Embed Size (px)
Citation preview

1
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E
BIOLOGIA EVOLUTIVA
FRANCIJARA ARAÚJO DA SILVA
Manaus, Amazonas
2019
GENOTOXICIDADE E CITOGENÔMICA COMPARATIVA EM PEIXES DE
IGARAPÉS POLUÍDOS DE MANAUS, AMAZONAS.

2
FRANCIJARA ARAÚJO DA SILVA
ORIENTADORA: Dra. Maria Claudia Gross – UNILA
COORIENTADORA: Dra. Eliana Feldberg – INPA
COORIENTADORA: Dra. Gislene Almeida Carvalho-Zilse – INPA
Manaus, Amazonas
2019
Tese apresentada ao Programa de Pós-
Graduação do Instituto Nacional de Pesquisas
da Amazônia, como requisito para a obtenção
do título de Doutora em Genética, Conservação
e Biologia Evolutiva.
GENOTOXICIDADE E CITOGENÔMICA COMPARATIVA EM PEIXES DE
IGARAPÉS POLUÍDOS DE MANAUS, AMAZONAS.

3

4
Ficha catalográfica
Sinopse:
Os ambientes aquáticos urbanos amazônicos sofrem com alteração ambiental
devido às atividades antrópicas. Consequentemente, substâncias que são lançadas
diariamente nestes locais podem ocasionar danos teciduais e danos no material genético
de células somáticas e germinativas. Para tanto, foram estudados os perfis limnológicos
abióticos, os efeitos genotóxicos, os perfis de metilação e a localização de sequências
repetitivas de DNA em espécies de peixes. No geral, a estrutura, organização e função do
genoma de peixes, que sobrevivem nestes locais, apresentam-se modificadas em espécies
expostas à poluição ambiental, apesar de algumas, terem desenvolvido sistemas
sofisticados para identificar e reparar o DNA danificado.
Palavras-chave: Plasticidade genômica; epigenética; poluição; Amazônia.

5
Dedico esta Tese à minha família, em
especial ao meu avô José Borges (in memorian)
que infelizmente não está presente neste
momento tão importante de minha vida.

6
Que a cada passo que eu der não me falte
foco, força e nem fé!
(Autor desconhecido)

7
A realização deste projeto foi possível devido:
Ao Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva
(GCBEv), do Instituto Nacional de Pesquisas da Amazônia (INPA).
Ao Laboratório de Genética Animal (LGA) do INPA sob coordenação da Dra. Eliana
Feldberg.
Ao Laboratório Temático de Biologia Molecular (LTBM) do INPA sob coordenação
da Dra. Jaqueline da Silva Batista
Ao Laboratório de Citogenômica Animal (LACA) da Universidade Federal do
Amazonas (UFAM), sob antiga coordenação da Dra. Maria Claudia Gross.
À Central Analítica da UFAM sob coordenação da Dra. Teresa Cristina Souza de
Oliveira.
À Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM) pela
concessão da bolsa de estudo durante a realização deste trabalho.
Apoio Financeiro: Processo 23038.009447/2013-45, número do auxílio 3295/2013,
Programa Pró-Amazônia: Biodiversidade e Sustentabilidade Edital Nº 047 / 2012.
Coordenadora: Maria Claudia Gross.
Apoio Financeiro: INCT – Centro de Estudos das Adaptações da Biota Aquática
ADAPTA-AMAZÔNIA/Fase II, Chamada Pública MCTI/CNPq/FAPS Nº 16/2014.
Coordenadora por área: Eliana Feldberg.

8
Agradecimentos
Ufa! Eis que chega minha última obrigação para finalizar a redação desta tese. Perdi as
contas de quantas vezes desejei que este momento chegasse, entretanto aqui estou eu, sem
saber o que escrever, mas ao mesmo tempo posso dizer, não será apenas uma ou duas pessoas,
mas sim VÁRIAS pessoas a quem agradecer. Então, OBRIGADA A TODOS!
Acima de tudo e de todos, agradeço primeiramente a Deus por estar realizando mais
uma etapa de minha vida, que por sinal, não foi nada fácil. Sem ele nada seria possível.
Obrigada Senhor pela força concedida para superar momentos tão difíceis e pelas lágrimas
que precisei derramar para chegar até aqui.
Agradeço à minha família, que representa meu Porto Seguro e minha principal
motivação para continuar buscando algo melhor para todos nós. Em especial, agradeço aos
meus pais Francisco e Joana pelo esforço que fizeram e continuam fazendo para que esta
realização se tornasse possível. Serei eternamente grata e espero retribuir todo este amor
incondicional da melhor forma possível. Agradeço aos meus irmãos Jussara, Júnior e Flávio, e
ao mais novo membro da família, meu cunhado Rubem, pelos incentivos, conselhos, broncas,
e pela força durante esta longa caminhada. Incluo aqui também as minhas meninas Lyla e
Pipoca que continuam sendo a alegria da casa e pelo amor incondicional, mesmo nos
momentos de raiva e estresse. Sem dúvida, o cachorro é o melhor amigo do homem. Amo
todos vocês!
À minha orientadora Dra. Maria Claudia Gross, a quem admiro como pessoa e como
profissional. São nove anos de orientação recheados de ensinamentos, conselhos, risadas,
críticas, incentivos, amizade/maternidade e, além disso, confiança. Olha só, veja no que eu me
tornei graças a você! Obrigada por ter me acolhido no LACA, um laboratório construído com
muito esforço e dedicação sua, é triste falar, mas ele acabou quando você fechou a porta pela
última vez (como faz para conter as lágrimas?). Seu retorno para o Paraná, na metade do meu
Doutorado, não foi nada fácil, mas isso me tornou uma pessoa mais forte, o que me fez
crescer e amadurecer profissionalmente, de uma forma que nem eu acreditei. Nunca se
esqueça de que você foi o diferencial e o pontapé inicial para que eu pudesse chegar até aqui e
por nunca me deixar desistir, por isso, sou grata por você ter me proporcionado experiências
incríveis, tanto fora quanto dentro do LACA. Jamais esquecerei a sua mensagem dizendo:
“Francy, só pra te relembrar: tenho muito orgulho de você, você é o maior exemplo de
orientado que supera o orientador”. Claudia, muito obrigada por tudo!

9
Às minhas coorientadoras Dra. Eliana Feldberg e Dra. Gislene Almeida Carvalho-
Zilse, pela ajuda e pelos ensinamentos durante o trabalho. Obrigada pelas discussões, críticas,
sugestões e pela paciência durante minhas dificuldades. Em especial, agradeço à Dra. Eliana
Feldberg (Dona Boa), pois no momento que me encontrava desmotivada em terminar o
Doutorado, abriu as portas de seu laboratório, e me recebeu com todo carinho, tornando-se
uma mãezona científica para todos nós. Obrigada por ter me proporcionado novamente um
ambiente produtivo cientificamente e é claro, por sempre estar disposta a ler e reler
rapidamente meus artigos. Espero sempre retribuir todo este carinho da melhor forma
possível.
Ao coordenador do Núcleo Especial de Polícia Marítima (NEPOM/AM), Cláudio
Cesar da Silva, que permitiu a autorização para a realização das coletas no igarapé do
Tarumã-açu. Obrigada pelo apoio logístico oferecido, mas principalmente pela sua
empolgação com o trabalho que iríamos realizar. Você se tornou um grande amigo e um
parceiro com quem posso contar a qualquer momento.
Como não deixar de agradecer aos pescadores Agenor Negrão, Atemir Andrade, José
da Silva, Mario Luiz Picanço e Astrogildo Dias, pelas coletas que foram realizadas nos
igarapés poluídos de Manaus e no lago Catalão. Apesar das dificuldades logísticas, pois não
era fácil coletar nestes locais, muito menos evitar o contato direto com a contaminação. Eles
toparam embarcar nessa comigo e hoje são meus amigos, com quem posso contar para voltar
lá. Sou muito, muito grata pela ajuda de vocês.
À Dra. Tereza Cristina Souza de Oliveira e ao MSc. Milton Costa Viana, por todo o
ensinamento da Química Ambiental. Quando me deparei com esta área do conhecimento, sem
ter a mínima experiência, vocês me abraçaram e apoiaram nesta jornada. Obrigada, pois sem a
ajuda de vocês, não teríamos alcançado resultados excelentes. Ao professor Dr. Fabrício
Baccaro, pela sua grande ajuda nas análises estatísticas do trabalho. Obrigada pelos
esclarecimentos quanto à escolha do melhor teste a ser utilizado e pela sua gentileza e
paciência em nos ajudar.
À antiga família LACA: Carlos Schneider, Natália Carvalho, Erika Milena, José
Paulo, Vanessa Figliuolo, Sabrina Araújo, Sabrina Mitoso, Marcos Andrei, Phamela Barbosa,
Leonardo Goll, Édika Sabrina, Masseo Egídio e Ingrid Cândido. Obrigada pela incrível e
grande ajuda durante minhas coletas e procedimentos laboratoriais do Doutorado, com certeza
não teria dado conta sem vocês, pelos momentos vividos com cada um, pelos bate-papos

10
(científicos ou não), pelas brigas, brincadeiras, dificuldades, passeios e pelas festinhas fora e
dentro do laboratório. Fico feliz por ter convivido com cada geração do LACA. Além disso,
como deixar de agradecer ao super Mário Nunes? Cara sempre fechada (rs!), mas que nunca
me negou ajuda e pela sua grande paciência de sentar comigo e ensinar a mexer nos
programas computacionais. Obrigada Mário.
À família científica do LGA/INPA: Érika Milena, Alber Campos, Simone Soares, José
Souza, Leonardo Goll, Janice Quadros, Alex Viana, Leandro Marajó, Patrik Viana, Vanessa
Figliuolo, Marcelle Amorim, Ramon Favarato, Debora Lunkes e Juliana Moraes pelo
acolhimento e receptividade ao chegar. Foi uma mudança repentina, mas sem o carinho
recebido, talvez tivesse sido mais difícil. Obrigada pelo apoio, discussões, conversas,
saidinhas e pelo convívio diário. Em especial, agradeço à Érika Milena, minha parceira de
todas as horas. Obrigada pela paciência com as minhas “sofrências” e irritações, mas
principalmente por sempre estar disponível em me ajudar (desde as coletas até a correção da
tese), mana passamos por poucas e boas, heim! Não poderia deixar de agradecer ao Arlindo
Batista (famoso Lindinho), uma pessoa incrível, e que merece todo meu reconhecimento
dentro do laboratório. Nunca esquecerei seus conselhos diários e principalmente da conversa
que tivemos quando eu mais precisei, foi difícil segurar as lágrimas naquele momento, por
isso, obrigada Arlindo. Ao Conselho e Comissão de Bolsas do PPG-GCBEv/INPA, dos quais
fiz parte por dois anos consecutivos. Obrigada Jaqueline Batista, Vera Val, Eliana Feldberg,
Gislene Zilse e Luciana Fé pelo aprendizado, reuniões, conversas, risos e comemorações
durante este período. Em especial, agradeço demais à Lu (Luciana Fé), por ser tão querida.
Obrigada Luzinha pelas nossas conversas, missões e conselhos. E mais, obrigada ainda por ter
assumido sozinha nesta reta final o cargo de Representante em virtude da minha ausência.
Aos amigos que ganhei, perdi e aos que permaneceram firmes e fortes comigo nessa
trajetória árdua. Vocês me proporcionaram momentos incríveis e estiveram do meu lado nos
momentos mais difíceis. Obrigada em especial à Sandra Hernandez, Rafael Ferreira, Andréia
Santa Rita, Janaína Brasil, Fabiola Viegas, Érika Milena, Louise Matos, Thamires Alvarenga,
Israela de Souza, Alber Campos, Paola Rodrigues, Priscila Azarack, Maricele Fonseca, Joice
Farias, Janice Quadros, Emerson Garajau e Simone Soares pelas palavras de conforto,
conselhos, festas/saidinhas repentinas, apoio e broncas (que não foram poucas) quando de fato
precisei. Em especial, agradeço à Milena Ferreira por tudo, minha gratidão será eterna. Amo
vocês. Peço perdão aos que não foram citados, mas serei eternamente grata a todos!

11
Agradeço a uma pessoa em especial, que mesmo estando longe, sempre está por perto,
uma pessoa que nunca te nega nada independente da hora, lugar ou situação, como diz o
ditado: É pau pra toda obra. Victor Fonseca (Victinho) deixarei aqui uma passagem sua, pois
quando bate a saudade é ela que me conforta. “Hoje, te desejo o universo de felicidades e um
sucesso imenso na vida, uma das coisas que a gente nunca esquece é a amizade verdadeira, e
tu pra mim, és daquelas pessoas que eu só tenho a agradecer por ter conhecido. Sabe aquela
pessoa que a gente pode chegar e falar de tudo? Pois é, você é esta pessoa. Deu uma saudade
doida agora, vontade de sorrir contigo, comer kikão e de conversar besteiras, mas lembre-se,
nunca esqueço de ti, mesmo estando longe e ficando tempos sem dar sinal de vida, e é claro,
vou te perturbar para o resto da vida”. Obrigada Victinho, sinto muito sua falta!
Enfim, agradeço todas as pessoas que fazem, fizeram e as que continuam fazendo
parte da minha vida, podem ter certeza que de alguma forma vocês contribuíram para essa
grande conquista.
Obrigada!

12
Resumo
Os grandes avanços e modificações urbanas em Manaus, principal cidade da
Amazônia ocidental brasileira, começaram em 1970 com a implantação de um Polo Industrial
conhecido como Zona Franca de Manaus. Esse acelerado processo urbanístico levou à
ocupação de áreas urbanas impróprias, como margens de igarapés, que hoje, servem como
depósitos de lixo e recebem efluentes industriais e hospitalares. Várias destas substâncias
lançadas diariamente nos igarapés urbanos são capazes de ocasionar danos teciduais, no
material genético de células somáticas e germinativas, além da bioacumulação de
contaminantes na biota aquática. Desta forma, analisamos os efeitos genotóxicos e
investigamos a localização cromossômica de sequências de DNA repetitivo em peixes
provenientes de diferentes ambientes poluídos, visando inferir sobre eventos que propiciam a
permanência destes organismos nos igarapés urbanos na cidade de Manaus. Para tanto, foram
estudados (i) os perfis limnológicos abióticos, (ii) efeitos genotóxicos, (iii) perfis de metilação
e a (iv) localização física cromossômica de sequências de DNA repetitivo em seis espécies de
peixes amazônicas e duas exóticas, de três locais com níveis mais críticos de poluição
ambiental em Manaus (AM): igarapé do Mindu, igarapé do Quarenta e igarapé do Tarumã-
açu. Como controle negativo, as mesmas espécies foram coletadas no lago Catalão, localizado
na confluência dos rios Negro e Solimões. No geral, os resultados demonstram que análise de
micronúcleos, anormalidades nucleares eritrocíticas e o ensaio do cometa são biomarcadores
importantes para avaliar a saúde do ecossistema aquático, especialmente em espécies que
estão apresentando remodelamento genético e epigenético, em função da péssima qualidade
da água dos igarapés urbanos poluídos na cidade de Manaus-AM. Além disso, as espécies que
ocorrem nesses locais apresentam sequências ribossomais altamente complexas no genoma,
mediada por um aumento sistemático no número de cópias do DNA ribossomal.
Adicionalmente, o ambiente poluído pode influenciar na metilação do DNA, que teria
evoluído, inicialmente, como um mecanismo de defesa dos organismos contra a expressão de
elementos transponíveis. O aumento na quantidade de elementos transponíveis do tipo Rex
nos cromossomos, evidenciados neste estudo, representa sua influência na modelagem do
genoma das populações que vivem no ambiente poluído, onde a estrutura, organização e
função do genoma são modificadas em indivíduos expostos à poluição ambiental. Isso,
provavelmente, se deve às interações com os elementos transponíveis, os quais devem estar
sob constante evolução adaptativa e respondem à mudança do ambiente de forma particular
para cada espécie.

13
Abstract
Major advances and urban modifications in Manaus, the main city of the western
Brazilian Amazon, began in 1970 with the establishment of an Industrial Pole known as Free
Zone of Manaus. This accelerated urbanization process led to the occupation of improper
urban areas, such as streams, which today serve as garbage dumps and receive industrial and
hospital effluents. Several of these substances released daily in urban streams are capable of
causing tissue damage in somatic and germ cell genetic material, as well as bioaccumulation
of contaminants in the aquatic biota. Thus, we analyzed the genotoxic effects and investigated
the chromosomal location of repetitive DNA sequences in fish from different polluted
environments, aiming to infer events that favor the permanence of these organisms in urban
streams in the city of Manaus. To this end, we studied (i) abiotic limnological profiles, (ii)
genotoxic effects, (iii) methylation profiles and (iv) location of repetitive DNA sequences in
six species of Amazonian fish and two species of exotic fish from three sites with more
critical levels of environmental pollution in the city of Manaus (AM): Mindu stream,
Quarenta stream and Tarumã-açu stream. As a negative control, the same species were
collected in Catalão Lake, located at the confluence of the Negro and Solimões rivers.
Overall, the results show that micronucleus analysis, erythrocyte nuclear abnormalities and
the comet assay are important biomarkers for assessing the health of the aquatic ecosystem,
especially in species that are exhibiting genetic and epigenetic remodeling due to poor water
as a function of the water quality of the urban streams in the city of Manaus-AM, who suffer
from environmental pollution. Additionally, the polluted environment may influence DNA
methylation, which would have initially evolved as a defense mechanism of organisms
against expression of transposable elements. The increase in the amount of Rex transposable
elements on the chromosomes, evidenced in this study, represents its influence on the genome
modeling of populations living in the polluted environment, where the structure, organization
and function of the genome are modified in individuals exposed to environmental pollution.
This is probably due to interactions with the transposable elements, which must be under
constant adaptive evolution and respond to changing environments in a particular way for
each species.

14
SUMÁRIO
1. Introdução ............................................................................................................................. 22
1.1 Poluição dos igarapés: desafios e potencialidades na cidade de Manaus-AM.................................22
1.2 Implicações da interação entre ictiofauna e ambientes aquáticos urbanos......................................25
1.3 Como os peixes são capazes de habitar ambientes hostis, sejam eles naturais ou modificados pelo
homem?...................................................................................................................................................27
2. Objetivos ............................................................................................................................... 30
2.1 Geral ................................................................................................................................................30
2.2 Específicos ...................................................................................................................................... 30
3. Material e Métodos ............................................................................................................... 31
3.1 Área de estudo e amostragem...........................................................................................................31
3.2 Métodos ........................................................................................................................................... 34
3.2.1 Perfis limnológicos abióticos da água...........................................................................................34
3.2.2 Marcadores de genotoxicidade......................................................................................................34
3.2.3 Análise estatística..........................................................................................................................36
3.2.4 Obtenção de cromossomos mitóticos............................................................................................37
3.2.5 Detecção das regiões organizadoras de nucléolo...........................................................................38
3.2.6 Detecção da heterocromatina constitutiva.....................................................................................38
3.2.7 Extração de DNA...........................................................................................................................38
3.2.8 Amplificação de sequências de DNA por Reação em Cadeia da Polimerase...............................30
3.2.9 Sequenciamento dos fragmentos de DNA.....................................................................................40
3.2.10 Análise de reconstrução filogenética...........................................................................................41
3.2.11 Marcação por nick translation e Hibridização in situ por fluorescência.....................................41
3.2.12 Análise cariotípica.......................................................................................................................43
3.2.13 Polimorfismo Amplificado Sensível à Metilação........................................................................44
4. Resultados e discussão..........................................................................................................45
4.1 Artigo 1 - Efeitos da poluição ambiental no rDNAome de peixes amazônicos...............................46

15
4.1.1 Resumo..........................................................................................................................................47
4.1.2 Introdução......................................................................................................................................48
4.1.3 Material e metódos.........................................................................................................................48
4.1.4 Resultados......................................................................................................................................52
4.1.5 Discussão.......................................................................................................................................60
4.2 Artigo 2 - Ensaio HpaII/MspI-PCR para medir a metilação do DNA em Hoplosternum littorale
(Callichthyidae, Siluriformes) de ambiente poluído na bacia central da Amazônia..............................63
4.2.1 Resumo..........................................................................................................................................64
4.2.2 Introdução......................................................................................................................................65
4.2.3 Material e metódos.........................................................................................................................66
4.2.4 Resultados e Discussão..................................................................................................................70
4.3 Artigo 3 - Elementos transponíveis de DNA em peixes amazônicos: do aumento no genoma à
adaptação genética a ambientes estressantes..........................................................................................77
4.3.1 Resumo..........................................................................................................................................78
4.3.2 Introdução......................................................................................................................................79
4.3.3 Material e metódos........................................................................................................................80
4.3.4 Resultados......................................................................................................................................82
4.3.5 Discussão.......................................................................................................................................88
4.4 Artigo 4 -Diversidade genética e dinâmica de elementos repetitivos em peixes exóticos
introduzidos na região Amazônica.........................................................................................................91
4.4.1 Resumo..........................................................................................................................................92
4.4.2 Introdução......................................................................................................................................93
4.4.3 Material e metódos.........................................................................................................................94
4.4.4 Resultados......................................................................................................................................98
4.4.5 Discussão.....................................................................................................................................101
4.5 Artigo 5 - Contaminantes ambientais e seus impactos na integridade genética de espécies de peixes
na bacia Amazônica..............................................................................................................................105
4.5.1 Resumo........................................................................................................................................106
4.5.2 Introdução....................................................................................................................................107
4.5.3 Material e metódos.......................................................................................................................108

16
4.5.4 Resultados....................................................................................................................................113
4.5.5 Discussão.....................................................................................................................................125
5. Conclusão............................................................................................................................130
6. Referências..........................................................................................................................131

17
Lista de Tabelas
Introdução
Tabela 1: Espécies de peixes analisadas e seus respectivos locais de coleta na cidade de
Manaus-AM. As espécies com asterisco são as não-nativas da bacia
amazônica..................................................................................................................................32
Artigo 1
Tabela 1: Espécies analisadas e seus respectivos locais de coleta na cidade de Manaus-
AM............................................................................................................................................50
Tabela 2: Caracterização cariotípica das espécies coletadas nos igarapés da cidade de
Manaus-AM..............................................................................................................................56
Tabela 3: Panorama comparativo das sequências repetitivas de DNAr 18S e 5S observadas
em cromossomos mitóticos de cinco espécies de peixes amostradas em ambiente poluído e
não poluído na região de Manaus-AM......................................................................................58
Artigo 3
Tabela 1: Espécies analisadas e seus locais de coleta em ambientes poluídos e não poluídos,
sendo: a) igarapé do Mindu [3 05 22.0 S, 60 0 5.31 W]; b) igarapé do Quarenta [3 08 19.7 S,
60 0 56.7 W]; c) igarapé do Tarumã-açu [3 00 51.1 S, 60 3 33.2 W]; d) lago do Catalão [3 13
22.3 S, 59 55 14.5 W]...............................................................................................................81
Tabela 2: Panorama comparativo da distribuição das sequências de elementos transponíveis
Rex 1, Rex 3 e Rex 6 em cromossomos mitóticos de três espécies de peixes amostradas de
ambiente poluído e não poluído na região de Manaus-AM......................................................87
Artigo 4
Tabela 1: Espécies de peixes exóticas analisadas e seus respectivos locais de coleta na cidade
de Manaus-AM.........................................................................................................................96
Artigo 5
Tabela 1: Espécies de peixes analisadas e seus respectivos locais de coleta.........................111
Tabela 2: Caracterização dos perfis físico-químicos dos igarapés urbanos poluídos de
Manaus-AM nos períodos de seca e cheia e do ambiente natural, lago Catalão....................114
Tabela 3: Análise multivariada de variância permutacional comparando os locais..............115

18
Tabela 4: Média e desvio padrão de microúcleos (MN) por indivíduo para cada espécie
analisada e seu respectivo local de coleta na cidade Manaus-AM..........................................117
Tabela 5: Valor de p comparando anormalidades nucleares eritrocíticas (ANEs) entre os
igarapés analisados e entre as espécies utilizadas no presente estudo....................................119
Tabela 6: Média e desvio padrão da contagem dos níveis de cometa por indivíduo para cada
espécie analisada e seu respectivo local de coleta na cidade Manaus-AM.............................122
Tabela 7: Valor de p comparando o número de cometas entre os igarapés analisados e entre
as espécies utilizadas no presente estudo................................................................................123

19
Lista de Figuras
Introdução
Figura 1: Bacias hidrográficas da cidade de Manaus-AM. .....................................................24
Material e métodos
Figura 2: Imagem ilustrativa das espécies de peixes amazônicas e exóticas analisadas neste
estudo........................................................................................................................................33
Artigo 1
Figura 1: Árvore de Inferência Bayesiana derivada da análise das sequências do gene
mitocondrial (COI) de Hoplias malabaricus (Erythrinidae; Characiformes). A probabilidade
posterior e os valores de bootstrap são mostrados ao lado dos nós. A identidade do indivíduo e
sua localização geográfica é mostrada no final de cada ramo..................................................53
Figura 2: Distribuição da heterocromatina nos cariótipos das espécies analisadas de ambiente
poluído e não poluído, em destaque os cromossomos portadores da RON (caixas), sendo m =
metacêntrico; sm = submetacêntrico; st = subtelocêntrico; a = acrocêntrico. Barra de escala
igual a 10 μm.............................................................................................................................57
Figura 3: Cariótipos das espécies de ambiente poluído e não poluído na região de Manaus,
evidenciando sítios de DNAr 18S (sinal vermelho) e de DNAr 5S (sinal verde). Os
cromossomos foram contracorados com DAPI. Barra de escala igual a 20
μm.............................................................................................................................................59
Artigo 2
Figura 1: Mapa de restrição virtual preparado na ferramenta Bioedit para a detecção dos
sítios CCGG na sequência do DNAr 18S disponível na base de dados do NCBI (números de
acesso: AF021880.1).................................................................................................................68
Figura 2: Perfis esperados (interpretações) do efeito de clivagem por MspI e HpaII. O
esquema mostra exemplos de amostras de DNA com controle negativo (DNA não digerido) e
controle positivo (DNA digerido) para ambas as enzimas.......................................................69
Figura 3: Cariótipo de Hoplosternum littorale evidenciando os sítios de DNAr 18S (par 5)
(caixas) e sítios de DNAr 5S (pares 23 e 27). (a) Ambiente não poluído (lago Catalão); (b)

20
ambiente poluído (igarapés do Mindu e Quarenta). Barra de escala igual a 20 μm. Fonte: Silva
et al. 2016..................................................................................................................................71
Figura 4: Digestão de amostras de DNA com enzimas MspI e HpaII utilizando DNA
genômico como controle da digestão enzimática. Onde A: lago Catalão; B: igarapé do
Quarenta; C: igarapé do Mindu.................................................................................................72
Figura 5: Perfil de amplificação das sequências de DNAr 18S de espécimes de Hoplosternum
littorale dos igarapés do Quarenta e Mindu (poluídos) (B; C respectivamente e lago Catalão
(não poluído) A) na cidade de Manaus, Amazonas (Brasil) tratados com as enzimas
isosquizoméricas MspI e HpaII. C = amostra de controle não digerida. L = 100 pb escada
molecular (Thermo Fisher Scientific).......................................................................................73
Artigo 3
Figura 1: Cariótipo de Pterygoplichthys pardalis de indivíduos do ambiente poluído e não
poluído da região de Manaus evidenciando sítios de Rex 1 (sinal verde), Rex 3 (sinal
vermelho) e Rex 6 (sinal verde). Os cromossomos foram contracorados com DAPI. Barra de
escala igual a 20 μm..................................................................................................................84
Figura 2: Cariótipo de Semaprochilodus insignis de indivíduos do ambiente poluído e não
poluído da região de Manaus, evidenciando sítios de Rex 1 (sinal verde), Rex 3 (sinal
vermelho) e Rex 6 (sinal verde). Os cromossomos foram contracorados com DAPI. Barra de
escala igual a 20 μm..................................................................................................................85
Figura 3: Cariótipo de Cichlasoma amazonarum de indivíduos do ambiente poluído e não
poluído da região de Manaus, evidenciando sítios de Rex 1 (sinal verde), Rex 3 (sinal
vermelho) e Rex 6 (sinal verde). Os cromossomos foram contracorados com DAPI. Barra de
escala igual a 20 μm..................................................................................................................86
Artigo 4
Figura 1: Cariótipos das espécies Oreochromis niloticus (igarapés do Mindu e Tarumã-açu)
(A, B, C e D) e Trichopodus trichopterus (igarapés do Mindu e Quarenta) (E, F, G e H). (A e
E) Padrão de distribuição da heterocromatina; (A1 e E1) Cromossomos portadores da RON;
(B e F) Sítios de DNAr 18S (sinal vermelho) (B1: Padrão dos indivíduos do igarapé do
Mindu) e de DNAr 5S (sinal verde); (C e G) Localização física cromossômica do
retroelemento Rex 1 (sinal verde); (D e H) Localização física cromossômica do retroelemento

21
Rex 3 (sinal vermelho). Os cromossomos foram contracorados com DAPI. Barra de escala
igual a 20 μm...........................................................................................................................100
Artigo 5
Figura 1: Bacias hidrográficas da cidade de Manaus-AM. Marcação em azul (igarapé do
Tarumã-açu); marcação em vermelho (igarapé do Mindu) e marcação em amarelo (igarapé do
Quarenta).................................................................................................................................110
Figura 2: Classificação do ensaio do cometa de acordo com Hartmann (1995) e Brianezi
(2011)......................................................................................................................................113
Figura 3: Análise de componentes principais (ACP) das variáveis ambientais (pH,
temperatura, OD, CE e turbidez) dos igarapés do Mindu, Quarenta, Tarumã-açu e lago
Catalão....................................................................................................................................115
Figura 4: Esfregaço sanguíneo de Pterygoplichthys pardalis corado com Giemsa
evidenciando células eritrocíticas normais (A) e danos genotóxicos (B, C, D, E, F). As setas
evidenciam: Célula micronucleada (B); Núcleo em forma de rim (C); Núcleo lobado (D);
Núcleo segmentado (E); Núcleo vacuolado (F). Barra: 10 μm...............................................116
Figura 5: Frequências (%) de micronúcleos (MN) em eritrócitos de Hoplias malabaricus,
Semaprochilodus insignis, Pterygoplichthys pardalis, Oreochromis niloticus e Trichopodus
trichopterus com seus respectivos locais de coleta na cidade de Manaus-AM......................118
Figura 6: Gráfico de barras mostrando as médias de contagem de tipos de anormalidades
nucleares eritrocíticas (ANEs) entre os igarapés e entre as espécies coletas no ambiente
poluído e não poluído de Manaus-AM...................................................................................120
Figura 7: Níveis de cometas encontrado em Oreochromis niloticus (mesmo padrão
encontrado para Pterygoplichthys pardalis, Semaprochilodus insignis, Hoplias malabaricus e
Trichopodus trichopterus) evidenciando a classificação nas quatro categorias (cometa 1,
cometa 2, cometa 3 e cometa 4). Barra: 10 μm.......................................................................121
Figura 8: Gráfico de barras comparando as médias para cada categoria de cometa entre os
ambientes (poluído e não poluído) e entre as espécies utilizadas no presente estudo. Eixo x:
classificação dos cometas (X1: nível 1 ; X2: nível 2; X3: nível 3; X4: nível 4; eixo y: número
de cometas...............................................................................................................................124

22
1. Introdução
1.1 Poluição dos igarapés: desafios e potencialidades na cidade de Manaus-AM
A poluição aquática, de modo geral, passou a receber maior atenção quando foi
possível perceber consequências adversas em seus ecossistemas, bem como em seus
organismos (Fent 2004). A partir deste momento surgiu um interesse global em relação às
questões referentes à poluição de ambientes aquáticos, embora ainda hajam diversos países
produzindo cargas enormes de poluentes ambientais (Tang et al. 2014; Liu et al. 2018). É
notório que as atitudes comportamentais do homem, desde que se tornou parte dominante dos
sistemas, têm demostrado tendência em sentido contrário à manutenção do equilíbrio
ambiental (Sunjog et al. 2013).
No Brasil, concentra-se a maior biodiversidade do mundo, distribuída em diferentes
biomas (Lewinsohn e Prado 2005). Parte significante da biota aquática está presente nos
ecossistemas amazônicos, que apresenta uma densa rede hídrica constituída por inúmeros rios
e incontáveis igarapés (pequenos corpos d’agua) (Sioli 1984). Porém, nos últimos anos, a
região amazônica sofreu um grande avanço no desenvolvimento urbano, aumentando
significativamente a população de suas grandes metrópoles (Kuck 2013). E, com isso, as
cidades se voltaram para o desenvolvimento tecnológico, gerando outros problemas
associados com o crescimento populacional desordenado (Nelson e Lucivaldo 2011), dentre
os quais destaca-se a perda das características naturais dos ecossistemas aquáticos urbanos e
da dinâmica/estrutura de suas comunidades bióticas (Omar et al. 2015).
Em Manaus, principal cidade da Amazônia ocidental brasileira, seus grandes avanços
e modificações começaram no ano de 1970, com a implantação de um Polo Industrial
conhecido como Zona Franca de Manaus. Esse acelerado processo urbanístico, tanto
relacionado ao aumento de indústrias como da população, levou à ocupação de áreas urbanas
impróprias, tais como as margens dos igarapés (Bentes 2005). Consequentemente, a ausência
de políticas públicas efetivas resultou em impactos ambientais, destacando-se a retirada da
vegetação, despejo de esgotos domésticos e lançamento de efluentes industriais nos corpos
hídricos. Além disso, os igarapés urbanos consistem em depósitos de lixo propriamente dito e
receptores diretos de efluentes industriais. Isso em conjunto com elevadas taxas de
assoreamento decorrentes da retirada da mata ciliar, agravam o grau de poluição desses
igarapés, uma vez que todo material acumulado nas margens acaba se degradando de forma
errônea (Ferreira et al. 2012; Pio et al. 2014).

23
Em sua área urbana, a cidade é atravessada por três bacias hidrográficas (Figura 1).
Uma encontra-se inteiramente urbanizada e compreende as microbacias do igarapé do São
Raimundo e do Educandos, tendo como principais tributários os igarapés do Mindu e
Quarenta, respectivamente, que drenam áreas densamente habitadas e com atividades
domésticas (lixo e esgoto sanitário) e industriais em seu entorno, desde suas nascentes até sua
foz (Pinto et al. 2009; Lemos 2016). As demais bacias drenam principalmente a área rural,
tais como as microbacias do Tarumã-açu e do Puraquequara (Pinto et al. 2009; Ferreira et al.
2012). Além disso, o aterro sanitário da cidade encontra-se situado nas margens do igarapé do
Tarumã-açu, com armazenamento inapropriado de altas quantidades de lixo, provenientes do
Polo Industrial, lixo urbano e hospitalar (Do Valle 1998; Piedade et al. 2014).
Esse armazenamento de lixo tem sido apontado como a grande causa da poluição do
solo e dos corpos d’água, tanto subterrâneas quanto superficiais (Nelson e Lucivaldo 2011).
Ainda, uma característica deste aterro sanitário é a produção de chorume, um líquido
resultante da atividade enzimática sobre a matéria orgânica, que se destaca por escoar
livremente no sistema hídrico, transportando metais como Zinco (Zn), Cromo (Cr), Manganês
(Mn), Níquel (Ni), Cobalto (Co), Cobre (Cu), Chumbo (Pb) e Ferro (Fe), além de ser um
contaminante causador de efeitos adversos (Filgueiras et al. 2004). A presença de metais
representa um grande perigo à saúde de todo ecossistema aquático, pois não são degradáveis e
apresentam um grande potencial bioacumulativo quando lançados em grandes quantidades.
Isso se torna ainda mais preocupante com relação ao consumo dos peixes, que são utilizados
como fonte de alimentação por populações humanas, e consiste na principal fonte proteica dos
manauaras (Nuvolari et al. 2011).
Além do aumento na concentração de metais tóxicos, as atividades antropogênicas
podem causar diversos impactos ambientais, estando associadas com o aumento da
condutividade elétrica, variação da temperatura da água e diminuição dos valores de oxigênio
dissolvido, quando comparados com ambientes naturais (Couceiro et al. 2007; Martins et al.
2008). Uma vez que os organismos estão em interação com o meio em que vivem é possível
observar alterações na comunidade e distribuição das espécies, pois as mais sensíveis a essas
mudanças tendem a deixar este ambiente (Oliveira et al. 2010; Omar et al. 2015; Kroon et al.
2017).
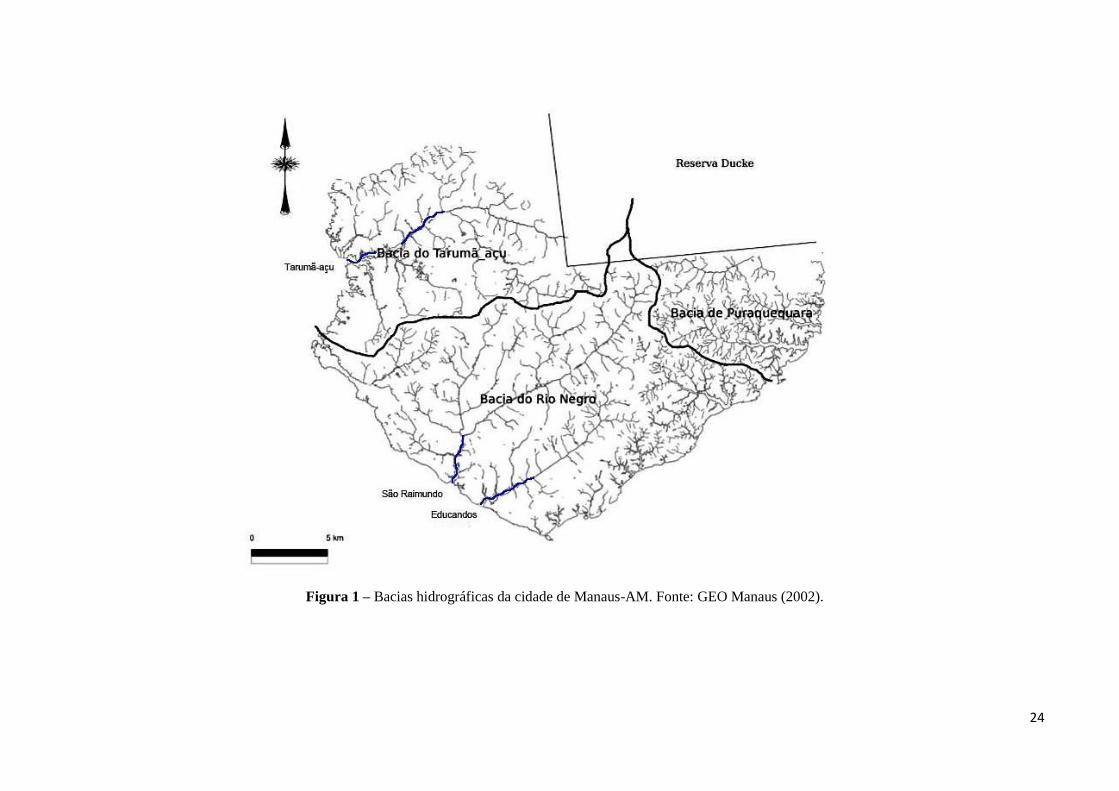
24
Figura 1 – Bacias hidrográficas da cidade de Manaus-AM. Fonte: GEO Manaus (2002).

