khazaal, K. Dentiho, M.T., Ribeiro, J.M. and Ørskov, E.R. 1993. A
comparision of gas production during incubation with rumen contents in vitro and nylon bag degradability as predictors of the apparent digestibility in vivo and the voluntary intake of hays. Anim. Prod. 57, 105-112.
6. Blummel, M. and Ørskov, E.R. 1993. Comparision of in vitro gas production and nylon bag degradability of roughages in predicting feed intake in cattle. Anim. Fd. Sci. Tech. 40, 109-119.

62
Hướng nghiên cứu 3. Động thái thoát qua dạ cỏ của thức ăn Hướng nghiên cứu 3.1. Tốc độ thoát ra khỏi qua dạ cỏ của thức ăn và ảnh hưởng của nó đến tỷ lệ tiêu hoá
Chúng ta đều đã rõ là khi lượng thức ăn ăn vào tăng lên, thức ăn đi qua dạ cỏ nhanh hơn và do đó quá trình lên men thức ăn ở dạ cỏ là không hoàn toàn. Phương pháp nylon bag cho ra một đường cong phân giải thức ăn, tỷ lệ phân giải thức ăn tăng theo thời gian hướng đến tỷ lệ phân giải thức ăn tiềm năng. Tuy nhiên tỷ lệ phân giải có hiệu quả (Effective degradability) phụ thuộc vào thời gian thức ăn lưu lại trong dạ cỏ, và có thể chỉ xác định được nếu đo được tốc độ thoát ra khỏi dạ cỏ của thức ăn. Bây giờ chúng ta sẽ xem làm thế nào để đo được tốc độ thoát ra khỏi dạ cỏ của thức ăn một cách tốt nhất.
Ở thời điểm chúng tôi nghiên cứu vấn đề này, Tiến sỹ Peter Uden từ Thụỵ Điển tới phòng thí nghiệm của chúng tôi. Ông đã làm việc với tiến sĩ Peter van Soest và họ đã phát triển một phương pháp đánh dấu sử dụng crom để đánh dấu xơ của khẩu phần thức ăn thô. Phương pháp này có thể được sử dụng cho thức ăn bổ sung protein hay không? Phương pháp này, sử dụng dichromate natri, nên thu lại chất nền không bị phân giải trong dạ cỏ. Chúng tôi đưa thức ăn bổ sung protein đã được sử lý vào dạ cỏ của cừu và cố gắng đo sự sụt giảm nồng độ của dichromate natri, nghĩa là đo tốc độ pha loãng tan của nó, bằng cách lấy mẫu từ dạ cỏ. Chúng tôi không thành công ngay từ đầu, kết quả chúng tôi nhận được quá biến động, nhưng sau đó chúng tôi nhớ đến công trình của Tiến sĩ Larry Grovum từ Canada. Ông đã chỉ ra rằng tốc độ thoát ra khỏi dạ cỏ của các mảnh thức ăn có thể được dự đoán một cách chính xác từ đồ thị giảm nồng độ của chất đánh dấu khẩu phần bài tiết ra từ phân. Một lý do cho sự biến động kết quả của chúng tôi là do bản chất không đồng nhất của chất chứa dạ cỏ. Chúng tôi sau đó đã thấy rằng khi chất chứa dạ cỏ được làm đồng nhất bằng cách nghiền khẩu phần, chúng tôi nhận được kết quả tương tự từ mẫu chất chứa dạ cỏ hay mẫu phân, nhưng mẫu phân lấy dễ hơn nhiều và không cần thiết phải dùng canula. Gia súc được cho ăn thức ăn đã xử lý với chất đánh dấu bằng cách cho uống hoặc ăn với rỉ mật. Một cán bộ nghiên cứu trẻ từ Bulgaria, Gancho ganev đã đén chỗ chúng tôi và là người đầu tiên cho thấy phương pháp này hoạt động tốt (1).
Đầu tiên chúng tôi xác nhận được rằng sự thoát ra khỏi dạ cỏ của các mảnh thức ăn nhỏ trong khẩu phần có thức ăn kích thước dài, nhanh hơn sự thoát ra khỏi dạ cỏ của các mảnh thức ăn nhỏ trong khẩu phần thức ăn tinh (1).
Mohamad Elimam thực hiện các nghiên cứu tiếp theo trong luận văn nghiên cứu sinh Tiến sỹ của mình (2, 3, 4, 5, 6). Nghiên cứu của Mohamad Elimam cho thấy tốc độ thoát ra khỏi dạ cỏ của các mảnh thức ăn nhỏ bị ảnh hưởng rất lớn bởi mức ăn (Bảng 3.1).
Bảng 3.1. Ảnh hưởng mức ăn đến tốc độ thoát ra khỏi dạ cỏ của bột cá và bột đậu
tương được đánh dấu bằng Chrom.

63
Cừu Bò sữa
(Mức ăn so với nhu cầu duy trì)
Tốc độ thoát ra khỏi dạ cỏ
(%/giờ)
(Mức ăn so với nhu cầu duy trì)
Tốc độ thoát ra khỏi dạ cỏ ( %/giờ)
0,5 0,8 1,5 6,5 1,0 1,8 2,0 7,2 1,5 2,6 2,5 9,1 2,0 3,2 3,0 8,8
Có thể thấy rằng tốc độ thoát ra khỏi dạ cỏ của protein dao động từ 1 đến
9%/giờ phụ thuộc vào mức nuôi dưỡng. Ở một vài bò sữa, tốc độ thoát ra khỏi dạ cỏ đôi khi lớn hơn 10%/giờ (5). Ở thí nghiệm khác, ảnh hưởng của bản chất của khẩu phần bò sữa (tỷ lệ phần cỏ khô dài so với thức ăn tinh) được nghiên cứu. Tỷ lệ phần cỏ dài so với thức ăn tinh là 1,0; 0,75; 0,50 và 0,25 của tổng DM khẩu phần. Tốc độ thoát ra khỏi dạ cỏ lần lượt là 7,9; 8,8; 8,6 và 6,4%/giờ, giá trị cuối cùng thấp hơn đáng kể so với 3 giá trị còn lại. Như vậy, kích cỡ của của cỏ khô rất quan trọng. Kích cỡ này biến động từ dài, 40 mm hoặc 20 mm, hoặc nghiền đến 5 mm và làm thành thức ăn viên. Với 4 kích cỡ như trên tốc độ thoát ra khỏi dạ cỏ lần lượt là 8,0; 8,6; 8,8 và 4,7%/giờ. Nghiền đã làm giảm tốc độ thoát ra khỏi dạ cỏ của các mảnh thức ăn bổ sung protein nhỏ, và ảnh hưởng của nghiền là đáng kể.
Dựa phần lớn vào các kết quả này chúng tôi đề nghị trong nhu cầu dinh dưỡng cho gia súc nhai lại của ARC rằng chỗ nào tỷ lệ phân giải đã được ước tính chúng ta nên sử dụng tốc độ thoát ra khỏi dạ cỏ 2,5 đến 8%/giờ cho mức nuôi dưỡng thấp ở bò có chửa, cho mức nuôi dưỡng cao bò thịt vỗ béo, và cho bò sữa.
Ở thí nghiệm tiếp theo chúng tôi cố gắng giữ môi trường dạ cỏ không đổi và thay đổi bản chất của thức ăn đánh dấu. Đây là một thí nghiệm lớn với 22 bò sữa được nuôi bằng khẩu phần trộn hoàn chỉnh: 50 % rơm sử lý ammonia và 50 % thức ăn tinh. Ba thức ăn đánh dấu được sử dụng gồm: rơm dài được xử lý ammonia, rơm sử lý ammonia đã nghiền (5 mm) và bột cá. Polyethylene glycol (PEG) cũng được dùng để xác định tốc độ thoát ra khỏi dạ cỏ. Kết quả chính của thí nghiệm được trình bày dưới đây:
Tốc độ thoát ra khỏi dạ cỏ: Mảnh rơm kích thước nhỏ 3,1%/giờ Rơm dài 2,8%/giờ Bột cá 7,0%/giờ Dung dịch (PEG) 14,6%/giờ Thời gian lưu lại tại dạ cỏ của các mảnh thức ăn kích thước nhỏ 47,6 giờ Thời gian lưu lại tại dạ cỏ của các mảnh thức ăn kích thước dài 53,2 giờ

64
Tốc độ thoát ra khỏi dạ cỏ không khác nhau đáng kể về mặt thống kê giữa
mảnh thức ăn kích thước dài và ngắn nhưng do pha dừng để mảnh thức ăn có kích thước dài giảm thành mảnh thức ăn có kích thước nhỏ, nên thời gian lưu lại tại dạ cỏ của các mảnh thức ăn kích thước dài lớn nhất. Một khía cạnh thú vị khác là sự khác nhau lớn giữa các mảnh xơ nhỏ và bột cá, không nghi ngờ gì nữa là do sự khác nhau về trọng lực riêng. Điều này có nghĩa là bổ xung protein kết hợp với xơ như bột linh lăng sẽ khác với bổ xung protein có từ hạt hoặc từ động vật hoặc từ bột cá. Cũng đáng luư ý là dịch lỏng thoát ra khỏi dạ cỏ với tốc độ gần gấp đôi ttốc độ này của chất rắn.
Một khía cạnh thú vị khác là có thể có liên quan giữa tốc độ thoát ra khỏi dạ cỏ của chất rắn và tỷ lệ tiêu hoá biểu kiến. Tương quan âm - 0,81 giữa tốc độ thoát ra khỏi dạ cỏ và tỷ lệ tiêu hoá biểu kiến là không mong đợi, có thể như vậy, nhưng kết quả này đảm bảo 1 lần nữa và tăng thêm niềm tin rằng phương pháp này tốt và sử dụng được. Kết quả trên dẫn chúng tôi tới một câu hỏi nghiên cứu khác là: sự khác nhau giữa các cá thể bò có chắc chắn khi cách kết hợp khẩu phần và mức ăn là khác và sự khác nhau đó có thể sử dụng để chọn lọc bò sử dụng tốt hơn thức ăn thô do có thời gian thức ăn lưu lại lâu hơn trong dạ cỏ? Để trả lời câ hỏi trên, chúng tôi chọn 3 con bò có tốc độ thoát ra khỏi dạ cỏ của thức ăn cao nhất và 3 bò có tốc độ thoát ra khỏi dạ cỏ của thức ăn thấp nhất và so sánh chúng khi cho ăn khẩu phần thức ăn thô thấp (LR), khẩu phần thức ăn thô cao (HR) ăn tự do hoặc ăn hạn chế: 70% mức ăn tự do (8), xem Bảng 3.2.
Bảng 3.2. Tốc độ thoát ra khỏi dạ cỏ của thức ăn và tỷ lệ tiêu hoá in vivo của khẩu phần có tỷ lệ thức ăn thô thấp và cao ở bò sữa được chọn lọc theo tốc độ thoát ra
khỏi dạ cỏ của thức ăn thấp (LO) và cao (HO).
Thức ăn thô thấp Thức ăn thô cao Chất khô ăn vào (kg/ngày) Ăn tự
do Ăn hạn
chế Ăn tự
do Ăn hạn
chế LO 14,9 10,6 10,7 7,6 HO 14,6 10,4 10,6 7,5
Tốc độ thoát ra khỏi dạ cỏ của thức ăn (%/giờ)
LO 2,98 2,57 2,75 2,54 HO 3,23 3,09 3,02 3,16
Tỷ lệ tiêu hoá DM (%) LO 68,8 73,1 65,7 65,5 HO 67,4 69,6 63,6 64,0
Ở đây có thể thấy rằng sự khác nhau là chăc chắn bất chấp mức ăn vào và
loại khẩu phần; và hơn thế nữa kết quả trên cho thấy sử dụng thức ăn rõ ràng tốt hơn khi tố độ thoát qua chậm. Thức ăn ăn vào cho 2 nhóm bò là tương tự nhau.

65
Liệu chúng ta có thể chọn bò tiêu hoá thức ăn thô tốt hơn hay không? Có. Nếu kế hợp chọn lọc bò tiêu hoá thức ăn thô tốt với dung tích dạ cỏ thì điều này là có thể. Tài liệu tham khảo cho phần 3
1. Ganev, G., Ørskov, E.R. and Smart, R.I. 1979. The effect of roughage or concentrate feeding and rumen retention time on total degradation of protein in the rumen. J. agric. Sci., Camb. 93, 651-656.
2. Elimam, M.E. and Ørskov, E.R. 1981. Determination of rate of ourflow of protein supplements from the rumen by measuring the rate of excretion of Cr treated protein supplements and PEG in the faeces. Anim. Prod. 32, 386.
3. Elimam, M.E. and Ørskov, E.R. 1982. Effect of feeding level of grass on the rate of outflow of protein supplements from the rumen of sheep. Proc. Nutr. Soc. 41, 29A.
4. Elimam, M.E. and Ørskov, E.R. 1982. The effect of level of feeding on rate of outflow of Cr treated protein supplements in dairy cows. Proc. Nutr. Soc. 41, 87A.
5. Elimam, M.E. and Ørskov, E.R. 1984. Factors affecting the outflow of protein supplements form the rumen. 1. Feeding level. Anim. Prod. 38, 45-51.
6. Elimam, M.E. and Ørskov, E.R. 1984. Estimation of rates of outflow of protein supplement from the rumen by determining the rate of excretion of chromium treated protein supplements in faeces. Anim. Prod. 39, 77-80.
7. Balch, C.C., Armstrong, D.C. Greenhalgh, J.F.D., Miller, E.L., Ørskov, E.R., Roy, J.H.B., Smith, R.H., and Taylor, J.C. Nutrient Requirement of Ruminant Livestock. Supplement 1, CAB Slough, London.
8. Ørskov, E.R., Oiwang, I. and Reid, G.W. 1988. A study on consistency of differences between cows in rumen outflow rate of fibrous particles and other substrates and consequences for digestibility and intake of roughages. Anim. Prod. 47, 45-51.

66
Hướng nghiên cứu 4. Phương pháp in vitro gas production Hướng nghiên cứu 4.1. Sự phát triển của phương pháp in vitro gas production
Sự phát triển của phương pháp nylon bag động, nghĩa là mô tả tốc độ cũng như tỷ lệ phân giải của các chất dinh dưỡng khác nhau, là một bước tiến lớn nhưng phương pháp này phụ thuộc vào cừu hoặc bò mổ lỗ dò. Chúng ta có thể có cách tiếp cận tương tự trong điều kiện in vitro đượckhông? Chúng tôi thấy rằng về tổng thể các hệ thống in vitro của Tilley& Terry đã đo được các chỉ tiêu tương tự: sự mất đi của các chất dinh dưỡng, nhưng phương pháp gas production do Menke phát triển ở Đức hấp dẫn vì phương pháp này đo đạc rất hiệu quả chỉ tiêu ngược lại: sự xuất hiện của các sản phẩm lên men hay một cách chính xác hơn, khí CH4 và CO2 thoát ra từ dung dịch đệm bởi các axít béo bay hơi sinh ra trong quá trình lên men. Michael Blummel - một sinh viên đến từ phòng thí nghiệm ở Hohenheim, Đức Sự đã giúp chúng tôi rất nhiều trong phát triển phương pháp của Menke. Tại thời điểm đó chúng tôi vừa mới kết thúc công trình nghiên cứu ước tính lượng thức ăn thô ăn vào từ các đặc điểm phân giải (Hướng nghiên cứu 2.5). Việc Michael Blummel đến phòng thiư nghiệm đã tạo cơ hội thử nghiệm động thái gas production, sử dụng cùng một công thức với phương pháp nylon bag: p = a + b (1-e-ct). Tuy nhiên ở đây p là lượng khí sinh ra ở thời điểm t, c là hằng số tốc độ và (a+b) là tổng số khí sinh ra. Liệu chúng tôi có thể dự đoán lượng ăn vào từ các đặc điểm phân giải sinh ra từ phương pháp sinh khí? Mẫu thức ăn được ủ với dung dịch đệm và dịch dạ cỏ trong một xylanh thuỷ tinh to, và công việc ử đây chỉ đơn giản là đo lượng khí sinh ra khi piston di chuyển lên trên và khí được sinh ra trong xylanh. Phươn pháp này có một ưu điểm hơn phương pháp nylon bag. Mỗi xylanh có thể được đo nhiều lần sau khi bắt đầu ủ thức ăn, trong khi đó với túi nylon, mỗi túi chỉ cho phép một lần đo.
Tương quan giữa lượng khí sinh ra với vật chất khô ăn vào có r = 0,88; giữa lượng khí sinh ra với chất khô tiêu hóa ăn vào có r = 0,93 và giữa lượng khí sinh ra với tăng trọng có r = 0,95. Kết quả này tốt gần tương đương với phương pháp nylon bag. Dĩ nhiên, cũng có một sự hoà hợp tuyệt vời giữa phương pháp nylon bag và phương pháp gas production. Tương quan giữa 2 phương pháp ở 8, 24, 48 và 72 giờ lần lợt có r = 0,93; 0,97; 0,97 và 0,97 (1).
Dường như vì thế phương pháp gas production đã có một tương lai lớn và sau đó Blummel bắt đầu khám phá khả năng sử dụng phương pháp này để xác định tỷ lệ giữa các axít béo bay hơi và sinh khối vi sinh vật.
Chúng tôi bắt đầu tham gia vào một dự án Châu Âu lớn với các nước vùng Địa Trung Hải như Bồ Đào Nha, Hy Lạp và Pháp. Sự tham gia này đã cho phép chúng tôi tiếp cận toàn bộ các loại thức ăn và đặc biệt các loài cây bụi còn rất mới với chúng tôi. Các loại cây thân gỗ như vậy thường chứa một lượng lớn các nhân tố kháng dinh dưỡng. Lúc này Khamal Khazal từ Lebanon tới làm việc với chúng tôi. Chúng tôi phát hiện ra rằng sự phân giải thức ăn khi sử dụng phương pháp

67
nylon bag và sinh khí thường không cho kết quả như nhau khi mẫu là những thức ăn có chứa các nhân tố kháng dinh dưỡng. Thức ăn càng nhiều nhân tố kháng dinh dưỡng, sự khác nhau về kết quả giữa hai phương pháp càng lớn (2). Lý do cho sự khác biệt này khá rõ. Các nhân tố kháng dinh dưỡng có trong mẫu thức ăn ủ trong nylon bag trong dạ cỏ bị pha loãng nên có ít hoặc không ảnh hưởng đến phân giải mẫu trừ khi bản thân gia súc mổ lỗ dò dạ cỏ cũng được nuôi với thức ăn có chứa các nhân tố kháng dinh dưỡng. Ngược ở phương pháp gas production các nhân tố kháng dinh dưỡng có trong mẫu thức ăn ảnh hưởng đến quá trình sinh khí vì tỷ lệ mẫu với dịch dạ cỏ lớn hơn nhiều.
Chúng tôi cũng quan sát thấy rằng các xác định về hóa học các nhân tố kháng dinh dưỡng nhóm fenolic, như fenolic chiết xuát, tanin đậm đặc và proanthocyanidins, không cho ra một chẩn đoán tốt về về ảnh hưởng của chất kháng dinh dưỡng vì một số tanin hoạt động mạnh trong khi một số khác thì không. Chúng tôi phân vân dùng polyvinylpolypyrolidone (PVP) có được không?, PVP đã từng được dùng để tạo phức hợp các nhân tố kháng dinh dưỡng cũng như để tách tanin từ các enzyme, có thể sử dụng để tạo phức hợp tanin trong xylanh do đó lượng khí sinh khí khi có hoặc không có PVP sẽ cho biết hàm lượng các nhân tố nhóm fenolic. Đúng là PVP có thể sử dụng được trong trường hợp (3) như các minh họa từ mọt số ví dụ trong bảng 4.1. Bảng 4.1. Ảnh hưởng của việc đưa PVP vào các mẫu cây bụi khác nhau đến lượng
khí sinh ra in vitro (3). Loài cây bụi Lượng khí sinh ra (ml) sau
24 giờ 48 giờ Areimisia campestris Không có PVP 25,1 30,5 Có PVP 29,5 34,9 % tăng 17 14 Teucrium polium Không có PVP 28,6 33,0 Có PVP 29,6 33,5 % tăng 4 1 Adenocarpus decorticans Không có PVP 35,1 38,7 Có PVP 35,5 39,3 % tăng 1 2 Quercus coccifera Không có PVP 14,8 21,3 Có PVP 18,5 26,2 % tăng 25 23
Rõ ràng có 2 loại cây bụi có hàm lượng các nhân tố kháng dinh dưỡng cao
là Aremisin và Quercum, các nhân tố này bị trung hoà khi phức hợp với PVP hình thành. Cũng rất thú vị thấy rằng việc tăng lượng khí sinh ra có tương quan yếu với các tanin đã được xác định về mặt hoá học. Dường như không phải tất cả các tanin đều ức chế quá trình lên men như nhau và chỉ các mẫu thức ăn được bảo vệ một

68
cách hiệu quả bởi tanin bây giờ đang được giải phóng và quá trình này được phản ánh ở sự tăng sản xuất axít béo bay hơi.
Sau đó nó chúng tôi thấy rằng (4) rằng nếu như không có hơp chất nào vượt trội hơn PVP trong tạo phức hợp các yếu tố kháng dinh dưỡng phenolic và không phổ biến hơn PVP thì nên sử dụng polyethylene glycol (PEG). Hiện nay PEG đã được sử dụng cho các phân tích in vitro các mẫu nghi ngờ có chứa các yếu tố kháng dinh dưỡng.
Đây là một ví dụ tốt bổ sung cho phương pháp in sacco và in vitro. Trong khi với thức ăn không chứa các nhân tố kháng dinh dưỡng, phương pháp nylon bag chắc chắn vượt trội hơn so với phương pháp sinh khí gas producton, phươn pháp nylon bag chậm hơn và cần có gia súc mổ lỗ dò với canule đủ lớn đẻ đưa các túi nylon vào dạ cỏ. Phương pháp sinh khí in vitro có lợi thế trong việc xác định các yếu tố kháng dinh dưỡng. Phương pháp sinh khí in vitro cũng có thể xác định được phần hoà tan ngay của thức ăn được cho là sẽ bị lên men hoàn toàn khi sử dụng phương pháp nylon bag, trên thực tế là phần được lên men hay không. Cuối cùng phương pháp này không phụ thuộc hoàn toàn vào gia súc mổ lỗ dò. Dịch dạ cỏ có thể lấy được bằng cách dùng bơm dạ dày hoặc chí ít từ cannule dạ cỏ rất nhỏ.
Tài liệu tham khảo cho 4 1. Blummel, M. and Ørskov, E.R. 1993. Comparision of in vitro gas
production and nylon bag degradability of roughages in predicting feed intake in cattle. Anim. Fd. Sci. Tech. 40, 109-119.
2. Khazaal, K.A., Markantonatos, X., Natis, A. and Ørskov, E.R. 1993. Changes with maturity in fibre composition and levels of extractable polyphenols in Greek browse: effects on in vitro gas production and in sacco dry matter degradation. J. Sci. Food Agric. 63, 237-244.
3. Zahedifar, M., Castro, F.B. and Ørskov, E.R. 1994. Effect of plant phenolic compounds extracted with either water or acetone solution on in vitro gas production. 45th Annual Meeting of the European Association for Animal Production, Edinburgh, N5. 17, 157.
4. Khazaal, K.A, Parissi, Z., Tsiouvaras, C., Natis, A. and Ørskov, E.R. 1996. Assessment of phenolics related antinutritive levels using the in vitro gas production technique: a comparision between different types of polyvinylpolyrrolidone or polyethylene glycol. J. Sci. Food and Agric. 71, 405-414.

69
Hướng nghiên cứu 5. Bảo vệ protein thoát khỏi sự phân giải dạ cỏ Hướng nghiên cứu 5.1. Phản xạ rãnh thực quản và tiêu hoá sau dạ cỏ
Chúng tôi đã chứng minh được ở cả trong các thí nghiệm tiêu hoá và các thí nghiệm sử dụng thức ăn năm 1967 và 1968 là protein bị phân giải rất mạnh trong dạ cỏ, các công trình nghiên cứu khác ở Úc, Pháp và công trình của Margaret Chalmers ở Viện Rowett cũng xác nhận kết quả trên. Mong muốn rát tự nhiên là tìm ra phương pháp bảo vệ protein tháot khỏi phân giải ở dạ cỏ. Tôi bị cuốn hút vào việc nghiên cứu xem có hay không một cơ chế tự nhiên mà gia súc non sử dụng để sữa được đổ trực tiếp vào dạ múi khế, phản xạ đóng rãnh thực quản, có thể sử dụng để đưa protein bổ sung trực tiếp tới dạ múi khế được không. Tôi đã nói chuyện với kỹ thuật viên X quang, David Benzie. David là người chụp X quang lâu năm. Ông ta là người rất có lập trường. Vào thời điểm đó ông ta đã bị một đột quị và đi lại không được tốt. Các ngón tay của ông bị cháy sém do tiếp xúc với bức xạ quá nhiều khi thao tác với các viên Radon phóng xạ từ rất lâu. Tôi nhớ rất rõ rằng ông Benzie, người ngưỡng mộ công việc của Wester (Wester I. 1930. Berl. Tierarztl Wsch. 46,1.) nói “Muối ở trong sữa đã kích thích đóng rãnh thực quản”. Nếu quả thât là như vậy thì điều chúng ta cần biết là muối nào, có thể chúng ta cho ăn thức ăn bổ sung protein bổ sung cùng với một số muối nhất định.
Chúng tôi tiến hành thí nghiệm để thử tất cả các loại muối bằng cách cho uống, cùng với barium sulphat, natri, kali và đồng thậm chí cũng được bổ sung vào thuốc tẩy giun sán để gây ảnh hưởng đến việc đóng rãnh tực quản. Không có muối nào gây ra việc đóng rãnh thực quản; đôi khi một lượng nhỏ muối nhìn thấy ở dạ múi khế nhưng phần lớn dịch vẫn vào dạ cỏ. Một thời gian dài chúng tôi đã không đi đến đâu cả. Tuy nhiên chúng tôi không dừng lại! Tôi đã nhìn thấy một con cừu được nuôi nhân tạo, thức ăn lỏng được cho ăn bằng một một cái chai gắn với núm vú cao xu. Tôi đã thử cho dung dịch muối bằng cách này và không nghi ngờ gì nữa tất cả dung dịch đã đi vào dạ múi khế, như David Benzie đã nói. Sau đó tôi tiến hành bước thí nghiệm khác, tôi bỏ muối ra và chỉ cho uống nước không. Ở đây tôi phải rất cẩn thận; có tể có một chút chủ quan khi đọc hình ảnh X quang và Benzie chắc chắn rằng đó là muối, sau đó khi ông Benzie tuyên bố 100% dịch cho uống đã ở dạ múi khế, tôi hỏi ông ta xem cái gì xảy ra nếu tôi chỉ sử dụng nước cho những con cừu đó. Ông ta nói “Không đáng để thử nữa, tôi đã thử rồi”. “Tốt” tôi nói, “Những con cừu đó chỉ có nước ở trong chai đúng không!”.
Bây giờ chúng tôi đã có một hệ thống làm việc tốt, câu hỏi là tại sao thức ăn lỏng hoặc nước chỉ đi vào dạ múi khế ở cừu cho uống bằng chai, nhưng thức ăn lỏng hoặc nước không đi vào dạ múi khế chai khi cừu được cho uống bằngphương pháp thông thường?. Chúng tôi đã bỏ núm vú ra, huấn luyện một vài cừu uống sữa từ máng và sau đó thay sữa bằng nước và cho uống từ máng, kết quả là nước cũng đi thẳng xuống dạ múi khế; tuy vậy nếu nước trong máng được cho cừu đang khát nứớc, nước sẽ xuống dạ cỏ. Chúng tôi cũng chỉ ra, dĩ nhiên vì đây là mục tiêu thực sự của thí nghiệm, rằng nếu như thức ăn bổ sung protein được bú từ chai chúng sẽ

70
đi trực tiếp đến múi khế, như vậy rõ ràng là các thức ăn bổ sung protein được bảo vệ hoàn toàn khỏi phân giải ở dạ cỏ bằng biện pháp do chính gia súc vận hành.
Vẫn còn một tình huống làm chúng tôi băn khoăn: tại sao thực quản đóng ở gia súc được huấn luyện mà không đóng ở gia súc không được huấn luyện khi cùng một loại chất lỏng đi qua cùng một loại thụ cảm thần kinh ở miệng và hầu họng? Phải có những yếu tố quyết định khác giống như phản xạ có điều kiện. Ở đây một lần nữa chúng tôi gọi cho Robin Kay (2), một người rất ngưỡng mộ các nghiên cứu của Pavlov. Robin Kay đã đặt một ống thông từ ngoài vào thực quản gia súc đã được huấn luyện uống từ chai. Bây giờ câu hỏi ở đây là: nếu chúng ta đưa một lượng dung dịch barium qua ống thông thì dung dịch sẽ đi đâu? Đúng như mong đợi nếu đưa dung dịch vào và cừu nhìn thấy chai, cừu sẵn sàng chấp nhận uống, dung dịch barium đi vào dạ múi khế, nếu cừu không nhìn thấy chai dung dịch barium đi vào dạ cỏ.
Đây là một quan sát rất hứng thú. Tuy nhiên, lần thử thứ tư hoặc thứ năm không thành công, dung dịch xuống dạ cỏ. Chúng tôi khá thất vọng phần. Chúng tôi đã sai ở đâu? Điều đã thực sự xảy ra lúc này là cừu đã học được cách phân biệt. Chúng học được rằng chúng không được uống khi ba người mặc áo blu trắng với

71
cái chai và một xilanh đầy barium hai lần một tuần xuất hiện! Thủ nghiệm chỉ có thể tiến hành hai lần một tuần vì barium phải được rửa sạch khỏi ruột trước khi thử nghiệm được lặp lại. Tôi đã phải tiếp tục cho ăn bằng chai hàng ngày và mọi thứ lại trở lai bình thường. Rõ ràng đây là phản xạ có điều kiện thực sự. Cừu không bao giờ nhận ra dung dịch thực ra là gì: nước, sữa, dịch dạ cỏ, chất thải sau biogas, beer, rượu vang hay bất cứ thức ăn bổ sung protein nào khác. Câu hỏi quan trọng là phản xạ sẽ hoạt động trong bao lâu, câu trả lời cho câu hỏi trên là phản xạ sẽ hoạt động dài nếu phản xạ được củng cố và sử dụng. Chúng tôi có những con dê cái đã có 5 lứa sữa tranh dành cái chai với lũ dê con. Nếu chúng ta dừng việc sử dụng phản xạ trong một giai đoạn – với một số gia súc chỉ một tuần là đủ – chúng ta không thể củng cố phản xạ được nữa. Chức năng của rãnh thực quản hoàn toàn độc lập với sự phát triển chức năng dạ cỏ.
Sự phát triển của phản xạ rãnh thực quản là sự phát triển rất sống động, cho đến bây giờ chúng tôi mới xâm nhập vào phần sau của đường tiêu hóa ở những gia súc với chức năng dạ cỏ bình thường.

72
Khai thác phản xạ rãnh thực quản
Rõ ràng protein là vấn đề đầu tiên trong nhiều việc phải làm. Ở thí nghiệm đầu tiên (3) chúng tôi cho một hỗn hợp protein thông qua ống thông thực quản vào dạ múi khế gia súc hoặc cho gia súc ăn một hỗn hợp protein ở dạng khô cùng với thức ăn tinh để thức ăn đi vào dạ múi khế. Kết quả chính của thí nghiệm được tóm tắt ở bảng 5.1(3); cân bằng N đã thực sự được cải thiện khi đua hỗn hợp protein vào dạ múi khế. Bảng 5.1. Cân bằng nitrogen ở 11 cừu được ăn protein bổ xung ở dạng được tiêu
hóa hoặc dạng khô như chất bổ xung vào khẩu phần cơ bản lúa mạch. Sử lý N ăn vào
(g/ngày) N nước tiểu
(g/ngày) N phân (g/ngày)
N giữ lại (g/ngày)
Protein khô 22,8 8,7 6,2 7,9 Protein dung dịch 22,9 6,5 6,6 9,8

73
SE 0,7 0,6 0,6 0,9 Bây giờ chúng ta xem nhiều loại protein bổ xung khác như bột cá, bột hạt hướng dương, men và bột đậu tương. Các kết quả chính được trình bầy trong bảng 5.2.
Bảng 5.2. Cân bằng nitơ khi các loại protein bổ xung khác nhau ở dạng lỏng được
đưa vào dạ múi khế. Khẩu phần N ăn vào
(g/ngày) N nước
tiểu (g/ngày)
N phân (g/ngày)
N giữ lại (g/ngày)
Lúa mạch và thức ăn tinh 14,3 4,4 4,8 5,1 + bột cá 20,2 6,4 4,3 9,4 + men 20,5 7,1 5,2 8,2 + bột hạt hướng dương 19,0 6,5 4,6 7,8 + bột đậu tương 18,9 5,8 5,2 7,9 SE 0,39 0,41 0,38 0,7
Rõ ràng cũng có sự khác nhau về giá trị sau dạ cỏ của các protein bổ sung
khác nhau nhưng đây là lần đầu tiên nó được chứng minh ở gia súc nhai lại, sụ khác nhau này được thể hiện ở lượng nitơ tích lũy khác nhau.
Ở đây tôi phải kể lại một việc tình cờ mà tôi luôn luôn nhớ tới với một sự hài lòng. Men chúng tôi sử dụng đợc sản xuất bởi BP. Đó là sản phẩm mới (Toprina) dựa trên cơ sở dầu đang rẻ, men này đang trở thành một đối thủ cạnh tranh lớn với tất cả các nguồn protein khác vào thời điểm đó. Dường như rất hấp dẫn để so sánh nó với các nguồn protein khác. Tôi gọi điện cho giám đốc của BP Nutrition và hỏi anh ta có quan tâm đến việc so sánh protein mới với các nguồn protein khác khi nó được bổ sung vào sau dạ cỏ. Ông nói là rất quan tâm và hỏi tôi cần bao nhiêu sản phẩm. Tôi nói 20 kg là thỏai mái. Một vài ngày sau một thùng 20 kg men được gửi đến phòng thí nghiệm của tôi. Vài ngày sau đó một nhân viên từ Uỷ Ban nghiên cứu Nông nghiệp đến thăm phòng thí nghiệm của tôi và chúng tôi đã thảo luận cái mà tôi đang nghiên cứu. Tôi cho anh ta thấy nguồn protein hấp dẫn mới mà tôi đã có.
Ngày tiếp theo tôi được gọi tới văn phòng giám đốc Viện nghiên cứu, Tiến sĩ Blaxter- có cái gì đó bất ổn đây - anh lấy protein đó ở đâu? Tôi nói tôi gọi điện cho giám đốc BP Nutrition. Sau đó Tiến sĩ Blaxter lấy xuống một tập tài liệu lớn và nói “Trong khoảng 18 tháng một nhóm ở Viện Rowett đã cố gắng có được nguồn protein bổ sung này cho các nghiên cứu cơ bản. Họ đã hứa sẽ có 200 g vào tháng 6, vậy làm cách nào anh lại có được 20 kg vào tháng 1?” Tôi xin lỗi tôi không biết nhưng tôi thấy hạnh phúc đưa ngay cho họ 200 g. Rõ ràng Tiến sĩ Blaxter đã đàm phán với quản lý của BP. Ông ta luôn muốn làm cái đó ngay lập tức trong khi tôi đi xuống tầng dứới!
Tuy vậy, đó chưa phải là kết thúc câu truyện. Chúng tôi đã sử dụng men

74
trong một vài thí nghiệm nhưng phát hiện thấy men đã nhiễm kẽm, khoảng 900 ppm. Chúng tôi gần như đã mạ kẽm những cừu thí nghiệm! Sau đó BP có một bài giảng tại trường đại học Aberdeen về thức ăn bổ sung protein mới mang tính cách mạng của họ. Ở thời gian đặt câu hỏi, tôi đã đề cập đến việc tôi đã tiến hành thí nghiệm với protein đó và hỏi tại sao nó chứa nhiều kẽm như vậy. Người giảng bài lùi lại và hỏi tôi từ đâu tới và mẻ protein nào tôi đang đề cập đến. Tôi đã cho ông ta cả hai thông tin trên. Ông ta nói ông ta sẽ xem xét nó. Ngày hôm sau tôi được gọi tới văn phòng của Tiến sỹ Blaxter và nhận được sự khiển trách. Bây giờ BP từ chối làm việc với chúng tôi. Tôi đã làm hỏng công trình nghiên cứu 2 năm của Viện do gây khó khăn cho công việc quảng cáo của BP. Sao tôi lại ngớ ngẩn thé nhỉ? Tôi cảm thấy thực sự hối tiếc. Tôi gọi điện cho giám đốc của BP Nutrition và nói với ông ta về kết quả thu đuwcj cho đến luc này và ước muốn của tôi làm một thí nghiệm thuyết cuối cùng để kết luận vấn đề. “Ngài cần bao nhiêu? Giám đốc của BP Nutrition hỏi. Tôi nói tôi cần 100 kg và 100 kg đã được gửi đến phòng thí nghiệm của tôi. Tôi gửi một văn bản tới Tiến sỹ Blaxter để nói với ông ta rằng tôi đã có 100 kg nhưng thực ra 50 kg là đủ và tôi có thể để dành 50 kg cho công trìh nghiên cứu 2 năm của nhóm protein! Tôi gặp Tiến sỹ Blaxter hai ngày sau đó và ông ta càu nhàu. Tôi không biết phải làm gì với anh đây! Ông ta bảo tôi. Lạy chúa thức ăn bổ sung protein mới mang tính cách mạng không bao giờ ra được thị trường do sự tăng giá dầu lửa.
Tôi làm thêm một thí nghiệm hết sức thú vị nữa (4) trong đó chúng tôi so sánh bột cá, casein và bột đậu tương hoặc ở dạng khô để vào dạ cỏ hoặc ở dạng lỏng để thoát qua dạ cỏ. Ở đây tôi đã đưa ra từ thoát qua (by-pass). Tôi ước tôi đã không bao đưa ra từ thoát qua vì nhiều đã sử dụng từ đó rất sai lầm, protein thoát qua không phải là protein đi qua dạ cỏ! Các kết quả quan trọng nhất được trình bày trong bảng 5.3.
Bảng 5.3. Ảnh hưởng của nguồn protein và cách cho ăn đến sử dụng protein khi
bổ xung vào khẩu phần lúa mạch cho cừu cai sữa sớm. Protein bổ sung
Phương pháp cho ăn
Nitơ ăn vào (g/ngày)
Nitơ nước tiểu
(g/ngày)
Nitơ phân (g/ngày)
Nitơ tích lũy
(g/ngày) Không - 12,1 3,0 4,8 4,3 Casein Khô 17,5 6,9 4,0 6,6 Casein Lỏng 17,3 5,8 4,3 7,2 Bột cá Khô 17,8 6,7 3,9 7,2 Bột cá Lỏng 17,5 5,1 4,5 7,9 Đậu tương Khô 17,4 6,6 4,4 6,3 Đậu tương Lỏng 17,3 5,4 5,1 6,8 SE 0,3 0,2 0,4
Kết quả cho thấy protein dạng lỏng cho kết quả tốt hơn đáng kể so với

75
protein dạng khô, chứng tỏ ảnh hưởng của phân giải dạ cỏ các thức ăn bỏ sung protein dạng khô. Bột cá được sử dụng tốt nhất dù bổ sung theo cách nào, phản ánh cả hai đặc tính: tỷ lệ phân giải thấp của bột cá ở dạ cỏ và thành phần amino axít tối ưu hơn các protein khác khi bột cá thoát qua dạ cỏ.
Công việc tiếp theo là nghiên cứu ảnh hưởng của việc cho ăn protein qua rãnh thực quản để protein thoát qua dạ cỏ, đây là cách bổ sung cho khẩu phần không gây ra hạn chế vật lý đối với lượng thức ăn ăn vào của khẩu phần cơ sở rơm lúa mạch bổ sung ure (5). Kết quả được trình bày trong bảng 5.4. Bảng 5.4. Ảnh hưởng của việc cho ăn thức ăn đậm đặc protein bột cá (FPC) bằng
chai đến lượng thức ăn ăn vào của khẩu phần lúa mạch ở các khối lượng sống khác nhau (LW) và đến tăng trọng trung bình.
Ăn vào (g/ngày) ở:
25 kg LW
35 kg LW 45 kg LW Tăng trọng trung bình (g/ngày)
Lúa mạch + urê 851 1078 1265 230 + 17 g FPC/ngày 994 1190 1415 300 + 34 g FPC/ngày 927 1196 1561 326 + 51 g FPC/ngày 1003 1241 1416 332 + 10g FPC/ngày 833 1062 1246 224
SE 48 49 43 22
Rất rõ ràng protein kích thích ăn vào trong khi bổ sung urê không kích thích như vậy. Nói một cách khác tốc độ trao đổi chất tăng có thể làm tăng tính ngon miệng và tăng trọng. Quan sát này chưa từng bao giờ được đề cập tới!
Bây giờ cái gì sẽ xảy ra nếu năng lượng được cho thoát qua dạ cỏ? Chúng tôi chọn lactose. Liệu lượng ăn vào có giảm theo lượng lactose cho vào dạ múi khế?
Bảng 5.5. Ảnh hưởng của việc bổ sung lactose bằng chai hoặc đưa vào thức ăn dạng khô trên cừu ăn khẩu phần lúa mạch (6).
Bổ sung Thức ăn cơ sở ăn
vào (g/ngày) Tổng lượng ăn vào
(g/ngày) Tăng trọng
(g/ngày) Không bổ sung 936 936 313 10% dung dịch lactose
828 936 325
20% dung dịch lactose
757 919 269
10% lactose khô 947 947 292 20% lactose khô 868 868 260

