Embed Size (px)
Citation preview

Stoffwechselhormone
- 1 -
Universität Ulm WS 2004/2005
Inhaltsverzeichnis
1. Theoretischer Hintergrund.......................................4
1.1. Definition des Begriffs „Hormon“............................4
1.2. Einteilungsmöglichkeiten von Hormonen...............4
a) Einteilung nach der Wirkungsweise
b) Einteilung nach Bildungs – und Wirkungsort
c)Einteilung nach chemischer Struktur
1.3. Molekulare Wirkungsmechanismen........................5
a) Genaktivierungsprinzip
b) Second – messenger – Prinzip
1.4. Hormondrüsen des Menschen.................................9
a) Drüsen und ihre Hormone
b) Hierarchie der Hormondrüsen
1.5. Hormone der Insekten.............................................10
1.6. Regulation des Energiestoffwechsels...................
a) beim Menschen
b) bei Inesekten

Stoffwechselhormone
- 2 -
1.7. Definition der „Lucusta-Einheit“............................
1.8. Aufgabenstellung des Versuchs............................
2. Material und Methoden............................................
2.1. Versuch 1: Dosis-Wirkungs-Kurve.......................
2.2. Versuch 2: Zeit-Wirkungs-Kurve............................
2.3. Präparation der Corpora cardiaca..........................
3. Ergebnisse................................................................
3.1. Versuch 1..................................................................
3.2. Versuch 2....................................................................
4. Diskussion.................................................................
4.1. Versuch 1...................................................................
4.2. Versuch 2...................................................................
4.3. Vergleich der Nullwerte...........................................
5. Quellenangaben........................................................

Stoffwechselhormone
- 3 -
1. Theoretischer Hintergrund
1.1. Definition des Begriffs "Hormon"
Hormone sind chemische Botenstoffe, die die metabolische Aktivität von Zellen
Geweben oder Organen beeinflussen. Sie werden von speziellen Drüsen oder Zellen
in kleinen Mengen produziert und auf verschiedenen Wegen zu ihren Zielorganen
transportiert (Blutbahn, Hämolymphe, Luft). Dort lösen Hormone nach Bindung an
einen spezifischen Rezeptor eine bestimmte Reaktion aus.
1.2. Einteilungsmöglichkeiten von Hormonen
a) Einteilung nach der Wirkungsweise
� Kinetisch: Wirkung auf Pigmentwanderung (Melatonin), Muskelkontraktion
(Adrenalin) und Drüsensekretion (Releasing-Hormone)
� Metabolisch: Wirkung auf den Kohlenhydrat – und Proteinhaushalt (Thyroxin,
Insulin) und den Elektrolyt – und Wasserhaushalt (ADH, Aldosteron)
� Morphogenetisch: Wirkung auf Wachstum und Differenzierung, zum Beispiel
Allgemeinwachstum (Wachstumshormone), Metarmophose (Thyroxin),
Differenzierung der primären und sekundären Geschlechtsmerkmale
(Androgene, Östrogene)
� Verhalten: Wirkung durch Einfluss auf die Funktionsweise des
Nervensystems (Östrogene, Androgene, Progesteron)

Stoffwechselhormone
- 4 -
b) Einteilung nach Bildungs – und Wirkungsort
Unterscheidung nach ihrem Bildungsort:
� Neurohormone: werden von neurosekretorischen Zellen produziert und
freigesetzt (hypothalamische Hormone, Neurohypophysenhormone)
� Glanduläre Hormone: Bildung in speziellen Hormondrüsen, z.B. Schilddrüse,
Bauchspeicheldrüse, Nebennieren
� Aglanduläre Hormone: Produktion durch bestimmte Gewebe mit endokriner
Funktion, Wirkung auf direkt benachbartes oder nahe gelegenes Zielgewebe
(Prostaglandine, Histamin)
� Mediatoren: hormonähnliche Stoffe, die von verschiedenen Zellen produziert
werden, aber nur eine lokale Wirkung zeigen
Unterscheidung nach ihrem Wirkungsort:
� Autokrin: Hormone, die die sezernierende Zelle selbst beeinflussen
� Parakrin: Wirkung auf direkt benachbarte Zellen
� Endokrin: Botenmoleküle werden über die Blutbahn zu weiter entfernt
liegendem Gewebe transportiert
� Spezielfall: wenn die Hormone von Nervenzellen gebildet und in die Blutbahn
abgegeben werden, spricht man vom neuroendokrinen System
� Exokrin: werden aus dem Organismus heraustransportiert (Bsp.: Pheromone)
c) Einteilung nach chemischer Struktur
� Amine (Aminosäureabkömmlinge): lassen sich von Aminosäuren ableiten,
Bsp.: Adrenalin, Thyroxin
� Fettsäureabkömmlinge: cyclisch ungesättigte Fettsäuren, die meistens aus
Lipiden der Plasmamembran gebildet werden, Bsp.: Prostaglandin

Stoffwechselhormone
- 5 -
� Steroide: cyclische Kohlenwasserstoffderivate, die aus Cholesterin
synthetisiert werden, Bsp.: Östrogen, Testosteron, Cortison
� Proteo – und Peptidhormone: bestehen aus kurzen oder längeren Amino-
säureketten, Bsp.: Insulin, Glukagon, Releasing-Hormone
1.3. Molekulare Wirkungsmechanismen
a) Genaktivierungsprinzip
Die Genaktivierung kann nur mit Hilfe von lipophilen Hormonen stattfinden, die die
Zellmembran der Zielzelle passieren können. Im Cytoplasma bindet das Hormon an
seinen spezifischen Rezeptor nach dem Schlüssel-Schloss-Prinzip und bildet einen
Hormon–Rezeptor–Komplex. Dieser kann nach einer Konformationsänderung die
Kernmembran passieren und so über ein Akzeptormolekül an bestimmte Bereiche
der DNA binden. Dort wirkt der Komplex als Transriptionsfaktor und schaltet
spezifische Gene an oder aus.
Abb. 1: Genaktivierungsprinzip
Campbell, Biologie, S.1002, 2.Auflage
b) Second – messenger – Prinzip

