Embed Size (px)
Citation preview


El Centro Internacional de Agricultura Tropical (CIAT) es una de las 16 organizaciones queinvestigan sobre alimentación y medio ambiente, conocidas como los centros Future Harvest(Cosecha del Futuro). Estos centros están localizados alrededor del mundo y haceninvestigación en colaboración con agricultores, científicos y formuladores de políticas, paracontribuir a la reducción de la pobreza y al aumento de la seguridad alimentaria sin dejarde proteger el acervo de recursos naturales. Los centros Future Harvest son financiadosprincipalmente por 58 países, fundaciones privadas y organizaciones regionales einternacionales, que constituyen el Grupo Consultivo para la Investigación AgrícolaInternacional (GCIAI).
En el 2002, el CIAT recibió el apoyo de los siguientes países donantes: Alemania,Australia, Bélgica, Brasil, Canadá, Colombia, España, Estados Unidos de América, Francia,Holanda, Irán, Italia, Japón, México, Noruega, Nueva Zelanda, Perú, Reino Unido, Repúblicade Sudáfrica, Suecia, Suiza y Tailandia.
Nuestro trabajo fue financiado también por las siguientes organizaciones y fundaciones:el Banco Asiático de Desarrollo (ADB), el Banco Mundial, el Centro Internacional deInvestigaciones para el Desarrollo (CIID), la Corporación Andina de Fomento (CAF), el FondoInternacional para el Desarrollo Agrícola (IFAD), la Fundación Kellogg, la Fundación Nippon,la Fundación Polar, la Fundación Rockefeller, la Fundación Wallace, la Organización de lasNaciones Unidas para la Agricultura y la Alimentación (FAO), el Programa de las NacionesUnidas para el Medio Ambiente (PNUMA) y la Unión Europea (UE).
El CIAT recibe también fondos para servicios de investigación y desarrollo que se prestan,bajo contrato, a un número creciente de clientes institucionales.
La información y las conclusiones contenidas en esta publicación no reflejannecesariamente los puntos de vista de los donantes.

ISBN 958-694-059-4
Centro Internacional de Agricultura TropicalInternational Center for Tropical Agriculture
El Arado Natural:Las Comunidades de
Macroinvertebrados del Suelo en las Sabanas
Neotropicales deColombia
Editado por:Juan J. Jiménez
yRichard J. Thomas

Centro Internacional de Agricultura Tropical (CIAT)International Center for Tropical AgricultureApartado Aéreo 6713Cali, Colombia
Fax: +57 (2) 4450073Correo electrónico: [email protected]
Publicación CIAT No. 336ISBN 958-694-059-4Tiraje: 500Impreso en ColombiaNoviembre 2003
El arado natural: Las comunidades de macroinvertebrados del suelo en las sabanasneotropicales de Colombia / editado por Juan J. Jiménez y Richard J. Thomas.Cali, CO : Centro Internacional de Agricultura Tropical (CIAT), 2003.444 p. -- (Publicación CIAT no. 336)ISBN 958-694-059-4
Descriptores AGROVOC en español:
1. Biología del suelo. 2. Biodiversidad. 3. Ecosistema. 4. Fauna del suelo.5. Lumbricidae. 6. Dinámica de la población. 7. Materia orgánica del suelo.8. Química del suelo. 9. Pastizales. 10. Suelo ácido. 11. Sabanas.12. Sostenibilidad. 13. Colombia. Descriptores AGROVOC en inglés:
1. Soil biology. 2. Biodiversity. 3. Ecosystems. 4. Soil fauna. 5. Lumbricidae.6. Population dynamics. 7. Soil organic matter. 8. Soil chemistry. 9. Pastures.10. Acid soils. 11. Savannas. 12. Sustainability. 13. Colombia.
I. Jiménez Jaén, Juan José. II. Thomas, Richard J. III. Centro Internacional deAgricultura Tropical. IV. Ser. Categoría de materia AGRIS: P34 Biología del suelo / Soil biology Clasificación LC: QH 84 .8 A7
Traducción de la versión inglesa “Nature’s plow: Soil macroinvertebrate communities in theneotropical savannas of Colombia”, editada por Juan J. Jiménez y Richard J. Thomas, CIAT,Cali, Colombia, 2001.(ISBN 958-694-038-1)
Derechos de Autor CIAT 2003. Todos los derechos reservados
El CIAT propicia la amplia diseminación de sus publicaciones impresas y electrónicas para queel público obtenga de ellas el máximo beneficio. Por tanto, en la mayoría de los casos, los colegasque trabajan en investigación y desarrollo no deben sentirse limitados en el uso de los materialesdel CIAT para fines no comerciales. Sin embargo, el Centro prohíbe la modificación de estosmateriales y espera recibir los créditos merecidos por ellos. Aunque el CIAT elabora suspublicaciones con sumo cuidado, no garantiza que sean exactas ni que contengan toda lainformación.

iii
Página
Prólogo vii
Prefacio ix
Capítulo
1 La macrofauna del suelo: Un recurso natural aprovechablepero poco conocido
J. J. Jiménez, T. Decaëns, R. J. Thomas y P. Lavelle 1
PARTE IImpacto de los Agroecosistemas en las Comunidades de
Macroinvertebrados del Suelo
2 Impacto del uso de la tierra en la macrofauna del suelo de losLlanos Orientales de Colombia
T. Decaëns, P. Lavelle, J. J. Jiménez, G. Escobar, G. Rippstein,J. Schneidmadl, J. I. Sanz, P. Hoyos y R. J. Thomas 21
3 Cuantificación de la macrofauna del suelo en una cuencahidrográfica de Colombia
A. Feijoo, E. B. Knapp, P. Lavelle y A. G. Moreno 46
PARTE IICiclo de Vida y Biología de las Lombrices de Tierra de
la Sabana
4 Las comunidades de lombrices en las sabanas nativasy en los pastizales introducidos de los Llanos Orientales deColombia
J. J. Jiménez, A. G. Moreno, T. Decaëns, P. Lavelle,M. J. Fisher y R. J. Thomas 57
5 Dinámica de poblaciones y estrategias adaptativas deMartiodrilus carimaguensis (Oligochaeta, Glossoscolecidae),una especie nativa de las sabanas bien drenadas de Colombia
J. J. Jiménez, A. G. Moreno, P. Lavelle y T. Decaëns 76
6 Estrategias reproductoras de tres especies de lombrices nativasde las sabanas de Carimagua (Colombia)
J. J. Jiménez, A. G. Moreno y P. Lavelle 87
Contenido

iv
El Arado Natural:…
Capítulo Página
7 Distribución vertical de las lombrices de tierra en los suelosde los “Llanos” de Colombia cubiertos por especies forrajeras
J. J. Jiménez y T. Decaëns 96
8 Distribución espacial de las lombrices en las sabanas desuelos ácidos de los Llanos Orientales de Colombia
J. J. Jiménez, J.-P. Rossi y P. Lavelle 115
9 Diferencias en la programación de la diapausa y en los patronesde estivación de algunas especies de lombriz de tierra del trópico
J. J. Jiménez, G. G. Brown, T. Decaëns, A. Feijoo y P. Lavelle 135
10 La dieta de una lombriz anécica de las sabanas colombianas:Una cuestión relacionada con las categorías ecológicas
L. Mariani, N. Bernier, J. J. Jiménez y T. Decaëns 154
PARTE IIIEfectos Producidos por los Ingenieros del
Ecosistema en las Propiedades Físicas del Suelo,a Escala de las Estructuras Biogénicas
11 Propiedades de las estructuras construidas por los ingenierosdel ecosistema en la superficie del suelo de una sabanacolombiana
T. Decaëns, J. H. Galvis y E. Amézquita 171
12 Dinámica de la degradación de las heces superficialesde la lombriz de tierra en áreas de sabana y de pastizalen los Llanos Orientales de Colombia
T. Decaëns 198
PARTE IVEfectos Producidos por los Ingenieros del
Ecosistema en la Materia Orgánica y en el Ciclo deNutrientes del Suelo, a Escala de las Estructuras Biogénicas
13 Dinámica del nitrógeno en las heces de las lombrices de tierrade la especie Martiodrilus carimaguensis Jiménez y Moreno,en un Oxisol de las sabanas de Colombia
A. F. Rangel, R. J. Thomas, J. J. Jiménez y T. Decaëns 219
14 Disponibilidad de fósforo en las heces de una especie anécicade lombriz de tierra de las sabanas de suelo Oxisol en Colombia
J. J. Jiménez, A. Cepeda, D. K. Friesen, T. Decaëns yA. Oberson 224
15 Materia orgánica del suelo contenida en las heces que produceuna comunidad de lombrices anécico-endógea de un pastizaltropical, en Colombia
G. Guggenberger, R. J. Thomas y W. Zech 238

v
Capítulo Página
16 Dinámica del carbono y del nitrógeno en las heces de diferenteedad depositadas por lombrices en áreas de sabana y depastizal en los Llanos Orientales de Colombia
T. Decaëns, A. F. Rangel, N. Asakawa y R. J. Thomas 255
17 Características del intercambio iónico en las heces de lalombriz de tierra anécica Martiodrilus carimaguensisJiménez y Moreno, en un Oxisol de una sabana de Colombia
A. F. Rangel, E. Madero, R. J. Thomas, D. K. Friesen yT. Decaëns 274
PARTE VEfectos Producidos por los Ingenieros delEcosistema en la Biodiversidad del Suelo,
a Escala de las Estructuras Biogénicas
18 Las comunidades de la macrofauna de la superficie del sueloasociadas con las heces de las lombrices de tierra en los LlanosOrientales de Colombia
T. Decaëns, L. Mariani y P. Lavelle 285
19 Efectos causados por las lombrices en el banco de semillaspermanente del suelo en sistemas de sabana y de pastizalen Colombia
T. Decaëns, L. Mariani, N. Betancourt y J. J. Jiménez 306
PARTE VIEfectos Producidos por los Ingenieros del
Ecosistema desde la Dimensión de la Muestra Unitariahasta la Parcela Experimental
20 Determinación, mediante la espectroscopía de RMN con 31P,del nivel de fósforo orgánico de un Oxisol, en pastizalesdel trópico derivados de la sabana nativa
G. Guggenberger, L. Haumaier, R. J. Thomas y W. Zech 333
21 Alteración de la lignina y de los carbohidratos en los agregadosde un Oxisol, en pastizales tropicales derivados de la sabananativa
G. Guggenberger, W. Zech y R. J. Thomas 349
22 Efecto de la exclusión de la lombriz anécica Martiodriluscarimaguensis Jiménez y Moreno en las propiedades del sueloy en el crecimiento de las plantas, en pastizales delos Llanos Orientales de Colombia
T. Decaëns, J. J. Jiménez y P. Lavelle 370
23 Estructura espacio-temporal de una comunidad de lombricesde tierra y heterogeneidad del suelo en un pastizal tropical enCarimagua, Colombia
T. Decaëns y J.-P. Rossi 380
Contenido

vi
El Arado Natural:…
Página
PARTE VIIConclusiones Generales y Prioridades para
la Investigación Futura
Capítulo
24 Conclusiones generales, resultados importantes dela investigación y necesidades futuras
J. J. Jiménez, T. Decaëns, R. J. Thomas, L. Mariani yP. Lavelle 407
Acrónimos y Abreviaturas Usados en el Texto 439

vii
Los suelos han sido estudiadosexhaustivamente respecto a susatributos químicos y físicos; sinembargo, los conocimientosadquiridos sobre sus propiedadesbiológicas, tanto de la microflora comode la macrofauna, son todavía muyescasos. Las razones que explicanesta escasez son muy sencillas: existe,primero, una gran diversidad desuelos; éstos albergan, a su vez, unsinnúmero de taxones vivientes debacterias, de hongos, de protozoariosy de animales invertebrados; y no estarea fácil, finalmente, aplicar al suelolos métodos que requiere su estudio.Esta deficiencia científica es críticaporque hay pruebas abundantes deque la comunidad de organismos delsuelo ofrece una extensa gama deservicios que son esenciales para elfuncionamiento de un ecosistema.
Entre los animales invertebradosque habitan en el suelo, las lombricesde tierra son, generalmente, el grupomás abundante considerando subiomasa. Las lombrices participan enla regulación de la dinámica de lamateria orgánica del suelo y del ciclobiogeoquímico de los nutrientes, en lapurificación del agua, en laneutralización de las toxinasgeneradas por los productosagroquímicos, y en la modificación dela estructura del suelo. Estosservicios son esenciales para elfuncionamiento de un ecosistema y
constituyen, además, un recursoimportante para el manejo sostenibley la durabilidad de los ecosistemasagrícolas.
La capacidad de que disponemospara gestionar estas funciones enpro de la agricultura sostenible sehalla seriamente limitada, en losagroecosistemas tropicales, por unaausencia de conocimientosfundamentales sobre la biodiversidadpresente en sus suelos, sobre labiología y la ecología básicas de lasdiferentes especies que constituyenesa biodiversidad, y sobre la funciónde dichas especies en elfuncionamiento del ecosistema.
Esta obra reúne diversosestudios realizados, principalmente,en las sabanas de Colombia, yconstituye uno de los trabajos máscompletos desarrollados hasta lafecha sobre cualquier agroecosistematropical. Como tal, la obra es unacontribución importante alconocimiento actual del papel quedesempeña la macrofauna en losprocesos funcionales del suelo.
Abrigamos la esperanza de queeste estudio sea el primero demuchos similares que nos permitiránmanejar los organismos del suelo, enforma tal que se beneficien de elloslos que labran la tierra y se protejan,al mismo tiempo, el ambiente y ladiversidad biológica del suelo.
Prólogo
Prof. Mike SwiftEx-director, Programa de Biología y Fertilidad de
los Suelos Tropicales (TSBF)Nairobi, Kenya

ix
Con el fin de entender mejor el papelvital que desempeña el suelo en elfuncionamiento de un ecosistema,hay que multiplicar lasoportunidades de combinar laciencia del suelo con la ecología y labiología. La ciencia del suelo seaparta de su paradigma originalcuya propuesta es la siguiente:superar las limitantes del suelo quereducen la producción agrícola,ajustándose así a los requerimientosnutricionales de las plantas,mediante la aplicación al suelo deinsumos externos como fertilizantesy otros agroquímicos. El nuevoparadigma, que actualmente seadopta en muchos sitios, seconcentra principalmente en losprocesos biológicos, los cualescomprenden el uso de germoplasmaadaptado a las condiciones adversaspresentes en el suelo, la estimulaciónde la actividad biológica del suelo, yla obtención de un óptimo ciclo denutrientes que permita que éstos seempleen del modo más eficienteposible. El papel que losorganismos, principalmente lamacrofauna, desempeñan en el sueloha pasado a ser un área importantede la investigación que pretendemejorar el manejo sostenible de lossuelos y desarrollar guías y pautaspara lograrlo.
Un grupo multi-institucionalubicado en el Centro Internacionalde Agricultura Tropical (CIAT), en
Colombia, ha desarrollado, comocontribución a este nuevo enfoque,una serie de estudios dirigidos a ladeterminación del papel de lamacrofauna del suelo —en especial,el que cumplen las lombrices detierra— en los ecosistemas naturalesy en los agroecosistemas de losLlanos Orientales de Colombia,región ésta caracterizada por lapresencia de sabanas de clima yvegetación bien definidos. Estosestudios comprenden un conjunto detrabajos comparativos de campo, unanálisis concienzudo de los datosrecolectados, y una fase deexperimentación in situ y ex situ quepermite probar varias de lashipótesis surgidas en la primera fasedel estudio. Los resultadosobtenidos se usaron después paragenerar un modelo de las funcionesde la macrofauna tanto en losecosistemas naturales como en losintervenidos por el hombre.
En este libro se compilan losresultados de la investigaciónrealizada por los colaboradores de ungrupo de exploración y estudio, quefueron publicados en diferentesrevistas científicas. Por esta razón,cada capítulo puede ser leído conindependencia de los otros. En laParte I del libro se describen laspoblaciones de la macrofauna delsuelo que se hallaron en diferentessistemas de uso de la tierra en lassabanas de Colombia, en particular
Prefacio

x
El Arado Natural:…
en el Centro Nacional deInvestigaciones (CNI) Carimagua.En la Parte II se describen labiología y la ecología de laslombrices en las sabanas,incluyendo aquí su diversidad, sudensidad, su biomasa, la dinámicade sus poblaciones, y lasestrategias adaptativas de lasdiferentes especies frente a laestacionalidad ambiental. De laParte III a la Parte VI del libro seanalizan los efectos que causa, endiversos procesos físicos, químicosy biológicos del suelo a diferentesescalas espaciales, un importante“ingeniero del ecosistema”.
Los lectores percibirán que,aunque el CNI Carimaguarepresenta un área fitogeográficacon similitudes en las extensassabanas de Colombia, losresultados obtenidos no sonextrapolables necesariamente aotras áreas fuera de la regiónmencionada, aunque éstas poseanun clima y una fisiogeografíasimilares.
Ahora bien, puesto que laliteratura sobre la macrofauna delsuelo en el medio tropical eslimitada, el trabajo aquí descritorepresenta un importante pasohacia el conocimiento de la biologíay de la ecología funcional de estosorganismos y, por ende, de suimportancia para el manejo de losrecursos naturales.
La mayoría de los trabajospresentados en esta obra sirvieron alproyecto Macrofauna financiado porla Unión Europea (EC-STD), el cualfue dirigido por el Profesor PatrickLavelle del Laboratorio de Ecología deSuelos Tropicales del Instituto deInvestigación para el Desarrollo (IRD,anteriormente ORSTOM) de Francia.Este proyecto congrega un grupo deinvestigadores de diferentesuniversidades europeas y de centrosde investigación situados en eltrópico.
En cada capítulo1 se mencionanlas instituciones que participaron enla investigación; de ellas, lasprincipales fueron el CIAT, elLaboratorio de Ecología de SuelosTropicales (IRD), el Consorcio para elManejo de Suelos Acidos (MAS) delprograma global del GCIAI sobre elManejo de Suelos, Agua y Nutrientes(SWNM), la Universidad Complutense(Madrid, España) y la Universidad deBayreuth (Alemania).
Esta obra puede considerarsetambién un complemento a lapublicación reciente titulada“Earthworm Management in TropicalAgroecosystems”, que fue editada porLavelle, Brussaard y Hendrix (1999).
1. La versión en español de este libro provienede la obra original en inglés, publicada en elaño 2001; de los 24 capítulos que contieneésta, 21 fueron traducidos por Juan J.Jiménez. (Nota del editor)

1
La Macrofauna del Suelo:…
CAPÍTULO 1
La Macrofauna del Suelo: Un RecursoNatural Aprovechable pero PocoConocido
J. J. Jiménez* , T. Decaëns**, R. J. Thomas* yP. Lavelle***
Un observador espontáneo de lanaturaleza se sorprendería al saberque los suelos albergan algunas delas comunidades biológicas másdiversificadas del planeta, ya quecontienen de 5 a 80 millones deespecies animales pertenecientes,principalmente, a los artrópodos(Giller et al. 1997). En un bosque dehayas de Europa, 1 g de suelo puedecontener hasta 40,000 especies debacterias (Tiedje 1995) y en 1 m2 deese suelo puede haber más de 1000especies de invertebrados (Schaefer ySchauermann 1990).
A pesar de este extraordinariodespliegue de formas de vida, lataxonomía de los organismos delsuelo se conoce aún de manerainsuficiente y hay muchos génerostaxonómicos cuyas especies no hanpodido aún ser identificadas niclasificadas (Brussaard et al. 1997;Giller 1996; Giller et al. 1997;Lavelle 1996). Se han descrito hastala fecha unas 3700 especies de
lombrices de tierra que son,probablemente, menos de la mitaddel número real de especiesexistentes (Fragoso et al. 1999;Reynolds 1994). Estedesconocimiento es mucho másnotorio respecto a los suelostropicales, que son actualmente losmás amenazados por los cambiosligados a una intensificación de laagricultura cuya secuela es ladisminución de la biodiversidad(Giller et al. 1997).
Los Organismos del Suelocomo Entidad
Funcional de DiversidadLa biodiversidad —en el sentido quele dan Wilson y Peter (1988), o sea,la diversidad biológica— se definecomo “la cantidad y la estructura dela información biológica contenidaen los ecosistemas vivos organizadosjerárquicamente” (Blondel 1995). Setrata de un atributo de los sistemasvivos que puede ser considerado bajodiferentes niveles de organización,desde los genes hasta la biosfera, lacual abarca, a su vez, desdepoblaciones de especies ycomunidades hasta paisajesecológicos (Solbrig 1991b, 1994). Unecosistema que se caracterice porpresentar una diversidad específica
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** Laboratoire d’Ecologie, UFR Sciences,Université de Rouen, F-76821 Mt SaintAignan Cedex, Francia.
*** Laboratoire d’Ecologie des Sols Tropicaux,Institut de Recherche pour leDéveloppement (IRD), 32 Av. HenriVaragnat, F-93143 Bondy Cedex, Francia.

2
El Arado Natural:…
alta se define por la pluralidad desus comunidades y por las relacionesexistentes entre los elementosconstitutivos de dicha pluralidad(Blondel 1995).
Petersen y Luxton (1982)recopilaron la mayor parte de losdatos obtenidos hasta la fechasobre las comunidades demicroorganismos, sobre losmacroinvertebrados y sobre losequivalentes energéticos de ambos,dentro del Programa BiológicoInternacional (IBP) de la UniónInternacional de Ciencias Biológicas(IUBS). Swift et al. (1979)contribuyeron también a sintetizarlos conocimientos disponibles sobrelos procesos de descomposición, ydesarrollaron una nueva concepciónde las interacciones que ocurrenentre los componentes biológicos,químicos y físicos de tales procesos.
Durante la última década, ladiversidad biológica ha sido un temaextenso de preocupación y deapuesta política debido a los cambioscausados en los ecosistemasterrestres y acuáticos por el hombrea diferentes escalas, desde el paisajeecológico hasta la biosfera (Blondel1995; Schulze y Mooney 1994;Solbrig 1991b). Estos cambios sonde naturaleza diversa: laintensificación agrícola, lamodificación del ciclo del carbono ydel nitrógeno, la contaminación, elefecto invernadero, la urbanización yla desertificación, entre otros (Asneret al. 1997; Pimm y Sugden 1994;Schulze y Mooney 1994; Solbrig1991b, 1994).
La pérdida de biodiversidad enun ecosistema conduce,inexorablemente, a una alteración desus funciones principales. Diversosautores han hallado una relacióndirecta entre la abundancia deespecies, la intensidad de ciertosprocesos fundamentales —como la
respiración, la descomposición, elalmacenamiento de nutrientes, laproductividad primaria— y laretención de agua (Asner et al. 1997;Pimm y Sugden 1994; Schulze yMooney 1994; Solbrig 1991b, 1994).Los suelos, por ejemplo, albergancomunidades complejas demicroinvertebrados que estimulanlos procesos de descomposición de lamateria orgánica (Coleman et al.1998; Setälä et al. 1991; Vedder etal. 1996).
En años más recientes, ciertosestudios han sugerido la posibleexistencia de especies redundantes ode equivalentes ecológicos (Lawton yBrown 1994). Estas especiesrealizan, en esencia, una mismafunción ecológica y, si una u otraespecie faltara, no se apreciaríancambios en el funcionamiento delecosistema: es una situación muysimilar a la de una fábrica que puedecontinuar operando aunque falte unode los trabajadores. Ahora bien, siun grupo funcional, es decir, unconjunto completo de especies quedesempeñan, cada una, una funcióndada equivalente (Blondel 1995)desapareciera, se afectaría sin dudael funcionamiento del ecosistema. Lapresencia de esta redundanciaconfiere a los ecosistemas ciertaestabilidad funcional frente a unadisminución accidental de ladiversidad específica de lascomunidades (Blondel 1995; Lawtony Brown 1994). Los resultadosexperimentales de numerososestudios recientes apoyan esta teoría(Grime 1997; Hooper y Vitousek1997; Tilman et al. 1996, 1997).
La Regulación Jerárquicade los Procesos del Suelo
El modelo jerárquico de laFigura 1-1 explica el funcionamientode los procesos del suelo mediante
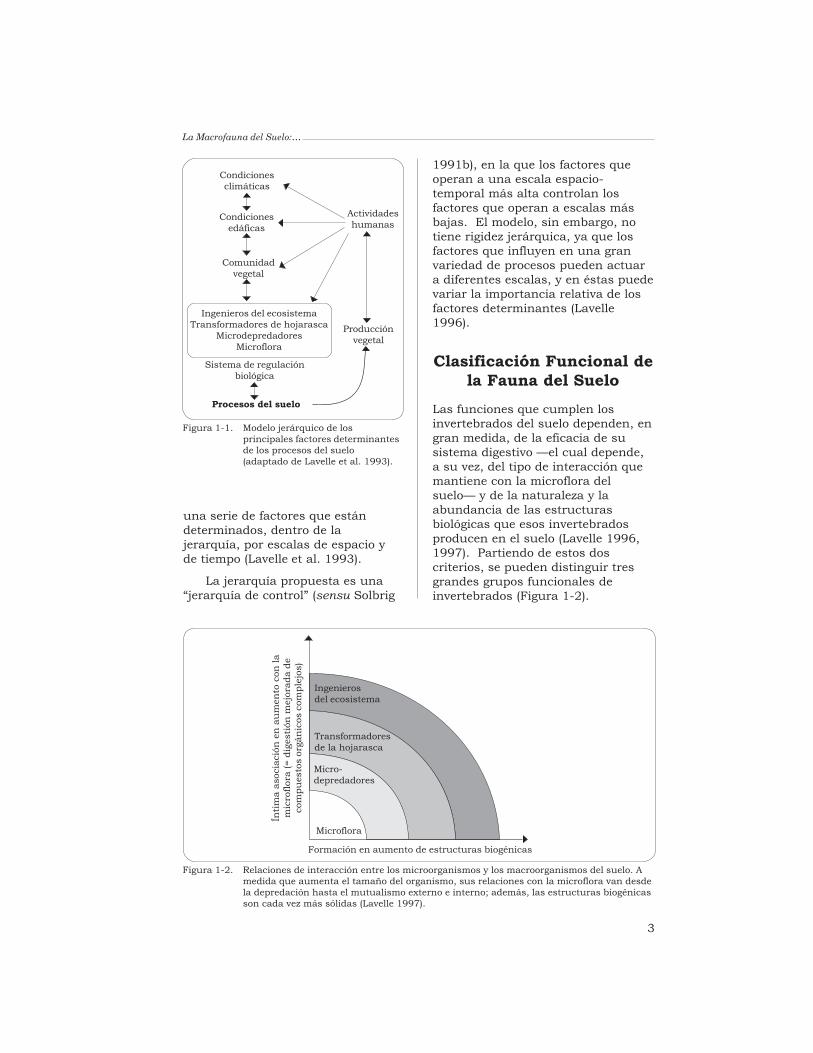
3
La Macrofauna del Suelo:…
una serie de factores que estándeterminados, dentro de lajerarquía, por escalas de espacio yde tiempo (Lavelle et al. 1993).
La jerarquía propuesta es una“jerarquía de control” (sensu Solbrig
1991b), en la que los factores queoperan a una escala espacio-temporal más alta controlan losfactores que operan a escalas másbajas. El modelo, sin embargo, notiene rigidez jerárquica, ya que losfactores que influyen en una granvariedad de procesos pueden actuara diferentes escalas, y en éstas puedevariar la importancia relativa de losfactores determinantes (Lavelle1996).
Clasificación Funcional dela Fauna del Suelo
Las funciones que cumplen losinvertebrados del suelo dependen, engran medida, de la eficacia de susistema digestivo —el cual depende,a su vez, del tipo de interacción quemantiene con la microflora delsuelo— y de la naturaleza y laabundancia de las estructurasbiológicas que esos invertebradosproducen en el suelo (Lavelle 1996,1997). Partiendo de estos doscriterios, se pueden distinguir tresgrandes grupos funcionales deinvertebrados (Figura 1-2).
Condicionesclimáticas
Condicionesedáficas
Comunidadvegetal
Ingenieros del ecosistemaTransformadores de hojarasca
MicrodepredadoresMicroflora
Actividadeshumanas
Producciónvegetal
Sistema de regulaciónbiológica
Procesos del suelo
Figura 1-1. Modelo jerárquico de losprincipales factores determinantesde los procesos del suelo(adaptado de Lavelle et al. 1993).
Formación en aumento de estructuras biogénicas
Ingenierosdel ecosistema
Transformadoresde la hojarasca
Micro-depredadores
Microflora
Figura 1-2. Relaciones de interacción entre los microorganismos y los macroorganismos del suelo. Amedida que aumenta el tamaño del organismo, sus relaciones con la microflora van desdela depredación hasta el mutualismo externo e interno; además, las estructuras biogénicasson cada vez más sólidas (Lavelle 1997).
Ínti
ma
asoc
iaci
ón e
n a
um
ento
con
lam
icro
flora
(= d
iges
tión
mej
orad
a de
com
pues
tos
orgá
nic
os c
ompl
ejos
)

4
El Arado Natural:…
MicrodepredadoresEste grupo incluye a losinvertebrados más pequeños, losprotozoos y los nematodos. Estosorganismos no producen ningunaestructura órgano-mineral (Lavelle1996, 1997) y su efecto principal esestimular la mineralización de lamateria orgánica (MO) (Coûteaux etal. 1991; Ingham et al. 1985).
Transformadores de lahojarascaEn este grupo se encuentran losrepresentantes de la mesofauna y departe de la macrofauna (Lavelle1996). Cuando estos invertebradosreingieren sus deyecciones, quesirven de incubadoras de lamicroflora, asimilan los metabolitosliberados por la acción microbiana.
Ingenieros del ecosistemaLos “ingenieros del ecosistema” o“ingenieros ecológicos” (sensu Joneset al. 1994) son aquellos organismosque producen estructuras físicas conlas cuales modifican ladisponibilidad o accesibilidad de unrecurso para otros organismos. Suactividad y la producción deestructuras biogénicas puedenmodificar la abundancia o laestructura de otras comunidades deorganismos (Jones et al. 1994,1997).
De las innumerables formas devida que habitan los suelos, sólo unpequeño número demacroinvertebrados (lombrices,termitas y hormigas) se distinguenpor su capacidad de horadar el sueloy producir una gran variedad deestructuras órgano-minerales:deyecciones, nidos, montículos,macroporos, galerías y cámaras.Estos organismos han sido descritoscomo ingenieros ecológicos del suelo,y las estructuras que producen han
sido llamadas “estructurasbiogénicas” (Anderson 1995; Lavelle1996, 1997). Se cree que la acciónfuncional de estas estructuras en elecosistema es importante y querepresentan sitios en que ocurrenalgunos procesos pedológicosfundamentales, como la estimulaciónde la actividad microbiana, laformación de la estructura del suelo,la dinámica de la MO, y elintercambio de agua y gas en el suelo(Anderson 1995; Beare y Lavelle1998; Lavelle 1996).
La visión clásica del estudio de labiología de cada especie de unacomunidad ha dado paso al estudiode las estructuras biogénicasproducidas por los ingenieros delecosistema. En efecto, este últimoconcepto y los dominios funcionalesasociados con las actividades de losingenieros han sido herramientasútiles para entender el papelfuncional de la biodiversidad y susefectos en el suelo (Jones et al. 1994;Lavelle 2000; Lavelle et al. 1997).Este concepto ha facilitado también lacomprensión de los efectos indirectosligados a las estructuras biogénicasque pueden existir entre el nivel delos macroinvertebrados del suelo y elde otros organismos de inferiortamaño.
Los dominios funcionales sonlugares específicos del suelo influidospor un regulador principal que puedeser biótico (p.ej., un ingeniero delecosistema o una raíz) o abiótico(p.ej., alternancia de períodos secos yhúmedos, o de frío y calor). En estossitios, caracterizados por el recursoorgánico que contienen (hojarasca uotro tipo de MO), el regulador bióticocrea una serie de estructuras, comodeyecciones, galerías y fisuras, queson ocupadas por invertebrados máspequeños y por microorganismos.Una comunidad biológica depende, enúltima instancia, de estos organismostan pequeños (Lavelle 2000).

5
La Macrofauna del Suelo:…
Los dominios funcionales sepueden identificar físicamente en elsuelo y cada estructura presente enel suelo es parte de un dominiofuncional. A veces, los límites de undominio son tan difusos que laidentificación de otros dominiospróximos o adyacentes resulta difícil.
En las comunidades deinvertebrados existen especies que,debido a su intensa acción mecánicay a su relación eficaz con lamicroflora, determinan laabundancia y la actividad de losorganismos que no poseen esasaptitudes. Por ejemplo, losmacroartrópodos y losmicroartrópodos, los enquitreidos ylos transformadores de la hojarascadependen de la actividad de unregulador principal. Esosorganismos, que son esencialmentelas lombrices, las hormigas y lastermitas, crean en el suelo susdominios funcionales, es decir, ladrilosfera, la mirmecosfera y latermitosfera, respectivamente.Existen otros dominios funcionales,como la rizosfera (el área deinfluencia de las raíces) (Hiltner1904) y la detritosfera (área deinfluencia de los artrópodos quetransforman la hojarasca) (Beare etal. 1994).
Aunque no todos los grupos deorganismos del suelo han sidoestudiados en detalle, cualquiercambio producido en las poblacionesde los ingenieros del ecosistematendrá, obviamente, consecuenciasdirectas en la diversidad y en laactividad de los grupos subordinadosa éstos. Por ejemplo, la actividad delas lombrices determina tanto laabundancia como las actividades delos microartrópodos (Loranger et al.1998) y de los nematodos (Boyer1998).
Las Lombrices y susEstructuras Biogénicas
como EjemploLas lombrices son el grupo demacrofauna del suelo másabundante en cuanto a biomasa serefiere (Lee 1985). Intervienen demanera directa o indirecta endiversos procesos físicos, químicos ybiológicos del suelo (Anderson 1988;Lavelle 1988). Las lombrices y losmacroinvertebrados, en general,intervienen mediante sus dominiosfuncionales en la regulación defunciones importantes del suelo.
Propiedades físicas del sueloLas lombrices ingieren, de maneraselectiva, una gran cantidad dematerial orgánico y mineral y suactividad conduce a la producciónde estructuras que influyen,directamente, en propiedades físicasdel suelo (Figura 1-3) como elaumento de la porosidad y de laaireación, el mejoramiento de laconductividad hidráulica, y unamejor estabilidad estructural queincluye la formación demacroagregados y microagregados(Aina 1984; Casenave y Valentin1988; Lavelle 1997; Lee 1985;Urbanek y Dolezal 1992).
Las lombrices influyen tambiénen la estructura física del sueloproduciendo grandes cantidades deagregados órgano-minerales en susturrículos y en las paredes de susgalerías. Los primeros soninestables cuando están frescospero, una vez secos, son másestables que los agregados del suelocircundante (Blanchart et al. 1993;Marinissen 1990, 1994; Shipitalo yProtz 1989). Asimismo, el tamañode los turrículos determina, en granmedida, los efectos causados en laestructura del suelo. En las sabanasde Costa de Marfil (Africa

6
El Arado Natural:…
Occidental), se han encontrado dostipos de turrículos producidos pordos grupos diferentes de lombrices,las especies “compactantes” (queproducen turrículos compactos degran tamaño) y las “decompactantes”(que producen turrículos pequeños)(Blanchart et al. 1997); de estosturrículos depende la formación y elmantenimiento de la estructura delsuelo (Blanchart 1998; Blanchart etal. 1999; Rossi 1998).
Dinámica de la MOLas lombrices digieren la MOempleando enzimas producidas por
ellas mismas y mediante lamicroflora mutualista que seencuentra en su tracto digestivo. Losefectos causados por las lombricesen la dinámica de la MO dependende las escalas espacio-temporalesconsideradas (Figura 1-4).
En un intervalo corto de tiempo,por ejemplo, de unas horas, ladigestión de la lombriz rompe losresiduos orgánicos y libera algunosnutrientes, como el nitrógeno (N) y elfósforo (P), que pueden entonces serasimilados por las plantas (Barois etal. 1987; Lavelle et al. 1992;Sharpley y Syers 1976). En períodosintermedios de tiempo (de días a
Figura 1-4. Efectos causados por las lombrices en la materia orgánica (MO) del suelo a diferentesniveles de una escala temporal y una espacial (Lavelle 1997).
Edad = Horas Días, semanas Meses Años, décadasEstructura = Contenido intestinal Heces frescas Heces viejas Perfíl de suelo
Efecto en la MO = Asimilación, Liberación de Protección Aceleracióntrituración nutrientes físíca del reciclaje
Propiedadeshídricas
Pedogénesis
Estructurasbiogénicas
Figura 1-3. Efectos causados por los ingenieros ecológicos en las propiedades físicas del suelo (Lavelle1997).
GaleríasCámaras
Bolas fecalesTurrículos
DomosepígeosNidos
Ingestión de grandescantidades de suelo
Selección departículas finas
Procesosbiológicos
Estructuradel suelo
Propiedadesfísicas

7
La Macrofauna del Suelo:…
meses), se altera la dinámica de laMO en el interior de las estructurasbiogénicas. En un período más largose produce un descenso en la tasa demineralización de la MO hasta llegara la inmovilización del proceso(Martin 1991). No se conoce aún elefecto general a largo plazo (de añosa décadas), porque no se hanrealizado experimentos tanprolongados en el tiempo (Lavelle1997).
Crecimiento de las plantasEl efecto de conjunto que causan laslombrices en tres aspectos: laestructura del suelo, la dinámica dela MO y la liberación de nutrientes,conduce generalmente a unaestimulación del crecimiento vegetal.La mayoría de los estudios muestranque los efectos son positivos (Curry yBoyle 1987; Derouard et al. 1997;Gilot-Villenave et al. 1996;Hoogerkamp1987; Hoogerkamp et al.1983; Pashanasi et al. 1992; Rose yWood 1980; Stephens et al. 1994;Stockdill 1982), aunque no todas lasplantas responden por igual y esarespuesta es proporcional a labiomasa de las lombrices. Larespuesta se considera ‘significativa’cuando dicha biomasa fresca essuperior a 30 g/m2 (Brown et al.1999).
Papel clave de las estructurasbiogénicasJones et al. (1994, 1997) handemostrado que la cantidad, lanaturaleza y la función de lasestructuras biogénicas producidaspor las lombrices y otros ingenierosdel ecosistema son muy importantes.
La abundancia y la diversidad delas estructuras biogénicas son, sinduda, dos aspectos importantes del
mantenimiento de las funciones delsuelo en el ecosistema (Lavelle 1996).Un ejemplo claro es el de los efectoscomplementarios de las lombricescompactantes y decompactantes(Blanchart et al. 1997; ver antes,p. 6). En un suelo que haya sidoalterado drásticamente y en el quesólo exista una especie de lombriz deefecto compactante, la producción degrandes cantidades de turrículosconduce a la compactación del suelo,la reducción de la tasa de infiltraciónde agua y la disminución delcrecimiento vegetal (Barros et al.1998; Blanchart et al. 1999; Chauvelet al. 1999; Rose y Wood 1980).
Las estructuras particularesproducidas por las lombricescausan, igualmente, efectos en ladiversidad y la abundancia de otrascomunidades de organismos menosmóviles (hipótesis de lasbiodiversidades ‘encajadas’ oanidadas de Lavelle, 1996). Laregulación de la actividad microbianapor acción de las lombrices ha sidodocumentada ampliamente (Barois yLavelle 1986; Daniel y Anderson1992; Scheu 1987; Trigo y Lavelle1993). El efecto de las estructurasbiogénicas de las lombrices en laactividad de la microfauna y lamesofauna ha sido demostradotambién en varios trabajos (versíntesis de Brown 1995).
Las Sabanas NeotropicalesLas sabanas neotropicales ocupanuna superficie de, aproximadamente,275 millones de hectáreas, quecomprende las de Brasil (250millones), Colombia (17 millones),Venezuela (10 millones), Guyana,Surinam, Guyana Francesa yParaguay, es decir, el 45% de lasuperficie total del continente

8
El Arado Natural:…
sudamericano (Cole 1986). Suposición en el planeta les confiere unclima caracterizado por temperaturasaltas y un período seco muy notorio,es decir, estas sabanas seencuentran, principalmente, entrelas selvas ecuatoriales y los desiertosde latitud media. La vegetación deuna sabana consiste en un estratoherbáceo continuo, que puede estararborizado o no estarlo; puede tenertambién arbustos de densidad yaltura variables. La presencia de lassabanas en dicha zona climática,donde la formación vegetaldominante debería ser la selvahúmeda tropical, es quizás elresultado de una época seca muyfuerte, de las diferencias edáficasexistentes y de la aparición frecuentede incendios y quemas (Blydenstein1967; Cochrane 1978).
La duración y la severidad de laépoca seca varían con cada tipo desabana, pero en todas ellas estánpresentes algunas especies vegetalesque poseen ciertas estrategiasadaptativas funcionales yestructurales, que les confierenresistencia a períodos secos cuyaduración es de 3 a 8 meses del año.Estas plantas pueden extraer aguadesde capas del suelo relativamenteprofundas y evadir además el estrésdebido a la escasez de aguadeshaciéndose de algunas hojas(estivación); de este modo sobrevivena un período seco (Cole 1986). Lassabanas constituyen, por tanto, unecosistema dinámico porquemuestran ritmos periódicos en quehay etapas de crecimiento yproductividad.
Las Sabanas de ColombiaLos “Llanos” colombianos, conocidoscomo “Llanos Orientales” deColombia, ocupan,aproximadamente, el 19% delterritorio nacional (16.9 millones de
hectáreas). Cerca del 80% de esasuperficie está cubierta por unavegetación herbácea (pastos) en laque se ha desarrollado unaganadería de tipo extensivo (Vera ySeré 1985), aunque el valor nutritivode esa vegetación nativa es pobre(Álvarez y Lascano 1987).
Aunque se cree que el fuego hasido el principal determinante de lafisionomía actual de las sabanas,porque las mantiene comoestructuras abiertas, la distribuciónque tienen hoy sugiere que noderivaron de selvas arrasadas por elfuego.
Las sabanas de los LlanosOrientales de Colombia se dividen encinco unidades fitogeográficas: elpiedemonte andino, las terrazasaluviales, la altillanura plana, laaltillanura disectada (o serranía) y lasabana de inundación (Botero 1989;Kleinheisterkamp y Häbich 1985).Las diferencias edáficas (elmicrorrelieve) y el grado deinundación de una sabanadeterminan la composición de suvegetación (Cole 1986).
SuelosLos suelos de los Llanos Orientalesde Colombia (Orinoquia colombiana)pueden dividirse en dos grandesgrupos: los Oxisoles y los Ultisoles(clasificación USDA). Los Oxisolesocupan el 75% de la superficie de lasabana en Colombia (Rippstein et al.1996). La presencia de una costralaterítica en el subsuelo que a vecesaflora a la superficie por la acción dela erosión es una de lascaracterísticas más relevantes de losLlanos. Esta costra está formadapor óxidos de hierro y aluminio ycondiciona, en gran medida, lafisionomía de las sabanas. Sólopocas especies arbóreas son capacesde penetrar la costra laterítica y, porello, las plantas herbáceas dominan

9
La Macrofauna del Suelo:…
la vegetación de sabana (Cole 1986).Los árboles que consiguen crecer enla sabana desarrollan un sistemaradicular extenso aunque superficial,por ejemplo, Curatella americana(Dilleniaceae), Byrsonima crassifolia(Malpighiaceae) y Bowdichiavirgiloides (Leguminosae).
El morichal, constituido por laspalmas moriche del género Mauritia(M. minor y M. flexuosa) es otrocomponente de la vegetación de lassabanas y se encuentra en elreborde de los esteros y en los‘bajos’.
ManejoLos Llanos Orientales de Colombiason explotados para la ganaderíaextensiva, aunque en ellos las tasasde producción animal y dereproducción son bajas, es decir, lasganancias de peso anuales oscilanentre los 15 y los 30 kg/ha de pesovivo (Kleinheisterkamp y Häbich1985; Lascano 1991; Paladines1975; Rippstein et al. 1996; Vera ySeré 1985; Vera et al. 1989). Estastasas de crecimiento estánasociadas, a su vez, con valoresbajos de carga animal que van de0.2 a 0.3 animal/ha (Guzmán y Vera1991).
La producción animal estárestringida principalmente por labaja calidad nutricional de lospastos de la sabana nativa que, porlo demás, solamente producenforraje al inicio de la época lluviosa(Rippstein et al. 1996). Rara vez usael productor pecuario un suplementomineral en la alimentación delganado. A medida que el pastizalcrece, su valor nutritivo disminuye ylos animales que lo pastanpresentan síntomas de deficienciaalimenticia. La quema periódica esuna práctica que busca eliminar elexceso de forraje seco acumulado yestimular el rebrote de las especies
nativas de mayor valor nutritivo(Rippstein et al. 1996). Este tipo demanejo determina la estructura delas comunidades vegetales de lassabanas.
Durante los últimos 20 años haaumentado en los Llanos la cantidadde tierra utilizada para la producciónintensiva de recursos alimenticios.Estos sistemas de producciónconsisten en pastizales mejoradosque se basan en gramíneasafricanas, Brachiaria spp. y Panicumspp., principalmente, ya seaasociadas con leguminosaspertenecientes a los génerosStylosanthes, Arachis y Pueraria o yaen siembra pura. Se han establecidotambién sistemas de cultivosanuales, arroz y soya generalmente,para condiciones de secano y conaplicaciones de insumos comofertilizantes y cal agrícola. Todosestos sistemas permiten un aumentorápido de la producción agrícola,pero su sostenibilidad y sudurabilidad a largo plazo están muyrestringidas.
Un factor limitante de ladurabilidad de los sistemas deproducción mencionados estárelacionado con el escaso número decultivares adaptados a lascondiciones físico-químicas de lossuelos ácidos. En el caso de losmonocultivos, las prácticas agrícolasempleadas conducen al deterioro delas propiedades físicas del suelo y ala aparición de problemas ligados aplagas, enfermedades y malezas.Los pastizales en que se asociangramíneas y leguminosas mantieneno mejoran las propiedades del suelo,aunque dependen de pequeñasaplicaciones de algún fertilizanteinorgánico, que pueden resultardemasiado costosas para la mayoríade los productores. La pocadurabilidad (contando períodoslargos) de las leguminosas en esasasociaciones es otra de las

10
El Arado Natural:…
limitaciones de estos sistemas(Friesen et al. 1996; Thomas et al.1995).
Antes de que se hiciera lainvestigación aquí presentada, no sehabía realizado ningún estudio sobrelos efectos que ejercen los cambioshechos en los sistemas de uso de latierra sobre las poblaciones y lascomunidades de la macrofauna delsuelo o de otros organismos de losLlanos Orientales de Colombia.
El Significado de“Intervención”
Los estudios que se hicieron en lassabanas fueron motivados por laexcesiva presión ejercida sobre laselva tropical, la cual es no sólo unreservorio de biodiversidad sino unactor decisivo en la regulación delciclo del carbono atmosférico y de losgases de efecto invernadero. Se creeque la intervención en las sabanaspuede ayudar a detener la elevadatasa de explotación de los bosquestropicales. La sustitución de lassabanas naturales por pastizales deespecies introducidas, generalmentegramíneas de África, es una técnicaagrícola ampliamente aceptada enlos Llanos Orientales y en lassabanas del “Cerrado” brasileño. Sinembargo, se sabe poco sobre losprocesos biológicos que sedesarrollan en estos ecosistemas;además, se desconoce la forma enque los cambios mencionadosalterarían dichos procesos y losservicios fundamentales delecosistema, por ejemplo, la cantidady la calidad del agua. Los estudioshechos en el Cerrado brasileño hanmostrado, además, que labiodiversidad de las sabanasneotropicales está, de hecho, másamenazada que la de los bosquesamazónicos (Smith et al. 1998).
El estudio de la diversidad y delos procesos ecológicos asociados conlos patrones de diversidad y con elfuncionamiento del ecosistema son elpunto de partida para comprender ymanejar los ecosistemas naturales ylos perturbados (Giller 1996). Lascomunidades de la fauna del sueloson, en general, sensibles a losfactores climáticos y edáficos quedeterminan tanto las reservasalimenticias disponibles como lascondiciones microclimáticas. Porello, las perturbaciones queexperimenten los ecosistemasnaturales alterarán las comunidadesde macroinvertebrados del suelo.
Hay que considerar la fauna delsuelo como un recurso natural quetiene un potencial de uso sostenibleen los sistemas de produccciónagrícola. Para lograr ese objetivo senecesitan dos acciones:
- conocer los ciclos de vida de lasdiferentes especies queconstituyen la comunidad deorganismos del suelo, y
- evaluar los efectos producidospor los ingenieros ecológicos enlas propiedades físicas, químicasy biológicas del suelo, adiferentes escalas espacio-temporales.
Una vez adquiridos estosconocimientos, se podrán desarrollarguías para el manejo de lasactividades de la macrofauna delsuelo en diferentes agroecosistemas.
En este trabajo se decidióevaluar el papel de las lombrices enel ecosistema descrito, porque sonorganismos representativos de lasabana que han sufrido una drásticamodificación, o aun la destrucción,de sus comunidades por lasprácticas agrícolas empleadas.Lavelle et al. (1989), Stork y Eggleton(1992) y Swift (1984) handocumentado los aspectos benéficos

11
La Macrofauna del Suelo:…
de la actividad biológica de laslombrices en diferentesagroecosistemas, y han señalado quelos efectos producidos dependen dela estructura funcional, es decir, dela composición ecológica de suscomunidades (Lavelle 1988). Laslombrices presentan algún grado deresistencia (o sea, capacidad derecuperación o ‘resiliencia’) a laperturbación, ya se trate deindividuos o de una población entera;sin embargo, en una comunidad, enla que hay diferentes poblaciones dediferentes especies, la diversidad delas especies y de la estructurafuncional pueden cambiar, lo quecausaría diferentes efectos en elecosistema. Esta capacidad derespuesta podría encerrar unatécnica que permita la“manipulación” de estascomunidades en beneficio delproductor agropecuario (Lavelle et al.1994), aunque debe mencionarseque, en algunos casos, la acción delas lombrices, unida a un mal uso delsistema de producción, puedencausar la degradación del suelo(Chauvel et al. 1999).
La Organización del LibroEl trabajo presentado en este librocomprende tres partes principales eincluye una síntesis al final:
Parte I. Impacto de losagroecosistemas en lascomunidades demacroinvertebrados del sueloAntes de desarrollar guías de uso dela macrofauna del suelo, hay queconocer bien la composición de ésta.Esta sección es, entonces, uninventario de las comunidades demacrofauna que encierran los suelosbajo diferentes prácticas de manejode la tierra.
Parte II. Ciclo de vida ybiología de las lombrices detierra de la sabanaLos capítulos de esta sección tratansobre la ecología y la biología de laslombrices, que son losrepresentantes principales de losmacroinvertebrados del suelo. Losdetalles de la ecología de estosimportantes organismos son unafuente de información excepcional ypoco frecuente sobre losagroecosistemas tropicales.
Partes III-VI. Efectosproducidos por los ingenierosecológicos en los procesos delsuelo a diferentes escalas deobservaciónEn esta sección se presentan losefectos que causan los ingenierosecológicos, en este caso laslombrices, en los procesos físicos,químicos y biológicos del suelo,desde la dimensión de lasestructuras biogénicas producidashasta la dimensión de la parcela.
Parte VII. Síntesis yperspectivasFinalmente, en un capítulo deconclusiones, se sintetizan losprincipales hallazgos de este estudioy se relacionan con las posibilidadesde uso de la macrofauna del suelo,con el fin de obtener mayorproducción vegetal y de mejorar lacalidad del ecosistema.
ReferenciasAina PO. 1984. Contribution of
earthworms to porosity and waterinfiltration in a tropical soil underforest and long-term cultivation.Pedobiologia 26:131-136.

12
El Arado Natural:…
Álvarez A; Lascano CE. 1987. Valornutritivo de la sabana bien drenadade los Llanos Orientales deColombia. Past Trop 9(3):9-17.
Anderson JM. 1988. Invertebrate-mediated transport processes insoils. Agric Ecosyst Environ24:5-19.
Anderson JM. 1995. Soil organisms asengineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linkingspecies and ecosystems. Chapmanand Hall, Londres. p 94-106.
Asner GP; Seastedt TR; Townsend AR.1997. The decoupling of terrestrialcarbon and nitrogen cycles: humaninfluences on land cover andnitrogen supply are altering naturalbiogeochemical links in thebiosphere. Bioscience 47:226-234.
Barois I; Lavelle P. 1986. Changes inrespiration rate and some physico-chemical properties of tropical soilduring gut transit throughPontoscolex corethrurus(Glossoscolecidae, Oligochaeta).Soil Biol Biochem 18:539-541.
Barois I; Verdier B; Kaiser P; Mariotti A;Rangel P; Lavelle P. 1987. Influenceof the tropical earthwormPontoscolex corethrurus(Glossoscolecidae) on the fixationand mineralization of nitrogen. En:Bonvicini Pagliai AM; Omodeo P,eds. On earthworms: selectedsymposia and monographs.Módena, Italia. p 151-158.
Barros E; Grimaldi M; Desjardins T;Sarrazin M; Chauvel A; Lavelle P.1998. Conversion of forest intopastures in Amazonia: effects onsoil macrofaunal diversity and soilwater dynamics. En: Proceedings ofthe 16th World Congress of SoilScience, 20-26 Agosto 1998,Montpellier, Francia. ISSS, AISS,IBG, SICS y AFES, Montpellier,Francia. Vol 1, p 203. (Disponibletambién en CD-ROM.)
Beare MH; Lavelle P. 1998. Regulation ofmicrobial activity and organicmatter dynamics bymacroorganisms: integratingbiological function in soil. En:Proceedings of the 16th WorldCongress of Soil Science,20-26 Agosto 1998. Montpellier,Francia. ISSS, AISS, IBG, SICS yAFES, Montpellier, Francia.(Disponible también en CD-ROM.)
Beare MH; Hendrix PF; Coleman DC.1994. Water-stable aggregates andorganic matter fractions inconventional and no-tillage soils.Soil Sci Soc Am J 58:777-786.
Blanchart E. 1998. Earthworms and soilstructure and physical properties inkaolinic and smectitic tropical soils.En: Proceedings of the 16th WorldCongress of Soil Science, 20-26Agosto 1998, Montpellier, Francia.ISSS, AISS, IBG, SICS y AFES,Montpellier, Francia. (Disponibletambién en CD-ROM.)
Blanchart E; Bruand A; Lavelle P. 1993.The physical structure of casts ofMillsonia anomala (Oligochaeta:Megascolecidae) in shrub savannasoils (Côte d’Ivoire). Geoderma56:119-132.
Blanchart E; Lavelle P; Braudeau E;Bissonnais YL; Valentin C. 1997.Regulation of soil structure bygeophagous earthworm activities inhumid savannas of Côte d’Ivoire.Soil Biol Biochem 29(3/4):431-439.
Blanchart E; Albrecht A; Alegre J;Duboisset A; Gilot-Villenave C;Pashanasi B; Lavelle P; BrussaardL. 1999. Effects of earthworms onsoil structure and physicalproperties. En: Lavelle P; BrussaardL; Hendrix PF, eds. Earthwormmanagement in tropicalagroecosystems. CAB International,Wallingford, Reino Unido.p 149-172.
Blondel J. 1995. Biogéographie:Approche ecologique et evolutive.Masson, París, Francia. 297 p.

13
La Macrofauna del Suelo:…
Blydenstein J. 1967. Tropical savannavegetation of the Llanos of Colombia.Ecology 48:1-15.
Botero BR. 1989. Manejo de explotacionesganaderas en las sabanas biendrenadas de los Llanos Orientales deColombia. Serie Boletines TécnicosNo. 2. Programa de PastosTropicales. CIAT, Cali, Colombia. 9 p.
Boyer J. 1998. Interactions biologiques(faune, ravageur, parasites,microflore) dans des sols souscultures en milieu tropical humide(Ile de la Reunion). Tesis. UniversitéParis VI, Francia.
Brown GG. 1995. How do earthwormsaffect microfloral and faunalcommunity diversity? Plant Soil170:209-231.
Brown GG; Pashanasi B; Gilot-Villenave C;Patrón JC; Senapati BK; Giri S;Barois I; Lavelle P; Blanchart E;Blakemore RJ; Spain AV; Boyer J.1999. Effects of earthworms on plantproduction. En: Lavelle P; BrussaardL; Hendrix PF, eds. Earthwormmanagement in tropicalagroecosystems. CAB International,Wallingford, Reino Unido. p 87-147.
Brussaard LJ; Behan-Pelletier VM; BignellDE; Brown VK; Didden W; FolgaraitP; Fragoso C; Freckman DW; GuptaVSR; Hattori T; Hawksworth DL;Klopatek C; Lavelle P; Malloch DW;Rusek J; Söderström B; Tiedje JM;Virginia RA. 1997. Biodiversity andecosystem functioning in the soil.Ambio 26(8):563-570.
Casenave A; Valentin C. 1988. Les états desurface: Une des clefs de l’hydrologiesahélienne. En: Demissie M; StoutGE, eds. Proceedings of the SahelForum on the State-of-the-Art ofHydrology and Hydrogeology in theArid and Semi-Arid Areas of Africa.Organización de las Naciones Unidaspara la Educación, la Ciencia y laCultura (UNESCO) y NorwegianNational Committee for Hydrology,Ouagadougou, Burkina Faso,Noviembre 7-12, 1988. p 61-72.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm. Nature398:32-33.
Cochrane TT. 1978. An ongoing appraisalof the savanna ecosystem of tropicalAmerica for beef cattle production.En: Sánchez PA; Tergas LE, eds.Producción de forrajes en los suelosácidos de los trópicos. CIAT, Cali,Colombia. p 1-12.
Cole M. 1986. The savannas: biogeographyand botany. Academic Press,Londres. 438 p.
Coleman DC; Heneghan L; Crossley DA Jr.1998. Biodiversity anddecomposition in temperate andtropical forests. En: Proceedings ofthe 16th World Congress of SoilScience, 20-26 Agosto 1998,Montpellier, Francia. ISSS, AISS,IBG, SICS y AFES, Montpellier,Francia. (Disponible también enCD-ROM.)
Coûteaux MM; Mousseau M; Célérier ML;Bottner P. 1991. Increasedatmospheric CO2 and litter quality:decomposition of sweet chestnut leafwith animal food webs of differentcomplexities. Oikos 61:54-64.
Curry JP; Boyle KE. 1987. Growth rates,establishment and effects on herbageyield of introduced earthworms ingrassland on reclaimed cutover peat.Biol Fertil Soils 3:95-98.
Daniel O; Anderson JM. 1992. Microbialbiomass and activity in contrastingsoil materials after passage throughthe gut of the earthworm Lumbricusrubellus Hoffmeister. Soil BiolBiochem 24:465-470.
Derouard L; Tondoh J; Vilcosqui L; LavelleP. 1997. Effects of earthwormintroduction on soil processes andplant growth. Soil Biol Biochem29:541-545.

14
El Arado Natural:…
Fragoso C; Lavelle P; Blanchart E;Senapati BK; Jiménez JJ; MartínezMA; Decaëns T; Tondoh J. 1999.Earthworm communities of tropicalagroecosystems: origin, structureand influence of managementpractices. En: Lavelle P; BrussaardL; Hendrix PF, eds. Earthwormmanagement in tropicalagroecosystems. CAB International,Wallingford, Reino Unido. p 27-55.
Friesen DK; Thomas RJ; Sanz JI; Rao IM;Amézquita E; Decaëns T; RippsteinG; Vera RR. 1996. Productive soilconserving/improving systems forthe tropical American savannas.Poster presented at the Congress onCrop Productivity andSustainability “Shaping the Future”,Nueva Delhi, India.
Giller PS. 1996. The diversity of soilcommunities, the “poor man’stropical rainforest”. BiodiversConserv 5:135-168.
Giller KE; Beare MH; Lavelle P; IzacA-MN; Swift MJ. 1997. Agriculturalintensification, soil biodiversity andagroecosystem function. Appl SoilEcol 6:3-16.
Gilot-Villenave C; Lavelle P; Granry F.1996. Effects of a tropicalgeophagous earthworm, Millsoniaanomala, on some soilcharacteristics, on maize-residuedecomposition and on maizeproduction in Ivory Coast. Appl SoilEcol 4:201-211.
Grime JP. 1997. Biodiversity andecosystem function: the debatedeepens. Science 277:1260-1261.
Guzmán S; Vera RR. 1991.Establecimiento de pasturas en losLlanos Orientales de Colombia. En:Lascano CE; Spain JM, eds.Establecimiento y renovación depasturas. Conceptos, experiencias yenfoque de la investigación. CIAT,Cali, Colombia. p 385-398.
Hiltner R. 1904. Uber neuere Erfahrungenund Probleme auf dem Gebiet derBodenbakteriologie und unterbesonderer Berücksichtingung derGründüngung und Brache. ArbDeutsche Landw Ges 98:59-78.
Hoogerkamp M. 1987. Effect ofearthworms on the productivity ofgrassland. An evaluation. En:Bonvicini Pagliai AM; Omodeo P, eds.On earthworms selected symposiaand monographs. Módena, Italia.p 485-495.
Hoogerkamp M; Rogaar H; Eijsackers HPJ.1983. Effect of earthworms ongrassland in recently reclaimedpolder soils in the Netherlands. En:Satchell JE, ed. Earthworm ecology:from Darwin to vermiculture.Chapman and Hall, Londres.p 85-105.
Hooper DU; Vitousek PM. 1997. The effectsof plant composition and diversity onecosystem processes. Science277:1302-1305.
Ingham RE; Trofymov JA; Ingham ER;Coleman DC. 1985. Interactions ofbacteria, fungi and their nematodegrazers: effects on nutrient cyclingand plant growth. Ecol Monogr55:119-140.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78(7):1946-1957.
Kleinheisterkamp I; Häbich G. 1985.Colombia: estudio biológico y técnico.En: Vera RR; Seré C, eds. Sistemasde producción pecuaria extensiva:Brasil, Colombia, Venezuela. Informefinal del proyecto ETES, 1978-1982.CIAT, Cali, Colombia. p 213-278.
Lascano CE. 1991. Managing the grazingresource for animal production insavannas of tropical America. TropGrassl 25:66-72.

15
La Macrofauna del Suelo:…
Lavelle P. 1988. Earthworm activities andthe soil system. Biol Fertil Soils6:237-251.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P. 2000. Ecological challenges forsoil science. Soil Sci 165:73-86.
Lavelle P; Barois I; Martin A; Zaidi Z;Schaefer R. 1989. Management ofearthworm populations inagroecosystems: a possible way tomaintain soil quality. En: ClarholmM; Bergström L, eds. Ecology ofarable land. Kluwer AcademicPublishers, La Haya, Países Bajos.p 109-122.
Lavelle P; Blanchart E; Martin A; SpainAV; Martin S. 1992. Impact of soilfauna on the properties of soils inthe humid tropics. En: Segoe S, ed.Myths and science in soils of thetropics. Special publication. SoilScience Society of America (SSSA),Madison, WI, Estados Unidos.p 157-185.
Lavelle P; Blanchart E; Martin A; MartinS; Spain AV; Toutain F; Barois I;Schaefer R. 1993. A hierarchicalmodel for decomposition interrestrial ecosystems: applicationto soils of the humid tropics.Biotropica 25(2):130-150.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B. 1994. Therelationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. Wiley & Sons,Chichester, Reino Unido.p 137-169.
Lavelle P; Bignell D; Lepage M.; WoltersV; Roger P; Ineson P; Heal OW;Dhillion S. 1997. Soil function in achanging world: the role ofecosystem engineers. Eur J Soil Biol33(4):159-193.
Lawton JH; Brown VK. 1994.Redundancy in ecosystems. En:Schulze ED; Mooney HA, eds.Biodiversity and ecosystemfunction. Springer-Verlag, Berlín.p 255-270.
Lee KE. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, NuevaYork. 411 p.
Loranger G; Ponge JF; Blanchart E;Lavelle P. 1998. Impact ofearthworms on the diversity ofmicroarthropods in a vertisol(Martinique). Biol Fertil Soils27(1):21-26.
Marinissen JCY. 1990. Mechanisms ofstabilization of casts and artificialcasts. Biol Fertil Soils 9:163-167.
Marinissen JCY. 1994. Earthwormpopulations and stability of soilstructure in a silt loam soil ofrecently reclaimed polder in theNetherlands. Agric Ecosyst Environ51:75-87.
Martin A. 1991. Short- and long-termeffects of the endogeic earthwormMillsonia anomala (Omodeo)(Megascolecidae, Oligochaeta) oftropical savannas, on soil organicmatter. Biol Fertil Soils 11:234-238.
Paladines O. 1975. El manejo y lautilización de las praderasnaturales en el trópico americano.En: El potencial para la producciónde ganado de carne en AméricaLatina. CIAT, Cali, Colombia.p 331-346.

16
El Arado Natural:…
Pashanasi B; Meléndez G; Szott L; LavelleP. 1992. Effect of inoculation withthe endogeic earthworm Pontoscolexcorethrurus (Glossoscolecidae) on Navailability, soil microbial biomassand the growth of three tropicalfruit tree seedlings in a potexperiment. Soil Biol Biochem24(12):1655-1659.
Petersen H; Luxton M. 1982. Acomparative analysis of soil faunapopulations and their role indecomposition processes. Oikos39(3):287-388.
Pimm SL; Sudgen AM. 1994. Tropicaldiversity and global change. Science263:933-934.
Reynolds J. 1994. Earthworms of theworld. Global Divers 4:11-16.
Rippstein G; Lascano CE; Decaëns T.1996. La production fourragèredans les savanes d’Amerique deSud intertropicale. Fourrages145:33-52.
Rose CJ; Wood AW. 1980. Someenvironmental factors affectingearthworm populations and sweetpotato production in the Tari Basin,Papua New Guinea Highlands.Papua New Guinea Agric J 31:1-10.
Rossi J-P. 1998. Rôle fonctionnel de ladistribution spatiale des vers deterre dans une savane humide deCôte d’Ivoire. Tesis Doctoral.Université Paris VI, Francia. 240 p.
Schaefer M; Schauermann J. 1990. Thesoil fauna of beech forests: acomparison between a mull and amoder soil. Pedobiologia34:299-314.
Scheu S. 1987. Microbial activity andnutrient dynamics in earthwormcasts (Lumbricidae). Biol Fertil Soils5:230-234.
Schulze ED; Mooney HA. 1994.Ecosystem function of biodiversity:a summary. En: Schulze ED;Mooney HA, eds. Biodiversity andecosystem function. Springer-Verlag, Berlín. p 497-510.
Setälä H; Tyynisma M; Martikainen E;Huhta V. 1991. Mineralisation of C,N, P in relation to decompositioncommunity structure in coniferousforest soil. Pedobiologia35:285-296.
Sharpley AN; Syers JK. 1976. Potentialrole of earthworm casts for thephosphorous enrichment of run-offwaters. Soil Biol Biochem8:341-346.
Shipitalo MJ; Protz R. 1989. Chemistryand micromorphology ofaggregation in earthworm casts.Geoderma 45:357-374.
Smith J; Winograd W; Gallopín G;Pachico D. 1998. Dynamics of theagricultural frontier in the Amazonand savannas of Brazil: analyzingthe impact of policy and technology.Environ Model Assess 3:31-46.
Solbrig OT. 1991a. Ecosystem complexityin time and space. En: Solbrig OT;Nicolis G, eds. Perspectives onbiological complexity. InternationalUnion of Biological Sciences (IUBS),París, Francia. p 163-188.
Solbrig OT. 1991b. The IUBS-SCOPE-UNESCO Program of Research inBiodiversity. En: Solbrig OT, ed.From genes to ecosystems: aresearch agenda for biodiversity.International Union of BiologicalSciences (IUBS), Cambridge, ReinoUnido. p 5-11.
Solbrig OT. 1994. Biodiversity: anintroduction. En: Solbrig OT;Emden HM; Oordt PGWJ, eds.Biodiversity and global change. CABInternational e International Unionof Biological Sciences (IUBS), París,Francia. p 13-20.
Stephens PM; Davoren CW; Doube BM;Ryder MH. 1994. Ability of theearthworms Aporrectodea rosea andA. trapezoides to increase plantgrowth and the foliar concentrationof elements in wheat (Triticumaestivum cv. Spear) in a sandy loamsoil. Biol Fertil Soils 18:150-154.

17
La Macrofauna del Suelo:…
Stockdill, SMJ. 1982. Effects ofintroduced earthworms on theproductivity of New Zealandpastures. Pedobiologia 24:29-35.
Stork NE; Eggleton P. 1992. Invertebratesas determinants and indicators ofsoil quality. Am J Alt Agric7(1, 2):38-47.
Swift MJ. 1984. Soil biological processesand tropical soil fertility: a proposalfor collaborative program ofresearch. Biol Int 5:1-38.
Swift MJ; Heal OW; Anderson JM. 1979.Decomposition in terrestrialecosystems. Blackwell Scientific,Oxford.
Thomas RJ; Fisher MJ; Ayarza M; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management:experimental basis forsustainability and environmentalquality. Advances in Soil ScienceSeries. Lewis Publishers, BocaRaton, FL, Estados Unidos.p 61-83.
Tiedje JM. 1995. Approaches to thecomprehensive evaluation ofProkaryote diversity of a habitat.En: Allsop D; Colwell RR;Hawksworth DL, eds. Microbialdiversity and ecosystem function.United Nations EnvironmentProgramme (UNEP) y CABInternational, Wallingford, ReinoUnido. p 73-82.
Tilman D; Wedin D; Knops J. 1996.Productivity and sustainabilityinfluenced by biodiversity ingrassland ecosystems. Nature379:718-720.
Tilman D; Knops J; Wedin D; Reich P;Ritchie M; Siemann E. 1997. Theinfluence of functional diversity andcomposition of ecosystemprocesses. Science 277:1300-1302.
Trigo D; Lavelle P. 1993. Changes inrespiration rate and somephysicochemical properties of soilduring gut transit throughAllolobophora molleri (Lumbricidae,Oligochaeta). Biol Fertil Soils15:185-188.
Urbanek J; Dolezal F. 1992. Review ofsome case studies on theabundance and the hydrolicefficiency of earthworm channels inCzechoslovak soils, with referenceto the subsurface pipe drainage.Soil Biol Biochem 24:1563-1571.
USDA (U.S. Department of Agriculture).1975. Soil taxonomy. Washington,DC. 754 p.
Vedder B; Kampichler C; Bachmann G;Bruchner A; Kandeler E. 1996.Impact of faunal complexity onmicrobial biomass and N turnoverin field mesocosms from a spruceforest soil. Biol Fertil Soils22:22-30.
Vera RR; Seré C. 1985. Brasil, Colombia,Venezuela. Los sistemas deproducción pecuaria extensiva deltrópico sudamericano. Análisiscomparativo. En: Sistemas deproducción pecuaria extensiva:Brasil, Colombia, Venezuela.Informe final del proyecto ETES,1978-1982. CIAT, Cali, Colombia.p 433-450.
Vera RR; Rivera B; Weniger JH. 1989.Composition of the diet of cattlegrazing integrated in sown tropicalpasture-savanna productionsystems. En: Seizième CongrèsInternational des Herbages, 4-15octubre 1989, Niza, Francia. Centrede Coopération Internationale enRecherche Agronomique pour leDéveloppement (CIRAD),Montpellier, Francia.
Wilson EO; Peter FM. 1988. Biodiversity.National Academy Press,Washington, DC.

PARTE I
Impacto de los Agroecosistemasen las Comunidades de
Macroinvertebrados del Suelo

21
Impacto del Uso de la Tierra…
CAPÍTULO 2
Impacto del Uso de la Tierra enla Macrofauna del Suelo de los LlanosOrientales de Colombia1
T. Decaëns*, P. Lavelle*, J. J. Jiménez**, G. Escobar***,G. Rippstein***, J. Schneidmadl***, J. I. Sanz***,
P. Hoyos*** y R. J. Thomas***
ResumenSe determinó, mediante lametodología TSBF, el efecto quecausan diferentes sistemas de usode la tierra en las comunidades demacroinvertebrados del suelo en lassabanas ácidas de Colombia. Losinvertebrados se identificaron hastael nivel de orden o familia, secontabilizaron y se agruparon engrandes unidades taxonómicas (UT),por ejemplo, lombrices, termitas,hormigas, coleópteros, arañas,miriápodos y “otros invertebrados”.La riqueza taxonómica y la densidadpoblacional más altas se registraronen el bosque de galería y en lasabana (valores medios de 4293 y2830 individuos/m2,respectivamente) y la biomasa tuvoen ambos un valor intermedio(13.6 y 15.3 g/m2, respectivamente).
Las termitas (47%) y las lombrices(31%) fueron los grupos másabundantes en la biomasa. La quemade la sabana redujo fuertemente lamacrofauna, aunque después de6 meses se habían recuperado losvalores iniciales: la biomasa y ladensidad no fueron diferentessignificativamente a las obtenidasantes de la quema, y la riquezataxonómica aumentó hasta 20 UT.
La biomasa y la riquezataxonómica de las comunidades demacroinvertebrados en los pastizalesintroducidos fueron altas mientrasque el valor de la densidadpoblacional fue intermedio. Lospastizales en que se asociabangramíneas africanas con leguminosasforrajeras presentaron los mayoresvalores de biomasa de lombrices (de22.9 a 51.1 g/m2), es decir, de 4 a10 veces más que los valoresobtenidos en la sabana. Laspoblaciones de lombrices de lospastizales estaban compuestas porlas especies nativas de la sabana yhubo en ellas una gran diversidad degrupos de macrofauna (de 26 a 32 UTrepresentadas). Los monocultivosanuales (arroz y yuca) exhibieron losvalores más bajos de biomasa (3.2 a4.3 g/m2) y de densidad (429 a592 individuos/m2) y su riquezataxonómica fue baja (18 UT). Lascomunidades de macroinvertebrados
1. Algunas partes de este trabajo han sidopublicadas en European Journal of SoilBiology 30, p 157-168, por Decaëns, Lavelle,Jiménez, Escobar, y Rippstein. Copyright ©1994 de Elsevier. Impreso con permiso de lacasa editorial.
* Laboratoire d’Ecologie des Sols Tropicaux,Institut de Recherche pour le Développement(IRD), 32 Av. Henri Varagnat, F-93143Bondy Cedex, Francia.
** Departamento de Biología Animal I, Facultadde Biología, Universidad Complutense,28040 Madrid, España.
*** CIAT, AA 6713, Cali, Colombia.

22
El Arado Natural:…
de las sabanas bien drenadas deColombia son muy sensibles a loscambios ambientales originados enla intensificación agrícola. Losresultados obtenidos en estosestudios permiten establecer variasalternativas de uso del suelo queconservan y estimulan lasactividades de la macrofauna delsuelo.
Palabras clave: biomasa, Colombia,densidad, intensificaciónagrícola, lombrices, macrofaunadel suelo, riqueza taxonómica,sabana, selva de galería, uso delsuelo
IntroducciónLos Llanos Orientales de Colombiason una vasta extensión de sabanapobremente explotada por unpastoreo extensivo. Laintensificación agrícola ha ocurridoya en las áreas más accesibles de esaregión. Los pastizales naturales ymejorados, y la rotación de loscultivos con pastizales mejorados,son algunas de las técnicas agrícolasempleadas en la actualidad; sinembargo, la sostenibilidad de estastécnicas y el impacto que causan enla calidad del suelo son todavía pococonocidos (Fisher et al. 1992). Losefectos que causan en el suelo laintroducción de gramíneas africanas,la simplificación de la comunidadvegetal o los diferentes niveles decarga animal han sido ignorados. Larápida degradación de extensasáreas de pastizales en las sabanaslatinoamericanas hacen temer a losecologistas que estos sistemas noson ni sostenibles ni capaces deconservar la biodiversidad.
Entre los diversos factores quedeterminan la calidad del suelo, losfactores biológicos reguladores, queson operados por los
macroorganismos del suelo (raícesvivas y macroinvertebrados), puedenser los más importantes (Lavelle et al.1994). Estos sistemas de regulacióntienen un papel clave en laconservación de la fertilidad del suelo,ya que afectan las propiedades físicasy químicas, la MO, el ciclo denutrientes y el crecimiento vegetal(Lavelle et al. 1994; Stork y Eggleton1992).
Los macroinvertebrados ejercendiferentes efectos en los procesos quedeterminan la fertilidad del suelo.Regulan, por ejemplo, lascomunidades de microorganismosresponsables de la mineralización y lahumificación y, en consecuencia,influyen en el ciclo de la MO y en ladisponibilidad de nutrientesasimilables por las plantas. A travésde su acción mecánica en el suelo,contribuyen a la formación deagregados estables que puedenproteger parte de la MO de unamineralización rápida y queconstituyen, por tanto, una reservade nutrientes potencialmentedisponibles para las plantas. Losmacroinvertebrados pueden modificarla textura y las propiedades físicas delsuelo en los horizontes superficialesen que habitan (Lavelle et al. 1994).Las actividades de la macrofaunafavorecen también el crecimiento delas plantas (Spain et al. 1992). Ladiversidad y la abundancia de lascomunidades de macroinvertebrados,así como la importancia relativa desus principales grupos, es decir, lastermitas, las lombrices y las hormigas,pueden utilizarse como indicadores dela calidad del suelo (Stork y Eggleton1992).
El presente estudio se realizó endos sitios de las sabanas colombianas:uno en la estación de investigaciónCarimagua (CIAT-CORPOICA) y el otroen una granja cerca a Matazul, en eldepartamento del Meta. Se consideróespecialmente el efecto que tenían

23
Impacto del Uso de la Tierra…
diferentes técnicas de manejo de lospastizales —como la quema de lasabana, la introducción degramíneas africanas (Brachiariadecumbens), su asociación conleguminosas (Pueraria phaseoloides,Arachis pintoi y Centrosemaacutifolium) y el uso de diferentesvalores de carga animal— en lamacrofauna del suelo. Por otro lado,se intentó probar la hipótesis de quelos pastizales crean “desiertosverdes” cuando son establecidos acosta de las selvas o de las sabanasnativas.
Materiales y Métodos
Características del área deestudio
Carimagua
La estación experimentalCarimagua (4°30' N, 71°30' O) estáubicada en la unidad fitogeográficadenominada “Altillanura plana”. Lavegetación característica consiste enpastos de sabana bien drenados enlas zonas altas (“altos”) y en selvasde galería en las zonas bajas(“bajos”).
El clima es subhúmedo tropicalcon una temperatura media anual de26 °C y una precipitación de2300 mm (media de los años1972-1992, datos del CIAT). Hay dos‘estaciones’ muy marcadas: unaépoca seca con una precipitaciónmedia mensual inferior a los100 mm, que transcurre desdediciembre a marzo; y una épocalluviosa cuya precipitación mediamensual supera los 100 mm y va deabril a noviembre.
En Carimagua se encuentran dostipos de suelos: los Oxisoles(Tropeptic Haplustox
Isohyperthermic) en los “altos” y losUltisoles (Ultic Aeric Plintaquox) enlos “bajos”. Presentan buenaspropiedades físicas (porosidad,retención de agua), pero sondeficientes en elementos químicos(CEC < 5 meq/100 g), altamenteácidos (pH < 5) y con una saturaciónde aluminio muy elevada (> 80%).
Granja MatazulLa granja (o finca) Matazul se
encuentra en el municipio de PuertoLópez, en el departamento del Meta,Colombia (4°9’ N, 72°39’ O),pertenece a la Altillanura plana, ytiene sabanas de Trachypogonvestitus Anders sobre un sueloOxisol. El clima es idéntico al deCarimagua, aunque la precipitaciónmedia anual es un poco mayor(2700 mm).
Elección de las prácticas deuso de la tierra
En CarimaguaEl muestreo se hizo en Oxisoles
franco-arcillosos cubiertos por lasabana nativa o por agroecosistemas(como pastizales y cultivos), durantela época lluviosa, desde mayo hastaagosto de 1993. Las muestras demacrofauna del suelo se tomaron en10 sistemas diferentes de uso de latierra:
I. Selva de galería nativa.
II. Sabana nativa protegida contrael fuego y dedicada a pastoreodurante más de 4 años.
III, IV, V. Sabana quemada y bajopastoreo, con carga animal baja(0.25 animales/ha), muestreada15 días después del fuego, DDF(III), 6 meses DDF (IV) y un añoDDF (V).

24
El Arado Natural:…
VI, VII. Sabana quemada y bajopastoreo, con carga animalmoderada (VI) y alta (VII) (0.5 y0.75 animales/ha,respectivamente), 16 meses DDF.
VIII. Pastizal de 15 años de edad deBrachiaria decumbens,establecido en la sabana nativa,con 1.67 animales/ha.
IX. Pastizal de 15 años de edadde B. decumbens + kudzú(Pueraria phaseoloides),establecido en la sabana nativa,con 1.67 animales/ha.
X. Cultivo de arroz de 2 meses deedad, sembrado con un nivel altode insumos en la sabana nativa.
XI. Cultivo de yuca de 8 meses deedad, plantado con un nivel altode insumos en un terreno en quese plantó yuca durante 20 añosen la sabana nativa.
El pastoreo extensivo tradicionalen pastizales naturales estárepresentado por los sitios III, IV, V,VI y VII, y el pastoreo intensivo enpastizales mejorados se muestreó enlos sitios VIII y IX. Los monocultivosanuales con niveles altos de insumoscorresponden a los sitios X y XI.
En Granja MatazulSe tomaron muestras en cinco
parcelas en que se emplearondiferentes sistemas de uso de latierra, en agosto de 1995:
Parcela 1: Sabana nativa que fuequemada 8 meses antes de tomarmuestras, y que ha sidopastoreada desde 1989 a razónde 0.25 animales/ha.
Parcela 2: Pastizal de 6 años deBrachiaria humidicola (Rendle),pastoreado de manera continua arazón de 1.5 animales/ha.
Parcela 3: Pastizal de 2 años deB. humidicola, pastoreado demanera continua a razón de2.17 animales/ha; fueestablecido inicialmente con uncultivo pionero de arrozsembrado en 1989 y se resembrócon arroz y pastos en 1993.
Parcela 4: Pastizal de 2 años en quese asociaban B. humidicola,Arachis pintoi Krap and Greg yCentrosema acutifolium Benth.,pastoreado de manera continuaa razón de 1.79 animales/ha.
Parcela 5: Monocultivo de arrozcultivado continuamente desde1989.
Método de muestreo y análisisde los datos
El muestreo se realizó siguiendo lametodología recomendada por elPrograma de Biología y Fertilidad delos Suelos Tropicales, TSBF(Anderson e Ingram 1993; Lavelle yPashanasi 1989). En Carimagua setomaron 10 muestras de 25 x 25 x30 cm a intervalos regulares de 5 m,a lo largo de un transecto cuyoorigen y dirección fueron escogidos alazar. Se empleó un marco metálicopara aislar el monolito de suelo, elcual fue extraído con una pala ydividido en cuatro capas sucesivas(hojarasca, 0-10, 10-20, 20-30 cm).En Matazul se tomaron ochomuestras en la sabana nativa y seisen el resto de las parcelas. Cadaestrato fue desmenuzado y revisadocuidadosamente en bandejasgrandes de 40 x 60 cm. Losmacroinvertebrados se recolectaron,se contaron, se pesaron y seguardaron en alcohol 75%, exceptolas lombrices, que fueronpreviamente fijadas en formalina al4% durante 2 ó 3 días.

25
Impacto del Uso de la Tierra…
En Carimagua, los invertebradosfueron identificados comopertenecientes a 57 UT amplias(orden o familia), se contaron y seagruparon luego en siete grandesgrupos: lombrices, termitas,hormigas, coleópteros, arañas,miriápodos y “otros invertebrados”.Se determinó la densidad y labiomasa de cada uno de estos sietegrupos; la biomasa se corrigióposteriormente debido a las pérdidasde peso debida al proceso de fijación(un 19% en las lombrices y lastermitas, un 9% en las hormigas, un11% en coleópteros, un 6% en arañasy miriápodos y un 13% en “otrosinvertebrados”).
Se realizó luego un análisis decomponentes principales (ACP) conlos programas GraphMu y MacMul(Thioulouse 1989, 1990) con el fin decomparar las comunidades demacroinvertebrados respecto a losdiferentes tipos de uso de la tierra eidentificar los factores determinantesde su distribución. Se emplearon39 variables que describen lamacrofauna, las propiedades físicas yquímicas del suelo, la carga animal ylas características de la vegetaciónherbácea. Los sitios X y XI no seincluyeron en el análisis porque no secontó con información sobre laspropiedades de sus suelos. Losresultados obtenidos en los diferentestratamientos se compararon con laprueba de t de Student.
En Matazul, los invertebrados secontaron, se pesaron y seidentificaron sólo hasta el nivel defamilia. Se emplearon criteriosmorfológicos (morfotipos) paraidentificar el número de especies delombrices, a diferencia de losmuestreos hechos en Carimagua,donde se hizo un trabajo másdetallado (Jiménez 1999). Los otrosgrupos se identificaron hasta el nivelde UT morfológicamente diferentes(morfotipos).
La riqueza taxonómica (S),definida como el número total de UTpresentes en un sistema, se calculócon dos niveles de precisión: uno, elnúmero de familias, y otro, elnúmero de morfotipos y especies.Todos los resultados provienen deuna compilación de 10 muestrasindividuales, excepto en el sitio XI,donde sólo se tomaron 4 muestras.
Resultados y Discusión
En CarimaguaLa lista de las UT observadas enCarimagua aparece en el Apéndice(p. 44). Los resultados del análisisde componentes principales (ACP)que se hizo con los datos de estaestación aparecen en la Figura 2-1 yen el Cuadro 2-1. Se extrajeron dosfactores que explicaron el 66.4% dela varianza total.
Factor IEste factor explica el 42.0% de la
varianza. Se define como el efectoglobal de la vegetación. Las selvas
Pastizalesmejorados
Factor II
Protegida
Sabana
Factor I
Quemaday pastoreada
Selva de galeríaI
IX
II
VII
VIII
VVIIVIII
Figura 2-1. Localización de los diferentes sitiosestudiados respecto a los dosprimeros factores extraídos en elACP.