76
SE 50 50 18
Bảng 5.5 cho thấy lượng thức ăn ăn vào của khẩu phần cơ sở giảm gần như tương đương với lượng lactose ăn vào từ chai. Không phát hiện thấy sự khác nhau đáng kể về tăng trọng.
Công việc tiếp theo là đánh giá xem chất béo được cung cấp dưới dạng khô hoặc lỏng bằng chai có ảnh hưởng gì đến lượng thức ăn ăn vào và tỷ lệ tiêu hoá của khẩu phần cỏ khô hay không. Chất béo sử dụng ở đây là mỡ động vật trộn với một lượng nhỏ sữa bột đã tách bơ. Mức mỡ động vật sử dụng là 70 và 140 g/ngày (6). Kết quả được trình bày ở bảng 5.6.
Bảng 5.6. Ảnh hưởng của việc bổ sung mỡ đến lượng cỏ khô ăn vào, tổng năng lượng tiêu hoá (DE) ăn vào/kgW0.75 và tỷ lệ tiêu hoá của cỏ.
Mỡ bổ sung Chất khô ăn
vào (g/ngày) DE
(MJ/ngày)
Tỷ lệ tiêu hóa (%) g/ngày Phương pháp
cho ăn 0 - 827 10,2 68,0
70 Khô 732 10,2 64,6 140 Khô 676 10,8 60,8 70 Lỏng 814 11,2 67,1 140 Lỏng 768 12,1 64,9 SE 40 0,6 2,3 Chúng ta đã biết rằng khi mỡ vào dạ cỏ, mỡ làm giảm tiêu hoá và giảm
lượng thức ăn thô ăn vào. Do vậy, chúng ta trông đợi rằng nếu bổ sung mỡ vòa khẩu phần cơ bản là thức ăn thô thì tốt hơn là để mỡ thát qua dạ cỏ. Điều này được minh họa một cách rõ ràng trong bảng 6.
Tiếp theo chúng tôi xem xét ảnh hưởng của việc cho glucose hoặc acetate vào dạ múi khế của dê đang vắt sữa (7). Chúng tôi đã huấn luyện 3 con dê bú sữa từ chai lúc mới sinh và duy trì phản xạ đóng rãnh thực quản suốt thời kỳ mang thai và tiết sữa. Những dê này được cho ăn khẩu phần cơ bản là thức ăn tinh (70% thức ăn tinh) sau đó chúng tôi thay thế 21% thức ăn tinh với dung dịch acetate (pH5) bằng cách trộn vào thức ăn hoặc cho uống glucose bằng chai. Tỷ lệ mỡ sữa cho nhóm đối chứng, nhóm bổ sung acetate và glucose lần lượt là 4,3; 5,4 và 3,8%; và năng suất sữa hàng ngày là 2591, 2609 và 1964g. Rõ ràng, glucose cho ăn thông qua dạ múi khế đã làm giảm cả tỷ lệ mỡ sữa và năng suất sữa. Kết quả dĩ nhiên sẽ là như vậy nhưng giúp để khẳng định rằng ở dê cũng giống nhu ở bò sữa, mỡ sữa giảm khi một lượng lớn tiền chất glucose hoặc glucose được hấp thu.
Thí nghiệm cuối cùng tôi muốn báo cáo về những mối quan tâm liên quan đến việc cho ăn sucrose dạng khô hoặc dạng lỏng bằng chai. Cừu được cho ăn 1 kg cỏ khô hằng ngày với 250g sucrose dạng khô hoặc bú từ chai. Chúng tôi trông đợi rằng sucrose sẽ được tiêu hoá kém ở phần sau dạ cỏ bởi vì gia súc nhai lại

77
không có men phân giải sucrose trong ruột non nhưng kết quả trình bày trong bảng 5.7 minh họa rõ ràng ảnh hưởng của việc cho ăn đến lên men trong dạ cỏ hoặc manh ràng. Ở đây chúng tôi hợp tác với Stanley Mann từ Bộ môn vi khuẩn học của Viện. Cừu được giết thịt sau 10 ngày cho ăn đường (8).
Bảng 5.7. Ảnh hưởng của việc bổ sung sucrose một cách thường xuyên đến khả năng lên men trong dạ cỏ và ruột tịt.
pH Tổng số vi khuẩn
nhìn thấy x 108/ml Dạ cỏ Manh
tràng Dạ cỏ Ruột tịt
Cỏ 6,6 6,9 5,9 0,7 Cỏ + sucrose khô 6,4 6,6 43,4 0,5 Khẩu phần cơ sở + sucrose lỏng 6,6 5,5 13,7 18,9 SE 0,2 0,2
Đây chỉ là một thí nghiệm nhỏ nhưng ảnh hưởng của bổ sung sucrose một
cách thường xuyên đến dịch manh tràng là rất đáng kể, điều đó khẳng định rằng sucrose đã được lên men ở manh tràng – phân đương nhiên rất mềm.
Hướng nghiên cứu về phản xạ rãnh thực quản rất thú vị. Trước hết hướng nghiên cứu này cho thấy thực quản đóng lại bởi phản xạ có điều kiện, phản xạ có điều kiện này có thể duy trì cho đến khi trưởng thành nếu được củng cố liên tục, phản xạ này không tái lập ngay được sau một thời gian gián đoạn không sử dụng. Khả năng của cừu phân biệt được các điều kiện báo hiệu chúng sắp sửa nhận được thức ăn từ chai và các tín hiệu thí nghiệm có chai nhưng sẽ không có thức ăn cũng thú vị và nói lên một vài điều về mối quan hệ giữa người chăn nuôi và gia súc, các quan hệ này có thể đo đếm được. Không nghi ngờ gì nữa có nhiều phản xạ có điều kiện khác ảnh hưởng đến dinh dưỡng cho gia súc nhưng còn chưa được xác định, ví dụ những phản xạ ảnh hưởng đến tiết dịch của dạ dày.
Từ quan điểm kỹ thuật, rõ ràng là protein được cho vào sau dạ cỏ (dạ múi khế), khi gia súc có nhu cầu protein lớn hơn lượng protein do vi sinh vật cung cấp, được sử dụng tốt hơn so với protein được thẳng vào dạ cỏ bằng cách cho ăn thông thường. Cho ăn năng lượng bổ sung vào sau dạ cỏ có ít ảnh hưởng mặc dù mỡ được sử dụng sau dạ cỏ tốt nhất, đặc biệt trong thời kỳ vắt sữa.
Tại sao có ít hoặc không có sự khai thác phản xạ rãnh thực quản trong thực tế? Trước hết, tôi không bao giờ biện hộ cho việc sử dụng phản xạ rãnh thực quản như một phương pháp trong thực tế trong nuôi dưỡng gia súc nhai lại. Nó cần quá nhiều chi phí về lao động. Nếu áp dụng cho bò vắt sữa, phản xạ phải được duy trì bởi vì gia súc là bê bú sữa, bê làm cho việc sử dụng phản xạ này càng không thực tế hơn. Tuy nhiên, phản xạ rãnh thực quản chứng tỏ là một công cụ tuyệt vời để sử dụng trong các thí nghiệm phân biệt vai trò của dạ cỏ và ruột non trong dinh dưỡng gia súc nhai lại và cung cấp một giải pháp lớn trong việc hiểu biết các hạn

78
chế của tiêu hoá sau dạ dày kép. Tài liệu tham khảo cho phần 5.1
1. Ørskov, E.R. and Benzie, D. 1969. Studies on the oesophageal groove reflex in sheep and on the potential use of the groove to prevent the fermentation of food in the rumen. Br. J. Nutr. 23, 415-420.
2. Ørskov, E.R., Benzie, D. and Kay, R.N.B. 1970. The effect of feeding procedure on closure of the oesophageal groove in sheep. Br. J. Nutr. 24, 785-795.
3. Ørskov, E.R. and Fraser, C. 1969. The effect on nitrogen retention in lambs of feeding protein supplements direct to the abomasum. Comparison of liquid and dry feeding and of various sources of protein. J. Agri. Sci. Camb. 73, 469-475.
4. Ørskov, E.R., Fraser, C. and Corse, E.L. 1970. The effect on protein utilization of feeding different protein supplements via the rumen or via the abomasum in young growing sheep. Br. J. Nutr. 24, 803-809.
5. Ørskov, E.R., Fraser, C., Corse, E.L. 1971. The effect of protein supplementation via the abomasum on the voluntary intake of concentrate by young growing sheep. Proc. Nutr. Soc. 30, 25A, 1971.
6. Bailey, P.C. and Ørskov, E.R. 1974. The effect of ruminal or postruminal digestion of lactose or fat on the voluntary intake and digestibility of dried grass by lambs. Proc. Nutr. Soc. 33, 45A.
7. Ørskov, E.R. and Fraser, C. 1974. Effect of changes in the glucogenic energy on yield of milk and milk fat in lactating goats. Proc. Nutr. Soc. 33, 9A.
8. Mann, S.O. and Ørskov, E.R. 1973. The effect of rumen and post rumen feeding of carbohydrates on the caecal mocroflora of sheep. J. Appl. Bact. 36, 475-484.

79
Hướng nghiên cứu 5. Bảo vệ protein khỏi sự phân giải dạ cỏ Hướng nghiên cứu 5.2. Nuôi dưỡng nhân tạo
Tôi không bao giờ nghĩ trước khi tôi đến làm việc tại Viện Rowett rằng
nuôi dưỡng nhân tạo đang đi vào kế hoạch công việc của tôi. Tuy nhiên, có những sự kiện làm cho nuôi dưỡng nhân tạo hấp dẫn và quan trọng. Trước hết, nếu chăn nuôi cừu thâm canh được xem là một hệ thống chăn nuôi khi đó cừu sẽ phải được nuôi một cách nhân tạo. Tuy nhiên, động lực chính cho nuôi dưỡng nhân tạo thực tế là: bây giờ chúng ta đã hiểu biết về phản xạ rãnh thực và có thể sử dụng nó trong các thí nghiệm của mình. Bởi vì nó là một phản xạ có điều kiện và không phụ thuộc vào thành phần sữa, chúng ta có thể thoải mái thay đổi thành phần sữa thay thế trong một phạm vi rộng. Ở cừu chúng ta có thể sử dụng phản xạ để đưa trực tiếp dịch lỏng của một loạt các chất dinh dưỡng không phải sữa như mỡ, protein và carbohydrate vào dạ múi khế và nghiên cứu tiêu hoá và sử dụng các chất dinh dưỡng này. Chúng ta có thể tiếp tục nghiên cứu như vậy lâu chừng nào chúng ta muốn mà không ức chế sự phát triển bình thường của dạ cỏ.
Chúng tôi đã chứng minh rằng chúng tôi có thể tiến hành nghiên cứu kiểu này, vì vậy ngài Blaxter đã cho chúng tôi một số chuồng cừu để thiết lập hệ thống cho uống sữa tự động. McPherson đã lắp đặt hệ thống này một cách rất thông minh. Sữa được lưu thông bởi 4 bơm bó trí xung quanh khu 16 chuồng cừu, mỗi chuồng có thiết bị đếm thời gian. Thiết bị đếm thời gian có thể giúp chúng tôi cho cừu ăn 4 loại sữa thay thế khác nhau và cung cấp một số lượng sữa thay thế nhất định với bao nhiêu lần chúng tôi muốn. Ở mỗi thời gian được định trước một đầu vú sẽ xuất hiện từ một chiếc hộp. Khi motor bắt đầu hoạt động, tất cả cừu hăm hở quay đầu ra hộp, chờ đợi đầu vú xuất hiện. Cừu cũng có thể uống nước một cách tự do. Đó thực sự là hệ thống thú vị, nó nhận được sự tán đồng của hoàng tử Philip trong một chuyến thăm Viện.
Nhiệm vụ đầu tiên của chúng tôi là cố gắng thay thế casein bằng protein bột cá và mỡ sữa bằng hỗn hợp mỡ lợn và dầu dừa (1). Kết quả chính được trình bày trong Bảng 5.8. Ảnh hưởng của thay thế mỡ sữa bằng mỡ lợn đã chyển thành dạng lỏng và dầu dừa không đáng ngạc nhiên lắm vì đã được quan sát trên bê. Điều gây ngạc nhiên hơn là casein hoặc protein sữa có thể được thay thế bằng protein bột cá thuỷ phân (FPH), mặc dù thực tế không thấy sự vón cục nhưng sự không vón cục là kết quả của cho cừu ăn 3 giờ một lần. Bảng 5.8. Ảnh hưởng của thay thế protein sữa bằng protein bột cá và thay thế mỡ
sữa bằng mỡ lợn + dầu dừa trong sữa thay thế cho cừu. Nguồn protein và cabonhydrate
Nguồn mỡ Tăng trọng (g/ngày)
Tỷ lệ tiêu hoá (%)
Protein Mỡ Sữa tách bơ khô Bơ 156 95,9 98,7

80
Sữa tách bơ khô Mỡ lợn + dầu dừa
148 93,4 96,5
Protein bột cá + lactose Bơ 136 95,5 98,9 Protein bột cá + lactose Mỡ lợn + dầu
dừa 119 92,8 96,2
Tiếp theo chúng tôi thử thay thế lactose bằng tinh bột thuỷ phân và chúng
tôi khá ngạc nhiên là không có vấn đề gì xảy ra. Tăng trọng không bị ảnh hưởng và tiêu hoá của tinh bột gần như hoàn toàn (2) (Bảng 5.9). Bảng 5.9. Ảnh hưởng của việc thay thế lactose bằng tinh bột thuỷ phân trong sữa
thay thế cho cừu. Nguồn carbonhydrate Tăng trọng
(g/ngày) Chuyển hoá
thức ăn Tỷ lệ tiêu hoá tinh bột (%)
17% lactose của tổng số năng lượng thuần (GE)
172 1,02 -
17% tinh bột thuỷ phân của tổng số năng lượng thuần(GE)
182 1,05 99,8
Chúng tôi đã tự hỏi, chúng tôi có thể đi với tinh bột thuỷ phân xa đến đâu,
liệu chúng tôi cũng có thể thay thế một phần mỡ hay không? Vâng, chúng tôi có thể, nhưng có một giới hạn (2). Trong Bảng 5.10, có thể thấy rằng chúng tôi đã bắt đầu với tỷ lệ tinh bột thấp bằng 17% năng lượng trao đổi hoặc năng lượng tiêu hoá, tương đương với năng lượng của lactose trong sữa cừu. Chúng tôi sau đó tăng dần lượng tinh bột và dừng lại khi đạt 41% năng lượng thô, hay khoảng 50% khối lượng của sữa khô thay thế, chúng tôi đã vượt quá khả năng tiêu hoá tinh bột của cừu chưa. Có thể thấy rõ là chúng tôi đã vượt quá khả năng tiêu hoá tinh bột của cừu: giá trị pH của phân thấp đi, khả năng sản xuất của cừu giảm, và một số cừu bắt đầu ỉa chảy nặng. Tuy nhiên, tinh bột chiếm tới 25% tổng năng lượng hoặc 33% chất khô thì không có vấn đề gì, như vậy không chỉ lactose mà thậm chí một số mỡ có thể thay thế bằng tinh bột thuỷ phân.
Bảng 5.10. Ảnh hưởng của thay thế một phần mỡ bằng tinh bột thuỷ phân trong sữa thay thế cho cừu (2).
% tinh bột của tổng năng lượng 17 25 33 41 % mỡ của tổng năng lượng 55 47 39 31 Tăng trọng 0-15 ngày 158 148 147 103 Tăng trọng 16-33 ngày 219 204 204 185 Kg DM/kg tăng trọng 0-15 ngày 1,09 1,07 1,20 1,76 Kg DM/kg tăng trọng 16-33 ngày 0,97 1,09 1,15 1,27 pH của phân Tuần 3 7,0 6,8 4,5 4,0

81
pH của phân Tuần 5 8,5 8,5 6,0 4,8
Câu hỏi tiếp theo là: chúng ta có thể thay thế hoàn toàn các thành phần sữa bằng các thành phần không phải sữa? Điều này rất hấp dẫn. Chúng tôi bỏ sữa hoàn toàn và làm sữa thay thế từ FPH (bột cá thủy phân), mỡ lợn, dầu dừa và tinh bột thuỷ phân và thấy năng sản xuất của cừu tương tự như với sữa thay thế làm từ casein, lactose và mỡ sữa (3). (Bảng 5.11).
Bảng 5.11. Ảnh hưởng của việc thay thế lactose, mỡ sữa và casein bằng tinh bột thuỷ phân, mỡ lợn, dầu dừa và protein bột cá thuỷ phân cho cừu nuôi nhân tạo
trong 33 ngày. Nguồn protein Casein Casein Casein Protein bột
cá Nguồn mỡ Bơ Bơ Mỡ lợn và dầu
dừa Mỡ lợn và dầu dừa
Nguồn carbonhydrate Lactose Tinh bột Tinh bột Tinh bột Tăng trọng (g/ngày) 177 183 154 170 Tỷ lệ chuyển hoá thức ăn 1,02 1,06 1,08 1,00 Tỷ lệ tiêu hoá % Vật chất khô 97,8 96,4 96,4 95,4 Nitrogen 96,4 94,9 94,4 93,6 Mỡ 98,9 98,3 98,2 97,7 Tinh bột - 99,8 99,8 99,9
Tăng trọng và tỷ lệ tiêu hoá của các thành phần tương tự cho tất cả các
khẩu phần. Như vậy chúng ta có thể thay thế hoàn toàn các thành phần sữa bằng các
thành phần không phải sữa. Cuối cùng chúng tôi muốn thay đổi nguồn protein (3). Chúng ta có thể sử dụng men (Toprina) để thay thế casein không? Ở đây rõ ràng chúng tôi đã gặp vấn đề (Bảng 5.12). Khả năng sản xuất của cừu giảm một cách tuyến tính khi casein đợc thay thế bằng men protein. Bảng 5.12. Ảnh hưởng của hay thế casein bàng men (Toprina) trong sữa thay thế
cho cừu cai sữa sớm trong 33 ngày. % Casein 100 67 33 0 % Men 0 33 67 100 Tăng trọng g/ngày 130 127 98 89 Chuyển hoá thức ăn 1,26 1,27 1,57 1,58 Tỷ lệ tiêu hoá của protein % 94,6 90,6 86,9 79,5
Dường như nuôi cừu nhân tạo không kinh tế, chúng tôi đã học được điều đó

82
từ sự mạo hiểm. Hussein Soliman là người chăn nuôi tuyệt vời và luận văn nghiên cứu sinh của anh ta dựa trên công việc này. Chúng tôi nghĩ điều quan trọng là cừu đợc cho ăn 3 giờ một lần với một lượng nhỏ do đó quá trình vón cục làm trì hoãn sự dịch chuyển của các chất dinh dưỡng vào tá tràng được khắc phục bằng cách cho ăn thờng xuyên. Chúng tôi không mất một con cừu nào trong các thí nghiệm do đó khi kết thúc thí nghiệm cừu mạnh khoẻ và được cai sữa chỉ với thức ăn khô. Tài liệu tham khảo cho phần 5.2
1. Soliman, H. and Ørskov, E.R. 1976. utilization of fish protein hydrolysate for artificial rearing of lambs. Proc. Nutr. Soc. 35, 91A-92A.
2. Soliman, H., Ørskov, E.R., Atkinson, T. and Smart, R.I. 1979. Utilization of partially hydrolysed starch in milk replacers by newborn lambs. J. Agric. Sci., Camb. 92, 343-349.
3. Soliman, H., Ørskov, E.R. and Mackie, I. 1979. Utilization of fish protein hydrolysates in milk substitutes for lambs. J. Agric. Sci., Camb. 93, 37-46.

83
Hướng nghiên cứu 5. Bảo vệ protein khỏi sự phân giải dạ cỏ Hướng nghiên cứu 5.3. Bảo vệ protein khỏi sự phân giải trong dạ cỏ
Sau khi thiết lập được kỹ thuật nylon bag là phương pháp ước đoán tỷ lệ phân giải protein trong dạ cỏ, chúng tôi hăng hái thử nghiệm càng nhiều càng tốt các thức ăn protein bao gồm cả men protein Toprina mới của BP như đã đề cập trước đó. Chúng tôi sớm xác định được rằng protein trong bột cá có tỷ lệ phân giải tương đối thấp nhưng tỷ lệ phân giải lại bị ảnh hưởng nhiều bởi các điều kiện chế biến (1). Chúng tôi cũng quan sát thấy rằng bột máu hầu như không bị phân giải. Sau khi đã suy xét các lý do tại sao bột máu hầu như không bị phân giải tôi cảm thấy rằng tôi có thể sử dụng máu gia súc để bao xung quanh các nguồn protein khác nhằm bảo vệ chúng! Tôi nhờ Colin Fraser lấy mẫu máu từ lợn và phun chúng vào bột đậu tương và làm khô. Việc phun này hoàn toàn thành công (1). Đây là kết quả của thí nghiệm tiếp theo (Bảng 5.13)(1).
Bảng 5.13. Ảnh hưởng của máu khô bảo vệ thức ăn giầu protein đến N mất đi của
mẫu ủ trong túi nylon ở dạ cỏ. Thời gian ủ trong dạ cỏ
Bột cá Bột lạc 4 h 8 h 4 h 8h
Bột máu (lít/kg) Nitơ mất đi (%) 0,0 56,5 65,0 64,0 78,0 0,25 35,5 48,2 37,6 49,6 0,50 23,9 31,5 22,6 28,3 1,00 24,4 28,5 16,3 22,5 1,50 19,9 26,4 11,9 16,5 3,00 11,2 14,5 8,9 13,6
Protein thưc ăn đã được bảo vệ. Khi tôi trình bày kết quả với ngài Kenneth
Blaxter, ông rất thích thú và đề nghị cấp bằng sáng chế. Ông mời các vị lãnh đạo và quan chức của Cục sáng chế tới và thuyết phục họ về bằng sáng chế. Tôi thấy nghiên cứu này không phải là công việc của tôi nữa! Chúng tôi làm thêm các thí nghiệm chi tiết nữa nhằm thoả mãn yêu cầu của Cục sáng chế, nhưng cuối cùng chẳng có bằng sáng chế nào cả! Tôi làm thêm một số thí nghiệm nữa. Tôi nhận được một số protein từ sữa: whey từ Đan Mạch và tôi cũng thử nghiệm lòng trắng trứng để xem việc bảo vệ protein có phải là do albumin hay không, và quả đúng là như vậy.
Vài năm sau đó, thực tế rất gần đây vào năm 1998, chúng tôi lại một lần nữa phải giải quyết vấn đề bằng một cách rất hấp dẫn. Castro (2) đã phát triển phương pháp dùng hơi nước xử lý rơm lúa mạch và phát triển một phương pháp tách hemicellulose, cellulose và lignin. Lignin rất tinh khiết và ở đây chúng tôi bắt đầu phát triển một ý tưởng, ý tưởng này sau đó chính là dự án nghiên cứu cho

84
nghiên cứu sinh tiến sỹ Zahidifar (3). Chúng ta đã biết tannin sẽ làm kết tủa protein và đôi khi protein bị kết tủa có thể bị phân giải ở pH dạ cỏ thấp. Bởi vì lignin có chứa các thành phần axít phenolic tương tự như tannin, nên lignin có thể bảo vệ protein chăng? Dúng vậy lignin có thể bảo vệ protein bằng cách rất thân thiện với môi trường. Protein bị kết tủa bởi lignin bị phân giải ở pH thấp trong dạ múi khế.
Bảng 5.14. Ảnh hưởng của việc lignin thuỷ phân đến sinh khi in vitro, nồng độ iso
axít (C4-C5) và ammonia sau 48 giờ ủ.
Lignin thuỷ phân Khí sinh ra+ (ml)+
Iso axít (mM/lit)*
Ammonia (mg/lit)
0 44,3 3,8 330 100 44,9 2,3 190 200 44,5 1,7 149 400 43,1 1,5 101
+ cỏ khô * casein và bột cellulose
Một vài kết quả được trình bày trong bảng 5.14. Có thể thấy rằng lignin không ức chế hoạt động của vi sinh vật vì không thấy có thay đổi tổng lượng khí sinh ra, nhưng protein đã được bảo vệ: nồng độ ammonia giảm, nồng độ iso axít cũng giảm. nồng độ iso axít là chỉ thị về quá trình lên men của protein.
Sử dụng một phế phẩm như lignin để bảo vệ protein rất hấp dẫn và không bị phản đối vì có hại cho sức khoẻ và môi trường như trường hợp của formaldehyde, Tài liệu tham khảo cho phần 5.3
1. Ørskov, E.R., Mills, C.F. and Robinson, J.J. 1980. The use of whole blood for the protection of organic materials from degradation in the rumen. Proc. Nutr. Soc. 39, 60A.
2. Castro, F.B., Hotten, P.M. and Ørskov, E.R. 1993. The potential of dilute axít hydrolysis as a treatment of improving the nutritional quality of industrial lignocellulosic by-product. Anim. Fd. Sci. and Tech. 42, 39-53.
3. Zahidifar, M., Castro, F.B. and Ørskov, E.R. 2001. effect of hydrolytic lignin on formation of protein lignin complexes and protein degradation by rumen microbes. Anim. Fd. Sci. and Tech. (in press).

85
Hướng nghiên cứu 6. Tiêu hoá tinh bột, đường (glucose) và sucrose
Hướng nghiên cứu 6.1. Tiêu hoá carbonhydrate sau dạ cỏ
Như tôi đã đề cập trước đây, trong khi tôi làm việc ở Mỹ tôi bắt đầu quan tâm đến việc tìm hiểu xem ở đâu trong đường tiêu hoá các khẩu phần khác nhau đang được tiêu hoá. Ước đoán mức lên men dạ cỏ từ hoá học lượng pháp luôn luôn cho thấy lên men dạ cỏ thấp hơn mức mong đợi. Hơn nữa, các nhà khoa học Hoa kỳ đã báo cáo rằng các giá trị về tinh bột thoát qua ở dạ cỏ cao hơn. Tôi muốn theo đuổi hướng nghiên cứu này và ở đây sự hợp tác của Bộ môn sinh lý của Viện với tiến sĩ Robin Kay đã rất thành công. Ông lắp các canule ở dạ cỏ, dạ múi khế và cuối hồi tràng cừu lần đầu tôi tin là thế, và tháo ra khi chúng tôi đi. Ở thí nghiệm đầu tiên chúng tôi chỉ đơn giản muốn xem liệu cho ăn lúa mạch đã cán dẹp (lúa mạch “rolled” là lúa mạch khô, sau đó ép hay cán mỏng ra) hoặc hỗn hợp lúa mạch “rolled” và cỏ khô có quan trọng không trong việc xác định mức lên men trong dạ cỏ. Các kết quả từ nghiên cứu cân bằng lên men cho thấy rằng điều đó là có thể (1). Bảng 6.1 là kết quả quan sát lượng tinh bột đi vào dạ múi khế và đoạn cuối hồi tràng (2).
Bảng 6.1. Ảnh hưởng mức nuôi dưỡng và tỷ lệ cỏ khô/lúa mạch đến vị trí tiêu hoá
tinh bột.
Khẩu phần Tinh bột ăn vào (% khẩu phần) Dạ múi khế Đoạn cuối hồi tràng
100% lúa mạch ăn tự do 6,8 1,4 lúa mạch ăn tự do 70% 4,4 1,6 60% lúa mạch + 40% cỏ khô ăn tự do 12,8 5,1 lúa mạch ăn tự do 70% 7,7 4,3
Phần lớn tinh bột biến mất khỏi dạ cỏ. Mức nuôi dưỡng chỉ có ảnh hưởng
nhỏ đến sự mất đi của tinh bột ở dạ cỏ và nhiều tinh bột từ lúa mạch thoát qua sự phân giải ở dạ cỏ khi lúa mạch được trộn với thức ăn thô. Tôi đã mong đợi các ảnh hưởng ngược lại, vì thế tôi đã không tìm thấy ở đây sự xác nhận các tính toán của tôi về cân bằng lên men. Bên cạnh đó, lượng tinh bột thoát khỏi sự lên men dạ cỏ thấp hơn nhiều so với các giá trị được các nhà khoa học Hoa Kỳ báo cáo. Lúc bấy giờ chúng tôi đã biết rằng khi có thức ăn kích thước dài và ngắn trong dạ cỏ, thức ăn kích thước dài trong dạ cỏ có khuynh hướng làm tăng sự thoát ra khỏi dạ cỏ của thức ăn kích thước nhỏ. Tinh bột thoát qua từ dạ cỏ phần lớn biến mất trong ruột non. Tôi cũng muốn biết phải chăng tinh bột ở ngô bị đập vỡ ra được sử dụng trong các thí nghiệm cúa các nghiên cứu tại Hoa Kỳ được tiêu hoá theo một cách khác với tiêu hóa tinh bột ở lúa mạch “rolled”; và tôi cũng muốn biết việc chế biến ngô sẽ ảnh hưởng nào không (xem Bảng 6.2).

86
Bảng 6.2. Ảnh hưởng của khẩu phần và chế biến đến tỷ lệ tinh bột khẩu phần đén được dạ múi khế của cừu.
Khẩu phần Tinh bột đi tới dạ múi khế (%) Lúa mạch “rolled” 6,2 Ngô nghiền và dát mỏng 5,4 Ngô nghiền 12,1 Ngô mảnh (ngô đập vỡ) 14,2
Rõ ràng khẩu phần và quá trình chế biến có ảnh hưởng. Nhiều tinh bột ở ngô thoát qua phân giải dạ cỏ hơn tinh bột ở lúa mạch trừ khi ngô đã được sử lý hơi nước và dát mỏng (ngô nghiền và dát mỏng).
Chúng tôi tiến hành thêm thí nghiệm nữa với ngô để xem cho thêm muối trong khẩu phần có ảnh hưởng gì đến sự thoát ra khỏi dạ cỏ của tinh bột. Chúng tôi so sánh 3 khẩu phần, lúa mạch, ngô mảnh, và ngô mảnh với 2% muối ở trong một thiết kế thí nghiệm ô vuông La tinh 3x3 (3). Muối không ảnh hưởng đến sự thoát ra khỏi dạ cỏ của tinh bột; phần trăm tinh bột thoát qua phân giải dạ cỏ là 8,8 với lúa mạch và 23% với ngô, kết quả này lớn hơn so với kết quả trước đó nhưng là do có một cừu cho kết quả khác thường. Ở cừu này 42% tinh bột thoát qua phân giải dạ cỏ trong khi ở cừu khác chỉ có 7% thoát qua. Điều này dĩ nhiên đã có ảnh hưởng rõ rệt đến tổng số tinh bột được tiêu hoá ở ruột non, 324 g ở một cừu và 21g ở cừu khác. Ở một cừu đã thấy 78g tinh bột đi tới đoạn cuối của hồi tràng, trong đó 57 g mất đi ở ruột già, phần còn lại được đào thải qua phân; ở cừu khác không có tinh bột đi đoạn cuối của hồi tràng. Tôi sẽ thảo luận việc ứng dụng quá trình lên men ở ruột già trong phần tiếp theo.
Chúng tôi sau đó đặt câu hỏi khả năng tiêu hoá tinh bột ở ruột non của cừu ra sao và enzyme nào đang hạn chế tiêu hoá tinh bột ở ruột non? Câu hỏi này là nội dung của luận án thạc sĩ của Bob Mayes (4).
Cừu ở các lứa tuổi khác nhau và cừu trưởng thành được đưa vào thí nghiệm. Một tinh bột lỏng đã hồ hóa được bơm vào cannula dạ múi khế với mức trung bình khoảng 300g/ngày. Trong số 300g đó, 150 đến 200g biến mất trong ruột non cho thấy cừu 40 kg có khả năng tiêu hóa khoảng 200g tinh bột/. Tuổi có ít hoặc không có ảnh hưởng đến tiêu hóa tinh bột và không có dấu hiệu thích nghi từ 10 ngày đầu đến 10 ngày cuối của 28 ngày thí nghiệm. Điều có lẽ thú vị nhất là thành phần alfa-glucoside của carbonhydrate đi tới đoạn cuối hồi tràng; xem bảng 6.3 (4). Bảng 6.3. Phân bố các phần glucose trong tổng số alfa-glucoside trong dịch hồi tràng g/kg.
Glucose Maltose Maltotriose Maltotetrose Oligosaccharides cao phân tử hơn
Tinh bột
121 174 168 183 197 223

87
Các kết quả này cho thấy: rất ít alfa-glucose từ tinh bột hồ hóa có măt như là đại diện của tinh bột tại đoạn cuối hồi tràng; trong khi các loại saccharide mạch ngắn lại có mặt ở đây; và amylase có thể không chỉ là nhân tố duy nhất hạn chế tiêu hoá tinh bột đã hồ hóa trong ruột non. Có quan hệ tuyến tính trực tiếp giữa pH hồi tràng và nồng độ alfa-glucoside cho thấy một vài thành phần của tinh bột bị lên men trong đoạn cuối hồi tràng. Nếu sử dụng tinh bột thô (chưa hồ hóa), hầu như chắc chắn tinh bột sẽ được tiêu hóa ít hơn ở đoạn ruột này.
Như vậy, dường như khả năng tiêu hoá tinh bột trong ruột non của cừu khá hạn chế. Một hạn chế nữa của ruột non ở cừu là khả năng hấp thu glucose. Chúng tôi đã nghiên cứu khả năng này bằng cách liên tục đưa glucose vào dạ múi khế với số lượng tăng từ 0 đến 500g/ngày ở cừu nặng khoảng 50kg. Khi hơn 300g glucose đưa vào mỗi ngày, glucose bắt đầu xuất hiện ở ruột già và khi hơn 400g/ngày được đưa vào, glucose bắt đầu xuất hiện trong phân. Việc liên tục đưa glucose vào được coi là cách tạo điều kiện tối ưu cho quá trình hấp thu. Với việc đưa glucose không liên tục vào ruột non bằng cách nuôi dưỡng bằng chai và sử dụng phản xạ rãnh thực quản, khả năng tiêu hóa tinh bột ở ruột non qua sát được là rất nhỏ (5).
Khả năng tiêu hoá sucrose cũng được thử nghiệm (6) bằng cả hai cách: sử dụng phản xạ rãnh thực quản và liên tục đưa sucrose vào dạ múi khế. Khi đưa sucrose liên tục vào dạ múi khế đạt mức trên 150g sucrose /ngày, phân bắt đầu rất mềm. Khoảng 30 đến 40% (của 150g sucrose) biến mất trước khi tới đoạn cuối hồi tràng, nhưng sự biến mất này có thể một it do quá trình lên men hoặc do không có hoạt động của enzyme sucrase trong dịch nhầy ruột non của cừu được cung cấp sucrose bằng chai trong 36 ngày.
Dường như khả năng tiêu hoá tinh bột trong ruột non của cừu khá hạn chế phần nào, và khả năng hấp thu glucose của ruột non cũng rất thấp so với nhu cầu năng lượng cho duy trì của chúng. Cừu không có khả năng tiêu hoá sucrose bằng enzyme. Tài liệu tham khảo cho phần 6.1
1. Ørskov, E.R., Flatt, W.P. and Moe, P.W. 1968. A fermentation balance approach to estimate of fermentation and efficiency of volatile fatty axít formation in ruminants. J. Dairy Sci. 51, 14-29.
2. Ørskov, E.R., Fraser, C. and Kay, R.N.B. 1969. Dietary factors influencing the digestion of starch in the rumen and small and large intestine of early weaned lambs. Br. J. Nutr. 23, 217-226.
3. Ørskov, E.R., Fraser, C. and McDonald, I. 1971. Digestion of concentrate in sheep. 3. Effect of rumen fermentation of barley and maize diets on protein digestion. Br. J. Nutr. 26, 477-486.
4. Mayes, R.W. and Ørskov, E.R. 1974. The utilization of gelled maize starch in the small intestine of sheep. Br. J. Nutr. 32, 143-153.
5. Ørskov, E.R., Mayes, R.W. and Penn, A. 1971. The capacity for removal of glucose from the intestine in mature sheep. Proc. Nutr. Soc. 30, 43A-44A.
6. Ørskov, E.R., Mayes, R.W. and Mann, S.O. 1972. Post ruminal digestion of

88
sucrose in sheep. Br. J. Nutr. 28, 425.

89
Hướng nghiên cứu 6. Tiêu hoá glucose, tinh bột và sucrose Hướng nghiên cứu 6. 2. Các nghiên cứu lên men tinh bột trong ruột già và ứng dụng trong xác định tỷ lệ tiêu hoá nitơ biểu
kiến.
Tôi đã không thực sự có ý định khám phá chức năng tiêu hoá của ruột già nhưng khi chúng tôi thực hiện một thí nghiệm khá lớn về vị trí tiêu hoá tinh bột (1) chúng tôi đã đo luợng tinh bột tới được đoạn cuối hồi tràng, nhưng đặc biệt hơn chúng tôi đã đặt cannula ở phần ruột tịt ở một số cừu do đó chúng tôi có thể lấy được mẫu phân và đo được tỷ lệ của các axít béo bay hơi.
Sự khác nhau đã xuất hiện khi chúng tôi so sánh các khẩu phần ngô và lúa mạch. Bảng 6.4 trình bày kết quả quan sát khi một cừu được cho ăn lúa mạch hoặc ngô làm vỡ thành mảnh. Rõ ràng tinh bột ảnh hưởng đến loại hình lên men ở ruột tịt. Bảng 6.4. Ảnh hưởng tỷ lệ tinh bột trong dạ múi khế đến loại hình lên men ở ruột tịt (Mol %). Khẩu phần Tinh bột trong
dạ múi khế (% vật chất
khô)
Axít acetic (%)
Axít propionic (%)
Axít butyric (%)
Lúa mạch 4,1 78,1 10,2 5,8 Ngô 20,1 57,1 12,4 19,4
Thí nghiệm tiếp theo chúng tôi thực hiện nhờ có một chút cơ hội. Hai cừu
đã được gắn cannula hồi tràng và manh tràng có sẵn ở bộ môn Sinh lý của Robin Ray, tôi thì có một sinh viên mùa hè, Meryl Foot, và một cái bơm truyền! Chúng tôi bơm tinh bột thô vào ruột già tăng dần từ 20g/ngày cho đến khi đạt 300g/ngày. Chúng tôi phân tích phân để xác định tinh bột và như thường lệ, phân tích cả nitơ. Khi cho khoảng 140g tinh bột thô/ngày vào, gần như tất cả tinh bột được lên men nhng với các mức cao hơn phần lớn phần tinh bột tăng lên xuất hiện trong phân. Tuy nhiên phần hấp dẫn nhất của thí nghiệm nhỏ đến lúc này là khi đưa tinh bột vào chúng tôi thấy có nitơ đi ra; nitơ trong phân tăng từ 5,8 đến 9,6g/ngày. Ảnh hưởng của bơm tinh bột đến tỷ lệ tiêu hóa nitơ biểu kiến (Cừu ăn khẩu phần cơ bản 900g cỏ ép viên hàng ngày) là làm cho tỷ lệ này giảm từ 69,3 xuống 48,7% đơn giản đây là kết quả của việc tăng quá trình lên men trong ruột già. Tôi không bao giờ nghĩ rằng chúng tôi lại có thể, trong thực tế, dóng lên hồi chuông khai tử cho việc sử dụng tỷ lệ tiêu hoá nitơ biểu kiến ở gia súc nhai lại. Chúng tôi đã làm giảm tỷ lệ tiêu hóa nitơ biểu kiến rất mạnh chỉ đơn giản bàng cách thay đổi vị trí tiêu hoá tinh bột. Khi nhìn lại, tôi nghĩ rằng tôi nên biết là tinh bột lên men trong ruột già chắc chắn dẫn tới bài tiết sinh khối vi sinh trong phân vì tiêu hoá vi sinh

90
vật trong ruột già tất nhỏ ở gia súc không ăn phân (non-coprophagic animals). Tuy nhiên, tôi có một vấn đề nho nhỏ. Bạn tôi, Victor Mason, đã tham gia
với Viện Rowett để xác định một cách chi tiết nguồn gốc và thành phần nitơ của phân và ở đây tôi đang đi vào lãnh địa của anh ta! Tôi vô tội - Tôi không có ý định làm như vậy! Chúng tôi quyết định hợp tác và viết một bài báo về bằng cách nào vị trí tiêu hoá đã có ảnh hưởng khác nhau đến nitơ trong phân (2). Nếu tinh bột được đưa vào dạ cỏ, tăng nitơ phân bao gồm vách tế bào vi sinh vật không tiêu hoá được trong khi nếu tinh bột lên men trong ruột già tăng nitơ phân bao gồm toàn bộ sinh khối vi sinh vật. Kết quả này được khẳng định nhờ các kỹ thuật phân tách của tiến sĩ Mason. Chúng tôi cũng khẳng định ảnh hưởng của vị trí tiêu hóa đến các sản phẩm lên men manh tràng và thành phần các axít béo bay hơi trong dịch phân (xem Bảng 2). Bảng 6.5. Ảnh hưởng của bơm tinh bột vào đoạn cuối ruột tịt đến các sản phẩm lên men của phân và manh tràng (Mol %).
Vị trí Tinh bột (g/ngày)
Axít acetic (%)
Axít propionic
(%)
Axít butyric (%)
Các axít phân tủe
lượng cao hơn (%)
Ruột tịt 0 71,6 11,6 5,7 11,1 300 63,4 10,2 17,7 8,7
Phân 0 68,8 17,5 6,2 7,5 300 66,5 6,9 18,9 7,7
Tỷ lệ axít butyric, đặc biệt, tăng cả ở chất chứa manh tràng và cả trong
phân.do lên men tinh bột. Ảnh hưởng đến tỷ lệ tiêu hoá biểu kiến nitơ được khẳng định thêm trong
một thí nghiệm khác trong đó cừu ăn ngô làm vỡ thành mảnh. Ở một cừu, một lượng lớn tinh bột thoát qua phân giải dạ cỏ do đó 60gtinh bột/ngày biến mất trong ruột già. Tỷ lệ tiêu hoá nitơ biểu kiến là 49% (3). Ở cừu khác, có ít hoặc không có tinh bột được lên men trong ruột già và tỷ lệ tiêu hoá nitơ biểu kiến là 75%.
Nếu sucrose được cho ăn bằng chai do đó sucrose thoát qua dạ cỏ, vi sinh vật manh tràng tăng đáng kể khi vì chỉ có một ít sucrose ra khỏi ruột non (4). Cuối cùng chúng tôi thấy rằng khi tăng lượng glucose bơm vào trong dạ múi khế (5) có tới khoảng 300g/ngày mât đi từ ruột non, khoảng 100g/ngày biến mất từ ruột già, và vượt quá 400g/ngày, một phần glucose sẽ vào phân, làm phân trở nên rất lỏng với pH nhỏ hơn 5.
Kết luận, sự đột nhập của chúng tôi vào quá trình tiêu hoá ruột già cho thấy khả năng lên men tinh bột và đường trong ruột già rất hạn chế, khoảng 2g/kg khối lợng sống. Một thay đổi nhỏ trong qui mô lên men ở ruột già có thể ảnh hưởng đáng kể đến tỷ lệ tiêu hoá protein biểu kiến, vì vậy các hệ thống nuôi dưỡng dựa trên các nhu cầu protein thô tiêu hoá biểu kiến không còn đứng vững được nữa. Về

91
phương diện này các thí nghiệm đã giúp hướng chúng tôi đến khái niệm mới về sử dụng nitơ và giúp hiểu ảnh hưởng qua lại giữa gia súc - vật chủ gia súc với hệ vi sinh vật trong nó. Bất cứ khi nào quá trình lên men trong ruột già tăng lên, lượng nước của phân tăng dẫn đến sự khử nước. Ngược lại khi gia súc nhai lại có dấu hiệu phân mềm, thường là tỷ lệ tiêu hoá nitơ biểu liến quá thấp, nhưng hoàn toàn tỷ lệ tiêu hoá thực của nitơ ở đây chẳng nói lên điều gì.
Tài liệu tham khảo cho phần 6.2 1. Ørskov, E.R., Fraser, C. and Kay, R.N.B. 1969. Dietary factors influencing
the digestion of starch in the rumen and small and large intestine of early weaned lambs. Br. J. Nutr. 23, 217-226.
2. Ørskov, E.R., Fraser, C., Masson, V.C. and Mann, S.O. 1970. The influence of starch digestion in the large intestine of sheep on caecel fermentation, caecal microflora and faecal nitrogen excretion. Br. J. Nutr. 24, 671-682.
3. Ørskov, E.R., Fraser, C. and McDonald, I. 1971. Digestion of concentrate in sheep. 3. Effect of rumen fermentation of barley and maize diets on protein digestion. Br. J. Nutr. 26, 477-486.
4. Ørskov, E.R., Mayes, R.W. and Mann, S.O. 1972. Post ruminal digestion of sucrose in sheep. Br. J. Nutr. 28, 425.
5. Ørskov, E.R., Mayes, R.W. and Penn, A. 1971. The capacity for removal of glucose from the intestine in mature sheep. Proc. Nutr. Soc. 30, 43A-44A.

