Embed Size (px)
Citation preview

UNIVERSITATEA ALEXANDRU IOAN CUZA DIN IAŞI FACULTATEA DE BIOLOGIE
PROIECT IDEI nr. 1073/2009, COD CNCSIS 85
IMPACTUL UNOR SUPERANTIGENE DE ORIGINE BACTERIANĂ ASUPRA STATUSULUI
FIZIOLOGIC AL ORGANISMULUI ANIMAL
- sinteză lucrare fază finală 2009 -
Director proiect,
Lector dr. Lucian HRIŢCU

1. STADIUL ACTUAL AL CERCETĂRILOR PRIVIND COMPLEXITATEA INTERACŢIUNILOR DINTRE SUPERANTIGENELE BACTERIENE ŞI
ORGANISMUL ANIMAL
Studiul efectelor superantigenelor bacteriene asupra organismului animal a suscitat interesul ştiinţific
datorită efectelor lor puternice de stimulare a activităţii limfocitelor T şi de iniţiere a unor condiţii
patofiziologice, incluzând şocul septic şi bolile autoimune [12].
Dintre toate superantigenele, cele mai intens studiate sunt enterotoxinele produse de bacteria Gram-
pozitivă - Staphylococcus aureus – în special enterotoxinele stafilococice A şi B (SEA / SEB) [12]. Ca activatori
ai celulelor T, SEA şi SEB acţionează într-un mod unic, în sensul că activarea limfocitelor T se realizează
independent de digestia intracelulară, incluzând moleculele clasei II MHC [26]. Activarea limfocitelor T se
realizează prin intermediul receptorilor T celulari (TCR) [6]. Numeroase studii experimentale, implicând
reactivitatea organismului animal faţă de SEA şi SEB, au demonstrat că această reactivitate este rezultatul unei
sinteze rapide de citokine, precum şi a factorului de necroză tumorală (TNF) şi interleukinei (IL-2) la 1-2 ore
după administrarea lor [22]. Studii anterioare au evidenţiat faptul în urma administrării SEA şi a SEB, la şoareci
şi la şobolani au loc alterări ale funcţiilor endocrine, neurobiologice şi comportamentale [9, 17]. De exemplu,
prin administrarea SEB la şoarecii BALB/cByJ, se produce stimularea axei hipotalamo-hipofizo-
corticosuprarenaliene (HHC), având ca rezultat creşterea nivelului plasmatic al corticotropinei (ACTH) şi al
corticosteronului. Acest proces este dependent de corticoliberina hipotalamică (CRH) şi de intensificarea ARNm
CRH în hipotalamus, precum şi în nucleul central al amigdalei. În mod similar, prin administrarea SEA la
şoarecii C57BL/6J se produce creşterea nivelului plasmatic al ACTH şi al corticosteronului [9]. Efectele
neuroendocrine şi comportamentale ale SEA si SEA se datorează specificităţii particulare a fiecărui antigen faţă
de diferiţi receptori TCR. Într-adevăr, în cazul celulelor murine, regiunea lanţului β al receptorului TCR este
codificată de cel puţin 20 de gene diferite care pot da naştere unor secvenţe unice de aminoacizi [5]. Acest lucru
va determina diferenţierea limfocitelor T (CD4 şi CD8), în subtipuri, pe baza contribuţiei genei Vβ în construcţia
TCR. Enterotoxinele stafilococice A şi B posedă afinităţi diferite pentru subtipurile Vβ (SEB stimulează
limfocitele T Vβ8, în timp ce SEA prezintă o afinitate ridicată pentru limfocitele T Vβ3 şi Vβ11). Mai mult,
efectele neuroendocrine ale SEA si SEB necesită prezenţa limfocitelor T funcţionale, lucru confirmat prin
imunosupresia cu ciclosporină A, aspect observat la şoarecii transgenici RAG-1, lipsiţi de limfocite funcţionale
[9]. Efectele SEB au fost evidenţiate prin studii ale comportamentului de ingestie. Astfel, administrarea SEB
determină o reducere semnificativă a consumării unei soluţii noi, iar efectul este menţinut de noutatea
contextului în care soluţia este prezentă. Expunerea animalelor de laborator la un context nou sau la excitanţi
gustativi determină activarea axei HHC şi creşterea latenţei de interacţiune cu un nou stimul [11]. SEB determină
efecte inhibitoare asupra proceselor de ingestie, inducând anxietate. Animalele tratate cu SEB manifestă o
reactivitate ridicată la stimulii înconjurători, ca în cazul răspunsurilor defensive.
Tot dintre superantigenele bacteriene, mai sunt studiate şi endotoxinele, cum ar fi lipopolizaharidele
bacteriene (LPS), care sunt constituenţi ai membranei bacteriilor Gram-negative (de exemplu, Escherichia coli).
LPS au fost utilizate în multe studii neuro-imunologice datorită abilităţii lor de a induce un răspuns rapid la

nivelul gazdei, într-o manieră doză-dependentă. LPS induc secreţia citokinelor (TNF, IL-1, IL-6), care dacă ar fi
injectate în formă lor pură ar mima unele efecte ale LPS în organismul gazdă [1].
Numeroase studii au evidenţiat efectele LPS asupra funcţiilor organismului animal. De exemplu, prin
administrarea LPS la şobolani, s-au constatat modificări ale reactivităţii faţă de stres, disfuncţii imunologice şi
creşteri ale susceptibilităţii la îmbolnăvire [3, 7, 14]. LPS pot induce alterări comportamentale, în special prin
intermediul citokinelor în timpul dezvoltării cortexului şi în special a hipocampului, o regiune importantă în
patologia comportamentală şi în bolile nervoase. Receptorii pentru citokine sunt distribuiţi la nivelul creierului,
având o densitate ridicată în hipocamp [13]. Din acest motiv, hipocampul pare să fie vulnerabil la alterările
imune [13]. Numeroase studii au raportat ca infectarea neonatală cu Escherichia coli este asociată cu alterări ale
proceselor cognitive (învăţarea şi memoria) [2]. Acest lucru poate fi evidenţiat numai dacă LPS sunt administrate
imediat după procesul de învăţare.
Identificarea relaţiilor neuro-imuno-endocrine va permite elaborarea unor noi modele experimentale de
studiu a acţiunii superantigenelor în raport cu imunitatea, metabolismul şi funcţiile cognitive, şi totodată va
facilita descifrarea parţială a mecanismelor complexe implicate în declanşarea bolilor autoimune şi
neurodegenerative.