Stoffwechselhormone
- 6 -
Im Gegensatz zu den lipophilen Hormonen können hydrophile Hormone die
Cytoplasmamembran nicht passieren. Sie wirken über einen intrazellulären
Botenstoff, der die jeweiligen Reaktionen in der Zelle in Gang setzt.
cAMP
Das zyklische Adnosinmonophosphat (cAMP) ist ein Beispiel für ein chemisches
Botenmolekül.
Die Aktivierung wird nun am Beispiel von Adrenalin erklärt:
Das Hormon (first messenger) bindet spezifisch an seinen membranständigen
Rezeptor in der jeweiligen Zielzelle. Dadurch entsteht ein Hormon-Rezeptor-
Komplex, der wiederum an ein bestimmtes Membranprotein bindet, ein sogenanntes
G–Protein. G–Proteine sind molekulare Überträgerstoffe, die eine aktivierende oder
eine hemmende Wirkung ausüben können. Durch die Bindung des G–Proteins mit
GTP (Guanintriphosphat) wird ein weiteres Protein, nämlich das Enzym
Adenylatcyclase, aktiviert. Die Adenylatcyclase setzt nun ATP zu cAMP um, welches
den second messenger darstellt.
Im nächsten Schritt beeinflusst cAMP ein weiteres Enzym, die cAMP-abhängige
Proteinkinase oder kurz A-Kinase. A-Kinasen bestehen aus einer regulatorischen
und einer katabolischen Untereinheit. Sie werden aktiviert, indem cAMP die
regulatorische Untereinheit entfernt, woraufhin die katalytische Untereinheit frei wird
und anschließend eine Enzymkaskade in Gang setzt. Nun findet die
Depolymerisation von Glycogen zu Glucose statt.

Stoffwechselhormone
- 7 -
Abb. 2: Sekond Messenger Prinzip
Campbell, Biologie, S.1003, 2.Auflage
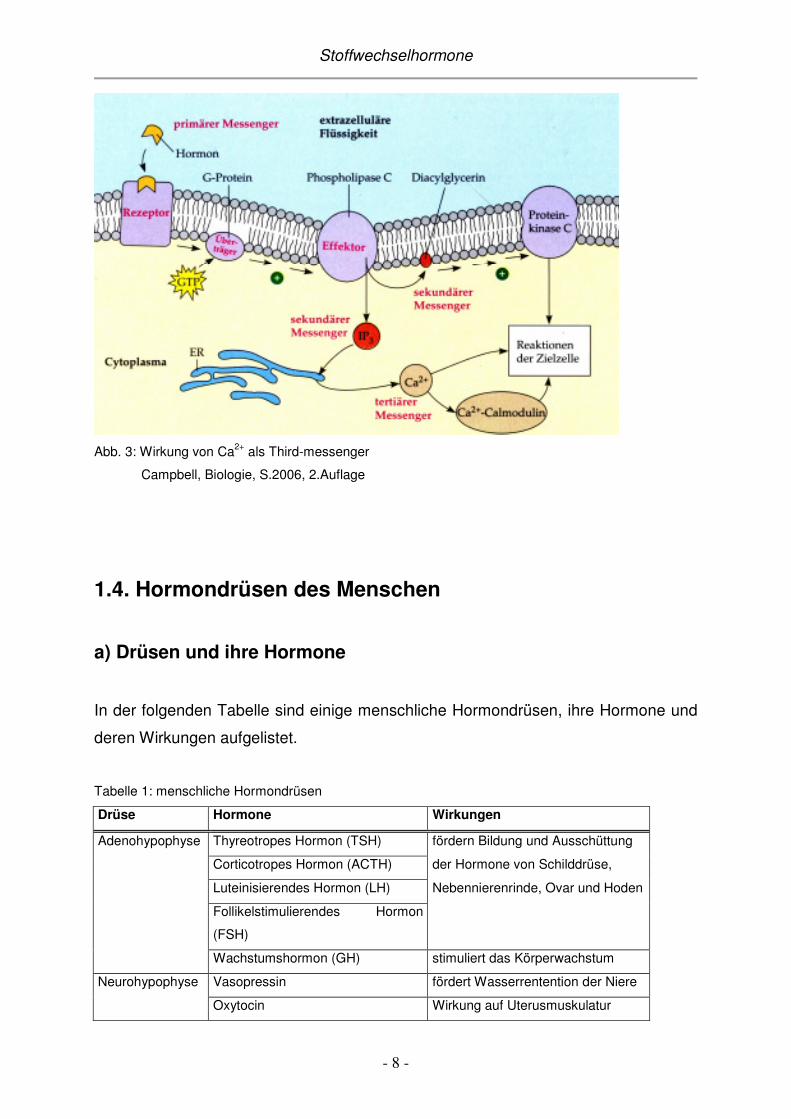
Ca2+
Neben den chemischen Botenmolekülen können auch verschiedene andere
Substanzen, wie zum Beispiel Calciumionen, für eine Signaltransduktion
verantwortlich sein.
Wieder bindet das Hormon (first messenger) an seinen spezifischen
Membranrezeptor und der Hormon-Rezeptor-Komplex aktiviert ein G-Protein. Das G-
Protein aktiviert daraufhin das Enzym Phospholipase C, welches Phospholipide in
Diacylglycerin (DAG) und Inositoltriphosphat (IP3) spaltet. Beide Stoffe wirken als
second messenger. DAG aktiviert das Enzym Proteinkinase C, das durch
Phosphorylierung bestimmter Proteine in der Zellmembran seine Wirkung entfaltet.
IP3 hingegen bewirkt eine intrazelluläre Erhöhung der Ca2+ - Konzentration, entweder
durch Aufnahme aus dem extrazellulären Medium oder durch Freisetzung von
Calciumionen aus dem ER. Ca2+ wirkt dabei als third messenger. Die Calciumionen
üben ihre Wirkung auf die Zielzelle nun direkt aus oder durch Bindung an das Protein
Calmodulin.
Ein Beispiel für diesen Mechanismus ist das Hormon Insulin.
Stoffwechselhormone
- 8 -
Abb. 3: Wirkung von Ca2+ als Third-messenger
Campbell, Biologie, S.2006, 2.Auflage
1.4. Hormondrüsen des Menschen
a) Drüsen und ihre Hormone
In der folgenden Tabelle sind einige menschliche Hormondrüsen, ihre Hormone und
deren Wirkungen aufgelistet.
Tabelle 1: menschliche Hormondrüsen
Drüse Hormone Wirkungen
Adenohypophyse Thyreotropes Hormon (TSH) fördern Bildung und Ausschüttung
Corticotropes Hormon (ACTH) der Hormone von Schilddrüse,
Luteinisierendes Hormon (LH) Nebennierenrinde, Ovar und Hoden
Follikelstimulierendes Hormon
(FSH)
Wachstumshormon (GH) stimuliert das Körperwachstum
Neurohypophyse Vasopressin fördert Wasserrentention der Niere
Oxytocin Wirkung auf Uterusmuskulatur