92
Hướng nghiên cứu 7. Chế biến ngũ cốc Hướng nghiên cứu 7. 1. Mỡ mềm và axít béo mạch nhánh
Tôi đã đề cập ở kết luận của thí nghiệm cừu thâm canh đầu tiên (Hướng nghiên cứu 1.1) rằng có một vấn đề: mỡ thân thịt của cừu mềm, thậm chí cả khi thân thịt đã được ướp lạnh. Điều này làm người bán thịt không thích chúng, vì không thể cắt để có miếng sườn cừu thịt rắn đẹp mắt. Khách hàng sẽ không thích miếng sườn cừu thịt mềm và nhớt. Tôi chắc rằng các axít béo chưa bão hòa đã gây nên hiện tượng trên vì như đã biết axít béo chưa bão hòa đông đặc ở nhiệt độ thấp hơn so với nhiệt đô đông đặc của axít béo bão hòa. Tôi mang mẫu mỡ tới Bộ môn sinh hóa chất béo và yêu cầu giúp đỡ. Trưởng Bộ môn sinh hóa chất béo sau đó nói rằng ông ta có nhiều việc quan trọng hơn để làm thay vì vội vã và loanh quanh thùng hoá chất; và thế là mẫu được cho vào cất giữ. Tuy nhiên, lén lút một chút, khi Trưởng Bộ môn sinh hóa chất béo đang nghỉ phép, tôi thuyết phục nhân viên kỹ thuật cao cấp phân tích một hoặc hai mẫu. Kết quả rất đáng kinh ngạc. Vấn đề không phải là do các axít béo chưa bão hòa, trong trường hợp này chúng ta vào thời điểm đó (nhưng không nhiều hy vọng) phải đảm báo với người tiêu dùng rằng thịt cừu như thế tốt cho sức khoẻ của họ. Thực ra, mẫu có rất nhiều các axít béo mạch lẻ và mạch nhánh, như trình bày trong Bảng 7.1 (1). Bảng 7.1. Tổng thành phần axít béo (mol/100 mol) của mỡ thân thịt cừu ăn lúa mạch cán dẹp và cỏ đóng viên. Axít béo Mạch lẻ
Mạch nhánh
Khẩu phần 14:0 16:0 16:1 18:0 18:1 18:2 15:0 + 17:0
Lúa mạch ”rolled”
4 22 2 7 40 4 7 12
Cỏ đóng khối 3 22 3 12 47 5 4 2
Các axít béo mạch nhánh, giống như các axít béo chưa bão hòa, trở nên rắn ở nhiệt độ thấp hơn và hàm lượng cao của chúng là nguyên nhân làm mỡ mềm. Chúng tôi chắc chắn không tranh luận axít béo mạch nhánh có tốt cho người tiêu dùng không, thực tế chúng tôi không chắc chắn rằng axít béo mạch nhánh không tốt cho người tiêu dùng. Các mạch CH3 nằm ở các điểm khác nhau dọc theo các phân tử axít béo. Khi Trưởng Bộ môn sinh hóa chất béo trở về ông ta đã rất kích động. Tôi được đề nghị không đề cập đến các axít béo mạch nhánh với bất cứ ai hoặc không viết về nó. Các nhà hoá học chất béo đã hăng hái xác định được hơn 100 axít béo mới. Axít béo trở thành chủ đề chính của họ trong một số năm và mang lại cho họ một số giải thưởng. Tuy nhiên sự khám phá axít béo mạch nhánh không giải quyết được vấn đề mỡ thân thịt. Chúng tôi phải tìm ra nguyên nhân, và

93
ở đây tôi đã có được sự trợ giúp lớn từ tiến sĩ Ted Weekes, một nhà sinh hoá tài giỏi. Chúng tôi ngồi và suy nghĩ về trao đổi chất của các axít béo bay hơi khác nhau được tạo ra ở dạ cỏ và kết luận rằng axít propionic chắc là thủ phạm. Axít này được sản xuất rất nhiều ở các khẩu phần thức ăn tinh. Nếu trao đổi chất axít propionic vượt quá khả năng của gan, axít propionic sẽ đi vào tuần hoàn ngoại biên và có thể cung cấp đơn vị hóa học đầu tiên cho tổng hợp mỡ và vì thế có một lượng axít béo C15 và C17 bất thường (xem Bảng 7.1). Nhưng các axít béo mạch nhánh thì sao? Quá trình trao đổi chất của axít propionic như sau:
Axít propionic axít methylmalonic axít succinic
Axít succinic dĩ nhiên là một thành phần chính trong vòng Krebs, nó cho
phép axít propionic đuwợc chuyển hoá thành glucose. Sự chuyển hoá axít methylmalonic thành axít succinic phụ thuộc vào vitamin B12 như một đồng nhân tố và chúng tôi ngay lập tức nghĩ rằng chúng tôi phải nghiên cứu sự thiếu hụt B12 hoặc cyanocobalamine. Sự thiếu hụt này có thể dẫn đến sự tích luỹ axít methylmalonic, axít methylmalonic đến lượt mình có thể cạnh tranh với axít malonic thông thờng để cho đi các đơn vị acetyl giúp kéo dài mạch axít béo. Nếu sự việc trên xảy ra một nhóm CH3 sẽ bị bị cho đi tại cùng thời gian đó và do đó tạo ta các axít béo phân nhánh.
Dường như có là hai hướng nghiên cứu có thể: hoặc nghiên cứu ảnh hưởng của axít propionic hoặc tìm xem có vấn đề gì với vitamin B12 hay không. Nghiên cứu do một sinh viên thạc sĩ ở Khoa Sinh lý đã cho thấy rằng lượng B12 được tổng hợp trong dạ cỏ ở khẩu phần cơ bản là lúa mạch ít hơn nhiều so với lượng B12 được tổng hợp trong dạ cỏ ở khẩu phần là cỏ khô. Đầu tiên, chúng tôi cố gắng kiểm tra cobalt trong khẩu phần có phải là yếu tố giới hạn không vì cobalt tạo nên lõi của phân tử cyanocobalamine. Chúng tôi cho cừu ăn một trong bốn khẩu phần cơ sở là lúa mạch: khẩu phần cơ sở là lúa mạch không bổ xung cobalt, khẩu phần cơ sở với cobalt bình thường, khẩu phần cơ sở với mức cobalt x 10, khẩu phần cơ sở với B12 hai lần một tuần (2).
Kết quả cho thấy rằng bổ sung cobalt và B12 không tạo nên sự khác biệt nào về tổng số axít béo mạch nhánh hay mạch lẻ trong mỡ, như vậy cobalt cũng như vitamin B12 không phải là vấn đề. Sau đó tôi tập trung chú ý sang chế biến ngũ cốc như là cách có thể nhằm giảm axít propionic bằng cách so sánh các thức ăn viên làm từ lúa mạch đã nghiền và cốn lại với với thức ăn viên làm từ lúa mạch nguyên hạt. Lúc đầu chúng tôi ngạc nhiên khi phát hiện ra rằng không có sự khác biệt giữa các khẩu phần trong thành phần axít béo bay hơi dạ cỏ, hay pH dạ cỏ, kết quả là, mỡ vẫn rắn. Tuy nhiên, lúa mạch nguyên hạt đã bị nghiền trong quá trình chế biến viên, ý tưởng nẩy ra từ đây là: sẽ rất hấp dẫn để xem cái gì sẽ xảy ra nếu lúa mạch không bị chế biến.
Có hai lo lắng ở đây: liệu cừu có ăn lúa mạch nguyên hạt? và nếu như chúng ăn thì tỷ lệ tiêu hoá sẽ như thế nào? Kết quả là tốt, lượng ăn vào tương tự như lượng ăn vào của lúa mạch viên. Tỷ lệ tiêu hoá của lúa mạch nguyên hạt là

94
78,0% và lúa mạch viên là 76,7% và không có sự khác nhau đáng kể. Chúng tôi khá kích động với kết quả này và suy nghĩ rất nhiều về axít béo bay hơi vì sẽ có những thuận lợi thực tế lớn lao khi cho cừu ăn lúa mạch nguyên hạt thay vì cho chúng ăn lúa mạch viên. Khi cừu đạt 40kg chúng được mổ thịt, nhưng trước đó chúng tôi lấy mẫu dạ cỏ và thấy rằng dịch dạ cỏ lỏng hơn nhiều và dễ lấy hơn từ cừu ăn lúa mạch nguyên hạt, một điều quan trọng hơn pH dạ cỏ của cừu ăn lúa mạch nguyên hạt là 6,2 trong khi pH dạ cỏ của cừu ăn lúa mạch viên là 5,2. Tôi biết sau đó chúng tôi đã nhận được nhiều hơn chúng tôi mong đợi. Gần như chắc chắn axít béo bay hơi sẽ khác nhưng trước tiên chúng tôi yêu cầu chuyên gia về thịt của chúng tôi đến lò mổ và phân loại thân thịt theo độ rắn mỡ, chuyên gia này không được biết về khẩu phần đã nuôi cừu. Tất cả cừu ăn ngũ cốc nguyên hạt có độ rắn của mỡ ở mức bình thường. Kết quả được trình bày trong bảng 7.2, được củng cố thêm sau đó khi axít béo bay hơi được phân tích (2). Bảng 7.2. Ảnh hưởng của nuôi dưỡng bằng lúa mạch nguyên hạt hoặc lúa mạch viên đến pH dạ cỏ, thành phần axít béo bay hơi (mol %), độ rắn mỡ thân thịt và khối lượng gan. Axít béo bay hơi dạ cỏ Lúa mạch
pH dạ cỏ
C2 C3 IC4 C4 Cao hơn
Khối lượng
gan (g)
Độ rắn
mỡ* Nguyên hạt
6,2 53 34 2 7 4 703 2,0
Viên 5,2 39 44 1 12 4 867 3,6 SE 0,1 2,6 2,2 0,3 2,4 0,4 41 0,4
*4 rất mềm và 1 rất cứng
Tôi chắc rằng người đọc có thể nhìn thấy rằng kết quả này quá hấp dẫn, trong một thí nghiệm nhỏ chúng tôi đã mở ra một lĩnh vực nghiên cứu hoàn toàn mới. Chế biến ngũ cốc trở thành một chủ đề sống động! Chúng tôi có thể đã có khả năng tiết kiệm cho ngành công nghiệp cừu nước Anh vài triệu bảng mỗi năm, và trong cùng một thời điểm có thể làm tăng sức khoẻ gia súc bằng cách giảm bệnh viêm dạ cỏ, và cải thiện chất lượng thân thịt. Câu hỏi hiển nhiên hiện ra trong đầu tôi là tại sao lại cho gia súc nhai lại nhỏ ăn ngũ cốc đã chế biến? Câu trả lời là cách đây lâu rồi người ta không đủ khả năng để sàng lọc các loại hạt cỏ dại có vỏ cứng và các hạt cứng thoát qua trực tiếp ruột và trải khắp các cánh đồng cùng với phân. Khi sàng lọc được cải thiện, chế biến đã trở thành một thực hành phổ biến, không có gì phải bàn cãi. Một quan sát khác gây tranh luận ủng hộ chế biến thức ăn viên là một số ngũ cốc nguyên hạt xuất hiện trong phân và nông dân thường giả thiết phần lớn thức ăn thoát qua ruột mà không được tiêu hoá khi họ nhìn thấy một hoặc hai hạt ngũ cốc còn nguyên hạt trong phân. Trong thực tế chúng tôi thấy điều này không phải như vậy. Tôi sẽ thảo luận việc chế biến hạt ngũ cốc một cách chi

95
tiết ở hướng nghiên cứu tiếp theo (7.3). Khi các axít béo phân nhánh được quan tâm chúng tôi thực hiện thêm 3 thí
nghiệm nữa. Trong một thí nghiệm chúng tôi xác định có hay không các loại ngũ cốc khác cho giá trị pH dạ cỏ và axít béo bay hơi tương đơng như các giá trị đã thấy ở khẩu phần lúa mạch. Các điểm chính được trình bày trong Bảng 7.3. Ảnh hưởng của tạo viên đến pH dạ cỏ và axít béo bay hơi giống nhau với tất cả các ngũ cốc đợc kiểm tra (2). Bảng 7.3. Ảnh hưởng của cho ăn lúa mạch, ngô, kiều mạch hoặc lúa mỳ dạng viên (P) hoặc nguyên hạt (L) đến pH dạ cỏ và thành phần axít acetic và propionic của cừu Ngũ cốc pH dạ cỏ Axít acetic Axít propionic Lúa mạch L 6,4 52,5 30,1 P 5,4 45,0 45,3 Ngô L 6,1 47,2 38,7 P 5,2 41,3 43,2 Kiều mạch L 6,7 65,0 18,6 P 6,1 53,2 37,5 Lúa mỳ L 5,9 52,3 32,2 P 5,0 34,2 42,6 SE 0,14 2,4 3,2
Thí nghiệm thứ hai được thực hiện trên dê để xem nếu như dê cũng sản
sinh các axít béo mạch nhánh khi được nuôi với các khẩu phần lúa mạch, dù ở bò điều này chúng tôi đã thấy không xẩy ra (3). Kết quả cho thấy rõ ràng là về phương diện này dê giống cừu. Dê non được vỗ béo bằng khẩu phần lúa mạch viên có 110mg/g axít béo mạch nhánh trên tổng axít beo các mô dưới da trong khi dê được ăn khẩu phần thức ăn thô chỉ có 30mg/g.
Cuối cùng một thí nghiệm nhỏ được thực hiện với cừu, cừu được cho ăn lúa mạch nguyên hạt, kiều mạch nguyên hạt hoặc các hỗn hợp trộn khác nhau của hai loại (4). Trong khi lúa mạch nguyên hạt hoàn toàn không loại các axít béo mạch nhánh và mạch lẻ như đã quan sát thấy ở lúa mạch viên, cho cừu ăn kiều mạch nguyên hạt cho kết quả tương tự như kết quả cho dê ăn khẩu phần thức ăn thô và cũng tạo ra mỡ dưới da bình thường. Trong Bảng 7.4 chỉ trình bày kết quả cho lúa mạch nguyên hạt và kiều mạch nguyên hạt vì các khẩu phần có tỷ lệ thành phần khác nhau cho kết quả trung gian như mong đợi. Bảng 7.4. Ảnh hưởng của việc nuôi dưỡng lúa mạch nguyên hoặc kiều mạch nguyên hạt cho cừu đến lên men dạ cỏ và mỡ thân thịt. pH dạ cỏ Axít acetic Axít
propionic Độ rắn của
mỡ*

96
Lúa mạch nguyên hạt 6,5 48,5 34,4 2,6 Kiều mạch nguyên hạt 6,8 59,5 22,6 1,0 SE 0,12 2,3 2,4 0,2
*4 = rất mềm 1= rất rắn
Chúng tôi cũng thấy rằng độ rắn của mỡ có tương quan âm với tỷ lệ axít
propionic tính bằng mol (r = 0,65). Nói cách khác kiều mạch nguyên hạt giống như thức ăn thô. Những vấn đề
thực tế về mỡ mềm và các axít béo mạch nhánh ở các khẩu phần hạt ngũ cốc như vậy là đã được giải quyết bằng cách phôí hợp các khẩu phần rẻ tiền hơn, nhưng vẫn còn có những câu hỏi: việc tạo mỡ mềm hoặc rắn đã xẩy ra như thế nào. Liệu vitamin B12, có một vai trò nào không và tại sao việc tạo mỡ mềm hoặc rắn không xảy ra ở bò ăn các khẩu phần tương tự, sản sinh một lượng axít béo dạ cỏ có thành phần tương tự?
Trong 15 năm vấn đề nêu trên đã không được ai nghiên cứu. Tôi tin rằng ít nhất một phần là vì đây không còn là vấn đề thực hành nữa. Tuy nhiên, một khi chúng tôi phát triển kỹ thuật xong kỹ thuật dinh dưỡng trực tiếp cho dạ dày (hướng nghiên cứu 8.2) chúng tôi có khả nănh kiểm soát hoàn toàn axít béo bay hơi dạ cỏ. Bây giờ chúng ta có thể hỏi ở tỷ lệ axít propionic nào bò sẽ bắt đầu sản sinh các axít béo mạch nhánh? Chúng tôi đã tăng lượng bơm vào dạ cỏ tới 70% phần trăm axít propionic (mol) nhưng không bao giờ tìm thấy điều gì! Do đó chúng tôi quyết định kiểm tra cừu một lần nữa để định nghĩa một cách chính xác hơn tỷ lệ axít propionic (mol), mà cao hơn tỷ lệ này bắt đầu có sự tích luỹ axít methylmalonic và có sản xuất các axít mạch nhánh. Ở đây chúng tôi đã gặp chút khó khăn, vì bây giờ chúng tôi không thể tạo ra sự ảnh hưởng này! chúng tôi bơm vào dạ cỏ tới 70% phần trăm axít propionic (mol) với tốc độ bơm lớn, nhưng không có dấu hiệu của các axít béo mạch nhánh, không có dấu hiệu của axít methylmalonic.
Giải pháp như chúng ta có thể thấy là khi nuôi cừu bằng các khẩu phần cơ bản là hạt ngũ cốc, một lượng lớn các đồng phân của vitamin B12 được tạo ra trong dạ cỏ và được hấp thu. Các đồng phân này có thể đã làm bão hoà các vị trí kết nối cho B12 trong cơ thể và do đó cạnh tranh với B12. Đưa thẳng B12 vào dạ cỏ như chúng tôi đã làm, vì thế đã không làm giảm được sự thiếu B12 vì B12 không thể gắn vào các vị trí kết nối. Trong các nghiên cứu bơm tẳng dinh dưỡng vào dạ dày, không có các đồng phân của B12 được tạo ra vì không có quá trình lên men trong dạ cỏ và B12 là một phần trong chất dinh dưỡng được đưa vào.
Tại sao ở bò lại không thấy xẩy ra các hiện tượng tương tự như ở cừu, đồng nghiệp của tôi tiến sĩ Price đã xác định các vị trí liên kết cho vitamin B12 ở bò khác hoàn toàn ở cừu và có thể chức năng của chúng là gắn kết các đòng phân của B12 riêng biệt do đó có mặt các đồng phân của B12 không làm suy yếu chức năng của B12 thực. Tình hình hiện còn chưa hoàn toàn rõ ràng, nhưng không nghi ngờ gì nữa sự có mặt của một lượng lớn đồng phân của B12 chú không phải thiếu hụt B12
là nguyên nhân làm mỡ mềm và có nhiều axít béo mạch nhánh.. Sự khác biệt giữa

97
cừu, dê và bò như trao đổi ở trên nhấn mạnh rằng có những khác biệt thú vị trong thích ứng sinh hoá giữa các loại gia súc nhai lại. Tài liệu tham khảo cho phần 7.1
1. Duncan, W.R.H., Ørskov, E.R. and Garton, G.A. 1972. Fatty axít composition of triglycerides of lambs fed on barley based diets. Proc. Nutr. Soc. 31, 19A-20A.
2. Ørskov, E.R., Fraser, C. and Gordon, J.G. 1974. Effect of processing of cereals on rumen fermentation, digestibility, rumination time and firmness of subcutaneous fat. Br. J. Nutr. 32, 59-69.
3. Duncan, W.R.H., Ørskov, E.R. and Garton, G.A. 1976. Fatty axít composition of triglycerides of goats fed on a barley rich diets. Proc. Nutr. Soc. 35, 89A-90A
4. Ørskov, E.R., Duncan, W.R.H. and Carnie, C.A. 1975. Cereal processing and food utilization by sheep. 3. The effect of replacing whole barley by whole oats on food utilization and firmness and composition of subcutaneous fat. Anim. Prod. 21, 51-58.
5. Ørskov, E.R. 1995. Utilization of short chain fatty axíts in ruminants. In: Physiological and Clinical Aspects of Short-chain Fatty Axíts (J.H. Cummings, J.L. Rombeau and Takashi Sakata eds.), Cambridge University Press, Cambridge, UK, pp. 243-256.

98
Hướng nghiên cứu 7. Chế biến ngũ cốc Hướng nghiên cứu 7. 2. Các vấn đề của viêm dạ cỏ
Như đã đề cập ở thí nghiệm 1, một vấn đề nảy sinh trong thí nghiệm lớn nuôi cừu thâm canh là viêm dạ cỏ: vách dạ cỏ sưng rộp, các lông nhung kết thành khối. Cũng không có gì ngạc nhiên lắm vì hiện tượng tương tự đã quan sát thấy ở bò, nhưng ngược với bò ở cừu không bao giờ thấy có dấu hiệu của áp xe gan. Bernard Fell, nhà bệnh học ở Viện Rowett, đã mô tả hiện tượng này ở bò và kết luận rằng một trong những nguyên nhân của các áp xe gan là do sự xâm nhập của các gai nhỏ từ ngũ cốc lúa mạch vào các biểu mô dạ cỏ, sự xâm nhập này cho phép vi sinh vật đi vào gan thông qua tĩnh mạch cửa. Tôi đã thảo luận vấn đề này vì chúng tôi thấy khó để chấp nhận rằng các gai hoặc râu lúa mạch gây ra áp xe gan vì tôi đã thấy vấn đề tương tự ở bò ăn ngô ở Hoa Kỳ. Chúng tôi quyết định thực hiện thí nghiệm một phần nhằm kiểm tra vách dạ cỏ cừu được vỗ béo bằng lúa mạch nghiền dạng viên hoặc ngô đập vỡ dạng viên (1). Nghiên cứu này không bao giờ được xuất bản nhưng không thấy có sự khác biệt giữa hai khẩu phần như vậy áp xe gan ở bò là do nguyên nhân khác chứ không phải là do gai và râu từ lúa mạch. Chúng tôi nghi ngờ lông bò có thể là nguyên nhân, cừu ăn ở khẩu phần ít xơ thường kéo lông từ cơ thể chúng hoặc gặm lông con khác, lông đã nuốt vào đã tạo thành những quả bóng lông trong dạ cỏ và đôi khi trong dạ múi khế. Lông bò nuốt vào lại khác, chúng không tạo thành các quả bóng lông mà xâm nhập vào vách dạ cỏ. Chúng tôi đã giải quyết vấn đề này đơn giản bằng cách trộn lông bò vào khẩu phần của cừu. Kết quả gây ấn tương mạnh; lông bò bò quấn vào nhau trong biểu mô phủ gần hết vách dạ cỏ. Cũng có những áp xe tìm thấy trong các mô của vật chủ. Rõ ràng lông bò làm cho viêm dạ cỏ trầm trọng thêm và có thể là nguyên nhân duy nhất của áp xe gan (2). Như vậy có thể giảm áp xe gan bằng cách xén tỉa bớt lông bò đi!
Đóng góp tiếp theo đến khi chúng tôi bắt đầu nghiên cứu ảnh hưởng của
chế biến ngũ cốc (3), trước hết do vấn đề mỡ mềm (Thí nghiệm 7.1). Trong thí nghiệm cuối cùng chúng tôi xác định lượng ăn vào và tỷ lệ tiêu hoá của lúa mạch nguyên hạt và lúa mạch viên ở cừu. Vào ngày trước khi gia súc được giết mổ Colin Fraser và tôi đã lấy mẫu dạ cỏ bằng bơm hút dịch dạ cỏ để xác định thành phần axít béo bay hơi và như thường lệ chúng tôi cũng xác định pH. Tôi đã sửng sốt khi thấy dịch dạ cỏ từ cừu cho ăn lúa mạch nguyên hạt khác hoàn toàn, lỏng hơn và dễ lấy ra. Quan trọng hơn, pH dạ cỏ trung bình 6,2 ở cừu ăn lúa mạch nguyên hạt và 5,2 ở cừu ăn lúa mạch viên. Dường như chúng tôi đã gần chắc chắn loại bỏ được viêm dạ cỏ. Tôi muốn chắc chắn tôi không thiên lệch. Tôi yêu cầu Colin Fraser đánh dấu dạ cỏ và tôi yêu cầu tiến sĩ Fell chia chúng thành hai nhóm: nhóm bình thường và nhóm có vách dạ cỏ bị viêm. Không nghi nghờ gì nữa - tất cả dạ cỏ cừu cho ăn lúa mạch nguyên hạt có lông nhung dạ cỏ không dính nhau dài và không bị viêm trong khi ở những cừu cho ăn lúa mạch viên, vách dạ cỏ dày

99
và các lông nhu kết thành cục. Như vậy cho ăn ngũ cốc nguyên hạt đã có thêm các lợi thế khác, cho ăn ngũ cốc nguyên hạt có vẻ sẽ loại bỏ được viêm dạ cỏ một rắc rối trong chăn nuôi cừu thâm canh. Chúng tôi xuất bản một bài báo khoa học “Ảnh hưởng của lúa mạch không chế biến đến viêm dạ cỏ ở cừu” nhằm can ngăn việc chế biến tạo viên thức ăn từ lúa mạch, hiện vào thời điểm đó đã trở thành một thông lệ trong nuôi dưỡng cừu. Chế biến tạo viên thức ăn từ lúa mạch là một quá trình chế biến không cần thiết, tốn kém và có hại. Những quan sát trên đã làm cho tôi rất hài lòng. Chúng tôi đã biết: viêm dạ cỏ xảy ra như thế nào và cái gì là quan trọng nhất, làm thế nào để tránh được viêm dạ cỏ đồng thời giảm giá thành của khẩu phần vì không cần chế biến thức ăn hạt.
Tài liệu tham khảo cho phần 7.2 1. Ørskov, E.R., Fraser, C., Gill, J.C. and Corse, E.L. 1971. The effect in an
intensive production system of type of cereal and time of weaning on the performance of lambs. Anim. Prod. 13, 485-492.
2. Fell, B.F. kay, M., Ørskov, E.R., Boyne, R. and Walker, J.T. 1972. The role of ingested animal hairs and plant spicules in the pathogenesis of rumenitis. Res. Vet. Sci. 13, 30-36.
3. Ørskov, E.R. and Fraser, C. 1972. Effect on type of rumen fermentation and digestibility of feeding whole as opposed to processed barley to sheep. Proc. Nutr. Soc. 31, 101A.
4. Ør0skov, E.R. 1973. The effect of not nrocessing barley on rumenitis in sheep. Res. Vet. Sci. 14, 110-112.

100
Hướng nghiên cứu 7. Chế biến ngũ cốc
Hướng nghiên cứu 7.3. Ngũ cốc nguyên hạt cho cừu và dê, và những ứng dụng trong công nghệ sản xuất thức ăn
Khám phá dùng thức ăn ngũ cốc nguyên hạt không chế biến nuôi cừu rất có lợi là một khám phá không mong đợi từ trước và rất đáng ngạc nhiên. Không những tỷ lệ tiêu hoá, lượng thức ăn ăn vào và khả năng sản xuất của cừu cho ăn ngũ cốc nguyên hạt tương tự như ở cừu cho ăn ngũ cốc đã chế biến, cho ăn ngũ cốc nguyên hạt còn loại bỏ các vấn đề về sức khoẻ đi kèm với viêm dạ cỏ và các vấn đề về chất lượng thịt xẻ do mỡ mềm (Thí nghiệm 7.1). Khám phá đã mở ra một lĩnh vực nghiên cứu mới cần được quan tâm. Hiệu quả chuyển hoá thức ăn của khẩu phần lúa mạch viên là 3,2 kg vật chất khô/kg tăng trọng trong khi với khẩu phần lúa mạch nguyên hạt hiệu quả chuyển hoá thức ăn chỉ là 3,0 (1). Bảng 7.5. Ảnh hưởng của chế biến lúa mạch, ngô, kiều mạch và lúa mỳ đến pH dạ cỏ, tỷ lệ axít acetic và propionic, chuyển hoá thức ăn, tăng trọng (LWG) và chất chứa ở ruột của cừu. Giai đoạn nuôi dưỡng từ 15 đến 35kg (LW).
Ngũ cốc Chế biến
pH dạ cỏ
Axít acetic (mol/100 mol VFA)
Axít propionic (mol/100 mol VFA)
Tỷ lệ tiêu hoá DM (%)
Chuyển hoá
thức ăn (kg
DM/kg LWG)
LW (g/ngày
)
Chất chứa ruột (% của LW)
Lúa mạch Viên 5,4 45,0 45,3 77,2 2,79 347 9,9
Nguyên hạt
6,4 52,5 30,1 81,1 2,75 340 14,1
Ngô Viên 5,2 41,3 43,2 82,1 2,62 346 8,8
Nguyên hạt
6,1 47,2 38,7 84,3 2,52 345 11,7
Kiều mạch Viên 6,1 53,2 37,5 67,5 3,33 238 20,8
Nguyên hạt
6,7 65,0 18,6 69,9 3,07 241 21,9
Lúa mỳ Viên 5,0 34,2 42,6 86,2 2,56 323 9,3
Nguyên hạt
5,9 52,3 32,2 82,7 2,97 303 10,5
SE 0,14 2,4 3,2 1,3 0,11 15 0,8
Câu hỏi đầu tiên của chúng tôi là: các kết quả trên lúa mach khi áp dụng cho các loại ngũ cốc khác có cho kết quả tương tự không? Chúng tôi đã so sánh lúa mạch, lúa mỳ, kiều mạch và ngô. Chúng tôi xây dựng khẩu phần dựa trên ngũ cốc viên và ngũ cốc nguyên hạt. Cả hai loại khẩu phần có 10% bột cá dựa trên tỷ lệ protein. Bảng 7.3 trình bày các kết quả chính (1,2,3).

101
Rõ ràng pH dạ cỏ chắc chắn cao hơn khoảng một đơn vị ở cừu cho ăn ngũ cốc nguyên hat, cho ăn ngũ cốc nguyên làm cho quá trình phân giải cellulose dễ dàng trong dạ cỏ. Tỷ lệ phân tử am axít acetic cũng cao hơn và tỷ lệ phân tử gam của propionic thấp hơn khi ngũ cốc được cho ăn nguyên hạt. Sự khác nhau ở tỷ lệ tiêu hoá, chuyển hoá thức ăn và tăng trọng nhỏ và không đáng kể; phù hợp với kết quả của chúng tôi về sử dụng axít béo bay hơi. Chất chứa ruột theo phần trăm của khối lượng sống cao hơn một chút khi nuôi dưỡng bằng thức ăn ngũ cốc nguyên hạt và sự khác nau này gàn như hoàn toàn do sự khác nhau về khối lượng chất chứa dạ cỏ.
Một so sánh thú vị cuối cùng được thực hiện (4) bằng cách vỗ béo cừu với khẩu phần kiều mạch nguyên hạt và lúa mạch nguyên hạt. Ở đây chúng tôi xem lại sự khác nhau về chất chứa dạ cỏ và giết thịt cừu ở khoảng 40 kg khi chúng được nuôi kiều mạch hoặc 35 kg khi chúng đợc nuôi với lúa mạch. Kết quả được trình bày ở bảng 7.6. Như có thể thấy trong bảng, chất chứa dạ cỏ ở cừu cho ăn kiều mạch nặng hơn chất chứa dạ cỏ ở cừu ăn yến mạch khoảng 5 kg. Điều thú vị ở đây là theo tính toán tổng chất hữu cơ tiêu hoá ăn vào để cừu đạt được khối lượng giết mổ ở khẩu phần kiều mạch cao hơn 10 kg so với tổng chất hữu cơ tiêu hoá ăn vào để cừu đạt được khối lượng giết mổ ở nhóm cừu ăn yến mạch (Bảng 7.6). Bảng 7.6. Ảnh hưởng của cho ăn lúa mạch nguyên hạt hoặc kiều mạch nguyên hạt đến chất chứa dạ cỏ, số ngày nuôi đẻ đạt tới khối lượng giết mổ và chất hữu cơ tiêu hoá ăn vào (DOM) cho duy trì và sản xuất. Khẩu phần
Khối lượng
sống lúc giết thịt
(kg)
Chất chứa ruột (kg)
Khối lượng
thân thịt (kg)
Ngày nuôi trước
giết mổ
Tổng lượng DOM (kg)
DOM cho duy
trì (kg)
DOM trên mức
duy trì (kg)
Lúa mạch nguyên
35,7 4,7 31,0 74 42,1 22,6 19,5
Kiều mạch nguyên
39,7 9,4 30,3 12,0 51,7 39,0 12,7
SE - 0,3 0,6 5 2,1 1,7 1,7
Tuy nhiên nếu chúng ta tính tổng chất hữu cơ tiêu hoá ăn vào cần cho duy trì với gỉa thiết là chúng cần 414 KJ/kg W0.75 cho duy trì chúg ta thấy là: do sự khác nhau ở số ngày cần để đạt đến khối lượng giết thịt, cừu vỗ béo ở khẩu phần kiều mạch cần 39,0 kg tổng chất hữu cơ tiêu hoá ăn vào trong khi cừu vỗ béo ở khẩu phần lúa mạch cần chỉ 22,6kg tổng chất hữu cơ tiêu hoá ăn vào cho nhu cầu duy trì. Tuy nhiên điều này có nghĩa rằng tổng chất hữu cơ tiêu hoá ăn vào còn lại (trên mức duy trì) ít hơn nhiều ở cừu được vỗ béo bằng kiều mạch so với tổng chất

102
hữu cơ tiêu hoá ăn vào còn lại ở cừu được vỗ béo bằng lúa mạch. Điều quan trọng là kết quả này cho thấy tính toán nhucầu duy trì dựa trên khối lượng sống có vấn đề vì là nhu cầu duy trì cho chất chứa dạ cỏ ít hơn so với nhu cầu duy trì cho kối lượng cơ thể. Không có khả năng là tổng chất hữu cơ tiêu hoá ăn vào cần cho vỗ béo cừu bằng kiều mạch thấp hơn tổng chất hữu cơ tiêu hoá ăn vào cần cho vỗ béo cừu bằng lúa mạch.
Rõ ràng là khi ngũ cốc dùng như là thành phần duy nhất trong khẩu phần cho gia súc nhai lại nhỏ, chúng nên ở dạng chưa chế biến (nguyên hạt). Nhưng bởi vì ngũ cốc thường được cho ăn như thức ăn bổ sung vào khẩu phần cơ sở là thức ăn thô, chúng ta cần đặt câu hỏi phải chăng khi đó nhiều ngũ cốc hơn thoát qua không bị tiêu hoá và do đó làm giảm tỷ lệ tiêu hoá? Nói một cách khác, chúng tôi cho rằng pH dạ cỏ cao hơn khi cho ăn ngũ cốc nguyên hạt có thể ít gây trở ngại cho tiêu hoá cellulose hơn pH thấp khi cho ăn ngũ cốc đã chế biến lên men rất nhanh.
Để có được ý tưởng đầu tiên về vấn đề trên, chúng tôi đã sử dụng phương pháp nylon bag lần đầu để xem xét môi trường dạ cỏ (xem hướng nghiên cứu 2.1). Ở đây chúng tôi sử dụng một mẫu tiêu chuẩn: cỏ khô, và thay đổi môi trường dạ cỏ; trước đó chúng tôi đã sử dụng phương pháp nylon bag chỉ với các mẫu thay đổi và một môi trường dạ cỏ tương đối tiêu chuẩn. Một số kết quả được trình bày trong Bảng 2.1 (trang 34). Kết quả cho thấy tỷ lệ phân giải cỏ khô rõ ràng cao hơn khi cừu ăn lúa mạch nguyên hạt là khẩu phần cơ sở so với khẩu phần cơ sở là lúa mạch viên. Tiếp theo chúng tôi bổ sung vào khẩu phần cơ sở là cỏ khô của cừu với các lượng lúa mạch viên hoặc nguyên hạt khác nhau và xác định lượng ăn vào của cỏ khô (5) (Bảng 7.7). Bảng 7.7. Ảnh hưởng của bổ sung lúa mạch đến lượng ăn vào và tỷ lệ tiêu hoá cỏ khô của cừu. Bổ xung Lượng bổ
xung Cỏ khô ăn
vào (g/ngày)
Tỷ lệ tiêu hoá (%) Tổng lượng DOM ăn vào
(g/ngày) OM ADF
Không - 991 69,8 80,5 692 Lúa mạch nguyên
310g/ngày 790 71,0 76,1 781
Lúa mạch nguyên
616g/ngày 642 73,1 73,3 910
Lúa mạch viên
309g/ngày 820 72,8 77,7 818
Lúa mạch viên
608g/ngày 472 75,4 67,5 814
OM: chất hữu cơ; DOM: chất hữu cơ tiêu hoá.
Ở đây nhìn chung chúng tôi thấy là có sự giảm nhẹ tỷ lệ tiêu hoá của lúa mạch

103
nguyên hạt so với tỷ lệ tiêu hoá của lúa mạch viên; và nhiều ngũ cốc xuất hiện ở phân. Tuy nhiên, sự giảm tỷ lệ tiêu hóa này đã được bù lại nhiều hơn vì tiêu hoá thức ăn thô tốt hơn và quan trọng tương đương hoặc quan trọng hơn là lượng ăn vào cao hơn đáng kể ở mức bổ sung cao. Có thể thấy rằng lượng thức ăn thô ăn vào bị giảm ở mức bổ sung lúa mạch viên cao và tổng lượng chất hữu cơ tiêu hoá ăn vào ở mức bổ sung lúa mạch viên cao không lớn hơn tổng lượng chất hữu cơ tiêu hoá ăn vào ở mức bổ sung lúa mạch viên thấp. Giảm lượng ăn vào của thức ăn thô và tỷ lệ tiêu hóa đã được bù lai hoàn toàn. Như vậy, có vẻ là không chế biến ngũ cốc cho gia súc nhai lại nhỏ có nhiều ưu thế hơn khi ngũ cốc được sử dụng để bổ sung cho khẩu phần cơ sở là thức ăn thô vì tránh được việc tỷ lệ tiêu hóa xơ giảm. Tài liệu tham khảo cho phần 7.3
1. Fraser, C. and Ørskov, E.R. 1974. Cereal processing and food utilization by sheep. 1. The effect of processing on utilization of barley by early weaned lambs. Anim. Prod. 18, 75-83.
2. Ørskov, E.R., Fraser, C. and McHattie, I. 1974. Cereal processing and food utilization by sheep. 2. A note on the effect of feeding unprocessed barley, maize, oats and wheat on food utilization by early weaned lambs. Anim. Prod. 18, 85-88.
3. Ørskov, E.R., Fraser, C. and Gordon, J.G. 1971. Effect of processing of cereals on rumen fermentation, digestibility, rumination time and firmness of subcutaneous fat. Br. J. Nutr. 32, 59-69.
4. Ørskov, E.R., Duncan, W.R.H. and Carnie, C.A. 1975. Cereal processing and food utilization by sheep. 3. The effect of replacing whole barley by whole oats on food utilization and firmness and composition of subcutaneous fat. Anim. Prod. 21, 51-58.
5. Ørskov, E.R., Fraser, C. 1975. The effect of processing of barley based supplements on rumen pH, rate of digestion and voluntary intake in sheep. Br. J. Nutr. 34, 493-500.