2. STABILIREA UNUI MODEL EXPERIMENTAL DE ANALIZĂ A RELAŢIEI
TEMPORALE SUPERANTIGEN-RASPUNS IMUN
2.1. Imunizarea animalelor de laborator (şobolani Wistar) prin administrarea acută sau cronică asuperantigenelor bacteriene şi analiza fazelor de dezvoltare a
răspunsului imun
2.1.1. Efectul administrării sistemice acute a superantigenelor bacteriene asupra răspunsului imun în şobolanii normali
Răspunsul neuroimunitar este caracterizat prin comunicarea bidirecţională dintre imunocitele periferice
şi sistemul nervos central. Una dintre particularităţile infecţiei cu LPS constă în declanşarea răspunsurilor febrile
şi secreţia corticosteronului, concomitent cu activarea sistemului imunitar, care va limita extinderea răspunsului
inflamator [8, 18]. Inhibarea acestor răspunsuri imune va afecta semnificativ morbiditatea şi mortalitatea asociată
infecţiei [8, 10, 15]. Lipopolizaharidul (LPS), bogat în acizi graşi, are activitate similară cu endotoxina
bacteriilor Gram-negative. Anticorpii, ca răspuns la infecţiile cu LPS apar la 7 zile de la imunizare, ating nivelul
maxim după 1 lună şi se menţin 2-3 luni după care dispar [24]. Astfel, administrarea acută a endotoxinelor
bacteriene - lipopolizaharidelor - în timpul perioadei neonatale poate altera semnificativ răspunsurile febrile,
imunologice, neuroendocrine, neurochimice şi comportamentale ale animalelor de experienţă [3, 4, 19, 21]. De
asemenea, LPS, derivat din bacteriile Gram-negative (Escherichia coli), reprezintă unul dintre stimulii imunitari,
a căror activitate este mediată de receptorii TLR-4 [23].
Materiale şi metode Animale Au fost utilizaţi 30 de şobolani Wistar, cântărind 200-250g la începutul experimentului. Animalele au
fost întreţinute într-o cameră specială (Laboratorul de Fiziologie animală, Facultatea de Biologie, Iaşi), cu
temperatură şi lumină controlate (22°C, ciclu de 12 ore începând cu ora 8), şi au avut acces liber la hrană şi apă
ad libitum. Şobolanii au fost trataţi în conformitate cu normele bioetice instituţionale în vigoare şi toate
procedurile efectuate au fost în concordanţă cu Directiva Consiliului European din 26 noiembrie 1986
(86/609/EEC).
Administrarea acută a LPS Pentru studiul implicării endotoxinelor bacteriene – lipopolizaharidelor (LPS) - în modularea
reactivităţii imunologice, s-au luat în studiu două loturi de animale.
Loturile de animale luate în studiu au fost formate din câte 15 şobolani, identici ca sex, greutate şi
vârstă. Un lot de animale a constituit lotul de control (n=15), injectat acut, o singură dată pe parcursul
experimentului, intraperitoneal (i.p.), cu o soluţie fiziologică sterilă, administrată într-un volum de
0,1ml/100g/şobolan. Un al doilea lot de animale (n=15), a fost imunizat prin injectarea intraperitoneală (i.p.) a
unei soluţii de LPS (lipopolizaharid de la Escherichia coli, serotipul 0111:B4, Sigma) (25µg/250µl soluţie salină
sterilă). Soluţia de LPS a fost administrată o singură dată (acut) pe parcursul experimentului.
Analiza fazei de dezvoltare a răspunsului imun s-a realizat în a 7-a zi de la imunizare. Toate animalele
luate în studiu au fost sacrificate în a 7-a zi de la imunizare. Probele de sânge au fost incubate timp de o oră la
temperatura camerei, apoi centrifugate pentru separarea serului (3000 rpm, timp de 10 minute). Serul a fost
utilizat pentru determinarea cantităţii totale de proteine serice, a globulinelor totale şi a raportului
albumină/globulină (metoda Weichselbaum – testul biuret pentru proteine). Loturile de animale luate în studiu au
fost cântărite la începutul experimentului, înaintea inducerii imunizării şi la finalul experimentului, pentru a
analiza evoluţia greutăţii lor în urma tratamentelor aplicate.
Datele obţinute au fost prelucrate şi interpretate statistic utilizând testul „T-test” a lui Student.
Rezultatele sunt exprimate prin medie ± E.S.M. (eroarea standard a mediei). S-au considerat semnificative din
punct de vedere statistic valorile lui p<0,05.
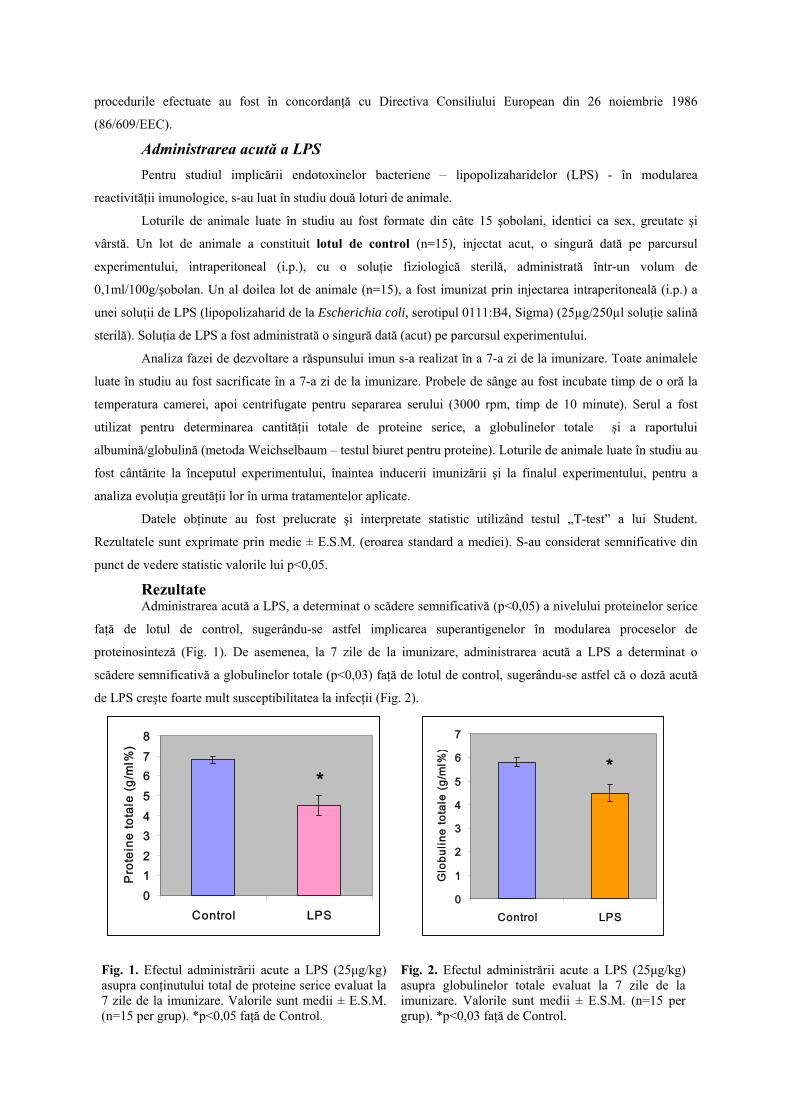
Rezultate Administrarea acută a LPS, a determinat o scădere semnificativă (p<0,05) a nivelului proteinelor serice
faţă de lotul de control, sugerându-se astfel implicarea superantigenelor în modularea proceselor de
proteinosinteză (Fig. 1). De asemenea, la 7 zile de la imunizare, administrarea acută a LPS a determinat o
scădere semnificativă a globulinelor totale (p<0,03) faţă de lotul de control, sugerându-se astfel că o doză acută
de LPS creşte foarte mult susceptibilitatea la infecţii (Fig. 2).
