Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Hypoxia-induced endogenous prostaglandin E2 negativelyregulates hypoxia-enhanced aberrant overgrowth of rheumatoidsynovial tissue
Hirofumi Mitomi • Hidehiro Yamada • Hiroshi Ito • Toshiko Nozaki Shibata •
Yoshioki Yamasaki • So Nomoto • Atsushi Kusaba • Hiroki Yamashita •
Shoichi Ozaki
Received: 3 August 2012 / Accepted: 26 October 2012 / Published online: 25 November 2012
� Japan College of Rheumatology 2012
Abstract
Objective During isometric exercise, the synovial joint
tissue is prone to hypoxia, which is further enhanced in the
presence of synovial inflammation. Hypoxia is also known to
induce inflammatory cascades, suggesting that periodic
hypoxia perpetuates synovitis in rheumatoid arthritis. We
previously established an ex vivo cellular model of rheu-
matoid arthritis using the synovial tissue-derived inflam-
matory cells, which reproduced aberrant synovial
overgrowth and pannus-like tissue development in vitro.
Using this model, we investigated the regulatory mechanism
of synovial cells against hypoxia in rheumatoid arthritis.
Methods Inflammatory cells that infiltrated synovial tis-
sue from patients with rheumatoid arthritis were collected
without enzyme digestion, and designated as synovial tis-
sue-derived inflammatory cells. Under normoxia or peri-
odic hypoxia twice a week, their single-cell suspension was
cultured in medium alone to observe an aberrant over-
growth of inflammatory tissue in vitro. Cytokines produced
in the culture supernatants were measured by enzyme-
linked immunosorbent assay kits.
Results Primary culture of the synovial tissue-derived
inflammatory cells under periodic hypoxia resulted in the
attenuation of the spontaneous growth of inflammatory tis-
sue in vitro compared to the culture under normoxia.
Endogenous prostaglandin E2 (PGE2) production was
enhanced under periodic hypoxia. When endogenous PGE2
was blocked by indomethacin, the aberrant tissue over-
growth was more enhanced under hypoxia than normoxia.
Indomethacin also enhanced the production of tumor
necrosis factor-a (TNF-a), macrophage colony-stimulating
factor (M-CSF), and matrix metalloproteinase-9 (MMP-9)
under periodic hypoxia compared to normoxia. The EP4-
specific antagonist reproduced the effect of indomethacin.
Exogenous PGE1 and EP4-specific agonist effectively
inhibited the aberrant overgrowth and the production of the
inflammatory mediators under periodic hypoxia as well as
normoxia.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10165-012-0794-7) contains supplementarymaterial, which is available to authorized users.
H. Mitomi � H. Yamada (&) � H. Ito � T. N. Shibata �Y. Yamasaki � S. Ozaki
Division of Rheumatology and Allergology, Department of
Internal Medicine, St. Marianna University School of Medicine,
2-16-1 Sugao, Miyamae-ku, Kawasaki 216-8511, Japan
e-mail: [email protected]
H. Mitomi
e-mail: [email protected]
H. Ito
e-mail: [email protected]
T. N. Shibata
e-mail: [email protected]
Y. Yamasaki
e-mail: [email protected]
S. Ozaki
e-mail: [email protected]
S. Nomoto
Department of Orthopedic Surgery and Rheumatology,
Saiseikai Yokohamashi Tobu Hospital, 3-6-1, Shimosueyoshi,
Tsurumi-ku, Yokohama 230-8765, Japan
e-mail: [email protected]
A. Kusaba � H. Yamashita
Institute of Rheumatology, Ebina General Hospital, 1320,
Kawaraguchi, Ebina 243-0433, Japan
e-mail: [email protected]
H. Yamashita
e-mail: [email protected]
123
Mod Rheumatol (2013) 23:1069–1075
DOI 10.1007/s10165-012-0794-7

Conclusions The enhancing effect of periodic hypoxia on
the aberrant overgrowth of rheumatoid synovial tissue was
effectively down-regulated by the simultaneously induced
endogenous PGE2.
Keywords Rheumatoid arthritis � Hypoxia � Synovial
overgrowth � PGE2 � NSAIDs
List of Abbreviations
RA Rheumatoid arthritis
IL Interleukin
PG Prostaglandin
TNF Tumor necrosis factor
M-CSF Macrophage colony-stimulating factor
MIP Macrophage inflammatory protein
VEGF Vascularendothelial growth factor
HIF-1a Hypoxia-inducible factor-1aMMP Matrix metalloproteinase
IFN Interferon
COX Cyclooxygenase
normo Normoxia
hypo Periodic hypoxia
Introduction
Inflammation in general is fundamentally a protective
response against cellular and tissue injury caused by diverse
pathological stimuli, and it is closely intertwined with the
process of repair. In some circumstances, inflammation and
tissue repair are not successfully completed and inflamma-
tion perpetuates chronically. Rheumatoid arthritis is char-
acterized by chronic inflammation of the synovial
membrane. A critical event during the chronic inflammation
is the transformation of the synovial membrane into an
aggressive granulation tissue (so-called pannus), resulting in
cartilage and bone destruction [1, 2]. The joint synovial
tissue is prone to hypoxia during exercise as well as ische-
mia–reperfusion injury, which is further enhanced in the
presence of synovial inflammation [3]. It is reported that
inflammatory effusions had lower oxygen tensions than non-
inflammatory effusions and the more severe histological
grading of synovial changes correlated with lower oxygen
tension [3]. It is also reported that low tissue oxygen pressure
levels in the inflamed synovial joint tissue was significantly
associated with the degree of synovitis and the level of TNF-
a, interleukin (IL)-1b, interferon (IFN)-c and the chemokine
macrophage inflammatory protein (MIP) 3a [4].