104
Hướng nghiên cứu 7. Chế biến ngũ cốc Hướng nghiên cứu 7.4. Những công nghệ đưa urê và khoáng vào trong ngũ cốc nguyên hạt và các áp dụng trong bảo quản ngũ cốc
Như đã được chứng minh ngũ cốc nguyên hạt tốt hơn ngũ cốc chế biến để nuôi gia súc nhai lại nhỏ, tuy nhiên có một vấn đề kỹ thuật xuất hiện ở đây. Ngũ cốc thường thiếu nitơ cho lên men ở dạ cỏ và đáp ứng của gia súc khi thiếu nitơ đã được quan sát (hướng nghiên cứu 1.3). Urê cú thể đưa vào trong thức ăn viên nhưng làm thế nào để những viên thức ăn đã có urê thêm vào với ngũ cốc nguyên hạt mà không bị tách rời ra và không gây ra ngộ độc urê khi cừu ăn urê ở dưới đáy máng ăn. Đầu tiên chứng tôi thử thêm rỉ mật vào ngũ cốc và sau đó bổ sung urê dạng hạt với hy vọng chúng sẽ dính vào ngũ cốc. Vấn đề là rỉ mật và urê đã không dính vào ngũ cốc. Khi rỉ mật khô đi, các hạt urê rời ra.
Một đêm, trong lúc rỗi chờ lấy mẫu từ cừu, tôi đang tìm kiếm xem điều gì sẽ xảy ra nếu 40% dung dịch urê được trộn với ngũ cốc nguyên hạt. Tụi mong đợi urê sẽ kết tinh lại bên ngoài hạt ngũ cốc, nhưng khi tôi trở lại vào sáng hôm sau tôi không thể biết được hạt ngũ cốc nào có urê, hạt ngũ cốc nào không có urê! Để chắc chắn rằng không phải là ai đó đã đùa tôi, chúng tôi đã phân tích ngũ cốc và phát hiện ra rằng urê đã bị hấp phụ hết. Thậm chí sau khi rửa vài lần, cũng không thể tách chiết lại được hơn 20% lượng urê đã dùng. Khi cho cừu ăn ngũ cốc được sử lý urê, tốc độ giải phóng urê thấp hơn nhiều so với tốc độ giải phóng urê được cho ăn khi phủ ngoài hạt ngũ cốc hoặc cho ăn dưới dạng thức ăn viên đã có urê. Đây là một kết quả tuyệt vời! so với sản phẩm đã được thương mại hóa có urê phân giải chậm: "Starea", ngũ cốc sử lý urê tốt hơn và rẻ hơn (1). Vậy khoáng, khoáng vi lượng và vitamin có đuợc hấp phụ tương tự không? Nếu chúng ta sử dụng, ở khả năng có thể, các dạng dung dịch khoáng, khoáng vi lượng và vitamin hòa tan chúng cũng được hấp phụ vào ngũ cốc nguyên hạt rất mạnh (2) do đó nhất định là chúng ta có thể tạo một khẩu phần hoàn chỉnh giống như ngũ cốc nguyên hạt nhưng có chứa những chất bổ xung cần thiết. Nếu cần bổ sung protein, protein phải được đưa vào khẩu phần dưới dạng viên kích cỡ 2mm, để dễ trộn với ngũ cốc, và vì đã trộn cừu không thể chọn thức ăn chúng muốn và thức ăn chúng không muốn ăn. Trong nhiều áp dụng như vỗ béo cừu, thức ăn ngũ cốc xử lý là dạng bổ sung rẻ nhất. Áp dụng để bảo quản ngũ cốc
Tôi cũng hơi lo lắng về những khuyến cáo ngay lập tức liên quan đến kỹ thuật đưa urê vào ngũ cốc bởi vì cho mỗi phần trăm urê bổ sung trong dung dịch 40% sẽ có 1,5% nước được đưa vào. Tôi sợ rằng thêm nước có thể làm cho ngũ cốc bị mốc, và như thế thì thật tai hại. Tuy nhiên, không có vấn đề gì xẩy ra, như chúng tôi đã thấy trong một thí nghiệm với soda (NaOH) xử lý ngũ cốc, vấn đề này sẽ được thảo luận sau (hướng nghiên cứu 7.5). Ở đây chúng tôi vấn đề còn

105
đang cần xem xét là urê có ổn định không trong ngũ cốc xử lý soda (NaOH) ở các độ ẩm khác nhau? Chúng tôi đã làm một thí nghiệm có ngũ cốc không sử lý urê, và ngũ cốc xử lý urê ở các độ ẩm khác nhau. Tôi mong đợi rằng ngũ cốc bảo quản với 18% độ ẩm hoặc cao hơn sẽ bị mốc sau khi bảo quản, sau khi bảo quản chúng tôi phát hiện ra rằng mốc chỉ xuất hiện ở ngũ cốc bảo quản với 18% độ ẩm hoặc cao hơn khi không xử lý hoặc xử lý hoặc xử lý với lượng urê thấp, các mẫu chứa urê, được xử lý với hơn 1% urê vẫn giữ không bị mốc (Bảng 7.8)(3).
Hàm lượng urê 1% hoặc cao hơn có thể bảo quản ngũ cốc tốt. Số lượng vi sinh vật ở mức an toàn cho việc bảo quản; nhưng nếu không có đủ urê, ngũ cốc bị hỏng nhiều.
Kết quả này thực sự là một ngạc nhiên hạnh phúc. Hoàn toàn không sợ phải bảo quản ngũ cốc với urê. Ngược lại urê đã giúp bảo quản ngũ cốc có độ ẩm cao. Như vậy, urê đã hoạt động phục vụ hai mục đích: là một nguồn nitơ phi protein và là chất bảo quản.
Bảng 7.8. Ảnh hưởng của urê đến bảo quản ngũ cốc có độ ẩm cao và hàm lượng nitơ sau 5 tháng bảo quản
Urê (g/kg) Chất khô của ngũ cốc
Hàm lượng nitơ (%) Vi khuẩn sống
(log/g)
Log khối lượng
khuẩn lạc nấm/ g
Ban đầu Sau 5 tháng
0 77,2 1,79 1,84 7,04 8,11 5 75,4 2,07 2,17 5,86 8,48
10 75,6 2,41 2,39 2,70 7,11 15 75,2 2,64 2,57 < 2,60 4,12 20 73,5 3,00 2,90 < 2,60 4,32
Tài liệu tham khảo cho phần 7.4
1. Ørskov, E.R., Smart, R. And Mehrez, A.Z. 1974. A method of including urea in whole grains. J. Agri. Sci., Camb. 83. 299-302.
2. Ørskov, E.R. and Grubb, D.A. 1977. The use of whole grain diets fortified with solutions of urea, minerals and vitamins for lambs. J. Anim. Sci. 2, 307-314.
3. Ørskov, E.R., Stewart, C.S. and Greenhalgh, J.F.D. 1979. The effect of sodium hydroxide and urea on some storage properties of moist grain. J. Agri. Sci., Camb. 92, 185-188.

106
Hướng nghiên cứu 7. Chế biến ngũ cốc Hướng nghiên cứu 7.5. Ngũ cốc xử lý kiềm cho bò
Phát minh ra sản phẩm được gọi là ngũ cốc soda hiện được hàng nghìn nông dân Anh, Mỹ và Châu âu, đặc biệt là nông dân nuôi bò sữa sử dụng, phát minh này có nền tảng khoa học là việc thành công dùng ngũ cốc nguyên hạt nuôi cừu và dê. Khi cho bò ăn ngũ cốc nguyên hạt, trừ một ngoại lệ có thể là kiều mạch, quá nhiều ngũ cốc nguyên hạt thoát qua không được tiêu hoá ở ruột vì vậy, ví dụ lúa mạch cũng chỉ có tỷ lệ tiêu hoá 65%. Rõ ràng là chế biến là cần thiết nhưng chúng ta cũng biết ưu điểm của việc lên men chậm và ưu điểm của duy trì cấu trúc của hạt thức ăn, vì thế thách thức ở đây là tìm ra phương pháp chế biến cho tỷ lệ tiêu hoá chấp nhận được và làm cho quá trình men chậm lại.
Để nghiên cứu tiêu hoá ngũ cốc, phương pháp túi nylon là của trời cho vì với phương pháp này chúng ta có thể xác định tốc độ phân giải của ngũ cốc. Tuy nhiên, ngũ cốc nguyên hạt trong túi không được nhai lại phân giải dạ cỏ và giữ nguyên là hạt hàng tuần lễ. Chúng tôi đã thử cán dẹt hạt ra và thay đổi khoảng cách giữa các trục lăn, nhưng vấn đề là nếu khoảng cách này quá rộng thì hạt nhỏ không bị động chạm đến và để chạm được đến những hạt nhỏ thì những hạt lớn hơn lại bị vỡ ra từng mảnh. Sau đó chúng tôi thử ép sử dụng những cái trục lăn có rãnh, những trục này làm vỡ hoặc bẻ cong hạt ngũ cốc rất hiệu quả. Làm kiểunày tốt hơn rất nhiều so với cán dẹt, nhưng ngũ cốc vẫn bị lên men quá nhanh và cho pH dạ cỏ thấp.
Sau đó tôi nghĩ, người nào khác nữa sử dụng ngũ cốc với số lượng lớn? Những người chưng cất rượu và làm bia ở Scotland chính là những người sử dụng ngũ cốc với số lượng lớn. Tôi gọi điện cho một trong số họ, và hỏi họ xem họ có sử dụng chế biến ngũ cốc để tăng sự nảy mẩm hoặc tạo mạch nha hay không. Họ rất hợp tác và gửi cho tôi 2 mẫu, một mẫu đã xử lý bằng giấy ráp để tăng sự nảy mầm, có lẽ bằng cách tăng sự hấp thu nước, mẫu còn lại được họ gọi là đã "rang". Mẫu được rang thực chất là được xử lý nhiệt hồng ngoại, một phương pháp xử lý bằng vi sóng. Trong thực tế phương pháp này tương tự như làm bỏng ngô nhưng chỉ thực hiện để có một chấm nhỏ tinh bột được trương phồng. Nhìn liếc qua ngũ cốc giống như chưa được xử lý nhưng khi nhìn gần mỗi hạt ngũ cốc đã có một điểm trắng, tại đó tinh bột từ bên trong hạt ngũ cốc đã trương phồng. Phương pháp xử lý bằng giấy ráp không tốt, các nghiên cứu dùng túi nylon cho thấy ngũ cốc xử lý bằng giấy ráp giống như ngũ cốc nguyên hạt chưa chế biến. Phương pháp "rang", tuy nhiên, rất hoàn hảo. Ngũ cốc xử lý bằng phương pháp này lên men chậm nhưng tiêu hoá hoàn toàn khi ủ trong túi nylon. Vậy tại sao không áp dụng phương pháp này? Một công ty thức ăn gia súc đã sử dụng công nghệ "rang" ngũ cốc, kết quả là về mặt dinh dưỡng công nghệ này rất tốt nhưng lại quá đắt để áp dụng trong sản xuất. Tuy nhiên, kết quả này đã giúp tôi một việc quan trọng. Trước tới nay tôi thường nghĩ rằng chế biến ngũ cốc phải là một quá trình cơ học cần dùng máy móc, nhưng không phải như vậy, có thể chế biến ngũ cốc bằng

107
nhiệt. Như vậy giờ đây trí tưởng tượng của người nghiên cứu được giải phóng để nghĩ tới các loại hình chế biến ngũ cốc khác không cần máy móc. Tại sao lại không cần máy móc? Nếu như bạn biết rằng tất cả những cái bạn cần là loại bỏ vỏ xơ của hạt để vi sinh vật tiếp xúc với cơ chất bên trong hạt ngũ cốc, vậy dùng các chất hóa học có thể là một giải pháp chăng? Kiềm, chất hóa học làm xơ phồng lên thì sao?. Tại sao không thử nhỉ? Tôi đã pha một dung dịch NaOH mạnh và trộn với lúa mạch, lúa mạch đã chuyển sang mầu nâu giống như rơm sử lý kiềm. Tuy nhiên, khi ngũ cốc xử lý được cho vào nước "vỏ xơ" rơi ra ngoài.
Lúa mạch sử lý soda được cho vào túi nylon và nó biến mất một cách từ từ. Sự cải thiện phân giải ở dạ cỏ thậm chí còn lớn hơn các phương pháp chế biến cơ học do tỷ lệ tiêu hoá vỏ xơ của hạt lớn hơn. Một loạt các câu hỏi bây giờ cần phải trả lời.
1. Gia súc sẽ ăn lúa mạch sử lý soda? 2. Bao nhiêu NaOH là cần thiết? 3. NaOH có tác động với các ngũ cốc khác không? 4. NaOH tương tác với thức ăn xơ như thế nào? 5. NaOH ảnh hưởng đến lên men dạ cỏ như thế nào? 6. Lúa mạch sử lý soda có ổn định không, có thể bổ sung urê được không và
lúa mạch sử lý soda bổ sung urê có ổn định không? 7. Lúa mạch sử lý soda có khả năng áp dụng thực tế được không và nông dân
sẽ sử dụng lúa mạch sử lý soda hay không? Chúng nghiên cứu các câu hỏi trên theo thứ tự. Trong thí nghiệm đầu tiên của chúng tôi lúa mạch sử lý kiềm được cho ăn
tự do, lượng ăn vào lúa mạch sử lý kiềm cao hơn 6kg/ngày và tương tự như lượng ăn vào của lúa mạch ép (1). Tỷ lệ tiêu hoá ước tính của chúng tôi cho lúa mạch nguyên hạt, lúa mạch ép và lúa mạch sử lý soda tương ứng là 67,2; 75,2 và 78,9 %. Ở đây tôi phải nói thêm rằng, chúng ta thường hàng để ngũ cốc ở ngoài không khí trong khoảng 48h, sau 48 h lượng CO2 đủ từ không khí đã được hấp thụ để chuyển NaOH thành Na2CO3 với pH 10 hoặc khoảng như vậy. Ở đây có thể có vấn đề. Nếu ngũ cốc chỉ mới được trộn với NaOH, một số tinh bột thuỷ phân sẽ có khuynh hướng thấm ra ngoài và gây ảnh hưởng kết dính vì vậy sau một vài giờ ngũ cốc xử lý trở thành thành một khối cứng. Tôi rất không thích khối cứng này, một lần một khối ngũ cốc đã chặn ngang máy trộn trục đứng ở Viện và chúng tôi đã phải dùng rìu để cậy khối cứng này ra! Việc tạo khối cứng có thể tránh được khi trộn ngũ cốc với kiềm và để trong một thời gian dài hơn. Tôi đã từng một lần đề cập đến điều này với một nhà khoa học Nam Phi, phương pháp tạo khối ngũ cốc là một phương pháp đáng mơ ước, khi tạo được các khối ngũ cốc sẽ không cần dngf túi để đựng ngũ cốc nữa.! Người Nam phi đã học phương pháp này và tôi tin rằng người ta bây giờ có thể đã mua những tảng ngũ cốc này!
Bao nhiêu NaOH cần dùng? Điều này đã khá rõ ràng, như mong đợi, lượng NaOH cần dùng phụ thuộc vào loại ngũ cốc (2). (Bảng 7.8) Bảng 7.8. Ảnh hưởng của NaOH (g/kg) đến tỷ lệ tiêu hóa chất khô in vivo (%) của

108
các ngũ cốc khác nhau khi dùng là khẩu phần duy nhất cho cừu.
NaOH (g/kg) Ngô Lúa mỳ Kiều mạch Lúa mạch 0 82,9 78,8 60,5 60,5
15 84,0 85,8 58,6 79,9 30 87,9 92,0 69,8 83,3 45 89,5 88,0 76,5 84,4
Viên 83,0 86,8 68,5 - SE 1,7 1,2 1,9 1,2
Với tất cả ngũ cốc tỷ lệ tiêu hóa chất khô in vivo lớn nhất với ngũ cốc xử lý
NaOH và lớn hơn tỷ lệ tiêu hóa chất khô in vivo của ngũ cốc chế biến cơ học do tỷ lệ tiêu hoá xơ cao hơn (2). Điều này đã xảy ra dù thực tế là một số ngũ cốc nguyên hạt không có vỏ xuất hiện trong phân. Kết quả cũng cho thấy rõ là: lượng NaOH cần dùng phụ thuộc vào hàm lượng xơ: cần ít nhất ở lúa mỳ, ngô và nhiều nhất ở kiều mạch. Khác nhau về tỷ lệ tiêu hoá giữa chế biến cơ học và xử lý NaOH cũng là lớn nhất với kiều mạch; 68,5% với kiều mạch cán dẹt và 76,5% với kiều mạch xử lý NaOH, nguyên nhân là do vỏ xơ của kiều mạch trở nên dễ tiêu hoá hơn nhiều khi xử lý NaOH (2).
Câu hỏi quan trọng nhất là: ngũ cốc xử lý NaOH tương tác với thức ăn xơ như thế nào? Chúng tôi nghiên cứu câu hỏi này bằng cách cho bò ăn một lượng cố định 4 kg chất khô, cùng với cỏ khô cho ăn tự do (Bảng 7.9)(3).
Bảng 7.9. Ảnh hưởng của phương pháp chế biến lúa mạch đến lượng ăn vào từ cỏ khô ở bò. Phương pháp chế biến Lượng cỏ khô ăn vào kg/ngày Xử lý NaOH (30g/kg) 3,4 ‘Rang’ 3,0 Ép cuốn 2,8 Ép 2,8 Nghiền 2,8 Nghiền và tạo viên 2,4 SE 0,11
Kết quả này với tôi là một trong những kết quả thuyết phục nhất. Chế biến
ngũ cốc càng kỹ, càng làm cho lượng ăn vào của cỏ khô giảm đi. Nếu xác định tỷ lệ tiêu hoá của cỏ khô tôi chắc chắn rằng nó cũng giảm.
Chúng tôi đã nghiên cứu ảnh hưởng của bảo quản ngũ cốc xử lý NaOH. Chúng tôi thấy rằng thậm chí chỉ với 2% NaOH cũng sẽ giữ cho số lượng vi sinh vật đủ thấp để cho phép bảo quản ngũ cốc trong 6 tháng ở độ ẩm 20% (4). Bảng 7.10 và kết quả cho những đống ngũ cốc lớn: đường kính 1,5m, đã bảo quản 4 tháng.

109
Bảng 7.10. Ảnh hưởng của xử lý lúa mạch bằng NaOH đến tổng số vi sinh vật lúa mạch và chồi mầm nấm sau 4 tháng bảo quản ở độ ẩm 21%.
NaOH g/kg Log vi khuẩn sống/g Log mầm chồi nấm/g 0 9,4 8,5 20 4,7 4,6 30 4,6 4,3 40 2,8 2,6 Như vậy, hình như xử lý NaOH có thể dùng bảo quản hạt cốc và chế biến
hạt cốc. Dù vậy vẫn còn có những vấn đề khi áp dụng vì kiềm tác động lên xơ sinh ra nhiệt trong đống ngũ cốc. Nhiệt làm cho hơi nước thoát ra khỏi đống ngũ cốc và đôi khi gây nên sự ngưng tụ hơi nước ở lớp ngũ cốc phía trên làm lớp này ẩm ướt và hấp thu CO2 do đó sau một thời gian pH giảm tới mức đủ cho nấm mốc phát triển. Việc này có thể tránh được bằng cách đặt rơm hoặc kiện rơm lên nóc của đống ngũ cốc do đó thay đổi bề mặt tương tác của không khí nóng và lạnh từ ngũ cốc đến rơm.
Ở thời điểm những thí nghiệm này được tiến hành, Home Grown Cereals Authority (Bộ phận phụ trách các hoạt động nghiên cứu, phát triển về cây ngũ cốc và hạt có dầu; trực thuộc Bộ Nông nghiệp Anh) cho chúng tôi một khoản tiền để nghiên cứu thêm với bò thịt và Bridget Barnes đến làm việc với chúng tôi (5,6,7). Kết luận chính từ công trình nghiên cứu này là sử dụng ngũ cốc xử lý NaOH tư-ơng tự như với ngũ cốc cán dẹp nhưng không có viêm dạ cỏ khi cho bò ăn ngũ cốc xử lý NaOH. Chúng tôi cũng lưu ý rằng cần nhiều NaOH bổ sung vào lúa mạch nếu lúa mạch cho ăn với thức ăn thô và cần ít NaOH bổ sung sung vào lúa mạch nếu lúa mạch thức ăn duy nhất vì tốc độ tỷ thoát qua nhanh hơn và do đó thời gian lưu lại ngắn hơn.
Trong một thí nghiệm chúng tôi cho bò ăn 1 lượng thức ăn ủ chua và một lượng lúa mạch cố định bằng cách cho thức ăn ủ trước, sau đó cho ăn lúa mạch hoặc trộn cả hai rồi cho ăn. Lúa mạch ở đây là dạng đã cán dẹp hoặc xử lý NaOH (7)(Bảng 7.11).
Bảng 7.11. Ảnh hưởng của việc cho ăn ủ chua trước, lúa mạch sau, hoặc cho ăn hỗn hợp. (Bò có khối lượng ban đầu 350kg). Phương thức chế biến
Cách cho ăn
Tăng trọng (kg/ngày)
Tổng lượng thức ăn tiêu
thụ (kg/ngày)
Ngày thí
nghiệm
Khối lượng thân thịt (kg)
Cán dẹp Trộn 1,15 8,0 92 253 Xử lý NaOH Trộn 1,20 8,2 90 256 Cán dẹp Ủ chua sau 0,89 5,8 125 247

110
đó ngũ cốc Xử lý NaOH Ử chua sau
đó ngũ cốc 0,78 6,0 125 252
SE 3,6
Kết quả rất thú vị. Thức ăn được ăn nhanh hơn khi trộn so với cho ăn thức ăn ủ chua trước vì bò cần nhiều thời gian thí nghiệm hơn. Điều mong đợi ở đây có thể là: chi phí cho duy trì sẽ nhiều hơn ở bò cần nuôi thêm 35 ngày nữa trong thí nghiệm ăn để ăn hết một lượng thức ăn tương tự và có khối lượng thịt xẻ tương tự. Cần có nghiên cứu thêm về vấn đề này.
Cuối cùng chúng tôi làm thí nghiệm với bò sữa, vì một ưu thế nữa khi sử dụng ngũ cốc xử lý NaOH là hoàn toàn có thể. Bình thường, nếu bò sữa ăn một lượng lớn ngũ cốc, pH dạ cỏ sẽ giảm và tỷ lệ axít propionic trong dạ cỏ sẽ cao lên, dẫn đến giảm mỡ sữa. Tuy nhiên, ngũ cốc xử lý soda có thể không làm giảm pH dạ cỏ do đó tỷ lệ axít acetic cao được duy trì và mỡ sữa không bị ảnh hưởng. Một thí nghiệm nhỏ được tiến hành với bò Friesian ở giữa chu kỳ sữa. Chúng được cho ăn hoặc lúa mạch xử lý NaOH hoặc lúa mạch cán dẹt để có 70 hoặc 80% ngũ cốc trong khẩu phần, cộng với khẩu phần thức ăn thô (Bảng 7.12)(8).
Ở đây có thể thấy rằng trong khi mỡ sữa giảm một cách rõ rệt khi cho ăn lúa mạch cán dẹp, điều này không xẩy ra với lúa mạch xử lý NaOH. Chúng tôi không bao giờ thử nghiệm nhưng tôi nghĩ rằng mỡ sữa sẽ được duy trì ở mức độ bình thường ngay cả khi không sử dụng thức ăn thô khi dùng lúa mạch xử lý NaOH. Bảng 7.12. Ảnh hưởng của tỷ lệ lúa mạch trong khẩu phần và phương thức chế biến đến axít béo bay hơi dạ cỏ, năng suất và mỡ sữa ở bò sữa. Khẩu phần Axít acetic
(phân tử gam %)
Axít propionic (phân tử gam %)
Axít butyric
(phân tử gam %)
Mỡ sữa (%) Năng suất sữa đã hiệu chỉnh theo mỡ sữa
(kg/ngày) Lúa mạch cán dẹp 70:30
56,0 31,0 10,8 3,30 14,7
Lúa mạch NaOH 70:30
64,0 18,0 15,6 3,90 15,5
Lúa mạch cán dẹp 80:20
49,5 37,8 9,5 2,84 13,0
Lúa mạch NaOH 80:20
61,4 19,0 16,7 3,85 14,6
SE 1,7 2,3 1,7 2,7 1,8
Một thí nghiệm lớn khác được thực hiện với bò sữa ở đầu chu kỳ sữa với 30 hoặc 50% rơm sử lý ammoniac trong khẩu phần hoàn chỉnh có lúa mạch cán dẹt

111
hoặc lúa mạch xử lý NaOH (Bảng 7.13)(9). Bảng 7.13. Ảnh hưởng của cho ăn 70 hoặc 50% lúa mạch cán dẹp hoặc xử lý NaOH trộn với rơm sử lý ammoniac đến lượng ăn vào và năng suất sữa bò sữa. Khẩu phần Tỷ lệ hức ăn
tinh thức ăn thô
Lượng ăn vào kg/ngày
Năng suất sữa kg/ngày
Năng suất sữa hiệu chỉnh theo mỡ sữa
kg/ngày Lúa mạch cán dẹp
70:30 16,5 23,5 21,9
Lúa mạch NaOH
70:30 18,8 24,9 23,6
Lúa mạch cán dẹp
50:50 15,6 21,5 21,0
Lúa mạch NaOH
50:50 16,8 26,5 26,3
Một lần nữa, lúa mạch xử lý NaOH vượt trội lúa cán dẹt, không nghi ngờ gì
nữa lúa mạch xử lý NaOH đã tạo ra hình thức lên men dạ cỏ phù hợp hơn cho sản xuất sữa.
Áp dụng thực tiễn
Mặc dù các cơ quan khuyến nông lúc đầu dường như lo ngại về ảnh hưởng của cho ăn ngũ cốc xử lý soda, nông dân đã sử dụng ngay kỹ thuật này, đặc biệt nông dân nuôi bò sữa cho bò ăn TMR (khẩu phần trộn hoàn toàn). Soda được bổ sung vào ngũ cốc ở dạng khô và nước được thêm vào để hoà tan NaOH trong ngũ cốc. Ngũ cốc với nước và NaOH được trộn trong khoảng 30 phút và giữ trong 48 giờ để phần lớn NaOH chuyển hoá thành Na2CO3. Một bất lợi khi sử dụng ngũ cốc xử lý soda là bò uống nhiều nước hơn và nước tiểu nhiều hơn.
Nhiều triệu tấn lúa mỳ và các loại ngũ cốc khác đang được xử lý với soda
hàng năm ỏ Anh và các nước khác. Tôi nhớ lại một lần tôi hỏi một nông dân ở một cuộc triển lãm: ông ta đã cho bò ăn thế nào?. Ông ta nói với tôi "Tôi sử dụng ngũ cốc soda”. Tôi đã tỏ ra thích thú và ông ta hỏi lại tôi: "Ông cũng làm như vậy chứ?".
Tài liệu tham khảo cho phần 7.5 1. Ørskov, E.R. and Greenhalgh, J.F.D. 1977. Alkali treatment as a method of
processing whole grains for cattle. J. Agri. Sci., Camb. 89, 253-255. 2. Ørskov, E.R., Macdearmid, A., Bernes, B.J., Grubb, D.A. and Lukins, B.A.
1979. Barley and oats treated with NaOH for cattle and sheep. Anim. Prod. 28, 432.
3. Ørskov, E.R., Hine, R.S. and Grubb, D.A. 1978. The effect of urea on

112
digestion and voluntary intake by sheep of diets supplemented with fat. Anim. Prod. 27, 241-245.
4. Ørskov, E.R., Stewart, C.S. and Greenhalgh, J.F.D. 1979. The effect of sodium hydroxide and urea on some storage properties of moist grain. J. Agri. Sci., Camb. 92, 185-188.
5. Ørskov, E.R., Macdearmid, A., Grubb, D.A. and Innes, G.M. 1981. Utilization of alkali treated grain. 1. Alkali treated grain in complete diets for steers and lambs. Anim. Fd. Sci. Tech. 6, 273-283.
6. Barnes, B.J. and Ørskov, E.R. 1981. Utilization of alkali treated grain. 2. Utilization by steers of diets based on hay and straw and mixed with either NaOH treated or rolled barley. Anim. Fd. Sci. tech. 6, 347-354.
7. Ørskov, E.R., Barnes, B.J., Macdearmid, A., Williams, P.E.V. and Innes, G.M. 1981. Utilization of alkali treated grain. 3. Utilization by steers of NaOH treated and rolled barley in silage based diets. Anim. Fd. Sci. Tech. 6, 355-365.
8. Ørskov, E.R. and Reid, G.W. 1979. The effect of proportion of concentrate and method of cereal processing on type of rumen fermentation and milk quality in Friesian cows. Proc. Nutr. Soc. 38, 131A.
9. Ørskov, E.R., Reid, G.W. and Tait, C.A.G. 1983. Ammonia treated straw and sodium-hydroxide treated or rolled barley for dairy cows. Anim. Prod. 36, 511.

113
Hướng nghiên cứu 8. Axít béo bay hơi
Hướng nghiên cứu 8.1. Sử dụng axít béo bay hơi – nghiên cứu đầu tiên Tôi muốn diễn tả hướng nghiên cứu này trong 2 giai đoạn bởi vì phần 2 chỉ
có thể nghiên cứu được sau khi có kỹ thuật dinh dưỡng trực tiếp cho dạ dày. Khi đến đại học ở Reading như một sinh viên sau tiến sĩ, năm 1961, tôi bắt
đầu nghiên cứu dinh dưỡng gia súc nhai lại. Người hướng dẫn tôi, Dave Allen, là một tiến sĩ trẻ từ Nottingham, ông đề nghị dự án nghiên cứu của tôi nên là nghiên cứu ảnh hưởng của sự dẫm đạp của cừu đến năng suất cỏ. Tôi cảm thấy đây là chủ đề buồn chán. Tuy nhiên, tôi tìm thấy công trình của Armstrong & Blaxter (1957) đăng trong tạp chí Dinh dưỡng của Anh, tập 11 đặc biệt hấp dẫn. Họ đã chỉ ra, rằng khi bổ xung axít béo bay hơi thường thấy ở dạ cỏ vào dạ cỏ, axít acetic được sử dụng một cách ít hiệu quả hơn axít propionic và axít butyric khi ở trên mức duy trì, họ thậm chí còn thấy rằng hỗn hợp axít béo bay hơi có hàm lượng axít acetic cao và thấp cho kết quả có thể dự đoán được trên cơ sở đánh giá các axít riêng biệt. Tôi cảm thấy rằng nếu điều này đúng là như vậy thì kết quả này sẽ là một sự phát triển cách mạng và có thể đem lại hiệu quả to lớn khi khai thác các phương pháp mới nhằm tăng hiệu quả cho chăn nuôi gia súc nhai lại.
Ở đây tôi phải tạm dừng lại và quay lại xem xét bởi vì những hiểu biết về axít béo bay hơi vẫn còn rất mới ở thời điểm đó. Vào những năm 1942-43 Joseph Barcroft và cộng sự từ Cambridge cuối cùng đã chứng minh được rằng axít béo bay hơi là nguồn năng lượng chính cho gia súc nhai lại chứ không phải là sinh khối vi sinh vật. Việc xác định axít béo bay hơi và đặc biệt là việc tách chúng ra có thể được thực hiện bằng phương pháp sắc ký, nhưng với trang thiết bị đang dùng thời đó, mỗi lần chạy mất khoảng 1 giờ. Một trong những quan sát đầu tiên là tỷ lệ axít acetic trong dịch dạ cỏ ở gia súc nhai lại ăn nhiều thức ăn thô luôn luôn cao hơn tỷ lệ ở gia súc ăn khẩu phần ít thức ăn thô hơn hoặc nhiều thức ăn tinh.
Như đã từng được chỉ ra nhiều lần trước đây năng lượng hấp thu từ khẩu phần thức ăn thô luôn được sử dụng ít hiệu quả hơn năng lượng hấp thu từ khẩu phần có lượng thức ăn tinh cao. Kellner đã thử xác định số lượng này ở thời điểm đầu thế kỷ 20 bằng cách tao ra một yếu tố hiệu chỉnh dựa trên xơ thô và ông gọi hiện tượng "Verdauringsarbeit" hoặc "công cần cho tiêu hoá" (Kellner in Die erhnaerung der Landwirtschaffichen Nutztiere 9th Ed Berlin Parey). Khái niệm "công cần cho tiêu hoá" không hoàn toàn đáng tin cậy khi thấy rằng nồng độ axít acetic dạ cỏ thường cao ở khẩu phần thức ăn thô, dường như khá logic để nghĩ rằng hiệu quả sử dụng ăng lượng thấp và sản xuất nhiều axít acetic có liên quan với nhau. Điều này cũng được ủng hộ bởi một thực tế là về mặt hóa sinh học axít acetic không thể chuyển hoá thành tiền chất glucose, như axít propionic. Axít butyric cũng không thể chuyển hoá thành tiền chất glucose mặc dù axít này được thấy có hiệu quả sử dụng tương đương như axít propionic, nhưng thực tế phiền phức này đã không nhận được nhiều quan tâm. Một sự phát triển khác được

114
Blaxter theo đuổi là xây dựng buồng hô hấp khép kín, không quá đắt để vận hành và sử dụng, giúp nghiên cứu quá trình trao đổi năng lượng dễ dàng hơn. Việc dễ dàng đưa cannullae vào dạ cỏ là một yếu tố khác giúp nghiên cứu bơm dinh dưỡng vào dạ cỏ kết hợp với nghiên cứu trao đổi năng lượng có thể thực hiện. Như vậy thực tế, sự kết hợp của các sự kiện khác nhau đồng thời xảy ra: (1) Nhận ra sự quan trọng của axít béo bay hơi, (2) xây dựng buồng hô hấp rẻ tiền, (3) những tiến bộ trong phẫu thuật, và (4) tách các axít béo bay hơi bằng sắc ký kết hợp với khả năng và quyết tâm của ngài Blaxter. Tóm lại, tôi cảm thấy nghiên cứu trao chất axít béo bay hơi đã đạt đợc một giai đoạn mới và đầy hứng thú.
Tôi có thể chỉ sử dụng những dụng cụ đơn giản. Chúng tôi có cừu và có thể mua muối canxi hoặc natri của các axít béo bay hơi. Chúng tôi không có kinh phí để phân tích thân thịt nhưng chúng tôi đo được độ dày mỡ, khối lượng thân thịt lạnh, etc. Công trình nghiên cứu đã được xuất bản trong 4 bài báo (1, 2, 3, 4) và có thể tóm tắt một cách ngắn gọn.
Chúng tôi không thể tìm ra bất cứ sự khác nhau nào trong việc sử dụng acetate, propionate hoặc butyrate được cho vào trong thức ăn cho cừu đang sinh trưởng. Đầu tiên tôi nghĩ đó là lỗi của tôi! Làm sao mà một nhà khoa học trẻ sử dụng các kỹ thuật kém tiên tiến lại có thể không đồg ý với các nhà khoa học như Blaxter và Armstrong? Tôi nhìn vào kết quả của tôi trong tuyệt vọng nhưng những kết quả này thực sự không ửng hộ các kết quả nghiên cứu của nhóm Blaxter. Khi viết luận văn tiến sĩ của mình, tôi đọc rất kỹ đến từng chi tiết các bài báo của Blaxter và Armstrong và phát hiện ra một sai lầm trong bài báo cuối cùng của họ. Họ đã bơm hai hỗn hợp axít béo bay hơi, có nồng độ axít acetic cao hoặc thấp, và cho rằng kết quả này củng cố các kết quả thu được trong một thí nghiệm khác khi họ bơm các axít béo bay hơi riêng rẽ. Tôi phát hiện ra rằng họ đã giả thiết là tỷ lệ năng lượng của axít béo bay hơi tương tự như thành phần phân tử lượng của chúng. Bởi vì axít acetic chứa 867 kj/mol, axít propionic 1520 kj/mol và axít butyric 2169 kj/mol, nên điều này rõ ràng không đúng và việc sử dụng các giá trị đúng đã làm suy yếu các kết luận của họ ở bài báo cuối cùng. Để chắc chắn hơn cho các kết luận của tôi, tôi đến gặp các đồng nghiệp của tôi ở Viện nghiên quốc gia về sữa để trao đổi, nhưng do danh tiếng của ngài Blaxter quá lớn, họ nói với tôi: nếu Blaxter đã tính toán thì chắc chắn kết quả tính toán là đúng. Tôi chắc chắn là sự việc không phải như vậy, nhưng ở đây tôi gặp phải một vấn đề nhỏ. Armstrong được chỉ định là người chấm luận án tiến sỹ từ cơ quan khác (External examiner) của tôi! Thật là không dễ dàng cho tôi vì cần thiết phải thảo luận kỹ sự khác nhau này trong luận văn của mình. Thầy hướng dẫn của tôi đã viết thư cho David Armstrong và giải thích các tính toán của tôi. David Armstrong trả lời rằng có vẻ chúng tôi đã đúng nhưng ông ta sẽ khẳng định sau. Ông ta đã không bao giờ làm điều đó! Chúng tôi đã có một bài thi vấn đáp tốt nhưng sai lầm của nhóm Blaxter đã không bao giờ được đề cập đến. Về sau khi tôi thảo luận sai lầm này với Kenneth Blaxter, ông bị mất tinh thần khi đã có một sai lầm như vậy! Tôi cảm thấy rằng sai lầm này nên được công bố trên tạp chí vì đây là một sai lầm quan trọng, nhưng điều đó đã không bao giờ xẩy ra.