*
012345678
Control LPS
Pro
tein
e to
tale
(g/m
l%)
*
0
1
2
3
4
5
6
7
Control LPS
Glo
bulin
e to
tale
(g/m
l%)
Fig. 1. Efectul administrării acute a LPS (25μg/kg) asupra conţinutului total de proteine serice evaluat la 7 zile de la imunizare. Valorile sunt medii ± E.S.M. (n=15 per grup). *p<0,05 faţă de Control.
Fig. 2. Efectul administrării acute a LPS (25μg/kg) asupra globulinelor totale evaluat la 7 zile de la imunizare. Valorile sunt medii ± E.S.M. (n=15 per grup). *p<0,03 faţă de Control.
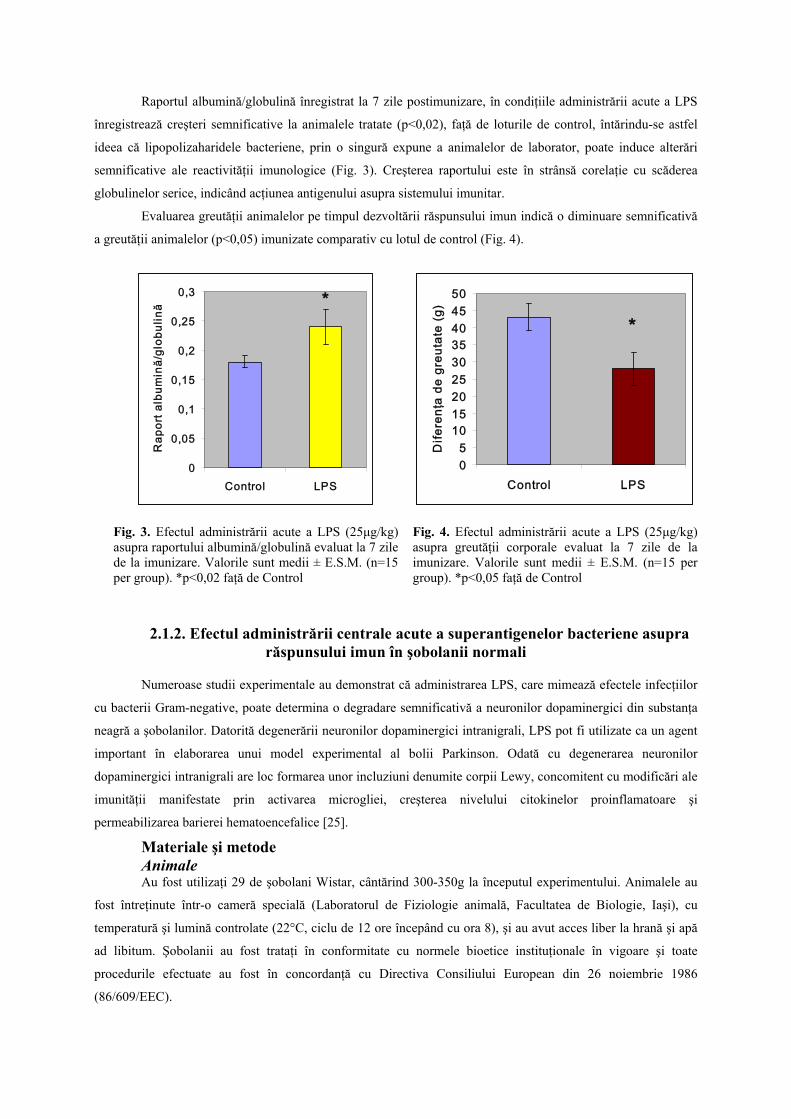
Raportul albumină/globulină înregistrat la 7 zile postimunizare, în condiţiile administrării acute a LPS
înregistrează creşteri semnificative la animalele tratate (p<0,02), faţă de loturile de control, întărindu-se astfel
ideea că lipopolizaharidele bacteriene, prin o singură expune a animalelor de laborator, poate induce alterări
semnificative ale reactivităţii imunologice (Fig. 3). Creşterea raportului este în strânsă corelaţie cu scăderea
globulinelor serice, indicând acţiunea antigenului asupra sistemului imunitar.
Evaluarea greutăţii animalelor pe timpul dezvoltării răspunsului imun indică o diminuare semnificativă
a greutăţii animalelor (p<0,05) imunizate comparativ cu lotul de control (Fig. 4).