26
El Arado Natural:…
de galería están claramenteseparadas de los pastizales,especialmente en los sitios quepresentan cobertura mínima, como lasabana a los 15 DDF o cuando tieneuna carga animal alta. Este factorsepara suelos con contenidos de limoaltos (63%), buena porosidad (60%),saturación de aluminio alta (88%) yelevados contenidos de Mg (0.2 meq/100 g) y de K (0.1 meq/100 g). Enestos suelos, la densidad y la riquezataxonómica de la macrofauna(número de taxones recolectados) sonaltas. Los miriápodos y otrosartrópodos (especialemente las larvasde la familia Cicadidae, los blátidos ylos isópodos) presentan un valor altode biomasa; las lombrices, lashormigas, las termitas y losmiriápodos son abundantes.
Factor IIEste factor explica el 24.4% de la
varianza y representa el efecto de lafertilidad del suelo. Este factorsepara con claridad la selva de galeríay las sabanas de los pastizalesmejorados. Aquí se encuentransuelos con poca resistencia a lapenetración (12-14 g/cm2) y con
Cuadro 2-1. Principales correlaciones entre las variables con los dos primeros factores extraídos delACP.
Factor I: 41.96% de la varianza total
Densidad de lombrices 0.74 Densidad (%) en la hojarasca 0.58Densidad de termitas 0.80 Contenido de Mg 0.75Densidad de hormigas 0.68 Contenido de K 0.86Densidad de miriápodos 0.98 Saturación de aluminio (%) 0.91Densidad de macrofauna 0.90 Arcilla (%) -0.91Biomasa de miriápodos 0.83 Infiltración de agua 0.97Biomasa de “otros” 0.83 Densidad del suelo -0.86Número de taxones 0.81 Porosidad 0.99
Factor II: 24.4% de la varianza total
Densidad de hormigas 0.85 pH 0.83Densidad de “otros” 0.68 Contenido de Ca 0.90Biomasa de lombrices 0.91 Resistencia a la penetración -0.77Biomasa de coleópteros 0.60 Leguminosa (%) 0.84Biomasa de macrofauna 0.92 Carga animal 0.88
contenido de Ca alto (0.53 meq/100 g), que presentan una vegetacióncon gran proporción de leguminosas(48%) y reciben una carga animal másalta (1.67 animales/ha). La biomasade macroinvertebrados esconsiderable a causa de la enormecontribución de las lombrices. Haypoca representación de las hormigas,aunque mucha de los coleópteros,cuya densidad y biomasa son altas.
Se pueden distinguir por tantocinco situaciones principales: la selvade galería, la sabana protegida delfuego, los pastizales naturales, lospastizales introducidos y losmonocultivos (Figura 2-1).
La selva de galería presentó lariqueza taxonómica y la densidadpoblacional más altas, aunque losvalores de su biomasa son de nivelintermedio (Cuadros 2-2 y 2-3). Losinvertebrados ocuparon,principalmente, los primeros 10 cmde suelo (84%) y sólo el 4% de ellos seencontró en la hojarasca(Figura 2-2). Las lombrices, lastermitas, las hormigas, los miriápodosy “otros artrópodos” presentaron lasdensidades más altas (Cuadro 2-3,Figura 2-3). El mayor componente de
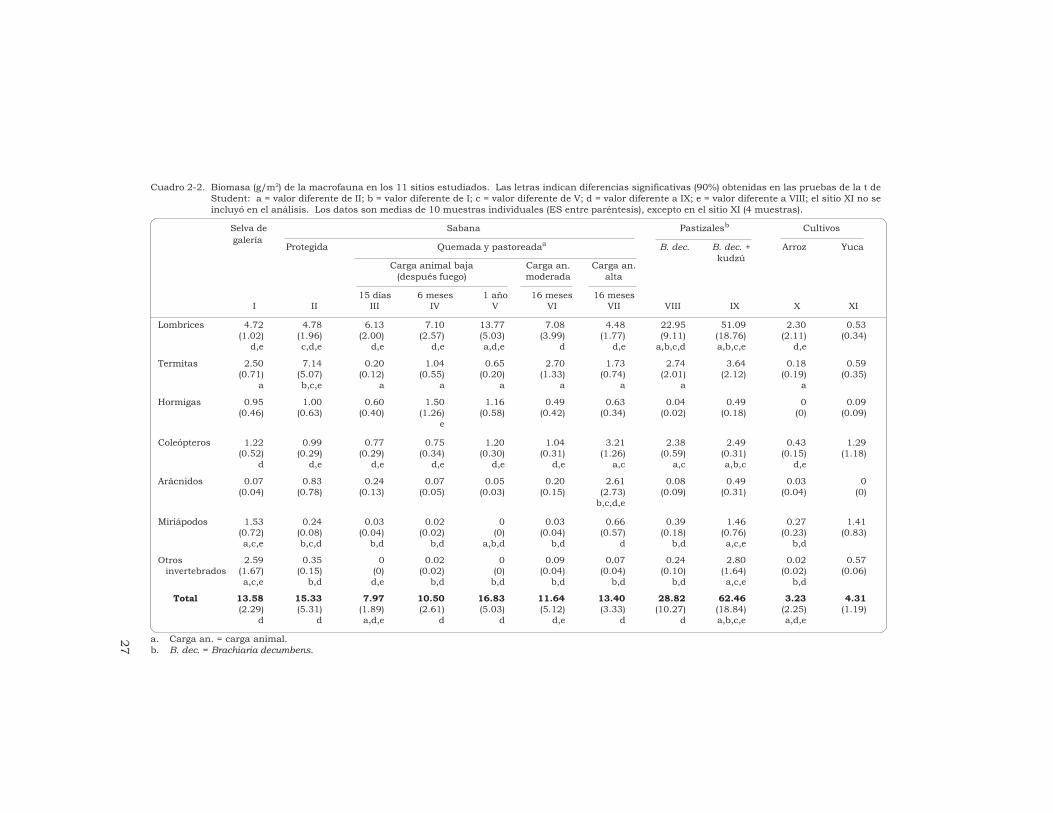
27
Impacto del U
so de la Tierra…
Cuadro 2-2. Biomasa (g/m2) de la macrofauna en los 11 sitios estudiados. Las letras indican diferencias significativas (90%) obtenidas en las pruebas de la t deStudent: a = valor diferente de II; b = valor diferente de I; c = valor diferente de V; d = valor diferente a IX; e = valor diferente a VIII; el sitio XI no seincluyó en el análisis. Los datos son medias de 10 muestras individuales (ES entre paréntesis), excepto en el sitio XI (4 muestras).
Selva de Sabana Pastizalesb Cultivos
Protegida Quemada y pastoreadaa B. dec. B. dec. + Arroz Yuca
Carga animal baja Carga an. Carga an.kudzú
(después fuego) moderada alta
15 días 6 meses 1 año 16 meses 16 mesesI II III IV V VI VII VIII IX X XI
Lombrices 4.72 4.78 6.13 7.10 13.77 7.08 4.48 22.95 51.09 2.30 0.53(1.02) (1.96) (2.00) (2.57) (5.03) (3.99) (1.77) (9.11) (18.76) (2.11) (0.34)
d,e c,d,e d,e d,e a,d,e d d,e a,b,c,d a,b,c,e d,e
Termitas 2.50 7.14 0.20 1.04 0.65 2.70 1.73 2.74 3.64 0.18 0.59(0.71) (5.07) (0.12) (0.55) (0.20) (1.33) (0.74) (2.01) (2.12) (0.19) (0.35)
a b,c,e a a a a a a a
Hormigas 0.95 1.00 0.60 1.50 1.16 0.49 0.63 0.04 0.49 0 0.09(0.46) (0.63) (0.40) (1.26) (0.58) (0.42) (0.34) (0.02) (0.18) (0) (0.09)
e
Coleópteros 1.22 0.99 0.77 0.75 1.20 1.04 3.21 2.38 2.49 0.43 1.29(0.52) (0.29) (0.29) (0.34) (0.30) (0.31) (1.26) (0.59) (0.31) (0.15) (1.18)
d d,e d,e d,e d,e d,e a,c a,c a,b,c d,e
Arácnidos 0.07 0.83 0.24 0.07 0.05 0.20 2.61 0.08 0.49 0.03 0(0.04) (0.78) (0.13) (0.05) (0.03) (0.15) (2.73) (0.09) (0.31) (0.04) (0)
b,c,d,e
Miriápodos 1.53 0.24 0.03 0.02 0 0.03 0.66 0.39 1.46 0.27 1.41(0.72) (0.08) (0.04) (0.02) (0) (0.04) (0.57) (0.18) (0.76) (0.23) (0.83)a,c,e b,c,d b,d b,d a,b,d b,d d b,d a,c,e b,d
Otros 2.59 0.35 0 0.02 0 0.09 0.07 0.24 2.80 0.02 0.57invertebrados (1.67) (0.15) (0) (0.02) (0) (0.04) (0.04) (0.10) (1.64) (0.02) (0.06)
a,c,e b,d d,e b,d b,d b,d b,d b,d a,c,e b,d
Total 13.58 15.33 7.97 10.50 16.83 11.64 13.40 28.82 62.46 3.23 4.31(2.29) (5.31) (1.89) (2.61) (5.03) (5.12) (3.33) (10.27) (18.84) (2.25) (1.19)
d d a,d,e d d d,e d d a,b,c,e a,d,e
a. Carga an. = carga animal.b. B. dec. = Brachiaria decumbens.
galería
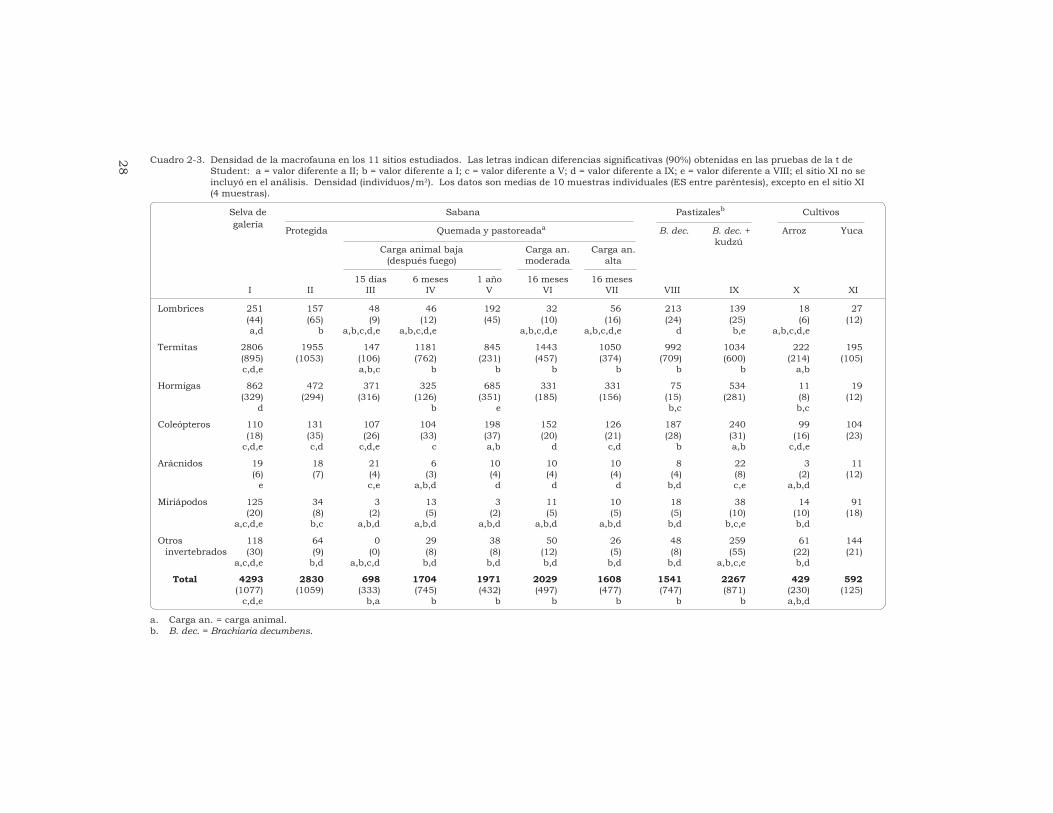
28
El A
rado Natural:…
Cuadro 2-3. Densidad de la macrofauna en los 11 sitios estudiados. Las letras indican diferencias significativas (90%) obtenidas en las pruebas de la t deStudent: a = valor diferente a II; b = valor diferente a I; c = valor diferente a V; d = valor diferente a IX; e = valor diferente a VIII; el sitio XI no seincluyó en el análisis. Densidad (individuos/m2). Los datos son medias de 10 muestras individuales (ES entre paréntesis), excepto en el sitio XI(4 muestras).
Selva de Sabana Pastizalesb Cultivos
Protegida Quemada y pastoreadaa B. dec. B. dec. + Arroz Yuca
Carga animal baja Carga an. Carga an.kudzú
(después fuego) moderada alta
15 días 6 meses 1 año 16 meses 16 mesesI II III IV V VI VII VIII IX X XI
Lombrices 251 157 48 46 192 32 56 213 139 18 27(44) (65) (9) (12) (45) (10) (16) (24) (25) (6) (12)a,d b a,b,c,d,e a,b,c,d,e a,b,c,d,e a,b,c,d,e d b,e a,b,c,d,e
Termitas 2806 1955 147 1181 845 1443 1050 992 1034 222 195(895) (1053) (106) (762) (231) (457) (374) (709) (600) (214) (105)c,d,e a,b,c b b b b b b a,b
Hormigas 862 472 371 325 685 331 331 75 534 11 19(329) (294) (316) (126) (351) (185) (156) (15) (281) (8) (12)
d b e b,c b,c
Coleópteros 110 131 107 104 198 152 126 187 240 99 104(18) (35) (26) (33) (37) (20) (21) (28) (31) (16) (23)
c,d,e c,d c,d,e c a,b d c,d b a,b c,d,e
Arácnidos 19 18 21 6 10 10 10 8 22 3 11(6) (7) (4) (3) (4) (4) (4) (4) (8) (2) (12)
e c,e a,b,d d d d b,d c,e a,b,d
Miriápodos 125 34 3 13 3 11 10 18 38 14 91(20) (8) (2) (5) (2) (5) (5) (5) (10) (10) (18)
a,c,d,e b,c a,b,d a,b,d a,b,d a,b,d a,b,d b,d b,c,e b,d
Otros 118 64 0 29 38 50 26 48 259 61 144invertebrados (30) (9) (0) (8) (8) (12) (5) (8) (55) (22) (21)
a,c,d,e b,d a,b,c,d b,d b,d b,d b,d b,d a,b,c,e b,d
Total 4293 2830 698 1704 1971 2029 1608 1541 2267 429 592(1077) (1059) (333) (745) (432) (497) (477) (747) (871) (230) (125)
c,d,e b,a b b b b b b a,b,d
a. Carga an. = carga animal.b. B. dec. = Brachiaria decumbens.
galería

29
Impacto del Uso de la Tierra…
Figura 2-2. Distribución vertical de la densidad de la macrofauna en 10 sitios estudiados.(Hojar. = hojarasca.)
I. Selva de galería
Hojar.
0-10
10-20
20-30
II. Sabana protegida
III. Sabana, 15 días despuésdel fuego
Hojar.
0-10
10-20
20-30
V. Sabana, 1 año después del fuego (carga animal baja)
Hojar.
0-10
10-20
20-30
Hojar.
0-10
10-20
20-30
IX. Pastizal de B. decumbens +kudzú
Hojar.
0-10
10-20
20-30
0 25 50 75 100%
VII. Sabana, 16 meses después del fuego (carga animal alta)
IV. Sabana, 6 meses despuésdel fuego
VI. Sabana, 16 meses despuésdel fuego (carga animalmoderada)
VIII. Pastizal de B. decumbens
X. Monocultivo de arroz
0 25 50 75 100%
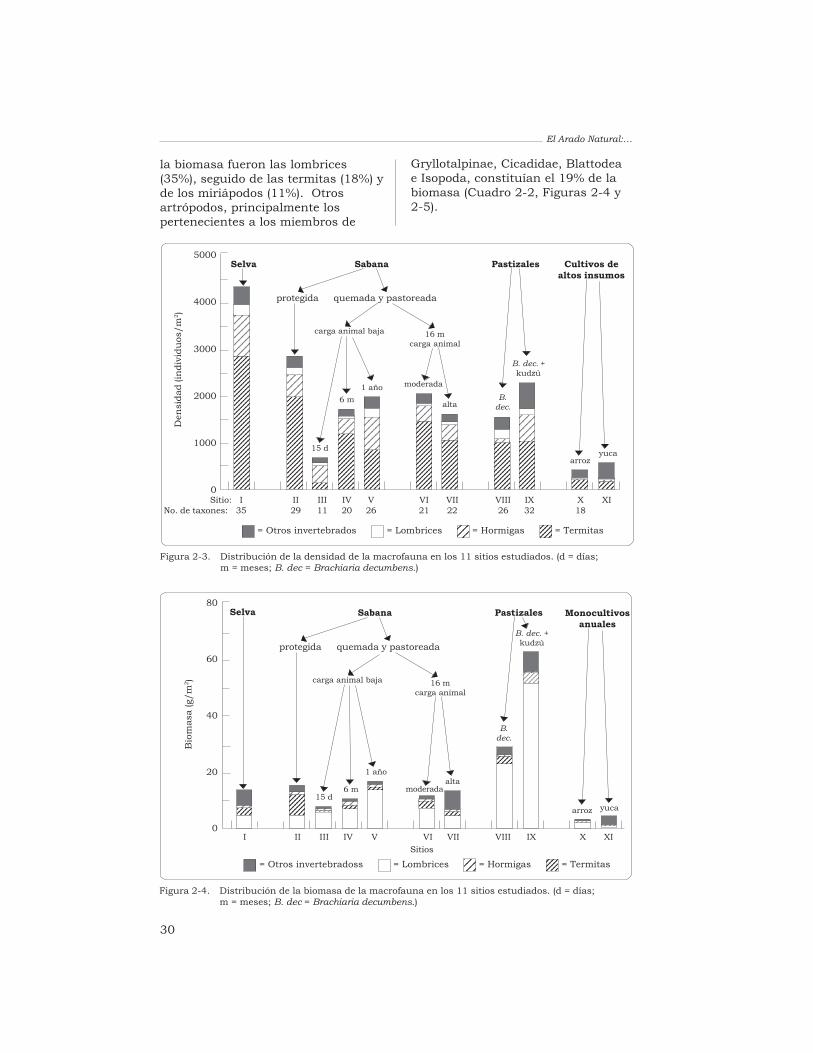
30
El Arado Natural:…
Figura 2-3. Distribución de la densidad de la macrofauna en los 11 sitios estudiados. (d = días;m = meses; B. dec = Brachiaria decumbens.)
123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234
123123123123123123123
123123
123412341234123412341234123412341234123412341234123412341234123412341234123412341234
1234123412341234
12341234
12345123451234512345123451234512345123451234512345123451234512345
123123123
123
1234123412341234123412341234123412341234
123412341234123412341234
12341234
1234512345
123123123
123123
1234123412341234123412341234123412341234123412341234123412341234
123123123123
123
123451234512345123451234512345123451234512345123451234512345
123412341234
1234
1234512345123451234512345123451234512345123451234512345
123123
123123
12341234123412341234123412341234123412341234
1234123412341234
12341234
123451234512345
1231231234512345
123123
5000
4000
3000
2000
1000
0
Sabana Pastizales
protegida
Selva
Den
sida
d (in
divi
duos
/m2 )
Sitio: I II III IV V VI VII VIII IX X XINo. de taxones: 35 29 11 20 26 21 22 26 32 18
= Otros invertebrados 123123123 = Lombrices
121212 = Hormigas
1234123412341234 = Termitas
Cultivos dealtos insumos
quemada y pastoreada
carga animal baja 16 mcarga animal
6 m
15 d
1 año moderada
altaB.
dec.
B. dec. +kudzú
arrozyuca
12345123451234512345
1234512345
123123
12345123451234512345
1234512345123451234512345
123123
12345123451234512345
12345123451234 1234
1234123412341234
12341234
123
123412341234123412341234123412341234
12341234
123123
1234512345123451234512345
123451234512345
123123
12341234123
123412341234123412341234123412341234123412341234123412341234
123412341234
123
123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234
1234123412341234
123
123451234512345
123412341234
Sabana Pastizales
protegida
Selva Monocultivosanuales
quemada y pastoreada
carga animal baja 16 mcarga animal
6 m15 d
1 año
moderadaalta
B.dec.
B. dec. +kudzú
arroz yuca
80
60
40
20
0I II III IV V VI VII VIII IX X XI
= Otros invertebradoss 123123123 = Lombrices
121212 = Hormigas
123123123123
= Termitas
Bio
mas
a (g
/m2 )
Sitios
la biomasa fueron las lombrices(35%), seguido de las termitas (18%) yde los miriápodos (11%). Otrosartrópodos, principalmente lospertenecientes a los miembros de
Gryllotalpinae, Cicadidae, Blattodeae Isopoda, constituían el 19% de labiomasa (Cuadro 2-2, Figuras 2-4 y2-5).
Figura 2-4. Distribución de la biomasa de la macrofauna en los 11 sitios estudiados. (d = días;m = meses; B. dec = Brachiaria decumbens.)
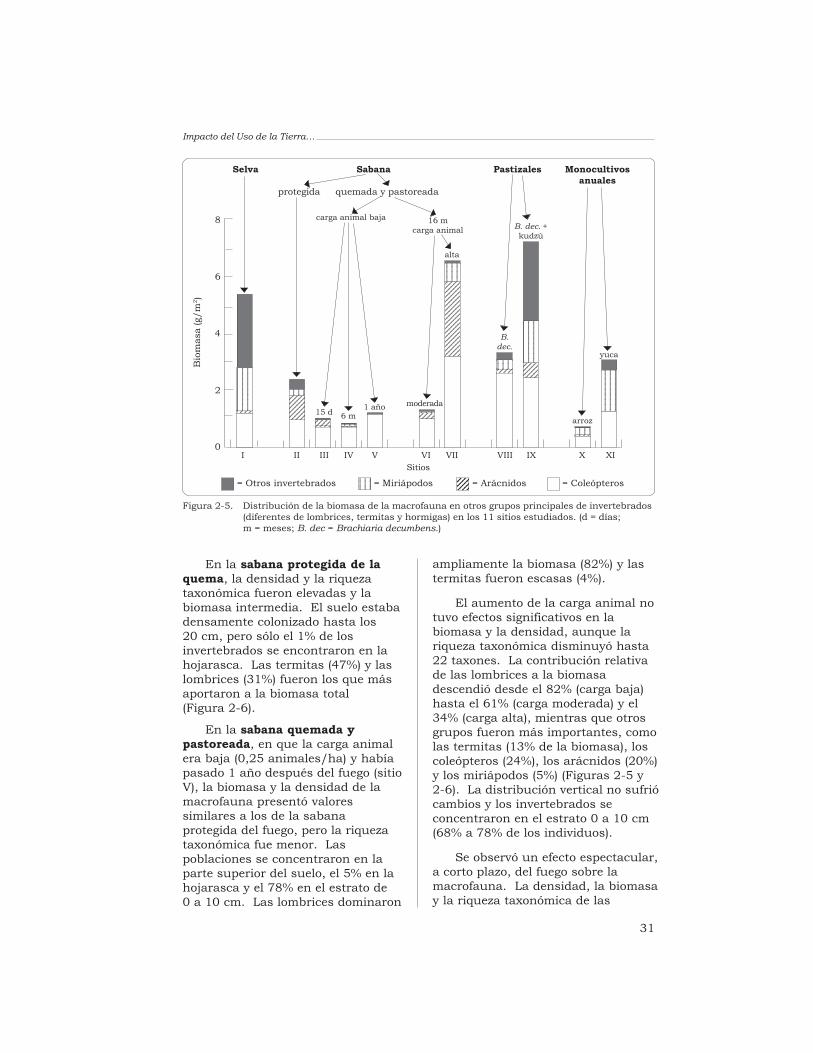
31
Impacto del Uso de la Tierra…
Figura 2-5. Distribución de la biomasa de la macrofauna en otros grupos principales de invertebrados(diferentes de lombrices, termitas y hormigas) en los 11 sitios estudiados. (d = días;m = meses; B. dec = Brachiaria decumbens.)
8
6
4
2
0
Bio
mas
a (g
/m2 )
I II III IV V VI VII VIII IX X XI
12345
1234123412341234123412341234123412341234
12345123451234512345123451234512345
12341234
123412341234 1234
12341234
123412341234 12345
1234512345
1234
123412341234123412341234123412341234123412341234123412341234123412341234
1234123412341234
12341234
123412341234
1234123412341234
123412341234123412341234123412341234
1234512345
12341234
123412341234123412341234123412341234
Sabana Pastizales
protegida
Selva Monocultivosanuales
quemada y pastoreada
carga animal baja 16 mcarga animal
6 m15 d 1 año moderada
alta
B.dec.
B. dec. +kudzú
arroz
yuca
= Otros invertebrados = Miriápodos 123123123123
= Arácnidos = Coleópteros
Sitios123123123
En la sabana protegida de laquema, la densidad y la riquezataxonómica fueron elevadas y labiomasa intermedia. El suelo estabadensamente colonizado hasta los20 cm, pero sólo el 1% de losinvertebrados se encontraron en lahojarasca. Las termitas (47%) y laslombrices (31%) fueron los que másaportaron a la biomasa total(Figura 2-6).
En la sabana quemada ypastoreada, en que la carga animalera baja (0,25 animales/ha) y habíapasado 1 año después del fuego (sitioV), la biomasa y la densidad de lamacrofauna presentó valoressimilares a los de la sabanaprotegida del fuego, pero la riquezataxonómica fue menor. Laspoblaciones se concentraron en laparte superior del suelo, el 5% en lahojarasca y el 78% en el estrato de0 a 10 cm. Las lombrices dominaron
ampliamente la biomasa (82%) y lastermitas fueron escasas (4%).
El aumento de la carga animal notuvo efectos significativos en labiomasa y la densidad, aunque lariqueza taxonómica disminuyó hasta22 taxones. La contribución relativade las lombrices a la biomasadescendió desde el 82% (carga baja)hasta el 61% (carga moderada) y el34% (carga alta), mientras que otrosgrupos fueron más importantes, comolas termitas (13% de la biomasa), loscoleópteros (24%), los arácnidos (20%)y los miriápodos (5%) (Figuras 2-5 y2-6). La distribución vertical no sufriócambios y los invertebrados seconcentraron en el estrato 0 a 10 cm(68% a 78% de los individuos).
Se observó un efecto espectacular,a corto plazo, del fuego sobre lamacrofauna. La densidad, la biomasay la riqueza taxonómica de las
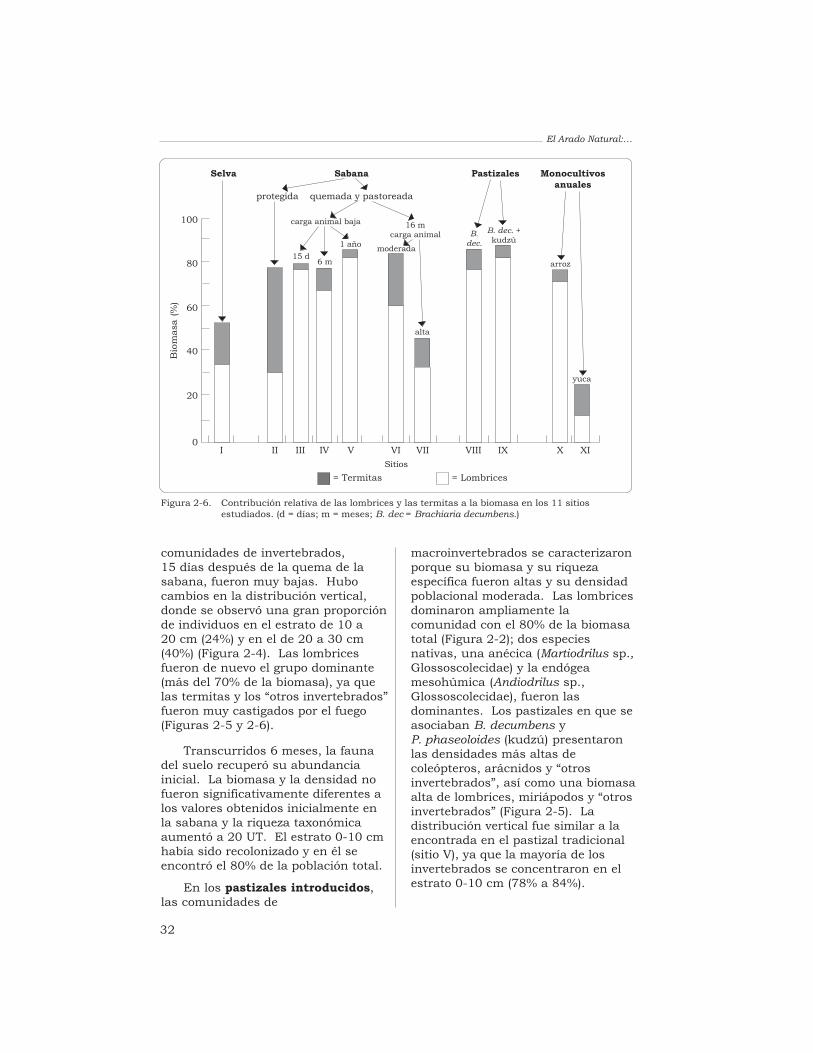
32
El Arado Natural:…
comunidades de invertebrados,15 días después de la quema de lasabana, fueron muy bajas. Hubocambios en la distribución vertical,donde se observó una gran proporciónde individuos en el estrato de 10 a20 cm (24%) y en el de 20 a 30 cm(40%) (Figura 2-4). Las lombricesfueron de nuevo el grupo dominante(más del 70% de la biomasa), ya quelas termitas y los “otros invertebrados”fueron muy castigados por el fuego(Figuras 2-5 y 2-6).
Transcurridos 6 meses, la faunadel suelo recuperó su abundanciainicial. La biomasa y la densidad nofueron significativamente diferentes alos valores obtenidos inicialmente enla sabana y la riqueza taxonómicaaumentó a 20 UT. El estrato 0-10 cmhabía sido recolonizado y en él seencontró el 80% de la población total.
En los pastizales introducidos,las comunidades de
macroinvertebrados se caracterizaronporque su biomasa y su riquezaespecífica fueron altas y su densidadpoblacional moderada. Las lombricesdominaron ampliamente lacomunidad con el 80% de la biomasatotal (Figura 2-2); dos especiesnativas, una anécica (Martiodrilus sp.,Glossoscolecidae) y la endógeamesohúmica (Andiodrilus sp.,Glossoscolecidae), fueron lasdominantes. Los pastizales en que seasociaban B. decumbens yP. phaseoloides (kudzú) presentaronlas densidades más altas decoleópteros, arácnidos y “otrosinvertebrados”, así como una biomasaalta de lombrices, miriápodos y “otrosinvertebrados” (Figura 2-5). Ladistribución vertical fue similar a laencontrada en el pastizal tradicional(sitio V), ya que la mayoría de losinvertebrados se concentraron en elestrato 0-10 cm (78% a 84%).
Figura 2-6. Contribución relativa de las lombrices y las termitas a la biomasa en los 11 sitiosestudiados. (d = días; m = meses; B. dec = Brachiaria decumbens.)
1234123412341234123412341234123412341234123412341234123412341234123412341234
1234512345123451234512345123451234512345123451234512345123451234512345123451234512345
12345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
1234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
12345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
1234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
1234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234123412341234
123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
12345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345123451234512345
1234123412341234123412341234
= Termitas = Lombrices
Sabana Pastizales
protegida
Selva Monocultivosanuales
quemada y pastoreada
carga animal baja 16 mcarga animal
6 m15 d
1 año moderada
alta
B.dec.
B. dec. +kudzú
arroz
yuca
I II III IV V VI VII VIII IX X XI
100
80
60
40
20
0
Bio
mas
a (%
)
Sitios

33
Impacto del Uso de la Tierra…
Los monocultivos anuales(arroz y yuca) de altos insumospresentaron los valores más bajos dedensidad, biomasa y riquezataxonómica. En el cultivo de arroz,las lombrices fueron las másabundantes en términos de biomasa(48%), y las termitas sólorepresentaron el 37% de ésta(Figura 2-6). La macrofauna seconcentró claramente en el estrato0-10 cm (87%).
En Granja MatazulLa comunidad de
macroinvertebrados de Matazul tuvola misma respuesta general a laintensificación agrícola que la deCarimagua: los pastizalespresentaron un aumento notable dela biomasa, mientras que en loscultivos anuales se redujeron tantola densidad como la biomasa de laspoblaciones de macroinvertebrados(Figuras 2-7 y 2-8). Sin embargo, yde modo general, las diferencias debiomasa y de densidad observadas
no son significativas según losanálisis estadísticos. Las lombricesson, también, menos abundantesque en Carimagua, especialmente enla sabana natural pastoreada y enlos pastizales introducidos.
La riqueza taxonómica (númerode familias de invertebrados) de lasabana nativa de Carimagua seredujo, en general, ligeramente en lospastizales extensivos e intensivos(Cuadro 2-4). Sin embargo, sólo elnúmero de familias ha disminuido(entre 3 y 8), mientras que el númerode morfotipos se mantuvo,generalmente, en todos los grupos deinvertebrados (3 a 13 morfotipos). Elpaso del fuego sobre la sabanadisminuye el número de UT (28morfotipos y 11 familias), aunque lariqueza inicial se recuperaparcialmente después de 6 meses(Cuadro 2-4).
La incorporación de cultivosanuales (monocultivo de arroz) en lasabana nativa entraña unadisminución de la riqueza
Figura 2-7. Biomasa de los macroinvertebrados del suelo en las cinco parcelas muestreadas deMatazul (Decaëns et al., datos sin publicar).
= Oligochaeta = Isoptera = Formicidae 123123 = Coleoptera
1234123412341234
= Araneida 1234123412341234
= Otros
123456
12345678123456781234567812345678123456781234567812345678123456781234567812345678
123456123456123456123456123456123456123456
1234567812345678
123456123456123456123456
12345678
123456123456123456123456123456123456
123456781234567812345678
1234561234567812345678
Pastizales intensivos Monocultivode arroz
B. humidicola
2 años
B. humidicola +A. pintoi
Sabananativa
2 años6 años
14
12
10
8
6
4
2
0
Bio
mas
a (g
/m2 )
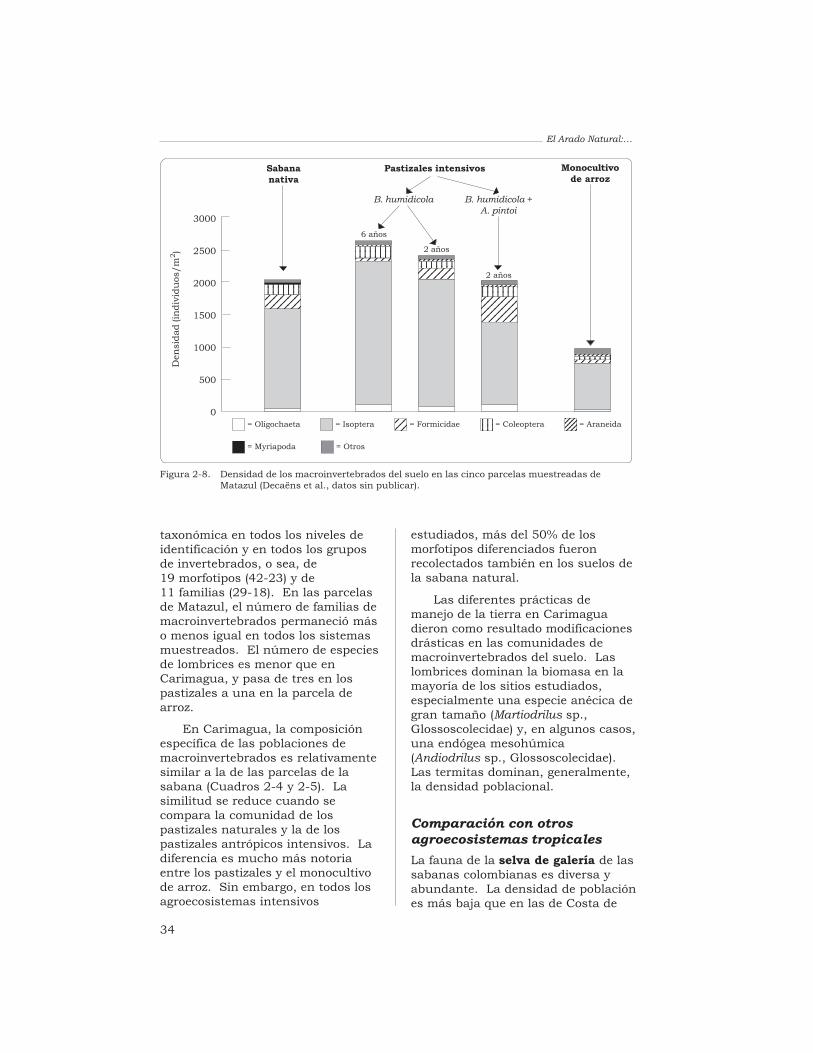
34
El Arado Natural:…
3000
2500
2000
1500
1000
500
0
Figura 2-8. Densidad de los macroinvertebrados del suelo en las cinco parcelas muestreadas deMatazul (Decaëns et al., datos sin publicar).
= Oligochaeta = Isoptera 121212 = Formicidae
123123123123
= Coleoptera 1234123412341234 = Araneida
= Myriapoda = Otros
Den
sida
d (in
divi
duos
/m2 )
123456789
123456123456123456123456
123456781234567812345678
123456781234567812345678123456123456
123456789
123456781234567812345678123456123456
123456781234567812345678123456123456
1234567812345678123456123456789123456789
Pastizales intensivos Monocultivode arroz
B. humidicola
2 años
B. humidicola +A. pintoi
Sabananativa
2 años
6 años123456789
taxonómica en todos los niveles deidentificación y en todos los gruposde invertebrados, o sea, de19 morfotipos (42-23) y de11 familias (29-18). En las parcelasde Matazul, el número de familias demacroinvertebrados permaneció máso menos igual en todos los sistemasmuestreados. El número de especiesde lombrices es menor que enCarimagua, y pasa de tres en lospastizales a una en la parcela dearroz.
En Carimagua, la composiciónespecífica de las poblaciones demacroinvertebrados es relativamentesimilar a la de las parcelas de lasabana (Cuadros 2-4 y 2-5). Lasimilitud se reduce cuando secompara la comunidad de lospastizales naturales y la de lospastizales antrópicos intensivos. Ladiferencia es mucho más notoriaentre los pastizales y el monocultivode arroz. Sin embargo, en todos losagroecosistemas intensivos
estudiados, más del 50% de losmorfotipos diferenciados fueronrecolectados también en los suelos dela sabana natural.
Las diferentes prácticas demanejo de la tierra en Carimaguadieron como resultado modificacionesdrásticas en las comunidades demacroinvertebrados del suelo. Laslombrices dominan la biomasa en lamayoría de los sitios estudiados,especialmente una especie anécica degran tamaño (Martiodrilus sp.,Glossoscolecidae) y, en algunos casos,una endógea mesohúmica(Andiodrilus sp., Glossoscolecidae).Las termitas dominan, generalmente,la densidad poblacional.
Comparación con otrosagroecosistemas tropicalesLa fauna de la selva de galería de lassabanas colombianas es diversa yabundante. La densidad de poblaciónes más baja que en las de Costa de
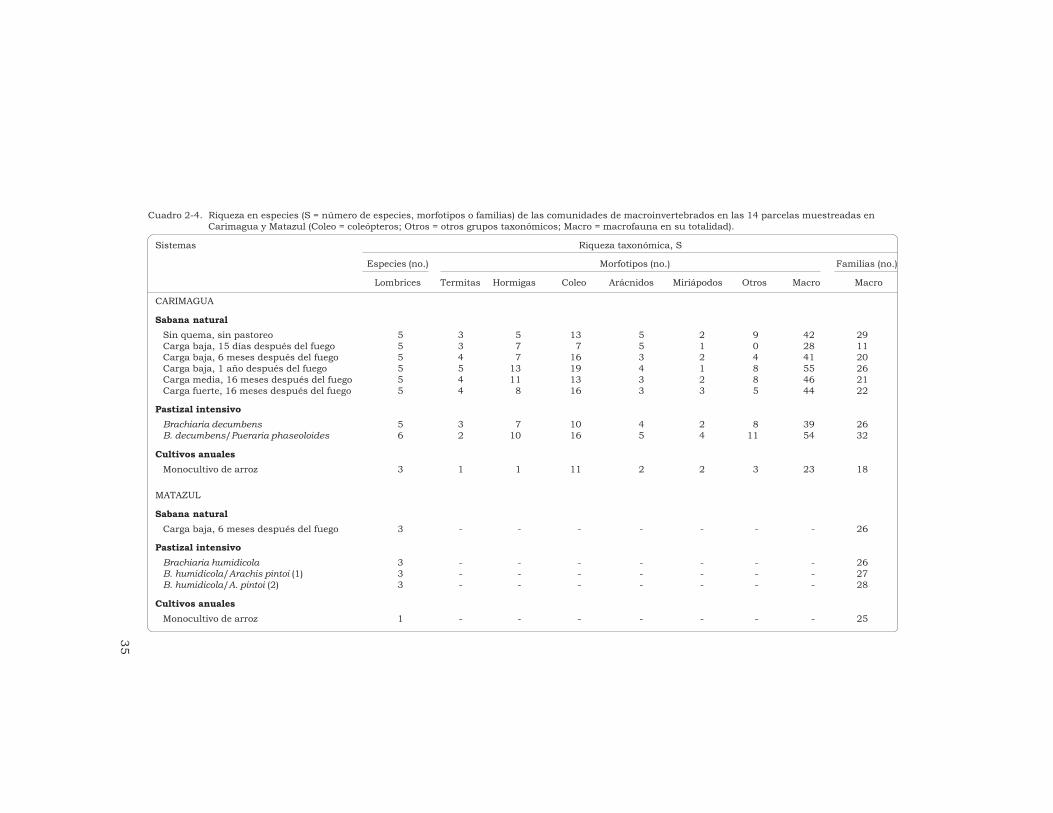
35
Impacto del U
so de la Tierra…
Cuadro 2-4. Riqueza en especies (S = número de especies, morfotipos o familias) de las comunidades de macroinvertebrados en las 14 parcelas muestreadas enCarimagua y Matazul (Coleo = coleópteros; Otros = otros grupos taxonómicos; Macro = macrofauna en su totalidad).
Sistemas Riqueza taxonómica, S
Especies (no.) Morfotipos (no.) Familias (no.)
Lombrices Termitas Hormigas Coleo Arácnidos Miriápodos Otros Macro Macro
CARIMAGUA
Sabana naturalSin quema, sin pastoreo 5 3 5 13 5 2 9 42 29Carga baja, 15 días después del fuego 5 3 7 7 5 1 0 28 11Carga baja, 6 meses después del fuego 5 4 7 16 3 2 4 41 20Carga baja, 1 año después del fuego 5 5 13 19 4 1 8 55 26Carga media, 16 meses después del fuego 5 4 11 13 3 2 8 46 21Carga fuerte, 16 meses después del fuego 5 4 8 16 3 3 5 44 22
Pastizal intensivoBrachiaria decumbens 5 3 7 10 4 2 8 39 26B. decumbens/Pueraria phaseoloides 6 2 10 16 5 4 11 54 32
Cultivos anualesMonocultivo de arroz 3 1 1 11 2 2 3 23 18
MATAZUL
Sabana naturalCarga baja, 6 meses después del fuego 3 - - - - - - - 26
Pastizal intensivoBrachiaria humidicola 3 - - - - - - - 26B. humidicola/Arachis pintoi (1) 3 - - - - - - - 27B. humidicola/A. pintoi (2) 3 - - - - - - - 28
Cultivos anualesMonocultivo de arroz 1 - - - - - - - 25

36
El Arado Natural:…
Cuadro 2-5. Indices de similitud de Sørensen calculados para nueve parcelas muestreadas enCarimagua y porcentaje de especies nativas observadas en los agroecosistemas intensivos(especie nativa = especie ya observada en el área de la sabana nativa; SN = sabana nativaprotegida del fuego; SP = sabana nativa pastoreada; 15d = 15 días después del fuego;6m = 6 meses después del fuego; 1a = 1 año después del fuego; CM = carga animalmedia; CA = carga animal alta; Bd = pastizal de Brachiaria decumbens; BdPp = pastizal deB. decumbens asociada con Pueraria phaseoloides; MR = monocultivo de arroz).
Prácticas de uso de la tierra SN SP15d SP6m SP1a CM CA Bd BdPp MRcomparadas
SN - 0.39 0.49 0.42 0.48 0.43 0.37 0.36 0.22
SP15d - - 0.42 0.40 0.22 0.41 0.30 0.25 0.25
SP6m - - - 0.43 0.47 0.46 0.35 0.32 0.29
SP1a - - - - 0.44 0.39 0.34 0.28 0.29
CM - - - - - 0.54 0.28 0.30 0.29
CA - - - - - - 0.31 0.33 0.31
Bd - - - - - - - 0.47 0.32
BdPp - - - - - - - - 0.21
Morfotipos de la sabana (%) 100 100 100 100 100 100 55 54 73
Marfil, comparable con la obtenidaen la Amazonia peruana y en losbosques de hayas de Europa, ysuperior a la de los bosquestropicales de México, Nigeria ySarawak. La biomasa es similar a laencontrada en los hayedos europeosy en algunos bosques de México yNigeria, de 2 a 6 veces superior a lade Sarawak, pero bastante inferior alos valores obtenidos en Perú y enCosta de Marfil (Cuadro 2-6 y susreferencias). Esto se debe a labiomasa de las lombrices, que es3 veces inferior al valor medio(12.9 g/m2) calculado por Fragoso yLavelle (1992) en 12 bosques deAmérica del Centro y del Sur, enÁfrica y en Asia. Los miriápodosrepresentan el 11% de la biomasa,como ocurre en Perú y México(Lavelle y Kohlman 1984; Lavelle yPashanasi 1989) y otros grupos,como Gryllotalpinae, Cicadidae,Isopoda o Blattodea, tienen relativaimportancia en Carimagua(Figura 2-5).
En la sabana protegida de laquema, la densidad y la biomasa dela macrofauna son
significativamente más bajas que enlas sabanas africanas de Costa deMarfil. La biomasa de las lombriceses de 3 a 10 veces más baja que enlas sabanas africanas (Cuadro 2-6) yrepresenta solamente el 31.2% de labiomasa total; las termitas fueron, encambio, el mayor componente de labiomasa (46.6%).
La práctica de manejotradicional no afecta ni a la biomasani a la densidad de la macrofauna,aunque reduce ligeramente lariqueza taxonómica. La biomasa esmucho más baja que en lospastizales tradicionales de Perú,mientras que la densidad depoblación es bastante similar(Cuadro 2-6). Este manejo afectamucho a las termitas; por ello, sucontribución a la biomasa se redujoa 3.9% de 46.6%, el valor que teníaen la sabana protegida del fuego.Las lombrices, en cambio, resultanfavorecidas por este tipo de manejo,ya que su biomasa es tres vecessuperior y representa el 82% de labiomasa total (Figura 2-4). Estosresultados confirman lo que seobservó anteriormente en América

37
Impacto del U
so de la Tierra…
Cuadro 2-6. Comparación de la densidad (individuos/m2) y biomasa (g/m2) de la macrofauna del suelo y de las lombrices de tierra en varios lugares estudiados.
Localidad Tipo de vegetación Macrofauna Lombrices Referencia
Biomasa Densidad Biomasa Densidad
Colombia Selva de galería 13.6 4294 4.7 251 Este estudioCosta de Marfil Selva tropical 74.2 5747 52.3 171.2 Gilot et al. 1995Nigeria Selva tropical 16.4 3119 10.2 34 Madge 1969México Selva tropical 16.4-18.9 888-3011 9.8-10.7 8-132 Lavelle y Kohlmann 1984;
Lavelle et al. 1981
Perú Selva tropical 24.1-53.9 4099-4303 11.9-28.2 85-120 Lavelle y Pashanasi 1989Sarawak Selva tropical 2.4-6.8 663-2579 0.4-1 24-42 Collins 1980Alemania Bosque de hayas 12.7 4035 10.7 205 Schaefer 1990
Colombia Sabana 15.3 1830 4.8 157 Este estudioCosta de Marfil Sabana 29.7-84.8 2015-10,905 17-48.6 188-400 Lavelle 1983;
Lavelle et al. 1992
Colombia Pastizal tradicional 8-16.8 698-2029 4.5-13.8 32-192 Este estudioPerú Pastizal tradicional 82.3-121.2 1768-2347 78-116.4 474-573 Lavelle y Pashanasi 1989
Colombia Pastizal mejorado 28.8-62.5 1541-2267 22.9-51.1 139-213 Este estudioPerú Pastizal mejorado 110.9-159.2 922-1546 103.2-153 546-740 Lavelle y Pashanasi 1989México Pastizal mejorado _ _ 35.8-55.5 620-948 Lavelle et al. 1981India Pastizal mejorado _ _ 30.2-56 17.4-800 Dash y Patra 1977;
Sepanati 1980

38
El Arado Natural:…
del Norte y en Costa de Marfil (James1988; Lavelle 1983), donde elpastoreo incrementa las poblacionesde lombrices.
La quema de la sabana destruye,al parecer, gran parte de lamacrofauna del suelo. La densidad,la biomasa y la riqueza taxonómicade la comunidad disminuyeronclaramente; las termitas parecen serel grupo más sensible (Figura 2-6).El fuego también afectó ladistribución vertical de lamacrofauna, ya que 15 días despuésde la quema sólo se encontró el 35%de los invertebrados en el estrato de0 a 10 cm (Figura 2-2). Los nivelesde macrofauna se recuperaronrápidamente y 6 meses después delfuego habían recobrado sus valoresiniciales de biomasa, densidad ydiversidad taxonómica; el estrato0-10 cm había sido recolonizado.Estas observaciones contradicen losresultados obtenidos por Athias et al.(1975) en Costa de Marfil, los cualesindican que habrá un efectoretrasado de la quema, siempre queno se observe un efecto inmediato.Es posible que la diferencia entreambos casos pueda provenir deltiempo en que se hace la quema. EnCosta de Marfil se ha observado quelas quemas realizadas durante laépoca seca, cuando la mayor partede la fauna del suelo está adaptadaal estrés de sequía, no afectan a lastermitas porque éstas viven a granprofundidad en el suelo o entermiteros de paredes muy gruesas(Josens 1972). En Carimagua noexiste, al parecer, preadaptación alestrés provocado por la quema.
El sobrepastoreo no afecta ni labiomasa ni la densidad poblacional,aunque reduce la riquezataxonómica. El aumento de la cargaanimal reduce la importancia relativade las lombrices, que aportan sólo el33.4% de la biomasa en lospastizales más pastoreados
(Figura 2-6). Esto puede deberse ala modificación que experimenta elmicroclima del suelo por ladisminución de la vegetaciónherbácea. Los coleópteros sonabundantes en estos pastizales(Figura 2-5) (7.1% y 24% de labiomasa en el pastizal con cargaanimal media y alta,respectivamente), pero son menosimportantes que en los pastizales deMéxico (de 25 a 48 g/m2) (Villalobosy Lavelle 1990). Una mayorincorporación de deposiciones delganado en el suelo puede explicaresta diferencia.
Las comunidades de macrofaunade los pastizales mejoradospresentan una densidad poblacionalmoderadamente abundante; subiomasa y su riqueza en especies sonaltas, aunque la biomasa es inferiora la reportada en otros pastizalestropicales. Las especies nativas de lasabana son el mayor componente deesta biomasa (80%) y tienen valoresabsolutos de 22.9 a 51.1 g/m2, unvalor equivalente al de otrospastizales de México o de la India,pero inferior al hallado en Perú(Cuadro 2-6). Este aumento debiomasa se debe, en realidad, a lamejor calidad de los desechos delsuelo, que están conformados porestiércol del ganado y por el materialaéreo y radical de la leguminosa. Laadición de hojas de leguminosa,ricas en nitrógeno, a la hojarasca delpastizal de B. decumbens + kudzú,mantiene la riqueza taxonómica eincrementa la biomasa hasta unvalor cuatro veces superior alobtenido en la sabana protegida dela quema.
Al revés de lo que ocurre en lospastizales establecidos donde huboantes selva tropical, las comunidadesde macrofauna parecen haber sidofavorecidas por el mejoramiento delas condiciones tróficas; sinembargo, no hay proliferación de

39
Impacto del Uso de la Tierra…
lombrices alóctonas, como ocurre enmuchos otros pastizales tropicales(Lavelle et al. 1981; Lavelle yPashanasi 1989). Otros grupos deimportancia son los isópodos, queindican la presencia de unmicroclima húmedo en el suelo, y loscoleópteros, a los que ciertamentefavorecen el estiércol del ganado y labiomasa de las raíces (Figura 2-5).
Los monocultivos de altosinsumos ocasionan un descensodrástico de la riqueza taxonómica, dela densidad y de la biomasapoblacional. Estos resultadosconfirman las observacionesrealizadas en Perú (Cuadro 2-6) ypueden ser explicados por la acciónnegativa del laboreo del suelo, por lafertilización (especialmente cuandoinduce una mayor acidificación en elsuelo), por la modificación delmicroclima del suelo, por la acciónde pesticidas no específicos y por lareducción de las reservas orgánicasde carbono disponibles en el suelo acausa de la destrucción de lavegetación perenne (House yParmelee 1985; Reddy y Goud 1987).
Los resultados obtenidos en lafinca de Matazul confirman lasobservaciones realizadaspreviamente en Carimagua. Enefecto, se observan allí las mismastendencias en el efecto positivo delos pastizales y en el negativo de loscultivos anuales, las pequeñas onulas diferencias entre las prácticasaplicadas en los pastizales, y laausencia de efectos en la riqueza enespecies de las comunidades deinvertebrados. La biomasa demacroinvertebrados obtenida en losLlanos Orientales es, en general,baja en comparación con los valoresmedios de biomasas calculados porLavelle et al. (1994). Las causas deestos resultados están ligadas, enparte, a ciertas propiedades físicasde los suelos (acidez, saturación dealuminio) o a la presencia de una
época seca muy definida que reducea 8 meses el período de actividad delos invertebrados.
La comparación de los datosobtenidos en Carimagua y enMatazul pone en evidencia la granvariabilidad, entre un sitio y otro, delas comunidades demacroinvertebrados. Se puede decir,en general, y para tipos de uso delsuelo similares en ambos sitios, quela biomasa de la macrofauna es másbaja en Matazul. Las especies delombrices de tierra no son lasmismas en los dos sitios (Jiménez,Decaëns, Feijoo, datos nopublicados) y la densidad es muchomás reducida en Matazul que enCarimagua. Estos resultadospueden explicarse por la cantidad deMO presente en los primeros 20 cmdel suelo de Matazul (Fisher et al.1994) o porque la tierra de Matazulse dedicó a fines agrícolas muchoantes que la de Carimagua.
La composición taxonómica de lacomunidad de macroinvertebradosde los pastizales intensivos deCarimagua difiere sensiblemente dela obtenida en la sabana. Sinembargo, la mayoría de losmorfotipos encontrados pertenecen ataxones que también se encontraronen la sabana, y son las especiesautóctonas las que parecenaprovechar el mejoramiento de lascondiciones tróficas. En el caso delas lombrices, por ejemplo, no hubouna proliferación de especiesalóctonas de distribuciónpantropical, como suele observarseen otros pastizales establecidos enregiones selváticas tropicales (Barroset al. 1998; Lavelle et al. 1981;Lavelle y Pashanasi 1989). Lariqueza en especies se ha mantenidoen la mayor parte de los grupos, aexcepción de las termitas y loscoleópteros. Ciertos grupos, comolas lombrices, muestran granadaptabilidad al mejoramiento de las

40
El Arado Natural:…
condiciones ambientales de lospastizales, estableciendo grandespoblaciones que dominan la biomasade la comunidad de invertebrados.
ConclusionesLas comunidades demacroinvertebrados de las sabanasbien drenadas de Colombia son muysensibles a los cambios ambientalesderivados de la intensificaciónagrícola. Se pueden distinguir trestipos de agroecosistemas que tienenefectos diferentes sobre lamacrofauna del suelo:
1. Pastizales naturalesextensivos, cuyos efectos en lamacrofauna son poco significativos ydonde la recolonización del suelodespués de la quema ocurre en untiempo relativamente corto. Laslombrices se benefician del pastoreoy del fuego, aunque su importanciadisminuye si hay sobrepastoreo. Lastermitas responden en formaabiertamente opuesta, de modo quela relación lombrices/termitas puedeconsiderarse un indicador sensibledel estado del ambiente.
2. Pastizales introducidos, quefavorecen a los principalescomponentes de los grupos demacroinvertebrados. La siembra deespecies forrajeras introducidas y elincremento de la producción animalen pastos mejorados hacen un fuerteimpacto en la macrofauna del suelo,especialmente en las poblaciones delombrices, cuya biomasa aumenta de4.8 a 51.1 g/m2. La asociaciónB. decumbens + kudzú es, al parecer,de alto valor respecto almantenimiento y al mejoramiento dela calidad del suelo y de labiodiversidad, ya que tambiénmantiene la riqueza taxonómica de lasabana nativa. Estos resultados seexplican por el mejoramiento de lacalidad de la hojarasca y por la gran
cantidad de deposiciones del ganadoque recibe el suelo. Por tanto, elestablecimiento de pastizalesmejorados sobre la sabana nativa notransforma el medio en un “desiertoverde” sino que, por el contrario,aumenta la actividad de lascomunidades locales de macrofauna.
3. Cultivos anuales, queafectan fuertemente las poblacionesde lombrices y de artrópodos,causando una reducciónespectacular de su biomasa, sudensidad y su riqueza taxonómica.Las prácticas agrícolas, como ellaboreo, la fertilización y laaplicación de herbicidas, así como lareducción de la biomasa radical y lamodificación de las condicionesmicroclimáticas tras la desapariciónde la cubierta vegetal, son losfactores que explican el anteriorfenómeno. La pérdida conjunta de laabundancia y la diversidad de dichascomunidades en los cultivos anualesconduce, invariablemente, a unapérdida de ciertas funcionesimportantes del suelo (Giller et al.1997; Lavelle 1996). En efecto, lamayor parte de los organismos delsuelo intervienen, en distinto grado,en la dinámica de la MO del suelo yen la regulación de la estructura deéste, las cuales determinan lafertilidad del suelo y la sostenibilidadde la producción vegetal (Lal 1988;Lavelle 1997; Lee y Foster 1991).
El estudio de alternativassostenibles para la producciónagrícola extensiva puede articularsecon el estudio de las comunidades deinvertebrados como indicadoressencillos de la estabilidad de losagroecosistemas. Para identificarsistemas alternativos que conservene incrementen la macrofauna, puedeayudar la comprensión de larespuesta de la macrofauna a lasperturbaciones inducidas por elhombre. El estudio de la dinámicaespacio-temporal de los

41
Impacto del Uso de la Tierra…
invertebrados “clave” (los ingenierosdel ecosistema) es de importanciacapital. Es necesario prestaratención a los fenómenos derecolonización que se presentan enlas poblaciones de macrofauna unavez ocurrida la perturbación, asícomo a la posible migración deespecies de un sistema favorable aotro desfavorable.
Los resultados obtenidos en esteestudio permiten identificar variasalternativas posibles para conservaro estimular las actividades de lamacrofauna del suelo:
- El efecto negativo de los cultivosanuales, por ejemplo, podría seratenuado si se reducen laintensidad y la frecuencia de laperturbación (disminución de lalabranza y de la aplicación depesticidas), o si se incrementanla cantidad y la calidad delrecurso energético utilizado porlos macroinvertebrados (p.ej.,cultivar leguminosas comocobertura del suelo, mantener losresiduos que dejan los cultivos).
- Los sistemas que integran fasescortas de cultivo en alternanciacon largos períodos de pastizal(3 a 5 años) parecen tambiénuna buena opción para mantenerlas poblaciones demacroinvertebrados y para traerotros beneficios a laspropiedades físicas y químicasdel suelo (Thomas et al. 1995).
- Con nuevos trabajos deinvestigación podrándeterminarse, en forma másprecisa, los efectos negativos opositivos de diferentes períodosde cultivo o de pastizal,respectivamente; estos efectosinfluyen en el tiempo requeridopara que una población efectivade macroinvertebrados serestablezca.
AgradecimientosAgradecemos a los trabajadores delPrograma de Sabana Nativa (CIAT/IEMVT) y de otros programas de laestación de Carimagua por la ayudaprestada en el trabajo de campo.Agradecemos a J.P. Rossi (ORSTOM)por los análisis estadísticosrealizados. Queremos dar las graciastambién a Rainer Herrmann por eltiempo que empleó en ayudarnos acompletar este trabajo después derealizado el suyo propio.
ReferenciasAnderson JM; Ingram J, eds. 1993.
Tropical soil biology and fertility: ahandbook of methods. 2a. ed.C.A.B., Oxford. 221p.
Athias F; Josens G; Lavelle P. 1975.Influence du feu de brousse annuelsur le peuplement endogé de lasavane de Lamto. En: Vanek J, ed.Progress in soil biology. Praga,República Checa. p 389-396.
Barros E; Grimaldi M; Desjardins T;Sarrazin M; Chauvel A; Lavelle P.1998. Conversion of forests intopastures in Amazonia: Effects onsoil macrofaunl diversity and soilwater dynamics. En: ISSS-AISS-IBG-SICS-AFES, eds. Proceedings ofthe 16th World Congress of SoilScience. Montpellier, Francia,agosto 1998.
Collins NM. 1980. The distribution of soilmacrofauna on the West Ridge ofGunung (Mount) Mulu, Sarawak.Oecologia 44:263-275.
Dash MC; Patra UC. 1977. Density,biomass and energy budget oftropical earthworm population froma grassland site in Orissa, India.Rev Ecol Biol Sol 14:461-471.

42
El Arado Natural:…
Fisher MJ; Lascano CE; Vera RR;Rippstein G. 1992. Integrating thenative savanna ressource withimproved pastures. En: CIAT,Pastures for tropical lowlands:CIAT’s contribution. Cali, Colombia.p 75-99.
Fisher MJ; Rao IM; Ayarza MJ; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage by introduceddeep-rooted grasses in the SouthAmerican savannas. Nature371:236-238.
Fragoso C; Lavelle P. 1992. Earthwormcommunities of tropical rain forests.Soil Biol Biochem 24:1397-1408.
Giller KE; Beare MH; Lavelle P; Izac A-MN; Swift MJ. 1997. Agriculturalintensification, soil biodiversity andagroecosystem function. Appl SoilEcol 6:3-16.
Gilot C; Lavelle P; Keli J; Kouassi P;Guillaume G. 1995. Biologicalactivity of soil under rubberplantation in Côte d’Ivoire. Acta ZoolFenn 196:186-189.
House GJ; Parmelee RW. 1985.Comparison of soil arthropods andearthworms from conventional andno-tillage agroecosystems. Soil TillRes 5:351-360.
James SW. 1988. The postfireenvironment and earthwormpopulations in tallgrass prairie.Ecology 69:476-483.
Jiménez JJ. 1999. Estructura de lascomunidades y dinámica de laspoblaciones de lombrices de tierraen sabanas naturales y perturbadasde Carimagua (Colombia). TesisDoctoral. Universidad Complutense,Madrid. 311 p.
Josens G. 1972. Etudes biologiques etécologiques des termites (Isoptera)de la savane de Lamto-Pakobo (Côted’Ivoire). Tesis de doctorado.Université Libre de Bruxelles.
Lal R. 1988. Effects of macrofauna on soilproperties in tropical systems. AgricEcosyst Environ 24:101-116.
Lavelle P. 1983. The soil fauna of tropicalsavannas; II: The earthworms. En:Bourlière F, eds. Tropical savannas.E.S.P.C., Amsterdam. p 485-504.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: Adaptive strategies thatdetermine ecosystem function.Adv Ecol Res 27:93-132.
Lavelle P; Kohlmann B. 1984. Etudequantitative de la macrofaune dusol dans une forêt tropicale duMexique (Bonampak, Chiapas).Pedobiologia 27:377-393.
Lavelle P; Maury ME; Serrano V. 1981.Estudio cuantitativo de la fauna delsuelo en la región de Laguna Verde,Veracruz: Época de lluvias. InstEcol Publ 6:75-105.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López D;Pashanasi B; Brussaard L. 1994.The relationship between soilmacrofauna and tropical soilfertility. En: Voomer PL; Swift MJ,eds. The biological management oftropical soil fertility. John Wiley &Sons, Chichester, Reino Unido.p 137-169.
Lee KE; Foster RC. 1991. Soil fauna andsoil structure. Aust J Soil Res29:745-775.
Madge DS. 1969. Field and laboratorystudies on the activities of twospecies of tropical earthworms.Pedobiologia 9:188-214.
Reddy MV; Goud AN. 1987. Impact ofinorganic fertilizers on theearthworm population density ofwetland rice (Oryza sativa)agroecosystems. Proc Nat Ecotoxic(1987), p 154-158.

43
Impacto del Uso de la Tierra…
Senapati BK. 1980. Aspects ofecophysiological studies on tropicalearthworms. Distribution,population dynamics, production,energetics, and their role indecomposing processes. Tesis.Sambalpur University, Sambalpur,India. 154 p.
Schaefer M. 1990. The soil fauna of abeech forest on limestone: trophicstructure and energy budget.Oecologia 82:128-136.
Spain AV; Lavelle P; Mariotti A. 1992.Stimulation of plant growth bytropical earthworms. Soil BiolBiochem 24:1629-1633.
Stork NE; Eggleton P. 1992. Invertebratesas determinants and indicators ofsoil quality. Am J Alt Agr 7:38-55.
Thioulouse J. 1989. Statistical analysisand graphical display ofmultivariate data on the Macintosh.Comp Appl Bio 5:287-292.
Thioulouse J. 1990. MacMul andGraphMu: two Macintosh programsfor the display and analysis ofmultivariate data. Comp Geosc16:1235-1240.
Thomas RJ; Fisher MJ; Ayarza MA; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA, eds.Soil management: experimentalbasis for sustainability andenvironmental quality. Advances inSoil Science Series, Lewis Pubs.,Boca Ratón, Estados Unidos.p 61-83.
Villalobos FJ; Lavelle P. 1990. The soilColeoptera community of a tropicalgrassland from Laguna Verde,Veracruz (Mexico). Rev Ecol Biol Sol27:73-93.
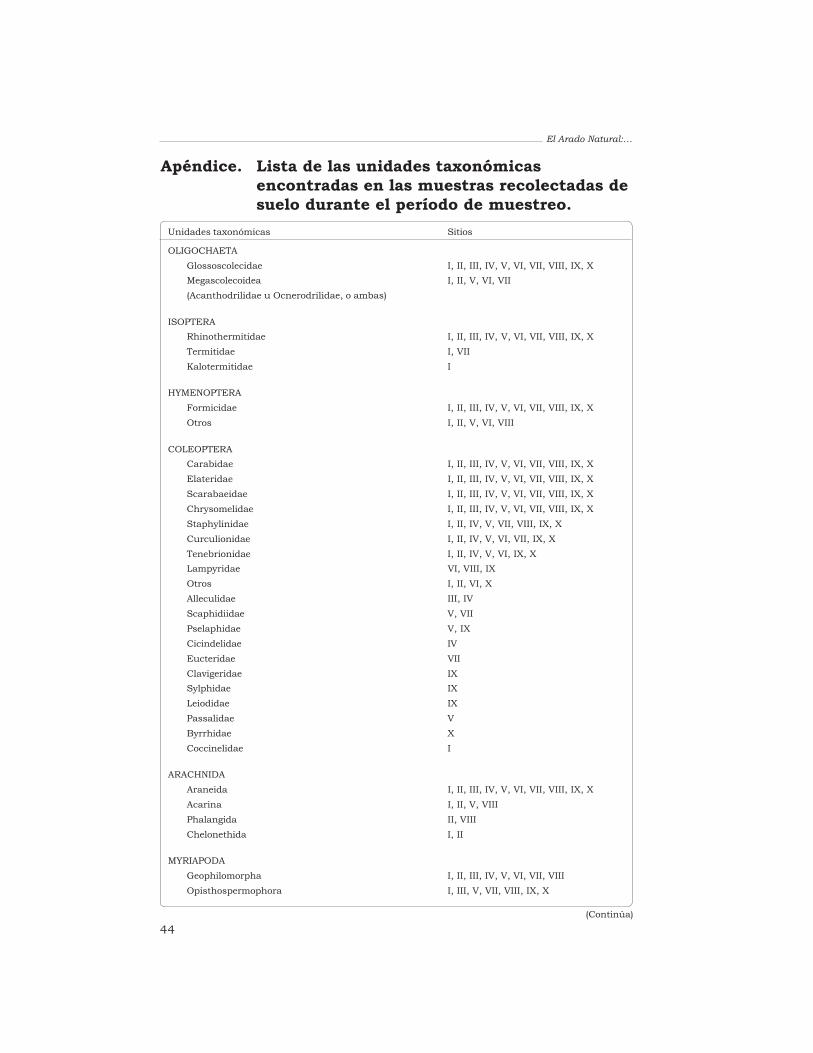
44
El Arado Natural:…
Apéndice. Lista de las unidades taxonómicasencontradas en las muestras recolectadas desuelo durante el período de muestreo.
Unidades taxonómicas Sitios
OLIGOCHAETAGlossoscolecidae I, II, III, IV, V, VI, VII, VIII, IX, XMegascolecoidea I, II, V, VI, VII(Acanthodrilidae u Ocnerodrilidae, o ambas)
ISOPTERARhinothermitidae I, II, III, IV, V, VI, VII, VIII, IX, XTermitidae I, VIIKalotermitidae I
HYMENOPTERAFormicidae I, II, III, IV, V, VI, VII, VIII, IX, XOtros I, II, V, VI, VIII
COLEOPTERACarabidae I, II, III, IV, V, VI, VII, VIII, IX, XElateridae I, II, III, IV, V, VI, VII, VIII, IX, XScarabaeidae I, II, III, IV, V, VI, VII, VIII, IX, XChrysomelidae I, II, III, IV, V, VI, VII, VIII, IX, XStaphylinidae I, II, IV, V, VII, VIII, IX, XCurculionidae I, II, IV, V, VI, VII, IX, XTenebrionidae I, II, IV, V, VI, IX, XLampyridae VI, VIII, IXOtros I, II, VI, XAlleculidae III, IVScaphidiidae V, VIIPselaphidae V, IXCicindelidae IVEucteridae VIIClavigeridae IXSylphidae IXLeiodidae IXPassalidae VByrrhidae XCoccinelidae I
ARACHNIDAAraneida I, II, III, IV, V, VI, VII, VIII, IX, XAcarina I, II, V, VIIIPhalangida II, VIIIChelonethida I, II
MYRIAPODAGeophilomorpha I, II, III, IV, V, VI, VII, VIIIOpisthospermophora I, III, V, VII, VIII, IX, X
(Continúa)

45
Impacto del Uso de la Tierra…
MYRIAPODAScolopendromorpha I, II, IV, V, VI, VIII, IXOtros V, VI, VIII, IX, XPolydesmida I, II, VIII, IX
NEMATODA I, II, IV, V, VI, VII, VIII, IX, X
HOMOPTERACercopidae VII, VIII, IXCicadidae I, IV, V, XCicadellidae II, IX
HEMIPTERAOtros I, II, IV, V, VI, VIICorimelaenidae VI, VIIReduvidae I, II, IXCydnidae II,
ORTHOPTERABlattidae I, II, V, VIII, IXGryllotalpinae IV, VIIGryllinae IXMantidae IX
LEPIDOPTERA I, IV, VII, IX
EMBIOPTERA I, V
CRUSTACEAIsopoda I, II, VIII, IX
THYSANURALepismatidae II
DIPTERA I, II, VI, VIII, IX, X
DERMAPTERA I
MOLLUSCAHelicidae I, VIII, IXLimacidae IX
DIPLURA I
Apéndice. (Continuación).
Unidades taxonómicas Sitios

46
El Arado Natural:…
ResumenDurante el período comprendidoentre abril y septiembre de 1994, secuantificaron la densidad demacrofauna, la biomasa total y ladiversidad de especies de laslombrices de tierra en diferentessistemas de uso de la tierra, en lacuenca del río Cabuyal. En estetrabajo se empleó la metodologíarecomendada en el Programa deBiología y Fertilidad de los SuelosTropicales (TSBF). La densidad demacrofauna, la biomasa y labiodiversidad de las lombrices varióconsiderablemente según el sistemaconsiderado y la profundidad del
suelo. El análisis factorial inicialpermitió extraer tres factores queexplicaron el 71.8% de la varianzatotal. Se observó una riquezataxonómica alta en la selva andina(98 unidades taxonómicas, UT), quedesciende abruptamente en lospastizales (va de 28 a 13 UT) yen las tierras cultivadas (de 31 a18 UT).
Palabras clave: Cabuyal, cuencahidrográfica, diversidad,lombrices, macrofauna del suelo,uso de la tierra
IntroducciónEn los ecosistemas tropicales,algunos procesos del suelo estándeterminados por los invertebrados.Algunos grupos clave, como laslombrices, las termitas, las hormigasy los artrópodos que se alimentan dehojarasca, afectan la estructurafísica del suelo e influyen en ladinámica de los nutrientes medianteel efecto que causan en lainmobilización y la humificación dela MO (Anderson y Flanagan 1989;Lavelle et al. 1992, 1994).
Sólo algunos estudios recienteshan sido enfocados hacia laimportancia de losmacroinvertebrados en las regionespaleotropical y neotropical, y hacia el
CAPÍTULO 3
Cuantificación de la Macrofauna delSuelo en una Cuenca Hidrográfica deColombia1,2
A. Feijoo*, E. B. Knapp*, P. Lavelle** yA. G. Moreno***
1. Publicado en Pedobiologia, vol. 43,p 513-517, por Feijoo et al. Copyright © 1999de Urban & Fischer Verlag. Impreso conpermiso de la casa editorial.
2. Aunque este trabajo no se relacionaespecíficamente con los Llanos Orientales, seha incluido en el libro con el fin de mostrarel impacto del uso de la tierra en la regiónandina. Eds.
* CIAT, Proyecto de Laderas, AA 6713, Cali,Colombia, y Universidad Nacional deColombia, AA 237, Palmira, Colombia.
** Laboratoire d’Ecologie et Biologie des SolsTropicaux, ORSTOM/Université Paris VI,32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.
*** Departamento de Biología Animal I(Zoología), Facultad de Biología, UniversidadComplutense, 28040 Madrid, España.

47
Cuantificación de la Macrofauna del Suelo...
impacto que causa en ellos unaperturbación ambiental restringida alos sistemas de uso de la tierra(Collins 1980; Decaëns et al. 1994;Lavelle y Kohlmann 1984; Lavelle yPashanasi 1989).
El programa de laderas del CIATha estado investigando en cuatrofrentes; en uno de ellos, conocidocomo ‘efectos de la degradación delsuelo’, tiene dos objetivos:
- desarrollar alternativas de manejode la tierra que mantengan labiodiversidad del suelo y quemejoren, al mismo tiempo, lafertilidad del suelo;
- emplear la macrofauna del suelocomo un indicador de laperturbación que éste reciba.
En este estudio se ha comparadola diversidad, la abundancia y labiomasa de la macrofauna del suelo,en áreas de ladera y bajo10 diferentes sistemas de uso.
Materiales y Métodos
Sitios del estudioEl estudio se realizó en la cuenca delrío Cabuyal, en los Andescolombianos. La cuenca ocupa unaextensión de 7000 ha y está situada a120 km de Cali (va de 76°33’ a76°30’ O y de 2°42’ a 2°52’ N). Lasmuestras se tomaron en las zonasintermedia (1450-1550 msnm) y alta(2000-2200 msnm) de la cuenca. Elestudio duró desde abril hastaseptiembre de 1994, durante la épocaseca, y se llevó a cabo en los 10 sitiossiguientes, que diferían según el tipode tierra y la frecuencia de uso deésta:
1. Selva secundaria (SS)
2. Selva de 40 años (S40)
3. Selva de más de 40 años (S40+)
4. Barbecho (B) (helechal conmalezas arbustivas)
5. Cultivo tradicional de café (CC)
6. Plantación de pino (PP)
7. Yuca asociada con frijol y maíz(YA)
8. Pastizal de kikuyo (Pennisetumclandestinum) (PK)
9. Pastizal de yaraguá (Melinisminutiflora) (PY)
10. Pastizal de Brachiaria humidicola(BH).
MétodosLas muestras se tomaron siguiendo lametodología del programa TSBF(Anderson e Ingram 1993). Losinvertebrados fueron agrupados enunidades taxonómicas (UT,Cuadro 3-1) y se cuantificó sudensidad y su biomasa. Las lombricesse conservaron en formalina al 5% y elresto de los invertebrados en alcoholal 70%. Las 23 UT procedentes de los10 sitios fueron sometidas a unanálisis de componentes principales(ACP, versión 609 de SAS). Tambiénse determinó el tipo de vegetación y eluso dado a la tierra.
ResultadosTres factores explicaron el 71.8% de lavarianza total de la macrofauna(Cuadro 3-1):
- El Factor I (43.6% de la varianza)se interpretó como el efecto de lavegetación densa y de la hojarascasobre la superficie del suelo. Ladiversidad de macroinvertebradosfue alta: habían termitas,miriápodos, arácnidos, dípteros,lepidópteros, isópodos,dermápteros, moluscos ehirudíneos.

48
El Arado Natural:…
Cuadro 3-1. Correlaciones obtenidas entre los factores extraídos del ACP aplicado a la densidad demacroinvertebrados.
Unidades taxonómicas (UT) Factor I (43.6%) Factor II (15.4%) Factor III (12.8%)
Diversidad 0.90 -0.14 0.32Termitas 0.95 0.05 0.12Miriápodos 0.88 0.004 -0.32Dípteros 0.88 0.04 -0.14Arácnidos 0.96 0.09 0.06Lepidópteros 0.95 0.01 0.15Isópodos 0.90 0.34 -0.09Moluscos 0.92 -0.14 0.08Hirudíneos 0.95 0.06 0.17Lombrices epígeas -0.17 0.83 0.08Lombrices endógeas -0.13 0.72 -0.15P. corethrurus -0.18 0.72 0.52Lombrices endógeas oligohúmicas 0.10 -0.58 -0.16Coleópteros 0.26 -0.77 0.12Blátidos 0.24 0.31 0.25Hormigas -0.08 -0.45 0.66Mermítidos -0.10 -0.21 0.71
- El Factor II explicó el 15.4% dela varianza y se consideró comoel efecto de la tala y la quema.Los sitios se caracterizaron por lapresencia de numerosas raíces ypor la sombra del follaje de losárboles, por las sucesionesseriadas, los cultivos perennes ylos pastizales de kikuyo. La
Cultivos y pastizales Sucesiones y pastizales
Sistemas
2 S40
Selva
5 Café
Yuca7
Brach10
Melinis9
3 S40+
4Barbecho Pennisetum
8
Selva
6 Pino
1
Plantación
Factor I
Factor II
Figura 3-1. Ordenamiento de los sitios estudiados según los dos primeros ejes extraídos del ACP.(Brach = Brachiaria humidicola.)
diversidad epígea, endógea yoligohúmica endógea fueintermedia, incluyendo aquí aPontoscolex corethrurus, especiesde coleópteros y de blátidos(Figura 3-1).
- El Factor III (12.8% de lavarianza) se interpretó como el

49
Cuantificación de la Macrofauna del Suelo...
efecto de la erosión que separa lossitios preparados con tracciónanimal, donde la degradación delsuelo era muy alta y ladiversidad, la abundancia y labiomasa de la macrofauna delsuelo eran bajas. Los organismosmás abundantes fueronP. corethrurus y los de la familiaMermithidae.
La selva secundaria (SS) poseeuna gran riqueza taxonómica (98 UT)y tiene una densidad poblacional(6790 individuos/m2) y una biomasa(98.6 g/m2) altas. Las comunidadesde coleópteros constituyeron el mayorcomponente de la biomasa (52.2%),seguido por el de las termitas (13.2%)y el de las lombrices (12.5%). Ladiversidad de lombrices fue alta y seregistraron las nueve especiessiguientes: Martiodrilus savanicola, M.agricola, Martiodrilus sp., Glossodrilussp., Onoreodrilus sp., Periscolex sp.,Thamnodrilus sp., Dichogaster sp. yuna especie sin identificar. La selvasecundaria de 40 años (S40) y la demás de 40 años (S40+) tuvieronvalores similares de diversidad (27 y31 especies, respectivamente), yvalores diferentes de densidadpoblacional (2056 y3932 individuos/m2) y de biomasa(27.04 y 91.42 g/m2),respectivamente. Las lombricesconstituyen el mayor componente deesta biomasa (para S40, 81%, y paraS40+, 26.6%) así como los coleópteros(para S40, 10.4%, y para S40+,20.6%). La diversidad de lombricesfue alta también en los dos sitios. EnS40 se encontraron las siete especiessiguientes: Martiodrilus heterostichon,Glossodrilus sp., Periscolex sp.,P. corethrurus, Dichogaster sp.,Amynthas gracilis y Periscolex sp.; enS40+ aparecieron cinco especies:Martiodrilus sp., M. agricola,Thamnodrilus sp., Periscolex sp.,A. corticis y Dichogaster sp.
Los sistemas de uso CC, YA y PPpresentaron diferencias en ladensidad poblacional de fauna(3352, 1187 y 870 individuos/m2,respectivamente); tanto en CCcomo en YA, la biomasa fuebastante alta (115.7 y 78.9 g/m2,respectivamente), mientras que enPP se obtuvieron los valores debiomasa más bajos de todos lossitios muestreados (3.2 g/m2). Encomparación con la selva secundaria(SS), la diversidad de estos sistemas,medida en UT, fue baja: CC = 31,YA = 18, PP = 15. Los invertebradosfueron más abundantes en losestratos de 0-10 cm y de 10-20 cm.En CC y YA las lombrices dominaronla biomasa (79.5% y 80.8%,respectivamente). Cuatro especiesnativas (M. savanicola, Periscolex sp.,P. corethrurus y A. gracilis) y unaexótica (P. corethrurus) seconcentraron en los primeros10 cm (62.1% en CC y 40.9% en YA),seguidas por los coleópteros (8.9% y17.1%, respectivamente). Laspoblaciones de invertebrados fueronbajas en PP, siendo los coleópteroslos más abundantes (73.98%). Sólouna especie de lombriz (Glossodrilussp.) se recolectó en este sitio ygeneralmente en los primeros 10 cmde suelo (40.9%).
La fauna se ubicó principalmenteen los primeros 10 cm de suelo enPK (78.4%), PY (62.9%) y BH (52.1%),y los valores de riqueza específicafueron, respectivamente, de 20, 28 y13 UT. La densidad fluctuó entre2038 individuos/m2 en PK y 6443 y1965 individuos/m2 en PY y BH,respectivamente; las hormigaspredominaron especialmente en PY(Ectatoma tuberculatum Dolichoderussp., Hypoponera sp., Solenopsis sp. yTranopelta sp.) y en PK (Linepithemasp., Pheidole sp., Neivamyrmex sp.,Acropyga sp.). La biomasa fue de67.9 g/m2 en PK, 33.9 en PY y

50
El Arado Natural:…
73.4 g/m2 en BH, siendo laslombrices y los coleópteros losprincipales contribuyentes (losporcentajes son respectivos) a labiomasa en PK (83.4%, 13.8%) y enBH (77.9%, 16%), mientras que loscoleópteros y las hormigas lo fueronen PY (64% y 21.5%,respectivamente). En PK seencontraron nueve especies delombrices, o sea, Martiodrilus sp.,M. agricola, M. savanicola,Glossodrilus sp., Thamnodrilus sp.,Dichogaster sp., A. gracilis, A. corticisy Dendrobaena octaedra; en PYcuatro especies: P. corethrurus,Glossodrilus sp., Periscolex yDichogaster sp.; y en BH una sola:P. corethrurus. En los tres pastizales,la mayor incidencia de lombricesocurrió en los primeros 10 cm: enPK 53.1%, en PY 63.3% y en BH73.8%. Los valores obtenidos enB (helechal en barbecho) fueronintermedios entre los de los gruposanteriores. La diversidad fue baja(19 UT), la densidad poblacional alta(4464 individuos/m2) y la biomasabaja (26.28 g/m2). La mayor partede la biomasa fue aportadaprincipalmente por las lombrices(64%) y las hormigas (17%). Lamacrofauna del suelo se concentróen los estratos de 0-10 (44.6%) y de10-20 cm (37.9%). Aquí seencontraron cinco especies delombrices: Martiodrilus sp.,Thamnodrilus sp., Glossodrilus sp.,A. corticis y Dendrobaena octaedra.
DiscusiónLos resultados obtenidos muestrancambios en la diversidad, laabundancia y la estructura funcionalde las comunidades de fauna delsuelo, según el sistema de uso de latierra que se considere en la cuencadel río Cabuyal.
- La riqueza específica disminuyesegún la degradación del suelo:
por ejemplo, en la selvasecundaria (SS) hay 98 UT y9 especies de lombrices, pero sólohay 13 UT y una especie delombriz en BH.
- En los sitios donde existe lasucesión natural, hay unatendencia a mantener labiodiversidad: por ejemplo, de19 UT y 5 especies de lombricesen S40, a 31 UT y 5 especies enla selva secundaria de más de40 años (S40+).
- El cultivo tradicional de café (CC)mostró valores altos de UT y deespecies de lombrices encomparación con los sistemas desucesión natural, mientras quelos valores en YA fueron bajos(18 y 1, respectivamente).
- En PK se encontraron 9 especiesde lombrices y 20 UT(Figura 3-2).
Fue posible distinguir variosorganismos indicadores de ambientesperturbados. En el caso de laslombrices de tierra, las siguientesespecies: A. corticis, A. gracilis,D. octaedra y P. corethrurus; por loscoleópteros, las siguientes:Dichotomius aff. septentrionalis,Heterogomphus chevrolatti,Oxisternom conspicillatum yPassalus sp.
En los ambientes con muchosresiduos vegetales en la superficie delsuelo (‘mulch’), los organismosepígeos y endógeos oligohúmicosfueron los más abundantes, mientrasque en los ambientes herbáceosabundaron principalmente losepígeos. La densidad y la biomasa enSS (Cuadro 3-2) fueron mucho másaltas (hasta 7 veces mayores) que enambientes similares del trópico deMéxico (Lavelle et al. 1981; Lavelle yKohlmann 1984), de Nigeria (Madge1969), de Sarawak (Collins 1980), de

51
Cuantificación de la Macrofauna del Suelo...
Figura 3-2. Unidades taxonómicas de la macrofauna del suelo y riqueza específica de lombrices en10 tipos de uso de la tierra.
27
7
Barbecho19
531
5
Selvasecundaria
P. patula15
1
98
9
M. minutiflora
P. clandestinum
B. humidicola
20
9
13
1
Café tradicional
Yuca asociada
31
4
18
1
Selva de más de 40 años
Selva de 40 años
28
4
= Selvas = Pastizales = Cultivos
= Plantación = Unidades taxonómicas = Lombrices de tierra
Perú (Lavelle y Pashanasi 1989) y dela estación Carimagua, en Colombia(Decaëns et al. 1994).
AgradecimientosNuestro agradecimiento mássincero a los siguientes científicos:Dr. Heimar Quintero, UniversidadNacional de Colombia; Rosa Aldana,Universidad del Valle, Colombia,Luis C. Pardo, INCIVA, Colombia; yJuan José Jiménez, UniversidadComplutense, Madrid. Nuestroreconocimiento especial a ManuelAntonio Trujillo, granjero de laregión, quien fue nuestro compañerodiario durante todo el estudio.
ReferenciasAnderson JM; Flanagan PW. 1989.
Biological processes regulatingorganic matter dynamics in tropicalsoils. En: Coleman DC; Oades JM;Uehara G, eds. Dynamics of soilorganic matter in tropicalecosystems. University of HawaiiPress, Honolulu, Hawaii.
Anderson J; Ingram J. eds. 1993. Tropicalsoil biology and fertility. Ahandbook of methods, 2a. ed. CABInternational, Wallingford, ReinoUnido. 221 p.
Collins NM. 1980. The distribution of soilmacrofauna of the west Ridge ofGunung (Mount) Mulu, Sarawak.Oecologia 44:263-275.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management in theOriental Llanos of Colombia. Eur JSoil Biol 30(4):157-168.
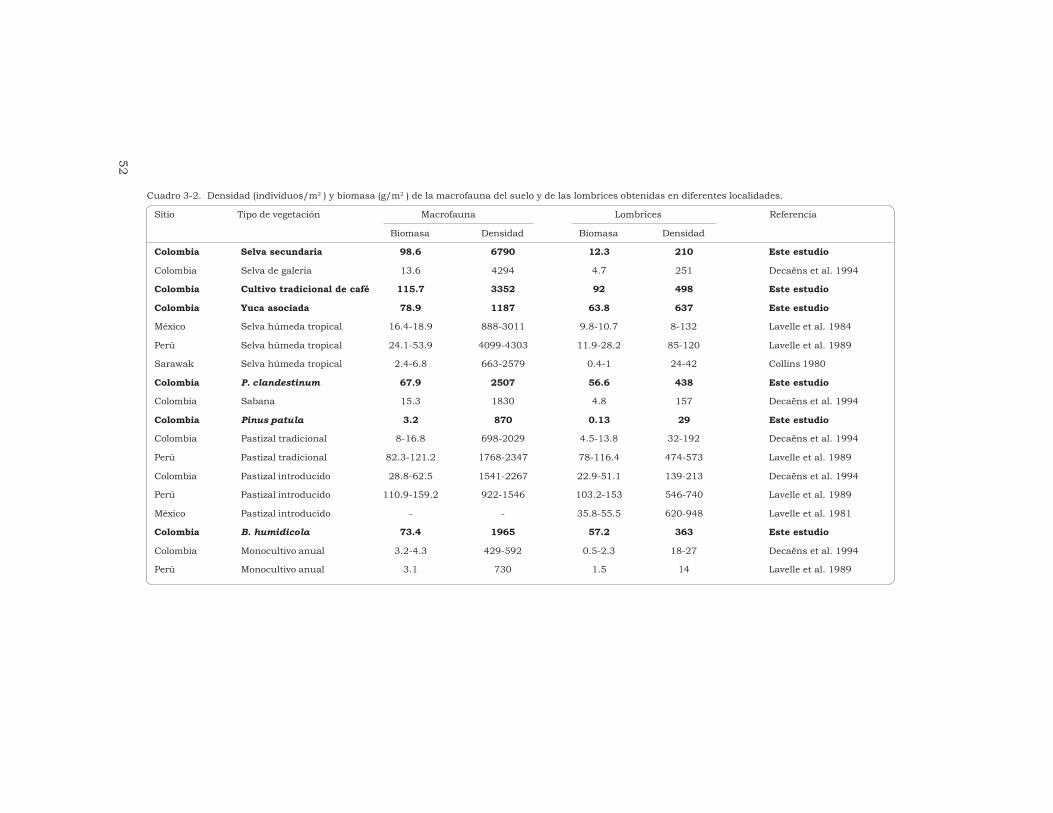
52
El A
rado Natural:…
Cuadro 3-2. Densidad (individuos/m2 ) y biomasa (g/m2 ) de la macrofauna del suelo y de las lombrices obtenidas en diferentes localidades.
Sitio Tipo de vegetación Macrofauna Lombrices Referencia
Biomasa Densidad Biomasa Densidad
Colombia Selva secundaria 98.6 6790 12.3 210 Este estudio
Colombia Selva de galería 13.6 4294 4.7 251 Decaëns et al. 1994
Colombia Cultivo tradicional de café 115.7 3352 92 498 Este estudio
Colombia Yuca asociada 78.9 1187 63.8 637 Este estudio
México Selva húmeda tropical 16.4-18.9 888-3011 9.8-10.7 8-132 Lavelle et al. 1984
Perú Selva húmeda tropical 24.1-53.9 4099-4303 11.9-28.2 85-120 Lavelle et al. 1989
Sarawak Selva húmeda tropical 2.4-6.8 663-2579 0.4-1 24-42 Collins 1980
Colombia P. clandestinum 67.9 2507 56.6 438 Este estudio
Colombia Sabana 15.3 1830 4.8 157 Decaëns et al. 1994
Colombia Pinus patula 3.2 870 0.13 29 Este estudio
Colombia Pastizal tradicional 8-16.8 698-2029 4.5-13.8 32-192 Decaëns et al. 1994
Perú Pastizal tradicional 82.3-121.2 1768-2347 78-116.4 474-573 Lavelle et al. 1989
Colombia Pastizal introducido 28.8-62.5 1541-2267 22.9-51.1 139-213 Decaëns et al. 1994
Perú Pastizal introducido 110.9-159.2 922-1546 103.2-153 546-740 Lavelle et al. 1989
México Pastizal introducido - - 35.8-55.5 620-948 Lavelle et al. 1981
Colombia B. humidicola 73.4 1965 57.2 363 Este estudio
Colombia Monocultivo anual 3.2-4.3 429-592 0.5-2.3 18-27 Decaëns et al. 1994
Perú Monocultivo anual 3.1 730 1.5 14 Lavelle et al. 1989

53
Cuantificación de la Macrofauna del Suelo...
Lavelle P. 1981. Stratégies dereproduction chez les vers de terre.Acta Oecol 2:117-133.
Lavelle P; Kohlman B. 1984. Étudequantitative de la macrofaune dusol dans un fôret tropicale duMexique (Bonampak, Chiapas).Pedobiologia 27:377-393.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lavelle P; Blanchart E; Martin A; SpainAV; Martin S. 1992. The impact ofsoil fauna on the properties of soilsin the humic tropics. En: Lal R;Sánchez P, eds. Myths and scienceof soil of the tropics. SpecialPublication No. 29. Soil ScienceSociety of America, Washington DC.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B; Brussaard L. 1994.The relationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. John Wiley &Sons, Chichester, Reino Unido.p 137-169.
Madge DS. 1969. Field and laboratorystudies on the activities of twospecies of tropical earthworms.Pedobiologia 9:119-129.