Stoffwechselhormone
- 9 -
Schilddrüse Thyroxin Regulation des
Trijodthyronin Grundumsatzes
Pankreas Insulin Regulation des Glukose-
Glukagon spiegels im Blut
Nebennierenmark Adrenalin und Erhöhung der Leistungsbereitschaft
Noradrenalin
Nebennierenrinde Clucocorticoide beeinflussen den Kohlenhydratstoff-
wechsel und die Immunreaktion
Mineralocorticoide Regulation des Mineral- und
Wasserhaushaltes
Eierstock Östrogene Ausbildung der weibl. Geschlechts-
merkmale
Gestagene regeln Ovarialzyklus und
Schwangerschaft
Hoden Androgene
Ausbildung der männl.
Geschlechts-
merkmale
b) Hierarchie der Hormondrüsen
Der Hypothalamus bildet die oberste Kontrollinstanz des Drüsensystems des
Körpers. Er empfängt Informationen aus allen Teilen des Körpers, vor allem aber von
anderen Gehirnregionen. Die hypothalamischen Hormone werden von zwei
verschiedenen Sorten von neurosekretorischen Zelle gebildet und dann zur
Hypophyse weitergeleitet.
Die Hypophyse (Hirnanhangsdrüse) ist die übergeordnete Hormondrüse, da sie
andere Hormondrüsen zur Hormonproduktion anregen kann, dabei aber selbst unter
hormoneller Kontrolle des Hypothalamus steht. Sie besteht aus zwei getrennten
Teilen, der Neurohypophyse und der Adenohypophyse.
Die Neurohypophyse ist ein Neurohämalorgan, d.h. die Axone der
neurosekretorischen Zellen des Hypothalamus sind so lang, dass sie bis in die
Neurohypophyse hineinreichen, wodurch die Hormone direkt übertragen werden

Stoffwechselhormone
- 10 -
können. Sie dient zur Speicherung und Sekretion der Hormone Adiuretin (ADH) und
Oxytocin.
Die Adenohypophyse wird durch Releasing-(RH) oder Inhibiting-Hormone (IH)
beeinflusst. Diese werden von neurosekretorischen Zellen mit kurzen Axonen
produziert und über ein Pfortadersystem zur Adenohypophyse transportiert. Sie
regen zur Bildung eigener Hormone an, zum Beispiel die vier tropischen Hormone
Thyreotropin (TSH), Adrenocorticotropin (ACTH), Follikel stimulierendes Hormon
(FSH) und Luteinisierendes Hormon (LH), die andere Hormondrüsen zur
Hormonproduktion anregen, oder das Wachstumshormon.
1.5. Hormone der Insekten
Hier sind Hormone für Metamorphose und Häutungszyklen, Fortpflanzung und
Diapause zu ständig. Daneben aber auch sowohl zur Regulation des Gehalts an
Metaboliten in der Hämolymphe als auch zur Osmoregulation.
Die „Hormondrüsen“ (s.Abb.4) lassen sich im Grunde in zwei Gruppen aufteilen, die
neurosekretorischen Zellen im Gehirn und die Neurohämalorgane im weiteren
Kopfbereich.
Abb.4 : Lage der endokrinen Organe bei einem Insekt
De Deuterocerebrum; Pr Protocerebrum; Tr Tritocerebrum
(aus: Vergleichende Tierphysiologie, Heldmaier/Neuweiler, S. 407, Abb. 9.4)
Die neurosekretorischen Zellen sind Nervenzellen, die Hormon produzieren. Diese
liegen im Zentralen Nervensystem und sind wie normale Nervenzellen aufgebaut.
Zusätzlich besitzen sie aber noch sogenannte neurosekretorische Granula.

Stoffwechselhormone
- 11 -
Das namengebende Neurosekret wird im Zellkörper gebildet und innerhalb der
Axone transportiert. Deren verdickten Enden bilden keine Synapsen wie normale
Nervenzellen, sondern enden in unmittelbarer Nähe zu den Neurohämalorganen.
Die Aufgabe dieser Organe besteht in der Speicherung und Kontrolle der Abgabe
des Neurosekrets. Zum Teil kann das Neurosekret aber auch direkt ins
Kreislaufsystem (und damit in die Hämolymphe) abgegeben werden.
Zu den Neurohämalorganen gehören zum einen die paarig angeordneten Corpora
cardiaca. Diese erhalten Neurosekret aus verschiedenen Regionen des
Protocerebrums und Tritocerebrums. Zusätzlich enthalten sie Zellen, die selbst
endokrine Funktion besitzen und Neurone, die über Axone mit den Corpora allata
verbunden sind.
Die Corpora allata ist ein herkömmliches endokrines Organ, d.h. die Zellen sind
darauf spezialisiert Botenstoffe zu synthetisieren und direkt ins Blut zu sekretieren.
Bei diversen Insekten ist die C. allata nicht mehr paarig angeordnet, sondern zu
einem einzigen Organ verschmolzen (C. allatum).
Das andere wichtige Organ ist die ebenfalls paarige Prothorakaldrüse. Bei
stammesgeschichtlich frühen Insekten ist sie eine kompakte endokrine Drüse, die im
Bereich des Kopfes liegt. Bei späteren Insekten dagegen liegt sie mehr im Thorax
und ist verbreitert.
In der nachstehenden Tabelle sind die wichtigsten Hormone und ihre Wirkungen
aufgelistet.
Tab.2 : Hormone der Insekten
Bildungsort Hormon Chem.
Struktur
Wirkung
Neurosekretorische
Zellen im Gehirn,
Ausschüttung über
Corpora cardiaca
Prothorakotrophes
Hormon (PTTH)
Protein Stimuliert Ecdyson
Synthese, startet
dadurch die
Vorbereitung der
Häutung
Prothorax-Drüse Ecdyson Steroid Häutungshormon,
verändert Stoffwechsel
der Epidermiszellen

Stoffwechselhormone
- 12 -
Corpora allata Juvenilhormon
(JH)
Terpenderivat Hoher JH-Gehalt erhält
Larvalstadien bei der
Häutung, niedriger Titer
verursacht
Imaginalhäutung
Neurosekretorische
Zellen im Gehirn,
Ausschüttung aus
den Corpora
cardiaca
Adipokinetisches
Hormon (AKH)
Protein Etwa 30 AKH´s bekannt,
mobilisieren
Speicherlipide,
Energieversorgung beim
Flug, gleichzeitig
Neurotransmitter,
stimuliert
lokomotorische Aktivität
Motoneurone Proctolin Protein Neuromodulator,
steigert Herzfrequenz,
Darmmotorik und
andere neuronal
ausgelöste
Muskelkontraktionen
Neurosekretische
Zellen im Gehirn
Bursicon Protein fördert
Cuticulaentwicklung,
Härten und Gerben der
Cuticula nach der
Häutung (vor allem nach
Imaginal-Häutung
2 Paar
neurosekretorische
Zellen im Gehirn
Eclosionshormon
(EH)
Protein Löst den eigentlichen
Schlüpfvorgang aus
1.6. Regulation des Energiestoffwechsels