*
0
0,05
0,1
0,15
0,2
0,25
0,3
Control LPS
Rap
ort a
lbum
ină/
glob
ulin
ă
*
05
101520253035404550
Control LPS
Dife
renţ
a de
gre
utat
e (g
)
Fig. 3. Efectul administrării acute a LPS (25μg/kg) asupra raportului albumină/globulină evaluat la 7 zile de la imunizare. Valorile sunt medii ± E.S.M. (n=15 per group). *p<0,02 faţă de Control
Fig. 4. Efectul administrării acute a LPS (25μg/kg) asupra greutăţii corporale evaluat la 7 zile de la imunizare. Valorile sunt medii ± E.S.M. (n=15 per group). *p<0,05 faţă de Control
2.1.2. Efectul administrării centrale acute a superantigenelor bacteriene asupra răspunsului imun în şobolanii normali
Numeroase studii experimentale au demonstrat că administrarea LPS, care mimează efectele infecţiilor
cu bacterii Gram-negative, poate determina o degradare semnificativă a neuronilor dopaminergici din substanţa
neagră a şobolanilor. Datorită degenerării neuronilor dopaminergici intranigrali, LPS pot fi utilizate ca un agent
important în elaborarea unui model experimental al bolii Parkinson. Odată cu degenerarea neuronilor
dopaminergici intranigrali are loc formarea unor incluziuni denumite corpii Lewy, concomitent cu modificări ale
imunităţii manifestate prin activarea microgliei, creşterea nivelului citokinelor proinflamatoare şi
permeabilizarea barierei hematoencefalice [25].
Materiale şi metode Animale Au fost utilizaţi 29 de şobolani Wistar, cântărind 300-350g la începutul experimentului. Animalele au
fost întreţinute într-o cameră specială (Laboratorul de Fiziologie animală, Facultatea de Biologie, Iaşi), cu
temperatură şi lumină controlate (22°C, ciclu de 12 ore începând cu ora 8), şi au avut acces liber la hrană şi apă
ad libitum. Şobolanii au fost trataţi în conformitate cu normele bioetice instituţionale în vigoare şi toate
procedurile efectuate au fost în concordanţă cu Directiva Consiliului European din 26 noiembrie 1986
(86/609/EEC).

Administrarea acută a LPS LPS a fost injectat stereotaxic în substanţa neagră (SN) a şobolanilor normali. Toate procedurile
chirurgicale s-au desfăşurat în condiţii aseptice, iar animalele au fost anesteziate cu nembutal (45 mg/kg, g.c.,
i.p., Sigma). S-a preparat o soluţie stoc de LPS (lipopolizaharid de la Escherichia coli, serotipul 0111: B4,
Sigma) în concentraţie de 1 µg/µl în soluţie fiziologică sterilă. Injectarea stereotaxică a LPS în SN s-a realizat cu
o microseringă Hamilton (10 µl), după următoarelor coordonate stereotaxice: 5,5 mm posterior de bregma; 2 mm
lateral de linia mediană; 7,4 mm ventral faţă de suprafaţa cortexului [16]. La fiecare injectare, acul seringei a fost
lăsat in situ, o perioadă de 5 minute, în vederea evitării refluxului substanţei injectate. Etapele realizării
intervenţiilor neurochirurgicale pot fi vizualizate în figura 5.
Şobolanii trataţi cu LPS au fost divizaţi în două loturi în funcţie de doza de LPS administrată (3 µg şi 10
µg). Loturile de control (fals-operate) au fost injectate stereotaxic în SN cu aceleaşi volume de soluţie
fiziologică sterilă. Analiza fazei de dezvoltare a răspunsului imun s-a realizat în a 7-a zi de la imunizare.
La 7 zile postimunizare, toţi şobolanii au fost anesteziaţi, rapid decapitaţi şi s-a realizat recoltarea
sângelui. Probele de sânge au fost incubate timp de o oră la temperatura camerei, apoi s-a realizat separarea
serului prin centrifugare la 3000 rpm, timp de 15 minute.
S-a utilizat metoda SDS-PAGE pentru analiza proteinelor solubile [14], precum şi densitometria în gel
pentru determinarea maselor moleculare şi cuantificarea cantităţilor de proteine folosind tehnica IMAGE
QUANT (GE Healthcare, Germania) [14].
Datele obţinute au fost prelucrate şi interpretate statistic utilizând testul „T-test” a lui Student.
Rezultatele sunt exprimate prin medie ± E.S.M. (eroarea standard a mediei). S-au considerat semnificative din
punct de vedere statistic valorile lui p<0,05.
Rezultate În probele de ser de la animalele tratate (şobolani trataţi cu LPS, 3 µg şi 10 µg) şi de la lotul de control
au putut fi separate şi vizualizate prin coloraţia Commasie în jur de 20 de fracţiuni proteice. Profilul
electroforetic al proteinelor plasmatice indică o mare heterogenitate (Fig. 6), cu 4 fracţiuni proteice
predominante, reprezentând circa 60-70% din conţinutul total de proteine. În funcţie de greutatea lor moleculară,
trei din aceste 4 proteine au fost recunoscute ca fiind albumine (61 kDa), gamma-globuline (lanţ greu – 51 kDa
şi lanţ uşor – 21 kDa). A patra bandă este o proteină necunoscută cu greutatea de 78 kDa luată în mod arbitrar şi
considerată a fi control.
S-a realizat o curbă etalon, utilizând albumina serică bovină (ASB), iar prin densitometria gelurilor s-a
putut cuantifica nivelul fiecărei fracţiuni proteice. Astfel, în tabelul 1 se poate observa că injectarea şobolanilor
cu LPS (3µg şi 10µg) a determinat o scădere semnificativă a albuminelor plasmatice (p<0,001) comparativ cu
loturile de control, şi o creştere semnificativă a nivelului gamma-globulinelor (lanţ greu - p<0,02 şi lanţul uşor -
p<0,004).

