Embed Size (px)
Citation preview

INDUKSI PERTUNASAN IN VITRO PADA JARINGAN
PUCUK APIKAL TANAMAN KURMA
(Phoenix dactylifera L.) cv. Barhee
HANINDYA WIDYAWATI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2015


PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Induksi Pertunasan In
vitro Pada Jaringan Pucuk Apikal Tanaman Kurma (Phoenix dactylifera L.) cv.
Barhee adalah benar karya saya dengan arahan dari komisi pembimbing dan belum
diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber
informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak
diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam
daftar pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, Mei 2015
Hanindya Widyawati
NIM G34100097

ABSTRAK
HANINDYA WIDYAWATI. Induksi Pertunasan In vitro Pada Jaringan Pucuk
Apikal Tanaman Kurma (Phoenix dactylifera L.) cv. Barhee. Dibimbing oleh DIAH
RATNADEWI dan ENCE DARMO JAYA SUPENA.
Kurma (Phoenix dactylifera L.) memiliki potensi untuk dibudidayakan di
Indonesia. Budidaya kurma secara komersial membutuhkan bibit tanaman dalam
jumlah yang besar. Teknik kultur jaringan dapat diharapkan untuk memproduksi
bibit tanaman kurma secara masal dalam waktu yang relatif singkat. Tujuan
penelitian ini ialah mengetahui konsentrasi BAP (Benzyl amino purin) yang dapat
menginduksi pertunasan pada eksplan kurma dan menentukan metode sterilisasi
eksplan yang efektif untuk jaringan pucuk apikal tanaman kurma. Eksplan yang
digunakan adalah pucuk apikal (1- 2 cm) yang dibelah empat secara vertikal. Tahap
pertama ialah menentukan metode sterilisasi eksplan yang efektif dari tiga metode
sterilisasi yang dicobakan. Tahap kedua ialah penanaman eksplan pada media
Murashige dan Skoog dengan lima perlakuan sitokinin BAP (0, 1, 2, 4, 6 mg/L).
Hasil penelitian menunjukkan media MS dengan BAP pada rentang konsentrasi 0
sampai 6 mg/L belum mampu menginduksi pertunasan pada jaringan tanaman
kurma kultivar Barhee ini. Metode sterilisasi 3, yaitu menggunakan Dithane M-45
0.1%, Benlate 0.1%, Agrept 0.2%, Bayclin 20%, Bayclin 10%, dan etanol 70%,
lebih baik dibandingkan metode sterilisasi lainnya.
Kata kunci: Kurma, kultur jaringan, tunas
ABSTRACT
HANINDYA WIDYAWATI. In vitro Shoot Induction on Explant of Date Palm
(Phoenix dactylifera L.) cv. Barhee. Supervised by DIAH RATNADEWI and
ENCE DARMO JAYA SUPENA.
Date palm (Phoenix dactylifera L.) has the potential to be cultivated in
Indonesia. Commercial cultivation of date palm requires large amounts of seed.
Tissue culture techniques can be used as a solution to produce date palm seedling
rapidly and efficiently. The purpose of this study was to determine the concentration
of BAP (Benzyl Amino Purin) which may induce shoots on dates explant and
determined the most effective method of sterilization of explants. The explant used
was apical shoot (1- 2 cm) that was then quartered vertically. The first stage was to
determine the effective method of sterilization of explants using three methods of
sterilization. The second stage was the cultivation of explants on Murashige and
Skoog media with five treatments of cytokinin BAP (0, 1, 2, 4, 6 mg/L). The results
showed that MS medium with BAP in the concentrations ranged 0 to 6 mg/L had
not been able to induce any new growth on the plant tissue of date palm cultivar
Barhee. The sterilization method 3, which used 20% Bayclin, 10% Bayclin, 0.1%
Agrept, 0.1% Dithane M-45, and 70% ethanol was better than the other sterilization
methods.
Keywords: Date palm, shoot, tissue culture

Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
INDUKSI PERTUNASAN IN VITRO PADA JARINGAN
PUCUK APIKAL TANAMAN KURMA
(Phoenix dactylifera L.) cv. Barhee
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2015
HANINDYA WIDYAWATI




PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas
segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian yang
dilaksanakan sejak bulan Desember 2013 sampai September 2014 ini ialah Induksi
Pertunasan In vitro Pada Jaringan Pucuk Apikal Tanaman Kurma (Phoenix
dactylifera L.) cv. Barhee.
Terima kasih penulis ucapkan kepada Ibu Prof Dr Ir Diah Ratnadewi, DEA.
dan Bapak Dr Ir Ence Darmo Jaya Supena, MSi. selaku pembimbing yang telah
memberikan bimbingan, saran, dan ilmu yang bermanfaat selama melaksanakan
penelitian dan penulisan karya ilmiah. Terima kasih kepada Ibu Dr Nisa Rachmania
Mubarik, MSi selaku penguji. Terima kasih penulis ucapkan kepada kedua orang
tua Ibu Ike dan Bapak Sukono tercinta, dan adik Haryo Satriaji, tante Wisye, tante
Bery serta seluruh keluarga besar yang telah memberikan dukungan, doa, semangat
dan bantuannya selama melaksanakan penelitian dan penulisan karya ilmiah ini.
