Embed Size (px)
Citation preview

IGF1 (Insulin-like growth factor 1)
aneb
od hypotalamu až k buňce
Radek Kučera
Plzeň 2015

2
Autoři: PharmDr. Radek Kučera, Ph.D. Imunoanalytická laboratoř, FN a LF v Plzni, Univerzita Karlova v Praze
Prof. MUDr. Ondřej Topolčan, CSc. Imunoanalytická laboratoř, FN a LF v Plzni, Univerzita Karlova v Praze
Doc. RNDr. Ladislav Pecen, CSc. Imunoanalytická laboratoř , FN a LF v Plzni, Univerzita Karlova v Praze Ústav informatiky, Akademie věd České Republiky
Doc. RNDr. Judita Kinkorová, CSc. Imunoanalytická laboratoř , FN a LF v Plzni, Univerzita Karlova v Praze
MUDr. Šárka Svobodová, Ph.D. Imunoanalytická laboratoř, FN a LF v Plzni, Univerzita Karlova v Praze III. interní klinika, Všeobecná fakultní nemocnice v Praze
MUDr. Ondřej Fiala, Ph.D. Onkologická a radioterapeutická klinika, FN a LF v Plzni, Univerzita Karlova v Praze Biomedicínské centrum, LF v Plzni, Univerzita Karlova v Praze Recenzent: Prof. RNDr. Richard Hampl, DrSc. Endokrinologický ústav AV, ČR
Jazyková úprava: Mgr. Josef Kučera
Tato kniha byl podpořena projektem Ministerstva školství
OPVK CZ (1.07/2.3.00/20.0040) a BBMRI_CZ 23754

3
OBSAH
Předmluva ............................................................................................................................. 4
1 Hypotalamo-hypofyzární systém řízení sekrece GH (R.Kučera) .................................... 5
1.1 IGF1 základní charakteristika .................................................................................10
1.2 IGF1 vazebné proteiny ...........................................................................................12
1.3 IGF1 receptory .......................................................................................................14
1.4 Signální dráha IGF1 ...............................................................................................15
2 IGF1, IGFBP3 - změny hladin v závislosti na věku a pohlaví (R. Kučera, L. Pecen) .....18
3 IGF1 a mechanizmus nádorové transformace (R. Kučera) ...........................................26
4 Systém IGF1 a vybrané nádorové diagnózy (R. Kučera, O. Topolčan, J. Kinkorová) ....28
4.1 Nádory kolorekta ....................................................................................................28
4.2 Nádory prsu............................................................................................................29
4.3 Karcinom prostaty ..................................................................................................31
4.4 Nádory plic .............................................................................................................32
4.5 Melanomy ..............................................................................................................33
4.6 Ovariální karcinom .................................................................................................34
4.7 Karcinom děložního čípku ......................................................................................34
4.8 Karcinom žaludku ...................................................................................................35
4.9 Nádory jater............................................................................................................35
4.10 Karcinom pankreatu ............................................................................................36
4.11 Nádory štítné žlázy .............................................................................................36
5 Systém IGF1 a kancerogeneze - vliv dalších faktorů (R. Kučera, Š. Svobodová) .........37
6 Blokátory růstových receptorů (R. Kučera, O. Fiala) .....................................................41
6.1 Monoklonální protilátky proti IGF1R .......................................................................41
6.2 Monoklonální protilátky proti ligandům IGF1R ........................................................42
6.3 Ihibitory IGF1 receptorové tyrozinkinázy .................................................................42
6.4 Kombinace IGF1R inhibitorů s chemoterapií nebo s dalšími inhibitory růstu ...........42
6.5 Prediktivní markery terapie, indikace a omezení ....................................................43
Seznam zkratek ....................................................................................................................44
Literatura ..............................................................................................................................47

4
Předmluva
Signální dráha IGF1 patří k nejdůležitějším růstovým drahám v organizmu.
Tato dráha byla studována řadu let v souvislosti s růstem a přežíváním buněk. Naše
publikace vychází z posledních poznatků a z našich vlastních zkušeností a mapuje
vlastně systém řízení růstu v organizmu – od hypotalamu až po buněčnou úroveň.
Největší důraz je kladen na informace o IG1. IGF1 je biomarker měnící se s věkem,
což dělá zkoumání a hodnocení poznatků o něm ještě o něco složitější. Jako
demonstrace náročnosti práce s takovým biomarkerem je zde zařazena kapitola o
referenčních hodnotách a způsobu jejich tvorby. Signální dráha IGF1 je léta
zkoumána pro její vliv na vznik a rozvoj nádorů. Tento vztah jsme zkoumali i v naší
laboratoři a v této publikaci je diskuzi na toto téma věnován značný prostor. IGF1
signální dráha je propojena s dalšími signálními drahami a je ovlivňována nejen
IGF1, ale i dalšími biomarkery, což dokládá komplexnost jejího působení. Veškeré
poznatky vyústily ve výrobu různých druhů blokátorů účinku IGF1. Tyto molekuly se
jeví jako nadějné pro budoucí využití v onkologické léčbě. Řada takových molekul je
dnes ve fázi klinického zkoušení. Dosavadní zkušenosti naznačují, že použití nově
vyvíjených léků bude přísně individuální, což zpětně klade značné nároky na
vymezení identifikačních a selekčních markerů, které by umožnily individualizaci
terapie a tím zajistily její funkčnost a maximální efektivitu léčby.
Přejeme příjemné čtení.
Autoři

5
1 Hypotalamo-hypofyzární systém řízení sekrece GH
(R.Kučera)
Signální dráha inzulínu podobného růstového faktoru 1 (IGF1, insuline-like
growth factor 1, somatomedin) patří k nejdůležitějším růstovým drahám v organizmu.
Tato dráha byla intenzivně studována od počátku 80. let minulého století. Z úrovně
séra se postupem času podařilo postoupit na mapování výskytu IGF1 ve tkáních.
Podařilo se zjistit, že na buněčné úrovni je účinek zprostředkován povrchovým
specifickým receptorem. Je popsána lokace genů, které kódují IGF1, jeho vazebné
proteiny a specifické receptory. Tyto poznatky nám umožnily pochopit fungování
systému IGF1 a jeho roli v růstu, diferenciaci a přežívání buněk. Na zvířecích
modelech byl dokonce prokázán přímý vliv funkce signální dráhy na délku života
daného organizmu.
Sekrece IGF1 je řízena růstovým hormonem (GH). Úvodem je tedy nutné
vysvětlit principy řízení sekrece GH hypotalamo-hypofyzárním systémem (Schéma
1).
Sekreční kaskáda řízení růstu začíná u hypotalamických hormonů liberinů a
statinů, které buď stimulují nebo inhibují sekreci hormonů adenohypofýzy, v našem
případě GH. Za stimulaci sekrece GH je odpovědný somatoliberin (GHRH, growth
hormone releasing hormone), který se skládá ze 44 aminokyselin. Biologický poločas
somatoliberinu je 50 minut. Somatoliberin se využívá k diferenciální diagnostice
poruch produkce GH. Podání somatoliberinu vede během několika minut ke zvýšení
sekrece GH, vrcholu sekrece je dosaženo po 30 minutách a účinek přetrvává 60-120
minut. Pokud je zaznamenána pozitivní odpověď organizmu na podání
somatoliberinu, porucha je hypotalamická, pokud po podání somatoliberinu nedojde
ke zvýšení produkce GH, porucha je hypofyzární. Při hypotalamických poruchách
produkce GH je možné podáváním somatoliberinu navodit fyziologickou pulzní
sekreci GH (1).

6
Schéma 1: Hypothalamo-hypofyzární systém řízení sekrece hormonů Sekreční kaskáda řízení růstu začíná u hypotalamických hormonů liberinů a statinů,
které buď stimulují nebo inhibují sekreci hormonů adenohypofýzy, v našem případě GH. Za stimulaci sekrece GH je odpovědný somatoliberin, inhibičně působí somatostatin. Transport GH v krvi probíhá pomocí vazby na specifický vazebný protein growth hormone binding protein (GHBP). GH stimuluje sekreci IGF1 a IGF2 v játrech.

7
Inhibičně působí somatostatin (GHIH, growth hormone inhibiting hormone),
který se skládá ze 14 aminokyselin a inhibuje sekreci nejen GH, ale i hormonu
stimulujícího produkci hormonů štítné žlázy - thyroid stimulating hormone (TSH).
Somatostatin je kromě hypothalamu syntetizován také v D-buňkách
Langerhansových ostrůvků pankreatu, v gastrointestinální sliznici a v
parafolikulárních buňkách (C-buňky) štítné žlázy. Somatostatin produkovaný ve
střevě má 28 aminokyselin a má silné inhibiční účinky na další hormony (např.
inzulín, glukagon, gastrín, sekretín). Biologický poločas somatostatinu je kolem 3
minut. Sekrece somatostatinu se zvyšuje při zvýšené hladině GH a IGF1. Pro klinické
využití je nativní somatotropin nevhodný díky krátkému poločasu (2). Pro léčbu se
využívá dlouhopůsobící syntetický analog somatostatinu. Klinicky se somatostatin
využívá především v terapii nadbytku GH při produkci GH nádory hypofýzy (např.
akromegalie).
Produkce GH patří mezi nejvýznamnější úkoly adenohypofýzy. Produkce GH
probíhá v somatotropních buňkách, které tvoří až 50% všech buněk adenohypofýzy.
GH vzniká odštěpením z větší molekuly pre-GH o molekulové hmotnosti 28 000. Tuto
molekulu je též možné detekovat v krvi, nemá však žádný prorůstový účinek. GH je
druhově specifický. Gen pro GH je umístěn na dlouhém raménku chromozomu 17. U
člověka je molekula GH složená ze 191 aminokyselin o molekulové hmotnosti 21
500. Prostorově je to lineární polypeptid, kdy jednořetězcová stočená molekula je
fixována dvěma disulfidickými můstky (3). Přehled základních údajů a fyziologických
funkcí somatoliberinu, somatostatinu a GH uvádí Tabulka 1.
Transport GH v krvi probíhá pomocí vazby na specifický vazebný protein
growth hormone binding protein (GHBP). GHBP je velkým fragmentem extracelulární
domény receptoru pro GH a jeho koncentrace je tedy úměrná počtu receptorů pro
GH v tkáních. Asi polovina množství GH v krvi je vázáná na GHBP, čímž vzniká
rezerva GH, která může vyrovnávat výkyvy v jeho sekreci, druhá polovina GH se
vyskytuje ve volné formě. Volný GH má biologický poločas 20–50 minut.

8
Tabulka 1: Základní charakteristika GHRH, GHIH a GH
Hormon Molekulová
hmotnost
Počet
aminokyselinFyziologická funkce
GHRH
5 040
44 Stimulace produkce GH
GHIH
1 638
(somatostatin-14)
14 (28)
14 - Inhibice produkce GH, TSH
28 – Inhibice produkce inzulínu,
glukagonu, gastrínu, sekretínu
GH
21 500
191
stimulace lineárního růstu
↑ proteosyntézy, ↓ proteokatabolizmu
↑ výdej glykogenu, antiinzulínový
účinek
Účinek GH na buňky je zprostředkován specifickým receptorem growth
hormone receptor (GHR). GHR je protein o 620 aminokyselinách patřící do rodiny
cytokinových receptorů. Molekula GH má dvě vazebná místa pro receptor. Po
navázání na jednu receptorovou podjednotku druhé vazebné místo přitáhne druhou
podjednotku receptoru, čímž vzniká homodimer, který je nezbytný pro správnou
funkci receptoru a následnou aktivaci intracelulární signální dráhy (4). Po aktivaci
receptoru pokračuje přenos signálu kaskádou enzymů a jejich substrátů (Schéma 2).
Fyziologická ranní koncentrace GH v krvi na lačno je u dospělých nižší než 5
ng/ml, někdy však téměř neměřitelná nebo nižší než 2 ng/ml. GH se do krve
vyplavuje v pulzech. Sekrece GH je spojená se spánkem a závisí na věku. Vrcholu
dosahuje za 1–4 hodiny po usnutí. Tyto noční sekreční pulzy činí asi 70% celkového
množství GH vylučovaného za den a jsou vyšší u dětí, s věkem se snižují. Produkce
GH je u zdravých dospělých 40 µg/den, u dospívající 700 µg/den. U dětí do třech
měsíců nemá sekrece GH vztah ke spánku, v pozdějším dětském věku je však
spánková sekrece GH vyšší než v dospělosti.
Biologický účinek GH spočívá především ve stimulaci lineárního růstu,
přičemž většina růstového účinku není způsobena přímým účinkem GH, ale je
zprostředkována IGF1 (nebo IGF2), jehož sekreci v játrech GH stimuluje.
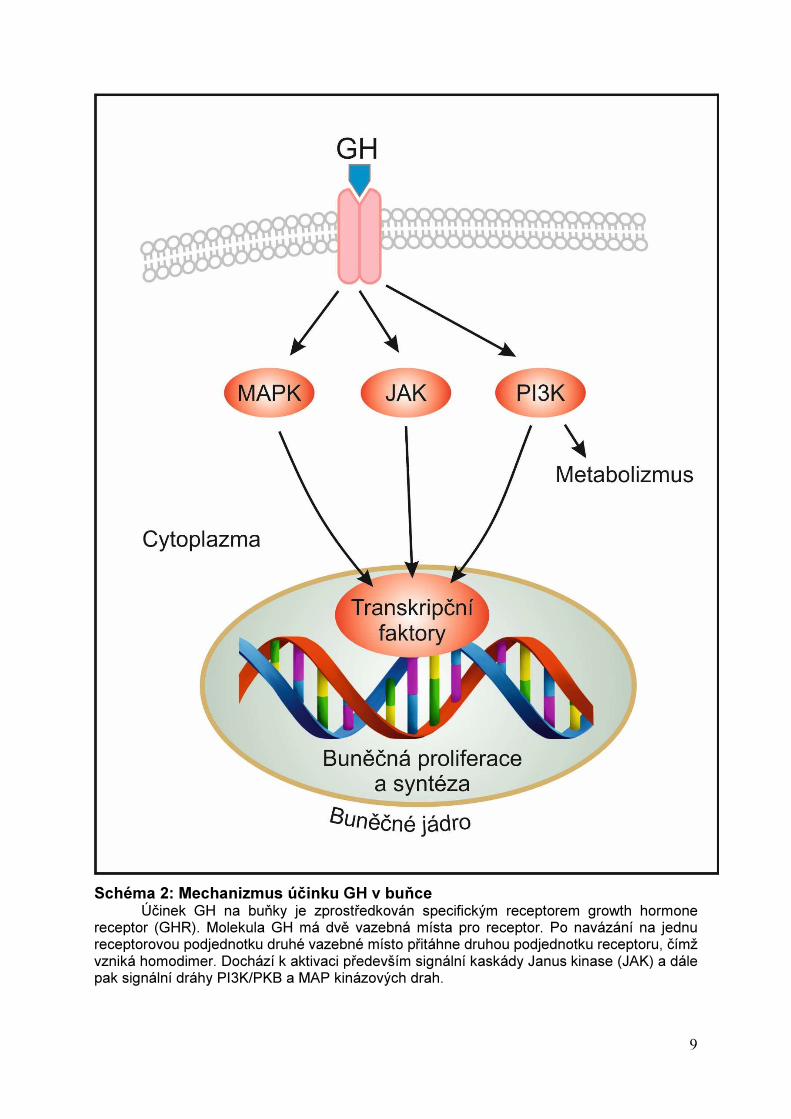
9
Schéma 2: Mechanizmus účinku GH v buňce Účinek GH na buňky je zprostředkován specifickým receptorem growth hormone
receptor (GHR). Molekula GH má dvě vazebná místa pro receptor. Po navázání na jednu receptorovou podjednotku druhé vazebné místo přitáhne druhou podjednotku receptoru, čímž vzniká homodimer. Dochází k aktivaci především signální kaskády Janus kinase (JAK) a dále pak signální dráhy PI3K/PKB a MAP kinázových drah.