PARTE II
Ciclo de Vida y Biología de lasLombrices de Tierra de la Sabana

57
Las Comunidades de Lombrices en las Sabanas Nativas…
CAPÍTULO 4
Las Comunidades de Lombrices en lasSabanas Nativas y en los PastizalesIntroducidos de los Llanos Orientalesde Colombia1
J. J. Jiménez* , A. G. Moreno*, T. Decaëns**, P. Lavelle***,M. J. Fisher† y R. J. Thomas†
ResumenA lo largo de 18 meses se evaluaronla estructura y la variaciónestacional de las comunidades delombrices de una sabana natural yde un pastizal introducido, en unOxisol de Colombia. En cada uno deestos sistemas se delimitó un área de90 x 90 m y cada mes se extrajeronde ella al azar 5 muestras de 1 m2 x0.5 m y 10 muestras de 20 x 20 x20 cm, en un diseño de bloquesestratificados. La riqueza específicafue similar en las dos parcelasestudiadas (siete especies), mientrasque el índice de diversidad H’(Shannon y Weaner 1949) fue muydiferente, es decir, 2.89 en la sabanay 1.29 en el pastizal. Las diferenciasobservadas en la estructura de la
comunidad de lombrices respecto aambos sistemas son la explicación deestos resultados. La densidadmedia anual de la sabana fue de49.8 individuos/m2 y osciló entre10.8 y 135.8 individuos/m2; labiomasa fue de 3.3 g/m2 (método deextracción manual), con valores queoscilaron entre 0.9 y 11.5 g/m2. Enel pastizal introducido, la densidadmedia anual fue de 80.1 individuos/m2 y fluctuó entre 24 y215.8 individuos/m2; la biomasa fuemás de 10 veces superior y oscilóentre 29.2 y 110.4 g/m2. Esto sedebió, principalmente, a la presenciade una especie anécica de grantamaño, Martiodrilus carimaguensisJiménez y Moreno, perteneciente a lafamilia Glossoscolecidae, la cualresultó favorecida al sustituir lasabana nativa por el pastizalintroducido. Las especies endógeasdominaron la comunidad delombrices de la sabana, donde el88% de la biomasa total de lombricesdel pastizal era aportada por M.carimaguensis. La densidad y labiomasa total de las lombrices fueronsignificativamente diferentes en losdos sistemas estudiados (t-test).Estos resultados indican unarespuesta clara de las lombrices a lospastizales introducidos, un sistemaagropastoril que ha sido adoptadoprogresivamente en las sabanashúmedas neotropicales.
1. Publicado en Biology and Fertility of Soils,vol. 28, p 101-110, por Jiménez et al.Copyright © 1998 de Springer. Impreso conpermiso de la casa editorial.
* Departamento de Biología Animal I(Zoología), Facultad de Biología, UniversidadComplutense de Madrid, 28040 Madrid,España.
** Laboratoire d’Ecologie Vegetal, UFRSciences/Unidad de Suelos y Nutrición dePlantas, CIAT, Université de Rouen, 76821Mont Saint Aignan Cx, Francia.
*** Laboratoire d’Ecologie et Biologie des SolsTropicaux, IRD (ex-ORSTOM), 72, routed’Aulnay, 93143, Bondy Cx, Francia.
† Programa de Trópico Bajo (actualmente,Unidad de Suelos y Nutrición de Plantas),CIAT, AA 6713, Cali, Colombia.

58
El Arado Natural:…
Palabras clave: abundancia,biomasa, comunidad delombrices, diversidad, pastizal,manejo de tierra, sabanastropicales
IntroducciónLas sabanas tropicales, cuyaextensión es de casi 250 millones dehectáreas, constituyen uno de losecosistemas predominantes enAmérica del Sur. Las sabanas son,por definición, comunidadesvegetales con una capa herbáceadominante y una capa arbóreadiscontinua, la cual puede constituirel 10% o el 50% de toda la cubiertavegetal (Johnson y Tothill 1985).
Los Oxisoles cubren un área deltrópico superior al 60%, y presentanvalores de pH y contenidos denutrientes bajos (Sánchez y Salinas1983). Estos suelos son el resultadode largos procesos pedogénicos quehan llevado al establecimiento deuna fauna tolerante al medio ácido,muy abundante y activa (Lavelle etal. 1995). Los Llanos Orientales deColombia, cubiertos por sabanasácidas, se dedican a sistemasextensivos de pastoreo en inmensasfincas cuya productividad animal esbaja; en algunas áreas interviene elhombre sembrando pastizalesintroducidos a los que se aplicanfertilizantes (Fisher et al. 1995).
El número de especies de unacomunidad de lombrices es,generalmente, menor después desustituir el ecosistema original, yaque los recursos explotados por lacomunidad se vuelven uniformes y,por tanto, la biodiversidad disminuye(Lavelle 1986). En los suelos ácidostropicales, los pastizales derivados deselvas o de sabanas puedenpresentar biomasas altas demacroinvertebrados del suelo(Decaëns et al. 1994; Lavelle y
Pashanasi 1989). Las especiesexóticas tienden a colonizar estosnuevos hábitat y a eliminar lasespecies endémicas que no consiguenadaptarse a las nuevas condiciones.La gran cantidad de poblacionesexóticas puede ser un factor de ladegradación del suelo. Por ejemplo,las poblaciones de Pontoscolexcorethrurus (Glossoscolecidae), unaespecie de distribución pantropical,compactan el suelo y a vecesocasionan problemas (Barros et al.1996; Pashanasi et al. 1992; Rose yWood 1980).
Hay un gran vacío de informaciónsobre la composición de especies y laestructura de las comunidades delombrices en las sabanas tropicales.Sólo unos pocos estudios hanconsiderado dicho tema: Dash yPatra (1977) y Senapati (1980)realizaron contribuciones importantesal estudio de las lombrices de la India;Lavelle (1978) y Lavelle et al. (1981)estudiaron la fauna oligoquetológicade las sabanas de Costa de Marfil yMéxico, respectivamente.
El objetivo del presente estudiofue evaluar el efecto que causan lospastizales introducidos en laestructura y en la abundancia de lascomunidades de lombrices de tierrade la sabana. La investigación serealizó en la estación experimental deCarimagua, en los Llanos Orientalesde Colombia, donde investiganCORPOICA y el CIAT. Un muestreopreliminar de las comunidades demacrofauna del suelo mostró unarespuesta espectacular de laspoblaciones de macroinvertebrados alos cambios inducidos por el manejode la tierra (Decaëns et al. 1994). Lasabana natural, que posee unacomunidad vegetal muy diversa peropoco productiva, es remplazada porun sistema mucho más productivo enel cual se asocian dos especies cuyaproducción de forraje es de altacalidad. La biomasa de lombrices en

59
Las Comunidades de Lombrices en las Sabanas Nativas…
la sabana sin pastoreo fue de4.8 g/m2, mientras que en el pastizalde gramínea con leguminosa seincrementó hasta 50.0 g/m2
(Cuadro 4-1).
Con el fin de observar ladinámica de las diferentespoblaciones de lombrices de tierra,se hizo también un muestreo de suscomunidades durante los 18 mesesde este estudio.
Materiales y Métodos
Descripción del sitio de trabajo
El área de estudio se sitúa en elCentro Nacional de InvestigacionesAgropecuarias (CNIA) de Carimagua,
en los Llanos Orientales de Colombia(4°37’ N, 71°19’ O y 175 msnm). Lamedia anual de precipitación y detemperatura es 2280 mm y 26 °C,respectivamente, y la época seca va dediciembre a marzo (Figura 4-1). Lavegetación está dominada por sabanasisohipertérmicas bien drenadassituadas sobre Oxisoles infértiles, enlos ‘altos’, y sobre Ultisoles, en los‘bajos’. Los primeros se caracterizanpor su acidez alta (pH en agua = 4.5),una saturación alta de Al (> 90%) y porcontenidos bajos de Ca, Mg y Kintercambiables (Lascano y Estrada1989). Los factores químicos quecontribuyen a la infertilidad del sueloácido y los efectos que causan en elcrecimiento de las plantas sonbastante complejos; entre ellos estánla toxicidad debida al Al, el bajocontenido de P y las tasas bajas de
Cuadro 4-1. Biomasa total de lombrices (LB) y de macrofauna (MC) obtenida en diferentes ecosistemasde Carimagua mediante el muestreo TSBF (Decaëns et al. 1994).
Ecosistema Biomasa LB Biomasa MC Biomasa LB(g peso fresco/m2) (g peso fresco/m2) (%)
Pastizal introducido 51.1 62.5 82Sabana nativa 4.8 15.3 31Selva de galería 4.7 13.6 35Monocultivo anual 2.3 3.2 71
Tem
pera
tura
(°C
)
30
25
20
15
10
5
0E F M A M J J A S O N D
400
300
200
100
0
Figura 4-1. Media mensual de temperatura y de precipitación en Carimagua, durante 22 años(1973-1995).FUENTE: Base de datos del CIAT.
Prec
ipit
ació
n (m
m)
Temperatura PrecipitaciónMeses

60
El Arado Natural:…
mineralización del N (Rao et al.1993). Los suelos de la zonaestudiada se pueden definir como deun Haplustox típico, finos, caoliníticoe isohipertérmico (suelos franco-arcillosos). Las especies vegetalespredominantes son Andropogonbicornis, Gymnopogon sp., Panicumsp., Trachypogon sp. y Axonopus sp.
Se evaluaron dos sistemas demanejo que presentaban fuertescontrastes: una sabana natural y unpastizal de 17 años en que seasociaron una gramínea africanaexótica, Brachiaria decumbens cv.Basilisk, y una leguminosa herbáceaforrajera, la especie Puerariaphaseoloides CIAT 9900 (el “kudzú”),adaptadas ambas a las condicionesde los suelos ácidos. El pastizal seestableció en un área ocupada antespor la sabana, y se fertilizó con 44 P,40 K, 14 Mg y 22 S en el momentode la siembra y con 10 P, 9 K,92.5 Ca, 9 Mg y 11.5 S cada 2 años(las cifras en kg/ha) en los 9 añossiguientes (Lascano y Estrada 1989).La carga animal en el pastizal fue de1 animal/ha en la época seca y2 animales/ha en la época lluviosa.Las principales características físicasy químicas de los suelos donde se
hicieron los muestreos aparecen enel Cuadro 4-2. No hay diferenciassignificativas aparentes entre los dossitios muestreados respecto a suscondiciones físicas y químicas, yaque se localizan en la misma unidadedafológica (IGAC 1974).
Muestreo de lombricesLos muestreos se realizaron cadames en ambos sistemas desde marzode 1994 hasta septiembre de 1995,excepto en junio de 1994. En cadasistema se delimitó un área o lote de90 x 90 m que fue dividido enparcelas cuadradas regulares de10 x 10 m hasta obtener un númerode 81 unidades de muestreo. Laextracción de las lombrices se hizopor combinación de una recolecciónmanual directa de muestras de suelode 1 m2 x 0.5 m de profundidad y dellavado y tamizado de monolitos de20 x 20 x 20 cm de profundidad(Lavelle 1978). En cada lote (ysistema) se tomaron al mes cincomuestras al azar de 1 m2 y10 muestras de lavado y tamizado(excepto en marzo de 1994, dondeéstas fueron 3 y 6, respectivamente).
Cuadro 4-2. Principales propiedades físicas y químicas de los sistemas estudiados (Análisis CIAT). Losdatos son medias ± desviación típica (dt).
Propiedades del suelo Sabana nativa Pastizal mejorado
Carbono (%)a 3.44 ± 1.67 3.41 ± 1.46P (µg/g suelo; Bray II) 1.26 ± 0.38 2.91 ± 2.22P (µg/g suelo)b 3.85 ± 0.85 4.82 ± 2.04pH 3.97 ± 1.55 4.97 ± 0.07Al (meq/100 g suelo ) 2.42 ± 0.12 1.90 ± 0.19Ca (meq/100 g suelo ) 0.36 ± 0.26 0.89 ± 0.20Mg (meq/100 g suelo ) 1.22 ± 2.43 0.33 ± 0.27K (meq/100 g suelo ) 0.32 ± 0.68 0.71 ± 1.37H (meq/100 g suelo ) 2.01 ± 2.17 1.44 ± 2.44N total (µg/g suelo ) 1538.87 ± 250.77 1670.37 ± 116.03P total (µg/g suelo ) 184.75 ± 8.74 226.75 ± 12.83Textura Arcillosa Arcillosa
a. Walkey-Black modificado (con calor).b. Bray II modificado.

61
Las Comunidades de Lombrices en las Sabanas Nativas…
La profundidad de muestreovarió estacionalmente a causa de lasmigraciones verticales de algunasespecies, como Martiodriluscarimaguensis (Glossoscolecidae), lacual se encuentra normalmente a80 cm de profundidad durante laépoca seca (Jiménez et al. 1998).Con el fin de facilitar la separaciónde los diferentes estratos analizadosy evitar la migración lateral dealgunas lombrices fuera delmonolito, se cavó una zanjaalrededor de éste. La muestra fuesubdividida en capas de 10 cm degrosor y todas las lombrices fueronrecogidas manualmente, lavadas enagua y fijadas en formol al 4% (10%de la dilución comercial). Laslombrices fueron separadas segúnsu especie y también en adultos,jóvenes e inmaduros, y fueron luegocontadas y pesadas en cada grupo(el peso fresco en formol es, enpromedio, un 15% menor que supeso vivo).
Cálculo del peso de laslombricesUna de las desventajas del muestreode extracción manual es que granparte de los individuos,principalmente los más grandes,quedan fragmentados, por lo que esbastante difícil conocer su pesoindividual. Se han buscado algunasrelaciones que permitan establecer elpeso completo de un individuo apartir de algunas variablesbiométricas de la parte anterior desu cuerpo de lombriz. El diámetropreclitelar máximo se usó paraestimar el peso total de losindividuos incompletos. La relaciónencontrada era o bien lineal oexponencial, y se hacían lascorrecciones pertinentes (Figura 4-2).
Medida de la humedad del sueloEn cada capa analizada manualmenteen el monolito de 1 m2 se tomaronmuestras de suelo que fueronintroducidas en bolsas plásticas yllevadas al laboratorio. De cada bolsase tomaron seis submuestras deaproximadamente 100 g de pesohúmedo y se secaron al horno duranteun mínimo de 4 días. El contenido deagua en el suelo se calculó según laexpresión:
[(Peso húmedo del suelo – peso secodel suelo)/Peso seco del suelo] x 100
Deposición de hecessuperficialesLas heces superficiales depositadaspor la especie anécica de gran tamañoM. carimaguensis (Glossoscolecidae),que tienen forma de torre y sedistinguen muy bien de lasdeposiciones de otras especies, fueroncontadas en ambos sistemas con el finde calcular la densidad poblacional.Las heces se retiraron de la unidad demuestreo escogida poco antes de laextracción del bloque de 1 m2 y seclasificaron en diferentes categorías:secas, frescas y frescas no recientes(de más de un día). Cada mes, en dosde las cinco muestras de 1 m2 serecogían todas las heces, se secaban yse pesaban para evaluar suproducción en ambos sistemas de usode la tierra.
Resultados
Riqueza específica, diversidad yequitatividadEn la región de Carimagua se hanrecolectado 21 especies de lombrices.La riqueza en especies de los dossistemas estudiados fue muy similar:8 especies en la sabana nativa y 7 enel pastizal (Cuadro 4-3). Andiorrhinus
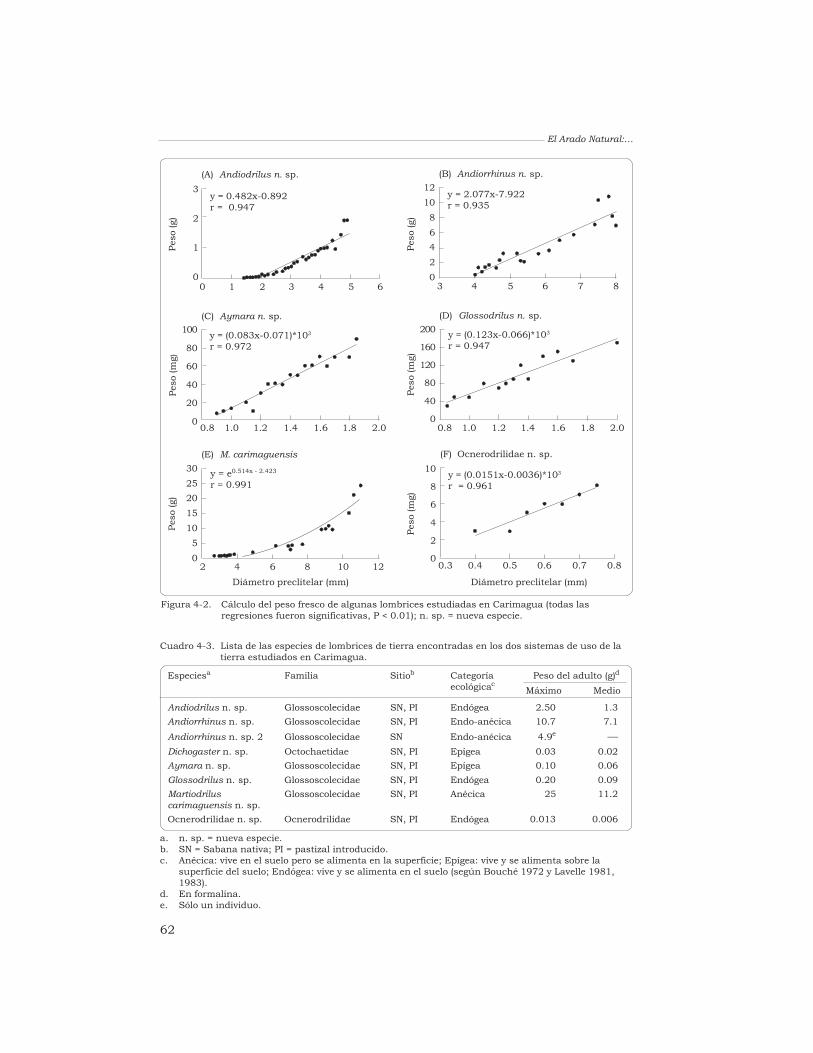
62
El Arado Natural:…
Cuadro 4-3. Lista de las especies de lombrices de tierra encontradas en los dos sistemas de uso de latierra estudiados en Carimagua.
Especiesa Familia Sitiob Categoría Peso del adulto (g)decológicac
Máximo Medio
Andiodrilus n. sp. Glossoscolecidae SN, PI Endógea 2.50 1.3Andiorrhinus n. sp. Glossoscolecidae SN, PI Endo-anécica 10.7 7.1
Andiorrhinus n. sp. 2 Glossoscolecidae SN Endo-anécica 4.9e —Dichogaster n. sp. Octochaetidae SN, PI Epígea 0.03 0.02Aymara n. sp. Glossoscolecidae SN, PI Epígea 0.10 0.06Glossodrilus n. sp. Glossoscolecidae SN, PI Endógea 0.20 0.09Martiodrilus Glossoscolecidae SN, PI Anécica 25 11.2carimaguensis n. sp.Ocnerodrilidae n. sp. Ocnerodrilidae SN, PI Endógea 0.013 0.006
a. n. sp. = nueva especie.b. SN = Sabana nativa; PI = pastizal introducido.c. Anécica: vive en el suelo pero se alimenta en la superficie; Epígea: vive y se alimenta sobre la
superficie del suelo; Endógea: vive y se alimenta en el suelo (según Bouché 1972 y Lavelle 1981,1983).
d. En formalina.e. Sólo un individuo.
Figura 4-2. Cálculo del peso fresco de algunas lombrices estudiadas en Carimagua (todas lasregresiones fueron significativas, P < 0.01); n. sp. = nueva especie.
Peso
(g)
3
2
1
00 1 2 3 4 5 6
y = 0.482x-0.892r = 0.947
Peso
(g)
302520151050
y = e0.514x - 2.423
r = 0.991
Diámetro preclitelar (mm)
Peso
(mg)
100
80
60
40
20
0
y = (0.083x-0.071)*103
r = 0.972
200
160
120
80
40
0
y = (0.123x-0.066)*103
r = 0.947
10
8
6
4
2
0
y = (0.0151x-0.0036)*103
r = 0.961
Diámetro preclitelar (mm)
121086420
y = 2.077x-7.922r = 0.935
3 4 5 6 7 8
0.8 1.0 1.2 1.4 1.6 1.8 2.0 0.8 1.0 1.2 1.4 1.6 1.8 2.0
2 4 6 8 10 12 0.3 0.4 0.5 0.6 0.7 0.8
Peso
(mg)
Peso
(mg)
Peso
(g)
(A) Andiodrilus n. sp.
(E) M. carimaguensis
(C) Aymara n. sp. (D) Glossodrilus n. sp.
(F) Ocnerodrilidae n. sp.
(B) Andiorrhinus n. sp.

63
Las Comunidades de Lombrices en las Sabanas Nativas…
n. sp. 2 fue la única especie de lasabana nativa que no se encontró enel pastizal, aunque es muy rara en lasabana pues sólo se registró en ellaun solo individuo durante todo elperiodo de muestreo. No aparecieronespecies exóticas en el pastizal y las7 recolectadas provenían de lasabana.
El índice de diversidad se calculómediante valores del promedio anualde biomasa, ya que es éste elprocedimiento recomendado cuandose dispone de esos datos (Barbault1992; Ludwig y Reynolds 1988). Elíndice de Shannon transformado quese obtuvo para la sabana fue 2.89 yel obtenido para el pastizal fue 1.29;los valores respectivos deequitatividad, H’/lnS (Pielou 1975),fueron 0.51 y 0.27.
La familia Glossoscolecidae,endémica en el neotrópico, fue lamás abundante; de ella seencontraron 6 especies: Andiodrilusn. sp., Andiorrhinus n. sp. 1,Andiorrhinus n. sp. 2, M.carimaguensis, Glossodrilus n. sp. yAymara n. sp. Se recolectaron dosespecies más, pertenecientes una ala familia Octochaetidae y la otra a lafamilia Ocnerodrilidae.
La comunidad de lombrices deCarimagua está compuesta porespecies cuyo tamaño y funciónecológica son diversos; dos especiesson epígeas, cuatro endógeas y unaanécica. En los estratos profundosdel suelo, donde el contenido de MOes menor, se encontraron ejemplaresde la familia Ocnerodrilidae en lasheces depositadas por individuosgrandes de M. carimaguensis en elinterior de galerías verticales. Estaespecie disgrega estas masascompactas de heces en estructurasgranulares más finas.
Densidad y biomasaEl valor medio anual de la densidady de la biomasa obtenidas en lasabana fueron 49.8 individuos/m2 y3.26 g/m2, respectivamente,mientras que en el pastizal ladensidad fue de 80.1 individuos/m2
y la biomasa de 57.1 g/m2, es decir,más de 10 veces el valor respectivoen la sabana. Las diferencias entrelos sistemas en cuanto a la densidady la biomasa fueron significativas(P < 0.002 y P < 0.001) a causa,principalmente, del notableincremento que experimentan laspoblaciones de M. carimaguensiscuando la sabana se sustituye porun pastizal mejorado (Figura 4-3).
Las diferencias entre las parcelasde los dos sistemas respecto a losvalores del promedio mensual de lahumedad del suelo fueronsignificativos (P = 0.028, t-test). Lasabana natural estaba más húmedaque el pastizal mejorado (Cuadro 4-4)y los valores de densidad y debiomasa estuvieron fuertementecorrelacionados (al 1% designificación) con la humedad mediadel suelo, excepto los valores debiomasa obtenidos en la sabananatural (éstos lo están a un 5%). Porel contrario, las diferencias enhumedad del suelo a lo largo de losestratos, entre ambos sistemas deuso de la tierra, no fueronsignificativas (ANOVA).
Los valores mensuales dedensidad y biomasa estuvieronestrechamente ligados con lahumedad del suelo, y la presencia deuna época seca de 4 meses deduración disminuyó drásticamente ladensidad poblacional. La densidadmínima de lombrices en la sabanase obtuvo en febrero de 1995(10.8 individuos/m2) y la máxima enjulio de 1994 (135.8). En el pastizal,la densidad mínima se obtuvotambién en febrero de 1995
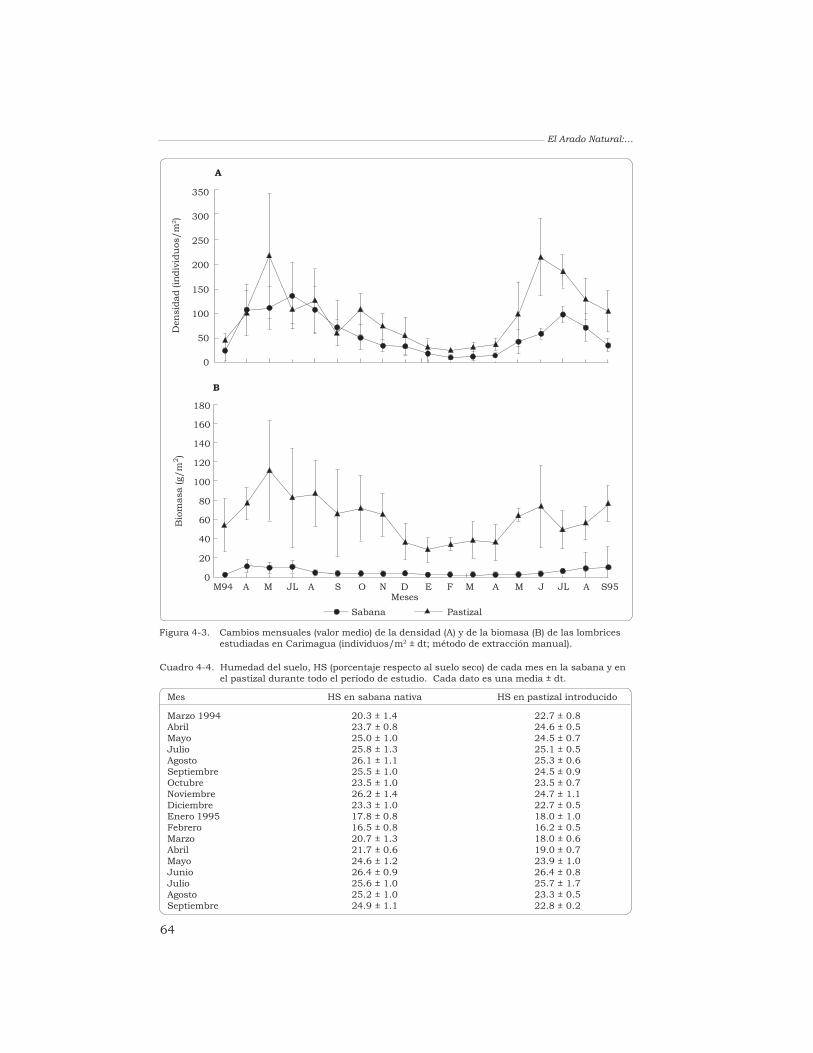
64
El Arado Natural:…
Cuadro 4-4. Humedad del suelo, HS (porcentaje respecto al suelo seco) de cada mes en la sabana y enel pastizal durante todo el período de estudio. Cada dato es una media ± dt.
Mes HS en sabana nativa HS en pastizal introducido
Marzo 1994 20.3 ± 1.4 22.7 ± 0.8Abril 23.7 ± 0.8 24.6 ± 0.5Mayo 25.0 ± 1.0 24.5 ± 0.7Julio 25.8 ± 1.3 25.1 ± 0.5Agosto 26.1 ± 1.1 25.3 ± 0.6Septiembre 25.5 ± 1.0 24.5 ± 0.9Octubre 23.5 ± 1.0 23.5 ± 0.7Noviembre 26.2 ± 1.4 24.7 ± 1.1Diciembre 23.3 ± 1.0 22.7 ± 0.5Enero 1995 17.8 ± 0.8 18.0 ± 1.0Febrero 16.5 ± 0.8 16.2 ± 0.5Marzo 20.7 ± 1.3 18.0 ± 0.6Abril 21.7 ± 0.6 19.0 ± 0.7Mayo 24.6 ± 1.2 23.9 ± 1.0Junio 26.4 ± 0.9 26.4 ± 0.8Julio 25.6 ± 1.0 25.7 ± 1.7Agosto 25.2 ± 1.0 23.3 ± 0.5Septiembre 24.9 ± 1.1 22.8 ± 0.2
Meses
350
300
250
200
150
100
50
0
Den
sida
d (in
divi
duos
/m2 )
180
160
140
120
100
80
60
40
20
0
Bio
mas
a (g
/m2 )
Figura 4-3. Cambios mensuales (valor medio) de la densidad (A) y de la biomasa (B) de las lombricesestudiadas en Carimagua (individuos/m2 ± dt; método de extracción manual).
A
Sabana Pastizal
B
M94 A M JL A S O N D E F M A M J JL A S95

65
Las Comunidades de Lombrices en las Sabanas Nativas…
(24 individuos/m2) y la máxima enmayo de 1994 (215.8). La biomasaen la sabana fue mínima en marzode 1994 (0.74 g/m2) y máxima enseptiembre de 1995 (11.5 g/m2). Enel pastizal, la biomasa fue mínima enenero de 1995 (29.2 g/m2) y máximaen mayo de 1994 (110.4 g/m2). Enel segundo año de muestreo, seobservó un retraso en el inicio de laactividad de las lombrices, ya que laépoca lluviosa comenzó casi un mesmás tarde de lo habitual. Enseptiembre de 1994, la profundidadde muestreo se incrementó paraseguir tras las migracionesverticales de las poblaciones deM. carimaguensis, que comienzan dejulio en adelante; en agosto de 1994,la densidad y la biomasa fueron talvez subvaloradas puesto que,probablemente, parte de la poblaciónhabía comenzado ya a migrar hacialos horizontes más profundos.
Se encontraron tambiéndiferencias en la estructura de lacomunidad de lombrices. En lasabana nativa, la especie másabundante, en términos de biomasa,fue Glossodrilus n. sp., una especieendógea de tamaño mediano ydelgada, que vive a una profundidadmedia de 7 cm; las especiesendógeas aportaron el 80% de labiomasa total contabilizada. En elpastizal, M. carimaguensis fue elprincipal componente de la biomasa(con un 88% del total) y las especiesendógeas, en este caso, sólocontribuyeron con el 12% a labiomasa total de lombrices(Figura 4-4). En el pastizal, sólo tresespecies incrementaron suspoblaciones significativamente encomparación con las de la sabananatural, y ellas son: Andiodrilus n.sp., Andiorrhinus n. sp. 1 y M.carimaguensis (Cuadro 4-5).
Deposición de hecessuperficialesM. carimaguensis deposita, sobre lasuperficie del suelo, heces grandescon forma de torre que llegan amedir hasta 10 cm de altura y 5 cmde ancho. Esta especie de grantamaño, cuyos adultos miden, enpromedio, 9.3 mm de diámetro y194 mm de longitud, habita en
Figura 4-4. Contribución relativa anual de lasdiferentes categorías ecológicas delombrices a la biomasa total deéstas. El tamaño de los círculos esproporcional a la biomasaobtenida.
Sabana
Epígea2.4%
Endógeas83.2%
Anécica14.4%
Pastizal
Anécica87.9%
Epígea0.1%
Endógeas12.0%
Cuadro 4-5. Diferencias encontradas en ladensidad (letra normal) y en labiomasa (letra cursiva) de la mediaanual de lombrices por m2 en losdos sistemas de uso de la tierraestudiados.
Especies Sabana Pastizal
Andiodrilus n. sp. 2.3 5.00.82 2.39*
Andiorrhinus n. sp. 0 0.6*0 1.78*
Aymara n. sp. 13.1 6.60.25 0.15
Glossodrilus n. sp. 50.1 51.23.04 4.43
M. carimaguensis 0.2 18.2**0.47 50.74**
Ocnerodrilidae n. sp. 14.9 22.60.08 0.13
*, ** = P < 0.05, P < 0.001.

66
El Arado Natural:…
galerías verticales que no sonpermanentes.
La producción de estas hecesvarió mucho de la sabana al pastizalpues casi la totalidad de la superficiede éste último estaba cubierta porellas. Las heces frescas indicaban lapresencia de un individuo activo. Seencontró una relación significativa(P<0.01) entre el número de hecesfrescas y el número de individuos deesta especie encontrados en losprimeros 10 cm del suelo durante laépoca lluviosa (Figura 4-5). Elcoeficiente r estuvo muy próximo a1, lo que indica que cada deposiciónfresca es depositada por unindividuo que está activo en sugalería.
La producción de heces mostrótambién fluctuaciones estacionalesmuy notorias: fue máxima al iniciode la época lluviosa y nula durantela época seca (Figura 4-6). Elnúmero de heces frescas obtenidasen el pastizal fue 11 veces mayorque las halladas en la sabana(Cuadro 4-6). Sin embargo, en el
Figura 4-5. Correlación entre el número deheces frescas y la abundancia deindividuos de M. carimaguensis enun pastizal introducido (P < 0.01).Los datos fueron obtenidosdurante la época lluviosa, desdeabril hasta diciembre de 1994.
Abu
nda
nci
a (n
o./m
2 )
r = 0.907
Heces frescas (no./m2) 0 2 4 6 8 10 12 14 16
20
18
16
14
12
10
8
6
4
2
0
transcurso de un año, desde octubrede 1994 hasta septiembre de 1995, elpeso total de las heces recolectadas,frescas y secas, fue de 31.3 t/ha enla sabana y de 37.7 t/ha en elpastizal.
Variación estacional de laactividadLas sabanas tropicales secaracterizan por una notoriaestacionalidad de las condicionesambientales. En Carimagua, lahumedad del suelo descendía avalores que fluctuaban entre el12.6% y el 16.5% (pF 4.2 = 15.4%) enlos primeros 10 cm de suelo (pesoseco) durante la época seca. El valormínimo obtenido durante dichaépoca en el pastizal fue 13.5%.Durante la época de lluvias, losvalores más altos obtenidos en lasabana y en el pastizal fueron 34.7%y 34.6% y los valores medios fueron26% y 24.7%, respectivamente.Respecto a las lombrices, tanto en lassabanas naturales como en lospastizales se observaron diferenciasen su actividad, según la especie.Todas las lombrices de tierraestuvieron inactivas durante la épocaseca, desde diciembre hasta marzo.Puesto que el contenido de agua delsuelo también varió con laprofundidad, esto explica lasdiferencias en el ciclo estacional delas diferentes especies.
Andiodrilus n. sp. y Andiorrhinusn. sp. 1, dos especies endógeas, nopresentaron, aparentemente,ninguna estrategia o forma deresistencia a la época seca.Descendieron simplemente unasdecenas de centímetros en el suelo(hasta la capa de 30-50 cm) yentraron en una fase de quiescencia.Muchos individuos murieron y lossupervivientes empezaron areproducirse al inicio de la época delluvias para poder incrementar sus
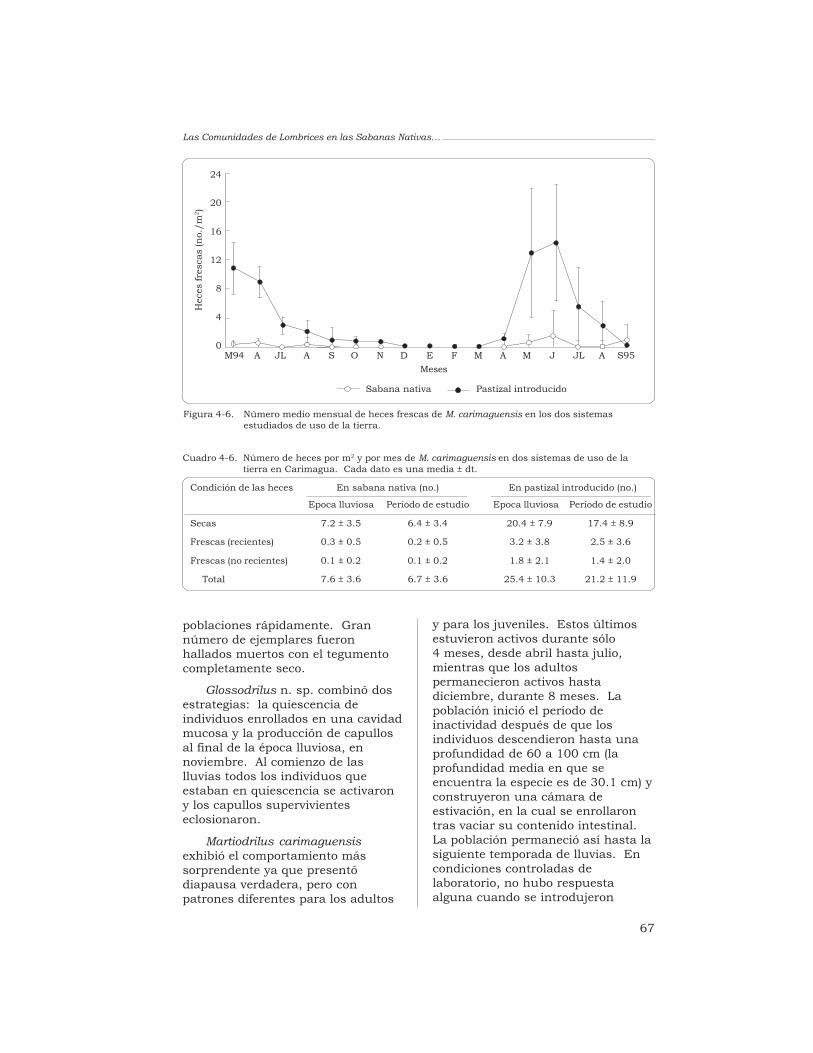
67
Las Comunidades de Lombrices en las Sabanas Nativas…
poblaciones rápidamente. Grannúmero de ejemplares fueronhallados muertos con el tegumentocompletamente seco.
Glossodrilus n. sp. combinó dosestrategias: la quiescencia deindividuos enrollados en una cavidadmucosa y la producción de capullosal final de la época lluviosa, ennoviembre. Al comienzo de laslluvias todos los individuos queestaban en quiescencia se activarony los capullos supervivienteseclosionaron.
Martiodrilus carimaguensisexhibió el comportamiento mássorprendente ya que presentódiapausa verdadera, pero conpatrones diferentes para los adultos
y para los juveniles. Estos últimosestuvieron activos durante sólo4 meses, desde abril hasta julio,mientras que los adultospermanecieron activos hastadiciembre, durante 8 meses. Lapoblación inició el período deinactividad después de que losindividuos descendieron hasta unaprofundidad de 60 a 100 cm (laprofundidad media en que seencuentra la especie es de 30.1 cm) yconstruyeron una cámara deestivación, en la cual se enrollarontras vaciar su contenido intestinal.La población permaneció así hasta lasiguiente temporada de lluvias. Encondiciones controladas delaboratorio, no hubo respuestaalguna cuando se introdujeron
Cuadro 4-6. Número de heces por m2 y por mes de M. carimaguensis en dos sistemas de uso de latierra en Carimagua. Cada dato es una media ± dt.
Condición de las heces En sabana nativa (no.) En pastizal introducido (no.)
Epoca lluviosa Período de estudio Epoca lluviosa Período de estudio
Secas 7.2 ± 3.5 6.4 ± 3.4 20.4 ± 7.9 17.4 ± 8.9
Frescas (recientes) 0.3 ± 0.5 0.2 ± 0.5 3.2 ± 3.8 2.5 ± 3.6
Frescas (no recientes) 0.1 ± 0.2 0.1 ± 0.2 1.8 ± 2.1 1.4 ± 2.0
Total 7.6 ± 3.6 6.7 ± 3.6 25.4 ± 10.3 21.2 ± 11.9
M94 A JL A S O N D E F M A M J JL A S95
24
20
16
12
8
4
0
Hec
es fr
esca
s (n
o./m
2 )
Sabana nativa Pastizal introducido
Figura 4-6. Número medio mensual de heces frescas de M. carimaguensis en los dos sistemasestudiados de uso de la tierra.
Meses

68
El Arado Natural:…
individuos inactivos en un suelohumedecido hasta la capacidad decampo, lo que indica la presencia deun fenómeno de diapausafisiológicamente inducida. Otracaracterística importante de lalombriz que estiva es que,generalmente, sella el final de sugalería con varios tabiques formadoscon heces para evitar la pérdida dehumedad del tegumento, la cual esvital para mantener una tasamínima de respiración cutánea. Lacombinación de estos dos patronesde comportamiento permite a laespecie reducir un riesgo demortalidad que sería alto, dadas lascondiciones tan desfavorables de laépoca seca.
Los capullos de esta especieanécica son ovales, de color marrónclaro y miden 25 x 15 mm; laslombrices los depositan entre los20 y los 50 cm de profundidad,excepto aquellos adultos y juvenilesque se encuentren a mayorprofundidad durante su diapausa.De cada capullo surgen dosindividuos de 760 mg de peso mediocada uno (n = 33) los cuales, duranteel período de agosto a diciembredescienden rápidamente a las capasinferiores a inactivarse (Jiménez etal. 1998). La relación existente entreel peso del capullo y el del adulto esla más alta obtenida hasta la fecha
en los estudios de las comunidadesde lombrices (Cuadro 4-7).
La pequeña especie oligohúmicade la familia Ocnerodrilidae (20 a30 mm de longitud) se encontróinactiva a gran profundidad durantetoda la época seca. En la épocalluviosa se alimentaba de las hecesde M. carimaguensis, ingiriéndolas yformando agregados mucho máspequeños. Esta especie se alimentótambién de otros recursos orgánicos,por ejemplo, algunos nidos decoleópteros de la subfamiliaScarabeinae.
DiscusiónDos objetivos perseguía este trabajo:uno, contribuir al estudio de laslombrices de las sabanas de Américadel Sur, y otro, investigar los efectosdel establecimiento de pastizalesintroducidos en la comunidad delombrices que habita un suelo desabana cuya fertilidad se suponebaja. Se estudiaron estos efectos, enespecial, porque se sembraronespecies vegetales muy productivasque aumentaban la disponibilidad dela hojarasca y la MO del suelo(Sombroek et al. 1993; Thomas et al.1995).
Hay estudios sobre la ecología delas lombrices en los ecosistemas
Cuadro 4-7. Aspectos cuantitativos de la reproducción de algunas especies anécicas de latitudestempladas y tropicales.
Especies Categoría Peso del Peso del Fecundidad PC/PA Referenciaecológica adulto, capullo,
PA (mg) PC (mg)
Millsonia lamtoiana Epianécica 32,000 320 3.1 0.01 Lavelle 1978
Lampito mauritii Epianécica (?) 1220 26 9.3 0.02 Senapati 1980
Allolobophora longa Anécica 2150 60 8 0.03 Evans y Guild1948
Lumbricus terrestris Epianécica 5230 57.1 - - Evans y Guild1948
M. carimaguensis Anécica 9580 1804 0.49 0.19 Este estudio

69
Las Comunidades de Lombrices en las Sabanas Nativas…
tropicales que han considerado tantola función que cumplen las especiesendémicas como su capacidad deadaptación cuando los ecosistemasnaturales son sustituidos por laacción humana (Fragoso 1993;Lavelle 1978; Lavelle y Pashanasi1989; Nemeth 1981; Senapati 1980).En Carimagua se presenta unasituación muy diferente de laencontrada en muchos pastizalesneotropicales que han sustituido laselva primaria: en éstos, lacomunidad de lombrices estádominada por una especie exóticaque es endógea mesohúmicaneotropical y pertenece a la familiaGlossoscolecidae, es decir,Pontoscolex corethrurus Müller 1857.Tal es el caso de Yurimaguas en Perú(Lavelle y Pashanasi 1989), deFlorencia en Colombia (Jiménez, obs.pers.), de Manaos en Brasil, deMéxico y Costa Rica (Fragoso yLavelle 1992) y de Ecuador (Lavelle,com. pers.)
Los pastizales son similares a lassabanas, en términos funcionales(Lavelle et al. 1994). Cuandoaquéllos se establecen en áreasselváticas de alta precipitación,aparecen, generalmente, especiesalóctonas de lombrices, por ejemplo,P. corethrurus Müller 1857 yPolypheretima elongata Perrier 1872.Los pastizales establecidos en suelosde sabana, ya estén conformadossólo por gramíneas o ya asociadoscon leguminosas forrajeras, son máspropensos a conservar la comunidadnativa de lombrices que aquellosestablecidos en sitios cubiertos antespor el bosque húmedo tropical.
Aunque en Carimagua laintroducción del pastizal mejoradoes, sin duda, una perturbación delmedio, se conserva en él la riquezade especies de lombrices que poseíaoriginalmente la sabana, tanto si elpasto está solo en el pastizal oasociado con leguminosas. Ahora
bien, la diversidad del pastizal esmenor que la de la sabana cuando secalcula el índice H’ de Shannon(Shannon y Weaver 1949) y no todaslas especies de la sabana resultaronfavorecidas por la adopción delsistema mejorado.
La drilosfera, uno de los cuatrocomponentes de los sistemasbiológicos de regulación del suelo(Lavelle et al. 1993), desempeña unpapel importante en los pastizales,donde está muy desarrollada y esmuy activa. Lee (1985) y Syers ySpringett (1984) resaltan los altosvalores de densidad y de biomasa delombrices que se han hallado en esosecosistemas. Kouassi (1987) estudióla macrofauna endógea de lassabanas y de los pastizales de Costade Marfil y encontró que laslombrices fueron el principalcomponente de la biomasa total deambos sistemas, con algunasdiferencias entre ellos:
– en los pastizales, la biomasa delombrices varió desde 6.5 g/m2,en la época seca, hasta70.9 g/m2 en la época lluviosa;
– en las sabanas, la variaciónrespectiva de la biomasa fue de3.0 y 26.3 g/m2.
El número de lombrices de lasabana nativa de Carimagua, que seencontró y estudió, es bajo si secompara con el de otras sabanastropicales (Cuadro 4-8), pero labiomasa del pastizal es altacomparada con la de otros lugarestropicales; sin embargo, estascomparaciones deben hacerse concierta cautela ya que se emplearonmétodos diferentes de extracción delombrices en esos lugares. En elCuadro 4-9 se comparan la densidady la biomasa de las lombrices deCarimagua, así como la contribuciónrelativa de las diferentes especies aambos parámetros (abundancia y
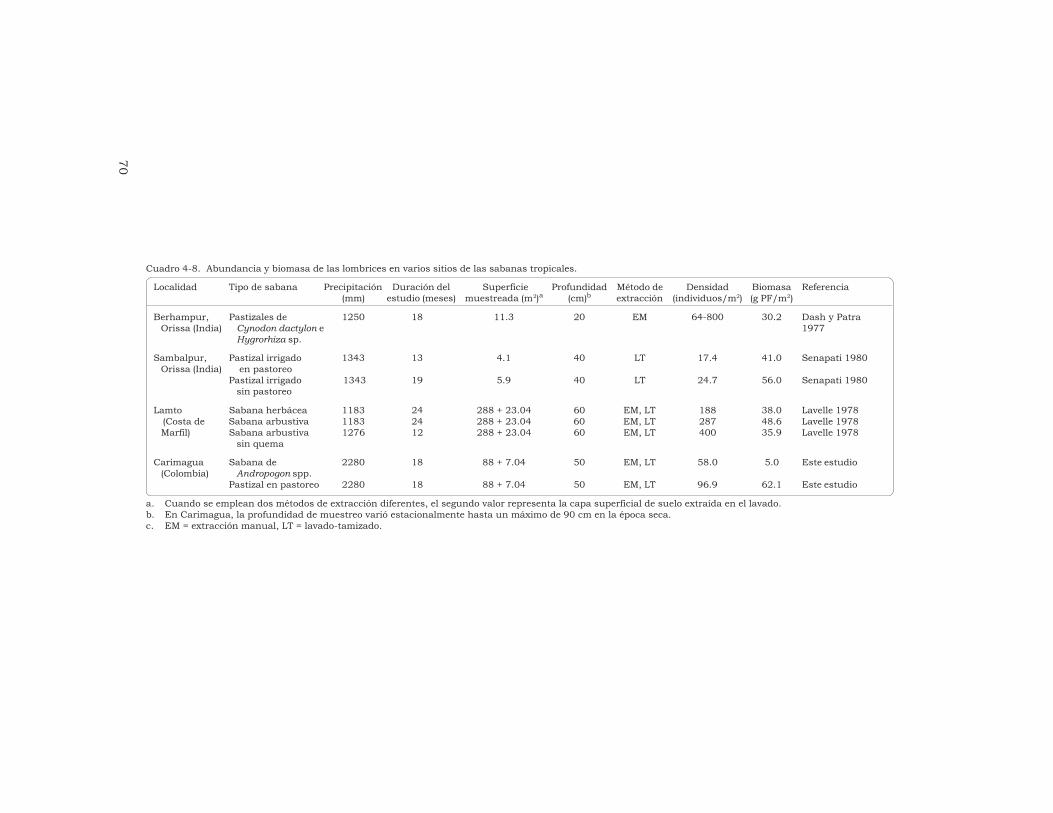
70
El A
rado Natural:…
Cuadro 4-8. Abundancia y biomasa de las lombrices en varios sitios de las sabanas tropicales.
Localidad Tipo de sabana Precipitación Duración del Superficie Profundidad Método de Densidad Biomasa Referencia(mm) estudio (meses) muestreada (m2)a (cm)b extracción (individuos/m2) (g PF/m2)
Berhampur, Pastizales de 1250 18 11.3 20 EM 64-800 30.2 Dash y PatraOrissa (India) Cynodon dactylon e 1977
Hygrorhiza sp.
Sambalpur, Pastizal irrigado 1343 13 4.1 40 LT 17.4 41.0 Senapati 1980Orissa (India) en pastoreo
Pastizal irrigado 1343 19 5.9 40 LT 24.7 56.0 Senapati 1980sin pastoreo
Lamto Sabana herbácea 1183 24 288 + 23.04 60 EM, LT 188 38.0 Lavelle 1978(Costa de Sabana arbustiva 1183 24 288 + 23.04 60 EM, LT 287 48.6 Lavelle 1978Marfil) Sabana arbustiva 1276 12 288 + 23.04 60 EM, LT 400 35.9 Lavelle 1978
sin quema
Carimagua Sabana de 2280 18 88 + 7.04 50 EM, LT 58.0 5.0 Este estudio(Colombia) Andropogon spp.
Pastizal en pastoreo 2280 18 88 + 7.04 50 EM, LT 96.9 62.1 Este estudio
a. Cuando se emplean dos métodos de extracción diferentes, el segundo valor representa la capa superficial de suelo extraída en el lavado.b. En Carimagua, la profundidad de muestreo varió estacionalmente hasta un máximo de 90 cm en la época seca.c. EM = extracción manual, LT = lavado-tamizado.
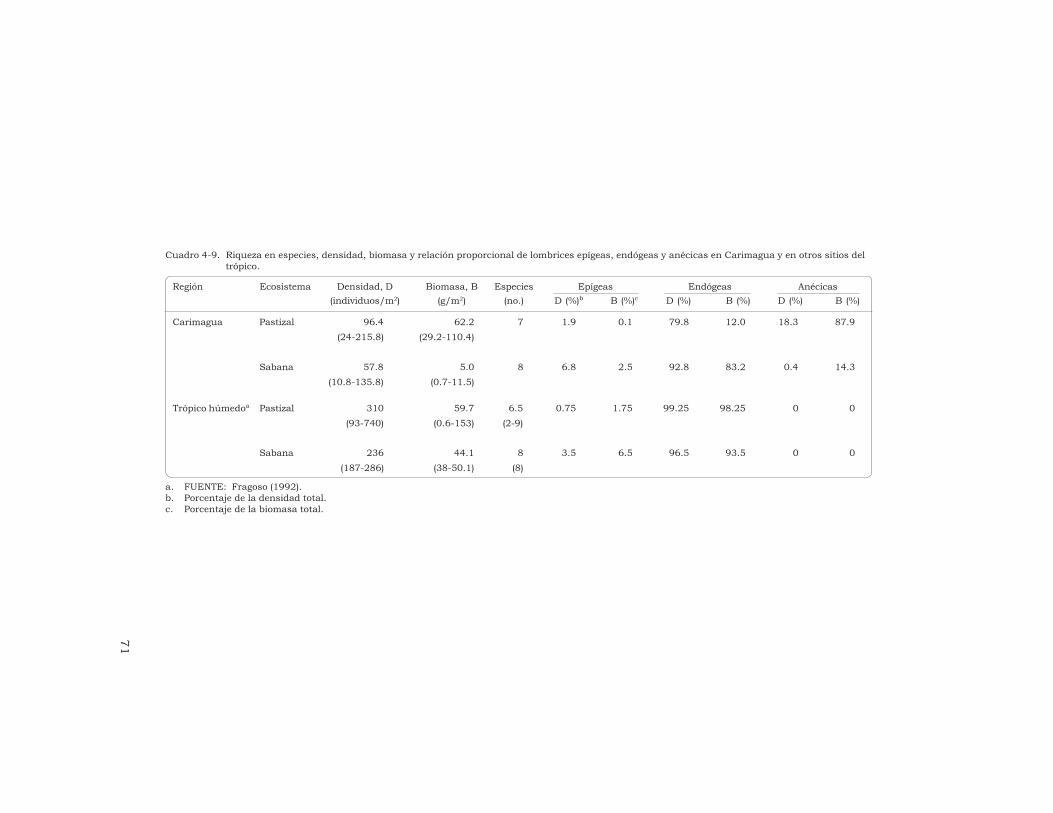
71
Las Com
unidades de Lombrices en las Sabanas N
ativas…
Cuadro 4-9. Riqueza en especies, densidad, biomasa y relación proporcional de lombrices epígeas, endógeas y anécicas en Carimagua y en otros sitios deltrópico.
Región Ecosistema Densidad, D Biomasa, B Especies Epígeas Endógeas Anécicas(individuos/m2) (g/m2) (no.) D (%)b B (%)c D (%) B (%) D (%) B (%)
Carimagua Pastizal 96.4 62.2 7 1.9 0.1 79.8 12.0 18.3 87.9(24-215.8) (29.2-110.4)
Sabana 57.8 5.0 8 6.8 2.5 92.8 83.2 0.4 14.3(10.8-135.8) (0.7-11.5)
Trópico húmedoa Pastizal 310 59.7 6.5 0.75 1.75 99.25 98.25 0 0(93-740) (0.6-153) (2-9)
Sabana 236 44.1 8 3.5 6.5 96.5 93.5 0 0(187-286) (38-50.1) (8)
a. FUENTE: Fragoso (1992).b. Porcentaje de la densidad total.c. Porcentaje de la biomasa total.

72
El Arado Natural:…
biomasa totales), con los valoresobtenidos en otros sitios del trópicohúmedo.
Barois et al. (1988) y Lavelle yPashanasi (1989) han reportadovalores de biomasa de 1000 a3000 kg/ha en los pastizalesmejorados. Ambas comunidadesestudiadas estaban dominadas porpoblaciones de una única especieexótica que aportaba del 80% al100% de la biomasa total. EnCarimagua, en cambio, la biomasadebida a las lombrices fue de620 kg/ha y una especie anécicacontribuyó con el 88% del valor deesa biomasa total.
La deposición de heces de M.carimaguensis estuvo limitada a laépoca lluviosa, un resultado queGates (1961) y Madge (1969)reportaron también para otrasespecies tropicales de lombrices.Según Edwards y Bohlen (1996), elrecuento de las heces superficialesda una idea de la actividad de laslombrices aunque no es, enabsoluto, un método válido de laabundancia real y de la actividadtotal de las lombrices. En esteestudio se demuestra que existe unarelación muy estrecha entre elnúmero de heces frescas depositadaspor M. carimaguensis en la superficiedel suelo y el número de individuosque habitan en los primeros 10 cmde suelo. Por tanto, la densidad depoblación puede calcularse, enalgunos casos, a partir del númerode heces frescas. Evans y Guild(1947) observaron también unafuerte correlación entre dos especiesde lombrícidos, Allolobophora longa yA. caliginosa. Por otro lado, Lavelle(1978) indicó que el 1.7% y el 3.5%del suelo total ingerido,respectivamente, por Millsoniaanomala (Megascolecidae) y porlombrices pequeñas de la familiaEudrilidae, se deposita como hecessuperficiales.
Aunque se encontraron grandesdiferencias respecto a la deposiciónde heces frescas de M. carimaguensisen uno u otro de los sistemasestudiados, el peso total de las hecesno fue muy diferente en ambossistemas. Cabria una explicaciónalterna para este resultado: o existequizás en el pastizal mejorado unareingestión de las heces después dela incubación microbiana que ocurreen ellas —lo que es considerado porSwift et al. (1979) como un “rumenexterno”— o esa especie es másactiva en la sabana y produce asímayor cantidad de heces por unidadde biomasa. Lavelle et al. (1989)mostraron, por su parte, que laslombrices ingerían mayor cantidadde suelo cuando se alimentaban ensuelos poco fértiles.
Es preciso investigar lo siguiente:la mayoría de los pastizales delneotrópico dan pruebas evidentes desu progresiva degradación, y locontrario ocurre en Carimagua,donde no hay hasta la fecha ningunaprueba de esa degradación. Esteinterrogante permanece aún sinrespuesta y se sabe muy poco sobrelos procesos que contribuyen a lasostenibilidad o a la degradación delos pastizales en el trópico (Fisher etal. 1995). Por otra parte, hay queconsiderar la posible contribución delas especies endémicas de lombricesadaptables a los cambios queacompañan el establecimiento de unpastizal, con el fin de diseñarprácticas agrícolas que causen unimpacto dañino mínimo.
AgradecimientosEste estudio fue incorporado a unesquema de colaboración en lainvestigación en que intervinieron elproyecto Macrofauna STD-3 —unprograma internacionalsubvencionado por la Comunidad

73
Las Comunidades de Lombrices en las Sabanas Nativas…
Europea y enfocado a la biología, laecología y las posibilidades demanejo de las lombrices presentes enlos ecosistemas naturales yperturbados del trópico— y elPrograma de Trópico Bajo(actualmente, Unidad de Suelos yNutrición de Plantas) del CentroInternacional de Agricultura Tropical(CIAT), situado en Cali, Colombia.Este estudio fue subvencionado, enparte, por una beca proveniente delprograma MACROFAUNA.
Los autores agradecen al CIAT,especialmente a todo el personal delantiguo Programa de Trópico Bajo,por el apoyo científico, técnico yhumano recibido. Expresan tambiénsu sincero agradecimiento por lavaliosa ayuda de los trabajadores deCarimagua en el duro trabajorealizado. Finalmente, el primerautor desea agradecer loscomentarios útiles que recibió de dosopiniones anónimas emitidas sobreel manuscrito preliminar.
ReferenciasBarbault R. 1992. Ecologie des
peuplements: Structure, dynamiqueet évolution. Masson, París. 273 p.
Barois I; Cadet P; Albrecht A; Lavelle P.1988. Systèmes de culture et faunedes sols: Quelques données. En:Feller C, ed. Fertilité des sols dansles agricultures paysannescaribéenes; Effets des restitutionsorganiques. ORSTOM-Martinique.París, Francia.
Barros ME; Blanchart E; Neves A;Desjardins T; Chauvel A; Lavelle P.1996. Relação entre a macrofauna eagregação do solo em tres sistemasna Amazonia central. En: Solo/Suelo, XII CongressoLatinoamericano de Ciencia doSolo, Aguas de Lindoia, Brasil.
Bouché MB 1972. Lombriciens de France.Ecologie et systematique. I.N.R.A.,París. 671 p.
Dash MC; Patra UC. 1977. Density,biomass and energy budget of atropical earthworm population froma grassland site in Orissa, India.Rev Ecol Biol Sol 14:461-471.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30(4):157-168.
Edwards CA; Bohlen PJ. 1996. Biologyand ecology of earthworms. 3a. ed.Chapman and Hall, Reino Unido.426 p.
Evans AC; Guild WJ. 1947. Method ofstudying the burrowing activity ofearthworms. Ann Mag Nat Hist11(14):643-650.
Fisher MJ; Rao IM; Thomas RJ; LascanoC. 1995. Grasslands in the well-watered tropical lowlands. En:Hodgson J; Illius AW, eds. Theecology and management of grazingsystems. CAB International, ReinoUnido. p 393-425.
Fragoso C. 1992. EWDATA BASE: asurvey of communities and selectedspecies of earthworms for theirpotential use in tropical agriculturalsystems. En: Lavelle P, ed.Conservation of soil fertility in lowinput agricultural systems of thehumid tropics by manipulatingearthworm communities. FinalReport, EEC/STD2 Programme.Comunidad Económica Europea(EEC), Bruselas, Bélgica.
Fragoso C. 1993. Les peuplements devers de terre dans l’est et sud-estdu Mexique. Thèse de doctorat,Université Paris 6. 228 p. + anexos.
Fragoso C; Lavelle P. 1992. Earthwormcommunities of tropical rainforests. Soil Biol Biochem24(12):1397-1408.
Gates G. 1961. Ecology of someearthworms with special referenceto seasonal activity. Am Midl Nat66:61-86.

74
El Arado Natural:…
IGAC (Instituto Geográfico AgustínCodazzi). 1974. Reconocimientogeneral de los suelos del CentroNacional de InvestigacionesAgropecuarias Carimagua(Departamento del Meta). Bogotá,DC.
Jiménez JJ; Moreno AG. Martiodriluscarimaguensis (Oligochaeta,Glossoscolecidae), una nuevaespecie de lombriz de tierra paraColombia. Megadrilogica. (Enrevisión.)
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Johnson RW; Tothill JC. 1985. Definitionand broad geographical outline ofsavanna lands. En: Tothill JC; MottJJ, eds. Ecology and managementof the world’s savannas. CAB, ReinoUnido. p 1-13.
Kouassi PK. 1987. Étude comparative dela macrofaune endogéed’écosystemes guinéens naturels ettransformés de Côte d’Ivoire. Tesisde doctorado. Université d’Abidjan,Costa de Marfil. 129 p.
Lascano CL; Estrada J. 1989. Long-termproductivity of legume-based andpure grass pastures in the EasternPlains of Colombia. En: Proceedingsof the XVI International GrasslandCongress, Niza, Francia. p 1179-1180.
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfonctions dans l’ècosystème. Tesisde doctorado. París VI. Publ LabZool ENS 12. 301 p.
Lavelle P. 1981. Strategies dereproduction chez les vers de terre.Acta Oecol 2:117-133.
Lavelle P. 1983. The soil fauna of tropicalsavannas. II. The earthworms. En:Bourlière F, ed. Tropical savannas.Elsevier, Amsterdam. p 485-504.
Lavelle P. 1986. Associations mutualistesavec la microflore du sol et richessespécifique sous les tropiques:l’hypothèse du premier maillon.Ecologie générale. C R Acad Sci SerIII 302(1):11-14.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lavelle P; Chauvel A; Fragoso C. 1995.Faunal activity in acid soils. En:Date RA, ed. Plant soil interactionsat low pH. Kluwer AcademicPublishers, Holanda. p 201-211.
Lavelle P; Maury ME; Serrano V. 1981.Estudio cuantitativo de la fauna delsuelo en la región de Laguna Verde(Veracruz, México); época de lluvias.En: Reyes P, ed. EstudiosEcológicos en el Trópico Mexicano.Instituto de Ecología, México.p 65-100.
Lavelle P; Schaefer R; Zaidi Z. 1989. Soilingestion and growth in Millsoniaanomala, a tropical earthworm, asinfluenced by the quality of theorganic matter ingested.Pedobiologia 33:379-388.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B; Brussaard L. 1994.The relationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. Wiley,Chichester, Reino Unido.p 137-169.
Lavelle P; Blanchart E; Martin A; MartinS; Spain A; Toutain F; Barois I;Schaefer R. 1993. A hierarchicalmodel for decomposition interrestrial ecosystems: applicationto soils of the humid tropics.Biotropica 25(2):130-150.

75
Las Comunidades de Lombrices en las Sabanas Nativas…
Lee K. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, NuevaYork. 411 p.
Ludwig JA; Reynolds JF. 1988. Statisticalecology: a primer on methods andcomputing. John Wiley and Sons,Nueva York. 337 p.
Madge DS. 1969. Field and laboratorystudies on the activities of twospecies of tropical earthworms.Pedobiologia 9(8):188-214.
Nemeth A. 1981. Estudio ecológico de laslombrices de tierra (Oligochaeta) enecosistemas de bosque húmedotropical en San Carlos de Río Negro,Territorio Federal Amazonas. Tesis.Universidad Central de Venezuela.92 p.
Pashanasi B; Meléndez G; Szott L; LavelleP. 1992. Effect of inoculation withthe endogeic earthworm Pontoscolexcorethrurus (Glossoscolecidae) on Navailability, soil microbial biomassand the growth of three tropicalfruit tree seedlings in a potexperiment. Soil Biol Biochem24:1655-1660.
Pielou EC. 1975. Ecological diversity.Wiley, Nueva York.
Rao IM; Zeigler RS; Vera R; Sarkarung S.1993. Selection and breeding foracid-soil tolerance in crops. Uplandrice and tropical forages as casestudies. Bioscience 43:454-465.
Rose CJ; Wood AW. 1980. Someenvironmental factors affectingearthworm populations and sweetpotato production on the TariBasin, Papua New Guineahighlands. Papua New Guinea AgricJ 31:1-13.
Sánchez P; Salinas JG. 1983. Low inputtechnology for managing Oxisolsand Ultisols in tropical America.Adv Agron 34:279-405.
Senapati BK. 1980. Aspects ofecophysiological studies on tropicalearthworms: distribution,population dynamics, production,energetics and their role in thedecomposition process. Tesis(Ph.D.). Sambalpur University,India. 154 p.
Shannon CE; Weaver W. 1949. Themathematical theory ofcommunication. University ofIllinois Press, Urbana, IL, EstadosUnidos.
Sombroek WG; Nachtergaele FO; Hebel A.1993. Amounts, dynamics andsequestering of carbon in tropicaland subtropical soils. Ambio22:417-426.
Swift MJ; Heal OW; Anderson JM. 1979.Decomposition in terrestrialecosystems: studies in ecology,Vol. 5. Blackwell, Oxford. 372 p.
Syers JK; Springett JA. 1984.Earthworms and soil fertility. PlantSoil 76:93-104.
Thomas R; Fisher MJ; Ayarza M; Sanz JI.1995. The role of forage grasses andlegumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. Advances inSoil Science Series, Lewispublishers, Boca Ratón, EstadosUnidos. p 61-83.

76
El Arado Natural:…
ResumenMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae) esuna lombriz nativa, grande, dehábito anécico, gris oscura por eldorso, que se encontró en lassabanas ácidas, tanto naturalescomo perturbadas, de los LlanosOrientales de Colombia. Desde abrilde 1994 hasta septiembre de 1995(excepto en junio de 1994) se estudióla dinámica de poblaciones de estaespecie en una sabana nativa y enun pastizal introducido sometido apastoreo durante 17 años, en que seasociaron una gramínea y unaleguminosa forrajera. La densidad yla biomasa fueron mayores en elpastizal que en la sabana. Seencontraron también diferencias enel número de heces frescasdepositadas sobre la superficie del
suelo: en el pastizal introducidohabía 38.4 x 103 de estas heces porhectárea, o sea, 11 veces más que localculado para la sabana. Se hallóuna correlación muy estrecha entreel número de individuos deM. carimaguensis y el de hecesfrescas superficiales. Se encontraronun patrón de adaptación a laestación seca en la población adultay otro diferente en la poblaciónjuvenil de esta especie. Los adultosson activos durante 8 meses,mientras que los juveniles inician unperíodo de diapausa de 3 a 4 mesesantes del final de la etapa deactividad de los adultos. Ladistribución vertical de la poblaciónde lombrices de tierra reflejó tambiénlos fuertes cambios estacionales delecosistema.
Palabras clave: dinámica depoblaciones, estrategiasadaptativas, Glossoscolecidae,Martiodrilus carimaguensis,Oligochaeta, sabanas
IntroducciónLa información disponible sobre labiología de las lombrices tropicaleses escasa (Dash y Patra 1977;Lavelle 1978). Pocas especies hansido estudiadas en detalle, tanto enlos ecosistemas de latitudestempladas como en los tropicales; de
CAPÍTULO 5
Dinámica de Poblaciones y EstrategiasAdaptativas de Martiodriluscarimaguensis (Oligochaeta,Glossoscolecidae), una Especie Nativade las Sabanas Bien Drenadas deColombia1
J. J. Jiménez* , A. G. Moreno*, P. Lavelle** y T. Decaëns***
1. Publicado en Applied Soil Ecology 9(1-3),p 153-160, por Jiménez et al. Copyright ©1998 de Elsevier. Impreso con permiso de lacasa editorial.
* Departamento de Biología Animal I(Zoología), Facultad de Biología, UniversidadComplutense de Madrid, 28040, Madrid,España.
** Laboratoire d’Ecologie et Biologie des SolsTropicaux, ORSTOM, 72, route d’Aulnay,93143 Bondy Cedex, Francia.
*** Laboratoire d’Ecologie Vegetal, UFRSciences/Unidad de Suelos y Nutrición dePlantas, CIAT, Université de Rouen, 76821Mont Saint Aignan Cedex, Francia.