Penulis juga mengucapkan terima kasih kepada Ibu Ucu, Ibu Dewi, Pak Asep, Pak
Jaka selaku laboran yang telah banyak memberikan bantuan selama pengamatan di
laboratorium, terima kasih kepada Yurika, Nita, Efah, Feni, Nurlatiefah, Ina, Catur
Putri, Naili, Kak Cut, Kak Sasa, serta seluruh teman seperjuangan di Biologi 47 atas
segala dukungan dan kebersamaan selama ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Mei 2015
Hanindya Widyawati

DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
BAHAN dan METODE 2
Waktu dan Tempat 2
Bahan dan Alat 2
Metode Sterilisasi Eksplan Kurma 3
Inisiasi dan Induksi Tunas Kurma 3
HASIL 4
Efektivitas Metode Sterilisasi 4
Tahap Induksi dan Inisiasi Tunas Kurma 4
PEMBAHASAN 6
SIMPULAN 7
DAFTAR PUSTAKA 8
LAMPIRAN 10
RIWAYAT HIDUP 12

DAFTAR TABEL
1 Perbandingan prosedur sterilisasi pada tiap metode sterilisasi 3 2 Perbandingan efektivitas metode sterilisasi eksplan kurma
berdasarkan tingkat kontaminasi 4 3 Data kualitatif penampilan, pertumbuhan dan perkembangan
kultur kurma pada 16 MST 5
4 Pengaruh perlakuan terhadap kondisi kultur kurma pada 16 MST 6


PENDAHULUAN
Latar Belakang
Kurma (Phoenix dactylifera L.) merupakan salah satu anggota keluarga
Arecaceae yang memiliki potensi tinggi untuk dikembangkan di Indonesia karena
beriklim tropis dan selalu mendapatkan sinar matahari sepanjang tahun. Tanaman
ini memiliki peranan penting dalam kehidupan ekonomi dan sosial masyarakat di
daerah kering dan semi-kering di dunia (Mahmoudi et al. 2008).
Hampir setiap bagian dari kurma cv. Barhee memiliki manfaat kecuali
bagian akar, buahnya dapat dikonsumsi karena kandungan karbohidrat sebesar 70-
80%, protein 1-3%, Vitamin A, Vitamin B1, dan Vitamin B2. Batang pohon kurma
dapat digunakan sebagai bahan bangunan, daun dan tangkai daunnya dimanfaatkan
sebagai sumber pulp selulosa, bijinya dapat diolah menjadi pakan ternak
(Mahmoudi et al. 2008).
Permintaan akan kurma semakin meningkat untuk konsumsi pangan bukan
saja di bulan Ramadhan. Buah kurma menjadi mata dagang ekspor di pasaran
internasional. Berdasarkan data Badan Pusat Statistik (BPS 2013), total impor
kurma di Indonesia pada tahun 2012 mencapai 16.200 ton dengan nilai US$ 20 juta.
Barhee merupakan kultivar unggulan yang banyak dicari di pasaran internasional,
serta buahnya dapat dikonsumsi pada tahap semi-matang. Pada tahap ini, buahnya
berwarna kuning keemasan dengan tekstur renyah dan rasa yang manis (Fki et al.
2011).
Budidaya kurma secara komersial memerlukan bibit dalam jumlah yang
besar. Perluasan budidaya tanaman kurma secara tradisional masih dibatasi oleh
kemampuan tanaman untuk menghasilkan bibit baru dalam jumlah banyak,
seragam, dan dalam waktu singkat. Teknik kultur jaringan dapat diharapkan untuk
memproduksi bibit secara masal dalam waktu yang relatif singkat. Teknik kultur
jaringan memerlukan bahan eksplan berupa bagian-bagian tanaman karena sel-sel
tanaman memiliki sifat totipoten. Totipotensi merupakan kemampuan sel tumbuhan
yang telah berdiferensiasi untuk menjadi sel embrionik kembali, kemudian
berkembang menjadi tumbuhan baru yang lengkap (Salisbury dan Ross 1995).
Bahan eksplan yang paling baik digunakan ialah yang memiliki sifat meristematik.
Bahan tanaman kurma yang dapat dijadikan eksplan berupa tunas, batang muda,
daun muda, dan bunga (Khan dan Bibi 2012). Perbanyakan tanaman secara in vitro
diawali dengan memperoleh bahan tanaman aseptik yang akan digunakan untuk
perbanyakan bibit. Oleh karena itu, diperlukan proses sterilisasi yang tepat untuk
mematikan mikroorganisme yang terdapat pada eksplan sehingga tidak
mengganggu pertumbuhan kultur.
Penggunaan zat pengatur tumbuh dalam kultur jaringan tanaman sangat
penting, yaitu untuk merangsang pertumbuhan tunas, akar, kalus, atau
embriogenesis somatik. Penggunaan zat pengatur tumbuh di dalam kultur jaringan
tergantung pada tujuan atau arah pertumbuhan tanaman yang diinginkan (Zaer dan
Mapes 1982). Menurut Lestari (2011), penggunaan auksin dan sitokinin pada
konsentrasi yang tepat, akan memacu organogenesis dalam pembentukan tunas.
Benzyl Amino Purin (BAP) merupakan salah satu zat pengatur tumbuh yang biasa
digunakan untuk memacu pembentukan tunas dengan daya aktivitas yang kuat

2
untuk mendorong proses pembelahan sel (George dan Sherrington 1984). Oleh
karena itu, diperlukan penelitian untuk mengetahui konsentrasi BAP yang tepat
untuk menginduksi pertunasan pada kurma.
Tisserat dan DeMason (1980) berhasil mengkulturkan tanaman kurma
melalui kultur jaringan. Setelah itu teknik perbanyakan tanaman kurma melalui
kultur jaringan semakin berkembang; keberhasilan didapat melalui teknik
organogenesis (Sharma et al. 1980, Hegazy dan Aboshama 2010) dan
embriogenesis somatik (Al-Khateeb 2008). Salah satu varietas kurma yang berhasil
dikulturkan melalui organogenesis langsung adalah kultivar Barhee (Zaid dan Wet
2002). Zaid dan Wet (2002) melakukan induksi tunas kurma cv. Barhee
menggunakan pucuk apikal yang ditanam dalam media MS (Murashige dan skoog)
yang diberi BAP 2 mg/L dan 2,4-D 1 mg/L. Sharma et al. (1980) telah berhasil
menumbuhkan tunas kurma melalui organogenesis langsung, dengan menggunakan
eksplan pucuk apikal kurma cv. Khalas pada media MS yang dilengkapi NAA 1
mg/L, BAP 3 mg/L, dan 2-iP 3 mg/L.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui konsentrasi BAP yang dapat
menginduksi pertunasan pada kurma cv. Barhee, dan mencari metode sterilisasi
eksplan yang efektif agar dapat digunakan sebagai acuan dalam melakukan
sterilisasi eksplan secara tepat untuk tanaman kurma.