10
Obecně lze říci o účincích GH na organizmus toto:
1. GH zvyšuje proteosyntézu tím, že zvyšuje vychytávání aminokyselin a
přímo zvyšuje transkripci a translaci mRNA.
2. GH snižuje proteokatabolizmus tím, že přímo způsobuje uvolnění mastných
kyselin z tukové tkáně.
3. GH zvyšuje výdej glykogenu z jater a ve svalech inhibuje vychytávání
glukózy a její další využití (inzulinorezistence), což je základem jeho diabetogenního
účinku. GH tak může zhoršovat klinický diabetes a při nadprodukci GH (tumor
adenohypofýzy) onemocní diabetem až 25% pacientů s tímto tumorem. GH sice
přímo nestimuluje sekreci inzulínu, ale hyperglykémie, kterou vyvolává, sekundárně
stimuluje pankreas a může nakonec vést až k vyčerpání β-buněk a rozvoji diabetu.
Vztah růstové signální dráhy IGF1 a inzulinorezistence je v současné době
intenzivně studován a bude podrobněji zmíněn v dalších kapitolách.
Úloha GH v prenatálním růstu je pravděpodobně pouze okrajová. Předokládá
se, že během intrauterinního života je růst plodu řízen prostřednictvím systému IGF1
a IGF2. Tyto růstové faktory jsou regulovány prostřednictvím transportu glukózy
placentou. Osa glukóza-inzulín-IGF1,IGF2 se tak primárně uplatňuje v prenatálním
růstu. Postnatální růst je regulován osou GH-IGF1, a proto po narození dochází k
vzestupu hladiny IGF1. Hladiny IGF2 se však během postnatálního života nemění.
1.1 IGF1 základní charakteristika
IGF1 společně též s inzulínu podobným růstovým faktorem 2 (IGF2, insuline-
like growth factor 2) jsou peptidy, které se fyziologicky podílejí na řízení růstu,
metabolizmu, přežívání a diferenciaci buněk a jsou regulovány růstovým hormonem
(GH, growth hormone). IGF2 se uplatňuje především v prenatálním období a IGF1
v období postnatálním (Tabulka 2). IGF1 a IGF2 se skládají z malých peptidů, které
jsou z 50% homologní s proinzulínem (5). Oba typy IGF jsou syntetizovány v mnoha
fetálních a dospělých tkáních a jsou obsažené dokonce i v mateřském mléce.
Až 95 % obou typů IGF cirkuluje v krvi vázáno na vazebné proteiny. IGF1 a
IGF2 mají své specifické vazebné proteiny (IGFBPs, insulin-like growth factor binding
proteins). Biologický poločas volných frakcí IGF1 a IGF2 se odhaduje na méně než
10 minut. Komplex IGF1/IGFBP3 či IGF2/IGFBP3 má biologický poločas asi 12
hodin.

11
IGF1 je bazický peptid s nízkou molekulovou hmotností 7 649. Je tvořen
jednoduchým řetězcem o 70 aminokyselinách. IGF1 je produkován ve velkém
množství v játrech (přibližně 10 mg/den). Tato produkce je pevně řízena GH. IGF1 je
produkován lokálně i v mnoha jiných tkáních např. ledvin, srdce, plic, tukové tkáni a
tkáních různých žláz. IGF1 je produkován také chondroblasty, fibroblasty a
osteoklasty. Na orgánové úrovni IGF1 účinkuje autokrinním i parakrinním
mechanizmem a je schopen komunikovat s ostatními místně produkovanými
růstovými faktory a steroidními hormony (6). Účinek v buňkách je zprostředkován
vlastním IGF1 receptorem. Syntézu IGF1 zvyšují hormony štítné žlázy a mírně též
prolaktin. Estrogeny a kortikoidy naopak omezují produkci IGF1. Sekrece IGF1
nezávisí jenom na účinku GH, ale také na výživě. Po dobu hladovění hladina IGF1
klesá a nereaguje ani na podání GH. Hladina IGF1 je vhodným ukazatelem
realimentace při malnutričních stavech. Snížené hladiny IGF1 jsou pozorovány po
odstranění hypofýzy.
Tabulka 2: Základní charakteristika IGF1 a IGF2
Růstový faktor Molekulová
hmotnost
Počet
aminokyselinFyziologická funkce
IGF1
7 464
70
Řízení růstu, metabolizmu, přežívání a diferenciace buněk
(především v postnatálním vývoji)
IGF2
7 400
67
Řízení růstu, metabolizmu, přežívání a diferenciace buněk
(především v prenatálním vývoji)
Z diagnostického hlediska je IGF1 přínosným vyšetřením zejména při
sledování akromegalií a deficitu růstového hormonu. Jeho výhodou je to, že na rozdíl
od sekrece GH, která probíhá pulzně, je jeho hladina v séru dlouhodobě konstantní.
U akromegalie jsou hladiny zvýšené a u deficitu GH jsou hladiny IGF1 snížené.
Důležitost IGF1 pro růst a vývoj organizmu byla zkoumána na zvířecích modelech
narušením genu pro IGF1 tzv. knock out (K.O.) metodou. IGF1 K.O. myši vykazovaly
ve srovnání s normálními jedinci o 40% sníženou porodní hmotnost, velmi vysokou
perinatální úmrtnost, opožděnou osifikaci, málo rozvinuté svalstvo a plicní tkáň,
stejně jako neplodnost (7).

12
IGF2 je neutrální peptid o molekulové hmotnosti 7400 Da, jehož řetězec je
tvořený 67 aminokyselinami. IGF2 je strukturálně blízký IGF1, ale je méně závislý na
regulaci GH. Účinek je zprostředkován specifickým IGF2 receptorem. IGF2 byl
dlouho považována za plodovou formu IGF1. Dnes již víme, že je toto rozdělení
překonané, avšak přesto je nesporné, že IGF2 hraje velmi důležitou roli během
embryogeneze. Normální hladiny v séru u zdravých lidí se pohybují od 400 do 900
μg/l.
Byly prováděny studie s cíleným narušením genu pro IGF2. IGF2 K.O. myši
měly o 40% nižší porodní hmotnost ve srovnání s normálními jedinci (8). Tyto myši
se na rozdíl od IG1 K.O. myší vyvinuly v normální jedince s trpasličím vzrůstem.
1.2 IGF1 vazebné proteiny
IGFBPs se nacházejí v různých tělních tekutinách a tkáních. IGFBPs jsou
syntetizovány v játrech a mohou být regulovány mimo jiné i tím, že IGF1 přímo
podporuje proteolytickou degradaci některých IGFBPs (IGFBP4) do fragmentů, které
nevážou IGF1, a tím IGF1 vlastně zvyšuje svojí vlastní biologickou dostupnost v
tělních tekutinách.
IGFBP zvyšují poločas IGF1 v séru a inhibují nebo podporují působení IGF1
vazbou na receptory cílových buněk. Navíc tyto vazebné proteiny fungují jako
zásobárna IGF1 v mezibuněčném prostoru. Některé IGFBP brání in vitro růstu
buněk. Buňky malobuněčného karcinomu plic produkují vazebné proteiny pro IGF1,
které se liší od těch nalezených v játrech. To podporuje teorii, že by mohly fungovat
jako prostředníci v autokrinní nebo parakrinní regulaci růstu těchto nádorových
buněk. V současné době je známo nejméně šest různých IGFBP, které se liší v jejich
vazebné účinnosti (Tabulka 3). Všeobecně vžitá je řada IGFBP1 až IGFBP6, přičemž
v literatuře můžeme dále najít i IGFBP7 a IGFBP8, které vznikly přejmenováním již
objevených proteinů, když bylo zjištěno, že tyto váží IGF1 popř. IGF2.
IGFBP1 se nachází převážně v placentě a plodové vodě. Zvýšené hladiny se
nacházejí u novorozenců. Nižší hladiny pak u ostatní populace. IGFBP1 má stejnou
afinitu k IGF1 i IGF2. IGFBP2 se nachází především v mozkomíšním moku a má
vysokou vazebnou afinitu k IGF2. Dominantní vazebný protein v séru je IGFBP3,
který vykazuje podobnou afinitu k IGF1 a IGF2. IGFBP3 inhibuje sekreci folikuly
stimulujícího hormonu (FSH). Výrazně snížené hladiny IGFBP3 jsou pozorovány u

13
pacientů s nedostatkem růstového hormonu, zatímco výrazně zvýšené hladiny jsou
pozorovány u pacientů s vysokými hladinami růstového hormonu (např.
akromegalie). IGFBP4 je převládající vazebná bílkovina IGFs v lidských
osteoblastech. Je totožná s bílkovinou známou jako inhibitor růstu buněk rakoviny
tlustého střeva (colon cancer cell growth inhibitor). IGFBP4 je cílem působení
IGFBP4 specifické proteázy indukované IGFs. Tato posttranslační úprava IGFBP4
zvyšuje tkáňovou dostupnost IGF1 a IGF2. IGFBP5 se vyskytuje nejvíce v ledvinách,
ale také např. v osteoblastech. IGFBP5 vykazuje poměrně nízké afinity k IGF1 a
IGF2. To zvyšuje mitogeneze v přítomnosti IGF1 nebo IGF2 a ale také v
nepřítomnosti exogenních nebo endogenních IGFs, díky snadnému uvolňování
z vazby na IGFBP5 a následné interakci s receptorem. IGFBP6 se vyskytuje v
mozkomíšním moku a má vysokou vazebnou afinitu k IGF2 (9).
Tabulka 3: Základní charakteristika vazebných proteinů IGFBPs
Vazebný protein Molekulová
hmotnost
Vazebná
afinita Výskyt
IGFBP1 34 000 IGF1, IGF2 placenta, plodová voda
IGFBP2 27 000 IGF2 mozkomíšní mok
IGFBP3 53 000 IGF1, IGF2 sérum
IGFBP4 26 000 IGF1, IGF2 osteoblasty
IGFBP5 23 000 IGF1, IGF2
nízká afinita ledviny, osteoblasty
IGFBP6 22 800 IGF2 mozkomíšní mok
V literatuře můžeme dále najít i IGFBP7 a IGFBP8. Název IGFBP7 navrhl
v roce 1996 Oh a jeho tým jako označení pro bílkovinu mac25 popsanou již v roce
1993 Murphym. V roce 1996 u ní byla totiž zjištěna schopnost vázat IGF1 a IGF2.
V porovnání s IGFBP3 je však afinita IGFBP7 k IGF1 a IGF2 100x nižší. Vysoká
exprese IGFBP7 (mac25) byla zjištěna ve stárnoucích epiteliálních buňkách mléčné
žlázy. Na druhou stranu IGFBP7 téměř chybí v estrogen receptor pozitivní tkáni
karcinomu prsu. Autoři označují IGFBP7 jako nádorový supresor. Označení IGFBP8
bylo navrženo v roce 1997 pro lidský tkáňový pojivový růstový faktor CTGF

14
(connective tissue growth factor) po zjištění, že tento faktor má zachovaný IGFBP
motiv a s nízkou afinitou váže IGFs. IGFBP8 byl nalezen v lidské kostní tkáni. Tento
protein má mnohem vyšší afinitu k IGF2 než k IGF1 a potencuje proliferační vliv IGF2
na kostní buňky.
1.3 IGF1 receptory
Účinek IGF1 (IGF2) na buňku se odehrává prostřednictvím specifických
receptorů (Tabulka 4). IGF1 se také váže s receptorem pro inzulín (IR) a naopak.
Heterologní ligand má však k receptoru 100x nižší afinitu. Dále se IGF1 může vázat
na hybridní receptory tvořené z poloviny IR a z poloviny IGF1R podjednotkou (10).
Receptory IGF1 patří do skupiny receptorových protein tyrosin kináz (PTK), které
fosforylují své substrátové bílkoviny na tyrosinových zbytcích. Patří sem receptory
pro většinu peptidických růstových faktorů, které se uplatňují v regulaci růstu a
diferenciace buněk.
IGF1 receptor (IGF1R, CD221) je transmembránový glykoprotein o
molekulové hmotnosti 350 000. Skládá se ze dvou α-podjednotek a dvou β-
podjednotek. Zatímco α-řetězce jsou lokalizovány extracelulárně a váží IGF1, β-
řetězce procházejí membránou do buňky. IGF1R je ze 60 % homologní s receptorem
pro inzulín. Po vazbě ligandu na receptor dojde k autofosforylaci tyrozinu na β-
podjednotce (11). IGF1R je životně důležitý pro přežití buněk, což dokládá IGF1R
K.O. fenotyp u myší. Narušení IGF1R genu vyústilo ve více než 50% snížení
hmotnosti K.O. zvířat oproti normální populaci. Tato zvířata zemřela v
bezprostředním poporodním období na respirační selhání a byly zjištěny
generalizované vývojové abnormality, hypoplazie, abnormální kožní formace,
opožděný vývoj kostí a abnormální morfologie centrálního nervového systému (12).
IGF2 receptor (IGF2R, CD222) je monomerní protein o molekulové hmotnosti
250 000. Jedná se o vazebný protein s vysokou afinitou pro IGF2 a velmi nízkou
afinitu k IGF1 a nulovou afinitou k inzulínu. Cytoplazmatická doména receptoru nemá
vlastní tyrosinkinázovou aktivitu a nemá schopnost přenášet signál. IGF2R není
zapojen do vnitrobuněčné signalizace. IGF2 receptor cíleně váže IGF2, a tím snižuje
biologickou dostupnost vysoce mitogenního IGF2. Některé z účinků IGF2 jsou
zprostředkovány především izoformním inzulínovým receptorem IR-A. IGF2R je
shodný s kation nezávislým manóza-6-fosfát (M6P) receptorem. Je to vlastně
15
multifunkční proteinový receptor, který váže IGF2 na povrchu buněk a manózou-6-
fosfátem (M6P) označené proteiny v Golgiho aparátu uvnitř buňky. IGF2/M6PR jsou
zmutované nebo úplně chybí v buňkách několika druhů rakoviny, což vede k názoru,
že IGF2/M6PR má roli nádorového supresoru (13).
Inzulínový receptor existuje ve dvou izoformách IR-A a IR-B. IR-A je vyjádřen
především v centrálním nervovém systému, krvetvorných buňkách, plodové tkáni a
různých nádorových buňkách. IR-B je přítomen především v tukové tkáni, játrech a
svalech. Hybridní receptory, které vážou IGF1, jsou složeny z jedné poloviny z IR a
z druhé poloviny z IGF1R. Vzhledem k izoformám IR mohou tedy existovat IR-
A/IGF1R a IR-B/IGF1R (14).
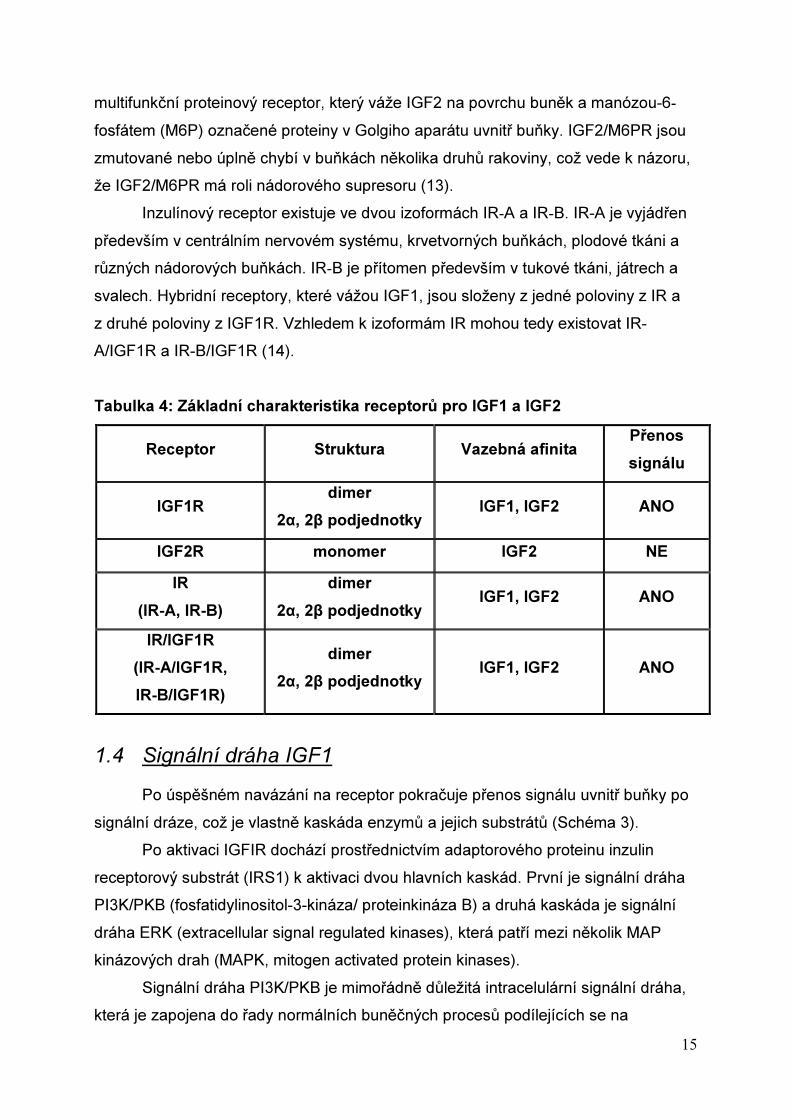
Tabulka 4: Základní charakteristika receptorů pro IGF1 a IGF2
Receptor Struktura Vazebná afinita Přenos
signálu
IGF1R dimer
2α, 2β podjednotky IGF1, IGF2 ANO
IGF2R monomer IGF2 NE
IR
(IR-A, IR-B)
dimer
2α, 2β podjednotky IGF1, IGF2 ANO
IR/IGF1R
(IR-A/IGF1R,
IR-B/IGF1R)
dimer
2α, 2β podjednotky IGF1, IGF2 ANO
1.4 Signální dráha IGF1
Po úspěšném navázání na receptor pokračuje přenos signálu uvnitř buňky po
signální dráze, což je vlastně kaskáda enzymů a jejich substrátů (Schéma 3).
Po aktivaci IGFIR dochází prostřednictvím adaptorového proteinu inzulin
receptorový substrát (IRS1) k aktivaci dvou hlavních kaskád. První je signální dráha
PI3K/PKB (fosfatidylinositol-3-kináza/ proteinkináza B) a druhá kaskáda je signální
dráha ERK (extracellular signal regulated kinases), která patří mezi několik MAP
kinázových drah (MAPK, mitogen activated protein kinases).
Signální dráha PI3K/PKB je mimořádně důležitá intracelulární signální dráha,
která je zapojena do řady normálních buněčných procesů podílejících se na