25
Dessa forma, a maior ameaça à biota aquática de igarapés urbanos da cidade de
Manaus-AM é a perda da qualidade da água e a modificação de habitats que vem sendo
ocasionada por ações antrópicas (Viana 2018). No entanto, a maior parte dos estudos
disponíveis para a biota aquática destes igarapés envolve aspectos com enfoque ecológico e
físico-químicos de suas águas (Sioli 1985; Catarino e Zuanon 2010; Santana 2015; 2016).
Porém, estudos que envolvam aspectos da análise de perfis limnológicos e de investigação
dos efeitos genotóxicos e genômicos/citogenômicos continuam escassos, sendo estes estudos
extremamente necessários para melhor compreender o impacto e os processos adaptativos da
biota aquática nestes ambientes.
1.2 Implicações da interação entre ictiofauna e ambientes aquáticos urbanos
A geração de esgotos, resíduos sólidos e efluentes industriais é inerente às sociedades
organizadas. Tais dejetos, bem como seus subprodutos, precisam ser tratados para minimizar
os impactos de seus descartes à saúde ambiental (Siqueira et al. 2009; Gouveia 2012). De
acordo com Rutherford et al. (2000) e Sisinno (2002), para evitar que efluentes descartados
rotineiramente nos corpos d’água causem efeitos tóxicos à biota aquática e à saúde humana,
faz-se necessário que seu controle incorpore também a realização de bioensaios, ou seja, uma
avaliação de biomarcadores de toxicidade e de controle da poluição.
Para tanto, os bioensaios buscam identificar interações entre poluentes presentes no
ambiente e em organismos vivos por meio de biomarcadores (Arias 2008; Mannarino et al.
2013). Estes são capazes de detectar precocemente efeitos da contaminação ambiental sobre
os organismos, antevendo possíveis impactos sobre a saúde humana. Mas é necessário esperar
até que a pressão da poluição deixe a marca no nível do ecossistema? A presença de
estressores no ambiente (principalmente poluentes orgânicos e inorgânicos) pode influenciar a
integridade das moléculas de DNA nos organismos aquáticos, podendo ter consequências nos
níveis individuais e populacionais (Kolarevic et al. 2016).
Substâncias que são lançadas diariamente nos corpos d’água de igarapés urbanos
amazônicos podem até não demonstrar efeitos agudos nos organismos que estão expostos,
como no caso de poluição severa que pode resultar no desaparecimento de tais organismos.
Entretanto, a exposição à poluição em curto prazo é capaz de ocasionar danos teciduais, danos
no material genético de células somáticas e germinativas, além da bioacumulação de
contaminantes (Zapata et al. 2016; Thellmann et al. 2017).

26
Testes com organismos indicadores para toxicidade e genotoxicidade estão sendo
realizados constantemente e se tornaram indispensáveis por fornecer alertas precoces para
alterações e degradações ambientais, induzidas por agentes genotóxicos (Pawar 2012), já que
análises físico-químicas identificam apenas a presença e as respectivas concentrações dos
diferentes poluentes (Matsumoto et al. 2006). Dentre as metodologias utilizadas para avaliar
danos causados por substâncias xenobióticas, destacam-se o teste do micronúcleo, análise de
anormalidades nucleares e o ensaio do cometa (Bombail et al. 2001; Rocha et al. 2009;
Barbosa et al. 2010; Zapata et al. 2016).
O teste do micronúcleo é considerado uma técnica vantajosa e simples, sendo possível
observá-los em todos os tipos de eritrócitos, como resposta complexa entre a exposição
genotóxica e a eficiência dos mecanismos fisiológicos de defesa dos organismos (Hayashi
2016). Os micronúcleos (MN) são denominações dadas a pequenos corpúsculos de cromatina
que se encontram dispersos no citoplasma de eritrócitos, sendo originados de fragmentos
cromossômicos ou cromossomos inteiros que não foram incorporados nos núcleos das
células-filhas durante a divisão celular (Schmid 1975; Obiakor et al. 2012).
Em ambientes naturais da região amazônica, como no rio Madeira, a contaminação por
mercúrio (Hg) foi avaliada pelo teste do micronúcleo em eritrócitos de três espécies de
diferentes níveis tróficos: Prochilodus nigricans Spix & Agassiz 1829 (detritívoro),
Mylossoma duriventris (Cuvier, 1818) (onívoro) e Hoplias malabaricus (Block 1794)
(piscívoro), onde foi possível observar altas frequências de micronúcleos em H. malabaricus,
indicando uma interação de fatores ambientais (hábitos não migratórios) e biológicos (danos
genéticos) (Porto et al. 2005). Deste modo, a exposição a agentes genotóxicos pode resultar
em um funcionamento incorreto da maquinaria celular, fenômeno este que pode levar ao
desenvolvimento de um processo cancerígeno ou até mesmo a morte celular, e caso não
ocorra o reparo do DNA danificado, consequências biológicas em suas células, órgãos ou
organismo inteiro pode ser ocasionado (Lee e Steinert 2003; Bombassaro 2007; Kolarevic et
al. 2016).
Esse tipo de teste, em combinação com as análises de anormalidades nucleares, tem
sido extensivamente aplicado entre os ensaios atualmente disponíveis, além de serem
amplamente recomendados para estudos de biomonitoramento ambiental empregando os
peixes como bioindicadores, com intuito de avaliar a qualidade de recursos hídricos e os
efeitos da poluição sobre os organismos (Pantaleão 2008; Hussain et al. 2018). Essas
anormalidades nucleares são formadas quando determinada quantidade de material genético

27
fica levemente atrasada na mitose, fazendo com que o núcleo resultante não seja oval, mas
apresente uma saliência de cromatina, conhecida como anormalidades nucleares eritrocíticas
(ANEs) (Souza e Fontanelli 2006; Kirschbaum et al. 2009; Braham et al. 2017).
Outro teste utilizado como ferramenta na avaliação genotóxica é o ensaio do cometa,
que avalia danos no DNA a nível de células individuais, resultantes da exposição a um agente
genotóxico, medindo a migração do DNA de células em gel de agarose, numa corrida de
eletroforese (Singh et al. 1988; Hussain et al. 2018). Este ensaio tem sido recomendado na
avaliação de despejos industriais, domésticos e agrícolas, indução de danos e reparo no DNA,
assim como em estudos que visam o biomonitoramento de populações expostas a aplicações
clínicas (Collins 2014; Lapuente et al. 2015). Ainda, é adequado para o monitoramento
ambiental, analisando peixes em seu próprio ambiente. Contudo, até o momento, não existem
informações acerca de danos no material genético de peixes relacionados a agentes
genotóxicos, expostos à poluição nos igarapés urbanos de Manaus-AM, tampouco de como
eles são capazes de lidar com ambientes hostis.
1.3 Como os peixes são capazes de habitar ambientes hostis, sejam eles naturais ou
modificados pelo homem?
Na Amazônia, supõe-se que a variabilidade de ambientes favoreceu o estabelecimento
de uma grande diversidade de peixes, os quais possuem uma forte relação com o ambiente em
que vivem (Dagosta e De Pinna 2019). Assim, espécies de peixes desenvolveram, durante o
seu processo evolutivo, uma grande plasticidade genotípica e fenotípica, gerando linhagens
múltiplas, que ocasionalmente levam à especiação (Chen et al. 2015; Braz-Mota et al. 2018;
Heinrichs-Caldas et al. 2019). Atualmente admite-se que a variabilidade genômica é o fator
essencial que garante uma rápida adaptação dos organismos ao ambiente aquático (Gross et
al. 2010; Schneider et al. 2013a; Silva et al. 2016).
Populações ambientalmente impactadas têm sido alvo de investigações com o uso de
marcadores genéticos nucleares e mitocondriais, com foco na variabilidade genética, uma vez
que o declínio desta variabilidade pode reduzir a capacidade de uma população em se adaptar
às mudanças ambientais (Lande 1988; Arif e Khan 2009). O gene citocromo c oxidase
subunidade I (COI), presente no genoma mitocondrial e que codifica parte de uma enzima
terminal da cadeia respiratória da mitocôndria apresenta-se como um gene conservado
intraespecificamente, o que o torna um valioso marcador na identificação de espécies. É
comumente utilizado em estudos de níveis e padrões de distribuição da variabilidade genética

28
entre as populações, devido a suas características de herança materna, clonal e taxa
mutacional (De Carvalho et al. 2011; Machado 2016).
Além disso, trabalhos realizados com peixes amazônicos vêm associando processos
adaptativos com a variabilidade genética, principalmente a encontrada em regiões de DNAs
repetitivos, por estarem menos sujeitos a sofrerem pressão seletiva (Gross et al. 2010; Santos
et al. 2016; Silva et al. 2016). Sabe-se que no genoma de eucariotos existe uma grande
quantidade de sequências repetitivas de DNA, que diferem no número de nucleotídeos e que,
por isso, tornam-se importantes ferramentas em análises genéticas (Caixeta et al. 2006;
Suntronpong et al. 2017). Estas sequências são consideradas importantes na evolução
estrutural, dinâmica e funcional do genoma de peixes (Symonová e Howell 2018) e podem
estar organizadas de acordo com sua distribuição no genoma, sendo encontradas tanto in
tandem (como os DNAs satélites, DNAs ribossomais e as sequências teloméricas) quanto
dispersas (com destaque para os elementos transponíveis) (Kuhn et al. 2012; Suntronpong et
al. 2017).
Os DNAs ribossomais (DNAr) codificam quatro genes que são essenciais para a
função ribossômica: o RNAr 5S; 28S; 18S e 5,8S e organizam-se em duas famílias
multigênicas: 5S e 45S, estando esta última relacionada com a região organizadora de
nucléolo (RON). Uma característica intrigante destas sequências ribossomais é que, apesar de
serem consideradas conservadas (Averbeck e Eickbush 2005), o DNAr tende a alterar o seu
número de cópias (Wang et al. 2017) e a sua posição nos cromossomos das espécies
(Sochorová et al. 2018). Em condições fisiológicas normais metade das cópias dos DNAr é
inativada por meio de uma combinação de mecanismos epigenéticos, incluindo tempo de
replicação tardia (Schlesinger et al. 2009), fatores de repressão específicos (Tan et al. 2012) e
metilação do promotor do DNAr (Bacalini et al. 2014). A quantidade de trabalhos disponíveis
na literatura contendo dados citogenéticos de DNAr tem aumentado de forma constante nos
últimos anos (Sochorová et al. 2018), tanto que em peixes a localização de genes ribossomais
revela uma importância biológica dentro do genoma, o que pode resultar na formação de um
“rDNAome” consistindo em mais de 20.000 cópias empregadas em diferentes funções de
codificação extra (Symonová e Howell 2018).
Estudos com peixes amazônicos vêm detectando mudanças na localização
cromossômica e tamanho de arranjos específicos no DNAr. Esta suposição é reforçada pelo
fato de que três espécies de acará-disco (Symphysodon spp.) apresentaram variabilidade dos

29
sítios de DNAr 18S devido à presença de translocações, envolvendo regiões heterocromáticas
ricas em elementos transponíveis (TEs) (Gross et al. 2010). Ainda, Schneider et al. (2013b)
observaram múltiplos sítios de 18S e variações no padrão de 5S em algumas espécies de
ciclídeos neotropicais, sugerindo que grande parte desta variabilidade encontrada seja devido
a diferentes classes de sequências repetitivas. No trabalho de Terencio et al. (2012) também
foi possível observar múltiplos sítios de 5S em duas espécies de jaraquis do gênero
Semaprochilodus, indicando que alguns destes sítios são derivados de pseudogenes, como
consequência de duplicações e transposições de DNA pela associação destas sequências com
elementos transponíveis.
Eventualmente os TEs, que são sequências capazes de deslocar-se e inserir-se em
outros locais do genoma, têm impulsionado a evolução e a plasticidade por influenciar
diretamente na evolução da regulação transcricional (Steige et al. 2017), estabilidade
cromossômica (Farré et al. 2016) e na metilação diferencial do DNA (Grandi et al. 2015).
Estes podem ser classificados em retrotransposons (Classe I) e transposons (Classe II), com
base em seus mecanismos de transposição. Dentro de cada Classe, os TEs estão subdivididos
em ordens de acordo com seus mecanismos de inserção, estrutura e proteínas que são
codificadas; e em superfamílias conforme sua replicação; e, em famílias e subfamílias, com
base na conservação destas sequências (Chalopin et al. 2015; Lanciano e Mirouze 2018).
Em contrapartida, os TEs são vulneráveis a mudanças ambientais, o que pode
influenciar significativamente no número de cópias dentro do genoma (Cowley e Oakey 2013;
Barbosa et al. 2014), ou seja, TEs ativados por estresse podem gerar uma fonte de variação
genômica bruta e, em tempos de evolução biológica, influenciar na capacidade do organismo
de se adaptar a mudanças em seu ambiente e conquistar novos nichos ecológicos (Schaack et
al. 2010). Desta forma, realizar uma abordagem transversal à plasticidade genotípica de
peixes amazônicos, que ocorrem em ambientes totalmente contrastantes, proporciona uma
melhor visão da sua organização genômica e evolução, sendo esta abordagem de fundamental
importância para elucidar o papel dos agentes propiciadores dos mecanismos adaptativos da
ictiofauna, diante de estresses naturais ou antrópicos.

30
2. Objetivos
2.1 Geral
Analisar os efeitos genotóxicos e investigar, comparativamente, a localização
cromossômica de sequências de DNA repetitivo em peixes, provenientes de diferentes
ambientes poluídos, visando inferir sobre eventos que propiciam sua permanência nos
igarapés urbanos na cidade de Manaus, Amazonas.
2.1 Específicos
a) Identificar as espécies e verificar se há estruturação genética nos diferentes ambientes.
b) Comparar a macroestrutura cariotípica e o mapeamento de sequências de DNA
repetitivo das espécies de peixes coletadas nos igarapés poluídos em relação ao grupo
controle (não poluído).
c) Investigar o perfil de metilação do gene DNAr 18S de uma espécie de peixe que habita
ambientes inóspitos na bacia amazônica.
d) Avaliar a qualidade da água nos igarapés poluídos de Manaus-AM, comparando com
um ambiente não poluído.
e) Analisar a incidência de micronúcleo, tipos de anormalidades nucleares eritrocíticas e
danos de DNA de peixes que vivem nos dois ambientes.

31
3. Material e Métodos
3.1 Área de estudo e amostragem
Foram estudados os perfis limnológicos abióticos de três locais com níveis mais
críticos de poluição ambiental, em três igarapés na cidade de Manaus-AM. Oito espécies de
peixes foram coletadas nestes locais, sendo seis amazônicas (Pterygoplichthys pardalis,
Hoplosternum littorale, Hoplias malabaricus, Semaprochilodus insignis, Pygocentrus
nattereri e Cichlasoma amazonarum) e duas exóticas (Trichopodus trichopterus e
Oreochromis niloticus), pertencentes às ordens Siluriformes, Characiformes e Perciformes
(Tabela 1; Figura 2). As coletas ocorreram nos meses de novembro de 2015 e maio de 2016:
1- Igarapé do Mindu [3 05 22.0 S, 60 0 5.31 W]; 2- Igarapé do Quarenta [3 08 19.7 S, 60 0
56.7 W]; 3- Igarapé do Tarumã-açu [3 00 51.1 S, 60 3 33.2 W], com pontos a jusante e a
montante (três pontos por igarapé). Como controle negativo, as mesmas espécies coletadas no
ambiente poluído foram coletadas em outubro de 2017, no lago Catalão [3 13 22.3 S, 59 55
14.5 W], localizado na confluência dos rios Negro e Solimões que, por apresentar um grande
volume de água, dificulta a detecção de altas concentrações de poluentes químicos (Pinto et
al. 2009).
Os exemplares foram capturados com uso de tarrafas, redes e rapichés, sendo estes
acondicionados em recipientes, contendo a água do próprio ambiente onde foram capturados e
encaminhados vivos ao Laboratório de Citogenômica Animal (LACA) da Universidade
Federal do Amazonas (UFAM) e ao Laboratório de Genética Animal (LGA) do Instituto
Nacional de Pesquisas da Amazônia (INPA) para a realização das técnicas propostas. Todas
as coletas foram realizadas sob a permissão do Instituto Chico Mendes de Conservação da
Biodiversidade (ICMBio/SISBIO número: 7634-1/2015). Os exemplares coletados foram
eutanasiados, seguindo as Diretrizes da Prática da Eutanásia do Concea (Concea, 2013) e com
aprovação da Comissão de Ética no Uso de Animais (CEUA) (no. 011/2017) do INPA. Após
a realização dos procedimentos, os exemplares testemunho foram depositados na Coleção de
Peixes do INPA.
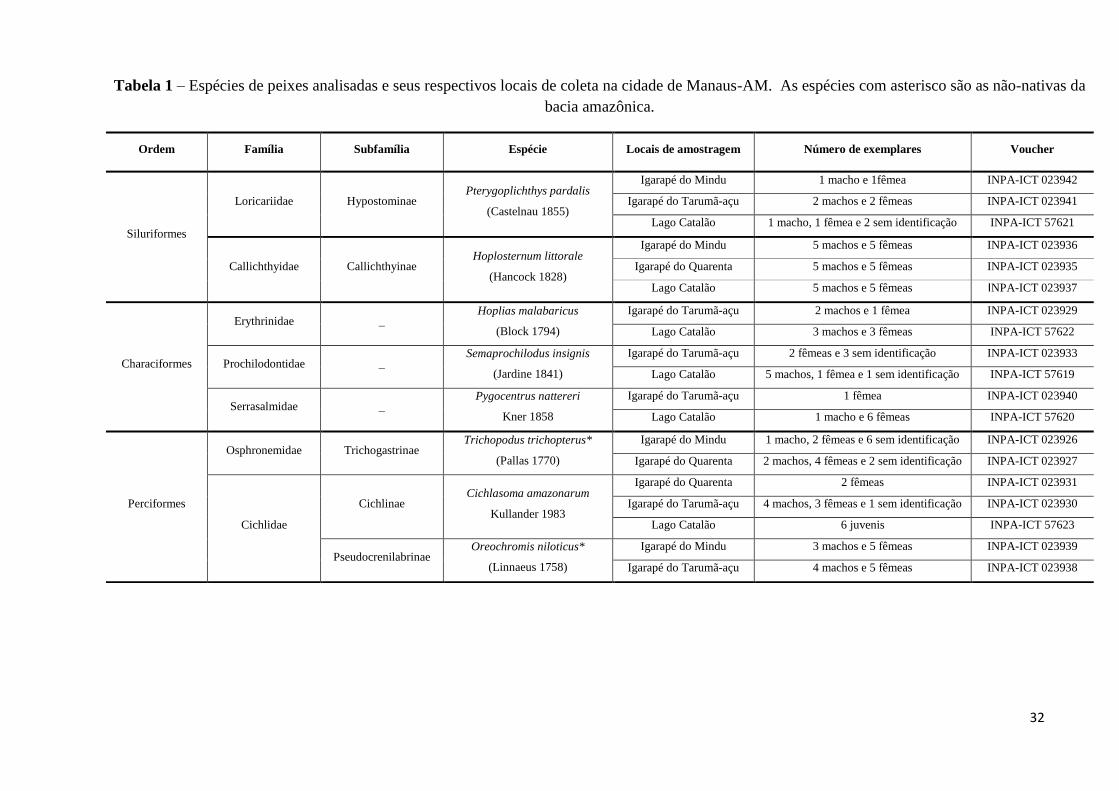
32
Tabela 1 – Espécies de peixes analisadas e seus respectivos locais de coleta na cidade de Manaus-AM. As espécies com asterisco são as não-nativas da
bacia amazônica.
Ordem Família Subfamília Espécie Locais de amostragem Número de exemplares Voucher
Siluriformes
Loricariidae Hypostominae Pterygoplichthys pardalis
(Castelnau 1855)
Igarapé do Mindu 1 macho e 1fêmea INPA-ICT 023942
Igarapé do Tarumã-açu 2 machos e 2 fêmeas INPA-ICT 023941
Lago Catalão 1 macho, 1 fêmea e 2 sem identificação INPA-ICT 57621
Callichthyidae Callichthyinae Hoplosternum littorale
(Hancock 1828)
Igarapé do Mindu 5 machos e 5 fêmeas INPA-ICT 023936
Igarapé do Quarenta 5 machos e 5 fêmeas INPA-ICT 023935
Lago Catalão 5 machos e 5 fêmeas INPA-ICT 023937
Characiformes
Erythrinidae _ Hoplias malabaricus
(Block 1794)
Igarapé do Tarumã-açu 2 machos e 1 fêmea INPA-ICT 023929
Lago Catalão 3 machos e 3 fêmeas INPA-ICT 57622
Prochilodontidae _ Semaprochilodus insignis
(Jardine 1841)
Igarapé do Tarumã-açu 2 fêmeas e 3 sem identificação INPA-ICT 023933
Lago Catalão 5 machos, 1 fêmea e 1 sem identificação INPA-ICT 57619
Serrasalmidae _ Pygocentrus nattereri
Kner 1858
Igarapé do Tarumã-açu 1 fêmea INPA-ICT 023940
Lago Catalão 1 macho e 6 fêmeas INPA-ICT 57620
Perciformes
Osphronemidae Trichogastrinae Trichopodus trichopterus*
(Pallas 1770)
Igarapé do Mindu 1 macho, 2 fêmeas e 6 sem identificação INPA-ICT 023926
Igarapé do Quarenta 2 machos, 4 fêmeas e 2 sem identificação INPA-ICT 023927
Cichlidae
Cichlinae Cichlasoma amazonarum
Kullander 1983
Igarapé do Quarenta 2 fêmeas INPA-ICT 023931
Igarapé do Tarumã-açu 4 machos, 3 fêmeas e 1 sem identificação INPA-ICT 023930
Lago Catalão 6 juvenis INPA-ICT 57623
Pseudocrenilabrinae Oreochromis niloticus*
(Linnaeus 1758)
Igarapé do Mindu 3 machos e 5 fêmeas INPA-ICT 023939
Igarapé do Tarumã-açu 4 machos e 5 fêmeas INPA-ICT 023938

33
1cm 1cm 1cm
1cm
1cm 1cm
1cm 1cm
Pterygoplichthys pardalis
(Acarí-Bodó)
Hoplosternum littorale
(Tamoatá)
Hoplias malabaricus
(Traíra)
Semaprochilodus insignis
(Jaraqui)
Pygocentrus nattereri
(Piranha vermelha)
Trichopodus trichopterus
(Gourami azul)
Oreochromis niloticus
(Tilápia do Nilo)
Cichlasoma amazonarum
(Acará)
Siluriformes
Characiformes
Perciformes
Figura 2 – Espécies de peixes amazônicas e exóticas analisadas neste estudo.

34
3.2 Métodos
3.2.1 Perfis limnológicos abióticos da água
As coletas de amostras de água foram realizadas com o auxílio de uma garrafa de Van
Dorn e armazenadas em frascos de Winkler, em duplicatas à temperatura ambiente, até a
realização das análises físico-químicas nos Laboratórios de Métodos Espectroscópicos UV-
IV-AA (LAMESP) e de Cromatografia Gasosa (LABCG) na Central Analítica da
Universidade Federal do Amazonas (UFAM) sob coordenação da Dra. Teresa Cristina Souza
de Oliveira. Parâmetros limnológicos como: temperatura (°C), pH, condutividade elétrica
(CE) e turbidez foram medidos nos locais de coleta, com uso de um pHmetro portátil (modelo
370 pH meter - JENWAY) e medidores de CE/temperatura (modelo 55-25 FT - YSI);
turbidez com o uso de turbidímetro (modelo HI98703 - HANNA), pelo método
Nefelométrico, sendo estes parâmetros analisados em triplicata. Em laboratório, foi
determinado o oxigênio dissolvido (OD) utilizando o método de Winkler. Todos os métodos
analíticos seguiram o Standard Methods 22ª Edição (APHA 2012), estando os métodos e
referências, detalhados no quadro abaixo:
Quadro 1 – Detalhamento dos parâmetros com seus respectivos métodos e referências.
Parâmetro Método Referência
OD Winkler APHA-5220-B
CE Potenciométrico APHA-2510-B
Turbidez Nefelométrico APHA-2130-B
pH Potenciométrico APHA-4500-H+
Todos os aparelhos e equipamentos foram previamente calibrados e devidamente
operados conforme seus respectivos manuais. Após as análises em triplicata, foram realizados
os cálculos da média e desvio padrão para cada local estudado e os resultados foram
confrontados com os parâmetros estabelecidos na Resolução do Conselho Nacional do Meio
Ambiente (CONAMA 410/2009 e 430/2011) (BRASIL 2005), em que se aplicam os limites
permitidos para a qualidade da água doce, segundo a classe 2 e com características físicas e
químicas de ambientes naturais da Amazônia (Da Silva et al. 2016).
3.2.2 Marcadores de genotoxicidade

35
Teste do micronúcleo (MN) e anormalidades nucleares eritrocíticas (ANEs)
No laboratório, sangue de todos os indivíduos foi coletado com o auxílio de uma
seringa com agulha, utilizando anticoagulante. Uma gota de sangue de cada indivíduo foi
distendida sobre uma lâmina limpa, com o auxílio de outra lâmina. Para cada animal foram
preparadas duas lâminas de esfregaço sanguíneo. Após sua secagem, em torno de 12 horas, as
lâminas foram então fixadas em Etanol PA (100%) por 30 minutos, secas ao ar e
posteriormente coradas com Giemsa 5% (diluído em tampão fosfato pH 6,8) por 10 minutos,
sendo lavadas posteriormente com água destilada e secas novamente ao ar para posterior
análise em microscópio óptico. Em resolução de 1000x foram contabilizados 1000 eritrócitos
por lâmina em coloração convencional e posterior anotação do número de micronúcleos e
ANEs encontrados (Bucker et al. 2006). Os micronúcleos foram identificados, seguindo o
critério proposto por Fenech et al. (2003) e Freitas et al. (2005) sendo considerados como
micronúcleos: partículas que não excediam 1/3 do diâmetro total do núcleo principal, não
estavam conectados e nem ligados ao núcleo e permaneciam no citoplasma, formato redondo
com bordas distinguíveis e mesma intensidade de coloração e refringência do núcleo
principal. A diferenciação das ANEs foi realizada de acordo com Carrasco et al. (1990) com
algumas modificações, sendo então classificadas:
Forma de rim: núcleos com uma pequena invaginação em forma de fenda bem definida, de
largura uniforme na membrana nuclear que se estende a uma profundidade apreciável para
dentro do núcleo.
Lobado: núcleos que apresentam formato irregular em consequência de evaginações, que
podem ser visualizadas como pequenas a grandes projeções da membrana nuclear, podendo
formar dois ou mais lobos conectados.
Segmentado: núcleos cuja estrutura aparece dividida em duas partes nitidamente separadas e
delimitadas pela membrana nuclear. A região compreendida entre os segmentos parece estar
destituída de material cromatínico.
Vacuolado: núcleos que apresentam uma região clara, destituída de qualquer material
cromatínico visível no seu interior.
Ensaio do cometa
Para o ensaio do cometa foi utilizada a técnica descrita por Singh et al. (1988) com
algumas modificações. Lâminas foram previamente tratadas com agarose 1,5% (normal),

36
diluída em tampão PBS e secas em temperatura ambiente, por um período de 24 horas.
Posteriormente, 10 μL de sangue de cada indivíduo (diluído em solução de RPMI) foram
misturados em 95 μL de agarose low melting point (LMP), diluída em tampão PBS (pH 7,4)
aquecida a 37 °C. Em seguida, 105 μL desta solução foi gotejada em uma lâmina preparada
com agarose 1,5% (normal) sendo coberta por uma lamínula até a solidificação.
Posteriormente as lâminas foram incubadas em solução gelada de lise (2,5M NaCl, 100mM
EDTA, 10mM Tris, 1% de Triton X-100, 10% DMSO) por pelo menos 1 hora. Logo após,
foram colocadas em posição horizontal em uma cuba de eletroforese contendo tampão de
eletroforese (300mM NaOH, 1mM EDTA e pH 13) por 20 minutos e logo em seguida, feita a
corrida de eletroforese a 25 V e 300mA por 15 minutos, seguindo os critérios propostos por
Gontijo et al. (2003). Após, as lâminas foram lavadas em solução neutralizadora (0,4M Tris,
pH 7,5), sendo este procedimento repetido por mais três vezes em intervalos de 5 minutos.
Para a coloração, as lâminas foram fixadas em Etanol PA (100%), secas ao ar e coradas com
nitrato de Prata (AgNO3) por 10 minutos. Foram contabilizados 50 eritrócitos por indivíduo,
em microscópio óptico e analisados no programa de imagens em PC de domínio público entre
plataformas para o ensaio do cometa CAPS_1.2.3b1.exe. Os danos foram classificados em
categorias, que variam de 0 a 4, referentes à extensão de dano no DNA: 0 = sem danos (<5
%); 1 = baixo nível de danos (5-20 %); 2 = médio nível de danos (20-40 %); 3 = alto nível de
danos (40-95 %); 4 = totalmente danificados (>95 %) (Hartmann 1995; Brianezi 2011).
3.2.3 Análise estatística
Para as variáveis ambientais foi realizada uma análise de componentes principais
(ACP) onde foram utilizados fatores de carregamento (Dancey e Reidy 2006) e análise de
agrupamento (Cluster) para ordenar os dados e uma PERMANOVA (análise multivariada por
permutação) para testar se existia diferença entre os igarapés baseado nas variáveis
limnológicas. Para os testes genéticos foi usado o teste de Kruskall-Wallis com comparações
múltiplas utilizando correção de Dunn, com significância estatística p<0,05 para comparar
métricas genéticas entre igarapés, sendo realizada uma análise por espécie para cada métrica
genética. Para os testes de comparação, foram executados no SigmaStat 3.5 e para elaboração
de gráficos no SigmaPlot 11.0. Todas as análises estatísticas foram realizadas utilizando o
pacote estatístico R (R Core Development Team 2016).

37
3.2.4 Obtenção de cromossomos mitóticos
Após a coleta de sangue, uma solução de fermento biológico foi aplicada em todos os
indivíduos para se obter um maior número de células em divisão (Oliveira et al. 1988). Esta
solução foi preparada com 0,3 g de fermento biológico comercial, 0,3 g de açúcar e 10 mL de
água destilada, sendo esta mantida em estufa (37 °C) por um período de 15-20 minutos, e
posteriormente injetada na região dorso-lateral dos peixes, na proporção de 1mL para cada
100 g de peso do animal, por um período de 24 horas.
As preparações citogenéticas foram obtidas seguindo-se a metodologia descrita por
Bertollo et al. (1978), com algumas modificações, que consistiram em injetar
intraperitonealmente colchicina 0,0125% na proporção de 1 mL para cada 100 g de peso do
animal. Os peixes permaneceram em aquário bem aerado por 40 minutos. Em seguida foram
sacrificados por imersão em água gelada e a porção anterior do rim, órgão hematopoiético em
peixes, foi retirada, sendo transferida para uma solução hipotônica de KCl 0,075 M (6-8 mL).
O tecido foi divulsionado com o auxílio de uma seringa de vidro e o sobrenadante (suspensão
celular) foi transferido com o auxílio de uma pipeta Pasteur para um tubo de centrífuga. A
suspensão celular obtida foi incubada em estufa a 37 °C por 30 minutos. Transcorrido este
tempo a suspensão foi pré-fixada com 6 gotas de metanol e ácido acético (3:1) e
ressuspendida. Após 5 minutos, foi adicionado fixador até encher o tubo. A suspensão foi
centrifugada por 10 minutos a 900 rpm. O sobrenadante foi desprezado e 6 mL de fixador
foram adicionados, sendo o material ressuspendido novamente e centrifugado por 10 minutos
a 900 rpm. Novamente o sobrenadante foi desprezado e 6 mL de fixador foram adicionados, e
essa lavagem foi repetida por mais duas vezes. Após a última centrifugação e eliminação do
sobrenadante, 1,5 mL de fixador foram adicionados e o material foi ressuspendido com
cuidado. Essa suspensão celular foi então estocada em microtubo e mantida em freezer para
posterior utilização.
Para a preparação das lâminas, estas foram lavadas, secas ao ar e posteriormente
imersas em água destilada a 55 ºC, em banho-maria. Após cinco minutos as lâminas foram
retiradas da água de forma a manter uma película de água sobre a sua superfície, na qual foi
gotejada a suspensão celular em diferentes regiões. As lâminas secaram diretamente ao ar.
3.2.5 Detecção das regiões organizadoras de nucléolo (RONs)
Para a detecção das regiões organizadoras de nucléolo (RONs) foi utilizada a técnica
descrita por Howell e Black (1980). Sobre as lâminas contendo suspensão celular foram

38
adicionadas 2 a 3 gotas de uma solução coloidal de gelatina (2 g de gelatina comercial sem
sabor, dissolvida em 100 mL de água destilada, acrescentando 1 mL de ácido fórmico) e,
sobre cada gota de gelatina, duas gotas de solução aquosa de nitrato de Prata (AgNO3) a 50%,
agitando-se levemente a lâmina. Esta foi coberta com lamínula e colocada em câmara úmida,
em banho-maria a 60 °C durante 3 a 8 minutos. Após o tempo apropriado, quando a lâmina
adquiriu uma coloração marrom dourada, foi lavada em água destilada, o que permitiu que a
lamínula fosse retirada naturalmente pela própria água e secas ao ar.
3.2.6 Detecção da heterocromatina constitutiva (Banda C)
Para a detecção da heterocromatina constitutiva (banda C) foi utilizada a técnica
descrita por Sumner (1972), que consistiu em tratar a lâmina preparada segundo a técnica
descrita para cromossomos mitóticos com HCl 0,2N à 42 °C por 2 minutos. Em seguida, a
lâmina foi lavada rapidamente em água destilada à temperatura ambiente e seca ao ar.
Posteriormente a mesma foi incubada por 1 minuto em solução de hidróxido de bário a 5%,
recém preparada e filtrada a 42 °C. A ação do hidróxido de bário foi interrompida, imergindo
rapidamente a lâmina em solução de HCl 0,2N (temperatura ambiente) e lavada em água
destilada. Após seca, a lâmina foi incubada em solução 2xSSC (cloreto de sódio 0,3M e
citrato trisódico 0,03M, pH 6,8) em banho-maria a 60 °C, por 15 minutos, sendo lavada em
água destilada, secas ao ar e coradas com solução de Giemsa (diluída a 5% em tampão fosfato
0,06M e pH 6,8) durante 10 minutos, lavadas em água destilada e secas ao ar.
3.2.7 Extração de DNA
O DNA genômico foi extraído do tecido muscular dos exemplares de peixes, usando o
kit Wizard® Genomic Purification (Promega), de acordo com o protocolo do fabricante e o
protocolo básico de Sambrook e Russel (2001), com algumas modificações. Para este
protocolo, os tecidos foram tratados com tampão de lise (Tris-HCl pH 8,0 em 10 mM, NaCl
0,3 M, EDTA 10 mM), posteriormente foram acrescentados: 15 μL de proteinase K (10
mg/mL) e 6 μL de RNAse A (10 mg/mL). As amostras foram incubadas para que o tecido
fosse totalmente digerido. A seguir foram feitas lavagens sucessivas com fenol clorofórmio e
clorofórmio hidratado (500 μL de cada um destes reagentes). Após a lise, o DNA foi separado
das proteínas por precipitação salina juntamente com centrifugação a 14000 rpm. O
sobrenadante foi então precipitado com 600 μL de isopropanol 100% gelado, também com o
auxílio de centrifugação. Ao final, o DNA foi hidratado com aproximadamente 100 μL de
água milli-Q, dependendo do tamanho do pellet (DNA precipitado) formado. Para possibilitar