BAHAN dan METODE
Waktu dan Tempat
Penelitian ini dilaksanakan mulai bulan Desember 2013 hingga September
2014 di Laboratorium Penelitian Kultur Jaringan Tanaman, Departemen Biologi,
FMIPA, Institut Pertanian Bogor.
Bahan dan Alat
Bahan tanaman yang digunakan yaitu tanaman kurma cv. Barhee umur dua
tahun (tanaman muda) dari Jonggol Farm, Bogor. Zat pengatur tumbuh yang
digunakan berupa BAP (Benzyl Amino Purin) dan NAA (Naphtalene Acetic Acid).
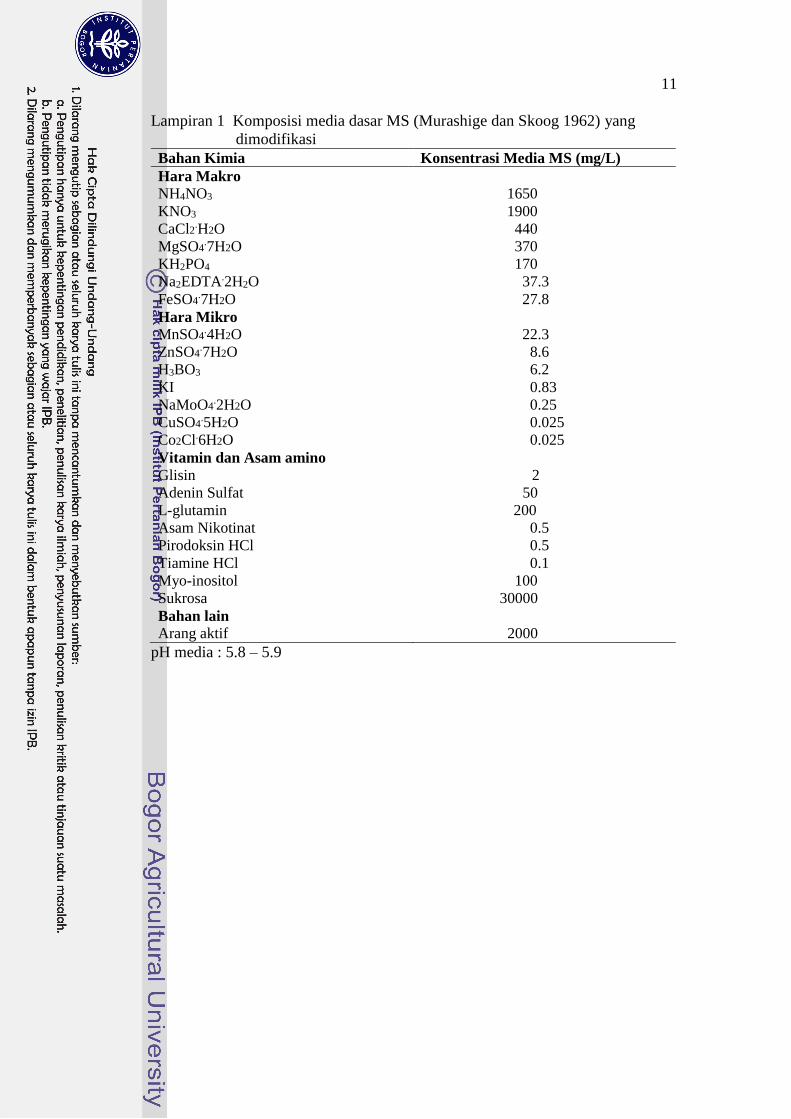
Media yang dipakai ialah Murashige dan Skoog (MS) dengan penambahan asam
amino L-glutamin dan Adenin Sulfat (Lampiran 1).
Bahan sterilisasi eksplan yang digunakan berupa larutan fungisida Dithane
M-45 (bahan aktif Mankozeb 80%) dan Benlate (bahan aktif Benomyl 50%),
larutan bakterisida (Agrept), etanol 70%, larutan Bayclin (bahan aktif NaOCl
5.25%), HgCl2, tween 80 serta akuades steril sebagai bahan pembilas. Bahan kimia
lain yang digunakan berupa arang aktif.
Alat yang digunakan antara lain autoklaf, timbangan, pH meter, laminar air
flow cabinet (LAFC), serta alat-alat diseksi.
3
Metode Sterilisasi Eksplan Kurma
Pada penelitian ini digunakan tiga macam metode sterilisasi yang tersaji
dalam Tabel 1. Mula-mula batang tanaman sepanjang ± 5 cm dibuang pelepah daun
dan akarnya, kemudian diisolasi bagian pucuk apikalnya, lalu dicuci dengan sabun
dan dibilas dengan air keran. Bahan tanaman yang telah dibilas kemudian
disterilisasi melalui berbagai tahap perendaman dalam bahan sterilan. Setiap
perpindahan tahap, bahan tanaman dibilas tiga kali dengan akuades steril.