16
proliferaci, metabolizmu, růstu a přežití buněk. Abnormality této dráhy, způsobené
izoformami a mutacemi různých regulačních komponent, vedou k některým formám
nádorových onemocnění (15). Aktivace PI3K vede ke zrušení inhibičního účinku
regulační podjednotky, která obratem přeměňuje membránový fosfatidylinositol-3,5-
bifosfát (PIP2) na trifosfátovou formu (PIP3). PIP3 následně indukuje fosforylaci PKB
prostřednictvím proteinu PDK1. Komplex PI3K je za normálních okolností negativně
regulován specifickou fosfatázou PTEN. PTEN působí jako negativní regulátor PI3K
dráhy tím, že hydrolyzuje PIP3 na PIP2.
V důsledku ztráty funkce PTEN mutací nebo epigenetickou modifikací se ale
mohou vyskytnout alterace vedoucí k hyperaktivaci PI3K. Ztráta funkce PTEN
podporuje akumulaci PIP3, která vede k deregulaci přenosu signálu přes kaskádu
PI3K/PKB. PKB se účastní ještě řady dalších signálních kaskád a zasahuje tedy do
celé řady buněčných dějů (proteosyntéza, přežívání, proliferace, glukózový
metabolizmus apod.) (16). Aktivace deregulované signální dráhy PI3K/PKB poskytuje
buňce signály pro neomezený růst a přežití.
Druhá signální dráha je kaskáda ERK (extracellular signal regulated kinases).
Po aktivaci IRS1 se signál přenáší přes GTPasu Ras a kinázy Raf a MEK, které
konečně vedou k aktivaci MAP kináz (mitogen activated protein kinases), konkrétně
ERK. Skupina enzymů nazývaných MAP kinázy reguluje expresi genu a případně
proliferaci a diferenciaci buňky (17).
Složitost IGF1 signalizace dokládá fenotyp myší s chybějícím genem pro
IGF1R. Například chybění tohoto genu v buňkách prostaty vedlo k aktivaci ERK
dráhy a způsobilo buněčné proliferace a hyperplazie. Toto zjištění dokazuje, že MAP
kinázové dráhy jsou aktivovány dalšími IGF1R nezávislými mechanizmy. Dále
přítomnost IGF1R signální dráhy usnadnila růst nádorů s poruchou činnosti enzymu
p53 (enzym regulující apoptózu) (18).

17
Schéma 3: Mechanizmus účinku IGFs Biologický účinek IGFs je zprostředkován IGF1 receptorem (IGF1R). IGF1 a IGF2 se
vážou na extracelulární doménu IGF1R. Tyrosin kinasové receptory po aktivaci autofosforylují na cytoplasmatické doméně a přes aktivaci adaptorového proteinu inzulin receptorového substrátu (IRS1/2) se aktivují dvě hlavní signální kaskády. Signální dráha PI3K (fosfatidylinositol-3-kináza)/PKB (proteinkináza B) a signální dráha ERK (extracellular signal regulated kinases), která je jednou z několika MAP kinázových drah (MAPK mitogen activated protein kinases).

18
Poznatky o komplexním působení signální dráhy v živých organizmech vedly
ke studiím ovlivnění dlouhověkosti prováděných na zvířecích modelech. Cílenými
mutacemi v signální dráze IGF1 se povedlo docílit zvýšení průměrné délky života o
20 – 70% (19). Podobná zjištění byla učiněna i u nižších živočišných druhů, než jsou
obratlovci, což znamená, že signální dráha IGF1 je vývojově jednou z nejstarších
růstových drah v živých organizmech vůbec (20).
Můžeme se tedy domnívat, že důkladné poznání signální dráhy IGF1 by
mohlo vést k odhalení mechanizmů stárnutí a možnosti ovlivnění délky lidského
života, avšak údaje o studiu podobných mutací ve vztahu k délce života u lidí
prozatím chybí.
2 IGF1, IGFBP3 - změny hladin v závislosti na věku a
pohlaví (R. Kučera, L. Pecen)
Sérové hladiny IGF1 a IGFBP3 se mění s věkem. Jejich hladiny se začínají
zvyšovat ihned po narození. Nejvyšší koncentrace nacházíme u dětí v pubertě.
V pozdějších letech hladiny IGF1 klesají, přibližně o 10 procent za každých 10 let.
Nejnižší hladiny se vyskytují ve stáří. K nejvýraznějšímu poklesu IGF1 dochází ve
věku 70 let a výše (21). Hladiny IGFBP3 víceméně kopírují vývoj hladin IGF1
v průběhu života. Pokles IGFBP3 s věkem je ale výrazně menší než u IGF1 (22).
V odborné literatuře se ještě kromě IGF1 a IGFBP3 objevuje též vypočítávaný
parametr, a to poměr IGF1/IGFBP3. Tento poměr je využíván především u
nádorových onemocnění jako index biologické dostupnosti IGF1 v séru, kdy se
předpokládá, že vyšší dostupnost IGF1 spíše zvyšuje riziko nádorové transformace a
nižší dostupnost IGF1 způsobená zvýšenou přítomností IGFBP3 působí spíše
protektivním způsobem (23). Poměr IGF1/IGFBP3 s věkem též klesá, což je dáno
poklesem obou biomarkerů, které jsou základem pro výpočet tohoto parametru.
Hladiny IGF1 jsou dobře zmapovány od narození až po nejvyšší věkové
skupiny. Existují práce, které se věnují pouze hladinám IGF1 u dětí a navrhují
referenční hladiny měnící se každý rok, což je pro období, kdy se se produkce
sledovaného biomarkeru dynamicky mění, samozřejmě vhodné (24). Hladiny IGF1
dosahují svého vrcholu kolem 15. roku života. Přibližně ve věku 18 - 20 let nastává

19
období, kdy hladiny IGF1 začínají pomalu klesat a není nutné ani účelné uvádět
referenční hodnoty, které se mění každý rok. Řada prací navrhuje normální hodnoty
se změnou v intervalu 5 let (25).
V imunoanalytické laboratoři ve FN v Plzni jsme též provedli studii na souboru
1022 zdravých osob s cílem navrhnout referenční rozmezí pro IGF1, IGFBP3 a
poměr IGF1/IGFBP3 (26). Došli jsme k závěru, že od 20 let věku je robustnějším
modelem změna referenčních hodnot v intervalu 10 let, vzhledem k poměrně značné
individuální variabilitě v poklesu hladin IGF1 s věkem. Při tvorbě modelů poklesu
IGF1 s věkem záleží též na statistickém přístupu. Pokud zvolíme nelineární model
regrese (Graf 2), můžeme detailněji postihnout určité rozdíly v rychlosti poklesu
hladin v jednotlivých dekádách života. Lineární modely jsou však dle našich
zkušeností robustnější, co se týká individuální variability poklesu IGF1 (Graf 1). Míra
nelinearity poklesu IGF1 s věkem je ale poměrně malá, až na výraznější pokles po
70-ti letech věku. Něco jiného je IGFBP3, kde můžeme vysledovat nelineární pokles,
a to především u žen. Pokles IGFBP3 u žen se stává mnohem výraznější až kolem
60 let věku. Co se týká závislosti hladin IGF1 na pohlaví, studie provedené v tomto
ohledu nedošly k úplně jednotným závěrům. Zajímavé je, že studie prováděné
v posledních letech se spíše přiklánějí k rozdělení referenčních hodnot podle pohlaví
(27, 28) v porovnání se staršími studiemi, které závislost na pohlaví neuvádějí (29).
V našem případě hladiny IGF1 byly statisticky významně vyšší u mužů než u žen a
rozdíly v IGF1 mezí pohlavími klesaly s věkem. U IGFBP3 jsou rozdíly mezi
pohlavími mnohem menší než u IGF1, přičemž pokles u žen je poněkud strmější než
u mužů. Pro poměr IGF1/IGFBP3 jsme zjistili podobný průběh jako u IGF1.
Na základě našich zkušeností jsme došli k závěru, že nejlépe vystihující model
referenčních hodnot lze získat pomocí lineární regrese závislé na věku a pohlaví.
Také výše zmíněné referenční meze po intervalech 10 let věku musí být určovány
zvlášť dle pohlaví (30).
Pro příklad uvádíme tabulky a grafy (Tabulka 5-7, Graf 1-6), které jsme získali
při tvorbě referenčních hodnot pro IGF1, IGFBP3 a poměr IGF1/IGFBP3 v české
populaci s použitím radioimunoanalytických souprav IGF1a IGFBP3 (Immunotech,
Praha, ČR).

20
Tabulka 5: Referenční hodnoty IGF1, IGFBP3, poměr IGF1/IGFBP3 v ČR - muži
IGF1 in males
Age category
N Mean Minimum 2.5th Pctl Lower
Quartile Median Upper Quartile 97.5th Pctl Maximum
20-29 52 242.37 84.43 100.80 202.10 239.70 282.60 400.00 403.10
30-39 62 202.65 100.40 101.90 164.60 192.15 242.70 341.21 341.50
40-49 67 167.66 75.25 103.70 140.00 163.60 188.40 264.90 268.30
50-59 95 160.82 46.60 77.14 132.60 155.83 183.20 296.60 339.30
60-69 75 143.55 37.42 39.30 105.60 145.40 182.70 241.20 264.60
70-79 66 110.15 19.70 33.85 67.70 101.50 147.70 219.60 290.10
80+ 50 96.18 11.20 39.50 65.80 99.30 126.00 150.00 185.30
IGFBP3 in males
Age category
N Mean Minimum 2.5th Pctl Lower
Quartile Median
Upper Quartile
97.5th Pctl Maximum
20-29 52 2879.4 1887.5 1922.0 2510.5 2838.8 3096.5 3959.0 4621.0
30-39 62 2688.7 1238.0 1927.0 2440.0 2651.0 2879.5 3967.5 4142.0
40-49 67 2590.2 1711.0 1966.5 2340.0 2585.5 2769.5 3486.0 3545.0
50-59 95 2547.7 1816.0 1863.0 2288.5 2515.0 2723.5 3514.0 3580.5
60-69 75 2520.1 1204.5 1572.0 2097.5 2474.0 3015.0 3423.5 3802.0
70-79 66 2492.9 1063.5 1631.5 1959.0 2433.8 2867.5 3812.0 3828.5
80+ 50 2499.3 1401.0 1425.0 1893.0 2356.3 3001.0 4570.5 4570.5
IGF1/IGFBP3 molar ratio in males
Age
category N Mean Minimum 2.5th Pctl
Lower Quartile
Median Upper
Quartile 97.5th Pctl Maximum
20-29 52 0.02418 0.00877 0.01389 0.01910 0.02378 0.02939 0.03546 0.03992
30-39 62 0.02147 0.01155 0.01508 0.01730 0.02115 0.02414 0.03407 0.04077
40-49 67 0.01815 0.00907 0.00912 0.01591 0.01725 0.02110 0.02778 0.02963
50-59 95 0.01785 0.01019 0.01108 0.01426 0.01751 0.02050 0.02645 0.02908
60-69 75 0.01671 0.00573 0.00673 0.01308 0.01626 0.01974 0.02973 0.03018
70-79 66 0.01296 0.00376 0.00416 0.00691 0.01219 0.01689 0.02568 0.02935
80+ 50 0.01133 0.00242 0.00388 0.00775 0.01098 0.01355 0.02157 0.02603

21
Tabulka 6: Referenční hodnoty IGF1, IGFBP3, poměr IGF1/IGFBP3 v ČR - ženy
IGF1 in females
Age
category N Mean Minimum
2.5th Pctl
Lower Quartile
Median Upper
Quartile 97.5th Pctl Maximum
20-29 53 188.02 45.26 80.03 143.80 186.10 211.30 359.70 416.00
30-39 58 167.67 71.00 84.72 131.20 161.10 200.40 281.00 315.90
40-49 64 150.55 61.07 66.88 128.40 149.18 167.70 277.40 299.50
50-59 79 135.78 27.03 54.00 96.49 133.90 162.10 249.20 258.20
60-69 75 134.46 29.97 57.80 96.00 130.30 164.00 250.90 283.30
70-79 129 83.55 15.20 16.30 52.90 76.40 112.50 175.80 315.30
80+ 97 74.12 13.30 15.77 39.30 69.30 99.40 170.00 298.10
IGFBP3 in females
Age
category N Mean Minimum 2.5th Pctl
Lower Quartile
Median Upper
Quartile 97.5th Pctl Maximum
20-29 53 3109.8 2334.0 2353.5 2833.5 3043.5 3442.0 3904.0 4332.5
30-39 58 3025.4 1674.0 1961.5 2757.5 3025.5 3348.5 3862.5 4148.5
40-49 64 3044.5 1851.0 2245.5 2700.0 3020.0 3323.0 3910.5 4051.0
50-59 79 2970.0 1379.0 1680.5 2631.0 3014.5 3264.0 3849.0 4085.5
60-69 75 2826.2 2073.0 2135.8 2449.3 2815.5 3145.5 3864.5 3875.5
70-79 129 2513.6 758.5 875.5 1952.8 2531.0 2981.8 4105.5 6760.0
80+ 97 2109.8 775.0 1082.5 1520.5 2076.0 2572.5 3736.0 4341.5
IGF1/IGFBP3 molar ratio in females
Age
category N Mean Minimum 2.5th Pctl
Lower Quartile
Median Upper
Quartile 97.5th Pctl Maximum
20-29 53 0.01674 0.00442 0.00727 0.01280 0.01665 0.01960 0.02726 0.03122
30-39 58 0.01575 0.00738 0.00754 0.01181 0.01455 0.01935 0.02822 0.03218
40-49 64 0.01379 0.00548 0.00720 0.01123 0.01319 0.01562 0.02336 0.02658
50-59 79 0.01362 0.00517 0.00749 0.01024 0.01325 0.01690 0.02221 0.02544
60-69 75 0.01365 0.00355 0.00452 0.00946 0.01427 0.01635 0.02348 0.02359
70-79 129 0.00934 0.00167 0.00247 0.00578 0.00876 0.01146 0.02295 0.04531
80+ 97 0.00965 0.00286 0.00310 0.00623 0.00870 0.01272 0.01812 0.03249