39
a análise da quantidade e integridade do material, o DNA extraído foi quantificado por
comparação com marcador de concentração conhecida, em eletroforese padrão (com tampão
Tris-Borato-EDTA 0,5X e corrida a 70 V por 40 minutos) em gel de agarose 0,8% e corado
com GelRed Acid Gel Stain Biotium (1:500). A visualização e análise do DNA no gel foram
feitas no fotodocumentador Easy Doc 100 (BioAgency), o qual possui acoplado um
transluminador de luz ultravioleta (260 nM). Adicionalmente, quantificações em
espectrofotômetro NanoVue Plus (GE Healthcare) foram realizadas.
3.2.8 Amplificação de sequências de DNA por Reação em Cadeia da Polimerase (PCR)
Gene mitocondrial citocromo c oxidase subunidade I (COI) e sequências repetitivas de
DNA s Rex1, Rex3 e Rex6 e DNAs ribossomais 5S e 18S foram amplificados por PCR (Saiki
et al. 1988) utilizando os primers:
- COI: (FishF2 5’-GTAAAACGACGGCCAGTCGACTAATCATAAAGATATCGGCAC-
3’, FishR2 5’-CAGGAAACAGCTATGACACTTCAGGGTGACCGAAGAATCAGAA -3’,
VF2 5’-GTAAAACGACGGCCAGTCAACCAACCACAAAGACATTGGCAC-3’ e VR1d
5’- CAGGAAACAGCTATGACTAGACTTCTGGGTGGCCRAARAAYCA-3’) (Ivanova et
al. 2007);
- Rex 1: (RTX1-F1 5’TTC TCC AGT GCC TTC AAC ACC e RTX1-R3 5’TCC CTC AGC
AGA AAG AGT CTG CTC) (Volff et al. 1999, 2000, 2001a);
- Rex 3: (RTX3-F3 5’CGG TGA YAA AGG GCA GCC CTG e RTX3-R3 5’TGG CAG
ACN GGG GTG GTG GT) (Volff et al. 1999, 2000, 2001a);
- Rex 6: (Rex 6-Medf1 5’TAA AGC ATA CAT GGA GCG CCAC e Rex 6-Medf2 5’AGG
AAC ATG TGT GCA GAA TATG) em combinação com os primers (Rex 6-Medr1 5’GGT
CCT CTA CCA GAG GCC TGGG ou Rex 6-Medr2 5’GGT GGT TTC TCC TCC AAG
CTCG (Volff et al. 2001b);
- DNAr 5S: 5Sf (5’- CGA GGTCGGGCCTGGTTAGTA-3’) e 5Sr (5’-
CTTCYGAGATCAGACGAGATC-3’ (Carvalho et al. 2016);
- DNAr 18S: 18Sf (5’-CCG CTT TGG TGA CTC TTG AT-3’) e 18Sr (5’-CCG AGGACC
TCA CTA AAC CA-3’) (Gross et al. 2010).
Os ciclos de amplificação seguiram as seguintes etapas:

40
a) COI: 73 °C por 1 minuto (desnaturação inicial), 35 ciclos de 10 segundos a 93 °C
(desnaturação), 45 segundos a 51 ºC (ligação dos primers), 1 minuto a 72 ºC
(extensão); 10 minutos a 72 ºC (extensão final).
b) Rex 1, 3 e 6: 2 minutos a 95 ºC (desnaturação inicial); 35 ciclos de 1 minuto a 95 ºC
(desnaturação), 40 segundos a 55 ºC (ligação dos primers), 2 minutos a 72 ºC
(extensão); 5 minutos a 72 ºC (extensão final);
c) DNAr 5S: 1 minutos a 94 ºC (desnaturação inicial); 30 ciclos de 1 minuto a 94 °C
(desnaturação), 1 minuto a 59 °C (ligação dos primers), 1 minuto e 30 segundos a 72
°C (extensão), e 5 minutos a 72 °C (extensão final);
d) DNAr 18S: 2 minutos a 94 ºC (desnaturação inicial); 35 ciclos de 1 minuto a 95 ºC
(desnaturação), 1 minuto a 55 ºC (ligação dos primers), 1 minuto e 40 segundos a 72
ºC (extensão); 7 minutos a 72 ºC (extensão final).
Os PCRs para o gene COI foram realizados contendo 1,2 μL de dNTP (1,25 mM), 1,5
μL de tampão 10x, 1,5 μL de cada primer, 1,2 μL de MgCl2 (50 mM), 0,3 μL de DNA Taq
Polimerase (5 U / μL), 1,0 μL de DNA, e Milli-Q para completar o volume de 15 μL. Para as
sequências repetitivas foram utilizados 1 μL de DNA genômico (100 ng); 1,5 μL de Tampão
10X com cloreto de magnésio (1,5 mM); 0,15 μL de Taq DNA polimerase (5U/μL); 3,0 μL de
dNTP (1 mM); 0,6 μL de cada primer (5 mM); água milli-Q para completar o volume de 15
μL. Todas as reações foram feitas em um termociclador Eppendorf - Mastercycler Gradient.
Os produtos de PCR foram verificados e quantificados por comparação com o marcador Low
Mass DNA Ladder (Invitrogen) em eletroforese em gel de agarose 1,0% corado com GelRed
Acid Gel Stain (Biotium 1:500). A visualização e análise do DNA no gel foram feitas no
fotodocumentador Easy Doc 100 (BioAgency), o qual possui acoplado um transluminador de
luz ultravioleta (260 nM). Adicionalmente, quantificações em espectrofotômetro NanoVue
Plus (GE Healthcare) foram realizadas.
3.2.9 Sequenciamento dos fragmentos de DNA
Para confirmar e identificar produtos isolados, alguns fragmentos de DNA foram
sequenciados usando os procedimentos descritos no protocolo do Kit ABI PRISM® Big
DyeTM Terminator Cycle Sequencing Ready Reaction (Applied Biosystems). Para a
realização do sequenciamento foram utilizados entre 150 e 300 ng de DNA, 5 pmol de cada
primer em reações separadas; 4 μL do premix (kit) e água mili-Q para completar o volume de
10 μL. Essas reações foram processadas em um termociclador em 30 ciclos sendo:
desnaturação a 95 °C por 20 segundos, anelamento a 50 °C por 15 segundos e extensão a 72

41
°C por 1 minuto e 30 segundos. Depois de amplificados, os fragmentos foram submetidos a
um tratamento de precipitação para a eliminação do produto não incorporado durante a reação
de sequenciamento e logo após foi realizada a leitura automática do fragmento no
sequenciador automático ABI 3500 (Applied Biosystem) nas condições de injeção e corrida
recomendadas pelo fabricante. O alinhamento das sequências geradas foi realizado através da
ferramenta MAFFT, implementada no software Geneious 7.0.6 (Kearse et al. 2012) e editadas
manualmente. Para identificação de possíveis homologias as sequências nucleotídicas obtidas
foram submetidas a buscas online BLAST (“Basic Local Alignment Search Tool”) (Altschul
et al. 1990) através do National Center for Biotechnology Information (NCBI) (USA),
“website” (https://www.ncbi.nlm.nih.gov) com a ferramenta BLASTn. As sequências geradas
foram depositadas no banco de dados do GenBank e os números de acessos encontram-se em
cada capítulo no tópico Resultados e Discussão.
3.2.10 Análises de reconstrução filogenética
A relação entre as sequências de COI foi baseada, inicialmente, no método de
agrupamento de vizinhos, utilizando o modelo de substituição Kimura 2 parâmetros
implementado no programa MEGA 6 (Tamura et al. 2013). O modelo evolutivo para as
análises de Inferência Bayesiana (IB) e Máxima Verossimilhança (MV) foi gerado através do
programa JModeltest (Posada 2008). A análise de IB foi realizada no programa MrBayes
3.1.2 (Ronquist e Huelsenbeck 2003), com quatro cadeias de 10 milhões de gerações,
amostrada a cada 100 gerações e burnin de 25%. Análise de MV foi realizada no programa
Garli 2.0 (Zwickl 2006), com bootstrap de 1000 réplicas e as árvores obtidas foram
sumarizadas por consenso de maioria no programa SumTrees. As árvores obtidas de MV e IB
foram visualizadas e editadas no programa Figtree versão 1.4.3 (Rambaut 2016).
3.2.11 Marcação por nick translation e Hibridização in situ por fluorescência (FISH)
Os produtos de PCR referentes aos marcadores citogenéticos (Rex 1, Rex 3, Rex 6,
DNAr 5S e DNAr 18S) foram marcados pelo método de nick translation com biotina-14-
dATP (Biotin Nick Translation mix; Invitrogen) e digoxigenina11-dUTP (Dig-Nick
Translation mix; Roche), seguindo as instruções do fabricante. Os anticorpos estreptavidina-
Alexa Fluor 488 e antidigoxigenina rodamina (Roche) foram usados para detecção e
amplificação do sinal.

42
Para a técnica de FISH, utilizando sondas homologas, foram realizadas seguindo o
protocolo de Pinkel et al. (1986), com 77% de estringência (2,5 ng/μL de sonda marcada,
50% formamida, 10% sulfato dextrano, 2XSSC a 37 °C por 18h) algumas modificações:
Tratamento das lâminas
As lâminas contendo preparações cromossômicas foram lavadas em tampão PBS 1x
por 5 minutos em temperatura ambiente. Posteriormente foram desidratadas em série
alcoólicas gelada (70%, 85% e 100%) durante 5 minutos cada. Em seguida tratadas com 90
μL de RNase 10 μg/mL (5 μL de RNase 10 mg/mL e 975 μL de 2xSSC) por 1 hora em
câmara úmida a 37 ºC. Posteriormente lavadas três vezes em 2xSSC durante 5 minutos cada.
Após isso as lâminas foram lavadas em PBS 1x durante 5 minutos.
Fixação
As lâminas foram fixadas em formaldeído 1% em PBS 1x/50mM MgCl2 durante 10
minutos à temperatura ambiente. Posteriormente lavadas em PBS 1x por 5 minutos. Após,
foram desidratadas em série alcoólicas geladas (70%, 85% e 100%) durante 5 minutos cada.
Pré-hibridização
As lâminas foram desnaturadas em formamida 70% em 2xSSC a 70 ºC por 5 minutos
e novamente desidratadas em etanol gelado 70%, 85% e 100% por 5 minutos cada.
Solução de hibridização
Em um tubo eppendorf foram adicionados 5 μL de sonda, 25 μL de formamida, 10 μL
de sulfato de dextrano 50% e 5 μL de 20xSSC. A sonda então foi desnaturada a 99 °C por 10
minutos e transferida imediatamente ao gelo.
Hibridização
Foram colocados 40 μL de solução de hibridização sobre uma lamínula e a lâmina
invertida sobre a lamínula. As lâminas foram mantidas com o material voltado para baixo em
câmara úmida (H2O destilada) a 37 °C por cerca de 24 horas.
Lavagens
As lamínulas foram removidas. Em seguida as lâminas foram lavadas duas vezes em
formamida 15% a 42°C durante 10 minutos cada e lavadas novamente em solução Tween
0,5% durante 5 minutos a temperatura ambiente.

43
Detecção
As lâminas foram incubadas em tampão NFDM por 15 minutos. Após, foram lavadas
duas vezes com solução Tween 5% por 5 minutos a temperatura ambiente. Foram colocadas
sobre cada lâmina 20 µL de anti digoxigenina-rodamina e 100 µL de solução contendo
tampão NFDM e estreptavidina (2 μL de estreptavidina estoque em 998 μL de NFDM). Em
seguida foram cobertas com lamínula e deixadas por 60 minutos em câmara úmida com água
destilada. Posteriormente as lamínulas foram removidas e as lâminas lavadas três vezes em
solução Tween 5% por 2 minutos a temperatura ambiente cada e desidratadas em série
alcoólica 70, 85 e 100% durante 5 min cada.
Montagem das lâminas
Foi adicionado a cada lâmina solução de DAPI diluído em antifade VectaShield
Vector (20 μL de antifade e 1 μL de DAPI).
3.2.12 Análise cariotípica
Após a análise e contagem dos cromossomos ao microscópio óptico foi estabelecido o
número diploide modal (30 células/indivíduo). As melhores metáfases obtidas com as técnicas
clássicas (RON e banda C) foram fotografadas, com objetiva de imersão em câmera digital
Leica DFC 3000G. As lâminas submetidas à técnica molecular (FISH) foram analisadas e
capturadas em um fotomicroscópio de epifluorescência Olympus BX51, em objetiva de
imersão. Para a montagem dos cariótipos foi utilizado o programa Adobe Photoshop 7.0,
versão CS5, a partir de cromossomos metafásicos mitóticos, os quais foram recortados e
emparelhados. Todos os cromossomos foram medidos utilizando o programa livre ImageJ e
organizados em ordem decrescente de tamanho. A morfologia cromossômica foi determinada
de acordo com os critérios de relação de braços (RB=BM/Bm, onde BM = braço maior e Bm
= braço menor) segundo Levan et al. (1964). Os cromossomos foram separados em
metacêntricos (m), que apresentam RB=1,00 a 1,70, submetacêntricos (sm) RB=1,71 a 3,00,
subtelocêntricos (st) RB=3,01 a 7,00 e acrocêntricos (a) RB > 7,00. O número fundamental
(NF) foi determinado de acordo com o número de braços cromossômicos.
3.2.13 Polimorfismo Amplificado Sensível à Metilação (MSAP)
Para uma investigação sobre o perfil de metilação do gene DNAr 18S na espécie
Hoplosternum littorale, utilizada como modelo para a realização da técnica, amostras de
tecido muscular foram digeridas com enzimas de restrição MspI e HpaII, seguindo as

44
instruções do fabricante e utilizando o DNA genômico como controle da digestão enzimática.
A enzima HpaII digere sequências CCGG não metiladas ao passo que MspI digere sítios
CCGG, independentemente da ocorrência de metilação ou não. Para o preparo das reações
foram utilizados DNA genômico (50-100 ng/ μL), Tampão 1X, Enzima (1U/ μL) e água milli-
Q para completar o volume. A digestão durou cerca de 1 hora a 37 ºC, seguida de 15 minutos
a 65 ºC para interrupção da ação das endonucleases. Posteriormente, os produtos da digestão
foram comparados por eletroforese padrão (com tampão Tris-Borato-EDTA 0,5X e corrida a
70 V por 40 minutos) em gel de agarose 1,7% e corado com GelRed (Biotium).
Posteriormente, os fragmentos digeridos foram então amplificados por PCR, utilizando os
primers de acordo com Gross et al. (2010) (citados na sessão 3.2.9), sendo os produtos da
PCR visualizados novamente em gel de agarose 1,7% corados com GelRed (Biotium) com
auxílio de transluminador de luz ultravioleta (260 nM) acoplado a fotodocumentador Easy
Doc 100 (BioAgency).

45
4. Resultados e discussão
Os resultados obtidos no presente estudo, juntamente com a discussão dos dados são
apresentados na forma de cinco artigos científicos, conforme mencionados a seguir:
4.1 Artigo 1 – Efeitos da poluição ambiental no rDNAome de peixes amazônicos.
4.2 Artigo 2 – Ensaio HpaII/MspI-PCR para medir a metilação do DNA em Hoplosternum
littorale (Siluriformes, Callichthyidae) de ambiente poluído na bacia central da Amazônia.
4.3 Artigo 3 – Elementos transponíveis de DNA em peixes amazônicos: do aumento no
genoma à adaptação genética a ambientes estressantes.
4.4 Artigo 4 – Diversidade genética e dinâmica de elementos repetitivos em peixes exóticos
introduzidos na região Amazônica.
4.5 Artigo 5 – Contaminantes ambientais e seus impactos na integridade genética de espécies
de peixes na bacia Amazônica.

46
4.1 Artigo 1
Efeitos da poluição ambiental no rDNAome de peixes amazônicos

47
4.1.1 Resumo
A poluição é um problema ambiental crescente em todo o mundo e o impacto das
atividades humanas na biodiversidade e na variabilidade genética das populações naturais é
cada vez mais preocupante, uma vez que os processos adaptativos dependem dessa
variabilidade, em particular a encontrada no DNA repetitivo. No presente estudo, o DNA
mitocondrial (COI) e a distribuição de sequências repetitivas de DNA (DNAr 18S e 5S) foram
analisadas em populações de peixes que habitam águas poluídas e não poluídas na bacia
Amazônica. Os resultados indicam sequências ribossomais altamente complexas no genoma
de peixes de ambientes poluídos, pois estão envolvidas principalmente na manutenção da
integridade do genoma, mediada por um aumento sistemático no número de cópias do DNA
ribossômico em resposta a mudanças nas condições ambientais.
Palavras-chave: DNA repetitivo; gene ribossomal; heterocromatina; COI; ações antrópicas.

48
4.1.2 Introdução
O baixo rio Negro e seus afluentes no norte da bacia amazônica são usados por
populações urbanas e ribeirinhas locais para transporte, pesca e recreação, além de fornecer
água potável para a cidade de Manaus, a capital do estado brasileiro do Amazonas, metrópole
na bacia amazônica ocidental. As principais mudanças socioambientais começaram na cidade
na década de 1970, após o estabelecimento da Zona Industrial local (Couceiro et al. 2007).
Isso resultou em um processo acelerado de urbanização, envolvendo não apenas a
implantação de instalações industriais, mas também o crescimento populacional, que muitas
vezes levou à ocupação desordenada de ambientes inadequados para o desenvolvimento
urbano, como as margens de córregos (Santana e Barroncas 2007). Esses corpos d’água foram
posteriormente utilizados como lixeiras, além de receberem efluentes industriais e
hospitalares (Santana e Barroncas 2007; Pio et al. 2014).
Esses processos têm degradado a qualidade do ecossistema e têm impactado a
estrutura e a dinâmica das comunidades bióticas residentes (Omar et al. 2015), principalmente
a dos peixes (Oliveira et al. 2010), além de provocar mudanças nos ecossistemas como a
qualidade da água, que se tornou cada vez mais poluída (Pinto et al. 2009; Silva et al. 2019e).
Os poluentes liberados em corpos hídricos podem ter efeitos nocivos sobre os organismos que
habitam esses ambientes e aqueles que os consomem (Omar et al. 2015; Kroon et al. 2017),
além de favorecerem a bioacumulação de contaminantes, e provocando alterações do tecido e
material genético de células somáticas e germinativas (Almeida-Val e Farias 1996; Freitas e
Siqueira-Souza 2009).
O mapeamento de sequências repetitivas de DNA nos cromossomos de várias espécies
de peixes que habitam ambientes impactados também revelou a importância biológica desses
elementos no genoma (Symonová e Howell 2018), onde essas sequências parecem
desempenhar um papel fundamental em um gama de processos adaptativos (Ziemniczak et al.
2014; Terencio et al. 2015). Neste contexto, cinco espécies de peixes foram identificadas, por
meio da análise das sequências de COI e comparadas entre ambientes aquáticos poluídos e
não poluídos na região de Manaus, para verificar a possível existência de diferenças na
organização genômica.
4.1.3 Material e Métodos

49
Áreas de estudo e amostragem
Para uma análise comparativa, indivíduos de cinco espécies de peixes (Tabela 1)
foram coletados em três locais poluídos na bacia amazônica, nos meses de novembro de 2015
e maio de 2016: 1- Igarapé do Mindu [3 05 22.0 S, 60 0 5.31 W]; 2- Igarapé do Quarenta [3
08 19.7 S, 60 0 56.7 W]; 3- Igarapé do Tarumã-açu [3 00 51.1 S, 60 3 33.2 W]. Estes corpos
d’água drenam áreas densamente habitadas na cidade de Manaus e estão sujeitos à intensa
poluição ambiental causada por efluentes domésticos, industriais e armazenamento
inapropriado de lixo proveniente do Polo Industrial, lixo urbano e hospitalar (Pinto et al.
2009).
Como controle negativo foram utilizadas amostras das mesmas espécies, coletadas no
lago Catalão [3 13 22.3 S, 59 55 14.5 W], localizado na confluência dos rios Negro e
Solimões. Este lago possui água misturada dos dois rios, embora suas características físico-
químicas sejam típicas da água branca do Solimões e são consideradas não poluídas (Pinto et
al. 2009). Após a realização dos procedimentos, os exemplares testemunho foram depositados
na Coleção de Peixes do Instituto Nacional de Pesquisas da Amazônia (INPA).

50
Tabela 1 – Espécies analisadas e seus respectivos locais de coleta na cidade de Manaus-AM.
Ordem Família Espécie Locais de amostragem Número de exemplares Voucher
Siluriformes Loricariidae Pterygoplichthys pardalis
(Castelnau, 1855)
Igarapé do Mindu 1 macho e 1fêmea INPA-ICT 023942
Igarapé do Tarumã-açu 2 machos e 2 fêmeas INPA-ICT 023941
Lago Catalão 1 macho, 1 fêmea e 2 sem
identificação sexual INPA-ICT 57621
Characiformes
Erythrinidae Hoplias malabaricus
(Block, 1794)
Igarapé do Tarumã-açu 2 machos e 1 fêmea INPA-ICT 023929
Lago Catalão 3 machos e 3 fêmeas INPA-ICT 57622
Prochilodontidae Semaprochilodus insignis
(Jardine, 1841)
Igarapé do Tarumã-açu 2 fêmeas e 3 sem identificação
sexual INPA-ICT 023933
Lago Catalão 5 machos, 1 fêmea e 1 sem
identificação sexual INPA-ICT 57619
Serrasalmidae Pygocentrus nattereri
Kner, 1858
Igarapé do Tarumã-açu 1 fêmea INPA-ICT 023940
Lago Catalão 1 macho e 6 fêmeas INPA-ICT 57620
Perciformes Cichlidae Cichlasoma amazonarum
Kullander, 1983
Igarapé do Quarenta 2 fêmeas INPA-ICT 023931
Igarapé do Tarumã-açu 4 machos, 3 fêmeas e 1 sem
identificação sexual INPA-ICT 023930
Lago Catalão 6 juvenis INPA-ICT 57623

51
Análises citogenéticas e moleculares
Técnicas citogenéticas clássicas
Cromossomos mitóticos foram obtidos a partir de células renais, utilizando o protocolo
descrito por Bertollo et al. (1978). Para a detecção da heterocromatina constitutiva (Banda C)
foi utilizada a técnica descrita por Sumner (1972) e das regiões organizadoras de nucléolo
(RONs) aplicou-se a técnica descrita por Howell e Black (1980).
Técnicas moleculares
O DNA genômico de todos os exemplares foi extraído segundo o protocolo Fenol-
Clorofórmio de Sambrook e Russel (2001). O coquetel de primers descrito por Ivanova et al.
(2007) foi usado para amplificar o gene mitocondrial citocromo c oxidase subunidade I (COI)
pela Reação em Cadeia da Polimerase (PCR - Saiki et al. 1988): FishF2 5’-
GTAAAACGACGGCCAGTCGACTAATCATAAAGATATCGGCAC-3’, FishR2 5’-
CAGGAAACAGCTATGACACTTCAGGGTGACCGAAGAATCAGAA -3’, VF2 5’-
GTAAAACGACGGCCAGTCAACCAACCACAAAGACATTGGCAC-3’, e VR1d 5’-
CAGGAAACAGCTATGACTAGACTTCTGGGTGGCCRAARAAYCA-3’.
Os produtos de PCR foram sequenciados usando o kit Big Dye (Applied Biosystems)
no sequenciador automático ABI 3500. O alinhamento das sequências geradas foi realizado
usando-se a ferramenta MAFFT, implementada no software Geneious 11.0.3 (Kearse et al.
2012) e editadas manualmente. As sequências nucleotídicas foram comparadas com
sequências depositadas na base de dados do NCBI (https://www.ncbi.nlm.nih.gov) usando a
ferramenta BLASTn. As relações entre as sequências de COI foram baseadas inicialmente no
método de agrupamento vizinho, usando o modelo de substituição de 2 parâmetros Kimura,
executado em MEGA 6. Todas as sequências de COI foram depositadas no GenBank sob os
seguintes números de acesso: MH973416–24 (Hoplias malabaricus), MH973425-40
(Cichlasoma amazonarum), MH973441-52 (Pterygoplichthys pardalis), MH973453-60
(Pygocentrus nattereri) e MH991706-17 (Semaprochilodus insignis).
Os modelos evolutivos para as abordagens Inferência Bayesiana (BI) e Máxima
Verossimilhança (ML) foram gerados por JModeltest (Posada 2008). As análises de BI foram
executadas no MrBayes 3.1.2 (Ronquist e Huelsenbeck 2003), com quatro cadeias de 10
milhões de gerações, amostradas a cada 100 gerações, com um burnin inicial de 25%. As
análises de ML foram realizadas em Garli 2.0 (Zwickl 2006), com um bootstrap de 1.000

52
réplicas e as árvores resultantes sendo resumidas pelo consenso majoritário em SumTrees. As
árvores ML e BI foram visualizadas e editadas no Figtree 1.4.3 (Rambaut 2016).
A técnica de Hibridização in situ Fluorescente (FISH) foi utilizada para mapear as
sequências repetitivas de DNAr 18S e 5S nos cromossomos, com sondas homólogas obtidas
por amplificação por PCR, utilizando os primers específicos para cada sequência. Para o 18S
rDNA os primers foram 18Sf (5'-CCG CTT TGG TGA CTC TTG AT-3 ') e 18Sr (5'-CCG
AGGACC TCA CTAACAC-3') (Gross et al. 2010), enquanto para o rDNA 5S os primers
foram 5Sf (5'- CGA GGTCGGGCCTGGTTAGTA-3') e 5Sr (5'-
CTTCYGAGATCAGACGAGATC-3' (Carvalho et al. 2016). Os produtos de PCR dos
marcadores citogenéticos foram marcados por nick translation com biotina-14-dATP (Biotin
Nick Translation mix; Invitrogen) e digoxigenina11-dUTP (Dig-Nick Translation mix;
Roche), seguindo as instruções do fabricante. Os anticorpos estreptavidina-Alexa Fluor
488/NFDM e antidigoxigenina rodamina (Roche) foram usados para detecção e amplificação
do sinal. A FISH utilizando sondas homólogas, foram realizadas seguindo Pinkel et al.
(1986), com 77% de estringência (2,5 ng/µL de sonda marcada, 50% formamida, 10% sulfato
dextrano, 2XSSC a 37 °C por 18 h). Os cromossomos foram contracorados com DAPI (4’, 6-
diamidino-2-fenilindol) (2 µg/mL), em meio de montagem Vectashield (Vector).
4.1.4 Resultados
DNA mitocondrial
Um único haplótipo COI foi recuperado em P. pardalis (522 pb) e P. nattereri (511
pb). Em S. insignis (540 pb) e C. amazonarum (522 pb), três haplótipos com dois sítios
polimórficos foram observados em cada uma, mas nenhuma estruturação foi encontrada nas
árvores filogenéticas obtidas para as duas espécies. Em H. malabaricus (522 pb), no entanto,
variabilidade genética foi encontrada entre os indivíduos analisados, resultando em uma
matriz com 21 sítios polimórficos e nove haplótipos. Apesar dessa variabilidade, as análises
filogenéticas não separaram os indivíduos por ambiente (igarapé do Tarumã-açu e lago do
Catalão), visto que um dos clados recuperados nesta análise incluiu os indivíduos do Catalão,
juntamente com um indivíduo do Tarumã-açu (1045), enquanto o outro clado incluiu os
outros dois indivíduos (1043 e 1044) do Tarumã-açu (Figura 1).
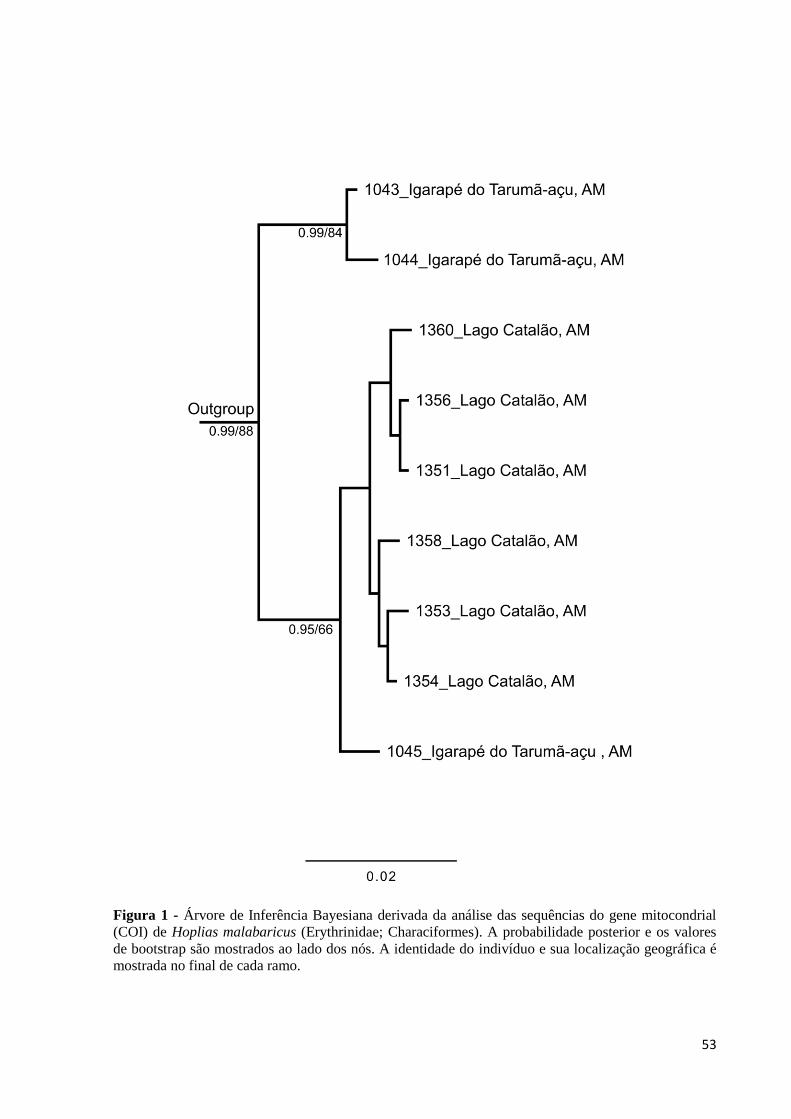
53
Figura 1 - Árvore de Inferência Bayesiana derivada da análise das sequências do gene mitocondrial
(COI) de Hoplias malabaricus (Erythrinidae; Characiformes). A probabilidade posterior e os valores
de bootstrap são mostrados ao lado dos nós. A identidade do indivíduo e sua localização geográfica é
mostrada no final de cada ramo.

54
Caracterização cariotípica
Nenhum cromossomo sexual diferenciado foi observado em nenhuma das cinco
espécies estudadas. A variação inter e intraespecífica nos cariótipos definidos pelas técnicas
citogenéticas clássicas foi consistente com o padrão esperado para as três ordens
representadas, ou seja, os Siluriformes, Characiformes e Perciformes (Tabela 2; Figura 2). A
distribuição da heterocromatina constitutiva (Banda C) e das RONs nos cariótipos das cinco
espécies, incluindo indivíduos de ambientes poluídos e não poluídos, é mostrada na Figura 2.
O mapeamento cromossômico das sequências repetitivas de DNAr 18S e 5S é mostrado na
Tabela 3. Em P. pardalis, o sítio do DNAr 18S está localizado no par 11 do cromossomo,
coincidente com a RON (Figura 3). Em todas as outras espécies, a FISH com sonda DNAr
18S revelou um grande número de sítios ribossomais, distribuídos ao longo do genoma, com
diferentes padrões sendo encontrados nos dois ambientes analisados (poluído e não poluído).
Múltiplos sítios de DNAr 18S foram observados em H. malabaricus com variação
sistemática entre ambiente poluído e não poluído. Nos indivíduos do ambiente poluído
(igarapé do Tarumã-açu) foram observados sítios DNAr 18S na região centromérica dos pares
cromossômicos 1, 12 e 17, e em um dos homólogos do par 3. Esses sinais coincidiram com as
RONs nos pares 1, 12 e 17. Nos indivíduos do lago Catalão, em contrapartida, os sítios de
DNAr 18S foram observados na região terminal dos pares cromossômicos 2, 4, 5 e 17, e na
região intersticial do par 16, com as RONs sendo encontradas nos pares 4, 5 e 17 (Figura 3).
Vale destacar que a impregnação pelo nitrato de Prata (AgNO3) impregnou regiões
heterocromáticas ricas em resíduos ácidos nos pares cromossômicos 4 e 5 em Hoplias
malabaricus de ambientes não poluídos.
Nos indivíduos de S. insignis provenientes do igarapé do Tarumã-açu, múltiplos sítios
de DNAr 18S foram observados na maioria dos cromossomos, com blocos
compartimentalizados na região centromérica, enquanto que nos indivíduos do lago Catalão
(ambiente natural) os sinais de DNAr 18S foram observados apenas no par 2 do cromossomo
(Figura 3). Nos indivíduos de P. nattereri oriundos do igarapé do Tarumã-açu, marcações
conspícuas foram detectadas em diferentes regiões de alguns pares cromossômicos, enquanto
que nos indivíduos do lago Catalão, as marcações foram localizadas nas regiões terminais dos
pares 26, 27, 28, 29, e 30 (Figura 3). Em C. amazonarum foram detectados sítios de DNAr
18S na maioria dos cromossomos dos indivíduos dos igarapés do Quarenta e Tarumã-açu,

55
além do lago Catalão, com um bloco evidente no par 2 coincidente com a RON para os
indivíduos provenientes do ambiente poluído (Figura 3).
Diferenças na distribuição e localização dos sítios de DNAr 5S foram observadas entre
os ambientes poluído e não poluído para H. malabaricus e S. insignis (Figura 3). As demais
espécies apresentaram marcação simples e localizada nos mesmos pares cromossômicos entre
os ambientes analisados. Em P. nattereri, o DNAr 5S foi localizado no par cromossômico 2,
enquanto em P. pardalis, no par 11. Os sítios em C. amazonarum no par 10 mostraram-se
presentes em ambos os ambientes (poluído e não poluído), porém a localização deste gene em
um dos homólogos do par cromossômico 19 esteve presente somente nos indivíduos do
ambiente poluído (Figura 3).
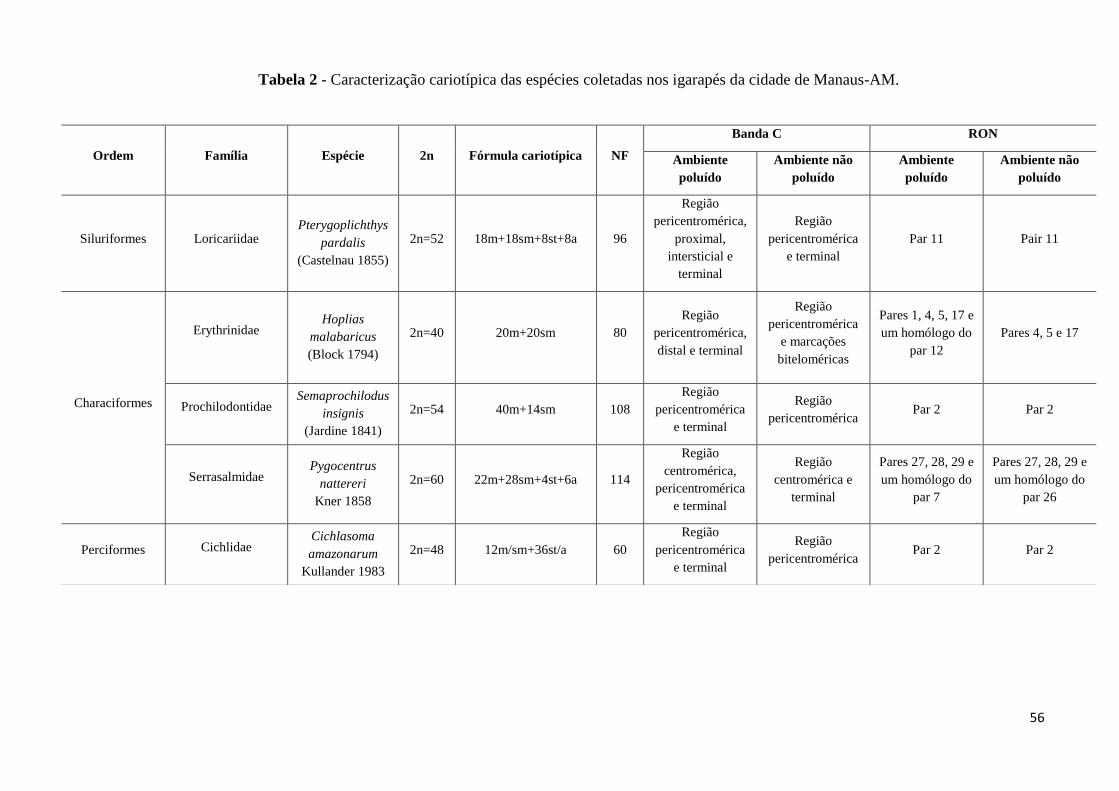
56
Tabela 2 - Caracterização cariotípica das espécies coletadas nos igarapés da cidade de Manaus-AM.
Ordem Família Espécie 2n Fórmula cariotípica NF
Banda C RON
Ambiente
poluído
Ambiente não
poluído
Ambiente
poluído
Ambiente não
poluído
Siluriformes Loricariidae Pterygoplichthys
pardalis
(Castelnau 1855)
2n=52 18m+18sm+8st+8a 96
Região
pericentromérica,
proximal,
intersticial e
terminal
Região
pericentromérica
e terminal
Par 11 Pair 11
Characiformes
Erythrinidae Hoplias
malabaricus
(Block 1794)
2n=40 20m+20sm 80
Região
pericentromérica,
distal e terminal
Região
pericentromérica
e marcações
biteloméricas
Pares 1, 4, 5, 17 e
um homólogo do
par 12
Pares 4, 5 e 17
Prochilodontidae Semaprochilodus
insignis
(Jardine 1841)
2n=54 40m+14sm 108
Região
pericentromérica
e terminal
Região
pericentromérica Par 2 Par 2
Serrasalmidae Pygocentrus
nattereri
Kner 1858
2n=60 22m+28sm+4st+6a 114
Região
centromérica,
pericentromérica
e terminal
Região
centromérica e
terminal
Pares 27, 28, 29 e
um homólogo do
par 7
Pares 27, 28, 29 e
um homólogo do
par 26
Perciformes Cichlidae Cichlasoma
amazonarum
Kullander 1983
2n=48 12m/sm+36st/a 60
Região
pericentromérica
e terminal
Região
pericentromérica Par 2 Par 2

57
Figura 2 – Distribuição da heterocromatina nos cariótipos das espécies analisadas de ambiente poluído e
não poluído, em destaque os cromossomos portadores da RON (caixas), sendo m = metacêntrico; sm =
submetacêntrico; st = subtelocêntrico; a = acrocêntrico. Barra de escala igual a 10 μm.