Stoffwechselhormone
- 13 -
Der Energiestoffwechsel wird neben anderen Faktoren auch hormonell geregelt.
Dabei ist es wichtig, dass die Höhe des Hormonspiegels überwacht und reguliert
werden kann. Im einfachsten Falle hemmt oder fördert die Substratkonzentration
selbst die Hormonbildung. Weitere Möglichkeiten wurden schon im Abschnitt 1.4.
erläutert.
a) beim Menschen
Chemische Energie kann auf verschiedene Weisen gewonnen werden, aerob oder
anaerob, und zwar wird dabei immer aus Nährstoffen ATP produziert.
Gärung als Abbau von Zuckermolekülen ohne Sauerstoff wird nur in
Ausnahmezuständen genutzt. Dazu zählt die Milchsäuregärung im Muskel, die zu
Muskelkater und Muskelerschöpfung führen kann. Dabei wird Pyruvat vom NADH
direkt zu Lactat reduziert. Dieses wird später vom Blut zur Leber befördert und in den
Zellen wieder zu Pyruvat umgesetzt. Die Milchsäuregärung ist aber auch
dahingehend negativ zu bewerten, dass hier nur zwei ATP Moleküle (der universelle
Energieüberträger der Zelle) entstehen.
Die normale Energiegewinnung ist die unter Sauerstoffverbrauch. Die dabei
ablaufenden Prozesse werden Glykolyse, Citratcyclus und Atmungskette genannt
und laufen im Mitochondrium ab.
Die Glykolyse entspricht der Milchsäuregärung, d. h. auch hier wird Glucose zu
Pyruvat oxidiert. Dieses wird dann allerdings im Citratcyclus weiter oxidiert, wodurch
die Energie die noch im Pyruvat steckt auch noch für den Körper nutzbar wird.
Bei der anschließenden Atmungskette (wozu auch die oxidative Phosphorilierung
gehört) wird nun der größte Teil der Energie gewonnen. Denn dabei oxidiert NADH
durch Luftsauerstoff und freiwerdende Energie wird zur ATP Synthese eingesetzt.
Dabei werden insgesamt pro Molekül Glucose immerhin 36 ATP Moleküle frei
(s.Abb.5).

Stoffwechselhormone
- 14 -
Abb.5: Eine Zusammenfassung der Zellatmung (aus: N.A.Campbell; Biologie, S.190, Abb. 9.15)
Als Beispiel für die Regulation des Energiestoffwechsels seien hier die
antagonistischen Hormone Insulin und Glucagon erwähnt, die von den
Langerhansschen Zellen ausgeschüttet werden. Um den Stoffwechsel im
Gleichgewicht zu halten, sollte der Blutglucosespiegel bei ungefähr 0,9 mg/ml liegen.
Insulin wird ausgeschüttet, sobald der Blutzuckergehalt über diesen Wert steigt.
Neben anderen Auswirkungen werden dadurch fast alle Körperzellen, ausgenommen
die Gehirnzellen, angeregt Glucose aus dem Blut aufzunehmen. Falls aber der Wert
unter diesen Punkt sinkt, wird Glucagon ausgeschüttet und z.B. die Leber angeregt
eingelagertes Glycogen abzubauen und als Glucose wieder ins Blut abzugeben.
Zur Energiespeicherung wird Glucose nicht nur in der Leber sondern auch in den
Muskeln in Form von Glycogen eingelagert, während das Fettgewebe es zu Fett
umbaut.
b) bei Insekten
Die Flugmuskeln der Insekten gewinnen ihre Energie stets aerob. Da die
Flugmuskeln selber keine großen Energiereserven speichern können, müssen
energiereiche Moleküle von der Hämolymphe geliefert werden.

Stoffwechselhormone
- 15 -
Hierbei kann man zwischen Aminosäuren-, Kohlenhydrat- und Fettfliegern
unterscheiden. Das Gewebe mit Speicherkapazität und der entsprechenden
Stoffwechselaktivität ist der Fettkörper der Insekten.
Nur wenige Arten wie die Tsetse-Fliege Glossina morsitans oder der Kartoffelkäfer
Leptinotarsa decemlineata nutzen Aminosäuren (genauer Prolin) als Energielieferant
im Flugstoffwechsel. Das Prolin entsteht im Fettkörper durch den Abbau von
Fettsäuren bzw. aus Acetyl-CoA, im Citratcyclus unter Beteiligung von Alanin.
Danach wird das Prolin über die Hämolymphe in den Flugmuskel transportiert und
hier oxidativ abgebaut. Dabei entsteht wieder Alanin, das über die Hämolymphe
zurück in den Fettkörper gelangt und damit zur erneuten Synthese von Prolin zur
Verfügung gestellt wird.
Als Kohlenhydrat-Flieger gelten zum Beispiel die Schmetterlinge. Denn sie haben
meist nur kurze Flüge von Blüte zu Blüte zu bewältigen und nehmen dabei
zuckerhaltigen Nektar auf. Dieser wird zum Disaccharid Trehalose umgebaut, da dies
der Transportzucker in der Hämolymphe ist. In den Flugmuskeln angekommen wird
die Trehalose vom Enzym Trehalase zu Glucose abgebaut, so dass in den Muskeln
die Glykolyse ablaufen kann.
Der von uns betrachtete Fettflieger ist die Wanderheuschrecke Locusta migratoria,
die zu den Langstreckenfliegern zählt. Um die Energie dafür aufbringen zu können,
wird in den ersten Flugminuten auf schneller verfügbare Energie aus Kohlenhydraten
(Trehalose) zurückgegriffen.
Der anfängliche Kohlenhydratstoffwechsel wird durch das Aminhormon Octopamin
gesteuert und entspricht damit dem blutzuckersteigernden Hormon Adrenalin der
Wirbeltiere. Nach 10-20 Minuten übernehmen, zum Ende der ersten Flugphase,
Peptidhormone die Regulation des Stoffwechsels und der Energiebereitstellung.
Diese werden in der Corpora cardiaca synthetisiert und im hinteren Hirnbereich
gespeichert. Über die Verbindung mit dem Nervi Corporis zum Gehirn wird die
Exozytose des AKH induziert. Durch die Hämolymphe gelangt es zum Fettkörper und
bindet dort an seine Rezeptoren. Wie in Abb. ersichtlich, löst dieser Vorgang in der