Fig. 5. Etapele administrării stereotaxice a LPS (foto originale)
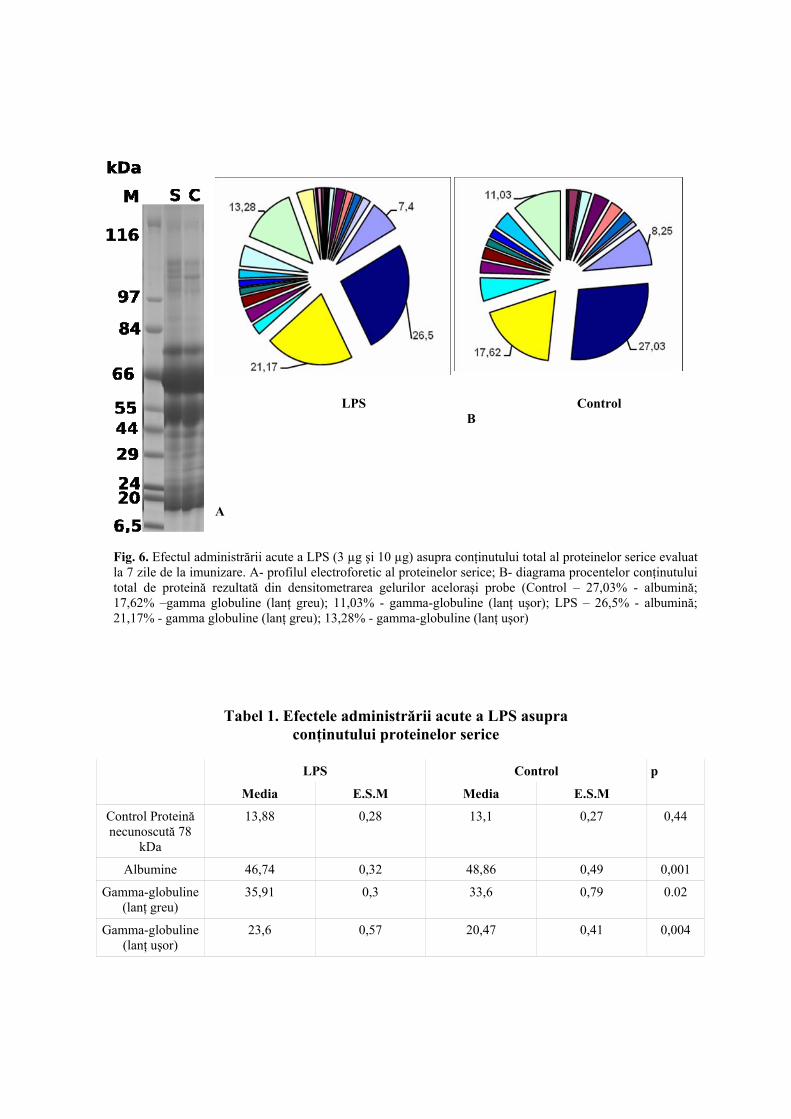
Fig. 6. Efectul administrării acute a LPS (3 µg şi 10 µg) asupra conţinutului total al proteinelor serice evaluat la 7 zile de la imunizare. A- profilul electroforetic al proteinelor serice; B- diagrama procentelor conţinutului total de proteină rezultată din densitometrarea gelurilor aceloraşi probe (Control – 27,03% - albumină; 17,62% –gamma globuline (lanţ greu); 11,03% - gamma-globuline (lanţ uşor); LPS – 26,5% - albumină; 21,17% - gamma globuline (lanţ greu); 13,28% - gamma-globuline (lanţ uşor)
Tabel 1. Efectele administrării acute a LPS asupra conţinutului proteinelor serice
LPS Control
Media E.S.M Media E.S.M
p
Control Proteină necunoscută 78
kDa
13,88 0,28 13,1 0,27 0,44
Albumine 46,74 0,32 48,86 0,49 0,001
Gamma-globuline (lanţ greu)
35,91 0,3 33,6 0,79 0.02
Gamma-globuline (lanţ uşor)
23,6 0,57 20,47 0,41 0,004
LPS Control B
A

2.1.3. Efectul administrării sistemice cronice a superantigenelor bacteriene asupra răspunsului imun în şobolanii normali
După administrarea acută a LPS, animalele vor dezvolta un răspuns imun adaptativ având ca efect
diminuarea activităţii sistemului imunitar.
Materiale şi metode
Animale Au fost utilizaţi 30 de şobolani Wistar, cântărind 180-230g la începutul experimentului. Animalele au
fost întreţinute într-o cameră specială (Laboratorul de Fiziologie animală, Facultatea de Biologie, Iaşi), cu
temperatură şi lumină controlate (22°C, ciclu de 12 ore începând cu ora 8), şi au avut acces liber la hrană şi apă
ad libitum. Şobolanii au fost trataţi în conformitate cu normele bioetice instituţionale în vigoare şi toate
procedurile efectuate au fost în concordanţă cu Directiva Consiliului European din 26 noiembrie 1986
(86/609/EEC). Administrarea cronică a LPS Pentru studiul implicării endotoxinelor bacteriene – lipopolizaharidelor – în modularea reactivităţii
imunologice, s-au luat în studiu două loturi de animale.
Loturile de animale luate în studiu au fost formate din câte 15 şobolani, identici ca sex, greutate şi
vârstă. Un lot de animale a constituit lotul de control (n=15), injectat cronic, intraperitoneal (i.p.), zilnic, timp
de 7 zile, cu o soluţie fiziologică sterilă, administrată într-un volum de 0,1ml/100g/şobolan.
Un al doilea lot de animale (n=15), a fost imunizat prin injectarea intraperitoneală (i.p.) a unei soluţii de
LPS (lipopolizaharid de la Escherichia coli, serotipul 0111:B4, Sigma) (250µg/kg soluţie salină sterilă). Soluţia
de LPS a fost administrată cronic, timp de 7 zile consecutive, pe parcursul experimentului.
Analiza fazei de dezvoltare a răspunsului imun s-a realizat în a 7-a zi de la imunizare. Toate animalele
luate în studiu au fost sacrificate în a 7-a zi de la imunizare. Probele de sânge au fost incubate timp de oră la
temperatura camerei, apoi centrifugate pentru separarea serului (3000 rpm, timp de 10 minute). Serul a fost
utilizat pentru determinarea cantităţii totale de proteine serice, a globulinelor totale şi a raportului
albumină/globulină (metoda Weichselbaum – testul biuret pentru proteine). Loturile de animale luate în studiu au
fost cântărite la începutul experimentului, înaintea inducerii imunizării şi la finalul experimentului, pentru a
analiza evoluţia greutăţii lor în urma tratamentelor aplicate.
Datele obţinute au fost prelucrate şi interpretate statistic utilizând testul „T-test” a lui Student.
Rezultatele sunt exprimate prin medie ± E.S.M. (eroarea standard a mediei). S-au considerat semnificative din
punct de vedere statistic valorile lui p<0,05.