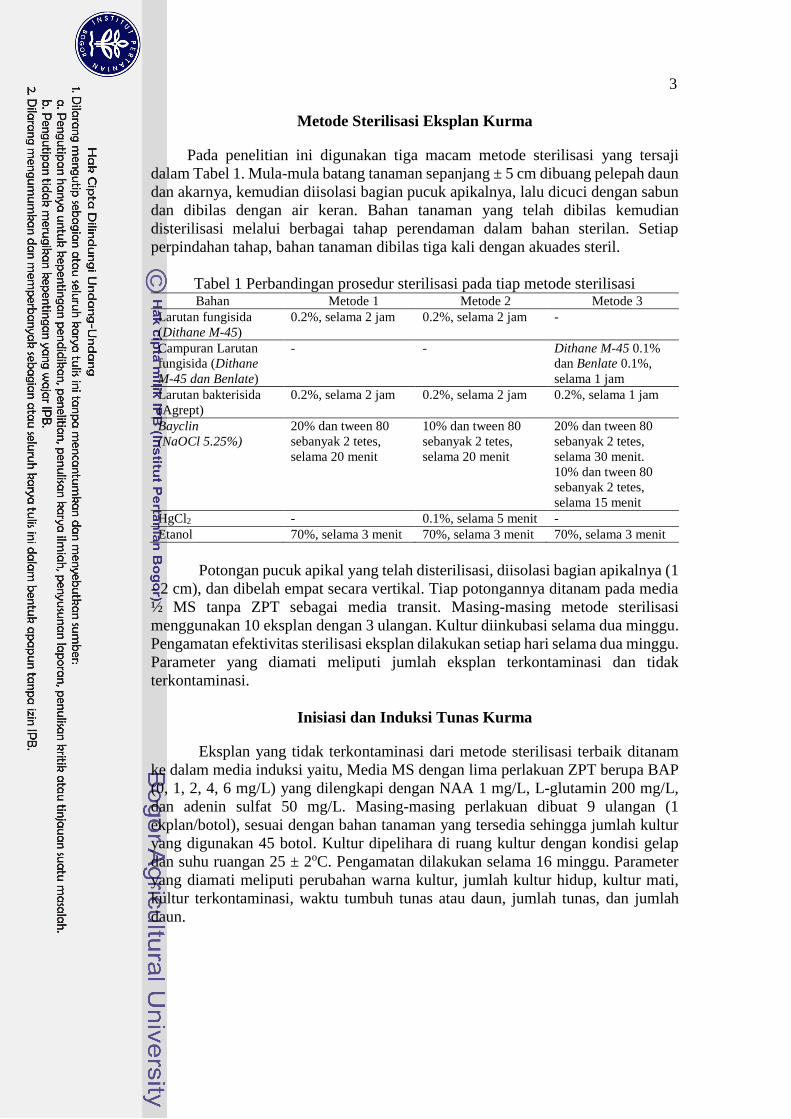
Tabel 1 Perbandingan prosedur sterilisasi pada tiap metode sterilisasi Bahan Metode 1 Metode 2 Metode 3
Larutan fungisida
(Dithane M-45)
0.2%, selama 2 jam 0.2%, selama 2 jam -
Campuran Larutan
fungisida (Dithane
M-45 dan Benlate)
- - Dithane M-45 0.1%
dan Benlate 0.1%,
selama 1 jam
Larutan bakterisida
(Agrept)
0.2%, selama 2 jam 0.2%, selama 2 jam 0.2%, selama 1 jam
Bayclin
(NaOCl 5.25%)
20% dan tween 80
sebanyak 2 tetes,
selama 20 menit
10% dan tween 80
sebanyak 2 tetes,
selama 20 menit
20% dan tween 80
sebanyak 2 tetes,
selama 30 menit.
10% dan tween 80
sebanyak 2 tetes,
selama 15 menit
HgCl2 - 0.1%, selama 5 menit -
Etanol 70%, selama 3 menit 70%, selama 3 menit 70%, selama 3 menit
Potongan pucuk apikal yang telah disterilisasi, diisolasi bagian apikalnya (1
- 2 cm), dan dibelah empat secara vertikal. Tiap potongannya ditanam pada media
½ MS tanpa ZPT sebagai media transit. Masing-masing metode sterilisasi
menggunakan 10 eksplan dengan 3 ulangan. Kultur diinkubasi selama dua minggu.
Pengamatan efektivitas sterilisasi eksplan dilakukan setiap hari selama dua minggu.
Parameter yang diamati meliputi jumlah eksplan terkontaminasi dan tidak
terkontaminasi.
Inisiasi dan Induksi Tunas Kurma
Eksplan yang tidak terkontaminasi dari metode sterilisasi terbaik ditanam
ke dalam media induksi yaitu, Media MS dengan lima perlakuan ZPT berupa BAP
(0, 1, 2, 4, 6 mg/L) yang dilengkapi dengan NAA 1 mg/L, L-glutamin 200 mg/L,
dan adenin sulfat 50 mg/L. Masing-masing perlakuan dibuat 9 ulangan (1
ekplan/botol), sesuai dengan bahan tanaman yang tersedia sehingga jumlah kultur
yang digunakan 45 botol. Kultur dipelihara di ruang kultur dengan kondisi gelap
dan suhu ruangan 25 ± 2oC. Pengamatan dilakukan selama 16 minggu. Parameter
yang diamati meliputi perubahan warna kultur, jumlah kultur hidup, kultur mati,
kultur terkontaminasi, waktu tumbuh tunas atau daun, jumlah tunas, dan jumlah
daun.