115
Trong thí nghiệm cuối cùng của chúng tôi (4) chúng tôi đã sử dụng khẩu phần có tỷ lệ thức ăn thô nhiều hoặc ít - và bổ xung muối của axít béo bay hơi vào cả hai khẩu phần. Như trình bày trong bảng 8.1, chúng tôi thấy có sự khác nhau trong sử dụng năng lượng tiêu hoá giữa hai khẩu phần và không có sai khác đáng kể về thống kê giữa các muối của axít béo bay hơi, ít nhất là trên cơ sở đánh giá tăng trọng trong khối lượng thân thịt. Bảng 8.1. Ảnh hưởng của cho ăn các muối axít béo bay hơi đến năng suất của cừu được nuôi với khẩu phần thức ăn thô cao (HR) hoặc thức ăn thô thấp (LR). Khối lượng ban đầu 25kg. Khẩu phần ME ăn vào
(MJ/ngày) Khối lượng kết thúc
(kg) Khối lượng thân
thịt (kg)
HR 7,37 33,6 14,0 HR + acetate 8,65 36,4 16,0 HR + propionate 8,69 36,6 16,6 HR + butyrate 8,65 35,4 16,1 LR 7,58 34,2 15,8 LR + acetate 8,86 39,1 18,9 LR + propionate 8,90 37,1 18,3 LR + butyrate 8,86 37,7 18,3 SE 0,60 0,45
Tôi đã có một kết luận quan trong khác, ít nhất là cho tôi, là nếu sự sai khác
giữa các khẩu phần thức ăn thô và thức ăn tinh không liên quan đến axít béo bay hơi, thì sự khác nhau phải là do sự tạo thành axít béo bay hơi chứ không phải do sự sử dụng axít béo bay hơi. Kết luận này trở nên rất quan trọng với tôi khi tôi đến làm việc với Platt tại phòng thí nghiệm về năng lượng trao đổi, của Bộ nông nghiệp Hoa kỳ (USDA), Beltsville, Myraland. Trong thời gian đó, tiến bộ về vi sinh vật học được Robert Hungate dẫn dắt, Robert Hungate vừa mới xuất bản cuốn sách của mình với một số phương trình nổi tiếng nhưng đơn giản:
C6H12O6 + 2H2O 2 CH3COOH + 4 H2 C6H12O6 + H2 2 CH3CH2COOH + 2 H2O C6H12O6 + 2 H2O CH3CH2CH2-COOH + 2 H2O 4H2 + CO2 CH4 + 2 H2O
Tôi rất hứng thú với cuốn sách này. Phải chăng chúng ta nên xem xét sự hình tạo axít béo bay hơi? Rõ ràng sự hình tạo luôn di kèm với tạo ra hydro và do đó sẽ có nhiều metan được tạo ra. Tôi đã thực hiện rất nhiều phép tính và cùng với Flatt & Moe chúng tôi đã viết một bài báo (5) về kỹ thuật cân bằng lên men dùng để ước tính lượng axít béo bay hơi được tạo ra. Lượng metan được tạo ra có thể được tính toán là M = 1/2pa - 1/4pp + 1/4 pb, ở đây pa và pp va pb là số mol axít acetic, propionic và axít butyric. Khi biết được lượng khí metan tạo ra và tỷ lệ

116
phân tử lượng của axít béo bay hơi, chúng ta có thể ước tính được có bao nhiêu phần trăm carbonhydrate tiêu hóa lên men trong dạ cỏ. Về tổng thể sự khác nhau trong lượng metan tạo ra giữa các khẩu phần thức ăn thô và thức ăn tinh không thể giải thích hoàn toàn sự khác nhau thông thường về hiệu quả sử dụng năng lượng giữa các khẩu phần thức ăn thô và thức ăn tinh. Tuy nhiên, khi năng lượng trao đổi (ME) được tính toán từ năng lượng tiêu hoá (DE), chắc chắn là không chính xác để giả thiết rằng metan tạo ra là không đổi cho tất cả các loại khẩu phần.
Chúng tôi thực hiện một thí nghiệm tại Beltssville để so sánh ảnh hưởng của axít acetic và propionic bơm vào trong bò sữa. Thí nghiệm này được thực hiện trong buồng hô hấp mở gián tiếp, thiết bị hiện đại nhất có vào lúc ấy. Kết quả rõ ràng. Khi tính đến lượng nhiệt tạo ra, không có sự khác nhau về sử dụng axít acetic và axít propionic, và axít acetic và axít propionic đuợc sử dụng để chuyển thành năng lượng trao đổi của khẩu phần cơ sở với với hiệu quả tương tự (6). Các kết quả nổi trội nhất được tóm tắt trong bảng 8.2.
Bảng 8.2. Lượng năng lượng trao đổi đưa vào cơ thể, lượng nhiệt tạo ra, năng lượng của sữa và cân bằng năng lượng trong mô của cơ thể ở bò sữa được đưa axít acetic hoặc axít propionic thay thế 15% năng lượng trao đổi. Khẩu phần Năng lượng
ăn vào (MJ/ngày)
Nhiệt tạo ra (MJ/ngày)
Năng lượng trong sữa (MJ/ngày)
Cân bằng năng lượng trong
mô của cơ thể (MJ/ngày)
Cơ sở 130 71 43 15 Cơ sở với axít acetic
121 67 39 14
Cơ sở với axít propionic
128 71 32 26
SE 1,6 2,9 2,9 2,1
Trong khi bơm acetate và propionic gây ra sự khác nhau đáng kể và có những thay đổi ngược chiều về năng lượng trong sữa và năng lượng trong mô của cơ thể, nhưng năng lượng ăn vào và cân bằng năng lượng trong mô của cơ thể không khác nhau một cách đáng kể về mặt thống kê, nhiệt tạo ra cũng không sai khác đáng kể về thống kê.
Trong khi nghiên cứu này quan trọng hơn nghiên cứu ở Viện Hannah, sách giáo khoa đã đầy những kết luận từ các nghiên cứu trước đây. Axít acetic bị kết tội là được sử dụng một cách không hiệu quả. Vấn đề thực sự là rất khó cung cấp bằng chứng thuyết phục, vì người ta có thể chỉ đưa một lượng axít bổ xung tương đối nhỏ, khoảng 15% năng lượng trao đổi. Tại sao lại chỉ 15% này? Lý do là nếu hơn 15 % tính ngon miệng của khẩu phần cơ sở sẽ giảm. Các kết quả nghiên cứu của Armtrong và Blaxter đã được chấp nhận một cách không phê phán mặc dù tính toán sai. Các công ty thương mại cạnh tranh lẫn nhau để tìm ra hoá chất có thể

117
giảm axít acetic đáng kinh sợ này. Thành công nhất là chất coccidiostat monensin. Hoá chất này làm giảm tạo khí metan một chút và giảm tỷ lệ axít acetic. Tuy nhiên, ảnh hưởng của việc giảm tạo khí metan trong sử dụng năng lượng khó có thể phát hiện được. Ảnh hưởng chính của coccidiostat monensin là ảnh hưởng của một kháng sinh, nhưng nhờ ảnh hưởng của nó đến axít acetic nó được cơ quan quản lý dược phẩm và thực phẩm Hoa kỳ chấp nhận như là chất ức chế metan! và vẫn đang được sử dụng rộng rãi.
Trong khi bơm axít acetic không ảnh hưởng đến lượng nhiệt tạo ra, chúng làm tăng tỷ lệ mỡ sữa trong khi bơm axít propionic lại làm giảm mỡ sữa. Kết quả này đúng như mong đợi vì thí nghiệm trước đó đã cho thấy: các khẩu phần thức ăn tinh nhiều làm giảm mỡ sữa - một kết quả có thể do nồng độ axít propionic cao. Kết quả này sau đó đã được khẳng định trong nghiên cứu bơn axít vào dạ cỏ. Các câu chuyện về axít béo bay hơi khác từ Beltsville
Khi tôi tới Beltsville các nhà khoa học ở đây đang ở trong giai đoạn cuối của một thí nghiệm lớn. Một loạt các khẩu phần khác nhau về tỷ lệ thức ăn tinh được dùng để nuôi bò sữa, hàm lượng năng lượng trao đổi của khẩu phần được ghi chép chi tiết. Thí nghiệm do Bill Flatt, người tổ chức thí nghiệm tốt nhất mà tôi từng làm việc, thiết kế. Trong suốt thời gian thí nghiệm, mẫu dạ cỏ được lấy bằng bơm dạ dày và được bảo quản trong tủ lạnh sâu. Nhiệm vụ của tôi là phân tích những mẫu này. Bởi vì đây là công việc thường ngày, tôi muốn nó được thực hiện một nhanh nhất có thể và khi tôi có máy sắc ký khí tôi làm việc ngày đêm để phân tích hết số mẫu. Mỗi lần phân tích mất khoảng 15 phút - một tiến bộ, trong khi với thiết bị sắc ký của chúng tôi ở Đại học Reading mỗi lần phân tích mất ít nhất 1giờ. Chúng tôi ghi lại đỉnh cho axít acetic, propionic, isobutyric, butyric, isocvaleric và valeric và ngay khi đỉnh axít valeric xuất hiện một mẫu mới được bơm vào máy.
Một đêm khi tôi đang cố gắng đưa các mẫu vào máy, tôi ngủ quên và khi thức dậy tôi thấy có một một đỉnh khá lớn khác xuất hiện sau đỉnh của axít valeric khoảng 15 phút. Chúng tôi xác định đó là axít caproic. Mẫu có axít caproic được lấy từ bò ăn khẩu phần có chứa glucose và cellulo công nghiệp. Đôi khi tỷ lệ axít caproic vượt quá 10%. Chúng tôi phát hiện thấy axít caproic bất cứ khi nào bò được ăn khẩu phần này, nhưng tại sao?
Sau đó tôi đọc được một bài báo về vi sinh vật, bài báo cho rằng nếu butyrate và ethanol có mặt trong môi trường kỵ khí thì axít caproic sẽ được tạo ra. Ngay sau đó chúng tôi nhận được một cột của máy sắc ký khí có thể xác định ethanol và lương ethanol vừa đủ cũng xuất hiện trong các mẫu dạ cỏ (7, 8). Tôi, sau đó đã đặt giả thiết với Roger Hemken, Đại học Maryland rằng axít caproic hình như đã làm tăng mỡ sữa, do vậy tỷ lệ mỡ sữa có thể tăng khi bơm alcohol vào dạ dầy! Chúng tôi đã thiết kế một thí nghiệm bơm dinh dưỡng vào dạ dầy tại Park college và chắc chắn rằng mỡ sữa sẽ tăng cùng với sự tăng lên của axít caproic trong dịch dạ cỏ (xem Bảng 8.3).
Niềm tin của tôi vào hoá sinh đã được đền đáp! Kết quả thí nghiệm đã được tôi công bố trên tạp chí cùng với Roger Hamken (9) người đã theo đuổi hướng nghiên cứu cùng với một nghiên cứu sinh tiến sĩ.

118
Bảng 8.3. Ảnh hưởng của việc bơm ethanol đến tỷ lệ mỡ sữa và tỷ lệ axít caproic trong dịch dạ cỏ ở 3 bò vắt sữa ở chu kỳ.
Ngày Khẩu phần cơ sở (kg/ngày)
Ethanol (g/ngày)
Tỷ lệ axít caproic Mỡ sữa (%)
1-14 9,1 0,0 Rất ít 2,27 19-42 8,2 875 6,4 3,37 43-46 8,2 0,0 Rất ít 2,92
Viện Rowett
Đây là kết thúc của phần 1 về axít béo bay hơi, vì năm 1966 tôi chấp nhận đề nghị của tiến sĩ Blaxter về làm việc tại Viện Rowett, Aberdeeen, để phát triển một hệ thống nuôi cừu thâm canh (hướng nghên cứu 1.1). Cừu cái cần phải trở thành những cái máy đẻ và cừu được nuôi thâm canh do đó chúng có ít nhất 3 lần chửa/ 2 năm. Tài liệu tham khảo cho phần 8.1
1. Ørskov, E.R. and Allen, D.M. 1966. Utilization of salt of volatile fatty axíts by growing sheep. 1. Acetate, propionate and butyrate as sources of energy for young growing lambs. Br. J. Nutr. 20, 295-305.
2. Ørskov, E.R., Hovell, F.D.DeB. and Allen, D.M. 1966. Utilization of salt of volatile fatty axíts by growing sheep. 2. Effect of stage of maturity and hormone implantation on the utilization of volatile fatty axíts as sources of energy for growth and fattening. Br. J. Nutr. 20, 307-315.
3. Ørskov, E.R. and Allen, D.M. 1966. Utilization of salt of volatile fatty axíts by growing sheep. 3. Effect of frequency of feeding on the utilization of acetate and propionate by young growing lambs. Br. J. Nutr. 20, 509-517.
4. Ørskov, E.R. and Allen, D.M. 1966. Utilization of salt of volatile fatty axíts by growing sheep. 4. Effect of type of rumen fermentation of the basal diet on the utilization of salt of volatile fatty axíts for nitrogen retention and body gains. Br. J. Nutr. 20, 519-532.
5. Ørskov, E.R., Flatt, W.P. and Moe, P.W. 1968. A fermentation balance approach to estimate extent of fermentation and efficiency of volatile fatty axít formation in ruminants. J. Dairy Sci. 51, 14-29.
6. Ørskov, E.R., Flatt, W.P., Moe, P.W., Munson, A.W. and Hemken, R.W. 1969. The influence of ruminal infusion of volatile fatty axíts on milk yield and composition and energy utilization by lactating cows. Br. J. Nutr. 23, 443-453.
7. Ørskov, E.R., Flatt, W.P., Moe, P.W. and Oltjen, R.R. 1967. Caproic axít and ethanol in ruminal ingesta of cattle receiving purified diets. J. Dairy Sci. 50, 239-242.
8. Ørskov, E.R. and Oltjen, R.R. 1967. Influence of carbohydraten and

119
nitrogen sources on the rumen volatile fatty axíts and ethanol of cattle fed purified diets. J. Nutr. 93, 222-228.
9. Ørskov, E.R., Hemken, R.W. and Moore, L.A. 1967. Effect of ethanol infusion on milk fat content and composition and on volatile fatty axíts in rumen liquor. J. Dairy Sci. 50, 692-697.

120
Hướng nghiên cứu 8. Axít béo bay hơi
Hướng nghiên cứu 8.2. Sự phát triển kỹ thuật dinh dưỡng trực tiếp cho dạ dày.
Theo tôi, trong tất cả các kỹ thuật mà tôi chịu trách nhiệm phát triển, kỹ
thuật dinh dưỡng trực tiếp cho dạ dày là kỹ thuật quan trọng nhất. Khi kỹ thuật này đang được đưa vào sử dụng, chúng ta như lại một lần nữa có thêm cánh để bay! Bao nhiêu điều trước đây không thể làm được đã trở nên có thể nhờ kỹ thuật này.. Ở đây chúng ta sẽ xem cái gì đã xẩy ra.
Tiếp theo những thành công nhất định của chúng tôi trong việc khai thác phản xạ đóng mở rãnh thực quản và trong phát triển những nghiên cứu tiêu hoá động sử dụng túi nylon tôi cảm thấy lúc này chúng tôi có đủ khả năng mạo hiểm theo đuổi các kỹ thuật mà nhiều nhà nghiên cứu đã thất bại. Dinh dưỡng trực tiếp cho dạ dày là nuôi gia súc nhai lại chỉ bằng cách bơm chất dinh dưỡng lỏng vào dạ dày, cho phép chúng ta duy trì gia súc nhai lại dựa vào axít béo bay hơi, protein và vitamin nghĩa là các sản phẩm cuối cùng cuả lên men vi sinh vật, nhưng với những trở ngại và sự không ổn định do lên men thức ăn của vi sinh vật trong dạ cỏ bị loại bỏ.
Vấn đề động lực chính cho nghiên cứu ở đây là là câu chuyện axít béo bay hơi, câu chuyện đã làm tôi thất vọng 15 năm trước. Tôi chắc chắn rằng kết luận của chúng tôi là đúng, nghĩa là không có sự khác nhau về hiệu quả sử dụng axít béo bay hơi, nhng bởi vì chỉ có một số lượng hạn chế axít béo bay hơi bổ sung được đưa vào khẩu phần của gia súc nuôi bình thường, vì vậy không thể chứng minh kết luận này một cách thoả đáng và sách giáo khoa vẫn còn giữ lại các kết quả về sự sử dụng không hiệu quả axít acetatic.
Cảm giác của tôi là nếu như kỹ thuật thất bại, chúng tôi ít nhất có được sự an ủi vì đã biết rằng mọi người cũng đã thất bại! Tôi đã có một sự giúp đỡ tuyệt vời ở David Grubb do đó chúng tôi bắt đầu thử nghiệm. Đầu tiên, chúng tôi theo logic rằng: nếu pH dạ cỏ là bình thường từ 6 đến 7, dịch dinh dưỡng bơm vào cũng nên có pH từ 6 đến 7. Tuy nhiên điều này là không chíng xác hoặc nó không hoạt động, vì ngay khi chúng tôi đạt đến mức dung dịch dinh dưỡng bơm vào đủ cung cấp dinh dưỡng cho khoảng một nửa nhu cầu duy trì, chúng tôi đã gặp vấn đề: gia súc ốm và nước tiểu bài tiết ra không bình thường do quá nhiều natri. Sau đó chúng tôi cố gắng bơm axít tự do đây là cách axít béo bay hơi được sản xuất. Cách này cũng không được, ngay khi chúng tôi đạt được mức duy trì, pH trong dạ cỏ giảm đột ngột và gia súc bị axidosis (axít dạ cỏ).
Trong tâm trạng thất vọng tôi đã đủ có khôn ngoan để hỏi gia súc (cách nói ẩn dụ) " Bạn làm cái đó như thế nào trên trái đất này?" Làm thế nào gia súc sản xuất nhiều axít béo bay như vậy mà vẫn giữ được pH dạ cỏ từ 6 đến 7? Điều này rất quan trọng và khi hồi tưởng lại đây là câu hỏi hiển nhiên nhất mà tôi đã không hỏi! Dựa trên những nghiên cứu trước của Dobson ở Viện Rowett gia súc đáp lại một cách đơn giản, cũng một cách nói ẩn dụ, chúng tôi sản xuất axít tự do, sau đó

121
axít tự do bị trung hoà thành anion và bơi qua bể dung dịch đệm rồi rời khỏi dạ cỏ phần lớn ở dạng axít tự do một lần nữa. Do đó tất cả điều các ông cần là phải thay thế dung dịch đệm đã roiừ khỏi dạ cỏ đi xuống phần đường tiêu hóa phía dới. Bình thường dung dịch đệm do nước bọt cung cấp nhưng khi gia súc không ăn gì cả sự tiết nước bọt giảm rất mạnh và cần phải được cung cấp hoặc chúng tôi cần tìm một cái gì đó để gia súc nhai!
Sau đó mọi việc đã rất dễ dàng. Chúng tôi chọn cách đưa khoáng đa lượng vào dạ cỏ vì chúng cung cấp chất đệm, khoáng đa lượng, khoáng vi lượng, vitamins và protein cùng được đưa vào dạ múi khế. Sau đó chúng tôi có 3 ống, một ống để đưa axít béo bay hơi vào dạ cỏ, một ống đưa nước bọt hoặc nước bọt nhân tạo vào dạ cỏ, ống còn lại dùng để đưa protein, cùng với khoáng vi lượng và vitamins vào dạ múi khế. Gordon Wenham là nhà giải phẫu xuất sắc; trong 40 con cừu chúng tôi chuẩn bị cho thí nghiệm lớn đầu tiên, không có con nào bị chết trong quá trình phẫu thuật cũng như trong thí nghiệm. Bill Corrigal bác sĩ thú y đã tự hào được hợp tác nghiên cứu và chụi trách nhiệm về tất cả các vấn đề tiềm tàng có thể gây nhiễm trùng. Trong khi kỹ thuật được phát triển để nghiên cứu sử dụng axít béo bay hơi, kỹ thuật cũng mở ra nhiều khả năng khác. Tôi không nghĩ có kỹ thuật nào khác trong dinh dưỡng gia súc nhai lại có ảnh hưởng sâu sắc trong nâng cao hiểu biết của chúng ta về trao đổi chất ở gia súc nhai lại.
Ở gia súc cho dinh dưỡng trực tiếp vào dạ cỏ thường thiếu các kích thích vào vách dạ cỏ, các kích thích này bình thường do thức ăn thô ăn vào đảm nhiệm, vì lý do này chúng tôi đặt các bàn chải cọ xong chảo bằng nhựa plastic vào dạ cỏ qua lỗ dò dạ cỏ (canula). Ở cừu các bàn chải được đưa vào từng cái một để tránh bị ợ lên thực quản do vận động của dạ cỏ. Ở bò chúng tôi buộc nửa tá bàn chải lại với nhau để chắc chắn rằng chúng không bị ợ lên. Chúng tôi cũng đã thử cho gia súc nhai những vật không tiêu hoá được để có phản dạ tiết nước bọt đủ điều chỉnh pH dạ cỏ mà không cần cung cấp thêm chất đệm. Những vật không tiêu hoá được đôi khi đã hoạt động tốt nhưng chúng tôi thấy không đáng tin cậy khi sử dụng những vật không tiêu hoá được trong việc kiểm soát pH dạ cỏ (1, 2).
Bàn chải đã giữ cho vách dạ cỏ khoẻ mạnh và lông nhung phát triển tốt. Điều này nói với chúng ta rằng trong khi gia súc nhai lại không có nhu cầu dinh dưỡng về chất xơ, nhưng chắc chắn chúng có nhu cầu về chất cọ xát rắn. Thông thường xơ là nguồn chất cọ xát rắn rẻ nhất. Nếu như cấu trúc hoặc tính cọ xát của chất xơ bị phá huỷ nó không còn tác dụng cọ xát kích thích vách dạ cỏ.
Bây giờ chúng tôi đã đã kiểm soát được tình hình. Chúng tôi đã tự do thay đổi thành phần axít béo bay hơi đưa vào dạ cỏ theo ý muốn. Chúng tôi dĩ nhiên đã có sự tự do thay đổi số lượng và điều quan trọng nhất tự do thay đổi tỷ lệ protein: năng lượng ý muốn. Điều này cực kỳ quan trọng vì khi có sự lên men vi sinh vật trong đièu kiện cho ăn bình thường (không phải đưa chất dinh dưỡng trực tiếp vào dạ dầy) khả năng thay đổi tỷ lệ năng lượng/protein là rất hạn chế. Nếu thiếu vi sinh vật trong dạ cỏ gia súc sẽ ngừng ăn. Chúng tôi có thể là lần đầu nghiên cứu khẩu phần không có protein, và có được thông tin về nhu cầu protein cơ bản của vật chủ. Chúng tôi có thể đạt được trạng thái sau hấp thu trong một vài giờ thay vì

122
trong một vài ngày. Chúng tôi có thể chỉ bơm một mình protein và một mình năng lượng vào dạ múi khế Như tôi đã nói ở phần đầu, kỹ thuật này đã chắp thêm đôi cánh cho chúng tôi một lần nữa!
Tài liệu tham khảo cho phần 8.2
1. Ørskov, E.R., Grubb, D.A. Wenham, G. and Corrigall, W. 1979. The sustennance of growing and fattening ruminants by intragastric infusion of volatile fatty axíts and protein. Br. J. Nutr. 41, 553-558.
2. MacLeod, N.A., Corrigall, W., Stirton, R.A. and Ørskov, E.R. 1982. Intragastric infusion of nutrients in cattle. Br. J. Nutr. 47, 547-552.

123
Hướng nghiên cứu 8. Axít béo bay hơi
Hướng nghiên cứu 8.3. Sử dụng axít béo bay hơi - Công trình cuối cùng
Tôi gọi phần sau này là axít béo bay hơi phần 2 một cách có chủ ý vì tôi đã phải để lại chủ đề này 15 năm không nghiên cứu sau khi tôi đến Viện Rowett. Bản thân tôi ít nhất đã rất hài lòng vì tôi đã đưa ra bằng chứng là không có sự khác nhau thực sự trong việc sử dụng năng lượng của axít béo bay hơi như tôi đã trình bày ở phần 1 axít béo bay hơi (Hướng nghiên cứu 8.1). Trong sách giáo khoa vẫn đầy thông tin cho rằng axít acetic được sử dụng không hiệu quả. Các công ty đã cố gắng giảm tỷ lệ axít acetic và bị nhầm lẫn với những thay đổi không đáng kể trong sản sinh khí mêtan ở dạ cỏ. Thuốc kháng sinh đã trở thành chất bổ sung vào thức ăn gia súc với quan niệm là kháng sinh sẽ làm giảm lượng acetate sinh ra và hiện chúng vẫn đang được sử dụng ở một số nước.
Năm 1977 khi tôi giải thích trong một chương về dinh dưỡng trực tiếp cho dạ dày tôi cảm thấy rằng chúng tôi có thể có đủ khả năng nếu cần thiết chịu một thất bại trong nỗ lực phát triển kỹ thuật dinh dưỡng trực tiếp cho dạ dày, nhưng câu chuyện axít béo bay hơi chỉ có thể thực sự được giải quyết nếu chúng ta có thể kiểm soát hoàn toàn số lượng và tỷ lệ các axít béo bay hơi được hấp thu. Như vậy sự phát triển của kỹ thuật dinh dưỡng trực tiếp cho dạ dày được tiếp thêm sức mạnh bởi chính sự thất bại hoàn toàn của câu chuyện về axít béo bay hơi, sự thất bại đã dẫn dắt khoa học đi theo hướng sai lầm, nhưng lại có vẻ không thể dừng lại được. Khi kỹ thuật đã hoàn hảo chúng tôi ngay lập tức dùng nó để nghiên cứu với 40 cừu được lắp một cannulla dạ cỏ loại nhỏ và một ống thông dạ múi khế (1).
Chúng tôi sử dụng 4 hỗn hợp các axít béo bay hơi và 2 mức bơm dinh dưỡng, một mức theo ước tính cung cấp đủ năng lượng cho duy trì và một mức gấp 2 lần năng lượng duy trì, với 2 mức như vậy cho phép chúng tôi tính toán hiệu quả sử dụng năng lượng ít nhất là trên mức duy trì. Chúng tôi thực tế đã sử dụng nhiều mức hơn, nhưng đã có một số vấn đề xẩy ra khi đạt được mức năng lượng gấp 2 lần năng lượng duy trì ở trường hợp tỷ lệ axít acetic cao hơn, nhưng bốn mức axit được trình bày dưới đây đại diện cho phạm vi thực tế tại đó tỷ lệ các axít béo bay hơi đợc phát hiện. Các kết quả quan trọng được trình bày ở bảng dưới đây (Bảng 8.4).
Bảng 8.4. Ước tính nhu cầu duy trì và hiệu quả sử dụng hỗn hợp axít béo bay hơi trên mức duy trì ở cừu.
Thành phần phân tử Cân bằng năng lượng
(kJ/Kg W0,75
Hiệu quả sử dụng năng lượng trên mức duy trì
(Kf)
KF ở nhu cầu duy trì cố định ở mức 450 kj/ Kg
W0,75
Axít acetic Axít propionic
Axít butyric
450 450 100 420 61 64

124
550 350 100 470 60 57 650 250 100 460 63 61 750 150 100 460 63 62
Như mong đợi, không có dấu hiệu của sự khác biệt nào cả. Chúng tôi cũng
đã sử dụng 85% axít acetic nhưng không thể nhận đạt được mức bơm vào trên 1,5 lần mức duy trì. Kf ở đây là 61. Hình như hiệu quả sử dụng axít béo bay hơi tối đa là khoảng 60% và dĩ nhiên không phải là tỷ lệ các axít béo làm cho thiệu quả sử dụng năng lượng trao đổi thay đổi từ 30-60% như ước tính trong các tài liệu tham khảo. Trong nghiên cứu tiếp sau nghiên cứu trên chúng tôi thấy hiệu quả sử dụng axít béo bay hơi tương tự là 64%. Sau đó chúng tôi tiến hành các thí nghiệm tương tự với bò nhưng sử dụng một phạm vi các tỷ lệ axít béo bay hơi lớn hơn nhiều, thay đổi từ 16 đến 91 phần trăm phân tử lượng axít acetic từ 0 đến 75% axít propionic và với tỷ lệ không đổi 9% axít butyric, tất cả đều ở mức 1,5 lần nhu cầu duy trì (2). Ở đây chúng tôi xác định các sản phẩm của trao đổi chất trong nước tiểu, huyết tươngg cũng như đo lượng nhiệt sinh ra.
Các kết luận chính được liệt kê ở dưới đây: 1. Dưới 75% phần trăm phân tử lượng axít acetic, mọi thứ đều ổn định cả các
sản phẩm của trao đổi chất và lượng nhiệt sỉnh ra. 2. Khi tỷ lệ axít acetíc từ 75% đến 91%, BOH butyrate huyết tương tăng từ
khoảng 0,4 đến 5 mmol/lít, nitơ nước tiểu tăng khoảng 20%, nhiệt sinh ra giảm khoảng 5%, insulin huyết tương giảm, glucose huyết tương giảm, nồng độ axít béo tự do trong huyết tương giảm.
3. Không có axít acetic trong nước tiểu ở mức 91% axít acetic. Nói một cách khác, vượt ra ngoài tỷ lệ axít béo bay hơi sinh lý bình thường,
trao đổi chất sẽ gặp vấn đề và như mong đợi glucose không đủ, nhiệt sinh ra giảm do quá trình oxy hóa các sản phẩm trao đổi chất: BOH butyrate và axít acetic kém. Vậy, chúng ta có thể chắc chắn rằng sự khác nhau trong sử dụng axít béo bay hơi đã không có vai trò gì trong sự khác biệt về sử dụng năng lượng trao đổi của thức ăn thô và thức ăn tinh.
Cái gì là nguyên nhân gây ra sự khác nhau trong sử dụng năng lượng trao đổi trong thức ăn thô hoặc thức ăn tinh? Well Juan Ku Vera đã nghiên cứu các nguyên nhân này và đã xác định chi phí cho nhai lại ăn và đứng là 11,4; 9,3 và 4,6 J/kg khối lượng/phút. Well Juan Ku Vera đã thực hiện nghiên cứu này bằng cách cho bò được đưa dinh dưỡng trực tiếp vào da dày ăn những thức ăn không tiêu hoá được. Vậy, có thể tính toán một cách dễ dàng rằng các hoạt động trên kết hợp với thời gian ăn, thời gian nhai lại và đứng lên để ăn là nguyên nhân của những khác nhau trong sử dụng năng lượng trao đổi của thức ăn thô và thức ăn tinh. Vào cuối thế kỷ 20 Kellner gọi sự khác nhau đó là "Verdaurungsarbeit" hoặc "công cho tiêu hoá", ông ta đã không quá sai lầm. Như vậy tăng hiệu quả sử dụng thức ăn thô có thể được hoàn thành bằng cách giảm thời gian ăn, ví dụ, nghiền thức ăn thô. Nghiền thức ăn thô, tuy vậy, khá đắt đỏ, và làm giảm tỷ lệ tiêu hoá thức ăn thô và giảm tiết nước bọt, giảm tỷ lệ tiêu hoá thức ăn thô và giảm tiết nước bọt sau đó sẽ

125
làm giảm tiêu hoá cellulo như đã thảo luận trong hướng nghiên cứu 2.1 và vì vậy nói chung nghiền thức ăn thô không phải là một lựa chọn kinh tế.
Chúng tôi đã tiến hành một thí nghiệm thú vị khác. Vì chúng tôi đã kiểm soát được tỷ lệ axít béo bay hơi trong dung dịch dinh dưỡng bơm vào dạ dày, chúng tôi có thể đặt câu hỏi: tỷ lệ axít béo bay hơi thấy trong dạ cỏ đại diện cho tỷ lệ axít béo bay hơi được tạo ra? Và có yếu tố nào nữa không có thể ảnh hưởng đến sự khác nhau về tỷ lệ axít béo bay hơi thấy trong dạ cỏ và tỷ lệ axít béo bay hơi được tạo ra? Chúng tôi để pH dạ cỏ biến động bằng cách giảm bơm dịch đệm và giữ lượng axít béo bay hơi bơm vào ổn định (Bảng 8.5)(4). Bảng 8.5. Ảnh hưởng của pH đến tỷ lệ axít béo bay hơi đo được trong dạ cỏ từ 1 tỷ lệ axít béo bay hơi cố địng trong dung dịch dinh dưỡng bơm vào
Tỷ lệ axít béo bay hơi trong dung dịch dinh dưỡng bơm vào (%)
pH dạ cỏ 6,6 5,5 7,2*
Axít acetic % 62,9 66,4 72,5 62,9 Axít propionic % 26,2 25,1 20,5 26,2 Axít butyric % 10,8 8,5 7,0 10,8
*Tính toán bằng hồi quy để biết pH thích hợp tại đó tỷ lệ phân tử lượng axít béo bay hơi sẽ tương tự như tỷ lệ này trong dung dịch dinh dưỡng bơm vào.
Rõ ràng tỷ lệ phân tử lượng axít béo bay hơi bị ảnh hưởng bởi pH do sự
khác nhau trong tốc độ hấp thu và ở pH thấp tỷ lệ phân tử lượng của axít acetíc được ước tính cao hơn 10 % và tỷ lệ phân tử lượng của axít propionic và butyric được ước tính thấp. Chúng ta phải rất cẩn trọng khi giả thiết rằng tỷ lệ phân tử lượng các axít béo bay hơi được phát hiện đại diện cho tỷ lệ phân tử lượng các axít béo bay hơi được sản xuất. Chúng tôi, trong một công trình chưa xuất bản, đã thay đổi áp suất thẩm thấu dạ cỏ nhưng thấy không có ảnh hưởng khác nhau của áp suất thẩm thấu dạ cỏ đến tốc độ hấp thu axít béo bay hơi.
Tài liệu tham khảo cho phần 8.3
1. Ørskov, E.R., Grubb, D.A., Smith, J.S., Webster, A.J.F. and Corrigall, W. 1979. Efficiency of utilization of volatile fatty axíts for maintenance and energy retention by sheep. Br. J. Nutr. 41, 541-551.
2. Ørskov, E.R., MacLeod, N.A. and Nakashima, Y. 1991. Effect of different volatile fatty axíts mixtures on energy metabolism in cattle. J. Anim. Sci. 69, 3389-3397.
3. Ku-Vera, J.C., MacLeod, N.A. and Ørskov, E.R. 1989. Energy exchanges of cattle nourished by intragastric infusion of nutrients. In: “Energy Metabolism of Farm Animals”. Y. van der Honeny and W.H. Close (eds). pp. 271-274. Proc. 11th Symp. Lunteren (EAAP 43), Pudoc, Wageningen.
4. MacLeod, N.A., Ørskov, E.R. and Atkinson, T. 1984. The effect of pH on the relative proportions of ruminal volatile fatty axíts in sheep sustained by

126
intragastric infusions. J. Agric. Sci. Camb. 103, 459-462.

127
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ
Hướng nghiên cứu 9.1. Protein và ảnh hưởng của protein đến huy động mỡ trợ giúp cho sinh trưởng
Trong phần đề cập đến nhu cầu protein của cừu cai sữa sớm (Hướng nghiên cứu 1.1), chúng ta trở lại với vấn đề là cừu nuôi ở mức dinh dưỡng tương đối thấp béo hơn ở cùng khối lượng giết mổ so với cừu nuôi vỗ béo ở mức dinh dưỡng cao. Không thấy có sự giảm mỡ khi tăng protein lên ở khẩu phần có mức dinh dưỡng thấp nhưng có sự đáp ứng với protein khẩu phần ở mức dinh dưỡng cao. Chúng tôi nhận ra, từ những nghiên cứu trên gia súc mổ lỗ dò là ở mức dinh dưỡng thấp, protein gần như bị phân giải hết trong dạ cỏ và do đó không có khả năng đáp ứng với protein. Kết quả là cừu nuôi với khẩu phần dinh dưỡng thấp thiếu protein ngay cả khi khẩu phần có đến 20% protein thô. Điều này đặt ra câu hỏi toàn bộ khái niệm đã được chấp nhận trước đây về hệ đường cong trong đó nhu cầu protein tăng lên cùng với mức dinh dưỡng tăng có còn đúng chăng? những đường cong như vậy ghi lại phần dinh dưỡng được cung cấp hơn là nhu cầu dinh dưỡng. Sự phát minh ra kỹ thuật dinh dưỡng trực tiếp cho dạ dày (Hướng nghiên cứu 8.2) có nghĩa là chúng ta bây giờ có thể đưa những câu hỏi vào nghiên cứu. Chúng ta đã có khả năng chứng minh rằng cừu, bò cạn sữa và bò đực non thiến tất cả đều đạt được cân bằng protein dương khi chỉ đưa một mình protein riêng vào dạ dày. Vì protein có khá ít năng lượng, gia súc ở trong tình trạng cân bằng năng lượng âm nghiêm trọng, cân bằng protein dương đã huy động mỡ tích lũy của cơ thể một cách hiệu quả như một nguồn năng lượng cho quá trình tích lũy protein – Đây hoàn toàn là một sự thay đổi về khái niệm!
Vậy liệu gia súc ở trong tình trạng cân bằng năng lượng âm nghiêm trọng, cân bằng protein dương huy động mỡ tích lũy của cơ thể cho sinh trưởng có xẩy ra trong thực tế không? Ở đây các nghiên cứu của chúng tôi về tỷ lệ phân giải (Hướng nghiên cứu 1.2) trở nên hữu ích, bây giờ chúng ta có thể cho gia súc ăn các nguồn protein, chúng ta đã biết phần lớn protein của nguồn thức ăn đó không bị phân giải ở dạ cỏ. Chúng tôi đã tiến hành thí nghiệm giết mổ so sánh (1) để xem xét nếu các câu hỏi trên có đúng là như vậy không?. Cừu được cho ăn hai mức dinh dưỡng thấp và cao, có hoặc không bổ sung 125g bột cá/ngày, trong thời gian 92 ngày. Khẩu phần cơ sơ là rơm xử lý NaOH với urê bổ sung (Bảng 9.1). Bảng 9.1. Ảnh hưởng của việc bổ sung bột cá vòa khẩu phần của cừu ở cân bằng năng lượng âm đến tăng trọng và thay đổi thành phần cơ thể.
Sử lý ME ăn vào
(KJ/ngày)
Tăng trọng
(g/ngày)
Tăng trọng thân thịt
(kg)
Tăng trọng (kg)
Mỡ tăng (kg)
Protein tăng (kg)
Ít rơm 235 -47 -5,1 -4,3 -3,5 -0,5 Ít rơm + bột cá 307 3 0,6 0,1 -1,5 0,5

128
Nhiều rơm 456 0 0,8 0 -1,4 -0,1 Nhiều rơm + bột cá 458 68 4,2 6,1 -0,9 0,9
SE 6 8 0,6 1,2 0,5 0,1 *Nhu cầu năng lượng duy trì khoảng 410KJME/kgW0.75
Kết quả rất rõ ràng. Mỡ dự trữ của cơ thể đã được huy động như là nguồn năng lượng cho tăng trọng khi protein có sẵn. Tăng khối lượng 6,1kg sau 92 ngày đã đạt được cùng với giảm mỡ trong cơ thể.
Tôi không cảm thấy những phát hiện này là đặc biệt quan trọng ở Anh nhưng chúng có thể rất quan trọng với một số nơi trên thế giới có những biến động lớn trong việc cung cấp chất dinh dưỡng cho gia súc. Thường có quá nhiều bò hay cừu vỗ béo được bán ra thị trường ở cuối mùa mưa trong khi trong mùa khô giá bán cao nhưng có ít bò bán ra thi trường. Ở đây đã có 1 phương pháp có thể nuôi gia súc ở một mức nuôi dưỡng thấp mà chúng vẫn có thể chịu đựng trong một thời gian với một chút sụt cân hoặc không, thậm chí còn có tăng trọng khi có đủ protein.
Trong khi tiềm năng rộng hơn của các phương pháp nuôi dưỡng này vẫn cần được kiểm chứng một cách đầy đủ hơn, một sự kiện thú vị bất chợt xảy ra làm cho phương pháp này phù hợp với Vương quốc liên hiệp Anh: thảm hoạ Chernobyl. Nổ nhà máy điện nguyên tử ở Kiev là cho một lượng nhỏ caeseim phóng xạ rơi xuống Scotland. Trong một thời gian, cừu từ một vùng của Scotland không bán được, và lúc lệnh cấm được dỡ bỏ cừu quá béo và không thích hợp cho việc trợ giá, vì trợ giá chỉ khi giá cừu sản xuất cao hơn gái bán ngoài thị trường. May mắn là số cừu này được để lại không bị hủy và nếu quá béo chúng được gửi trỏ lại trang trại nơi chúng xuất phát. Người ta đã hỏi chúng tôi: Bây giờ chúng ta phải làm gì?. Câu trả lời thật đơn giản, đưa chúng trở lại trang trại, cho chúng ăn rơm, và cũng bổ xung protein: 50g bột cá mỗi ngày. Khẩu phần giảm béo đã có tác dụng. Những người nông dân rất dễ chịu và thực sự đã quay lại nói với tôi rằng cừu đã tăng trọng và đã giảm béo tới mức có thể chấp nhận được.
Sau đó chúng tôi đã thực hiện một thí nghiệm kết hợp với Trường Nông nghiệp Scotland (2) để chúng minh điều này. Cừu quá béo được cho ăn rơm và bổ xung hoặc không bổ xung bột cá như trình bày ở Bảng 9.2. Bảng 9.2. Ảnh hưởng của nuôi dưỡng cừu quá béo bằng rơm hoặc bổ sung bột cá trong thời gian 14, 28 và 42 ngày đến khối lượng thân thịt của
Thời gian nuôi Không có bột cá Có bột cá 14 21,8 22,4 28 20,4 21,9 42 18,3 21,6
Ở đây cừu cho ăn bột cá duy trì khối lượng thân thịt và giảm mỡ nên thịt
đạt chất lượng tốt trong khi cừu cho ăn rơm giảm cả mỡ và phần nạc nên đã giảm

129
3,5 kg khối lượg thân thịt. Hiện tượng tăng trọng bù thực sự là do thiếu và đủ protein (3,4,5). Cừu
được nuôi duy trì càng lâu, chúng càng đáp ứng nhanh hơn với bổ sung protein ví dụ bột cá. Chúng ta cũng có thể cũng phân biệt giữa đầy đủ protein và tăng trưởng bù. Theo cách đó gia súc trưởng thành nuôi thiếu năng lượng sẽ giảm protein trong cơ thể vì protein vi sinh vật không đủ cho nhu cầu duy trì protein. Khi năng lượng và protein vi sinh vật gián tiếp tăng lên, hoặc bổ xung protein không bị phân giải, gia súc nhanh chóng lấp bù các thiếu hụt protein trong cơ thể trong một giai đoạn cho đến khi protein trong cơ thể hoàn toàn đủ. Gia súc non, tuy nhiên, nếu nuôi thiếu dinh dưỡng sẽ có khối lượng thấp hơn gia súc non được nuôi đầy đủ ở cùng độ tuổi. Khi việc cung cấp protein được phục hồi những gia súc trước đó được nuôi thiếu dinh dưỡng sẽ thể hiện cả hiện tượng đủ protein và sinh trưởng bù.
Ảnh hưởng của tình trạng protein đã được nghiên cứu thêm. Cừu sẽ không đủ protein do không bơm chất dinh dưỡng có protein trong 10 ngày. Trước khi protein trong cơ thể giảm, protein được duy trì ở 714mg N/kgW0.75 trong khi sau khi giảm chúng duy trì ở mức 1103mg N/ kgW0.75 nếu bơm vào dạ dầy 2430 mg N/ kgW0.75.
Nghiên cứu cùng về quan hệ giữa năng lượng và protein là xem xét xem có ảnh hưởng nào không của năng lượng ngoại sinh đến đáp ứng của gia súc với protein khi gia súc đã tích lũy mỡ trong cơ thể.
Kết luận chung ở đây là năng lượng ngoại sinh ít hoặc không có ảnh hưởng gì. Sự thiếu hụt glucose lúc đói làm tăng hiệu quả sử dụng protein lên trên 100% khi chỉ có protein được bơm vào dạ dầy vì một phần protein được sử dụng như tiền chất của glucose. Tuy nhiên, nếu mức betahydroxybutyrate trong huyết thanh ở mức bình thường, nghĩa là không có sự thiếu hụt glucose thì cung cấp năng lượng ngoại sinh ít có hoặc không có ảnh hưởng gì đến tích luỹ protein. Kết quả này cho thấy việc thiết lập hệ đường cong kết nối cung cấp năng lượng với tích luỹ nitơ là do một thực tế là lượng năng lượng cung cấp cho gia súc nuôi bằng các thức ăn bình thường cũng xác định được lượng protein vi sinh vật tạo ra. Tuy thế, cúng ta đã thấy rằng cừu gầy ít đáp ứng với năng lượng ngoại sinh trong khi cừu béo không có đáp ứng gì cả (6, 7). Ở đây nên nhớ, như đã đề cập trong hướng nghiên cứu 9.4 về trao đổi đói, rằng bò hình như có nhu cầu glucose cao hơn cừu. Để kết luận, có thể nói rằng:
1. Chúng ta thực sự ít biết về nhu cầu protein thực sự của gia súc nhai lại vật chủ.
2. Chúng ta biết rằng nhu cầu protein bị ảnh hưởng rất nhiều bởi dinh dưỡng trước thời điểm xác định nhu cầu hoặc tình trạng protein của gia súc trước đó, do đó tính nhu cầu protein trên khối lượng gia súc là hoàn toàn chưa đầy đủ. Mối quan hệ năng lượng-protein là cơ sở bảo đảm chỉ khi việc cung cấp
năng lượng nội sinh cũng được xem xét. Gia súc nhai lại nhanh chóng thay đổi từ năng lượng ngoại sinh sang thành năng lượng nội sinh do đó trong phần lớn trường hợp tích luỹ protein không tăng do tăng cung cấp năng lượng ngoại sinh.