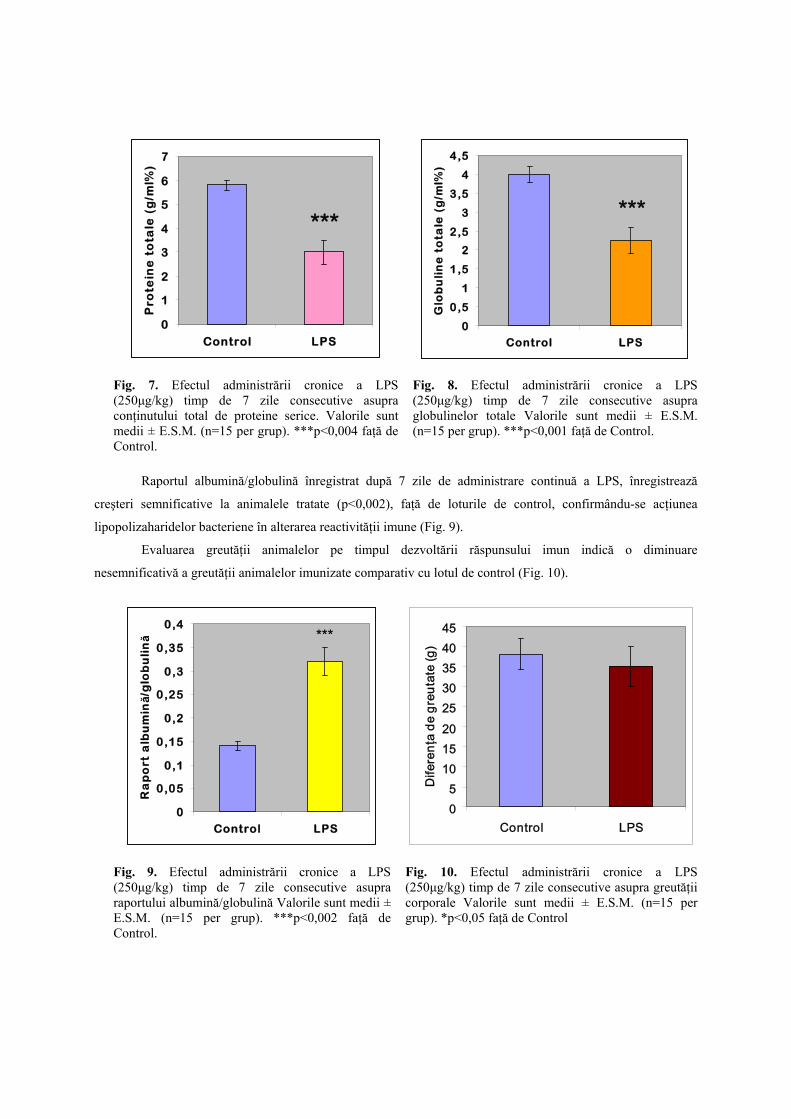
Rezultate Administrarea cronică a LPS, a determinat o scădere semnificativă (p<0,004) a nivelului proteinelor
serice faţă de lotul de control, sugerându-se astfel supresia semnificativă a proceselor de proteinosinteză (Fig. 7).
De asemenea, la 7 zile de la imunizare, administrarea cronică a LPS a determinat o scădere semnificativă a
globulinelor totale (p<0,001) faţă de lotul de control, sugerându-se importanţa majoră a lipopolizaharidelor
bacteriene în supresia vegetativă a reactivităţii imunitare concomitent cu exacerbarea degradării reactivităţii
imunitare (Fig. 8).
***
0
1
2
3
4
5
6
7
Control LPS
Pro
tein
e t
ota
le (
g/m
l%)
***
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
Control LPS
Glo
bu
lin
e t
ota
le (
g/m
l%)
Fig. 7. Efectul administrării cronice a LPS (250μg/kg) timp de 7 zile consecutive asupra conţinutului total de proteine serice. Valorile sunt medii ± E.S.M. (n=15 per grup). ***p<0,004 faţă de Control.
Fig. 8. Efectul administrării cronice a LPS (250μg/kg) timp de 7 zile consecutive asupra globulinelor totale Valorile sunt medii ± E.S.M. (n=15 per grup). ***p<0,001 faţă de Control.
Raportul albumină/globulină înregistrat după 7 zile de administrare continuă a LPS, înregistrează
creşteri semnificative la animalele tratate (p<0,002), faţă de loturile de control, confirmându-se acţiunea
lipopolizaharidelor bacteriene în alterarea reactivităţii imune (Fig. 9).
Evaluarea greutăţii animalelor pe timpul dezvoltării răspunsului imun indică o diminuare
nesemnificativă a greutăţii animalelor imunizate comparativ cu lotul de control (Fig. 10).
***
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Control LPS
Ra
po
rt a
lbu
mină/
glo
bu
lină
05
1015202530354045
Control LPS
Dife
renţ
a de
gre
utat
e (g
)
Fig. 9. Efectul administrării cronice a LPS (250μg/kg) timp de 7 zile consecutive asupra raportului albumină/globulină Valorile sunt medii ± E.S.M. (n=15 per grup). ***p<0,002 faţă de Control.