4
HASIL
Efektivitas Metode Sterilisasi
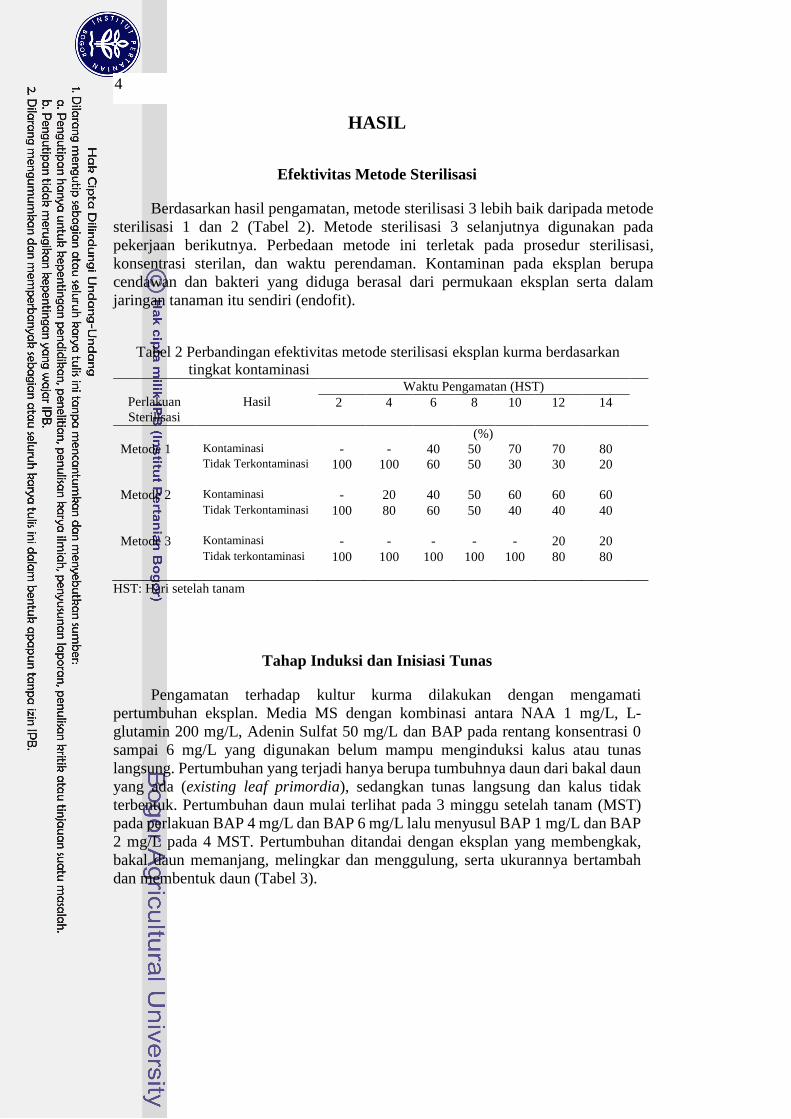
Berdasarkan hasil pengamatan, metode sterilisasi 3 lebih baik daripada metode
sterilisasi 1 dan 2 (Tabel 2). Metode sterilisasi 3 selanjutnya digunakan pada
pekerjaan berikutnya. Perbedaan metode ini terletak pada prosedur sterilisasi,
konsentrasi sterilan, dan waktu perendaman. Kontaminan pada eksplan berupa
cendawan dan bakteri yang diduga berasal dari permukaan eksplan serta dalam
jaringan tanaman itu sendiri (endofit).
Tabel 2 Perbandingan efektivitas metode sterilisasi eksplan kurma berdasarkan
tingkat kontaminasi
Perlakuan
Sterilisasi
Hasil
Waktu Pengamatan (HST)
2
4
6
8
10
12
14
(%)
Metode 1 Kontaminasi - - 40 50 70 70 80
Tidak Terkontaminasi 100 100 60 50 30 30 20
Metode 2 Kontaminasi - 20 40 50 60 60 60
Tidak Terkontaminasi 100 80 60 50 40 40 40
Metode 3 Kontaminasi - - - - - 20 20
Tidak terkontaminasi 100 100 100 100 100 80 80
HST: Hari setelah tanam
Tahap Induksi dan Inisiasi Tunas
Pengamatan terhadap kultur kurma dilakukan dengan mengamati
pertumbuhan eksplan. Media MS dengan kombinasi antara NAA 1 mg/L, L-
glutamin 200 mg/L, Adenin Sulfat 50 mg/L dan BAP pada rentang konsentrasi 0
sampai 6 mg/L yang digunakan belum mampu menginduksi kalus atau tunas
langsung. Pertumbuhan yang terjadi hanya berupa tumbuhnya daun dari bakal daun
yang ada (existing leaf primordia), sedangkan tunas langsung dan kalus tidak
terbentuk. Pertumbuhan daun mulai terlihat pada 3 minggu setelah tanam (MST)
pada perlakuan BAP 4 mg/L dan BAP 6 mg/L lalu menyusul BAP 1 mg/L dan BAP
2 mg/L pada 4 MST. Pertumbuhan ditandai dengan eksplan yang membengkak,
bakal daun memanjang, melingkar dan menggulung, serta ukurannya bertambah
dan membentuk daun (Tabel 3).
5
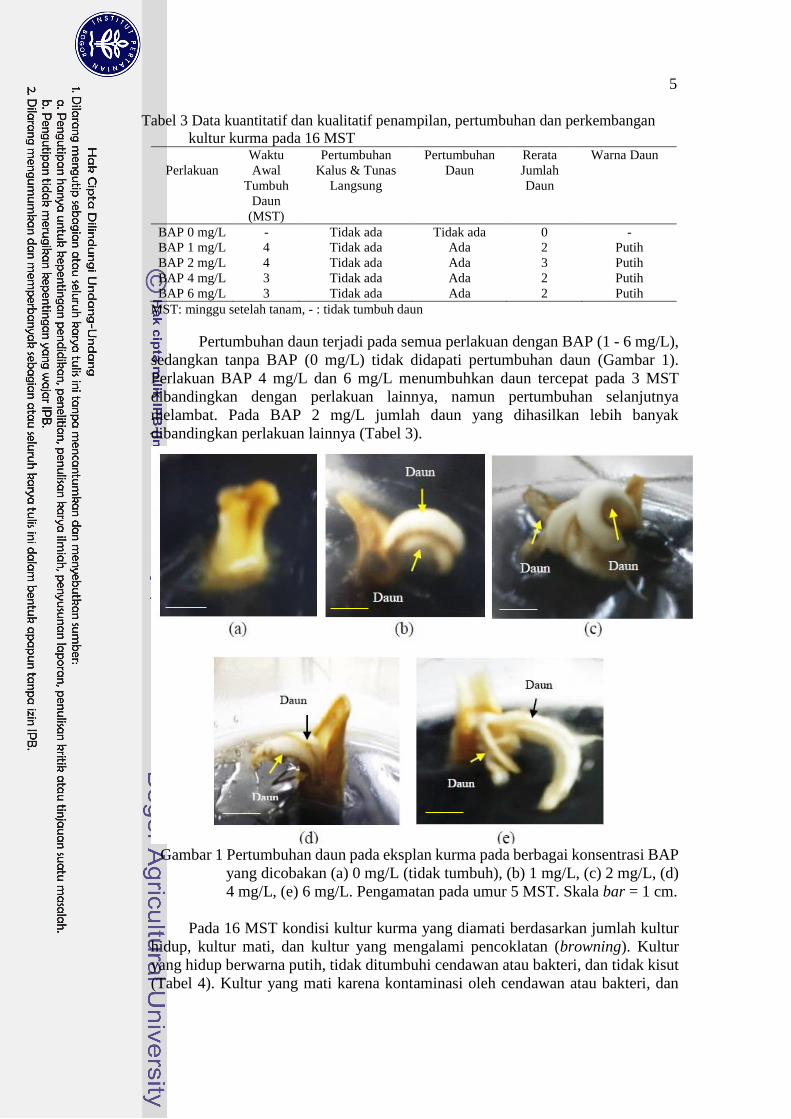
Tabel 3 Data kuantitatif dan kualitatif penampilan, pertumbuhan dan perkembangan
kultur kurma pada 16 MST
Perlakuan
Waktu
Awal
Tumbuh
Daun
(MST)
Pertumbuhan
Kalus & Tunas
Langsung
Pertumbuhan
Daun
Rerata
Jumlah
Daun
Warna Daun
BAP 0 mg/L - Tidak ada Tidak ada 0 -
BAP 1 mg/L 4 Tidak ada Ada 2 Putih
BAP 2 mg/L 4 Tidak ada Ada 3 Putih
BAP 4 mg/L 3 Tidak ada Ada 2 Putih
BAP 6 mg/L 3 Tidak ada Ada 2 Putih
MST: minggu setelah tanam, - : tidak tumbuh daun
Pertumbuhan daun terjadi pada semua perlakuan dengan BAP (1 - 6 mg/L),
sedangkan tanpa BAP (0 mg/L) tidak didapati pertumbuhan daun (Gambar 1).