Cytokine networks and cell–cell interaction, as well as
other inflammatory mediators, such as prostanoids, con-
tribute to the development of pannus tissue. This complex
system of rheumatoid synovitis includes both positive and
negative feedback regulation of the inflammatory response.
Therefore, a human cell model that represents this complex
will be useful to study the role of hypoxia in the patho-
genesis of rheumatoid arthritis. We previously established
an ex vivo cellular model using the synovial tissue-derived
inflammatory cells, which reproduced pannus-like tissue
growth in vitro [5]. Using this model, we demonstrated that
periodic hypoxia enhanced aberrant synovial overgrowth,
which was negatively regulated by the simultaneously
induced PGE2 in the hypoxic condition.
Materials and methods
Reagents
PGE1 was purchased from Sigma-Aldrich (St. Louis, MO).
Indomethacin was from Wako (Osaka, Japan). The EP4-
specific agonist (ONO-AE1-329 and ONO-AE1-437) and
EP4-specific antagonist (ONO-AE3-208) were provided
from Ono Pharmaceuticals co (Osaka, Japan). In mem-
branes of Chinese hamster ovary cells expressing the
respective mouse EP receptors, the above EP agonist had
the following Ki values in 3H-PGE2 binding assays: ONO-
AE1-329, 0.0097 lM at the EP4 receptor; and ONO-AE1-
437, 0.0007 lM at the EP4 receptor [6, 7, 8]. The 50 %
maximum response concentration values were as follows:
ONO-AE3-329, 0.0031 lM at the EP4 receptor ([10 lM at
the EP1 receptor, 1.2 lM at the EP2 receptor, and 5.8 lM at
the EP3 receptor); ONO-AE1-437, 0.0007 lM at the EP4
receptor ([10 lM at the EP1 receptor, 0.62 lM at the EP2
receptor, and 0.056 lM at the EP3 receptor). The Ki values
of EP4 receptor antagonist ONO-AE3-208 were 0.0013 lM
for the EP4 receptor, 0.03 lM for the EP3 receptor, and
[10 lM for the mouse EP1, EP2, DP receptors [9], and it
antagonized both an increase of intracellular cAMP induced
by 100 nM PGE2 with a 50 % inhibition concentration
(IC50) value of 1.7 nM and an increase of cytosolic calcium
induced by 10 nM PGE2 with an IC50 value of 120 nM.
Synovial tissue specimens
Synovial tissue specimens were obtained from patients who
fulfilled the 1987 revised classification criteria of the
American College of Rheumatology who underwent knee
joint replacement. In compliance with institutional policies,
informed consent was obtained from all patients. The ethics
committee of each institution approved this study.
In vitro reconstruction of inflammatory tissue
by synovial tissue-derived inflammatory cells
Synovial tissue-derived inflammatory cells were prepared as
previously described [5]. In brief, synovial tissue specimen
1070 Mod Rheumatol (2013) 23:1069–1075
123
were cut into small pieces and cultured in 100-mm dishes
containing RPMI 1640 (Asahi Technoglass, Chiba, Japan)
with 10 % fetal calf serum (FCS) and 100 units/ml of peni-
cillin G sodium and 100 lg/ml streptomycin sulfate (Gibco
BRL, Grand Island, NY). After 1–3 days’ incubation, tissue
was removed and single cells were collected by vigorous
pipetting. Cell suspensions were washed once and viable cells
were collected onto Lymphocyte Separation Solution (Nac-
alai Tesque, Kyoto, Japan). Single suspension of synovial
tissue-derived inflammatory cells were seeded at a density of
5 9 105/well in 48-well culture plates and cultured in Dul-
becco’s modified Eagle’s medium (DMEM; Gibco BRL)
containing 10 % FCS, 100 units/ml of penicillin G sodium
and 100 lg/ml streptomycin sulfate. The culture was
observed for morphologic changes under an inverted phase-
contrast microscope twice a week for 4 weeks. When cul-
tured in DMEM/10 % FCS in the absence or presence of
EP4-specific antagonist (100 ng/ml) or indomethacin (1 lM)
under hypoxic condition or normoxic condition, synovial
tissue-derived inflammatory cells started to aggregate,
forming foci within a few days. Further culturing resulted in a
3-dimensional (3-D) growth, which ultimately produced
macroscopic tissue 2 mm in size within 4 weeks. Morpho-
logic changes were semiquantitatively scored on a scale of
0–4, according to the degree of tissue development, where
0 = no cellular foci or aggregations, 1 = formation of cel-
lular foci or aggregations, 2 = further growth of cellular
aggregations, 3 = further 3-D growth with a multilayered
structure, and 4 = development of macroscopic tissue. A
cumulative tissue growth score was calculated by the total
sum of the tissue growth scores obtained twice a week for
4 weeks of culture. Half of the supernatants were collected
twice a week and replaced with fresh medium or the addition
of a dose of EP4-specific antagonist or indomethacin.