58
Tabela 3 – Panorama comparativo das sequências repetitivas de DNAr 18S e 5S observadas em cromossomos mitóticos de cinco espécies de peixes
amostradas em ambiente poluído e não poluído na região de Manaus-AM.
Ordem Família Espécie 2n DNAr 18S DNAr 5S
Ambiente poluído Ambiente não poluído Ambiente poluído Ambiente não poluído
Siluriformes Loricariidae Pterygoplichthys
pardalis
(Castelnau, 1855)
2n=52 Par 11 Par 11 Par 11 Par 11
Characiformes
Erythrinidae Hoplias
malabaricus
(Block, 1794)
2n=40 Pares 1, 12,17 e um
homólogo do par 3 Pares 2, 4, 5, 16 e 17 Pares 14 e 18 Pares 13 e 17
Prochilodontidae Semaprochilodus
insignis
(Jardine, 1841)
2n=54
Pares 2, 4, 6, 8, 13, 14, 15,
16, 18, 21, 23, 25 e um
homólogo dos pares 1 e 5
Par 2
Pares 2, 5, 21, 22, 23,
26 e um homólogo
do par 9 e do par 27
Pares 2, 3, 11, 15, 17, 23
e 26
Serrasalmidae Pygocentrus
nattereri
Kner, 1858
2n=60
Pares 3, 12, 20, 22, 23, 27 e
um homólogo dos pares 7,
25, 29 e 30
Pares 26, 27, 28, 29 e 30 Par 2 Par 2
Perciformes Cichlidae Cichlasoma
amazonarum
Kullander, 1983
2n =48 Não apresentou marcação
nos pares 18, 19, 20, 21 e 22
Não apresentou
marcação no par 23
Par 10 e um
homólogo do par 19 Par 10

59
Figura 3 – Cariótipos das espécies de ambiente poluído e não poluído na região de Manaus,
evidenciando sítios de DNAr 18S (sinal vermelho) e de DNAr 5S (sinal verde). Os cromossomos
foram contracorados com DAPI. Barra de escala igual a 20 μm.

60
4.1.5 Discussão
As populações de peixes estão sendo profundamente impactadas pela poluição
ambiental em toda a região Neotropical (Barroca 2012) e a capacidade das diferentes espécies
de persistirem em ecossistemas contaminados depende fundamentalmente de sua capacidade
adaptativa, que está intimamente ligada à sua variabilidade genética (Gross et al. 2010; Silva
et al. 2016; Heinrichs-Caldas et al. 2019).
Diferentes populações de espécies de peixes que habitam ambientes poluídos e não
poluídos podem apresentar níveis relativamente altos de variação em seu DNA mitocondrial
(Schizas et al. 2001). No presente estudo, no entanto, nenhuma variação sistemática foi
encontrada entre os locais, com as sequências de COI, e nem haplótipos associados a um
ambiente específico, o que mostra que a variação nos sítios ribossômicos não está relacionada
com a presença de diferentes unidades evolutivas de cada espécie, específicas para cada
ambiente.
Assim, a variação encontrada no mapeamento cromossômico físico das espécies
estudadas (Figura 3) não foi relacionada sistematicamente a qualquer variação nas sequências
de COI, mas sim a fatores ambientais, dado que diferenças evidentes na distribuição dos
marcadores citogenéticos (sequências repetitivas) foram encontradas entre ambiente poluído e
não poluído (Figuras 2 e 3). As sequências repetitivas parecem evoluir de forma relativamente
rápida e o mapeamento cromossômico desses elementos permite a definição dos diferentes
estágios de diversificação do genoma, mesmo aqueles que são indistinguíveis
morfologicamente (Mehner et al. 2010; Symonová et al. 2013).
Em peixes, a localização dos genes ribossomais em cromossomos de indivíduos
provenientes de diferentes condições ambientais tem destacado sua importância biológica no
genoma, o que pode resultar na formação de um “rDNAome” (Symonová e Howell 2018), o
que tem provado ser uma ferramenta extremamente útil para a compreensão da diversificação
das populações, da especiação e do estabelecimento da diversidade (Costa et al. 2016). No
presente estudo foram encontradas diferenças no número de cópias do DNAr 18S entre
indivíduos de ambiente poluído e não poluído (Figura 3). Além disso, os sítios apresentaram-
se colocalizados com heterocromatina constitutiva, indicando que as mesmas pressões
evolutivas podem estar contribuindo para a propagação desses dois componentes (Cabral-de-
Mello et al. 2011; Lima-Filho et al. 2014a).

61
A heterocromatina constitutiva, que inicialmente foi considerada como lixo genômico,
mostra-se altamente dinâmica, sendo ativa na transcrição e desempenhando um papel
importante na estabilidade do genoma (Postepska-Igielska e Grummt 2014). O aumento da
heterocromatina, distribuída em diferentes regiões do genoma e colocalizada com grandes
blocos de DNAr 18S, como observado no par 2 de C. amazonarum (Figura 3), por exemplo,
pode ser explicada por um processo de heterocromatinização e/ou adição de heterocromatina,
como resultado de eventos epigenéticos, desencadeados pelo contato desses peixes com o
ambiente poluído de Manaus. Em outras palavras, como marca estrutural de sequências
repetitivas, a heterocromatina assegura a integridade genômica por neutralizar o rearranjo
mutagênico, causado pela recombinação homóloga excessiva e/ou eventos de transposição
descontrolados, desencadeados por elementos transponíveis (TEs) (Postepska-Igielska e
Grummt 2014).
O aumento observado no número de cópias do DNAr 18S nos indivíduos de ambiente
poluído pode estar eventualmente relacionado ao sistema de amplificação do gene ribossomal
(como explicado por Kobayashi 2016), que pode ser ajustado para manter ou, quando
necessário, recuperar um número espécie-específico de cópias de DNAr. Isso provavelmente
explicaria a instabilidade do DNAr 18S observada no genoma dos indivíduos de P. nattereri,
S. insignis e H. malabaricus do ambiente poluído, em comparação com aqueles do habitat não
poluído (Figura 3). Os dados obtidos no presente estudo para o marcador DNAr 18S para o
ambiente não poluído foram consistentes com os da literatura (Nakayama et al. 2012;
Terencio et al. 2012; Guimarães et al. 2017), indicando que houve de fato uma divergência no
DNAr nos indivíduos do ambiente estressor, que pode refletir níveis diferenciados de funções
celulares essenciais.
O DNAr e os nucléolos são elementos essenciais que integram sinais ambientais e
intracelulares, além de sensores de estresse celular (Grummt 2013; Tsekrekou et al. 2017).
Em outras palavras, condições ambientais adversas podem favorecer quebras cromossômicas,
principalmente em regiões altamente repetitivas, devido à extrema fragilidade destes locais
(Jacobina et al. 2013). Como esses genes funcionam como sensores primários dos danos
causados por fatores ambientais (Kobayashi 2008; Gross et al. 2010b), número e posição do
DNAr 18S variaram nos indivíduos do ambiente poluído, além de apresentarem um
heteromorfismo de tamanho nos sítios transcricionalmente ativos (Figuras 2 e 3). Isso indica
uma duplicação ou dispersão dessas sequências nos indivíduos do ambiente poluído, mediada
pela mobilização de elementos transponíveis (TEs), o que pode causar a reestruturação das

62
modificações genéticas e a manutenção da integridade do genoma, necessária para uma rápida
diversificação (Zeh et al. 2009; Coan e Martins 2018).
Neste contexto, o sistema especializado para a amplificação do gene do DNAr evoluiu
para manter um número ideal de repetições dos genes ribossomais no genoma dos peixes, em
resposta à mudança do ambiente. No entanto, estudos anteriores indicam a ocorrência de
eventos independentes de dispersão dos genes 18S e 5S no genoma de diferentes grupos de
peixes (Britton-Davidian et al. 2012; Sochorová et al. 2018). No presente estudo, em geral, o
número de sítios de DNAr 5S foi relativamente estável entre as diferentes espécies, com a
presença de apenas um ou dois sítios na maioria dos casos, exceto em S. insignis. Essa
organização pode representar uma estratégia de proteção contra eventos de transposição e
permutação (Galetti e Martins 2004; Martins et al. 2004; Silva et al. 2019a).
Apesar desse arranjo conservador, eventos estocásticos podem resultar na ampla
dispersão das sequências de DNAr 5S no genoma (Affonso e Galetti 2005; Lima-Filho et al.
2014b) e essa variabilidade pode estar relacionada à presença de espaçadores não transcritos
(NTS), que normalmente estão associados a outros tipos de DNA repetitivo (Rebordinos et al.
2013; Da Silva et al. 2016). Isso explicaria a quinta marcação de 5S observada em C.
amazonarum e os múltiplos sítios encontrados em S. insignis (Figura 3). Também foi
observada sintenia entre os genes ribossomais em P. pardalis (Figura 3) e S. insignis, que
podem refletir uma provável condição adaptativa para a organização dessas famílias
multigênicas no genoma dessas espécies (Gornung 2013; Amorin et al. 2016).
Em geral, as espécies aqui estudadas se apresentaram com pouca variação no gene
COI, exceto em H. malabaricus. A extensão e a complexidade da organização das sequências
ribossomais no genoma de peixes expostos ao ambiente poluído também indicam que essas
sequências estão envolvidas principalmente na manutenção da integridade do genoma, que é
mediada pelo aumento do número de cópias do DNAr em resposta a mudanças ambientais.
Essa variação no número de cópias pode ter consequências evolutivas, como a formação de
barreiras reprodutivas e antecedentes cromossômicos, o que levaria à diferenciação do
genoma e, em última instância, à especiação. Tal possibilidade expõe a necessidade de
explorar recursos disponíveis em abordagens citogenômicas integrativas com intuito de
elucidar os fatores que podem estar envolvidos na evolução e organização do genoma
funcional dos peixes.

63
4.2 Artigo 2
Ensaio HpaII/MspI-PCR para medir a metilação do DNA em Hoplosternum
littorale (Callichthyidae, Siluriformes) de ambiente poluído na bacia central
da Amazônia

64
4.2.1 Resumo
A HpaII/MspI-PCR é um método baseado na sensibilidade de enzimas de restrição à
metilação em seus locais de clivagem, que permite a comparação do status de metilação do
DNA de diferentes organismos, com base nos padrões diferenciais de digestão. Antes do
presente estudo, nada era conhecido sobre o perfil de metilação de sequências específicas de
DNA em peixes amazônicos, com base em enzimas sensíveis à metilação. Diante disso, o
presente estudo investigou o perfil de metilação do gene DNAr 18S de um peixe conhecido
como tamoatá (Hoplosternum littorale, Siluriformes, Callichthyidae) que habita ambientes
relativamente inóspitos na bacia amazônica. Nossos achados, revelaram um perfil de
metilação diferencial nos sítios de DNAr 18S, indicando que o ambiente poluído pode
influenciar na metilação do DNA e que, esta característica teria evoluído, inicialmente, como
um mecanismo de defesa dos organismos contra a expressão de elementos transponíveis.
Palavras-chave: DNA repetitivo; epigenética; enzimas de restrição; poluição; tamoatá.

65
4.2.2 Introdução
Peixes da região amazônica adquiriram considerável plasticidade genotípica e
fenotípica durante sua história evolutiva, o que sustenta sua rápida adaptação às diversas
condições encontradas nesse ambiente complexo (Braz-Mota et al. 2018; Heinrichs-Caldas et
al. 2019). Essa capacidade é especialmente importante em ambientes urbanos impactados pela
descarga de efluentes domésticos e industriais (Santana e Barroncas, 2007; Silva et al. 2016;
Silva et al. 2019a).
Uma das espécies que consegue sobreviver em ambientes poluídos na cidade de
Manaus (capital do estado brasileiro Amazonas) é Hoplosternum littorale (Siluriformes,
Callichthyidae), conhecido localmente como tamoatá. Nesta espécie foram encontrados
polimorfismos de heterocromatina associados aos genes ribossomais 18S, sendo a
heterocromatinização/adição de heterocromatina possíveis eventos epigenéticos como uma
resposta adaptativa a estressores de ambientes (Silva et al. 2016; Silva et al. 2019a;c). Além
da heterocromatinização e adição de heterocromatina, que envolve a modificação das histonas
e o remodelamento da cromatina, outros mecanismos com padrão de herança epigenética
podem ocorrer, tais como RNAs não-codificantes e metilação do DNA, que tem atraído
atenção considerável nos últimos anos, na tentativa de entender a relação entre o ambiente e o
genoma em vários táxons (Head 2014; Ruiz-Hernandez et al. 2015; Brander et al. 2017).
Evidências em uma variedade de organismos eucarióticos indicam que a metilação do
DNA, que ocorre através de uma ligação covalente de um grupo metil (-CH3) na posição 5 do
anel pirimídico de uma citosina, tem efeitos epigenéticos e mutagênicos, além de papel
fundamental na expressão gênica (Ruiz-Hernandez et al. 2015; Muthusamy et al. 2016). Em
geral, os peixes tendem a ter uma porcentagem maior de metilação global do DNA em
comparação com outros vertebrados, como aves e mamíferos (Head et al. 2014). Porém, a
avaliação da metilação de sítios específicos também pode ser usada para compreender o papel
de genes específicos após a exposição a certas condições ambientais (Martin e Fry 2018).
Um dos métodos clássicos, comumente usado para a detecção de 5mC em genes
específicos, é o ensaio HpaII / MspI-PCR sensível à metilação, que é baseado na sensibilidade
de enzimas de restrição à metilação em seus locais de clivagem, seguido pela amplificação de
fragmentos de restrição (Reyna-Lopez et al.1997; Fraga e Esteller 2002). Estes
isosquizômeros geralmente são utilizados aos pares, tais como as enzimas MspI / HpaII, que
reconhecem ambos o mesmo local 5'-CCGG. No entanto, a enzima HpaII é sensível se uma

66
ou ambas as citosinas presentes forem metiladas, embora possa clivar sequências
hemimetiladas, ao contrário da enzima MspI, que cliva o sítio C5m
CGG, hemimetilado ou com
ambas as cadeias metiladas (Busslinger et al. 1983 McClelland et al. 1994), mas não cliva os
sítios 5m
C5m
CGG ou 5m
CCGG (Fulneček e Kovařík 2014).
Os genes ribossomais são regiões gênicas repetitivas, arranjadas in tandem em série de
unidades transcricionais. Especificamente, o gene 18S é uma sequência moderadamente
repetitiva, responsável pela formação do RNAr 18S, o qual faz parte da subunidade menor do
ribossomo das células eucarióticas. Esse gene contém múltiplos sítios CCGG palindrômicos,
que estão sujeitos à metilação pelo DNA metil transferases ou DNMTs (Kucharski et al.
2008). Como o Hoplosternum littorale é encontrado em diversos ambientes amazônicos e os
mecanismos epigenéticos parecem estar associados ao gene ribossômico 18S, o presente
estudo investigou e comparou o perfil de metilação desse gene em indivíduos de H. littorale
de ambientes poluídos e não poluídos na bacia amazônica.
4.2.3 Material e Métodos
Trinta indivíduos de Hoplosternum littorale foram coletados com autorização do
Instituto Chico Mendes de Conservação da Biodiversidade (ICMBIO - Licença Permanente nº
7634-1/2015). Os indivíduos foram capturados com o uso de tarrafas, redes e rapiches,
armazenados em recipientes contendo água do ambiente onde foram capturados e enviados ao
Laboratório de Genética Animal do Instituto Nacional de Pesquisas da Amazônia (INPA) para
realização dos procedimentos. Os peixes foram eutanasiados seguindo as diretrizes para
Prática da Eutanásia do Conselho Nacional de Controle de Experimentação Animal - Concea.
Amostras de tecido muscular foram coletadas para a extração do DNA. Os procedimentos
foram previamente aprovados pelo Comitê de Ética em Uso Animal (CEUA) do Instituto
Nacional de Pesquisas da Amazônia (nº 0111/2017).
O DNA genômico foi extraído do tecido muscular de H. littorale, utilizando o
protocolo descrito por Sambrook e Russell (2001). Dez indivíduos foram coletados em um
ambiente não poluído (lago Catalão [3 13 22.3 S, 59 55 14.5 W]), localizado na confluência
dos rios Negro e Solimões e vinte indivíduos de dois igarapés urbanos da cidade de Manaus,
estado do Amazonas. Os dois igarapés, Mindu [3 05 22.0 S, 60 0 5.31 W] e o Quarenta [3 08
19.7 S, 60 0 56.7 W] são considerados poluídos por efluentes industriais e por resíduos
orgânicos em suas águas (Pinto et al. 2009; Piedade et al. 2014). Os exemplares testemunho

67
foram depositados na coleção de peixes do INPA sob números: INPA-ICT 023935; INPA-
ICT 023936; INPA-ICT 023937.
O DNA (5 ng/μL) foi digerido com as enzimas MspI e HpaII, seguindo as instruções do
fabricante e utilizando o DNA genômico como controle da digestão enzimática. As amostras
foram incubadas a 37 ºC por 1 h, seguidas de 15 min a 65 ºC, para interromper a ação das
endonucleases. Os produtos da digestão foram comparados por eletroforese padrão (com
tampão Tris-Borato-EDTA 0,5X e corrida a 70 V por 40 minutos) em gel de agarose 1,7% e
corados com GelRed (Biotium). Um mapa de restrição virtual foi preparado pela primeira vez
na ferramenta Bioedit para a detecção dos sítios CCGG na sequência do DNAr 18S disponível
no banco de dados do NCBI (números de acesso: AF021880.1) (Figura 1).
Na sequência, na Figura 2, é possível observar o esquema de interpretação com as
enzimas HpaII/MspI-PCR. Os fragmentos resultantes da digestão pelas enzimas MspI e HpaII
foram então amplificados por Reação em Cadeia da Polimerase (PCR - Saiki et al. 1988),
utilizando os seguintes primers: 18S - 18Sf (5'-CCG CTT TGG TGA CTC TTG AT-3′) e
18Sr (5′-CCG AGGACC CTA CTA AAC CA-3) (Gross et al. 2010). As reações de PCR
foram feitas em um termociclador Eppendorf-Mastercycler Gradient, para um volume final de
15 μL (1 μL de DNA genômico (100 ng), 1,5 μL de Tampão 10X com cloreto de magnésio
(1,5 mM), 0,15 μL de Taq DNA polimerase (5U / μL), 3,0 μL de dNTP (1 mM), 0,6 μL de
cada primer (5 mM), água milli-Q para completar o volume). Os produtos de PCR foram
visualizados em gel de agarose 1,7% corados com GelRed (Biotium) utilizando um
transluminador de luz ultravioleta (260 nM) acoplado a um fotodocumentador Easy Doc 100
(BioAgency).

68
Figura 1 - Mapa de restrição virtual preparado na ferramenta Bioedit para a detecção dos sítios
CCGG na sequência do DNAr 18S disponível na base de dados do NCBI (números de acesso:
AF021880.1).

69
Figura 2 - Perfis esperados (interpretações) do efeito de clivagem por MspI e HpaII. O esquema
mostra exemplos de amostras de DNA com controle negativo (DNA não digerido) e controle
positivo (DNA digerido) para ambas as enzimas.

70
4.2.4 Resultados e Discussão
O mapa de restrição virtual da sequência de pares de bases de 1.872 do gene RNAr 18S
revelou 19 sítios "CCGG" entre as regiões de anelamento dos primers (Figura 1).
Citogeneticamente, os locos do DNAr 18S nos indivíduos de H. littorale do ambiente não
poluído (lago Catalão) e poluído (igarapés do Mindu e Quarenta) da região amazônica foram
restritos à região terminal dos braços curtos de ambos os homólogos do par cromossômico 5,
com apenas um heteromorfismo de tamanho revelado nos indivíduos dos ambientes poluídos
(Silva et al. 2016) (Figura 3).
No presente estudo, com base nos perfis esperados do efeito de clivagem por MspI e
HpaII (Figura 4), analisamos a digestão de todas as amostras de diferentes locais e
observamos que o gene DNAr 18S apresentou sítios metilados, confirmados por digestões
positivas usando ambas as enzimas (Figura 5), indicando que existe alguma estratégia
evolutiva para regulação epigenética nesta espécie. A sequência do 18S foi amplificada em
amostras de controle (não digerido pelas enzimas MspI e HpaII) dos ambientes não poluído e
poluído (Figura 5).
No igarapé do Mindu (ambiente poluído) (Figura 5C) uma banda fraca é visível em
alguns indivíduos, digeridos com HpaII, o que pode representar a hipometilação de alguns
sítios 18S, uma vez que esta enzima não corta palíndromos metilados. A ausência de clivagem
em algumas fitas moldes do gene indica que alguns sítios foram metilados. Em outros
indivíduos, bandas de intensidade semelhante foram observadas no controle e na amostra
digerida pela HpaII, o que pode ser explicado pela não digestão por esta enzima de sítios
CCGG, que seriam metilados na posição C5m
CGG (Reyna-Lopez et al. 1997). O palíndromo
com 5m
C nesta posição interna pode ser digerido pela enzima MspI e, consequentemente, não
teriam amplificação positiva.

71
Figura 3 - Cariótipo de Hoplosternum littorale evidenciando os sítios de DNAr 18S (par 5) (caixas) e
sítios de DNAr 5S (pares 23 e 27). (a) Ambiente não poluído (lago Catalão); (b) ambiente poluído
(igarapés do Mindu e Quarenta). Barra de escala igual a 20 μm. Fonte: Silva et al. 2016.

72
Figura 4 - Digestão de amostras de DNA com enzimas MspI e HpaII utilizando DNA genômico como controle da
digestão enzimática. Onde A: lago Catalão; B: igarapé do Quarenta; C: igarapé do Mindu.
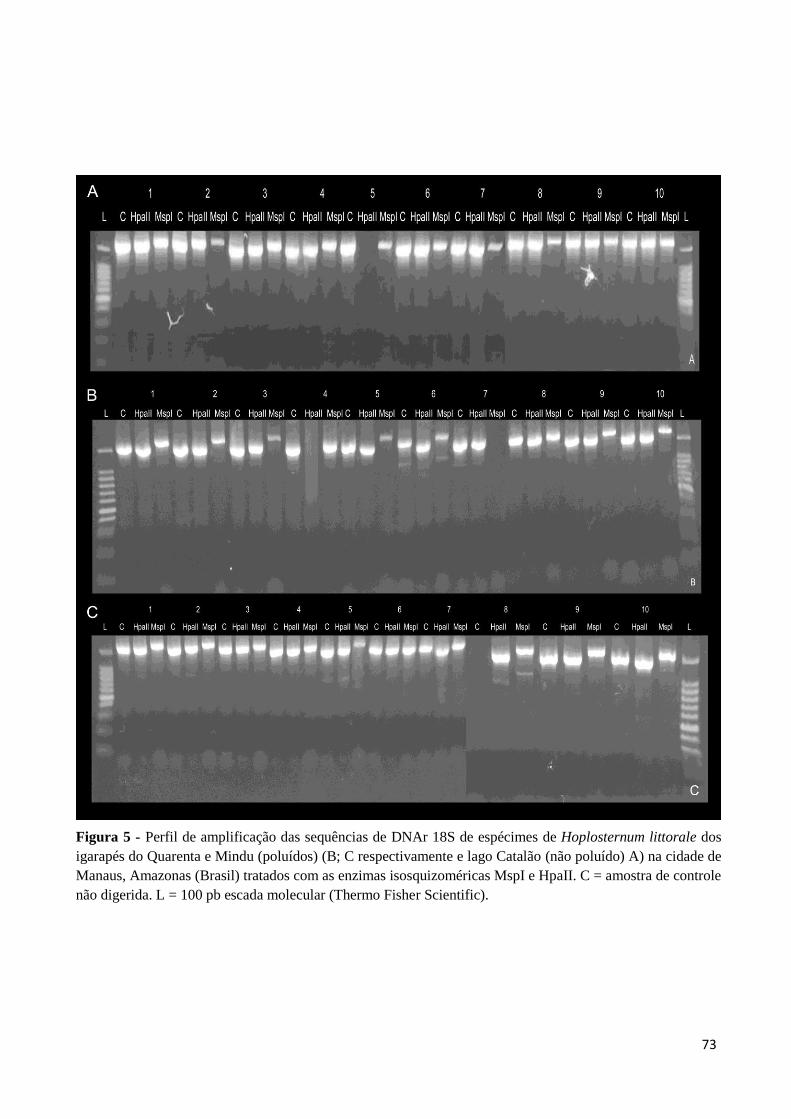
73
Figura 5 - Perfil de amplificação das sequências de DNAr 18S de espécimes de Hoplosternum littorale dos
igarapés do Quarenta e Mindu (poluídos) (B; C respectivamente e lago Catalão (não poluído) A) na cidade de
Manaus, Amazonas (Brasil) tratados com as enzimas isosquizoméricas MspI e HpaII. C = amostra de controle
não digerida. L = 100 pb escada molecular (Thermo Fisher Scientific).

74
Diversos estudos têm evidenciado que os padrões de metilação do DNA podem ser
alterados por fatores dietéticos assim como por condições externas estressantes (Murayama et
al. 2008; Solis et al. 2012). Em mamíferos, por exemplo, as alterações na metilação do DNA
nos sítios de DNAr têm uma grande capacidade de regular o tempo de vida celular (Srivastava
et al. 2016). Em peixes, os processos epigenéticos que estabelecem padrões de metilação do
DNA e que são sensíveis ao estresse ambiental são bem descritos, em particular no zebrafish
(Danio rerio), um organismo modelo para estudos epigenéticos e ecotoxicológicos (Head
2014).
A variação observada no perfil de metilação dos indivíduos do igarapé do Quarenta
(Figura 4B) indica a metilação em sítios CCGG na posição C5m
CGG no gene 18S de alguns
indivíduos, confirmada pela ausência (não amplificada) de uma banda após a digestão pela
enzima HpaII. Outros indivíduos, no entanto, a presença de uma banda fraca no gel da
amostra digerida pela enzima MspI indica que alguns locais podem estar hipermetilados, ou
seja, a citosina interna é metilada (C5m
CGG), e provavelmente a citosina externa também se
encontra metilada (5m
C5m
CGG), já que este fragmento não é passível de digestão pela MspI e
pode resultar na amplificação do 18S (Fulneček e Kovařík 2014).
O perfil de amplificação para os indivíduos do ambiente natural, lago Catalão,
considerado um local isento de poluição (Pinto et al. 2009) indica que o DNAr 18S parece
estar menos metilado, quando comparado com bandas amplificadas de indivíduos de ambiente
poluído. A frequência de aparecimento desses palíndromos duplamente metilados no genoma
de vertebrados não foi por nós elucidada, embora se saiba que no genoma de planta, os
palíndromos duplamente metilados são muito comuns nas regiões de heterocromatina, e a
frequência dos mesmos parece seguir a seguinte ordem: Cm
CGG> m
Cm
CGG> m
CCGG
(Fulneček e Kovařík 2014). A base biológica da variação observada com os resultados
encontrados no presente estudo não é clara, porém, a metilação desses locos pode ser
fortemente influenciada por fatores ambientais e está frequentemente associada à repressão
transcricional (Murayama et al. 2008; Bacalini et al. 2014).
Portanto, para tentar elucidar a natureza da variação observada para apenas dois
indivíduos, repetimos o experimento para uma amostragem maior, e 10 indivíduos foram
respectivamente digeridos por HpaII e MspI. Os resultados obtidos com a digestão indicaram
que os genomas dos indivíduos do Catalão apresentam-se menos metilados, já que os rastros
de digestão são mais visíveis para as amostras desse ambiente. Nos ambientes poluídos
(igarapés do Quarenta e Mindu) foram visualizados poucos rastros de digestão,

75
principalmente para a enzima HpaII o que indica maior nível de metilação global do genoma
(Figura 5).
A partir da análise da amplificação, após a digestão enzimática nos indivíduos do
ambiente não poluído (lago Catalão), observou-se amplificação em todos os indivíduos
digeridos com exceção do indivíduo 5 para HpaII cuja amplificação falhou. Note-se, no
entanto, que a amplificação nos indivíduos digeridos com MspI é diferenciada da
amplificação dos controles e dos indivíduos digeridos com HpaII. A banda de menor
intensidade observada nessas amostras digeridas pela MspI pode ser devido ao menor número
de DNA template disponível para amplificação, uma vez que a enzima MspI digere o DNA de
forma mais eficiente.
Para os indivíduos, 1, 3, 4, 9 e 10 amostras do padrão de amplificação foram
igualmente intensas indicando alto nível de metilação das amostras. Nas amostras
provenientes do igarapé do Quarenta, os resultados foram similares aos encontrados no lago
Catalão, e nas amostras digeridas pela MspI a amplificação foi menos intensa indicando
também um menor número de cópias disponíveis para amplificação. Para o indivíduo número
7 dessa mesma localidade, não houve amplificação na digestão por MspI indicando que a
metilação provavelmente encontra-se na citosina externa do palindrome CCGG. Nas amostras
coletadas no igarapé do Mindu, todas as amostras foram igualmente amplificadas, indicando
um alto grau de metilação (Figura 5).
A partir da análise dos resultados obtidos no presente trabalho podemos concluir que as
amostras coletadas nos três ambientes estudados são em sua maioria altamente metiladas.
Nossos resultados não nos permitem inferir a natureza precisa desse alto grau de metilação,
mas um dos possíveis mecanismos pode ser a ação de agentes poluidores, já que no igarapé do
Quarenta, de acordo com relatos da literatura, apresenta contaminação por resíduos industriais
e no igarapé do Mindu a contaminação se dá por rejeitos domésticos e orgânicos (Pinto et al.
2009). Esse perfil diferenciado de poluição pode estar correlacionado com as diferenças no
perfil de metilação observado para as amostras desses ambientes.
O DNAr 18S é um gene que se apresenta em diversas cópias pelo genoma, no genoma
humano apresenta-se com 300 a 400 cópias e destas aproximadamente a metade delas são
inativadas pela metilação (Bacalini et al. 2014). Portanto, a presença de amostras metiladas no
ambiente natural do lago Catalão pode corresponder às cópias normalmente metiladas pelas
vias epigenéticas convencionais. Por outro lado, a hipermetilação de amostras de ambiente

76
poluído pode ser um indício da ação deletéria da poluição nesses organismos. Esta é uma
hipótese que deverá ser testada em trabalhos futuros.
A regulação epigenética dos sítios de DNA ribossomais desempenha um papel
fundamental na biogênese do ribossomo, e a inativação desses sítios pela metilação pode ter
um efeito indireto sobre o processo de tradução gênica no organismo. Além disso, a metilação
do DNA pode contribuir para a estabilidade do genoma ao reprimir elementos genéticos
prejudiciais, além de ser um dos mecanismos mais comuns envolvidos na repressão dos
retrotransposons (Yoder et al. 1997; Schaeffer e Lyko 2010). Em H. littorale, o DNAr 18S
está colocalizado com o retroelemento Rex 3, evidenciando um acúmulo/aumento deste
retrotransposon nesta espécie (Silva et al. 2016). Neste caso, parece provável que a metilação
do DNA surgiu inicialmente como um mecanismo de defesa usado pelo organismo contra a
expressão de elementos de transposição, e que, a repressão da transcrição pela metilação do
DNA em sítios de 18S seria uma maneira de evitar a ativação dos elementos transponíveis
incorporados nas regiões promotoras desse gene.
Nossos resultados indicam que existem diferenças sutis nos perfis de metilação nos
sítios de DNAr 18S das amostras provenientes do Catalão, Mindu e Quarenta. Se essas
diferenças são realmente produzidas pela ação de agentes poluidores é uma questão ainda em
aberto que deverá ser alvo de futuras abordagens utilizando metodologias mais resolutivas,
como o sequenciamento por bissulfeto. No entanto, entender como de fato a poluição gera
impactos na integridade genética em diferentes espécies de peixes se torna uma importante
ferramenta no entendimento desta dinâmica.

77
4.3 Artigo 3
Elementos transponíveis de DNA em peixes amazônicos: do aumento no
genoma à adaptação genética a ambientes estressantes
Artigo será enviado para publicação no periódico Cytogenetic and Genome Research.

78
4.3.1 Resumo
Os elementos transponíveis têm impulsionado evolução e plasticidade do genoma de
diversas maneiras em uma gama de organismos, e diferentes tipos de estresses bióticos e
abióticos podem estimular a expressão ou a transposição destes elementos móveis. Aqui
analisamos citogeneticamente populações naturais de peixes da mesma espécie vivendo em
diferentes condições ambientais a fim de verificar a influência e a organização de elementos
transponíveis no genoma destas espécies. Foi possível observar um comportamento
diferencial dos marcadores Rex 1, Rex 3 e Rex 6 nos cromossomos de indivíduos de uma
mesma espécie, mas provenientes de diferentes ambientes, poluído e não poluído. É notório o
aumento da quantidade dos elementos transponíveis do tipo Rex nos cromossomos e sua
influência na modelagem do genoma das populações que vivem no ambiente poluído, o qual
deve estar sob constante evolução adaptativa.
Palavras-chave: Amazônia; instabilidade cromossômica; plasticidade genômica; poluição;
retrotransposon.