Stoffwechselhormone
- 16 -
Zelle, über die second- Messenger cAMP und Ca2+, die Aktivierung der Lipase aus,
welche die gespeicherten Triacylglyceride in Diacylglyceride spaltet. Diese
Diglyceride werden an der Membranoberfläche des Fettkörpers mit dem (in der
Hämolymphe vorhandenen) Transportprotein HDLp und dem Apoprotein ApoLp-III
zu einem Transportkomplex zusammengelagert. Dieser ganze Komplex wird als
LDLp bezeichnet und übernimmt die Beförderung der Diglyceride zu den
Muskelzellen.
Dort bindet der Transportkomplex an das membranständige Enzym Lipophorin-
Lipase, welche den Abbau katalysiert und dabei die Fettsäuren von Glycerin spaltet.
Die Fettsäuren wiederum werden in die Muskelzelle aufgenommen und oxidiert,
während die restlichen Bestandteile des Komplexes in der Hämolymphe
zurückbleiben und somit wieder der Komplexbildung zur Verfügung stehen. Der
Verlauf des Fettstoffwechsels ist in Abb. 6 dargestellt.
Abb.6: Die physiologische Steuerung des Fettstoffwechsels
AKH Adipokinetisches Hormon; ApoLp-III ApollpophorinIII; CC Corpora cardiaca; DG
Diacylglyceride; FFa freie Fettsäuren; HDLp High-Density-Lipophorin; LDLp Low-Density-
Lipophorin; TG Triacylglyceride
(aus: Skript zum Tierphysiologischen Anfängerpraktikum)
Die Regulation dieses Stoffwechselprozesses gelingt nur über die Zusammenarbeit
aller beteiligten Proteine. So inhibiert freies ApoLp-III in der Hämolymphe zusätzlich
die Lipophorin- Lipase, was einen Mindestgehalt an Fett in der Hämolymphe sichert.
Der Umstand, dass Fette osmotisch inaktiv speicherbar sind, und beim oxidativen
Abbau der Fettsäuren metabolisches Wasser entsteht, führt zu einem geringen
Wasserbedarf der Tiere. Außerdem besitzen Fette ein hohes ATP-Äquivalent von

Stoffwechselhormone
- 17 -
0,65 mol ATP pro g Fett. Diese Fakten sind für Langstreckenflieger von besonderer
Bedeutung und Nutzen.
1.7. Definition der „Lucusta-Einheit“
Unter der Lucusta-Einheit versteht man die Menge an AKH, die benötigt wird, um die
halbe Fettkonzentration in der Hämolymphe zu erreichen. Diese AKH Menge
mobilisiert eine bestimmte Menge Fett in Form von Diglyceriden.
1.8. Aufgabenstellung des Versuchs
In diesem Versuch wird untersucht, in wieweit sich das Hormon AKH auf den
Fettmetabolismus der Wanderheuschrecke Locusta migratoria auswirkt. Dies wird
anhand der Abhängigkeit von der Zeit und der Dosis auf die Fettkonzentration der
Hämolymphe analysiert. Außerdem werden am Ende aus sieben
Wanderheuschrecken die Corpora cardiaca´s herauspräpariert, damit ein Corpora
cardiaca Aliquot für die nächste Gruppe bereit steht.
2. Material und Methoden
2.1. Versuch 1: Dosis-Wirkungs-Kurve
Vor Versuchsbeginn wurde vier Wanderheuschrecken (Locusta migratoria) je 5µl
einer unterschiedlich hoch konzentrierten Lösung eines Corpora cardiaca Extraktes,
welches das adipokinetische Hormon (AKH) enthält, von der Betreuerin injiziert.
Dafür wurden sieben Corpora cardiaca aus dem Versuch der vorherigen Woche
verwendet und mit Wasser verdünnt, damit man die Mengen von 0,009, 0,0175,
0,035 und 0,07 als Anteile der extrahierten Hormondüsen bekommt. Außerdem wird
einem Tier als Blindwert 5µl bidestilliertes H20 injiziert.

Stoffwechselhormone
- 18 -
60 Minuten nach der Injektion wird dem Tier Hämolymphe entnommen, um den darin
vorhandenen Fettgehalt zu bestimmen. Hierfür wird das Tier auf den Rücken gelegt,
festgehalten und der Kopf mit einer Pipettenspitze nach hinten gehalten. An der
Übergansregion von Caput und Thorax, an der der Chitinpanzer sehr dünn ist, wird
die Wanderheuschrecke mit einem Skalpell perforiert. Durch Ausübung von Druck
auf den Thorax bildet sich an der angeschnittenen Stelle eine Tropfen Hämolymphe,
der dann abpipettiert werden kann. Hierbei werden jedem Tier zur
Doppelbestimmung zweimal 10µl Hämolymphe entnommen, wobei darauf
aufgepasst werden muss, dass keine Fettzellen aus dem nahe liegende Fettkörper
mit aufgenommen werden.
Die entnommene Hämolymphe wird zu je 1ml Isopropanol gegeben und sofort mit
Hilfe eines Rüttlers gemischt. Ebenfalls werden ein Triglycerid-Standard (Triolein,
3mg/ml), ein Diglycerid- Standard (Dipalmitin, 3mg/ml) und reines Wasser als
Leerwert mit je 1ml Isopropanol vermischt (Pipettierschema siehe Tabelle 3).
Tabelle 3: Pipettierschema zur Dosis-Wirkungs-Kurve
Isopropanol Hämolymphe Standard H20
Probe 1ml 10µl
Standard 1ml 10µl
Leerwert 1ml 10µl
Zu jeder Probe werden nun 200µl Kalilauge (KOH) gegeben, mit dem Rüttler
vermischt und in einem Wasserbad 5 Minuten bei 60°C inkubiert. Hierbei reagieren
die Di- und Triglyceride mit dem KOH zu Glycerin und Fettsäuren. Um die Reaktion
zu stoppen, werden die Reagenzien in Eis abgekühlt.
Anschließend wird zu jeder Probe 200µl Natriumperjodad hinzugefügt, geschüttelt
und 10 min bei Raumtemperatur stehen gelassen. Dabei wird das Glycerin durch
Natriumperjodad in Formaldehyd gespalten.
Zuletzt pipettiert man die Farbreagenz (NH4+ und Acetylaceton) in jedes
Reagenzglas, vermischt den Inhalt im Rüttler, setzt Glaskugel auf die Reagenzgläser
und inkubiert sie eine halbe Stunde bei 60°C. Dadurch reagiert das Formaldehyd zu
Diacetyldihydrolutidin, wodurch eine Farbveränderung (gelb) auftritt.