Supernatants were frozen at -80 �C until assayed.
Hypoxic culture conditions
In each experiment, synovial tissue-derived inflammatory
cells from one donor were seeded in two 48 well culture
plates. One plate was incubated under a normoxic condi-
tion in a 5 % CO2 incubator (Sanyo Co., Osaka, Japan) as a
control, another plate was exposed to a 1 % O2 condition
for 24 h twice a week in a hypoxic chamber, PERSONAL
CO2 MULTI GAS INCUBATOR (ASTEC Co., Fukuoka,
Japan) which create an atmosphere composed of 1 % O2,
5 % CO2 and 94 % N2. Joint movements in daily life
frequently result in the hypoxia-reperfusion of synovial
tissue especially in patients with rheumatoid arthritis. In
order to mimic this condition, we cultured the synovial
cells under periodic hypoxia. Another reason was the fact
that continuous hypoxia during 4 weeks culture resulted in
the loss of cell viability in our pilot experiments.
Cytokine assay
Supernatants obtained from each course of cell cultures
were stored at -80 �C until assayed, and levels of vascular
endothelial growth factor (VEGF), PGE2, TNF-a, M-CSF
(all from R&D Systems, Minneapolis, MN, USA), and
matrix metalloproteinase 9 (MMP-9; Amersham Biosci-
ences, Buckinghamshire UK) released into the culture
supernatants were measured using enzyme-linked immu-
nosorbent assay kits according to the manufacturers’
recommendations.
Result
Periodic hypoxia induces VEGF release
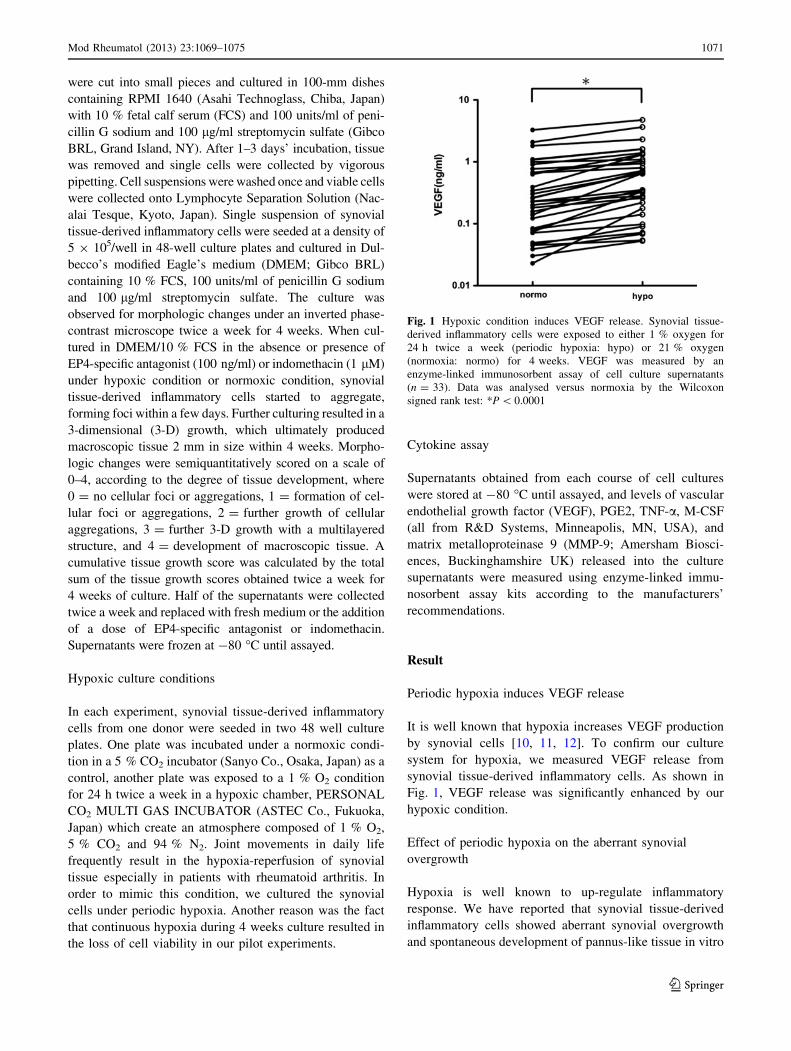
It is well known that hypoxia increases VEGF production
by synovial cells [10, 11, 12]. To confirm our culture
system for hypoxia, we measured VEGF release from
synovial tissue-derived inflammatory cells. As shown in
Fig. 1, VEGF release was significantly enhanced by our
hypoxic condition.