22
Tabulka 7: Referenční hodnoty IGF1, IGFBP3, poměr IGF1/IGFBP3 v ČR – celá populace
IGF1 in the whole population
Age category
N Mean Minimum 2.5th Pctl
Lower Quartile
Median Upper
Quartile 97.5th Pctl Maximum
20-29 105 214.93 45.26 84.43 172.10 207.40 253.60 400.00 416.00
30-39 120 185.74 71.00 90.10 144.47 181.85 223.10 312.50 341.50
40-49 131 159.30 61.07 75.94 132.80 155.50 179.80 268.30 299.50
50-59 174 149.45 27.03 61.40 115.30 148.15 172.60 263.95 339.30
60-69 150 139.00 29.97 49.20 101.80 139.55 171.10 241.20 283.30
70-79 195 92.55 15.20 19.70 55.60 83.90 119.50 199.10 315.30
80+ 147 81.62 11.20 15.77 46.30 74.60 108.90 170.00 298.10
IGFBP3 in the whole population
Age category
N Mean Minimum 2.5th Pctl
Lower Quartile
Median Upper
Quartile
97.5th Pctl Maximum
20-29 105 2995.7 1887.5 2110.5 2734.5 2962.5 3291.5 3959.0 4621.0
30-39 120 2858.7 1238.0 1927.0 2573.5 2820.5 3207.0 3967.5 4148.5
40-49 131 2819.6 1711.0 1966.5 2461.5 2755.5 3188.5 3828.0 4051.0
50-59 174 2776.3 1379.0 1816.0 2448.5 2713.0 3115.0 3731.5 4085.5
60-69 150 2656.2 1204.5 1733.5 2262.0 2650.3 3033.0 3831.0 3875.5
70-79 195 2507.2 758.5 941.5 1959.0 2465.8 2932.0 4068.0 6760.0
80+ 147 2242.6 775.0 1088.5 1672.0 2178.0 2680.0 3913.5 4570.5
IGF1/IGFBP3 molar ratio in the whole population
Age category
N Mean Minimum 2.5th Pctl
Lower Quartile
Median Upper
Quartile
97.5th Pctl Maximum
20-29 105 0.02042 0.00442 0.00740 0.01549 0.01919 0.02518 0.03439 0.03992
30-39 120 0.01858 0.00738 0.00887 0.01454 0.01799 0.02263 0.03218 0.04077
40-49 131 0.01595 0.00548 0.00766 0.01268 0.01562 0.01934 0.02710 0.02963
50-59 174 0.01556 0.00517 0.00757 0.01232 0.01546 0.01879 0.02544 0.02908
60-69 150 0.01535 0.00355 0.00573 0.01161 0.01501 0.01888 0.02876 0.03018
70-79 195 0.01046 0.00167 0.00262 0.00612 0.00938 0.01289 0.02500 0.04531
80+ 147 0.01022 0.00242 0.00310 0.00720 0.00937 0.01289 0.02157 0.03249

23
Graf 1: Lineární regresní model pro IGF1 – muži, ženy
Graf 2: Lineární regresní model pro IGFBP3 – muži, ženy
24
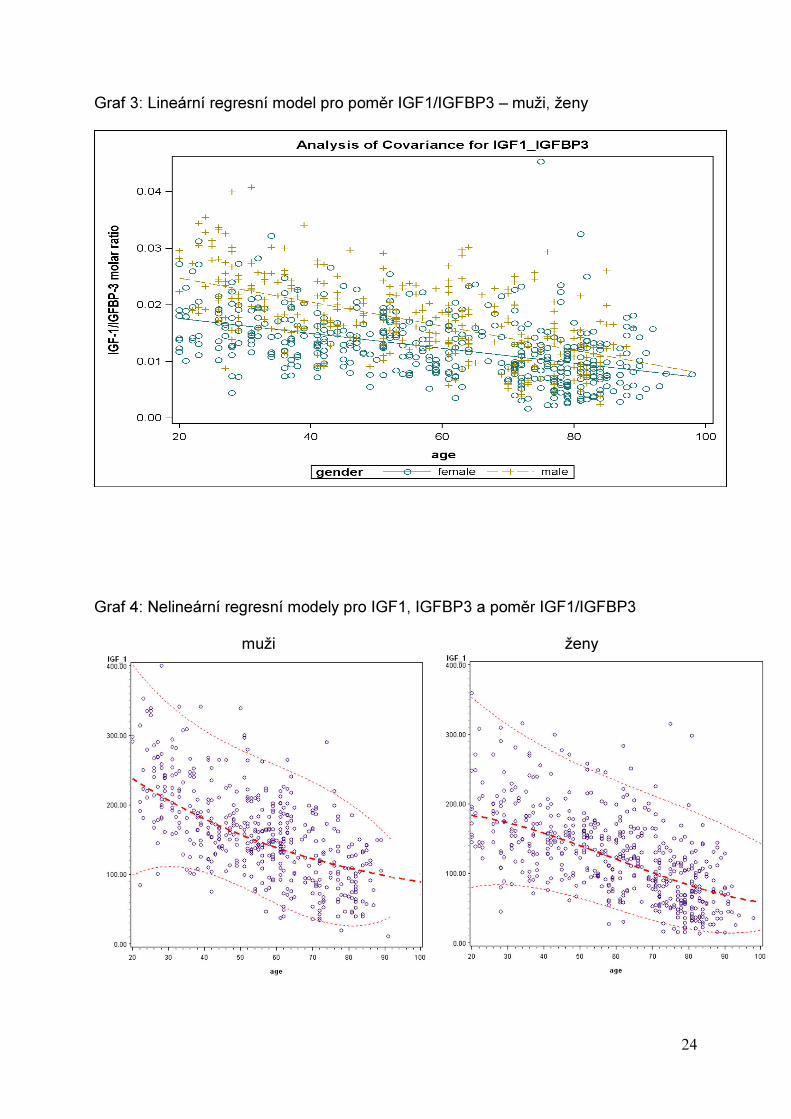
Graf 3: Lineární regresní model pro poměr IGF1/IGFBP3 – muži, ženy
Graf 4: Nelineární regresní modely pro IGF1, IGFBP3 a poměr IGF1/IGFBP3
muži ženy

25
Graf 5: Nelineární regresní modely pro IGFBP3
muži ženy
Graf 6: Nelineární regresní modely pro poměr IGF1/IGFBP3
muži ženy

26
Biomarker měnící se s věkem představuje vždy tak trochu výzvu při
zpracování dat z provedených studií a při tvorbě referenčních hodnot. Také při
porovnávání dvou souborů pacientů s naměřenými hodnotami IGF1 je vždy nutné
provádět pomocí statistických metod pečlivé srovnání věkového rozložení
porovnávaných skupin. Když se věkové složení statisticky významně liší, je nutné
vždy provést věkovou adjustaci naměřených hodnot. Pokud se takový přístup
neuplatňuje, je možné se u takových biomarkerů, jakým je IGF1, snadno dobrat
nesprávných závěrů na správně naměřených výsledcích (31).
3 IGF1 a mechanizmus nádorové transformace (R. Kučera)
Jak již bylo uvedeno v předchozích kapitolách, IGF1 a IGF2 spolu se svými
vazebnými proteiny a specifickými receptory se podílejí na fungování signální dráhy
IGF1.
Z mechanizmu účinku IGF1 na buňky se usuzuje, že vysoké sérové hladiny
IGF1 zvyšují riziko vzniku nádorů a svým dalším působením zvyšují proliferaci a
riziko maligního zvratu. Naproti tomu vysoké hladiny IGFBP3 by měly riziko snižovat
a růst buněk omezovat. Přes vysoký počet provedených studií není ale možné přímo
zkorelovat sérové hladiny IGF1 či IGFBP3 s rozvojem nádoru.
Nezávisle na hladinách IGF1 a IGFBP3 je možné říci, že nádorová
transformace buněk je možná pouze v případě přítomnosti funkčních IGF1R.
Klíčovou úlohu IGF1R v onkogenní transformaci dokládají výsledky pokusů, které
ukazují, že fibroblasty získané z myších embryí zbavené IGF1R, nemohou být
změněny působením některých z řady známých onkogenů. Znovuzavedení
funkčního IGF1R činí buňky opět vnímavé k transformační činnosti onkogenů (32).
Nadměrná exogenní exprese IGF1R ve fibroblastech vede ke vzniku vysoce
transformovaného fenotypu, který způsobuje tvorbu nádorů u nahých myší. Na
druhou stranu, zablokování IGF1R signální dráhy pomocí IGF1R protilátek vede ke
značnému snížení progrese melanomu, karcinomu prsu, karcinomu tlustého střeva,
neuroblastomu a jiných nádorových buněk.
Zajímavé je, že schopnost onkogenů způsobit transformaci závisí na jejich
schopnosti fosforylace IGF1R. Neporušená tyrosin kinázová doména je zásadní

27
podmínkou toho, aby IGF1R vykonával své silné antiapoptotické a transformační
činnosti (33).
Základní podmínky přenosu signálu do buňky tedy jsou:
- přítomnost funkčních receptorů
- neporušená tyrosin kinázová (TK) doména
- fosforylace TK domény
Též jako velmi významná se ukázala autokrinní či parakrinní role IGF1 a IGF2.
Zvýšení místní produkce IGF1 nebo IGF2 nebo jejich kombinace byly zjištěny u
různých nádorových onemocnění a většinou pozitivně korelovaly s progresí nádorů
(34). Co se týče exprese IGF1R je jejich prognostický význam pro riziko nádorových
onemocnění prozatím nejasný v důsledku rozporných výsledků některých studií. Tyto
rozpory mohou být způsobeny rozdílnými metodikami včetně techniky konzervace
tkání, velikostí vzorků a způsobem analýzy dat.
Jiná situace nastává, hodnotíme-li vztah IGF1R k nádorovým metastázám.
IGF1R signalizace hraje důležitou roli v klíčových fázích metastatického procesu jako
je buněčná adheze, migrace, angiogeneze a podpora růstu vzdálených metastáz.
Další in vivo studie ukázaly, že funkční poruchy IGF1R přispívají jednak k potlačení
růstu primárního nádoru a především výrazně napomáhají poklesu nádorových
metastáz. Buňky karcinomu tlustého střeva bez přítomného funkčního IGF1R na
jejich cytoplasmatických membránách nevytvořily jaterní metastázy po přímém
vstřikování do jater, což naznačuje, že IGF1R může regulovat vznik nádorových
metastáz nezávisle na růstu nádoru (35).
Rizikové faktory nádorové transformace v souvislosti s produkcí a
mechanizmem účinku IGF1 tedy můžeme shrnout na:
Extracelulární rizikové faktory
- lokální extrahepatální produkce IGF1
- autokrinní a parakrinní funkce IGF1
- zvýšená sérová hladina IGF1
- intracelulární rizikové faktory
- mutace v signální dráze
- genové mutace

28
4 Systém IGF1 a vybrané nádorové diagnózy
(R. Kučera, O. Topolčan, J. Kinkorová)
Po dohledání a přečtení velkého množství studií jsme zjistili, že téměř u všech
nádorových onemocnění byly sledovány jednotlivé složky signální dráhy IGF1 a to
jak v séru, tak na buněčné úrovni. Vybranými nádory jsme se pak zabývali detailněji.
4.1 Nádory kolorekta
Celá řada studií se věnuje vztahu cirkulujícího IGF1 a kolorektálního
adenomu. Poměrně rozsáhlá multicentrická studie měřila sérové hladiny IGF1 a jeho
vazebných proteinů a současně hladiny C-peptidu jako markeru sekrece inzulínu u
554 nemocných pacientů s adenomem a 786 kontrol s normální endoskopií. Nízké
hladiny IGFBP1 a vysoké sérové hladiny C-peptidu byly statisticky významně spojeny
s rizikem kolorektálního adenomu. IGF1 nehrál v riziku dle této studie žádnou
významnou roli. Tato studie naznačuje, že hyperinzulinémie působí jako etiologický
faktor v kolorektální karcinogenezi (36). Tento fakt není zas tak překvapující
v souvislosti se zjištěním, že IGF1 se dokáže vázat na receptor pro inzulín (IR) a
inzulín se váže na IGF1R. Heterologní ligand má samozřejmě k danému receptoru
nižší afinitu. Dále se IGF1 a inzulín mohou vázat na hybridní receptory tvořené
z poloviny IR a z poloviny IGF1R podjednotkou. Inzulín se tedy stává dalším
významným prvkem vstupujícím do IGF1 signální dráhy (37).
Zajímavé výsledky přineslo sledování hladin IGF1 v závislosti na opakovaném
výskytu kolorektálního adenomu. Sérové hladiny IGF1 byly měřeny při prvním
záchytu adenomu u 299 mužů. Hladiny IGF1 byly významně pozitivně asociovány s
přítomností adenomů. V kontrastu s tímto zjištěním byly koncentrace IGF1 nepřímo
spojené s opakovaným nálezem kolorektálního adenomu. Autoři vyslovují teorii, že
jakmile je odstraněn adenom, vyšší koncentrace IGF1 snižují pravděpodobnost
vzniku nových lézí v konečníku (38). Jiní autoři prokázali u primárního adenomu
zvýšení rizika u vysokého IGF1 a snížení rizika u vysokého IGFBP3. U opakovaného
výskytu adenomu však zjistili autoři nejdelší intervaly relapsů u vysokých hladin
IGFBP3 i IGF1 (39). Správné fungování osy IGF1 bude tedy zřejmě nutné pro
zachování odolnosti buněk tak, jak to bylo prokázáno v případě keratinocytů, což je
podrobněji zmíněno v oddílu pojednávajícím o melanomech.

29
Autoři studie zabývající se hodnocením vztahu rizika vzniku adenomů a
sérových hladin IGFBP3 a rizika vzniku adenomů a tkáňové expese IGFBP3 mRNA
nezjistili žádný významný vztah mezi sérovými hladinami IGFBP3 a rizikem
adenomu, ale nalezli negativní korelaci mezi nižší tkáňovou expresí IGFBP3 mRNA a
rizikem adenomů (40). Stejný tým zveřejnil nedávno práci o zvýšeném riziku
kolorektálního adenomu u pacientů s hyperinzulinémií při snížení IGF1 mRNA a
změnách poměru izoforem inzulinového receptoru IR-A/IR-B ve tkáních (41).
Studie zabývající se expresí IGF1R jako základního prvku pro fungování
signální dráhy IGF1 přinesly zajímavá zjištění. Starší studie uvádí zvýšení exprese
IGF1R během progrese z kolorektálního adenom přes adenokarcinom a metastázy.
IGF1R exprese korelovala se stadiem onemocnění. Novější studie na toto téma
uvádí, že IGF1R je vyjádřen ve vysoké míře v časných stadiích nádorové aberace. U
pokročilých nádorů s nízkou diferenciací byly nalezeny nízké exprese IGF1R (42).
U pacientů s kolorektálním karcinomem bylo zjištěno, že vyšší sérové hladiny
IGFBP3 se nacházejí u pacientů s delším přežitím (43). Jiní autoři poukazují na
spojitost vyšší plazmatické hladiny IGFBP1 a vyšší míry přežití, zatímco hladiny IGF1
a IGFBP3 neměly žádný vztah k délce přežití pacientů (44).
Samostatnou kapitolou jsou pacienti s akromegalií. Zvýšená produkce
růstového hormonu u těchto pacientů způsobuje též zvýšení plazmatických hladin
IGF1. U pacientů s akromegalií se vyskytují ve zvýšené míře adenomatózní polypy
tlustého střeva. Pacienti s akromegalií mají dvojnásobně zvýšené riziko vzniku
karcinomu tlustého střeva (45).
U pacientů léčených růstovým hormonem se vzhledem na mechanizmus
působení IGF1 uvažovalo o možném zvýšeném riziku nádorů střevní sliznice (46).
Zvýšení rizika pro vznik nádorů tlustého střeva u pacientů léčených růstovým
hormonem se však neprokázalo.
4.2 Nádory prsu
Nádory prsu patří k těm nádorům, u kterých je fungování IGF1 systému
nejvíce prostudováno. Pokud porovnáme celou signální soustavu IGF1 u pacientek
s karcinomem prsu a u zdravých žen, tak vidíme, že mnoho součástí osy IGF1
vykazuje změny a to jak v krevním oběhu pacientek s nádory, tak i v nádorové tkáni
samotné.