Stoffwechselhormone
- 19 -
Die entstandenen Lösungen werden nun in Küvetten umgeschüttet und bei 410nm
photometriert, wobei die Extinktion gemessen wird.
2.2. Versuch 2: Zeit-Wirkungs-Kurve
Fünf Wanderheuschrecken wurden vor dem Versuch eine Menge von 0,07
Rohextrakt der Corpora cardiaca von der Betreuerin injiziert. Nach unterschiedlichen
Zeiten (10min, 30min, 60min, 90min und 120min) wird dann Hämolymphe
entnommen und der jeweilige Fettgehalt, wie in Versuch 1 beschrieben, bestimmt.
Außerdem wird der Fettgehalt der Hämolymphe eines Ruhetiers, welchem nichts
injiziert wurde, untersucht.
2.3. Präparation der Corpora cardiaca
Die Corpora cardiaca wird aus den Köpfen von 7 Wanderheuschrecken unter dem
Stereobinocular präpariert, welche davor vom restlichen Körper abgetrennt wurden.
Die Hormondrüse ist halbmonförmig und liegt dorsal hinter dem Gehirn, hinter den
Mundwerkzeugen und Oberschlundganglion.
Die sieben Drüsen werden in reinem Wasser homogenisiert, so dass das Gewebe
aufgeschlossen wird und in Eppendorfgefäße gegeben. Dann wird nochmals
homogenisiert, wobei als Lösungsmittel Methanol verwendet wird und wieder in
Eppendorfgefäße umgefüllt. Diese werden dann in einer Biofuge 5min bei 13000U
zentrifugiert. Der Überstand wird abgenommen und über Nacht eingeengt, so dass
das für Insekten schädliche Methanol verdampfen kann. Das gewonnene Rohextrakt
der Corpora cardiaca kann dann von dem nächsten Kurs verwendet werden.
3. Ergebnisse
3.1. Versuch 1
Um die Konzentration der Lösungen berechnen zu können, wird zuerst der
Absorptionskoeffizient ε mit Hilfe des Lambert-Beerschen-Gesetztes berechnet, das
wie folgt lautet:
∆E=c*d*ε � c*d
E∆=ε
Stoffwechselhormone
- 20 -
Dieses Gesetz besagt, dass die Extinktion E∆ der Konzentration c des
absorbierenden Stoffes und der Schichtdicke d der Lösung proportional ist.
Der Absorptionskoeffizient ist ε, er ist abhängig von der Art der Substanz und der
Wellenlänge.
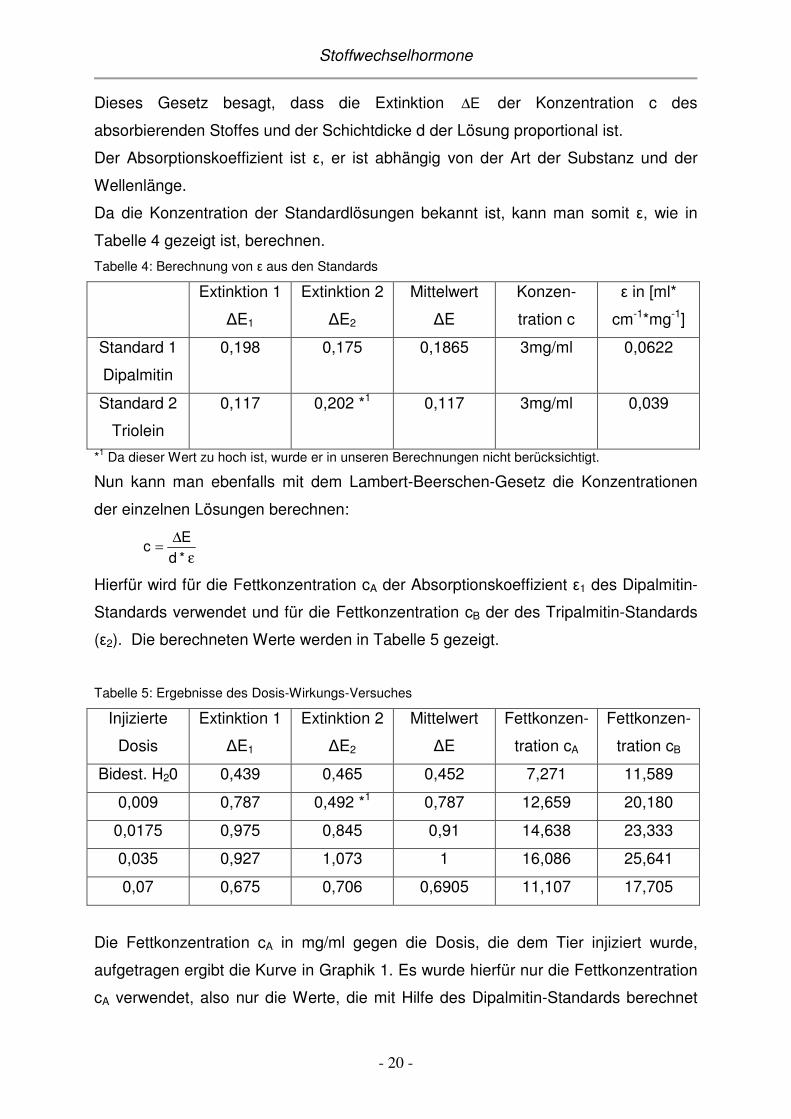
Da die Konzentration der Standardlösungen bekannt ist, kann man somit ε, wie in
Tabelle 4 gezeigt ist, berechnen.
Tabelle 4: Berechnung von ε aus den Standards