Effect of periodic hypoxia on the aberrant synovial
overgrowth
Hypoxia is well known to up-regulate inflammatory
response. We have reported that synovial tissue-derived
inflammatory cells showed aberrant synovial overgrowth
and spontaneous development of pannus-like tissue in vitro
Fig. 1 Hypoxic condition induces VEGF release. Synovial tissue-
derived inflammatory cells were exposed to either 1 % oxygen for
24 h twice a week (periodic hypoxia: hypo) or 21 % oxygen
(normoxia: normo) for 4 weeks. VEGF was measured by an
enzyme-linked immunosorbent assay of cell culture supernatants
(n = 33). Data was analysed versus normoxia by the Wilcoxon
signed rank test: *P \ 0.0001
Mod Rheumatol (2013) 23:1069–1075 1071
123
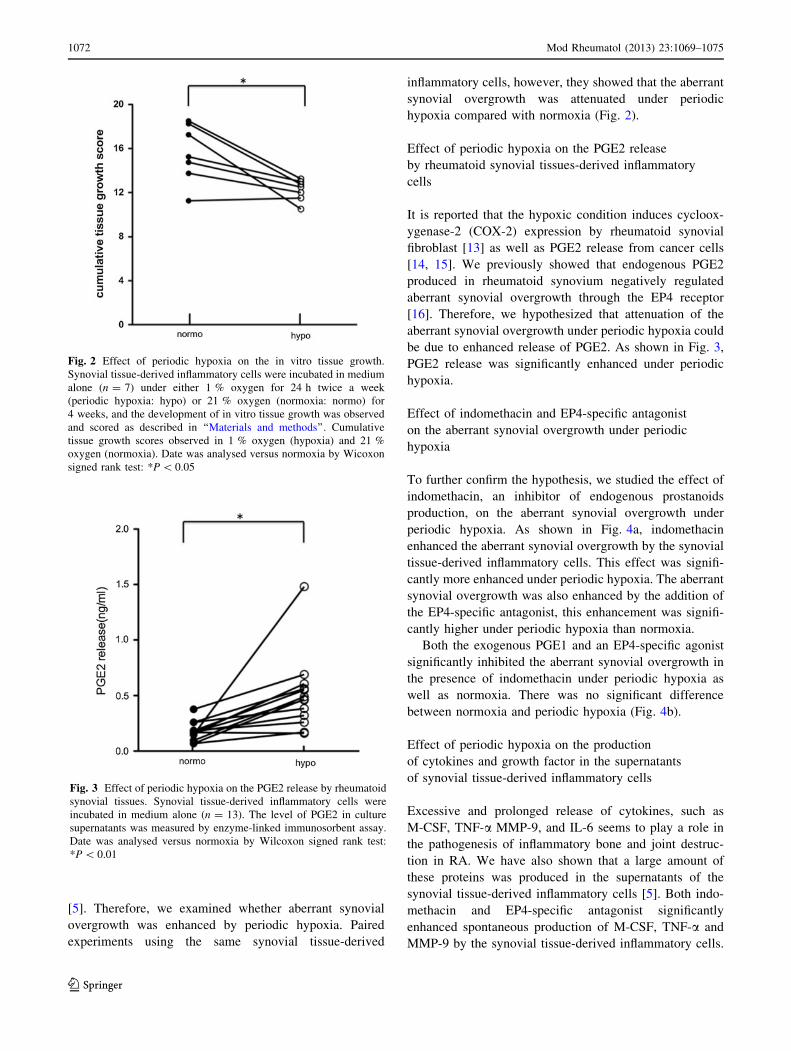
[5]. Therefore, we examined whether aberrant synovial
overgrowth was enhanced by periodic hypoxia. Paired
experiments using the same synovial tissue-derived
inflammatory cells, however, they showed that the aberrant
synovial overgrowth was attenuated under periodic
hypoxia compared with normoxia (Fig. 2).
Effect of periodic hypoxia on the PGE2 release
by rheumatoid synovial tissues-derived inflammatory
cells
It is reported that the hypoxic condition induces cycloox-
ygenase-2 (COX-2) expression by rheumatoid synovial
fibroblast [13] as well as PGE2 release from cancer cells
[14, 15]. We previously showed that endogenous PGE2
produced in rheumatoid synovium negatively regulated
aberrant synovial overgrowth through the EP4 receptor
[16]. Therefore, we hypothesized that attenuation of the
aberrant synovial overgrowth under periodic hypoxia could
be due to enhanced release of PGE2. As shown in Fig. 3,
PGE2 release was significantly enhanced under periodic
hypoxia.