30
V posledních letech bylo provedeno množství studií s velkými počty nemocných i
zdravých žen, nebo se někteří autoři zaměřili na souhrnné hodnocení výsledků z více
studií.
Někteří autoři nacházejí pozitivní korelaci mezi zvýšenými hladinami IGF1 a
IGFBP3 a rizikem karcinomu prsu u žen před menopauzou (47).
Studie provedená v Austrálii, která zahrnovala 423 případů rakoviny prsu a
1901 kontrol, ukázala, že zvýšená hladina IGF1 a IGFBP3 byla pozitivně spojena s
rizikem rakoviny prsu u pacientek starších než 50 let, ale ne u mladších žen (48).
Evropská studie The European Prospective Investigation into Cancer and Nutrition
(EPIC), která analyzovala data od 1081 pacientek s rakovinou prsu a 2098 zdravých
žen, došla k závěru, že vysoké hladiny IGF1 nebo IGFBP3 jsou spojeny se 40%
zvýšením rizika rakoviny prsu u žen starších než 50 let, ale ne u mladších žen (49).
Studie provedená na populaci žen v různých oblastech Japonska ukázala
nevýznamnou roli IGF1 v riziku karcinomu prsu. Vyšší hladiny IGFBP3 snižovaly
riziko u žen před menopauzou, u žen po menopauze se žádný vztah k diagnóze
prokázat nepodařilo (50).
Čínští autoři provedli hodnocení dat z téměř stovky studií. Na základě tohoto
zhodnocení došli k závěru, že zvýšené sérové hladiny IGF1 pozitivně korelují se
zvýšeným rizikem karcinomu prsu (51). Americká studie populace žen do 45let
zkoumala závislost hladin IGF1, IGFBP1, IGFBP3, GH a rizika rakoviny prsu, ale
žádné statisticky významné závislosti nalezeny nebyly (52).
Kromě epidemiologických dat se experimentální důkazy též dotýkají role IGF
systému v etiologii rakoviny prsu. Aktivace IGF1R chrání buňky rakoviny prsu před
apoptózou. Gen IGF1R je vysoce vyjádřený v buňkách karcinomu prsu. Přesný
biologický význam však není úplně jasný. Nižší výskyt IGFIR nacházíme v benigních
lézích a normální prsní tkáni v porovnání s maligní tkání. Nicméně ve vysokých
koncentracích byl nalezen IGF1R v tkáních dobře a středně diferencovaných
karcinomů prsu, ale v mnohem nižších koncentracích u špatně diferencovaných
karcinomů. Někteří autoři se domnívají, že vysoké hladiny IGF1R zhoršují prognózu,
zatímco jiní považují vysokou úroveň IGF1R za příznivý prognostický ukazatel.
Hladiny IGF1R byly signifikantně vyšší u nádorů s genetickou mutací supresorového
genu BRCA1 (Breast Cancer 1) ve srovnání s nádory bez této mutace (53, 54).
Hladiny některých komponent signální kaskády IGF, včetně IGF1R, byly častěji

31
zvýšené v normální a nádorové tkáni u žen se silnou rodinnou anamnézou rakoviny
prsu než u žen bez této anamnézy.
Zajímavým zjištěním je, že v buňkách získaných z biopsií nádorů prsu je
možné vysledovat expresi určitých genů jako výsledek působení signální kaskády
IGF1. Exprese IGF1 indukovaných genů významně koreluje s agresivitou nádoru.
Nádorové buňky bylo možno rozdělit do dvou skupin. Ve skupině s vyšší expresí IGF
indukovaných genů bylo přežití významně kratší než ve skupině s expresí nižší (55).
Tato zjištění podporují teorii, kterou vyslovují někteří autoři i v souvislosti s nádory
kolorekta.
ER (estrogen receptor) pozitivní klony buněk měly zvýšenou úroveň IGF1R,
klony ER negativní měly úroveň sníženou. Vysoké hladiny IGF1R vedly k recidivě
karcinomu prsu po radioterapii a mohou též přispívat k rozvoji rezistence na léčbu
Herceptinem (trastuzumab - monoklonální protilátka proti HER2 receptoru) (56).
Zablokování IGF1R vede ke snížení proliferace nádorových buněk karcinomu
prsu. Sledování výchozích hladin IGF1 a IGFBP3 a jejich změn v závislosti na
různých léčebných strategiích karcinomu prsu však neprokázalo jednoznačný vztah
mezi délkou přežití a výchozími hodnotami či jejich změnami v průběhu terapie (57).
4.3 Karcinom prostaty
Množství údajů naznačuje, že signální systém IGF1 hraje důležitou roli při
vzniku a progresi karcinomu prostaty. Vyšší hladiny IGF1 v oběhu zvyšují riziko
karcinomu prostaty, zatímco vyšší koncentrace IGFBP3 riziko snižují (58). Další
studie ukázala, že cirkulující hladiny IGF1 a IGFBP3 byly signifikantně vyšší u
pacientů s karcinomem prostaty oproti zdravým mužům (59). Zajímavým zjištěním je,
že poměr IGF1/PSA měl lepší výpovědní hodnotu ohledně predikce rizika karcinomu
prostaty než samotné měření hladin PSA nebo IGF1 (60). Rozsáhlá britská studie
uvádí, že u pacientů s karcinomem prostaty byly spolu se zvýšeným PSA nalezeny
snížené hladiny IGF2 a IGFBP3. U IGF1 nebyly zjištěny žádné změny hladin při
porovnání se zdravými muži (61).
Studie zaměřené na mechanizmus vzniku karcinomu prostaty poukazují na
významnou roli autokrinní produkce IGF1. Karcinom prostaty vyvolaný vysokými
dávkami testosteronu u potkanů byl podporován zvýšenou autokrinní produkcí IGF1.
Exprese transgenů IGF1 u myší vedla k aktivaci IGF1R a spontánní tvorbě tumorů

32
prostaty (62). Studie 54 bioptických vzorků prostaty nalezla výraznou upregulaci
IGF1R na úrovni proteinů i mRNA v primárním karcinomu prostaty v porovnání
s benigním tumorem prostaty. Též byla nalezena zvýšená exprese inzulin
receptorového substrátu 1(IRS1) v maligních biopsiích (63). Ve 12 spárovaných
biopsiích primárního karcinomu prostaty a následné kostní metastázy byla zjišťována
exprese IGF1R, IRS1 a PTEN. Ve 4 případech byla úroveň exprese IGF1R a IRS1
nižší u metastáz než v primárním nádoru. Ve 3 těchto metastázách nebyla prokázána
přítomnost PTEN. Tento nález byl poměrně neobvyklý, protože v 8 z 12 případů byla
zjištěna nadměrná exprese IGF1R a IRS1 v primární i metastatické biopsii (64).
Studie zaměřená na hledání IGF1R ve zmrazených tkáňových řezech prostaty
nalezla zvýšený výskyt IGF1R v normální prostatické tkáni, ve tkáni karcinomu
prostaty a metastáz, ale nenalezla téměř žádné IGF1R ve tkáni benigních tumorů
prostaty (65).
4.4 Nádory plic
Ve studii provedené v Luisianě v roce 1999 byla sledována sérová hladina
IGF1, IGF2 a IGFBP3 u 204 pacientů s nově diagnostikovanou rakovinou plic a u
218 kontrolních osob. Vysoké hladiny IGF1 byly spojeny se zvýšeným rizikem
rakoviny. Izolované hodnocení hladin IGFBP3 neukázalo žádnou spojitost s rizikem
rakoviny plic. Pokud bylo IGFBP3 hodnoceno společně s IGF1, byly vysoké hladiny
IGFBP3 spojeny s nižším rizikem rakoviny plic. Hladiny IGF2 neměly žádnou
souvislost s rizikem rakoviny plic. Novější studie zabývající se sérovými hladinami
IGF1 a IGFBP3 ve vztahu k riziku nádorů plic zjistila, že zvýšené hladiny IGF1 riziko
významně nemění, ale vyšší hladiny IGFBP3 riziko snižují (66).
U rizika plicních nádorů hraje významnou roli autokrinní a parakrinní působení
IGF1. Při zkoumání vzorků bronchiální tkáně bylo zjištěno, že hladiny IGF1 a IGF2
byly výrazně vyšší ve vzorcích tkáně s vysokým obsahem dysplazií než v těch, které
obsahovaly pouze běžný epitel (67). Tento nález ještě umocňuje zvýšená exprese
IGF1R na buňkách dysplazií. Transformaci buněk výrazně potlačuje inaktivace
IGF1R a tím inhibice účinku IGF1.
Zajímavé výsledky poskytla studie, kdy se autoři zaměřili na zkoumání role
IGF1R při vzniku a rozvoji plicního nádoru. Na zvířecím modelu transgenních myší se
autorům podařilo zvýšenou expresí IGF1R vyvolat změny v plicní tkáni, které vedly

33
k rozvoji nádorů. Při blokaci IGF1R nedošlo k pozitivní odpovědi u všech nádorů, ale
menší část pokračovala nadále v růstu. Tyto nálezy naznačují, že některé nádory,
které jsou iniciovány nadměrnou expresí IGF1R, mohou dalšími mutacemi získat
nezávislost na IGF1R signalizaci (68).
K podobným závěrům dospěla studie prognostického významu exprese
IGF1R u pacientů s nemalobuněčným karcinomem plic (69).
4.5 Melanomy
Sérové hladiny IGF1 a IGFBP3 u maligních melanomů byly zkoumány
v poměrně malém počtu studií. U pacientů s melanomy byly nalezeny jak snížené
hladiny IGF1, tak i zvýšené hladiny. Hladiny IGFBP3 jsou u melanomů popisovány
převážně zvýšené, nicméně jako efektivnější se jeví zkoumání exprese jednotlivých
součástí IGF1 systému přímo ve tkáních (70).
Podle studií prováděných na buněčných kulturách buňky melanomů reagují
v časném nádorovém stadiu na stimulaci pomocí IGF1 zvýšeným přežitím, migrací a
růstem. V pozdně primárních nebo metastatických lézích již není reakce na stimulaci
IGF1 příliš výrazná. Ve studii zkoumající 36 biopsií melanomů byla vysoká exprese
IGF1R spojena se zvýšeným počtem metastáz a kratším přežitím pacientů (71).
Hladina sérového IGF1 též pozitivně koreluje s metastatickým postižením
lymfatických uzlin (72). Tato korelace však není tak silná, jako když zkoumáme
expresi IGF1R přímo ve tkáních.
Pomocí zkoumání signální dráhy IGF1 v klonech melanomových buněk byly
vysvětleny antiapoptotické účinky IGF1 a vliv na ochranu mitochondrií před jejich
funkčním poškozením, což zaručilo melanomovým buňkám odolnost a úspěšné
přežívání (73).
Signální dráha IGF1má však zásadní význam pro zachování správné
odolnosti keratinocytů na ultrafialové záření. Tato odolnost je důležitá pro potlačení
onkogenních transformací v kůži. U stárnoucí kůže nacházíme sníženou expresi
IGF1R, což koreluje s faktem, že 80% rakoviny kůže se vyskytuje u pacientů starších
60ti let. Pokud je IGF1R neaktivní v době UV ozáření, keratinocyty mají menší šanci
na přežití a velké procento z keratinocytů projde apoptózou. Nicméně některé
přežívající keratinocyty s poškozenou DNA se můžou i nadále množit s možností
karcinogenních mutací (74).

34
4.6 Ovariální karcinom
Část autorů uvádí, že zvýšené hladiny IGF1 zvyšují riziko ovariálního
karcinomu, zatímco vyšší hladiny IGFBP3 toto riziko mírně snižují. Určitým předělem
se zdá být věk cca 55 let. Hodnocení hladin IGF1 u žen starších než 55 let nepřineslo
žádné změny rizika v závislosti na hladinách IGF1 a IGFBP3. Na druhou stranu v
několika retrospektivních studiích byly koncentrace IGF1 v séru u žen s maligním
nádorem vaječníků nižší než u kontrolní skupiny (75, 76).
Při experimentech zaměřených na chování signální dráhy IGF1 in vitro se
poměrně často používají klony ovariálních nádorů. Na buněčných klonech
nádorových linií karcinomu vaječníků byla prokázána autokrinní produkce IGF1 a
IGF2 a tvorba IGF1R, čímž se prokázal vznik tzv. autokrinní růstové smyčky. RNA
analýzy odhalily přítomnost IGF1 a IGF1R mRNA ve 100% čerstvě izolovaných
nádorových vzorků. Studie mapující úlohu IGF2 zjistila výrazně vyšší IGF2 mRNA ve
109 vzorcích nádorové tkáně vaječníků ve srovnání s osmi vzorky normální tkáně.
Kromě toho byla vysoká IGF2 genové exprese spojena s pokročilým stadiem
onemocnění a kratším přežitím. Zablokování IGF1R vede ke snížení proliferace
buněk karcinomu ovarií (77).
4.7 Karcinom děložního čípku
U karcinomu děložního čípku byly nalezeny zvýšené hladiny IGF2 a normální
hladiny IGF1. Zvýšené hladiny IGF2 může blokovat IGFBP5. Autoři též popisují
zvýšenou expresi mRNA po stimulaci linií nádorových buněk epidermálním růstovým
faktorem (EGF). Na základě těchto výsledků autoři vyslovili teorii zkříženého
působení systémů IGF a EGF. Autokrinní produkce IGF2 může zprostředkovat
mitogenní účinek EGF v buňkách rakoviny děložního čípku (78).
Významnou roli autokrinní produkce IGF2 v patogenezi karcinomu děložního
čípku potvrzuje studie bioptických vzorků nádorové a normální tkáně. Ve tkáni
karcinomu děložního čípku byly nalezeny významně zvýšené hladiny oproti normální
tkáni. Autoři také nalezli významný vztah mezi IGF2 hladinami u nádorů a metastáz v
pánevních lymfatických uzlinách. Koncentrace IGFBP3 vykazovaly výrazný pokles v
pokročilém stádiu onemocnění (79).

35
4.8 Karcinom žaludku
U pacientů s karcinomem žaludku byly hodnoceny sérové hladiny v čase
zjištění diagnózy. Hladiny IGF1 byly snížené, hladiny IGF2 lehce zvýšené a hladiny
IGFBP3 nevykazovaly žádný rozdíl ve srovnání s kontrolní skupinou (80). V novější
studii 210 případů rakoviny žaludku nepřineslo hodnocení sérových hladin IGF1,
IGF2, a IGFBP3 zjištění žádných významných závislostí co se týče rizika karcinomu
žaludku (81).
Při hodnocení role signální dráhy IGF1 v souvislosti s karcinomem žaludku
bylo zjištěno, že IGF1 signalizace podporuje růst a přežití buněk karcinomu žaludku.
Xenotransplantát lidských buněk karcinomu žaludku do peritonea myší byl sledován
po aplikaci rekombinantního adenoviru, který způsobí expresi okleštěného IGF1R,
který nepřenáší signál do buňky. Zablokováním signální dráhy IGF1 došlo u myší
k výraznému zvýšení apoptózy indukované radioterapií a chemoterapií. Zvýšená
exprese IGF1R ve tkáních karcinomu žaludku je negativním prognostickým faktorem
(82).
4.9 Nádory jater
Téměř všechny hepatocelulární karcinomy se objevují v souvislosti s
chronickým poškozením jater, zánětem a zvýšeným dělením hepatocytů. Následující
regenerační procesy a fibróza vedou k cirhóze jater, při níž se zvyšuje frekvence
mutací v hepatocytech a případně vzniká maligní bujení. Rozvoj hepatocelulárního
karcinomu (HCC) na základě hyperaktivace signální dráhy IGF1 byl jasně prokázán
studiemi na zvířecím modelu (83).
Jaterní buňky produkují IGF1, někteří autoři spojují schopnost jaterních buněk
produkovat IGF1 a tedy měření jeho sérových hladin s mírou jaterního poškození
nádorovým procesem. Studie zaměřená na produkci IGF1 jako parametru poškození
jaterních buněk a prognostického ukazatele u pacientů s HCC prokázala snižování
hladiny IGF1 spolu se závažností onemocnění (84). Studie, která porovnávala
hladiny sérového IGF1 u pacientů s HCC a u pacientů s hepatitidou typu B a C,
odhalila výrazně nižší sérové koncentrace IGF1 u pacientů s HCC (85). Další studie
porovnávala hladiny IGF1 u 65ti pacientů s HCC a skupinou zdravých osob. Výrazně
nižší sérové hladiny IGF1 byly zjištěny u pacientů trpících HCC (86).