Extinktion 1
∆E1
Extinktion 2
∆E2
Mittelwert
∆E
Konzen-
tration c
ε in [ml*
cm-1*mg-1]
Standard 1
Dipalmitin
0,198 0,175 0,1865 3mg/ml 0,0622
Standard 2
Triolein
0,117 0,202 *1 0,117 3mg/ml 0,039
*1 Da dieser Wert zu hoch ist, wurde er in unseren Berechnungen nicht berücksichtigt.
Nun kann man ebenfalls mit dem Lambert-Beerschen-Gesetz die Konzentrationen
der einzelnen Lösungen berechnen:
ε
∆=
*d
Ec
Hierfür wird für die Fettkonzentration cA der Absorptionskoeffizient ε1 des Dipalmitin-
Standards verwendet und für die Fettkonzentration cB der des Tripalmitin-Standards
(ε2). Die berechneten Werte werden in Tabelle 5 gezeigt.
Tabelle 5: Ergebnisse des Dosis-Wirkungs-Versuches
Injizierte
Dosis
Extinktion 1
∆E1
Extinktion 2
∆E2
Mittelwert
∆E
Fettkonzen-
tration cA
Fettkonzen-
tration cB
Bidest. H20 0,439 0,465 0,452 7,271 11,589
0,009 0,787 0,492 *1 0,787 12,659 20,180
0,0175 0,975 0,845 0,91 14,638 23,333
0,035 0,927 1,073 1 16,086 25,641
0,07 0,675 0,706 0,6905 11,107 17,705
Die Fettkonzentration cA in mg/ml gegen die Dosis, die dem Tier injiziert wurde,
aufgetragen ergibt die Kurve in Graphik 1. Es wurde hierfür nur die Fettkonzentration
cA verwendet, also nur die Werte, die mit Hilfe des Dipalmitin-Standards berechnet
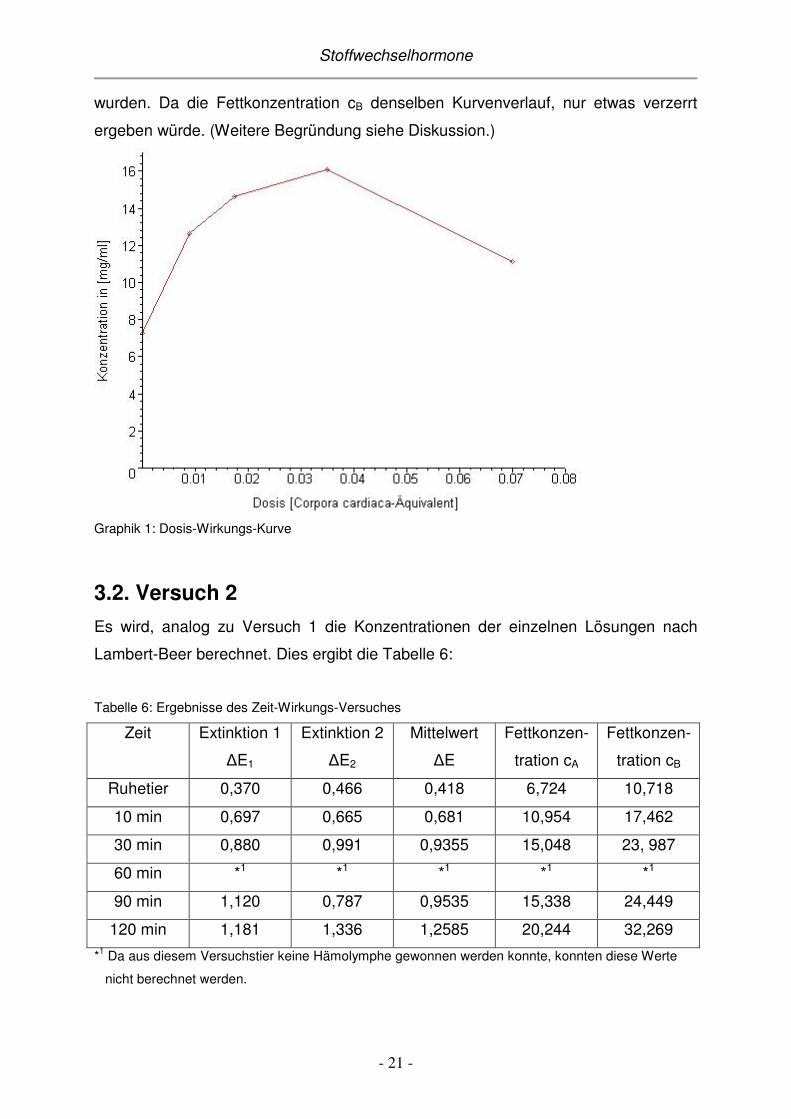
Stoffwechselhormone
- 21 -
wurden. Da die Fettkonzentration cB denselben Kurvenverlauf, nur etwas verzerrt
ergeben würde. (Weitere Begründung siehe Diskussion.)
Graphik 1: Dosis-Wirkungs-Kurve
3.2. Versuch 2
Es wird, analog zu Versuch 1 die Konzentrationen der einzelnen Lösungen nach
Lambert-Beer berechnet. Dies ergibt die Tabelle 6:
Tabelle 6: Ergebnisse des Zeit-Wirkungs-Versuches
Zeit Extinktion 1
∆E1
Extinktion 2
∆E2
Mittelwert
∆E
Fettkonzen-
tration cA
Fettkonzen-
tration cB
Ruhetier 0,370 0,466 0,418 6,724 10,718
10 min 0,697 0,665 0,681 10,954 17,462
30 min 0,880 0,991 0,9355 15,048 23, 987
60 min *1 *1 *1 *1 *1
90 min 1,120 0,787 0,9535 15,338 24,449
120 min 1,181 1,336 1,2585 20,244 32,269
*1 Da aus diesem Versuchstier keine Hämolymphe gewonnen werden konnte, konnten diese Werte
nicht berechnet werden.