Effect of indomethacin and EP4-specific antagonist
on the aberrant synovial overgrowth under periodic
hypoxia
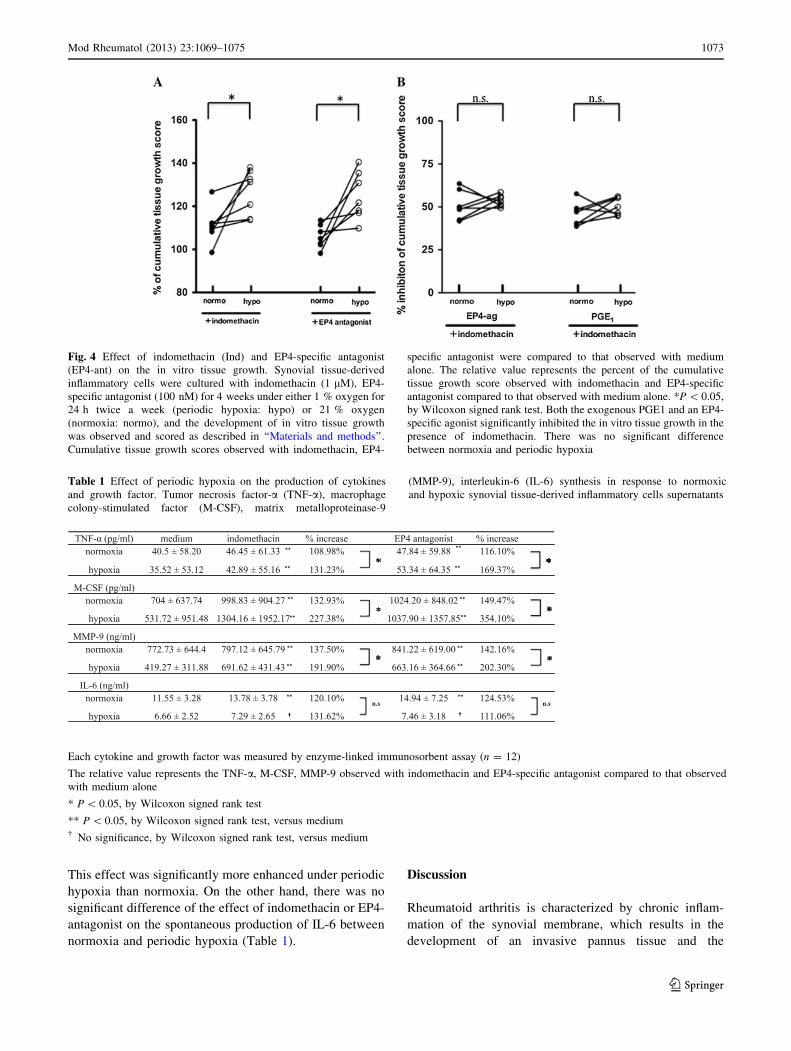
To further confirm the hypothesis, we studied the effect of
indomethacin, an inhibitor of endogenous prostanoids
production, on the aberrant synovial overgrowth under
periodic hypoxia. As shown in Fig. 4a, indomethacin
enhanced the aberrant synovial overgrowth by the synovial
tissue-derived inflammatory cells. This effect was signifi-
cantly more enhanced under periodic hypoxia. The aberrant
synovial overgrowth was also enhanced by the addition of
the EP4-specific antagonist, this enhancement was signifi-
cantly higher under periodic hypoxia than normoxia.
Both the exogenous PGE1 and an EP4-specific agonist
significantly inhibited the aberrant synovial overgrowth in
the presence of indomethacin under periodic hypoxia as
well as normoxia. There was no significant difference
between normoxia and periodic hypoxia (Fig. 4b).
Effect of periodic hypoxia on the production
of cytokines and growth factor in the supernatants
of synovial tissue-derived inflammatory cells
Excessive and prolonged release of cytokines, such as
M-CSF, TNF-a MMP-9, and IL-6 seems to play a role in
the pathogenesis of inflammatory bone and joint destruc-
tion in RA. We have also shown that a large amount of
these proteins was produced in the supernatants of the
synovial tissue-derived inflammatory cells [5]. Both indo-
methacin and EP4-specific antagonist significantly
enhanced spontaneous production of M-CSF, TNF-a and
MMP-9 by the synovial tissue-derived inflammatory cells.
Fig. 2 Effect of periodic hypoxia on the in vitro tissue growth.
Synovial tissue-derived inflammatory cells were incubated in medium
alone (n = 7) under either 1 % oxygen for 24 h twice a week
(periodic hypoxia: hypo) or 21 % oxygen (normoxia: normo) for
4 weeks, and the development of in vitro tissue growth was observed
and scored as described in ‘‘Materials and methods’’. Cumulative
tissue growth scores observed in 1 % oxygen (hypoxia) and 21 %
oxygen (normoxia). Date was analysed versus normoxia by Wicoxon
signed rank test: *P \ 0.05
Fig. 3 Effect of periodic hypoxia on the PGE2 release by rheumatoid
synovial tissues. Synovial tissue-derived inflammatory cells were
incubated in medium alone (n = 13). The level of PGE2 in culture
supernatants was measured by enzyme-linked immunosorbent assay.
Date was analysed versus normoxia by Wilcoxon signed rank test:
*P \ 0.01
1072 Mod Rheumatol (2013) 23:1069–1075
123
This effect was significantly more enhanced under periodic
hypoxia than normoxia. On the other hand, there was no
significant difference of the effect of indomethacin or EP4-
antagonist on the spontaneous production of IL-6 between
normoxia and periodic hypoxia (Table 1).