79
4.3.2 Introdução
Os elementos transponíveis (TEs) têm impulsionado a evolução e plasticidade do
genoma de diferentes organismos, uma vez que promovem mutações de grande magnitude e
influenciam a evolução da regulação transcricional, metilação diferencial do DNA e a
estabilidade cromossômica (Rey et al. 2016; Suh et al. 2017). Evidências sugerem que
inserções individuais de TEs podem ter grandes efeitos fenotípicos, incluindo os deletérios,
como doenças (revisado por Hancks e Kazazian 2012), mas também os benéficos, como a
adaptação de diversos organismos (Casacuberta e Gonzalez 2013; Stapley et al. 2015).
Como os organismos são constantemente desafiados por alterações no ambiente, a
evolução adaptativa desempenha um papel importante na determinação do destino das
espécies, como resposta a condições ambientais variáveis (Hoffmann e Sgro 2011;
Casacuberta e Gonzalez 2013). Desta forma, diferentes tipos de estresses bióticos e abióticos
podem estimular a expressão ou a transposição de TEs em uma ampla gama de organismos
(Silva et al. 2016; Suh et al. 2017). Com isso, os TEs representam entidades genéticas
capazes de se inserir em diferentes porções do genoma e induzir alterações no funcionamento
de genes com os quais se associam (de la Veja et al. 2007; Philippsen 2014), estando os TEs
propensos a desempenhar um papel relevante na adaptação, devido sua capacidade de gerar
mutações, além de responderem e serem susceptíveis a mudanças ambientais (Hua Van 2011;
Casacuberta e Gonzalez 2013; Stapley et al. 2015).
Um estudo realizado com três populações de Hoplias malabaricus, provenientes de
diferentes ambientes aquáticos da bacia amazônica (rios de águas brancas, rios de águas claras
e rios de águas pretas), revelou padrões distintos para o retrotransposon Rex 3, indicando
padrões de acordo com o ambiente e uma possível existência com processos adaptativos
locais da espécie em diferentes condições ambientais (Santos et al. 2016). Para Hoplosternum
littorale, verificou-se com o mapeamento cromossômico do retrotransposon Rex 3, padrões
distintos de distribuição do elemento nos cromossomos de indivíduos de ambiente poluído e
ambiente não poluído (Silva et al. 2016).
Considerando que os peixes teleósteos exibem uma das maiores diversidades de
elementos transponíveis entre os vertebrados (Chalopin et al. 2015; Sotero-Caio et al. 2017) e
que os eventos de transposição são uma das principais causas de instabilidade e de rearranjos
cromossômicos, o objetivo deste estudo foi mapear cromossomicamente sequências
repetitivas de DNA em espécies de peixes provenientes de populações naturais, que vivem

80
tanto em ambiente poluído quanto não poluído, a fim de verificar o comportamento e a
organização de TEs no genoma destas espécies nestes ambientes.
4.3.3 Material e Métodos
Locais de estudo e amostragem
Para uma análise comparativa, três espécies de peixes de diferentes ordens (Tabela 1)
foram coletadas em dois ambientes: a) poluído urbano – locais que drenam áreas densamente
habitadas na cidade de Manaus, capital do Estado do Amazonas-AM e apresentam uma
intensa poluição ambiental, proveniente de atividades domésticas, industriais e do
armazenamento inapropriado de lixo, oriundo do Polo Industrial, lixo urbano e hospitalar; b)
não poluído - confluência dos rios Negro e Solimões (Pinto et al. 2009). Estas mesmas
espécies de peixes, utilizadas no estudo anterior, não apresentam variabilidade no gene COI
em função do ambiente.
Todas as coletas foram realizadas sob a permissão do Instituto Chico Mendes de
Conservação da Biodiversidade (ICMBio/SISBIO número: 7634-1/2015). Os exemplares
coletados foram eutanasiados seguindo as Diretrizes da Prática da Eutanásia do Concea
(Concea, 2013) e com aprovação da Comissão de Ética no Uso de Animais (CEUA) (no.
011/2017) do Instituto Nacional de Pesquisas da Amazônia (INPA). Após a realização dos
procedimentos, os exemplares testemunhos foram depositados na Coleção de Peixes do
Instituto Nacional de Pesquisas da Amazônia - INPA (Tabela 1).
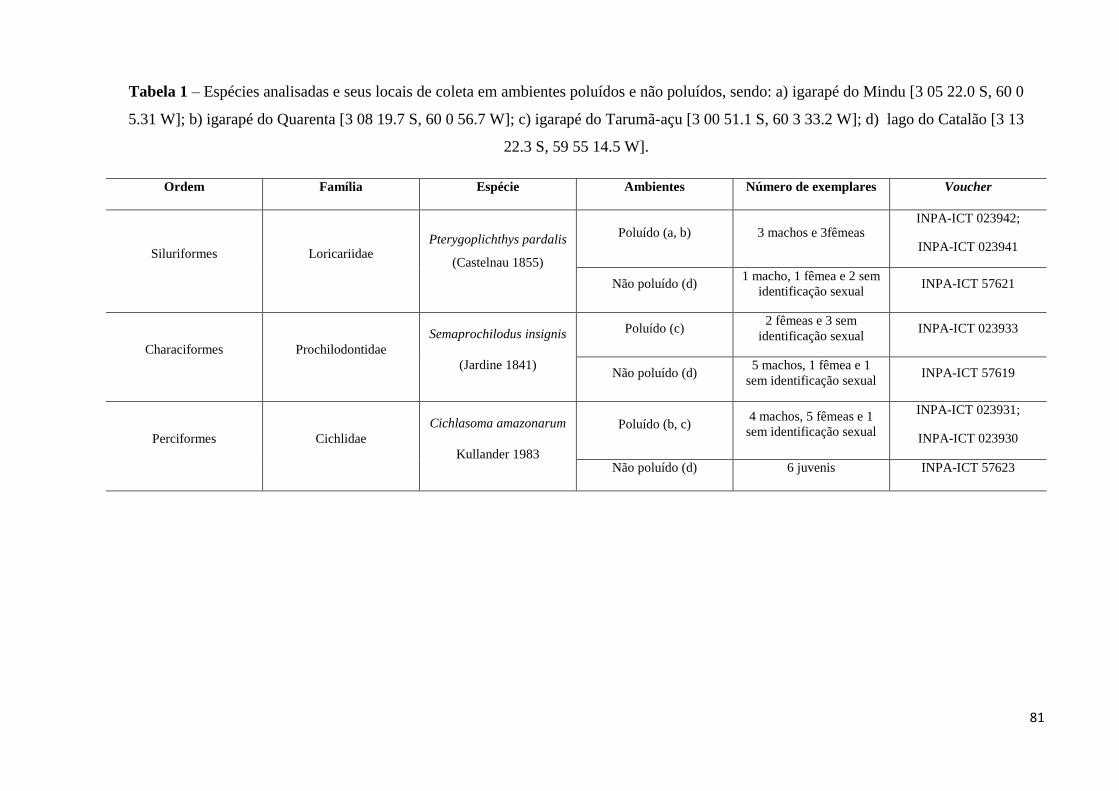
81
Tabela 1 – Espécies analisadas e seus locais de coleta em ambientes poluídos e não poluídos, sendo: a) igarapé do Mindu [3 05 22.0 S, 60 0
5.31 W]; b) igarapé do Quarenta [3 08 19.7 S, 60 0 56.7 W]; c) igarapé do Tarumã-açu [3 00 51.1 S, 60 3 33.2 W]; d) lago do Catalão [3 13
22.3 S, 59 55 14.5 W].
Ordem Família Espécie Ambientes Número de exemplares Voucher
Siluriformes Loricariidae Pterygoplichthys pardalis
(Castelnau 1855)
Poluído (a, b) 3 machos e 3fêmeas INPA-ICT 023942;
INPA-ICT 023941
Não poluído (d) 1 macho, 1 fêmea e 2 sem
identificação sexual INPA-ICT 57621
Characiformes Prochilodontidae
Semaprochilodus insignis
(Jardine 1841)
Poluído (c) 2 fêmeas e 3 sem
identificação sexual INPA-ICT 023933
Não poluído (d) 5 machos, 1 fêmea e 1
sem identificação sexual INPA-ICT 57619
Perciformes Cichlidae
Cichlasoma amazonarum
Kullander 1983
Poluído (b, c) 4 machos, 5 fêmeas e 1
sem identificação sexual
INPA-ICT 023931;
INPA-ICT 023930
Não poluído (d) 6 juvenis INPA-ICT 57623

82
Técnicas moleculares
Cromossomos mitóticos foram obtidos a partir de células renais, utilizando o protocolo
descrito por Bertollo et al. (1978). As sondas de TEs já identificadas e caracterizadas foram
obtidas com a utilização do DNA genômico, extraído do tecido muscular de todos os
exemplares, usando o kit Wizard® Genomic Purification (Promega), de acordo com o
protocolo do fabricante. Para a localização física cromossômica foram mapeados os TEs de
Classe I (retrotransposons: Rex 1, Rex 3 e Rex 6) com sondas homólogas obtidas a partir da
amplificação via Reação em Cadeia da Polimerase (PCR) utilizando-se os seguintes primers:
-Rex 1 (RTX1-F1: 5’-TTC TCC AGT GCC TTC AAC ACC-3’ e RTX1-R3: 5’-TCC CTC
AGC AGA AAG AGT CTG CTC-3’) (Volff et al. 1999; 2000; 2001a);
-Rex 3 (RTX3-F3: 5’-CGG TGA YAA AGG GCA GCC CTG-3’ e RTX3-R3: 5’-TGG CAG
ACN GGG GTG GTG GT-3’) (Volff et al. 1999; 2000; 2001a);
-Rex 6 (Rex 6-Medf1: 5’-TAA AGC ATA CAT GGA GCG CCAC-3’ e Rex 6-Medf2: 5’-
AGG AAC ATG TGT GCA GAA TATG-3’) em combinação com os primers (Rex 6-Medr1:
5’-GGT CCT CTA CCA GAG GCC TGGG-3’ ou Rex 6-Medr2: 5’-GGT GGT TTC TCC
TCC AAG CTCG-3’ (Volff et al. 2001b).
Os produtos obtidos via PCR foram marcados por nick translation com biotina-14-
dATP (Biotin Nick Translation mix; Invitrogen) e digoxigenina11-dUTP (Dig-Nick
Translation mix; Roche), seguindo as instruções do fabricante. Os anticorpos estreptavidina-
Alexa Fluor 488 e antidigoxigenina rodamina (Roche) foram usados para detecção e
amplificação do sinal. Hibridizações in situ fluorescente, utilizando sondas homólogas, foram
realizadas, com 77% de estringência (2,5 ng/µL de sonda marcada, 50% formamida, 10%
sulfato dextrano, 2XSSC a 37 °C por 18 h). Os cromossomos foram contracorados com DAPI
(4’, 6-diamidino-2-fenilindol) (2 µg/mL), em meio de montagem Vectashield (Vector). Os
cromossomos foram analisados em microscópio de epifluorescência Olympus BX51 e as
imagens capturadas com câmera digital (Olympus DP71), usando o software Image-Pro MC
6.3.
4.3.4 Resultados
A localização física cromossômica dos elementos transponíveis Rex 1, Rex 3 e Rex 6
em Pterygoplichthys pardalis (Siluriformes), Semaprochilodus insignis (Characiformes) e
Cichlasoma amazonarum (Perciformes) não revelou diferenças entre os indivíduos, quando

83
coletados em mais de uma localidade nos ambientes poluídos e nem entre machos e fêmeas.
Entretanto, diferenças foram observadas em relação ao tipo de ambiente (ambiente poluído e
ambiente não poluído) em que as espécies foram coletadas (Figura 1, 2 e 3). Foi evidenciado
um aumento de sinais de hibridização de sequências de TEs nos cromossomos de indivíduos
provenientes do ambiente poluído (igarapé do Quarenta, Mindu e Tarumã-açu), quando
comparados com os indivíduos do ambiente natural (lago Catalão). Estes dados comparativos
em relação ao tipo de ambiente (poluído e não poluído), espécies analisadas (P. pardalis, S.
insignis e C. amazonarum), marcadores citogenéticos (Rex 1, Rex 3 e Rex 6), tipo de
marcação (disperso ou compartimentalizado), presença ou não evidente além do aumento
destas marcações podem ser observados na Tabela 2 e Figuras 1, 2 e 3.

84
Figura 1 - Cariótipo de Pterygoplichthys pardalis de indivíduos do ambiente poluído e não poluído da
região de Manaus evidenciando sítios de Rex 1 (sinal verde), Rex 3 (sinal vermelho) e Rex 6 (sinal
verde). Os cromossomos foram contracorados com DAPI. Barra de escala igual a 20 μm.

85
Figura 2 - Cariótipo de Semaprochilodus insignis de indivíduos do ambiente poluído e não poluído
da região de Manaus, evidenciando sítios de Rex 1 (sinal verde), Rex 3 (sinal vermelho) e Rex 6
(sinal verde). Os cromossomos foram contracorados com DAPI. Barra de escala igual a 20 μm.

86
Figura 3 - Cariótipo de Cichlasoma amazonarum de indivíduos do ambiente poluído e não
poluído da região de Manaus, evidenciando sítios de Rex 1 (sinal verde), Rex 3 (sinal vermelho) e
Rex 6 (sinal verde). Os cromossomos foram contracorados com DAPI. Barra de escala igual a 20
μm.

87
Ordem Espécie
Rex 1 Rex 3 Rex 6
Ambiente poluído Ambiente não poluído Ambiente poluído Ambiente não poluído Ambiente poluído Ambiente não poluído
Siluriformes Pterygoplichthys
pardalis
(Castelnau 1855)
Presente e disperso Presente e disperso Presente e disperso Presente e disperso Presente, aumento e
compartimentalizado Não evidente
Characiformes Semaprochilodus
insignis
(Jardine 1841)
Presente, aumento e
compartimentalizado
Presente e
compartimentalizado
Presente e
disperso/compartimen-
talizado
Presente e Disperso Presente, aumento e
compartimentalizado Não evidente
Perciformes Cichlasoma
amazonarum
Kullander 1983
Presente, aumento e
compartimentalizado
Presente e
compartimentalizado
Presente, aumento e
disperso/compartimen-
talizado
Presente e
compartimentalizado
Presente, aumento e
disperso/compartimen-
talizado
Presente e disperso
Tabela 2 – Panorama comparativo da distribuição das sequências de elementos transponíveis Rex 1, Rex 3 e Rex 6 em
cromossomos mitóticos de três espécies de peixes amostradas de ambiente poluído e não poluído na região de Manaus-AM.

88
4.3.5 Discussão
Mudanças ambientais como contaminação, alterações de temperatura, hipóxia e outras
alterações que representam uma condição de estresse ambiental podem estimular a transcrição
de retrotransposons. Em organismos aquáticos como camarão e peixe, as condições da água
parecem ser determinantes na expressão diferencial de retrotransposons (Barbosa et al. 2014).
De modo geral, observamos neste estudo um aumento de Rex 6 em P. pardalis (Figura 1);
Rex 1 e Rex 6 em S. insignis (Figura 2) e Rex 1, Rex 3 e Rex 6 em C. amazonarum (Figura 3)
provenientes de ambiente poluído, em comparação ao ambiente não poluído, evidenciando um
comportamento diferencial destas sequências sob diferentes condições ambientais.
Interessantemente, o retroelemento Rex 6 apresentou maior sensibilidade às variações
ambientais, nas três espécies analisadas, destacando-se como um bom marcador para
ambientes inóspitos. Ao compararmos a distribuição dos elementos da família Rex com
marcadores clássicos e moleculares, percebemos que em P. pardalis há colocalização do Rex
6 com blocos heterocromáticos, além do primeiro par cromossômico do complemento possuir
maior concentração de heterocromatina em indivíduos do ambiente poluído. Ainda grande
número de marcações de Rex 1 está presente neste par cromossômico.
Em S. insignis, o padrão de localização física cromossômica, bem como o número de
sítios encontrados para Rex 1 e Rex 3 de indivíduos provenientes do lago Catalão (ambiente
não poluído) corroboraram os dados observados por Terencio et al. (2013) para a mesma
espécie. O Rex 1 está compartimentalizado na região centromérica de poucos cromossomos e
Rex 3 na região subterminal e terminal da maioria dos cromossomos. Porém, nos indivíduos
de ambiente poluído é notavelmente observado um maior número de marcações de Rex1, Rex
3 e Rex 6, distribuídas na maioria dos pares cromossômicos entre as espécies (Figura 2). Estas
marcações estiveram associadas com os sítios adicionais de DNAr 18S (colocalizados com as
regiões de heterocromatina), encontrados nos mesmos indivíduos do ambiente poluído (Silva
et al. 2019a).
Com relação à espécie C. amazonarum, o Rex 1 está colocalizado com a
heterocromatina, tanto no ambiente poluído quanto no não poluído (Silva et al. 2019a).
Marcações adicionais de Rex 3 estão presentes em regiões de eucromatina e de Rex 6 tanto
em regiões de eucromatina quanto de heterocromatina nos indivíduos de ambiente poluído
(Figura 3). Rex 1, Rex 3 e Rex 6 estão colocalizados com o bloco de DNAr 18S amplificado
no par 2 dos indivíduos do ambiente poluído, bem como com possíveis pseudogenes de DNAr

89
18S, indicando a dispersão destes sítios. Já o DNAr 5S (par 10) (Silva et al. 2019a) está
ladeado pelos três retrotransposons.
Com isso, seria possível pensarmos que, a partir da quantidade de Rex em um
cariótipo de uma dada espécie, conseguiríamos supor que ela pertence a um ambiente poluído
ou não? As condições ambientais podem alterar a atividade de TEs, alterar seu número de
cópias, bem como causar grandes alterações e instabilidade no genoma, principalmente por
meio de modificações no silenciamento epigenético (Coan e Martins 2018).
Consequentemente, os TEs podem ter um impacto na expressão gênica como resposta ao
estresse ambiental (ver Negi et al. 2016; Lanciano e Mirouze 2018). Em conjunto, nossos
achados configuram dados sólidos com o fato de que a ativação das sequências de Rex, em
resposta à poluição ambiental, pode estar relacionada a mudanças em seu status de metilação
e na susceptibilidade do hospedeiro (Casacuberta e González 2013), ou seja, podemos inferir
que as alterações aqui observadas no comportamento dos TEs, relacionadas ao ambiente
poluído, correspondem à ativação destas sequências de Rex nas especies aqui analisadas em
algum momento da sua história evolutiva.
Quando incorporados em regiões gênicas, os TEs geram diferentes tipos de rearranjos
genômicos por meio de transposições, que levam a inserções de genes, deleções, inversões e
duplicações (Alzohairy et al. 2013). Isto explicaria a discrepância que foi observada entre o
número de cópias do gene ribossomal DNAr 18S nestas mesmas espécies, provenientes de
ambiente poluído, quando comparadas com as do ambiente natural (Silva et al. 2019a), uma
vez que o mapeamento destes sítios mostrou colocalização com sequências de TEs, utilizadas
neste estudo. Sendo assim, TEs influenciados pelo estresse ambiental podem gerar a
diversidade bruta que as espécies necessitam para sobreviver nestas condições ao longo do
tempo evolutivo (Casacuberta e González 2013; Brawand et al. 2014).
Após eventos de invasão de TEs no genoma, a criação de novas variações genéticas
pode levar a um aumento na resistência a xenobióticos em diferentes organismos (Zeh et al.
2009; Casacuberta e González 2013). Ao regular a atividade transcricional de genes, os TEs
podem ter um forte impacto no fenótipo dos organismos hospedeiros (Lisch 2013). Porém, até
que ponto eles podem estar envolvidos na origem das características adaptativas? Diversos
trabalhos afirmam que após eventos de invasão, a adaptação ocorre em resposta a pressões
seletivas no novo ambiente (Lee 2002; Brawand et al. 2014) e tais organismos podem
responder com mudanças em suas características biológicas (Frederico et al. 2016) mas,
principalmente pela plasticidade genômica (Silva et al. 2016; Heinrichs-Caldas et al. 2019).

90
Recentemente, o dimorfismo da mariposa Biston betularia foi explicado pela presença
de TEs no primeiro íntron do córtex, um gene que está envolvido na via de pigmentação
(Van't Hof et al. 2016). Assim, a inserção de TEs contribuiu para uma vantagem seletiva da
espécie. Outro exemplo, foi verificado na instabilidade genômica diferencial, mediada pelo
acúmulo/aumento do retroelemento Rex 3, em uma espécie de peixe (Hoplosternum littorale)
que sobrevive em ambiente poluído. O trabalho revelou padrões distintos de marcadores
citogenéticos clássicos e moleculares intercalados com o retroelemento, indicando que tal
sequência pode desempenhar um papel funcional nos processos de adaptabilidade desta
espécie ao ambiente poluído (Silva et al. 2016), o que corrobora os dados encontrados neste
estudo.
Isto é particularmente importante se considerarmos que os ambientes urbanos
amazônicos vêm sofrendo alterações em consequência de ações de efluentes domésticos e
industriais, que também contribuem para o estabelecimento do complexo ecossistema
aquático amazônico (Santana e Barroncas 2007). Portanto, entender a atividade de TEs é, de
fato, um aspecto importante na elucidação da base molecular de mudanças fenotípicas
decorrentes das alterações ambientais (Lanciano e Mirouze 2018). Aqui, demonstramos que o
aumento e a influência dos TEs na modelagem do genoma nas espécies que vivem no
ambiente poluído é clara, quando comparados ao ambiente natural na região amazônica e que
seus impactos podem estar em constante evolução, bem como envolvidos na evolução
adaptativa das espécies de peixes que vivem em diferentes tipos de ambientes.

91
4.4 Artigo 4
Diversidade genética e dinâmica de elementos repetitivos em peixes exóticos
introduzidos na região Amazônica
Genetic diversity and dynamics of repetitive DNA elements of exotic fish introduced into the
Brazilian Amazon region. Artigo aceito para publicação como Short Notes - Adaptations of
Aquatic Biota, 2019.

92
4.4.1 Resumo
A adaptação rápida de espécies exóticas, introduzidas a novos habitats, confunde os
biólogos evolutivos há décadas. Embora essas espécies compartilhem uma série de
características ecológicas, pouco se sabe sobre os possíveis mecanismos genômicos ou
genéticos que possam contribuir para seu sucesso adaptativo. Neste contexto, marcadores
moleculares e citogenéticos foram utilizados no presente estudo para investigar uma possível
diferenciação genética de duas espécies exóticas de peixes (Oreochromis niloticus e
Trichopodus trichopterus) que podem determinar sua capacidade de adaptação a ambientes
poluídos na bacia amazônica. Os resultados das análises revelaram baixa diversidade genética
nas espécies estudadas, embora uma certa variação tenha sido encontrada nos sítios de DNAs
ribossomais, bem como a adição de heterocromatina e regiões de heterocromatinização
flanqueadas por elementos transponíveis (ETs), o que indica que a estrutura, organização e
função do genoma foram modificadas em indivíduos expostos à poluição ambiental,
provavelmente através de interações com os TEs.
Palavras-chave: DNA ribossomal, elemento transponível, heterocromatina, gene
mitocondrial, poluição.

93
4.4.2 Introdução
O ser humano vem modificando a distribuição das espécies no planeta de maneira
descontrolada. Em relação aos peixes, a introdução de espécies exóticas em ambientes nos
quais elas não existiam é, de fato, mediada pela atividade humana e algumas dessas espécies
tornam-se abundantes no ambiente onde foram introduzidas (Gardener et al. 2012). Em
consequência causam sérios problemas como redução da biota aquática local, mudanças na
estrutura e funcionamento das comunidades e dos ecossistemas aquáticos, além de se
tornarem poluentes biológicos (Souza et al. 2009; Abreu e Durigan 2011).
Entre as atividades humanas, apontadas como as principais causas no mecanismo de
dispersão e introdução de espécies exóticas em novos ambientes, incluem a pesca recreativa,
aquicultura e aquariofilia. Estas atividades podem modificar as características da comunidade
por meio de uma homogeneização antropogênica, que resulta na exclusão de espécies nativas,
por competição ou predação (Souza et al. 2009) e no aumento das espécies invasoras, devido
seu potencial de adaptação rápida, que as permitem se estabelecer e reproduzir mesmo em
situações de estresse (Novak 2007; Prentis et al. 2008). Os ciclídeos africanos, em particular
os da tribo Tilapini, são o grupo de peixes exóticos mais amplamente introduzidos no Brasil
(Sampaio et al. 2017), mas também existem registros da ocorrência de tricogastrinas asiáticas
no país (Cole et al. 1999). Nos córregos poluídos de Manaus, capital do Estado do Amazonas,
maior cidade da Amazônia brasileira ocidental, são encontrados tanto Oreochromis niloticus
(Cichlidae, Pseudocrenilabrinae), um ciclídeo africano, como Trichopodus trichopterus
(Osphronemidae, Trichogastrinae), um peixe ornamental endêmico do Sudeste Asiático.
Entretanto, para que possam enfrentar o novo ambiente, estas espécies devem ter uma
plasticidade tanto fenotípica quanto genotípica, desenvolvida durante seu processo evolutivo,
gerando linhagens múltiplas que, eventualmente, levam à especiação e à formação de
complexos de espécies (Chen et al. 2015; Braz-Mota et al. 2018; Heinrichs-Caldas et al.
2019). Diversos estudos com marcadores genéticos nucleares e mitocondriais já evidenciaram
que a variabilidade genética é o fator que garante uma rápida adaptação dos organismos a um
novo ambiente (Arif e Khan 2009). Outro elemento que tem sido considerado como um dos
principais responsáveis por esta plasticidade são os elementos repetitivos de DNA, dentre eles
os elementos transponíveis (ETs) e sítios ribossomais (Primack 2008; Casacuberta e González
2013, Silva et al. 2019a, b).

94
Os ETs são capazes de se mover dentro do genoma, podem alterar a atividade de
alguns genes, gerar duplicação, polimorfismos e variabilidade no número de cópias dentro e
entre espécies (Mita e Boeke 2016). Além disso, seus sítios de inserção influenciam na
estabilidade do genoma e na expressão gênica (Jurka et al. 2007; Bridier-Nahmias et al.
2015), uma vez que a transposição destes elementos móveis não é um processo aleatório,
podendo ser controlada por sinais celulares ou se tornar sensível e susceptível a mudanças
ambientais (Hua-Van et al. 2011; Silva 2019b). Além disso, os DNA ribossomais são
consideradas sequências importantes na evolução estrutural, dinâmica e funcional no genoma
de peixes, podendo estar associadas a rearranjos cromossômicos, variações cariotípicas e
desempenhando papéis fundamentais em seus processos adaptativos (Barros et al. 2017; Silva
et al. 2019a).
Embora variações intensas e repentinas no ambiente estejam em um contexto negativo,
tais perturbações podem facilitar uma adaptação rápida, afetando a estrutura, organização e
função do genoma por meio de interações com ETs, especialmente em populações com baixa
diversidade genética (Stapley et al. 2015). Desta forma, variações no ambiente podem
influenciar significativamente o número de cópias dos ETs e também seus níveis de expressão
(Hua-Van et al. 2011; Cowley e Oakey 2013). Em peixes, a família Rex de elementos
transponíveis é a mais estudada e estes são componentes importantes da plasticidade do
genoma e da resposta do organismo ao estresse ambiental (Rey et al. 2016; Suh et al. 2017).
Evidências de correlações de polimorfismo genético com heterogeneidade ambiental e
estresse já foram vistas em diferentes espécies, populações e indivíduos (Nevo 2001). A
instabilidade genômica diferencial, medida pelo aumento e acúmulo do retroelemento Rex 3,
foi registrada em uma comparação de populações de uma espécie de peixe (Hoplosternum
litoralle), encontrada em ambiente poluído com aquelas de ambiente não poluído (Silva et al.
2016). Neste contexto, o presente estudo avaliou a diversidade genética e mapeou
cromossomicamente elementos repetitivos de DNA (DNAr 18S, DNAr 5S, Rex 1 e Rex 3) em
cromossomos mitóticos de Tilápia do Nilo (Oreochromis niloticus) e do Gourami azul
(Trichopodus trichopterus), provenientes de ambientes poluídos na bacia amazônica
brasileira, com o objetivo de verificar se existem variações nos indivíduos que estão
adaptados a este ambiente exótico e antropizado.
4.4.3 Material e Métodos
Locais de coleta e amostragem

95
Indivíduos de O. niloticus e T. trichopterus foram coletados em três locais poluídos na
bacia amazônica: 1- Igarapé do Mindu [3 05 22,0 S, 60 05 31 W], 2- Igarapé do Quarenta [3
08 19,7 S, 60 56,7 W] e 3- Igarapé do Tarumã-açu [3 00 51,1 S, 60 3 33,2 W]. Estes corpos
de água drenam áreas densamente habitadas na cidade de Manaus-AM e apresentam uma
intensa poluição ambiental devido a atividades domésticas, industriais, armazenamento
inapropriado de lixo proveniente do Polo Industrial, lixo urbano e hospitalar (Pinto et al.
2009; Silva et al. 2019e). No total, dezessete indivíduos da espécie O. niloticus e dezoito
indivíduos da espécie T. trichopterus foram coletados sob a permissão do Instituto Chico
Mendes de Conservação da Biodiversidade (ICMBio/SISBIO número: 7634-1/2015)
(Tabela1).

96
Tabela 1 – Espécies de peixes exóticas analisadas e seus respectivos locais de coleta na cidade de Manaus-AM.
Ordem Família Subfamília Espécie Locais de coleta Número de
exemplares Voucher
Perciformes
Cichlidae Pseudocrenilabrinae
Oreochromis
niloticus
(Linnaeus, 1758)
Igarapé do Mindu 8 (três machos e cinco
fêmeas) INPA-ICT 023939
Igarapé do Tarumã-açu 9 (quatro machos e
cinco fêmeas) INPA-ICT 023938
Osphronemidae Trichogastrinae
Trichopodus
trichopterus
( Pallas, 1970)
Igarapé do Mindu
9 (um macho, duas
fêmeas e seis sem
identificação sexual)
INPA-ICT 023926
Igarapé do Quarenta
8 (dois machos, quatro
fêmeas e dois sem
identificação sexual)
INPA-ICT 023927

97
Marcadores moleculares e análises citogenéticas
Técnicas citogenéticas clássicas
Os exemplares coletados foram eutanasiados seguindo as Diretrizes da Prática da
Eutanásia do Concea (Concea 2013) e com aprovação da Comissão de Ética no Uso de
Animais (CEUA) (no. 011/2017) do Instituto Nacional de Pesquisas da Amazônia (INPA). A
identificação das espécies foi realizada pelo especialista Jansen Alfredo Sampaio Zuanon e,
após a realização dos procedimentos citogenéticos, os exemplares testemunho foram
depositados na Coleção de Peixes do INPA, conforme vouchers na Tabela 1. Cromossomos
mitóticos foram obtidos a partir de células renais, utilizando o protocolo descrito por Bertollo
et al. (1978). Para a detecção da heterocromatina constitutiva (Banda C) foi utilizada a técnica
descrita por Sumner (1972), enquanto para as regiões organizadoras de nucléolo (RONs)
aplicou-se a técnica descrita por Howell e Black (1980).
Técnicas moleculares
O DNA genômico foi extraído segundo o protocolo de Fenol-Clorofórmio de
Sambrook e Russel (2001). O gene mitocondrial citocromo c oxidase subunidade I (COI) foi
amplificado por Reação em Cadeia da Polimerase (PCR - Saiki et al. 1988) utilizando
coquetéis de primers descrito por Ivanova et al. (2007):
- (FishF2 5’-GTAAAACGACGGCCAGTCGACTAATCATAAAGATATCGGCAC-3’,
FishR2 5’-CAGGAAACAGCTATGACACTTCAGGGTGACCGAAGAATCAGAA -3’,
VF2 5’-GTAAAACGACGGCCAGTCAACCAACCACAAAGACATTGGCAC-3’ e VR1d
5’- CAGGAAACAGCTATGACTAGACTTCTGGGTGGCCRAARAAYCA-3’).
Os produtos de PCR foram sequenciados usando o kit Big Dye (Applied Biosystems)
no sequenciador automático ABI 3500. O alinhamento das sequências geradas foi realizado
através da ferramenta MAFFT, implementada no software Geneious 7.0.6 (Kearse et al. 2012)
e editadas manualmente. Para a localização física cromossômica dos marcadores citogenéticos
(DNAr 18S, DNAr 5S, Rex 1 e Rex 3), já identificados e caracterizados, foram utilizados os
seguintes primers para amplificação via PCR:
-DNAr 18S - 18Sf (5’-CCG CTT TGG TGA CTC TTG AT-3’) e 18Sr (5’-CCG AGGACC
TCA CTA AAC CA-3’) (Gross et al. 2010);
-DNAr 5S - 5Sf (5’- CGA GGTCGGGCCTGGTTAGTA-3’) e 5Sr (5’-
CTTCYGAGATCAGACGAGATC-3’ (Carvalho et al. 2016);

98
-Rex 1 - (RTX1-F1 5’TTC TCC AGT GCC TTC AAC ACC e RTX1-R3 5’TCC CTC AGC
AGA AAG AGT CTG CTC) (Volff et al. 1999; 2000);
-Rex 3 - (RTX3-F3 5’CGG TGA YAA AGG GCA GCC CTG e RTX3-R3 5’TGG CAG
ACN GGG GTG GTG GT) (Volff et al. 1999; 2000)
Os produtos de PCR dos marcadores citogenéticos foram marcados por nick
translation com biotina-14-dATP (Biotin Nick Translation mix; Invitrogen) e
digoxigenina11-dUTP (Dig-Nick Translation mix; Roche), seguindo as instruções do
fabricante. Hibridizações in situ fluorescentes, utilizando sondas homólogas, foram realizadas
seguindo o protocolo descrito por Pinkel et al. (1986), com 77% de estringência (2,5 ng/µL de
sonda marcada, 50% formamida, 10% sulfato dextrano, 2XSSC a 37 °C por 18 h). Os
cromossomos foram contracorados com DAPI (4’, 6-diamidino-2-fenilindol) (2 µg/mL), em
meio de montagem Vectashield (Vector).
Os cromossomos foram analisados em microscópio de epifluorescência Olympus
BX51 e as imagens capturadas com câmera digital (Olympus DP71), usando o software
Image-Pro MC 6.3. Os cariótipos foram organizados utilizando o programa Adobe Photoshop
7.0, versão CS5. Os cromossomos foram medidos no programa Image J e classificados de
acordo com a nomenclatura proposta por Levan et al. (1964). Para determinação do número
fundamental (NF) de braços cromossômicos, de acordo com a classificação de Thompson
(1979) foram considerados cromossomos meta/submetacêntricos (m/sm) como portadores de
dois braços e subtelo/acrocêntricos (st/a) portadores de um braço.
4.4.4 Resultados
Sequências de DNA mitocondrial
Foi observada baixa diversidade genética em O. niloticus, proveniente dos igarapés do
Mindu e Tarumã-açu e em T. trichopterus, proveniente dos igarapés do Mindu e Quarenta,
sendo recuperado um único haplótipo COI por espécie, independentemente do local de coleta
(Tabela 1). As sequências de COI apresentaram 650 pb e foram depositadas no GenBank,
com os seguintes números de acesso: MG714797 a MG714814 para O. niloticus e MG714779
a MG714796 para T. trichopterus. A frequência nucleotídica das sequências foram:
A=23,54%, C=30,15%, G=18,00% e T=28,31% para O. niloticus e A=23,78%, C=26,52%,
G=18,29% e T=31,40% para T. trichopterus.
Caracterização cromossômica das espécies exóticas

99
Em O. niloticus não foram observadas diferenças cariotípicas entre os indivíduos
provenientes dos igarapés do Mindu e Tarumã-açu, que apresentaram número diploide de
2n=44 cromossomos, com a presença de um grande par st/a no complemento, fórmula
cariotípica de 2m + 42st/a, NF igual a 46 e RONs múltiplas, localizadas terminalmente nos
braços longos do par 6 e nos braços curtos do par 8 (Figura 1A). Com relação à distribuição
da heterocromatina, blocos foram evidenciados na região pericentromérica de todos
cromossomos e adição de blocos tênues na região terminal de alguns cromossomos, revelando
pouca diferença entre os indivíduos dos igarapés analisados. No entanto, acúmulo de
heterocromatina foi perceptível no braço longo do maior par st/a de todos os indivíduos
analisados (Figura 1A). O DNAr 18S foi coincidente com as RONs (pares 6 e 8). Porém, uma
quinta marcação foi evidenciada em um dos homólogos do par 17 dos indivíduos do igarapé
Tarumã-açu, diferentemente dos indivíduos do igarapé do Mindu, que apresentaram uma
marcação extra, localizada em um dos homólogos do maior par st/a (par 1). Sítios de DNAr
5S foram localizados na região distal do par 8 e pericentromérica do par 16 (Figura 1B). Com
relação aos retroelementos Rex 1 e Rex 3, ambos apresentaram uma distribuição dispersa no
genoma da espécie, com marcações ao longo de braços cromossômicos, sendo evidente
principalmente, em toda extensão do maior par st/a do complemento (Figura 1C; D).
Trichopodus trichopterus apresenta 2n=46 cromossomos, em uma série gradual de
st/a, NF igual a 46 e RON simples, localizada na região terminal do braço longo do par 2
(Figura 1E). Quanto à heterocromatina, variabilidade intra e interindividual foi revelada em
indivíduos de ambos locais de coleta, havendo blocos conspícuos nos primeiros pares do
complemento nos indivíduos do igarapé do Quarenta, quando comparados com indivíduos
coletados no igarapé do Mindu (Figura 1E). Sítio de DNAr 18S foi localizado em um único
par cromossômico da espécie, sendo coincidente com o par 2 da RON (Figura 1F). DNAr 5S
foi observado na região pericentromérica do par 4 e uma terceira marcação localizada na
região pericentromérica de um dos homólogos do par 5 de todos os indivíduos analisados
(Figura 1F). Quanto aos Rex 1 e Rex 3, blocos compartimentalizados foram evidenciados na
maioria dos pares cromossômicos e com sítios invadindo a extensão de braços
cromossômicos, principalmente nos primeiros pares do complemento (Figura 1G; H).

100
Figura 1 - Cariótipos das espécies Oreochromis niloticus (igarapés do Mindu e Tarumã-açu) (A, B, C e D) e
Trichopodus trichopterus (igarapés do Mindu e Quarenta) (E, F, G e H). (A e E) Padrão de distribuição da
heterocromatina; (A1 e E1) Cromossomos portadores da RON; (B e F) Sítios de DNAr 18S (sinal vermelho)
(B1: Padrão dos indivíduos do igarapé do Mindu) e de DNAr 5S (sinal verde); (C e G) Localização física
cromossômica do retroelemento Rex 1 (sinal verde); (D e H) Localização física cromossômica do retroelemento
Rex 3 (sinal vermelho). Os cromossomos foram contracorados com DAPI. Barra de escala igual a 20 μm.

101
4.4.5 Discussão
O sucesso da invasão de espécies exóticas pode ser influenciado pelas modificações
genéticas dentro de suas populações (Reznick e Ghalambor 2001; Lee 2002). Neste contexto,
o gene COI, que é conhecido por ser altamente variável, pode ser considerado útil para a
análise de linhagens e a identificação de populações estreitamente relacionadas (Lefébure et
al. 2006; Witt et al. 2006). No presente estudo, as sequências de COI revelaram baixos níveis
de diversidade genética nas populações de ambas as espécies. Diante disso, parece improvável
que a variação encontrada na organização genômica dos indivíduos coletados nos diferentes
igarapés de Manaus esteja relacionada à presença de mais de uma linhagem genética.
Em T. trichopterus foi possível observar um aumento da heterocromatina
principalmente nos primeiros pares do complemento dos indivíduos provenientes do igarapé
do Quarenta, diferente dos indivíduos do igarapé do Mindu (Figura 1E), o que poderia estar
associado com diferentes fontes de contaminação, uma vez que um único haplótipo COI foi
recuperado para a espécie. Resultados semelhantes foram obtidos, utilizando o tamoatá
(Hoplosternum littorale) de ambiente poluído e não poluído também de Manaus, AM, sendo
observada uma instabilidade genômica promovida pelo ambiente, com padrões distintos de
heterocromatina ricas em ETs em função de padrões distintos de contaminação (Gross et al.
2012; Silva et al. 2016).
Em contrapartida O. niloticus não apresentou diferenças no padrão heterocromático
entre os diferentes ambientes amostrados no presente trabalho. Porém, houveram diferenças
se considerado o padrão descrito para a espécie (Chew et al. 2002; Poletto et al. 2010;
Mazzuchelli et al. 2012), sendo perceptível um aumento em porções heterocromáticas de
alguns pares, bem como um acúmulo no braço longo do maior par st/a de todos os indivíduos
(Figura 1A).
Regiões heterocromáticas normalmente atuam na segregação cromossômica e na
regulação da expressão gênica associadas às respostas ambientais (Grewal e Jia 2007;
Varriale et al. 2008; Buhler 2009). Tanto para O. niloticus quanto para T. trichopterus, a
heterocromatina se mostrou colocalizada com os ETs analisados, o que indica a preferência de
ETs ativos a certos padrões de inserção (Green et al. 2012) e esses padrões podem acarretar
em certas implicações, como no processo de heterocromatinização e na adição de
heterocromatina, com intuito de minimizar os efeitos deletérios de transposição (Boeke e
Devine 1998). Evidentemente, os ETs mobilizados podem reestruturar o genoma, gerando

102
modificações genéticas necessárias para uma rápida diversificação (Oliver e Greene 2009;
Zeh et al. 2009).
O padrão de distribuição dos retroelementos Rex 1 (Figura 1C; G) e Rex 3 (Figura 1D;
H) obtidos no presente estudo indicam que O. niloticus e T. trichopterus de ambientes
poluídos apresentaram uma composição e intensidade muito maior de marcações, se
comparado com os descritos na literatura (Volff et al. 1999; Valente et al. 2011). Diversos
trabalhos afirmam que após eventos de invasão, a adaptação ocorre em resposta a pressões
seletivas no novo ambiente (Lee 2002; Brawand et al 2014) e tais organismos podem
responder com mudanças em suas características biológicas (Carroll et al. 2001; Frederico et
al. 2016) mas, principalmente pela plasticidade genômica (Weinig 2000; Chevin et al. 2010).
Nesse contexto, os TEs promovem a evolução e a plasticidade do genoma dos peixes
através da facilitação de mutações em grande escala, contribuindo para a variabilidade
genética desses organismos e, em última análise, para seu sucesso ecológico (Hua-Van et al.
2011; Philippsen 2014). Os TEs influenciados pelo estresse ambiental podem, assim, gerar a
diversidade bruta que uma espécie exige para sobreviver sob essas condições ao longo de uma
escala de tempo evolutiva (Casacuberta e González 2013; Brawand et al. 2014). Isso
explicaria o aumento e a disseminação desses elementos nos genomas de ambas as espécies
analisadas no presente estudo, após sua introdução em ambientes poluídos na região
amazônica. Isto demonstra claramente que a atividade de TE é induzida pela exposição ao
novo ambiente, criando-se assim, novas variações genéticas (Naito et al. 2006; Mills et al.
2007).
Com relação aos DNAs ribossomais, estas regiões genômicas têm sido consideradas
instáveis, frágeis e susceptíveis a danos ocasionados por diversos fatores, inclusive ambientais
(Kobayashi 2006; 2008). A variabilidade dos sítios de DNAr, presente em peixes amazônicos,
tem sido atribuída a: a) ocorrência de translocações, envolvendo regiões heterocromáticas
ricas em ETs (Gross et al. 2010); b) diferentes classes de sequências repetitivas, presentes em
regiões de heterocromatina (Schneider et al. 2013a); c) alguns sítios derivados de
pseudogenes, como consequência de duplicações e transposições de DNA e pela associação
destas sequências com ETs (Terencio et al. 2012).