Dinámica de Poblaciones y Estrategias Adaptativas...
77
ellas, la mayoría han sidointroducidas por el hombre, porejemplo, Pontoscolex corethrurusMüller y Dichogaster bolaui(Michaelsen). En la mayor parte delas áreas perturbadas, lascomunidades de lombrices localesdesaparecen cuando son remplazadaspor especies introducidas (Bohlen etal. 1995; Lavelle y Pashanasi 1989;Lavelle et al. 1987).
Ahora bien, poca atención se haprestado al papel que hacen lasespecies nativas que se adaptan biena las perturbaciones experimentadaspor los ecosistemas naturales. EnCarimagua, los pastizalesintroducidos en las sabanasnaturales han conservado labiodiversidad original de laslombrices y su biomasa llegó a 51.1 gPF/m2 desde los 4.8 que tenía lasabana nativa (Decaëns et al. 1994).Esta clase de manejo que se le dio ala tierra favoreció a una especie,Martiodrilus carimaguensis Jiménez yMoreno, cuya densidad y biomasaaumentaron significativamente(P < 0.001).
M. carimaguensis es una especieanécica grande, perteneciente a lafamilia Glossoscolecidae; tiene, enpromedio, 9.3 mm de diámetro y194.3 mm de longitud y un pesofresco (en formalina al 4%) de 11.2 g(n = 29). El color del cuerpo deladulto es gris oscuro por la partedorsal y gris claro por la ventral(Jiménez y Moreno, en revisión).
Materiales y Métodos
Sitio del estudioLa zona en que se hizo este estudio seencuentra en la estaciónexperimental Carimagua de CIAT-CORPOICA (resultado de un convenioentre el Centro Internacional de
Agricultura Tropical y la CorporaciónColombiana de InvestigaciónAgropecuaria), la cual está asentadaen las sabanas isohipertérmicas biendrenadas de los Llanos Orientales deColombia (4°37' N y 71°19' O, a175 msnm). Como medias anuales,la precipitación es de 2280 mm y latemperatura de 26 °C; hay unaépoca seca entre diciembre y marzo.Los suelos son de dos tipos:Oxisoles de baja fertilidad en laszonas altas de la sabana (los “altos”)y Ultisoles en las zonas bajas (los“bajos”). Los Oxisoles secaracterizan por su acidez alta(pH(H2O) = 4.5), una saturación alta deAl (> 90%) y un contenido bajo de loscationes intercambiables Ca, Mg y K.Los factores químicos quecontribuyen a la infertilidad del sueloácido y a la aparición de los efectosderivados de él en el crecimientovegetal son bastante complejos; entreellos están la toxicidad debida al Al,el bajo contenido de P y las tasasbajas de mineralización del N (Rao etal. 1993).
Se evaluaron dos sistemasdiferentes de uso de la tierra quepresentan contrastes: una sabananatural (SN) sin ningunaintervención humana, en la que lasplantas Andropogon bicornis,Gymnopogon sp., Panicum spp.,Trachypogon spp. e Imperata sp.dominaban la vegetación, y unpastizal mejorado (PM) de 17 años enque se asociaban la gramíneaafricana Brachiaria decumbens cv.Basilisk y la leguminosa herbáceaforrajera Pueraria phaseoloides CIAT9900 (“kudzú”).
Muestreo de lombricesEn cada sistema se delimitó un áreade 90 x 90 m, que fue dividida encuadrados regulares de 10 x 10 m.Durante 17 meses, las lombrices serecogieron cada mes manualmente

78
El Arado Natural:…
de 5 monolitos de suelo de 1 x 1 x0.5 m (según Lavelle 1978), quefueron extraídos en 5 cuadradosescogidos al azar en cada sistema.La profundidad de muestreo cambióestacionalmente a causa de lasmigraciones verticales de la especieestudiada. La muestra se subdividíaen capas de 10 cm y todas laslombrices recogidas en cada capa selavaban en agua y se fijaban enformalina al 4%. En el laboratorio seseparaban los adultos (que teníanclitelo y glándulas sexualesasociadas) de los individuos jóvenes(sin clitelo ni glándulas); ambos secontaban y se pesaban, lo mismoque los capullos.
Deposición de hecessuperficialesComo especie anécica,M. carimaguensis puede depositar,sobre la superficie del suelo, hecesde gran tamaño con forma de torreque pueden alcanzar 15 cm dealtura. En ambos sistemas demanejo de la tierra se escogieron alazar dos monolitos de 1 m2, y enellos se contaron estas hecessuperficiales, fácilmentedistinguibles de otras deposiciones, yse clasificaron en dos categorías:frescas y secas. Las heces sesecaron al horno a 60 ºC durante72 horas y se pesaron.
Resultados
Densidad y biomasaLa densidad media de laspoblaciones de M. carimaguensisvarió en la sabana desde0 individuos/m2 en noviembre de1994 hasta 1.06 individuos/m2 enseptiembre de 1995; en el pastizalvarió desde 11 individuos/m2 enseptiembre de 1994 hasta
23.2 individuos/m2 en mayo de 1995(Figura 5-1). Los valores más altosde densidad se registraron alcomienzo de la época lluviosa y elincremento de la densidad enoctubre de 1994 se debió a laeclosión de nuevos jóvenes. Ladensidad media en el pastizalintroducido y en la sabana nativafue, respectivamente, 17.9 y0.2 individuos/m2.
En la sabana, la biomasa oscilóentre 0.24 g/m2 (en marzo de 1994)y 8.76 g/m2 (en septiembre de 1995).En el pastizal, la biomasa variódesde 26.5 g/m2 (enero de 1995)hasta 94.8 g/m2 (mayo de 1994).Hubo una correlación significativaentre la biomasa media mensual y laprecipitación, aunque sólo en elpastizal introducido (r = 0.821;P < 0.01). Los valores de biomasamás altos se obtuvieron al comienzode la época lluviosa, cuando toda lapoblación estaba activa. Durantejulio y agosto, la biomasa descendióporque los individuos juveniles seinactivaron. Al final de la temporadade lluvias, la biomasa se redujo enun 50% porque los adultosdescendieron a cierta profundidad enel suelo y se inactivaron tras vaciarsu contenido intestinal.
En la sabana nativa,M. carimaguensis representó el15.1% de la biomasa total delombrices y en el pastizal introducidoel 85.1%. Otras cinco especiesnativas, cuatro endógeas y unaepígea, aportaron el resto de labiomasa.
No se encontró ninguna relaciónentre la densidad media mensual depoblación y la humedad del suelo; sehalló, en cambio, correlaciónsignificativa entre la humedad delsuelo y la biomasa (r = 0.651;P < 0.01) y entre dicha humedad y elporcentaje de actividad de lapoblación total (r = 0.673; P < 0.01).
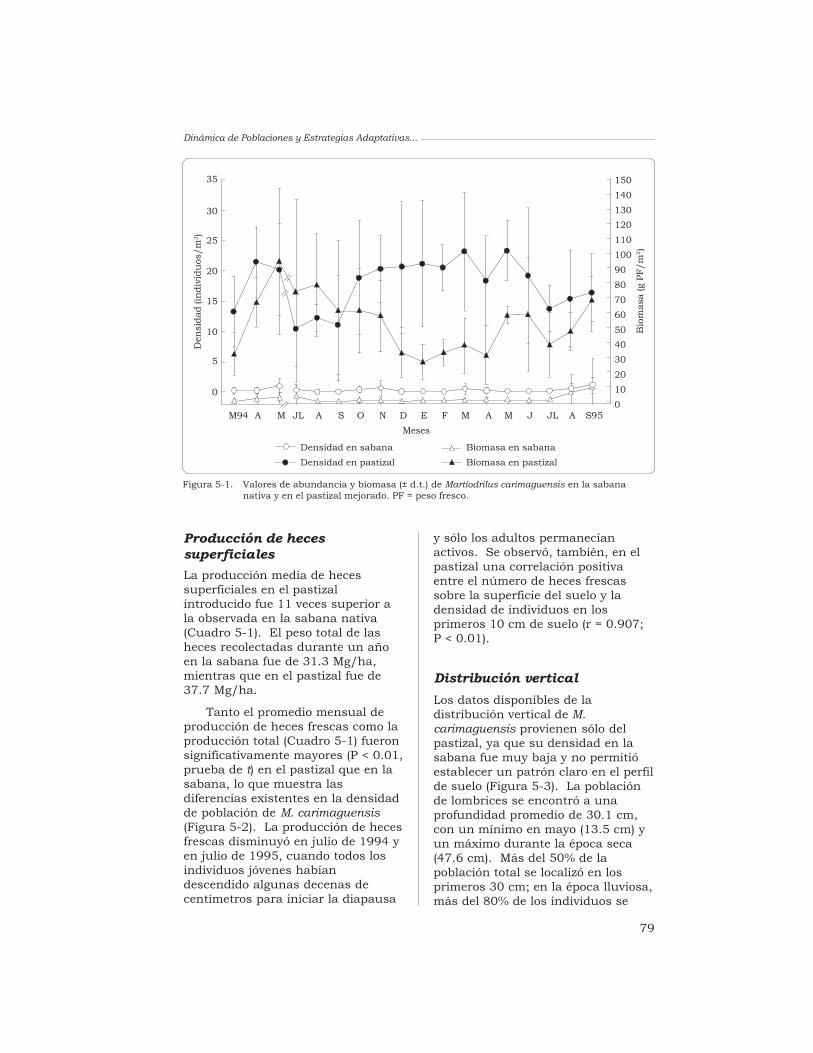
Dinámica de Poblaciones y Estrategias Adaptativas...
79
Producción de hecessuperficialesLa producción media de hecessuperficiales en el pastizalintroducido fue 11 veces superior ala observada en la sabana nativa(Cuadro 5-1). El peso total de lasheces recolectadas durante un añoen la sabana fue de 31.3 Mg/ha,mientras que en el pastizal fue de37.7 Mg/ha.
Tanto el promedio mensual deproducción de heces frescas como laproducción total (Cuadro 5-1) fueronsignificativamente mayores (P < 0.01,prueba de t) en el pastizal que en lasabana, lo que muestra lasdiferencias existentes en la densidadde población de M. carimaguensis(Figura 5-2). La producción de hecesfrescas disminuyó en julio de 1994 yen julio de 1995, cuando todos losindividuos jóvenes habíandescendido algunas decenas decentímetros para iniciar la diapausa
y sólo los adultos permanecíanactivos. Se observó, también, en elpastizal una correlación positivaentre el número de heces frescassobre la superficie del suelo y ladensidad de individuos en losprimeros 10 cm de suelo (r = 0.907;P < 0.01).
Distribución verticalLos datos disponibles de ladistribución vertical de M.carimaguensis provienen sólo delpastizal, ya que su densidad en lasabana fue muy baja y no permitióestablecer un patrón claro en el perfilde suelo (Figura 5-3). La poblaciónde lombrices se encontró a unaprofundidad promedio de 30.1 cm,con un mínimo en mayo (13.5 cm) yun máximo durante la época seca(47.6 cm). Más del 50% de lapoblación total se localizó en losprimeros 30 cm; en la época lluviosa,más del 80% de los individuos se
1501401301201101009080706050403020100
Bio
mas
a (g
PF/
m2 )
M94 A M JL A S O N D E F M A M J JL A S95
35
30
25
20
15
10
5
0
Figura 5-1. Valores de abundancia y biomasa (± d.t.) de Martiodrilus carimaguensis en la sabananativa y en el pastizal mejorado. PF = peso fresco.
Densidad en sabana Biomasa en sabanaDensidad en pastizal Biomasa en pastizal
Meses
Den
sida
d (in
divi
duos
/m2 )

80
El Arado Natural:…
encontraban próximos a lasuperficie, donde el contenido demateria orgánica era mayor. Loscapullos fueron depositados a unaprofundidad de 20 a 50 cm.
Adaptación a la época secaMartiodrilus carimaguensis exhibióun comportamiento interesante queincluyó un fenómeno de diapausaverdadera, aunque con patronesdiferentes entre los adultos y losindividuos jóvenes. Estos últimosestuvieron activos sólo durante4 meses, de abril a julio, de modoque iniciaron la diapausa muchoantes que los adultos: éstos, en
efecto, permanecieron activos hastadiciembre (8 meses). La inactividadcomenzó cuando los individuosdescendieron hasta una profundidadde 60 a 100 cm; cada uno de ellosconstruyó una cámara de estivaciónal final de su galeríasemipermanente, en la que seenrollaron tras vaciar su contenidointestinal y suspendieron todaactividad hasta el inicio de lasiguiente época lluviosa.Generalmente, el final de la galeríaestaba sellado con varios tabiqueselaborados con heces para impedir lapérdida de humedad del tegumento,la cual es vital para mantener unatasa mínima de respiración. El
Cuadro 5-1. Producción media mensual de heces de Martiodrilus carimaguensis por m2 (valores ± d.t.)en los dos sistemas estudiados (sabana y pastizal).
Condición Sabana nativa Pastizal mejorado
Epoca lluviosa Período Epoca lluviosa Períodode estudio de estudio
Heces secas 7.15 ± 3.5 6.38 ± 3.4 20.38 ± 7.9 17.35 ± 8.9
Heces frescas recientes 0.31 ± 0.5 0.23 ± 0.5 3.25 ± 3.8 2.48 ± 3.6
Heces frescas no recientesa 0.12 ± 0.2 0.09 ± 0.21 1.77 ± 2.1 1.39 ± 2.0
Total 7.58 ± 3.6 6.72 ± 3.6 25.43 ± 10.3 21.21 ± 11.9
a. Deposición fresca con la base seca.
de las heces
Figura 5-2. Valor medio mensual (± d.t.) de la cantidad de heces frescas de M. carimaguensis por m2.
Hec
es fr
esca
s/m
2
24
20
16
12
8
4
0A94 M JL A S O N D E F M A M J JL A S95
MesesSabana nativa Pastizal introducido
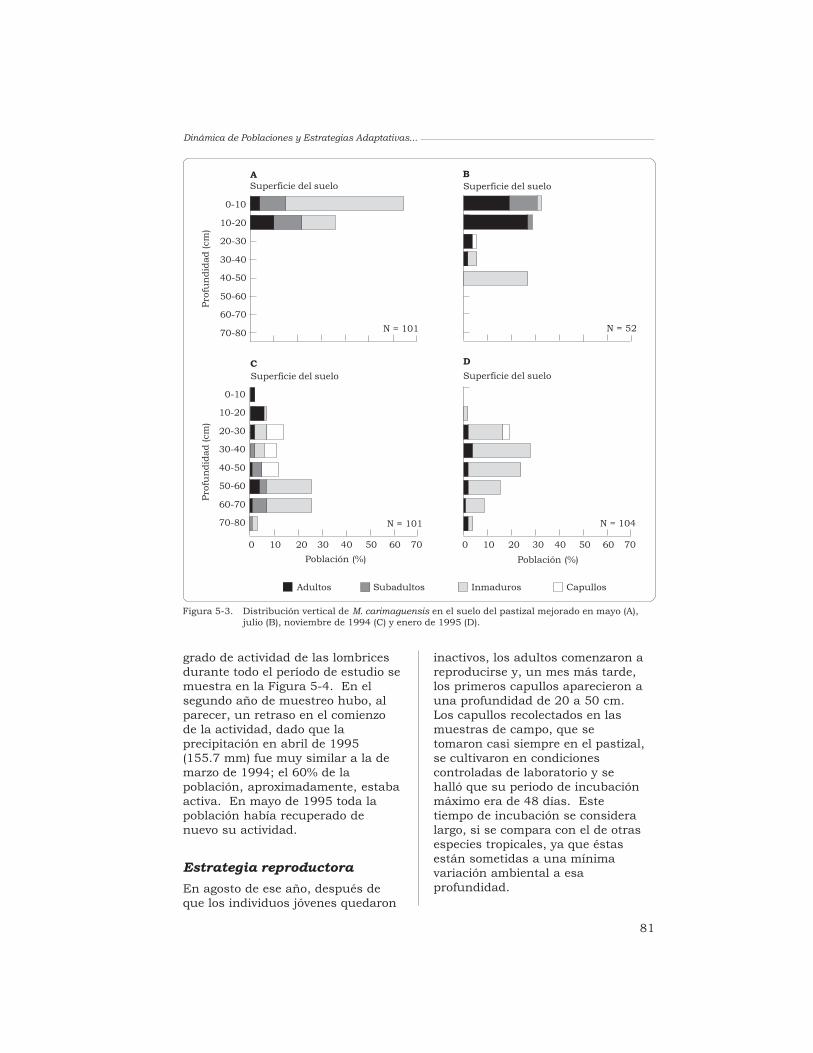
Dinámica de Poblaciones y Estrategias Adaptativas...
81
grado de actividad de las lombricesdurante todo el período de estudio semuestra en la Figura 5-4. En elsegundo año de muestreo hubo, alparecer, un retraso en el comienzode la actividad, dado que laprecipitación en abril de 1995(155.7 mm) fue muy similar a la demarzo de 1994; el 60% de lapoblación, aproximadamente, estabaactiva. En mayo de 1995 toda lapoblación había recuperado denuevo su actividad.
Estrategia reproductoraEn agosto de ese año, después deque los individuos jóvenes quedaron
inactivos, los adultos comenzaron areproducirse y, un mes más tarde,los primeros capullos aparecieron auna profundidad de 20 a 50 cm.Los capullos recolectados en lasmuestras de campo, que setomaron casi siempre en el pastizal,se cultivaron en condicionescontroladas de laboratorio y sehalló que su periodo de incubaciónmáximo era de 48 días. Estetiempo de incubación se consideralargo, si se compara con el de otrasespecies tropicales, ya que éstasestán sometidas a una mínimavariación ambiental a esaprofundidad.
Población (%)
Prof
un
dida
d (c
m)
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
Prof
un
dida
d (c
m)
0 10 20 30 40 50 60 70 0 10 20 30 40 50 60 70Población (%)
A
C
B
N = 101 N = 52
N = 104N = 101
Adultos Subadultos Inmaduros Capullos
Figura 5-3. Distribución vertical de M. carimaguensis en el suelo del pastizal mejorado en mayo (A),julio (B), noviembre de 1994 (C) y enero de 1995 (D).
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
D
Superficie del suelo
Superficie del suelo
Superficie del suelo
Superficie del suelo
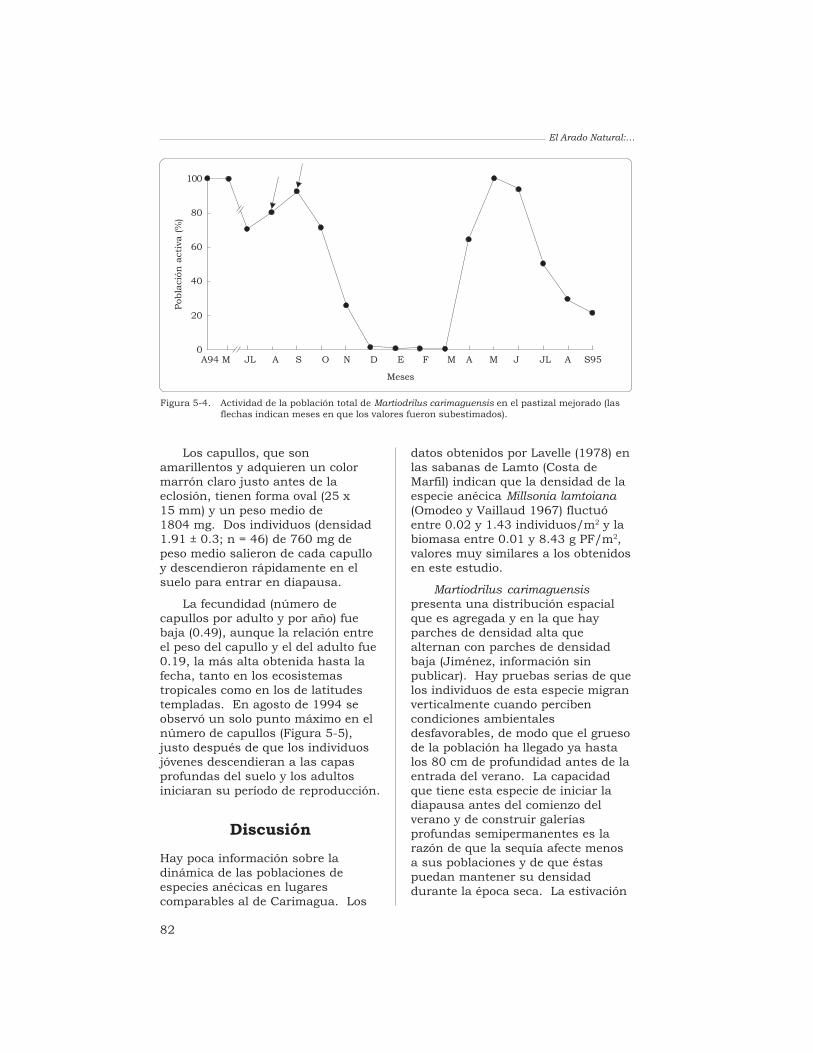
82
El Arado Natural:…
Los capullos, que sonamarillentos y adquieren un colormarrón claro justo antes de laeclosión, tienen forma oval (25 x15 mm) y un peso medio de1804 mg. Dos individuos (densidad1.91 ± 0.3; n = 46) de 760 mg depeso medio salieron de cada capulloy descendieron rápidamente en elsuelo para entrar en diapausa.
La fecundidad (número decapullos por adulto y por año) fuebaja (0.49), aunque la relación entreel peso del capullo y el del adulto fue0.19, la más alta obtenida hasta lafecha, tanto en los ecosistemastropicales como en los de latitudestempladas. En agosto de 1994 seobservó un solo punto máximo en elnúmero de capullos (Figura 5-5),justo después de que los individuosjóvenes descendieran a las capasprofundas del suelo y los adultosiniciaran su período de reproducción.
DiscusiónHay poca información sobre ladinámica de las poblaciones deespecies anécicas en lugarescomparables al de Carimagua. Los
datos obtenidos por Lavelle (1978) enlas sabanas de Lamto (Costa deMarfil) indican que la densidad de laespecie anécica Millsonia lamtoiana(Omodeo y Vaillaud 1967) fluctuóentre 0.02 y 1.43 individuos/m2 y labiomasa entre 0.01 y 8.43 g PF/m2,valores muy similares a los obtenidosen este estudio.
Martiodrilus carimaguensispresenta una distribución espacialque es agregada y en la que hayparches de densidad alta quealternan con parches de densidadbaja (Jiménez, información sinpublicar). Hay pruebas serias de quelos individuos de esta especie migranverticalmente cuando percibencondiciones ambientalesdesfavorables, de modo que el gruesode la población ha llegado ya hastalos 80 cm de profundidad antes de laentrada del verano. La capacidadque tiene esta especie de iniciar ladiapausa antes del comienzo delverano y de construir galeríasprofundas semipermanentes es larazón de que la sequía afecte menosa sus poblaciones y de que éstaspuedan mantener su densidaddurante la época seca. La estivación
A94 M JL A S O N D E F M A M J JL A S95
100
80
60
40
20
0
Meses
Pobl
ació
n a
ctiv
a (%
)
Figura 5-4. Actividad de la población total de Martiodrilus carimaguensis en el pastizal mejorado (lasflechas indican meses en que los valores fueron subestimados).
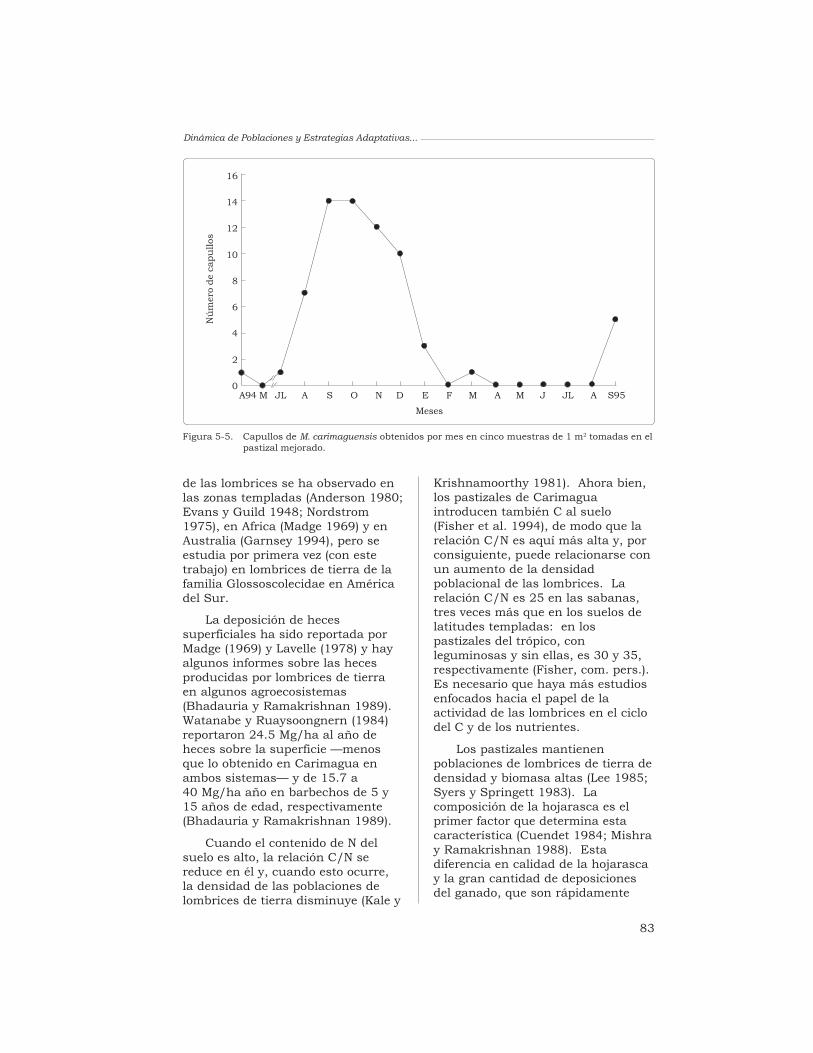
Dinámica de Poblaciones y Estrategias Adaptativas...
83
de las lombrices se ha observado enlas zonas templadas (Anderson 1980;Evans y Guild 1948; Nordstrom1975), en Africa (Madge 1969) y enAustralia (Garnsey 1994), pero seestudia por primera vez (con estetrabajo) en lombrices de tierra de lafamilia Glossoscolecidae en Américadel Sur.
La deposición de hecessuperficiales ha sido reportada porMadge (1969) y Lavelle (1978) y hayalgunos informes sobre las hecesproducidas por lombrices de tierraen algunos agroecosistemas(Bhadauria y Ramakrishnan 1989).Watanabe y Ruaysoongnern (1984)reportaron 24.5 Mg/ha al año deheces sobre la superficie —menosque lo obtenido en Carimagua enambos sistemas— y de 15.7 a40 Mg/ha año en barbechos de 5 y15 años de edad, respectivamente(Bhadauria y Ramakrishnan 1989).
Cuando el contenido de N delsuelo es alto, la relación C/N sereduce en él y, cuando esto ocurre,la densidad de las poblaciones delombrices de tierra disminuye (Kale y
Krishnamoorthy 1981). Ahora bien,los pastizales de Carimaguaintroducen también C al suelo(Fisher et al. 1994), de modo que larelación C/N es aquí más alta y, porconsiguiente, puede relacionarse conun aumento de la densidadpoblacional de las lombrices. Larelación C/N es 25 en las sabanas,tres veces más que en los suelos delatitudes templadas: en lospastizales del trópico, conleguminosas y sin ellas, es 30 y 35,respectivamente (Fisher, com. pers.).Es necesario que haya más estudiosenfocados hacia el papel de laactividad de las lombrices en el ciclodel C y de los nutrientes.
Los pastizales mantienenpoblaciones de lombrices de tierra dedensidad y biomasa altas (Lee 1985;Syers y Springett 1983). Lacomposición de la hojarasca es elprimer factor que determina estacaracterística (Cuendet 1984; Mishray Ramakrishnan 1988). Estadiferencia en calidad de la hojarascay la gran cantidad de deposicionesdel ganado, que son rápidamente
A94 M JL A S O N D E F M A M J JL A S95
16
14
12
10
8
6
4
2
0
Meses
Nú
mer
o de
cap
ullo
s
Figura 5-5. Capullos de M. carimaguensis obtenidos por mes en cinco muestras de 1 m2 tomadas en elpastizal mejorado.

84
El Arado Natural:…
incorporadas en el suelo por estaespecie de lombriz y por algunosescarabajos, serían los factoresresponsables del incremento de laactividad de M. carimaguensis en lospastizales mejorados.
Hay que tener en cuenta elcomportamiento anécico de M.carimaguensis, que es de granimportancia en relación con el ciclodel C y de otros nutrientes. Elpastizal que contiene unaleguminosa necesita el nitrógenosuministrado por ésta. Se haobservado también que las raíces dela gramínea y de la leguminosacolonizan las heces situadas en elinterior de las galerías y que unalombriz oligohúmica pequeña, de lafamilia Ocnerodrilidae, se alimentade estas heces (Jiménez et al. 1994).No hay datos, hasta la fecha, sobrelas interrelaciones que hay entre laslombrices, las heces depositadas ensus galerías y las raíces que seaproximan a a estas heces en elperfil del suelo; tampoco los haysobre el papel de las lombrices en elciclo del C y de los nutrientes, en lospastizales en que se asociangramíneas y leguminosas. Losautores creen que estos estudiosserán muy relevantes si puedendemostrar que M. carimaguensis esresponsable, en parte, delcrecimiento de estas raíces y queestaría contribuyendo, por tanto, a laacumulación de C en el suelo.
Aunque en la sabana nativa esbaja la densidad de esta especie ypocas las heces frescas halladas ensu superficie, el peso total de lasheces recolectadas en ella duranteun año no difiere mucho del obtenidoen un pastizal mejorado. Es posibleque, en este último sistema, laintensa actividad de las lombrices setraduzca en una reingestión de suspropias heces después de que éstas
han experimentado una incubaciónmicrobiana (Swift et al. 1979).
Finalmente, los pastizalesintroducidos presentan una fuerteactividad macrobiológica ymicrobiológica que mejora suspropiedades físicas y químicas, esdecir, los parámetros de calidad delsuelo. En estudios desarrolladosrecientemente en Carimagua, se haconsiderado la posibilidad demanejar los agroecosistemas demanera que las áreas donde laactividad de las lombrices es intensa,o sea, los pastizales introducidos, sesitúen adyacentes a los cultivosanuales para facilitar la difusión depoblaciones de lombrices hacia elloscon el fin de que los colonicen.Lavelle et al. (1994) han logradoavanzar en la comprensión de lasactividades de la fauna del suelolocal y en el uso potencial de lasespecies nativas en losagroecosistemas tropicales; convieneahora sugerir que se hagan másestudios para considerar lainfluencia de dicha actividadbiológica en las propiedades físicas yquímicas del suelo.
AgradecimientosEste trabajo fue sostenido por unasubvención para investigación delproyecto Macrofauna (STD3,programa de la UE), por la cualestamos profundamente agradecidos.Queremos agradecer también al CIAT(Centro Internacional de AgriculturaTropical), especialmente al antiguoPrograma de Trópico Bajo, por suapoyo humano y técnico y por lasdiscusiones científicas surgidas deeste estudio. Nuestro más profundoagradecimiento para la gente deCarimagua y, especialmente, para losoperarios de campo por suinestimable ayuda.

Dinámica de Poblaciones y Estrategias Adaptativas...
85
ReferenciasAnderson C. 1980. The influence of
climatic conditions on activity andvertical distribution of earthwormsin a Danish arable soil. K Vet-Landbohojsk. Arsskrift 57-68.
Bhadauria T; Ramakrishnan PS. 1989.Earthworm population dynamicsand contribution to nutrient cyclingduring cropping and fallow phasesof shifting agriculture (Jhum) inNorth-east India. J Appl Ecol26:505-520.
Bohlen PJ; Edwards WM; Edwards CA.1995. Earthworm communitystructure and diversity inexperimental agriculturalwatersheds in Northeastern Ohio.Plant Soil 170:233-239.
Bouché M. 1972. Lombriciens de France.Ecologie et Systematique. I.N.R.A.,París. 671 p.
Cuendet G. 1984. A comparative study ofthe earthworm population of fourdifferent woodland types in Wythamwoods, Oxford. Pedobiologia26:421-439.
Dash MC; Patra UC. 1977. Density,biomass and energy budget of atropical earthworm population froma grassland site in Orissa, India.Rev Ecol Biol Sol 14:461-471.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30(4):157-168.
Evans AC; Guild WJ. 1948. Studies onthe relationships betweenearthworms and soil fertility. IV. Onthe life cycles of some BritishLumbricidae. Ann Appl Biol35(4):473-493.
Fisher MJ; Rao IM; Ayarza MA; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage by introduceddeep-rooted grasses in the SouthAmerican savannas. Nature371:236-238.
Garnsey RB. 1994. Seasonal activity andaestivation of Lumbricidearthworms in the Midlands ofTasmania. Aust J Soil Res32:1355-1367.
Jiménez JJ; Decaëns T; Moreno AG;Rossi J-P; Lavelle P. 1994.Dynamics and short-term effects ofearthworms in natural andmanaged savannas of the EasternPlains of Colombia. En: Lavelle P,ed. Conservation of soil fertility inlow-input agricultural systems ofthe humid tropics by manipulatingearthworm communities(Macrofauna Project, STD2 EC).p 52-61.
Jiménez JJ; Moreno AG. Martiodriluscarimaguensis sp. nov. (Oligochaeta,Glossoscolecidae), una nuevaespecie de lombriz de tierra paraColombia. Megadrilogica. (Enrevisión.)
Kale RD; Krishnamoorthy RV. 1981. Whataffects the abundance and diversityof earthworms in soils? Proc IndianAcad Sci 90(1):117-121.
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfonctions dans l’ècosystème. Tesis(Doctorado). París VI. Publ. Lab.Zool. ENS 12, París. 301 p.
Lavelle P. 1981. Stratégies dereproduction chez les vers de terre.Acta Oecol 2(2):117-133.
Lavelle P. 1983. The soil fauna of tropicalsavannas. II. The earthworms. En:Bourlière F, ed. Tropical savannas.Elsevier Scientific PublishingCompany, Países Bajos. p 485-504.
Lavelle P; Barois I; Cruz I; Fragoso C;Hernández A; Pineda A; Rangel P.1987. Adaptive strategies of P.corethrurus (Glossoscolecidae,Oligochaeta), a peregrinegeophagous earthworm of thehumid tropics. Biol Fertil Soils5:188-194.

86
El Arado Natural:…
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B; Brussard L. 1994.The relationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. John Wiley &Sons, Chichester, Reino Unido.p 137-169.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lee K. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, NuevaYork. 411 p.
Madge DS. 1969. Field and laboratorystudies on the activities of twospecies of tropical earthworms.Pedobiologia 9:188-214.
Mishra KC; Ramakrishnan PS. 1988.Earthworm population dynamics indifferent jhum fallows developedafter slash and burn agriculture innorth-eastern India. Proc IndianAcad Sci 97(4):309-318.
Nordström S. 1975. Seasonal activity oflumbricids in Southern Sweden.Oikos 26:307-315.
Omodeo P; Vaillaud M. 1967. Lesoligochètes de la Savanna deGpakobo en Côte d’Ivoire. Bull IFAN29 Sér A 3:925-944.
Rao IM; Zeigler RS; Vera R; Sakarung S.1993. Selection and breeding foracid-soil tolerance in crops: uplandrice and tropical forages as casestudies. Bioscience 43:454-465.
Sahu SK; Mishra SK; Senapati BK. 1988.Population biology and reproductivestrategie of Dichogaster bolaui(Oligochaeta: Octochaetidae) in twotropical agroecosystems. ProcIndian Acad Sci 97(3):239-250.
Senapati BK. 1980. Aspects ofecophysiological studies on tropicalearthworms (distribution,population dynamics, production,energetics and their role in thedecomposition process). Tesis(Ph.D.). Sambalpur University,India. 154 p.
Swift MJ; Heal OW; Anderson JM. 1979.Decomposition in terrestrialecosystems: Studies in ecology,Vol. 5. Blackwell, Oxford. 372 p.
Syers JK; Springett JA. 1983. Earthwormecology in grassland soils. En:Satchell JE, ed. Earthworm ecology.Chapman and Hall, Londres.p 67-84.
Watanabe H; Ruaysoongnern S. 1984.Cast production by themegascolecid earthworm Pheretimasp. in North-eastern Thailand.Pedobiologia 26:37-44.

Estrategias Reproductoras...
87
ResumenEste trabajo presenta informaciónsobre la biología de la reproducciónde tres especies de lombrices detierra de la familia Glossoscolecidae,que son nativas de los LlanosOrientales de Colombia. Ladinámica estacional de lareproducción varió entre las especiesestudiadas que presentaban uno odos períodos de reproducción poraño. Las lombrices depositaban loscapullos a diferentes profundidades,siendo la máxima 50 cm. Larelación entre el peso del capullo y eldel adulto fue la misma (un 6%) endos especies de lombrices endógeas,mientras que la relación observadaen una especie anécica (alrededor de16%) ha sido la más alta obtenidahasta la fecha para cualquierlombriz del trópico o de las zonastempladas.
Palabras clave: capullo, ciclos devida, distribución vertical,
estrategias reproductoras,lombrices
IntroducciónLos estudios realizados sobrediferentes aspectos de las estrategiasreproductoras de las lombrices detierra son los siguientes:Stephenson (1930), Walsh (1936),Evans y Guild (1948), Gerard (1967),Satchell (1967), Bouché (1972),Reynolds (1973), Nowak (1975),Phillipson y Bolton (1977), Lavelle(1971, 1978, 1979), Senapati (1980)y Garnsey (1994), entre otros.
Bouché (1977) y Lavelle (1977)establecieron una relación muyestrecha entre las estrategiasreproductoras y las categoríasecológicas de las lombrices. Lasepígeas, que se alimentan y viven enla hojarasca, sobrevivengeneralmente al estrés ambientalporque producen capullos. Lasendógeas, que viven en el suelo y sealimentan de él, presentan unaforma de resistencia llamadaquiescencia durante las condicionesambientales desfavorables y entranrápidamente en la fase reproductoracuando éstas son favorables.Finalmente, las anécicas, que vivenen el suelo pero se alimentan en lasuperficie, emplean dos mecanismospara evitar la mortalidad de la
CAPÍTULO 6
Estrategias Reproductoras de TresEspecies de Lombrices Nativas de lasSabanas de Carimagua (Colombia)1
J. J. Jiménez* , A. G. Moreno* y P. Lavelle**
1. Publicado en Pedobiologia 43, p 851-858,por Jiménez et al. Copyright © 1999 deUrban & Fischer Verlag. Impreso conpermiso de la casa editorial.
* Departamento de Biología Animal I(Zoología), Facultad de Biología, UniversidadComplutense de Madrid, 28040, Madrid,España.
** Laboratoire d’Ecologie et Biologie des SolsTropicaux, IRD (ex-ORSTOM), 32 Av. HenriVaragnat, F-93143 Bondy Cedex, Francia.

88
El Arado Natural:…
población: la diapausa, que puedeser diferente para los adultos y losindividuos juveniles, y los capullos,que aparecen al final de la épocalluviosa (Jiménez et al. 1998a).
Son relativamente escasosactualmente los conocimientosbásicos sobre la biología y la ecologíade las lombrices de tierra de la regiónneotropical del planeta, porque sehan emprendido muy pocos estudiossobre este tema. Németh (1981)estudió la fauna de lombrices de laselva tropical de Venezuela, Fragoso(1993) la de ambientes naturales yperturbados del sur de México,Feijoo (sin publicar) la de las laderasdel departamento de Cauca enColombia, y Muñoz-Pedreros (1997)la de Chile.
En un artículo previo (Jiménez etal. 1998a) se dan algunos datossobre la estrategia reproductora deMartiodrilus carimaguensis Jiménez yMoreno (en revisión). En este trabajose presentan, más detalladamente, elciclo de vida y la dinámicareproductora de tres especies nativasde los Llanos Orientales deColombia, Andiodrilus sp.,Glossodrilus sp. y M. carimaguensis.
Materiales y Métodos
Sitio del estudioEl sitio del estudio se halla en lassabanas isohipertérmicas biendrenadas de los Llanos Orientales deColombia, en la estación deinvestigación Carimagua. El valormedio anual de la precipitación y dela temperatura es 2280 mm y 26 °C,respectivamente, y la época seca seextiende de diciembre a marzo. Lossuelos son Oxisoles caracterizadospor su alta acidez [pH (H2O) 4.5],saturación de Al > 90%, y uncontenido bajo de los cationes
intercambiables Ca, Mg y K. Seevaluaron dos sistemas de uso de latierra que presentan fuertescontrastes: la sabana nativaherbácea y el pastizal mejorado enque se sembró una asociación degramínea y leguminosa (Brachiariadecumbens cv. Basilisk y Puerariaphaseoloides CIAT 9900). La cargaanimal de este sistema era de1 animal/ha en la época seca y de2 animales/ha en la época lluviosa;ningún tipo de manejo se empleó enel primer sistema.
Muestreo de lombricesEl diseño experimental se basa en unmuestreo estratificado al azar. Enambos sistemas de manejo sedelimitó un área de 90 x 90 m, quese dividió en cuadrículas regularesde 10 x 10 m. Durante 17 meses(desde abril de 1994 hastaseptiembre de 1995, excepto en juniode 1994) se emplearon dos métodosfísicos de extracción de lombrices: laextracción manual de cinco muestrasde suelo de 1 x 1 x 0.5 m cada mes,y el lavado y tamizado de10 muestras de suelo de 20 x 20 x20 cm cada mes (según Lavelle1978). Los monolitos fuerondivididos en capas de 10 cm degrosor para determinar el patrón dedistribución vertical de las lombrices,y todos los capullos recolectados encada capa fueron lavados en agua yllevados al laboratorio. Los detallesdel muestreo pueden consultarse enLavelle (1978) y Jiménez et al.(1998a, 1998b).
Incubación de los capullosLos capullos obtenidos en lasmuestras de campo se pesaron y secolocaron en placas de Petri sobrepapel de filtro húmedo. Los cultivosse rehumedecían todos los días y sepesaban los individuos que salían al

Estrategias Reproductoras...
89
hacer eclosión los capullos. Latemperatura del laboratorio semantuvo constante entre 26 y 28 ºC.
ResultadosLa cópula nunca pudo observarseporque ocurre, probablemente,durante la noche, tiempo en que noera posible salir de la estaciónexperimental. Se evaluó, por tanto,mediante métodos indirectos.
Capullos: morfología, tamañoy período de incubaciónLos capullos producidos porGlossodrilus sp. eran blanquecinos yesféricos (Cuadro 6-1) y tenían unextremo puntiagudo y extendido. Sutamaño medio era de 3.0 x 2.2 mm ysu peso fresco in vivo de 6.2 mg.Ninguno de los 79 capullosrecolectados en el campo pudoeclosionar bajo condicionescontroladas de laboratorio, por loque el período de incubación y elnúmero y el peso de los individuosrecién salidos de ellos no han podidodeterminarse para esta especie.
Los capullos de Andiodrilus sp.eran también esféricos, blanquecinosy algo translúcidos; su diámetromedio era de 5 mm y su peso fresco78.9 mg. Un solo zigoto podíaobservarse a través de cada capullo,el cual tenía un período deincubación medio de 13 días y unpeso fresco de 55.7 mg al momentode la eclosión. Aproximadamente, el50% del número total de capullosrecolectados no eclosionó en ellaboratorio. El 71% del peso total deun capullo correspondía al peso de lalombriz.
El capullo de M. carimaguensisera amarillento, oval y de grantamaño (23.6 x 14 mm) y su pesomedio alcanzó los 1,808 mg. Decada capullo salían dos embriones(sólo un capullo arrojó un soloembrión) cuyo peso medio era de760 mg (Figura 6-1). El períodomedio de incubación en esta especie,contando todos los capullos, fue de23.5 días; en cada capullo, el 80%del peso era aportado por losembriones. La tasa de eclosiónde capullos en el laboratorio fue de73.9%.
Cuadro 6-1. Principales características biológicas de los capullos de tres especies de lombrices detierra de Carimaguaa.
Características Glossodrilus n. sp. Andiodrilus n. sp. Martiodriluscarimaguensis
Capullos estudiados 79 88 46
Morfología Esférico Esférico Oval
Tamaño (mm) 3.0x2.2 6 23.6x14
Peso fresco in vivo (mg) 6.2 ± 1.4 78.9 ± 22.2 1808 ± 414.5(4.6-8.1) (30-130) (890-3020)
Período de incubación (días) — 12.8 ± 6.3 23.3 ± 12.9(1-28) (1-48)
Individuos por capullo — 1.02 ± 0.15 1.91 ± 0.29
Peso de individuos al 5b 55.7 ± 22.9 760 ± 219.4eclosionar capullo (mg) (20-120) (270-1760)
Peso adulto/peso capullo (%) 80.6 71 79.7
Tasa de eclosión en lab. (%) 0 48.3 73.9
a. Medias ± desviación típica (mínimo y máximo entre paréntesis). — = No fue determinado.b. Datos de individuos hallados en el campo.

90
El Arado Natural:…
Dinámica estacionalLa producción de capullos enCarimagua indicó que las lombricestendían a producirlos tanto alcomienzo como al final de la épocalluviosa, cuando las fluctuaciones dela humedad del suelo eran altas.Glossodrilus sp. y M. carimaguensispresentaron un solo período dereproducción anual, mientras queAndiodrilus sp. exhibió una dinámicaestacional bimodal (Figura 6-2).
Glossodrilus sp. puso suscapullos al final del período delluvias. En octubre de 1994 seencontraron los primeros capullos enla parcela de sabana y su númeromáximo se registró en enero de1995, cuando toda la población,principalmente los inmaduros,estaba inactiva. En el pastizal, encambio, los primeros capullos seencontraron en septiembre de 1994 yllegaron a un máximo en febrero de1995. La mayoría de los capulloseclosionan al inicio de la épocalluviosa. Esto explica la presencia delos picos máximos en la curva deaparición de capullos, que seobservaron durante los meses deverano.
Los capullos de Andiodrilus sp. seencontraron en dos períodosdiferentes durante la temporada delluvias. En la sabana se recolectaroncapullos principalmente al comienzo yen la mitad de la época húmeda.Durante el segundo año de estudioaparecieron dos picos (en la curva deabundancia) en los mismos meses,aunque el número de capullosregistrado fue más bajo que el delprimer año. En el pastizal seencontraron más capullos que en lasabana. Los picos máximos de lacurva se observaron en mayo yoctubre de 1994, y en mayo-junio yagosto del segundo año.
Martiodrilus carimaguensisdepositó sus capullos al final de laépoca húmeda y el número máximode éstos se registró en septiembre-octubre. En el siguiente año seencontraron capullos en marzo, loque prueba que algunos de ellos noeclosionan hasta la siguientetemporada de lluvias.
Distribución verticalA partir del número total de capullosrecolectados en cada capa se hacalculado la profundidad media a laque éstos fueron depositados(Figura 6-3). Las especies Andiodrilussp. y Glossodrilus sp. se encuentrancerca de la superficie del suelo, demodo que la mayoría de sus capullosse hallaban en los primeros 20 cm delperfil. La profundidad media a la queGlossodrilus sp. depositó los capullosen la sabana y en el pastizal fue de8.8 cm y de 12.4 cm,respectivamente. Estos datos sebasan en el número de capullosobtenidos por el método de lavado ytamizado. Algunos capullos seencontraron por debajo de los 20 cmde profundidad en las muestras de1 m2, lo que hizo necesaria unacorrección en el porcentaje decapullos encontrados.
1400
1200
1000
800
600
400
200
0
Figura 6-1. Gráfica encajada (box-plot) delpeso de los individuos reciénnacidos de Martiodriluscarimaguensis en la que aparecenla media (línea punteada) y lospercentiles 5% y 95%.
Peso individualmás bajo
Peso
rec
ién
nac
idos
(mg)
Peso individualmás alto
○ ○ ○ ○
○ ○ ○ ○
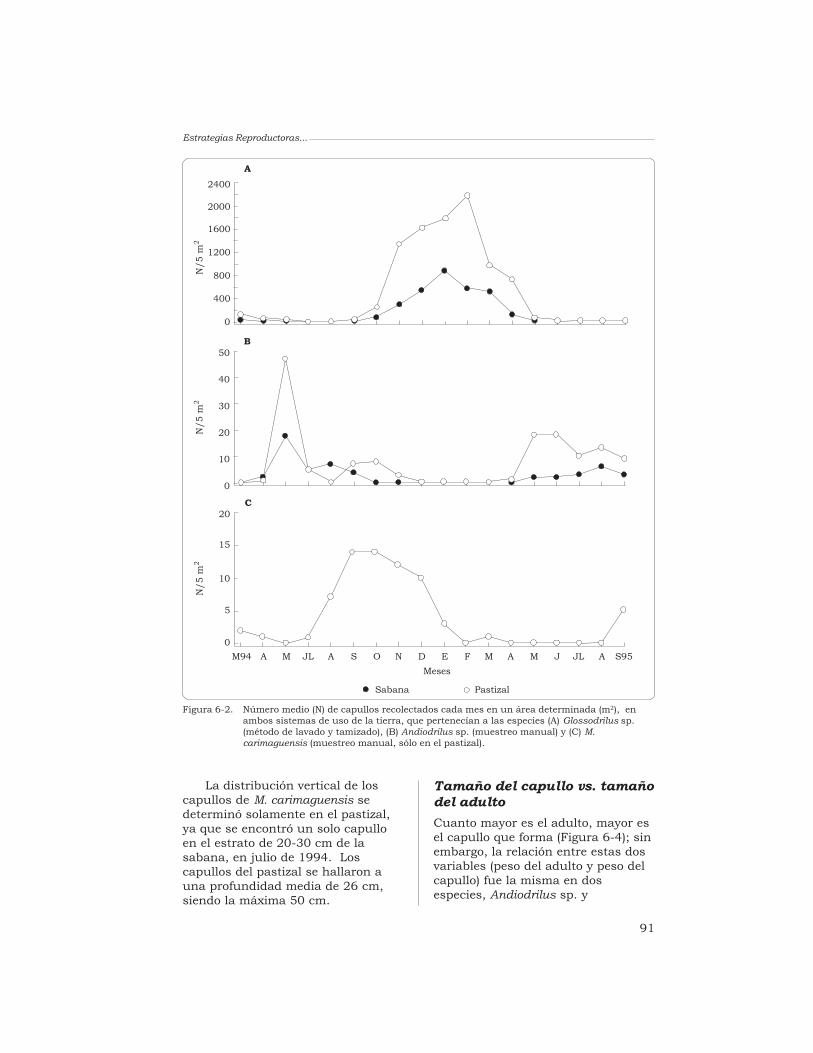
Estrategias Reproductoras...
91
La distribución vertical de loscapullos de M. carimaguensis sedeterminó solamente en el pastizal,ya que se encontró un solo capulloen el estrato de 20-30 cm de lasabana, en julio de 1994. Loscapullos del pastizal se hallaron auna profundidad media de 26 cm,siendo la máxima 50 cm.
Tamaño del capullo vs. tamañodel adultoCuanto mayor es el adulto, mayor esel capullo que forma (Figura 6-4); sinembargo, la relación entre estas dosvariables (peso del adulto y peso delcapullo) fue la misma en dosespecies, Andiodrilus sp. y
M94 A M JL A S O N D E F M A M J JL A S95
20
15
10
5
0
N/5
m2
Figura 6-2. Número medio (N) de capullos recolectados cada mes en un área determinada (m2), enambos sistemas de uso de la tierra, que pertenecían a las especies (A) Glossodrilus sp.(método de lavado y tamizado), (B) Andiodrilus sp. (muestreo manual) y (C) M.carimaguensis (muestreo manual, sólo en el pastizal).
2400
2000
1600
1200
800
400
0
N/5
m2
50
40
30
20
10
0
N/5
m2
Meses
Sabana Pastizal
A
B
C
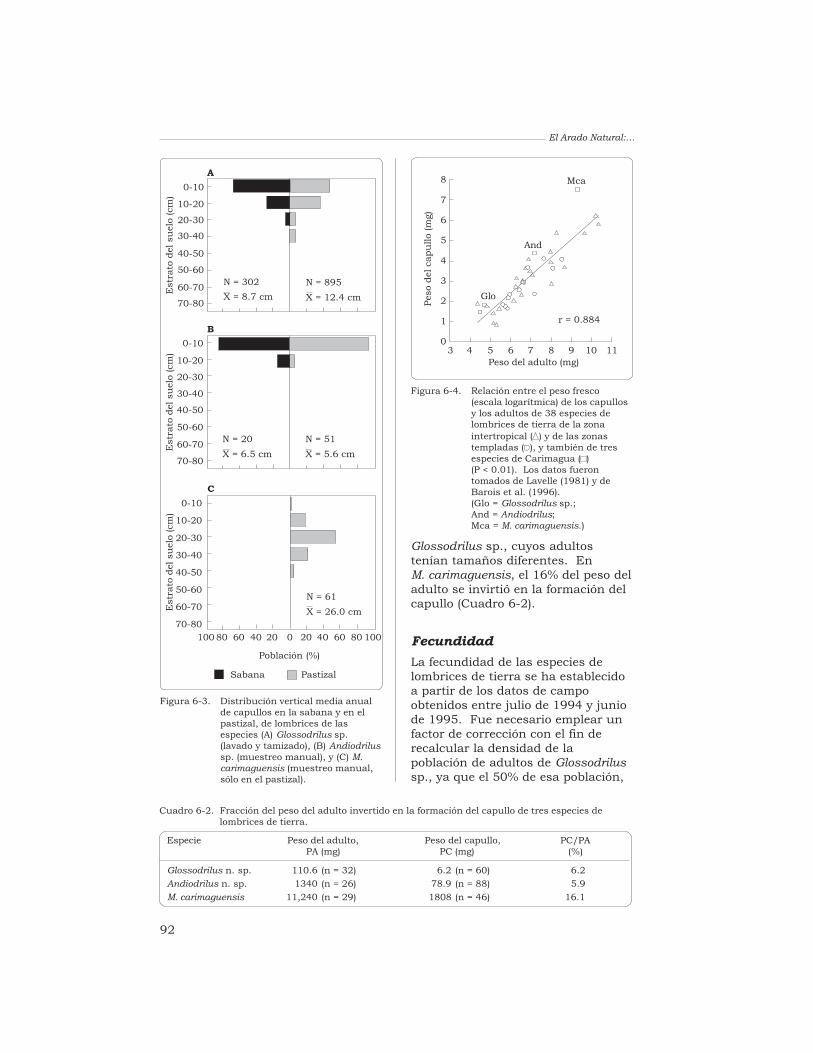
92
El Arado Natural:…
Glossodrilus sp., cuyos adultostenían tamaños diferentes. EnM. carimaguensis, el 16% del peso deladulto se invirtió en la formación delcapullo (Cuadro 6-2).
FecundidadLa fecundidad de las especies delombrices de tierra se ha establecidoa partir de los datos de campoobtenidos entre julio de 1994 y juniode 1995. Fue necesario emplear unfactor de corrección con el fin derecalcular la densidad de lapoblación de adultos de Glossodrilussp., ya que el 50% de esa población,
Cuadro 6-2. Fracción del peso del adulto invertido en la formación del capullo de tres especies delombrices de tierra.
Especie Peso del adulto, Peso del capullo, PC/PAPA (mg) PC (mg) (%)
Glossodrilus n. sp. 110.6 (n = 32) 6.2 (n = 60) 6.2Andiodrilus n. sp. 1340 (n = 26) 78.9 (n = 88) 5.9M. carimaguensis 11,240 (n = 29) 1808 (n = 46) 16.1
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
Est
rato
del
su
elo
(cm
)
100 80 60 40 20 0 20 40 60 80 100
N = 302X = 8.7 cm
N = 895X = 12.4 cm
Est
rato
del
su
elo
(cm
)
N = 20X = 6.5 cm
N = 51X = 5.6 cm
Est
rato
del
su
elo
(cm
)
N = 61X = 26.0 cm
A
B
C
Población (%)
Sabana Pastizal
Figura 6-3. Distribución vertical media anualde capullos en la sabana y en elpastizal, de lombrices de lasespecies (A) Glossodrilus sp.(lavado y tamizado), (B) Andiodrilussp. (muestreo manual), y (C) M.carimaguensis (muestreo manual,sólo en el pastizal).
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
3 4 5 6 7 8 9 10 11
Peso
del
cap
ullo
(mg)
8
7
6
5
4
3
2
1
0
Peso del adulto (mg)
Glo
And
Mca
r = 0.884
Figura 6-4. Relación entre el peso fresco(escala logarítmica) de los capullosy los adultos de 38 especies delombrices de tierra de la zonaintertropical ( ) y de las zonastempladas ( ), y también de tresespecies de Carimagua ( )(P < 0.01). Los datos fuerontomados de Lavelle (1981) y deBarois et al. (1996).(Glo = Glossodrilus sp.;And = Andiodrilus;Mca = M. carimaguensis.)

Estrategias Reproductoras...
93
aproximadamente, no pudorecogerse de los monolitos de 1 m2
cuando se hizo el muestreo manual(Jiménez, información sin publicar).La fecundidad más alta, es decir, elmayor número de capullosproducidos por un adulto en un añose registró en Glossodrilus sp. Estaespecie produjo 9.9 y 13 capullos/adulto al año en la sabana y en elpastizal, respectivamente.Andiodrilus sp. y M. carimaguensispresentaron valores más bajos: parala primera especie se calcularon0.61 y 0.50 capullos/adulto alaño en la sabana y en el pastizal,respectivamente, y paraM. carimaguensis 0.25 en el pastizal;de hecho, este valor era 0.49, o sea,el producto de 0.25 por 1.91individuos que contenía cadacapullo.
DiscusiónLos factores que afectan la actividadreproductora de las lombrices detierra son la temperatura y lahumedad del suelo (Evans y Guild1948). Michalis y Panidis (1993)concluyeron que ambos factoresregulan el patrón reproductor deOctodrilus complanatus (Dugés) yReinecke y Kriel (1980) realizaronobservaciones similares en la lombrizEisenia fetida. En Carimagua, lasfuertes variaciones climáticasestacionales restringen la postura decapullos a sólo 8 meses del año, yaque en el resto del año la comunidadde lombrices está inactiva. Seencontraron en este estudio dosestrategias diferentes y generales: ladeposición continua de Andiodrilussp. y la deposición al final de laépoca lluviosa seguida de diapausade los adultos (en M. carimaguensis)o de muerte de los adultos (enGlossodrilus sp.).
El tamaño del capullo no serelaciona siempre con el del adulto
(Edwards y Bohlen 1996). Sinembargo, Lavelle (1981) encontróuna clara correlación positiva entreel tamaño de los adultos y el de loscapullos que éstos forman, en variasespecies tropicales y de latitudestempladas. La fuerte inversión deM. carimaguensis en su reproducciónpuede ser el resultado de unaestrategia adaptativa más compleja.El valor obtenido para esta especieha sido el más alto hasta la fecha, loque explica su posición en laFigura 6-4: el punto más alejado dela recta principal de regresión.
La mayoría de los capullos queno eclosionaron presentaron unainfección fungosa en su superficie.Esta afección pudo ser la causa delos valores bajos de eclosiónobtenidos en condiciones delaboratorio, aunque no es posibleasegurarlo: bien pudieron ser estoshongos el resultado de la inviabilidaddel capullo, como lo mencionóSenapati (1980) estudiando laslombrices de tierra de la India.
Existe una relación entre elnúmero de capullos producidos y sulocalización en el suelo (Satchell1967). Este autor concluye que lasespecies de lombrices que viven enambientes que propician unamortalidad elevada de sus individuospresentan una tasa mayor deproducción de capullos. Pues bien,Glossodrilus sp. fue la especie queprodujo, en este estudio, la mayorcantidad de capullos, y éstos sehallaban, principalmente, en losprimeros 10 cm del perfil del suelo.Muchos de estos capullos noeclosionarán y morirán, es decir, laespecie presenta una estrategia rtípica (Pianka 1970). Por elcontrario, M. carimaguensis es unaespecie que aplica la estrategia K, esdecir, produce mucho menoscapullos y es capaz de descenderhasta capas profundas del suelopara pasar por una diapausa.

94
El Arado Natural:…
AgradecimientosEste estudio formó parte del proyectoMacrofauna STD-3 de la ComunidadEuropea y del antiguo Programa deTrópico Bajo del Centro Internacionalde Agricultura Tropical (CIAT),situado en Cali, Colombia. El primerautor expresa su más profundoagradecimiento a T. Decaëns por suayuda en el trabajo de campo y porlos comentarios que emitiórelacionados con este estudio.
ReferenciasBarois I; Ángeles A; Blanchart E;
Brossard M; Fragoso C; Jiménez JJ;Martínez MA; Moreno AG; LattaudC; Lavelle P; Rossi J-P; SenapatiBK; Giri S; Tondoh J. 1996. Ecologyof species with large environmentaltolerance and/or extendeddistributions. En: Lavelle P, ed.Conservation of soil fertility in low-input agricultural systems of thehumid tropics by manipulatingearthworm communities. Informefinal del proyecto Macrofauna.
Bouché MB. 1972. Lombriciens deFrance. Ecologie et systematique.INRA Publ. 72-2, París. 671 p.
Bouché MB. 1977. Stratégieslombriciennes. Ecol Bull 25:122-132.
Edwards CA; Bohlen PJ. 1996. Biologyand ecology of earthworms. 3a. ed.Chapman and Hall, Londres, ReinoUnido. 426 p.
Evans AC; Guild WJ McL. 1948. Studieson the relationships betweenearthworms and soil fertility. IV: Onthe life cycles of some BritishLumbricidae. Ann Appl Biol35:471-484.
Fragoso C. 1993. Les peuplements devers de terre dans l’est et sud-estdu Mexique. Tesis (Doctorado).Université Paris VI. 228 p + anexos.
Garnsey RB. 1994. Seasonal activity andaestivation of lumbricid earthwormsin the Middlands of Tasmania. AustJ Soil Res 32:1355-1367.
Gerard BM. 1967. Factors affectingearthworms in pastures. J AnimEcol 36:235-252.
Jiménez JJ; Moreno AG. Martiodriluscarmaguensis (Oligochaeta,Glossoscolecidae), una nuevaespecie de lombriz de tierra paraColombia. Megadrilogica. (Enrevisión.)
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drained savannas of Colombia. ApplSoil Eco 9(1-3):153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998. Earthworm communities innative savannas and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.
Lavelle P. 1971. Recherches ecologiquesdans la savane de Lamto (Côted’Ivoire): production annuelle d’unver de terre Millsonia anomala,Omodeo. Terre Vie 2-71:240-254.
Lavelle P. 1977. Bilan energetique despopulations naturelles du ver deterre geophage Millsonia anomala(Oligochetae-Acanthodrilidae) dansla savane de Lamto (Côte d’Ivoire).Geo Eco Trop 1(2):149-157.
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfonctions dans l’ècosystème. Tesis(Doctorado). París VI. Publ Lab ZoolENS. 12. 301 p.
Lavelle P. 1979. Relations entre typesécologiques et profilsdémographiques chez les vers deterre de la savane de Lamto (Côted’Ivoire). Rev Ecol Biol Sol16:85-101.

Estrategias Reproductoras...
95
Lavelle P. 1981. Stratégies dereproduction chez les vers de terre.Acta Oecol 2(2):117-133.
Michalis K; Panidis S. 1993. Seasonalvariation in the reproductive activityof Octodrilus complanatus(Oligochaeta, Lumbricidae). Eur JSoil Biol 29(3-4):161-166.
Muñoz-Pedreros A; Ruiz E; Poblete C;Santelices M. 1997. Aspectos de labiología reproductiva delumbrícidos silvestres (Oligochaeta:Lumbricidae) en el sur de Chile. RevChil Hist Nat 70:101-108.
Németh A. 1981. Estudio ecológicopreliminar de las lombrices detierra (Oligochaeta) en ecosistemasde bosque húmedo tropical en SanCarlos de Río Negro, TerritorioFederal Amazonas. Tesis.Universidad Central de Venezuela.92 p.
Nowak E. 1975. Population density ofearthworms and some elements oftheir production in severalgrassland environmments. Ekol Pol23:459-491.
Phillipson J; Bolton PJ. 1977. Growthand cocoon production byAllolobophora rosea (Oligochaeta:Lumbricidae). Pedobiologia17:70-82.
Pianka ER. 1970. On r- and K-selection.Am Nat 104:592-597.
Reinecke AJ; Kriel JR. 1980. Influence oftemperature on the reproduction ofthe earthworm Eisenia fetida(Oligochaeta). S Afr J Zool16(2):96-100.
Reynolds JW. 1973. Earthworm(Annelida: Oligochaeta) ecology andsystematics. En: Dindal D, ed.Proceedings 1st SoilMicrocommunities Conference.Primera Reunión deMicrocomunidades del Suelo,Siracusa, N.Y. octubre 1971.p 95-120.
Satchell JE. 1967. Lumbricidae. En:Burges A; Raw F, eds. Soil biology.Academic Press, Londres.p 259-322.
Senapati BK. 1980. Aspects ofecophysiological studies on tropicalearthworms. Distribution,population dynamics, production,energetics and their role in thedecomposition process. Tesis(Ph.D.). Sambalpur University,India. 154 p.
Stephenson J. 1930. The Oligochaeta.Oxford, Clarendon Press.
Walsh AS. 1936. Lumbricus lore. TurtoxNews 14:57-60.

96
El Arado Natural:…
ResumenDurante 17 meses se estudió ladistribución vertical de las lombricesde tierra nativas en las sabanasnaturales y en las perturbadas cuyossuelos son Oxisoles de los LlanosOrientales de Colombia; el estudio sehizo en una sabana y en un pastizalde gramínea y leguminosa que habíasido pastoreado durante 17 años.Se encontraron diferentes patronesde estratificación vertical en todaslas especies evaluadas, y se observóuna fuerte migración de laspoblaciones hacia los estratos másprofundos del suelo durante la épocaseca. Aunque las especies másgrandes se encontraron a mayorprofundidad, la correlación entre eltamaño de la lombriz y laprofundidad media a la que ésta seencontraba no fue significativa(P > 0.05). Los hábitos de vida y lasestrategias adaptativas de la especiemás pequeña encontrada en los dossistemas de manejo estudiados,Ocnerodrilidae n. sp., contribuyeron
a estos resultados. Esta especieendógea está relacionada con lasacumulaciones orgánicas producidaspor una especie anécica y deberávalorarse, en futuros estudios,respecto al papel que desempeña en elfuncionamiento del ecosistema. Losindividuos adultos de la especieanécica se localizaron a unaprofundidad mayor que los inmaduros(P < 0.01). Hubo un efecto importantede la humedad del suelo en ladistribución vertical de las lombrices,aunque las diferencias encontradasen este parámetro entre losinmaduros y los adultos de la especieanécica Martiodrilus carimaguensisson también, probablemente, deorigen biótico. Este trabajo presenta,además, datos nuevos sobre labiología y la ecología de las especiesneotropicales estudiadas.
Palabras clave: distribución vertical,estrategias adaptativas, lombrices,perturbación, sabanas
Dedico este capítulo ala memoria de mi tío Juan Lobato yde mi tía Ramona Romero (J.J.J.)
IntroducciónLa distribución vertical de la fauna delsuelo es uno de los factores quedeterminan la partición del nicho enlas comunidades de macrofauna y
CAPÍTULO 7
Distribución Vertical de las Lombricesde Tierra en los Suelos de los “Llanos”de Colombia Cubiertos por EspeciesForrajeras1
J. J. Jiménez* y T. Decaëns**
1. Publicado en Biology and Fertility of Soils32(6), p 463-473, por Jiménez y Decaëns.Copyright © 2000 de Springer. Impreso conpermiso de la casa editorial.
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** Laboratoire d’Ecologie Vegetal, UFRSciences, Université de Rouen, F-76821Mont Saint Aignan Cedex, Francia.

Distribución Vertical de las Lombrices...
97
reducen la competencia tantointerespecífica como intraespecíficaentre los taxones. Pocos estudios sehan realizado en la fauna del suelosobre diferenciación del nichoecológico. Kaczmareck (1975) yCurry (1978) estudiaron la amplituddel nicho en los artrópodos del suelo,Vegter (1983) la estudiaron enalgunas especies de Collemboladistribuidas a lo largo de ungradiente de humedad, y Davidson(1985) y Lynch et al. (1980) lainvestigaron en hormigas granívoras.Krishnamoorthy (1985) encontró, enlas lombrices de tierra, un ciertogrado de diferenciación del nicho enrelación con la distribución vertical,observando el contenido de materiaorgánica y la humedad a diferentesprofundidades.
La distribución de las lombricesdentro del perfil del suelo estádeterminada por factores abióticos(como la humedad y la temperatura)y bióticos (su comportamiento)(Martin y Lavelle 1992). La humedaddel suelo es un factor esencial quedetermina el grado de actividad y lalocalización de las lombrices en elsuelo, ya que son animales derespiración cutánea (Lavelle 1983;Lee 1985) que extraen agua del sueloque ingieren (Barois y Lavelle 1986).
La distribución vertical de laslombrices ha sido menos estudiadaen los ecosistemas tropicales que enlos de latitudes templadas. Reddy yPasha (1993) estudiaron ladistribución vertical de las lombricesen pastizales semiáridos de la India;Senapati (1980) en potreros bajopastoreo y sin pastoreo de la India;Németh y Herrera (1982) en lossuelos ácidos de una selva húmedatropical de Venezuela; Lavelle (1978)en diferentes tipos de la sabana deLamto (Costa de Marfil); y Fragoso(1993) en ecosistemas naturales yalterados del sureste mexicano. Enlos ecosistemas tropicales,
caracterizados por una fuerteestacionalidad, las lombrices migranhacia las capas inferiores del suelopara iniciar un estado de estivaciónde varios tipos, por ejemplo, laquiescencia o diapausa (Fragoso1985, 1993; Jiménez et al. 1998a;Lavelle 1978).
Algunos autores han reportadotambién las diferencias en ladistribución vertical que presentanlos individuos inmaduros y losadultos. Satchell (1955) encontróque los adultos de algunas especiesde lumbrícidos europeos seencontraban a una profundidadmayor que los inmaduros. Estefenómeno ha sido observado tambiénpor Gerard (1967) en pastizales deGran Bretaña, donde los adultosmaduros descendían a mayorprofundidad antes de entrar enreposo. Piearce (1983) confirmótambién este proceso y halló que losestadios juveniles estaban muchomás cerca de la superficie del sueloque los adultos, mientras que en elnorte de España Mato et al. (1988)encontraron este mismo patrón endos lumbrícidos. En Tasmania,Garnsey (1994) halló que losindividuos juveniles se localizaban auna profundidad menor que losadultos al iniciarse la estivación. Lasrazones que explican, generalmente,este patrón son dos: una, laprofundidad a la que sondepositados los capullos y, por ende,a la que se encuentran los estadiosjuveniles al eclosionar aquéllos; yotra, la débil capacidad deexcavación de los individuosjuveniles pequeños en comparacióncon la de los adultos.
La distribución vertical de laslombrices en el suelo se usa,habitualmente, como criterio para suclasificación en categorías ecológicas(Bouché 1972; Lavelle 1981, 1988;Lee 1959, 1985). El tamaño de lalombriz es otro parámetro que

98
El Arado Natural:…
algunos autores han relacionadocon su distribución vertical, esdecir, que cuanto mayor es lalombriz mayor será la profundidada la que desciende (Piearce 1983).Lavelle (1978) encontró unarelación positiva entre ladistribución vertical y el tamaño delas lombrices en las sabanas deLamto (Costa de Marfil), y Fragoso(1993) la observó en los suelosácidos aluviales de Chiapas(México).
Hay un interés creciente en ladescripción de la biodiversidad delsuelo y en la comprensión de losprocesos que explican elfuncionamiento de las comunidadesque lo habitan, tanto en losecosistemas naturales como en losalterados por el hombre (Giller1996). En las sabanas deCarimagua, situadas en los LlanosOrientales de Colombia, se hanllevado a cabo estudios quepersiguen estos objetivos en unárea donde no se había realizado,con anterioridad ningún intento deconocer la fauna del suelo (Decaënset al. 1994, 1999a, 1999b; Jiménezet al. 1998a, 1998b, 1999).
La descripción de ladistribución vertical de laslombrices de tierra y su explicaciónson elementos necesarios para elestudio de las poblaciones de éstas,con el fin de poseer todas lasherramientas posibles paramanejar, en el futuro, el recursonatural representado por dichaslombrices. El objetivo de esteestudio fue establecer los patronesestacionales de distribución verticalde la comunidad de lombrices detierra en una sabana nativa y en unpastizal intensivo, localizados en lassabanas isohipertérmicas biendrenadas de Colombia. Se analiza ydiscute la relación que hay entredicha distribución, de un lado, y lahumedad del suelo y el tamaño de
las diferentes especies de lombrices,del otro.
Materiales y Métodos
Descripción del lugar deestudioEl estudio se realizó en la estaciónexperimental de Carimagua (acuerdoCIAT-CORPOICA) situada en lassabanas isohipertérmicas biendrenadas de los Llanos Orientales deColombia (4°37’ N, 71°19’ O y175 msnm). Los valores mediosanuales de precipitación y detemperatura son 2280 mm y 26 °C,respectivamente (datos del CIAT,desde 1972 hasta 1995); hay unaépoca seca muy fuerte de diciembrea marzo. Los suelos se hanclasificado como finos, caoliníticos,isohipertérmicos, Haplustox típicoUSDA (suelo franco arcilloso), loscuales se suponen de baja fertilidad.Se caracterizan por su acidez(pH = 4.5, en agua), su saturación deAl alta (> 90%) y por valores bajos delos cationes intercambiables Ca, Mgy K (Lascano y Estrada 1989).
Se estudiaron dos parcelasdiferentes:
- una sabana natural herbácea sinpastorear, en donde ocurrió unfuego accidental 2 años antes delinicio de los muestreos;
- un pastizal mantenido 17 añosbajo pastoreo, en el cual unagramínea procedente de Africa(Brachiaria decumbens cv.Basilisk) se asoció con unaleguminosa herbácea (Puerariaphaseoloides CIAT 9900) llamada“kudzú”.
El pastizal se fertilizó con lasdosis siguientes (en kg/ha): 44 de P,40 de K, 14 de Mg y 22 de S duranteel establecimiento, y con 10 de P,

Distribución Vertical de las Lombrices...
99
9 de K, 92.5 de Ca, 9 de Mg y11.5 de S cada segundo año, durantelos siguientes 9 años (Lascano yEstrada 1989). La carga animal enel pastizal introducido era de1 animal/ha en la época seca (el“verano”) y 2 animales/ha en laépoca lluviosa (el “invierno”).
Muestreo de lombricesTanto en la sabana como en elpastizal se tomaron muestras desuelo cada mes, desde marzo de1994 hasta septiembre de 1995. Enjunio de 1994 no fue posible hacerlodebido a la ausencia de personaldisponible. En cada uno de los dossistemas de manejo se realizó unmuestreo estratificado al azar en unárea de 90 x 90 m dividida encuadrantes regulares de 10 x 10 m,que fueron agrupados en cincozonas. Se aplicaron dos métodosfísicos (según Lavelle 1978):extracción manual en muestras de1 m2 x 0.5 m de profundidad, ylavado y tamizado en muestras de20 cm2 x 20 cm de profundidad.Cada mes y en cada sistema demanejo se tomaron al azar cincomuestras de 1 m2 y 10 muestras delavado y tamizado (excepto en marzode 1994, cuando fueron 3 y 6,respectivamente). Las muestrasempleadas para el proceso de lavadoy tamizado se tomaron a amboslados de la muestra de 1 m2.
Alrededor de las muestras de1 m2 se excavó una zanja para evitarla posible salida lateral de laslombrices y para facilitar laseparación de las diferentes capas desuelo de 10 cm de grosor cada una.La profundidad del muestreo varióestacionalmente porque algunasespecies, por ejemplo, Martiodriluscarimaguensis (Jiménez y Moreno, enrevisión), migraban verticalmente;esta especie presenta diapausa ysuele descender hasta 80 cm de
profundidad durante la época seca(Jiménez et al. 1998a). Todas laslombrices observadas fueronrecogidas, lavadas en agua y fijadasen formalina al 4% (10% de ladilución comercial). En el laboratoriose separaron según su especie, secontaron y se pesaronindividualmente los adultos (conclitelo presente), los subadultos (sinclitelo, pero con marcas sexualespresentes), los inmaduros (sin cliteloni marcas) y los capullos.
Humedad del sueloEn cada capa de suelo muestreadase tomaron, además, muestras delsuelo. El contenido de agua delsuelo (en porcentaje respecto al pesodel suelo seco) se calculó a partir deseis submuestras obtenidas en cadacapa, que se pesaron (unos 100 g desuelo húmedo) y se secaron al hornoa 60 ºC durante 4 días comomínimo.
Comunidad de lombrices en lasabana y en el pastizalLa comunidad de lombrices de lasabana y del pastizal estáconstituida por ocho especies nuevaspara la ciencia. Todas ellas sonnativas, incluso las del pastizal, ytienen tamaños y funcionesecológicas diversas (Jiménez et al.1998b). Aunque se mantiene lariqueza específica, el índice H’(Shannon y Weaner 1949) disminuyó(desde 2.89 hasta 1.29) cuando elecosistema natural fue remplazadopor el pastizal introducido; estadisminución se debió a una especieanécica que contribuye con el 88% ala biomasa total debida a laslombrices (Jiménez et al. 1998b).Aunque la densidad de las lombriceses muy similar en el pastizal y en lasabana (130.3 frente a 114.5individuos/m2), la biomasa de

100
El Arado Natural:…
aquéllas en el pastizal (59.4 g PF/m2)supera en más de 10 veces laregistrada en la sabana (4.8 g PF/m2). Los patrones de distribuciónvertical se establecieron solamenteen seis especies: Andiodrilus n. sp.(endógea mesohúmica), Andiorrhinusn. sp. (endoanécica), Aymara n. sp.(epígea), Glossodrilus n. sp. (endógeapolihúmica), M. carimaguensis(anécica) y Ocnerodrilidae n. sp.(endógea polihúmica).
Análisis estadísticosEl grado de correlación entre algunasvariables, por ejemplo, entre larelación longitud/diámetro (L/D) y larelación peso/diámetro (P/D), de unaparte, y la distribución vertical, deotra, o entre la densidad o labiomasa respecto a la humedad delsuelo, se obtuvo mediante unanálisis de regresión lineal simple (rde Pearson). Se emplearon tambiénla prueba de t y el análisis devarianza (ANOVA) con el fin deestablecer diferencias significativasentre las medias. Finalmente, seaplicó una prueba no paramétrica, elANOVA de Kruskal Wallis, paracomparar las variables bajo lapresunción de que el tamaño de lasmuestras era diferente. Se empleó elpaquete estadístico SPSS en todoslos casos y se aplicó el programa(software) “SigmaPlot JandelScientific” para la presentación delas figuras.
Resultados
Distribución vertical de lasespeciesLa evaluación del patrón dedistribución vertical de laspoblaciones de lombrices de tierrapermite definir las capas de sueloexplotadas por las diferentesespecies que integran la comunidadde lombrices. La distribuciónvertical media anual de las especiesdesde julio de 1994 hasta junio de1995, en los dos sistemas de manejoestudiados, se presenta en elCuadro 7-1. El sistema de uso de latierra no causó ningún efecto en ladistribución vertical de las especies,excepto en M. carimaguensis(ANOVA, P < 0.001). La humedadmedia del suelo, obtenida cada mes,fue significativamente diferente entrelos dos sistemas de uso (P = 0.028,prueba de t); sin embargo, lasdiferencias en el contenido dehumedad entre los estratos de suelode ambos sistemas no fueronsignificativas (ANOVA, P > 0.05).
Tres especies se encontraron auna profundidad media de 0 a10 cm: Andiodrilus n. sp., Aymaran. sp. y Glossodrilus n. sp.Andiorrhinus n. sp. se encontró auna profundidad media de 12 cm enel pastizal; no se observaronindividuos de esta especie en lasabana entre julio de 1994 y junio
Cuadro 7-1. Valor medio (cm) de la distribución vertical en que se encontraron algunas especies delombrices de Carimagua. Los valores seguidos de diferente letra indican diferenciassignificativas (ANOVA, P < 0.001).
Especies Categoría ecológica Sabana Pastizalnativa introducido
Andiodrilus n. sp. Endógea (mesohúmica) 9.5a 8.3aAndiorrhinus n. sp. Endo-anécica ND1 12.1Aymara n. sp. Epígea 10.8a 5.2aGlossodrilus n. sp. Endógea (polihúmica) 10.3a 6.7aMartiodrilus carimaguensis Anécica 39.3a 32.5bOcnerodrilidae n. sp. Endógea (polihúmica) 25.4a 22.7a
1. ND = no determinado.
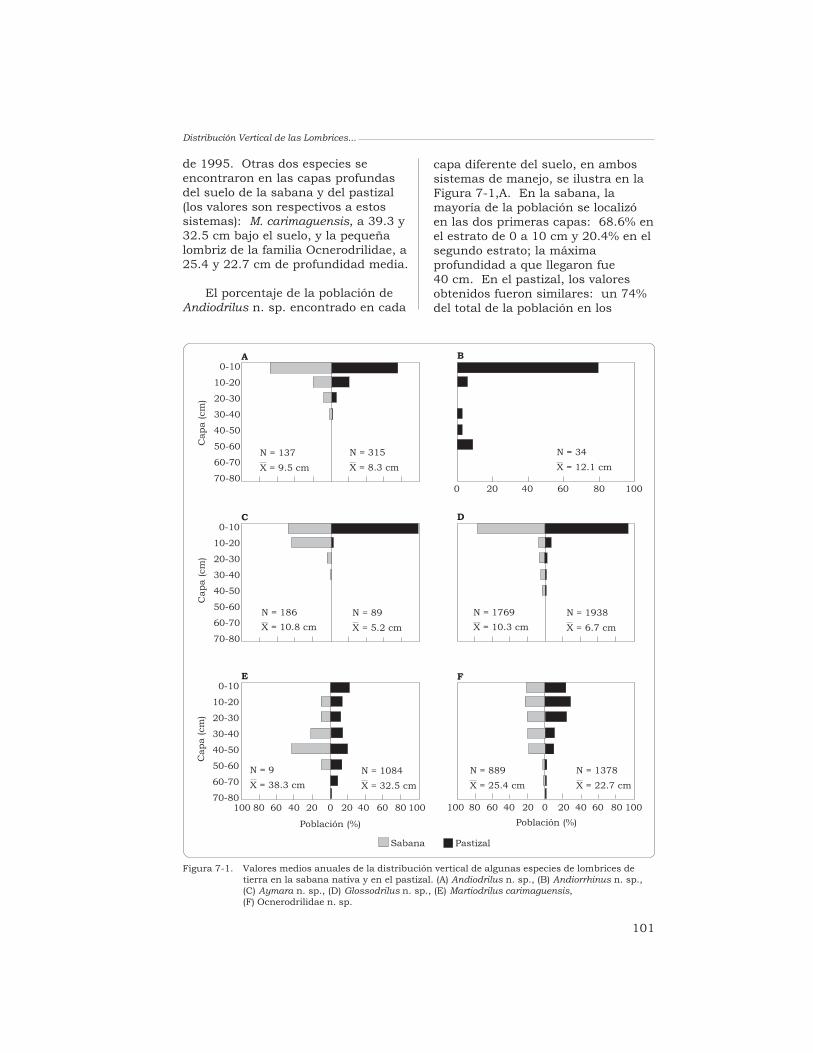
Distribución Vertical de las Lombrices...
101
de 1995. Otras dos especies seencontraron en las capas profundasdel suelo de la sabana y del pastizal(los valores son respectivos a estossistemas): M. carimaguensis, a 39.3 y32.5 cm bajo el suelo, y la pequeñalombriz de la familia Ocnerodrilidae, a25.4 y 22.7 cm de profundidad media.
El porcentaje de la población deAndiodrilus n. sp. encontrado en cada
capa diferente del suelo, en ambossistemas de manejo, se ilustra en laFigura 7-1,A. En la sabana, lamayoría de la población se localizóen las dos primeras capas: 68.6% enel estrato de 0 a 10 cm y 20.4% en elsegundo estrato; la máximaprofundidad a que llegaron fue40 cm. En el pastizal, los valoresobtenidos fueron similares: un 74%del total de la población en los
Cap
a (c
m)
A
Población (%)
Sabana Pastizal
Figura 7-1. Valores medios anuales de la distribución vertical de algunas especies de lombrices detierra en la sabana nativa y en el pastizal. (A) Andiodrilus n. sp., (B) Andiorrhinus n. sp.,(C) Aymara n. sp., (D) Glossodrilus n. sp., (E) Martiodrilus carimaguensis,(F) Ocnerodrilidae n. sp.
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
N = 137X = 9.5 cm
N = 315X = 8.3 cm
N = 34X = 12.1 cm
0 20 40 60 80 100
Cap
a (c
m)
C0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
N = 186X = 10.8 cm
N = 89X = 5.2 cm
Cap
a (c
m)
E0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
N = 9X = 38.3 cm
N = 1084X = 32.5 cm
100 80 60 40 20 0 20 40 60 80 100
D
N = 1769X = 10.3 cm
N = 1938X = 6.7 cm
F
N = 889X = 25.4 cm
N = 1378X = 22.7 cm
100 80 60 40 20 0 20 40 60 80 100Población (%)
B

102
El Arado Natural:…
primeros 10 cm y un 19.7% en lasegunda capa.
Andiorrhinus n. sp. no seencontró en la sabana, por lo que ladistribución vertical de esta especieestá representada por los valoresobtenidos en el pastizal(Figura 7-1,B). Los pocos individuosobservados indicaron que casi el80% del total de la población seencontraba en la primera capa, y quela profundidad máxima quealcanzaban era 60 cm.
En la sabana, Aymara n. sp. sedistribuyó por igual en las dosprimeras capas de suelo y sólo el5.4% se encontró en la capa de 20 a30 cm (Figura 7-1,C). En el pastizal,casi toda la población se encontró enlos primeros 10 cm de profundidad.
Glossodrilus n. sp. se encontró,principalmente, en las capas mássuperficiales del suelo. Durante laépoca seca, la máxima profundidaden la que se observaron lombrices deesta especie fue de 80 cm(Figura 7-1,D). En la sabana, el 76%de la población se halló en el estratode 0 a 10 cm y los demásporcentajes disminuían según laprofundidad. En el pastizal, el90.2% de los individuos seencontraron en los primeros 10 cm;algunos ejemplares aparecieronhasta en la capa de 70 a 80 cm.
La distribución vertical mediaanual de M. carimaguensis semuestra en la Figura 7-1,E. En lasabana se encontraron solamente9 individuos en un lapso de 1 año;por tanto, la comparación de estadistribución vertical de la especie conla del pastizal es tentativa. En elpastizal, las capas de 0 a 10 cm y de40 a 50 cm presentaron los valoresmás altos de población, es decir,21.3% y 19.2%, respectivamente.Estos resultados reflejan la variaciónestacional de la distribución verticalde esta especie. Porcentajes de
.
población aproximadamente igualesse observaron en cada una de lascapas restantes, hasta llegar a los60 cm de profundidad (los valoresoscilaron entre el 11.2% y el 13.4%).En las capas de 60 a 70 cm y de70 a 80 cm se encontraron el 8.2% yel 1.7%, respectivamente, de ladensidad total de la población.
Respecto a la especieOcnerodrilidae n. sp., cerca del 95%de su población, en la sabana, estabadistribuida homogéneamente hastalos 50 cm de profundidad(Figura 7-1,F). Menos del 5% deltotal se encontró a una profundidadmayor, cuyo valor máximo fue 80 cm.En el pastizal, el 22.6% de losindividuos se hallaron en la capa de0 a 10 cm y se obtuvieron valoressimilares en las dos capas siguientes.En las capas de 30 a 40 cm y de 40 a50 cm se encontró el 10.5% y el 9.9%de la población total, respectivamente,y sólo el 5% entre los 50 y los 80 cm.
Tamaño de la lombriz vs.distribución verticalAlgunos autores han establecidorelaciones entre algunas variablesbiométricas de las lombrices de tierray su distribución vertical en el perfildel suelo (Fragoso 1993; Lavelle1978). Piearce (1983) reveló laexistencia de una correlación positivaentre la longitud del cuerpo de lalombriz de tierra y la profundidad a laque excava el suelo.
Las relaciones L/D y P/D se hancorrelacionado con la distribuciónvertical media anual (Figura 7-2). Sinembargo, no se observarondiferencias significativas (P > 0.05)por la presencia de la especieOcnerodrilidae n. sp. Si esta especieno hubiera estado presente en lacomunidad de lombrices, lascorrelaciones habrían sidosignificativas, ya que ella estáasociada con los lugares donde el

Distribución Vertical de las Lombrices...
103
contenido de materia orgánica es alto,por ejemplo, donde se produce ladescomposición de la hojarasca y delas raíces (T. Decaëns, datos nopublicados), con los nidos de especiesde coleópteros y con las galerías de M.carimaguensis (Jiménez et al. 1998a).Estas galerías penetran
0 5 10 15 20 25 30 35
Peso
/Diá
met
ro
1.0
0.8
0.6
0.4
0.2
0
Profundidad media (cm)
Aym
r = 0.552
B
Glo
And And
AymGlo Ocn Ocn
Anr
Mca
Lon
gitu
d/D
iám
etro
60
50
40
30
20
10
Aym
r = 0.319
A
And
AndAym
OcnOcn
Mca
GloGlo
Anr
Figura 7-2. (A) Correlación entre la longitudrelativa (L/D) de la lombriz y laprofundidad media en que sehallan varias especies de lasabana ( ) y del pastizal ( );(B) Correlación entre el pesorelativo (P/D) de la lombriz y laprofundidad media en que seencuentran varias especies de lasabana ( ) y del pastizal ( ) (nosignificativo. (And = Andiodrilus n.sp.; Anr = Andiorrhinus n. sp.;Aym = Aymara n. sp.;Glo = Glossodrilus n. sp.;Mca = M. carimaguensis;Ocn = Ocnerodrilidae n. sp.)
profundamente en el suelo, de modoque Ocnerodrilidae n. sp., a pesar desu tamaño, puede encontrarse a granprofundidad. Por otra parte, lasespecies de lombrices más grandes noestán localizadas, necesariamente, enlas capas más profundas del suelo.
La clasificación ecológica de laslombrices desarrollada por Lee (1959,op. cit. en 1985) sitúa especies en lahojarasca (‘litter’), en la capasuperficial (‘topsoil’) del suelo, y en laszonas más profundas (‘subsoil’) delsuelo:
- Aymara n. sp. es una especie de lahojarasca con una relaciónL/D que oscila entre 33 en lasabana hasta 29.0 en el pastizal;
- Andiodrilus n. sp. y Glossodrilussp. son especies superficiales cuyarelación L/D es de 20 y 53,respectivamente;
- Andiorrhinus sp. y Ocnerodrilidaen. sp. son especies intermediasentre la superficie y los estratosprofundos y su relación L/Doscila entre 22.9 y 32.4,respectivamente, aunque lainclusión de la última especie endicha categoría ecológica no estámuy clara;
- M. carimaguensis es una especiesuperficial con una relación L/Dde 20.7.
Las relaciones P/D oscilaron entre7 x 10-3 en Ocnerodrilidae n. sp. hasta0.90 en M. carimaguensis(Cuadro 7-2).
Distribución vertical de laestivación en M. carimaguensisMartiodrilus carimaguensis es unaespecie nativa perteneciente a lafamilia Glossoscolecidae; susdimensiones medias, en estado adulto,son 9.3 mm de diámetro y 194.3 mmde longitud, y su peso fresco es de