Fig. 10. Efectul administrării cronice a LPS (250μg/kg) timp de 7 zile consecutive asupra greutăţii corporale Valorile sunt medii ± E.S.M. (n=15 per grup). *p<0,05 faţă de Control
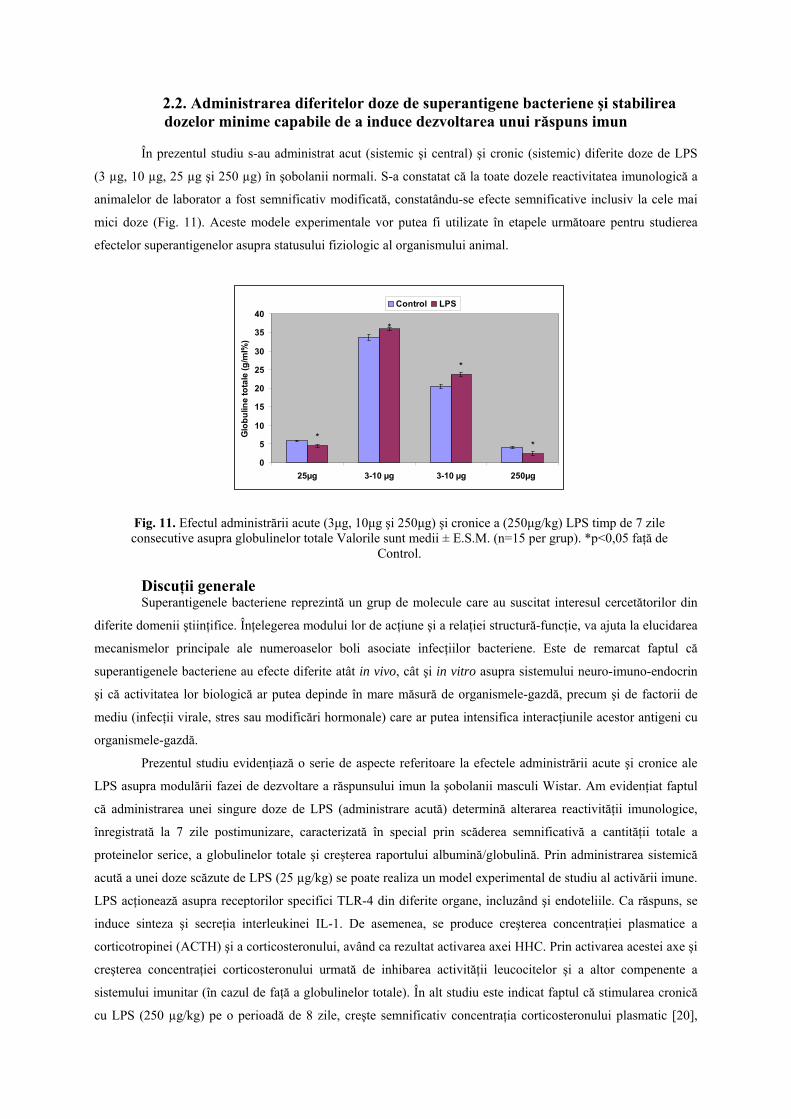
2.2. Administrarea diferitelor doze de superantigene bacteriene şi stabilirea dozelor minime capabile de a induce dezvoltarea unui răspuns imun
În prezentul studiu s-au administrat acut (sistemic şi central) şi cronic (sistemic) diferite doze de LPS
(3 µg, 10 µg, 25 µg şi 250 µg) în şobolanii normali. S-a constatat că la toate dozele reactivitatea imunologică a
animalelor de laborator a fost semnificativ modificată, constatându-se efecte semnificative inclusiv la cele mai
mici doze (Fig. 11). Aceste modele experimentale vor putea fi utilizate în etapele următoare pentru studierea
efectelor superantigenelor asupra statusului fiziologic al organismului animal.
*
*
*
*
0
5
10
15
20
25
30
35
40
25µg 3-10 µg 3-10 µg 250µg
Glo
bulin
e to
tale
(g/m
l%)
Control LPS
Fig. 11. Efectul administrării acute (3μg, 10μg şi 250μg) şi cronice a (250μg/kg) LPS timp de 7 zile consecutive asupra globulinelor totale Valorile sunt medii ± E.S.M. (n=15 per grup). *p<0,05 faţă de
Control.
Discuţii generale Superantigenele bacteriene reprezintă un grup de molecule care au suscitat interesul cercetătorilor din
diferite domenii ştiinţifice. Înţelegerea modului lor de acţiune şi a relaţiei structură-funcţie, va ajuta la elucidarea
mecanismelor principale ale numeroaselor boli asociate infecţiilor bacteriene. Este de remarcat faptul că
superantigenele bacteriene au efecte diferite atât in vivo, cât şi in vitro asupra sistemului neuro-imuno-endocrin
şi că activitatea lor biologică ar putea depinde în mare măsură de organismele-gazdă, precum şi de factorii de
mediu (infecţii virale, stres sau modificări hormonale) care ar putea intensifica interacţiunile acestor antigeni cu
organismele-gazdă.
Prezentul studiu evidenţiază o serie de aspecte referitoare la efectele administrării acute şi cronice ale
LPS asupra modulării fazei de dezvoltare a răspunsului imun la şobolanii masculi Wistar. Am evidenţiat faptul
că administrarea unei singure doze de LPS (administrare acută) determină alterarea reactivităţii imunologice,
înregistrată la 7 zile postimunizare, caracterizată în special prin scăderea semnificativă a cantităţii totale a
proteinelor serice, a globulinelor totale şi creşterea raportului albumină/globulină. Prin administrarea sistemică
acută a unei doze scăzute de LPS (25 µg/kg) se poate realiza un model experimental de studiu al activării imune.
LPS acţionează asupra receptorilor specifici TLR-4 din diferite organe, incluzând şi endoteliile. Ca răspuns, se
induce sinteza şi secreţia interleukinei IL-1. De asemenea, se produce creşterea concentraţiei plasmatice a
corticotropinei (ACTH) şi a corticosteronului, având ca rezultat activarea axei HHC. Prin activarea acestei axe şi
creşterea concentraţiei corticosteronului urmată de inhibarea activităţii leucocitelor şi a altor compenente a
sistemului imunitar (în cazul de faţă a globulinelor totale). În alt studiu este indicat faptul că stimularea cronică
cu LPS (250 µg/kg) pe o perioadă de 8 zile, creşte semnificativ concentraţia corticosteronului plasmatic [20],

având ca efect intensificarea stimulării axei HHC, concomitent cu intensificarea stresului. Din acest considerent,
în prezentul studiu, posibila scădere a nivelului globulinelor totale, cantităţii totale de proteine serice şi creşterea
raportului albunină/globulină, prin injectarea reperată (cronică) a LPS, timp de 7 zile, s-ar datora intensificării
stresului (stres cronic), ca răspuns al activării axei HHC.
Unii autori au demonstrat că administrarea LPS, poate determina o degradare semnificativă a neuronilor
dopaminergici din substanţa neagră a şobolanilor, determinând simptoame caracteristice maladiei Parkinson
[25]. O serie de maladii neurodegenerative (Parkinson, Alzheimer) sunt asociate cu activarea sistemului
imunitar. Astfel, în prezentul studiu prin injectarea acută a LPS (3µg şi 10 µg), a avut loc activarea sistemului
imunitar în sensul creşterii semnificative a nivelului globulinelor (lanţ greu şi uşor) la 7 zile postimunizare. De
remarcat este faptul ca prin acţiunea specifică a LPS asupra neuronilor dopaminergici din substanţa neagră a
şobolanilor, se poate crea un model experimental al bolii Parkinson. Înţelegerea proceselor în care aceste
răspunsuri imune determină neurodegenerescenţă poate servi la elaborarea unor strategii terapeutice
caracteristice unor maladii neurodegenerative (Parkinson, Alzheimer).
Concluzii Pe baza datelor obţinute în prezentul studiu, se poate afirma că prin administrarea superantigenelor se
poate constitui un model experimental de studiu al activării imune. Administrarea sistemică acută şi cronică a
superantigenelor altereză semnificativ reactivitatea imunologică, în timp ce administrarea centrală acută a LPS
intensifică răspunsul imun al animalelor de experienţă.