Perlakuan BAP 4 mg/L dan 6 mg/L menumbuhkan daun tercepat pada 3 MST
dibandingkan dengan perlakuan lainnya, namun pertumbuhan selanjutnya
melambat. Pada BAP 2 mg/L jumlah daun yang dihasilkan lebih banyak
dibandingkan perlakuan lainnya (Tabel 3).
Gambar 1 Pertumbuhan daun pada eksplan kurma pada berbagai konsentrasi BAP
yang dicobakan (a) 0 mg/L (tidak tumbuh), (b) 1 mg/L, (c) 2 mg/L, (d)
4 mg/L, (e) 6 mg/L. Pengamatan pada umur 5 MST. Skala bar = 1 cm.
Pada 16 MST kondisi kultur kurma yang diamati berdasarkan jumlah kultur
hidup, kultur mati, dan kultur yang mengalami pencoklatan (browning). Kultur
yang hidup berwarna putih, tidak ditumbuhi cendawan atau bakteri, dan tidak kisut
(Tabel 4). Kultur yang mati karena kontaminasi oleh cendawan atau bakteri, dan
6
lama kelamaan terlihat membusuk. Kultur yang mengalami pencoklatan (browning),
eksplan mencoklat mulai dari bagian ujung eksplan setelah pemotongan. Meskipun
kultur ini mencoklat, masih terlihat pertumbuhan berupa respons bagian meristem
apikal yang membesar namun pertumbuhannya lambat.
Tabel 4 Pengaruh perlakuan terhadap kondisi kultur kurma pada 16 MST
Perlakuan Hidup Mati Mencoklat Kultur Hidup (%)
BAP 0 mg/L 7 2 - 77.8
BAP 1 mg/L 4 4 1 44.4
BAP 2 mg/L 4 5 - 44.4
BAP 4 mg/L 4 4 1 44.4
BAP 6 mg/L 5 4 - 55.6
Pada pengamatan minggu ke-16 persentase kultur hidup yang didapatkan
melalui metode sterilisasi 3, berkisar antara 44.4% (BAP 1, 2, 4 mg/L) hingga
77.8% (BAP 0 mg/L) (Tabel 4). Pada BAP 0 mg/L kultur hidup, namun tidak ada
pertumbuhan tunas langsung maupun daun. Hal ini diduga karena pemberian auksin,
L-glutamin, adenin sulfat saja tanpa sitokinin tampaknya tidak mampu merangsang
pertumbuhan tunas maupun daun.
PEMBAHASAN
Bahan tanaman yang dijadikan eksplan harus dalam keadaan steril. Oleh
karena itu, sterilisasi eksplan merupakan tahap terpenting dalam kultur jaringan
tanaman terutama bila sumber eksplan berasal dari lapang. Sterilisasi bertujuan
mematikan bakteri dan cendawan yang berada di permukaan. Setiap tanaman
mempunyai respons spesifik terhadap bahan sterilan. Berdasarkan dosis bahan
sterilisasi dan waktu perendaman eksplan, metode sterilisasi 3 lebih baik, karena
menghasilkan kontaminasi yang rendah (20%) jika dibandingkan dengan metode
sterilisasi 1 dan 2 yang menghasilkan kontaminasi tinggi (80% dan 60%). Metode
sterilisasi 3 menghasilkan kontaminasi yang rendah karena penggunaan campuran
2 jenis larutan fungisida (Dithane M-45 0.1% dan Benlate 0.1%), serta Bayclin
dengan dosis bertingkat (20% dan 10%).
Dosis sterilan dan waktu perendaman eksplan bergantung pada dua hal, yaitu
ukuran eksplan dan jenis tanaman seperti tanaman herba atau berkayu. Menurut
Zulkarnain (2009) semakin besar ukuran eksplan, maka akan semakin besar
peluang terkontaminasi baik secara internal maupun eksternal, tetapi kemungkinan
keberhasilan proliferasi semakin besar. Aisyah dan Surachman (2011) menyatakan
bahwa keberhasilan sterilisasi juga dipengaruhi oleh kondisi lingkungan (musim
hujan atau kemarau). Pengambilan bahan tanaman saat musim hujan menyebabkan
tingkat kontaminasi yang tinggi karena terjadi kenaikan kelembapan tanah, dan
kelebihan air cenderung mendukung pertumbuhan jamur atau bakteri secara cepat
pada lingkungan tumbuh tempat pengambilan tanaman.