104
El Arado Natural:…
11.2 g (en formalina 4%; n = 29). Elcolor del cuerpo es gris oscuro en ellado dorsal y gris claro en el ladoventral (Jiménez et al. 1998a). Estaespecie es un anécico típico (sensuBouché 1972) que ha dado pruebasclaras de su migración vertical en elsuelo y que presenta diferencias,respecto al inicio de la diapausa, entrelos estados juveniles y los adultos:mientras éstos comienzan la diapausaal final de la época lluviosa, ennoviembre o diciembre, los juvenilesla inician 4 meses antes, en julio o enagosto (Jiménez et al. 1998a).
En general, se encontraron máslombrices en estivación en la capa de40 a 50 cm que en otras capas delsuelo, mientras que tanto los adultoscomo los subadultos se localizaban enla de 50 a 60 cm (Figura 7-3).Prescindiendo del estado demográficode toda la población, se obtuvo unarelación no lineal significativa entre elpeso de las lombrices en diapausa yla profundidad a la cual seencontraban (ANOVA Kruskal-Wallis,P = 0.011). Cuanto más grande es lalombriz, mayor es la profundidad a laque se encuentra (Figura 7-4);además, el peso medio de losindividuos en diapausa osciló entre1 y 2.5 g (4 g fue el máximo), que esbastante bajo si se compara con elpeso fresco del adulto. Esto se debe aque la lombriz expulsa su contenidointestinal antes de enrollarse en lacámara de estivación.
Figura 7-3. Distribución vertical de ladiapausa de acuerdo a la edad delas lombrices, en la especieM. carimaguensis en un pastizal.
Total individuos (no.)
Cap
a de
l su
elo
(cm
)
0-1010-2020-3030-4040-5050-6060-7070-8080-90
0 50 100 150 200 250 300
Superficie delsuelo
Adultos y subadultos Inmaduros
Figura 7-4. Relación entre el peso medio de losindividuos de M. carimaguensis enestivación y la diferenteprofundidad a que se encuentran.(Las barras indican la desviacióntípica; las cifras sobre las barrasindican número de individuos).
Peso medio (g)
Cap
a de
l su
elo
(cm
)
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5
7
66
140254
202
104
18
0-10
10-20
20-30
30-40
40-50
50-60
60-70
70-80
80-90
Superficie delsuelo
Cuadro 7-2. Características biométricas de la comunidad de lombrices estudiada en la sabana y en elpastizal (cada dato indica la media ± desviación típica).
Especie Longitud/Diámetro Peso/Diámetro Muestra (n) en:
Sabana Pastizal Sabana Pastizal Sabana Pastizal
Andiodrilus n. sp. 23.0 ± 3.5 20.2 ± 4.3 0.17 ± 0.1 0.17 ± 0.1 31 52Andiorrhinus n. sp. ND1 22.9 ± 0.7 ND 0.39 ± 0.2 ND 24Aymara n. sp. 29.0 ± 5.1 33.9 ± 5.3 0.02 ± 0.01 0.03 ± 0.01 64 23Glossodrilus n. sp. 52.9 ± 4.8 52.0 ± 6.2 0.06 ± 0.01 0.06 ± 0.01 68 99M. carimaguensis ND 20.7 ± 0.6 ND 0.90 ± 0.4 ND 99Ocnerodrilidae n. sp. 31.6 ± 1.0 32.4 ± 0.9 8.10-3 ± 0.002 7.10-3 ± 0.002 135 198
1. ND = no determinado.
. .
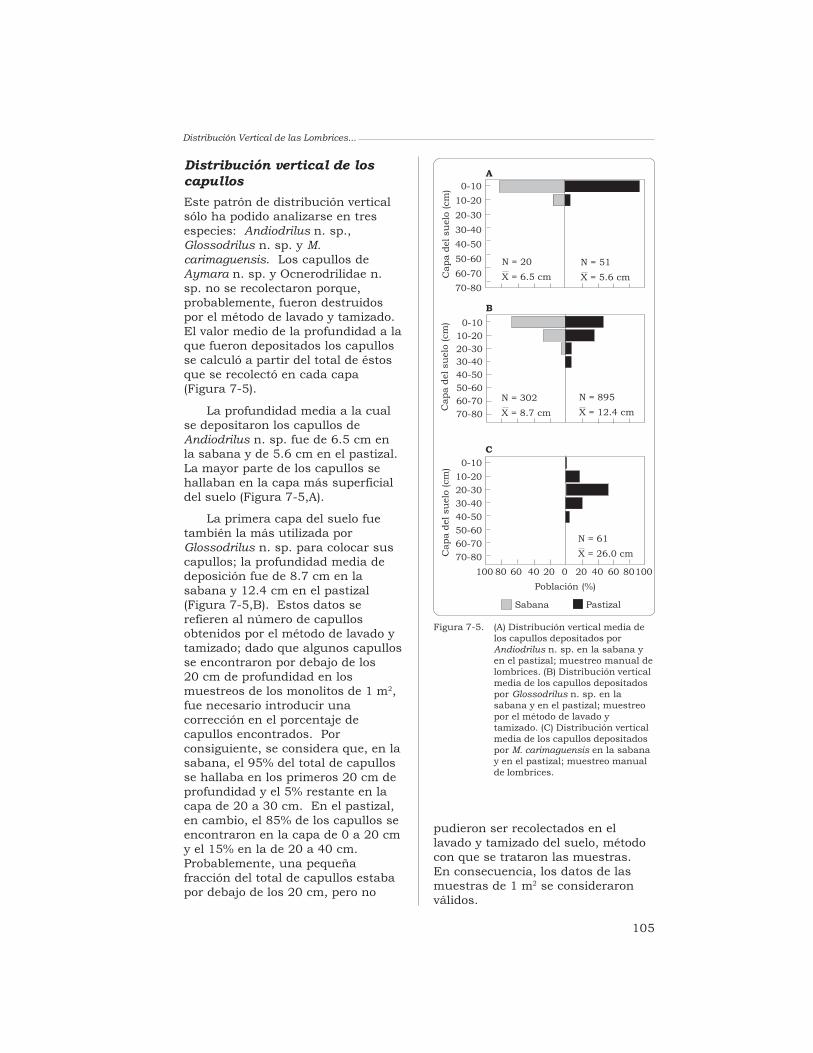
Distribución Vertical de las Lombrices...
105
Distribución vertical de loscapullosEste patrón de distribución verticalsólo ha podido analizarse en tresespecies: Andiodrilus n. sp.,Glossodrilus n. sp. y M.carimaguensis. Los capullos deAymara n. sp. y Ocnerodrilidae n.sp. no se recolectaron porque,probablemente, fueron destruidospor el método de lavado y tamizado.El valor medio de la profundidad a laque fueron depositados los capullosse calculó a partir del total de éstosque se recolectó en cada capa(Figura 7-5).
La profundidad media a la cualse depositaron los capullos deAndiodrilus n. sp. fue de 6.5 cm enla sabana y de 5.6 cm en el pastizal.La mayor parte de los capullos sehallaban en la capa más superficialdel suelo (Figura 7-5,A).
La primera capa del suelo fuetambién la más utilizada porGlossodrilus n. sp. para colocar suscapullos; la profundidad media dedeposición fue de 8.7 cm en lasabana y 12.4 cm en el pastizal(Figura 7-5,B). Estos datos serefieren al número de capullosobtenidos por el método de lavado ytamizado; dado que algunos capullosse encontraron por debajo de los20 cm de profundidad en losmuestreos de los monolitos de 1 m2,fue necesario introducir unacorrección en el porcentaje decapullos encontrados. Porconsiguiente, se considera que, en lasabana, el 95% del total de capullosse hallaba en los primeros 20 cm deprofundidad y el 5% restante en lacapa de 20 a 30 cm. En el pastizal,en cambio, el 85% de los capullos seencontraron en la capa de 0 a 20 cmy el 15% en la de 20 a 40 cm.Probablemente, una pequeñafracción del total de capullos estabapor debajo de los 20 cm, pero no
Figura 7-5. (A) Distribución vertical media delos capullos depositados porAndiodrilus n. sp. en la sabana yen el pastizal; muestreo manual delombrices. (B) Distribución verticalmedia de los capullos depositadospor Glossodrilus n. sp. en lasabana y en el pastizal; muestreopor el método de lavado ytamizado. (C) Distribución verticalmedia de los capullos depositadospor M. carimaguensis en la sabanay en el pastizal; muestreo manualde lombrices.
Cap
a de
l su
elo
(cm
)
A0-10
10-2020-3030-4040-5050-6060-7070-80
N = 20X = 6.5 cm
N = 51X = 5.6 cm
0-1010-2020-3030-4040-5050-6060-7070-80
Cap
a de
l su
elo
(cm
)
B
N = 302X = 8.7 cm
N = 895X = 12.4 cm
Población (%)
Cap
a de
l su
elo
(cm
)C
N = 61X = 26.0 cm
100 80 60 40 20 0 20 40 60 80100
0-1010-2020-3030-4040-5050-6060-7070-80
Sabana Pastizal
pudieron ser recolectados en ellavado y tamizado del suelo, métodocon que se trataron las muestras.En consecuencia, los datos de lasmuestras de 1 m2 se consideraronválidos.
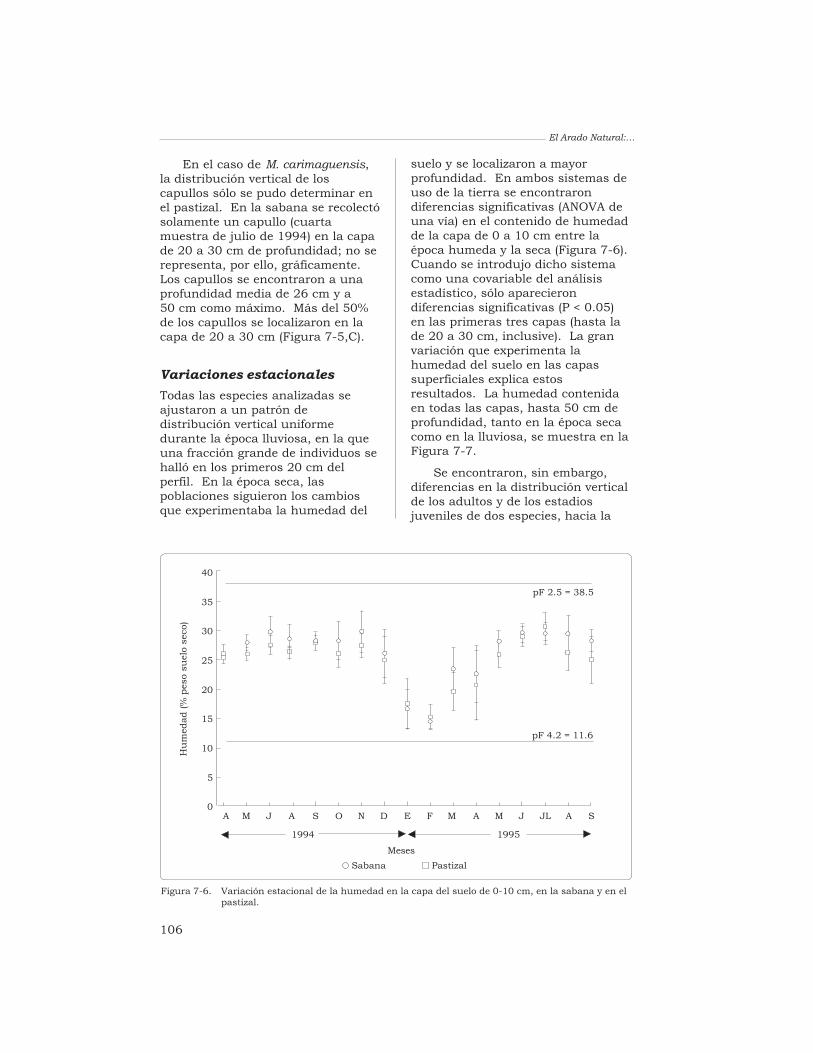
106
El Arado Natural:…
Figura 7-6. Variación estacional de la humedad en la capa del suelo de 0-10 cm, en la sabana y en elpastizal.
A M J A S O N D E F M A M J JL A S
40
35
30
25
20
15
10
5
0
Hu
med
ad (%
pes
o su
elo
seco
)
Meses Sabana Pastizal
pF 2.5 = 38.5
pF 4.2 = 11.6
19951994
En el caso de M. carimaguensis,la distribución vertical de loscapullos sólo se pudo determinar enel pastizal. En la sabana se recolectósolamente un capullo (cuartamuestra de julio de 1994) en la capade 20 a 30 cm de profundidad; no serepresenta, por ello, gráficamente.Los capullos se encontraron a unaprofundidad media de 26 cm y a50 cm como máximo. Más del 50%de los capullos se localizaron en lacapa de 20 a 30 cm (Figura 7-5,C).
Variaciones estacionalesTodas las especies analizadas seajustaron a un patrón dedistribución vertical uniformedurante la época lluviosa, en la queuna fracción grande de individuos sehalló en los primeros 20 cm delperfil. En la época seca, laspoblaciones siguieron los cambiosque experimentaba la humedad del
suelo y se localizaron a mayorprofundidad. En ambos sistemas deuso de la tierra se encontrarondiferencias significativas (ANOVA deuna vía) en el contenido de humedadde la capa de 0 a 10 cm entre laépoca humeda y la seca (Figura 7-6).Cuando se introdujo dicho sistemacomo una covariable del análisisestadístico, sólo aparecierondiferencias significativas (P < 0.05)en las primeras tres capas (hasta lade 20 a 30 cm, inclusive). La granvariación que experimenta lahumedad del suelo en las capassuperficiales explica estosresultados. La humedad contenidaen todas las capas, hasta 50 cm deprofundidad, tanto en la época secacomo en la lluviosa, se muestra en laFigura 7-7.
Se encontraron, sin embargo,diferencias en la distribución verticalde los adultos y de los estadiosjuveniles de dos especies, hacia la

Distribución Vertical de las Lombrices...
107
mitad de la época de lluvias: M.carimaguensis y Glossodrilus n. sp.
- Los individuos inmaduros deM. carimaguensis descendieronhacia las capas profundas delsuelo para iniciar la diapausa,4 meses antes que los adultos(Jiménez et al. 1998a). El patrónde distribución vertical deambas especies se ilustra en laFigura 7-8. Durante el primeraño de muestreo, al inicio de laépoca lluviosa, la mayor parte dela población de estas especies selocalizó en los primeros 20 cmdel perfil. Hacia la mitad dedicha época, además de loscambios ocurridos en el estadodemográfico de las poblaciones,se observó que, en las capas másprofundas del perfil, había unnúmero importante de individuosinactivos, especialmente de losinmaduros, de M. carimaguensis;los adultos de la especie, en
cambio, se mantenían todavíaactivos en los primeros 20 cm.
- Los individuos inmaduros deGlossodrilus n. sp. comenzaron adescender en el suelo de lasabana en agosto de 1994, y sunúmero aumentó con el tiempoen las capas más profundas. Enoctubre de 1994, la población deGlossodrilus n. sp. estabaconstituida completamente poradultos que preferían ubicarse enla capa de 0 a 10 cm. Encambio, la población deM. carimaguensis estaba dividida,al parecer, en dos grupos: losadultos aún se encontraban enlos primeros 20 cm y más abajohabía subadultos, individuosinmaduros y capullos (además dealgunos adultos). En enero de1995, no había ningún estímuloque hiciera retornar a suactividad a la comunidad delombrices (Jiménez et al. 2000);por ello, la población deGlossodrilus n. sp. estabacompuesta sólo de capullos eindividuos inmaduros, cada unode los cuales estivaba en elinterior de una esferatransparente de moco similar aun capullo (J. J. Jiménez, datossin publicar) en el estrato de 20 a40 cm. Algunos adultos ymuchos individuos inmaduros deM. carimaguensis se encontrabanentonces a una profundidad quevariaba entre 20 y 80 cm.
- Existió, al parecer, un retraso enel cese de actividades de lapoblación de Glossodrilus n. sp.que se hallaba en el pastizal, encomparación con la encontradaen la sabana. Cuando llegó laépoca de lluvias, todas estaslombrices se ubicaron de nuevoen el estrato de 0 a 10 cm. Sinembargo, los individuosinmaduros del pastizal
Figura 7-7. Humedad del suelo en diferentescapas del suelo en la épocalluviosa (julio de 1994) y en laépoca seca (febrero de 1995), en lasabana y en el pastizal estudiados.
12 14 16 18 20 22 24 26 28 30 32 34
Prof
un
dida
d (c
m)
0-10
10-20
20-30
30-40
40-50
Humedad (% peso suelo seco)
Sabana (época lluviosa)Pastizal (época lluviosa)Sabana (época seca)Pastizal (época seca)
Superficie
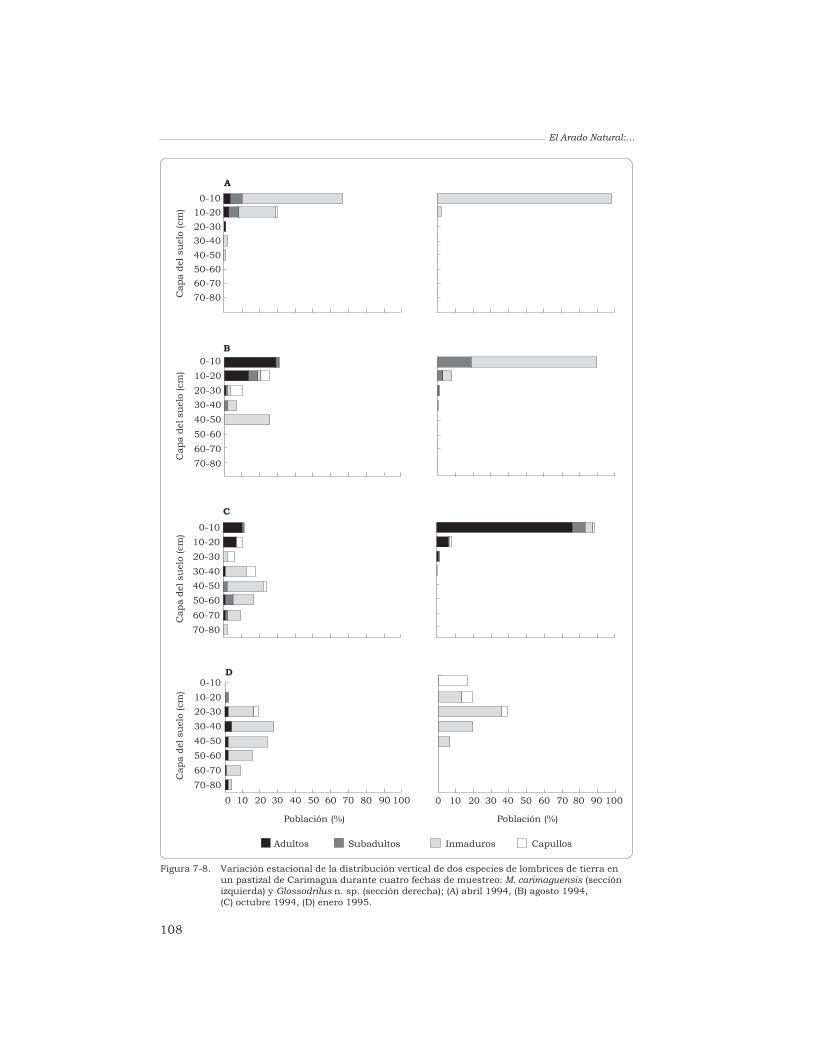
108
El Arado Natural:…
Figura 7-8. Variación estacional de la distribución vertical de dos especies de lombrices de tierra enun pastizal de Carimagua durante cuatro fechas de muestreo: M. carimaguensis (secciónizquierda) y Glossodrilus n. sp. (sección derecha); (A) abril 1994, (B) agosto 1994,(C) octubre 1994, (D) enero 1995.
Cap
a de
l su
elo
(cm
)A
0-1010-2020-3030-4040-5050-6060-7070-80
Cap
a de
l su
elo
(cm
)
B0-10
10-2020-3030-4040-5050-6060-7070-80
Cap
a de
l su
elo
(cm
)
C0-10
10-2020-3030-4040-5050-6060-7070-80
Población (%)
Cap
a de
l su
elo
(cm
)
D0-10
10-2020-3030-4040-5050-6060-7070-80
0 10 20 30 40 50 60 70 80 90 100 0 10 20 30 40 50 60 70 80 90 100
Población (%)
Adultos Subadultos Inmaduros Capullos

Distribución Vertical de las Lombrices...
109
estuvieron más meses en eseprimer estrato del suelo que losinmaduros de la sabana; al finalde la época lluviosa, toda lapoblación de esta especie estabaconformada por adultos.
Discusión
En general, la sustitución antrópicade un ecosistema original por unsistema diferente de uso de la tierracausa un gran impacto en elfuncionamiento de dicho ecosistema.Por ejemplo, las comunidades delombrices nativas son eliminadas odrásticamente reducidas cuando losecosistemas tropicales sonremplazados tanto por monocultivosanuales de alto nivel de insumosagrícolas o por pastizales de usointensivo (Barros et al. 1998; Fragoso1993; Fragoso y Lavelle 1992). Noobstante, la conversión de la sabanaen pastizal introducido no presentóefecto alguno en la distribuciónvertical de la comunidad delombrices estudiada en Carimagua.
La falta de una clara relaciónentre las variables biométricasempleadas, es decir, entre lasrelaciones L/D y P/D, y ladistribución vertical de las especiesde lombrices no difiere respecto a losresultados obtenidos en otrosestudios (Fragoso 1993; Lavelle1978; Németh y Herrera 1982); enéstos se reporta que las especies demayor tamaño se distribuyen en elperfil del suelo hasta unaprofundidad considerable. Losresultados de este trabajo puedenexplicarse por el hábito de vida de laespecie Ocnerodrilidae n. sp., unalombriz polihúmica (sensu Lavelle1981) muy pequeña (5 x 10-3 g PF y19.2 mm de longitud) que vive a unaprofundidad media de 25.4 cm en lasabana y de 22.7 cm en el pastizal.
Esta especie está, al menosparcialmente, asociada con lasgalerías horadadas porM. carimaguensis. Su distribuciónvertical dependerá, entonces, de lapresencia de galerías llenas de hecesde la especie anécica. Ocnerodrilidaen. sp. puede considerarse como unaespecie que pertenece a un grupo delombrices de tierra adaptadas aambientes muy particulares yespecíficos. Bouché (1972) propusoel término de lombrices ‘foleófilas’(“pholéophiles”, en francés) paraaquellas especies epígeas que seencuentran en las galerías de otrasespecies. Saussey (1956), Bouché(1970) y Piearce (1983) encontraronque la lombriz epígea Dendrobaenamammalis (Lumbricidae) habitaba lasgalerías de la endógea Allolobophoralonga (Lumbricidae). Ocnerodrilidaen. sp. sería un caso particular en elque una lombriz endógea estáasociada con las galerías construidaspor una lombriz anécica.
Las diferencias significativasobtenidas (P < 0.05) en el contenidode humedad de las tres primerascapas estudiadas del suelo, cuandoel sistema de uso de la tierra seincluyó como covariable, tendrían suorigen en varios factores. Porejemplo, el tipo de vegetación, lacantidad de hojarasca sobre lasuperficie del suelo, la distribuciónde las raíces en el suelo y laestructura física de éste, entre otros,pueden afectar las propiedadeshidráulicas del suelo, como sucontenido de agua, en un sistema oen otro.
Aunque la humedad del sueloejerce un efecto importante en ladistribución vertical de las lombrices,las diferencias encontradas entre losadultos y los individuos inmadurosde M. carimaguensis tienen,probablemente, una causa biótica.La pronta entrada en diapausa de los

110
El Arado Natural:…
inmaduros está fisiológicamenteinducida, ya que no hubo respuestaen ellos cuando fueron introducidosen un suelo humedecido hasta pF 2.8(Jiménez et al. 1998a). La razón deeste comportamiento sería unaestrategia adaptativa de las especiesgrandes que habitan la partesuperficial del suelo, con el fin deevitar densidades altas de individuosen las capas superiores durante elperíodo reproductor. Esto coincidecon Fragoso (1993), quien demostróque la competencia interespecíficapor nutrientes en las capassuperficiales del suelo puede ser unfactor que determine tanto el tamañocomo la distribución vertical de laslombrices endógeas.
Los resultados obtenidos en estey en otros estudios sugieren, comoconclusión obvia, que los factoresambientales influyen sobre el patrónde distribución vertical de lasespecies de lombrices de tierra,aunque hay factores de ordenfisiológico, en algunos casos, quepueden responder por dichocomportamiento. Los individuosinmaduros de M. carimaguensisdescienden en el suelo, en mitad dela época lluviosa, unas decenas decentímetros para iniciar la diapausa.Sería necesario emprender nuevosestudios en el futuro para encontrarlas razones que expliquen este patrónde comportamiento.
Lee (1959) clasificó la faunaoligoquetológica de Nueva Zelanda yconcluyó que las especies de lahojarasca presentaban una relaciónL/D que iba de 8 a 17, las de lascapas superficiales del suelo de 15 a40, y las de las capas más profundasde 20 a 130. Lavelle (1978) obtuvovalores similares en la familiaafricana Megascolecidae. EnCarimagua, Glossodrilus n. sp. esuna especie del suelo superficial,aunque es una excepción ya que surelación L/D es 53 (larga y fina).
Andiodrilus n. sp. y Glossodrilusn. sp. son especies que viven cerca dela superficie del suelo y que depositanlos capullos en los primeros 20 cm delperfil, principalmente en la capa de0 a 10 cm. Tras la eclosión de loscapullos, los individuos inmaduros deambas especies se localizan a esaprofundidad. A diferencia de ésta, M.carimaguensis es una especie anécicacuyos adultos depositan la mayorparte de sus capullos en la capa de20 a 30 cm; la profundidad media ala que se encontraron los capullos fuede 35 cm. Los individuos inmadurosrecién salidos de sus capullos al finalde la época lluviosa descienden en elsuelo para iniciar la diapausa(Jiménez et al. 1998a).
Tener un cuerpo de gran tamañopuede ser una adaptación pararesistir la sequía y para soportarlargos períodos de escasez dealimentos (Piearce 1983). Martiodriluscarimaguensis es la especie másgrande que haya sido encontrada enlas sabanas y en los pastizales deCarimagua y la estrategia queemplean sus individuos para superarla época seca es una de las mássorprendentes descubiertas hasta lafecha en los estudios sobrecomunidades de lombrices (Jiménezet al. 1998a). Una relación L/D altapuede ser importante para lograr unarespiración eficiente en un ambienteanaeróbico (Piearce 1983), como en elcaso de Allolobophora (sensu lato)oculata oculata (Lumbricidae) oHaplotaxis gordioides (Haplotaxidae).En Carimagua, Glossodrilus n. sp.presenta la mayor relación L/Dencontrada, lo que podría representaruna ventaja frente a otras especies,especialmente en Carimagua cuandolas lluvias fuertes inundan lasuperficie del suelo de los Llanosdurante cierto tiempo.
Las especies de lombrices quecoexisten en los sistemas de uso de latierra estudiados en Carimagua, tanto

Distribución Vertical de las Lombrices...
111
en el nativo como en el de pasturasintroducidas, están muyespecializadas; esto limita laexclusión competitiva puesto quehacen un uso diferencial de losrecursos. Aunque dos especies queviven cerca de la superficie del suelo,Andiodrilus n. sp. y Glossodrilus n.sp., muestran un grado alto desuperposición de nicho en la épocade lluvias respecto a su distribuciónvertical, debe reducirse, al parecer,su competencia a lo largo de algúnotro eje dimensional, por ejemplo,por la diferencia de tamaño de lasespecies, por la variación estacionalde su densidad poblacional, por lasestrategias adaptativas, y por el ciclode vida (J. J. Jiménez, datos sinpublicar). De este modo, las especiesexplotan diferentes capas del suelo yse alimentan de agregados del suelode diversos tamaños y de detritusorgánicos cuya tasa dedescomposición es variable.
La estrategia adaptativa de M.carimaguensis conduce a lareducción de la amplitud temporaldel nicho. Los individuos inmadurosdescienden hacia las capasprofundas del suelo para comenzarla diapausa 4 meses antes que losadultos, quienes inician después superiodo de reproducción. Puesto quelos adultos invierten casi el 16% desu peso en la formación de loscapullos —el porcentaje más altoregistrado hasta la fecha (Jiménez etal. 1999)— tal cantidad de energíadebe provenir, al parecer, derecursos orgánicos muy energéticos,como la hojarasca. Un númeroelevado de individuos, tantoinmaduros como adultos, reunidos aun tiempo en el mismo nicho,reducirían este recurso alimenticioenormemente e intensificarían lacompetencia intraespecífica.
Tanto los factores abióticos comolos bióticos (no cuantificados en esteestudio) influyen en el grado de
competencia que hay entre lasespecies de lombrices y permiten unareducción de ésta mediante lapartición de los recursos empleados.La especie que no reduzca lacompetencia en la dimensión verticaldel suelo lo hará en una de las otrasdos dimensiones principales, o sea,la temporal o la trófica (Giller 1984).
Por otro lado, las interaccionesbióticas crean patrones en lapartición de los recursos (Giller1984). Martiodrilus carimaguensis escapaz de producir diversasestructuras biogénicas, por ejemplo,galerías, cámaras de estivación ygran cantidad de heces superficiales,las cuales afectan tanto el ambientecircundante (Decaëns et al. 1999b)como a otros organismos del suelo(Decaëns et al. 1999a) y al banco desemillas del suelo (Decaëns et al.2003). Por consiguiente, se debeconsiderar el aspecto funcional deesta especie que es, sin duda alguna,un ingeniero del ecosistema (sensuJones et al. 1994), ya que moldea,directa o indirectamente, losrecursos y los hace disponibles paraotras especies imprimiendo cambiosfísicos en los materiales bióticos oabióticos (Decaëns 2000; Decaëns etal. 2001). Martiodrilus carimaguensiselabora estructuras cuando sealimenta, excava sus galerías ydeposita sus heces; las dos últimaspueden convertirse en un nuevohábitat para otros organismos. Nosólo la especie Ocnerodrilidae n. sp.resulta favorecida por estos procesos,sino también las raíces, las cualesencuentran cantidades considerablesde recursos tróficos en dichasestructuras. Una conclusión de estecapítulo sería, por tanto, que ladiversidad de obras de los ingenierosdel suelo propicia la diversidad enotros niveles tróficos, lo que daríaorigen a una comunidad debiodiversidades encajadas (Lavelle1996).

112
El Arado Natural:…
AgradecimientosEste trabajo se realizó gracias a unasubvención para la investigación enel proyecto Macrofauna (CE-STD2).Los autores agradecen al ProfesorPatrick Lavelle (IRD, Francia) y a losDrs. Richard J. Thomas y Myles J.Fisher (Centro Internacional deAgricultura Tropical, CIAT, Cali,Colombia) por el ánimo recibido deellos y por el entusiasmo quemanifestaron en el estudio del papelde las lombrices en las sabanas y enlos pastizales de Carimagua, unaestación situada en los LlanosOrientales de Colombia. El primerautor quiere agradecer a Jean-PierreRossi (IRD, Francia) y a dos revisoresanónimos por sus comentarios y suscríticas a una primera versión deeste artículo.
ReferenciasBarois I; Lavelle P. 1986. Changes in
respiration rate and somephysicochemical properties of atropical soil during transit throughPontoscolex corethrurus(Glossoscolecidae, Oligochaeta). SoilBiol Biochem 18:539-541.
Barros ME; Grimaldi M; Desjardins T;Sarrazin M; Chauvel A; Lavelle P.1998. Conversion of forest intopastures in Amazonia: effects onsoil macrofaunal diversity and soilwater dynamics. En: ISSS-AISS-IBG-SICS, eds. Proceedings of the16th World Congress of Soil Science,20-26 agosto 1998, Montpellier,Francia. p 203.
Bouché MB. 1970. Relations entre lesstructures spatialles et fonctionellesdes écosystèmes ilustrées par lerôle pédobiologique des vers deterre. En: Pesson P, ed. La vie dansle sol. Gauthiers-Villars, París.p 187-209.
Bouché MB. 1972. Lombriciens deFrance: ecologie et systematique.I.N.R.A., París. 671 p.
Curry JP. 1978. Relationships betweenmicroarthropod communities andsoil and vegetational types. Sci ProcR Dublin Soc 6:131-141.
Davidson DW. 1985. An experimentalstudy of diffuse competition inharvester ants. Amer Nat125:500-506.
Decaëns T. 2000. Degradation dynamicsof surface earthworm casts ingrasslands of the eastern plains ofColombia. Biol Fertil Soils32(2):149-156.
Decaëns T; Galvis J; Amézquita E. 2001.Properties of those structurescreated by soil ecological engineersfrom the Colombian savannas. C RAcad Sci Sér III 324:465-478.
Decaëns T; Mariani L; Lavelle P. 1999a.Soil surface macrofaunalcommunities associated withearthworm casts in grasslands ofthe eastern plains of Colombia.Appl Soil Ecol 13:87-100.
Decaëns T; Jiménez JJ; Lavelle P. 1999b.Effect of exclusion of the anecicearthworm Martiodriluscarimaguensis Jiménez and Morenoon soil properties and plant growthin grasslands of the eastern plainsof Colombia. Pedobiologia43:835-841.
Decaëns T; Mariani L; Betancourt N;Jiménez JJ. 2003. Seed dispersionby surface casting activities ofearthworms in Colombiangrasslands. Acta Oecol24(4):175-185.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental llanos ofColombia. Eur J Soil Biol30(4):157-168.
Fragoso C. 1985. Ecología general de laslombrices terrestres (Oligochaeta:Annelida) de la región Boca delChajul, Selva Lacandona (Chiapas,México). Tesis. UNAM, México.

Distribución Vertical de las Lombrices...
113
Fragoso C. 1993. Les peuplements de versde terre dans l’est et sud-est duMexique. Tesis (Doctorado).Université Paris 6. 228 p + anexos.
Fragoso C; Lavelle P. 1992. Earthwormcommunities of tropical rain forests.Soil Biol Biochem 24(12):1397-1408.
Garnsey RB. 1994. Seasonal activity andaestivation of lumbricid earthwormsin the midlands of Tasmania. Aust JSoil Res 32:1355-1367.
Gerard BM. 1967. Factors affectingearthworms in pastures. J AnimEcol 36:235-252.
Giller PS. 1984. The community structureand the niche. Chapman and Hall,Londres, Reino Unido.
Giller PS. 1996. The diversity of soilcommunities, the “poor man’stropical rainforest”. Biodiv Cons5:135-168.
Jiménez JJ; Moreno AG. Martiodriluscarimaguensis (Oligochaeta,Glossoscolecidae), una nuevaespecie de lombriz de tierra paraColombia. Megadrilogica. (Enrevisión.)
Jiménez JJ; Moreno AG; Lavelle P. 1999.Reproductive strategies of threenative earthworm species from thesavannas of Carimagua (Colombia).Pedobiologia 43:851-858.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innatural savannas and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.
Jiménez JJ; Brown GG; Decaëns T; FeijooA; Lavelle P. 2000. Differences inthe timing of diapause and patternsof aestivation in some tropicalearthworms. Pedobiologia44(6):677-694.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Kaczmarek M. 1975. An analysis ofCollembola communities in differentpine forest environments. EkolPolska 23:265-293.
Krishnamoorthy RV. 1985. A comparativestudy of wormcast production byearthworm populations fromgrassland and woodland sites nearBangalore, India. Rev Ecol Biol Sol22(2):209-219.
Lascano CL; Estrada J. 1989. Long-termproductivity of legume-based andpure grass pastures in the EasternPlains of Colombia. En: The FrenchGrassland Society, ed. Proceedingsof the XVI International GrasslandCongress, 4-11 octubre 1989, Niza,Francia. p 1179-1180.
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfonctions dans l’ècosystème. ParisVI. Publ Lab Zool ENS, 12. 301 p.
Lavelle P. 1981. Strategies dereproduction chez les vers de terre.Acta Oecol 2(2):117-133.
Lavelle P. 1983. The soil fauna of tropicalsavannas; II: The earthworms. En:Bourlière F, ed. Tropical savannas.Elsevier, Amsterdam. p 485-504.
Lavelle P. 1988. Earthworm activities andthe soil system. Biol Fertil Soils6:237-251.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lee K. 1959. The earthworm fauna of NewZealand. New Zealand Departmentof Scientific and IndustrialResearch Bulletin 130.

114
El Arado Natural:…
Lee K. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, NuevaYork. 411 p.
Lynch J; Balinsky E; Vail S. 1980.Foraging patterns in threesympatric forest ant species,Prenolepis imparis, Paratrechinamelanderi and Aphaenogaster rudis(Hymenoptera: Formicidae) in thesoil and litter layers of a Marylandforest. Amer Midl Nat 119:31-44.
Martin S; Lavelle P. 1992. A simulationmodel of the vertical movements ofan earthwom population (Millsoniaanomala, Omodeo, Megascolecidae)in an African savanna (Lamto, IvoryCoast). Soil Biol Biochem24(12):1419-1424.
Mato S; Mascato R; TrigoD; Díaz-CosínDJ. 1988. Vertical distribution insoil of earthworms in Sierra delCaurel. 1. Species and vegetationtypes. Pedobiologia 32:193-200.
Nemeth A; Herrera R. 1982. Earthwormpopulations in a Venezuelan rainforest. Pedobiologia 23:437-443.
Piearce TG. 1983. Functional morphologyof lumbricid earthworms withspecial reference to locomotion. JNat Hist 17(1):95-111.
Reddy MV; Pasha M. 1993. Influence ofrainfall, temperature and some soilphysico-chemical variables onseasonal population structure andvertical distribution of earthwormsin two semi-arid tropical grasslandsoils. Int J Biometeor 37:19-26.
Satchell JE. 1955. Some aspects ofearthworm ecology. En: Kevan DKMcE, ed. Soil zoology. Butterworth,Londres. p 180-201.
Saussey M. 1956. Un cas decommensallisme chez leslombriciens. Bull Soc Fr81:411-413.
Senapati BK. 1980. Aspects ofecophysiological studies on tropicalearthworms: distribution,population dynamics, production,energetics and their role in thedecomposition process. Tesis(Ph.D.). Sambalpur University,India. 154 p.
Shannon CE; Weaner W. 1949. Themathematical theory ofcommunication. University ofIllinois Press, Urbana, IL, EstadosUnidos.
Vegter JJ. 1983. Food and habitatspecialisation in coexistingspringtails (Collembola,Ebntomobryidae). Pedobiologia25:253-262.

Distribución Espacial de las Lombrices...
115
ResumenMediante el cómputo de índices deagregación y empleando lageoestadística, se estudió ladistribución espacial horizontal(DEH) de las lombrices de tierra enuna sabana nativa y en un pastizalde 17 años de edad, en Carimagua(Colombia). Los índices Id deMorisita y b de Taylor se calcularona partir de los datos de campoobtenidos en muestras de suelo de1 m2 (monolitos) durante 17 meses;tanto los semivariogramas como losmapas de densidad se obtuvieron entres fechas diferentes mediante unmuestreo rápido de 64 bloques desuelo de 40 x 40 x 15 cm,distribuidos regularmente en losnodos de una red de 8 x 8 puntos (osea, 70 x 70 m).
En los dos sistemas de uso de latierra estudiados, se observó en laslombrices la tendencia a unadistribución contagiosa, es decir, su
DEH resultó agregada; se encontró,además, una relación multivariadaentre el tamaño de las lombrices ylos índices de agregación empleados.El sistema de uso de la tierra noafectó la distribución espacial de laslombrices, las cuales pertenecían acategorías ecológicas diferentes, erande tamaño variado y tenían supoblación distribuida en parches devarias decenas de metros. Lossemivariogramas obtenidosreflejaban, en ocasiones, el tamañode estos parches; en otras fechas demuestreo no se obtuvo ningúnpatrón espacial. La causa de estaausencia pudo ser la distancia quehabía entre los puntos de muestreo(10 m), de modo que si se cambia laescala de observación podríanaparecer patrones no visualizadosanteriormente. El objetivo de esteestudio fue, en primer lugar,determinar la intensidad de laagregación espacial de los individuosde algunas especies de lombrices dela zona neotropical y, en segundolugar, observar el movimiento de losparches de población en lasdimensiones del área estudiada.Este tipo de análisis espacial,empleado junto con otros métodos,puede ser muy útil para establecer ladinámica de las poblaciones delombrices de tierra en condicionesnaturales.
CAPÍTULO 8
Distribución Espacial de lasLombrices en las Sabanas de SuelosAcidos de los Llanos Orientales deColombia1
J. J. Jiménez* , J.-P. Rossi** y P. Lavelle**
1. Publicado en Applied Soil Ecology 17(3),p 267-278, por Jiménez et al. Copyright ©2001 de Elsevier. Impreso con permiso de lacasa editorial.
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** Laboratoire d’Ecologie et Biologie des SolsTropicaux, IRD, 32 Av. Henri Varagnat,F-93143 Bondy Cedex, Francia.

116
El Arado Natural:…
Palabras clave: análisis decomponentes principales,distribución espacial, ecología depoblaciones, geoestadística,índice de agregación, sabanas
IntroducciónMuchos estudios se han realizadorecientemente sobre la abundancia yla distribución espacial de diversostaxones del reino animal que vivenen el suelo. El análisisgeoestadístico se ha empleado paradeterminar la distribución espacialde microorganismos y de colémbolos(Fromm et al. 1993), nematodos(Delaville et al. 1996; Ettema et al.1998; Robertson y Freckman 1995;Rossi et al. 1996; Wallace y Hawkins1994) y de lombrices de tierra (Poiery Richter 1992; Rossi et al. 1995,1997; Stein et al. 1992).
Aunque se sabe que ladistribución de las lombrices en elsuelo es irregular y agregada (Guild1955; Satchell 1955; Svendsen1957), Phillipson et al. (1976)trataron de cuantificar, por primeravez, el patrón espacial de laspoblaciones de lombrices, trabajoque fue continuado másdetalladamente por Boag et al.(1994), Decaëns y Rossi (2001),Fragoso (1993), Lavelle (1978,1983a, 1988), Rossi (1998), Rossi yLavelle (1998) y Rossi et al. (1995,1997). Estos estudios mostraron quela fauna del suelo formaba,generalmente, estructuras espacialesa escala pequeña (< 100 m)(Robertson 1994).
El conocimiento actual de losfactores que controlan los patronesde distribución espacial observadosen la macrofauna del suelo, o queinfluyen en ellos, es muy escaso. Sepresume que los factores abióticosexplican, al menos en parte, laestructura espacial que presentan
los macroinvertebrados del suelo.Ahora bien, varios estudios hanseñalado la ausencia de relaciónentre los factores abióticos y ladistribución espacial de variostaxones de invertebrados del suelo.Por ejemplo, Rossi et al. (1997)mostraron que la distribuciónespacial de la lombriz endógea(familia Megascolecidae)Polypheretima elongata Perrier eraindependiente del gradiente de lamateria orgánica del suelo, surecurso alimenticio. Ettema et al.(1998) demostraron, en un estudiosobre la dinámica espacio-temporalde nematodos bacteriófagos, la faltade correlación entre los nematodos ylos patrones de distribución de surecurso en el suelo, aunqueexplicaron los resultados por la pocaedad de las parcelas estudiadas. Losorganismos parecen estardistribuidos según ciertos factoresabióticos expresados a una escaladeterminada (por ejemplo, la texturadel suelo o la abundancia denematodos fitoparásitos [Wallace etal. 193]), aunque Rossi y Quénéhervé(1998) mostraron que estos tipos derelaciones pueden ser, en algunoscasos, muy débiles. Los patrones dedistribución espacial de estasespecies están controlados,generalmente, por muchos factoresque actúan a diferentes escalasespaciales y temporales.
La cubierta vegetal es uno de losfactores que influyen en ladistribución espacial de losorganismos del suelo, dando comoresultado un mosaico horizontal deáreas con gradientes diferentes dedisponibilidad de nutrientes y decondiciones microclimáticas (Lavelle1983b). Dicho patrón es,probablemente, más notorio en losecosistemas de sabana que encualquier otro, en razón de la fuerteestacionalidad del medio, que afectatanto al gradiente de temperatura

Distribución Espacial de las Lombrices...
117
como al de humedad. Los sistemasde uso de la tierra y las prácticasagrícolas, por otro lado, afectandirectamente el patrón dedistribución en parches de losrecursos del suelo (Robertson et al.1993). A su vez, se supone que uncambio en la heterogeneidad de unrecurso del suelo causaría un efectopotencial muy fuerte en la estructurade la comunidad vegetal y en ladistribución de los organismos delsuelo (Tilman 1988). Asimismo, lacomposición de la comunidad vegetalestaría determinada por el tipo depráctica agrícola empleada y por elestado de los nutrientes del suelo(Miles 1985; Wardle y Lavelle 1997).Más aún, si la comunidad vegetalinfluye, por ejemplo, en lacomposición de las termitas y laslombrices (Lavelle et al. 1997),debería afectar también sudistribución espacial. El impacto deestos organismos en elfuncionamiento del suelo estácondicionado, obviamente, por elefecto per capita y también por lalocalización espacial de losindividuos. Cuando se valoran losefectos de los ingenieros delecosistema (sensu Jones et al. 1994),lo anterior cobra una relevanciaespecial.
El objetivo de este trabajo fueanalizar el efecto potencial queejercería el cambio de sabana nativaen pastizal mejorado —cambiodebido a la introducción degramíneas africanas y leguminosasherbáceas tropicales destinadas alpastoreo— en la distribución espacialde las lombrices de tierra y en suscaracteres morfológicos. Se examinótambién la relación existente, segúnel uso dado a la tierra, entre ladistribución espacial de las lombricesy los parámetros morfológicos deéstas, así como la utilidad de lastécnicas geoestadísticas paradescribir convenientemente ladistribución espacial de las
lombrices, dada la distancia que sedejó en el terreno entre una muestray otra.
Se calcularon diferentes índicesde agregación de las lombrices enuna sabana natural y en otraperturbada, con el fin de determinarel efecto de la perturbación en ladistribución espacial de suspoblaciones. Ahora bien, los índicesde agregación no consideran lalocalización real de las unidades opuntos de muestreo y, por tanto, nosuministran ninguna informaciónsobre la distribución espacial de losorganismos a una escala superior ala del tamaño de la unidad demuestreo (Rossi et al. 1995). Poresta razón se intentó caracterizarplenamente la distribución espacialde las lombrices empleando dostécnicas: el diseño de un esquemade muestreo que sea explícito ensentido espacial, y el análisisgeoestadístico aplicado alprocesamiento de los datos. Estemétodo permite valorar la coherenciade los patrones de distribuciónespacial así como la escala en queéstos se manifiestan (Robertson1994).
Materiales y Métodos
Descripción del sitio de estudioEl estudio se llevó a cabo en laestación experimental de Carimagua(del convenio CIAT-CORPOICA)situada en las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia(4°37’ N y 71°19’ O), a 175 msnm. Elvalor medio anual de la precipitacióny de la temperatura es 2280 mm y26 °C, respectivamente, y la épocaseca se extiende desde diciembrehasta marzo. Los suelos de esa áreason de dos tipos: Oxisoles de bajafertilidad en las zonas altas (los

118
El Arado Natural:…
“altos”) y Ultisoles en las sabanasbajas (los “bajos”). Los primeros secaracterizan por su acidez (pH 4.5,en agua), su alta saturación de Al(> 90%) y el valor bajo de suscationes intercambiables Ca, Mg y K.
La investigación se realizó en dossistemas de uso de uso de la tierraque contrastaban entre sí:
- una sabana nativa donde no sepracticaban la quema ni elpastoreo (manejo tradicional) ycuyas especies vegetales másabundantes eran Andropogonbicornis, Gymnopogon sp.,Panicum spp., Trachypogon spp.e Imperata sp.;
- un pastizal que llevaba 17 añossometido a pastoreo, en el que secombinaban una gramíneaafricana (Brachiaria decumbenscv. Basilisk) y una leguminosaherbácea tropical (Puerariaphaseoloides CIAT 9900)conocida como “kudzú”.
Historial biológico de laslombricesEn las parcelas estudiadas, lacomunidad de lombrices estabacompuesta por ocho especies nativasque también se encontraron en elpastizal introducido (Jiménez et al.1998b). La mayoría de estasespecies pertenece a la familiaGlossoscolecidae, la más abundanteen la región neotropical: Andiodrilussp. (endógea mesohúmica),Andiorrhinus sp. (endoanécica),Andiorrhinus sp. 2 (endoanécica?),Aymara sp. (epígea), Glossodrilus sp.(endógea polihúmica) y Martiodriluscarimaguensis (anécica). Hay dosgéneros nuevos pertenecientes uno ala familia Ocnerodrilidae (endógeapolihúmica) y otro a la familiaAcanthodrilidae (epígea) (Jiménez yMoreno, datos sin publicar). Haygrandes diferencias en la estructura
de las comunidades de lombricesencontradas en las parcelas desabana y de pastizal mejorado. En lacomunidad de sabana, las especiesendógeas constituyen el 83.2% de labiomasa de todas las lombrices, hayalrededor de 3.26 g PF/m2, yGlossodrilus sp. es la especie másabundante; en la comunidad depastizal, en cambio, las especiesendógeas sólo representan el 12% dela biomasa total de lombrices, hay57.1 g PF/m2, y la especie anécicaconstituye el 88% de esa biomasatotal (Jiménez et al. 1998b).
Muestreo de lombricesEn este estudio se emplearon dosestrategias de muestreo:
• Muestreo mensual. Durante17 meses (desde abril de 1994hasta septiembre de 1995,excepto en junio de 1994) serealizó un muestreo estratificadoal azar en ambos sistemas de usode la tierra. Las lombrices serecogieron manualmente decinco monolitos de suelo de1 x 1 x 0.5 m (Lavelle 1978) encada sistema y se llevaron allaboratorio; en ellas se midierontres variables: longitud (mm),diámetro preclitelar (mm) ypeso (g).
• Muestreo espacialmenteexplícito. En ambos sistemas(sabana y pastizal) se tomaronmuestras en 64 puntos ubicadosen los nodos de una tramaregular de 70 x 70 m. En cadapunto nodal se extraíarápidamente un monolito de40 x 40 x 15 cm, que seexaminaba manualmente paraanotar las especies presentes asícomo su categoría demográfica:adultos (con clitelo yprotuberancias sexualespresentes), individuos juveniles(subadultos, con protuberancias

Distribución Espacial de las Lombrices...
119
sexuales presentes pero sinclitelo visible), individuosinmaduros (sin clitelo niprotuberancias sexuales), ycapullos. El método, que seguíael esquema de Rossi y Lavelle(1993) debió ser adaptado acausa del gran tamaño de laespecie M. carimaguensis.Finalizado el muestreo, todas laslombrices fueron devueltas alagujero del suelo (en el nodo) dedonde se extrajeron. Con el finde evitar que, en diferentesfechas, se tomaran muestras enlos mismos puntos, las muestrassiguientes se tomaron siguiendoel curso de una espiral cuyoorigen estaba representado por elpunto muestreado en la primerafecha. Dado el tamaño de lasparcelas estudiadas (variashectáreas), la comparación deéstas con los puntos de muestreode las diferentes fechas hace queéstos se consideren idénticos.En los dos sistemas estudiados(sabana y pastizal) se tomaronmuestras en tres fechasdiferentes: para la sabananativa, en noviembre de 1993,noviembre de 1994 y mayo de1995; para el pastizal mejorado,en septiembre de 1993, octubrede 1994 y junio de 1995.
Análisis de los datos
Indices de agregaciónEn un primer intento de
determinar el patrón espacial de laslombrices se computaron dos índicesde agregación: el Id de Morisita(1959) y el b de Taylor (1961).
Indice de Morisita. Es precisoestablecer un índice que no seasensible a la media de la densidad depoblación cuando se compara laagregación espacial de poblacionesque presentan medias de densidad
muy diferentes. Morisita (1959,1971) propuso un índice deagregación independiente de lamedia de las muestras y del númerototal de individuos recolectados enlas muestras (∑x). Ahora bien, esteíndice está muy ligado al númerototal de unidades de muestreo. Elíndice se expresa así:
donde n es el número de unidadesde muestreo y Zi el recuento deindividuos en la unidad i demuestreo.
El índice de Morisita toma elvalor 1 cuando la distribución esaleatoria, un valor menor que 1 si ladistribución es regular, y un valormayor que 1 si la distribución esagregada. Un análisis de chi-cuadrado (χ2) permite decidir si ladistribución agregada es significativao no lo es (Elliot 1971).
Indice de Taylor. La LeyExponencial de Taylor (Taylor 1961,1984; Taylor et al. 1978) se basa enla relación empírica que se estableceentre la media m y la varianza s2 yse expresa mediante la siguientefunción exponencial:
s 2 = amb
Los parámetros a y b sonparámetros de población. Elparámetro b es una medidaintrínseca de la agregación de lapoblación y varía continuamentedesde cero, valor que toma en ladistribución regular (s2 = a, siendoa<1), pasando por el valor 1 en ladistribución al azar (s2 = m, siendoa = 1), hasta el valor ∞ en ladistribución muy agregada (Taylor1961).
El parámetro a es un factorescalar que depende del tamaño dela unidad de muestreo (Elliot 1971;

120
El Arado Natural:…
Taylor 1961). El exponente b seusa como índice de agregación; b seconsidera independiente de la mediapoblacional; de ese modo, es unvalor que mide la agregaciónindependientemente de los cambiosproducidos en la media de ladensidad poblacional (Taylor 1961,1984; Taylor et al. 1988).
Los parámetros a y b estándeterminados por la regresión linealexistente entre la media y lavarianza de las muestras, una vezejecutada la transformaciónlogarítmica de estas últimas:
log s2 = log a + b log mEl índice de agregación de la Ley
Exponencial de Taylor es propio decada especie (índice específico,Taylor 1961, 1988) y estárelacionado estrechamente con elciclo de vida de varias especies delombrices encontradas en Africa(Rossi y Lavelle 1998).
Análisis de ComponentesPrincipalesCon el fin de estudiar la relación
existente entre varios parámetrosbiológicos —por ejemplo, el peso deladulto (PA), la longitud del adulto(LA), el peso de los individuosinmaduros (PI), la longitud de losinmaduros (LI), la relación entre lalongitud y el diámetro (L/D), y larelación entre el peso y el diámetro(P/D)— de una parte, y ladistribución vertical anual (DV) juntocon los índices de agregación deMorisita y de Taylor, de la otra, sellevó a cabo un Análisis deComponentes Principales (ACP)(Webster y Oliver 1990). Paraconocer la influencia que ejercía elsistema de uso de la tierra en dichasvariables (o sea, en los índices deagregación y los caracteresbiológicos), cuyos valores seregistraron en las poblaciones de
lombrices presentes en las parcelasde pastizal y de sabana aquíestudiadas, estas parcelas seanalizaron conjuntamente.
GeoestadísticaLa Geoestadística, también
llamada Teoría de las VariablesRegionalizadas (Matheron 1965,1971), es una herramienta útil paradescribir la estructura espacial decualquier variable del suelo. Elaspecto central de la geoestadísticaes el semivariograma, o sea, lafunción que describe la evolución dela semi-varianza en relación con ladistancia entre los puntos demuestreo (Webster y McBratney1989). El semivariograma es la únicaherramienta sencilla realmenteimportante en la aplicación de lageoestadística a la distribuciónespacial de las variables del suelo(McBratney y Webster 1986). En lateoría de las variables regionalizadas,el parámetro (h) es la semi-varianzay se calcula a partir de la siguienteexpresión:
donde M (h) es el número de paresde muestras en cada intervalo dedistancia h (o sea, la distancia entredos puntos de muestreo), y Z (xi) yZ (xi+h) son los valores de la variableen dos puntos de muestreocualesquiera separados por unadistancia h.
El semivariograma es la gráficade la relación entre la semi-varianzay la distancia; la forma de esta curvaindica si la variable dependeespacialmente o no, es decir, si hayautocorrelación espacial. Cuantomás parecidos sean los valoresseparados por una distancia dada,menor será el valor de la semi-varianza. Esta aumenta,generalmente, al aumentar la
γ

Distribución Espacial de las Lombrices...
121
distancia y al disminuir la similitudentre los puntos de muestreo. Si lavariable no presenta una estructuraespacial, la semi-varianza fluctúaalrededor de la varianza de lamuestra y el semivariogramaobtenido será ‘plano’.
Los valores calculados de lasemi-varianza se adaptan a unmodelo teórico (McBratney y Webster1986; Rossi et al. 1995; Webster yOliver 1990) que se ajusta alsemivariograma calculado a partir delos valores obtenidos en lasmuestras. En general, sólo seemplean algunas funcionesautorizadas cuando elsemivariograma se ajusta a unmodelo; consultar a Rossi et al. 1995y a Webster 1985 para una revisióndel tema y de los modelos utilizadoshabitualmente. Si existeautocorrelación espacial, elsemivariograma alcanza un valormáximo (la varianza “sill” o ‘deumbral’) cuando se llega a una ciertadistancia entre los puntos demuestreo (el rango). El rango marcael límite de la dependencia espacial ode la autocorrelación de la variableanalizada en el semivariograma.Cuando las distancias son mayoresque el rango, la semi-varianzapermanece fija y genera así unameseta en el semivariograma. Elpunto en que hay intercepción delsemivariograma (punto de corte) es,generalmente, un valor diferente decero llamado la varianza “nugget” (osea, mínima, C0) que expresa lavariabilidad debida a patrones nopercibidos (y no determinados en elsemivariograma), es decir, los que sepresentan a una escala menor que ladistancia mínima que separa lospuntos de muestreo. La varianzamáxima (“sill”) menos la varianzaminima (“nugget”) es la varianzaespacial (C). Este valor correspondea la parte de la varianza total quepuede ser modelada por la
estructura espacial (McBratney yWebster 1986).
Los semivariogramas secalcularon aplicando el programainformático VAR 5 (Yost et al. 1989),que permite hacer una estimacióndel semivariograma y ajustar elmodelo mediante un procedimientomodificado de diferencias mínimascuadráticas (Cressie 1985).
Mapas de contornoLos mapas clásicos de contorno
fueron trazados con ayuda delcomando “gráfico de contorno” queposee el programa informáticoSigmaplot 4.0.
ResultadosRespecto al muestreo geoestadístico,la densidad de las poblaciones delombrices varió según la especie delombriz y según la fecha. En los dossistemas de uso de la tierraestudiados, la especie másabundante fue Glossodrilus sp. Enla sabana nativa, su densidad mediavarió de 18.9 individuos/m2
(noviembre de 1994) hasta 46.3individuos/m2 (noviembre de 1993);en el pastizal mejorado, esa densidadvarió de 36.9 individuos/m2 (1994)hasta 102.5 individuos/m2 (1995).La especie epígea Aymara sp. seregistró solamente en el muestreo de1995, en el cual su densidad fue de7.81 y 13.5 individuos/m2 en lasabana y en el pastizal,respectivamente. La densidad de M.carimaguensis y de Andiorrhinus sp.fue inferior a 1 individuo/m2, exceptoen el muestreo realizado en elpastizal en 1995, cuando la primeraespecie obtuvo 14.5 individuos/m2.La densidad de las lombricespertenecientes a Ocnerodrilidae fuetambién baja: en la sabana, su valorosciló desde 2 individuos/m2 en

122
El Arado Natural:…
1993, hasta 7 individuos/m2 en1995, y en el pastizal, desde 6.6individuos/m2 hasta 18.8individuos/m2 en 1994 y 1995,respectivamente. Finalmente, losvalores de densidad obtenidos paraAndiodrilus sp. oscilaron entre1.97 individuos/m2 (en 1993) hasta2.7 individuos/m2 (en 1995), en lasabana, y desde 3.5 individuos/m2
(en 1993) hasta 6.2 individuos/m2
(en 1995) en el pastizal. Los datosobtenidos sobre los diferentesestadios demográficos de todas lasespecies, tanto en la sabana como enel pastizal, aparecen resumidos enlos Cuadros 8-1 y 8-2.
La distribución espacial de lasespecies de lombrices se manifestócomo agregada. Tanto el índice Idcomo el b mostraron que todas lasespecies estaban fuertementeagrupadas en ambos sistemas, lasabana y el pastizal (Cuadro 8-3).Todos los coeficientes de regresiónfueron significativos a un nivel deconfianza del 1%, excepto en el casode M. carimaguensis (P < 0.05).Respecto al índice Id, las especiesAndiodrilus sp., M. carimaguensis yAymara sp. presentaron la mayoragregación en la sabana; en elpastizal, esta agregación alta seregistró en Andiodrilus sp.,Andiorrhinus sp. y Aymara sp. Todoslos valores obtenidos fueronsignificativos, es decir, diferentes de1, según la prueba de χ2.
El ACP mostró una relaciónmultivariada entre el tamaño de laslombrices y su agregación(Figura 8-1). El porcentaje de lavarianza total explicada por los dosprimeros ejes fue de 57.6% y 22.1%,respectivamente. Las variablesbiométricas y la distribución vertical,excepto la relación L/D, definieron eleje 1. El eje 2, por su parte,discrimina la relación L/D y los dosíndices de agregación empleados. Eleje 1 separó las especies grandes que
descienden en el suelo a bastanteprofundidad (M. carimaguensis) deaquellas más pequeñas que vivencerca de la superficie, mientras queel eje 2 separó las especies quepresentan una relación L/D alta(como Glossodrilus sp.) de aquéllasque tienen una distribución másagregada y una relación L/D menor(Andiodrilus sp.). El sistema de usode la tierra no tuvo un efectosignificativo en las variables, exceptoen la distribución vertical anual(prueba de t, P = 0.0123). Ladistribución de las especies sobre losejes factoriales 1 y 2 (Figura 8-2)indicó que el sistema de uso de latierra no ejerció ningún efectocoherente, de modo que ladistribución de las especies delombrices debe ser explicada porfactores biológicos.
La estructura espacial de losdiferentes estadios demográficos dela población de lombrices sólo pudovalorarse en algunossemivariogramas. Esto significa que,en la mayoría de los casos, ladistancia con que se separaron lospuntos de muestreo, es decir, 10 m,fue muy grande y no permitióestablecer correctamente laestructura espacial de la comunidadde lombrices estudiada. Es probableque los patrones de distribuciónagregados que indicaban los índicesse expresen, en la mayoría de loscasos, más allá de la escala de 10 m.Cuando los semivariogramasobtenidos fueron significativos,pertenecían, en su mayoría, al tipoesférico (“spherical”) (Figura 8-3) eindicaban que la variabilidadespacial se expresaba en escalas quevariaban desde 27.1 m hasta57.4 m (Cuadro 8-4).
Las poblaciones de lombricesestuvieron distribuidas, en ambossistemas de uso de la tierra, enparches de tamaño variable de unafecha de muestreo a otra. El tamaño
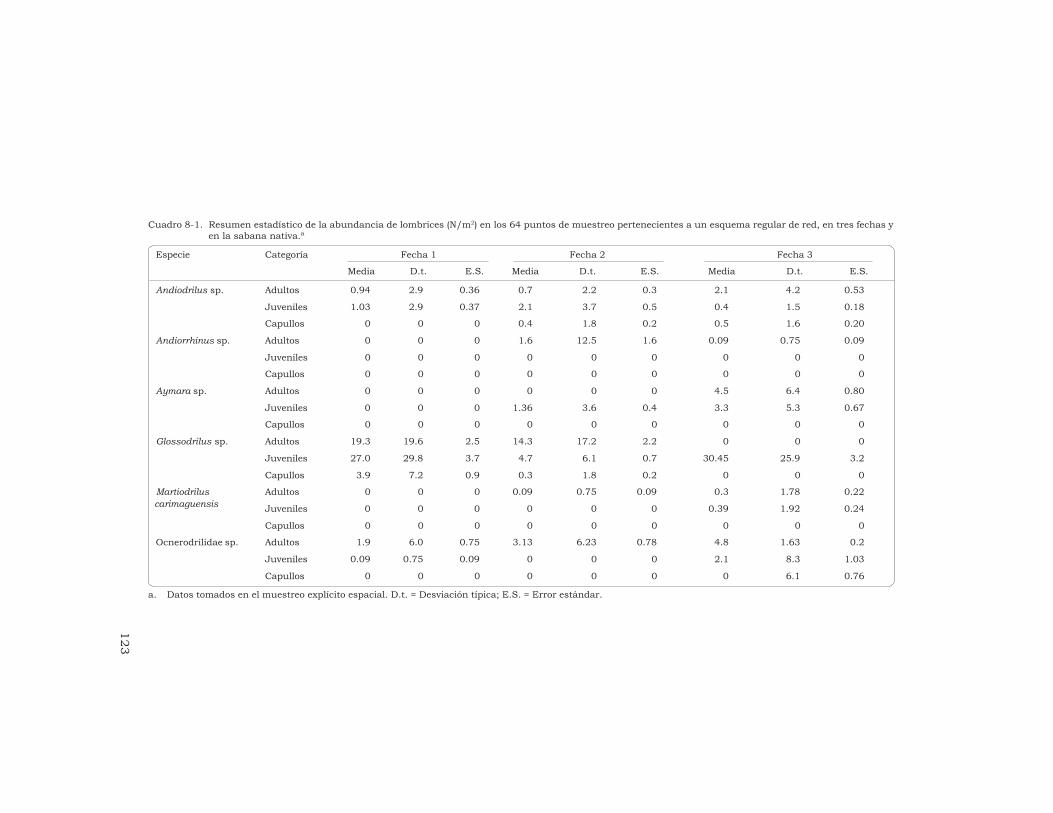
Distribución E
spacial de las Lombrices...
123
Cuadro 8-1. Resumen estadístico de la abundancia de lombrices (N/m2) en los 64 puntos de muestreo pertenecientes a un esquema regular de red, en tres fechas yen la sabana nativa.a
Especie Categoría Fecha 1 Fecha 2 Fecha 3
Media D.t. E.S. Media D.t. E.S. Media D.t. E.S.
Andiodrilus sp. Adultos 0.94 2.9 0.36 0.7 2.2 0.3 2.1 4.2 0.53
Juveniles 1.03 2.9 0.37 2.1 3.7 0.5 0.4 1.5 0.18
Capullos 0 0 0 0.4 1.8 0.2 0.5 1.6 0.20
Andiorrhinus sp. Adultos 0 0 0 1.6 12.5 1.6 0.09 0.75 0.09
Juveniles 0 0 0 0 0 0 0 0 0
Capullos 0 0 0 0 0 0 0 0 0
Aymara sp. Adultos 0 0 0 0 0 0 4.5 6.4 0.80
Juveniles 0 0 0 1.36 3.6 0.4 3.3 5.3 0.67
Capullos 0 0 0 0 0 0 0 0 0
Glossodrilus sp. Adultos 19.3 19.6 2.5 14.3 17.2 2.2 0 0 0
Juveniles 27.0 29.8 3.7 4.7 6.1 0.7 30.45 25.9 3.2
Capullos 3.9 7.2 0.9 0.3 1.8 0.2 0 0 0
Martiodrilus Adultos 0 0 0 0.09 0.75 0.09 0.3 1.78 0.22
Juveniles 0 0 0 0 0 0 0.39 1.92 0.24
Capullos 0 0 0 0 0 0 0 0 0
Ocnerodrilidae sp. Adultos 1.9 6.0 0.75 3.13 6.23 0.78 4.8 1.63 0.2
Juveniles 0.09 0.75 0.09 0 0 0 2.1 8.3 1.03
Capullos 0 0 0 0 0 0 0 6.1 0.76
a. Datos tomados en el muestreo explícito espacial. D.t. = Desviación típica; E.S. = Error estándar.
carimaguensis
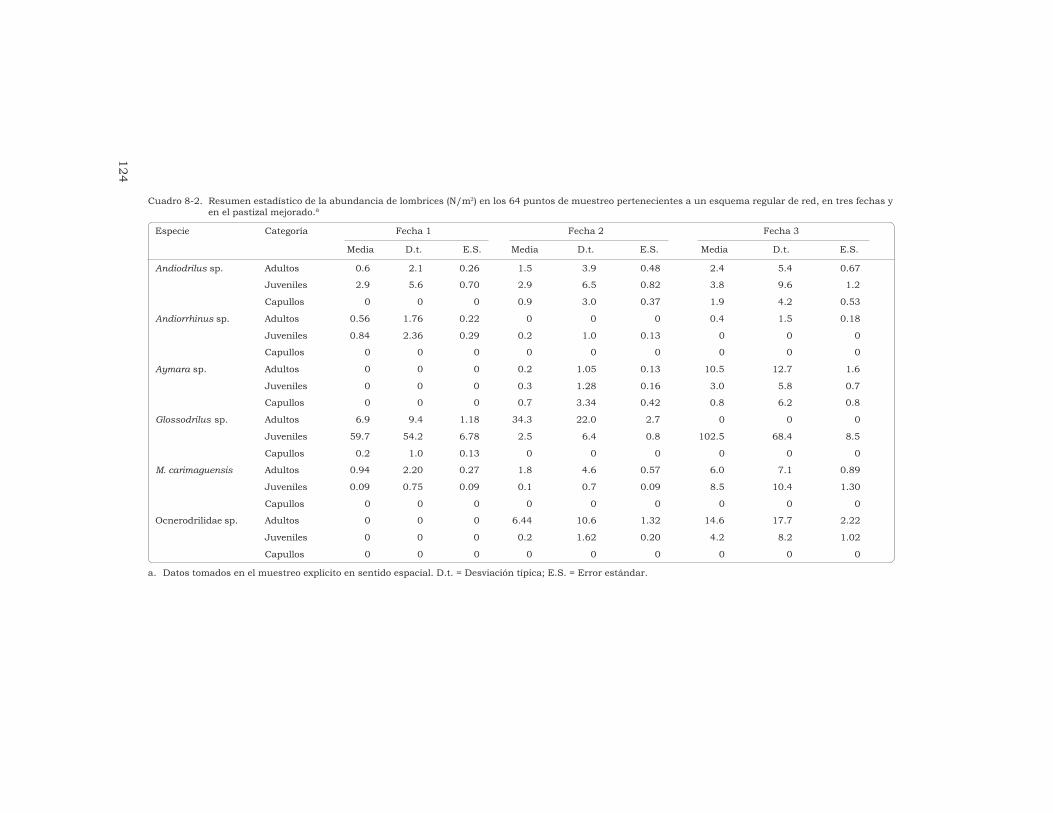
124
El A
rado Natural:…
Cuadro 8-2. Resumen estadístico de la abundancia de lombrices (N/m2) en los 64 puntos de muestreo pertenecientes a un esquema regular de red, en tres fechas yen el pastizal mejorado.a
Especie Categoría Fecha 1 Fecha 2 Fecha 3
Media D.t. E.S. Media D.t. E.S. Media D.t. E.S.
Andiodrilus sp. Adultos 0.6 2.1 0.26 1.5 3.9 0.48 2.4 5.4 0.67
Juveniles 2.9 5.6 0.70 2.9 6.5 0.82 3.8 9.6 1.2
Capullos 0 0 0 0.9 3.0 0.37 1.9 4.2 0.53
Andiorrhinus sp. Adultos 0.56 1.76 0.22 0 0 0 0.4 1.5 0.18
Juveniles 0.84 2.36 0.29 0.2 1.0 0.13 0 0 0
Capullos 0 0 0 0 0 0 0 0 0
Aymara sp. Adultos 0 0 0 0.2 1.05 0.13 10.5 12.7 1.6
Juveniles 0 0 0 0.3 1.28 0.16 3.0 5.8 0.7
Capullos 0 0 0 0.7 3.34 0.42 0.8 6.2 0.8
Glossodrilus sp. Adultos 6.9 9.4 1.18 34.3 22.0 2.7 0 0 0
Juveniles 59.7 54.2 6.78 2.5 6.4 0.8 102.5 68.4 8.5
Capullos 0.2 1.0 0.13 0 0 0 0 0 0
M. carimaguensis Adultos 0.94 2.20 0.27 1.8 4.6 0.57 6.0 7.1 0.89
Juveniles 0.09 0.75 0.09 0.1 0.7 0.09 8.5 10.4 1.30
Capullos 0 0 0 0 0 0 0 0 0
Ocnerodrilidae sp. Adultos 0 0 0 6.44 10.6 1.32 14.6 17.7 2.22
Juveniles 0 0 0 0.2 1.62 0.20 4.2 8.2 1.02
Capullos 0 0 0 0 0 0 0 0 0
a. Datos tomados en el muestreo explícito en sentido espacial. D.t. = Desviación típica; E.S. = Error estándar.

Distribución Espacial de las Lombrices...
125
Cuadro 8-3. Indices de agregación obtenidos para varias especies de lombrices de tierra en las parcelasde la sabana y del pastizal.
Especie Indice de Morisita Indice b de Taylor Valor ErrorSabana Pastizal Sabana Pastizal medio de b estándar
Andiodrilus sp. 2.903* 3.396* 1.889 1.704 1.797 0.131
Andiorrhinus sp. NDa 3.759* ND 1.801 1.801 NDAymara sp. 3.722* 3.505* 1.634 1.385 1.510 0.176Glossodrilus sp. 1.833* 2.160* 1.476 1.615 1.546 0.098M. carimaguensis 4.857* 1.137* 1.542 1.405 1.474 0.097Ocnerodrilidae sp. 2.604* 2.234* 1.579 1.603 1.591 0.017
a. ND = no determinado.* Indica un valor significativamente diferente de una distribución al azar (prueba de ‘chi´ cuadrado, χ2).
L/D
DV
Eje 1 (57.6%)
Eje
2 (2
2.1%
)
Indice Id
Indice b
LA
PA
PIP/D
LI
Figura 8-1. Mapa factorial del Análisis deComponentes Principales (ACP)ejecutado con algunas variablesbiométricas y con los dos índicesde agregación empleados.
11
-1-1
Eje 1 (57.6%)
M. carimaguensis(Pastizal)
Figura 8-2. Proyección de los objetos (las especies de lombrices de los dos sistemas, sabana y pastizal)en los dos primeros ejes extraídos del ACP.
Eje 2 (22.1%)
Glossodrilus sp.(Sabana)
Ocnerodrilidae sp.(Sabana)
Aymara sp.(Sabana)
Andiodrilus sp.(Pastizal)
Andiodrilus sp.(Sabana)
Andiorrhinus sp.(Pastizal)
Glossodrilus sp.(Pastizal)
Aymara sp. (Pastizal)
Ocnerodrilidae sp.(Pastizal)
Figura 8-3. Semivariograma esférico obtenidoen la especie Andiodrilus n. sp. conlos datos recolectados en elpastizal en junio de 1995.
3
2
1
0
Sem
i-va
rian
za
0 10 20 30 40 50Distancia (m)
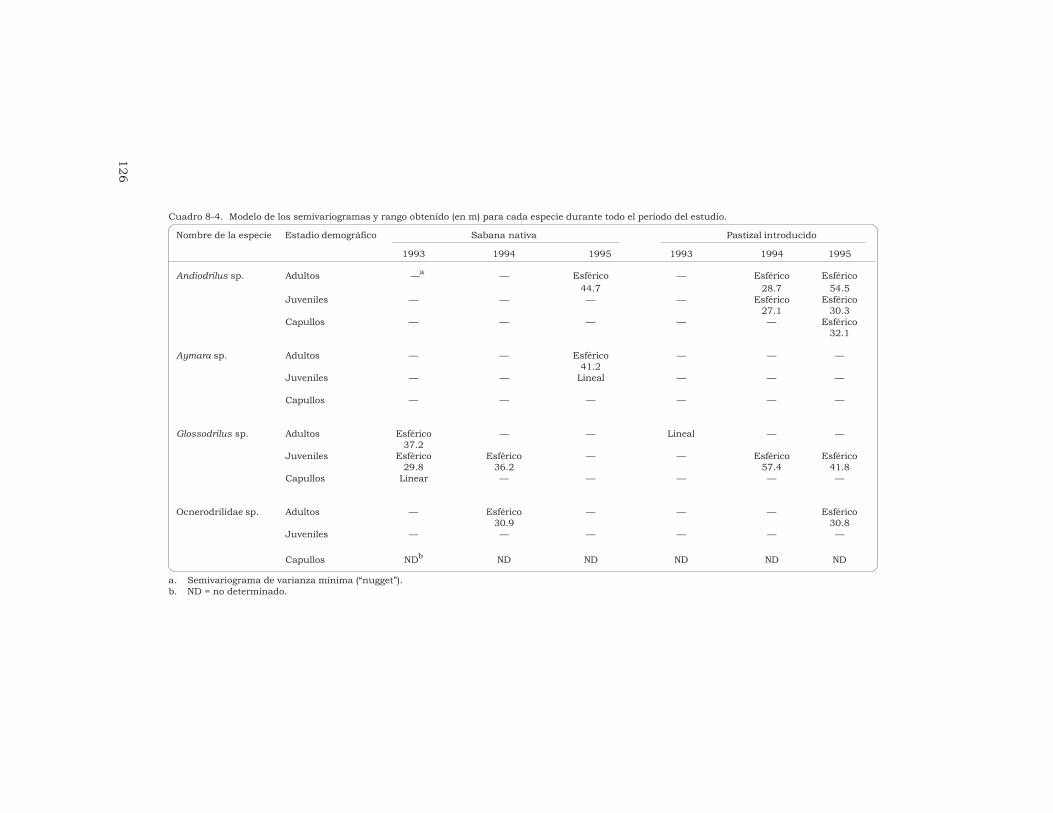
126
El A
rado Natural:…
Cuadro 8-4. Modelo de los semivariogramas y rango obtenido (en m) para cada especie durante todo el período del estudio.
Nombre de la especie Estadio demográfico Sabana nativa Pastizal introducido
1993 1994 1995 1993 1994 1995
Andiodrilus sp. Adultos —a — Esférico — Esférico Esférico44.7 28.7 54.5
Juveniles — — — — Esférico Esférico27.1 30.3
Capullos — — — — — Esférico32.1
Aymara sp. Adultos — — Esférico — — —41.2
Juveniles — — Lineal — — —
Capullos — — — — — —
Glossodrilus sp. Adultos Esférico — — Lineal — —37.2
Juveniles Esférico Esférico — — Esférico Esférico29.8 36.2 57.4 41.8
Capullos Linear — — — — —
Ocnerodrilidae sp. Adultos — Esférico — — — Esférico30.9 30.8
Juveniles — — — — — —
Capullos NDb ND ND ND ND ND
a. Semivariograma de varianza mínima (“nugget”).b. ND = no determinado.

Distribución Espacial de las Lombrices...
127
medio de los parches en que seagregaron las especies de lombricesfluctuaba entre 30 y 40 m, y a vecesera mayor. Por ejemplo, enseptiembre de 1993, casi todos losindividuos que constituían la especieGlossodrilus sp. en el pastizal eranjuveniles (Figura 8-4). En dichafigura se observan dos parches depoblación: uno de ellos es grande yse localiza en el lado izquierdo de laparcela (Figura 8-4,A) y el otro, unpoco menor que el primero, seencuentra en la esquina superiorderecha. En junio de 1995, latotalidad de la población estabaformada por individuos juveniles,
puesto que los adultos suelenaparecer al final de la época lluviosa(Jiménez et al. 1998b). Al parecer,el parche de población más grandese había movido hacia la izquierda yen la esquina superior derecha sedistinguían dos nuevos parches depoblación.
DiscusiónLas lombrices son organismos queadoptan una distribución espacialfuertemente agregada (Rossi yLavelle 1998). Los resultados deeste estudio coinciden con Piearce(1982), quien encontró que, sinconsiderar el índice de agregaciónempleado, las lombrices estabanmuy agrupadas en los suelos de lascostas noroccidentales de Inglaterra.Los valores obtenidos aquí para elíndice b de Taylor fueron muysimilares a los obtenidos por Tayloret al. (1978), quienes usaron losdatos de Gerard (1960). El valor altodel índice b obtenido en esteestudio para la especie Andiorrhinussp. puede explicarse por laabundancia de esta especie en losdos sistemas estudiados. Hairston(1959, citado en Lloyd [1967])concluye que los valores de losíndices de agregación son muy altospara las especies poco comunes yAndiorrhinus sp. es una especieescasa en Carimagua (Jiménez et al.1998b).
La evaluación de los índices deagregación depende de la escala demedición empleada y se aplica atodos los métodos que dan un valorcuantitativo a la varianza espacialde la distribución de cualquierorganismo (Horne y Schneider1995). Curtis y Mcintosh (1950) y,posteriormente, Pielou (1969) yGreig-Smith (1983) demostraron ladependencia de los índices deagregación respecto a la escalautilizada. Hurlbert (1990) concluyó,
Figura 8-4. Mapas de contorno que muestranla abundancia (N/m2) de individuosjuveniles de Glossodrilus sp. en elpastizal, en dos fechas diferentesde muestreo.
A
B
70
60
50
40
30
20
10
0
0 10 20 30 40 50 60 70
804040 40 40
12080
40
40
40
40 40
40
40
40
40
40
40
4040
40
40
80
80
80
80
80
80
80
80
80
80
80
40
120
120 120
120 120
120
120
160
160
160
160
200
240
80 160120
80
120
40
80
120
80
80
120
160
120
160 160 120
200
160200240
120
120 160
200240280
160
120
80
80 40
120
120
16012040
80 160 120
8040
160
120
80
4040
70
60
50
40
30
20
10
0
120 80
40

128
El Arado Natural:…
sin embargo, que dicha evaluaciónes una medida de la agregación sininterpretación alguna. En este caso,la comparación de los índices deagregación entre las poblaciones sóloserá válida cuando se realice en lamisma escala espacial.
Martiodrilus carimaguensis esuna especie cuyos individuos seencuentran activos en una mismagalería, al menos durante la primerasemana después de iniciada ladeposición de heces sobre lasuperficie del suelo (Decaëns 2000;Jiménez et al. 1998a). Esta especiehabita, de manera semipermanente,galerías verticales en forma de U(Jiménez y Decaëns, datos sinpublicar) y su comportamiento esafectado por la estacionalidadambiental. Lo normal es que lalombriz abandone la galería cuandoésta se satura de agua de lluvia, yaque la inundación ocasiona undéficit de O2 en el suelo (Jiménez etal. 1998a). Es posible que este tipode comportamiento ocasione losvalores bajos del índice Id obtenidosen el pastizal; es probable, también,que la caracterización de ladistribución espacial de lasdiferentes categorías demográficas,por ejemplo de los adultos y de losindividuos inmaduros, revelediferencias aún no percibidas.
Los dos índices empleados eneste estudio fueron una herramientavaliosa para determinar el patrónespacial y su correlación conparámetros biológicos, por ejemplo eltamaño de las lombrices. Este tipode relación se estableció también enel estudio realizado en unacomunidad de lombrices de lasabana africana de Lamto (Rossi yLavelle 1998). Estos dos autoresencontraron también que lasespecies más pequeñas presentabanuna fuerte agregación. EnCarimagua, las especies de tamañomedio y grande arrojaron los índices
de agregación más altos; por tanto,estas conclusiones deben extraerssecon sumo cuidado y es necesarioestudiar otras comunidades delombrices para establecer lospatrones que definan la agregaciónde las especies. Una explicaciónplausible de esta diferencia puedeser la ausencia de una lombrizanécica, como M. carimaguensis, enLamto. Coincidimos también con losautores mencionados en que estetipo de análisis debe emplearse paradefinir, de un modo más preciso, lascategorías ecológicas de laslombrices, especialmente si el índiceb es específico de cada especie,como sostuvieron Taylor et al. (1988).
Son varios los factoresresponsables del poco éxito obtenidoen el análisis geoestadístico:
1. Densidad. La densidad de unapoblación fluctúaestacionalmente a causa de lascondiciones ambientales. Elnúmero de lombrices al final dela época lluviosa (1993 y 1994)es un poco bajo en algunasespecies porque sus poblacionesiniciaron ya el período deinactividad.
2. Tamaño de las especies. Elmuestreo se realizómanualmente y no todos losindividuos son recolectadoscuando se aplica un método demuestreo manual. En algunasespecies, como en Aymara sp.,Glossodrilus sp. y Ocnerodrilidaen. gen. n. sp., queda sinrecolectar casi el 50% de lapoblación cuando se aplica estemétodo (Jiménez, datos sinpublicar).
3. Tamaño de la unidad demuestreo. Es posible que eltamaño elegido no fuerarealmente el óptimo como pararecolectar individuos de la

Distribución Espacial de las Lombrices...
129
especie anécica M.carimaguensis, que son de grantamaño y se retraen rápidamentehacia el interior de la galeríavertical.
4. Distancia entre las muestras.Las poblaciones de lombricespresentan una estructuraespacial agregada bastante clara,al menos en la escala de launidad de muestreo; esprobable, por tanto, que ladistancia mínima entre lospuntos de muestreo sea muygrande y no permita establecerun patrón claro de distribuciónespacial a una escala mayor quedicho valor mínimo. Losresultados de este estudiomuestran que algunas especiesestán agregadas en parches quetienen menos de 10 m dediámetro.
5. Estructura en mosaico. Martin(1991) desarrolló la hipótesis deque la especie Millsonia anomalaOmodeo presentabasubpoblaciones en diferentesfases demográficas. Es probableque los patrones observados enla comunidad de lombricesanalizada en ambos sistemas deuso de la tierra correspondan aun mosaico espacio-temporal deparches de población. Estahipótesis, sin embargo, debe serdemostrada en estudios futuros.
Los patrones de distribución ensentido horizontal de las lombricesson difíciles de establecer, ya quehan sido descritos en varias escalas,desde la local (Lavelle 1978;Phillipson et al. 1976) hasta laregional (Evans y Guild 1948;Németh 1981) y la geográfica(Bouché 1972; Fragoso 1993). Lavariabilidad de la distribuciónespacial horizontal de las lombricesva desde 1 m (Rossi 1998) hasta
50 m (Poier y Richter 1992). Laespecie endógea tropicalPolypheretima elongata Perrier sedistribuye en parches de poblaciónque miden de 20 a 30 m dediámetro, y presenta una notoriaoposición entre el patrón de losadultos y el de los individuosjuveniles (Rossi et al. 1997).Ninguno de los estadios demográficosde esta especie presentó unaautocorrelación con alguna variabledel suelo, por ejemplo la materiaorgánica o un gradiente en elcontenido de arcilla. En el estudioantes mencionado, los autoresexplican los patrones observadosbasándose en procesosdemográficos, como la competenciaintraespecífica que conduce a undescenso de la fecundidad en áreasdonde hay una densidad alta deadultos. El tamaño de los parchescoincide con el que reportaron Rossiy Lavelle (1998) y Rossi et al. (1997)para algunas especies de la sabanade Lamto (Costa de Marfil).Asimismo, el límite establecido parala distribución espacial de laslombrices de tierra en este estudio,que es el de parches menores que100 m, se ajusta al que describióRobertson (1994).
La estrategia de muestreoempleada en este estudio no fue muyútil al tratar de establecer, demanera precisa, la distribuciónespacial de las lombrices empleandoel análisis geoestadístico, inclusotras haber demostrado que lasespecies estaban agregadas (losíndices de agregación diferían de unadistribución al azar). La agregaciónes una característica de la poblaciónde lombrices de tierra, y fue fácildemostrarla empleando los dosíndices, Id y b; es necesario, sinembargo, aumentar la resolución delanálisis geoestadístico para mostrarel patrón de distribución real de laspoblaciones.

130
El Arado Natural:…
No se puede concluir, a partir delos resultados obtenidos, que ladistribución observada se debe afactores dinámicos, internos oexternos, incluidos en la poblaciónde lombrices. Rossi et al. (1997)establecieron la distribución espacialde P. elongata en las parcelas deproducción comercial de flores de laisla de Martinica, empleandoprocesos internos contenidos en lapoblación. En otro estudio, Rossi yQuénéhervé (1998) no encontraronninguna relación significativa entrelos nematodos, tanto de vida librecomo fitoparásitos, y variosparámetros de los suelos Vertisolesdel mismo lugar, excepto entre ladensidad del nematodo fitoparásitoHelicotylenchus retusus Siddiqui yBrown y el contenido de arcilla dedichos suelos, cuya correlación fuenegativa.
La perturbación de los sistemasecológicos es un factor determinanteen la estructuración de éstos puestoque crean un mosaico espacio-temporal de parches. Moloney yLevin (1996) sostienen que, engeneral, el patrón espacial de unaperturbación ha sido ignorado y sólose ha prestado atención a lacomprensión de los efectosprovocados por la tasa y laintensidad de la perturbación. Laestructura espacial de lasperturbaciones apenas ha sidoconsiderada en los estudiosrealizados sobre ese tema.Asimismo, la dinámica espacio-temporal de las variables del sueloen los sistemas perturbados, porejemplo la densidad de las lombrices,no ha sido estudiada enprofundidad, excepto por Decaëns yRossi (2001), Jiménez (1999) y Rossi(1998; datos sin publicar), quienessuministran información sobre estetema.
En este trabajo se ha resaltado lautilidad de las diferentes
herramientas estadísticas empleadasen el análisis espacial de laspoblaciones de lombrices. Lasprácticas del uso de la tierra, por suparte, no tuvieron un impacto claroen esa distribución, un hallazgo queresulta interesante. Este resultadoapoya la hipótesis de que ladistribución espacial de las lombricesestá determinada por factores queactúan dentro de sus poblaciones,aunque es necesario investigar máseste punto. Con ayuda de un métodonuevo para detectar modelos dedistribución espacial comunes en quese consideran diferentes fechas demuestreo (Jiménez et al., en revisión),se mostrará la estabilidad y ladistribución opuesta que presentanlas diferentes especies de lombrices.
AgradecimientosEste estudio fue incorporado alProyecto Macrofauna STD-2 de laUnión Europea. El primer autorquiere agradecer al CIAT (CentroInternacional de Agricultura Tropical)por el apoyo técnico y humanorecibido durante el tiempo delestudio, así como a T. Decaëns y ados evaluadores anónimos por susútiles comentarios a las primerasversiones de este trabajo.
ReferenciasBoag B; Legg RK; Neilson R; Palmer LF;
Hackett CA. 1994. The use ofTaylor’s Power Law to describe theaggregated distribution ofearthworms in permanent pastureand arable soil in Scotland.Pedobiologia 38:303-306.
Bouché MB. 1972. Lombriciens deFrance: ecologie et systematique.INRA, París. 671 p.
Cressie N. 1985. Fitting variogram modelsby weighted least squares. MathGeol 17:563-586.

Distribución Espacial de las Lombrices...
131
Curtis JT; Mcintosh RP. 1950. Theinterrelation of certain analytic andsynthetic phytosociologicalcharacters. Ecology 31:434-455.
Decaëns T. 2000. Degradation dynamicsof surface earthworm casts ingrasslands of the eastern plains ofColombia. Biol Fertil Soils 32:149-156.
Decaëns T; Rossi J-P. 2001.Spatiotemporal structure of anearthworm community and soilheterogeneity in a tropical pasture(Carimagua, Colombia). Ecography24:671-682.
Delaville L; Rossi J-P; Quénéhervé P.1996. Plant row and soil factorsinfluencing the micro-spatialpatterns of plant-parasiticnematodes on sugarcane inMartinique. Fundam Appl Nematol19(4):321-328.
Elliot JM. 1971. Some methods forstatistical analysis of sampling ofbenthic invertebrates. FreshwaterBiological Associates, The FerryHouse, Ambleside, Reino Unido.144 p.
Ettema CH; Coleman DC; Vellidis G;Lowrance R; Rathbun L. 1998.Spatiotemporal distribution ofbacterivorous nematodes and soilresources in a restored riparianwetland. Ecology 79(8):721-734.
Evans AC; Guild WJMcL. 1948. Studieson the relationships betweenearthworms and soil fertility; IV: Onthe life cycles of some BritishLumbricidae. Ann Appl Biol35:471-484.
Fragoso C. 1993. Les peuplements devers de terre dans l’est et sud-est duMexique. Tesis (Doctorado).Université Paris 6. 228 p. + anexos.
Fromm H; Winer K; Filser J; Hantschel R;Beese F. 1993. The influence of soiltype and cultivation system on thespatial distributions of the soilfauna and microorganisms andtheir interactions. Geoderma60:109-118.
Gerard BM. 1960. The biology of certainBritish earthworms in relation toenvironmental conditions. Tesis(Ph.D.). Londres, Reino Unido.
Greig-Smith P. 1983. Quantitative plantecology. University of CaliforniaPress, Berkeley, Estados Unidos.
Guild WJMcL. 1955. Earthworms and soilstructure. En: McKevan DK, ed. Soilzoology. Butterworths, Londres.p 83-98.
Horne JK; Schneider DC. 1995. Spatialvariance in ecology. Oikos 74:18-26.
Hurlbert SH. 1990. Spatial distribution ofmontane unicorn. Oikos 58:257-271.
Jiménez JJ. 1999. Estructura de lascomunidades y dinámica de laspoblaciones de lombrices de tierraen las sabanas naturales yperturbadas de Carimagua(Colombia). Tesis (Ph.D.).Universidad Complutense, Madrid.311 p.
Jiménez JJ; Decaëns T; Rossi J-P.Stability of the spatiotemporaldistribution and competition inneotropical earthwormassemblages. (En revisión.)
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Glossoscolecidae, Oligochaeta), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9(1-3):153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28(1):101-110.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.