Stoffwechselhormone
- 22 -
Die Fettkonzentration cA gegen die Zeit aufgetragen, ergibt die Graphik 2:
Graphik 2: Zeit-Wirkungs-Kurve
4. Diskussion
Bei beiden Versuchen wurden die Ergebnisse mit dem Absorptionskoeffizienten des
Dipalmitin-Standards berechnet, da in den Wanderheuschrecken Diglyceride und
nicht Triglyceride zur eigentlichen Energiegewinnung gespalten werden. Da Triolein
der gängigere Standard ist und deshalb Verwechslungsgefahr besteht, wurde er in
unseren Versuch miteinbezogen. Die höheren Fettkonzentrationen bei Triolein als
Standard lassen sich dadurch erklären, dass bei selber Menge verhältnismäßig
weniger Glycerinmoleküle enthalten sind, was bewirkt, dass weniger Farbstoff
gebildet wird, also die Extinktion geringer ist. Dies hat zur Folge, dass der
Extinktionskoeffizient ε2 niedriger ist als ε1, und somit bei der Berechnung der
Fettkonzentrationen höhere Werte für cB als für cA zustande kommen.

Stoffwechselhormone
- 23 -
4.1. Versuch 1
Bei dem Dosis-Wirkungsversuch wäre eine Sättigungskurve, die sich asymptotisch
einem Grenzwert nähert zu erwarten. Wenn weniger ADH - Hormone als
Rezeptoren am Fettkörper vorhanden sind, steigt die Fettkonzentration durch
Erhöhung der Hormonkonzentration an. Sind alle Rezeptoren mit Hormonen besetzt,
kann trotz Erhöhung der ADH Konzentration nicht mehr Hormon gebunden werden.
Es können also nicht mehr Triglyceride in Diglyceride gespalten werden und dadurch
hat auch die Menge an Glycerin in der Hämolymphe auf einem konstant hohen
Niveau erreicht.
Die Messwerte für Bidest. Wasser und für die Corpora cardiaca Äquivalente von
0,009, 0,0175 sowie 0,035 entsprechen diesen Erwartungen. Der Messwert für das
Corpora cardiaca Äquivalent von 0,07 ist deutlich zu niedrig, dadurch sinkt die Kurve
stark ab, anstatt ein Plateau bei konstantem Wert zu bilden.
Gründe für die zu niedrige Fettkonzentration könnten sein:
� Beim Pipettieren der Hämolymphe sind versehentlich Luftblasen entstanden,
so dass nur eine verringerte Menge an Hämolymphe untersucht werden
konnte.
� Es können Variationen bei der Hormonkonzentration auftreten, da kein reines
AKH verwendet wurde, sondern nur das Äquivalent aus der Corpora cardiaca.
Meint ihr hiermit Schwankungen innerhalb der Probe, weil die Lösung nicht
homogen war? Schwankungen zwischen den einzelnen Konzentrationen
(0,07, 0,035 usw.) können nicht auftreten, da es sich immer um eine 1:2
Verdünnung handelte. Es könnten also nur Fehler auftreten, wenn ich die
Probe vor der Weiterverdünnung nicht richtig vermischt habe. Die
Verwendung von reinem AKH würde an dieser Fehlerquelle aber auch nichts
ändern.
� Das Versuchstier könnte aufgrund der Haltung in einer kleinen Box nervös
geworden sein und versucht haben zu fliehen, was einen deutlich höheren
Fettumsatz zur Folge hat. Somit wäre nur noch eine geringe Menge an
Glycerin im Körper vorhanden und damit nachweisbar.

Stoffwechselhormone
- 24 -
4.2. Versuch 2
Bei dem Dosis-Zeit-Versuch wäre für die Fettkonzentration in Abhängigkeit von der
Zeit eine Glockenkurve zu erwarten, was bedeutet, dass die Kurve relativ steil
ansteigt, dann einen Höhepunkt erreicht und anschließend wieder abfällt. Der
Hochpunkt wäre ungefähr bei der Zeit 60min zu erwarten.
Die injizierte Menge an Corpora cardiaca bzw. ADH war bei allen Versuchstieren
gleich hoch bei 0,07. Somit kann also eine maximale Sättigung wie im Versuch
vorher gezeigt, erreicht werden. Die am Anfang sehr niedrige Fettkonzentration, die
dann ansteigt, bedeutet, dass es eine gewisse Zeit dauert, bis das Hormon seinem
Wirkungsort erreicht und bis die Fette gespalten sind, so dass Glycerin
nachgewiesen werden kann. Nach ca. 60 min ist das Optimum der Fettkonzentration
erreicht, das bedeutet, dass eine maximale Anzahl von ADH-Molekülen den
Fettkörper erreicht haben. Dies bewirkt, dass sich ein Gleichgewicht zwischen den
Molekülen, die den Fettkörper erreichen und dann wieder in diesem abgebaut
werden, einstellt. Da das ADH nach einer gewissen Zeit enzymatische abgebaut
wird, sinkt die Fettkonzentration folglich.
Die Messwerte der Fettkonzentration beim Ruhetier und bei den Tieren, bei denen
die Hämolymphe-Abnahme nach 10min, 30min und 90min erfolgte entsprechen
unseren gerade genannten Vermutungen. Da dem 60 min Tier keine Hämolymphe
entnommen werden konnte, ist in unserer Kurve kein Optimum vorhanden. Wir
vermuten, dass nach 60min die Fettkonzentration bei ca. 19-20mg/ml liegt. Die nach
120min gemessene Fettkonzentration ist viel zu hoch, da die Kurve zu diesem
Zeitpunkt nicht wie bei uns steigen, sondern schon abfallen sollte.
Die Gründe für den zu hohen Messwert können folgende sein:
� Bei der Entnahme von Hämolymphe könnte der Fettkörper mit angestochen
worden sein, so dass daraus zusätzlich Fett in unsere Probe gelangen konnte.
� Wahrscheinlicher hingegen ist, dass das Versuchstier kurz vor der
Hämolymphe-Entnahme unter Stress gelitten hat und so selbst eine hohe
Konzentration an AKH ausgeschüttet hat, wodurch zusätzlich Diglyceride
entstanden und gespalten wurden.

Stoffwechselhormone
- 25 -
� Es wäre auch möglich, dass unser Versuchstier lethargisch war und sich
überhaupt nicht bewegt hat, so dass kein Fett bis zur Entnahme verbraucht
wurde.
Allgemein sind die gemessenen Werte nicht unbedingt zuverlässig und repräsentativ,
da wir jeweils nur einem Tier Hämolymphe entnommen haben und da jede Messung
nur zweifach durchgeführt wurde. Um eine repräsentative Statistik zu bekommen,
müsste man die Versuche viel häufiger durchführen, wodurch exaktere Mittelwerte
entstehen würden und somit abweichende Werte sicherer als Ausreißer erkannt
werden können.
4.3. Vergleich der Nullwerte
Zum Abgleichen wurde einmal ein Ruhetier verwendet, das andere Mal ein Tier, dem
bidest. Wasser injiziert wurde. Dieses Tier steht unter demselben Stress durch die
Injektion, wie die anderen Versuchstiere. Das Ruhetier war diesem Stress nicht
ausgesetzt, ist also in dieser Hinsicht nicht ganz so repräsentativ. Da aber durch die
Wasserinjektion die Hämolymphe verdünnt wurde, wodurch der Fettgehalt in der
entnommenen Hämolymphe erniedrigt wird. Allgemein ist es schwer die beiden Tiere
zu vergleichen, da sie unter unterschiedlichen Bedingungen leben mussten und
jedes Tier sich anders auf Stress reagiert.
5.Quellenangaben
1. Versuchsskript „Kurs Stoffwechselhormone“, Anfängerpraktikum
Tierphysiologie, WS 2004/5
2. Zoologie Wehner Gering, Thieme Verlag, 23.Auflage, 1995
3. Biologie, Campbell, Spektrum Akademischer Verlag, 2.Auflage, 2000
4. Tierphysiologie, Eckert, Thieme Verlag, 4.Auflage, 2002
5. Penzlin H.; Lehrbuch der Tierphysiologie, 4. Auflage, 1989
6. Heldmaier/Neuweiler; Vergleichende Tierphysiologie, 2004





