130
Khái niệm mối quan hệ là do sự kết hợp giữa năng lượng ngoại sinh và protein vi sinh vật.
Tài liệu tham khảo cho phần 9.1 1. Fattet, I., Hovell. F.D.DeB., Ørskov, E.R., Kyle, K.J. and Smart, R.I. 1984.
Undernutrition in sheep. The effect of supplementation with protein on protein accretion. Br. J. Nutr. 52, 561-574.
2. Vipond, J.E., King, Magaret E., Ørskov, E.R. and Wetherill, G.Z. 1989. Effects of fish meal supplementation on performance of overfat lambs fed on barley straw to reduce carcass fatness. Anim. Prod. 48, 131-138.
3. Ørskov, E.R., Grubb, D.A. and MacLeod, N.A. 1983. basal urinary nitrogen excretion and growth response to supplemantal protein by lambs close to energy equilibrium. Br. J. Nutr. 50, 173-187.
4. Hovell. F.D.DeB., Ørskov, E.R., MacLeod, N.A. and McDonald, I. 1983. The effect of changes in the amount of energy infused as volatile fatty axíts on the nitrogen retention and creatinine excretion of lambs wholly nourished by intragastric infusion. Br. J. Nutr. 50, 331-343.
5. Hovell. F.D.DeB., Ørskov, E.R., Kyle, D.J. and MacLeod, N.A. 1987. Undernutrition in sheep. Nitrogen repletion by N-depleted sheep. Br. J. Nutr. 57, 77-88.
6. Chowdhury, S.A., Ørskov, E.R., Hovell. F.D.DeB., Scaife, J.R. and Mollison, G. 1997. Protein utilization during energy undernutrition in sheep sustained by intragastric infusion: effects of protein infusion level, with or without sub-maintenance amounts of energy from volatile fatty axíts, on energy and protein metabolism. Br. J. Nutr., 77, (4), 565-576.
7. Chowdhury, S.A., Ørskov, E.R., Hovell. F.D.DeB., Scaife, J.R. and Mollison, G. 1997. Protein utilization during energy undernutrition in sheep sustained by intrgastric infusion. Effect of body fatness on the protein metabolism of energy-restricted sheep. Br. J. Nutr., 78, No. 2, 273-282.

131
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ
Hướng nghiên cứu 9.2. Bài xuất nitơ cơ bản hoặc nhu cầu protein tối thiểu
Khi chúng tôi chuẩn bị cuốn sách của ARC về nhu cầu dinh dưỡng của gia súc nhai lại năm 1977, chúng tôi bắt buộc phải lấy nhu cầu protein nội sinh, sự bài xuất nitơ khi gia súc nhai lại ăn khẩu phần có tỷ lệ protein thấp, được báo cáo lần đầu vào năm 1977 (1, 2). Giá trị được chấp nhận là 100mg nitơ/kgW0.75/ngày. Chúng tôi đã quyết định tách nhu cầu protein thành nhu cầu protein cho vi sinh vật dạ cỏ và nhu cầu protein cho gia súc vật chủ và loại bỏ khái niệm nitơ phân trao đổi.
Với sự kiện phát triển được kỹ thuật dinh dưỡng trực tiếp cho dạ dày chúng tôi bây giờ đã có thể kiểm tra giá trị trên bằng kỹ thuật dinh dưỡng trực tiếp cho dạ dày. Chúng tôi bơm vào dạ dầy bò, bò đực thiến và cừu lượng axít béo bay hơi đủ cung cấp năng lượng cho duy trì (khoảng 450kJ/kgW0.75/ngày). Chúng tôi không bơm protein vào dạ cỏ, việc này đôi khi không thể thực hiện được ở gia súc nhai lại nuôi dưỡng bình thường (cho ăn). Các giá trị về bài xuất nitơ được trình bày trong Bảng 1. Chúng vào khoảng 350mg/kg0.75, cao hơn nhiều giá trị được chấp nhận là 100mg/kg0.75. Khi chúng tôi đã lặp lại thí nghiệm một vài lần tôi đến gặp ngài Kenneth Blaxter, giám đốc của chúng tôi, đồng thời cũng là chủ tịch Uỷ ban ARC về Nhu cầu dinh dưỡng. Tôi nói với ông "Chúng tôi có những sai sót, rất nghiêm trọng" và tôi là người có trách nhiệm chính về việc này.
Nói ngắn gọn vấn đề là thế này: nếu chúng ta tính toán sản xuất amino axít vi sinh vật (AA) ở gia súc nhận được 450 kj ME hàng ngày sử dụng yếu tố chúng tôi đã chấp nhận (1,32 g nitơ vi sinh vật/MJ năng lượng ME) và sử dụng các giá trị chúng ta đã tính được về hiệu quả sử dụng protein vi sinh vật là 0,8 cho AAN (Nitơ của axit amin); tỷ lệ tiêu hoá ở ruột non là 0,85 và tỷ lệ hấp thu axit amin là 0,8 chúng ta có: 0,450 x 1,32 x 0,8 x 0,85 x 0,8 = 323 mg/AA thuần khiết/kg0,75. Bây giờ nếu nhu cầu cơ bản trong khoảng 350mg/kg0.75, một gia súc được nuôi khẩu phần có năng lượng chỉ đủ cho duy trì sẽ khó sản xuất đủ protein vi sinh vật thoả mãn nhu cầu duy trì của nó, dĩ nhiên nếu nhu cầu thực sự chỉ là 100 mg/ kgW0.75 thì đây sẽ là một câu chuyện khác.
Ngài Kenneth Blaxter ngay lập tức họp Tiểu ban ARC về nhu cầu protein gia súc nhai lại và phần bổ sung chúng tôi đã xuất bản năm 1984 (4) tuy vậy phần bổ sung đã không thực sự được cải tiến thêm. Kể từ đó, nhiều thí nghiệm thực địa đã cho thấy gia súc nhai lại còn non được cho ăn khẩu phần đủ năng lượng duy trì, không cho ăn protein, protein vi sinh vật dạ cỏ là nguồn protein duy nhất của chúng có thể giảm khối lượng do mô bị mất đi, nhưng gia súc trưởng thành vẫn giữ được cân bằng.
Chúng tôi đã tiến hành quan rất nhiều lần bài xuất nitơ cơ bản ở cả bò và cừu và các giá trị cho mỗi thí nghiệm được trình bày trong Bảng 10.3. Có thể thấy rằng tính theo khối lượng trao đổi các giá trị này có vẻ thấp hơn một chút ở cừu (307 mg) so với bò (341 mg/kg0,75). Giá trị trung bình chung dao động từ 224 đến

132
429 mg và các giá trị cao nhất được ghi nhận ở những gia súc non nhất nhưng chúng tôi không hiểu rõ nguyên nhân của sự khác biệt này. Mỗi giá trị thường có độ lệch tiêu chuẩn khoảng 10 đến 15%. Các giá trị này bao gồm cả nitơ phân và nước tiểu nhưng nitơ phân không bao giờ chiếm hơn 10% tổng số nitơ. Như vậy phần lớn bài xuất nitơ cơ bản qua nước tiểu. Bảng 9.3. Bài xuất nitơ phân và nước tiểu ở gia súc được nuôi duy trì bằng cách chỉ bơm vào dạ dày axít béo bay hơi (không bơm protein) nhằm đáp ứng nhu cầu năng lượng cho duy trì.
Gia súc Bài xuất N cơ bản (mg/kgW0,75)
Tài liệu tham khảo
Bò sữa ở đầu chu kỳ sữa 340 3 Bò sữa ở giữa chu kỳ sữa 329 3 Bò sữa ở cuối chu kỳ sữa 283 3 Bò đực non 408 3 Bò đực già 320 3 Bò đực 345 5 Bò đực 409 6 Bò đực 365 7 Bò đực 265 8 Bò đực 352 *Ku Vera Bò đực 332 *Ku Vera Bò đực 354 *Ku Vera Cừu 429 9 Cừu 356 10 Cừu 386 6 Cừu 327 11 Cừu 250 11 Cừu 224 11 Cừu 320 12 Cừu 269 12 Cừu 264 13 Cừu 299 13
*J.C. Ku Vera, trao đổi năng lượng và nitơ ở bò đợc nuôi bằng bơm dinh dưỡng trực tiếp vào dạ dày. Luận án tiến sĩ, Trường đại học Aberdeen.
Hiệu quả thực tế của kết quả này đơn giản là nhìn chung gia súc được nuôi bằng khẩu phần chỉ đủ năng lượng cho duy trì sẽ chỉ sản xuất đủ protein vi sinh vật dạ cỏ để đáp ứng nhu cầu tối thiểu hoặc nội sinh của chúng, gia súc non ăn khẩu phần như trên sẽ có thiếu hụt nghĩa là chúng có thể giảm khối lượng trong khi vẫn có cân bằng năng lượng. Phải nhớ rằng, dĩ nhiên, gia súc nhai lại ăn khẩu phần như trên hoặc chúng tăng lượng mỡ lên và giảm mô cơ, sản xuất protein vi

133
sinh vật cũng không ổn định và biến thiên. Khám phá này đã đóng góp rất lớn cho hiểu biết của chúng ta về dinh dưỡng protein ở gia súc vật chủ, và khám phá này có thể đã không có được nếu không áp dụng kỹ thuật dinh dưỡng trực tiếp cho dạ dày. Tài liệu tham khảo cho phần 9.2
1. Roy, J.H.B., Balch, C.C., Miller, E.L., Ørskov, E.R. and Smith, R.H. 1977. Calculation of the N requirement for ruminants from nitrogen metabolism studies. 2nd Internatinal Symposium on Protein Metabolism and Nutrition (ed. S. Taminga) pp. 126-129 Centre for Agricultural Publishing, Wageningen, The Netherlands.
2. Miller, E.L., Balch, C.C., Ørskov, E.R., Roy, J.H.B. and Smith, R.H. 1977. Comparison of calculated N requirement for ruminants with the results from practical feeding trials. 2nd Internatinal Symposium on Protein Metabolism and Nutrition (ed. S. Taminga) pp. 126-129 Centre for Agricultural Publishing, Wageningen, The Netherlands.
3. Ørskov, E.R. and MacLeod, N.A. 1982. The determination of the minimal nitrogen excretion in steers and dairy cows and its physiological and practical implications. Br. J. Nutr. 47, 625-636.
4. Balch, C.C., Asmstrong, D.G., Greenhalgh, J.F.D., Miller, E.L., Ørskov, E.R., Roy, J.H.B., Smith, R.H. and Tayler, J.C. 1984. The nutrient Requirements of Ruminant Livestock. Supplement 1. Commonweath Agric. Bureaux, Slough, England.
5. Ørskov, E.R., MacLeod, N.A., Fahmy, S.T.M., Istasse, L. and Hovell. F.D.DeB. 1983. Investigation of nitrogen balance in dairy cows and steers nourished by intragstric infusion. Effect of submaintenance energy input with or without protein. Br. J. Nutr. 50, 99-107.
6. Hovell. F.D.DeB., Asplund, J.M., Ørskov, E.R., Ku Vera, J. and MacLeod, N.A. 1984. Basal and fasting N losses, and endogenous flow of nitrogen of ruminants nourished by intragastric infusion. Can. J. Anim. Sci. 64, 143-152.
7. Ku Vera, J., MacLeod, N.A. and Ørskov, E.R. 1989. energy exchanges of acttle nourished by intragastric infusion of nutrients. In: In: “Energy Metabolism of Farm Animals”. Y. van der Honeny and W.H. Close (eds). pp. 271-274. Proc. 11th Symp. Lunteren (EAAP 43), Pudoc, Wageningen.
8. Ørskov, E.R., Meehan, D.E., MacLeod, N.A., and Kyle, D.J. 1999. Effect of glucose supply on fasting N excretion and effect of level and type of volatile fatty axít infusion on response to protein infusion in cattle. Br. J. Nutr. 81, 389-393.
9. Ørskov, E.R., Grubb, D.A. and MacLeod, N.A. 1983. Basal urinary nitrogen excretion and growth response to supplemental protein by lambs close to energy equilibrium. Br. J. Nutr. 50, 173-187.
10. Hovell. F.D.DeB., Ørskov, E.R., Kyle, D.J. and MacLeod, N.A. 1987.

134
Undernutrition in sheep. Nitrogen repletion by N-depleted sheep. Br. J. Nutr. 57, 77-88.
11. Ku Vera, J., Ørskov, E.R., Hovell. F.D.DeB., and MacLeod, N.A. 1987. Effect of small amounts of glucose and different volatile fatty axít mixtures on fasting N excretion in lambs. 5th International Symposium on Protein Metabolism and Nutrition. p. 94. Rostock, German Democratic Republic.
12. Chowdhury, S.A., Ørskov, E.R., Hovell. F.D.DeB., Scaife, J.R. and Mollison, G. 1997. Protein utilization during energy undernutrition in sheep sustained by intragastric infusion: effects of protein infusion level, with or without sub-maintenance amounts of energy from volatile fatty axíts, on energy and protein metabolism. Br. J. Nutr., 77, (4), 565-576.
13. Chowdhury, S.A., Ørskov, E.R., Hovell. F.D.DeB., Scaife, J.R. and Mollison, G. 1997. Protein utilization during energy undernutrition in sheep sustained by intrgastric infusion. Effect of body fatness on the protein metabolism of energy-restricted sheep. Br. J. Nutr., 78, No. 2, 273-282.

135
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ Hướng nghiên cứu 9.3. Bài xuất nitơ nội sinh trong các phần khác nhau đường tiêu hóa
Việc chúng tôi sẽ tiến hành xác định nitơ nội sinh gần như chắc chắn phải xảy ra vì chúng tôi đã có kỹ thuật loại trừ nitơ khẩu phần và nitơ vi sinh vật, việc loại trừ này đã từng khó tiến hành ở gia súc nhai lại. Dĩ nhiên có tranh luận rằng nitơ nội sinh thoát xuống phía dưới đường tiêu hóa sẽ khác nếu gia súc được cho ăn khẩu phàn theo cách thông thường. Tuy nhiên Bernard Fell đã chỉ ra rằng chỉ số phân chia sinh trưởng của tế bào biểu mô dạ cỏ của gia súc được nuôi bằng phương pháp bơm dinh dưỡng trực tiếp vào dạ dày cũng giống như ở gia súc nuôi bình thường. Chúng tôi cũng đã xem xét thành phần của axít amin của nitơ phi ammonia tách ra từ dịch dạ cỏ và dịch dạ múi khế. Thành phần này gần tương tự như thành phàn axít amin của cơ thể, ngoại trừ cystine là 2,1 đến 2,6 g/16 g nitơ trong khi trong cơ thể tỷ lệ này là 1,0g/ 16 g nitơ, có thể do bản chất “sừng hóa” của tế bào biểu mô dạ cỏ. Cũng thấy có một lượng nhỏ axít diaminopilemic từ hệ vi sinh vật khu trú trong vách dạ cỏ. Khoảng 80% nitơ nội sinh chứa AA-N (1). Với hầu hết các thí nghiệm, nitơ thoát xuống được xác định ở gia súc được nuôi bằng cách bơm axít béo bay hơi vào dạ dày đủ để cuung cấp năng lượng cho duy trì hay 450 KJ/kgW0,75.
Kết quả chính từ một số thí nghiệm với giá trị trung bình được trình bày tóm tắt trong Bảng 9.4. Như mong đợi, có sự biến động đáng kể, tuy nhiên nitơ nội sinh dạ cỏ đóng góp vào phần thoát xuống dạ múi khế khoảng 50%. Nitơ phi ammonia nội sinh có nguồn gốc từ dạ cỏ chủ yếu từ tế bào biểu mô bong ra là từ 50 đến 80 mg nitơ/kg W0,75 trong khi lượng nitơ thoát xuóng từ dạ múi khế dao động từ 130 đến 190 mg do có sự bổ sung vào của enzyme tiêu hóa. Chúng tôi chỉ có một giá trị trung bình có được ở dịch hồi tràng cừu, giá trị này là 54 mg nitơ/kg W0,75, điều này cho thấy khoảng 2/3 lượng nitơ nội sinh từ dạ múi khế được tiêu hóa trong ruột non, đóng góp cho quá trình hấp thu axít amin. Bảng 9.4. Nitơ phi ammonia (g nitơ/kg W0,75) thoát xuống từ dạ cỏ, dạ múi khế, hồi tràng và xuất hiện trong phân, uớc tính bằng nồng độ x lượng dịch thoát xuống. Loài gia súc Dạ
cỏ Dạ múi khế Hồi tràng Phân Tài liệu tham
khảo Bò cái 66 ND ND ND 2 Bò đực thiến 66 ND ND ND 2 Bò đực thiến 86 130 ND ND 3 Bò cái 66 130 ND 25 4 Bò đực thiến 70 131 ND 21 4 Bò cái 54 ND ND ND 5

136
Cừu 85 195 ND ND 5 Cừu 76 181 ND ND 5 Cừu ND ND 54 ND 6 ND = không xác định
Chúng tôi cũng có một vài số liệu về nitơ phân ở đây từ các thí nghiệm được thiết kế để xác định lượng thoát xuống. Một lý do là gia súc không thải phân ra hàng ngày nhưng chúng tôi có nhiều quan sát về lượng nitơ phân bài xuất từ cá thí nghiệm xác định bài xuất nitơ cơ bản và chúng tôi thấy nitơ phân thường chiếm khoảng 10% nitơ cơ bản bài xuất ở khẩu phần không có nitơ, có nghĩa là từ 25 đến 35 mg nitơ/kgW0,75. Lượng nhỏ này có thể được coi là nitơ phân trao đổi thực.
Một quan sát cho thấy có mối tương quan dương giữa pH dạ múi khế và lượng thoát xuống của nitơ dạ múi khế; điều này có nghĩa là pH dạ múi khế càng kiềm, lượng nitơ nội sinh bài thải càng lớn.
Vậy áp dụng các kết quả nghiên cứu ở đây là gì? Một ứng dụng ở đây là: không thể có tình trạng hoàn toàn không có protein vì dạ cỏ và chất tiết dạ múi khế hầu hết là nguồn axít amin. Cũng có giá trị khi biết rằng khoảng một nửa nitơ nội sinh từ dạ múi khế có thể có nguồn gốc từ dạ cỏ nhưng cũng có thể phần tế bào biểu mô bong ra từ dạ cỏ sẽ được phân giải bởi vi sinh vật trong dạ cỏ trong các điều kiện nuôi dưỡng bình thường.
Tài liệu tham khảo cho phần 9.3 1. Ørskov, E.R., MacLeod, N.A and Kyle, D.J. 1986. Flow of nitrogen from
the rumen and abomasum in cattle and sheep given protein-free nutrients by intragastric infusion. Br. J. Nutr. 56, 241-248.
2. Ørskov, E.R. and MacLeod, N.A. 1982. The flow of N from the rumen of cows and steers maintained on intra-ruminal infusion of volatile fatty axíts. Proc. Nutr. Soc. 41, 76A.
3. Ørskov, E.R. and MacLeod, N.A. 1982. Flow of endogenous N from the rumen and abomasum of cattle given protein free nutrients. Proc. Nutr. Soc. 42, 61A.
4. Hovell. F.D.DeB., Asplund, J.M., Ørskov, E.R., Ku Vera, J. and MacLeod, N.A. 1984. Basal and fasting N losses, and endogenous flow of nitrogen of ruminants nourished by intragastric infusion. Can. J. Anim. Sci. 64, 143-152.
5. Ørskov, E.R., MacLeod, N.A and Kyle, D.J. 1986. Flow of nitrogen from the rumen and abomasum in cattle and sheep given protein-free nutrients by intragastric infusion. Br. J. Nutr. 56, 241-248.
6. Storm, E., Brown, D.S. and Ørskov, E.R. 1983. The nutritive value of rumen micro-organisms in ruminants. 3. The digestion of microbial amino and nucleic axíts in, and losses of endogenous nitrogen from, the small intestine of sheep. Br. J. Nutr. 50, 479-485.

137
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ Hướng nghiên cứu 9.4. Ảnh hưởng của nhịn đói đến trao đổi năng lượng, protein và nhu cầu glucose
Thâm nhập vào lĩnh vực trao đổi chất lúc đói chắc chắn có thể thực hiện được vì chúng ta có khả năng nuôi gia súc ở trạng thái duy trì bằng kỹ thuật bơm dinh dưỡng trực tiếp vào dạ dày. Bất chợt, chúng tôi đã có thể bắt đầu xem xét dinh dưỡng của gia súc vật chủ ở dạng hoàn hảo không bị phức tạp hóa bởi ảnh hưởng của lên men vi sinh vật trong dạ cỏ, ảnh hưởng này ở một vài khía cạnh đã phủ mây đen lên sự hiểu biết của chúng ta về dinh dưỡng gia súc nhai lại. Với sự giúp đỡ của các sinh viên và các nhà nghiên cứu đến trao đổi và làm việc tại Viện, chúng tôi bắt đầu tập trung một cách có hệ thống vào một số vấn đề mà kỹ thuật bơm dinh dưỡng trực tiếp vào dạ dày có thể giúp chúng tôi giải quyết được. Trao đổi đói và bài xuất nitơ
Có thể không dễ dàng hiểu được tại sao trao đổi đói có thể được nghiên cứu bằng cách sử dụng kỹ thuật bơm dinh dưỡng trực tiếp vào dạ dày. Tuy nhiên kỹ thuật này cho phép có thể đạt được trạng thái sau hấp thu một cách nhanh chóng, trong một vài giờ, trong khi gia súc được nuôi bình thường phải mất vài ngày để đạt tới trạng thái đó.
Quan sát đầu tiên của chúng tôi liên quan đến sự tăng đáng kể bài xuất nitơ khi nhịn đói, khác biệt nhiều với bài xuất nitơ cơ bản (có nghĩa là bài xuất ở mức dinh dưỡng duy trì). Tóm tắt một số giá trị trung bình được trình bày trong Bảng 9.5, cùng với kết quả bài xuất nitơ cơ bản khi có đo đạc. Trong khi giá trị trung bình chung cho bài xuất nitơ cơ bản là 307 mg nitơ/kgW0,75/ngày với cừu và 341 với bò, sự khác nhau của các giá trị trung bình khi nhịn đói là lớn hơn nhiều, 438 và 608 lần lượt cho cừu và bò. Nói một cách khác, nitơ bài xuất lúc đói cao hơn khoảng 50% so với nitơ bài xuất cơ bản ở cừu, nhưng với bò cao hơn tới 75%. Năm 1982, trong một cuộc hội thảo về trao đổi năng lượng ở Nauy (8) tôi đã tranh luận rằng chúng ta không nên sử dụng nhiệt sinh ra trong trao đổi đói làm cơ sở để năng lượng nữa. Một mặt tôi đồng ý với các nhà khoa học khác đã phê phán cơ sở này như John Webster nhưng ngoài ra còn vì những lý do khác. Tôi tranh luận rằng gia súc bị đói thực tế có khẩu phần rất thiếu hụt protein với mỡ nội sinh là nguồn năng lượng và protein nội sinh là nguồn tiền chất của glucose, khẩu phần này làm cho lượng nitơ bài xuất cao. Theo tôi, có vẻ không logíc khi một khẩu phần thiếu hụt được sử dụng làm cơ sở cho việc xác định các giá trị của khẩu phần cân bằng dinh dưỡng, tôi cho rằng protein bị dị hóa bắt buộc có thể là lý do hợp lý cho sự khác nhau giữa cái được gọi là Km (hiệu quả sử dụng năng lượng dưới mức duy trì) và Kf (hiệu quả sử dụng năng lượng trên mức duy trì). Các quan sát của tôi là đúng nhưng các lý do đưa ra không thật sự đúng! Đói trong thực tế do là khẩu phần thiếu glucose; kết quả là Beta-hydroxybutyrate huyết tương tăng lên, như đã thấy ở tất cả các thí nghiệm (tham khảo trong bảng), và nitơ bài xuất tăng

138
lên vì tiền chất glucose tăng lên trong quá trình dị hóa protein được sử dụng để hạn chế quá trình tái tổng hợp protein. Bảng 9.5. Nitơ bài xuất trong trạng thái trao đổi đói so với nitơ bài xuất cơ bản ở bò và cừu. Các giá trị được thể hiện theo mg/kgW0,75.
Loài gia súc Nhịn đói Cơ bản Tài liệu tham khảoCừu Cừu 511 386 1 Cừu 570 327 2 Cừu 325 250 2 Cừu 372 224 2 Cừu 479 320 3 Cừu 422 269 3 Cừu 423 264 4 Cừu 408 249 4 Cừu 438 286 5 Bò Bò 617 409 1 Bò 642 345 5 Bò 616 365 6 Bò 633 - 3 Bò 529 265 7 Bò 664 - * Ku Vera Bò 546 352 * Ku Vera Bò 620 332 * Ku Vera Bò 616 354 * Ku Vera
*J.C. Ku Vera, trao đổi năng lượng và nitơ ở bò đợc nuôi bằng bơm dinh dưỡng trực tiếp vào dạ dày. Luận án tiến sĩ, Trường đại học Aberdeen.
Các ứng dụng cho năng lượng trao đổi
Kết quả cả ở bò (6) và ở cừu (9) đều cho thấy rằng phần năng lượng đầu tiên bơm vào dạ dầy gia súc nhịn đói thực tế đã làm giảm sinh nhiệt khoảng 10%. Đây là kết quả gây ngạc nhiên vì từ các quan sát trước đây trong các nghiên cứu với axít béo bay hơi chúng tôi mong đợi có ít nhất 40% năng lượng của axít béo bay hơi bị tiêu hao dưới dạng nhiệt. Sự giảm này là rất rõ và chỉ dừng lại khi nitơ bài xuất và Beta-hydroxybutyrate trở lại mức bình thường. Năng lượng đưa vào phải ít nhất một phần là các chất có thể sinh glucose (gluconeogenic) như hỗn hợp axít béo bay hơi có propionate và thậm chí có casein. Axít acetic và mỡ không làm giảm đáng kể nitơ bài xuất khi đói (10). Điều này, đương nhiên có nghĩa là nhiệt sản xuất đói là môt cách đọc dẫn đến các sai lầm, nó làm tăng sự khác biệt giữa hiệu quả trên và dưới mức duy trì. Tiêu tốn thêm lượng nitơ bài xuất có thể được lý giải một phần sự khác nhau trong sinh nhiệt (9). Nếu gia súc được ăn khoảng 20

139
đến 30% năng lượng cần cho cân bằng năng lượng (duy trì) khi đó nhu cầu glucose cơ bản được đáp ứng, Beta-hydroxybutyrate huyết thanh bình thường, và không có sự khác nhau về hiệu quả sử dụng năng lượng ở trên và dưới mức duy trì có thể được ghi lại trong phạm vi các mức nuôi dưỡng có thế đạt được ở gia súc nhai lại, kết quả là hiệu quả sử dụng năng lượng dự trữ trong cơ thể hiệu quả hơn.
Bây giờ chúng tôi đã có thể xác định nhu cầu glucose cho duy trì hay chính xác hơn là nhu cầu cho tiền chất glucose.
Chúng tôi đã tiến hành xác định nhu cầu này ở bò năm 1989 (6) và sau đó năm 1999 (7). Kết quả được trình bày trong Bảng 9.6.
Bảng 9.6. Ảnh hưởng của việc bơm glucose vào dạ múi khế của bò nhịn đói đến bài xuất nitơ.
Ku Vera và cộng sự., 1989 (6) Orskov và cộng sự., 1999 (7) Glucose bơm vào
(g/kgW0,75) N bài xuất
(mg/kgW0,75) Glucose bơm vào
(g/kgW0,75) N bài xuất
(mg/kgW0,75) 0 620 0 529
0,6 541 1,5 494 1,3 401 2,5 364 2,4 366 3,5 329 4,8 332 4,5 300 14,2 303 5,5 261 28,5 278 6,5 268
Ở đây chúng ta thấy nhu cầu cho bò là khoảng 5g glucose hoặc glucose
đương lượng/kgW0,75/ngày, ở mức này nitơ bài xuất giảm xuống mức cơ bản. Tuy nhiên, quan sát của chúng tôi với cừu lại khác, chúng tôi phát hiện ra nhu cầu cho glucose ở cừu chỉ là 1 đến 2 g/ kgW0,75(2, 9). Điều này thể hiện sự khác nhau thú vị nữa giữa cừu và bò. Đây có thể chính là lý do tại sao bài xuất nitơ lúc đói dường như khá ổn định và cao hơn đáng kể ở bò so với ở cừu, như đã mô tả trước đó. Sẽ là rất thú vị khi nghiên cứu xem khía cạnh này của trao đổi chất có khác nhau không giữa bò được chọn lọc cho năng suất cao và lượng ăn vào nhiều và gia súc bản địa đã thích nghi do chọn lọc tự nhiên để có thể tồn tại với khẩu phần chủ yếu là thức ăn thô.
Kết quả này đưa đến các kết luậnsau: những quan sát từ các nghiên cứu của chúng tôi về trao đổi đói cần phải được đưa vào các hệ thống nuôi dưỡng không chắc chắn, hay các hệ thống dựa trên trao đổi chất đói để không nên áp dụng nữa. Các kết quả làm nổi bật sự khác nhau giữa bò và cừu cần nhiều nghiên cứu hơn nữa.
Tài liệu tham khảo cho phần 9.4 1. Hovell. F.D.DeB., Asplund, J.M., Ørskov, E.R., Ku Vera, J. and MacLeod,
N.A. 1984. Basal and fasting N losses, and endogenous flow of nitrogen of

140
ruminants nourished by intragastric infusion. Can. J. Anim. Sci. 64, 143-152.
2. Ku Vera, J., Ørskov, E.R., Hovell. F.D.DeB., and MacLeod, N.A. 1987. Effect of small amounts of glucose and different volatile fatty axít mixtures on fasting N excretion in lambs. 5th International Symposium on Protein Metabolism and Nutrition. p. 94. Rostock, German Democratic Republic.
3. Chowdhury, S.A., Ørskov, E.R., Hovell. F.D.DeB., Scaife, J.R. and Mollison, G. 1997. Protein utilization during energy undernutrition in sheep sustained by intragastric infusion: effects of protein infusion level, with or without sub-maintenance amounts of energy from volatile fatty axíts, on energy and protein metabolism. Br. J. Nutr., 77, (4), 565-576.
4. Chowdhury, S.A., Ørskov, E.R., Hovell. F.D.DeB., Scaife, J.R. and Mollison, G. 1997. Protein utilization during energy undernutrition in sheep sustained by intrgastric infusion. Effect of body fatness on the protein metabolism of energy-restricted sheep. Br. J. Nutr., 78, No. 2, 273-282.
5. Ørskov, E.R., MacLeod, N.A., Fahmy, S.T.M., Istasse, L. and Hovell. F.D.DeB. 1983. Investigation of nitrogen balance in dairy cows and steers nourished by intragstric infusion. Effect of submaintenance energy input with or without protein. Br. J. Nutr. 50, 99-107.
6. Ku Vera, J., MacLeod, N.A. and Ørskov, E.R. 1989. energy exchanges of acttle nourished by intragastric infusion of nutrients. In: In: “Energy Metabolism of Farm Animals”. Y. van der Honeny and W.H. Close (eds). pp. 271-274. Proc. 11th Symp. Lunteren (EAAP 43), Pudoc, Wageningen.
7. Ørskov, E.R., Meehan, D.E., MacLeod, N.A., and Kyle, D.J. 1999. Effect of glucose supply on fasting N excretion and effect of level and type of volatile fatty axít infusion on response to protein infusion in cattle. Br. J. Nutr. 81, 389-393.
8. Ørskov, E.R. 1982. Maintenance and growth in ruminants. Introductory commnets. In Energy Metabolism of Farm Animals. Eds. Ekern, A. and Sundstoll, F. pp 141-146. The Agricultural University of Warsaw Aas.
9. Chowdhury, S.A., and Ørskov, E.R. 1994. Implications of fasting on the energy metabolism and feed evaluation in ruminants. J. of Anim. and Feed Sci. 3, 161-169.
10. Asplund, J.M., Ørskov, E.R., Hovell. F.D.DeB. and MacLeod, N.A. 1985. The effect of intragastric infusion of glucose, lipids or acetate on fasting of N excretial and blood metabolites in sheep. Br. J. Nutr. 54, 189-195.

141
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ Hướng nghiên cứu 9.5. Sử dụng protein vi sinh vật và thành phần axít amin tối ưu cho sinh trưởng và sản xuất sữa
Lĩnh vực nghiên cứu này không được dự tính trước nhưng chúng tôi đã tiến hành nhiều nghiên cứu về nó! Một sinh viên trẻ thông minh có khiếu về cơ khí máy móc, Ebbe Storm, đã đến chỗ tôi để làm nghiên cứu sinh. Anh ta ôn tồn, hơi ngang bướng nhưng anh ta muốn đạt học vị tiến sĩ. Chúng tôi thảo luận xem anh ta sẽ làm gì và một câu hỏi hợp lý được đặt ra là: tại sao lại bơm casein trực tiếp vào dạ dầy trong khi gia súc nhai lại thực sự sử dụng protein vi sinh vật. Tôi đã chỉ ra rằng chưa ai từng tách được khoảng hơn 100g protein vi sinh vật để làm các thí nghiệm trên chuột. Không thể tách đủ protein vi sinh vật để bơm vào dạ dầy khi nhu cầu hàng ngày của cừu là 100g. Đây là một thách thức cho Ebbe Storm; nếu nó là điều không thể và sau trở thành điều có thể với anh ta thì sẽ rất tốt! Ebbe Storm đã đến thăm lò giết mổ lợn ở địa phơng và từ catalogue các sản phẩm của hãng Sharp và anh ta đã thấy rằng họ có một chiếc máy ly tâm lớn dùng để phân tách huyết tthanh. Ebbe Storm đã nói chuyện với các kỹ sư ở đây và biết rằng máy ly tâm hiện không xử dụng. Ebbe Storm đã mượn chiếc máy đó, thuê cần cẩu và vận chuyển máy đến cạnh lò giết mổ gia súc nhai lại. Ở lò mổ, chất chứa dạ cỏ được bỏ đi qua một cái máng xuống xe móc ở dưới tầng hầm. Anh ta đã thuyết phục được người quản lý cho lắp một chiếc vòi ở đáy của xe móc; vì như đã quan sát ở dạ cỏ, dịch dạ cỏ đọng lại ở đáy xe móc. Dịch dạ cỏ sau đó được lọc và cho đi qua một loạt các máy ly tâm cũng ở tầng hầm. Anh ta dùng điện 3 pha cho các máy ly tâm do đó chẳng mấy chốc đã tách được hơn 1kg/ngày chất khô vi sinh vật khá thuần khiết thích hợp cho thí nghiệm bơm protein vi sinh vât vào dạ dầy. mỗi ngày một phần protein vi sinh vật tách được được giữ lạnh ở -200C, đôi khi được cất trong tủ lạnh của anh ta, tất nhiên vợ anh ta không hài lòng! Chỉ trong vài tháng anh ta đã sản xuất ra hơn 70kg chất khô protein vi sinh vât (1). Thật đáng sợ!
Tôi chỉ được biết khi hệ thống đã hoạt động. Anh ta biến mất, làm tôi rất bực mình, khoảng 1 tuần sau anh ta gọi điện cho tôi từ lò mổ và nói: ông đến mà xem. Quá trình sản xuất được tiến hành tốt trước khi tin tức đến Viện. Sau đó có nhiều câu hỏi về vấn đề bảo hiểm - điều tôi chưa hề nghĩ tới. May mắn thay vào thời điểm đó, chúng tôi đã sản xuất đủ lượng protein vi sinh vật chúng tôi cần, nếu không chúng tôi có thể bị bắt buộc phải dừng lại!
Nitơ vi sinh vật chứa 15,4% DNA và RNA-N; 80,9% nitơ axít amin (AA-N) và phần còn lại là hỗn hợp nitơ chưa biết. Thành phần axít aim của protein vi sinh vật chúng tôi sản xuất tương tự như các số liệu đã công bố.
Bây giờ, lần đầu tiên, chúng tôi đã có thể nghiên cứu cách nghiêm túc sử dụng nitơ vi sinh vật ở gia súc nhai lại! Cừu đã được bơm vào dạ múi khế lượng nitơ vi sinh vật từ 0 đến 2500 mg/kgW0,75/ngày. Chúng tôi cũng lắp canula ở đoạn cuối hồi tràng do đó có thể xác định tỷ lệ tiêu hóa trong ruột non.

142
Các kết quả quan trọng nhất (1, 2 và 3) là:
Lượng nitơ vi sinh vật tích lũy 0,54 Nitơ axít tamin trong nitơ của vi sinh vật 0,80 Tỷ lệ tiêu hóa nitơ axít amin của protein vi sinh vật trong ruột non
0,85
Lượng tích lũy của nitơ axít amin vi sinh vật đã hấp thu 0,80 Một số axít amin có tỷ lệ tiêu hóa thấp hơn so với axít amin khác; như
diaminopimelic axít (0,37), histidine (0,68) và cysteine (0,73). Nghiên cứu này thực sự là một nhiệm vụ khổng lồ. Các kết quả có được
thật không dễ dàng và chưa hề được ai đó lặp lại cho đến thời điểm này! Tôi mong muốn cám ơn các nhân viên làm việc tại các lò mổ rất nhiều! Tôi chưa bao giờ có thể hiểu được tại sao chúng tôi có thể chịu đựng được khi phải làm việc ở dưới tầng hầm, nhưng đã khám phá ra rằng cài vòi lắp vào xe móc được sử dụng khi chúng tôi cũng không cần nó! Cái vòi làm cho nhiều chất rắn hơn cần được loại bỏ từ xe móc!
Thành phần amino axít tối ưu cho tích lũy
Hiệu quả sử dụng axít amin hấp thu có nguồn gốc vi sinh vật cho nitơ tích lũy là rất cao 80%; nhưng về mặt lý thuyết nếu thành phần axít amin là tối ưu, tỷ lệ này có thể đạt tới 100%. Ebbe Storm cũng muốn xác định các axít amin hạn chế. Những thí nghiệm như vậy chưa từng được tiến hành ở gia súc nhai lại. Có một điều lạ là các axít amin chứa sulphur như methionine là axít hạn chế đầu tiên đối với sản xuất thịt nhưng chưa từng được ai chứng minh cả.
Các phương pháp truyền thống xác định tỷ lệ amino axít tối ưu tốn nhiều thời gian để tiến hành và kết quả cuối cùng không thỏa đáng.
Thủ tục thông thường sẽ là: 1. Xác định sử dụng amino axít hấp thu từ nguồn protein cần nghiên cứu bằng
cách đo cân bằng nitơ. 2. Bổ sung bằng cách bơm vào dạ dầy hoặc bổ xung vào khẩu phần từng loại
trong 13 axít amin quan trọng để xác định axít amin nào cho đáp ứng với nitơ tích lũy.
3. Xác định mức độ tối ưu của axít amin hạn chế này bằng cách mô tả đáp ứng với các liều bổ sung khác nhau.
4. Xác định axít amin hạn chế thứ hai bằng cách giữ axít amin hạn chế thứ nhất ở mức độ tối ưu đã được xác định.
5. Xác định mức tối ưu của axít amin hạn chế thứ hai bằng cách mô tả đường cong (hay đồ thị) đáp ứng.
6. Sau đó xác định axít amin hạn chế thứ ba bằng cách tương tự. Một vấn đề xảy ra là nếu 2 axít amin hạn chế ở cùng một thời điểm. Phương pháp sẽ không cho bạn đi xa hơn nữa.