103
Em O. niloticus, além dos sítios de DNAr 18S coincidentes com as RONs, que
corroboram os dados obtidos por Martins et al. (2000; 2002) e Leonhardt et al. (2006), uma
quinta marcação foi evidenciada, a qual se diferenciou entre os locais de estudo (Figura 1B).
Estes dados não podem ser atribuídos à erros de hibridização com a técnica de FISH, uma vez
que este processo foi realizado sob condições de alta estringência e com replicatas utilizando
sondas homólogas. Assim, é provável que haja associação destes sítios com outros elementos
transponíveis (ETs). Este fato já foi relatado para Hoplosternum littorale (tamoatá),
provenientes de igarapés poluídos da cidade de Manaus (AM) (Silva et al. 2016).
Para T. trichopterus, uma terceira marcação do DNAr 5S se fez presente no genoma de
todos os indivíduos analisados (Figura 1F), discordando dos dados de Pazza et al. (2009), que
encontraram apenas um par de cromossomos marcados. Esta variabilidade encontrada nos
sítios de DNAr 18S e 5S nas duas espécies pode ser resultado de diversificação
cromossômica, em decorrência de rearranjos, causados pela ativação de ETs (Molina et al.
2014; Brawand et al. 2014; Chadha e Sharma 2014) uma vez que, mostraram-se colocalizados
com Rex 1 e Rex 3. Uma vez ativos, estes elementos podem contribuir com rearranjos
genômicos (Maumus et al. 2009), conferir a genes próximos ou colocalizados a capacidade de
responder às mudanças ambientais (Guo e Levin 2010; Servant et al. 2012), gerar novas redes
regulatórias (Naito et al. 2009; Ito et al. 2011) bem como, aumentar o número de cópias de
genes pelo genoma (Dai et al. 2007).
Conter ETs potencialmente ativos pode aumentar a capacidade do genoma de lidar
com certas mudanças ambientais (Casacuberta e González 2013; Chadha e Sharma 2014;
Brawand et al. 2014). Porém, ETs parecem ter desenvolvido mecanismos que direcionam sua
integração para regiões heterocromáticas dentro do genoma ou promovem sua
heterocromatinização (Cecco et al. 2013; Sotero-Caio et al. 2017).
Embora muitos estudos nas últimas décadas tenham revelado respostas moleculares ao
estresse ambiental, a maneira pela qual essas respostas ocorrem, em nível organizacional do
genoma, ainda é pouco compreendida (Casacuberta e Gonzalez 2013; Chadha e Sharma
2014). Os resultados do presente estudo, no entanto, indicam que os elementos de DNA
repetitivos, quando expostos a condições ambientais adversas, podem ser os principais
responsáveis pela adaptação dos genomas de O. niloticus e T. trichopterus, revelando uma
configuração única das sequências repetitivas do DNA ribossomal, além da heterocromatina,
e a heterocromatinização de regiões cromossômicas distintas. Isso indica que a atividade dos

104
TEs pode ser de fundamental importância para a capacidade de ambas espécies exóticas se
adaptarem rapidamente à poluição ambiental semelhante ao observado em espécies nativas.

105
4.5 Artigo 5
Contaminantes ambientais e seus impactos na integridade genética de
espécies de peixes na bacia amazônica
Artigo será enviado para publicação no periódico Journal Aquatic Toxicology.

106
4.5.1 Resumo
Organismos aquáticos, principalmente populações de peixes, estão sendo
profundamente impactadas pela poluição ambiental em toda a região Neotropical. Neste
contexto, análises da integridade do DNA de organismos aquáticos têm sido amplamente
aceitas como um método adequado para avaliar a contaminação genotóxica do ambiente, já
que análises físico-químicas da água identificam apenas a presença e as respectivas
concentrações de diferentes poluentes. Neste estudo avaliamos a integridade genética de
espécies de peixes que habitam ambientes antropizados e não antropizados da Amazônia
central, estabelecendo uma relação entre os dados citogenéticos encontrados e os parâmetros
físico-químicos destes ambientes aquáticos. Nossos resultados demonstram que a análise de
micronúcleos, de anormalidades nucleares eritrocíticas e teste do cometa são biomarcadores
importantes para avaliar a saúde do ecossistema aquático, especialmente em espécies que
estão apresentando remodelamento genético e epigenético, em função da qualidade da água
dos igarapés urbanos da cidade de Manaus-AM, que sofrem com a poluição ambiental.
Palavras-chave: Amazônia; Cometa; Micronúcleo; Qualidade da água; Poluição.

107
4.5.2 Introdução
A cidade de Manaus, capital do Amazonas, em sua área urbana, é atravessada por três
bacias hidrográficas. Duas microbacias encontram-se inteiramente urbanizadas, como as do
igarapé do São Raimundo e do Educandos que drenam áreas densamente habitadas e com
atividades domésticas (lixo e esgoto sanitário) e industriais em seu entorno, desde suas
nascentes até sua foz (Pinto et al. 2009; Nogueira et al. 2015). As demais drenam
principalmente a área rural, tais como as bacias do Tarumã-açu e do Puraquequara (Pinto et
al. 2009; Ferreira et al. 2012). O aterro sanitário da cidade de Manaus está situado nas
margens do igarapé do Tarumã-açu, com armazenamento inapropriado de altas quantidades
de lixo, provenientes do Polo Industrial, lixo urbano e hospitalar (Do Valle 1998; Piedade et
al. 2014).
De maneira geral, nas últimas décadas, a industrialização e as diferentes influências
antropogênicas resultaram em uma maior presença de contaminantes em ecossistemas
aquáticos (Sunjog et al. 2013). Como consequência, expressiva queda na qualidade da água e
perda da biodiversidade aquática tem sido observada (Goulart e Callisto 2003; Zappata et al.
2016; Kolarevic et al. 2017). Isso ocorre porque estas descargas poluidoras contêm uma
mistura de contaminantes orgânicos e inorgânicos, que podem acarretar múltiplos efeitos
biológicos, bem como induzir alterações genéticas e influenciar a integridade de moléculas de
DNA em organismos aquáticos, devido às interações com múltiplas sequências alvo nos
organismos expostos (Kolarević et al. 2016; Thellmann et al. 2017).
Estes organismos aquáticos estão sendo profundamente impactados pela poluição
ambiental em toda a região Neotropical (Barroca 2012), visto que a capacidade de diferentes
espécies de persistirem em ecossistemas contaminados depende de sua capacidade adaptativa,
que está relacionada à sua variabilidade genética (Gross et al. 2010; Silva et al. 2016; Silva et
al. 2019a). Isto foi possível observar com a variação encontrada nos sítios de DNAs
ribossomais nas espécies Hoplosternum littorale, Semaprochilodus insignis, Hoplias
malabaricus, Pygocentrus nattereri e Cichlasoma amazonarum evidenciando um aumento no
número de sítios de DNAs ribossomais, bem como a adição de heterocromatina, regiões de
heterocromatinização flanqueadas por TEs e por um comportamento diferencial dos
retrotransposons Rex 1, Rex 3 e Rex 6 nos cromossomos de indivíduos de uma mesma
espécie, mas provenientes de diferentes populações que habitam ambientes poluídos e não
poluídos. Isso demonstra uma instabilidade cromossômica no genoma dessas espécies, que

108
poderia ser um indicativo de perda de material genético (Silva et al. 2016; Silva et al. 2019a;
c; Silva 2019b).
Neste contexto, testes biológicos para toxicidade e genotoxicidade são indispensáveis
na avaliação das reações que os organismos vivos apresentam quando em contato com
poluição ambiental complexa e indicação dos efeitos sinergísticos potenciais de vários
poluentes (Matsumoto et al. 2006; Xavier et al. 2011). Isto porque, análises físico-químicas
da água identificam apenas a presença e as respectivas concentrações dos diferentes poluentes
(Matsumoto et al. 2006). Assim, a análise da integridade do DNA de organismos aquáticos
tem sido amplamente aceita como um método adequado para avaliar a contaminação
genotóxica do ambiente (Frenzilli et al. 2009). Dentre os principais testes biológicos para
toxicidade e genotoxicidade estão o teste do micronúcleo, anormalidades nucleares e o ensaio
do cometa (Bombail et al. 2001; Zappata et al. 2016). Estes testes biológicos mensuram
diferentes alterações genéticas e geram informações complementares, em um curto prazo de
tempo, na avaliação dos danos no DNA de peixes expostos a vários xenobióticos no ambiente
aquático (Dhawan et al. 2009; Sunjog et al. 2013).
Portanto, neste estudo avaliamos a integridade genética de espécies de peixes que
habitam ambientes antropizados e não antropizados da Amazônia central, estabelecendo uma
relação entre os dados citogenéticos encontrados e parâmetros físico-químicos destes
ambientes aquáticos. Assumimos como pressupostos que estas mesmas espécies acumulam
diferenças estruturais em seu genoma, em função do ambiente em que vivem, além de
apresentarem um grande valor comercial na região, por serem muito apreciados por
populações ribeirinhas e urbanas da Amazônia brasileira como fonte de alimentação.
4.5.3 Material e Métodos
Caracterização da área e amostragem do estudo
Amostras de água e espécies de peixes foram coletadas em quatro locais da bacia
amazônica central, sendo o lago Catalão [3 13 22.3 S, 59 55 14.5 W] considerado não
contaminado e os outros três locais considerados contaminados na bacia amazônica, drenando
áreas com grande densidade populacional na cidade de Manaus: 1- Igarapé do Mindu [3 05
22.0 S, 60 0 5.31 W]; 2- Igarapé do Quarenta [3 08 19.7 S, 60 0 56.7 W]; 3- Igarapé do
Tarumã-açu [3 00 51.1 S, 60 3 33.2 W] (Pinto et al. 2009; Piedade et al. 2014).

109
O igarapé do Mindu, principal tributário da microbacia do São Raimundo, apresenta
contaminantes oriundos de esgotos e lixos domésticos em suas águas. O igarapé do Quarenta,
um dos principais tributários da microbacia do Educandos, passa pelo Distrito Industrial da
cidade e recebe efluentes tanto industriais quanto domésticos. O igarapé do Tarumã-açu
recebe um aporte de chorume do único aterro sanitário da cidade (Figura 1) (Pinto et al. 2009;
Piedade et al. 2014). Para uma análise comparativa dos testes biológicos foram utilizadas
amostras das mesmas espécies coletadas nestes igarapés (ambiente poluído) e do lago Catalão,
considerado um ambiente não poluído, que fica localizado na confluência dos rios Negro e
Solimões, e apresenta água misturada, mas com características físico-químicas de água branca
(Pinto et al. 2009).
Cinco espécies de peixes, que apresentaram maior abundância nos igarapés poluídos e
utilizadas nos estudos anteriores (Silva et al. 2019a, d), foram escolhidas como organismos
modelo, sendo três espécies amazônicas (Pterygoplichthys pardalis, Hoplias malabaricus e
Semaprochilodus insignis) presentes nos dois tipos de ambiente (poluído e não poluído) e
duas espécies exóticas (Oreochromis niloticus e T. trichopterus) presente somente no
ambiente poluído (Tabela 1). As coletas foram realizadas sob a permissão do Instituto Chico
Mendes de Conservação da Biodiversidade (ICMBio/SISBIO número: 7634-1/2015). Os
exemplares coletados foram eutanasiados seguindo as Diretrizes da Prática da Eutanásia do
Concea (Concea 2013) e com aprovação da Comissão de Ética no Uso de Animais (CEUA)
(no. 011/2017) do Instituto Nacional de Pesquisas da Amazônia (INPA). Após a realização
dos procedimentos, os exemplares testemunhos foram depositados na Coleção de Peixes do
INPA.

110
Figura 1 – Bacias hidrográficas da cidade de Manaus-AM.
Figura 1 – Bacias hidrográficas da cidade de Manaus-AM. Marcação em azul (igarapé do Tarumã-açu);
marcação em vermelho (igarapé do Mindu) e marcação em amarelo (igarapé do Quarenta).

111
Ordem Família Espécie Locais de amostragem Número de exemplares Voucher
Siluriformes Loricariidae Pterygoplichthys pardalis
(Castelnau 1855)
Igarapé do Mindu 1 macho e 1fêmea INPA-ICT 023942
Igarapé do Tarumã-açu 2 machos e 2 fêmeas INPA-ICT 023941
Lago Catalão 1 macho, 1 fêmea e 2 sem
identificação sexual INPA-ICT 57621
Characiformes
Erythrinidae Hoplias malabaricus
(Block 1794)
Igarapé do Tarumã-açu 2 machos e 1 fêmea INPA-ICT 023929
Lago Catalão 3 machos e 3 fêmeas INPA-ICT 57622
Prochilodontidae Semaprochilodus insignis
(Jardine 1841)
Igarapé do Tarumã-açu 2 fêmeas e 3 sem identificação
sexual INPA-ICT 023933
Lago Catalão 5 machos, 1 fêmea e 1 sem
identificação sexual INPA-ICT 57619
Cichlidae Oreochromis niloticus
(Linnaeus 1758)
Igarapé do Mindu 3 machos e 5 fêmeas INPA-ICT 023939
Igarapé do Tarumã-açu 4 machos e 5 fêmeas INPA-ICT 023938
Osphronemidae Trichopodus trichopterus
(Pallas 1770)
Igarapé do Mindu 1 macho, 2 fêmeas e 6 sem
identificação sexual INPA-ICT 023926
Igarapé do Quarenta 2 machos, 4 fêmeas e 2 sem
identificação sexual INPA-ICT 023927
Tabela 1 - Espécies de peixes analisadas e seus respectivos locais de coleta.

112
Variáveis ambientais
As coletas de água nos dois tipos de ambiente ocorreram no período de 2015 a 2017
em três pontos georeferenciados para cada local de coleta. Dentre os parâmetros de qualidade
da água monitorados utilizou-se: Oxigênio dissolvido (OD), pH e condutividade elétrica (CE),
medidos nos locais de coleta, com uso de um pHmetro portátil (modelo 370 pH meter -
JENWAY) e medidores de CE/temperatura (modelo 55-25 FT - YSI); turbidez com o uso de
turbidímetro (modelo HI98703 - HANNA), pelo método Nefelométrico, sendo estes
parâmetros analisados em triplicata. Os métodos analíticos, bem como a preservação das
amostras seguiram os procedimentos definidos pelo Standard Methods (APHA 1998).
Teste do micronúcleo (MN) e anormalidades nucleares eritrocíticas (ANEs)
Após a coleta de sangue de todos os indivíduos foram preparadas duas lâminas de
esfregaços sanguíneos. As lâminas foram fixadas em etanol PA (100%) e coradas com
Giemsa 5% (diluído em tampão fosfato pH 6,8) para análise. Para estimar a quantidade total
de eritrócitos contendo MN e ANEs foram contabilizados 2000 eritrócitos (1000 células por
lâmina) por indivíduo (Bucker et al. 2006). Os MN foram identificados, seguindo o critério
proposto por Fenech et al. (2003) e Freitas et al. (2005) e a diferenciação das ANEs de acordo
com Carrasco et al. (1990).
Ensaio do cometa
Para o ensaio do cometa foi utilizada a técnica descrita por Singh et al. (1988) com
algumas modificações. Foram contabilizados 50 eritrócitos por indivíduo em microscópio
óptico e analisados no programa de imagens em PC de domínio público entre plataformas
para o ensaio do cometa CAPS_1.2.3b1.exe. Os danos foram classificados em categorias que
variam de 0 a 4, referentes à extensão de dano no DNA na cauda do cometa por uma regra de
três simples: 0 = sem danos (<5 %); 1 = baixo nível de danos (5-20 %); 2 = médio nível de
danos (20-40 %); 3 = alto nível de danos (40-95 %); 4 = totalmente danificados (>95 %)
(Hartmann 1995; Brianezi 2011) (Figura 2).

113
Análise estatística
Para as variáveis ambientais foi realizada uma análise de componentes principais
(ACP) para ordenar os dados e uma PERMANOVA (análise multivariada por permutação)
para testar se existia diferença entre os igarapés baseado nas variáveis da água. Para os testes
genéticos foi usado o teste de Kruskall-Wallis com comparações múltiplas, utilizando
correção de Dunn, com significância estatística p<0,05 para comparar métricas genéticas
entre igarapés, sendo realizada uma análise por espécie para cada métrica genética. Toda a
análise estatística foi realizada utilizando o pacote estatístico R (R Core Development Team
2016).
4.5.4 Resultados
Caracterização físico-química dos ambientes
Os dados limnológicos analisados estão apresentados na Tabela 2, incluindo as médias
e o desvio-padrão dos perfis físico-químicos para cada local amostrado durante as estações
seca e cheia, confrontando com os mesmos parâmetros estabelecidos nas Resoluções do
Conselho Nacional do Meio Ambiente (CONAMA 410/2009 e 430/2011). Parâmetros como
pH, OD e turbidez apresentaram-se fora dos parâmetros estabelecidos na resolução do
CONAMA para alguns locais de coleta.
Figura 2 – Classificação do ensaio do cometa de acordo com Hartmann (1995) e Brianezi (2011).

114
Tabela 2 - Caracterização dos perfis físico-químicos dos igarapés urbanos poluídos de Manaus-AM nos períodos de seca e
cheia e do ambiente natural, lago Catalão.
Parâmetros de
qualidade da
água
Igarapé do Mindu Igarapé do Quarenta Igarapé do Tarumã-açu
Lago Catalão Valores de Referência
CONAMA nº 430/2011 SECA CHEIA SECA CHEIA SECA CHEIA
pH 6,56±0,22 7,13±0,18 6,66±0,25 6,69±0,31 5,75±0,24 6,35±1,32 6,7 6,00 - 9,00
Temperatura (°C) 29,66±0,06 29,83±1,27 29,99±2,46 30,18±3,02 27,88±1,39 27,67±2,14 29,7 Máximo: 40 (°C)
OD (mgL-1) 3,16±3,30 2,94±2,89 3,98±1,95 3,85±2,52 4,76±1,30 5,77±3,67 3,7 ≥ 5,00 mg/bL-1
Condutividade
elétrica (µSmL-1) 179,55±140,02 343,96±68,10 261,72±72,61 221,30±94,81 30,69±17,28 25,27±19,83 26,0 *
Turbidez (NTU) 254,59±253,13 118,10±106,21 401,78±163,16 44,71±18,18 71,91±28,07 33,80±26,79 90,0 ≤100 NTU
*Valores em negrito representam os parâmetros que estão fora dos estabelecidos na resolução do CONAMA.

115
A análise de componentes principais (ACP) das variáveis ambientais (pH,
temperatura, OD, CE e turbidez) agrupou três conjuntos relevantes, comparando os ambientes
igarapé do Mindu, igarapé do Quarenta, igarapé do Tarumã-açu e lago Catalão (Tabela 3). O
conjunto 1 compilou os igarapés do Mindu e do Quarenta, considerados ambientes totalmente
poluídos. No conjunto 2 o agrupamento se deu com um ambiente cuja poluição ainda é
recente e o conjunto 3 com o ambiente não poluído (Figura 3).
Tabela 3 - Análise multivariada de variância permutacional comparando os locais.
Pares Modelo F Valor de p
(>F)
Igarapé do Mindu x Igarapé do Quarenta 0.102 0.907
Igarapé do Mindu x Igarapé do Tarumã-açu 7.738 0.010*
Igarapé do Mindu x Lago Catalão 8.970 0.010*
Igarapé do Quarenta x Igarapé do Tarumã-açu 7.713 0.010*
Igarapé do Quarenta x Lago Catalão 8.978 0.006*
Igarapé do Tarumã-açu x Lago Catalão 4.990 0.010*
-2 -1 0 1 2
-1
0
1
2
PC1
PC
2
Local
Mindu
Quarenta
Tarumã-açu
Catalão
Figura 3 - Análise de componentes principais (ACP) das variáveis ambientais (pH, temperatura,
OD, CE e turbidez) dos igarapés do Mindu, Quarenta, Tarumã-açu e lago Catalão.
*Valores significativos (p<0,05)

116
Marcadores de genotoxicidade
A análise de esfregaços sanguíneos das espécies coletadas nos ambientes considerados
poluídos e no ambiente não poluído evidenciou em maior número células com morfologia
nuclear característica (Figura 4A). Contudo, foram observadas nas cinco espécies analisadas
células micronucleadas (Figura 4B); núcleos em forma de rim (Figura 4C); núcleos lobados
(Figura 4D); núcleos segmentados (Figura 4E) e núcleos vacuolados (Figura 4F). Para os
testes, a média e o desvio padrão por espécie são detalhados na Tabela 4. Analisando a
frequência do MN, diferenças significativas foram encontradas para a espécie S. insignis de
ambiente poluído quando comparada com o grupo controle (p-value = 0.040). Para as demais
espécies a quantidade de MN foi similar entre o ambiente poluído e não poluído: H.
malabaricus (p-value = 0.413), P. pardalis (p-value = 0.465), O. niloticus (p-value = 0.986) e
T. trichopterus (p-value = 0.557) (Figura 5).
Figura 4 – Esfregaço sanguíneo de Pterygoplichthys pardalis corado com Giemsa evidenciando
células eritrocíticas normais (A) e danos genotóxicos (B, C, D, E, F). As setas evidenciam: Célula
micronucleada (B); Núcleo em forma de rim (C); Núcleo lobado (D); Núcleo segmentado (E);
Núcleo vacuolado (F). Barra: 10 μm.

117
Tabela 4 – Média e desvio padrão de microúcleos (MN) por indivíduo para cada espécie analisada e seu respectivo local de coleta na cidade
Manaus-AM.
Espécie Local de coleta Micronúcleo
(média e desvio
padrão)
Núcleos em
forma de rim
(média e desvio
padrão)
Núcleos lobados
(média e desvio
padrão)
Núcleos
segmentados
(média e desvio
padrão)
Núcleos
vacuolados
(média e desvio
padrão)
Hoplias malabaricus Igarapé do Tarumã-açu 1,33±1,53 7,33±4,51 1,0±1,0 0,67±0,58 0,67±1,15
Lago Catalão 1,91±1,04 0,5±1,95 0,82±0,75 0,36±0,67 0±0
Semaprochilodus insignis Igarapé Tarumã-açu 3,67±2,25 1,83±1,33 0,17±0,41 0,5±0,55 0±0
Lago Catalão 1,2±1,3 1,2±0,84 0,6±0,89 0±0 0,2±0,45
Pterygoplichthys pardalis
Igarapé do Mindu 7,0±6,24 0,67±0,58 0,33±0,58 0±0 0±0
Igarapé do Tarumã-açu 3,4±2,07 1,4±1,52 0,2±0,45 0±0 0±0
Lago Catalão 2,6±1,14 0,6±0,84 0,4±0,82 0,2±0,45 0,2±0,45
Oreochromis niloticus Igarapé do Mindu 5,3±4,4 1,5±1,43 0,3±0,48 0±0 0±0
Igarapé Tarumã-açu 4,93±3,0 2,3±1,94 0,44±1,12 0,48±1,12 0,19±0,48
Trichopodus trichopterus Igarapé do Mindu 8,64±7,57 7,86±4,82 2,14±2,03 0,21±0,58 11,86±9,64
Igarapé do Quarenta 5,3±3,33 5,3±3,89 0,6±0,7 0,8±1,03 1,5±2,32
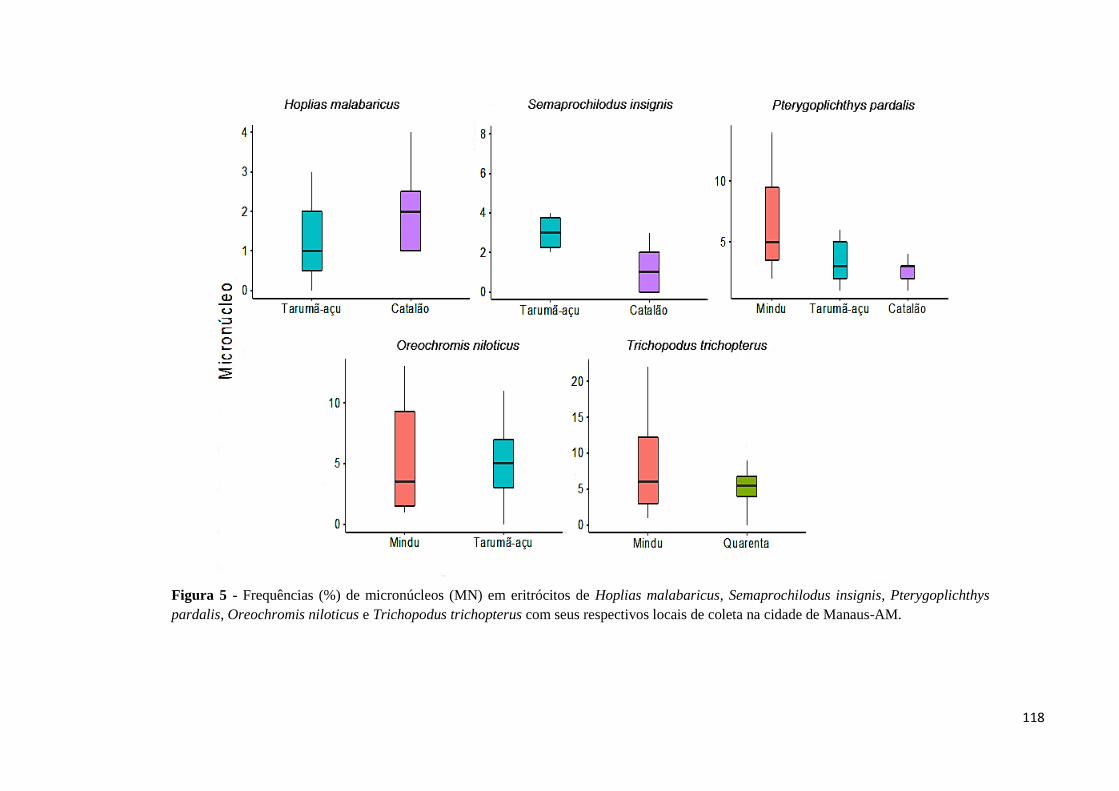
118
Figura 5 - Frequências (%) de micronúcleos (MN) em eritrócitos de Hoplias malabaricus, Semaprochilodus insignis, Pterygoplichthys
pardalis, Oreochromis niloticus e Trichopodus trichopterus com seus respectivos locais de coleta na cidade de Manaus-AM.
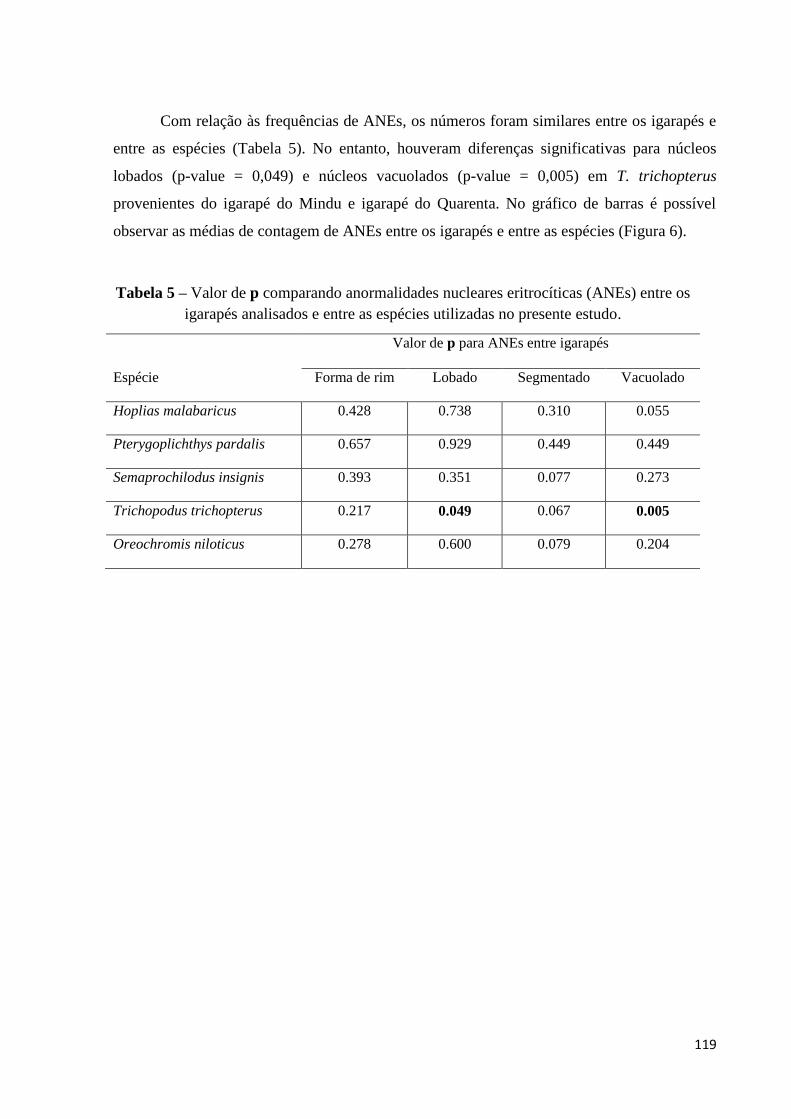
119
Com relação às frequências de ANEs, os números foram similares entre os igarapés e
entre as espécies (Tabela 5). No entanto, houveram diferenças significativas para núcleos
lobados (p-value = 0,049) e núcleos vacuolados (p-value = 0,005) em T. trichopterus
provenientes do igarapé do Mindu e igarapé do Quarenta. No gráfico de barras é possível
observar as médias de contagem de ANEs entre os igarapés e entre as espécies (Figura 6).
Tabela 5 – Valor de p comparando anormalidades nucleares eritrocíticas (ANEs) entre os
igarapés analisados e entre as espécies utilizadas no presente estudo.
Valor de p para ANEs entre igarapés
Espécie Forma de rim Lobado Segmentado Vacuolado
Hoplias malabaricus 0.428 0.738 0.310 0.055
Pterygoplichthys pardalis 0.657 0.929 0.449 0.449
Semaprochilodus insignis 0.393 0.351 0.077 0.273
Trichopodus trichopterus 0.217 0.049 0.067 0.005
Oreochromis niloticus 0.278 0.600 0.079 0.204

120
Figura 6 – Gráfico de barras mostrando as médias de contagem de tipos de anormalidades nucleares eritrocíticas (ANEs) entre os
igarapés e entre as espécies coletas no ambiente poluído e não poluído de Manaus-AM.

121
Na análise de cometas foram contabilizados e quantificados danos classificados em
quatro categorias para as espécies coletadas nos dois tipos de ambientes (poluído x não
poluído): 0 = sem danos (<5 %); 1 = baixo nível de danos (5-20 %); 2 = médio nível de danos
(20-40 %); 3 = alto nível de danos (40-95 %); 4 = totalmente danificados (>95 %) (Figura 7 e
Tabela 6).
Figura 7 – Níveis de cometas encontrado em Oreochromis niloticus (mesmo padrão
encontrado para Pterygoplichthys pardalis, Semaprochilodus insignis, Hoplias malabaricus e
Trichopodus trichopterus) evidenciando a classificação nas quatro categorias (cometa 1,
cometa 2, cometa 3 e cometa 4). Barra: 10 μm.

122
Tabela 6 – Média e desvio padrão da contagem dos níveis de cometa por indivíduo para cada espécie analisada e seu
respectivo local de coleta na cidade Manaus-AM.
Espécie Local de coleta Média da contagem dos níveis de cometa
0 1 2 3 4
Hoplias malabaricus Igarapé do Tarumã-açu 37,5±25,0 12,5±25,0 0±0 0±0 0±0
Lago Catalão 19,3±23,99 29,1±22,92 1,6±3,33 0±0 0±0
Pterygoplichthys pardalis
Igarapé do Tarumã-açu 14,8±13,99 19,0±13,45 12,8±9,12 2,8±5,72 0,6±1,34
Igarapé do Mindu 23,5±19,09 4,0±1,41 19,0±15,56 3,5±2,12 0±0
Lago Catalão 2,0±2,65 32,0±26,96 15,3±26,56 0±0 0,67±1,15
Semaprochilodus insignis Igarapé do Tarumã-açu 30,7±22,35 14,0±17,44 3,75±7,50 1,25±2,50 0,25±0,5
Lago Catalão 42,6±16,55 6,0±13,42 1,2±2,68 0,2±0,45 0±0
Trichopodus trichopterus Igarapé do Mindu 49,8±0,38 0,14±0,38 0±0 0±0 0±0
Igarapé do Quarenta 34,0±24,8 3,0±5,62 9,67±17,08 2,83±6,01 0,5±0,84
Oreochromis niloticus Igarapé do Mindu 30,0±24,98 19,6±24,42 0,33±0,58 0±0 0±0
Igarapé do Tarumã-açu 15,0±19,48 20,0±15,49 11,0±10,03 3,46±6,72 0,46±1,39

123
Houve diferença na frequência das respostas genotóxicas para a espécie O. niloticus,
coletada no igarapé do Tarumã-açu e igarapé do Mindu referente à categoria 2, que engloba o
nível médio de danos (20-40 %) (p-value = 0.035). Para as demais espécies o número de
cometas para cada categoria foi similar entre os igarapés e entre as espécies (Tabela 7 e
Figura 8).
Tabela 7 – Valor de p comparando o número de cometas entre os igarapés analisados e
entre as espécies utilizadas no presente estudo.
Espécie Valor de p (número de cometas entre igarapés)
0 1 2 3 4
Hoplias malabaricus 0.124 0.197 0.156 NA NA
Pterygoplichthys pardalis 0.090 0.513 0.620 0.125 0.732
Semaprochilodus insignis 0.382 0.382 0.737 0.737 0.263
Trichopodus trichopterus 0.334 0.334 0.111 0.111 0.111
Oreochromis niloticus 0.381 0.892 0.035 0.119 0.482
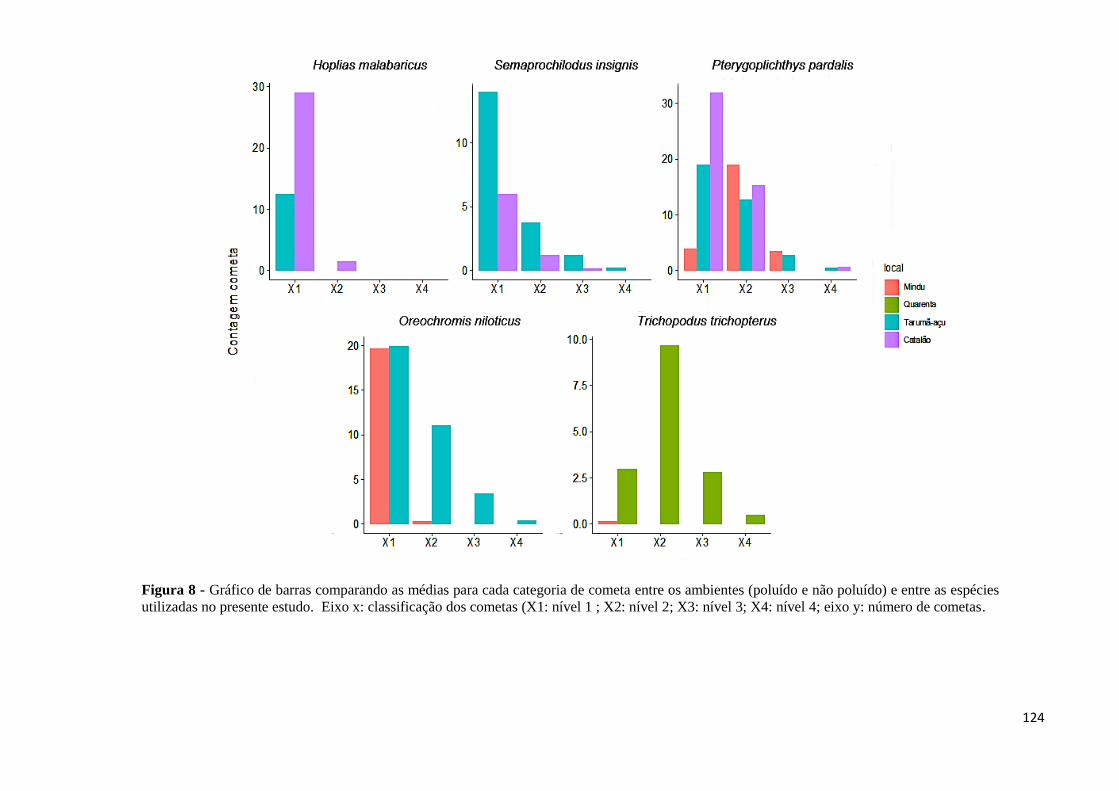
124
Figura 8 - Gráfico de barras comparando as médias para cada categoria de cometa entre os ambientes (poluído e não poluído) e entre as espécies
utilizadas no presente estudo. Eixo x: classificação dos cometas (X1: nível 1 ; X2: nível 2; X3: nível 3; X4: nível 4; eixo y: número de cometas.