132
El Arado Natural:…
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfonctions dans l’ècosystème. Tesis(Doctorado). Paris VI. Publ Lab ZoolENS, 12. 301 p.
Lavelle P. 1983a. The structure ofearthworm communities. En:Satchell JE, ed. Earthworm ecology:from Darwin to vermiculture.Chapman and Hall, Londres.p 449-466.
Lavelle P. 1983b. The soil fauna oftropical savannas. I. Thecommunity structure. En: BourlièreF, ed. Tropical savannas. Elsevier,Amsterdam. p 477-484.
Lavelle P. 1988 Assesing the abundanceand role of invertebratecommunities in tropical soils: Aimsand methods. J Afr Zool 102:275-283.
Lavelle P; Bignell D; Lepage M; Wolters V;Roger P; Ineson P; Heal OW;Dhillion S. 1997. Soil function in achanging world: the role ofinvertebrate ecosystem engineers.Eur J Soil Biol 33(4):159-193.
Lloyd M. 1967. Mean crowding. J AnimEcol 36:1-30.
Martin S. 1991. Modélisation de ladynamique et du rôle d’unepopulation de vers de terre(Millsonia anomala) dans lessavanes humides de Côte d’Ivoire.Tesis. Université Paris VI.
Matheron G. 1965. Les variablesregionalisées et leur estimation.Masson, París. 305 p.
Matheron G. 1971. The theory ofregionalized variables and itsapplication. Cahiers du Centre deMorphologie Mathématique, Nº 5.Fontainebleau, Francia.
McBratney AB; Webster R. 1986.Choosing functions forsemivariograms of soil propertiesand fitting them to samplingestimates. J Soil Sci 37:617-639.
Miles J. 1985. Soil in the ecosystem. En:Atkinson D; Fitter AH; Read DJ;Usher MB, eds. Ecologicalinteractions in soil, plants,microbes and animals. BlackwellScientific Publications, Oxford,Reino Unido. p 407-427.
Moloney KA; Levin SA. 1996. The effectsof disturbance architecture onlandscape-level populationdynamics. Ecology 77(2):375-394.
Morisita M. 1959. Measuring of thedispersion of individual andanalysis of distribution patterns.Mem Fac Sci Kyushu Univ Ser EBiol 2:215-235.
Morisita M. 1971. Composition of theId-index. Res Popul Ecol (Kyoto)13:1-27.
Németh A. 1981. Estudio ecológicopreliminar de las lombrices detierra (Oligochaeta) en ecosistemasde bosque húmedo tropical en SanCarlos de Río Negro, TerritorioFederal Amazonas. Tesis.Universidad Central de Venezuela.92 p.
Phillipson J; Abel R; Steel J; Woodell SRJ.1976. Earthworms and the factorsgoverning their distribution in anEnglish beechwood. Pedobiologia16:258-285.
Piearce TG. 1982. Recovery of earthwormpopulations following salt-waterflooding. Pedobiologia 24:91-100.
Pielou EC. 1969. An introduction tomathematical ecology. John Wiley,Nueva York, Estados Unidos.
Poier KR; Richter J. 1992. Spatialdistribution of earthworms and soilproperties in an arable loess soil.Soil Biol Biochem 24:1601-1608.
Robertson GP. 1994. The impact of soiland crop management practices onsoil spatial heterogeneity. En:Pankhurst CE; Doube BM; GuptaVVSR; Grace PR, eds. Soil biota:management in sustainable farmingsystems. CSIRO, Melbourne,Australia. p 156-161.

Distribución Espacial de las Lombrices...
133
Robertson GP; Crum JR; Ellis BG. 1993.The spatial variability of soilresources following long-termdisturbance. Oecologia 96:451-456.
Robertson GP; Freckman D. 1995. Thespatial distribution of nematodetrophic groups across a cultivatedecosystem. Ecology 76:1425-1432.
Rossi J-P. 1998. Rôle fonctionnel de ladistribution spatiale des vers deterre d’une savane humide de Côted’Ivoire. Tesis. Université Pierre etMarie Curie, París 6. 192 p.
Rossi J-P; Lavelle P. 1993. Statisticalprocedure for field surveys. En:Lavelle P, ed. Conservation of soilfertility in low-input agriculturalsystems by manipulatingearthworm communities. ProyectoEC Nº ERBTS3*CT920128. InformeNº 1. p 63-86.
Rossi J-P; Lavelle P. 1998. Earthwormaggregation in the savannas ofLamto (Côte d’Ivoire). Appl Soil Ecol7:195-199.
Rossi J-P; Quénéhervé P. 1998. Relatingspecies density to environmentalvariables in presence of spatialautocorrelation: a study case on soilnematodes distribution. Ecography21(2):117-123.
Rossi J-P; Delaville L; Quénéhervé P.1996. Microspatial structure of aplant-parasitic nematodecommunity in a sugarcane field inMartinique. Appl Soil Ecol 3:17-26.
Rossi J-P; Lavelle P; Albrecht A. 1997.Relationships between spatialpattern of the endogeic earthwormPolypheretima elongata and soilheterogeneity. Soil Biol Biochem29(3-4):485-488.
Rossi J-P; Lavelle P; Tondoh JE. 1995.Statistical tool for soil biology; X:Geostatistical analysis. Eur J SoilBiol 31(4):173-181.
Satchell JE. 1955. Some aspects ofearthworm ecology. In: McKevanDK, ed. Soil zoology. Butterworths,Londres, Reino Unido. p 180-201.
Stein A; Bekker RM; Blom JHC; RogaarH. 1992. Spatial variability ofearthworm populations in apermanent polder grassland. BiolFertil Soils 14:260-266.
Svendsen JA. 1957. The distribution ofLumbricidae in an area of Penninemoorland (Moor House NatureReserve). J Anim Ecol 26:423-439.
Taylor LR. 1961. Aggregation, varianceand the mean. Nature 189:732-735.
Taylor LR. 1984. Assessing andinterpreting the spatial distributionof insect populations. Annu RevEntomol 29:321-357.
Taylor LR; Woiwod IP; Perry JN. 1978.The density-dependence of spatialbehaviour and the rarity ofrandomness. J Anim Ecol 47:383-406.
Taylor LR; Perry JN; Woiwod IP; TaylorRAJ. 1988. Specificity of the spatialpower-law exponent in ecology andagriculture. Nature 332:721-722.
Tilman D. 1988. Plant strategies and thedynamics and structure of plantcommunities. Princeton UniversityPress, Princeton.
Wallace MK; Hawkins DM. 1994.Application of geostatistics in plantnematology. J Nematol 26:626-634.
Wallace MK; Rust RH; Hawkins DM;MacDonald DH. 1993. Correlationof edaphic factors with plant-parasitic nematode populationdensities in a forage field. J Nematol25:642-653.
Wardle D; Lavelle P. 1997. Linkagesbetween soil biota, plant litterquality and decomposition. En:Cadisch G; Giller KE, eds. Drivenby nature. CAB International,Wallingford, Reino Unido.p 107-125.
Webster R. 1985. Quantitative spatialanalysis of soil in the field. Adv SoilSci 3:1-70.

134
El Arado Natural:…
Webster R; McBratney AB. 1989. On theAkaike Information Criterion forchoosing models for variogramsof soil properties. J Soil Sci40:493-496.
Webster R; Oliver MA. 1990. Statisticalmethods in soil and land resourcesurvey. Oxford University Press,Londres, Reino Unido. 316 p.
Yost RS; Trangmar BB; Ndiaye JP;Yoshida NS. 1989. Geostatisticalsoftware for PC-DOS and MS-DOS.I. Semivariograms. Department ofAgronomy and Soil Science,University of Hawaii, Honolulu, HI,Estados Unidos.

Diferencias en la Programación de la Diapausa...
135
ResumenLa estivación es un período deinactividad que forma parte del ciclode vida de muchos organismos delsuelo. Gracias a la heterogeneidadgenética, fisiológica y ambiental delas lombrices de tierra, su estivaciónocurre de muchas maneras. En esteestudio se emplea el términoestivación en el sentido deinactividad de una población delombrices de tierra en cualquierépoca del año. Se muestran aquítambién las diferentes estrategias deestivación encontradas en algunaslombrices tropicales. Los resultadosobtenidos provienen de estudiosdetallados de las comunidades delombrices de tierra encontradas enlas sabanas de Colombia y México,en varios pastizales de este último
país, y en algunos agroecosistemasde Tanzania. Aunque todas lasespecies construían cámaras deestivación en las que se enrollaban enciertas épocas del año, tenían, sinembargo, diferencias en sus patronesde estivación. Por ejemplo, laslombrices de dos especiesneotropicales fueron halladas en elinterior de una esfera recubierta conuna secreción mucosa; las de otrasdos de dichas especies no formaronninguna estructura mucosa, perosellaron el extremo de su galería convarios tabiques construidos conheces; las de una especie africanaformaron su cámara de estivacióncon granos de arena grandes que seadherían al exterior de las lombricese impedían así que éstas tocaran lasuperficie interna de la cámara.
Este capítulo presenta unadescripción detallada, junto con losdibujos correspondientes, de lascámaras de estivación de cincoespecies de lombrices; trae ademásun análisis completo de los procesosque determinan el mecanismo de estepatrón de comportamiento en unaespecie anécica de Colombia. Elcomienzo de la estivación fuediferente para los adultos y losindividuos jóvenes de dos especiesneotropicales de la familiaGlossoscolecidae: Glossodrilus n. sp.y Martiodrilus carimaguensis. Los
CAPÍTULO 9
Diferencias en la Programación dela Diapausa y en los Patrones deEstivación de Algunas Especies deLombriz de Tierra del Trópico1
J. J. Jiménez* , G. G. Brown**, T. Decaëns***,A. Feijoo† y P. Lavelle**
1. Publicado en Pedobiologia 44(6), p 677-694,por Jiménez et al. Copyright © 2000 deUrban & Fischer Verlag. Impreso conpermiso de la casa editorial.
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** Laboratoire d’Ecologie et Biologie des SolsTropicaux, Université Pierre et Marie Curie,centre IRD d’Île de France, 32 Av. HenriVaragnat, F-93143 Bondy Cedex, Francia.
*** Laboratoire d’Ecologie, UFR Sciences etTechniques, Université de Rouen, F-76821Mont Saint Aignan Cedex, Francia.
† Proyecto de Laderas, CIAT, AA 6713, Cali,Colombia.

136
El Arado Natural:…
adultos de esta última especie, quees nativa de las sabanas deColombia, inician el proceso dediapausa después de horadar unagalería profunda en el suelo al finalde la época lluviosa; los individuosjóvenes inician ese proceso 4 mesesantes. Varios autores hanestablecido una relación entre lossegmentos nuevos añadidos alcuerpo de la lombriz y el período deestivación; en este estudio no sehalló ninguna relación de este tipoen la única especie que se estudiócon más detenimiento.
Palabras clave: diapausa, estivación,estrategias adaptativas,lombrices, sabanas
IntroducciónAlgunos organismos vivos tienencapacidad de entrar en una fase deinactividad o en una especie deletargo, cuando las condiciones delambiente son desfavorables. Laestivación2 es un período que formaparte del ciclo de vida de muchosorganismos del suelo y se puededefinir como la capacidad que tienende suspender su actividad duranteperíodos de tiempo variables. Casitodas las especies de lombrices detierra emplean este mecanismo comorespuesta a las fluctuacionesestacionales de la humedad y de latemperatura del suelo. Durante estafase de inactividad, los individuospermanecen en las capas profundasdel suelo inmóviles y sin alimentarse;pueden incluso experimentar lareabsorción de sus caracteressexuales primarios y secundarios.
De acuerdo con Bouché (1972,1984) y con Olive y Clark (1978, op.cit. Lee 1985), se distinguen tres
tipos de inactividad cuyos límites noestán definidos claramente; puedehaber, además, varios estadiosintermedios que dependen del estrésambiental (Lee 1985):
1. Quiescencia. Este procesoocurre cuando cambian lascondiciones ambientalesfavorables. Los individuosretoman la actividad cuando lascondiciones que iniciaron elproceso se vuelven favorablespara ellos. No construyencámaras de estivación ni seenroscan, sino que permanecenextendidos, por lo que puedenexperimentar una deshidrataciónmuy severa. Pueden vaciar elcontenido intestinal, pero suscaracteres sexuales no sonreabsorbidos.
2. Paradiapausa (= diapausafacultativa, Saussey 1966, op.cit.; Lee 1985). Este tipo deletargo es inducido por la pérdidade humedad del suelo. Lasupresión de los factores que loinducen propicia el reinicio de laactividad. Cada individuoconstruye una cámara deestivación y se enrolla en elladespués de vaciar su contenidodigestivo. Hay regresión de loscaracteres sexuales. En esteestado la deshidratación de laslombrices se reduce mucho.
3. Diapausa. Las condicionesadversas inducen, a nivelfisiológico, la diapausaobligatoria. Un factor ambientalle da origen, pero su final (o sea,el reinicio de la actividad) estádeterminado a nivel fisiológico. Adiferencia de la paradiapausa, nohay aquí respuesta de losindividuos cuando éstos sesometen a condiciones favorablesde un ambiente artificial, porejemplo, cuando las lombrices seintroducen en un suelo húmedo
2. Los autores se refieren a la inactividad deuna población animal en cualquier época delaño.

Diferencias en la Programación de la Diapausa...
137
hasta la capacidad de campo(pF 2.8). Los individuos quepasan por diapausa construyentambién una cámara deestivación, pierden peso porquevacían su contenido intestinal yno sufren deshidratación.También se produce, duranteesta fase, una reabsorción de loscaracteres sexuales y a veces laregeneración de algunos tejidos.
Muchos autores han descrito lasdiferentes estrategias que presentanlas lombrices al comenzar su fase dequiescencia o de diapausa (Abeloos yAvel 1928; Bouché 1972, 1984;Evans y Guild 1948; Lee 1951;Morgan y Winters 1991; Satchell1967; Saussey 1966). Sin embargo,en pocos estudios se ha ilustrado elpatrón de formación de las cámarasde estivación. El objetivo delpresente trabajo fue, de un lado,ilustrar gráficamente las diversascámaras de estivación observadas envarias especies tropicales dediferentes familias en doscontinentes y, del otro, establecer losdetalles de los mecanismos deestivación (inducción), tantofisiológicos como edáficos, en unalombriz anécica de las sabanas delos Llanos Orientales de Colombia.
Materiales y MétodosLa estivación de las lombrices detierra se estableció a partir tanto deestudios detallados de suscomunidades, que se realizarondurante 18 meses en Carimagua(Colombia) y en La Víbora (México),como de observaciones de campo enagroecosistemas cercanos a Gairo(Tanzania) y en Isla, Tuxpan yCarranza (México); esta investigaciónformó parte de tres proyectosinternacionales (financiados por laUE, IFS y AECI). Las lombrices quepresentaron estivación, así como sus
cámaras, se observaron durante unmuestreo manual de aquéllas adiferente profundidad, en monolitosde suelo de tamaño variable(Anderson e Ingram 1993; Lavelle1978).
Sitio experimental enColombiaEl trabajo principal de campo serealizó en la estación experimentalde Carimagua (CIAT-CORPOICA) enlos Llanos Orientales de Colombia(4°37’ N y 71°19’ O, 175 msnm). Elvalor medio anual de la precipitaciónfue de 2200 mm y el de latemperatura de 26 ºC (en el período1973-1995). La fuerteestacionalidad ambiental se reflejaen una época seca de 4 meses deduración. Los suelos se clasificancomo Oxisoles y Ultisoles infértiles(USDA 1975) y sobre ellos seasientan sabanas herbáceas conarbustos aislados y bosques degalería anastomosados; éstoscomprenden, principalmente,palmas del género Mauritia y estánasociados a los cursos de agua. Unaparcela de sabana natural y unpastizal introducido de gramínea yleguminosa dedicado al pastoreo seeligieron como sitios de muestreo.
Se tomaron muestras detalladasde las comunidades de lombricesdurante 18 meses (desde marzo de1994 hasta septiembre de 1995,excepto en junio de 1994); en esasmuestras se combinó el muestreomanual de bloques de suelo de1 m2 x 0.5 m, y el lavado ytamizado de monolitos pequeños de20 x 20 cm. Aunque la profundidadhabitual de muestreo fue de 50 cm,ésta varió según la época del año acausa de la migración vertical dealgunas especies de lombrices(Jiménez et al. 1998a). Los bloquesde suelo fueron divididos en capasde 10 cm de espesor y todas las

138
El Arado Natural:…
lombrices encontradas en ellas seintrodujeron en formol al 4% (segúnla metodología de Lavelle 1978). Elmuestreo finalizaba cuando, en unacapa determinada (nunca antes de50 cm) no se encontraba ningunalombriz. En algunos casos,especialmente en la época seca, laprofundidad de muestreo llegó a80 cm. Cada mes se extraían cincomonolitos de 1 m2 y 10 muestraspara lavado y tamizado, según undiseño de bloques estratificado ycompletamente al azar, para ambossistemas de uso de la tierra.
La comunidad de lombrices detierra de la sabana nativa y de lospastizales introducidos estabaconstituida por ocho especies, todasellas nativas (Cuadro 9-1). Lariqueza en especies es similar enambos sistemas, pero hay unaespecie en la sabana que no seencuentra en el pastizal. Sinembargo, los valores medios anualesde la densidad y de la biomasa de lacomunidad de lombrices de lasabana fue de 49.8 individuos/m2 yde 3.26 g pF/m2, respectivamente;estos valores fueron mucho másbajos que los obtenidos en el pastizal(80.1 individuos/m2 y 57.1 g pF/m2,respectivamente) a causa de laabundante población de Martiodrilus
carimaguensis en este último sistema.Esta especie contribuye allí con un88% a la biomasa total de lombricesde tierra (Jiménez et al. 1998b).
Sitios experimentales enMéxicoEstos sitios se localizan al oriente deMéxico, en el estado de Veracruz, enla franja de la costa atlántica (35 a80 msnm); presentan un climaestacional húmedo y caluroso, conuna precipitación anual superior a1200 mm. Hay, sin embargo, unaépoca seca de 4 a 6 meses,interrumpida intermitentemente porfuertes vientos procedentes del nortey por lluvias suaves llamadas“nortes.” En Isla y en La Víbora, lossuelos (un Ultisol y un Alfisol,respectivamente) son de texturaarenosa o franco-arenosa, pobres enmateria orgánica (< 1.5% C), cuyavegetación original era,probablemente, la de una sabanaherbácea y arbustiva. En amboslugares, la vegetación actual consisteen una gramínea nativa que sirve depastura. En La Víbora, el muestreose realizó mensualmente (desde juniode 1996 hasta septiembre de 1997);se extrajeron manualmente bloquesde suelo de 25 x 25 cm2, hasta una
Cuadro 9-1. Comunidad de lombrices de las sabanas y de los pastizales de Carimagua y tamañomáximo alcanzado por los adultos.
Especie Familia Categoría Peso adulto Longitud adulto ecológicaa (g)b (mm)c
Andiodrilus n. sp. Glossoscolecidae Endógea 1.3 ± 0.3 109 ± 14.4Andiorrhinus n. sp. 1 Glossoscolecidae Endo-anécica 7.1 ± 2.1 188 ± 27.6Andiorrhinus n. sp. 2 Glossoscolecidae Endo-anécica 4.9d –Aymara n. sp. Glossoscolecidae Epígea 0.06 ± 0.02 58.1 ± 10.3Glossodrilus n. sp. Glossoscolecidae Endógea 0.09 ± 0.02 83.9 ± 10.7Martiodrilus carimaguensis Glossoscolecidae Anécica 11.2 ± 2.1 194.3 ± 24.3Octochaetidae n. gen. Octochaetidae Epígea 0.02 ± 0.01 45.2 ± 8.1Ocnerodrilidae n. gen. Ocnerodrilidae Endógea 6.10-3 ± 2.10-3 22.8 ± 2.7
a. Anécica: vive en el suelo, pero se alimenta en la superficie; Epigea: vive y se alimenta en lasuperficie del suelo; Endógea: vive y se alimenta en el suelo (según Bouché 1972 y Lavelle 1981).
b. En formol.c. Los datos son medias ± desviación típica.d. Sólo un ejemplar.

Diferencias en la Programación de la Diapausa...
139
profundidad de 20 cm (Brown et al.sin publicar). Todas las lombricesencontradas se introdujeron enformol al 4% y se llevaron allaboratorio donde fueron pesadas.Se encontraron siete especies, unade ellas exótica y el resto nativas;tres de ellas eran nuevas para laciencia. En este capítulo se haráreferencia exclusivamente a losejemplares del género que pertenecea la familia Glossoscolecidae (gen.nov. 1). En Isla se tomaronmuestras en la época lluviosa(septiembre a octubre) y en la seca(mayo) de 1998 y 1999, mediantecalicatas rectangulares de 50 x25 cm que tenían de 40 a 60 cm deprofundidad.
En Carranza y Tuxpan, lossuelos son Vertisoles cuyo contenidode arcillas es alto (> 40%) y cuyariqueza en C (> 2.5%) es mayor queen los antes mencionados. Lavegetación original del sitio fue unbosque tropical semideciduo;actualmente, éste se ha convertidoen un pastizal de la especieintroducida Digitaria decumbens.Estos sitios fueron muestreados deigual manera y en las mismas fechasque los de Isla.
Otros sitios experimentales (enTanzania y en Colombia)El sitio de Tanzania se encuentracerca del pueblo de Rubeho (SE), aunos 900 msnm. La vegetaciónoriginal era el bosque de Miombo y elmuestreo (metodología TSBF;Anderson e Ingram 1993) se realizóen un campo de girasoles en 1996.El suelo es un Alfisol, de texturagruesa y con valores deC intermedios. El sitio, que seencuentra en el lado oeste de unacordillera orientada hacia el océano(al este), cerca de Gairo, secaracteriza por una época seca muyprolongada. La precipitación media
anual es de 700 mm que seregistran, en su mayor parte, entrenoviembre y febrero. Es bienconocida aquí la gran diversidad decienpiés (o milpiés), pero se sabepoco sobre la fauna relacionada conlas lombrices de tierra. Durante elmuestreo, el suelo estaba muy seco(8% p/p H2O). Las lombrices de lasmuestras se conservaron en formolal 4%.
El sitio de Colombia seencuentra en la cuenca del ríoCabuyal (departamento del Cauca),sobre las estribaciones de lacordillera andina, a 1400 msnm.Las muestras de lombrices de tierrase tomaron en un bosque secundariode más de 40 años, desde junio de1995 hasta febrero de 1997. Losvalores medios anuales deprecipitación y temperatura son1500 mm y 21 ºC, respectivamente,y el régimen de precipitación esbimodal. Hay dos épocas secas: dediciembre a febrero y de junio aagosto, y dos épocas lluviosas: demarzo a mayo y de septiembre anoviembre. Los suelos se definencomo Dystropept óxicos, provienende cenizas volcánicas, y son detextura fina, bien drenados yligeramente ácidos (pH = 5.3) (Feijoo,datos sin publicar).
Estivación vs. regeneración (enColombia)Se ha establecido una relación entrela estivación y la regeneración enM. carimaguensis, una especieanécica nativa que sobrevive a laépoca desfavorable del año mediantela diapausa. Se probó la hipótesisde que ciertas especies de lombricesincrementan el número desegmentos de su cuerpo durante esteperíodo de inactividad. Se contó, porello, el número de segmentos nuevosañadidos a la región posterior delcuerpo de las lombrices, tanto en los

140
El Arado Natural:…
adultos como en los individuossubadultos o immaduros que habíanpasado por la estivación.
Análisis de los datosLos análisis estadísticos se realizaroncon ayuda del paquete SPSS 6.0. Secalcularon las regresiones paraobtener las ecuaciones que mejor seajustaban a los datos. También seempleó el ANOVA de una sola vía deKruskal-Wallis (una prueba noparamétrica) empleando el númerode lombrices halladas a diferentesprofundidades en las muestras desuelo.
Resultados
Cámaras de estivaciónEn las lombrices de tierra dedistintas especies que serecolectaron durante la época secaen el interior de las cámaras deestivación se observaron diferentesestrategias adaptativas orientadas aevitar que se secaran los tejidos deesas lombrices. Estos diversospatrones de estivación se muestranen la Figura 9-1 (A-D):
A. Cámara de diapausa de unaespecie de gran tamaño (15 a30 cm de longitud) de lassabanas de Colombia: M.carimaguensis (Glossoscolecidae).
Cada individuo de esta especieconstruyó una cámara al final de lagalería en que habitaba de manerasemipermanente; se enroscó en lacámara después de vaciar sucontenido intestinal y suspendió suactividad hasta el comienzo de lasiguiente temporada de lluvias. Lainactivación se produjo después deque la especie descendió hasta unaprofundidad de 60 a 110 cm. Casisiempre, el extremo de la galería
estaba sellado con heces queformaban tabiques con el fin deevitar la pérdida de humedadcorporal de la lombriz, humedad quees vital para mantener una tasamínima de respiración. Hubodiferencias en el comienzo de laestivación de los adultos y de losindividuos jóvenes (Jiménez et al.1998a).
B. Cámara de estivación de laespecie pequeña (de 2 a 2.5 cm)feofílica (“feófila”) (Bouché 1972;Lee 1985) perteneciente aOcnerodrilidae n. gen. y de laespecie epígea Aymara n. sp.(Glossoscolecidae), halladas enCarimagua, Colombia.
Los individuos de estas especiesformaron una cámara de estivaciónal final de su galería pero no lasellaron, a diferencia de los deM. carimaguensis.
C. Cámara de estivación deGlossodrilus n. sp.(Glossoscolecidae), especie detamaño pequeño (de 8 a 10 cm)de los Llanos Orientales deColombia.
Estas lombrices se rodean deuna esfera mucosa cuya estructuraes muy similar a la de un capullo,aunque de mayor tamaño (unos5 mm de diámetro); los capullos deesta especie son muy pequeños,ovales, y miden 3.0 x 2.2 mm(Jiménez et al. 1999). La cámara nodebe confundirse con un capullocuando se practica un muestreo.
D. Cámara de estivación de unalombriz de tamaño intermedio(de 10 a 15 cm) de Tanzania aúnno clasificada, y paraChuniodrilus zielae Omodeo 1963(Eudrilidae) de Lamto, en Costade Marfil.
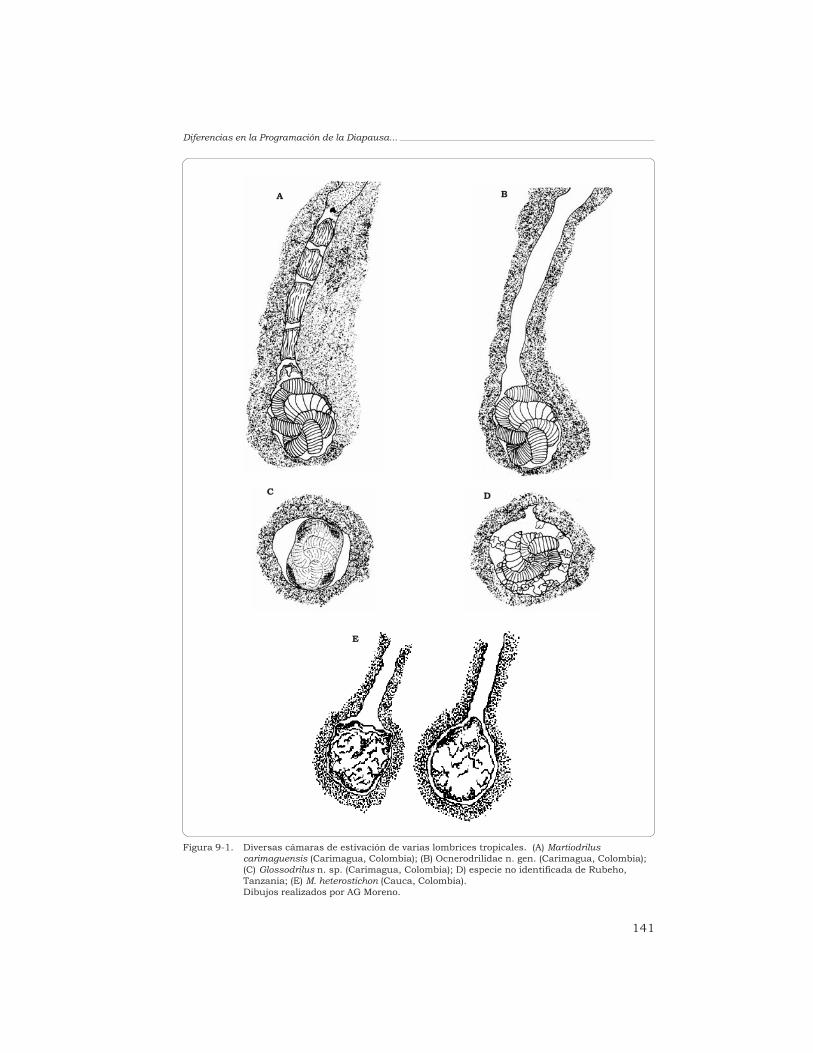
Diferencias en la Programación de la Diapausa...
141
Figura 9-1. Diversas cámaras de estivación de varias lombrices tropicales. (A) Martiodriluscarimaguensis (Carimagua, Colombia); (B) Ocnerodrilidae n. gen. (Carimagua, Colombia);(C) Glossodrilus n. sp. (Carimagua, Colombia); D) especie no identificada de Rubeho,Tanzania; (E) M. heterostichon (Cauca, Colombia).Dibujos realizados por AG Moreno.
A B
C D
E

142
El Arado Natural:…
Estas lombrices hacen que losgranos grandes de arena quedenadheridos a la superficie interna dela cámara para evitar,probablemente, que se seque eltegumento de su cuerpo, expuesto aperder humedad hacia dichasuperficie interna de suelo. Hace yamás de un siglo, Darwin (1881) yHensen (1877) observaronfenómenos similares. Esa superficieinterna está recubierta con moco(que le da un aspecto brillante) yprobablemente con arcillas que lalombriz compactaría con la presiónde su cuerpo. Este recubrimientoayudó a evitar también la pérdida deagua hacia el suelo seco circundante.
E. Cámara de estivación de unalombriz de tamaño intermedio(de 8 a 12 cm de longitud), aúnno descrita, pero perteneciente ala subfamilia Acanthodrilinae, deIsla, México.
Esta especie formó una cámara auna profundidad de 30 a 50 cm y lausó incluso al final de la épocalluviosa cuando el medio estaba tanseco que podía inducir el proceso deestivación. Las cámaras eranestructuras discretas que podíansepararse fácilmente del suelocircundante y su interior estabarecubierto con una capa mucosa yarcillosa. Para construir estacámara, la lombriz debió seleccionarpartículas arcillosas de un suelo detextura franco-arenosa y depositarlasluego de manera que formaran unaesfera completa, con la cual secubrió para enroscarse en suinterior.
F. Cámara de estivación de ungénero nuevo de lombriz de tierrade tamaño intermedio (de 8 a12 cm) perteneciente aGlossoscolecidae gen. nov. 1, deLa Víbora, México.
Esta cámara puede considerarseintermedia entre la que construye la
especie de Isla y la del género nuevode la familia Ocnerodrilidae deColombia. La lombriz formó unacámara cuyas paredes internas,recubiertas con moco (yprobablemente con arcillas) yligeramente compactadas, lepermitieron enrollarse como unovillo. La cámara como tal quedabacerrada, aunque esta especie noforma estructuras discretas como laespecie hallada en Isla.
G. Cámara de estivación deMartiodrilus heterostichonSchmarda 1861(Glossoscolecidae), deldepartamento del Cauca,Colombia.
Esta especie endógeaoligohúmica construye una cámara a30 cm de profundidad (Figura 9-1,E),como mínimo, que usa incluso alfinal de la época lluviosa si lahumedad del suelo desciende amenos de 22% (p/p H2O), nivel alque se induce la diapausa. Lacámara observada era unaestructura discreta que podíasepararse completamente del restodel suelo y estaba recubierta en suinterior con moco y partículasarcillosas. Para construir estacámara, la lombriz debió seleccionarpartículas arcillosas de un suelo detextura franco-arenosa y depositarlasluego de manera que formaran unaestructura globosa, con la cual secubrió para enroscarse en suinterior. Cuando empieza la épocalluviosa, la lombriz abandona lacámara en cuyo interior hay heces, yasciende hacia los primeros 20 cmdel suelo. Es muy común encontrarestas cámaras llenas de heces, quetambién se encuentran en lasgalerías que abre la lombriz.
Patrones de estivaciónSe han observado diferentesestrategias de estivación en las
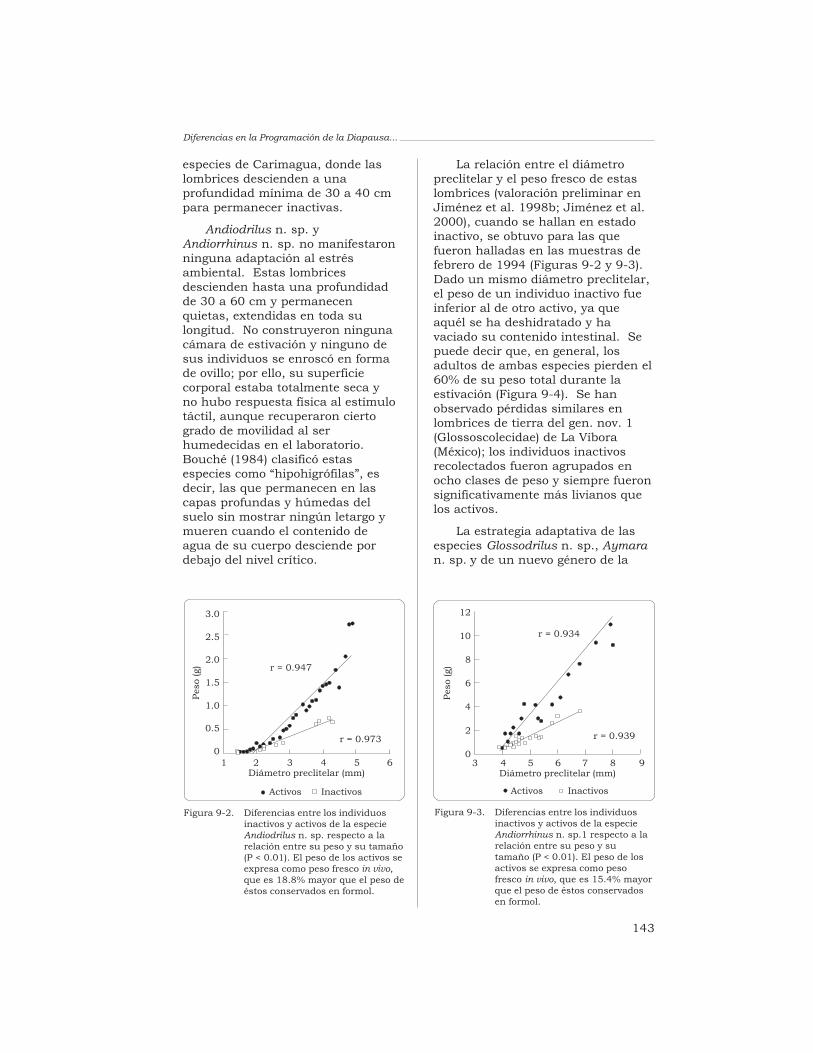
Diferencias en la Programación de la Diapausa...
143
especies de Carimagua, donde laslombrices descienden a unaprofundidad mínima de 30 a 40 cmpara permanecer inactivas.
Andiodrilus n. sp. yAndiorrhinus n. sp. no manifestaronninguna adaptación al estrésambiental. Estas lombricesdescienden hasta una profundidadde 30 a 60 cm y permanecenquietas, extendidas en toda sulongitud. No construyeron ningunacámara de estivación y ninguno desus individuos se enroscó en formade ovillo; por ello, su superficiecorporal estaba totalmente seca yno hubo respuesta física al estímulotáctil, aunque recuperaron ciertogrado de movilidad al serhumedecidas en el laboratorio.Bouché (1984) clasificó estasespecies como “hipohigrófilas”, esdecir, las que permanecen en lascapas profundas y húmedas delsuelo sin mostrar ningún letargo ymueren cuando el contenido deagua de su cuerpo desciende pordebajo del nivel crítico.
La relación entre el diámetropreclitelar y el peso fresco de estaslombrices (valoración preliminar enJiménez et al. 1998b; Jiménez et al.2000), cuando se hallan en estadoinactivo, se obtuvo para las quefueron halladas en las muestras defebrero de 1994 (Figuras 9-2 y 9-3).Dado un mismo diámetro preclitelar,el peso de un individuo inactivo fueinferior al de otro activo, ya queaquél se ha deshidratado y havaciado su contenido intestinal. Sepuede decir que, en general, losadultos de ambas especies pierden el60% de su peso total durante laestivación (Figura 9-4). Se hanobservado pérdidas similares enlombrices de tierra del gen. nov. 1(Glossoscolecidae) de La Víbora(México); los individuos inactivosrecolectados fueron agrupados enocho clases de peso y siempre fueronsignificativamente más livianos quelos activos.
La estrategia adaptativa de lasespecies Glossodrilus n. sp., Aymaran. sp. y de un nuevo género de la
Activos Inactivos
Figura 9-2. Diferencias entre los individuosinactivos y activos de la especieAndiodrilus n. sp. respecto a larelación entre su peso y su tamaño(P < 0.01). El peso de los activos seexpresa como peso fresco in vivo,que es 18.8% mayor que el peso deéstos conservados en formol.
Peso
(g)
1 2 3 4 5 6Diámetro preclitelar (mm)
3.0
2.5
2.0
1.5
1.0
0.5
0
r = 0.947
r = 0.973
Figura 9-3. Diferencias entre los individuosinactivos y activos de la especieAndiorrhinus n. sp.1 respecto a larelación entre su peso y sutamaño (P < 0.01). El peso de losactivos se expresa como pesofresco in vivo, que es 15.4% mayorque el peso de éstos conservadosen formol.
Peso
(g)
3 4 5 6 7 8 9Diámetro preclitelar (mm)
12
10
8
6
4
2
0
Activos Inactivos
r = 0.934
r = 0.939
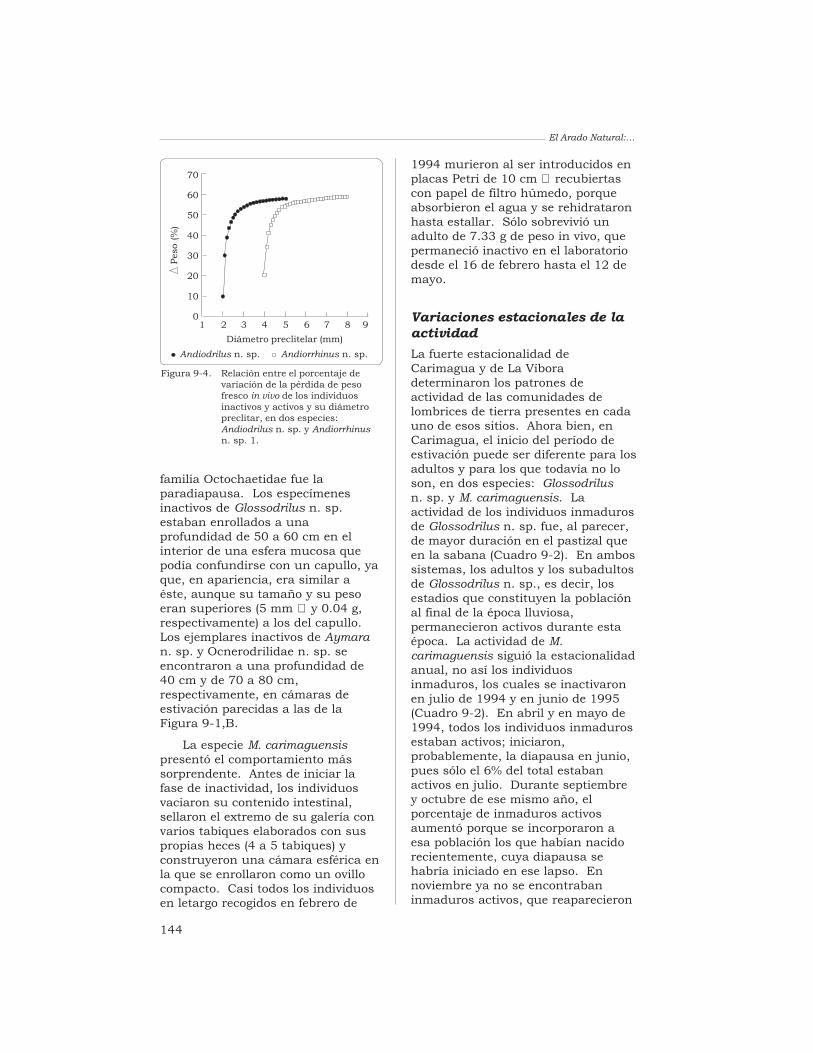
144
El Arado Natural:…
familia Octochaetidae fue laparadiapausa. Los especímenesinactivos de Glossodrilus n. sp.estaban enrollados a unaprofundidad de 50 a 60 cm en elinterior de una esfera mucosa quepodía confundirse con un capullo, yaque, en apariencia, era similar aéste, aunque su tamaño y su pesoeran superiores (5 mm ∅ y 0.04 g,respectivamente) a los del capullo.Los ejemplares inactivos de Aymaran. sp. y Ocnerodrilidae n. sp. seencontraron a una profundidad de40 cm y de 70 a 80 cm,respectivamente, en cámaras deestivación parecidas a las de laFigura 9-1,B.
La especie M. carimaguensispresentó el comportamiento mássorprendente. Antes de iniciar lafase de inactividad, los individuosvaciaron su contenido intestinal,sellaron el extremo de su galería convarios tabiques elaborados con suspropias heces (4 a 5 tabiques) yconstruyeron una cámara esférica enla que se enrollaron como un ovillocompacto. Casi todos los individuosen letargo recogidos en febrero de
1994 murieron al ser introducidos enplacas Petri de 10 cm ∅ recubiertascon papel de filtro húmedo, porqueabsorbieron el agua y se rehidrataronhasta estallar. Sólo sobrevivió unadulto de 7.33 g de peso in vivo, quepermaneció inactivo en el laboratoriodesde el 16 de febrero hasta el 12 demayo.
Variaciones estacionales de laactividadLa fuerte estacionalidad deCarimagua y de La Víboradeterminaron los patrones deactividad de las comunidades delombrices de tierra presentes en cadauno de esos sitios. Ahora bien, enCarimagua, el inicio del período deestivación puede ser diferente para losadultos y para los que todavía no loson, en dos especies: Glossodrilusn. sp. y M. carimaguensis. Laactividad de los individuos inmadurosde Glossodrilus n. sp. fue, al parecer,de mayor duración en el pastizal queen la sabana (Cuadro 9-2). En ambossistemas, los adultos y los subadultosde Glossodrilus n. sp., es decir, losestadios que constituyen la poblaciónal final de la época lluviosa,permanecieron activos durante estaépoca. La actividad de M.carimaguensis siguió la estacionalidadanual, no así los individuosinmaduros, los cuales se inactivaronen julio de 1994 y en junio de 1995(Cuadro 9-2). En abril y en mayo de1994, todos los individuos inmadurosestaban activos; iniciaron,probablemente, la diapausa en junio,pues sólo el 6% del total estabanactivos en julio. Durante septiembrey octubre de ese mismo año, elporcentaje de inmaduros activosaumentó porque se incorporaron aesa población los que habían nacidorecientemente, cuya diapausa sehabría iniciado en ese lapso. Ennoviembre ya no se encontrabaninmaduros activos, que reaparecieron
Andiodrilus n. sp. Andiorrhinus n. sp.
Figura 9-4. Relación entre el porcentaje devariación de la pérdida de pesofresco in vivo de los individuosinactivos y activos y su diámetropreclitar, en dos especies:Andiodrilus n. sp. y Andiorrhinusn. sp. 1.
Peso
(%
)
1 2 3 4 5 6 7 8 9Diámetro preclitelar (mm)
70
60
50
40
30
20
10
0

Diferencias en la Programación de la Diapausa...
145
al comienzo de la siguiente temporadade lluvias. En abril de 1995, el 62%de los individuos inmaduros estabanactivos de nuevo y al mes siguiente el100%. Estos iniciaron su diapausaen junio, de modo que la población deinmaduros activos descendió a un3.8% en agosto. Hubo diferenciassignificativas (P < 0.01, ANOVAKruskal-Wallis) en el peso medio delos individuos que se hallaban enestivación, según la capa de suelo queocupaban (Figura 9-5).
En La Víbora, desde diciembrehasta junio todos los individuos delgen. nov. 1 se hallaban estivando auna profundidad mayor que 15 cm,generalmente, la cual dependía de laprofundidad del horizonte A. Duranteeste período, la humedad del suelofue inferior al punto demarchitamiento (pF 4.2, 4% H2O,p/p). Los primeros individuos activosse encontraron a finales de junio —enconexión con la llegada de lasprimeras lluvias— y su actividad
Cuadro 9-2. Variación estacional de la actividad de los individuos inmaduros (Inm) y no inmaduros(N Inm) de Glossodrilus n. sp. en la sabana y en el pastizal y de los inmaduros (Inm),subadultos (Su) y adultos (Ad) de M. carimaguensis en el pastizal.
Mes de Glossodrilus n. sp. M. carimaguensis Sabana Pastizal Pastizal
Inm N Inm Inm N Inm Inm Su Ad
En 1994Abril 100 –a 100 – 100 100 100Mayo 100 – 100 – 100 100 100Julio 100 – 100 – 6 100 100Agosto 97 100 100 – 5 84 100Septiembre 88 100 100 100 11 – 100Octubre 19 100 100 100 12 18 90Noviembre 7 100 91 100 0 0 70Diciembre 8 100 53 100 0 0 8
En 1995Enero 0 0 0 – 0 – 0Febrero 0 – 0 – 0 – 0Marzo 34 – 0 – 0 – 0Abril 85 – 90 – 62 – 82Mayo 100 – 100 – 100 100 100Junio 100 – 100 – 96 100 100Julio 100 – 100 – 43 – 100Agosto 100 – 100 – 4 – 80Septiembre 92 – 100 100 20 – 100
a. El signo – indica que no se recolectó ningún ejemplar durante el mes de muestreo; en el caso deGlossodrilus n. sp., porque los adultos aparecen hacia el final de la época lluviosa.
Figura 9-5. Relación entre la distribuciónvertical en diferentes capas delsuelo (cm de espesor) y el pesomedio (± desviación estándar) delos individuos de M. carimaguensisen estivación (ANOVA de Kruskal-Wallis, P < 0.01). Los númerossobre cada punto indican el totalde individuos encontrados en cadacapa.
Peso medio (g)
Cap
a de
l su
elo
(cm
)
0-1010-2020-3030-4040-5050-6060-7070-8080-90
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5
Superficie delsuelo
7
66140
254202
10418
continuó hasta finales de diciembre—o hasta el tiempo en que finalizaba laépoca de lluvias.
muestreo

146
El Arado Natural:…
Estivación vs. regeneración enM. carimaguensisDe todas las lombrices de tierraencontradas en estivación (790), sóloel 6.6% presentó regeneración; larelación proporcional equivalentepara los individuos inmaduros fue de5.7% (Cuadro 9-3). No seencontraron diferencias significativasen el número de segmentos nuevosañadidos según el estado dedesarrollo de la lombriz (Kolmogorov-Smirnov, P = 0.85). Asimismo, no seencontró ninguna relación entre elnúmero de segmentos nuevos de laslombrices que estaban estivando y elpeso de éstas (Figura 9-6).
El porcentaje de adultos consegmentos nuevos totalmenteformados estaría señalando una tasade ‘fragmentación individual’ de laslombrices totalmente desarrolladas de
M. carimaguensis. El valor 8.8%corresponde a la pérdida de la parteposterior del cuerpo por causasnaturales, por ejemplo, ladepredación; cuando este valor seaplica a los individuos subadultos, latasa mencionada es de 9.9% (18.7%menos 8.8%, Cuadro 9-3). Este valores casi el doble del que se obtuvopara los individuos inmaduros(5.7%). Si se considera toda lapoblación (790 individuos), losadultos y los subadultos consegmentos nuevos sólo representanel 1.8% de ese total, del cualrepresentan los individuosinmaduros en iguales condiciones el4.8%.
La relación entre el número desegmentos nuevos añadidos y el pesode las lombrices en estivación semuestra en la Figura 9-7. La
Figura 9-6. Regresión entre el número desegmentos nuevos añadidos y elpeso de los individuos inmadurosen estivación de M. carimaguensis(n.s., P > 0.05).
Seg
men
tos
nu
evos
(no.
)
0 1 2 3 4 5
Peso (g)
120
100
80
60
40
20
0
r = 0.114
Figura 9-7. Regresión entre el número desegmentos nuevos añadidos yel peso de los individuosinmaduros (< 2 g) en estivación deM. carimaguensis (P < 0.05).
Seg
men
tos
nu
evos
(no.
)
0 0.5 1.0 1.5 2.0
Peso (g)
80
70
60
50
40
30
20
10
0
r = 0.518
Cuadro 9-3 Número de lombrices de la especie M. carimaguensis (N) halladas en diapausa y quepresentaban regeneración.
Lombrices en estivación Regeneración presente N2/N1 (%)
N1 % N2 %
Adultos 91 11.5 8 15.2 8.8Subadultos 32 4.0 6 11.5 18.7Inmaduros 667 84.4 38 73.1 5.7
Total 790 100 52 100

Diferencias en la Programación de la Diapausa...
147
regresión no fue significativa,excepto cuando se consideraron laslombrices cuyo peso era inferior a2 g (P < 0.05). Estos datos indicanque los individuos de M.carimaguensis agregarán nuevossegmentos según su peso individual,y que ese aumento será mayorcuando tengan un peso inferior a2 g.
DiscusiónLa estivación de las lombrices se haestudiado en la zona templada deEuropa y en Australia (Anderson1980; Evans y Guild 1948; Garnsey1994; Gerard 1967; Nordström1975); en regiones de Africa tropical,para las especies Hyperiodrilusafricanus Beddard 1893 y Eudriluseugeniae Kinberg 1867 (Madge1969); en Africa del Sur, paraAporrectodea rosea Savigny yAllolobophora trapezoides Eisen 1874(Reinecke y Ljungström 1969); y enIndia, para Octochaetona surensisMichaelsen (Senapati 1980). En estetrabajo se estudian por primera vez yen detalle las lombrices de tierraneotropicales.
La duración de la diapausa de M.carimaguensis en el laboratorio fuesimilar a la que presentó encondiciones naturales (Jiménez et al.1998b). Este patrón fue observadotambién por Saussey y Debout(1984) en Aporrectodea giardiRibaucourt (Lumbricidae). Gates(1961) reportó, por su parte, quealgunas especies asiáticas entran enuna fase de quiescencia antes deque termine la época lluviosa, ypermanecen inactivas aunque seanintroducidas en un ambiente en quela humedad les sea favorable; estoindica, por tanto, la presencia deuna diapausa obligatoria.
Reinecke (1983) reportó tambiénque el inicio de la diapausa era
diferente para los adultos y para losindividuos inmaduros de la especiesurafricana Microchaetus modestus(Glossoscolecidae), aunque noidentificó la señal que determinaesas diferencias. Gerard (1967)encontró un patrón opuesto alencontrado en este estudio enalgunos pastizales británicos, dondelas lombrices de tierra, cuyo peso erasuperior a 3 g, se encontrabanquiescentes y a mayor profundidadque los individuos recién nacidos, loscuales se mantenían activos y cercade la superficie del suelo.
Se pueden proponer, por tanto,dos hipótesis sobre la adición desegmentos en las lombrices de tierradurante su período de estivación:
- que las lombrices presentan elmismo número de segmentosdurante toda su vida;
- que algunos individuospresentan, en cambio, unnúmero variable de segmentosdurante su crecimiento y puedenincrementar ese número en losperíodos de estivación.
Si esta última hipótesis resultaracierta, indicaría una estrategiaadaptativa intraespecífica por lasiguiente razón: estas lombricespodrían ingerir una cantidad mayorde suelo que permitiría,especialmente si los recursosorgánicos son limitados —contenidosde C y N bajos, por ejemplo— comoocurre en las sabanas de suelosácidos, que se incrementara laasimilación de nutrientes en sutracto digestivo.
Los adultos de Eisenia fetidaSavigny 1826 presentan un númerode segmentos similar al de losindividuos recién nacidos (Sun yPratt 1931). Moment (1953) sostieneque la adquisición de nuevossegmentos finaliza en el momento dela eclosión y que, si ocurre una

148
El Arado Natural:…
amputación caudal, la lombrizproducirá para repararla el mismonúmero de segmentos que teníaantes. No obstante, el número desegmentos de los adultos puede sersuperior al de los individuos reciénnacidos, como ocurre en las especiesAllolobophora terrestris f. longaSavigny y A. nocturna (Evans y Guild1947) y en A. terrestris f. typica(Saussey 1971b).
Evans (1946) informó quealgunas especies adquierensegmentos adicionales durante elcrecimiento (otras no los producen).Abeloos y Avel (1928) y Saussey(1971) han establecido relacionesentre el período de estivación de laslombrices de tierra y su regeneraciónen la especie Allolobophora terrestrisf. typica, y Piearce (1983) en laespecie A. longa. En este últimoestudio no se encontró ningunarelación entre la regeneración y laestivación en los individuos de unade las especies investigadas con másdetenimiento.
Hay que realizar más estudiospara determinar si la regeneracióndurante la estivación es unaestrategia que les permite a laslombrices aumentar el número desegmentos o es, simplemente, unmecanismo de remplazo de la parteamputada. Es posible que ambosprocesos ocurran y esto explicaría laausencia de relación en los adultosde M. carimaguensis, aunque lasegunda hipótesis no ha sidoprobada todavía. Por otra parte, losindividuos inmaduros, cuyo pesoosciló entre 0.5 y 1 g, tenían unmayor número de segmentos nuevosque los que pesaron entre 1 y 2 g.Estos resultados podrían explicarsepor la extrema fragilidad de losindividuos inmaduros (Edwards yBohlen 1996), cuyas partes caudalesse rompen fácilmente —o son másatacadas por algunos depredadoresinvertebrados— cuando descienden a
estivar, lo que no ocurre entre losadultos y los subadultos.
La incorporación de segmentosnuevos implica, o bien uncrecimiento continuo (sin ningunaamputación) o bien una regeneraciónhipermérica tras producirse unaamputación caudal, como encontróSaussey (1971b) en A. terrestris.Omodeo (1962) afirma que laregeneración no se debe a unareparación caudal sino que es unafunción normal del crecimiento. Haytres veces más segmentos nuevos enlos adultos de A. caliginosa Savignyque en los individuos inmaduros dela especie. Sería muy extraño que,en este caso, les fuera amputada laparte caudal a las lombrices antes dellegar al estado adulto. Vedovini(1968, 1969) sostiene también estahipótesis en otras cuatro especies:Eophila dollfusi Tétry, E. dugesiRosa, A. caliginosa f. typica (Savigny)y E. sanaryensis Tétry. Saussey(1971b) demostró que hubo unaumento caudal en 4 individuos deun total de 27 a los que se indujoartificialmente la diapausa.
La formación de segmentosnuevos en la región caudal de A.terrestris ocurre siempre durante ladiapausa, en condiciones naturales(Abeloos y Avel 1928). Saussey(1971) demostró que los adultos deA. terrestris f. typica tienen mássegmentos nuevos que las lombricesrecién nacidas, aunque el porcentajede individuos que experimentaron laregeneración fue muy similar tantoen los inmaduros (57.1%) como enlos adultos (55%).
La diapausa puede considerarseuna inversión realizada en un estilode vida alternativo que ofrece suspropios beneficios, costos ycompensaciones, y no es tanto unahuida de las condicionesestacionales desfavorables(Bradshaw et al. 1998). Cuando el

Diferencias en la Programación de la Diapausa...
149
alimento disponible escasea, seactiva la diapausa en una granvariedad de organismos (Beck 1980;Gilbert y Schreiber 1998; Harper1977); un efecto igual produce elfotoperíodo en algunos insectos(Tauber et al. 1986) y en algunosácaros (Lees 1953). Tanto latemperatura como el fotoperíodo danseñales que inducen la diapausa enlas lombrices de tierra (Reinecke1983; Saussey y Debout 1984). Enla lombriz paleártica anécicaAporrectodea longa (Ude), los factoresque determinan el inicio de ladiapausa son la temperatura delsuelo, el estrés hídrico y elfotoperíodo, o sea, cuando laduración del día es superior a15.3 horas (Heidet y Bouché 1991).Morgan y Winters (1991) observaronun aumento en las reservasenergéticas de lípidos antes de iniciarla diapausa en la especie A. longa.Quizás los adultos segregan unasustancia inductora de la diapausaque es la responsable de la prontaentrada de los inmaduros en estafase. Una gran cantidad de estasustancia se produciríaespecialmente cuando los individuosinmaduros se inactivan en la mitadde la época de lluvias. Estosupondría, obviamente, un enormeesfuerzo energético para aquellasespecies que viven en suelos dondelos recursos orgánicos son bajos.
Después de la producción de loscapullos, los adultos de M.carimaguensis descienden a lascapas profundas del suelo tantocomo lo permite su peso corporal(Jiménez y Decaëns 2000). Estaespecie inicia la diapausa después deque sus adultos pierden el 20% delpeso fresco, al final de la épocalluviosa. En aquellos sitios donde laprecipitación es escasa o dondeexiste una estacionalidad muydefinida, como en las sabanasneotropicales, los mecanismos de
esta índole son, sin duda, esencialespara mantener la densidad de laspoblaciones de lombrices a lo largodel tiempo. Por tanto, la diapausadesempeña, al parecer, un papelmuy importante en el control delciclo de vida de M. carimaguensis.En las lombrices de tierra, ladiapausa estacional constituye unafase de regresión sexual, de ahorroenergético (Gallisian 1971) y decrecimiento postcaudal (Saussey ySong 1981). Forma parte no sólo dela regulación del ciclo de lasactividades trófica y sexual (Bouché1984), sino también de la densidadpoblacional.
Es poco probable que lascondiciones abióticas sean las quepropicien el inicio de la diapausa; encambio, la evasión de la competenciaintraespecífica e interespecífica, asícomo la depredación, pueden ser losfactores responsables de esecomienzo. Se ha observado tambiénque estos procesos son importantesen otros taxones, por ejemplo, enalgunas especies de copépodos(Santer y Lampert 1995). Quizás laexplicación posible delcomportamiento de M.carimaguiensis respecto a ladiapausa se encuentre en lanecesidad de reducir la competencia,en los hábitos reproductivos, en losfactores ambientales y aun en laescasez de alimento (disponibilidadde hojarasca); sin embargo, todosestos fenómenos deben serestudiados con más profundidad eninvestigaciones futuras.
La humedad del suelo es el factorresponsable de los patronesestacionales observados en ladistribución vertical de lascomunidades de lombrices de tierra,aunque las diferencias observadasentre los adultos y los individuosinmaduros de M. carimaguensispueden tener también un origenbiótico. La diapausa anticipada de

150
El Arado Natural:…
los inmaduros responde a unainducción fisiológica (Jiménez et al.1998b), pero no hay aún unaevidencia experimental sobre lanaturaleza de los factores quedesencadenan ese fenómeno. Ahorabien, una explicación plausible seríala regulación de la densidad depoblación de las lombrices, ya que seajusta especialmente a esta especieque necesita 3 años, al menos, paraalcanzar la madurez sexual(Jiménez, datos sin publicar). Laestrategia adaptativa de las especiesmás grandes, que explotan las capassuperficiales del suelo para evitaruna concentración elevada deindividuos de la misma especie, yreducir así la competencia intra-específica, podría ser la que regulaeste tipo de comportamiento en M.carimaguensis. El fotoperíodo, comofactor de inducción de la diapausaen las lombrices (Begon et al. 1996;Saussey y Debout 1984), puedeafectar a los adultos y a losindividuos inmaduros de mododiferente. Cabe la observación deque esta diversidad de hipótesis yrespuestas requiere más estudios,principalmente de carácterfisiológico, que permitan definir lasdiferencias que existen entre losesquemas o patrones de estivaciónde las lombrices de tierra.
AgradecimientosLos autores agradecen a CarlosFragoso y a Isabelle Barois por suscríticas a la primera versión delmanuscrito, y a varios estudiantes yoperarios de campo por la ayudaprestada para este estudio.Agradecen también a A.G. Morenopor los dibujos de las diferentescámaras de estivación. Lainfraestructura y los fondos pararealizar el estudio provinieron
principalmente del CIAT y del IRD(antiguo ORSTOM). Agradecenademás el apoyo recibido, durantevarias fases de este trabajo, delPrograma de Biología y Fertilidad delos Suelos Tropicales (TSBF), de laUniversidad Complutense (España),del Instituto de Ecología (México) yde la Agencia Española deCooperación Internacional (AECI).Agradecen, finalmente, por loscomentarios y las sugerencias útilesde dos revisores anónimos.
ReferenciasAbeloos M; Avel M. 1928. Un cas de
périodicité du pouvoir régénérateur:la régénération de la queu chez lesLombriciens Allolobophora terrestriset A. caliginosa. C R Soc Biol99:737-738.
Anderson C. 1980. The influence ofclimatic conditions on activity andvertical distribution of earthwormsin a Danish arable soil. K Vet-Landbohojsk Arssk 57-68.
Anderson JM; Ingram JSI. 1993. Tropicalsoil biology and fertility: ahandbook of methods. 2a. ed.C.A.B. International, Londres,Reino Unido. 221 p.
Beck SD. 1980. Insect photoperiodism.2a. ed. Academic Press, NuevaYork, Estados Unidos.
Begon M; Harper JL; Townsend CR.1996. Ecology: individuals,populations and communities. 3a.ed. Blackwell Science, Oxford,Reino Unido. 1068 p.
Bouché MB. 1972. Lombriciens deFrance: ecologie et systematique.INRA, París, Francia. 671 p.
Bouché MB. 1984. Les modalitésd’adaptation des lombriciénnes à lasécheresse. Bull Soc Bot Fr131:319-327.

Diferencias en la Programación de la Diapausa...
151
Bradshaw WE; Armbruter PA; HolzapfelCM. 1998. Fitness consequences ofhibernal diapause in the pitcher-plant mosquito Wyeomyia smithii.Ecology 79:1458-1462.
Darwin C. 1881. The formation ofvegetable mould through the actionof worms, with observations of theirhabits. Murray, Londres.
Edwards CA; Bohlen PJ. 1996. Biologyand ecology of earthworms. 3a. ed.Chapman and Hall, Londres, ReinoUnido. 426 p.
Evans AC. 1946. Distribution of numbersof segments in earthworms and itssignificance. Nature 158:98.
Evans AC; Guild WJ. 1947. Studies onthe relationships betwenearthworms and soil fertility; I:Biological studies in the field. AnnAppl Biol 34:307-330.
Evans AC; Guild WJ. 1948. Studies onthe relationships betweenearthworms and soil fertility; IV: Onthe life cycles of some BritishLumbricidae. Ann Appl Biol35:473-493.
Gallisian A. 1971. Diapause etrégénération postérieure chez lelombricide Eophila dollfussi (Tétry).Tesis. Université de Provence,Francia. 243 p.
Garnsey RB. 1994. Seasonal activity andaestivation of lumbricid earthwormsin the midlands of Tasmania. Aust JSoil Res 32:1355-1367.
Gates GE. 1961. Ecology of someearthworms with special referenceto seasonal activity. Am Midl Nat66:61-86.
Gerard BM. 1967. Factors affectingearthworms in pastures. J AnimEcol 36:235-252.
Gilbert JJ; Schreiber DK. 1998. Asexualdiapause induced by food limitationin the rotifer Synchaeta pectinata.Ecology 79:1371-1381.
Harper JL. 1977. Population biology ofplants. Academic Press, NuevaYork, Estados Unidos.
Heidet J-C; Bouché MB. 1991. Regulationde l’activité lombricienne: influencede la temperature, de laphotoperiode et de l’humidité insitu sur l’indice de mobilité deNicodrilus longus longus (Ude)(Lumbricidae, Oligochaeta). En:Veeresh GK; Rajogopal D;Viraktamath CA, eds. Advances inmanagement and conservation ofsoil fauna. Oxford and IBH, NuevaDelhi, India. p 643-655.
Hensen V. 1877. Die Tätigkeit desRegenwurms (Lumbricus terrestrisL.) für die Fruchtbarkeit desErdbodens. Z Wiss Zool28:354-364.
Jiménez JJ; Decaëns T. 2000. Verticaldistribution of earthworms ingrassland soils of the ColombianLlanos. Biol Fertil Soils 32:463-473.
Jiménez JJ; Mamolar E; Lavelle P. 2000.Biometric relationships inearthworms (Oligochaeta). Eur JSoil Biol 36:45-50.
Jiménez JJ; Moreno AG; Lavelle P. 1999.Reproductive strategies of threenative earthworm species from thesavannas of Carimagua (Colombia).Pedobiologia 43:851-858.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis n. sp.(Glossoscolecidae, Oligochaeta), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innative savanna and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.

152
El Arado Natural:…
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfonctions dans l’ècosystème. Tesis(Doctorado). Paris VI Publ Lab ZoolENS 12. 301 p.
Lavelle P. 1981. Stratégies dereproduction chez les vers de terre.Acta Oecologica 2:117-133.
Lee KE. 1951. Role of earthworms in NewZealand soil. Tuatara 4:22-27.
Lee KE. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, NuevaYork. 411 p.
Lees AD. 1953. Environmental factorscontrolling the evocation andtermination of diapause in thefruit tree red spider miteMetatetranychus ulmi Koch (Acarina,Tetranychidae). Ann Appl Biol40:449-486.
Madge DS. 1969. Field and laboratorystudies on the activities of twospecies of tropical earthworms.Pedobiologia 9:188-214.
Moment GB. 1953. On the way a commonearthworm, Eisenia foetida, grows inlength. J Morphol 93:489-507.
Morgan AJ; Winters C. 1991. Diapause inthe earthworm Aporrectodea longa:morphological and quantitative X-ray microanalysis of cryosectionedchloragogenous tissue. ScanningMicrosc 5:219-228.
Nordström S. 1975. Seasonal activity oflumbricids in Southern Sweden.Oikos 26:307-315.
Omodeo P. 1962. Oligochetes des Alpes;I. Mem Mus Civ Stor Nat Verona10:71-96.
Piearce TG. 1983. Functional morphologyof lumbricid earthworms, withspecial reference to locomotion. JNat Hist 17:95-111.
Reinecke AJ. 1983. The ecology ofearthworms in southern Africa. En:Satchell JE, ed. Earthworm ecology:from Darwin to vermiculture.Chapman and Hall, Londres.p 195-207.
Reinecke AJ; Ljunstrom PO. 1969. Anecological study of the earthwormsfrom the banks of Mooi River inPotchefstroom, South Africa.Pedobiologia 9:106-111.
Santer B; Lampert W. 1995. Summerdiapause in cyclopoid copepods:adaptive response to a foodbottleneck? J Anim Ecol64:600-613.
Satchell JE. 1967. Lumbricidae. En:Burgess A; Raw F, eds. Soil biology.Academic Press, Londres.p 259-322.
Saussey M. 1966. Contribution à l’étudedes phénomène de diapause et derégénération caudale chez A. ictericaSav. Mém Soc Lin Norm 1:1-158.
Saussey M. 1971. Une techniqued’élevage des lombriciens dans lesconditions naturelles et sesapplications: comportementd’Allolobophora terrestris (Savigny) f.typica après amputation de lamoitié postérieure du corps. Animhors série:557-569.
Saussey M. 1971b. Étude preliminaire duproblème de la croissance post-embryonnaire chez Allolobophoraterrestris (Savigny) f. typica(Oligochète, Lombricien). Bull SocLinn Normandie 102:93-104.
Saussey M; Debout G. 1984. Nouvellesdonnées sur le determinisme de ladiapause de Nicodrilus giardi(Ribacourt) (Oligochète,Lombricien). C R Acad Sci ParisSerie III 299(2):35-38.
Saussey M; Song MJ. 1981. Le problèmedes néoformations caudalessuccessives chez Nicodrilus giardi(Ribaucourt) (Oligochète,Lombricien). C R Acad Sci ParisSerie III 292:49-53.

Diferencias en la Programación de la Diapausa...
153
Senapati BK. 1980. Aspects ofecophysiological studies on tropicalearthworms: Distribution,population dynamics, production,energetics and their role in thedecomposition process. Tesis(Ph.D.). School of Life Science,Sambalpur University, India. 154 p.
Sun KH; Pratt KC. 1931 Do earthwormsgrow by adding segments? Am Nat65:31-48.
Tauber MJ; Tauber CA; Masaki S. 1986.Seasonal adaptation of insects.Oxford University Press, NuevaYork, Estados Unidos.
USDA Soil Survey Staff .1975. Soiltaxonomy. Washington, D.C. 754 p.
Vedovini A. 1968. Observations sur lesEophila (Lumbricidae) de Provence.Bull Soc Zool France 93:647-652.
Vedovini A. 1969. Contribution à l’étudedes variations de l’espèceAllolobophora caliginosa(Lombricidae). Bull Soc Zool France94:657-662.

154
El Arado Natural:…
ResumenSe puede emplear una clasificaciónecológica basada en una serie decaracteres morfológicos paradescribir las funciones quedesempeñan las lombrices de tierraen el suelo, las cuales, en realidad,rara vez han sido verificadas. Seestudiaron las mollejas deMartiodrilus carimaguensis(Glossoscolecidae) para determinar sila morfología anécica correspondía ala dieta de la lombriz (ingestión dehojarasca y material órgano-mineraldel suelo). Mediante la observacióndirecta del contenido de la molleja de13 adultos con una lupa binocular yun microscopio de contraste de fase,se determinó que el porcentaje (envolumen) de fragmentos vegetales,raíces, materia orgánica (MO) amorfay partículas minerales era de 55, 6.3,
5.0 y 35% (v/v), respectivamente. El80% de los fragmentos vegetalescontenidos en la molleja eranmicroscópicos y, en su mayoría,estaban incorporados en la materiaórgano-mineral de las heces de laslombrices, que se conservaban bien ensus mollejas. Las característicaspredominantes de la dieta deM. carimaguensis fueron la coprofagiay la necrorizofagia. Ahora bien, lasheces producidas indicaban una granvariabilidad en su dieta, hecho quepodría relacionarse con la adopcióntransitoria de una dieta típicamenteanécica durante ciertos períodos detiempo.
Palabras clave: contenido intestinal,coprofagia, dieta, gruposecológicos, lombrices anécicas,sabana, suelos tropicales
IntroducciónLas lombrices constituyen un recursopotencial de gran interés en lasostenibilidad de la agricultura, yaque participan activamente en laregulación de las propiedades físicasdel suelo, en la dinámica de la MO delentorno y en el crecimiento de lasplantas (Lavelle et al. 1999). Lastécnicas de inoculación con lombricesde tierra, en condiciones naturales,proporcionan beneficios, aunque sucosto es muy alto; son, por tanto, una
CAPÍTULO 10
La Dieta de una Lombriz Anécica delas Sabanas Colombianas: UnaCuestión Relacionada con lasCategorías Ecológicas1
L. Mariani* , N. Bernier**, J. J. Jiménez*** yT. Decaëns†
1. Publicado en Comptes Rendus de l’Académiedes Sciences, Séries III 324(8), p 733-742,por Mariani et al. Copyright © 2001 deElsevier. Impreso con permiso de la casaeditorial.
* Laboratoire d’Ecologie et Biologie des SolsTropicaux, IRD, 32 Av. Henri Varagnat,F-93143 Bondy Cedex, Francia.
** Laboratoire d’Ecologie Générale, MuséumNational d’Histoire Naturelle, 4 Av. du PetitChâteau, F-91800, Brunoy, Francia.
*** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
† Laboratoire d’Ecologie, UFR Sciences etTechniques, Université de Rouen, F-76821,Mont Saint Aignan Cedex, Francia.

La Dieta de una Lombriz Anécica...
155
inversión que sólo debería hacerse encultivos de alto valor comercial(Senapati et al. 1999). Estastécnicas no son aplicables a loscultivos más extendidos, como loscereales; por ello, el manejo indirectode las poblaciones demacroinvertebrados del suelo pareceser la solución, ya que emplea unaserie de prácticas agronómicas en uncontexto espacial. Ahora bien, unaetapa esencial de ese manejo es elconocimiento sólido de los hábitosalimenticios de las especies delombrices elegidas, para comprendery, en ocasiones, incrementar lacapacidad de carga de lombrices delas áreas cultivadas.
El régimen alimenticio de laslombrices es un componente esencialde su clasificación funcional.Bouché (1977) propuso una tipologíabasada en los caracteresmorfológicos de las lombrices y en surelación con los hábitos alimenticiosde éstas y, hasta cierto punto, con lafunción que se supone que ellasdesempeñan en el suelo. Laslombrices epígeas, homogéneamentepigmentadas y fácilmentereconocibles, viven entre la hojarascao justo en la capa de suelo situadabajo ella, y se alimentan de restosvegetales. Las lombrices endógeasno tienen pigmentación, presentanun sistema muscular apenasdesarrollado y se alimentan,principalmente, del suelo y dealgunas raíces. Las lombricesanécicas, cuya pigmentación antero-dorsal es muy notoria, presentan unsistema muscular muy desarrolladoque está en consonancia con suactividad de introducir hojarasca enel suelo. La información sobre loshábitos alimenticios de las lombricesanécicas está restringida a seisespecies solamente, ninguna de ellasperteneciente a una región tropical(Bernier 1998; Bolton y Phillipson1976; Bouché y Kretzschmar 1974;
Ferrière 1980; James y Cunningham1989; James y Seastedt 1986; Judas1992; Kakajondo 1984; Piearce1972). Considerando la visión delmanejo ecológico de losagroecosistemas, es importanteestablecer el alcance que tienen loscriterios morfológicos definidos porBouché (1977) como indicadoresacertados del régimen alimenticio delas lombrices de tierra. Asimismo, esnecesario conocer tanto el origencomo el destino de la MO ingeridapor una lombriz, una vez que aquéllaha pasado por su tracto digestivo.
La lombriz anécica Martiodriluscarimaguensis (Glossoscolecidae) esuna especie nativa de las sabanas delos Llanos Orientales de Colombia.Sus individuos son de tamañogrande: 9 mm de diámetro, 190 mmde longitud y 11 g de peso fresco enestado adulto (Jiménez et al. 1998a).Se escogió esta especie para elestudio porque adquiere, en lospastizales introducidos, una biomasamuy alta y porque es capaz decolonizar con regularidad los cultivosintensivos adyacentes (Decaëns et al.1994; Jiménez et al. 1998b). Lalombriz forma galerías sub-verticales,abiertas a la superficie del suelo,donde deposita heces de grantamaño (o turrículos) que puedenresistir las fuerzas de desagregacióndurante varios meses (Decaëns2000). Estos turrículos constituyenuna fuente de nutrientes asimilablescuando están frescos y una reservade MO una vez que se secan(Decaëns et al. 1999c). Variosestudios han mostrado el efectopositivo que estos turrículos ejercenen la biomasa radical, así como en ladensidad de algunos grupos demacroinvertebrados epígeos yanécicos (Decaëns et al. 1999b).Cuando se excluyó a M.carimaguensis de pequeñas parcelasexperimentales durante 18 meses,los resultados fueron la

156
El Arado Natural:…
compactación del suelo, unareducción del contenido de C delsuelo y de la biomasa vegetal, unincremento de la saturación de Al yun porcentaje más alto de malashierbas (Decaëns et al. 1999a).
Materiales y Métodos
Sitio del estudioEste estudio se realizó en la estaciónexperimental de Carimagua(CORPOICA-CIAT) situada en losLlanos Orientales de Colombia(4°37’ N, 71°19’ O) a 175 msnm. Lavegetación dominante corresponde ala de una sabana herbácea noinundable. El valor medio anual dela precipitación y de la temperaturaes de 2280 mm y 26 °C,respectivamente (datos del CIAT) yhay una época seca muy definida dediciembre a marzo. Los suelos sonOxisoles bien estructurados, de pocafertilidad, con una acidez [pH (H2O)4.5] alta, y con una saturación dealuminio alta (más del 90% delcomplejo de absorción del suelo estásaturado con Al3+); además, sucontenido de cationesintercambiables (Ca2+, Mg2+ y K+) esbajo (datos del CIAT).
Las parcelas en que sedesarrolló el muestreo pertenecen aun ensayo agropastorilmultidisciplinario cuyo objetivo esestudiar el efecto que ejerce laintensificación agrícola en elfuncionamiento del suelo en lassabanas de América del Sur. Endicho ensayo se prueban numerososy diferentes agroecosistemas, desdelos cultivos anuales que requierenaplicaciones altas de insumos hastalos pastizales de uso intensivo; lasabana nativa se toma como testigo.
Se escogieron, por tanto, dossitios para llevar a cabo esteestudio:
• Una sabana natural que no hasido pastoreada, que se quemacada año antes del comienzo dela época seca, y dondeTrachypogon vestitus Anderss,Paspalum pectinatum Nees,Axonopus aureus Beauv.,Schyzachyrium hirtiflorum Ness,Gymnopogon foliosus Nees eHyptis conferta Pohl ex Benthson las gramíneas másabundantes.
• Un pastizal introducido en que seasocian la gramínea alóctonaBrachiaria humidicola Rendle ylas leguminosas Arachis pintoiKrap. & Greg., Desmodiumovalifolium Guill. & Perr. yStylosanthes capitata Vog.
En el pastizal introducido, lasgramíneas desarrollaron un sistemaradical profundo, aunque susmacollas se dispersen sobre lasuperficie del suelo, en el cualaparecen raíces superficialesespontáneas (Fisher et al. 1994). Elsuelo del pastizal está enriquecido enC, es decir, tiene 3.7% frente a2.83% de la sabana en los primeros5 cm del perfil (Decaëns et al.1999c). Durante la época lluviosa,la carga animal es de 2 UA/ha en elpastizal, mientras que en la sabananativa es de 1 UA por 2 ó 3 ha(1 UA = 250 kg). La capacidad decarga del pastizal para mantenerlombrices es cinco veces mayor quela de la sabana [1.6 individuos/m2
frente a 0.4 individuos/m2 (Jiménezet al. 1998b) que equivalen a 24 y6 g/m2, respectivamente, debiomasa].
Muestreo de las lombrices y dela fracción liviana de MO en elsuelo y en la hojarascaDurante el mes de agosto de 1999,se recolectaron individuos adultos encada parcela mediante la aplicación

La Dieta de una Lombriz Anécica...
157
directa de una solución de formol(0.2%) dentro de la galería de lalombriz. Estas lombrices seguardaron luego en una solución quecontenía, en partes iguales, formol al4% y alcohol al 70% y, finalmente, seconservaron en alcohol al 70%. Laslombrices se midieron y se pesarondespués. Todas se recolectaron en lasprimeras horas de la mañana durante2 semanas.
La hojarasca se obtuvo del interiorde dos superficies circulares de0.377 m2 cada una, concéntricas,delimitadas por dos aros metálicossituados alrededor de la galería de lalombriz. La superficie interior era uncírculo de 50 cm de diámetro y laexterior otro alejado de 50 a 70 cm dela entrada de la galería. La cantidadde hojarasca recolectada hacia elinterior del primer círculo seconsideró hojarasca de la galería,mientras que la recolectada entre éstey el círculo exterior se tomó comotestigo. La hojarasca se lavó despuéscon agua para quitar las partículas desuelo adheridas a ella y se secó en unhorno a 75 °C durante 2 días.
Se recolectaron también muestrasde suelo con un cilindro de aluminiode 10 cm de diámetro y 15 cm dealtura, a 5 m de la galería de donde sehabía extraído previamente la lombriz.La MO del suelo —en este caso, lafracción ligera mayor que 0.125 mm—se aisló mediante la técnica de lavadoy tamizado en cuatro clases según eltamaño: > 2mm, 1 a 2 mm, 0.5 a1 mm y 0.125 a 0.5 mm. Cada claseo subfracción se secó en un horno a75 °C durante 2 días y se pesó.
Se hizo también una colección dereferencia con las especies vegetalesmás abundantes en las dos parcelasestudiadas; tanto la parte aérea comolas raíces de las plantas recolectadasfueron conservadas en alcohol al70%.
Contenido de la mollejaSe aisló el contenido intestinal de8 lombrices adultas del pastizalintroducido y de 5 de la sabananativa y se analizó ya sea bajo unalupa binocular (magnificación 40X)cuando se trataba de materiaórgano-mineral mayor que 500 mm oya bajo un microscopio de contrastede fase (magnificación 630X) parapercibir los fragmentos órgano-minerales más pequeños (Bernier1998). El material que, una vezingerido por la lombriz, está menosmodificado es el que se encuentra enla molleja (Ferriére 1980).
Las observaciones realizadasbajo la lupa permitieron separar elmaterial vegetal, los fragmentosminerales (contenido de Fe), losagregados órgano-minerales (masaórgano-mineral de textura fina conelementos no identificables), elcarbón y las deyeccionesholorgánicas (heces). La materiavegetal macroscópica se clasificósegún su origen (de raíces, de parteaérea y sin determinar). Fue posibleobservar las estructuras órgano-minerales porque habían sidoconservadas en etanol al 95%; éstasse clasificaron como agregados biendefinidos procedentes de pequeñasbolas fecales depositadas por laslombrices, como agregadosangulares, y como agregados sinclasificar (debido, probablemente, ala manipulación y a la conservación).Bajo el microscopio se identificaronla materia vegetal libre, el carbón, lasestructuras minerales, la MO amorfaaislada y la incluida en complejosorgano-minerales, y la MO vegetal,amorfa y mineral (Bernier 1980). Elporcentaje volumétrico de cadacategoría se calculó por el método deconteo de puntos en una rejillatransparente. Cada punto de lamuestra se seleccionó al azar porquela imagen se enfoca primero en la redy después en el portaobjetos. En

158
El Arado Natural:…
ambos casos se contabilizan cercade 100 puntos por cadaportaobjetos analizado. Ahora bien,este método puede sobreestimar laproporción de elementos alargados oaplastados, lo que se compensa,supuestamente, por la disposiciónrelativamente aleatoria de estoselementos sobre el portaobjetos. Seregistraron en 12 clases de tamañolos fragmentos que oscilaban entre0.8 y 0.08 mm en el microscopio(para todos los componentes,excepto los amorfos) y entre 0.025 y3 mm en la lupa. Estas dos escalasde observación se superponenparcialmente porque las clasescomprendidas entre 0.025 y0.05 mm y entre 0.05 y 1 mm soncomunes a ambas escalas. Laspruebas de comparación empleadasinducen a elegir las observacionesrealizadas bajo el microscopio en laclase de 0.025 a 0.05 mm, ya quebajo la lupa ocurre unasubestimación importante. En laclase superior, en cambio, laelección de las observacionesrealizadas bajo la lupa es arbitrariaporque las dos medidas no sonestadísticamente diferentes.
El contenido de la molleja quees, generalmente, menor que 1 cm3,se observó bajo la lupa, mientrasque en el microscopio se empleó unaalícuota órgano-mineral extraída alazar de numerosos agregados. Elporcentaje de fragmentos observadoen el microscopio fue corregido auna prorrata de los valores de losagregados observados en la lupa.
La colección de plantas dereferencia se empleó para identificarel origen de los fragmentos vegetalesque se observaron bajo la lupa, esdecir, si provenían de la parte aéreao de la raíz.
Análisis estadísticosEl procedimiento Box-Cox permitetransformar los datos para reducir laasimetría y la kurtosis de lasdistribuciones (Sokal y Rohlf 1995)mediante el programa estadístico R(Legendre y Vadour 1995). Se aplicóuna serie de pruebas estadísticas noparamétricas a algunas de lasvariables analizadas, por dosrazones: las muestras eran pocas yla presunción de la homogeneidad delas varianzas no se cumplió(Scherrer 1984). Se empleó laprueba de U Wilcoxon-Mann-Whitney para comparar las muestrasque provenían de las dos parcelas ylas clases (según tamaño) comunesque se midieron en el microscopio yen la lupa. Se emplearon laspruebas de Wilcoxon de muestras nopareadas para comparar algunasvariables (los porcentajes defragmentos de raíz y de parte aérea).Se hicieron correlaciones noparamétricas (de rango) con el fin deexplicar la variabilidad intergrupos eintragrupos, entre la cantidad dehojarasca, la fracción liviana de laMO, la masa corporal de losindividuos y las características de lasmollejas.
Estas pruebas no paramétricasse realizan con variables o pares devariables y no emplean ningúncriterio para manejar múltiplescomparaciones partiendo de unconjunto especial de datos. Ahorabien, el riesgo de la primera especie(α) para k pruebas simultáneas esde 1- (1- αk), lo que da un valor de0.4 en lugar del nivel 0.05 para lascorrelaciones entre las cincovariables (Scherrer 1984). Paraejecutar correctamente las pruebasestadísticas se pueden seguir doscaminos: (Legendre P., Université deMontreal, com. pers.), o bien probarla hipótesis general incluyendo todaslas pruebas simultáneas trasefectuar cada prueba sin corrección,