Kontaminan yang muncul pada eksplan terdiri atas cendawan dan bakteri.
Kontaminan yang berasal dari cendawan lebih banyak dan sangat merugikan
eksplan hingga mengalami kematian. Kontaminasi oleh cendawan dicirikan dengan
adanya hifa putih yang tumbuh pada media kultur dari eksplan. Kondisi media
kultur yang lembab dan banyak mengandung nutrisi menyebabkan pertumbuhan

7
cendawan lebih cepat daripada pertumbuhan eksplannya. Cendawan yang
menyerang eksplan lama-kelamaan menutupi eksplan, yang akhirnya menyebabkan
kematian eksplan. Kontaminasi cendawan diduga berasal dari permukaan eksplan
dan dalam jaringan tanaman itu sendiri (endofit). eksplan. Cendawan endofit adalah
cendawan yang mengoloni jaringan tumbuhan sehat tanpa menimbulkan gejala
penyakit (Ramdan et al. 2013). Cendawan endofit hidup dalam jaringan tumbuhan
pada periode tertentu dan mampu membentuk koloni dalam jaringan tumbuhan.
Cendawan endofit yang berasosiasi dengan tanaman kurma antara lain Bauveria
bassiana, Lecanicillium dimorphum dan Lecanicillium c.f. psalliotae (Gomez et al.
2006).
Kontaminasi bakteri pada kultur dicirikan dengan adanya lendir berwarna
putih agak bening pada bagian pangkal eksplan hingga menyebar ke media tanam.
Kontaminasi bakteri diduga berasal dari permukaan eksplan dan dalam jaringan
tanaman itu sendiri (endofit). Bakteri endofit adalah bakteri yang hidup
bersimbiosis mutualis dengan jaringan tanaman. Pada kultur in vitro, bakteri endofit
bersifat merugikan karena dapat menghambat pertumbuhan eksplan (Malfanova
2013). Jika di dalam eksplan terdapat bakteri endofit, maka bakteri tersebut akan
keluar ke media tumbuh dan bersaing dengan eksplan untuk memenuhi kebutuhan
nutrisinya. Eksplan yang tidak dapat bersaing akan didominasi oleh pertumbuhan
bakteri endofit hingga mengakibatkan kematian pada kultur. Keberadaan bakteri
endofit menyebabkan eksplan tidak dapat dibersihkan dengan sterilisasi
permukaan. Pemberian sterilan dan waktu perendaman pada penelitian ini tidak
dapat mematikan bakteri yang berada di dalam jaringan eksplan, meskipun sampai
telah menyebabkan klorosis pada eksplan.
Media MS dengan BAP pada rentang konsentrasi 0 sampai 6 mg/L, yang
dilengkapi dengan NAA 1 mg/L, L-glutamin 200 mg/L, dan adenin sulfat 50 mg/L
belum mampu untuk menginduksi tunas langsung atau kalus pada kultur kurma.
Induksi pertumbuhan daun ditandai dengan eksplan yang membengkak, bakal daun
memanjang, melingkar dan menggulung, ukurannya bertambah dan membentuk
daun. Hal ini diduga karena kombinasi dosis antara BAP, NAA, L-glutamin dan
adenin sulfat belum tepat. Pada perlakuan BAP 4 mg/L dan 6 mg/L pertumbuhan
daun menurun. Hal ini diduga pemberian BAP dengan konsentrasi 4 mg/L – 6 mg/L
terlalu tinggi bagi eksplan sehingga pertumbuhan daun menjadi terhambat. Pada
kontrol (BAP 0 mg/L) tidak didapati pertumbuhan eksplan karena nampaknya
pemberian auksin saja tanpa sitokinin tidak mampu merangsang pertumbuhan tunas
langsung maupun daun. Perlakuan BAP 4 mg/L dan BAP 6 mg/L menumbuhkan
daun pada 3 MST, lebih cepat dibandingkan dengan perlakuan lainnya, namun
pertumbuhan selanjutnya melambat. Hal ini sesuai dengan hasil penelitian Junaid
dan Saeed (2009) yang menyatakan bahwa laju pertumbuhan tunas dan daun kurma
kultivar Khalas akan berkurang ketika konsentrasi BAP lebih dari 3 mg/L.
SIMPULAN
Media MS dengan kombinasi BAP pada rentang konsentrasi 0 sampai 6 mg/L,
dengan NAA 1 mg/L, L-glutamin 200 mg/L, dan adenin sulfat 50 mg/L belum
mampu menginduksi pertumbuhan tunas pada kurma kultivar Barhee. Metode
sterilisasi 3 yang menggunakan campuran 2 jenis larutan fungisida (Dithane M-45

8
0.1% dan Benlate 0.1%), dan Bayclin dengan dosis bertingkat (20% dan 10%) lebih
efektif, karena tingkat kontaminasinya rendah (20%) dibandingkan dengan metode
sterilisasi 1 dan 2 (80% dan 60%).
DAFTAR PUSTAKA
Aisyah S, Surachman D. 2011. Teknik sterilisasi rimpang jahe sebagai bahan
perbanyakan tanaman jahe sehat secara in vitro. Bul Teknik Pertan. 16(1):34-
36.
Al-Khateeb A. 2008. Comparison effects of sucrose and date palm syrup on
somatic embryogenesis of date palm (Phoenix dactylifera L.). Am J Biochem
Biotechnol. 4(1):19-23.
[BPS] Badan Pusat Statistik. 2013. Data Ekspor dan Impor Buah Tahun 2012.
Jakarta (ID): BPS.
Fki L, Bouaziz N, Kriaa W, Masmoudi BR, Bouzid GR, Alain R, Drira N. 2011.
Multiple bud culture of ‘Barhee’ date palm (Phoenix dactylifera L.) and
physiological status of regenerated plants. J Plant Physiol. 168:1694-1700.
George EF, Sherrington PD. 1984. Plant Propagation by Tissue Culture, Handbook
and Directory of Comercial Laboratories. Basingstoke (GB): Easter Pr.