36
Ve studii bioptických vzorků pacientů s HCC byla snížená exprese IGFBP1,
IGFBP3 a IGFBP4 (87). V další studii 11 pacientů s hepatoblastomem byla zjištěna
snížená exprese IGFBP1 v nádorové tkáni oproti normální jaterní tkáni. V této studii
bylo dále zjištěno snížení IGFBP2 v nádorové tkáni. Autoři formulovali teorii, že z
důvodu snížené exprese vazebných proteinů je zvýšená nabídka biologicky aktivních
IGF1 a IGF2 a tyto pak potencují proliferační účinky (88).
4.10 Karcinom pankreatu
Ve čtyřech amerických prospektivních studiích bylo zhodnoceno 144 případů
rakoviny slinivky břišní. Byly hodnoceny sérové hladiny IGF1, IGFBP1, IGFBP3 a C-
peptidu jako ukazatele produkce inzulínu. Nízké sérové hladiny IGFBP1 byly
významně spojeny se zvýšeným rizikem rakoviny slinivky (89). Ve finské studii
hodnotili autoři vztah IGF1 a IGFBP3 k rakovině pankreatu u mužů kuřáků (90). Bylo
hodnoceno 93 případů adenokarcinomů u finských mužů - kuřáků ve věku 50-69 let.
Sérové hladiny IGF1 a IGFBP3 byly srovnávány s hladinami u 400 náhodně
vybraných kontrol. Sérové koncentrace IGF1 a IGFBP3 ani poměr IGF1/IGFBP3
nebyl významně spojen s adenokarcinomem pankreatu.
Při studiu IGF1R a jeho signální dráhy ve vztahu k rakovině slinivky byly do
tkáňových kultur buněk karcinomu pankreatu přidány specifické inhibitory IGF1R a
PI3K. Došlo tím k zablokování přenosu signálu v kaskádě PI3K/PKB. Oba inhibitory
způsobily apoptózu v rakovinných buňkách (91).
4.11 Nádory štítné žlázy
U adenomů štítné žlázy jsou popisovány nižší sérové hladiny IGF1 ve
srovnání se zdravou populací (92).
Zvýšená exprese IGF1R koreluje u nádorů štítné žlázy se špatnou prognózou.
U IGF1R nacházíme zjevnou souvislost mezi jejich expresí a diferenciací nádorových
buněk. U dobře a středně diferencovaných karcinomů nacházíme významnou expresi
těchto receptorů, zatímco u anaplastických karcinomů již exprese chybí. Tento jev
autoři vysvětlují tím, že u anaplastických buněk je aktivováno dostatečné množství
signálních prorůstových drah a dráha zprostředkovaná IGF1R již není zapotřebí (93).

37
5 Systém IGF1 a kancerogeneze - vliv dalších faktorů
(R. Kučera, Š. Svobodová)
Jak již bylo popsáno v úvodní kapitole, GH zvyšuje výdej glykogenu z jater a
ve svalech inhibuje vychytávání glukózy a její další využití, což je základem jeho
diabetogenního účinku. GH sice přímo nestimuluje sekreci inzulínu, ale
hyperglykémie, kterou GH samotný a zejména pak prostřednictvím IGF1 vyvolává,
sekundárně stimuluje pankreas a může nakonec vést až k vyčerpání β-buněk a
rozvoji diabetu.
Inzulín je produktem β-buněk Langerhansových ostrůvků pankreatu. Je to
dimer tvořený řetězci A a B, které jsou spojeny dvěma disulfidickými můstky. Inzulín
je generován proteolytickým štěpením z jeho prekurzoru - proinzulínu. Vzniká tak
inzulín a C-peptid, který je využívám diagnosticky, protože jeho biologický poločas
(30 minut) je delší než biologický poločas inzulínu (5-10 minut). V reakci na stimulaci
β-buňky uvolňují inzulín a C-peptid v ekvimolárním množstvím do krve s malými
množstvími proinzulínu. Inzulín je sekretován primárně v závislosti na zvýšené
koncentraci glukózy v krvi. Hlavní funkcí inzulínu je regulovat energetický
metabolizmus stimulují buněčnou proliferaci a anabolické pochody jako funkce
dostupné energie (94).
Vztah růstové signální dráhy IGF1 a zvýšené hladiny inzulínu je v současné
době předmětem zájmu řady studií. Hyperinzulinizmus a následně diabetes, dále
obezita, hypertenze a kardiovaskulární onemocnění jsou atributy metabolického
syndromu, který je v současnosti stále rostoucím zdravotním problémem
v celosvětovém měřítku (95) V poslední době je metabolický syndrom dáván do
souvislosti se zvýšeným rizikem některých druhů nádorových onemocnění. Hledání
takové souvislosti má racionální základ. V současnosti víme, že inzulín a IGF1
dokáží navzájem využívat své receptory či dokonce existují hybridní receptory
IR/IGF1R, jak uvádí Tabulka 4. Je tedy předpoklad, že zvýšené hladiny inzulínu
budou ovlivňovat růstové dráhy v buňce podobným mechanizmem jako IGF1. Různé
signální dráhy se v buňce vzájemně ovlivňují, proto se v současné době zaměřuje
pozornost na „styčné body“ (enzymy a substráty) těchto signálních drah (96) V této
kapitole bychom chtěli stručně zmínit pohled na riziko kancerogeneze z pohledu

38
produkce a využití inzulínu a dalších dvou působků – leptinu a adiponektinu, které
s níže diskutovanou problematikou souvisí.
Leptin je protein tvořený v adipocytech v tukové tkáni. Patří mezi tzv.
adipokiny - takto se souhrnně nazývají látky, které jsou tvořeny tukovými buňkami.
Leptin je prvním objeveným adipokinem. Jeho hlavní funkcí je udržování energetické
rovnováhy. Omezuje příjem potravy a zvyšuje energetický výdej. Sérové hladiny
leptinu korelují se zásobami tuku v organizmu (97). Zvýšené hladiny leptinu začaly
být dávány do souvislosti s rozvojem nádorových onemocnění po zjištění, že leptin
stimuluje buněčnou proliferaci a potlačuje apoptózu stimulací stejných signálních
drah jako IGF1. Leptin je též produkován lokálně ve tkáních a působí též svým
autokrinním a parakrinním efektem. Jako další účinek leptinu bylo popsáno
zprostředkování efektu některých cytokinů a up-regulace signální dráhy VEGF
v některých typech nádorů (98).
Adiponektin je další protein ze skupiny adipokinů. Adiponektin tvoří největší
procento působků tvořených v tukové tkáni. Adiponektin chrání před rozvojem
aterosklerotických změn, potlačuje produkci kyslíkových radikálů při zvýšených
hladinách glukózy, zlepšuje inzulínovou rezistenci ve svalech, snižuje hladinu
adhezivních molekul, má protizánětlivý efekt (99). Adiponektin má antikarcinogenní
účinek. Tento účinek je vysvětlován inhibicí proliferace endoteliálních buněk a
indukcí kaspázou zprostředkované apoptózy, což bylo prokázáno studiemi in-vitro
(100). Stručná charakteristika výše uvedených biomarkerů uvádí Tabulka 8.
Tabulka 8: Základní charakteristika izulínu, leptinu a adiponektinu
Biomarker Molekulová
hmotnost
Počet
aminokyselinFyziologická funkce
Inzulín 5 800 51
Ve svalech aktivace transportu glukózy do buněk, v játrech podpora
tvorby glykogenu, v tukové tkáni podpora tvorby tuků
Leptin 16 200 146 Regulace tukových zásob v organizmu, prorůstový,
antiapoptotické efekt
Adiponektin 180 000 244 Regulace metabolizmu glukózy a procesu oxidaci mastných

39
Schéma 4 znázorňuje mechanizmus, jakým ovlivňují inzulín, IGF1, leptin a
adiponektin proliferační, prorůstové, antiapoptotické nebo na druhé straně
proapoptotické systémy v buňce.
Biologický účinek leptinu, IGF1, inzulínu a adiponektinu je zprostředkován
receptory. Leptin, IGF1 a inzulín aktivují dvě hlavní prorůstové signální dráhy.
Signální dráhu PI3K (phosphatidylinositol-3-kinase)/PKB (proteinkinase B) a signální
dráhu ERK (extracellular signal regulated kinases), což je jedna z MAP kinázových
drah (MAPK mitogen activated protein kinases) (101).
IGF1 je hlavní aktivátor těchto signálních drah. Mechanizmus fungování
signální dráhy PI3K/PKB a signální dráhy ERK (extracellular signal regulated
kinases) byl již detailně popsán. V této kapitole můžeme doplnit, že obě dráhy je
možná aktivovat též pomocí inzulínového receptoru a receptoru pro leptin, který je
přítomen v centrálním nervovém systému, ale také v periferních tkáních (102).
Adiponektin zvyšuje využití inzulínu v buňkách podporou účinnosti
inzulínového receptoru. Snížené hladiny adiponektinu vedou k rozvoji inzulínové
rezistence a k chronické hyperinzulinémii. Zvýšené hladiny inzulínu působí společně
s IGF1 zvýšenou stimulaci prorůstových drah, kdy hlavní úlohu hraje receptor pro
IGF1. Adiponektin snižuje též hladinu leptinu a jeho receptoru v neoplastických
buňkách. Adiponektin podporuje apoptózu indukcí enzymu kaspázy. Signální dráhy
PI3K/PKB a ERK naopak kaspázu inhibují a apoptózu tím blokují (103).

40
Schéma 4: Biologický účinek leptinu, IGF1, inzulínu a adiponektinu je zprostředkován receptory. Leptin, IGF1 a inzulín aktivují dvě hlavní prorůstové signální dráhy. Signální dráhu PI3K (phosphatidylinositol-3-kinase)/PKB (proteinkinase B) a signální dráhu ERK (extracellular signal regulated kinases), což je jedna z MAP kinázových drah (MAPK mitogen activated protein kinases). Adiponektin zvyšuje využití inzulínu v buňkách podporou účinnosti inzulínového receptoru.

41
6 Blokátory růstových receptorů (R. Kučera, O. Fiala)
V posledních letech byly prozkoumány a detailně popsány funkce signální
dráhy IGF1. Na základě těchto poznatků bylo možné vyvinout inhibitory růstu
zaměřené proti určitým složkám této signální dráhy, které jsou dalším možným
nástrojem onkologické léčby (104). V současné době zatím ještě není žádný
z doposud vyvinutých inhibitorů v klinickém použití. Více než deset nových léčiv je
však ve stadiu klinických zkoušek. Tato nově vyvinutá léčiva je možné rozdělit
v podstatě do tří skupin:
1 - monoklonální protilátky proti IGF1R
2 - monoklonální protilátky proti ligandům IGF1R (tj. proti IGF1 a IGF2)
3 - inhibitory IGF1 receptorové tyrozinkinázy
Klinická účinnost této skupiny inhibitorů byla prokázána u některých vybraných
typů nádorů, avšak řada klinických studií však nedosáhla příliš povzbudivých
výsledků. Možné příčiny neúspěchu tkví ve složitosti receptorového systému IGF1,
kdy je tento částečně sdílen s inzulínem (Tabulka 4) a dále pak existence paralelních
prorůstových drah, které mohou v případě zablokování IGF1R převzít úlohu
přenašečů prorůstových, proliferativních a antiapoptotických signálů (105). Jako
řešení tohoto problému se samozřejmě nabízí možnost kombinace s již známými
inhibitory růstu, které jsou již v klinickém použití. Tyto možnosti kombinací jsou
v současné době též předmětem klinických zkoušek.
6.1 Monoklonální protilátky proti IGF1R
Zkušenosti z klinických studií ukazují poměrně dobrou snášenlivost
monoklonálních protilátek proti IGF1R v monoterapii. Nejčastějším nežádoucím
účinkem byla až u 20% pacientů hyperglykémie. Ve většině případů však byla
zvládnutelná s nasazením perorálních antidiabetik (106).
Zkoušky s povzbudivým výsledkem proběhly u Ewingova sarkomu,
neuroendokrinních nádorů, hepatocelulárního karcinomu a karcinomu prostaty.

42
6.2 Monoklonální protilátky proti ligandům IGF1R
V této skupině je prozatím zaznamenáno poměrně málo zkušeností. Prozatím
jedna studie prokázala dobrou snášenlivost, hyperglykémie se projevila pouze u
dvou pacientů, avšak nebylo dosaženo pozitivní odezvy na léčbu (107). Jako další
možnost využití inhibitorů IGF1 se jeví kombinovaná terapie při léčbě karcinomu
prsu.
6.3 Inhibitory IGF1 receptorové tyrozinkinázy
U této skupiny inhibitorů růstu se v klinických zkouškách objevilo několik
závažnějších nežádoucích účinků ve formě těžší hyperglykémie, ovlivnění srdečního
rytmu a vzestupu jaterních enzymů ALT a AST. Určitých pozitivních výsledků bylo
dosaženo u adrenokortikálních a kolorektálních karcinomů (108). Další možnosti
kombinované terapie se zkoušejí s inhibitory EGFR.
6.4 Kombinace IGF1R inhibitorů s chemoterapií nebo s dalšími inhibitory růstu
Při prohledání medicínské databáze PubMed je možné zjistit, že klinických
zkoušek inhibitorů signální dráhy IGF1 s dalšími onkologickými léčivy je prováděno
poměrně velké množství.
Jedná se o kombinaci s klasickou chemoterapií, dále pak kombinace
s hormonální anti estrogenovou terapií. Co se týká kombinovaných terapií s ostatními
inhibitory růstu, je možné dohledat kombinace s inhibitory EGFR, mTOR a MAPK
inhibitory.
Co se týká výsledků těchto klinických studií, je možné prozatím shrnout tak, že
určitých úspěchů bylo dosaženo, avšak teprve další studie ukáží, zda naděje
vkládané do tohoto typu onkologické léčby na základě slibných výsledků experimentů
provedených in vitro, budou naplněny i při použití v klinické praxi.

43
6.5 Prediktivní markery terapie, indikace a omezení
Zkušenosti s inhibitory růstové dráhy IGF1 ukazují, že zásadní klinický přínos
léčby je omezen na menší skupinu pacientů, která musí být přesně definována.
Zásadním úkolem pro blízkou budoucnost je tedy vymezení identifikačních a
selekčních markerů, které by umožnily individualizaci terapie a tím zajistily její
funkčnost a maximální efektivitu u pacientů, kteří budou podstupovat léčbu tímto
druhem inhibitorů růstu. V současné době bohužel žádné spolehlivé prediktivní
markery pro tento druh terapie neexistují. Je to dáno zřejmě složitostí celého
systému signální dráhy IGF1 a též jejího propojení s ostatními signálními drahami.
Zapojení jiných ligandů (inzulín) nebo aktivace alternativních růstových cest pomocí
EGFR, VEGFR či receptoru pro leptin může způsobit vytvoření rychlé rezistence a
následné selhání léčby.
Předběžné údaje založené na omezeném počtu pacientů potvrzují, že pouze
přísná individualizace terapie zaručí léčebný úspěch. Pokud dokážeme překonat
omezení spočívající v této individualizaci, můžeme říci, že vývoj nových molekul
inhibitorů signální dráhy IGF1 je pozitivním příslibem do budoucna pro řadu pacientů.