La Dieta de una Lombriz Anécica...
159
o bien corregir el nivel designificación de cada prueba. Ennuestro caso, se ha probado lahipótesis global de la diferencia entrela media multidimensional de losdos grupos (parcelas o clases segúntamaño) con la prueba (test) deMantel: ésta consiste en lacomparación de una matriz quecontiene la similitud entre todas lasobservaciones y una matriz modeloque representa la división de lasobservaciones en dos grupos(Legendre y Fortín 1989; Legendre yLegendre 1998). Esta pruebaequivale a un análisis de la varianzamultivariante no-paramétrica. Lassimilitudes entre las observacionesse calculan a partir del coeficientecuantitativo y simétrico de Gower(Legendre y Legendre 1998, ecuación7.20 p. 259). Para las correlacionesparamétricas no existe ningunaprueba disponible; en este caso seha empleado la corrección múltiplede Bonferroni para pruebasmúltiples, modificada por Hochbergy que se desarrolla en dos etapas(Hochberg 1998; Legendre yLegendre 1998). En general, se tratade probar la hipótesis nula en que,del conjunto de pruebas simultáneasrealizadas al nivel α, hay al menosuna prueba significativa al nivel deα/(número de pruebas simultáneas).Después, y para probar la hipótesisnula en cada prueba, se ordenanen orden creciente las kprobabilidades, se corrigenempleando el factor k-i + 1 (dondei es el nivel de probabilidad), y secomparan con α.
Para la representación de lasvariables se escogieron la mediana,los quartiles y la desviación porquerepresentan mucho mejor ladistribución cuando las variables nose ajustan a una distribuciónnormal. Por consiguiente, en el casode una fuerte asimetría en ladistribución de los datos, la suma de
las medianas podría ser ligeramentesuperior a 100.
ResultadosEl contenido de las mollejas,conservado en alcohol y observadobajo la lupa, se mostró como unamasa órgano-mineral de textura fina(que se analizará luego bajo elmicroscopio), en la que aparecieronfragmentos macroscópicos denaturaleza mineral y vegetal. Estamasa órgano-mineral puede estar obien claramente moldeada o bienformada por un conjunto de gránulosque están dentro de la molleja.Estos se distinguen por su forma yson, de hecho, agregados biendefinidos. Estos agregados fueronmoldeados por otros animales y elalto contenido de suelo superficialindica que se trata de deyecciones delombrices más pequeñas. Dentro dela masa descrita se encontrarontambién pequeños fragmentos decarbón. La estructura de la masaera más o menos desagregada, dadala cantidad de fragmentos vegetalesde la muestra y el proceso demanipulación a que fue sometida.
Pese a la gran variabilidadindividual observada (Figura 10-1),se pudo constatar que los agregadosórgano-minerales de los 13individuos analizados (mediana =77%) fueron predominantes; losfragmentos vegetales y minerales> 50 µm eran escasos, y presentabanmedianas de 14% v/v y 11% v/v,respectivamente. Estos agregadosórgano-minerales contenían grandescantidades de fragmentos vegetales< 50 µm (44% v/v) y su contenidototal de MO fue mayor que el de losconstituyentes macroscópicos ymicroscópicos juntos (Figura 10-1,B).La MO liviana representó el 63% delvolumen total del contenido de lasmollejas, mientras que se obtuvo un
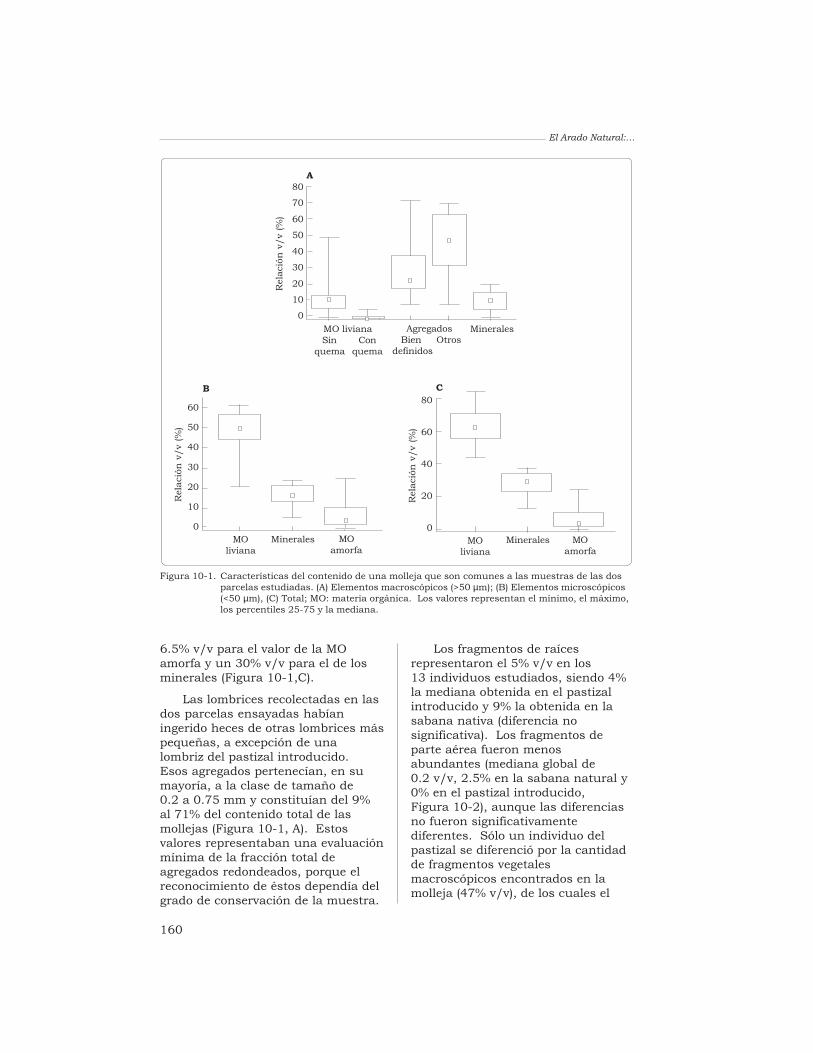
160
El Arado Natural:…
Rel
ació
n v
/v (%
)
A
MO livianaSin Con
quema quema
Rel
ació
n v
/v (%
)
80
70
60
50
40
30
20
10
0Agregados
Bien Otrosdefinidos
Minerales
MOliviana
Rel
ació
n v
/v (%
)
60
50
40
30
20
10
0MO
amorfaMinerales
B
MOliviana
80
60
40
20
0MO
amorfaMinerales
C
Figura 10-1. Características del contenido de una molleja que son comunes a las muestras de las dosparcelas estudiadas. (A) Elementos macroscópicos (>50 µm); (B) Elementos microscópicos(<50 µm), (C) Total; MO: materia orgánica. Los valores representan el mínimo, el máximo,los percentiles 25-75 y la mediana.
6.5% v/v para el valor de la MOamorfa y un 30% v/v para el de losminerales (Figura 10-1,C).
Las lombrices recolectadas en lasdos parcelas ensayadas habíaningerido heces de otras lombrices máspequeñas, a excepción de unalombriz del pastizal introducido.Esos agregados pertenecían, en sumayoría, a la clase de tamaño de0.2 a 0.75 mm y constituían del 9%al 71% del contenido total de lasmollejas (Figura 10-1, A). Estosvalores representaban una evaluaciónmínima de la fracción total deagregados redondeados, porque elreconocimiento de éstos dependía delgrado de conservación de la muestra.
Los fragmentos de raícesrepresentaron el 5% v/v en los13 individuos estudiados, siendo 4%la mediana obtenida en el pastizalintroducido y 9% la obtenida en lasabana nativa (diferencia nosignificativa). Los fragmentos departe aérea fueron menosabundantes (mediana global de0.2 v/v, 2.5% en la sabana natural y0% en el pastizal introducido,Figura 10-2), aunque las diferenciasno fueron significativamentediferentes. Sólo un individuo delpastizal se diferenció por la cantidadde fragmentos vegetalesmacroscópicos encontrados en lamolleja (47% v/v), de los cuales el
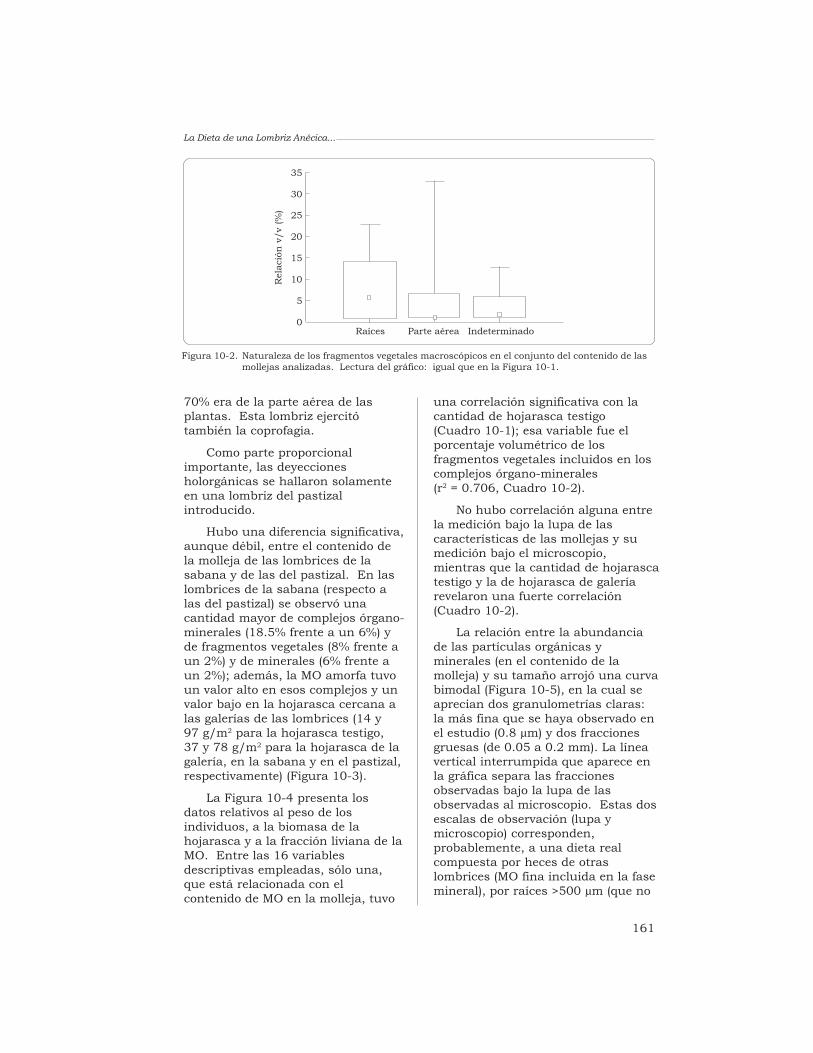
La Dieta de una Lombriz Anécica...
161
Rel
ació
n v
/v (%
)
35
30
25
20
15
10
5
0IndeterminadoParte aéreaRaíces
Figura 10-2. Naturaleza de los fragmentos vegetales macroscópicos en el conjunto del contenido de lasmollejas analizadas. Lectura del gráfico: igual que en la Figura 10-1.
70% era de la parte aérea de lasplantas. Esta lombriz ejercitótambién la coprofagia.
Como parte proporcionalimportante, las deyeccionesholorgánicas se hallaron solamenteen una lombriz del pastizalintroducido.
Hubo una diferencia significativa,aunque débil, entre el contenido dela molleja de las lombrices de lasabana y de las del pastizal. En laslombrices de la sabana (respecto alas del pastizal) se observó unacantidad mayor de complejos órgano-minerales (18.5% frente a un 6%) yde fragmentos vegetales (8% frente aun 2%) y de minerales (6% frente aun 2%); además, la MO amorfa tuvoun valor alto en esos complejos y unvalor bajo en la hojarasca cercana alas galerías de las lombrices (14 y97 g/m2 para la hojarasca testigo,37 y 78 g/m2 para la hojarasca de lagalería, en la sabana y en el pastizal,respectivamente) (Figura 10-3).
La Figura 10-4 presenta losdatos relativos al peso de losindividuos, a la biomasa de lahojarasca y a la fracción liviana de laMO. Entre las 16 variablesdescriptivas empleadas, sólo una,que está relacionada con elcontenido de MO en la molleja, tuvo
una correlación significativa con lacantidad de hojarasca testigo(Cuadro 10-1); esa variable fue elporcentaje volumétrico de losfragmentos vegetales incluidos en loscomplejos órgano-minerales(r2 = 0.706, Cuadro 10-2).
No hubo correlación alguna entrela medición bajo la lupa de lascaracterísticas de las mollejas y sumedición bajo el microscopio,mientras que la cantidad de hojarascatestigo y la de hojarasca de galeríarevelaron una fuerte correlación(Cuadro 10-2).
La relación entre la abundanciade las partículas orgánicas yminerales (en el contenido de lamolleja) y su tamaño arrojó una curvabimodal (Figura 10-5), en la cual seaprecian dos granulometrías claras:la más fina que se haya observado enel estudio (0.8 µm) y dos fraccionesgruesas (de 0.05 a 0.2 mm). La líneavertical interrumpida que aparece enla gráfica separa las fraccionesobservadas bajo la lupa de lasobservadas al microscopio. Estas dosescalas de observación (lupa ymicroscopio) corresponden,probablemente, a una dieta realcompuesta por heces de otraslombrices (MO fina incluida en la fasemineral), por raíces >500 µm (que no
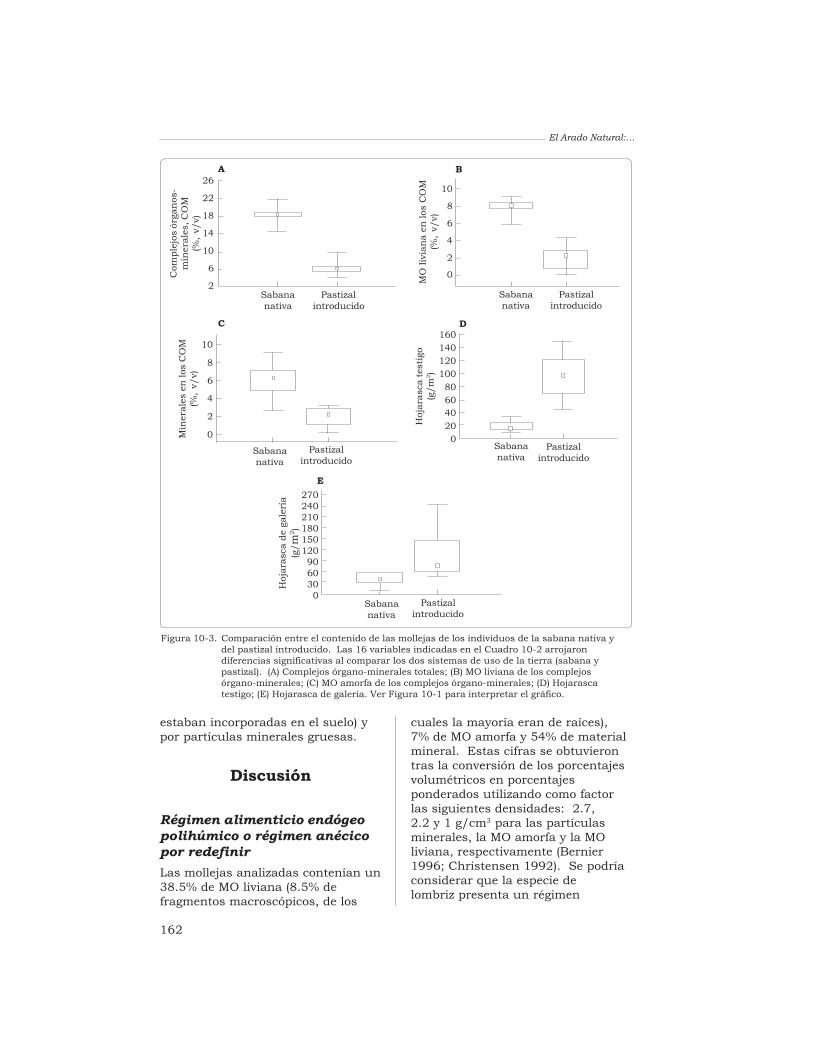
162
El Arado Natural:…
Sabananativa
Com
plej
os ó
rgan
os-
min
eral
es, C
OM
(%,
v/v)
26
22
18
14
10
6
2Pastizal
introducido
A
Sabananativa
MO
livi
ana
en lo
s C
OM
(%,
v/v)
10
8
6
4
2
0
Pastizalintroducido
B
Sabananativa
Min
eral
es e
n lo
s C
OM
(%,
v/v)
10
8
6
4
2
0Pastizal
introducidoSabananativa
Hoj
aras
ca te
stig
o(g
/m2 )
160140120100806040200
Pastizalintroducido
D
Hoj
aras
ca d
e ga
lerí
a(g
/m2 )
2702402101801501209060300
E
Sabananativa
Pastizalintroducido
C
Figura 10-3. Comparación entre el contenido de las mollejas de los individuos de la sabana nativa ydel pastizal introducido. Las 16 variables indicadas en el Cuadro 10-2 arrojarondiferencias significativas al comparar los dos sistemas de uso de la tierra (sabana ypastizal). (A) Complejos órgano-minerales totales; (B) MO liviana de los complejosórgano-minerales; (C) MO amorfa de los complejos órgano-minerales; (D) Hojarascatestigo; (E) Hojarasca de galería. Ver Figura 10-1 para interpretar el gráfico.
estaban incorporadas en el suelo) ypor partículas minerales gruesas.
Discusión
Régimen alimenticio endógeopolihúmico o régimen anécicopor redefinirLas mollejas analizadas contenían un38.5% de MO liviana (8.5% defragmentos macroscópicos, de los
cuales la mayoría eran de raíces),7% de MO amorfa y 54% de materialmineral. Estas cifras se obtuvierontras la conversión de los porcentajesvolumétricos en porcentajesponderados utilizando como factorlas siguientes densidades: 2.7,2.2 y 1 g/cm3 para las partículasminerales, la MO amorfa y la MOliviana, respectivamente (Bernier1996; Christensen 1992). Se podríaconsiderar que la especie delombriz presenta un régimen

La Dieta de una Lombriz Anécica...
163
Hojarascatestigo
Peso
(g)
240
180
120
60
0Hojarascade galería
A
Masade los
individuos
28
24
20
16
12
8
4
0MO
liviana delsuelo
B
Peso
(g)
Figura 10-4. Biomasa de los individuos y características de los sistemas. (A) Masa de la hojarasca(g/m2); (B) Masa de los individuos (en gramos) y de la MO liviana del suelo (mg/g suelo).Ver Figura 10-1 para la interpretación.
mg/
g de
su
elo
Cuadro 10-2. Correlaciones significativas entrelas variables descritas en elCuadro 10-1.
Hojarascade galería
Hojarasca testigo 0.90***
Complejos órgano-minerales -0.84*
* = prueba significativa al 5%; *** = pruebasignificativa al 0.5%.
alimentario anécico si el 77% de losfragmentos vegetales no sonmicroscópicos ni están yaincorporados en los agregadosórgano-minerales. La identificaciónde la mayoría de estos agregadoscomo deyecciones de lombricesimplica que otras lombrices habíanincorporado este material al suelo.En consecuencia, la ingestión
Cuadro 10-1. Lista de las 16 variables empleadas en las correlaciones y en la comparación de lasparcelas estudiadas.
Observaciones bajo la lupa Fragmentos vegetalesAgregadosMineralesRaíz, fragmentos macroscópicosParte aérea, fragmentos macroscópicos
Observaciones bajo el microscopio Fragmentos vegetales y bacterias, total(%, en volumen) Minerales, contenido total
Fragmentos vegetales y bacterias en los complejosórgano-mineralesMinerales en los complejos órgano-mineralesMO amorfa en los complejos órgano-minerales
Complejos órgano-minerales, total
MO amorfa libre
MO amorfa total
MO total
Gramos, g Peso de los individuos en alcohol
mg/g de suelo Fragmentos vegetales del suelo
g/m2 Hojarasca testigo
Hojarasca de galería
(%, en volumen)
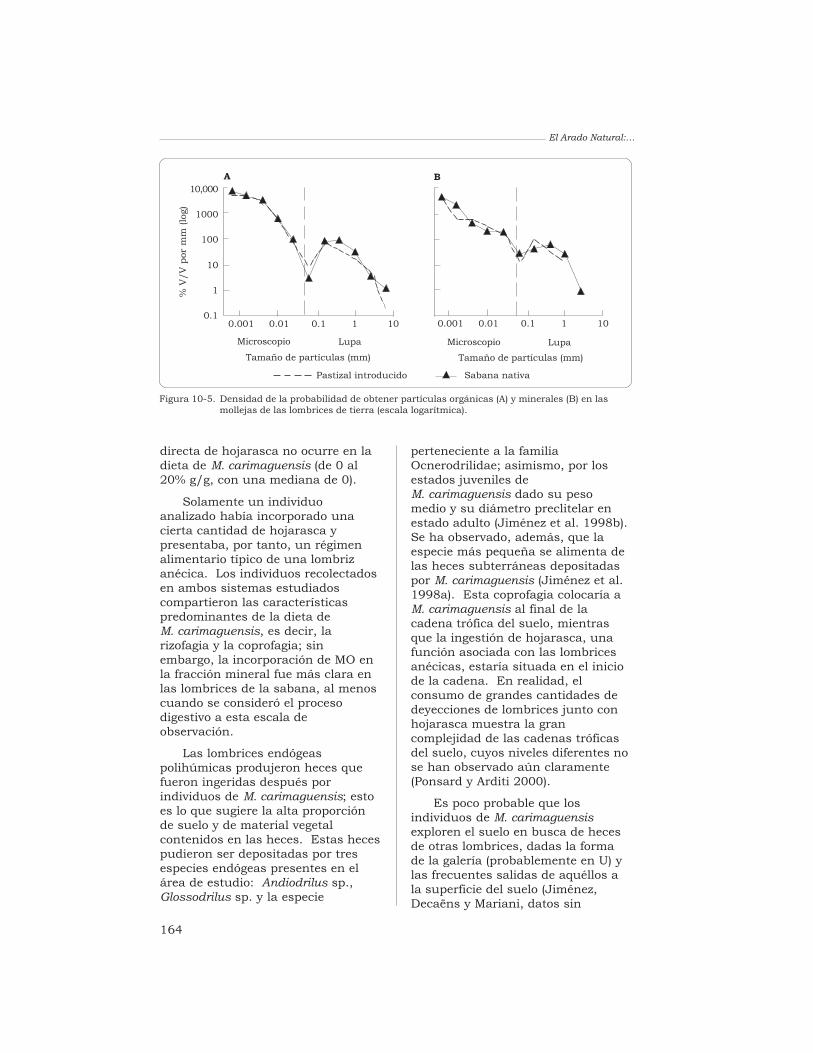
164
El Arado Natural:…
directa de hojarasca no ocurre en ladieta de M. carimaguensis (de 0 al20% g/g, con una mediana de 0).
Solamente un individuoanalizado había incorporado unacierta cantidad de hojarasca ypresentaba, por tanto, un régimenalimentario típico de una lombrizanécica. Los individuos recolectadosen ambos sistemas estudiadoscompartieron las característicaspredominantes de la dieta deM. carimaguensis, es decir, larizofagia y la coprofagia; sinembargo, la incorporación de MO enla fracción mineral fue más clara enlas lombrices de la sabana, al menoscuando se consideró el procesodigestivo a esta escala deobservación.
Las lombrices endógeaspolihúmicas produjeron heces quefueron ingeridas después porindividuos de M. carimaguensis; estoes lo que sugiere la alta proporciónde suelo y de material vegetalcontenidos en las heces. Estas hecespudieron ser depositadas por tresespecies endógeas presentes en elárea de estudio: Andiodrilus sp.,Glossodrilus sp. y la especie
perteneciente a la familiaOcnerodrilidae; asimismo, por losestados juveniles deM. carimaguensis dado su pesomedio y su diámetro preclitelar enestado adulto (Jiménez et al. 1998b).Se ha observado, además, que laespecie más pequeña se alimenta delas heces subterráneas depositadaspor M. carimaguensis (Jiménez et al.1998a). Esta coprofagia colocaría aM. carimaguensis al final de lacadena trófica del suelo, mientrasque la ingestión de hojarasca, unafunción asociada con las lombricesanécicas, estaría situada en el iniciode la cadena. En realidad, elconsumo de grandes cantidades dedeyecciones de lombrices junto conhojarasca muestra la grancomplejidad de las cadenas tróficasdel suelo, cuyos niveles diferentes nose han observado aún claramente(Ponsard y Arditi 2000).
Es poco probable que losindividuos de M. carimaguensisexploren el suelo en busca de hecesde otras lombrices, dadas la formade la galería (probablemente en U) ylas frecuentes salidas de aquéllos ala superficie del suelo (Jiménez,Decaëns y Mariani, datos sin
Figura 10-5. Densidad de la probabilidad de obtener partículas orgánicas (A) y minerales (B) en lasmollejas de las lombrices de tierra (escala logarítmica).
% V
/V p
or m
m (l
og)
10,000
1000
100
10
1
0.10.001 0.01 0.1 1 10
A
Microscopio Lupa
Tamaño de partículas (mm)
B
Tamaño de partículas (mm)
Pastizal introducido Sabana nativa
Microscopio Lupa
0.001 0.01 0.1 1 10

La Dieta de una Lombriz Anécica...
165
publicar). Hay además un númerorelativamente alto de heces en lasuperficie del suelo que fuerondepositadas por lombrices deespecies endógeas, como lo muestrala cartografía superficial de lasestructuras biogénicas en la mismaparcela de sabana nativa del estudio(Mariani, Decaëns y Jiménez, datossin publicar). De hecho, esteestudio revela que las hecessuperficiales de Glossodrilus sp. yde la especie de Ocnerodrilidae sonmuy escasas, mientras que las deAndiodrilus sp. cubren el 2.2% de lasuperficie total del suelo, aunque laheterogeneidad espacial presente esbastante alta (132% CV). Ahorabien, es necesario realizar estudiossobre la composición de lahojarasca, del humus y de ladistribución vertical de las heces,para comprender los factores quedeterminan la ingestión de unamasa órgano-mineral por parte deM. carimaguensis y poder atribuirlea esta acción un significadoecológico. Si se confirma lahipótesis de la ingestión selectiva deuna masa órgano-mineral en lasuperficie del suelo, se habráencontrado entonces uncomportamiento anécico típico,aunque con un sentido ecológicodiferente. Este hecho sería laconsecuencia de la adaptación deuna especie anécica a la ausenciade hojarasca en las sabanasdurante la quema periódica deéstas.
Variabilidad estacional delrégimen alimentario enM. carimaguensisEl análisis de las heces superficialesda resultados poco coherentesfrente a los datos obtenidos en esteestudio; además, se ha probadoclaramente que M. carimaguensisincorpora hojarasca a la masaórgano-mineral (Guggenberger et al.
1995, 1996). Ahora bien, esosfragmentos de hojarasca pudieronhaber sido incluidos durante ladeyección de heces que hace lalombriz en la superficie del suelo.En realidad, estas heces contienendos tipos de fragmentos vegetales:los primeros, grandes y rígidos,indican que no fueron ingeridos porla lombriz sino cubiertos,probablemente, por las heces de éstaen el momento de la deposición; lossegundos son fragmentos muy finos(de algunos mm), visibles después deque las gotas de lluvia destruyen laenvoltura fina (cortex) de ladeyección (Mariani et al., datos sinpublicar) y que, probablemente,pasaron por el intestino de lalombriz. Otras deyecciones,recogidas en las mismas parcelas yen las mismas fechas que las de esteestudio, contenían más fragmentosde semillas que las muestras desuelo (Decaëns et al. 2003). Comoresultado de lo anterior, la especieM. carimaguensis estaríaseleccionando su alimento, lo que nose confirmó en este estudio, quetrataba del contenido de susmollejas. Al analizar las seccionesfinas cortadas en las heces frescasde M. carimaguensis que serecolectaron en el pastizal y en lasabana nativa, se halló una granheterogeneidad en la distribución delos fragmentos vegetales finos dentrode las heces (Decaëns 2000). Dichaheterogeneidad se podría explicarmediante la variabilidad observadaen el régimen alimenticio de laslombrices de M. carimaguensis segúnla escala de tiempo en que seformaron las heces, es decir, en unpromedio de 3 días y con un máximode 3 semanas (Decaëns 2000;Decaëns y Jiménez, com. pers.).Dada esta hipótesis, las lombrices deM. carimaguensis podrían adoptar,periódicamente, un régimen anécicotípico.

166
El Arado Natural:…
ConclusionesEste estudio demuestra que la dietaalimenticia de los individuos deM. carimaguensis no contiene,simplemente, una mezcla de suelo yde hojarasca, ya que estas lombricespueden alimentarse también deraíces y de las heces de otraslombrices. Hace falta un análisisdetallado del humus, de la hojarascay del contenido intestinal de laslombrices, así como un estudio delciclo nictameral, para lograr dosobjetivos: descubrir el agente queincorpora la hojarasca a la partemineral del suelo y la forma en quelo hace, y determinar el tipo derelación que existe entre M.carimaguensis y las otras especies dela macrofauna del suelo.
AgradecimientosLos autores agradecen a P. Lavelledel LEST (IRD/Universidad de ParísVI, Francia) por sus comentariosútiles sobre una versión previa deeste manuscrito; asimismo, a lagente de la estación de Carimagua,Colombia, por el trabajo realizadodurante la temporada de muestreo.Agradecen también a P. Margerie, dela Universidad de Rouen, Francia,por su ayuda en la identificación delos fragmentos vegetales.
ReferenciasBernier N. 1996. Altitudinal changes in
humus form dynamics in a spruceforest at the montane level. PlantSoil 178:1-28.
Bernier N. 1998. Earthworm feedingactivity and development of thehumus profile. Biol Fertil Soils26:215-223.
Bolton PJ; Phillipson J. 1976. Burrowing,feeding, egestion and energy budget ofAllolobophora rosea (Savigny)(Lumbricidæ). Oecologia 23:225-245.
Bouché MB. 1977. Stratégies lombriciennes.En: Lohm U; Persson T, eds. Soilorganisms as components ofecosystems. Ecol Bull 25:122-132.
Bouché MB; Kretzschmar A. 1974. Fonctiondes lombriciens; II: Recherchesméthodologiques pour l’analyse du solingéré (étude du peuplement de lastation R.C.P.-165/P.B.I.). Rev EcolBiol Sol 11:127-139.
Christensen BT. 1992. Physical fractionationof soil and organic matter in primaryparticle size and density separates.Adv Soil Sci 20:1-90.
Decaëns T. 2000. Degradation dynamics ofsurface earthworm casts in grasslandsof the Eastern Plains of Colombia. BiolFertil Soils 32:149-156.
Decaëns T; Jiménez JJ; Lavelle P. 1999a.Effect of exclusion of the anecicearthworm Martiodrilus carimaguensisJiménez and Moreno on soil propertiesand plant growth in grasslands of theEastern Plains of Colombia.Pedobiologia 43:835-841.
Decaëns T; Mariani L; Lavelle P. 1999b. Soilsurface macrofaunal communitiesassociated with earthworm casts ingrasslands of the Eastern Plains ofColombia. Appl Soil Ecol 13:87-100.
Decaëns T; Rangel AF; Asakawa N; ThomasR. 1999c. Carbon and nitrogendynamics in in-situ ageing earthwormcasts in grasslands of the EasternPlains of Colombia. Biol Fertil Soils30:20-28.
Decaëns T; Mariani L; Betancourt N;Jiménez JJ. 2003. Seed dispersion bysurface casting activities ofearthworms in Colombian grasslands.Acta Oecol 24(4):175-185.
Decaëns T; Lavelle P; Jiménez JJ; EscobarG; Rippstein G. 1994. Impact of landmanagement on soil macrofauna inthe Oriental Llanos of Colombia. Eur JSoil Biol 30:157-168.

La Dieta de una Lombriz Anécica...
167
Ferrière G. 1980. Fonctions deslombriciens; VII: Une méthoded’analyse de la matière organiquevégétale ingérée. Pedobiologia20:263-273.
Fisher MJ; Rao IM; Ayarza MA; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage byintroduced deep-rooted grasses inthe South American savannas.Nature 371:236-238.
Guggenberger G; Thomas RJ; Zech W.1996. Soil organic matter withinearthworm casts of an anecic-endogeic tropical pasturecommunity, Colombia. Appl SoilEcol 3:263-274.
Guggenberger G; Zech W; Thomas RJ.1995. Lignin and carbohydratealteration in particle-size separatesof an oxisol under tropical pasturesfollowing native savanna. Soil BiolBiochem 27:1629-1638.
Hochberg Y. 1988. A sharper Bonferroniprocedure for multiple tests ofsignificance. Biometrika75:800-802.
James SW; Cunningham MR. 1989.Feeding ecology of someearthworms in Kansas tallgrassprairie. Am Midl Nat 121:78-83.
James SW; Seastedt TR. 1986. Nitrogenmineralization by native andintroduced earthworms: effects onbig bluestem growth. Ecology67:1094-1097.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Glossoscolecidae, Oligochaeta), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9(1-3):153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28(1):101-110.
Judas M. 1992. Gut content analysis ofearthworms (Lumbricidae) in abeechwood. Soil Biol Biochem24:1413-1417.
Kanyonyo Kakajondo JB. 1984. Ecologiealimentaire du ver de terredetritivore Millsonia lamtoiana(Oligochaeta, Acanthodrilidae) dansla savane de Lamto (Côte d’Ivoire),Tesis (3er. ciclo). Université de ParisVI.
Lavelle P; Brussaard L; Hendrix P. (eds).1999. Earthworm management intropical agroecosystems. CABIPublishing. Oxon y Nueva York.300 p.
Legendre L; Legendre P. 1998. Numericalecology: developments inenvironmental modelling 20. 2a. ed.Elsevier, Amsterdam.
Piearce TG. 1972. The calcium relationsof selected Lumbricidae. J AnimEcol 41:167-188.
Ponsard S; Arditi R. 2000. What canstable isotopes (δ15N and δ13C) tellabout the food web of soilmacroinvertebrates? Ecology81:852-864.
Scherrer B. 1984. Biostatistique. GaëtanMorin éditeur. Montreal, Canadá.
Senapati BK; Lavelle P; Giri S; PashanasiB; Alegre J; Decaëns T; Jiménez JJ;Albrecht A; Blanchart E; MahieuxM; Rousseaux L; Thomas RJ;Panigrahi PK; Venkatachalam M.1999. In-soil earthwormtechnologies for tropicalecosystems. En: Lavelle P;Brussaard L; Hendrix P, eds.Earthworm management in tropicalagrosystems. CABI Publishing,Oxon y Nueva York. p 199-238.

Propiedades de las Estructuras...
169
PARTE III
Efectos Producidos por los Ingenierosdel Ecosistema en las PropiedadesFísicas del Suelo, a Escala de las
Estructuras Biogénicas

Propiedades de las Estructuras...
171
ResumenSe tomaron muestras de 14 tipos deestructuras físicas construidas porlos ingenieros del ecosistema sobrela superficie del suelo de los LlanosOrientales de Colombia. Seidentificaron, además, losinvertebrados que produjeron esasestructuras. Se realizaron análisisfísicos (de tamaño de agregados y deestabilidad, de densidad aparente) yquímicos (de contenidos totales de C,N y P y de pH). Se identificaron asítres grandes grupos de estructuras:(a) las heces de lombrices quepresentaban las siguientescaracterísticas: una densidadaparente (DA) alta (1.3 a 1.4 g/cm3),agregados de gran tamaño (7 a10 mm), altos contenidos de C (3% a4%) y nutrientes asimilables; (b) lostermiteros, cuya DA era baja (0.6 a0.9 g/cm3), estaban formados poragregados de gran tamaño (8 a9 mm), y tenían altos contenidos de
C (3.5% a 10%) y de nutrientesasimilables; (c) las galeríassuperficiales elaboradas por termitasy los hormigueros, que sonligeramente compactos (0.4 a0.7 g/cm3) y granulares (tamaño deagregados <1.5 mm), tienen uncontenido bajo de C orgánico (menosde 1.5%) y de nutrientes asimilables.Estos resultados indican que seproduce una gran diversidad deestructuras biogénicas en los suelosestudiados y sugieren una posibleclasificación funcional de losorganismos ingenieros, que sebasaría en los diferentes atributosfuncionales de dichas estructuras.
Palabras clave: Colombia, ecologíadel suelo, estructuras biogénicas,estructura del suelo, hormigas,ingenieros del ecosistema,lombrices, materia orgánica,sabana, termitas
IntroducciónLos “ingenieros del ecosistema”, o“ingenieros ecológicos” (sensu Joneset al. 1994) se definen comoorganismos que manifiestan lacapacidad de estructurar físicamenteel medio en el cual viven, mediante laproducción de artefactos oestructuras biogénicas. Con ellas,los organismos ingenieros puedenafectar la disponibilidad o la
CAPÍTULO 11
Propiedades de las EstructurasConstruidas por los Ingenieros delEcosistema en la Superficie del Suelode una Sabana Colombiana1
T. Decaëns* , J. H. Galvis** y E. Amézquita**
1. Publicado en Comptes Rendues de l’Académiedes Sciences, Série III 324(5), p 465-478, porDecaëns et al. Copyright © 2001 de Elsevier.Impreso con permiso de la casa editorial.
* Laboratoire d’Écologie des Sols Tropicaux delInstitut de Recherche pour le Développement(IRD)/Unidad de Suelos y Nutrición dePlantas (CIAT), IRD-Bondy, 32 Av. HenriVaragnat, F-93143 Bondy Cedex, Francia.
** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.

172
El Arado Natural:…
accesibilidad de uno o más recursosutilizados por otros organismos. Susactividades, entre ellas la producciónde las estructuras biogénicas, puedenmodificar, por tanto, la abundancia ola estructura comunitaria de otraspoblaciones de organismos sin queello implique ninguna relación directade tipo trófico (o sea, de predación, deparasitismo, de mutualismo o decompetencia) entre los organismos(Jones et al. 1994, 1997).
Las lombrices de tierra, lastermitas y las hormigas son losgrupos que principalmente seconsideran ingenieros del suelo;además, la escala espacio-temporalde las estructuras biogénicas queellos producen (p.ej., turrículos,galerías, domos) puede ayudar aevaluar el impacto de dichasestructuras en el suelo y en losorganismos que viven en él (Andersone Ingram 1993; Lavelle 1997). Lanaturaleza físicoquímica, laabundancia, el tamaño y, en especial,la dinámica de la producción y ladegradación de estas estructuras son,por tanto, parámetros que permitenevaluar el tipo y la amplitud de losefectos indirectos que los ingenierosecológicos ejercen en el ambiente(Jones et al. 1994, 1997). Lasestructuras biogénicas son el reflejode ciertos atributos funcionales de lasespecies que las producen, atributosque están ligados directamente a ladefinición de ingenieros delecosistema; asimismo, la descripciónde dichas estructuras puede servirpara hacer una clasificaciónfuncional de los ingenieros del suelo.
El objetivo de este estudio esclasificar los ingenieros ecológicospartiendo de la naturaleza de lasestructuras físicas que ellosproducen. Con tal fin, se handescrito las estructuras biogénicasmás comunes observadas sobre lasuperficie del suelo de una sabanabien drenada de Colombia. Se ha
establecido así una tipología deestas estructuras partiendo de12 variables que describen lanaturaleza física de los agregadosproducidos [densidad aparente (DA),tamaño de los agregados], suestabilidad estructural, y sunaturaleza orgánica y química (pH,C orgánico, N, P, Ca, K, Mg, P, Al ysaturación de aluminio).
Materiales y Métodos
Area del estudioEste estudio se realizó en el CentroNacional de InvestigacionesAgropecuarias de Carimagua (delconvenio CORPOICA-CIAT), situado a4°37’ N y 71°19’ O, en la regiónfitogeográfica de las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia.El clima es de tipo subhúmedotropical, el valor medio anual de latemperatura es de 26 °C, el de laprecipitación es de 2300 mm, y hayuna época seca muy fuerte denoviembre a marzo. La vegetaciónestá determinada por la topografía:hay sabanas herbáceas en las áreasmás altas (los “altos”) y tambiénbosques de galería y sabanasinundables en las partes bajas (los“bajos”). Los suelos son ferralíticos ycaoliníticos (Oxisoles) en las áreasmás altas; en las zonas inundablesaparecen ferrisoles y plintitas(Ultisoles). Los dos tipos de suelos secaracterizan por su estructuragranular y por tener una fertilidadquímica muy baja: pH (H2O) < 5,saturación de aluminio >80%, yCIC < 5 meq/100 g (datos del CIAT).
Parcelas experimentalesLas observaciones y el muestreo serealizaron en una sabana de sueloferralítico. La vegetación es propiade una sabana natural no

Propiedades de las Estructuras...
173
pastoreada, y está dominada porTrachypogon vestitus Anders. y porPaspalum pectinatum Nees. Laparcela se manejó siguiendo elmétodo tradicional en que la quemase realiza cada año durante laestación seca (en febrero). Lasmuestras se tomaron en 1996 alinicio de la época lluviosa (durante elmes de mayo), período de máximaactividad de la macrofauna del suelo.
Identificación y descripciónmorfológicaLas parcelas fueron minuciosamenterevisadas (se empleó 1 hora en cadaparcela) por tres personas. Todas lasestructuras biogénicas encontradassobre la superficie de la sabananatural fueron descritas y losmacroinvertebrados responsables desu formación se identificaron hastael nivel más preciso posible (familia,género o especie). Con este muestreono se pretendía elaborar una listaexhaustiva de todas las estructurasbiogénicas producidas en la sabana,sino hacer una descripción detalladade las más representativas dondehubiera ese tipo de vegetación.
Se describió el color, el tamaño,la forma y el aspecto general de lasestructuras en cada especieinventariada. Se usaron formasgeométricas simples con el fin deevaluar fácilmente el volumen desuelo trasladado, en cada tipo deestructura, hasta la superficie delterreno (ver Apéndice 1). Alrededorde 500 g de material fresco serecolectaron en cada una de lasestructuras encontradas.
Análisis químicosEl contenido de C orgánico y el deelementos minerales se determinaronen las estructuras biogénicas cuyapresencia permitía la recolección dematerial abundante. Se muestreó el
estrato superficial del suelo, de 0 a10 cm de profundidad, con la ayudade un cilindro de 5 cm de diámetro y10 cm de altura. Los análisis seefectuaron siguiendo los métodosestándar recomendados por elprograma TSBF (Programa deBiología y Fertilidad de los SuelosTropicales) (Anderson e Ingram1993).
Los carbohidratos hidrosolublesson un factor importante en laestabilización de los agregados delsuelo (Gijsman y Thomas 1995;Haynes y Swift 1990; Tisdall 1994).Se midió entonces su concentracióntanto en la capa de 0 a 10 cm dedichos agregados como en un gruposelecto y representativo deestructuras biogénicas: 2 tipos deturrículos de lombrices, 2 determiteros y 2 de hormigueros; todosellos fueron escogidos en razón de suabundancia en la superficie del sueloestudiado. Las estructuras sesecaron al aire y fueron tamizadaspara obtener tamaños de 0.250 y2 mm. Se tomaron luego 2 g de cadamuestra (suelo superficial yestructuras) que fueron agitadosdurante 8 horas en 20 ml de aguacaliente (80 °C). La soluciónobtenida se centrifugó durante6 minutos a 13,000 rpm y elsobrenadante se recogió y secentrifugó también en las mismascondiciones. Los carbohidratos sedeterminaron mediante un métodocolorimétrico estándar (Cheschire1979).
Densidad aparenteSe evaluó la DA de todas lasestructuras biogénicas a partir demuestras secadas a 75 °C durante48 horas. En las estructurasblandas (los hormigueros), estadensidad se determinó a partir delvolumen ocupado por la muestra enuna probeta graduada. En las

174
El Arado Natural:…
estructuras duras (los turrículos delas lombrices y los termiteros), sedeterminó mediante el método delagua desplazada por la muestra, unavez impermeabilizada ésta con papelde parafina (‘Parafilm’). La DA delsuelo de la sabana se calculó al pesarun volumen conocido del mismo, esdecir, una muestra de suelo obtenidacon un cilindro de 5 cm de diámetro y10 cm de alto, previamente secado a75 °C durante 48 horas.
Distribución de los agregadosen clases de tamaño yestabilidad estructuralLos agregados de todas lasestructuras biogénicas sedistribuyeron en clases según sutamaño. Se recolectaron en lasabana, como mínimo, cuatromuestras de cada estructura descrita.Se extrajeron, además, ocho cilindrosde suelo (8 cm de diámetro y 10 cmde altura) con el fin de comparar losagregados de las estructuras con losde la capa superficial del suelo.
La distribución de los agregadosen clases de tamaño se evaluómediante el método de tamizado enseco (Blanchart 1990; Kemper yRousenau 1986), utilizadoanteriormente en estudios sobre lossuelos de las sabanas colombianas(Gijsman 1996; Gijsman y Thomas1996). Las muestras se secaron alaire durante 4 días y luego se dejaroncaer desde una altura de 2 m sobreuna superficie dura con el fin dedisociar los agregados. Acontinuación, se tamizaron 20 vecesa través de un juego completo detamices de 0.053, 0.125, 0.250, 0.5,1, 2, 5 y 10 mm; cada una de estasfracciones se secó durante 48 horas a75 °C antes de ser pesada. Ladistribución de los agregados de unamuestra completa se expresó como eldiámetro ponderado medio (DPM)según las siguientes ecuaciones:
i
n
ii wxDPM
1y la fracción de los agregados:
totalmuestra
itamizi M
Mw )(
donde,ix es el diámetro medio de cada
fracción de agregados; es el peso seco de las
partículas retenidas en el tamiz i; es el peso seco de la
muestra total.
La estabilidad de los agregadosen agua (EAA) se determinó en lasmuestras evaluadas para calcular laconcentración de los carbohidratossolubles en agua. El métodoempleado supone la aplicación deuna fuerza destructiva de ciertamagnitud (Gijsman y Thomas 1996;Kemper y Rousenau 1986). Setoman 5 g de agregados (Ø de 2 a5 cm), se secan al aire y luego sehumedecen colocándolos sobre unacapa de arena húmeda a unasucción de 1 cm de agua durantemenos de 45 minutos. La muestrase coloca luego sobre un tamiz de1 mm de malla, se sumerge en aguay, con la ayuda de un aparatoautomático, se agita de arriba abajo(amplitud: 3 cm) durante 3 minutos,a razón de 34 oscilaciones porminuto. El suelo que permanecesobre el tamiz se seca luego a 105 °Cy se pesa.
Se calculó también la cantidadde arena de la muestra total y delsuelo retenido por el tamiz de 1 mm.Para lograrlo, los agregados sedispersaron en una solución al 5%de hexametafosfato antes de serpasados por el tamiz de 1 mm demalla. La EAA se expresa como elpeso de los agregados estables(corregido según su contenido dearena) dividido por el peso de la
[1]
[2]
Mtamiz i
Mmuestr total

Propiedades de las Estructuras...
175
muestra inicial (corregido según elcontenido de arena), del modosiguiente:
EAA(%) =Mm. estable Marena en m. estable ×100
Mm. total Marena en m. total
donde, es el peso seco de la
muestra inicial (m. = muestra);
es el peso seco de laspartículas retenidas en el tamizdespués de ser dispersadas enagua;
y sonlos pesos secos de la arenacontenida en la muestra inicial yde la arena dispersada en agua,respectivamente.
Análisis estadísticoLas comparaciones entre las mediasse obtuvieron mediante un ANOVA ycon las pruebas PLSD de Fisher, aun nivel de significación de 0.05.
La tipología de las estructuras seestableció mediante un análisis decomponentes principales (ACP). Seutilizaron los parámetros químicos yfísicos que se calcularon en cada unade las estructuras identificadas(propiedades químicas, DA y DPM).De las estructuras producidas por lashormigas sólo se emplearon aquellasen las que se disponía de un juegocompleto de datos. La matrizutilizada contenía 11 filas (objetos =11 estructuras muestreadas) y 12columnas (variables = característicasfísicas y químicas). El análisis serealizó con la ayuda del paqueteinformático ADE-4 (Thioulouse et al.1997).
Se emplearon regresiones linealespara evaluar las relaciones quepudieran existir entre el contenido decarbohidratos de las muestras y laEAA. La normalidad de los datos se
había verificado previamentemediante la prueba de Kolmogorov-Smirnov (Lilliefors 1967), ejecutadasiguiendo el programa estadísticoVerNorm 3.0 del paquete“R package” (Legendre y Vadour1991). La normalidad de los datosse aceptó con un nivel designificación de 0.05 para todos losjuegos de datos.
Resultados
Identificaciónmicromorfológica ydescripcionesSe identificaron 14 especies demacroinvertebrados comoresponsables de la formación de lasestructuras biogénicas superficialeshalladas en la sabana nativa. Deéstas se recolectaron ocho tipos dehormigueros (Fotos 11-1 y 11-2),tres tipos de domos epígeosproducidos por las termitas —de loscuales uno parece construido conuna pasta acartonada y los otrosdos con una mezcla órgano-mineral(Fotos 11-3 y 11-4)—, un tipo deplaca superficial (o galeríasuperficial sólida) producida por unaespecie de termita (Foto 11-5), y dostipos de turrículos producidos porlombrices de tierra (Fotos 11-5 y11-6). En el Cuadro 11-1 se indicanlas 14 especies demacroinvertebrados y se describenbrevemente las estructuras que elloselaboran. En el Apéndice 2 sepresentan las demás estructurasbiogénicas encontradas sobre lasuperficie de la sabana deCarimagua (Fotos 11-8A a 11-15A).
Desde un punto de vistomicromorfológico, una bioestructurabiogénica puede considerarse segúnlas siguientes definiciones de Bal(1973):
totalmM .
Mm estable.
Marena en m total. Marena en m estable.

176
El Arado Natural:…
Foto 11-2. Montículo epígeo y entrada de una colonia de hormigas Camponotus sp. en la sabananativa (escala: longitud de la foto = 50 cm).
Foto 11-1. Montículo epígeo y entrada de una colonia de hormigas Trachymyrmex sp. en la sabananativa (escala: longitud de la foto = 40 cm).

Propiedades de las Estructuras...
177
Foto 11-4. Montículo epígeo de una coloniade termitas Spinitermes sp. en lasabana nativa (escala: longitudde la foto = 1 metro).
Foto 11-3. Montículo epígeo de una coloniade termitas Velocitermes sp. enla sabana nativa (escala:longitud de la foto = 30 cm).

178
El Arado Natural:…
Foto 11-6. Turrículos de M. carimaguensis en la sabana nativa (escala: longitud de la foto = 20 cm).
Foto 11-5. Placas de Ruptitermes sp. y turrículos de Andiodrilus sp. en la sabana nativa (escala:longitud de la foto = 30 cm).
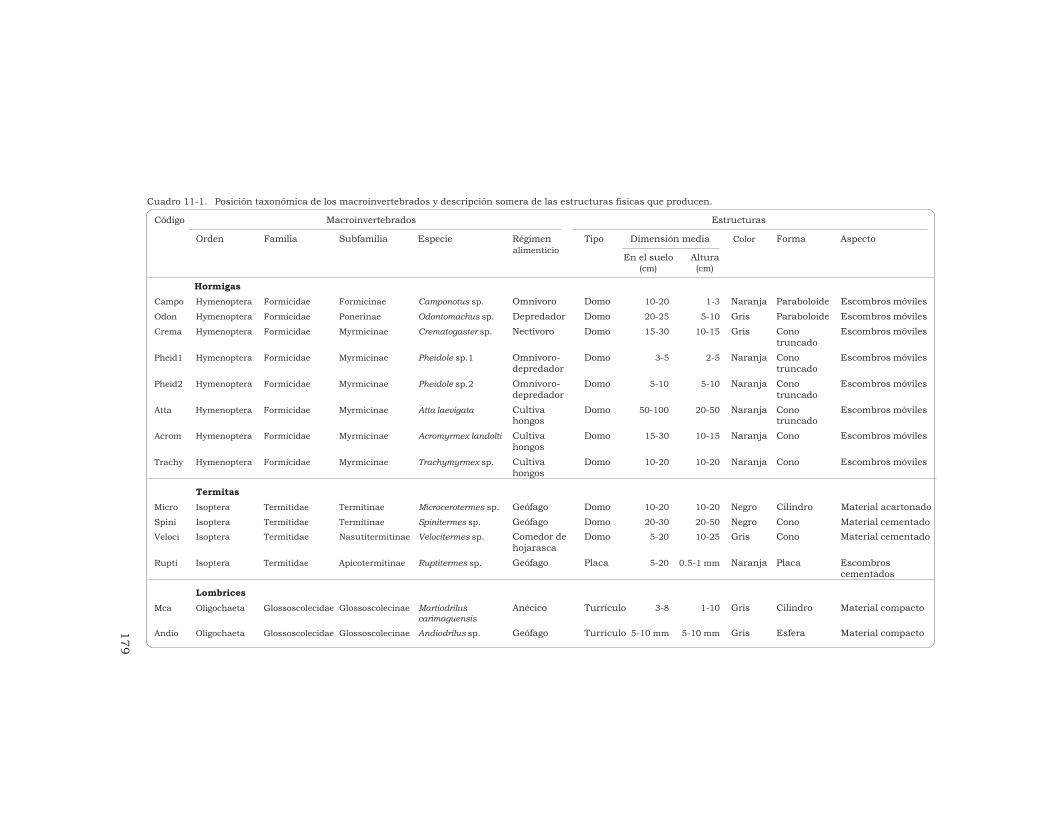
Propiedades de las Estructuras...
179
Cuadro 11-1. Posición taxonómica de los macroinvertebrados y descripción somera de las estructuras físicas que producen.
Código Macroinvertebrados Estructuras
Orden Familia Subfamilia Especie Régimen Tipo Dimensión media Color Forma Aspecto alimenticio
En el suelo Altura(cm) (cm)
HormigasCampo Hymenoptera Formicidae Formicinae Camponotus sp. Omnívoro Domo 10-20 1-3 Naranja Paraboloide Escombros móvilesOdon Hymenoptera Formicidae Ponerinae Odontomachus sp. Depredador Domo 20-25 5-10 Gris Paraboloide Escombros móvilesCrema Hymenoptera Formicidae Myrmicinae Crematogaster sp. Nectívoro Domo 15-30 10-15 Gris Cono Escombros móviles
truncadoPheid1 Hymenoptera Formicidae Myrmicinae Pheidole sp.1 Omnívoro- Domo 3-5 2-5 Naranja Cono Escombros móviles
depredador truncadoPheid2 Hymenoptera Formicidae Myrmicinae Pheidole sp.2 Omnívoro- Domo 5-10 5-10 Naranja Cono Escombros móviles
depredador truncadoAtta Hymenoptera Formicidae Myrmicinae Atta laevigata Cultiva Domo 50-100 20-50 Naranja Cono Escombros móviles
hongos truncadoAcrom Hymenoptera Formicidae Myrmicinae Acromyrmex landolti Cultiva Domo 15-30 10-15 Naranja Cono Escombros móviles
hongosTrachy Hymenoptera Formicidae Myrmicinae Trachymyrmex sp. Cultiva Domo 10-20 10-20 Naranja Cono Escombros móviles
hongos
TermitasMicro Isoptera Termitidae Termitinae Microcerotermes sp. Geófago Domo 10-20 10-20 Negro Cilindro Material acartonadoSpini Isoptera Termitidae Termitinae Spinitermes sp. Geófago Domo 20-30 20-50 Negro Cono Material cementadoVeloci Isoptera Termitidae Nasutitermitinae Velocitermes sp. Comedor de Domo 5-20 10-25 Gris Cono Material cementado
hojarascaRupti Isoptera Termitidae Apicotermitinae Ruptitermes sp. Geófago Placa 5-20 0.5-1 mm Naranja Placa Escombros
cementados
LombricesMca Oligochaeta Glossoscolecidae Glossoscolecinae Martiodrilus Anécico Turrículo 3-8 1-10 Gris Cilindro Material compacto
carimaguensis
Andio Oligochaeta Glossoscolecidae Glossoscolecinae Andiodrilus sp. Geófago Turrículo 5-10 mm 5-10 mm Gris Esfera Material compacto

180
El Arado Natural:…
• El resultado de la ß-acumulaciónde agregados del sueloprovenientes de los horizontesprofundos y transportados porlos invertebrados a la superficiedel suelo (el terraplén de unhormiguero).
• Un ‘agrotúbulo’, es decir, unaestructura tubular constituidapor agregados cementados queno muestran ninguna orientacióndireccional (las placas, o canales,superficiales de las termitas).
• Un ‘modexis’, es decir, unaestructura tridimensional eindividual formada porexcrementos producidos por unanimal (las heces de laslombrices de tierra). Losturrículos de Martiodriluscarimaguensis podríanclasificarse, con mucha mayorprecisión, como ‘modexotúbulos’,porque presentan un huecotubular central que correspondea la entrada de la galería de lalombriz.
• Una red tridimensional de‘escatotúbulos’, es decir, deestructuras tubularesconstituidas por excrementos deinvertebrados y que no estánclaramente individualizadasunas con respecto a las otras (losdomos epígeos de los termiteros).
Propiedades químicasLos turrículos de las lombrices y lostermiteros presentan, en general,contenidos de C orgánico y deelementos minerales que superan losdel suelo superficial de la sabana;por ejemplo, +8.6% y +248.3% de Corgánico en los turrículos deAndiodrilus sp. y en los domos deMicrocerotermes sp., respectivamente(Figura 11-1). El pH aumentóigualmente, mientras que la
concentración de Al disminuyó enforma significativa. Los hormiguerosy las placas producidas porRuptitermes sp. presentaron, encambio, contenidos de C orgánicobastante inferiores a los del suelosuperficial, o sea, de –47.6% en lashormigas del género Pheidole y de–69.2% en las de Atta laevigataSmith y Acromyrmex landolti Forel.El pH y el contenido de los elementosminerales no cambiaron o fueroninferiores respecto a los del suelo,mientras que la saturación de Aldisminuyó ligeramente.
La concentración decarbohidratos hidrosolubles en loshormigueros y en los turrículos deAndiodrilus sp. fue equivalente oinferior a la del suelo superficial dela sabana, es decir, –29.7% enTrachymyrmex sp. y –70.3% enA. laevigata (Figura 11-2A). Encambio, los agregados queconstituyen los turrículos deM. carimaguensis y los termiterostenían concentraciones dos vecessuperiores a las del suelo, es decir,+12.8% en los de M. carimaguensis,+78.4% en los de Spinitermes sp. y+132.4% en los de Velocitermes sp.
Propiedades físicasLa DA de las estructuras biogénicasvarió mucho según la especie deinvertebrados considerada. Laslombrices de tierra, por ejemplo,producen turrículos compactos cuyaDA es superior a 1.30 g/cm3, esdecir, de 10% a 20% superior a ladel estrato 0-10 cm del suelo de lasabana (Cuadro 11-2). Las termitasy las hormigas, en cambio, producenestructuras menos compactas que elsuelo circundante, o sea, con unaDA de 0.90 g/cm3 o inferior a ésta.
La distribución en clases detamaño de los agregados‘bioformados’ fue muy diferente. Losturrículos de las lombrices y los
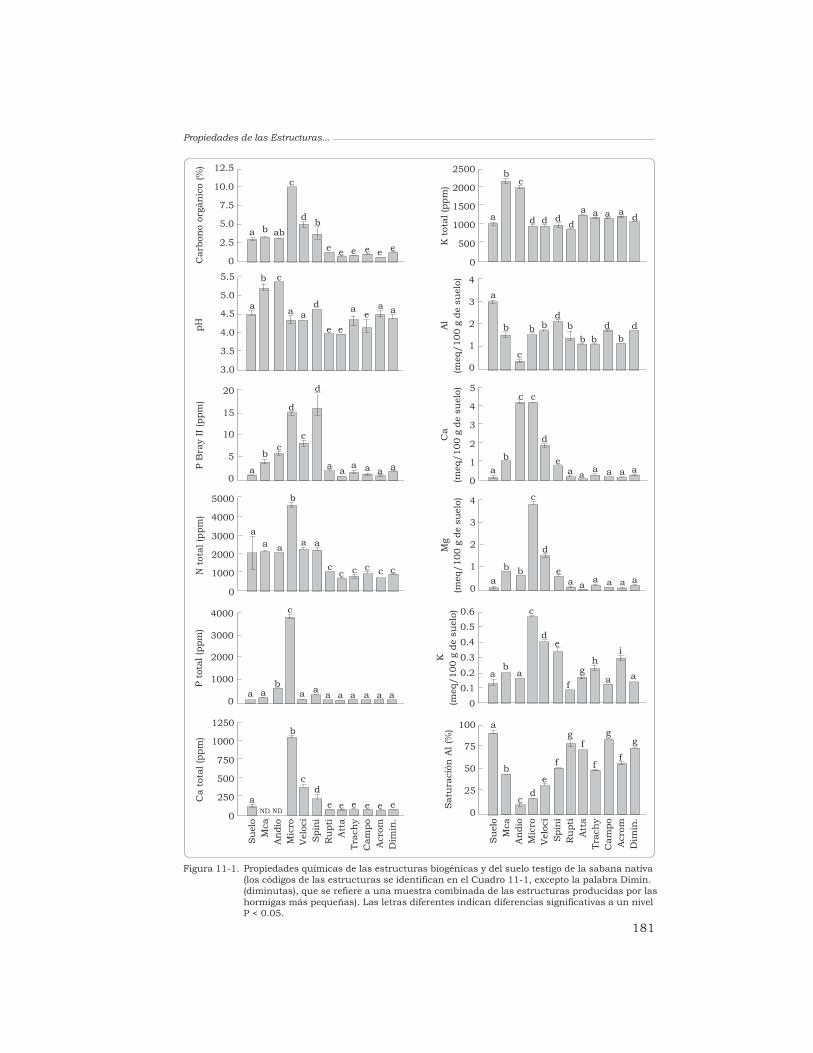
Propiedades de las Estructuras...
181
12.5
10.0
7.5
5.0
2.5
0Car
bon
o or
gán
ico
(%)
a b ab
c
d b
e e e e e e
2500
2000
1500
1000
500
0
K t
otal
(ppm
)
a
bc
d d d da a a a d
5.5
5.0
4.5
4.0
3.5
3.0
pH
a a ad
e e
a ea a
4
3
2
1
0
Al
(meq
/100
g d
e su
elo)
a
b
c
b bd
bb b
db
d
b c
20
15
10
5
0
P B
ray
II (p
pm)
a
bc
d
c
a a a a a a
5
4
3
2
1
0
Ca
(meq
/100
g d
e su
elo)
ab
c
d
ea a
dc
a a a a
5000
4000
3000
2000
1000
0
N t
otal
(ppm
)
aa a a
cc c c c c
4
3
2
1
0
Mg
(meq
/100
g d
e su
elo)b
a
ab
d
ea aa a a a
b
c
4000
3000
2000
1000
0
P to
tal (
ppm
)
0.6
0.5
0.4
0.3
0.2
0.1
0
K(m
eq/1
00 g
de
suel
o)
ab
a
de
fg
h
a
i
a
a a a a a aa a a ab
c c
1250
1000
750
500
250
0
Suel
oM
caA
ndio
Mic
roV
eloc
iSp
ini
Ru
pti
Att
aTr
achy
Cam
poA
crom
Dim
in.
Ca
tota
l (pp
m)
aND ND
b
cd
e e e e e e
100
75
50
25
0
Suel
oM
caA
ndio
Mic
roV
eloc
iSp
ini
Ru
pti
Att
aTr
achy
Cam
poA
crom
Dim
in.
Sat
ura
ción
Al (
%) a
b
cd
e
f
gf
f
g
f
g
Figura 11-1. Propiedades químicas de las estructuras biogénicas y del suelo testigo de la sabana nativa(los códigos de las estructuras se identifican en el Cuadro 11-1, excepto la palabra Dimin.(diminutas), que se refiere a una muestra combinada de las estructuras producidas por lashormigas más pequeñas). Las letras diferentes indican diferencias significativas a un nivelP < 0.05.
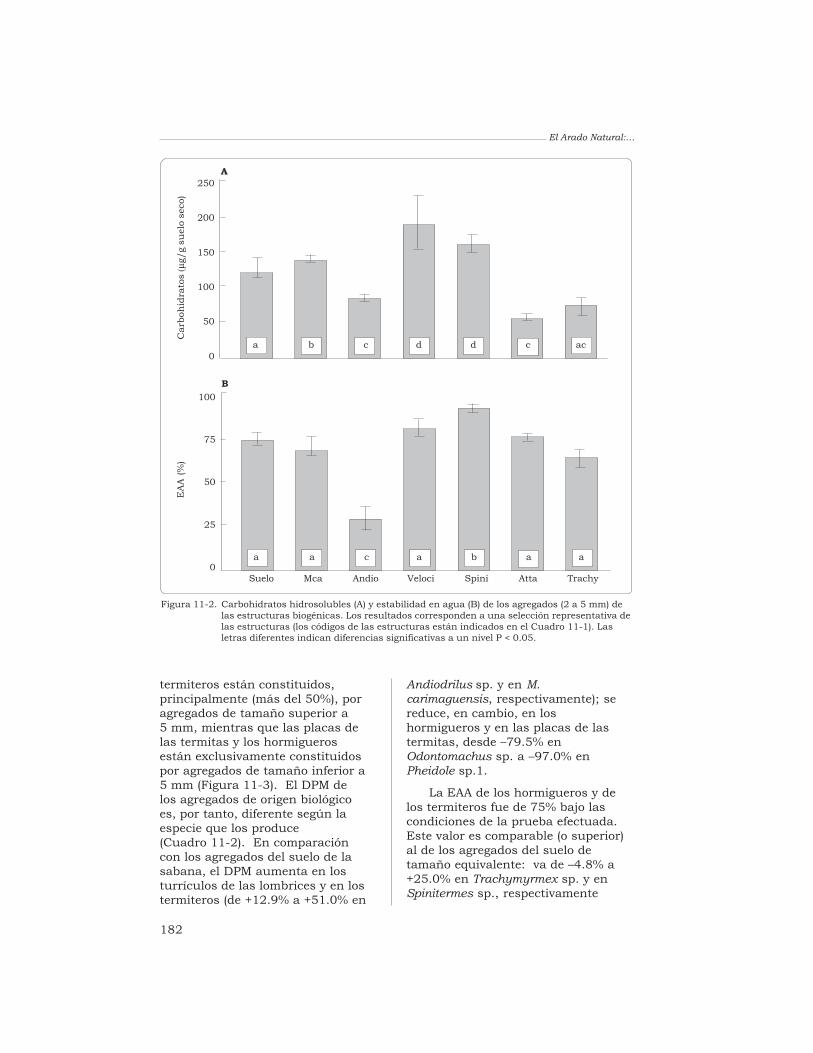
182
El Arado Natural:…
Car
boh
idra
tos
(µg/
g su
elo
seco
)250
200
150
100
50
0
A
a b c d d c ac
Suelo Mca Andio Veloci Spini Atta Trachy
100
75
50
25
0
B
a a c a b a a
EA
A (
%)
Figura 11-2. Carbohidratos hidrosolubles (A) y estabilidad en agua (B) de los agregados (2 a 5 mm) delas estructuras biogénicas. Los resultados corresponden a una selección representativa delas estructuras (los códigos de las estructuras están indicados en el Cuadro 11-1). Lasletras diferentes indican diferencias significativas a un nivel P < 0.05.
termiteros están constituidos,principalmente (más del 50%), poragregados de tamaño superior a5 mm, mientras que las placas delas termitas y los hormiguerosestán exclusivamente constituidospor agregados de tamaño inferior a5 mm (Figura 11-3). El DPM delos agregados de origen biológicoes, por tanto, diferente según laespecie que los produce(Cuadro 11-2). En comparacióncon los agregados del suelo de lasabana, el DPM aumenta en losturrículos de las lombrices y en lostermiteros (de +12.9% a +51.0% en
Andiodrilus sp. y en M.carimaguensis, respectivamente); sereduce, en cambio, en loshormigueros y en las placas de lastermitas, desde –79.5% enOdontomachus sp. a –97.0% enPheidole sp.1.
La EAA de los hormigueros y delos termiteros fue de 75% bajo lascondiciones de la prueba efectuada.Este valor es comparable (o superior)al de los agregados del suelo detamaño equivalente: va de –4.8% a+25.0% en Trachymyrmex sp. y enSpinitermes sp., respectivamente
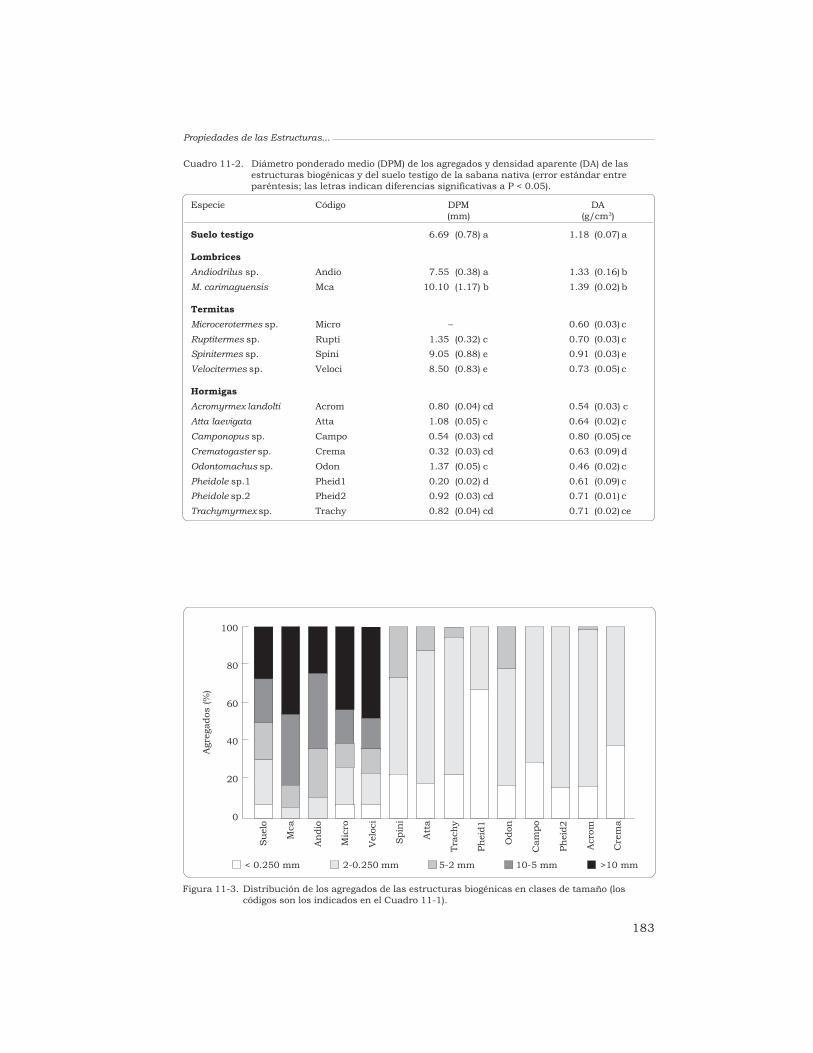
Propiedades de las Estructuras...
183
Suel
o
Mca
And
io
Mic
ro
Vel
oci
Spin
i
Att
a
Trac
hy
Phei
d1
Odo
n
Cam
po
Phei
d2
Acr
om
Cre
ma
< 0.250 mm 2-0.250 mm 5-2 mm 10-5 mm >10 mm
Figura 11-3. Distribución de los agregados de las estructuras biogénicas en clases de tamaño (loscódigos son los indicados en el Cuadro 11-1).
100
80
60
40
20
0
Agr
egad
os (%
)
Cuadro 11-2. Diámetro ponderado medio (DPM) de los agregados y densidad aparente (DA) de lasestructuras biogénicas y del suelo testigo de la sabana nativa (error estándar entreparéntesis; las letras indican diferencias significativas a P < 0.05).
Especie Código DPM DA(mm) (g/cm3)
Suelo testigo 6.69 (0.78) a 1.18 (0.07) a
LombricesAndiodrilus sp. Andio 7.55 (0.38) a 1.33 (0.16) bM. carimaguensis Mca 10.10 (1.17) b 1.39 (0.02) b
TermitasMicrocerotermes sp. Micro – 0.60 (0.03) cRuptitermes sp. Rupti 1.35 (0.32) c 0.70 (0.03) cSpinitermes sp. Spini 9.05 (0.88) e 0.91 (0.03) eVelocitermes sp. Veloci 8.50 (0.83) e 0.73 (0.05) c
HormigasAcromyrmex landolti Acrom 0.80 (0.04) cd 0.54 (0.03) cAtta laevigata Atta 1.08 (0.05) c 0.64 (0.02) cCamponopus sp. Campo 0.54 (0.03) cd 0.80 (0.05) ceCrematogaster sp. Crema 0.32 (0.03) cd 0.63 (0.09) dOdontomachus sp. Odon 1.37 (0.05) c 0.46 (0.02) cPheidole sp.1 Pheid1 0.20 (0.02) d 0.61 (0.09) cPheidole sp.2 Pheid2 0.92 (0.03) cd 0.71 (0.01) cTrachymyrmex sp. Trachy 0.82 (0.04) cd 0.71 (0.02) ce
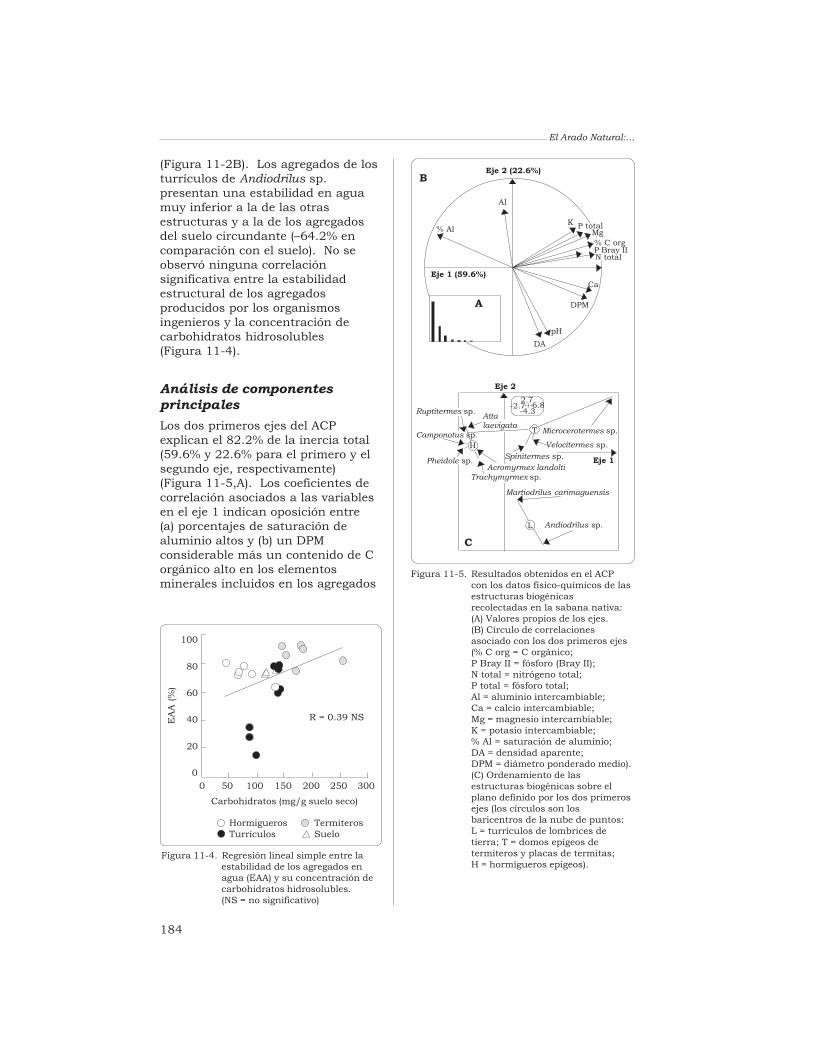
184
El Arado Natural:…
Eje 2 (22.6%)
DA
Eje 1 (59.6%)
A
B
Al
% AlK
Mg% C orgP Bray IIN total
Ca
DPM
pH
Velocitermes sp.
Microcerotermes sp.
Spinitermes sp.
Martiodrilus carimaguensis
Andiodrilus sp.
2.7-2.7
-4.36.8
Eje 2
H
T
L
Eje 1
C
Figura 11-5. Resultados obtenidos en el ACPcon los datos físico-químicos de lasestructuras biogénicasrecolectadas en la sabana nativa:(A) Valores propios de los ejes.(B) Círculo de correlacionesasociado con los dos primeros ejes(% C org = C orgánico;P Bray II = fósforo (Bray II);N total = nitrógeno total;P total = fósforo total;Al = aluminio intercambiable;Ca = calcio intercambiable;Mg = magnesio intercambiable;K = potasio intercambiable;% Al = saturación de aluminio;DA = densidad aparente;DPM = diámetro ponderado medio).(C) Ordenamiento de lasestructuras biogénicas sobre elplano definido por los dos primerosejes (los círculos son losbaricentros de la nube de puntos:L = turrículos de lombrices detierra; T = domos epígeos determiteros y placas de termitas;H = hormigueros epígeos).
P total
Attalaevigata
Acromyrmex landoltiTrachymyrmex sp.
Ruptitermes sp.
Camponotus sp.
Pheidole sp.
(Figura 11-2B). Los agregados de losturrículos de Andiodrilus sp.presentan una estabilidad en aguamuy inferior a la de las otrasestructuras y a la de los agregadosdel suelo circundante (–64.2% encomparación con el suelo). No seobservó ninguna correlaciónsignificativa entre la estabilidadestructural de los agregadosproducidos por los organismosingenieros y la concentración decarbohidratos hidrosolubles(Figura 11-4).
Análisis de componentesprincipalesLos dos primeros ejes del ACPexplican el 82.2% de la inercia total(59.6% y 22.6% para el primero y elsegundo eje, respectivamente)(Figura 11-5,A). Los coeficientes decorrelación asociados a las variablesen el eje 1 indican oposición entre(a) porcentajes de saturación dealuminio altos y (b) un DPMconsiderable más un contenido de Corgánico alto en los elementosminerales incluidos en los agregados
Figura 11-4. Regresión lineal simple entre laestabilidad de los agregados enagua (EAA) y su concentración decarbohidratos hidrosolubles.(NS = no significativo)
EA
A (
%)
0 50 100 150 200 250 300
Carbohidratos (mg/g suelo seco)
100
80
60
40
20
0
R = 0.39 NS
Hormigueros TermiterosTurrículos Suelo

Propiedades de las Estructuras...
185
(Figura 11-5,B). La representaciónde los objetos sobre el eje 1 muestrauna notoria oposición en los casossigientes: (a) entre los hormigueros ylas placas de las termitas; (b) entrelos termiteros y los turrículos de laslombrices (Figura 11-5,C). El eje 2 sedefine por las siguientes oposiciones:(a) de un lado, la saturación de Al;(b) del otro, la DA y el pH de lasestructuras (Figura 11-5,B). Tambiénen este eje hay oposición entre lostermiteros y los hormigueros, de unlado, y los turrículos compactos delas lombrices de tierra, del otro(Figura 11-5,C).
Discusión
Contenido de materia orgánicay de elementos mineralesLa fuerte concentración de C orgánicoy de elementos minerales que seregistró en las heces de las lombricesde tierra de Carimagua es común ennumerosas especies de oligoquetosque producen turrículos sobre lasuperficie del suelo (Lee 1985). Esteresultado, corroborado porobservaciones hechas en las heces deM. carimaguensis (Rangel et al. 1999),se explica por los hábitos alimenticiosde las especies estudiadas. SegúnJiménez et al. (1998), Andiodrilus sp.y M. carimaguensis pertenecen,respectivamente, a las siguientescategorías ecológicas: endógeapolihúmica (sensu Lavelle 1979) yanécica (sensu Bouché 1977),respectivamente, en las cuales laslombrices tienen la particularidad deingerir, de modo selectivo, unsustrato más rico en materiaorgánica (MO) que el del suelocircundante (suelo orgánico omaterial vegetal fresco o ambos).
Las termitas construyen domosepígeos utilizando, generalmente,
suelo que cementan con cantidadesvariables de secreciones salivales yexcrementos ricos en MO (Kooyman yOnck 1987; Lee y Foster 1991; Lee yWood 1971; Wood y Sands 1978). Enciertos casos, una parte considerablede las paredes del termitero estáconstituida por un materialacartonado muy rico en C orgánico (elde Microcerotermes sp., por ejemplo).La abundancia de MO en las hecesexplica las diferencias en laconcentración de C y de elementosminerales observadas entre lostermiteros y el suelo (Black y Okwakol1997; Lee y Wood 1971; Lobry-de-Bruyn y Conacher 1990; Wood ySands 1978). Las paredes o tabiquesde las galerías aéreas de Ruptitermessp. recubren los accesos hacia lasfuentes de alimentación; aquéllasdesempeñan, probablemente, unpapel protector frente al calor, a losdepredadores y a los competidores(Kooyman y Onck 1987). A diferenciade lo que figura generalmente en laliteratura (Lee y Wood 1971; Lobry-de-Bruyn y Conacher 1995), lasparedes de las muestras tomadas eneste estudio no están enriquecidascon MO ni con bases intercambiables.
Los domos epígeos producidos porlas hormigas, a diferencia de lasestructuras producidas por lastermitas y por las lombrices de tierraque se consideraron en este estudio,eran todos de color amarillo claro. Eséste un suelo que proviene de loshorizontes inferiores del perfil y estransportado a la superficie, donde seacumula poco a poco; representa asíuna medida de la excavación de losespacios subterráneos del nido (Elmes1991; Petal 1978). Este suelo espobre y no experimenta ningunamodificación importante al sertrasladado; tampoco está enriquecidocon MO, dado su bajo contenido de Cy de elementos minerales.

186
El Arado Natural:…
Tamaño y estabilidad de losagregadosLos turrículos de M. carimaguensis yde Andiodrilus sp. son, comparadoscon el suelo circundante, estructurascompactas constituidas por agregadosde gran tamaño, que corresponden altipo globular descrito por Lee (1985).Estos agregados tienen unaestabilidad equivalente o inferior a lade los agregados del suelo, unresultado sorprendente porque losturrículos de las lombrices de tierrason, generalmente, más estables quelos agregados de suelo de tamañocomparable (Lee 1985). Un estudioreciente demostró la gran estabilidadestructural de los turrículos de M.carimaguensis (Guggenberger et al.1996). Este resultado está, quizás,relacionado con el empleo, en esteestudio, de turrículos que habían sidodepositados recientemente. Enrealidad, las deposiciones de laslombrices de tierra adquierenestabilidad estructural en el curso desu envejecimiento (Blanchart et al.1993; Hindell et al. 1997a, 1997b;Marinissen 1990), probablemente porla acción conjunta de los siguientesprocesos:
- procesos físicos, como elendurecimiento de las heces auna humedad constante o bajo elefecto de la alternancia de faseshúmedas y secas (Hindell et al.1997b; Marinissen 1990);
- procesos biológicos, como eldesarrollo de hongos sobre lasuperficie de los agregados o laproducción de polisacáridos deorigen microbiano (Marinissen1990; Shipitalo y Protz 1989;Tisdall 1994).
Las bolas fecales depositadas porlas termitas están constituidas poragregados órgano-minerales detamaño inferior a los que producenlas lombrices de tierra (Garnier-Sillam
1991). Usan estas bolas, junto consus secreciones salivales, paracementar las partículas del suelo enla edificación de las paredes deltermitero. Durante este proceso, laporosidad del suelo disminuye y elmaterial reorganizado será muchomás compacto que el suelo original(Lal 1988; Lobry-de-Bruyn yConacher 1990). Ahora bien, en elpresente trabajo se halló que lasnumerosas galerías de los termiteroslos convertían en estructuras muycavernosas, lo que explica la baja DAobservada en ellos. Es interesanteconstatar que las paredes de lostermiteros, después de recibir ungolpe, se fragmentan enmacroagregados de tamañoimportante. Se puede imaginar que elmismo fenómeno ocurrenaturalmente, es decir, que elimpacto de las gotas de lluviaconduce a una lenta redistribución delos macroagregados en la superficiedel suelo. Estos agregados son muyestables y permiten pensar que losefectos que tales procesos ejercen enla estabilidad del suelo sonsignificativos a una escala espacio-temporal importante. La estabilidadde los agregados producidos por lastermitas ha sido ya señalada en otrosestudios, aunque los procesos queella implica no han sido identificadoscon precisión (Garnier-Sillam 1991;Lal 1988; Lobry-de-Bruyn y Conacher1990).
El estudio de la estructura de losturrículos de las lombrices y de lastermitas se ha intensificado a lo largode los últimos 10 años; no obstante,la estabilidad de los agregadosproducidos por la actividad de lashormigas apenas ha atraído laatención de la mayoría de los biólogos(Folgarait 1998; Lobry-de-Bruyn yConacher 1990). Los hormigueros dela sabana de Carimagua estánconstituidos por agregados libres, queson transportados por los insectos a

Propiedades de las Estructuras...
187
la superficie desde los horizontesprofundos del suelo y apilados,simplemente, los unos sobre losotros. El tamaño medio de losagregados es proporcional al tamañode las mandíbulas de las hormigasque los transportan (Weber 1979) ysu estabilidad estructural escomparable a la de los agregados detamaño similar del suelo superficial.Los agregados no están cementadospor ningún pegamento orgánico, queles daría una estructura de tipogranular, ni están unidos entre sí;por ello, la presencia de losnumerosos espacios que los separanexplica la baja DA de las estructurasformadas.
Los carbohidratos hidrosolublesse consideran componentesimportantes en los mecanismos deestabilización de los agregados delsuelo en las sabanas colombianas(Gijsman y Thomas 1996). Entre loscarbohidratos implicados en laagregación del suelo están los querepresentan una fracción específica eimportante que se deriva,principalmente, del metabolismomicrobiano y, quizás, de algunostejidos vegetales (Haynes y Swift1990; Tisdall 1991, 1994). Sinembargo, los resultados de esteestudio no muestran ningunacorrelación entre la estabilidadestructural de los agregadosbiogénicos y la presencia de talescompuestos. Este resultado indicaque no son los carbohidratos los queintervienen en la estabilidad de losagregados (Gijsman y Thomas 1996)sino que hay otros mecanismosimplicados, algunos de carácterfísico. Por ejemplo, elendurecimiento de las estructuras acausa de la variación en la humedadpodría desempeñar un papelimportante en la superficie del suelo(Hindell et al. 1997b; Marinissen1990).
Significación funcional de lasestructurasLa comparación de las característicasfísicoquímicas de las estructurasbiogénicas enumeradas en esteestudio con las características delsuelo circundante permiteclasificarlas en tres gruposprincipales:
1. Estructuras más compactas queel suelo, enriquecidas con Corgánico y elementos minerales, yconstituidas por agregados degran tamaño. Corresponden, eneste estudio, a los turrículos detipo globular construidos por lasdos especies de lombrices detierra estudiadas.
2. Estructuras menos compactasque el suelo, enriquecidas con Corgánico y elementos minerales, yconstituidas por agregados degran tamaño. Corresponden, eneste estudio, a los domos epígeosde los termiteros.
3. Estructuras de mínimacompactación (construidas consuelo de las capas másprofundas), de tipo granular, cuyocontenido de C orgánico esinferior al del suelo superficial; suconcentración de elementosminerales es, en general, inferioro equivalente a la de dicho suelo,y están constituidas poragregados muy estables detamaño pequeño. Estánrepresentadas aquí por loshormigueros y las placas epígeasde Ruptitermes sp.
A partir de esta tipología deestructuras biogénicas, es posiblededucir el impacto que causan en elsuelo las especies que las producen.En efecto, la naturaleza de dichasestructuras puede considerarse comoel reflejo de ciertos atributosfuncionales de las especies que lasconstruyen (Lavelle 1996). Así, las

188
El Arado Natural:…
especies que producen lasestructuras de tipos 1 y 2 (termitas ylombrices de tierra) acumulan Corgánico en la superficie del suelo einfluyen, probablemente, en ladinámica de la MO y en la tasa deliberación de los elementos mineralesasimilables por las plantas (Black yOkwakol 1997; Lavelle et al. 1998;Lobry-de-Bruyn y Conacher 1990).
Las estructuras pertenecientes aestos dos primeros grupos secaracterizan, además, por su grantamaño y por la clase de agregadosque las constituyen, rasgos quecontrastan con el tamaño muyreducido de las estructuras de tipo 3(placas de termitas y hormigueros).La producción de agregados decaracterísticas físicoquímicasdiversas podría contribuir a unaregulación eficiente de la estructuradel suelo. Este mecanismo ha sidoya descrito, por ejemplo, en lassabanas de Costa de Marfil, dondelas especies más pequeñas delombrices fragmentan los turrículosproducidos por las especies de mayortamaño, evitando así su acumulaciónexcesiva sobre la superficie del suelo(Blanchart et al. 1997; Rossi 1998).En Carimagua existe una regulacióndel mismo tipo: las termitas(Nasutermitinae) aceleranvisiblemente la cinética dedegradación de los grandes turrículosproducidos por M. carimaguensis(Decaëns 2000). Estudios recientesllevados a cabo en los pastizales de lacuenca amazónica han mostrado queuna población abundante delombrices de tierra puede conducir auna compactación considerable delsuelo, ya que éstas no están allíasociadas con alguna especie capazde fragmentar los turrículos hastareducirlos a agregados de menortamaño (Chauvel et al. 1999).
Las estructuras biogénicassuperficiales influyen,probablemente, en la estructura del
suelo desde el momento en que sedisgregan para que estos fragmentosse dispersen sobre el suelo, al cual seincorporarán progresivamente. Esprobable que los agregados sueltosque se observaron, en este estudio,sobre la superficie del suelocorrespondan, esencialmente, afragmentos de estructuras biogénicasque estarían en el proceso deincorporación al suelo. La hipótesisse apoya en una observación sencilla:la forma de estos agregados es, en sumayoría, la de turrículos pequeños deAndiodrilus sp. o de fragmentosreconocibles de turrículos de M.carimaguensis (Foto 11-7). Es fácilverificar en el terreno que la mayorparte de las estructuras biogénicas sedisocian, efectivamente, en agregadoslibres, y que éstos quedan dispersossobre el suelo.
ConclusionesLos resultados de este estudio sonuna prueba clara de la diversidad deestructuras producidas por losingenieros ecológicos sobre lasuperficie del suelo de una sabananatural. Se identificaron tres tipos deestructuras partiendo de suspropiedades físicoquímicas:
- las estructuras compactas, ricasen MO (turrículos de lombrices);
- las estructuras poco compactas,ricas en MO (termiteros); y
- las estructuras sin compactación,granulares y pobres en MO (placasde termitas y hormigueros).
El análisis multivariado reflejó lagran diversidad de estructurasproducidas sobre la superficie delsuelo y dejó entrever tanto laposibilidad como la complejidad deuna clasificación funcional de losorganismos ingenieros, la cual tendríaen cuenta, a un mismo tiempo, las

Propiedades de las Estructuras...
189
diferentes funciones manifestadas porestas estructuras.
Este estudio se ha limitado adescribir las estructuras superficialesproducidas por los ingenierosecológicos. La tipología en élpresentada no ha considerado, enrealidad, un gran número deestructuras producidas en el interiordel suelo. Son numerosos losinvertebrados del suelo capaces deproducir estructuras endógeas (nidos,galerías, agregados) que puedeninfluir de manera específica sobre losprocesos del suelo. Por ejemplo, enCarimagua existe una especie delombriz endógea polihúmica(Ocnerodrilidae sp.) que prefiereingerir los turrículos compactos de M.carimaguensis y produce turrículosgranulares de tamaño pequeño(Jiménez et al. 1998). Otra especie detamaño mediano, Glossodrilus sp.,presenta a veces un comportamientomuy similar. Estas especiespertenecen al grupo de ingeniero que
produce estructuras granulares muypoco compactas. Ahora bien, dadoque los turrículos de estas lombricesde tierra pueden ser más ricos en MOque el material empleado en loshormigueros y en las placas de lastermitas, hay que establecer unanueva categoría constituida poringenieros que producen estructuraspoco compactas, granulares y ricasen MO.
La definición de los gruposfuncionales de los ingenierosecológicos del suelo se apoya en latipología de las estructuras que ellosproducen. Es necesario describircompletamente la totalidad de lasestructuras biogénicas producidaspor estos organismos —por ejemplo,agregados constituidos tanto en lasuperficie del suelo como a través desu perfil, galerías, nidos endógeos—descripción que permitirá definir conprecisión las funciones de losingenieros. Contribuirá también esadescripción a establecer con exactitud
Foto 11-7. Superficie del suelo de la sabana nativa cubierta por agregados libres provenientes de lasestructuras biogénicas (escala: longitud de la foto = 40 cm).