Gomez VS, Lopez LV, Jansson HB, Salinas J. 2006. Endophytic colonization of
date palm (Phoenix dactylifera L.) leaves by entomopathogenic fungi. J
Micron. 37:624-632.
Hegazy AE, Aboshama HM. 2010. An efficient novel pathway discovered in date
palm micropropagation. Acta Hort. 882: 167-176.
Junaid A, Saeed AK. 2009. In vitro micropropagation of ‘Khalas’ date palm
(Phoenix dactylifera L.), an important fruit Plant. J Fruit Orna Plant Res.
17:15-27.
Khan S, Bibi T. 2012. Direct shoot regeneratif system for date palm (Phoenix
dactylifera L.) cv. Dhakki as a means of micropropagation. J Bot. 44(6):
1965-1971.
Lestari GE. 2011. Peranan zat pengatur tumbuh dalam perbanyakan tanaman
melalui kultur jaringan. Jur Agro Biogen. 7(1): 63-68.
Mahmoudi H, Hosseininia G, Azadi H, Fatemi M. 2008. Enchanting date palm
processing, marketing and set control through organic culture. J Organic
Systems. 3(2) : 29-39
Malfanova NV. 2013. Endophytic Bacteria with Plant Growth Promoting and
Biocontrol Abilities. Leiden (NL): Leiden University.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bio assays
with tobacco tissue culture. Physiol Plant. 15: 473.
Ramdan EP, Widodo, Tondok ET, Wiyono S, Hidayat SH. 2013. Cendawan endofit
nonpatogen asal tanaman cabai dan potensinya sebagai agens pemacu
pertumbuhan. J Fitopatol Indones 9(5): 139-144.
Salisbury FB, Ross CW. 1995. Fisiologi Tumbuhan Jilid 3. Lukman DR,
Sumaryono, penerjemah; Niksolihin S, editor. Bandung (ID): ITB Press.
Terjemahan dari: Plant Physiology. Ed ke-4.
Sharma DR, Kumari R, Chowdhury JB. 1980. In vitro culture of female date palm
(Phoenix dactylifera L.) tissues. Euphytica. 29: 169-174.

9
Tisserat B, DeMason DA. 1980. A histological study of the development of
adventive embryos in organ culture of Phoenix dactylifera L. Ann Bot. 45:
465-472.
Zaid A, Wet DPF. 2002. Date palm propagation. J Date Palm 2: 73-79.
Zaer JB, Mapes MO. 1982. Action of growth regeneration. Bul Tissue Cultu Forest.
1 : 231-235.
Zulkarnain H. 2009. Kultur Jaringan Tanaman sebagai Solusi Perbanyakan
Tanaman. Jakarta (ID): Bumi Aksara.

10
LAMPIRAN
11
Lampiran 1 Komposisi media dasar MS (Murashige dan Skoog 1962) yang
dimodifikasi
Bahan Kimia Konsentrasi Media MS (mg/L)
Hara Makro NH4NO3 1650
KNO3 1900
CaCl2.H2O 440
MgSO4.7H2O 370
KH2PO4 170
Na2EDTA.2H2O 37.3
FeSO4.7H2O 27.8
Hara Mikro MnSO4
.4H2O 22.3
ZnSO4.7H2O 8.6
H3BO3 6.2
KI 0.83
NaMoO4.2H2O 0.25
CuSO4.5H2O 0.025
Co2Cl.6H2O 0.025
Vitamin dan Asam amino Glisin 2
Adenin Sulfat 50
L-glutamin 200
Asam Nikotinat 0.5
Pirodoksin HCl 0.5
Tiamine HCl 0.1
Myo-inositol 100
Sukrosa 30000
Bahan lain
Arang aktif 2000
pH media : 5.8 – 5.9

12
RIWAYAT HIDUP
Penulis dilahirkan di Tangerang pada tanggal 25 September 1992 dari
pasangan Sukono, ST dan Sri Areka Dhayaningtias. Penulis merupakan anak
pertama dari dua bersaudara. Penulis menyelesaikan pendidikan TK di Bhakti
Atomita Batan Indah pada tahun 1997, menyelesaikan pendidikan dasar di SD
Negeri Batan Indah pada tahun 2004, dan menyelesaikan pendidikan lanjutan
menengah pertama di SMP Negeri 8 Tangerang Selatan, Puspiptek pada tahun 2007.
Penulis menyelesaikan pendidikan menengah atas di SMA Negeri 2 Tangerang
Selatan dan lulus pada tahun 2010. Melalui jalur Ujian Talenta Masuk IPB (UTMI)
penulis melanjutkan pendidikannya di Departemen Biologi, Fakultas Matematika
dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor pada tahun 2010.
Selama mengikuti perkuliahan, penulis menjadi asisten praktikum Biologi
Dasar TPB pada tahun ajaran 2012/2013 dan 2013/2014, Fisiologi Tumbuhan Dasar
dan Kultur Jaringan Tanaman pada tahun ajaran 2013/2014, Pendidikan Agama
Islam TPB pada tahun ajaran 2013/2014. Penulis juga aktif sebagai bendahara II
Departemen KOMINFO BEM FMIPA Kabinet FMIPA Bersatu 2011/2012,
sekretaris Badan Pengawas HIMABIO 2011/2012-2012/2013, dan staf Humas
organisasi eksternal kampus Garuda Keadilan Bogor 2011/2012. Selain itu penulis
juga aktif di berbagai kepanitiaan yang diselenggarakan di IPB.
Selama menempuh studi di Departemen Biologi, penulis melakukan
penelitian dalam studi lapangan mengenai Reduksi Nitrat di Ekosistem Akuatik
Taman Nasional Gunung Gede Pangrango pada tahun 2012 dan praktik lapangan
di Pusat Aplikasi Teknologi Isotop dan Radiasi – BATAN mengenai Seleksi Galur
Mutan Padi Dataran Tinggi Toleran Cekaman Kekeringan secara In vitro pada
tahun 2013.





