125
4.5.5 Discussão
Embora já existam dados que permitem afirmar que a degradação dos igarapés de
Manaus é resultante do crescimento desordenado e do rápido aumento da cidade, é necessário
conhecer melhor como essas modificações ambientais, decorrentes dessa evolução urbana,
afetam as populações que os habitam (Santos-Silva e Silva 1993).
A caracterização destes locais, utilizando testes estatísticos comparativos, revelou
índices estimados de qualidade da água com baixa variação espacial e sazonal, quando
confrontados com as resoluções do CONAMA 410/2009 e 430/2011 (Tabela 1). Isso foi
observado tanto para o igarapé do Mindu (principal tributário da microbacia do São
Raimundo), que recebe contaminantes oriundos de esgotos e lixos domésticos, quanto para o
igarapé do Quarenta (um dos principais tributários da microbacia do Educandos), que passa
pelo Distrito Industrial da cidade, recebendo efluentes tanto industriais quanto domésticos
(Pinto et al. 2009; Thomas et al. 2014a).
A ausência de variação das variáveis avaliadas (pH, temperatura, OD, CE e turbidez)
com relação à sazonalidade e a desconformidade com os parâmetros estabelecidos pelo
CONAMA (2011), provavelmente se deve ao fato dos dos igarapés do Mindu e Quarenta,
apresentarem uma intensa alteração ambiental. Essas alteração são decorrentes de atividades
antrópicas, chegando a atingir total descaracterização destes locais, em virtude do pequeno
porte e pouca capacidade de autodepuração das cargas poluidoras (Nakamoto 2014; Viana
2018). A semelhança entre as variáveis físico-químicas avaliadas entre esses dois pode ser
observada na análise de ACP (Figura 1).
Por outro lado, a relação na análise de ACP do igarapé do Tarumã-açu e lago Catalão
(Figura 1) evidencia que os igarapés da bacia do Tarumã-açu ainda apresentam água com
características mais próximas das consideradas naturais para estes ecossistemas (Rodrigues et
al. 2009), embora já se observam processos de ocupação em áreas próximas a suas margens.
Alguns estudos, no entanto, apontam que o igarapé do Tarumã-açu que corta a cidade pela sua
zona Oeste e deságua no rio Negro vem sofrendo processo de contaminação diariamente tanto
por esgotos domésticos, como pelo fato de ter em sua bacia de drenagem a localização do
aterro controlado da rodovia AM-010 (Santana et al. 2007).
Porém, esta influência antrópica ainda não é muito acentuada pelo fato da densidade
populacional ser, por enquanto, relativamente pequena (IBGE, 2010). Isto explicaria o
agrupamento deste igarapé ao ambiente natural, pela análise de ACP e que particularmente no

126
igarapé do Tarumã-açu, os parâmetros físico-químicos analisados parecem sofrer influência
da sazonalidade, pois apesar de alguns valores observados na seca estarem fora dos valores
exigidos pelo CONAMA (2011), como pH e OD, esta situação tende a se regularizar no
período da cheia Durante esse período há maior incidência de chuva e aumento da vazão da
água, ocasionando maior turbulência da mesma elevando valores de pH e Od (Melo et al.,
2005; CETESB, 2007) (Tabela 1).
Além disso, o monitoramento físico e químico da água é pouco eficiente na detecção
de alterações na diversidade de hábitats e microhábitats e insuficiente na determinação das
consequências da alteração da qualidade da água sobre as comunidades biológicas. Em
contrapartida, a avaliação genotóxica em grupos biológicos representa um teste na avaliação
de como a exposição a xenobióticos pode afetar a saúde e o status de populações naturais e
invasoras (Zapata et al. 2016). Isto foi comprovado por meio da maior ocorrência de MN e
ANEs (média por indivíduos) em ambiente poluído (Tabela 4 e 5).
Nossos dados revelaram diferenças significativas de MN (p-value = 0.040) para S.
insignis entre o ambiente poluído e não poluído, resultado que indica perdas severas de
material genético (Xavier et al. 2011; Araldi et al. 2015). Porém, os dados observados para o
ensaio do cometa (Tabela 7) não revelaram diferenças entre os ambientes. Alguns estudos
descrevem que uma eficiente resposta observada em MN não é necessariamente refletida na
mesma intensidade no ensaio do cometa, uma vez que o MN e o cometa avaliam diferentes
aspectos do dano ao DNA. O MN avalia efeitos clastogênicos (provocam quebras no material
genético) e aneugênicos (que refletem danos às fibras do fuso mitótico), enquanto o ensaio do
cometa mensura quebras de fitas simples e duplas de DNA, locais instáveis alcalinos, DNA-
DNA e crosslinks DNA-proteína (Bombail et al. 2001; Klobucar, 2008; Thomas et al. 2014b).
Isto explicaria os dados observados para esta espécie quanto ao ensaio do cometa, já que é
possível que alguns dos danos tenham sido reparados.
As células são equipadas com sistemas complexos e sofisticados para o reparo do
DNA danificado, tolerância a diferentes danos, pontos de verificação do ciclo celular e vias de
morte celular, que funcionam coletivamente para reduzir as consequências deletérias. Com
isso, as células respondem, instilando vias robustas de resposta ao dano no DNA, que
permitem tempo suficiente para que as vias especializadas de reparo removam fisicamente o
dano. Pelo menos cinco principais vias de reparo de DNA como: reparo de excisão de base,
reparo de excisão de nucleotídeos, reparo de desemparelhamento, recombinação homóloga e
junção não homóloga, são ativas em diferentes estágios do ciclo celular, permitindo que as

127
células reparem os danos para manter a estabilidade genética nas células (Chatterjee e Walker
2017).
Para as demais espécies, os danos no DNA, permaneceram relativamente constantes,
indicando que as espécies P. pardalis, H. malabaricus, O. niloticus e T. trichopterus
desenvolveram sistemas sofisticados para identificar e reparar o DNA danificado (Ribeiro et
al. 2003). O mecanismo de reparo do DNA na célula corrige, por exemplo, quebras
bifilamentares por meio da junção de pontas quebradas, o que deve contribuir para a baixa
frequência de MN e ANEs em situações normais. Ou seja, MN e ANEs podem então ocorrer
mediante problemas que afetem o bom funcionamento do sistema de reparo do DNA (Ferreira
2013). Isto, explicaria os dados encontrados para as baixas frequências de MN e ANEs entre
os ambientes (Tabela 5 e Figura 5), com excessão de uma espécie exótica (T. trichopterus)
que apresentou diferenças significativas entre os dois ambientes poluídos (Mindu e Quarenta)
para núcleos lobados (p-value = 0,049) e núcleos vacuolados (p-value = 0,005), representando
um indício de que a contaminação pode estar influenciando diretamente no citoesqueleto e na
organização da cromatina no envelope nuclear.
As células eucarióticas possuem moléculas protéicas que se organizam em complexos
funcionais de diferentes níves, entre eles está o citoesqueleto, que possui milhares de
monômeros idênticos, polimerizados em filamentos lineares, divididos em queratinas tipo I e
II, vimentina, proteína glial fibrilar ácida, desmina e neurofilamentos (Oliveira 2000) e o
envelope nuclear, uma estrutura forma por uma dupla membrana com proteínas específicas
que se ligam a lâmina nuclear, que são responsáveis pela manutenção da integridade do
envoltório nuclear, ancoragem da cromatina durante a interfase e remontagem da membrana
nuclear após a mitose (Rolls et al. 1999).
Evidências tem demonstrado que a vimentina, uma proteína da família dos filamentos
intermediários do citoesqueleto, geralmente encontrada em células de origem mesenquimal,
tem tido sua expressão relacionada à morfologia nuclear e na localização da membrana
nuclear durante a mitose, onde a ausência destes filamentos representa invaginações no núcleo
(Sarria et al. 1994). Além disso, o envelope nuclear de células mitóticas passa por uma série
de mudanças estruturais que alteram sua morfologia em decorrência do crescimento dos
microtúbulos (Georgatos et al. 1997).
Contudo, essa associação de ANEs com possíveis alterações dos elementos que
constituem o citoesqueleto e do envelope nuclear, pode se tornar uma importante descoberta

128
de pontes cromatínicas entre o núcleo e estruturas associadas (Oliveira 2000), bem como, o
estudo de proteínas associadas ao citoesqueleto e do envelope nuclear podem esclarecer a
associação de suas estruturas com as alterações nucleares. Porém, como o mecanismo de
formação de ANEs não é bem conhecido, fica uma dúvida quanto a serem estruturas distintas,
o que poderia representar etapas diferentes de um mesmo processo relacionado a
genotoxicidade.
Neste estudo, embora tenham sido observadas diferenças baixas e não significativas de
MN e ANEs em O. niloticus, uma espécie exótica coletada em dois locais poluídos, o ensaio
do cometa foi utilizado com sucesso, mostrando diferenças significativas nesta espécie,
referente à categoria 2: médio nível de danos (20-40 %) (p-value = 0.035). Interessante pensar
para as duas espécies exóticas, coletadas neste estudo, como ótimos indicadores sensíveis de
exposição, uma vez que, diferenças significativas foram observadas utilizando os testes de
genotoxicidade entre ambientes considerados poluídos, o que poderia relacionar estas
diferenças com a localidade onde vivem, ou seja, com o tipo de poluição que estes igarapés
urbanos apresentam em suas águas.
Em relação à uma análise qualitativa para o ensaio do cometa é possível observar
maior dano ao DNA nos ambientes poluídos, quando comparados com o ambiente não
poluído (Figura 8). No entanto, a categoria 1 = baixo nível de danos (5-20 %) e a categoria 2
= médio nível de danos (20-40 %) foram maiores no ambiente natural para as espécies H.
malabaricus e P. pardalis. Isto pode ser explicado provavelmente pelas características da
água, que se apresentam pouco oxigenadas, com pH ácido (Tabela 1) e altos valores de nitrato
e amônia (Brito et al. 2014), indicando que estes fatores podem ser a causa de privações de
nutrientes, estresse oxidativo e danos no DNA, que não podem ser corrigidos pelo sistema de
reparo da célula (Oliveira 2015).
Sendo assim, nossos resultados demonstram a utilidade de MN, ANEs e cometa como
biomarcadores adicionais para avaliar a saúde do ecossistema aquático, bem como a
importância de se avaliar diferentes espécies de peixes. As espécies exóticas parecem exibir
maior prevalência em resposta à genotoxicidade, como indicado por ANEs e cometa, contudo,
isso não significa que não esteja havendo o remodelamento genético e epigenético nas outras
espécies, provenientes de igarapés urbanos da cidade de Manaus-AM, as quais vêm sofrendo
fortes mudanças e impactos em virtude da ação humana. Isto porque, as sequências
ribossomais são altamente complexas no genoma de peixes de ambiente poluído, com um
perfil de metilação diferencial no gene 18S entre os ambientes, bem como a adição de

129
heterocromatina e regiões de heterocromatinização, ladeadas por elementos transponíveis,
além do aumento na quantidade de sequências de elementos transponíveis nos cromossomos
que estão influenciando na modelagem do genoma das populações que vivem em locais
contaminados e que a resposta à mudança do ambiente é particular de cada espécie. Neste
contexto, é preciso investir urgentemente em ações de conscientização, que devem ter foco
especial nas populações que vivem as margens dos igarapés e que já banalizaram a prática de
descartar nas águas o lixo que produzem. Por outro lado, cabe ao poder público oferecer
alternativas adequadas de descarte a essas famílias. No entanto, mudar a triste realidade atual
não é tarefa fácil, é missão para muitos anos com aplicação de um plano inteligente.

130
5 Conclusões
Danos no DNA permaneceram relativamente constantes, indicando que algumas
espécies desenvolveram sistemas sofisticados para identificar e reparar o DNA
danificado, impedindo assim a ocorrência exacerbada de mutações. Além disso, a
contaminação pode estar influenciando diretamente no citoesqueleto e na
organização da cromatina do envelope nuclear.
Sequências ribossomais são altamente complexas no genoma de peixes de
ambientes poluídos, além de estarem envolvidas na manutenção da integridade do
genoma, mediada por um aumento no número de cópias do DNA ribossomal em
resposta a mudanças nas condições ambientais.
O aumento na quantidade dos elementos transponíveis nos cromossomos de peixes,
acarreta em uma modelagem do genoma em populações que vivem em ambientes
poluídos, cujos efeitos, devem estar sob constante evolução adaptativa das espécies
de peixes que vivem nestes ambientes.
A estrutura, organização e função do genoma apresentam-se modificadas em
indivíduos expostos à poluição ambiental, provavelmente em decorrência a
interações com os elementos transponíveis.
A contaminação pode influenciar na metilação do DNA, gerando perfis de
metilação diferencial como encontrado nos sítios de DNAr 18S, que teriam
evoluído nas espécies, inicialmente, como um mecanismo de defesa contra a
expressão de elementos transponíveis.

131
6 Referências
Abreu, R.C.R.; Durigan, G. 2011. Changes in the plant community of a Brazilian grassland
savannah after 22 years of invasion by Pinus elliottii Engelm. Plant Ecology & Diversity, 4:
269-278.
Affonso, P.R.A.; Galetti Jr., P.M. 2005. Chromosomal diversification of reef fishes from
genus Centropyge (Perciformes, Pomacanthidae). Genetica, 123: 227-233.
Almeida-Val, V.M.F.; Farias, I. P. 1996. Metabolic adjustments to chronic hypoxia. In: Val,
A. L.; Almeida-Val, V. M. F.; Randall, D. J. (Org.). Physiology and Biochemistry of the
Fishes of the Amazon. Instituto Nacional de Pesquisas da Amazônia, Manaus, Amazonas. p.
257-271.
Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. 1990. Basic local alignment
search tool. Journal of Molecular Biology, 215: 403-410.
Alzohairy, A.M.; Gyulai, G; Jansen, R.K.; Bahieldin, A. 2013. Transposable elements
domesticated and neofunctionalized by eukaryotic genomes. Plasmid, 69: 1-15.
Amorin, K.D.J.; Cioffi, M.B.; Bertollo, L.A.C.; Soares, R.X.; Souza, A.S.; Costa, G.W.W.F.;
Molina, W.F. 2016. Co-located 18S/5S rDNA arrays: an ancient and unusual chromosomal
trait in Julidini species (Labridae, Perciformes). Comparative Cytogenetics, 10(4): 555-570.
Araldi, R.P.; Melo, T.C.; Mendes, T.B.; Sá, P.L.; Nozima, B.H.N.; Ito, E.T.; Carvalho, R.F.;
Souza, E.B.; Stocco, R.C.;. 2015. Using the comet and micronucleus assays for genotoxicity
studies: A Review. Biomedicine & Pharmacotherapy 72: 74-82.
Arias, A.R.L.; Inacio, A.F.; Novo, L.A.; Albuquerque, C.; Moreira, J.C. 2008.
Multibiomarker approach in fish to assess the impact of pollution in a large Brazilian River,
Paraiba do Sul. Environ Pollut, 156(3): 974-979.
Arif, I.; Khan, H. 2009. Molecular markers for biodiversity analysis of wildlife animals: a
brief review. Animal Biodiv Conser, 32: 9-17.
Averbeck, K.T.; Eickbush, T.H. 2005. Monitoring the mode and tempo of concerted evolution
in the Drosophila melanogaster rDNA locus. Genetics, 171(4): 1837-1846.
Bacalini, M.G.; Pacilli, A.; Giuliani, C.; Penzo, M.; Treré, D.; Pirazzini, C.; Salvioli, S.;
Franceschi, C.; Montanaro, L.; Garagnani, P. 2014. The nucleolar size is associated to the
methylation status of ribosomal DNA in breast carcinomas. BMC Cancer 14: 361.
Barbosa, J.S.; Cabral, T.M.; Ferreira, D.N.; Agnez-Lima, L.F.; Batistuzzo, M.S.R. 2010.
Genotoxicity assessment in aquatic environment impacted by the presence of heavy metals.
Ecotoxicology and Environmental Safety, 73: 320-325.
Barbosa, C.M; Mareco, E.M.; Silva, M.D.P.; Martins, C.; Alves-Costa, F.A. 2014.
Differential expression of a retrotransposable element, Rex6, in Colossoma macropomum fish
from different Amazonian environments, Mobile Genetic Elements, 4(4): e30003.

132
Barroca, T.M. 2012. Análise da variabilidade genética de Prochilodus spp. (Prochilodontidae)
das bacias dos rios Paraopeba, Pará e Grande, utilizando marcador de complexas repetições
hipervariáveis. Tese de Doutorado, Pós-Graduação em Genética. Instituto de Ciências
Biológicas da Universidade Federal de Minas Gerais, p. 150.
Barros, A.V.; Wolski, M.A.V.; Nogaroto, V.; Almeida, M.C.; Moreira-Filho, O.; Vicari, M.R.
2017. Fragile sites, dysfunctional telomere and chromosome fusions: What is 5S rDNA role?
Gene, 608: 20-27.
Baršiene, J.; Rybakovas, A.; Lang, T.; Andreike, ˙N.L.; Michailovas, A. 2013. Environmental
genotoxicity and cytotoxicity levels in fish from the North Sea offshore region and Atlantic
coastal waters. Mar. Poll. Bull., 68(1-2): 106-116.
Bentes, N. 2005. Manaus: realidade e constrastes sociais. Editora Valer, Manaus, Amazonas.
Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. 1978. Cytotaxonomic considerations on
Hoplias lacerdae (Pisces, Erytrinidae). Revista Brasileira de Genética, 1: 103-120.
Boeke, J.D.; Devine, S.E. 1998. Yeast retrotransposons: finding a nice quiet neighborhood.
Cell, 93: 1087-1089.
Bombail, V.A.D.; Gordon, E.; Batty, J. 2001. Application of the comet and micronucleus
assays to butterfish (Pholis gunnellus) erythrocytes from the Firth of Forth. Chemosphere, 44:
383-392.
Bombassaro, J.U. 2007. Análise genotóxica do complexo fosfatase ácida púrpura símile Fe
Cu OHH2BPBPMP em leucócitos totais e sangue total humano. Trabalho de conclusão de
curso - Curso de Medicina da Universidade do Extremo Sul Catarinense, UNESC. Criciúma,
p. 51.
Braham, R.P.; Blazer, V.S.; Shaw, C.H.; Mazik, P.M. 2017. Micronuclei and other
erythrocyte nuclear abnormalities in fishes from the Great Lakes Basin, USA. Environmental
and Molecular Mutagenesis, 58: 570-581.
Brander, S.M.; Biales, A.D.; Connonc, R.E. 2017. The Role of Epigenomics in Aquatic
Toxicology. Environmental Toxicology and Chemistry, 36(10): 2565- 2573.
Brasil. Resolução N° 357, de 17 de março de 2005. Dispõe sobre a classificação dos corpos
de água e diretrizes ambientais para seu enquadramento, bem como estabelece as condições e
padrões de lançamento de efluentes, e dá outras providências. Brasília, DF, 2005.
Brawand, D.; Wagner, C.E.; Li, Y.I.; Malinsky, M.; Keller, I.; Fan, S.; Simakov, O.; Ng,
A.Y.; Lim, Z.W.; Bezault, E. 2014. The genomic substrate for adaptive radiation in African
cichlid fish. Nature, 513: 375-381.
Braz-Mota, S.; Campos, D.F.; Maccormack, T.J.; Duarte, R.M.; Val, A.L.; Almeida-Val,
V.M.F. 2018. Mechanisms of toxic action of copper and copper nanoparticles in two amazon
fish species: dwarf cichlid (Apistogramma agassizii) and cardinal tetra (Paracheirodon
axelrodi). Sci. Total Environ, 630: 1168-1180.

133
Brianezi, G. 2011. Desenvolvimento de um método de análise digital para o teste do cometa
corado pela prata. Dissertação de Mestrado. Pós-Graduação em Patologia. Botucatu, São
Paulo. p. 101.
Bridier-Nahmias, A.; Tchalikian-Cosson, A.; Baller, J.A.; Menouni, R.; Fayol, H.; Flores, A.;
Saïb, A.; Werner, M.; Voytas, D.F.; Lesage, P. 2015. Retrotransposons. An RNA polymerase
III subunit determines sites of retrotransposon integration. Science, 348: 585-588.
Brito, J.G.; Alves, L.F.; Espirito Santo, H.M.V. 2014. Seasonal and spatial variations in
limnological conditions of a floodplain lake (Lake Catalão) connected to both the Solimões
and Negro Rivers, Central Amazonia. Acta Amazonica, 44(1): 121-134.
Britton-Davidian, J.; Cazaux, B.; Catalan, J. 2012. Chromosomal dynamics of nucleolar
organizer regions (NORs) in the house mouse: microevolutionary insights. Heredity, 108(1):
68-74.
Bucker, A.; Carvalho, W.; Alves-Gomes, J.A. 2006. Avaliação da mutagênese e
genotoxicidade em Eigenmannia virescens (Teleostei: Gymnotiformes) expostos ao benzeno.
Acta Amazonica, 36(3): 357-364.
Bühler, M. 2009. RNA turnover and chromatin-dependent gene silencing. Chromosoma, 118:
141-151.
Busslinger, M.; de Boer, E.; Wright, S.; Grosveld, F.G.; Flavell, R.A. 1983. The sequence
GGCmCGG is resistant to MspI cleavage. Nucleic Acids Res., 11: 3559-3569.
Cabral-de-Mello, D.C.; Oliveira, S.G.; de Moura, R.C.; Martins, C. 2011. Chromosomal
organization of the 18S and 5S rRNAs and histone H3 genes in Scarabaeinae coleopterans:
insights into the evolutionary dynamics of multigene families and heterochromatin. BMC
genetics, 12(1): 88.
Caixeta, E.T.; Oliveira, A.C.B.; Brito, G.G.; Sakiyama, N.S. 2006. Tipos de Marcadores
Moleculares. In: Borem, A.; Caixeta, E.T. (ed) Marcadores moleculares. Viçosa. p. 130.
Carrasco, K.R.; Tilbury, K.L.; Myers, M.S. 1990. Assessment of the piscine micronucleus test
as an in situ biological indicator of chemical contaminant effects. Canadian Journal of
Fisheries and Aquatic Sciences, 47: 2123-2136.
Carroll, S.P.; Dingle, H.; Famula, T.R.; Fox, C.W. 2001. Genetic architecture of adaptive
differentiation in evolving host races of the soapberry bug, Jadera haematoloma. Genetica,
112: 257-272.
Carvalho, N.D.M.; Pinheiro, V.S.S.; Carmo, E.J.; Goll, L.G.; Schneider, C.H.; Gross, M.C.
2016. The Organization of Repetitive DNA in the Genomes of Amazonian Lizard Species in
the Family Teiidae. Cytogenetic and Genome Research, 147: 161-168.
Casacuberta, E.; Gonzalez, J. 2013. The impact of transposable elements in environmental
adaptation. Molecular Ecology, 22: 1503-1517.

134
Catarino, M.F.; Zuanon, J. 2010. Feeding ecology of the leaf fish Monocirrhus polyacanthus
(Perciformes: Polycentridae) in a terra firme stream in the Brazilian Amazon. Neotropical
Ichthyology, 8 (1): 183-186.
Cecco, M.D.; Criscione, S.W.; Peckham, E.J.; Hillenmeyer, S.; Hamm, E.A.; Manivannan, J.
et al. 2013. Genomes of replicatively senescent cells undergo global epigenetic changes
leading to gene silencing and activation of transposable elements. Aging Cell, 12: 247-256.
Censo Demográfico. 2010. Características gerais da população, religião e pessoas com
deficiência. Rio de Janeiro: IBGE. Acesso em: junho. 2019.
Chadha, S.; Sharma, M. 2014. Transposable Elements as stress adaptive capacitors induce
genomic instability in Fungal Pathogen Magnaporthe oryzae. PLoS One, 9: e94415.
Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J-N. 2015. Comparative Analysis of
transposable elements Highlights mobilome diversity and evolution in vertebrates. Genome
Biol. Evol. 7(2):567-580.
Chatterjee, N.; Walker, G.C. 2017. Mechanisms of DNA Damage, Repair, and Mutagenesis.
Environmental and Molecular Mutagenesis 00:00-00.
Chen, X.; Chen, K.; Yang, H.; Qu, Q.; Feng, Y.; Bai, J. 2015. Complete mitochondrial
genome of the Trichogaster leeri (Anabantoidei: Osphronemidae). Mitochondrial DNA, 1736:
1-2.
Chevin, L.M.; Lande, R.; Mace, G.M. 2010. Adaptation, plasticity, and extinction in a
changing environment: towards a predictive theory. PLoS Biology, 8: e1000357.
Chew, J.K.; Oliveira, C.; Wright, J.M.; Dobson, M.J. 2002. Molecular and cytogenetic
analysis of the telomeric (TTAGGG)n repetitive sequence in the Nile tilapia, Oreochromis
niloticus (Teleostei: Cichlidae). Chromosoma, 111: 45-52.
Coan, R.L.B.; Martins, C. 2018. Landscape of Transposable Elements focusing on the B
chromosome of the Cichlid Fish Astatotilapia latifasciata. Genes, 9: 269.
Cole, B.; Tamaru, C.S.; Bailey, R. 1999. A manual for commercial production of the gourami,
Trichogaster trichopterus, a temporary paired spawner. Honolulu: Center for Tropical and
Subtropical Aquaculture Publication, 135: 28.
Collins, A.R. 2014. Measuring oxidative damage to DNA and its repair with the comet assay.
Biochim. Biophys. Acta, 1840(2):794-800.
Companhia de Tecnologia de Saneamento Ambiental (CETESB). 2007. Qualidade das águas
interiores no estado de São Paulo. Secretaria do meio ambiente. Companhia Ambiental do
Estado de São Paulo. Relatórios 2008.
Conselho Nacional de Controle de Experimentação Animal. 2013. Ministério da Ciência,
Tecnologia, Inovações e Comunicações. Acesso em:
https://www.mctic.gov.br/mctic/opencms/institucional/concea.
Costa, F.O., Henzler, C.M.; Lunt, D.H.; Whiteley, N.M.; Rock, J. 2016. Probing marine
Gammarus (Amphipoda) taxonomy with DNA barcodes. Syst. Biodivers, 7(4):365-379.

135
Couceiro, S.R.M.; Hamada, N.; Luz, S.L.B.; Forsberg, B.R.; Pimentel, T.P. 2007.
Deforestation and sewage effects on aquatic macroinvertebrates in urban streams in Manaus,
Amazonas, Brazil. Hydrobiologia, 575: 271-284.
Cowley, M.; Oakey, R.J. 2013. Transposable elements rewire and finetune the transcriptome.
PLoS Genet, 9(1): 1003234.
Dagosta, F.C.P.; Pinna, M.C.C. 2019. The fishes of the Amazon: distribution and
biogeographical patterns, with a comprehensive list of species. Bulletin of the American
Museum of Natural History, no. 431.
Da Silva, M.; Barbosa, P.; Artoni, R.F.; Feldberg, E. 2016. Evolutionary Dynamics of 5S
rDNA and recurrent association of Transposable Elements in electric fish of the Family
Gymnotidae (Gymnotiformes): The Case of Gymnotus mamiraua. Cytogenetic and Genome
Research, 149(4): 297-303.
Dai, J.; Xie, W.; Brady, T.L.; Gao, J.; Voytas, D.F. 2007. Phosphorylation regulates
integration of the yeast Ty5 retrotransposon into heterochromatin. Mol. Cell, 27: 289–299.
Dancey, C.P.; Reidy, J. 2006. Estatística sem Matemática para Psicologia: usando SPSS para
Windows. [Tradução VIALI, L.]. 3a ed. Porto Alegre: Artmed. p. 608.
De Carvalho, D.C.; Oliveira, D.A.A.; Pompeu, P.S.; Leal, C.G.; Oliveira, C.; Hanner, R.
2011. Deep barcode divergence in Brazilian freshwater fishes: the case of the São Francisco
River basin. Mitochondrial DNA, 22: 80-86.
De la Veja, E.; Degnan, B.M.; Hall, M.R.; Wilson, K.J. 2007. Differential expression of
immune-related genes and transposable elements in black tiger shrimp (Penaeus monodon)
exposed to a range of environmental stressors. Fish & Shellfish Immunology, 23:1072-1088.
Dhawan, A.; Bajpayee, M.; Parmar, D. 2009. Comet assay: a reliable tool for the assessment
of DNA damage in different models. Cell Biol Toxicol.,25(1):5-32.
Do Valle, A. 1998. Tarumã tem equilíbrio ambiental ameaçado. A Crítica, 12/06/98, A7,
Manaus/AM.
Farré, M.; Narayan, J.; Slavov, G.T.; Damas, J.; Auvil, L.; Li, C.; Larkin, D.M. 2016. Novel
insights into chromosome evolution in birds, archosaurs, and reptiles. Genome Biology and
Evolution, 8: 2442-2451.
Fenech, M.; Cheng, W.P.; Kirsch-Volders, M.; Holland, N.; Bonassi, S.; Zeiger, E. 2003.
Humn project: detailed description of the scoring criteria for the cytokinesis-block
micronucleus assay using isolated human lymphocyte cultures. Mutation Research, 534: 65-
75.
Fent, K. 2004. Ecotoxicological effects at contaminated sites. Toxicology, 205: 223-240.
Ferreira, R.B.; Dantas, R.B.; Tonini, J.F.R. 2012. Distribuição espacial e sazonal de anfíbios
em quatro poças na região serrana do Espírito Santo, sudeste do Brasil: influência de
corredores florestais. Iheringia, Série Zoologia, Porto Alegre, 102(2): 163-169.

136
Ferreira, M.R.S. 2013. Perfil genotóxico e bioacumulação de metais em Hoplias malabaricus
(Bloch, 1794) de tributários dos rios Negro e Solimões. Dissertação de Mestrado. Programa
de Pós-Graduação em Genética, Conservação e Biologia Evolutiva do Instituto Nacional de
Pesquisas da Amazônia, p. 103.
Filgueiras, A.V.; Lavilla, I.; Bendicho, C. 2004. Evaluation of distribution, mobility and
binding behaviour of heavy metals in surficial sediments of Louro River (Galicia, Spain)
using chemometric analysis: a case study. Science of the Total Environment, 330: 115-129.
Fraga, M.F.; Esteller, M. 2002. DNA Methylation: A Profile of Methods and Applications.
BioTechniques, 33: 632-649.
Frederico, R.G.; Olden, J.D.; Zuanon, J. 2016. Climate change sensitivity of threatened, and
largely unprotected, Amazonian fishes. Aquatic Conservation: Marine and Freshwater
Ecosystems, 26: 91-102.
Freitas, V.S.; Lopes, M.A.; Meireles, J.R.C.; Reis, L.C.; Eneida, M.M. 2005. Efeitos
genotóxicos de fatores considerados de risco para o câncer bucal. Revista Baiana de Saúde
Pública, 29(2): 189-199.
Freitas, C.E.C.; Siqueira-Souza, F.K. 2009. O uso de peixes como bioindicador ambiental em
áreas de várzea da bacia amazônica. Revista Agrogeoambiental, (2009): 1-7.
Fulneček, J.; Kovařík, A. 2014. How to interpret Methylation Sensitive Amplified
Polymorphism (MSAP) profiles? BMC Genetics, 15: 2.
Frenzilli, G.; Nigro, M.; Lyons, BP. 2009. The Comet assay for the evaluation of genotoxic
impact in aquatic environments. Mutation Research, 681(1):80-92.
Galetti, Jr., P.M.; Martins, C. 2004. Contribuição da hibridização in situ para conhecimento
dos cromossomos de peixes. In: Guerra, M. (Org) FISH: Conceitos e aplicações na
citogenética. Sociedade Brasileira de Genética, Ribeirão Preto, Brasil. p. 61-88.
Gardener, M.R.; Bustamante, R.O.; Herrera, I.; Durigan, G.; Pivello, V.R.; Moro, M.F.; Stoll,
A.; Langdon, B.; Baruch, Z.; Rico, A.; Arredondo-Nuñez, A.; Flores, S. 2012. Plant invasions
research in Latin America: fast track to a more focused agenda. Plant Ecology & Diversity, 1:
1-8.
Georgatos, S.D.; Pyrpasopoulou, A.; Theooropoulos, P.A. 1997. Nuclear envelope breakdown
in mammalian cells involves stepwise lamina disassembly and microtubule-drive deformation
of the nuclear membrane. Journal of Cell Science, 110:2129-2140.
Gontijo, A.M.M.C.; Tice, R. 2003. Teste do cometa para a detecção de dano no DNA e reparo
em células individualizadas. In: Ribeiro, L. R.; Salvadori, D. M. F.; Marques, E. K. (Org.).
Mutagênese Ambiental. Canoas: Ulbra. p. 173-200.
Gornung, E. 2013. Twenty years of physical mapping of major ribosomal RNA genes across
the teleosts: a review of research. Cytogenetic and Genome Research, 141: 90-102.
Goulart, M.; Callisto, M. 2003. Bioindicadores de qualidade de água como ferramenta em
estudos de impacto ambiental. Revista da FAPAM, ano 2, no 1.

137
Gouveia, N. 2012. Resíduos sólidos urbanos: impactos socioambientais e perspectivas do
manejo sustentável com inclusão social. Cad Saude Colet, 17(6): 1503-1510.
Grandi, F.C.; Rosser, J.M.; Newkirk, S.J.; Yin, J.; Jiang, X.; Xing, Z.; Ye, P. 2015.
Retrotransposition creates sloping shores: A graded influence of hypomethylated CpG islands
on flanking CpG sites. Genome Research, 25: 1135-1146.
Green, B.; Bouchier, C.; Fairhead, C.; Craig, N.L.; Cormack, B.P. 2012. Insertion site
preference of Mu, Tn5, and Tn7 transposons. Mobile DNA, 3: 3.
Grewal, S.I.S.; Jia, S. 2007. Heterochromatin revisited. Nature Reviews Genetics, 8: 35-46.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Martins, C.; Feldberg, E. 2010a. Variability of
18S rDNA locus among Symphysodon fishes: chromosomal rearrangements. Journal of Fish
Biology, 76: 1117-1127.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Porto, J.I.R.; Martins, C., Feldberg, E. 2010b.
Comparative cytogenetic analysis of the genus Symphysodon (Discus Fishes, Cichlidae):
Chromosomal characteristics of retrotransposons and minor ribosomal DNA. Cytogenetic and
Genome Research, 127(1): 43-53.
Gross, M.C.; Schneider, C.H.; Terencio, M.L.; Ribeiro, L.B.; Moraes, B.N.G.; Azevedo,
M.B.; Silva, F.A.; Feldberg, E. 2012. Comparative analyses of the Rex 3 repetitive elements
in fish species from different Amazonian environments. In: Workshop Adaptations of Aquatic
Biota, Manaus. Short Notes - Adaptations of Aquatic Biota. Manaus, AM.
Grummt, I. 2013. The nucleolus—Guardian of cellular homeostasis and genome integrity.
Chromosoma, 122: 487-497.
Guimarães, E.M.C.; Carvalho, N.D.M.; Schneider, C.H.; Feldberg, E.; Gross, M.C. 2017.
Karyotypic Comparison of Hoplias malabaricus (Bloch, 1794) (Characiformes, Erythrinidae)
in Central Amazon. Zebrafish, 14(1): 80-89.
Guo, Y.; Levin, H.L. 2010. High-throughput sequencing of retrotransposon integration
provides a saturated profile of target activity in Schizosaccharomyces pombe. Genome Res.,
20: 239-248.
Hancks, D.C.; Kazazian, H.H. 2012. Active human retrotransposons: Variation and disease.
Current Opinion in Genetics & Development, 22: 191-203.
Hartmann, A.; Herkommer, K.; Gluck, M.; Speit, G. 1995. DNA-damaging effect of
cyclophosphamide on human blood cells in vivo and in vitro studied with the single-cell gel
test (comet assay). Environ Mol Mutagen., 25(3):180-187.
Hayash, M. 2016. The micronucleus test—most widely used in vivo genotoxicity test. Genes
and Environment, 38: 18.
Head, J.A. 2014. Patterns of DNA Methylation in Animals: An Ecotoxicological Perspective.
Integrative and Comparative Biology, 54(1):77-86.

138
Head, J.A.; Mittal, K.; Basu, N. 2014. Application of the LUminometric Methylation Assay
(LUMA) to ecological species: tissue quality requirements and a survey of DNA methylation
levels in animals. Mol Ecol Resour., 14(5): 943-52.
Heinrichs-caldas, W.; Campos, D.F.; Paula-Silva, M.N.; Almeida-Val, V.M.F. 2019. Oxygen-
dependent distinct expression of hif-1α gene in aerobic and anaerobic tissues of the amazon
oscar, Astronotus crassipinnis. In: Comparative Biochemistry and Physiology B-Biochemistry
& Molecular Biology, 227:31-38.
Hoffmann, A.A.; Sgro, C.M. 2011. Climate change and evolutionary adaptation. Nature, 470:
479-485.
Howell, W.M.; Black, D.A. 1980. Controlled silver staining of nucleolus organizer regions
with a protective colloidal developer: a 1-step method. Experientia, 3: 1014-1015.
Hua-Van, A.; Le Rouzic, A.; Boutin, T.S.; Filee, J.; Capy, P. 2011. The struggle for life of the
genome’s selfish architects. Biol Direct, 6: 19.
Hussain, B.; Sultana, T.; Sultana, S.; Masoud, M.S.; Ahmed, Z.; Mahboob, S. 2018. Fish eco-
genotoxicology: Comet and micronucleus assay in fish erythrocytes as in situ biomarker of
freshwater pollution. Saudi Journal of Biological Sciences, 25: 393-398.
Ito, H.; Gaubert, H.; Bucher, E.; Mirouze, M.; Vaillant, I.; Paszkowski, J. 2011. A siRNA
pathway prevents transgenerational retrotransposition in plants subjected to stress. Nature,
472: 115-119.
Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. 2007. Universal primer cocktails
for fish DNA barcoding. Molecular Ecology Notes, 7: 544-548.
Jacobina, U.P.; Vicari, M.R.; Martinez, P.A.; Cioffi, M.B.; Bertollo, L.A.C.; Molina W.F.
2013. Atlantic moonfishes: independent pathways of karyotypic and morphological
differentiation. Helgoland Marine Research, 67(3): 499-506.
Jurka, J.; Kapitonov, V.; Kohany, O.; Jurka, M.V. 2007. Repetitive sequences in complex
genomes: structure and evolution. Annual Review of Genomics and Human Genetics, 8: 241-
259.
Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S. 2012. Geneious
Basic: an integrated and extendable desktop software platform for the organization and
analysis of sequence data. Bioinformatics, 28: 1647-1649.
Kirschbaum, A.A., Seriani, R., Ranzani-Paiva, M.J.T., Abessa, D.M.S., Pereira, C.D.S., 2009.
Cytogenotoxicity biomarkers in fat snook Centropomus parallelus from Cananéiaand São
Vicente estuaries, SP. Brazil. Gen. Mol. Biol. 32 (1), 151-154.
Klobucar, G.; Pavlica, M.; Erben, R.; Papes, D. 2008. Application of the micronucleus and
comet assays to mussel Dreissena polymorpha haemocytes for genotoxicity monitoring of
freshwater environments. Aquatic Toxicology, 64: 15-23.