143
Ebbe Storm quan tâm đến một cách tiếp cận khác. Thay vì đặt câu hỏi axít amin nào là hạn chế nhất, anh ta đặt câu hỏi axít amin nào dư thừa nhất. Anh ta đã dùng phương pháp đã được sử dụng từ trước nhưng không phải theo cách như mô tả ở trên. Chúng ta đã xác định rằng sử dụng axít amin hấp thu từ vi sinh vật dạ cỏ là 0,80. Hiệu quả này có thể tăng lên gần 1,00 nếu thành phần axít amin là tối ưu, do đó lượng nitơ tích lũy tăng thêm 25%: (1,0 - 0,8) : 0,8 x 100 = 25%
Nitơ tích lũy dĩ nhiên có thể tăng thêm 25% nếu thêm 25% proteinvi sinh vật nguyên bản vì nó sẽ cung cấp thêm 25% các axít amin không thay thế đang bị giới hạn.
Chúng tôi đã chấp nhận phương pháp này, bơm protein vi sinh vật theo mức định trước (Ro) và sau đó bơm thêm 25% để đạt mức R1. Tiếp theo, thay vì bổ sung thêm 25% protein vi sinh vật, chúng tôi bơm hỗn hợp axít amin tinh khiết có thành phần tương tự thành phần axít amin đã hấp thu của protein vi sinh vật và xem nếu axít amin tinh khiết vẫn tuân theo quan hệ hồi qui tuyến tính như chúng tôi đã thấy khi dùng protein vi sinh vật, và quan hệ vẫn đúng là quan hệ hồi qui tuyến tính. Tiếp theo chúng tôi loại bỏ từng axít amin một theo thứ tự. Khi axít amin hạn chế đầu tiên được loại bỏ, nitơ tích lũy sẽ là Ro, vì giá trị tích lũy 0,8 đã được xác định cho axít amin hạn chế đầu tiên. Điều quan trọng hơn, khi loại bỏ từng axít amin theo thứ tự, đáp ứng với nitơ tích lũy sẽ chỉ ra cách chính xác điểm mà tại điểm đó axít amin trở nên hạn chế. Nếu không có đáp ứng, axít amin này không phải là axít amin hạn chế. Bây giờ chúng tôi đã có thể tính toán quang phổ tối ưu của axít amin hấp thu cần thiết để có giá trị tích lũy nitơ của axít amin hấp thu gần 1,0. Chỉ có 4 axít amin đáp ứng đáng kể và chúng được trình bày trong bảng dưới đây (Bảng 9.7). Với cừu methionine là axít amin hạn chế đầu tiên (1,0) sau đó là lysine (0,73) histidine (0,32) và arginine (0,26).
Sau đó Diane Fraser tiến hành thí nghiệm tương tự với bò sữa (6). Tuy nhiên ở đây, chúng tôi đã phải quay trở lại với casein vì chúng tôi đã dùng hết protein vi sinh vật! Tuy nhiên quang phổ hấp thu tối ưu dĩ nhiên tương tự dù casein hay protein vi sinh vật được sử dụng làm protein cơ bản. Hình như histidine quan trọng hơn với bò sữa so với cừu.
Bảng 9.7. Tỷ lệ axít amin hấp thu tối ưu (g axít amin/100g axít amin) của các axít amin hạn chế trong protein vi sinh vật ở cừu (4) và bò sữa (5) Cystein +
methionine Lysine Arginine Histidine
Cừu 4,3 9,9 5,5 1,5 Bò sữa 4,1 9,5 4,7 2,2
Sự kiện methionine là axít amin hạn chế đầu tiên, dĩ nhiên, không có nghĩa
là thành phần methionine phải lớn nhất trong món cocktail bổ sung axít amin, vì dựa trên thành phần bổ sung ban đầu; tỷ lệ theo khối lượng của lysine lớn hơn nhiều, như thấy trong Bảng 9.8.

144
Bảng 9.8. Tính toán thành phần khối lượng trong cocktail (dung dịch axít amin bơm vào dạ dầy) bổ sung axít amin axít amin cho cừu. Cystein +
methionineLysine Arginine Histidine
Axít amin tối ưu từ lượng axít amin hấp thu (g/100 g axít amin)
2,80 7,20 4,88 1,29
Thành phần axít amin trong dung dịch axít amin bơm vào dạ dầy
0,87 2,10 1,30 0,35
Thành phần axít amin hạn chế 1,00 0,73 0,26 0,32 Thành phần axít amin trong dung dịch axít amin bơm vào dạ dầy tối ưu
0,87 1,53 0,33 0,11
Phần trăm (%) theo khối lượng trong dung dịch axít amin bơm vào dạ dầy
30,6 53,8 11,6 3,9
Kết quả ở đây có hậu quả logíc là bổ sung axít amin phải quan tâm đến
thành phần axít amin của nguyên liệu cơ sở. Bổ sung axít amin quá nhiều sẽ không kinh tế và không làm tăng năng suất. Cũng đã thấy rằng bổ xung một mình methionine chỉ cho một đáp ứng nhỏ. Để bổ xung có hiệu quả thành phần axít amin bổ sung phải có đủ lượng hợp lý của mỗi axít amin hạn chế. Vẫn còn có cơ hội để thu được các ước lượng chính xác hơn về thành phần axít amin bổ sung, phương pháp được chúng tôi phát triển cho cung cấp khả năng xác định quang phổ axít amin tối ưu cho các mục đích sản xuất khác nhau với tiêu tốn ít lao động hơn và chính xác hơn so với các phương pháp xác định axít amin hạn chế trước đó.
Tài liệu tham khảo cho phần 9.5 1. Storm, E., and Ørskov, E.R. 1983. The nutritive value of rumen mocro-
organisms in ruminants. 1. Large-scale isolation and chemical composition of rumen micro-organisms. Br. J. Nutr. 50, 463-470.
2. Storm, E., Brown, Ørskov, E.R. and Smart, R. 1983. The nutritive value of rumen mocro-organisms in ruminants. 2. The apparent digestibility and net utilization of microbial N for growing lambs. Br. J. Nutr. 50, 471-478.
3. Storm, E., Brown, D.S. and Ørskov, E.R. 1983. The nutritive value of rumen mocro-organisms in ruminants. 3. The digestion of microbial amino and nucleic axíts in, and losses of endogenous nitrogen from, the small intestine of sheep. Br. J. Nutr. 50, 479-485.
4. Storm, E., and Ørskov, E.R. 1983. The nutritive value of rumen mocro-organisms in ruminants. 4. The limiting axít amins of microbial protein in growing sheep determined by a new approach. Br. J. Nutr. 52, 613-620.
5. Fraser, D.L., Ørskov, E.R., Whitelaw, F.G. and MacLeod, N.A. 1991. limiting axít amins in dairy cows given casein as the sole source of protein.

145
Livestock Prod. Sci. 28, 235-252. 6. Fraser, D.L., Ørskov, E.R., Whitelaw, F.G. and MacLeod, N.A. 1990. the
efficiency of casein N utilization in dairy cows. Livestock Prod. Sci. 25, 67-78.

146
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ Hướng nghiên cứu 9.6. Sử dụng dẫn xuất purine nước tiểu từ phân giải nucleic axít để xác định lượng protein do vi sinh vật sản xuất
Khi kỹ thuật dinh dưỡng trực tiếp cho dạ dầy đã được thiết lập vững chắc,
chúng tôi mang kỹ thuật này đến hội thảo trao đổi chất dạ cỏ tại INRA, Pháp, năm 1979 và trình bày kỹ thuật này tại phần kỹ thuật do Ebbe Storm là người tổ chức chính. Ở đây chúng tôi gặp Anna Antoniewitz đến từ Balan. Kỹ thuật này thực sự là cái mà cô đang tìm kiếm để xác định bài xuất cơ bản của allantoin ở cừu mà không có sự đóng góp của vi sinh vật khi ở dạ cỏ hoạt động bình thường. Cô ấy đã quay trở về nhà ở Krakov, đưa ra kỹ thuật và thu được các số liệu về bài xuất allantoin. Đưa ra kỹ thuật là một thành công và cô xứng đáng được ca ngợi khi nghiên cứu ở Balan vào thời điểm đó không phải dễ dàng. Chúng tôi đã có rất nhiều thứ để nghiên cứu nhưng vì chúng tôi đã thu nước tiểu từ bò và cừu trong các nghiên cứu sử dụng kỹ thuật dinh dưỡng trực tiếp cho dạ dầy chúng tôi nghĩ rằng chúng tôi có thể cũng xác định purine hoặc đầu tiên xác định allantoin trước. Một sinh viên thạc sĩ: Simba Sibanda đã làm công việc này (1) và ngay lập tức thông báo có những sự khác nhau lớn trong bài xuất dẫn xuất purine (PD) giữa cừu và bò, với bò bài xuất dẫn xuất purine khoảng 4 đến 5 lần (tính trên khối lượng trao đổi) cao hơn ở cừu. Khi bơm protein vi sinh vật vào dạ múi khế, bài xuất dẫn xuất purine tăng tuyến tính với lượng bơm vào.
Đối với những người mới bắt đầu nghiên cứu, tôi phải giải thích ở đây rằng dẫn xuất purine, hypoxanthine, xanthine, axít uric và allantoin là những sản phẩm bài xuất từ oxy hoá purine. Allantoin là sản phẩm cuối cùng. Khả năng sử dụng dẫn xuất purine trong nước tiểu làm chất đánh dấu cho sản xuất protein vi sinh vật đã được J H Topps và R E Elliot (Nature 205, 498) phát hiện năm 1965 nhưng không được sử dụng vì ở thời điểm đó không thể xác định bài xuất nội sinh của dẫn xuất purine sinh ra từ axít nucleic của bản thân vật chủ. Không thể phân biệt giữa dẫn xuất purine nội sinh và dẫn xuất purine của vi sinh vật. Với kỹ thuật dinh dưỡng trực tiếp cho dạ dầy, bây giờ chúng ta có thể nghiên cứu bài xuất nội sinh của dẫn xuất purine từ gia súc vật chủ.
Cuộc rà soát tiếp theo của chúng tôi với dẫn xuất purine là khi Tiến sĩ Fujihara từ Nhật bản đến làm việc ở chỗ chúng tôi trong kỳ nghỉ phép. Chúng tôi đã có một số vấn đề trong việc xác định hypoxanthine, xanthine và axít uric nhưng chúng tôi cũng biết rằng xanthine oxidase và uricase có thể chuyển chúng thành allantoin. Ở cừu một lượng đáng kể và biến động dẫn xuất purine nhưng không pahỉ là allantoin được tìm thấy, trong khi ở bò allantoin chiếm khoảng 90% dẫn xuất purine. Fujihara đã thành công chuyển PD thành allatoin, sự chuyển này làm cho việc xác định dẫn xuất purine đơn giản hơn, và cung cấp bằng chúng rằng rằng casein bơm vào dạ dày đã có ít hoặc không ảnh hưởng đến việc bài xuất dẫn

147
xuất purine (2). Fujihara cũng chứng minh rất rõ ràng rằng: có sự khác nhau trong bài xuất dẫn xuất purine nội sinh ở bò và cừu, bài xuất dẫn xuất purine ở bò cao hơn khoảng 3 lần so với bài xuất này ở cừu do liên quan đến khối lượng trao đổi. Như vậy rõ ràng là nếu muốn dùng dẫn xuất purine làm chất đánh dấu cho sản xuất protein vi sinh vật, cần phát triển những phương trình khác nhau cho bò và cừu.
Những tiến bộ thực sự trong bài xuất dẫn xuất purine đã đạt được khi X.B. Chen - một nghiên cứu sinh tiến sĩ đến từ Trung Quốc. Anh ta đã khảng định rằng sự khác nhau trong bài xuất dẫn xuất purine nội sinh giữa cừu (168 mol/kgW0.75) và bò (564 mol/kgW0.75) (3) là rất lớn và từ các nghiên cứu enzyme anh ta tìm ra lý do hợp lý cho sự khác nhau trong bài xuất dẫn xuất purine nội sinh giữa các loài, đó là sự có mặt của xanthine oxidase (XO) trong tất cả các mô bào của bò nhưng xanthine oxidase (XO) lại chỉ có trong gan của cừu. Nói cách khác khả năng hypoxanthine được chuyển thành xanthine và được bài xuất dưới dạng dẫn xuất purine ở bò cao hơn cừu. Sự hiện diện của XO trong mô của ruột non ở bò cũng cho thấy rằng các sản phẩm được hấp thu của axít nucleic có nguồn gốc từ vi sinh vật không thể được sử dụng trong trao đổi axít nucleic ở bò vì chúng có thể đã đến gan ở dạng đã chuyển thành dẫn xuất purine trong khi ở cừu chúng có thể được sử dụng trong trao đổi axít nucleic (4).
Để xác định tổng số protein vi sinh vật sinh ra trong dạ cỏ, một thí nghiệm lớn được thiết lập trong đó protein vi sinh vật được bơm vào trong dạ múi khế của bò được nuôi bằng kỹ thuật dinh dưỡng trực tiếp cho dạ dầy. Lượng thu hồi trung bình của purine đưa vào dưới dạng xuất purine nước tiểu chỉ là 77%, như vậy hình như có thể một số purine được bài xuất bởi các con đường khác, một trong số đó là nước bọt. Purine bài xuất qua nước bọt có thể chiếm tới 10% lượng purine bị mất, vì allatoin trong dạ cỏ bị phá huỷ một cách nhanh chóng bởi vi sinh vật và không thoát ra khỏi dạ cỏ với số lượng lớn (5).
Một vấn đề thực tế với phương pháp dẫn xuất purine là cần thiết phải thu toàn bộ nước tiểu và do đó khả năng sử dụng phương pháp lấy mẫu điểm (một phần) với creatinine là chất đánh dấu nội sinh cần được đánh giá (6, 7). Lấy mẫu điểm dĩ nhiên giảm độ chính xác vì bài xuất reatinine thay đổi giữa các gia súc. Phương pháp lấy mẫu điểm đã hoạt đọnh tốt khi gia súc được cho ăn một hoặc một vài bữa/ngày. Khả năng khác là sử dụng nồng độ dẫn xuất purine huyết thanh để nghiên cứu, tuy nhiên sự thái hóa nhanh chóng của purine trong thận làm cho việc sử dụng nồng độ dẫn xuất purine huyết thanh để nghiên cứu không chính xác và ít giá trị thực tiễn.
Chúng tôi đã mở rộng các quan sát ở trâu. Cô Samaraweera từ Sri Lanka đã đến vùng tam giác dừa cùng với người cha đã nghỉ hưu để thu mẫu nước tiểu từ trâu cái (9). Các mẫu đó đã gây sốc với những giải thích trước đây về sự khác nhau giữa cừu và bò. Trâu có sự bài xuất dẫn xuất purine nội sinh tương tự dẫn xuất purine nội sinh ở cừu trao đổi đói nhưng có một quang phổ xanthine oxidase (XO) của mô bào tương tự với quang phổ XO của mô bào ở bò. Tuy nhiên lượng protein vi sinh vật xác định bằng dẫn xuất purine ở trâu thấp hơn. Câu chuyện này

148
vẫn chưa kết thúc, có thể tốc độ lọc ở cầu thận thấp hơn ở trâu. Lạc đà không bướu và có thể lạc đà nói chung bài xuất purine nội sinh tương tự cừu và có thành phần allatoin và axít uric nước tiểu cũng tương tự nhưng nồng độ axít uric huyết thanh cao hơn nhiều so với các loài khác. Chúng tôi vẫn có những khó khăn ở trâu. Giả thuyết ở thời điểm đó là sự khác nhau về tốc độ lọc của cầu thận giữa các loại gia súc có thể đã cho phép dẫn xuất purine ở lại lâu hơn trong huyết thanh ở một số loài và do đó đã tăng khả năng đi vào dạ cỏ hoặc bộ phận dưới của đường tiêu hóa nơi chúng sẽ bị phân huỷ bởi vi sinh vật. Câu chuyện về purine được theo đuổi trong hợp tác với Cơ quan Năng lợng Nguyên tử Quốc tế tại các phòng thí nghiệm ở Tây Ban Nha, Malaysia, Indonesian, Việt nam, Sri Lanka, Trung Quốc, Venezuela và Thổ Nhĩ Kỳ.
Thông qua lượng bài xuất dẫn xuất purine ghi chép được, kết quả chỉ ra rằng (10) mức nuôi dưỡng cao hơn sẽ có khuynh hướng tăng sản xuất protein vi sinh vật cho mỗi đơn vị thức ăn được lên men. Người ta giả định rằng do nhu càu duy trì vi sinh vật tăng lên khi thời gian thức ăn lưu lại dạ cỏ dài ra.
Sử dụng trong thực tế phương pháp dùng dẫn xuất purine vẫn cần được quyết định. Ví dụ, vì sản xuất protein vi sinh vật chụi ảnh hưởng của lượng thức ăn ăn vào, tỷ lệ purine/creatinine trong mẫu nước tiểu sử dụng pương pháp lấy mẫu điểm có thể dùng như một công cụ ước tính lượng thức ăn ăn vào ở đồng cỏ.
Tài liệu tham khảo cho phần 9.6
1. Sibanda, S., Topps, J.H., Storm, E. and Ørskov, E.R. 1982. The excretion of allantoin by ruminants in relation to protein entering the abomasum. Proc. Nutr. Soc. 41, 75A.
2. Fuhjihara, T., Ørskov, E.R. and Kyle, D.J. 1987. The effect of protein infusion on urinary excretion of purine derivatives in ruminants nourished by intragastric nutrition. J. Agric. Sci. Camb. 109, 7-12.
3. Chen, X.B., Ørskov, E.R. and Hovell, F.D.DeB. 1990. Excretion of purine derivates by ruminants: endogenous excretion, differences between cattle and sheep. Br. J. Nutr. 63, 121-129.
4. Chen, X.B., Hovell, F.D.DeB., Ørskov, E.R. and Brown, D.S. 1990. Excretion of purine derivatives by ruminants: effect of exogenous nucleic axít supply on purine derivate excretion by sheep. Br. J. Nutr. 63, 131-142.
5. Chen, X.B., Hovell, F.D.DeB. and Ørskov, E.R. 1990. Excretion of purine derivatives by ruminants: recycling of allantoin into the rumen via saliva and its fate in the gut. Br. J. Nutr. 63, 197-205.
6. Chen, X.B., Mejia, A.T., Kyle, D.J. and Ørskov, E.R. 1993. Purine derivatives in spot urine samples and plasma of sheep: a possible index of microbial protein supply. Anim. Prod. 56, 420 (abst.).
7. Chen, X.B., Mejia, A.T., Kyle, D.J. and Ørskov, E.R. 1995. Evaluation of the use of the purine derivative: creatinine ratio in spot urine and plasma samples as an index of microbial protein supply in ruminants: studies in sheep. J. of agric. Sci. Camb. 125, 137-143.

149
8. Chen, X.B., Kyle, D.J., Ørskov, E.R. and Hovell, F.D.DeB. 1991. Renal clearance of plasma allantoin in sheep. Exp. Physiol. 76, 59-65.
9. Chen, X.B., Samaraweera, L., Kyle, D.J., Ørskov, E.R. and Abeygunawardene, H. 1996. Urinary excretion of purine derivatives and tissue xanthine oxidase (E.C. 1.2.3.2) activity in buffaloes (Bubalis bubalis) with special reference to differences between buffaloes and Bos taurus cattle. Br. J. Nutr., 75, No. 3, 397-407.
10. Chen, X.B., Chen, Y.K., Franklin, M.F., Ørskov, E.R. and Shand, W.J. 1992. The Effect of Feed Intake and Body Weight on Purine Derivative Excretion and Microbial Protein Supply in Sheep. J. Anim. Sci. 1992. 70, 1543-1552.

150
Hướng nghiên cứu 9. Dinh dưỡng protein cho gia súc vật chủ Hướng nghiên cứu 9.7. Các thí nghiệm khác với dinh dưỡng trực tiếp cho dạ dày liên quan đến tái sử dụng urê, và chất đánh dấu để nghiên cứu tình trạng protein và lượng thức ăn ăn vào Động thái trao đổi chất và cơ chế tái sử dụng urê Có vẻ hơi ngạc nhiên một chút khi nghĩ rằng kỹ thuật dinh dưỡng trực tiếp cho dạ dầy lại có thể dùng để nghiên cứu tái sử dụng urê và động thái trao đổi urê. Whitelaw (1, 2) cho thấy thực sự động thái của dòng urê là như nhau ở gia súc nuôi theo cách thông thường và gia súc được nuôi bằng kỹ thuật dinh dưỡng trực tiếp cho dạ dầy. Khi nồng độ urê huyết tương tăng lên, urê vào ruột tăng lên và nồng độ ammonia dạ cỏ tăng lên ở cừu, cho dù chúng được nuôi dưỡng bình thường hay được nuôi dưỡng bằng kỹ thuật dinh dưỡng trực tiếp cho dạ dầy. Trong thực tế điều này không đáng ngạc ngạc nhiên lắm vì R.J. Wallace (3) đã chỉ ra rằng khu hệ vi sinh vật trong dạ cỏ phân giải urê một cách mạnh mẽ dù gia súc có được cho ăn hay không. Các quần xã vi sinh vật nhất định sinh sống trong vách dạ cỏ bao gồm những vi sinh vật kỵ khí ngẫu nhiên, phân giải urê rất mạnh. Whitelaw cũng cho thấy rằng nếu hoạt động phân giải urê của vi sinh vật dạ cỏ bị loại bỏ, nồng độ urê trong dạ cỏ sẽ tương tự nồng độ urê trong huyết tương.
Urê di chuyển qua vách dạ cỏ rõ ràng tương ứng với nồng độ urê huyết tương. Nghiên cứu ở đâu đó đã cho thấy nồng độ urê trong nước bọt đi vào dạ cỏ cũng tương ứng với nồng độ urê huyết tương. Kiểm soát tích cực hơn việc tái sử dụng urê xảy ra trong thận, thông qua tốc độ lọc của cầu thận và tái hấp thu ở ống tiểu cầu. Tốc độ lọc của cầu thận và tái hấp thu ở ống tiểu cầu có thể khác nhau giữa các giống gia súc và bị ảnh hưởng bởi lượng nước uống vào.
Sự phụ thuộc của urê đi vào dạ cỏ vào nồng độ urê huyết tương thực tế có một số bất lợi. Các khẩu phần có một lượng nitơ quá cao, dẫn đến nồng độ urê cao trong huyết tương, nồng độ urê cao trong huyết tương sẽ làm cho nồng độ urê của dòng urê đi vào và ra khỏi dạ cỏ cũng cao, kết quả là urê có thể được tổng hợp ở gan vài lần trước khi được bài tiết và vì vậy gây tốn kém năng lượng. Điều này cũng có thể có nghĩa là gia súc năng xuất cao phải có tốc độ lọc tiểu cầu cao để chắc chắn nitơ thừ được thải ra càng nhanh càng tốt. Nói một cách khác, gia súc bản địa năng suất thấp với tốc độ lọc tiểu cầu thấp có thể có khả năng tái sử dụng nitơ tốt hơn và cũng chụi đựng tốt hơn với những khẩu phần xơ cao và nitơ thấp. Sự tiết nước bọt
Đo đạc lượng nước bọt bài tiết là cực kỳ khó khăn. Đặt canula tuyến mang tai là rất đơn giản nhưng đặt canula ở một tuyến sẽ có thể được bù lại bằng việc nhai nhiều hơn và do đó bài tiết nước bọt nhiều hơn ở tuyến bên kia. Có thể xác định lượng nước bọt tiết ra một cách gián tiếp hay không? Kỹ thuật dinh dưỡng trực tiếp cho dạ dày có thể giúp giải quyết vấn đề này hay không? Hình như, trái ngược lại, bài tiết nước bọt bị giảm rất nhiều ở những gia súc được nuôi bằng dinh

151
dưỡng trực tiếp cho dạ dày vì việc bài tiết nước bọt liên quan với việc ăn và nhai lại nhưng khi gia súc nuôi bằng dinh dưỡng trực tiếp cho dạ dày không có việc ăn và nhai lại. Một báo cáo đã ghi chép một cố gắng ước tính bài tiết nước bọt từ P nước bọt chảy xuống dạ cỏ, vì ở đây không có sự hấp thu của P từ dạ cỏ. Ở gia súc nuôi bình thường, vấn đề là gần như không thể có một khẩu phần không có P vì thế P trong dạ cỏ có được là từ cả nước bọt và khẩu phần. Nếu chúng ta đưa dinh dưỡng trực tiếp vào dạ dầy để nuôi gia súc và không đưa P vào, thì P chỉ từ một nguồn duy nhất là từ nước bọt và chúng ta có thể đo đươc nồng độ P và do đó ước tính lượng nước bọt tiết ra (S). Chúng ta cũng có thể đo được lượng dịch thu nhận (L), đó là lượng dịch đã bơm vào. Chúng ta cũng có thể xác định dịch lỏng thoát qua từ dạ cỏ (LO) và do đó có thể tính toán lượng nước lưu thông giữa dạ cỏ và máu. Như vậy chúng ta có nước bọt + dịch bơm = dịch lỏng thoát qua từ dạ cỏ + dịch lỏng chảy qua vách dạ cỏ (LF) và LF có thể tính toán được. LF phụ thuộc vào áp suất thẩm thấu của dạ cỏ, và vì vậy có thể tính toán một cách độc lập. khi xác lập được quan hệ giữa dòng chảy và áp suất thẩm thấu. Zhao (4) đã làm một thí nghiệm trong đó áp suất thẩm thấu của dạ cỏ biến động từ 149 đến 338 m.osmol/l và đã xác định được quan hệ giữa dòng chảy và áp suất thẩm thấu gần gióng với phương trình dưới đây:
S + L = LO + LF, trong đó LF chưa biết. Bước tiếp theo bây giờ là tìm phần chưa biết là nước bọt tiết ra ở gia súc
nuôi bình thường. Như vậy ở gia súc bình thường phần không biết là nước bọt tiết ra và phương trình trên bây giờ là: nước bọt tiết ra + nước trong nước uống và thức ăn + nước sinh ra từ sự lên men = dịch lỏng thoát qua từ dạ cỏ + dịch lỏng chảy qua vách dạ cỏ.
S+L = LO + LF, trong đó S chưa biết. Mt sinh viên tiến sĩ, Miroslav Djuric (5)đã làm một thí nghiệm để xác định
S. Trong thí nghiêm này 4 khẩu phần được thiết kế để tạo ra sự khác nhau trong tiết nước bọt do khác nhau ở thời gian ăn và nhai lại. Khẩu phần thí nghiệm bao gồm cỏ khô được nghiền mịn và cỏ khô chặt cùng hai khẩu phần trung gian có tỷ lệ cỏ khô nghiền và cỏ khô chặt tương ứng là 2:1 và 1:2. Kết quả được trình bày ở Bảng 9.9.
Bảng 9.9. Ảnh hưởng cấu trúc khẩu phần đến lượng nước bọt bài tiết tính được ở cừu. Cỏ khô chặt 0 37 67 100 Cỏ khô nghiền mịn 100 67 37 0 Nước tiêu thụ: Nước uống (lít/ngày) 2,79 2,20 2,43 2,35 Nước trong thức ăn (lít/ngày) 0,18 0,18 0,18 0,18 Nước do lên men (lít/ngày) 0,02 0,03 0,03 0,02 Áp suất thẩm thấu dạ cỏ (mosmol/kg nước) 354 349 325 309 Dung tích dạ cỏ (lít) 4,53 5,52 5,17 6,00

152
Tốc độ thoát qua dạ cỏ (lít/ngày) 8,03 9,07 9,67 10,32 Lưu lượng nước, dạ cỏ đến máu - 0,82 - 0,77 0,32 1,28 Bài tiết nước bọt (lít/ngày) 4,22 5,91 7,36 9,05
Dung tích dạ cỏ và tốc độ thoát qua dạ cỏ được xác định bằng dung dịch
các chất đánh dấu. Kết quả hình như rất nhậy và có thể đưa vào phương pháp ước tính tổng lượng nước bọt bài tiết chính xác ở gia súc nuôi bình thường vì dung tích dạ cỏ, áp suất thẩm thấu và tốc độ dịch thoát qua tất cả đề có thể xác định được mà không cần mổ xẻ. Bài xuất các dẫn xuất của axít benzoic như các chất đánh dấu tiềm tàng để tính lượng ăn vào của các thức ăn nhiều xơ và các đặc điểm đặc trưng cho cây bụi
Sở hữu một công cụ nghiên cứu, như chúng ta đã thấy từ trước, đôi khi gợi ý một sự phát triển xa hơn, sự phát triển thêm này chính là cái mà chúng tôi gọi ở đây là các chất đánh dấu từ nước tiểu khác. Chúng tôi đã có máy nghi sắc ký lỏng cao áp do đó chúng tôi có thể phân tích dẫn xuất purine nước tiểu một cách nhanh chóng. X B Chen đã quan sát thấy một đỉnh lớn trong nước tiểu của cừu ăn cỏ nhưng hoàn toàn không có đỉnh này trong nước tiểu cừu nuôi bằng phương pháp dinh dưỡng trực tiếp cho dạ dày và chỉ thấy một đỉnh nhỏ ở cừu ăn lúa mạch. Đỉnh này được xác định là axít hippuric. Điều này cho chúng ta một đầu mối: liệu chúng ta có thể sử dụng đỉnh này để xác định lượng thức ăn thô nhiều xơ ăn vào hay không? Chúng tôi thấy thực sự có mối quan hệ chặt chẽ giữa lượng thức ăn thô nhiều xơ ăn vào và lượng axít hippuric bài xuất, vì axít hipporic là sản phẩm bài xuất của dẫn xuất axít benzzoic sau sự biến đổi bởi vi sinh vật và gia súc vật chủ. Từ những kết quả đã xuất bản của Pagella trong luận văn tiến sĩ của mình và được nhắc lại một phần (6) hình như trong khi tổng số dẫn xuất purine nước tiểu có tương quan chặt chẽ với lượng ăn vào của chất hữu cơ tiêu hoá, vì nó phản ánh lượng ăn vào của carbonhydrate lên men do mối quan hệ giữa việc sản xuất protein vi sinh vật và năng lượng lên men, bài xuất axít hyppiric có thể cho thấy phần nào của chất hữu cơ tiêu hoá có chứa thức ăn thô giầu cellulose tiêu hoá được bởi vì nó xuất phát phần lớn từ các tiền chất lignin. Như vậy cho đến lúc này xem như đã có hai chất đánh dấu từ nước tiểu được tập hợp lại, tuy nhiên cần có nghiên cứu chi tiết hơn.
Đặc điểm đăc trưng cho các cây bụi
Được khuyến khích một lần nữa sau khi tìm ra được chất đánh dấu từ nước tiểu giúp ước tính được lượng thức ăn ăn vào của thức ăn giầu xơ, chúng tôi tiến xa hơn và tìm kiếm chất đánh dấu cho thực vật – các thức ăn của các loài gia súc ăn chồi ngọn (dê). Đây là chủ đề cho luận văn tiến sĩ của Brenda Keir và cho đến lúc này,các nghiên cứu đang tiến triển tốt. Nhiều thực vật cho thấy những quang phổ của các đỉnh trong nước tiểu rất đặc trưng cho từng loại thực vật. Những quang phổ này có thể được sử dụng trong các thí nghiệm nghiên cứu khẩu phần

153
cho con người để xem xét lượng rau quả ăn vào ghi chép được có thực được phản ánh ở nước tiểu hay không. Các chất đánh dấu từ nước tiểu đang trong giai đoạn mới phát triển và trong một vài năm tới nhiều chất đánh dấu từ nước tiểu hơn nữa sẽ được tìm ra vì những lượng rất nhỏ có thể được phát hiện bằng các kỹ thuật như HPLC và quang phổ rộng.
Phát triển thêm về các chất đánh dấu trong dinh dưỡng
Dựa trên kinh nghiệm của chúng tôi về chất đánh dấu trong dinh dưỡng cho protein và ở một phạm vi nào đó là chất đánh dấu trong dinh dưỡng cho năng lượng được cung cấp, ví dụ dẫn xuất purine và dẫn xuất axít benzoic, liệu chúng ta cũng có thể tìm chất đánh dấu cho nhu cầu dinh dưỡng hay không? Chúng tôi vẫn còn vấn đề về xác định nhu cầu protein của gia súc vật chủ, và bằng cách nào nhu cầu protein của gia súc vật chủ bị ảnh hưởng lớn bởi dinh dưỡng giai đoạn trước?. Chúng ta có thể tìm các chất đánh dấu huyết trong huyết thanh hay nước tiểu để giúp đỡ chúng ta hay không? Một lần nữa chúng ta đã có lợi thế của kỹ thuật dinh dưỡng trực tiếp cho dạ dày, kỹ thuật này cho phép chúng ta kiểm soát chính xác mức độ giảm hay đầy đủ. Chúng tôi sớm nhận ra rằng những chất đánh dấu từ nước tiểu ít được sử dụng vì chúng đơn giản chỉ phản ánh tình trạng dinh dưỡng tức thì và không phản ánh được nhu cầu protein.
Chúng tôi quyết định xác định IGF-1 huyết tuơng, fibronectin huyết tương (FN) và tổng số protein huyết tương ở cừu. Chúng tôi sau đó đã làm một thí nghiệm cung cấp thiếu và cung cấp đầy đủ nitơ. FN đáp ứng khá nhạy cảm với sự thay đổi đột ngột trong cung cấp nitơ nhưng không phản ánh đượ tình trạng dinh dưỡng nitơ.
Thực tế IGF-1 cho kết quả gần tương tự. Khi thiếu nitơ xảy ra IGF-1 giảm và khi bắt đầu đủ nitơ IGF-1 tăng ngay lập tức. Một số gia súc được phép tăng lượng nitơ cơ thể tới 33% và ở một nghiệm thức khác chúng mất tới 22% lượng nitơ ban đầu của cơ thể. Thành phần nitơ đầu tiên của cơ thể được tính toán từ tổng số nitơ cơ thể xác định khi giết mổ ở cuối của thí nghiệm, nitơ tăng thêm hoặc mất đi được xác định qua cân bằng nitơ.
Nồng độ protein tổng số của huyết tương (TP) cho đến thời điểm đó là chỉ thị tốt nhất của tình trạng protein. Giá trị của nó được cải thiện nếu thể tích máu được xác định. Trong thời kỳ thiếu nitơ cả thể tích máu và TP giảm.
Shimin Liu (7) thấy ở cừu TP giảm 10g/lít có thể đại diện cho 8% tổng số protein cơ thể mất đi. Tác giả cũng thấy TP nhạy cảm với thiếu protein hơn so với khi protein được cung cấp trở lại. Ở những gia súc đã thiếu protein, TP và dung tích máu thay đổi từ từ về phía các giá trị TP và dung tích máu lúc được ăn đầy đủ
Chúng tôi cần những nghiên cứu chi tiết hơn về TP như là một chất đánh dấu nhưng nhờ sự chính xác để tạo ra đủ hay thiếu protein trong cơ thể với kỹ thuật dinh dưỡng trực tiếp cho dạ dầy, việc đo đạc đơn giản này đã xác định chất đánh dấu mạnh nhất chỉ thị cho tình trạng protein của cơ thể. Tầm quan trọng của tình trạng protein của cơ thể trong xác định nhu cầu protein không thể quá nhấn mạnh. Khối lượng của gia súc và tình trạng sinh lý thực sự thường ít quan trọng

154
hơn tình trạng protein của cơ thể trong phản ánh nhu cầu. Trong thực tế chúng ta vẫn chưa hiểu biết về nhu cầu đâyd đủ. Ví dụ, bằng cách nào một gia súc thiếu protein lại có thể nhanh chóng trở thành đủ protein? Có thể nào bò trong giai đoạn tăng trưởng bù có thể tăng trọng 5kg/ngày nếu có đủ protein đưa vào ruột non? Đơn giản là chúng ta không biết.
Một chút đo đạc thêm nữa về protein huyết tương kết hợp với khối lượng và tình trạng sinh lý của gia súc có thể đưa chúng ta đến gần hơn với việc hiểu biết nhu cầu protein thực và đáp ứng tăng trưởng tiềm năng của gia súc. Hiểu biết nhu cầu protein thực và đáp ứng tăng trưởng tiềm năng của gia súc cho phép kiểm soát chặt chẽ hơn hiệu quả sử dụng protein và do đó một cách gián tiếp kiểm soát chặt chẽ hơn việc bài xuất protein, bài xuất protein trong chăn nuôi thâm canh tạo ra một lượng lớn chất gây ô nhiễm môi trường.
Tài liệu tham khảo cho phần 9.7 1. Whitelaw, F.G., Milne, J.S. and Ørskov, E.R. 1987. Urea metabolism in
sheep nourished by intragastric infusion. 5th International Symposium on Protein Metabolism and Nutrition. p. 82. Rostock, German Democratic Republic.
2. Whitelaw, F.G., Milne, J.S., Ørskov, E.R., Stansfield, R. and Franklin, M.F. 1990. Urea metabolism in sheep given conventional feeds or nourished by intragastric infusion. Exp. Physiol. 75, 239-254.
3. Wallace, R.J., Cheng, K.J., Dinsdale, D. and Ørskov, E.R. 1979. An independent microbial flora of the epithelium and its role in the ecomicrobiology of the rumen. Nature. 279, 424-426.
4. Zhao, G.Y., Duric, M., MacLeod, N.A., Ørskov, E.R., Hovell. F.D.DeB. and Feng, Y.L. 1995. The use of intragastric infusion to study sliva secretion and the relationship between rumen osmotic pressure and water transport. Br. J. Nutr. 73, 155-161.
5. Duric, M., Zhao, G.Y., Ørskov, E.R. and Chen, X.B. 1994. Indirect measurement of saliva secretion in sheep fed diets of different structures and the effect of such diets on ruminal fluid kinetics and fermentation pattern. Exp. Phisiol. 79, 823-830.
6. Pagella, J.H., Chen, X.B., MacLeod, N.A., Ørskov, E.R. and Dewey, P.J.S. 1997. Excretion of benzoic axít derivates in urine of sheep given intraruminal infusions of 3-phenylpropionic and cyclohexanecarboxylic axíts. Br. J. Nutr. 77, (4) 577-592.
7. Liu, S.M., MacLeod, N.A., Luo, Q.J., Chen, X.B., Kyle, D.J., Nicol, P., Harbron, C.G. and Ørskov, E.R. 1997. The effects of acute and chronic protein depletion and accretion on plasma concentrations of insulin-like growth factor-1, fibronectin and total protein for ruminants nourished by intragstric infusion of nutrients. Br. J. Nutr. 78, No. 3, 411-426.
Hướng nghiên cứu 10. Sinh lý tiết sữa

155
Hướng nghiên cứu 10.1 Ảnh hưởng của protein và glucose đến huy động mỡ ở bò sữa trong giai đoạn đầu vắt sữa.
Khi tôi được trao nhiệm vụ nghiên cứu về bò sữa năm 1977 ở Viện Rowett câu hỏi đầu tiên tôi muốn giải quyết là một vấn đề tranh cãi ở thời điểm đó. Cái gì đã hạn chế sản xuất sữa trong giai đoạn đầu vắt sữa, protein hay năng lượng từ glucose? Ý kiến xung quanh vấn đề này ở thời điểm đó là protein bị phân giải để cung cấp tiền chất glucose bởi vì người ta giả thiết rằng ở thời kỳ này có sự thiếu hụt glucose.
Có vẻ đây là một là một việc đơn giản. Tất cả những cái mà chúng tôi muốn là một ống thông (Catheter) để bơm dung dịch glucose hoặc casein vào trong dạ múi khế. Robin Kay đã lắp các ống thông vào dạ múi khế khoảng 2 ngày sau khi đẻ và ống thông được xâu thành chuỗi dưới da và chỉ bộc lộ ra ngoài ở lưng do đó gia súc không thể động chạm đến chúng. Chúng tôi nuôi bò với 10 lít sữa/ngày, và biết rằng chúng có khả năng cho năng suất sữa nhiều hơn, và chúng tôi đặt vấn đề một cách đơn giản, liệu đó là protein hoặc glucose mà bạn cần? (1). Thực tế bò cho 14 lít sữa ở khẩu phần đối chứng và ở tình trạng cân bằng năng lượng âm. Bảng 10.1. Đáp ứng của bò sữa khi bơm nước, 300g glucose hoặc 300g casein vào dạ múi khế.
Nghiệm thức
Năng suất sữa (kg/ngày) Protein sữa (%) Mỡ sữa (%)
Nước 14,5 3,07 4,93 Glucose 14,0 3,13 4,72 Casein 17,0 3,51 4,72 SE 0,4 0,06 0,02
Thí nghiệm đầu tiên (Bảng 10.1) cho thấy có sự đáp ứng với casein nhưng
không có đáp ứng với glucose. Chúng tôi quyết định tiếp tục mô tả đường cong đáp ứng với lượng casein hoặc glucose tăng dần. Chúng tôi sử dụng 4 con bò cái khác, gắn ống thông dạ múi kế, những bò này được nuôi để có thể sản xuất khoảng 10 lít sữa/ngày và chắc chắn có cân bằng năng lượng âm (Bảng 10.2). Bảng 10.2. Ảnh hưởng của tăng casein và giảm glucose bơm vào dạ múi khế đến năng suất sữa và thằnh phần sữa trong giai đoạn đầu vắt sữa.
Casein (g/ngày)
Glucose (g/ngày)
Năng suất sữa
(kg/ngày)
Mỡ sữa (%)
Protein sữa
(g/ngày)
Mỡ sữa hiệu chỉnh (FCM)
(Kg/ngày)
Cân bằng năng lượng (MJ/ngày)
0 750 16,8 4,82 4,23 18,9 -20,5