Bibliografie
[1] A.K. Abbas, A.H. Lichtman, J.S. Pober, Cytokines, Cell. Mol. Immunol. 3 (1997) 250-277. [2] S.D. Bilbo, L.H. Levkoff, J.H. Mahoney, L.R. Watkins, J.W. Rudy, S.F. Maier, Neonatal infection
induces memory impairments following an immune challenge in adulthood, Behav Neurosci 119 (2005) 293-301.
[3] L. Boisse, A. Mouihate, S. Ellis, Q.J. Pittman, Long-term alterations in neuroimmune responses after neonatal exposure to lipopolysaccharide, J Neurosci 24 (2004) 4928-4934.
[4] S. Ellis, A. Mouihate, Q.J. Pittman, Early life immune challenge alters innate immune responses to lipopolysaccharide: implications for host defense as adults FASEB J 19 (2005) 1519-1521.
[5] R. Gonzalez-Quintial, R. Baccala, R.S. Balderas, A.N. Theofilopoulos, V beta gene repertoire in the aging mouse: a developmental perspective, Int. Rev. Immunol. 12 (1995) 27-40.
[6] S.C. Hong, G. Waterbury, C.A. Janeway Jr., Different superantigens interact with distinct sites in the Vbeta domain of a single T cell receptor, J. Exp. Med. 183 (1996) 1437-1446.
[7] L. Hritcu, M. Stefan, Acute lypopolisaccharide adminstration impaired imuune responsiveness in normal rats., Analele stiintifice ale Universitatii Alexandru Ioan Cuza, Sectiunea Genetica si Biologie Moleculara TOM X (2009) 63-68.
[8] Q. Jiang, A.S. Cross, I.S. Singh, T.T. Chen, R.M. Viscardi, J.D. Hasday, Febrile Core Temperature Is Essential for Optimal Host Defense in Bacterial Peritonitis, Infect. Immun. 68 (2000) 1265-1270.
[9] N. Kawashima, A.W. Kusnecov, Effects of staphylococcal enterotoxin A on pituitary–adrenal activation and neophobic behavior in the C57BL/6 mouse, J. Neuroimmunol. 123 (2002) 41-49.
[10] M.J. Kluger, W. Kozak, C.A. Conn, L.R. Leon, D. Soszynski, "Role of Fever in Disease", Ann.N.Y.Acad.Sci. 856 (1998) 224-233.
[11] C. Kopp, E. Vogel, M.C. Rettori, P. Delagrange, B. Guardiola-Lemaitre, R. Misslin, Effects of melatonin on neophobic responses in different strains of mice, Pharmacol. Biochem. Behav. 63 (1999) 521-526.
[12] M. Llewelyn, J. Cohen, Superantigens: microbial agents that corrupt immunity, Lancet Infect. Dis. 2 (2002) 156-162.
[13] A.M. Lynch, C. Walsh, A. Delaney, Y. Nolan, V.A. Campbell, M.A. Lynch, Lipopolissacharide-induced increase in signalling in hippocampus is abrogated by IL-10-a role for IL-1 beta?, J Neurochem 88 (2004) 635-646.

[14] M. Mihasan, L. Hritcu, V. Artenie, A. Ciobica, M. Stefan, L. Gorgan, Bacterial lypopolisaccharide enhanced immunological responsiveness in exposed rats, Analele stiintifice ale Universitatii Alexandru Ioan Cuza, Sectiunea Genetica si Biologie Moleculara TOM X (2009) 41-44.
[15] S. Nadeau, S. Rivest, Glucocorticoids Play a Fundamental Role in Protecting the Brain during Innate Immune Response J Neurosci 23 (2003) 5536-5544.
[16] G. Paxinos, C. Watson, A.P. 2005, Sidney, The Rat Brain in Stereotaxic Coordinates, Academic Press, Sidney, 2005.
[17] A. Rossi-George, F. LeBlanc, T. Kaneta, D. Urbach, W.A. Kusnecov, Effects of bacterial superantigens on behavior of mice in the elevated plus maze and light–dark box, Brain Behav Immun 18 (2004) 46-54.
[18] B. Schobitz, J.M.H.M. Reul, F. Holsboer, The role of the hypothalamic-pituitary-adrenocortical system during inflammatory conditions, Critical Rev Neurobiol. 8 (1994) 263-291.
[19] N. Shanks, R.J. Windle, P.A. Perks, M.S. Harbuz, D.S. Jessop, C.D. Ingram, S.L. Lightman, Early-life exposure to endotoxin alters hypothalamic-pituitary-adrenal function and predisposition to inflammation, Proc Natl Acad Sci U S A 97 (2000) 5645-5650.
[20] L. Soto, A. Martín, S. Millán, E. Vara, A. López-Calderón, Effects of endotoxin lipopolysaccharide administration on the somatotropic axis, J Endocrinol. 159 (1998) 239-246.
[21] S.J. Spencer, J.G. Heida, Q.J. Pittman, Early life immune challenge - effects on behavioural indices of adult rat fear and anxiety, Behav. Br. Res. 164 (2005) 231-238.
[22] A. Sundstedt, M. Dohlsten, G. Hedlund, I. Hoiden, M. Bjorklund, T. Kalland, Superantigens anergize cytokine production but not cytotoxicity in vivo, Immunology 82 (1994) 117-125.
[23] R.I. Tapping, S. Akashi, K. Miyake, P.J. Godowski, P.S. Tobias, Toll-Like Receptor 4, But Not Toll-Like Receptor 2, Is a Signaling Receptor for Escherichia and Salmonella Lipopolysaccharides, J. Immunol. 165 (2000) 5780-5787.
[24] F. Toma Sacarea, Bacteriologie medicala, Ed. Universitatii Oradea, 2006. [25] S. Wang, J.Y. Yan, Y.K. Lo, P.M. Carvey, Z. Ling, Dopaminergic and serotoninergic deficiencies in
young adult rats prenatally exposed to the bacterial lipopolysaccharide, Brain Res. 1265 (2009) 196-204.
[26] J.J. Yagi, S. Rath, C.A. Janeway Jr., Control of T cell responses to staphylococcal enterotoxins by stimulator cell MHC class II polymorphism, J. Immunol. 147 (1991) 1398-1405.





