139
Kobayashi, T. 2006. Strategies to maintain the stability of the ribosomal RNA gene repeats –
Collaboration of recombination, cohesion, and condensation. Genes & Genetic Systems, 81:
155-161.
Kobayashi, T. 2008. A new role of the rDNA and nucleolus in the nucleus – rDNA instability
maintains genome integrity. Bioessays, 30: 267-272.
Kobayashi, T. 2016. Genome Instability of Repetitive Sequence: Lesson from the Ribosomal
RNA Gene Repeat. In: DNA Replication, Recombination and Repair. Hanaoka, F., Sugasawa,
K., Eds.; Springer: Tokyo, Japan, p. 235-247.
Kolarević, S.; Aborgiba, M.; Kračun-Kolarević, M.; Kostić, J.; Simonović, P.; Simić, V;
Milošković, A.; Reischer, G. et al. 2017. Evaluation of Genotoxic Pressure along the Sava
River. PLoS ONE 11(9): e0162450.
Kroon, F.; Streten, C.; Harries, S. 2017. A protocol for identifying suitable biomarkers to
assess fish health: A systematic review. PLoS One, 12(4): e0174762.
Kucharski, R.; Maleszka, J.; Foret, S.; Maleszka, R. 2008. Nutritional control of reproductive
status in honeybees via DNA methylation. Science, 319(5871): 1827-1830.
Kuck, J. 2013. CAU/AM. CONSELHO DE ARQUITETURA E URBANISMO DA
AMAZÔNIA. Disponivel em: <http://www.cauam.org.br/wp-
content/uploads/2013/07/CARTA_CAU_AM2.pdf>. Acesso em: 23 NOVEMBRO 2016.
Kuhn, G.C.S.; Kuttler, H.; Moreira-Filho, O.; Heslop-Harrison, J.S. 2012. The 1.688
repetitive DNA of Drosophila: concerted evolution at different genomic scales and
association with genes. Mol Biol Evol, 29: 7-11.
Lanciano, S.; Mirouze, M. 2018. Transposable elements: all mobile, all different, some stress
responsive, some adaptive? Current Opinion in Genetics & Development, 49:106-114.
Lande, R. 1988. Genetics and demography in biological conservation. Science, 241: 1455-
1460.
Lapuente, J.; Lourenço, J.; Mendo, S.A.; Borràs, M; Martins, M.G.; Costa, P.M.; Pacheco, M.
2015. The Comet Assay and its applications in the field of ecotoxicology: a mature tool that
continues to expand its perspectives. Front. Genet., 6: 180.
Lee, C.E. 2002. Evolutionary genetics of invasive species Review. Trends in Ecology &
Evolution, 17: 9-11.
Lee, R.F.; Steinert, S. 2003. Use of the single Cell gel electrophoresis/comet assay for
detecting DNA damage in aquatic (marine and freshwater) animals. Mutation
Research/Review in Mutation Research, 544(1): 43-64.
Lefébure, T.; Douady, C.J.; Gouy, M.; Trontelj, P.; Briolay, J.; Gibert, J. 2006.
Phylogeography of a subterranean amphipod reveals cryptic diversity and dynamic evolution
in extreme environments. Mol Ecol, 5: 1797-806.

140
Lemos, L.S.M. 2016. Áreas de risco nas bacias hidrográficas urbanizadas de Manaus.
Dissertação de Mestrado do Programa de Pós-Graduação em Geografia da Universidade
Federal do Amazonas. p. 191.
Leonhardt, C.M.R.; Caetano, L.G.; Fenocchio, A.; Filho, M.C.; Leonhardt, J.H. 2006.
Karyotypical characterization from stock of Nile tilapia, Oreochromis niloticus, at Londrina
State University, PR, Brazil, through several techniques of chromosomes band. Semana:
Ciências Agrárias, Londrina 27(4): 679-684.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric position on
chromosomes. Hereditas, 52: 201-220.
Lewinsohn, T. M.; Prado, P. I. 2005. Quantas espécies há no Brasil? Megadiversidade, 1(1):
36- 42.
Lima-Filho, P.A.; Amorim, K.D.J.; Cioffi, M.B.; Bertollo, L.A.C.; Molina, W.F. 2014a.
Chromosomal mapping of repetitive DNAs in Gobionellus oceanicus and G. stomatus
(Gobiidae; Perciformes): A shared XX/XY system and an unusual distribution of 5S rDNA
sites on the Y chromosome. Cytogenetic and Genome Research, 144(4): 333-40.
Lima-Filho, P.A.; Bertollo, L.A.C.; Cioffi, M.B.; Costa, G.W.W.F.; Molina, W.F. 2014b.
Karyotype divergence and spreading of 5S rDNA sequences between genomes of two species:
Darter and emerald gobies (Ctenogobius, Gobiidae). Cytogenetic and Genome Research,
142(3): 197-203.
Lisch DR: 2013. How important are transposons for plant evolution? Nat Rev Genet, 14:49-
61.
Liu, W.Y; Yu, Z.B.; Qiu, H.Y.; Wang, J.B.; Chen, X.Y.; Chen, K. 2018. Association between
ambient air pollutants and preterm birth in Ningbo, China: a time-series study. BMC
Pediatrics, 18: 305.
Machado, V.N. 2016. Diversidade genética dos peixes Serrasalmídeos na Amazônia. Tese de
Doutorado do Programa de Pós-Graduação em Biodiversidade e Biotecnologia da Amazônia
Legal da Universidade Federal do Amazonas, p 120.
Mannarino, C.F.; Moreira, J.C.; Ferreira, J.A.; Arias, A.R.L. 2013. Avaliação de impactos do
efluente do tratamento combinado de lixiviado de aterro de resíduos sólidos urbanos e esgoto
doméstico sobre a biota aquática. Ciência & Saúde Coletiva, 18(113): 3235-3243.
Martin, E.M.; Fry, C. 2018. Environmental Influences on the Epigenome: Exposure
Associated DNA Methylation in Human Populations. Annu. Rev. Public Health, 39: 309-333.
Martins, C.; Wasko, A.P.; Oliveira, C.; Wright, J.M. 2000. Nucleotide sequence of 5S rDNA
and localization of the ribosomal RNA genes to metaphase chromosomes of the Tilapiine
cichlid fish, Oreochromis niloticus. Hereditas, 133: 39-46.
Martins, C.; Wasko, A.P.; Oliveira, C.; Porto-Foresti, F.; Parise-Maltempi, P.P.; Wright, J.M.;
Foresti, F. 2002. Dynamics of 5S rDNA in the tilapia (Oreochromis niloticus) genome:
Repeat units, inverted sequences, pseudogenes and chromosome loci. Cytogenetic and
Genome Research, 98: 78-85.

141
Martins, C.; Oliveira, C.; Wasko, A.P.; Wright, J.M. 2004. Physical mapping of the Nile
tilapia (Oreochromis niloticus) genome by fluorescent in situ hybridization of repetitive
DNAs to metaphase chromosomes - A review. Aquaculture, 231: 37-49.
Martins, R.T.; Stephan, N.N.C.; Alves, R.G. 2008. Tubificidae (Annelida: Oligochaeta) as an
indicator of water quality in an urban stream in southeast Brazil. Acta Limnologica
Brasiliensia, 20: 221-226.
Matsumoto, S.T.; Mantovani, M.S.; Malaguttii, M.I.A.; Dias, A.L.; Fonseca, I.C.; Marin-
Morales, M.A. 2006. Genotoxicity and mutagenicity of water contaminated with tannery
effluents, as evaluated by the micronucleus test and comet assay using the fish Oreochromis
niloticus and chromosome aberrations in onion root-tips. Genetics and Molecular Biology,
29(1): 148-158.
Maumus, F.; Allen, A.E.; Mhiri, C.; Hu, H.; Jabbari, K.; Vardi, A.; Grandbastien, M.A.;
Bowler C. 2009. Potential impact of stress activated retrotransposons on genome evolution in
a marine diatom. BMC Genomics, 10(624): 1-19.
Mazzuchelli, J.; Kocher, T.D.; Yang, F.; Martins, C. 2012. Integrating cytogenetics and
genomics in comparative evolutionary studies of cichlid fish. BMC Genomics, 13: 463.
McClelland, M.; Nelson, M.; Raschke, E. 1994. Effect of site-specific modification on
restriction endonucleases and DNA modification methyltransferases. Nucleic acids research,
22: 3640-3659.
Mehner, T.; Pohlmann, K.; Elkin, C.; Monaghan, M.T.; Nitz, B.; Freyhof, J. 2010. Genetic
population structure of sympatric and allopatric populations of Baltic ciscoes (Coregonus
albula complex, Teleostei, Coregonidae). BMC Evol. Biol., 10: 85.
Melo, E.G.F.; Silva, M.S.R.; Miranda, S.A.F. 2005. Influência Antrópica sobre Águas de
Igarapés na Cidade de Manaus – Amazonas. Caminhos de Geografia, 5(16):40-47.
Mills, R.E.; Bennett, E.A.; Iskow, R.C.; Devine, S.E. 2007. Which transposable elements are
active in the human genome? Trends in Genetics, 23: 183-191.
Mita, P.; Boeke, J.D. 2016. How retrotransposons shape genome regulation. Current Opinion
in Genetics and Development, 37: 90-100.
Molina, W.F.; Pacheco, G.A.; Filho, W.M.B. 2014. Padrões citogenéticos de duas espécies de
ciclídeos de bacias do semi-árido do Brasil: Crenicichla menezesi e Cichlasoma oriental.
Biota Amazônia, 4(4):33-39.
Murayama, A.; Ohmori, K.; Fujimura, A.; Minami, H.; Yasuzawa-Tanaka, K.; Kuroda, T.; et
al. 2008. Epigenetic control of rDNA loci in response to intracellular energy status. Cell, 133:
627-639.
Muthusamy, M.; Uma, S.; Backiyarani, S.; Saraswathi, M.S.; Chandrasekar, A. 2016.
Transcriptomic Changes of Drought-Tolerant and Sensitive Banana Cultivars Exposed to
Drought Stress. Front. Plant Sci., 7: 1609.

142
Naito, K.; Cho, E.; Yang, G.; Campbell, M.A. Yano, K.; Okumoto, Y. 2006. Dramatic
amplification of a rice transposable element during recent domestication. Proceedings of the
National Academy of Sciences, 103: 17620-17625.
Naito, K.; Zhang, F.; Tsukiyama, T.; Saito, H.; Hancock, C.N.; Richardson, A.O.; Okumoto,
Y.; Tanisaka, T.; Wessler, S.R. 2009. Unexpected consequences of a sudden and massive
transposon amplification on rice gene expression. Nature, 461: 1130-1134.
Nakamoto, R.D.C.F. 2014. Meio Ambiente: recursos naturais e sua finitude. Meio Ambiente:
recursos naturais e sua finitude, São Paulo/SP, p. 30.
Nakayama, C.M.; Feldberg, E.; Bertollo, L.A.C. 2012. Karyotype differentiation and
cytotaxonomic considerations in species of Serrasalmidae (Characiformes) from the Amazon
basin. Neotropical Ichthyology, 10(1): 53-58.
Negi, P.; Rai, A.N.; Suprasanna, P.; Moving. 2016. Through the stressed genome: emerging
regulatory roles for transposons in plant stress response. Front Plant Sci, 7:1448.
Nelson, P.; Lucivaldo, J. 2016. Dados estatísticos – Igarapés Manaus. Equipe Vencer, 2011.
Disponivel em: <http://equipevencer.blogspot.com.br/2011/08/dados-estatisticos-igarapes-
manaus.html>. Acesso em: 08 novembro 2016.
Nevo, E. 2001. Evolution of genome–phenome diversity under environmental stress.
Proceedings of the National Academy of Sciences, 98: 6233-6240.
Nogueira, E.M., Yanai, A.M.; Fonseca, F.O.; Fearnside, P.M. 2015. Carbon stock loss from
deforestation through 2013 in Brazilian Amazonia. Global Change Biology, 21: 1271-1292.
Novak, S.J. 2007. The role of evolution in the invasion process. Proceedings of the National
Academy of Sciences of the United States of America, 104: 3671-3672.
Nuvolari, A.; Martinelli, A.; Telles, D.D.; Ribeiro, J.T.; Miyashita, N.J.; Rodrigues, R.B.; De
Araújo, R. 2011. Esgoto Sanitário - Coleta, Transporte, Tratamento e Reúso Agrícola. In:
Nuvolari, A. Esgoto Sanitário. 2. ed. São Paulo: Blucher.
Obiakor M. O.; Okonkwo J.C.; Nnabude P.C.; Ezeonyejiaku C.D. 2012. Eco-genotoxicology:
Micronucleus Assay in Fish Erythrocytes as In situ Aquatic Pollution Biomarker: a Review. J
Anim Sci Adv, 2(1): 123-133.
Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F.; Toledo-Filho, S.A. 1988. Supernumerary
Chromosomes, Robertsonian rearrangement and multiple NORs in Corydora aeneas (Pisces,
Siluriformes, Callichthyidae). Caryologia, 41: 227-236.
Oliveira, V.; Martins, R.; Alves, R. 2010. Evaluation of water quality of an urban stream in
Southeastern Brazil using Chironomidae larvae (Insecta: Diptera). Neotropical Entomology,
39: 873-878.
Oliveira, V., Martins, R., Alves, R., 2010. Evaluation of water quality of an urban stream in
Southeastern Brazil using Chironomidae larvae (Insecta: Diptera). Neotrop. Entomol. 39, 873-
878.

143
Oliveira, V.C.S. 2015. Análise citogenética e genotóxica de Caiman crocodilus (Linnaeus,
1758) provenientes de ambiente natural e antropizado dos arredores de Manaus. Dissertação
de Mestrado. Pós-Graduação em Genética, Conservação e Biologia Evolutiva. Manaus,
Amazonas, p. 77.
Oliver, K.R.; Greene, W.K. 2009. Transposable elements: powerful facilitators of evolution.
BioEssays, 31: 703-714.
Omar, W.A.; Mikhail, W.Z.A.; Abdo, H.M.;
Abou El Defan, T.A.; Poraas, M.M.
2015.
Avaliação do Risco Ecológico da Poluição Metálicos ao longo do Grande Setor do Cairo no
Rio Nilo, no Egito, Usando Tilapia do Nilo, Oreochromis niloticus, como Bioindicador. J
Toxicol., 167: 319.
Pantaleão, S.M. 2008. Impacto genotóxico de poluentes químicos presentes na água e
sedimento do Rio Japaratuba (Sergipe). In: Pascalicchio, A.E. 2002. Contaminação por
metais pesados. Ed. Annablume, São Paulo/SP, p. 132.
Pawar, D.H. 2012. River water pollution, an environmental crisis a case study of Panchaganga
River of Kolhapur city. Int. J. Ecol. Develop Sum, 9(1): 131-133.
Pazza, R.; Kavalco, K.F.; Penteado, P.R.; Kavalco, S.A.F.; Almeida-Toledo, L.F. 2009. Gene
mapping of 18S and 5S rDNA genes in the karyotype of the three-spot gourami Trichogaster
trichopterus (Perciformes, Osphronemidae). Zebrafish, 6: 219-222.
Philippsen, G.S. 2014. Estudo da influência de elementos transponíveis no genoma de algas
C. reinhardtii e V. carteri. Tese de Doutorado do Pós-Graduação em Física do Instituto de
Física de São Carlos, Universidade de São Paulo.
Piedade, M.T.F.; Val, V.M.F.A.; Lopes, A.; Henrique, H.S.; Fé, L.M.L.; Wittmann, F. 2014.
Organismos aquáticos e de áreas úmidas em uma Amazônia em transição. Ciência e Cultura,
66(3): 34-40.
Pinkel, D.; Straume, T.; Gray, J.W. 1986. Cytogenetic analysis using quantitative, high-
sensitivity, fluorescence hybridization. Proceedings of the Natural Academy of Science, 83:
2934-2938.
Pinto, A.G.N.; Horbe, A.M.C.; Silva, M.S.R.; Miranda, A.F.; Pascoaloato, D.; Santos, H.M.C.
2009. Efeitos da ação antrópica sobre a hidrogeoquímica do rio Negro na orla de Manaus,
AM. Acta Amazonica, 39(3): 627-638.
Pio, M.C.S.; Souza, K.S.; Santana, G.P. 2014. Capacidade da Lemna aequinoctialis para
acumular metais pesados de água contaminada. Acta Amazonica, 43(2): 203-210.
Poletto, A.B.; Ferreira, I.A.; Cabral-de-Mello, D.C.; Nakajima, R.T.; Mazzuchelli, J.; Ribeiro,
H.B. et al. 2010. Chromosome differentiation patterns during cichlid fish evolution. BMC
Genet. 11: 50.
Porto, J.I.R.; Araújo, C.S.O.; Feldberg, E. 2005. Mutagenic effects of mercury pollution as
revealed by micronucleus test on three Amazonian fish species. Environmental Research, 97:
287-292.

144
Posada, D. 2008. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol., 25(7): 1253-
1256.
Postepska-Igielska, A.; Grummt, I. 2014. NoRC silences rRNA genes, telomeres, and
centromeres. Cell Cycle, 13(4): 493-494.
Prentis, P.J.; Wilson, J.R.U.; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. 2008. Adaptive
evolution in invasive species. Trends in Plant Science, 16: 288-294.
Primack R:A. 2008. Primer of Conservation Biology. 4th ed. Sinauer Associates, Sunderland.
R Core Team. 2016. A language and environment for statistical computing. R Foundation for
Statistical Computing, Vienna, Austria, Available at: www.R-project.org/ Accessed February
3, 2019.
Rambaut, A. 2016. Fig Tree, 1.4.3 2006-2016. http://tree.bio.ed.ac.uk/software/figtree/.
Rebordinos, L.; Cross, I.; Merlo, A. 2013. High evolutionary dynamism in 5S rDNA of fish:
state of the art. Cytogenet Genome Res, 141: 103-113.
Rey, O.; Danchin, E.; Mirouze, M.; Loot, C.; Blanchet, S. 2016. Adaptation to Global
Change: A Transposable Element–Epigenetics Perspective. Trends in Ecology & Evolution,
MonthYear, Vol.xx, No.yy.
Reyna-Lopez, G.E.; Simpson, J.; Ruiz-Herrera, J. 1997. Differences in DNA methylation
patterns are detectable during the dimorphic transition of fungi by amplification of restriction
polymorphisms. Mol Gen Genet, 253(6): 703-710.
Reznick, D.N.; Ghalambor, C.K. 2001. The population ecology of contemporary adaptations:
what empirical studies reveal about the conditions that promote adaptive evolution. Genetica,
112: 183-198.
Ribeiro, L.R.; Salvadori, D.M.F.; Marques, E.K. 2003. Mutagênese Ambiental. Editora da
ULBRA. Canoas-RS. p. 356.
Rocha, P.S.; Luvizotto, J.L.; Kosmehl, T.; Böttcher, M.; Storch, V.; Braunbeck, T.; Hollert,
H. 2009. Sediment genotoxicity in the Tietê River (São Paulo, Brazil): In vitro comet assay
versus in situ micronucleus assay studies. Ecotoxicology and Environmental Safety, 72: 1842-
1848.
Rodrigues, D.O.; Silva, S.L.R.; Silva, M.S.R. 2009. Avaliação Ecotoxicológica Preliminar das
Águas das Bacias Hidrográficas dos rios Tarumã, São Raimundo e Educandos. Acta
Amazônica, 39(4):935-942.
Rolls, M.M.; Stein, P.A.; Taylor, S.S.; Há, E.; Mckeon, F.; Rapoport, T.A. 1999. A visual
screen of a GFP-fusion library identifies a new type of nuclear envelope membrane protein.
Jorunal of Cell Biology, 146(1):29-43.
Ronquist, F.; Huelsenbeck, J.F. 2003. MrBayes: Bayesian phylogenetic inference under
mixed models. Bioinformatics, 19(12): 1572-1574.

145
Ruiz-Hernandez, A.; Kuo, C.C.; Rentero-Garrido, P.; Tang, W.Y.; Redon, J.; Ordovas, J.M.;
Tellez-Plaza, M. 2015. Environmental chemicals and DNA methylation in adults: a
systematic review of the epidemiologic evidence. Clinical Epigenetics, 7(1): 55.
Rutherford, L.A.; Matthews, S.L.; Doe, K.G.; Julien, G.R.L. 2000. Aquatic Toxicity and
Environmental Impact of Leachate Discharges from a Municipal Landfill. Water Qual. Res. J.
Canada, 35(1): 39-57.
Saiki, R.K.; Gelfand, D.H.; Stoffel, S.; Scharf, S.J.; Higuchi, R.; Horn, G.T.; Mullis, K. B.;
Erlich, H.A. 1988. Primer-directed enzymatic amplification of DNA with a thermo stable
DNA polymerase. Science, 239: 487-491.
Sambrook, J.; Russell, D.W. 2001. Molecular cloning: a laboratory manual. Cold Spring
Harbor Press, Vol. I. Cold Spring Harbor, NY. p. 633-664.
Sampaio, W.M.S.; Belei, F.; Giongo, P.; Dergam, J.Á.; Orsi, M.L. 2017. Heterotilapia
buttikoferi (Hubrecht, 1881) (Perciformes: Cichlidae), an introduced exotic fish in the upper
Paraná River basin. Check List, 13: 245-250.
Santana, G.P., Barroncas, P.S.R., 2007. Estudo de metais pesados (Co, Cu, Fe, Cr, Ni, Mn, Pb
e Zn) na Bacia do Tarumã-Açu Manaus – (AM). Acta Amazon, 37(1):111-118.
Santana, G.P. 2015. Sediment-distributed metal from the Manaus Industrial District (MID)
Region (AM - Brazil). Journal of Chemical Engineering and Chemistry, 1(2): 55-64.
Santana, G.P. 2016. Heavy metal distribution in the sediment and Hoplosternum littorale
from Manaus Industrial district. Journal of chemical Eng, 2(2): 70-81.
Santos Silva, E.; Silva, C. 1993. A expasão de Manaus como Exemplo do Processo de
Extinçao dos Igarapés. Bases Científicas para Estrategias de Preservaçao e
Desenovolvimento da Amazônia, v.2.
Santos, F.A.; Marques, D.F.; Terencio, M.L.; Feldberg, E.; Rodrigues, L.R.R. 2016.
Cytogenetic variation of repetitive DNA elements in Hoplias malabaricus (Characiformes -
Erythrinidae) from white, black and clear water rivers of the Amazon basin. Genetics and
Molecular Biology, 39:40-48.
Sarria, A.J.; Lieber, J.G.; Nordee, S.K.; Evans, R.M. 1994. The presence or absence of a
vitamin-type intermediate filament network affects the shape of the nucleus in human SW-13
cells. Journal of Cell Science, 107:1593-1607.
Schaack, S.; Gilbert, C.; Feschotte, C. 2010. Promiscuous DNA: horizontal transfer of
transposable elements and why it matters for eukaryotic evolution? Trends Ecol. Evol., 25:
537-546.
Schaeffer, M.; Lyko, F. 2010. Lack of Evidence for DNA Methylation of Invader
Retroelements in Drosophila and Implications for Dnmt2? Mediated Epigenetic Regulation.
Nat. Genet., 42: 920-921.

146
Schizas, N.V.; Chandler, G.T.; Coull, B.C.; Klosterhaus, S.L.; Quattro, J.M. 2001.
Differential survival of three mitochondrial lineages of a marine benthic copepod exposed to a
pesticide mixture. Environ Sci Technol. 35(535): 53-8.
Schlesinger, S.; Selig, S.; Bergman, Y.; Cedar, H. 2009. Allelic inactivation of rDNA loci.
Genes Dev, 23: 2437-2447.
Schmid, W. 1975. The micronucleus test for cytogenetics analysis. In: Hollaender, A. (Eds).
Principles and Methods for Their Detection. vol. 4. Plenum Press, New York, p. 31-53.
Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Do Carmo, E.J.; Martins, C.; Feldberg, E.
2013a. Evolutionary dynamics of retrotransposable elements Rex 1, Rex 3 and Rex 6 in
Neotropical cichlid genomes. BMC Evolutionary Biology (Online), 13: 152.
Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Artoni, R.F.; Vicari, M.R.; Martins, C.;
Feldberg, E. 2013b. Chromosomal evolution of Neotropical cichlids: the role of repetitive
DNA sequences in the organization and structure of karyotype. Reviews in Fish Biology and
Fisheries, 23: 201-214.
Servant, G.; Pinson, B.; Tchalikian-Cosson, A.; Coulpier, F.; Lemoine, S.; Pennetier, C.; et al.
2012. Tye7 regulates yeast Ty1 retrotransposon sense and antisense transcription in response
to adenylic nucleotides stress. Nucleic acids Research, 40: 5271-5282.
Silva, F.A.; Schneider, C.H.; Feldberg, E.; Baccaro, F.B.; Carvalho, N.D.M.; Gross, M.C.
2016. Genomic organization under different environmental conditions: Hoplosternum littorale
as a Model. Zebrafish, 13(3): 197-208.
Silva, F.A.; Feldberg, F.; Carvalho, N.D.M.; Rangel, S.M.H.; Schneider, C.H.; Carvalho-
Zilse, G.A.; Silva, V.F.; Gross, M.C. 2019a. Effects of environmental pollution on the
rDNAomics of Amazonian fish. Environmental Pollution, 252:180-187.
Silva, 2019b. Elementos transponíveis de DNA em peixes amazônicos: do aumento no
genoma à adaptação a ambientes estressantes. Tese de Doutorado, Pós-Graduação em
Genética, Conservação e Biologia Evolutiva. Instituto Nacional de Pesquisas da Amazônia, p
153.
Silva, F.A.; Schneider, C.H.; Feldberg, H.; Guimarães, E.M.C.; Figliuolo, V.S.P.; Silva, V.F.;
Carvalho-Zilse, G.C.; Gross, M.C. 2019c. Genetic diversity and dynamics of repetitive DNA
elements of exotic fish introduced into the Brazilian Amazon region. Short Notes -
Adaptations of Aquatic Biota, 2019 (in progress).
Silva, F.A.; Feldberg, E.; Goll, L.G.; Carvalho-Zilse, G.A.; Gross, M.C.; Schneider, C.H.;
Matoso, D.A. 2019d. An HpaII/MspI-PCR assay to measure methylation of DNA in
Hoplosternum littorale (Callichthyidae, Siluriformes) from a polluted environment in the
central Amazon basin. Environmental Technology & Innovation, 14:100354.
Singh, N.P.; McCoy, M.T.; Tice, R.R.; Scheider, E.L. 1988. A simple technique for
quantification of low levels of DNA damage in individual cells. Experimental Cell Research,
175(1): 184-191.

147
Sioli, H. 1984. The Amazon and its main effluents: Hydrography, morphology of the river
courses, and river types. In: Sioli, H. (ed). The Amazon: limnology and landscape ecology of a
mighty tropical river and its basin. Netherlands, W. Junk Publications. 127-166.
Sioli, H. 1985. Amazônia: Fundamentos da ecologia da maior região de florestas tropicais.
Rio de Janeiro: Vozes, 72p.
Siqueira, M.M.; Moraes, M.S. 2009. Saúde coletiva, resíduos sólidos urbanos e catadores de
lixo. Cad Saude Colet, 14(6): 2115-2122.
Sisinno, C.L.S. 2002. Destino dos Resíduos Sólidos Urbanos e Industriais no Estado do Rio
de Janeiro: Avaliação da Toxicidade dos Resíduos e suas Implicações para o Ambiente e
para a Saúde Humana [tese]. Rio de Janeiro: Fiocruz.
Sochorová, S.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. 2018. Evolutionary trends
in animal ribosomal DNA loci: introduction to a new online database. Chromosoma, 127:
141-150.
Solis, E.; Zdravkovic, I.; Tomlinson, I.D.; Noskov, S.Y.; Rosenthal, S.J.; De Felice, L.J. 2012.
4-(4-(Dimethylamino)phenyl)-1-methylpyridinium (APP+): Is a Fluorescent Substrate for the
Human Serotonin Transporter? J Biol Chem., 287(12): 8852-8863.
Sotero-Caio, C.G.; Platt, R.N.; Suh, A.; Ray, D.A. 2017. Evolution and diversity of
transposable elements in vertebrate genomes. Genome Biology and Evolution, 9: 161-177.
Souza, T.D.S.; Fontanelli, C.S. 2006. Micronucleus test and observation of nuclear alterations
in erythrocytes of Nile tilapia exposed to waters affected by refinery effluents. Mutation
Research, 605: 87-93.
Souza, R.C.C.L.; Calazans, S.H.; Silva, E.P. 2009. Impacto das Espécies Invasoras no
Ambiente Aquático. Marine Biotechnology, 11: 35-41.
Srivastava, R.; Srivastava, R.; Ahn, S.H. 2016. The Epigenetic Pathways to Ribosomal DNA
Silencing. The epigenetic pathways to ribosomal DNA silencing. Microbiol Mol Biol Rev, 80:
545-563.
Stapley, J.; Santure, A.W.; Dennis, S.R. 2015. Transposable elements as agents of rapid
adaptation may explain the genetic paradox of invasive species. Molecular Ecology, 24: 2241-
2252.
Steige, K.A.; Laenen, B.; Reimegard, J.; Scofield, D.G.; Slotte, T. 2017. Genomic analysis
reveals major determinants of cis-regulatory variation in Capsella grandiflora. Proceedings of
the National Academy of Sciences of the United States of America, 114(5): 1087-1092.
Suh, A.; Smeds, L.; Ellegren, H. 2017. Abundant recent activity of retrovirus-like
retrotransposons within and among flycatcher species implies a rich source of structural
variation in songbird genomes. Molecular Ecology.,7:1-13.
Sumner, A.T. 1972. A simple technique for demonstrating centromeric heterochromatic.
Experimental Cell Research, 75: 304-306.

148
Sunjog, K.; Kolarević, S.; Héberger, K.; Gačić, Z.; Knežević-Vukčević, J.; Vuković-Gačić,
B.; et al. 2013. Comparison of comet assay parameters for estimation of genotoxicity by sum
of ranking differences. Anal Bioanal Chem., 14: 4879-4885.
Suntronpong, A.; Thapana, W.; Twilprawat, P.; Prakhongcheep, O.; Somyong, S.; Muaai, N.;
Peyachoknagul, S.; Srikulnath. K. 2017. Karyological characterization and identification of
four repetitive element groups (the 18S – 28S rRNA gene, telomeric sequences, microsatellite
repeat motifs, Rex retroelements) of the Asian swamp eel (Monopterus albus). Comparative
Cytogenetics, 11(3): 435-462.
Symonová, R.; Majtanová, Z.; Sember, A.; Staaks, G.B.O.; Bohlen, J.; Freyhof, J.; Rabová
M.; Rab, P. 2013. Genome differentiation in a species pair of coregonine fishes: an extremely
rapid speciation driven by stress activated retrotransposons mediating extensive ribosomal
DNA multiplications. BMC Evol Biol, 13: 42.
Symonová, R.W.; Howell, W.M. 2018. Vertebrate Genome Evolution in the Light of Fish
Cytogenomics and rDNAomics. Genes, 9: 96.
Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. 2013. MEGA6: Molecular
Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution, 30:2725-2729.
Tan, B.C.M.; Yang, C.C.; Hsieh, C.L.; Chou, Y.H.; Zhong, C.Z.; Yung, B.Y.M.; Liu, H.
2012. Epigeneitc silencing of ribosomal RNA genes by Mybbp1a. J Biomed Sci, 19:57.
Tang, W. et al. 2014. Heavy metal pollution characteristics of surface sediments in different
aquatic ecosystems in eastern china: A comprehensive understanding. PLoS ONE 9, e108996.
Terencio, M. L.; Schneider, C. H; Gross, M. C.; Feldberg, E. 2012. Stable karyotypes: A
general rule for the fish of the family Prochilodontidae? Hydrobiologia 686:147-156.
Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Vicari, M.R.; Farias, I.P.; Passos, K.B.;
Feldberg, E. 2013. Evolutionary Dynamics of Repetitive DNA in Semaprochilodus
(Characiformes, Prochilodontidae): A Fish Model for Sex Chromosome Differentiation.
Sexual Development, 7:325-333.
Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Do Carmo, E.J.; Nogaroto, V.; Almeida, M.C.;
Artoni, R.F.; Vicari, M.R.; Feldberg, E. 2015. Repetitive sequences: the hidden diversity of
heterochromatin in prochilodontid fish. Comp. Cytogenet., 9(4): 465-481.
Thellmann, P.; Kuch, B.; Wurm, K.; Köhler, H.R.; Triebskorn, R. 2017. Water quality
assessment in the “German River of the years 2014/2015”: how a case study on the impact of
a storm water sedimentation basin displayed impairment of fish health in the Argen River
(Southern Germany). Environ Sci Eur 29:10.
Thomas, K.V.; Silva, F.M.A.; Langford, K.H.; Souza, A.D.L.; Nizzeto, L.; Waichman, A.V.
2014a. Screening for selected human pharmaceuticals and cocaine in the urban streams of
Manaus, Amazonas, Brazil. J Am Water Resour Assoc, 50:1-7.
Thomas, E.G.; Šrut, M.; Štambuk, A.; KlobuIar, G.I.V.; Seitz, A.; Griebeler, E.M. 2014b.
Effects of Freshwater Pollution on the Genetics of Zebra Mussels (Dreissena polymorpha) at
the Molecular and Population Level. Hindawi Publishing Corporation, 11:795-481.

149
Thompson, K.W. 1979. Cytotaxonomy of 41 species of Neotropical Cichlidae. Copeia, 4:
679-691.
Tsekrekou, M., Stratigi, K., Chatzinikolaou, G., 2017. The Nucleolus: In Genome
Maintenance and Repair. Int. J. Mol. Sci. 18, 1411.
Valente, G.T.; Mazzuchelli, J.; Ferreira, I.A.; Poletto, A.B.; Fantinatti, B.E.A.; Martins, C.
2011. Cytogenetic mapping of the retroelements Rex1, Rex3 and Rex6 among Cichlid fish:
New insights on the chromosomal distribution of transposable elements. Cytogenetic Gen
Res., 133:34-42.
Van’t Hof, A.E.; Campagne, P.; Rigden, D.J.; Yung, C.J.; Lingley, J.; Quail, M.A.; Hall, N.;
Darby, A.C.; Saccheri, I.J. 2016. The industrial melanism mutation in British peppered moths
is a transposable element. Nature, 534:102-105.
Varriale, A.; Torelli, G.; Bernardi, G. 2008. Compositional properties and thermal adaptation
of 18S rRNA in vertebrates. RNA, 14:1492-1500.
Viana, M.C. 2018. Estudo de contaminantes em água, sólidos totais suspensos e sedimentos
de igarapés da zona urbana de Manaus-Amazonas. Dissertação de Mestrado pelo Programa de
Pós-Graduação em Química pela Universidade Federal do Amazonas, 108p.
Volff, J.N.; Korting, C.; Sweeney, K.; Schartl, M. 1999. The non-LTR retrotransposon Rex3
from the fish Xiphophorus is widespread among teleosts. Molecular Biology and Evolution,
16:1427-1438.
Volff, J.N., Korting, C., Schartl, M. 2000. Multiple lineages of the non-LTR retrotransposon
Rex1 with varying success in invading fish genomes. Molecular Biology and Evolution, 17:
1673-1684.
Volff, J.N.; Hornung, U.; Schartl, M. 2001a. Fish retroposons related to the Penelope element
of Drosophila virilis define a new group of retrotransposable elements. Molecular Genetics
and Genomics, 265:711-720.
Volff, J.N., Körting, C., Froschauer, A., Sweeney, K., Schartl, M. 2001b. Non-LTR
retrotransposons encoding a restriction enzyme-like endonuclease in vertebrates. Journal of
Molecular Evolution, 52:351-360.
Wang, M.; Lemos, B.; Eng, C. 2017. Ribosomal DNA copy number amplification and loss in
human cancers is linked to tumor genetic context, nucleolus activity, and proliferation. PLoS
Genet 13(9):e1006994.
Weinig, C. 2000. Plasticity versus canalization: population differences in the timing of shade-
avoidance responses. Evolution, 54: 441-451.
Witt, J.D.S.; Threloff, D.L.; Hebert, P.D.N. 2006. DNA barcoding reveals extraordinary
cryptic diversity in an amphipod genus: implications for desert spring conservation. Mol
Ecol., 15: 3073-3082.

150
Xavier, B.O.; Passos, L.; Oliveira, R.C.; Mielli, A.C. 2011. Avaliação do Efeito Genotóxico
de Diferentes Tipos de Solos para Biomonitoramento com Tradescantia Pallida. Genética na
Escola.
Yoder, J.A.; Walsh, C.P.; Bestor, T.H. 1997. Cytosine methylation and the ecology of
intragenomic parasites. Trends Gener., 13, 335-340.
Zapata, L. M.;.Bock, B. C.;Orozco, L. Y.; Palacio, J. A. 2016. Application of the
micronucleus test and comet assay in Trachemys callirostris erythrocytes as a model for in
situ genotoxic monitoring. Ecotoxicology and Environmental Safety, 127:108-116.
Zeh, D.W., Zeh, J.A., Ishida, Y. 2009. Transposable elements and an epigenetic basis for
punctuated equilibria. BioEssays 31, 715-726.
Ziemniczak, K.; Traldi, J.B.; Nogaroto, V.; Almeida, M.C.; Artoni, R.F.; Moreira-Filho, O.;
Vicari, M.R. 2014. In situ Localization of (GATA)n and (TTAGGG)n Repeated DNAs and W
Sex Chromosome Differentiation in Parodontidae (Actinopterygii: Characiformes).
Cytogenet. Genome Res., 144:325-332.
Zwickl, D.J. 2006. Genetic Algorithm Approaches for the Phylogenetic Analysis of Large
Biological Sequence Datasets under the Maximum Likelihood Criterion. Ph.D. dissertation.
The University of Texas at Austin, p. 1-125.





