156
250 500 19,8 4,98 566 22,7 -30,5 500 250 21,6 5,10 644 25,2 -38,5 750 0 21,4 5,48 675 26,1 -41,0 SE 0,8 0,12 38
Kết quả là một kết luận công bằng: không có sự đáp ứng với glucose nhưng
có sự đáp ứng rõ ràng với casein như vậy protein đã kích thích làm cho cân bằng năng lượng âm cao lên, và điều đó đã kích thích huy động mỡ.
Tại thời điểm đó chúng tôi đã biết thay đổi cung cấp protein bằng cách thay đổi tỷ lệ phân giải protein khẩu phần trong dạ cỏ (Hướng nghiên cứu 1.2). Nếu protein khẩu phần có thể kích thích huy động mỡ (Hướng nghiên cứu 10.1), đáp ứng với protein không bị phân giải sẽ là lớn nhất khi năng lượng cung cấp là ít nhất so với nhu cầu cho sữa. Chúg tôi đã kiểm tra giả thuyết trên bằng 1 thí nghiệm sử dụng lạc (G) hoặc bột cá (F) ở mức cao (H) và thấp (L) trong khẩu phần. Năng suất sữa với các khẩu phần HG, HF, LG và LF tương ứng là 26,3; 26,0; 21,2 và 23,9 kg/ngày. năng lượng trao đổi ăn vào lần lượt là 174, 188, 137 và 141 MJ/ngày. Kết quả này một lần nữa khẳng định các kết quả trước đó. Ở mức năng lượng trao đổi ăn vào cao, lượng sữa đầy đủ được sản xuất từ lên men dạ cỏ. Ở khẩu phần năng lượng ăn vào thấp không phải như vậy, và do đó có sự đáp ứng với bột cá.
Một vấn đề chúng tôi phải quan tâm là nếu protein kích thích tăng huy động mỡ cơ thể có thể dẫn đến hội chứng ketosis. Chúng tôi quyết định tiến hành thêm một thí nghiệm bằng cách cho bò ở thời kỳ cho sữa sớm ăn khẩu phần có thức ăn thô cao (70% thức ăn ủ) nhưng với bột cá từ 0, 4, 8 và 12% của toàn bộ khẩu phần (Bảng 10.3) (3).
Bảng 10.3. Ảnh hưởng của bột cá trong khẩu phần hỗn hợp hoàn chỉnh cho bò sữa ở tuần vắt sữa thứ 2 đến năng suất sữa, thành phần sữa và cân bằng năng lượng tính được
Bột cá (% theo DM)
Năng suất sữa (kg/ngày)
FCM (kg/ngày)
Mỡ sữa (%)
Cân bằng năng lượng tính được
(MJ/ngày) 0 23,6 28,5 5,4 -77 4 26,1 29,2 4,8 -64 8 27,1 32,0 5,2 -80
12 28,5 34,9 5,5 -97 SE 1,8 1,8 0,3 Kết quả rất đáng ngạc nhiên. Bò đáp ứng tuyến tính với bột cá đưa vào
nhưng ME ăn vào chỉ đủ cho sản xuất khoảng 12 kg sữa hiệu chỉnh/ngày và thậm chí không có bột cá chúng cũng cho cho 28kg sữa/ngày và tăng đến 35kg

157
sữa/ngày. Với 12% bột cá 2 bò phát triển ketosis nhưng chúng đã huy động 2 đến 3 kg mỡ cơ thể/ngày và cho khoảng 25kg sữa hiệu chỉnh/ngày với năng lượng lấy từ mỡ cơ thể. Khẩu phần 12% bột cá không được tiếp tục cho ăn nữa nhưng năng suất vẫn như vậy cho đến 15 tuần (bảng 10.4).
Bảng 10.4. Ảnh hưởng của bột cá trong khẩu phần bò sữa đến năng suất sữa và cân bằng năng lượng từ 2 đến 15 tuần sau khi sinh.
Bột cá Năng suất sữa
(kg/ngày)
Sữa hiệu chỉnh (kg/ngày)
Mỡ sữa (%)
Cân bằng năng lượng tính được (MJ/ngày)
0 21,9 23,5 4,5 -32,3 4 25,2 25,6 4,1 -39,3 8 27,4 28,0 4,3 -43,8
SE 1,2 1,2 0,2
Ở đây chúng tôi không gặp vấn đề gì liên quan đến khả năng sinh sản hoặc ketosis mặc dù thực tế lượng mỡ mất đi tính được là hơn 1kg/ngày cho 15 tuần.
Câu hỏi khác chúng tôi cũng quan tâm là bò có nhạy cảm không với cung cấp protein ở cuối chu kỳ sữa. Nghiên cứu này là một phần công việc của sinh viên tiến sĩ người Bỉ của tôi Louis Istasse (4). Các kết quả chính được trình bày trong Bảng 10.5. Bảng 10.5. Ảnh hưởng của việc bơm casein vào dạ múi khế ở bò sữa trong giai đoạn đầu và cuối chu kỳ sữa
Đầu chu kỳ Cuối chu kỳ Casein
bơm vào (g/ngày)
Năng suất sữa
(kg/ngày)
Năng suất mỡ sữa (g/ngày)
Casein bơm vào
(g/ngày)
Năng suất sữa
(kg/ngày)
Năng suất mỡ sữa (g/ngày)
0 18,4 770 0 7,4 322 232 19,5 851 314 7,5 308 670 20,1 846 679 7,8 354 1009 21,4 832 1019 8,0 320
Như đã dự đoán có sự đáp ứng với protein ở đầu chu kỳ nhưng không có
đáp ứng đáng kể nào ở cuối chu kỳ sữa.
Bảng 10.6. Ảnh hưởng của bơm casein hoặc glucose vào dạ múi khế bò sữa đến năng suất sữa và sinh nhiệt.

158
Casein (g/ngày)
Năng suất sữa
(kg/ngày)
NL thức ăn (MJ/ngày)
NL casein (MJ/ngày)
NL sữa (MJ/ngày)
Nitơ sữa (g/ngày)
Mỡ sữa (g/ngày)
Sinh nhiệt (MJ/ngày)
0 13,0 168,0 0 45,1 60,8 599 59,2
200 15,3 166,7 4,3 52,4 73,5 700 61,8
400 16,5 171,4 8,6 56,6 80,7 745 65,2
600 17,2 169,1 12,9 56,2 85,6 719 64,4
Glucose (g/ngày 800)
12,3 - - - 593 514 -
SE 0,76 3,0 3,8 46 1,4
Đã đến lúc kết thúc và để có kết luận chắc chắn về câu chuyện glucose,
đồng nghiệp của tôi tiến sĩ Whitelaw (5) đã tiến hành một thí nghiệm khác. Ông bơm các mức casein khác nhau và cùng một mức glucose vào trong dạ múi khế bò sữa ở đầu chu kỳ sữa. Ảnh hưởng của glucose tương tự như ở đối chứng, không có đáp ứng, trong khi năng suất sữa, mỡ và protein sữa đáp ứng tuyến tính với casein (Bảng 10.1). Chúng tôi cũng đo trao đổi năng lượng và thật thú vị là bơm 4,3 MJ casein đã tạo ra sự tăng năng lượng sữa 7,3 MJ cho thấy casein đã kích thích huy động mỡ như thế nào. Sinh nhiệt chỉ tăng chút ít do việc bơm casein vào. Casein bơm vào được chuyển hoá vào sữa và được sử dụng với hiệu suất 64%.
Kết luận của thí nghiệm này là bò sữa ở đầu chu kỳ sữa và cân bằng năng lượng âm đáp ứng rất nhạy cảm với cung cấp protein vì các mô của cơ thể được huy động ở mức protein thấp. Thất bại trong việc cung cấp protein ở giai đoạn đầu chu kỳ sữa dẫn đến việc mất đi các mô nạc, không có đỉnh sữa nhưng có sự giảm năng suất sữa ngay từ tuần đầu sau khi sinh. Điều này dẫn đến chậm động dục trở lại.
Đáp ứng về năng suất sữa với cung cấp năng lượng ngoại sinh ở thời kỳ đầu chu kỳ sữa chủ yếu do lượng protein vi sinh vật điều khiển, luợng protein vi sinh vật này được tạo ra do tăng lượng thức ăn được lên men trong dạ cỏ. Protein có thể cũng kích thích huy động mỡ và có thể ở dưới những mức cực đoan dẫn đến ketosis nhưng trong thí nghiệm của chúng tôi ketosis chỉ xảy ra khi năng suất sữa cao hơn 26kg/ngày được cung cấp năng lượng từ huy động mỡ. Ở giai đoạn cuối chu kỳ sữa không có sự nhạy cảm như vậy với cung cấp proetin. Năng suất sữa không có vẻ bị hạn chế bởi tiền chất glucose. Tài liệu tham khảo cho phần 10.1.
1. Orskov, E.R., Grubb, D.A. and Kay, R.N.B. 1977. Effect of postruminal glucose or protein supplementation on milk yield and composition in friesian cows in early lactation and negative energy balance. Br. J. Nutr. 38, 397-405.
2. ỉrskov, E.R., Reid, G.W. and McDonald, I. 1981. The effect of protein degradability and food intake on milk yield and composition in cows in early

159
lactation. Br. J. Nutr. 45, 547-555. 3. ỉrskov, E.R., Reid, G.W. and Tait, C.A.G. 1987. Effect of fish meal on the
mobilization of body energy in dairy cows. Anim. Prod. 45, 3, 345-358. 4. Istasse, L., Brewer, A.C. and ỉrskov, E.R. 1986. Effect of stage of lactation
on the response of dairy cows to abomasal infusion of casein. Livestock Prod. Sci. 15, 97-102.
5. Whitelaw, F.G., Milne, J.S., ỉrskov, E.R. and Smith, J.S. 1986. The nitrogen and energy metabolism of lactating cows given abomasal infusion of casein. Br. J. Nutr. 55, 537-556.

160
Hướng nghiên cứu 10. Sinh lý tiết sữa Hướng nghiên cứu 10.2. Ảnh hưởng của bơm glucose, axít propionic hoặc
casein liên tục hoặc theo giai đoạn đến insulin, đường, glucose, axít propionic trong huyết tương và thành phần sữa.
Khả năng làm việc của Louis Istasse là vô tận. Ông đã thực hiện một số
công trình tuyệt vời về sự tương tác của thức ăn (Hướng nghiên cứu 2.1), nhưng ông cũng quan tâm đến dinh dưỡng trực tiếp cho dạ dày ở bò sữa (1). Trong thực tế ông đã phải ở với chúng thường xuyên, ngày và đêm. Louis Istasse đã cung cấp cho bò sữa một dinh dưỡng bơm vào dạ cỏ lớn đủ để chúng cho năng suất hơn 25 kg sữa/ngày, đôi khi tôi nghĩ là không thể làm được việc này. Bò cũng được cho ăn một ít thức ăn thô -2 kg hỗn hợp cỏ khô và rơm ủ urê với một chút củ cải đường. Không nghi nghờ gì việc nhai và nhai lại thức ăn thô này kích thích tiết nước bọt, điều này giúp đệm axít béo bơm vào, nhưng tôi lại một lần nữa ngạc nhiên – làm thế nào để những bò được bơm nhiều dinh dưỡng như vậy vào dạ cỏ lại có thể sẵn sàng ăn bất cứ thứ gì? Nhưng chúng đã ăn, và thậm chí tỷ lệ tiêu hoá chất hữu cơ ở chúng đạt 60% như ở bò bình thường. Năng lượng trao đổi của thức ăn khô ăn vào là 17MJ/ngày trong khi năng lượng trao đổi ăn vào từ dung dịch dinh dưỡng bơm vào dạ cỏ là 113MJ/ngày. Điều này có nghĩa là lên men dạ cỏ có thể vẫn tiếp tục ngay cả khi 85% năng lượng trao đổi được bơm vào dưới dạng axít béo bay hơi.
Một khẩu phần trộn hoàn toàn và một khẩu phần trong đó thức ăn tinh được cho ăn 2 lần một ngày sẽ cho ra lượng axít béo bay hơi, insulin và đường huyết rất khác nhau. Những ảnh hưởng này được kích thích khi hoặc tiếp tục bơm dinh dưỡng hoặc bơm hỗn hợp 70% axít propionic (15% năng lượng), ở hai, 3 giờ bơm liên tục rồi dừng. Một số kết quả trình bày trong Bảng 10.7. Bảng 10.7. Ảnh hưởng của bơm axít propionic liên tục hay theo giai đoạn đến đường huyết và glucose. Bơm liên tục Bơm theo giai đoạn
(3 h) Insulin (mM/l) 31,9 94,6 Glucose (mg/l) 699 684 Axít propionic (mmol/l) 0,039 0,228
Rõ ràng là một số axít propionic đã đi qua gan vào máu ngoại biên và đã
ảnh hưởng lớn đến nồng độ insulin mặc dù không có ảnh hưởng đến glucose huyết tương. Có một khuynh hướng là mỡ sữa và năng suất sữa giảm trong khoảng thời gian ngừng bơm axít propionic qua mạch máu. Ở cuối chu kỳ sữa không có ảnh hưởng này đến năng suất và thành phần sữa.

161
Ở đây tôi phải đề cập rằng Ken Lough cho thấy có sự tăng rõ rệt các axít mạch lẻ của mỡ sữa khi tỷ lệ phân tử lượng của axít propionic trong dịch dinh dưỡn bơm vào tăng (2). Trong thực tế tăng tỷ lệ phân tử lượng của propionate từ 25 đến 45 phần trăm làm tăng axít béo mạch lẻ từ 3 đến 10 phần trăm và việc tăng này luôn kết hợp với việc giảm tỷ lệ mỡ sữa. Một điều cũng thú vị là tỷ lệ axít béo mạch nhánh trong mỡ sữa, như iso và ante-iso axít chủ yếu có nguồn gốc từ vi sinh vật, giảm từ 3,3% ở bò sữa nuôi bình thường xuống dưới 1% ở bò bơm dinh dưỡng trực tiếp vào dạ dầy. Đây là một kết quả không mong đợi, nhưng đã giúp khẳng định nguồn gốc của các axít mạch nhánh vì có ít các hoạt động của vi sinh vật dạ cỏ ở bò nuôi bằng kỹ thuật bơm dinh dưỡng trực tiếp vào dạ dầy (Hướng nghiên cứu 8.2).
Ở một thí nghiệm được tiến hành công phu khác (3) Istasse đã so sánh những ảnh hưởng của bơm liên tục và theo giai đoạn propionic và glucose và casein đến insulin huyết tương và glucose huyết tương (Bảng 10.8). Glucose được bơm liên tục hoặc theo giai đoạn vào dạ múi khế nhằm cung cấp lượng năng lượng tương đương với lượng năng lượng của axít propionic bơm vào.
Bảng 10.8. Ảnh hưởng của của bơm liên tục hoặc gián đoạn axít propionic,
glucose (tương đương với lượng năng lượng của axít propionic bơm vào), và casein hoặc một dung dịch dinh dưỡng không có protein Insulin huyết tương
(mu/l) Glucose huyết tương (mg/l)
1100 h 1400 h 1100 h 1400 hAxít béo bay hơi & casein liên tục 25,7 24,8 685 648
Propionic theo giai đoạn 17,3 89,5 661 626
Glucose liên tục 25,0 32,5 696 698
Glucose theo giai đoạn 20,1 120,0 668 987
Casein theo giai đoạn 23,9 48,9 677 608
Không có casein 22,4 25,7 674 670 SED 3,4 16,2 30 41
*1400 h tuơng ứng thời gian kết thúc của các giai đoạn bơm gián đoạn
Việc bơm axít propionic gián đoạn một lần nữa tạo ra một đáp ứng insilin
mạnh mẽ mà không gây ra sự thay đổi nào về hàm lượng glucose máu. Nói một cách khác sự thay đổi hàm lượng insulin hình như không liên quan gì đến glucose máu nhưng luôn đi liền với axít propionic đã đi vào tuần hoàn ngoại biên. Với việc bơm gián đoạn glucose, insulin huyết tương phản ánh sự tăng glucose huyết tương. Với casein có sự tăng nhỏ nhưng đáng kể insulin huyết tương và giảm nhẹ glucose huyết tương. Liệu đáp ứng insulin là nhờ có các axít amin trong huyết tương? Nitơ amoniac tự do trong huyết tương tăng. Các vấn đề này vẫn chưa được

162
hiểu một cách đầy đủ nhưng chúng tôi đã phải để chúng lại dù không muốn vì Louis Istate phải quay về Bỉ.
Tài liệu tham khảo cho phần 10.2.
1. Istasse, L., Hovell. F.D.DeB., MacLeod, N.A. and Ørskov, E.R. 1987. The effects of continuous or intermittent infusion of propionic axít on plasma insulin and milk yield in dairy cows nourished by intragastric infusion of nutrients. Livestock Prod. Sci. 16, 201-214.
2. Lough, A.K., Ørskov, E.R., Calder, A.G. and Coutts, L. 1983. Composition of milk lipids from lactating cows maintained by intragastric infusion. Proc. Nutr. Soc. 42, 125A.
3. Istasse, L., MacLeod, N.A., Goodall, E.D. and Ørskov, E.R. 1987. Effects on plasma insulin of intermittent infusions of propionic axít, glucose or casein into the alimentary tract of non-lactating cows maintained on a liquid diet. Br. J. Nutr. 58, 139-148.

163
Hướng nghiên cứu 11. Chăn nuôi trong mối quan hệ hữu cơ bền vững với thực vật và đất Hướng nghiên cứu 11.1. Tương tác với các nước đang phát triển
Do một phần khả năng của chúng tôi trong đánh giá và nâng cao chất lượng thức ăn thô sử dụng kỹ thuật túi nylon (nylon bag) và do một số nghiên cứu khởi đầu ở Bangladesh và Cộng hoà Dominican, tôi ngày càng quan tâm nhiều hơn đến hoàn cảnh khó khăn của những nông dân nhỏ nuôi gia súc bản địa bằng thức ăn thô. Sự quan tâm này còn bắt đầu từ những quan sát rằng bò ở Bangladesh có dung tích đường tiêu hóa bằng 25 đến 33% khối lượng cơ thể, trong khi ở bò Holstein giá trị này là 15% (1). Tôi cũng đã đến thăm Bulgaria và Nam tư (cũ) trong các chuyến công tác của FAO.
Năm 1986 tôi nhận đợc một bức điện từ giáo sư Jasiorowski, trưởng bộ phận Thú y và Chăn nuôi gia súc của FAO đề nghị tôi về làm việc tại. Tôi không nộp đơn xin làm việc tại đây, nhưng tôi đã đến làm việc tại FAO với sự đồng ý của giáo sư James, Giám đốc Viện nghiên cứu Rowett là: tôi sẽ làm tư vấn cho FAO 6 tháng, nhưng công việc đã kéo dài hơn 2 năm. FAO đã chấp nhận và công việc của tôi đã bắt đầu tại các nước đang phát triển, làm việc ở các nước đang phát triển từ đó đã trở thành công việc của tôi. Trong thực tế, trung bình kể từ khi làm việc cho FAO, mỗi năm tôi đến khoảng 15 nước.
Công việc này đã cho tôi cơ hội giảng bài ở các cuộc họp và ở các trường đại học ở rất nhiều nước. Tôi cũng tổ chức các khoá học về dinh dưỡng ở các nước như Cuba, Indonesia, Zimbabwe, Sudan, Việt nam, Thổ Nhĩ Kỳ, Syria, Trung Quốc và Argentina. Không nghi nghờ gì các khoá học về dinh dưỡng đã ảnh hưởng đến mục tiêu nghiên cứu của chúng tôi. Do quản lý rất nhiều dự án phát triển, chủ yếu của FAO và Viện năng lượng Nguyên tử Quốc tế (IAEA) tôi đã hiểu được nhu cầu đào tạo cán bộ tại chỗ về những kỹ thuật phù hợp. Hiểu biết này đã dẫn đến việc thành lập chính thức Đơn vị nghiên cứu nguồn thức ăn quốc tế, chỉ với 6 năm Đơn vị nghiên cứu này tổ chức đào tạo ngắn hạn cho khoảng 60 người từ hơn 50 quốc gia khác nhau.
Đã có nhiều dự án hướng tới đánh giá thức ăn, nâng cao chất lượng thức ăn, thức ăn bổ xung, sản xuất sữa. Một số dự án thất bại do thiếu kiến thức về nhu cầu tại chỗ. Các dự án khác đã thành công như dự án bắt đầu ở Trung quốc về sử dụng urê nâng cao chất lượng rơm vào năm 1987. Zhenhai và Tingshuang (1999) báo cáo rằng 14 triệu nông hộ nhỏ đã xử lý urê cho 45 triệu tấn rơm năm (Kỷ yếu hội nghị quóc tế lần thứ ba về nâng cao năng suất chăn nuôi từ các nguồn lực địa phương, Chengdu Sishuan, Trung quốc).
Tôi đã nhận ra rằng mục đích của chăn nuôi là rất khác nhau từ hệ thống chăn nuôi tự cấp tự túc của nông dân, ở đây quản lý rủi ro quan trong hơn quản ý rủi ro ở các hệ thống chăn nuôi theo định hướng thị trường. Viglizzo từ Argentina (2) và tôi cùng đưa ra một bản tóm tắt trong Bảng 11.1 dưới đây.

164
Bảng 11.1. Một số yếu tố để phát triển các chiến lược nghiên cứu chăn nuôi giảm rủi ro. So sánh mục tiêu của hệ thống chăn nuôi theo định hướng thị trường và chăn nuôi tự cấp tự túc của nông dân Các hệ thống chăn nuôi
theo định hướng thị trường
Các hệ thống chăn nuôi tự cung tự cấp
Các mục tiêu tổng thể cho nông dân chăn nuôi
Lợi nhuận cao nhất Có nhiều tiền mặt Năng suất
Rủi ro ít nhất Hỗi trợ gia đình Ổn định và bền vững
Vai trò của nhà khoa học
Thiết kế hệ thống Quản lý hệ sinh thái
Mục tiêu trước mắt Đồng nhất di truyền Tăng năng suất Tiềm năng Gia súc chuên dụng Trao đổi dinh dưỡng
Đa dạng sinh học Duy trì và cải thiện Tiềm năng Gia súc kiêm dụng Dự trữ dinh dưỡng
Phương pháp luận Chuyên sâu Bền vững
Cách tiếp cận khoa học Một chuyên ngành Đa ngành và liên ngành Các giá trị thống kê sinh học được nhấn mạnh
Trug bình và ảnh hưởng chủ yếu
Phương sai và các tương tác
Ở đây chúng ta có thể thấy rằng hầu hết các nỗ lực khoa học nhắm hướng
đến các hệ thống chăn nuôi theo định hướng thị trường trong khi ở phạm vi toàn cầu các hệ thống chăn nuôi theo định hướng thị trường ít quan trọng nhất.
Tôi cũng nhận thấy tính phức tạp của gia súc kiêm dụng. Chiến lược giống sẽ hư thế nào khi có tới 8 sản phẩm chăn nuôi? Giống và nuôi dưỡng để sinh tồn chắc chắn là quan trọng. Dưới đây xin đưa ra môt ví dụ từ phỏng vấn các nhóm nông dân ở vùng núi đá Zambezi ở Zimbabwe. Sử dụng phương pháp đánh giá có sự tham gia, nông dân được hỏi về tên các sản phẩm từ gia súc của họ - bò, dê và gia cầm (Bảng 11.2).
Sau đó họ được đề nghị phân hạng các sản phẩm theo quan điểm nhận thức của họ về mức độ quan trọng - bằng cách đưa cho họ 20 hạt để họ xếp thứ tự quan trọng.
Bảng 11.2. Phân hạng các sản phẩm chăn nuôi của nông dân từ các cộng đồng khác nhau xung quanh vùng núi đá Zambezi ở Zimbabwe. Mỗi số thể hiện phân hạng của 20 khả năng Sức Bán An ninh Của hồi môn Thịt Sữa Lễ tiệc Phân

165
kéo của cô dâu Bò 6 3 3 3 1 2 1 1 Gia cầm - 5 3 - 7* - 4 2 Dê - 5 3 3 5 2 2 1
*Bao gồm trứng. Nguồn: Manuchi, G., Mashingaidze, K. vaf Orskov, E.R. không xuất bản.
. Ở đây chúng ta thấy rằng với bò, cày kéo là quan trọng nhất và an ninh
cũng rất quan trọng. Điều hấp dẫn ở đây là thịt và sữa đựơc phân hạng về tầm quan trọng thấp hơn của hồi môn của cô dâu (Bảng 11.2).
Chọn lọc cho tốc độ tăng trọng cao hơn và tăng lượng sữa sản xuất không thể giúp được cộng đồng này, tuy rằng chọn lọc cho tốc độ tăng trọng cao hơn và tăng lượng sữa sản xuất lại là mục tiêu các nhà khoa học theo đuổi.
Tôi cũng nhận ra rằng chăn nuôi trong nhiều trường hợp cần được xem xét trong mối quan hệ hữu cơ bền vững với thực vật và đất và không nên xem xét chăn nuôi chỉ dưới quan điểm chuyên sâu một ngành. Cách nhìn nhận này được minh họa ở Bảng 11.3 (3) dưới đây nơi mà bò ở Sri Lanka được chăn thả trong vườn dừa với cuòng độ chăn thả cao. Ở đây chúng ta thấy rằng chăn thả và sự quay vòng nhanh hơn của sinh khối đã làm tăng năng suất dừa lên khoảng 20%. Sản phẩm lớn nhất về chăn nuôi có thể bị bỏ quên nếu chúng ta không xác định năng xuất dừa. chăn thả và sự quay vòng nhanh hơn của sinh khối cũng giúp cải tạo đất. Bổ sung rơm lúa và cám gạo cho bò đã có ảnh hưởng tích cực đến khả năng suất gia súc (sữa và sinh sản), năng suất dừa và đô mẫu mỡ của đất. Rõ ràng chăn nuôi cân bằng tích cực với đất và cây trồng. Bảng 11.3. Ảnh huởng của chăn thả bò dưới tán dừa đến năng suất dừa, năng suất của 12 bò chăn thả và đặc tính của đất. UG: không chăn thả; G: chăn thả; GS: chăn thả bổ sung rơm lúa; GSS: chăn thả và bổ sung rơm lúa, cám gạo Năng suất dừa
(quả/cây/năm) Cùi dừa khô (kg/cây/năm)
Bê sinh ra/năm/bò
Năng suất sữa (kg/năm)
Ngày vắt sữa
Khả năng giữ nước của đất
(Mm/m) UG 41,1 11,1 - - 16,9 G 47,9 13,3 0,3 181 201 18,3 GS 50,6 14,1 0,6 405 264 18,9 GSS 57,4 16,7 0,9 664 282 17,6 SE 2,0 0,5 0,03 25 4
Cuối cùng tôi đã học được từ những người nông dân ở Châu Phi và đặc biệt
ở Châu Á (Việt nam, Philipines, Trung Quốc): có thể tách chiết được bao nhiêu từ sinh khối để sử dụng. Ở đây phân không phải là chất bỏ đi mà là nguồn sinh khối giá trị để sản xuất khí đốt sinh học, thức ăn và thậm chí nước thải từ bể biogas đ-ược sử dụng làm phân bón, thức ăn cho cá và nước từ các hồ nuôi cá được sử dụng để tưới cho cây trồng. Khi vịt được nuôi ở Việt nam, phân của chúng dùng để nuôi

166
cá trong các ao cá. Nông dân Kikuyo ở Kenya nuôi gà để lấy trứng và thịt và phân gà được sử dụng bổ sung vào thân cây ngô làm thức ăn cho bò. Các kết quả này được minh họa trong sơ dưới đây.
Tôi gọi sơ đồ này là quản lý tổng thể nguồn lợi
Suy nghĩ một cách nghiêm túc cho thấy quá nhiều sinh khối đang bị lãng phí. Chúng ta có thể nuôi được nhiều người hơn nếu quan tâm hơn đến việc sử dụng sinh khối. Không cần biến đổi gen để tăng năng suất! Chỉ cần một sự thay đổi từ hiệu quả sử dụng lao động sang hiệu quả sử dụng nguồn lợi. Có nhiều thách thức khoa học có thể giúp mở khoá cánh cửa kho dự trữ sinh khối to lớn hiện đang bị lãng phí.
Chúng ta được đào tạo để suy nghĩ chuyên sâu, chúng ta được đào tạo để suy nghĩ về việc sử dụng tối đa thức ăn để có ít chất dinh dưỡng trong phân và chúng ta diễn đạt quan hệ này là chuyển hoá thức ăn. Chúng ta không quan tâm đến việc phân sẽ đi như thế nào. Trong lý thuyết về quản lý tổng thể nguồn lợi, chất lượng phân có thể rất quan trọng ví dụ nếu phân gia cầm được sử dụng làm thức ăn cho gia súc nhai lại và sau đó phân gia súc nhai lại lại được sử dụng cho sản xuất khí sinh học và nuôi cá hoặc nuôi giun đất.
Ở Việt nam giá trị của cá từ ao nuôi cá được bón phân thường cao hơn chăn nuôi lợn để sản xuất phân. Khoa học có thể giúp chúng ta chọn ra các hệ thống
Phụ phẩm chất lượng cao
Phụ phẩm chất lượng thấp
Gia súc dạ dầy đơn
Chất thải
Phân Ao cá Khí sinh học
Gia súc nhai lại
Chất thải
Nước thải sau Biogas
Ao cá Phân
Ao cá
Phân

167
quản lý nguồn lợi thích hợp nhất cho các vùng khí hậu khác nhau, các môi trường kinh tế xã hội khác nhau. Nghiên cứu tập trung vào quản lý tổng thể nguồn lợi có thể có nhiều ứng dụng trong xuất thức ăn. Một điểm nữa cũng đáng phải chỉ ra ở đây là sử dụng tổng thể nguồn lợi không gây ô nhiễm môi trường.
Tài liệu tham khảo cho phần 11.1. 1. Mould, F.L., Saadullah, M., Haque, M., Davis, C., Dolberg, F. and Ørskov,
E.R. 1982. Investigation of some of the physilogical factors influencing intake and digestion of rice straw by native cattle of Bangladesh. Trop. Anim. Prod. 7, 174-181.
2. Ørskov, E.R. and Viglizzo, E.F. 1994. The Role of Animals in Spreading Farmers’ Ricks: a New Paradigm for Animal Science. Outlock on Agriculture. 23 (2) 81-89.
3. Pathirana, K.K., Mangalika, U.L.P., Liyanage, M. De S. and Ørskov, E.R. 1996. Effect of grazing and supplementation in a coconut plantation on cattle production, coconut yield and soil fertility. Outlock on Agriculture. 25, 3, 187-192.

168
Hướng nghiên cứu 12. Bây giờ chúng ta đang ở đâu?
Trong khi sự nghiệp nghiên cứu của tôi có thể đang giảm dần, ý chí của tôi đang tiếp tục khuyến khích các nhà khoa học trẻ nhiệt tình với nghiên cứu chăn nuôi. Trong những năm gần đây ở Châu Âu vấn đề tối ưu sinh học có thể ít quan trọng với nông dân hơn hệ thống trợ giúp. Sự nỗ lực cho việc quay vòng nhanh ở bò cho đến lúc này đã có những ảnh hưởng không mong muốn đến chăn nuôi bền vững dựa trên thức ăn thô do đó một số gia súc đã được chọn lọc theo hướng tăng dung tích đường tiêu hóa để có thể ăn và chứa một lượng lớn thức ăn có dung tích lớn bằng cách tăng tỷ lệ loại thải ở bò thịt. Cũng đã có khuynh hướng chọn lọc chống lại khuynh hướng tích luỹ mỡ - một phương pháp sống còn tiết kiệm thức ăn khi nguồn cung dinh dưỡng biến động. Bền vững được hiểu là chăn nuôi gia súc dựa vào chăn thả, thức ăn ủ chua và phế phụ phầm cây trồng, do đó chúng ta phải quan tâm nhiều hơn đến khả năng tiết kiệm thức ăn của bò bằng cách tích luỹ mỡ như đã trình bày ở hướng nghiên cứu 10.1 và khả năng của chúng chứa được một lượng lớn thức ăn có dung tích cồng kềnh. Ở phần lớn các nước đang phát triển khi môi trường còn đang trong tầm kiểm soát, gia súc thường thích nghi cao nhưng khả năng thích nghi bị suy yếu bởi sức ép liên tục của các nước phương Tây để xuất khẩu cái được gọi là chăn nuôi cải tiến đã được chọn lọc trong các điều kiện kiểm soát môi trường.
Hiện vẫn còn các khả năng hiện hữu nhưng quan trọng là các nhà khoa học cần phải nhận thức về những mục tiêu lâu dài cho nông nghiệp ở các hệ sinh thái khác nhau. Cũng cần thiết là các nhà khoa học phải tham gia vào việc hoạch định các mục tiêu vì họ biết, hoặc sẽ hiểu biết tốt hơn các nhà chính trị và các nhà lập kế hoạch cái gì là khả thi. Nên tập trung nhiều hơn vào sử dụng có hiệu quả các chất dinh dưỡng từ phế phụ phẩm trồng trọt và nên ít tập trung vào các nỗ lực biến đổi gen không thích hợp, vì tập trung vào các nỗ lực biến đổi gen không thích hợp sẽ sinh ra nghèo đói thay vì xóa đói giảm nghèo. Ở nhiều phương diện nghiên cứu chăn nuôi trong những năm gần đây thu hút nhiều nhân tài vật lực hơn ở các nước đang phát triển vì nó gần với thực tế hơn. Chúng ta hy vọng rằng nghiên cứu chăn nuôi ở các nước phát triển cũng sẽ quay trở lại gần với thực tế hơn!
Kết luận
Tôi hy vọng rằng các câu truyện và các chuyến thăm quan khoa học kể ra ở đây sẽ giúp truyền cảm hứng cho những người đang nghĩ đến việc coi nghiên cứu nông nghiệp như một nghề nghiệp của mình.
Tôi đã cố gắng diễn tả bằng cách nào một thí nghiệm và các kết quả của nó dẫn tới thí nghiệm khác và bằng cách nào một thí nghiệm và các kết quả của nó đem đến một nền tảng suy nghĩ mới dẫn đến việc đặt ra các câu hỏi chưa hề được nghĩ đến trước đó.
Cảm giác một người thành công trong phát triển những kỹ thuật, phương

169
pháp mới như phản xạ rãnh thực quản, kỹ thuật dinh dưỡng trực tiếp cho dạ dầy, hoặc kỹ thuật chế biến ngũ cốc, giống như sự kích động khi lần đầu đạt tới đỉnh một ngọn núi và nhìn ra xung quanh. Những viễn cảnh mới mới mở ra và lý do ban đầu để đến được đỉnh núi có thể ít thích hợp hơn so với cái bây giờ có thể làm.
Cũng như vậy, một người mới đến với những tài năng mới có thể khiến một dự án đi theo hướng không đúng như kế hoạch ban đầu. Tôi đã hiếm có cơ hội chọn nhân viên, và có thể đó lại không phải là một điều tồi tệ. Đón chào sinh viên từ khắp nơi trên thế giới với những quan tâm, kỹ năng và tài năng khác nhau có thể thích hợp hơn để chọn những nhân viên làm việc lâu dài với mục tiêu có chủ tâm và định trước.
Phần thưởng khác, ít nhất là cho tôi, chính là sự hữu ích của của các tri thức mới chúng tôi đã đạt được đối với khách hàng của chúng tôi. Đúng là như nhận được giải thưởng khi nhìn thấy những người nông dân áp dụng những kỹ thuật do nhóm chúng tôi nghiên cứu, thậm chí nếu họ không biết kỹ thuật đang sủ dụng do ai nghiên cứu ra thì đó vẫn là một giải thưởng. Trong một dịp tình cờ ở triển lãm nông nghiệp tôi đang nhìn những bò sữa triển lãm và nói chuyện với một nông dân về cách ông ta nuôi bò. Ông ta đã nói với tôi về việc sử dụng ngũ cốc xử lý soda và ông ta thật sự hài lòng hài về ngũ cốc xử lý soda. Khi tôi thể hiện sự quan tâm và hiểu biết về việc này, ông ta ngay lập tức hỏi: anh cũng sử dụng phương pháp đó chứ? Một người chăn nuôi cừu đang cần tư vấn về nuôi dưỡng kể cho tôi về sự hài lòng của ông ta khi sử dụng ngũ cốc nguyên hạt và bột cá ép viên và hỏi xem ở Viện chúng tôi có nuôi cừu với ngũ cốc nguyên hạt và bột cá ép viên như ông ta không! Mặc dù họ không biết ai đã nghiên cứu ra kỹ thuật, nhưng việc họ áp dụng kỹ thuật đã là một phần thưởng ít nhất là với tôi, phần thưởng này có giá trị hơn cả những bài báo được chấp nhận cho xuất bản ở các tạp chí quốc tế.
Vậy sau đó phải chăng nghiên cứu đã được định hướng theo nhu cầu của nông dân? ở đây tôi phải nói, không, không thực sự như vậy. Một cách chung nhất trong khi tôi ủng hộ cách tiếp cận này ỏ các nhà khoa học, các nhà khoa học cần có gắng hiểu các vấn đề của nông dân và thiết lập các nghiên cứu để giải quyết chúng, những nghiên cứu diễn tả ở đây không được định hướng trực tiếp bởi nhu cầu khách hàng của chúng ta nhưng chúng được định hướng nhiều hơn bởi ham muốn hiểu biết sự tác động qua lại phức tạp giữa thức ăn và gia súc. Trong suốt thời gian qua, có thể nền tảng hiểu biết của tôi đã giúp tôi biết cách làm thế nào để một phát kiến mới có thể áp dụng trực tiếp vào trang trại. Dinh dưỡng trực tiếp cho dạ dày hình như khác biệt xa với chăn nuôi gia súc, tuy vậy kỹ thuật sẽ mang lại nhiều kết quả cho dinh dưỡng gia súc và vì vậy cho chăn nuôi.
Cuối cùng tôi đó có cơ hội tốt có được những người đồng nghiệp xuất sắc luôn luôn hạnh phúc thảo luận và hợp tác. Không thể kể hết tên của họ nhưng ngài Keneth Blaxter là nguồn cảm hứng và là nhà trí thức lớn. Các đồng nghiệp như Robin Kay, Ian McDonald, John Robinson, Vernon Fowler, Dickin Hovell, George Whitelaw, John Wallace, Colin Sterwart và các trợ lý kỹ thuật của tôi như Colin Fraser, Elizabeth Corse, David Grubb, George Reid, Neil McLeod, Alan McDearmid, Charlie Tait, David Kyle, một số lượng lớn sinh viên thạc sĩ, tiến sĩ,

170
công nhân và nhiều người khác nữa xứng đáng nhận được sự khen ngợi cho các nghiên cứu được mô tả ở đây.

171
ĐOẠN BÌA NGOÀI CUỐI
Trong cuốn sách này GS. Orskov đã miêu tả rất nhiều các dự án nghiên cứu và phát triển
chăn nuôi mà ông đã tham gia. Ông đã giải thích làm thế nào mà từ một dự án này lại dẫn
đến một dự án khác và thỉnh thoảng phần kết thúc của hướng nghiên cứu lại không được
nhận ra ngay từ lúc bắt đầu. Sự khởi đầu của hướng nghiên cứu và hướng đi của nó
không những chịu ảnh hưởng của các mục tiêu hợp lý mà còn chịu ảnh hưởng của tính
cách con người, các khả năng về kỹ thuật, các quan sát tình cờ và tài năng của các sinh
viên và nhân viên kỹ thuật. Các yếu tố này không bao giờ xuất hiện trong các ấn phẩm
khoa học. Các hướng nghiên cứu, do vậy, mang một nội hàm khoa học, kỹ thuật to lớn và
cũng hàm chứa các yếu tố con người trong nó. Hy vọng rằng những nội hàm này sẽ mang
lại niềm say mê cho bạn đọc