Discussion
Rheumatoid arthritis is characterized by chronic inflam-
mation of the synovial membrane, which results in the
development of an invasive pannus tissue and the
Fig. 4 Effect of indomethacin (Ind) and EP4-specific antagonist
(EP4-ant) on the in vitro tissue growth. Synovial tissue-derived
inflammatory cells were cultured with indomethacin (1 lM), EP4-
specific antagonist (100 nM) for 4 weeks under either 1 % oxygen for
24 h twice a week (periodic hypoxia: hypo) or 21 % oxygen
(normoxia: normo), and the development of in vitro tissue growth
was observed and scored as described in ‘‘Materials and methods’’.
Cumulative tissue growth scores observed with indomethacin, EP4-
specific antagonist were compared to that observed with medium
alone. The relative value represents the percent of the cumulative
tissue growth score observed with indomethacin and EP4-specific
antagonist compared to that observed with medium alone. *P \ 0.05,
by Wilcoxon signed rank test. Both the exogenous PGE1 and an EP4-
specific agonist significantly inhibited the in vitro tissue growth in the
presence of indomethacin. There was no significant difference
between normoxia and periodic hypoxia
Table 1 Effect of periodic hypoxia on the production of cytokines
and growth factor. Tumor necrosis factor-a (TNF-a), macrophage
colony-stimulated factor (M-CSF), matrix metalloproteinase-9
(MMP-9), interleukin-6 (IL-6) synthesis in response to normoxic
and hypoxic synovial tissue-derived inflammatory cells supernatants
Each cytokine and growth factor was measured by enzyme-linked immunosorbent assay (n = 12)
The relative value represents the TNF-a, M-CSF, MMP-9 observed with indomethacin and EP4-specific antagonist compared to that observed
with medium alone
* P \ 0.05, by Wilcoxon signed rank test
** P \ 0.05, by Wilcoxon signed rank test, versus medium� No significance, by Wilcoxon signed rank test, versus medium
Mod Rheumatol (2013) 23:1069–1075 1073
123

subsequent destruction of cartilage and bone. The hyper-
plasia of the synovial lining layer results in microenvi-
ronmental alteration causing hypoxia and hypoperfusion.
The microenvironment of the inflamed joint is character-
ized by a low partial pressure of oxygen. Low oxygen
tension measurements were first recorded in the synovial
fluid of patients with rheumatoid arthritis [17], and sub-
sequent studies demonstrated decreased oxygen tension
and glucose levels alongside raised carbon dioxide, lactate
and acetate levels, consistent with anaerobic metabolism
[18, 19].
It is suggested that hypoxia plays a pivotal role in the
development and persistence of synovial inflammation and
subsequent joint destruction in both the adjuvant-induced
arthritis model [20] and rheumatoid synovial cells through
the induction of hypoxia-inducible factor-1a (HIF-1a) and
inflammatory cytokines [21–24]. However, little is known
about the net effects of hypoxia, cytokine and cell–cell
interaction that might contribute to the development of
pannus tissue. Therefore, a human cell model that repre-
sents this complex system of the aberrant synovial over-
growth will be useful to study the role of hypoxia in the
pathogenesis of rheumatoid arthritis. We have reported that
synovial tissue-derived inflammatory cells showed spon-
taneous development of the aberrant synovial overgrowth
and pannus-like tissue in vitro [5].
The objective of our study was, therefore, to examine the
potential in vivo consequences of periodic hypoxia in
rheumatoid arthritis in terms of synovial proliferation and
pannus growth, by periodically exposing synovial tissue-
derived inflammatory cells to 1 % oxygen (hypoxia). Using
a human cellular model of pannus, we have demonstrated
that periodic hypoxia induced negative feedback regulation
by PGE2 production, as well as proinflammatory cascades
including TNF-a, M-CSF and MMP-9. The positive effect
of hypoxia on aberrant synovial overgrowth by the synovial
tissue-derived inflammatory cells was effectively down-
regulated by the simultaneously induced endogenous PGE2.
Inhibition of either endogenous prostanoids by indo-
methacin or EP4 receptor by the EP4-specific antagonist
resulted in the enhancement of aberrant synovial over-
growth. Therefore, the net effects of periodic hypoxia may
depend upon the balance between the positive and negative
regulatory responses.
There is a controversy in the effects of hypoxia on PGE2
production. Demasi et al. [13] reported that hypoxia
enhanced IL-1b-stimulated COX-2 expression and PGE2
production by fibroblast-like synoviocytes. Choi et al. [25]
reported that hypoxia decreased IL-1b-stimulated PGE2
production by fibroblast-like synoviocytes even though it
increased expression of COX-2. The major differences
between their and our results may be the fact that we
measured spontaneous production of PGE2 during the
mixed cell–cell interaction. Choi et al. [25] also reported
that hypoxia did not affect IL-1b-stimulated IL-6 and IL-8
production by fibroblast-like synoviocytes. This is consis-
tent with our finding that periodic hypoxia did not show
any effect on the production of IL-6. On the other hand,
M-CSF production was significantly enhanced by periodic
hypoxia in the present study. Our preliminary experiments
indicated that M-CSF production was best correlated with
an in vitro tissue growth score among other cytokines such
as TNF-a, IL-6 and IL-8. Furthermore, it was reported that
the M-CSF level correlated with disease activity in rheu-
matoid arthritis [26].