44
Seznam zkratek
Některé zkratky uvádíme vysvětlené pouze v angličtině. Je to proto, že zejména
pro intracelulární elementy signálních drah nejsou vžité české ekvivalenty a dle
našich zkušeností pro pochopení příslušného schématu anglické vysvětlivky zcela
postačují.
ACTH - adrenokortikotropní hormon
Akt - proteinkináza B (PKB)
ALT - alaninaminotransferáza
AST - aspartátaminotransferáza
BRCA1 - supresorový gen Breast Cancer 1
CD221 - receptor IGF1 (IGF1R)
CD222 - receptor IGF2 (IGF2R)
CTGF - connective tissue growth factor, lidský tkáňový pojivový růstový faktor
(IGFBP8)
EPIC - Evropská studie The European Prospective Investigation into Cancer and
Nutrition
GH - growth hormone, somatotropin, růstový hormon (STH)
GHRH - growth hormone releasing hormone, somatoliberin
GHIH - growth hormone inhibiting hormone, somatostatin
GHBP - growth hormone binding protein, vazebný protein růstového hormonu
HCC – hepatocelulární karcinom
HER2 - human epidermal growth factor receptor 2
IGF1 - insulin like growth factor 1, inzulínu podobný růstový faktor 1
IGF2 - insulin like growth factor 1, inzulínu podobný růstový faktor 2
IGFBP1 - insulin like growth factor binding protein 1, IGF1 a IGF2 vazebný protein
IGFBP2 - insulin like growth factor binding protein 2, IGF2 vazebný protein
IGFBP3 - insulin like growth factor binding protein 3, IGF1 a IGF2 vazebný protein
IGFBP4 - insulin like growth factor binding protein 4, IGF1 a IGF2 vazebný protein
IGFBP5 - insulin like growth factor binding protein 5, IGF1 a IGF2 vazebný protein
IGFBP6 - insulin like growth factor binding protein 6, IGF2 vazebný protein

45
IGFBP7 - insulin like growth factor binding protein 6, IGF1 a IGF2 vazebný protein,
též možné najít pod původním názvem mac25
IGFBP8 - insulin like growth factor binding protein 6, IGF2 vazebný protein, též
možné najít pod názvem CTGF
IGF1R - receptor IGF1 (CD221)
IGF2R - receptor IGF2 (CD222)
IRS1 - inzulín receptorový substrát
casp - kaspáza
ER - estrogenový receptor
ERK - extracellular signal regulated kinases
EGF - epidermal growth factor, epidermální růstový faktor
EGFR - receptor EGF
FSH - folitropin
IR - inzulínový receptor
IR-A - inzulínový receptor, izoforma A
IR-B - inzulínový receptor, izoforma B
IR-A/IGF1R - hybridní receptor pro inzulín a IGF1
IR-B/IGF1R - hybridní receptor pro inzulín a IGF1
JAK - Janus kinase
LH - lutropin
M6P - manóza-6-fosfát
mac25 - insulin like growth factor binding protein 6, IGF1 a IGF2 vazebný protein
MEK - serine/threonine-specific protein kinase
MAPK - mitogen-activated protein kinase
mRNA – messenger ribonucleic acid
mTOR - mammalian target of rapamycin
p53 - enzym regulující apoptózu
PIP2 - fosfatidylinositol-3,5-biphosphate
PIP3 - fosfatidylinositol-3,5-triphosphate
PI3K - phosphatidylinositol-3-kinase
PDK1 - pyruvate dehydrogenase kinase isozyme 1
PSA - prostatický specifický antigen
PTEN - phosphatase and tensin homolog
PRL - prolaktin

46
PKB - proteinkináza B (Akt)
Raf - rapidly accelerated fibrosarcoma kinase
Ras - small GTPase
STH - somatotropin, growth hormone, růstový hormon (GH)
T3 - trijodtyronin
T4 - tyroxin
TSH - thyroid stimulating hormone, tyreotropní hormon
VEGF - vascular endothelial growth factor, vaskulární endoteliální růstový faktor
VEGFR - VEGF receptor

47
Literatura
1. Nyberg F: Growth hormone in the brain: characteristics of specific brain
targets for the hormone and their functional significance. Front
Neuroendocrinol. 2000 Oct;21(4):330-48.
2. Maruff P, Falleti M: Cognitive function in growth hormone deficiency and
growth hormone replacement. Horm Res. 2005;64 Suppl 3:100-8.
3. Nyberg F, Hallberg M: Growth hormone and cognitive function. Nat Rev
Endocrinol. 2013 Jun;9(6):357-65.
4. Scarth JP: Modulation of the growth hormone-insulin-like growth factor (GH-
IGF) axis by pharmaceutical, nutraceutical and environmental xenobiotics: an
emerging role for xenobiotic-metabolizing enzymes and the transcription
factors regulating their expression. A review. Xenobiotica. 2006 Feb-Mar;3 (2-
3):119-218.
5. Hwa V, Oh Y, Rosenfeld RG: The insulin-like growth factor-binding protein
(IGFBP) superfamily. Endocr. Rev. 1999 20(6): 761–87.
6. Dupont J, LeRoith D., Insulin and insulin-like growth factor I receptors:
similarities and differences in signal transduction, Horm Res. 55 Suppl 2:22-6.
7. Powell-Braxton L, Hollingshead P, Warburton C, Dowd M, Pitts-Meek S,
Dalton D, Gillett N, Stewart TA., IGF-I is required for normal embryonic growth
in mice, Genes Dev. 1993 7:(12B), 2609-2617.
8. DeChiara TM, Efstratiadis A, Robertson EJ., A growth-deficiency phenotype in
heterozygous mice carrying an insulin-like growth factor II gene disrupted by
targeting, Nature 1990; 345(6270):78-80.
9. Iams WT, Lovly CM. Molecular Pathways: Clinical Applications and Future
Direction of Insulin-like Growth Factor-1 Receptor Pathway Blockade. Clin
Cancer Res. 2015 Oct 1;21(19):4270-7.
10. Ben Abdelaziz R, Ben Chehida A, Azzouz H, Boudabbous H, Lascols O, Ben
Turkia H, Tebib N. A novel homozygous missense mutation in the insulin
receptor gene results in an atypical presentation of Rabson-Mendenhall
syndrome. Eur J Med Genet. 2015 Dec 9;59(1):16-19.
11. Riedemann J., Macaulay VM., IGFIR signaling and its inhibition, Endocrine
Related Cancer 2006, Suppl.1:33-43.

48
12. Hishiya A, Ito M, Aburatani H, Motoyama N, Ikeda K, Watanabe K. Ataxia
telangiectasia mutated (Atm) knockout mice as a model of osteopenia due to
impaired bone formation. Bone. 2005 Oct;37(4):497-503.
13. Renehan AG, Zwahlen M, Minder C, O'Dwyer ST, Shalet SM, Egger M.
Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk:
systematic review and meta-regression analysis. Lancet. 2004;363:1346-
1353.
14. Bruchim I, Sarfstein R, Werner H. The IGF Hormonal Network in Endometrial
Cancer: Functions, Regulation, and Targeting Approaches. Front Endocrinol
(Lausanne). 2014 May 19;5:76.Werner H, LeRoith D. The role of the insulin-
like growth factor system in human cancer. Adv Cancer Res. 1996;68:183-
223.
15. McCampbell AS, Broaddus RR, Loose DS, Davies PJ. Overexpression of the
insulin-like growth factor I receptor and activation of the AKT pathway in
hyperplastic endometrium. Clin Cancer Res, Nov 1;12(21):6373-8, 2006.
16. Monti S., Proietti-Pannunzi L., Sciarra A., Lolli F., Falasca P., Poggi M., Celi
FS., Toscano V., The IGF axis in prostate cancer. 2007, Curr Pharm Des
13:719-27. Samani AA, Yakar S, LeRoith D, Brodt P. The role of the IGF
system in cancer growth and metastasis: overview and recent insights. Endocr
Rev. 2007;28:20-47.
17. Vorwerk P, Wex H, Hohmann B, Mohnike K, Schmidt U, Mittler U. Expression
of components of the IGF signalling system in childhood acute lymphoblastic
leukaemia. Mol Pathol. 2002.
18. Sutherland BW, Knoblaugh SE, Kaplan-Lefko PJ, Wang F, Holzenberger M,
Greenberg NM. Conditional deletion of insulin-like growth factor-I receptor in
prostate epithelium. Cancer Res. 2008 May 1;68(9):3495-504.
19. Mari D. Role of the IGF/insulin system in longevity. Minerva Endocrinol
2011;36:181-5.
20. Bartke A, Brown-Borg H. Life extension in the dwarf mouse. Curr Top Dev
Biol. 2004;63:189-225.
21. Leite DB, Meirelles RM, Mandarim-de-Lacerda CA, Matos HJ, Bernardo-Filho
M. Serum insulin-like growth factor-I adult reference values for an automated
chemiluminescence immunoassay system. Afr. J Biotechnol 2011;10:18027-
33.

49
22. Friedrich N, Alte D, Völzke H, Spilcke-Liss E, Lüdemann J, Lerch MM, et al.
Reference ranges of serum IGF-1 and IGFBP-3 levels in a general adult
population: Results of the Study of Health in Pomerania (SHIP). Growth Horm
IGF Res 2008;18:228-37.
23. Sandhu MS, Dunger DB, Giovannucci EL. Insulin, insulin-like growth factor-I
(IGF-I), IGF binding proteins, their biologic interactions, and colorectal cancer.
J Natl Cancer Inst. 2002 Jul 3;94(13):972-80.
24. Alberti C, Chevenne D, Mercat I, Josserand E, Armoogum-Boizeau P, Tichet
J, Léger J. Serum concentrations of insulin-like growth factor (IGF)-1 and IGF
binding protein-3 (IGFBP-3), IGF-1/IGFBP-3 ratio, and markers of bone
turnover: reference values for French children and adolescents and z-score
comparability with other references. Clin Chem. 2011 Oct;57(10):1424-35.
25. Rosario PW. Normal values of serum IGF-1 in adults: results from a Brazilian
population. Arq Bras Endocrinol Metabol 2010;54(5):477-81.
26. Kucera R, Topolcan O, Pecen L, Kinkorova J, Svobodova S, Windrichova J,
Fuchsova R. Reference values of IGF1, IGFBP3 and IGF1/IGFBP3 ratio in
adult population in the Czech Republic. Clin Chim Acta. 2015 Apr 15;444:271.
27. Friedrich N, Wolthers OD, Arafat AM, Emeny RT, Spranger J, Roswall J,
Kratzsch J, Grabe HJ, Hübener C, Pfeiffer AF, Döring A, Bielohuby M,
Dahlgren J, Frystyk J, Wallaschofski H, Bidlingmaier M. Age- and sex-specific
reference intervals across life span for insulin-like growth factor binding protein
3 (IGFBP-3) and the IGF-I to IGFBP-3 ratio measured by new automated
chemiluminescence assays. J Clin Endocrinol Metab 2014;99:1675-86.
28. Guven B, Can M, Mungan G, Acіkgoz S. Reference values for serum levels of
insulin-like growth factor 1 (IGF-1) and IGF-binding protein 3 (IGFBP-3) in the
West Black Sea region of Turkey. Scand J Clin Lab Invest 2013;73:135-40.
29. Lin CM, Huang YL, Lin ZY. Influence of gender on serum growth hormone,
insulin-like growth factor-I and its binding protein-3 during aging. Yonsei Med J
2009;50:407-13.
30. Wright EM, Royston P. Simplified estimation of age-specific reference interval
for skewed data. Stat Med 1997;16:2785-803.
31. Kucera R, Cerna M, Narsanska A, Svobodova S, Strakova M, Vrzalova J,
Fuchsova R, Treskova I, Kydlicek T, Treska V, Pecen L, Topolcan O, Pazdiora

50
P. Growth factors and breast tumors, comparison of selected growth factors
with traditional tumor markers. Anticancer Res. 2011 Dec;31(12):4653-6.
32. Ketha H, Singh RJ. Clinical assays for quantitation of insulin-like-growth-
factor-1 (IGF1). Methods. 2015 Jun 15;81:93-8.
33. Bach LA. Endothelial cells and the IGF system. J Mol Endocrinol. 2015
Feb;54(1):R1-13.
34. Liu X, Resch J, Rush T, Lobner D. Functional upregulation of system xc- by
fibroblast growth factor-2. Neuropharmacology. 2012 Feb;62(2):901-6.
35. Peterson JE, Kulik G, Jelinek T, Reuter CW, Shannon JA, Weber MJ. Src
phosphorylates the insulin-like growth factor type I receptor on the
autophosphorylation sites. Requirement for transformation by src. J Biol
Chem. 1996 Dec 6;271(49):31562–31571.
36. Samani AA, Yakar S, LeRoith D, Brodt P. The role of the IGF system in cancer
growth and metastasis: overview and recent insights. Endocr Rev.
2007;28:20–47.
37. Schnarr B, Strunz K, Ohsam J, Benner A, Wacker J, Mayer D. Down-
regulation of insulin-like growth factor-I receptor and insulin receptor substrate-
1 expression in advanced human breast cancer. Int J Cancer. 2000;89:506–
513.
38. Le Marchand L, Wang H, Rinaldi S, Kaaks R, Vogt TM, Yokochi L, Decker R.,
Associations of plasma C-peptide and IGFBP-1 levels with risk of colorectal
adenoma in a multiethnic population., Cancer Epidemiol Biomarkers
Prev.Jun;19(6):1471-7.Epub, 2010.
39. Nakae J., Kido Y., Accili D., Distinct and overlapping functions of insulin and
IGF-I receptors, Endocrine Rev 22:818-835, 2001.
40. Jacobs CI.., A review of the role of insulin-like growth factor 2 in malignancy
and its potential as a modifier of radiation sensitivity., Clin Oncol.,
Jun;20(5):345-52., 2008.
41. Flood A, Mai V, Pfeiffer R, Kahle L, Remaley AT, Rosen CJ, Lanza E,
Schatzkin A., The effects of a high-fruit and -vegetable, high-fiber, low-fat
dietary intervention on serum concentrations of insulin, glucose, IGF-I and
IGFBP-3., Eur J Clin Nutr. Feb;62(2):186-196, 2008.
42. Santoro MA, Andres SF, Galanko JA, Sandler RS, Keku TO, Lund PK.
Reduced insulin-like growth factor I receptor and altered insulin receptor

51
isoform mRNAs in normal mucosa predict colorectal adenoma risk. Cancer
Epidemiol Biomarkers Prev. 2014 Oct;23(10):2093-100.
43. Allison AS, McIntyre MA, McArdle C, Habib FK., The insulin-like growth factor
type 1 receptor and colorectal neoplasia: insights into invasion, Hum Pathol.
2007 Nov;38(11):1590-602.
44. Haydon AM, Macinnis RJ, English DR, Morris H, Giles GG. Physical activity,
insulin-like growth factor 1, insulin-like growth factor binding protein 3, and
survival from colorectal cancer. Gut. 2006 May;55(5):689-94.
45. Wolpin BM, et al. Circulating insulin-like growth factor binding protein-1 and
the risk of pancreatic cancer. Cancer Res. 2007;67:7923–8.
46. Renehan AG, Brennan BM., Acromegaly, growth hormone and cancer
risk.,Best Pract Res Clin Endocrinol Metab. 2008:22(4):639-57.
47. Sugumar A, Liu YC, Xia Q, Koh YS, Matsuo K., Insulin-like growth factor
(IGF)-I and IGF-binding protein 3 and the risk of premenopausal breast
cancer: a meta-analysis of literature., Int J Cancer 2004, 111(2):293-7.
48. Baglietto L, English DR, Hopper JL, Morris HA, Tilley WD, Giles GG.,
Circulating insulin-like growth factor-I and binding protein-3 and the risk of
breast cancer., Cancer Epidemiol Biomarkers 2007,16(4):763-8.
49. Rinaldi S, and all., IGF-I, IGFBP-3 and breast cancer risk in women: The
European Prospective Investigation into Cancer and Nutrition (EPIC)., Endocr
Relat Cancer 2006,13(2):593-605.
50. Sakauchi F, Nojima M, Mori M, Wakai K, Suzuki S, Tamakoshi A, Ito Y,
Watanabe Y, Inaba Y, Tajima K, Nakachi K., Serum insulin-like growth factors
I and II, insulin-like growth factor binding protein-3 and risk of breast cancer in
the Japan Collaborative Cohort study. Asian Pac J Cancer 2009,10 Suppl:51-
5.
51. Chen W, Wang S, Tian T, Bai J, Hu Z, Xu Y, Dong J, Chen F, Wang X, Shen
H. Phenotypes and genotypes of insulin-like growth factor 1, IGF-binding
protein-3 and cancer risk: evidence from 96 studies. Eur J Hum Genet. 2009
Dec;17(12):1668-75.
52. Schernhammer ES, Holly JM, Hunter DJ, Pollak MN, Hankinson SE. Insulin-
like growth factor-I, its binding proteins (IGFBP-1 and IGFBP-3), and growth
hormone and breast cancer risk in The Nurses Health Study II. Endocr Relat
Cancer. 2006, 13(2):583-92.