The recent study clearly demonstrated that COX-2
specific (as well as the bispecific) NSAIDs, by reducing the
levels of PGE2, were able to increase expression of the key
proinflammatory cytokine TNF-a in rheumatoid synovium
[27]. We also showed that endogenous PGE2 inhibition by
NSAIDs enhanced aberrant synovial overgrowth and
osteoclastic activity [16].
Proinflammatory cytokines, including TNF-a, IL-1, and
IL-6 play a critical role in the progression of synovitis and
joint destruction mainly through activation of NF-jB [28–
30], while they directly induce COX-2 and PGE2 expres-
sion [29]. PGE2 up-regulates COX-2 expression via EP2/
EP4 receptors and the cyclic adenosine monophosphate-
dependent signaling pathway [31], which in turn modulates
the production of the proinflammatory molecules. The link
between proinflammatory molecules and PGE2 could have
considerable importance in the regulation of inflammatory
cells of rheumatoid arthritis. The paracrine and autocrine
feedback mechanisms via COX-2, PGE2, EP2 and EP4
could help to avoid the potential pathological damage
caused by the excessive production of inflammatory
mediators in response to various biological stimuli such as
periodic hypoxia in rheumatoid arthritis.
Acknowledgments The authors are grateful to Dr. Kuniomi
Yamasaki for continuing encouragement and his financial support for
our work, and we also thank Kiyomi Matsuo for the excellent tech-
nical assistance.
Conflict of interest Hidehiro Yamada received research funds from
Ono Pharmaceuticals Co. All other authors have declared no conflicts
of interest.
References
1. Harris ED Jr. Rheumatoid arthritis. Pathophysiology and impli-
cations for therapy. N Engl J Med. 1990;322:1277–89.
2. Mansson B, Carey D, Alini M, Ionescu M, Rosenberg LC, Poole
AR, et al. Cartilage and bone metabolism in rheumatoid arthritis.
Differences between rapid and slow progression of disease
identified by serum markers of cartilage metabolism. J Clin
Invest. 1995;95:1071–7.
1074 Mod Rheumatol (2013) 23:1069–1075
123

3. Woodruff T, Blake DR, Freeman J, Andrews FJ, Salt P, Lunec J.
Is chronic synovitis an example of reperfusion injury? Ann
Rheum Dis. 1986;45:608–11.
4. Ng CT, Biniecka M, Kennedy A, McCormick J, Fitzgerald O,
Bresnihan B, et al. Synovial tissue hypoxia and inflammation
in vivo. Ann Rheum Dis. 2010;69:1389–95.
5. Nozaki T, Takahashi K, Ishii O, Endo S, Hioki K, Mori T, et al.
Development of an ex vivo cellular model of rheumatoid arthritis:
critical role of CD14-positive monocyte/macrophages in the
development of pannus tissue. Arthritis Rheum. 2007;56:
2875–85.
6. Suzawa T, Miyaura C, Inada M, Maruyama T, Sugimoto Y,
Ushikubi F, et al. The role of prostaglandin E receptor subtypes
(EP1, EP2, EP3, and EP4) in bone resorption: an analysis using
specific agonists for the respective EPs. Endocrinology. 2000;
141:1554–9.
7. Miyamoto M, Ito H, Mukai S, Kobayashi T, Yamamoto H, Ko-
bayashi M, et al. Simultaneous stimulation of EP2 and EP4 is
essential to the effect of prostaglandin E2 in chondrocyte dif-
ferentiation. Osteoarthritis Cartilage. 2003;11:644–52.
8. Jones RL, Qian YM, Chan KM, Yim AP. Characterization of a
prostanoid EP3-receptor in guinea-pig aorta: partial agonist
action of the non-prostanoid ONO-AP-324. Br J Pharmacol.
1998;125:1288–96.
9. Kabashima K, Saji T, Murata T, Nagamachi M, Matsuoka T, Segi
E, et al. The prostaglandin receptor EP4 suppresses colitis,
mucosal damage and CD4 cell activation in the gut. J Clin Invest.
2002;109:883–93.
10. Akhavani MA, Madden L, Buysschaert I, Sivakumar B, Kang N,
Paleolog EM. Hypoxia upregulates angiogenesis and synovial
cell migration in rheumatoid arthritis. Arthritis Res Ther. 2009;
11:R64.
11. Ke J, Liu Y, Long X, Li J, Fang W, Meng Q, et al. Up-regulation
of vascular endothelial growth factor in synovial fibroblasts from
human temporomandibular joint by hypoxia. J Oral Pathol Med.
2007;36:290–6.
12. del Rey MJ, Izquierdo E, Caja S, Usategui A, Santiago B, Gal-
indo M, et al. Human inflammatory synovial fibroblasts induce
enhanced myeloid cell recruitment and angiogenesis through a
hypoxia-inducible transcription factor 1alpha/vascular endothe-
lial growth factor-mediated pathway in immunodeficient mice.
Arthritis Rheum. 2009;60:2926–34.