52
53. Maor S, Mayer D, Yarden RI, Lee AV, Sarfstein R, Werner H, Papa MZ.,
Estrogen receptor regulates insulin-like growth factor-I receptor gene
expression in breast tumor cells: involvement of transcription factor Sp1, J
Endocrinol. 2006, 191(3):605-12.
54. Maor S., Yosepoxich A, and all., Elevated insulin-like growth factor-I receptor
(IGF-IR) levels in primary breast cancer tumours associated with BSCA1
mutations., Cancer Lett 2007, 257:236-43.
55. Rajski M, Zanetti-Dällenbach R, Vogel B, Herrmann R, Rochlitz C, Buess M.,
IGF-I induced genes in stromal fibroblasts predict the clinical outcome of
breast and lung cancer patients, BMC Med. 2010, 5;8-11.
56. Werner H, Bruchim I., Basic and clinical significance of IGF-I-induced
signatures in cancer., BMC Med. 2010, 5;18-22.
57. Chapman JA, Pritchard KI, Goss PE, Ingle JN, Muss HB, Dent SF,
Vandenberg TA, Findlay B, Gelmon KA, Wilson CF, Shepherd LE, Pollak MN.
Competing risks of death in younger and older postmenopausal breast cancer
patients. World J Clin Oncol. 2014 Dec 10;5(5):1088-96.
58. Monti S., Proietti-Pannunzi L., Sciarra A., Lolli F., Falasca P., Poggi M., Celi
FS., Toscano V., The IGF axis in prostate cancer., Curr Pharm Des 13:719-27,
2007.
59. Shi R, Berkel HJ, Yu H., Insulin-like growth factor-I and prostate cancer: a
meta-analysis., Br J Cancer.;85(7):991-6., 2001.
60. Djavan B, Bursa B, Seitz C, Soeregi G, Remzi M, Basharkhah A, Wolfram R,
Marberger M., Insulin-like growth factor 1 (IGF-1), IGF-1 density, and IGF-
1/PSA ratio for prostate cancer detection., Urology.;54(4):603-6. 1999.
61. Rowlands MA, Gunnell D, Harris R, Vatten LJ, Holly JM, Martin RM.,
Circulating insulin-like growth factor peptides and prostate cancer risk: a
systematic review and meta-analysis, Int J Cancer.;124(10):2416-29. Review,
2009.
62. Weiss S, Henle P, Bidlingmaier M, Moghaddam A, Kasten P, Zimmermann G.,
Systemic response of the GH/IGF-I axis in timely versus delayed fracture
healing, Growth Horm IGF Res.;18(3):205-12., 2008.
63. Yakar S, LeRoith D, Brodt P. The role of the growth hormone/insulin-like
growth factor axis in tumor growth and progression: lessons from animal
models. Cytokine & Growth Factor Reviews. 2005;16:407–420.

53
64. Nickerson T, Chang F, Lorimer D, Smeekens SP, Sawyers CL, Pollak M. In
vivo progression of LAPC-9 and LNCaP prostate cancer models to androgen
independence is associated with increased expression of insulin-like growth
factor I (IGF-I) and IGF-I receptor (IGF-IR) Cancer Res. 2001;61(16):6276–
6280.
65. Ryan CJ, Haqq CM, Simko J, Nonaka DF, Chan JM, Weinberg V, Small EJ,
Goldfine ID., Expression of insulin-like growth factor-1 receptor in local and
metastatic prostate cancer, Urol Oncol. 2007, 25(2):134-40.
66. Chen B, Liu S, Xu W, Wang X, Zhao W, Wu J., IGF-I and IGFBP-3 and the
risk of lung cancer: a meta-analysis based on nested case-control studies., J
Exp Clin Cancer Res. 2009, 24;28:89.
67. Kim SY, Toretsky JA, Scher D, Helman LJ., The role of IGF-1R in pediatric
malignancies, Oncologist.;14(1):83-91 Review, 2009.
68. Linnerth NM, Siwicky MD, Campbell CI, Watson KL, Petrik JJ, Whitsett JA,
Moorehead RA., Type I insulin-like growth factor receptor induces pulmonary
tumorigenesis., Neoplasia.;11(7):672-82., 2009.
69. Dilli UD, Yildırim M, Suren D, Alikanoglu A, Kaya V, Goktas S, Yildiz M, Sezer
C, Gunduz S. Lack of any prognostic role of insulin-like growth factor-1
receptor in non-small cell lung cancer. Asian Pac J Cancer Prev.
2014;15(14):5753-7.
70. Park SL, Setiawan VW, Kanetsky PA, Zhang ZF, Wilkens LR, Kolonel LN, Le
Marchand L. Serum insulin-like growth factor-I and insulin-like growth factor
binding protein-3 levels with risk of malignant melanoma. Cancer Causes
Control. 2011 Sep;22(9):1267-75.
71. All-Ericsson C, Girnita L, Seregard S, Bartolazzi A, Jager MJ, Larsson O.,
Insulin-like growth factor-1 receptor in uveal melanoma: a predictor for
metastatic disease and a potential therapeutic target., Invest Ophthalmol Vis
Sci. ;43(1):1-8., 2002.
72. Kucera R, Treskova I, Vrzalova J, Svobodova S, Topolcan O, Fuchsova R,
Rousarova M, Treska V, Kydlicek T. Evaluation of IGF1 serum levels in
malignant melanoma and healthy subjects. Anticancer Res. 2014
Sep;34(9):5217-20.
73. Hilmi C, Larribere L, Giuliano S, Bille K, Ortonne JP, Ballotti R, Bertolotto C.
IGF1 promotes resistance to apoptosis in melanoma cells through an

54
increased expression of BCL2, BCL-X(L), and survivin. J Invest Dermatol.
2008 Jun;128(6):1499-505.
74. Lewis DA, Travers JB, Somani AK, Spandau DF., The IGF-1/IGF-1R signaling
axis in the skin: a new role for the dermis in aging-associated skin cancer,
Oncogene.;29(10):1475-85., 2010.
75. Lukanova A, Lundin E, Toniolo P, Micheli A, Akhmedkhanov A, Rinaldi S, Muti
P, Lenner P, Biessy C, Krogh V, Zeleniuch-Jacquotte A, Berrino F, Hallmans
G, Riboli E, Kaaks R., Circulating levels of insulin-like growth factor-I and risk
of ovarian cancer., Int J Cancer. 2002, 20;101(6):549-54.
76. Peeters PH, Lukanova A, Allen N, Berrino F, Key T, and. all., Serum IGF-I, its
major binding protein (IGFBP-3) and epithelial ovarian cancer risk: the
European Prospective Investigation into Cancer and Nutrition (EPIC), Endocr
Relat Cancer. 2007, 14(1):81-90.
77. Gotlieb WH, Bruchim I, Gu J, Shi Y, Camirand A, Blouin MJ, Zhao Y, Pollak
MN.,Insulin-like growth factor receptor I targeting in epithelial ovarian cancer,
Gynecol Oncol. 2006, 100(2):389-96.
78. Steller MA, Delgado CH, Bartels CJ, Woodworth CD, Zou Z., Overexpression
of the insulin-like growth factor-1 receptor and autocrine stimulation in human
cervical cancer cells., Cancer Res.;56(8):1761-5.,1996.
79. Mathur SP, Mathur RS, Gray EA, Lane D, Underwood PG, Kohler M,
Creasman WT., Serum vascular endothelial growth factor C (VEGF-C) as a
specific biomarker for advanced cervical cancer: Relationship to insulin-like
growth factor II (IGF-II), IGF binding protein 3 (IGF-BP3) and VEGF-A,
Gynecol Oncol.;98(3):467-83. 2005.
80. Yi HK, Hwang PH, Yang DH, Kang CW, Lee DY. Expression of the insulin-like
growth factors (IGFs) and the IGF-binding proteins (IGFBPs) in human gastric
cancer cells. Eur J Cancer.;37:2257–2263., 2001.
81. Yatsuya H, Toyoshima H, Tamakoshi K, Tamakoshi A, Kondo T, Hayakawa N,
Sakata K, Kikuchi S, Hoshiyama Y, Fujino Y, Mizoue T, Tokui N, Yoshimura
T., Serum levels of insulin-like growth factor I, II, and binding protein 3,
transforming growth factor beta-1, soluble fas ligand and superoxide
dismutase activity in stomach cancer cases and their controls in the JACC
Study. J Epidemiol;15 Suppl 2:S120-5., 2005.

55
82. Matsubara J, Yamada Y, Nakajima TE, Kato K, Hamaguchi T, Shirao K,
Shimada Y, Shimoda T., Clinical significance of insulin-like growth factor type
1 receptor and epidermal growth factor receptor in patients with advanced
gastric cancer, Oncology.;74(1-2):76-83, 2008.
83. Chung W, Kim M, de la Monte S, Longato L, Carlson R, Slagle BL, Dong X,
Wands JR. Activation of signal transduction pathways during hepatic
oncogenesis. Cancer Lett. 2015 Oct 1. pii: S0304-3835(15)00607-2.
84. Kaseb AO, Abbruzzese JL, Vauthey JN, Aloia TA, Abdalla EK, Hassan MM,
Lin E, Xiao L, El-Deeb AS, Rashid A, Morris JS. I-CLIP: improved stratification
of advanced hepatocellular carcinoma patients by integrating plasma IGF-1
into CLIP score. Oncology. 2011;80(5-6):373-81.
85. Stuver SO, Kuper H, Tzonou A, Lagiou P, Spanos E, Hsieh CC, Mantzoros C,
Trichopoulos D. Insulin-like growth factor 1 in hepatocellular carcinoma and
metastatic liver cancer in men. Int J Cancer. 2000;87:118–121.
86. Su WW, Lee KT, Yeh YT, Soon MS, Wang CL, Yu ML, Wang SN. Association
of circulating insulin-like growth factor 1 with hepatocellular carcinoma: one
cross-sectional correlation study. J Clin Lab Anal. 2010;24:195–200.
87. Luo SM, Tan WM, Deng WX, Zhuang SM, Luo JW. Expression of albumin,
IGF-1, IGFBP-3 in tumor tissues and adjacent non-tumor tissues of
hepatocellular carcinoma patients with cirrhosis. World J Gastroenterol.
2005;11:4272–4276.
88. Gray SG, Eriksson T, Ekström C, Holm S, von Schweinitz D, Kogner P,
Sandstedt B, Pietsch T, Ekström TJ., Altered expression of members of the
IGF-axis in hepatoblastomas, Br J Cancer.;82(9):1561-7., 2000.
89. Wolpin BM, Michaud DS, Giovannucci EL, Schernhammer ES, Stampfer MJ,
Manson JE, Cochrane BB, Rohan TE, Ma J, Pollak MN, Fuchs CS. Circulating
insulin-like growth factor axis and the risk of pancreatic cancer in four
prospective cohorts. Br J Cancer. 2007 Jul 2;97(1):98-104.
90. Stolzenberg-Solomon RZ, Limburg P, Pollak M, Taylor PR, Virtamo J, Albanes
D. Insulin-like growth factor (IGF)-1, IGF-binding protein-3, and pancreatic
cancer in male smokers. Cancer Epidemiol Biomarkers Prev. 2004
Mar;13(3):438-44.

56
91. Tomizawa M, Shinozaki F, Sugiyama T, Yamamoto S, Sueishi M, Yoshida T.,
Insulin-like growth factor-I receptor in proliferation and motility of pancreatic
cancer, World J Gastroenterol. 2010, 16:1854-8.
92. Vesely D, Astl J, Lastuvka P, Matucha P, Sterzl I, Betka J., Serum levels of
IGF-I, HGF, TGFbeta1, bFGF and VEGF in thyroid gland tumors, Physiol Res.
2004, 53(1):83-9.
93. Chakravarty G, Santillan AA, Galer C, Adams HP, El-Naggar AK, Jasser SA,
Mohsin S, Mondal D, Clayman GL, Myers JN., Phosphorylated insulin like
growth factor-I receptor expression and its clinico-pathological significance in
histologic subtypes of human thyroid cancer, Exp Biol Med. 2009, 234(4):372-
83.
94. Rhodes, Ch.J., Shoelson, S., Halban, P.A.: Insulin biosynthesis, processing,
and chemistry. In: Joslin´s Diabetes mellitus. Eds.: Kahn CR. et al., Lippincott
Williams&Wilkins, Philadelphia, 2005, 66-82.
95. Calle EE, Rodriguez C, Walker-Thurmond K, Thun MJ: Overweight, obesity,
and mortality from cancer in a prospectively studied cohort of U.S. adults. N
Engl J Med. 2003 Apr 24;348(17):1625-38.
96. Harish K, Dharmalingam M, Himanshu M.: Study Protocol: insulin and its role
in cancer. BMC Endocr Disord. 2007, Oct 22;7-10.
97. Ambati S., Kim H. K., Yang J. Y. Effects of leptin on apoptosis and
adipogenesis in 3T3-L1 adipocytes. Biochemical pharmacology , 2007, 73:
378 – 384.
98. Gonzalez-Perez RR, Xu Y, Guo S, Watters A, Zhou W, Leibovich SJ. Leptin
upregulates VEGF in breast cancer via canonic and non-canonical signalling
pathways and NFkappaB/HIF-1alpha activation. Cell Signal. 2010;22:1350-62
99. Gable D. R., Hurel S.J., Humphries S.E. Adiponectin and its gene variants as
risk factors for insulin resistance, the metabolic syndrome and cardiovascular
disease. Atherosclerosis, 2006, 188: 231–244.
100. Brakenhielm E, Veitonmaki N, Cao R, Kihara S, Matsuzawa Y, Zhivotovsky B,
Funahashi T, Cao Y: Adiponectin-induced antiangiogenesis and antitumor
activity involve caspase-mediated endothelial cell apoptosis. Proc Natl Acad
Sci. 2004, 101: 2476–2481.

57
101. Sutherland BW, Knoblaugh SE, Kaplan-Lefko PJ, Wang F, Holzenberger M,
Greenberg NM., Conditional deletion of insulin-like growth factor-I receptor in
prostate epithelium, Cancer Res. 2008, 68(9):3495-504.
102. Endo H, Hosono K, Uchiyama T, Sakai E, Sugiyama M, Takahashi H,
Nakajima N, Wada K, Takeda K, Nakagama H, Nakajima A. Leptin acts as a
growth factor for colorectal tumours at stages subsequent to tumour initiation
in murine colon carcinogenesis. Gut. 2011, 60(10): 1363-71.
103. Nishida M., Funahashi T and ShimomuraI.: Pathophysiological significance of
adiponectin. Med Mol Morphol. 2007, 40:55–67.
104. Pollak M. Insulin and insulin-like growth factor signalling in neoplasia. Nat Rev
Cancer. 2008;8:915–928.
105. Frasca F, Pandini G, Sciacca L, et al. et al. The role of insulin receptors and
IGF-I receptors in cancer and other diseases. Arch Physiol Biochem.
2008;114:23–37.
106. Tolcher AW, Sarantopoulos J, Patnaik A, et al. et al. A phase I
pharmacokinetic and pharmacodynamic study of AMG 479, a fully human
monoclonal antibody against insulin-like growth factor type 1 receptor. J Clin
Oncol. 2009;27:5800–5807.
107. Haluska P, Huang J, Lam B, et al. et al. MEDI-573 as a novel approach to
IGF-1R and IR-A signaling inhibition by blocking IGF ligands: Phase I PK/PD,
safety data, and disease linkage studies in breast cancer. J Clin Oncol.
2011;29(suppl 27):abstr 271.
108. Carden CP, Frentzas S, Langham M, et al. et al. Preliminary activity in
adrenocortical tumor (ACC) in phase I dose escalation study of intermittent
oral dosing of OSI-906, a small-molecule insulin-like growth factor-1 receptor
(IGF-1R) tyrosine kinase inhibitor in patients with advanced solid tumors. J
Clin Oncol. 2009;27(suppl):abstr 3544.






![zcu.czkke.zcu.cz/old_web/_files/projekty/enazp/25/IUT/165... · Web viewv živé buňce to jsou enzymy [4, str. 168] (druh bílkoviny, která podstatně urychluje chemickou reakci](https://img.pdfslide.tips/doc/110x75/610454d7b9b90726f3151100/zcuczkkezcuczoldwebfilesprojektyenazp25iut165-web-view-v-iv.jpg)






