13. Demasi M, Cleland LG, Cook-Johnson RJ, James MJ. Effects of
hypoxia on the expression and activity of cyclooxygenase 2 in
fibroblast-like synoviocytes: interactions with monocyte-derived
soluble mediators. Arthritis Rheum. 2004;50:2441–9.
14. Lee JJ, Natsuizaka M, Ohashi S, Wong GS, Takaoka M, Mic-
haylira CZ, et al. Hypoxia activates the cyclooxygenase-2-pros-
taglandin E synthase axis. Carcinogenesis. 2010;31:427–34.
15. Liu XH, Kirschenbaum A, Lu M, Yao S, Dosoretz A, Holland JF,
et al. Prostaglandin E2 induces hypoxia-inducible factor-1alpha
stabilization and nuclear localization in a human prostate cancer
cell line. J Biol Chem. 2002;277:50081–6.
16. Shibata-Nozaki T, Ito H, Mitomi H, Akaogi J, Komagata T,
Kanaji T, et al. Endogenous prostaglandin E(2) inhibits aberrant
overgrowth of rheumatoid synovial tissue and the development of
osteoclast activity through EP4 receptor. Arthritis Rheum. 2011;
63:2595–605.
17. Lund-Olesen K. Oxygen tension in synovial fluids. Arthritis
Rheum. 1970;13:769–76.
18. Treuhaft PS, DJ MC. Synovial fluid pH, lactate, oxygen and
carbon dioxide partial pressure in various joint diseases. Arthritis
Rheum. 1971;14:475–84.
19. Ahlqvist J. A hypothesis on the pathogenesis of rheumatoid and
other non-specific synovitides. IV A. The possible intermediate
role of local hypoxia and metabolic alterations. Med Hypotheses.
1984;13:257–302.
20. Peters CL, Morris CJ, Mapp PI, Blake DR, Lewis CE, Winrow
VR. The transcription factors hypoxia-inducible factor 1alpha
and Ets-1 colocalize in the hypoxic synovium of inflamed joints
in adjuvant-induced arthritis. Arthritis Rheum. 2004;50:291–6.
21. Knowles HJ, Athanasou NA. Acute hypoxia and osteoclast
activity: a balance between enhanced resorption and increased
apoptosis. J Pathol. 2009;218:256–64.
22. Utting JC, Flanagan AM, Brandao-Burch A, Orriss IR, Arnett
TR. Hypoxia stimulates osteoclast formation from human
peripheral blood. Cell Biochem Funct. 2010;28:374–80.
23. Muz B, Khan MN, Kiriakidis S, Paleolog EM. Hypoxia. The role
of hypoxia and HIF-dependent signalling events in rheumatoid
arthritis. Arthritis Res Ther. 2009;11:201.
24. Cha HS, Ahn KS, Jeon CH, Kim J, Song YW, Koh EM. Influence
of hypoxia on the expression of matrix metalloproteinase-1, -3
and tissue inhibitor of metalloproteinase-1 in rheumatoid synovial
fibroblasts. Clin Exp Rheumatol. 2003;21:593–8.
25. Choi HM, da Oh H, Bang JS, Yang HI, Yoo MC, Kim KS.
Differential effect of IL-1beta and TNFalpha on the production of
IL-6, IL-8 and PGE2 in fibroblast-like synoviocytes and THP-1
macrophages. Rheumatol Int. 2010;30:1025–33.
26. Yang PT, Kasai H, Xiao WG, Zhao LJ, He LM, Yamashita A,
et al. Increased expression of macrophage colony-stimulating
factor in ankylosing spondylitis and rheumatoid arthritis. Ann
Rheum Dis. 2006;65:1671–2.
27. Page TH, Turner JJ, Brown AC, Timms EM, Inglis JJ, Brennan
FM, et al. Nonsteroidal anti-inflammatory drugs increase TNF
production in rheumatoid synovial membrane cultures and whole
blood. J Immunol. 2010;185:3694–701.
28. Gomez PF, Pillinger MH, Attur M, Marjanovic N, Dave M, Park
J, et al. Resolution of inflammation: prostaglandin E2 dissociates
nuclear trafficking of individual NF-kappaB subunits (p65, p50)
in stimulated rheumatoid synovial fibroblasts. J Immunol.
2005;175:6924–30.
29. Barnes PJ, Karin M. Nuclear factor-kappaB: a pivotal transcrip-
tion factor in chronic inflammatory diseases. N Engl J Med.
1997;336:1066–71.
30. Brown KD, Claudio E, Siebenlist U. The roles of the classical and
alternative nuclear factor-kappaB pathways: potential implica-
tions for autoimmunity and rheumatoid arthritis. Arthritis Res
Ther. 2008;10:212.
31. Hinz B, Brune K, Pahl A. Prostaglandin E(2) upregulates
cyclooxygenase-2 expression in lipopolysaccharide-stimulated
RAW 264.7 macrophages. Biochem Biophys Res Commun.
2000;272:744–8.
Mod Rheumatol (2013) 23:1069–1075 1075
123





