190
El Arado Natural:…
aquellas características de lasestructuras que podrían emplearsepara definir los atributos funcionalesde una especie determinada deingenieros. Por ejemplo: ¿lasespecies que producen estructurascompactas ejercen, sistemáticamente,un efecto ‘compactador’ en el suelo(sensu Blanchart et al. 1997)? Por suparte, ¿aquellas especies queproducen estructuras pococompactas, causan un efecto‘descompactador’ en el suelo? Lasespecies que concentran MO en susestructuras ¿ejercen un efecto en ladinámica de la MO? Si lo hacen,¿cuál sería este efecto? Finalmente,¿qué impactos causan estasestructuras en las condiciones de vidade otros organismos del suelo?
AgradecimientosLos autores agradecen a R.J. Thomas,D.K. Friesen y C.G. Meléndez (CIAT)por el apoyo logístico recibido.A M.P. Hernández (CIAT), P. Eggletony J.J. Jiménez (CIAT) por laidentificación de las hormigas, lastermitas y las lombrices de tierra.A N. Asakawa (CIAT) por los análisisquímicos. A A.V. Spain (CSIRO),E. Blanchart (IRD) y P. Folgarait(Universidad Nacional de Quilmes)por los comentarios emitidos acercade una primera versión delmanuscrito de este trabajo.A I. Poudevigne y H. Decaëns(Université de Rouen) por la críticaconstructiva del texto final.
ReferenciasAnderson JM; Ingram J, eds. 1993.
Tropical soil biology and fertility. Ahandbook of methods. CABInternational, Oxford.
Bal L. 1973. Micromorphological analysisof soils. Lower organization oforganic soil materials. NetherlandsSoil Survey Institute, Wageningen,Holanda.
Black HIJ; Okwakol MJN. 1997.Agricultural intensification, soilbiodiversity and agroecosystemfunction in the tropics: the role oftermites. Appl Soil Ecol 6:37-53.
Blanchart E. 1990. Rôle fonctionnel desvers de terre dans la formation de lastructure des sols de la savane deLamto (Côte d’Ivoire). Tesis (Ph.D.).Université de Rennes I, Francia.
Blanchart E; Bruand A; Lavelle P. 1993.The physical structure of casts ofMillsonia anomala (Oligochaeta:Megascolecidae) in shrub savannasoils (Côte d’Ivoire). Geoderma56:119-132.
Blanchart E; Lavelle P; Braudeau E;Bissonnais YL; Valentin C. 1997.Regulation of soil structure bygeophagous earthworm activities inhumid savannas of Côte d’Ivoire.Soil Biol Biochem 29:431-439.
Bouché MB. 1977. Stratégieslombriciennes. En: Lohm U; PerssonT, eds. Soil organisms ascomponent of ecosystems. Ecol Bull25:122-132.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm.Nature 398:32-33.
Cheschire MV. 1979. Nature and originsof carbohydrates in soils. AcademicPress, Londres.
Decaëns T. 2000. Degradation dynamicsof surface earthworm casts ingrasslands of the eastern plains ofColombia. Biol Fertil Soils32:149-156.
Elmes GW. 1991. Ant colonies andenvironmental disturbance. SympZool Soc Lond 63:15-32.

Propiedades de las Estructuras...
191
Folgarait P. 1998. Ant biodiversity and itsrelationships to ecosystemfunctioning: a review. BiodivConserv 7:1121-1244.
Garnier-Sillam E. 1991. The pedologicalrole of fungus-growing termites(Termitidae: Macrotermitidae) intropical environments, with specialreference to Macrotermes muelleri.Sociobiology 15:181-196.
Gijsman AJ. 1996. Soil aggregatestability and soil organic matterfractions under agropastoralsystems established in nativesavanna. Aust J Soil Res34:891-907.
Gijsman AJ; Thomas RJ. 1996. Aggregatesize distribution and stability of anOxisol under legume-based andpure grass pasture in the easternColombian savannas. Aust J SoilRes 33:153-165.
Guggenberger G; Thomas RJ; Zech W.Soil organic matter withinearthworm casts of an anecic-endogeic tropical pasturecommunity, Colombia. Appl SoilEcol 3:263-274.
Haynes RJ; Swift RS. 1990. Stability ofsoil aggregates in relation toorganic constituents and soil watercontent. J Soil Sci 41:73-83.
Hindell RP; McKenzie BM; Tisdall JM.1997a. Destabilization of soilduring the production of earthworm(Lumbricidae) and artificial casts.Biol Fertil Soil 24:153-163.
Hindell RP; McKenzie BM; Tisdall JM.1997b. Influence of drying andageing on the stabilization ofearthworm (Lumbricidae) casts.Biol Fertil Soil 25:27-35.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78:1946-1957.
Kemper WD; Rousenau RC. 1986.Aggregate stability and sizedistribution. En: Klute A, ed.Methods of soil analysis. AmericanSociety of Agronomy (ASA) and SoilScience Society of America (SSSA),Madison, WI, Estados Unidos.p 425-442.
Kooyman C; Onck RFM. 1987. Theinteractions between termiteactivity, agricultural practices andsoil characteristics in Kisii District,Kenya. Wageningen AgriculturalUniversity, Holanda.
Lal R. 1988. Effects of macrofauna on soilproperties in tropical systems. AgricEcosyst Environ 24:101-116.
Lavelle P. 1979. Relations entre typesécologiques et profilsdémographiques chez les vers deterre de la savane de Lamto (Côted’Ivoire). Rev Ecol Biol Sol16:85-101.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Intern33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P; Pashanasi B; Charpentier F;Gilot C; Rossi J-P; Derouard L;Andre J; Ponge JP; Bernier N. 1998.Large-scale effects of earthworms onsoil organic matter and nutrientdynamics. En: Edwards CA, ed.Earthworm ecology. St. LuciesPress, Boca Ratón, FL, EstadosUnidos. p 103-122.
Lee KE. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, Orlando,FL, Estados Unidos.

192
El Arado Natural:…
Lee KE; Foster RC. 1991. Soil fauna andsoil structure. Aust J Soil Res29:745-775.
Lee KE; Wood TG. Physical and chemicaleffects on soils of some Australiantermites, and their pedologicalsignificance. Pedobiologia11:376-409.
Legendre P; Vaudor A. 1991. The Rpackage: multidimentionalanalysis, spatial analysis.Université de Montréal, Canadá.
Lilliefors HW. 1967. The Kolmogorov-Smirnov test for normality withmean and variance unknown. J AmStat Assoc 62:399-402.
Lobry-de-Bruyn LA; Conacher AJ. 1990.The role of termites and ants in soilmodification: a review. Aust J SoilRes 28:55-93.
Lobry-de-Bruyn LA; Conacher AJ. 1995.Soil modification by termites in theCentral Wheatbelt of WesternAustralia. Aust J Soil Res33:179-193.
Marinissen JCY. 1990. Mechanisms ofstabilization of casts and artificialcasts. Biol Fertil Soils 9:163-167.
Petal J. 1978. The role of ants inecosystems. En: Brian MV, ed.Production ecology of ants andtermites. Cambridge UniversityPress, Reino Unido. p 293-325.
Rangel AF; Thomas RJ; Jiménez JJ;Decaëns T. 1999. Nitrogendynamics associated withearthworm casts of Martiodriluscarimaguensis Jiménez and Morenoin a Colombian savanna Oxisol.Pedobiologia 43:557-560.
Rossi J-P. 1998. Rôle fonctionnel de ladistribution spatiale des vers deterre dans une savane humide deCôte d’Ivoire. Tesis (Ph.D.).Université Pierre et Marie Curie-Paris VI, Francia.
Shipitalo MJ; Protz R. 1989. Chemistryand micromorphology ofaggregation in earthworm casts.Geoderma 45:357-374.
Thioulouse J; Chessel D; Dolédec S;Olivier JM. ADE-4: a multivariateanalysis and graphical displaysoftware. Stat Comput 7:5-83.
Tisdall JM. 1991. Fungal hyphae andstructural stability of soil. Aust JSoil Res 29:729-743.
Tisdall JM. 1994. Possible role of soilmicroorganisms in aggregation insoils. Plant Soil 159:115-121.
Weber NA. 1972. Gardening ants, theAttines. Mem Am Phil Soc 92:1-146.
Wood TG; Sands WA. 1978. The role oftermites in ecosystems. En: BrianMV, ed. Production ecology of antsand termites. Cambridge UniversityPress, Reino Unido. p 245-292.

Propiedades de las Estructuras...
193
V = π R2l
Cono: Cono truncado: Esfera:
Placa: Paraboloide: Cilindro:
h
rr
lh
r
h
r
h
r
R
R
Apéndice 1. Formas geométricas empleadas en laevaluación del volumen de las estructurasbiogénicas halladas en la superficie delsuelo.
H
V = π r2hV = π r2h2
V = π r2h3 V = 4 πR3 3V = π H (R2 + r2 + Rr)3

194
El Arado Natural:…
Apéndice 2. Otras estructuras biogénicas encontradasen la superficie de los suelos de la sabanade Carimagua.
Foto 11-8A. Montículo epígeo de una colonia de hormigas Atta laevigata en la sabana nativa (escala:longitud de la foto = 3 m).
Foto 11-9A. Montículo epígeo y entrada de una colonia de hormigas Pheidole sp.1 en la sabana nativa(escala: longitud de la foto = 10 cm).
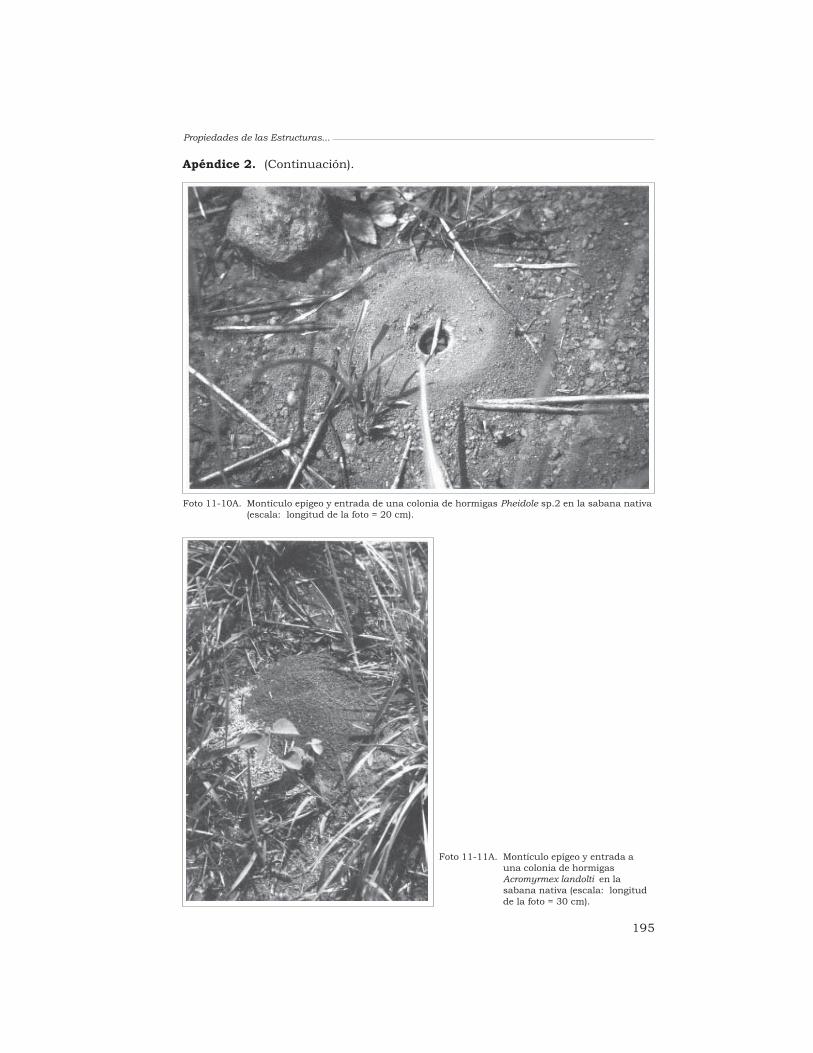
Propiedades de las Estructuras...
195
Foto 11-10A. Montículo epígeo y entrada de una colonia de hormigas Pheidole sp.2 en la sabana nativa(escala: longitud de la foto = 20 cm).
Foto 11-11A. Montículo epígeo y entrada auna colonia de hormigasAcromyrmex landolti en lasabana nativa (escala: longitudde la foto = 30 cm).
Apéndice 2. (Continuación).

196
El Arado Natural:…
Foto 11-12A. Montículo epígeo y entrada de una colonia de Crematogaster sp. en la sabana nativa(escala: longitud de la foto = 30 cm).
Foto 11-13A. Montículo epígeo de una colonia de termitas Microcerotermes sp. en la sabana nativa(escala: longitud de la foto = 30 cm).
Apéndice 2. (Continuación).

Propiedades de las Estructuras...
197
Foto 11-14A. Desagregación y fragmentación total de un montículo epígeo de una colonia de termitasSpinitermes sp. y dispersión de los agregados libres sobre la superficie del suelo de lasabana nativa (escala: longitud de la foto = 50 cm).
Foto 11-15A. Turrículos de M. carimaguensis parcialmente fragmentados en agregados libres que seesparcen sobre la superficie del suelo de la sabana nativa (escala: longitud de lafoto = 30 cm).
Apéndice 2. (Continuación).

198
El Arado Natural:…
ResumenLa definición de ingenieros delecosistema se aplica plenamente alas lombrices de tierra. Las hecesque éstas producen, por ejemplo,son estructuras de gran importanciaen la regulación de algunos procesosdel suelo. No obstante, la vidamedia y la tasa de degradación deestas estructuras se han estudiadomuy poco. En este trabajo sedeterminó, en una parcela de sabananativa y en un pastizal intensivo, ladinámica de la desaparición de lasheces depositadas en la superficiedel suelo por la especie de lombrizanécica Martiodrilus carimaguensis ylos cambios producidos en laspropiedades físicas de dichas heces.En ambos sistemas de uso de latierra, las heces estaban formadaspor una superposición de capas quelas lombrices depositaban durantevarios días. La vida media de lasheces varió de 2 a 11 meses en unpastizal bajo pastoreo y en un
pastizal protegido, respectivamente,y fue de 5 meses en la sabana sinpastoreo. Las heces secaspermanecieron durante más de unaño sobre la superficie del suelo.
La desaparición de las heces sedebió, principalmente, a los impactosproducidos por las gotas de lluvia yal efecto del pisoteo del ganado. Ladensidad aparente (DA) de las hecesfrescas fue mayor (+17%, P < 0.05)que la del suelo circundante en lasabana o equivalente a la del suelo(-1%, P > 0.05) en el pastizal. Losagregados de las heces frescas erande tamaño mayor que los del sueloen ambos sistemas de uso de latierra (+70%, P < 0.05). La DA y eltamaño de los agregadosdisminuyeron con el envejecimientode las heces, y son –29% y –24%para la primera y –68% y –80% parael segundo, en el pastizal y en lasabana, respectivamente. Se observótambién que algunosmacroinvertebrados horadaban lasheces y eran parcialmenteresponsables de la degradación físicade éstas a lo largo del tiempo.
Palabras clave: heces superficialesde lombrices, ingenieros delecosistema, Martiodriluscarimaguensis, propiedadesfísicas, sabana
CAPÍTULO 12
Dinámica de la Degradación de lasHeces Superficiales de la Lombriz deTierra en Areas de Sabana y dePastizal en los Llanos Orientales deColombia1
T. Decaëns*
1. Publicado en Biology and Fertility of Soils32(2), p 149-156, por Decaëns. Copyright ©2000 de Springer. Impreso con permiso de lacasa editorial.
* Laboratoire d’Écologie des Sols Tropicaux(IRD)/Unidad Suelos y Nutrición de Plantas(CIAT), IRD-Bondy, 32 Av. Henri Varagnat,F-93143 Bondy Cedex, Francia.

Dinámica de la Degradación...
199
IntroducciónEn numerosos estudios se handescrito las propiedades físicas delas heces de las lombrices de tierra.El tamaño y la forma de talesestructuras están determinadas, engran parte, por el tamaño y loshábitos alimenticios de las especiesque las producen, pero también loestán por la textura del sueloingerido (ver la revisión de Barois etal. 1999 y de Lee 1985). Se handescrito dos grandes grupos deheces: las globulares, es decir, lasque están formadas por agregadoscompactos y estables de grantamaño (van de unos milímetroshasta unos poco centímetros), y lasgranulares, o sea, las que estánformadas por agregados de tamañopequeño (menores que 1 mm) yligeramente estables (Lee 1985). Laproducción combinada de estos dostipos de heces por una comunidadcompuesta por diversas lombrices detierra regula la estructura del suelode manera favorable, ya que lasespecies de menor tamaño soncapaces de romper los agregadosmás compactos producidos por lasespecies más grandes (Blanchart etal. 1997; Rossi 1998).
Las lombrices de tierra seconsideran uno de los grupos másimportantes entre los ingenierosecológicos del suelo (sensu Jones etal. 1994), es decir, de losinvertebrados del suelo que regulanvarios procesos del suelo mediante laproducción de estructuras físicas(canales, agregados, montículos, etc.)(Anderson 1995; Lavelle 1996, 1997).Aunque se conoce bien laimportancia, en términosfuncionales, de las heces de laslombrices en la regulación de dichosprocesos, la vida media y la dinámicade la degradación de estasestructuras son aún bastantedesconocidas. Por consiguiente,
tiene mucha importancia lainvestigación de estos dos aspectospara comprender el impacto de losingenieros del ecosistema en elambiente (Jones et al. 1997).
El objetivo de este estudio fuedescribir la dinámica de ladegradación de las hecessuperficiales producidas porindividuos de la especie anécicaMartiodrilus carimaguensis Jiménez yMoreno en la sabana nativa y en unpastizal intensivo de los LlanosOrientales de Colombia. Se midió laDA y el tamaño de los agregadosbiogénicos con el fin de evaluar elimpacto de dicha dinámica en laestructura del suelo. Losmacroorganismos, o sea, losmacroinvertebrados y las raicillas, semuestrearon dentro de las hecespara establecer el impacto deaquéllos en la degradación de éstas.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua (4°37’ N,71°19’ O) de CIAT-CORPOICA, quepertenece a la unidad fitogeográficade las sabanas isohipertérmicas biendrenadas de los Llanos Orientales deColombia. El clima es de tiposubhúmedo tropical; en él, latemperatura registra un valor medioanual de 26 °C y la precipitación de2300 mm. Hay además una épocaseca muy definida que dura desdenoviembre-diciembre hasta marzo decada año (datos del CIAT). En laFigura 12-1 se presenta laacumulación de la precipitación porperíodos de 15 días. La vegetacióndel sitio de estudio varía con latopografía: hay sabanas abiertas enlas áreas altas (o ‘altos’) y bosques degalería o sabanas inundables en lasáreas bajas (o ‘bajos’). Los suelos

200
El Arado Natural:…
son Oxisoles (Tropeptic Haplustoxisohipertérmicos) en los altos yUltisoles (Ultic Aeric Plintaquox) enlos bajos. Ambos tipos de suelospresentan propiedades físicasfavorables (como agregación,porosidad, retención de humedad),acidez alta [pH (H2O) < 5], y unafertilidad química muy baja(saturación de Al > 80%, CIC< 5 meq/100 g).
Parcelas experimentalesEl experimento se realizó en dossistemas diferentes de uso de latierra ubicados en un Oxisol biendrenado. Las parcelasexperimentales de ambos sistemas,que distaban entre sí 60 m, son lassiguientes:
- Una parcela de sabana nativacubierta de Trachypogon vestitusAnders, protegida de la quema ydel pastoreo durante 4 años, yexplotada a la manera tradicionalque incluye una quema anualdurante la época seca. Estaparcela fue quemada en febrerode 1996 y después quedó
protegida de la quema durante elexperimento.
- Un pastizal de 3 años en que sesembraron Brachiaria humidicola(Rendle), Arachis pintoi Krap. yGreg., Stylosanthes capitata Vog. yCentrosema acutifolium Benth., yque fue pastoreado por ganado deraza cebú, con una carga animalde 2.0 animales/ha.
Diseño experimentalDe las ocho especies de lombrices detierra presentes en la sabana nativadel área de estudio (Jiménez et al.1998a) se escogió M. carimaguensis,porque es la única que deposita unacantidad grande de heces en lasuperficie del suelo. Las hecesproducidas por esta especie sonestructuras grandes (hasta de 15 cmde altura), con forma de torre, cuyaproducción dura, generalmente,varios días (los turrículos). Estaespecie, además, se adapta muy biena algunos agroecosistemas (pastizalesintensivos, en especial) y puedeconsiderarse como un agenteimportante en el manejo biológico dela fertilidad y de la estructura delsuelo (Jiménez et al. 1998b).
El experimento comenzó en mayode 1996, al inicio de la época delluvias, durante el pico de máximaactividad de las lombrices (Jiménez etal. 1998a). Se consideraron dossituaciones diferentes en el pastizal:(a) heces expuestas al pisoteo delganado y (b) heces protegidas delpisoteo de los animales mediantejaulas metálicas. En ambos sistemas(sabana y pastizal) y en cada una delas situaciones (a y b) se identificaronindividualmente con pequeñas placasmetálicas 300 heces deM. carimaguensis, las cuales sedistribuyeron en cuatro grupos de75 heces cada uno, relativamentecercanos uno de otro (a unos 30 cmen el pastizal y a 2 m en la sabana).
Figura 12-1. Precipitación acumulada enperíodos de 15 días (en cadaintervalo de 30) durante el tiempodel estudio.
0 30 60 90 120 150180210 240 270 300 330
250
200
150
100
50
0Prec
ipit
ació
n a
cum
ula
da (m
m)
Tiempo desde el comienzode la deposición (días)
Primeraquincena
Segundaquincena

Dinámica de la Degradación...
201
Sólo se escogieron heces frescaspequeñas, es decir, las que sehallaban al inicio del proceso dedeposición. Este procedimientopermitió asegurar la presencia de laslombrices en las galerías junto a lasheces marcadas y evitar, además,cualquier efecto acumulativo en lasheces como resultado de la presenciaprolongada de las lombrices antesdel inicio del experimento.
Clasificación de las hecesLas heces se observaron a los 0, 1, 2,3, 4, 7, 14, 21, 30, 45, 60, 105, 150,195, 240 y 330 días después delinicio de la deposición. En cadafecha y en cada situación (sabana opastizal, protegido o expuesto alpisoteo del ganado), seinspeccionaron todas las heces y seasignaron a uno de los cuatro grupossiguientes, según su aspecto físico:(a) heces frescas (en estado dedeposición), (b) heces secas eintactas, (c) heces secas yfracturadas, (d) heces destruidas odesaparecidas (o de ambos aspectos).
Propiedades físicas de lashecesSe establecieron las propiedadesfísicas de las heces halladas en lasabana nativa y en el pastizal,ambos protegidos del pisoteo delganado. Se tomaron muestras de lasheces a los 0, 1, 2, 7 y 14 días yluego a los 1, 2, 3.5, 5, 6.5, 8 y11 meses después de la deposición.En cada fecha se recolectaron dosheces al azar de cada grupo de 75, esdecir, 8 heces por fecha.
La DA de las heces se determinóen fragmentos de unos 10 g, de loscuales se tomaron, en cada fecha,una media de 12 fragmentos porcada grupo de heces; la densidad secalculó por el método deldesplazamiento de agua (las
muestras fueron selladaspreviamente con Parafilm).
La distribución de los agregadosen clases según su tamaño se evaluópor el método estándar del tamizadoen húmedo (Blanchart 1990; Kempery Rousenau 1986). Las heces sesecaron al aire durante 4 días y sedejaron caer sobre una superficiedura para disociar los agregados.Estos fueron luego tamizados en unacolumna de tamices de 0.053, 0.125,0.250, 0.5, 1, 2, 5 y 10 mm de luz enel entramado. Cada fracción se secóen el horno a 75 °C durante 48 horasy después se pesó. La distribuciónde los agregados en clases según sutamaño se relacionó con su diámetroponderal medio (DPM), comosugieren Kemper y Rousenau (1986):
y la fracción de agregados:
donde es el diámetro medio de losagregados de cada fracción; Mtamiz esel peso seco de las partículasretenidas en el tamiz i; esel peso seco de la muestra total.
Se hicieron cortes finos en hecesfrescas y en heces de 5 meses deedad, que habían sido impregnadasen resina de poliéster Se hicierontres cortes en cada una de las dosfechas de muestreo. Los cortes seobservaron en una lupa binocular debaja magnificación (X3) paradescribir la estructura interna de lasheces y calcular, “a groso modo”, larelación proporcional de losdiferentes materiales que lasconstituían. Este proceso secomplementó con observaciones bajola lupa binocular (X4) de lamorfología externa de muestras noimpregnadas secadas al aire.
i
n
iiwxDPM
1
totalmuestra
tamizi M
Mw )(
ix
Mmuestra total

202
El Arado Natural:…
Muestras de raíces y demacroinvertebradosSe tomaron muestras de otras dosheces de cada grupo a los 0, 7, 14,30, 60, 105, 150, 195, 240 y330 días después de haber sidodepositadas, con el fin de determinarla biomasa de las raicillas y de losmacroinvertebrados (o sea, aquéllamayor que 2 mm) que estuvieranpresente en las heces. Losmacroinvertebrados se extrajeronmanualmente y las heces se lavaronen un tamiz de 0.053 mm de luz en latrama para extraer las raíces. Lasraíces vivas se separaronmanualmente, según su color, de lasmuertas y de los fragmentos dehojarasca. Los macroinvertebradosse guardaron en alcohol al 70%.Tanto las raíces como losmacroinvertebrados se secaron alhorno a 75 ºC durante 48 horas yluego se pesaron.
Propiedades físicas del sueloSe tomaron muestras del suelo en dosfechas (julio de 1996 y julio de 1997)para evaluar el DPM de los agregadosdel suelo y su DA. Para el DPM setomaron tres muestras al azar encada parcela experimental con uncilindro de aluminio de 10 cm dediámetro y 10 cm de altura. El suelose secó durante 48 horas y, medianteel tamizado en húmedo (antesdescrito), se distribuyeron losagregados en clases según sutamaño. Se tomaron seis muestrascon un cilindro de aluminio de 5 cmde diámetro y 10 cm de altura paraevaluar la DA del suelo. Lasmuestras se secaron a 70 °C en elhorno durante 48 horas y después sepesaron.
Análisis estadísticoSe empleó el test de Kolmogorov-Smirnov (Lilliefors 1967) para
comprobar si la frecuencia de losdatos sigue una distribución normal.Estos se transformaron antes delanálisis, siempre que era necesario,para reducir la asimetría de ladistribución mencionada. Lanormalización de los datos seconsiguió con la transformación Box-Cox (Sokal y Rohlf 1995) que es lasiguiente:
y = (xδ 1)/δLa prueba de normalidad y la
transformación se obtuvieronempleando el programa VerNorm 3.0del “R package” (Legendre y Vaudor1991).
Para probar que la edad dedeposición de las heces y el sistemade uso de la tierra ejercían un efectoen la biomasa de las raíces, de losmacroinvertebrados y de losmacroorganismos (raíces +macroinvertebrados) presentes en lasheces, así como en el DPM y en la DAde las mismas heces, se llevó a caboun ANOVA de dos vías. Los nivelesde probabilidad se ajustaronempleando la corrección deBonferroni para pruebas múltiplesencajadas (Cooper 1968); así, losniveles de 0.05, 0.01 y 0.001quedaron, respectivamente, como0.005 [= 0.05/(2 × 5)], 0.001 [= 0.01/(2 × 5)] y 0.0001 [= 0.001/(2 × 5)]. Lacomparación de las medias se hizocon la prueba de Fisher a un nivel designificación de 0.05.
El efecto que la colonización delas heces por los macroorganismoscausaba en el DPM de las mismasheces y en su DA se probó medianteregresiones lineales simples. Lasignificación de los coeficientes r seprobó en los niveles de significación0.05, 0.01 y 0.001.
ResultadosLas heces superficiales de losindividuos de M. carimaguensis son

Dinámica de la Degradación...
203
globulares (sensu Lee 1985) y estánformadas por agregados de grantamaño que semejan una torre (elturrículo) (Jiménez et al. 1998b).Tienen un diámetro que oscila entre3 y 6 cm, su altura fluctúa entre 2 y10 cm y, en promedio, su peso secoes de 25 g. Las heces más grandesllegan a 15 cm de altura y pesanhasta 400 g (peso seco).Generalmente, las lombricesdepositan las heces sobre la entradade la propia galería durante variosdías y, a veces, durante algunassemanas. Depositan capassucesivas de deyecciones que tienenunos pocos centímetros de diámetroy de altura (Foto 12-1). Las hecesestán constituidas, por tanto, poruna superposición de capas dediferentes edades, donde las másviejas (las secas y duras) se localizanen la base y las más recientes(frescas y blandas) en la partesuperior. Las heces no se secantotalmente hasta que las lombriceshayan abandonado definitivamente
sus galerías (Decaëns et al. 1999a).Las heces secas son unaacumulación de capas circulares queconforman una estructurasubcilíndrica compuesta pordiferentes agregados. Pasados unosmeses, las heces se rompen en formaprogresiva, probablemente por elimpacto de la lluvia y por el pisoteodel ganado, y los agregados sedispersan sobre la superficie delsuelo.
En las tres situacionesestudiadas, una fracción importantede las heces permaneció húmedadurante los 4 días que siguieron alcomienzo de la deposición(Figura 12-2). Durante este tiempo,las heces están expuestas a unarápida degradación por la acción delas gotas de lluvia, aunque lacontinua producción de heces por lalombriz lleva a la formación de unturrículo estable. La cantidad deheces secas aumentó más adelante yalcanzó el 100% de su poblacióntotal después de 7 días, en la
Foto 12-1. Turrículo fresco de Martiodrilus carimaguensis sobre la superficie del suelo de la sabananativa.
1 cm
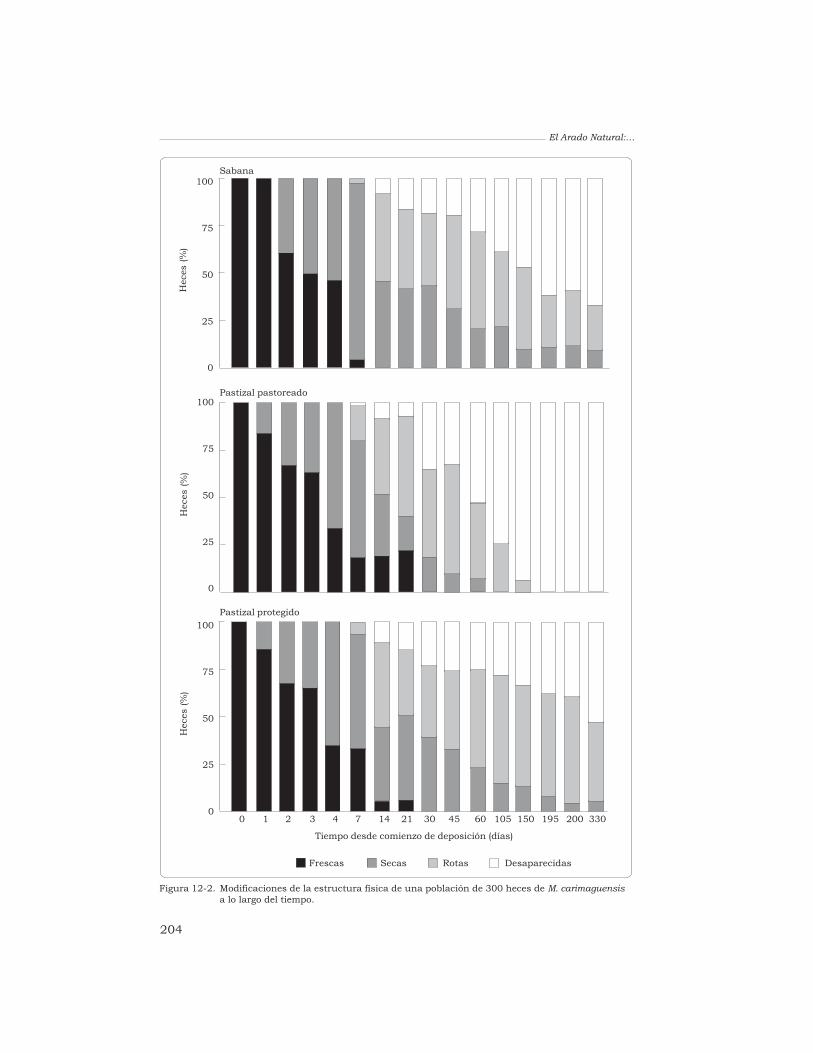
204
El Arado Natural:…
Figura 12-2. Modificaciones de la estructura física de una población de 300 heces de M. carimaguensisa lo largo del tiempo.
100
75
50
25
0
0 1 2 3 4 7 14 21 30 45 60 105 150 195 200 330
Hec
es (%
)
100
75
50
25
0
Hec
es (%
)
100
75
50
25
0
Hec
es (%
)
Tiempo desde comienzo de deposición (días)
Frescas Secas Rotas Desaparecidas
Pastizal pastoreado
Pastizal protegido
Sabana
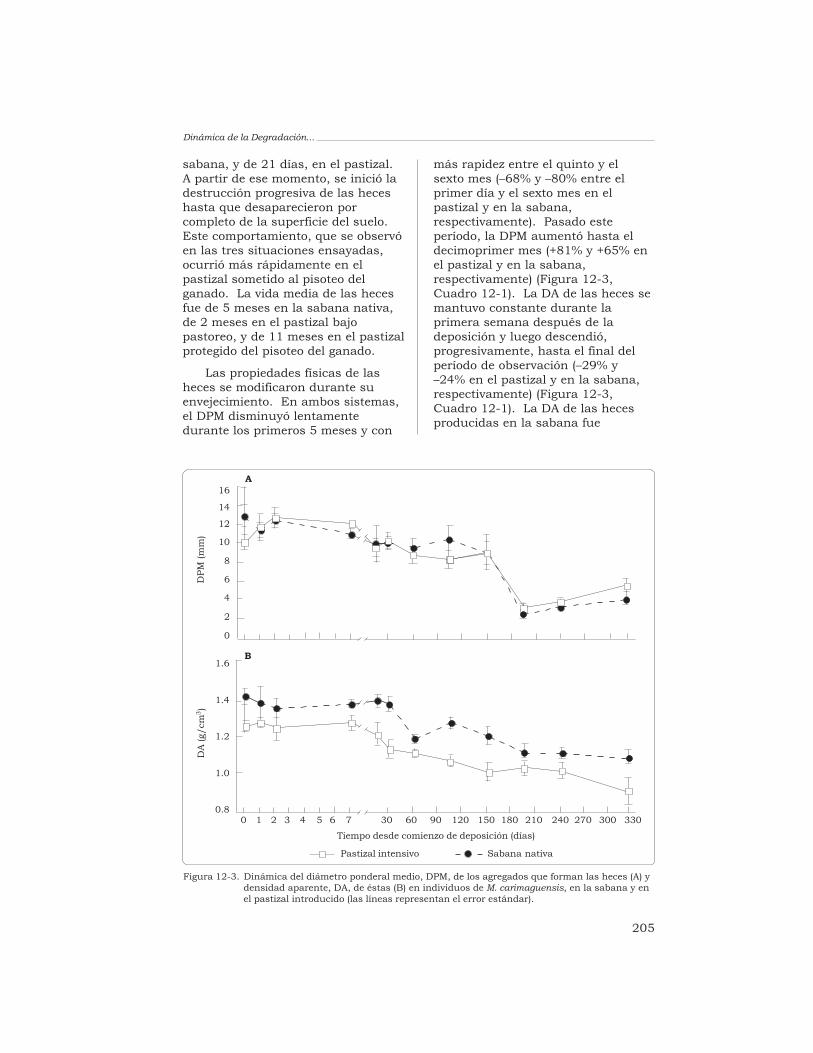
Dinámica de la Degradación...
205
sabana, y de 21 días, en el pastizal.A partir de ese momento, se inició ladestrucción progresiva de las heceshasta que desaparecieron porcompleto de la superficie del suelo.Este comportamiento, que se observóen las tres situaciones ensayadas,ocurrió más rápidamente en elpastizal sometido al pisoteo delganado. La vida media de las hecesfue de 5 meses en la sabana nativa,de 2 meses en el pastizal bajopastoreo, y de 11 meses en el pastizalprotegido del pisoteo del ganado.
Las propiedades físicas de lasheces se modificaron durante suenvejecimiento. En ambos sistemas,el DPM disminuyó lentamentedurante los primeros 5 meses y con
más rapidez entre el quinto y elsexto mes (–68% y –80% entre elprimer día y el sexto mes en elpastizal y en la sabana,respectivamente). Pasado esteperíodo, la DPM aumentó hasta eldecimoprimer mes (+81% y +65% enel pastizal y en la sabana,respectivamente) (Figura 12-3,Cuadro 12-1). La DA de las heces semantuvo constante durante laprimera semana después de ladeposición y luego descendió,progresivamente, hasta el final delperíodo de observación (–29% y–24% en el pastizal y en la sabana,respectivamente) (Figura 12-3,Cuadro 12-1). La DA de las hecesproducidas en la sabana fue
Figura 12-3. Dinámica del diámetro ponderal medio, DPM, de los agregados que forman las heces (A) ydensidad aparente, DA, de éstas (B) en individuos de M. carimaguensis, en la sabana y enel pastizal introducido (las líneas representan el error estándar).
Tiempo desde comienzo de deposición (días)
0 1 2 3 4 5 6 7 30 60 90 120 150 180 210 240 270 300 330
16
14
12
10
8
6
4
2
0
DPM
(mm
)
1.6
1.4
1.2
1.0
0.8
DA
(g/c
m3)
A
Pastizal intensivo Sabana nativa
B
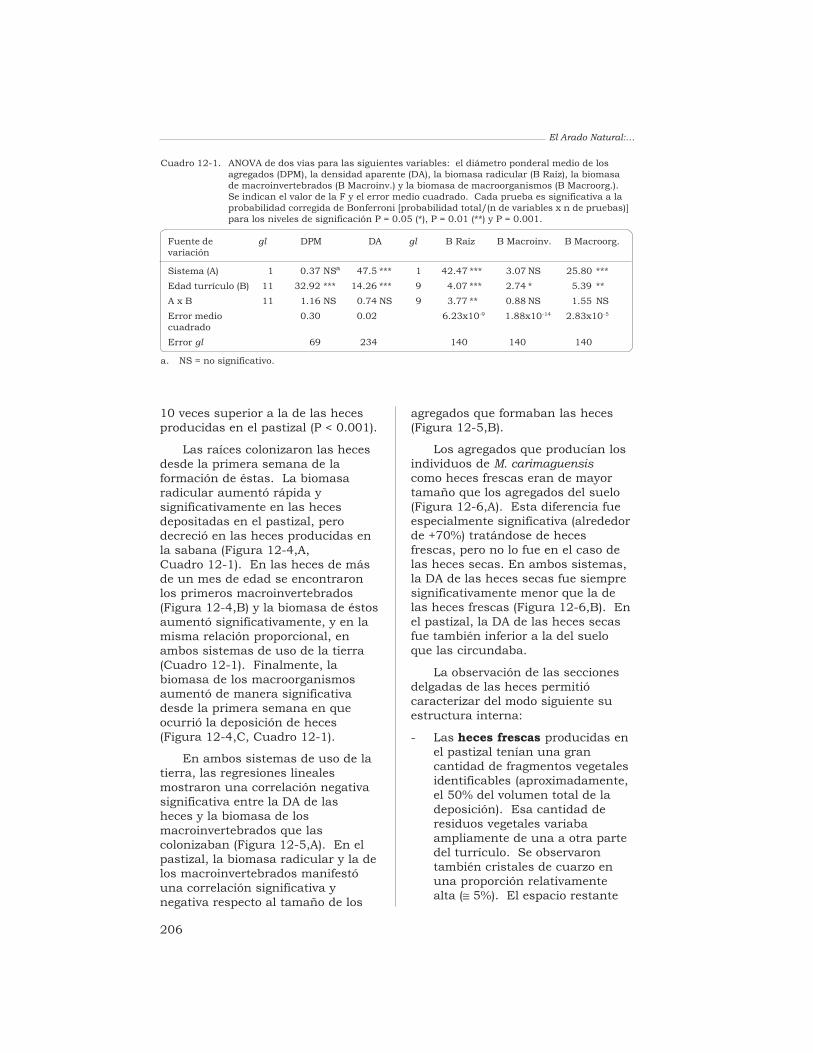
206
El Arado Natural:…
Cuadro 12-1. ANOVA de dos vías para las siguientes variables: el diámetro ponderal medio de losagregados (DPM), la densidad aparente (DA), la biomasa radicular (B Raíz), la biomasade macroinvertebrados (B Macroinv.) y la biomasa de macroorganismos (B Macroorg.).Se indican el valor de la F y el error medio cuadrado. Cada prueba es significativa a laprobabilidad corregida de Bonferroni [probabilidad total/(n de variables x n de pruebas)]para los niveles de significación P = 0.05 (*), P = 0.01 (**) y P = 0.001.
Fuente de gl DPM DA gl B Raíz B Macroinv. B Macroorg.variación
Sistema (A) 1 0.37 NSa 47.5 *** 1 42.47 *** 3.07 NS 25.80 ***Edad turrículo (B) 11 32.92 *** 14.26 *** 9 4.07 *** 2.74 * 5.39 **A x B 11 1.16 NS 0.74 NS 9 3.77 ** 0.88 NS 1.55 NSError medio 0.30 0.02 6.23x10-9 1.88x10-14 2.83x10-5
cuadradoError gl 69 234 140 140 140
a. NS = no significativo.
10 veces superior a la de las hecesproducidas en el pastizal (P < 0.001).
Las raíces colonizaron las hecesdesde la primera semana de laformación de éstas. La biomasaradicular aumentó rápida ysignificativamente en las hecesdepositadas en el pastizal, perodecreció en las heces producidas enla sabana (Figura 12-4,A,Cuadro 12-1). En las heces de másde un mes de edad se encontraronlos primeros macroinvertebrados(Figura 12-4,B) y la biomasa de éstosaumentó significativamente, y en lamisma relación proporcional, enambos sistemas de uso de la tierra(Cuadro 12-1). Finalmente, labiomasa de los macroorganismosaumentó de manera significativadesde la primera semana en queocurrió la deposición de heces(Figura 12-4,C, Cuadro 12-1).
En ambos sistemas de uso de latierra, las regresiones linealesmostraron una correlación negativasignificativa entre la DA de lasheces y la biomasa de losmacroinvertebrados que lascolonizaban (Figura 12-5,A). En elpastizal, la biomasa radicular y la delos macroinvertebrados manifestóuna correlación significativa ynegativa respecto al tamaño de los
agregados que formaban las heces(Figura 12-5,B).
Los agregados que producían losindividuos de M. carimaguensiscomo heces frescas eran de mayortamaño que los agregados del suelo(Figura 12-6,A). Esta diferencia fueespecialmente significativa (alrededorde +70%) tratándose de hecesfrescas, pero no lo fue en el caso delas heces secas. En ambos sistemas,la DA de las heces secas fue siempresignificativamente menor que la delas heces frescas (Figura 12-6,B). Enel pastizal, la DA de las heces secasfue también inferior a la del sueloque las circundaba.
La observación de las seccionesdelgadas de las heces permitiócaracterizar del modo siguiente suestructura interna:
- Las heces frescas producidas enel pastizal tenían una grancantidad de fragmentos vegetalesidentificables (aproximadamente,el 50% del volumen total de ladeposición). Esa cantidad deresiduos vegetales variabaampliamente de una a otra partedel turrículo. Se observarontambién cristales de cuarzo enuna proporción relativamentealta (≅ 5%). El espacio restante
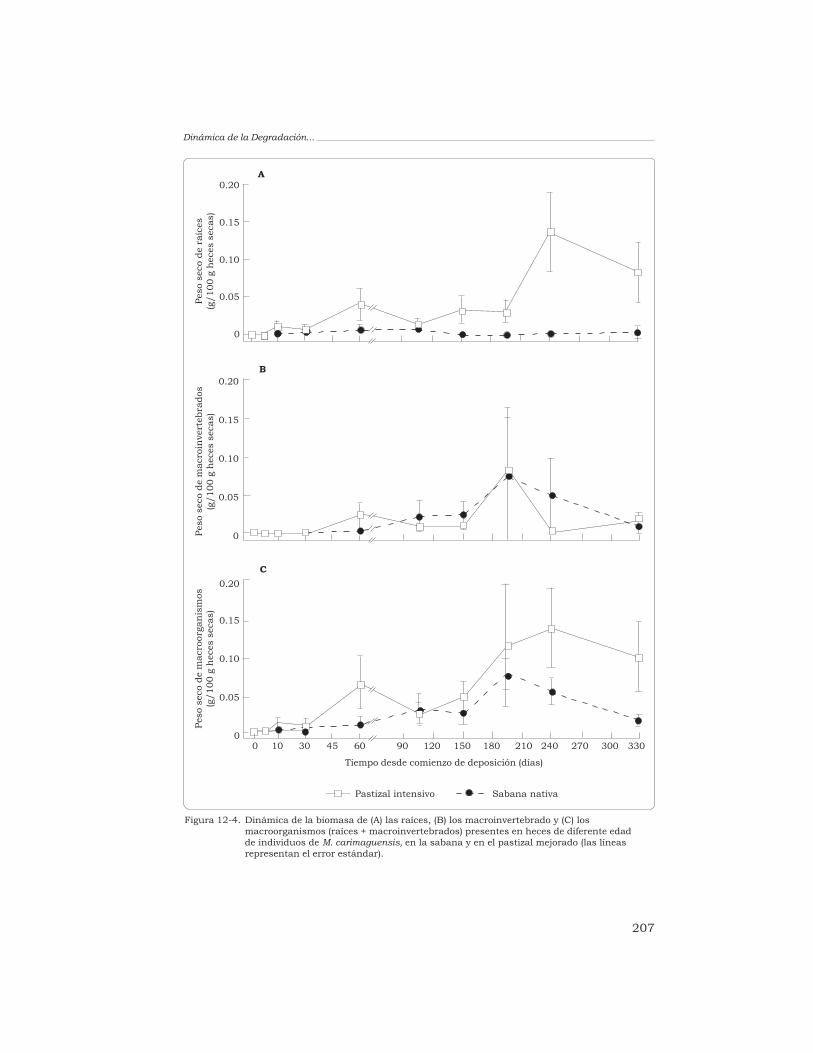
Dinámica de la Degradación...
207
0.20
0.15
0.10
0.05
0
Peso
sec
o de
raí
ces
(g/1
00 g
hec
es s
ecas
)
0.20
0.15
0.10
0.05
0Peso
sec
o de
mac
roin
vert
ebra
dos
(g/1
00 g
hec
es s
ecas
)
A
B
0 10 30 45 60 90 120 150 180 210 240 270 300 330
Peso
sec
o de
mac
roor
gan
ism
os(g
/100
g h
eces
sec
as)
0.20
0.15
0.10
0.05
0
C
Pastizal intensivo Sabana nativa
Tiempo desde comienzo de deposición (días)
Figura 12-4. Dinámica de la biomasa de (A) las raíces, (B) los macroinvertebrado y (C) losmacroorganismos (raíces + macroinvertebrados) presentes en heces de diferente edadde individuos de M. carimaguensis, en la sabana y en el pastizal mejorado (las líneasrepresentan el error estándar).
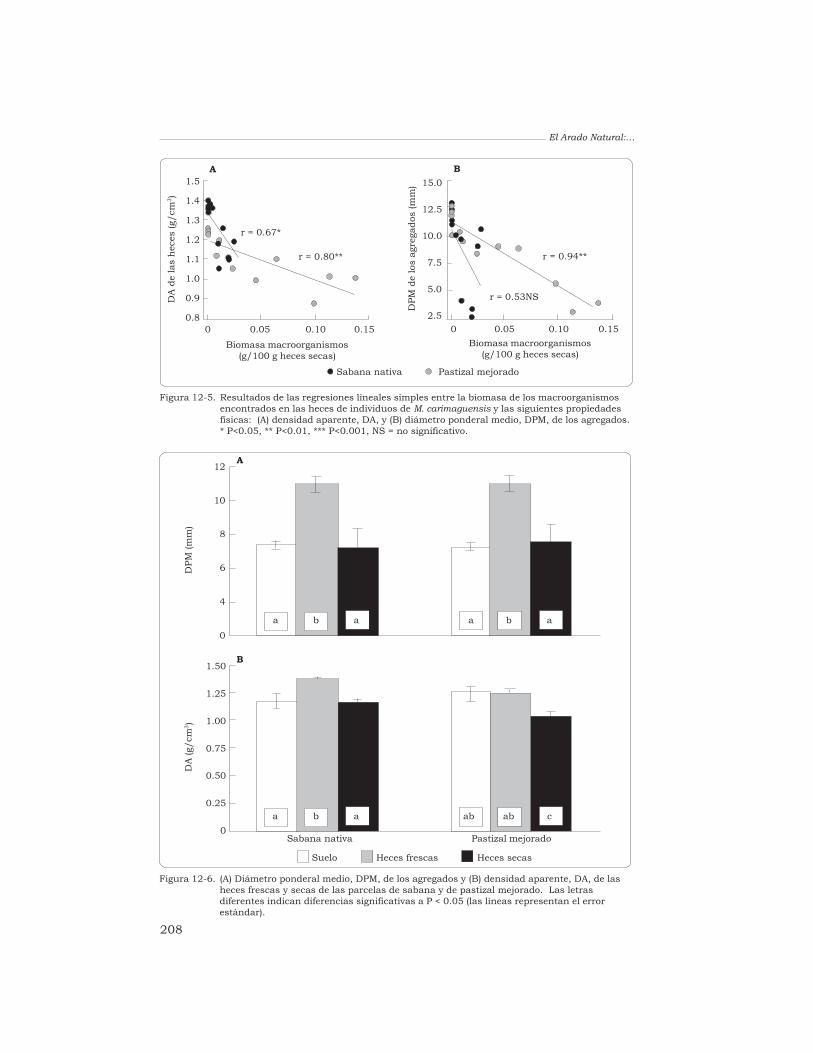
208
El Arado Natural:…
Figura 12-5. Resultados de las regresiones lineales simples entre la biomasa de los macroorganismosencontrados en las heces de individuos de M. carimaguensis y las siguientes propiedadesfísicas: (A) densidad aparente, DA, y (B) diámetro ponderal medio, DPM, de los agregados.* P<0.05, ** P<0.01, *** P<0.001, NS = no significativo.
Biomasa macroorganismos(g/100 g heces secas)
DA
de
las
hec
es (g
/cm
3 )1.5
1.4
1.3
1.2
1.1
1.0
0.9
0.80 0.05 0.10 0.15
r = 0.67*
A
r = 0.80**
Biomasa macroorganismos(g/100 g heces secas)
DPM
de
los
agre
gado
s (m
m) 15.0
12.5
10.0
7.5
5.0
2.5 0 0.05 0.10 0.15
r = 0.53NS
B
r = 0.94**
Figura 12-6. (A) Diámetro ponderal medio, DPM, de los agregados y (B) densidad aparente, DA, de lasheces frescas y secas de las parcelas de sabana y de pastizal mejorado. Las letrasdiferentes indican diferencias significativas a P < 0.05 (las líneas representan el errorestándar).
DPM
(mm
)
12
10
8
6
4
0
a b a a b a
A
DA
(g/c
m3 )
1.50
1.25
1.00
0.75
0.50
0.25
0
B
Sabana nativa Pastizal mejorado
Sabana nativa Pastizal mejorado
Suelo Heces frescas Heces secas
a b a ab ab c
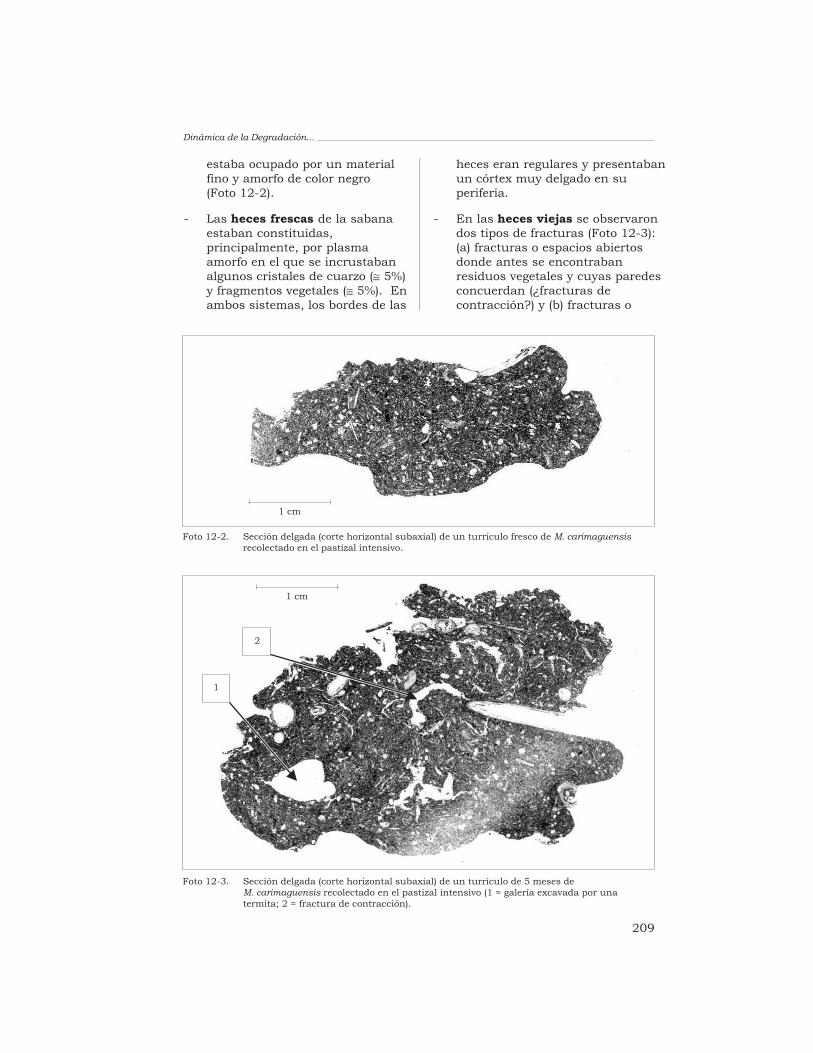
Dinámica de la Degradación...
209
Foto 12-2. Sección delgada (corte horizontal subaxial) de un turrículo fresco de M. carimaguensisrecolectado en el pastizal intensivo.
Foto 12-3. Sección delgada (corte horizontal subaxial) de un turrículo de 5 meses deM. carimaguensis recolectado en el pastizal intensivo (1 = galería excavada por unatermita; 2 = fractura de contracción).
2
1
1 cm
1 cm
estaba ocupado por un materialfino y amorfo de color negro(Foto 12-2).
- Las heces frescas de la sabanaestaban constituidas,principalmente, por plasmaamorfo en el que se incrustabanalgunos cristales de cuarzo (≅ 5%)y fragmentos vegetales (≅ 5%). Enambos sistemas, los bordes de las
heces eran regulares y presentabanun córtex muy delgado en superiferia.
- En las heces viejas se observarondos tipos de fracturas (Foto 12-3):(a) fracturas o espacios abiertosdonde antes se encontrabanresiduos vegetales y cuyas paredesconcuerdan (¿fracturas decontracción?) y (b) fracturas o
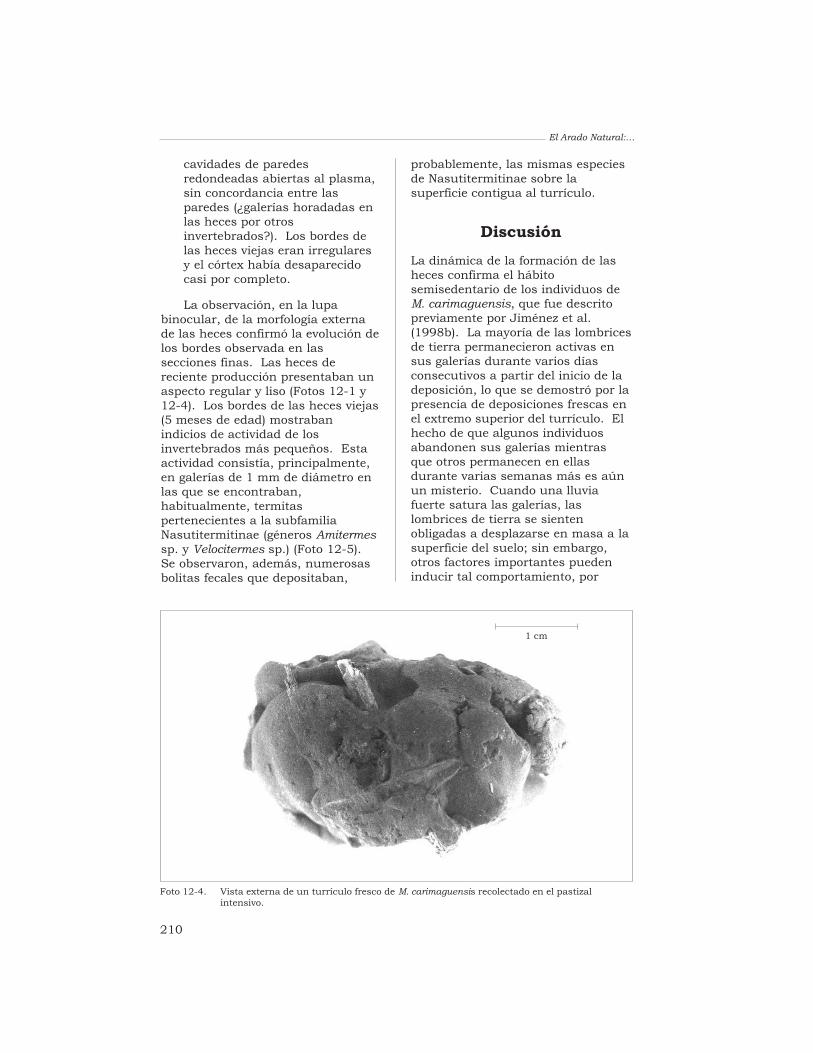
210
El Arado Natural:…
cavidades de paredesredondeadas abiertas al plasma,sin concordancia entre lasparedes (¿galerías horadadas enlas heces por otrosinvertebrados?). Los bordes delas heces viejas eran irregularesy el córtex había desaparecidocasi por completo.
La observación, en la lupabinocular, de la morfología externade las heces confirmó la evolución delos bordes observada en lassecciones finas. Las heces dereciente producción presentaban unaspecto regular y liso (Fotos 12-1 y12-4). Los bordes de las heces viejas(5 meses de edad) mostrabanindicios de actividad de losinvertebrados más pequeños. Estaactividad consistía, principalmente,en galerías de 1 mm de diámetro enlas que se encontraban,habitualmente, termitaspertenecientes a la subfamiliaNasutitermitinae (géneros Amitermessp. y Velocitermes sp.) (Foto 12-5).Se observaron, además, numerosasbolitas fecales que depositaban,
probablemente, las mismas especiesde Nasutitermitinae sobre lasuperficie contigua al turrículo.
DiscusiónLa dinámica de la formación de lasheces confirma el hábitosemisedentario de los individuos deM. carimaguensis, que fue descritopreviamente por Jiménez et al.(1998b). La mayoría de las lombricesde tierra permanecieron activas ensus galerías durante varios díasconsecutivos a partir del inicio de ladeposición, lo que se demostró por lapresencia de deposiciones frescas enel extremo superior del turrículo. Elhecho de que algunos individuosabandonen sus galerías mientrasque otros permanecen en ellasdurante varias semanas más es aúnun misterio. Cuando una lluviafuerte satura las galerías, laslombrices de tierra se sientenobligadas a desplazarse en masa a lasuperficie del suelo; sin embargo,otros factores importantes puedeninducir tal comportamiento, por
Foto 12-4. Vista externa de un turrículo fresco de M. carimaguensis recolectado en el pastizalintensivo.
1 cm
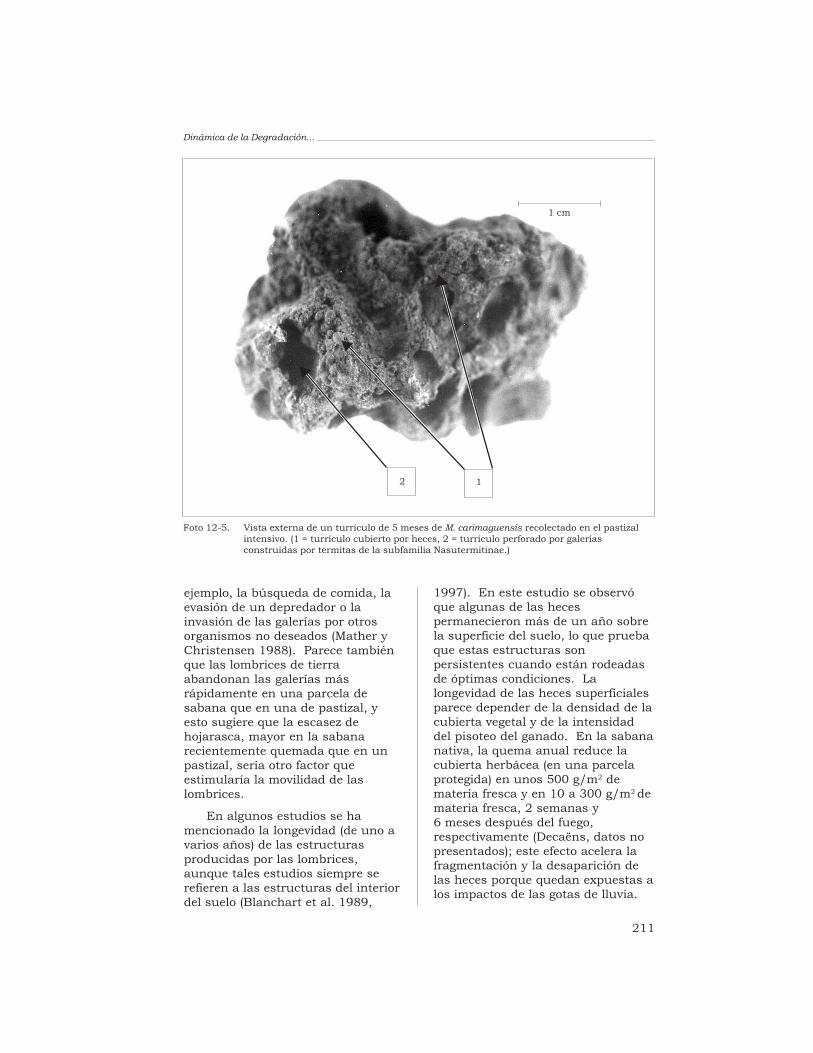
Dinámica de la Degradación...
211
ejemplo, la búsqueda de comida, laevasión de un depredador o lainvasión de las galerías por otrosorganismos no deseados (Mather yChristensen 1988). Parece tambiénque las lombrices de tierraabandonan las galerías másrápidamente en una parcela desabana que en una de pastizal, yesto sugiere que la escasez dehojarasca, mayor en la sabanarecientemente quemada que en unpastizal, sería otro factor queestimularía la movilidad de laslombrices.
En algunos estudios se hamencionado la longevidad (de uno avarios años) de las estructurasproducidas por las lombrices,aunque tales estudios siempre serefieren a las estructuras del interiordel suelo (Blanchart et al. 1989,
1997). En este estudio se observóque algunas de las hecespermanecieron más de un año sobrela superficie del suelo, lo que pruebaque estas estructuras sonpersistentes cuando están rodeadasde óptimas condiciones. Lalongevidad de las heces superficialesparece depender de la densidad de lacubierta vegetal y de la intensidaddel pisoteo del ganado. En la sabananativa, la quema anual reduce lacubierta herbácea (en una parcelaprotegida) en unos 500 g/m2 demateria fresca y en 10 a 300 g/m2 demateria fresca, 2 semanas y6 meses después del fuego,respectivamente (Decaëns, datos nopresentados); este efecto acelera lafragmentación y la desaparición delas heces porque quedan expuestas alos impactos de las gotas de lluvia.
Foto 12-5. Vista externa de un turrículo de 5 meses de M. carimaguensis recolectado en el pastizalintensivo. (1 = turrículo cubierto por heces, 2 = turrículo perforado por galeríasconstruidas por termitas de la subfamilia Nasutermitinae.)
2 1
1 cm

212
El Arado Natural:…
En parcelas recién quemadas, laslluvias fuertes pueden generar talescorrentía que arrastran las heceshacia los bajos. Este fenómeno seobservó justo al inicio delexperimento, cuando se produjo unafuerte tormenta (75 mm en 2 horas)que ocasionó la desaparición de lamitad de los turrículos marcados enla sabana nativa; no se observó elmismo efecto en el pastizal. En esteúltimo, la cubierta vegetal es gruesa(de 400 a 500 g/m2 de materiafresca) y permanente (Decaëns, datosno presentados) y, aunque evita laescorrentía, no puede impedir que elpisoteo del ganado acelere ladestrucción de los turrículos. Esnecesario investigar más el efecto detales procesos en la erosión y en laspérdidas de suelo derivadas de laescorrentía.
La presencia de una cantidadconsiderable de residuos orgánicosen los turrículos del pastizal explicala baja DA de éstos en comparacióncon los depositados en la sabana.En ausencia de tales fragmentos, lacompactación del suelo durante laformación de las heces puedeexplicarse por las siguientes razones:
- formación de uniones órgano-minerales durante el recorridodel suelo por el tracto digestivode las lombrices;
- reabsorción de agua en la parteposterior del intestino de laslombrices;
- compactación mecánica en elpigidio durante la excreción de ladeposición (Blanchart et al.1993).
La abundancia relativa defragmentos vegetales en las heceshalladas en el pastizal demuestratambién que los individuos deM. carimaguensis ingieren, al menosocasionalmente, hojarasca fresca
además de suelo. La ausencia dehojarasca en la superficie de lasabana quemada es, ciertamente, larazón de que se hayan observado tanescasos fragmentos vegetales en lasheces depositadas en la parcela desabana. Guggenberger et al. (1996)sostienen que la materia orgánica(MO) confiere mayor estabilidadestructural a las heces de losindividuos de M. carimaguensis que alas muestras del suelo circundante aellas. Esa MO, por su parte, esprotegida en las heces de lamineralización rápida, de modo quela producción de turrículos puedecontribuir a la formación de unareserva de C en el suelo (Lavelle yMartin 1992, Lavelle et al. 1998;Martin 1991).
El córtex presente en la periferiade las heces ha sido descrito en laespecie Millsonia anomala Omodeo(Megascolecidae) y su formación sedebe a la readsorción de agua en laparte final de los tractos (procesodigestivo), acción que atrae laspartículas finas hacia la periferia delas heces antes de su excreción(Blanchart et al. 1993). El córtexpodría reforzar la estabilidadestructural de las heces y crear lascondiciones anaeróbicas queconducen a una menormineralización de la MO contenidaen las heces (Blanchart et al. 1993;Elliott et al. 1990).
La reducción del tamaño de losagregados y de su DA se debe enparte, probablemente, a los cambiosde temperatura y de humedad queestimulan la formación de fisuras decontracción y debilitan la estructurade las deposiciones. Asimismo, lacolonización de las heces por algunosmacroorganismos(macroinvertebrados y raicillas)propicia la formación de galerías ycámaras en su interior. Estefenómeno se debe, principalmente, a

Dinámica de la Degradación...
213
la acción de especies del géneroNasutitermes (Isoptera:Nasutitermitinae), que horadan elinterior de las heces y lasfragmentan en agregados máspequeños. Otros estudios handemostrado también que las especiespequeñas son capaces de romper lasheces de las especies más grandes yreducirlas a agregados máspequeños y débiles (Blanchart et al.1997; Derouard et al. 1997; Rossi1998). Esta situación de equilibrioentre la producción y la destrucciónde las heces grandes por especies dediferentes tamaños tiene comoresultado una regulación activa de laestructura del suelo. El procesoimpide la acumulación de hecesgrandes y compactas sobre lasuperficie del suelo la cual, enalgunos casos, conduce a laformación de una capa impermeablecuyos efectos en el crecimiento de lasplantas son negativos (Blanchart etal. 1999; Chauvel et al. 1999; Rose yWood 1980).
El aumento del DPM observadoal final del experimento es un efectoartificial del diseño experimental.Con el tiempo, las heces menosestables desaparecían y las quequedaban disponibles para elmuestreo eran las más resistentes ala fragmentación. En consecuencia,la estabilidad y el tamaño de losagregados habrían sidosobreestimados en las últimas fechasdel experimento. La mismadificultad tendría la medición de laDA, lo que lleva a concluir que ladisminución real de la DA puede sermayor que la indicada en losresultados.
Los agregados que producen losindividuos de M. carimaguensis sonmás estables en agua que losagregados del suelo de tamañoequivalente (Guggenberger et al.1996). Esa producción puedeconsiderarse, por tanto, como un
factor determinante en la formaciónde una estructura estable de suelo(Blanchart et al. 1997; Marinissen1994). En los sistemas estudiados,las poblaciones de M. carimaguensispueden depositar al año, en lasabana y en el pastizal,respectivamente, 10 y 50 t/ha deheces secas superficiales, así como5 y 60 t/ha de heces subterráneas(Decaëns et al. 1999a). Porconsiguiente, esta especie puedeconvertir anualmente enmacroagregados del 2% al 11% delos primeros 10 cm del suelo. Estasapreciaciones apoyan algunasconclusiones obtenidas en estudiosprevios, en las que se refleja el papelque podían desempeñar laslombrices de tierra en la formaciónde una macroestructura estable enlos suelos de los pastizales deCarimagua (Gijsman 1996; Gijsmany Thomas 1995).
En conclusión, la persistencia delos turrículos producidos por laespecie anécica M. carimaguensisparece estar determinada por dosfactores antagónicos: (a) la densidadde la vegetación y de la capa dehojarasca, que protegen las heces delos impactos de la lluvia, y (b) elpisoteo del ganado, que acelera ladestrucción de las heces. Laformación de fisuras defragmentación en las heces y laactividad de los invertebrados máspequeños, que forman galerías en elinterior de ellas, pueden afectartambién, con el paso del tiempo, laestructura de estas deposiciones.
La participación de losmacroinvertebrados de menortamaño en la fragmentación de lasestructuras producidas por lasespecies de lombrices de tierragrandes apoya la hipótesis de queuna diversidad funcional altaconduce a una estabilidad mayor delos ecosistemas (Lawton y Brown1994, revisión de Blondel 1995). En

214
El Arado Natural:…
efecto, la conservación de unafauna funcionalmente diversa en elsuelo permite a las especiespequeñas de macroinvertebradosregular eficientemente lasestructuras producidas por lasespecies más grandes. Por último,esta combinación de efectos quecontrastan entre sí conduce a laregulación biogénica de laestructura del suelo y de laspropiedades físicas asociadas conella.
Martiodrilus carimaguensispuede considerarse un ingeniero delecosistema que regula importantesprocesos del suelo mediante lasheces que produce. Empleando laescala de las estructurasproducidas, el suelo estáconformado por macroagregadoscuyo tamaño y DA estándeterminados. La producción deuna cantidad importante de hecespuede causar, por tanto, efectossignificativos en la estructura, enlas propiedades físicas y en ladinámica de la MO del suelo, pero ala escala propia del perfil de éste.Estos efectos pueden modificar,indirectamente, los recursosespaciales (el espacio entre losporos) y tróficos (o sea, la MO) deotros invertebrados del suelo einfluir, de este modo, en el tamañoy en la organización de suscomunidades. Se han observado yadichos efectos en ciertos grupos demacroinvertebrados (Decaëns et al.1999b), pero es necesarioemprender otros estudios paracomprender, de manera másprecisa, la forma en que laslombrices de tierra pueden afectarla estructura de la comunidad delsuelo tomado como un todo.
AgradecimientosLos autores expresan suagradecimiento a R.J. Thomas,D.K. Friesen, E. Amézquita yC.G. Meléndez (CIAT) por el apoyotécnico recibido; a J.J. Jiménez(Universidad Complutense deMadrid) y a P. Lavelle, L. Mariani yJ.P. Rossi (IRD) por sus comentariosútiles a una primera versión de estecapítulo. Este trabajo fue financiadoparcialmente por el Consorcio parael Manejo de Suelos Acidos(Managing Acid Soils Consortium),perteneciente al Programa deManejo de Suelos, Agua yNutrientes (SWNM, en inglés).
ReferenciasAnderson JM. 1995. Soil organisms as
engineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linkingspecies and ecosystems. Chapmany Hall, Londres. p 94-106.
Barois I; Lavelle P; Brossard M; TondohJ; Martinez MA; Rossi J-P;Senapati B; Angeles A; Fragoso C;Jiménez JJ; Decaëns T; Lattaud C;Kanyonyo J; Blanchart E; ChapuisL; Brown GG; Moreno AG. 1999.Ecology of earthworm species withlarge environmental tolerance and/or extended distributions. En:Lavelle P; Brussaard L; Hendrix P,eds. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido. p 57-85.
Blanchart E. 1990. Rôle fonctionneldes vers de terre dans la formationde la structure des sols de lasavane de Lamto (Côte d’Ivoire).Tesis (Doctorado). Université deRennes I.
Blanchart E; Bruand A; Lavelle P. 1993.The physical structure of casts ofMillsonia anomala (Oligochaeta:Megascolecidae) in shrub savannasoils (Côte d’Ivoire). Geoderma56:119-132.

Dinámica de la Degradación...
215
Blanchart E; Lavelle P; Spain AV. 1989.Effects of two species of tropicalearthworms (Oligochaeta:Eudrilidae) on the size distributionof aggregates in an African soil. RevEcol Biol Sols 26:417-425.
Blanchart E; Lavelle P; Braudeau E;Bissonnais YL; Valentin C. 1997.Regulation of soil structure bygeophagous earthworm activities inhumid savannas of Côte d’Ivoire.Soil Biol Biochem 29:431-439.
Blanchart E; Albrecht A; Alegre J;Duboisset A; Villenave C; PashanasiB; Lavelle P; Brussaard L. 1999.Effects of earthworms on soilstructure and physical properties.En: Lavelle P; Brussaard L; HendrixP, eds. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido. p 149-172.
Blondel J. 1995. Biogéographie: approcheecologique et evolutive. Masson,París, Francia.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm.Nature 398:32-33.
Cooper DW. 1968. The significancelevel in multiple tests madesimultaneously. Heredity23:614-617.
Decaëns T; Rangel AF; Asakawa N;Thomas RJ. 1999a. Carbon andnitrogen dynamics in ageingearthworm casts in grasslands ofthe Eastern Plains of Colombia. BiolFertil Soils 30:20-28.
Decaëns T; Mariani L; Lavelle P. 1999b.Soil surface macrofaunalcommunities associated withearthworm casts in grasslands ofthe Eastern Plains of Colombia.Appl Soil Ecol 13:87-100.
Derouard L; Tondoh J; Vilcosqui L;Lavelle P. 1997. Effects ofearthworm introduction on soilprocesses and plant growth. SoilBiol Biochem 29:541-545.
Elliott PW; Knight D; Anderson JM. 1990.Denitrification in earthworm castsand soil from pastures underdifferent fertilizer and drainageregimes. Soil Biol Biochem22:601-605.
Gijsman AJ. 1996. Soil aggregate stabilityand soil organic matter fractionsunder agropastoral systemsestablished in native savanna. AustJ Soil Res 34:891-907.
Gijsman AJ; Thomas RJ. 1995. Aggregatesize distribution and stability of anOxisol under legume-based andpure grass pasture in the EasternColombian savannas. Aust J SoilRes 33:153-165.
Guggenberger G; Thomas RJ; Zech W.1996. Soil organic matter withinearthworm casts of an anecic-endogeic tropical pasturecommunity, Colombia. Appl SoilEcol 3:263-274.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998a. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998b. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78:1946-1957.

216
El Arado Natural:…
Kemper WD; Rousenau RC. 1986.Aggregate stability and sizedistribution. En: Klute A, ed.Methods of soil analysis. AmericanSociety of Agronomy/Soil ScienceSociety of America, Madison,Estados Unidos. p 425-442.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humidtropics. Soil Biol Biochem24:1491-1498.
Lavelle P; Pashanasi B; Charpentier F;Gilot C; Rossi J-P; Derouard L;Andre J; Ponge JP; Bernier N. 1998.Large-scale effects of earthwormson soil organic matter and nutrientdynamics. En: Edwards CA, ed.Earthworm ecology. St. LuciesPress, Boca Ratón, Estados Unidos.p 103-122.
Lawton JH; Brown VK. 1994.Redundancy in ecosystems. En:Schulze ED; Mooney HA, eds.Biodiversity and ecosystemfunction. Springer-Verlag, Berlín.p 255-270.
Lee KE. 1985. Earthworms. Their ecologyand relationships with soils andland use. Academic Press, Orlando,FL, Estados Unidos.
Legendre P; Vaudor A. 1991. The RPackage: multidimentional analysis,spatial analysis. Université deMontréal, Département desSciences Biologiques, Montreal,Quebec.
Lilliefors HW. 1967. The Kolmogorov-Smirnov test for normality withmean and variance unknown. J AmStat Assoc 62:399-402.
Marinissen JCY. 1994. Earthwormpopulations and stability of soilstructure in a silt loam soil ofrecently reclaimed polder in theNetherlands. Agric Ecosyst Environ51:75-87.
Martin A. 1991. Short- and long-termeffects of the endogeic earthwormMillsonia anomala (Omodeo)(Megascolecidae: Oligochaeta) oftropical savannas, on soil organicmatter. Biol Fertil Soils 11:134-138.
Mather JG; Christensen O. 1988. Surfacemigration of earthworms ingrassland. Pedobiologia 36:51-57.
Rose CJ; Wood AW. 1980. Someenvironmental factors affectingearthworm populations and sweetpotato production in the Tari Basin,Papua New Guinea Highlands.Papua New Guinea Agric J 31:1-10.
Rossi J-P. 1998. Rôle fonctionnel de ladistribution spatiale des vers deterre dans une savane humide deCôte d’Ivoire. Tesis (Doctorado).Université Paris VI-Pierre et MarieCurie.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed. WHFreeman, Nueva York, EstadosUnidos.

Dinámica del Nitrógeno...
217
PARTE IV
Efectos Producidos por los Ingenierosdel Ecosistema en la Materia
Orgánica y en el Ciclo deNutrientes del Suelo, a Escala de las
Estructuras Biogénicas

Dinámica del Nitrógeno...
219
ResumenSe estudiaron los efectos producidospor una especie de lombriz anécicade gran tamaño (Martiodriluscarimaguensis Jiménez y Moreno) enla dinámica del nitrógeno mineral enparcelas de sabana nativa y depastizal mejorado de los LlanosOrientales de Colombia. Se comparóel C total, el N total y el contenido deNH4 y de NO3 en las heces de laslombrices y en el suelo no ingerido.Las muestras se tomaron endiferentes fechas hasta pasados20 días después de la deposición delas heces. El contenido total de C yde N fue varias veces superior en lasheces que en el suelo no ingerido entodos los sistemas ensayados. Elnivel de NH4 fue de 5.2 a 28.3 vecesmás alto en las heces de laslombrices que en el suelo, según el
sistema de uso de la tierra ensayado.El N mineral aumentó rápidamenteen las heces en los primeros días quesiguieron a la deposición de éstas ydecreció más tarde, aunque alcanzóvalores 4.5 veces superiores a losobtenidos en muestras de suelo. ElN incorporado en compuestosminerales por acción de laslombrices de tierra equivale a unaaplicación de 40 a 63 kg/ha de Nal año.
Palabras clave: dinámica de la MOdel suelo, dinámica del N,lombrices de tierra, Martiodriluscarimaguensis, suelos ácidos
IntroducciónLa actividad desarrollada por laslombrices es uno de los principalesreguladores de la dinámica de lamateria orgánica del suelo (MO) ycon esa actividad mejoran la calidaddel suelo (Lavelle et al. 1989). Variosestudios han demostrado que lasheces de las lombrices de tierra sonmás ricas en N que el suelo que lascircunda (Syers et al. 1979). Elpapel que las lombrices desempeñanen la dinámica del N depende tantode la cantidad de suelo ingeridocomo de la calidad del material queellas ingieren. En los LlanosOrientales de Colombia, Jiménez etal. (1998) calcularon que la
CAPÍTULO 13
Dinámica del Nitrógeno en las Hecesde las Lombrices de Tierra de laEspecie Martiodrilus carimaguensisJiménez y Moreno, en un Oxisol delas Sabanas de Colombia1
A. F. Rangel* , R. J. Thomas*, J. J. Jiménez** yT. Decaëns***
1. Publicado en Pedobiologia 43, p 557-560, porRangel et al. Copyright © 1999 de Urban &Fischer Verlag. Impreso con permiso de lacasa editorial.
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** Departamento de Biología Animal I(Zoología), Facultad de Biología, UniversidadComplutense de Madrid, 28040, Madrid,España.
*** Laboratoire d’ Ecologie des Sols Tropicaux,IRD-Bondy, 32 Av. Henri Varagnat, F-93143Bondy Cedex, Francia.
+
+
–

220
El Arado Natural:…
producción de heces secas (en lasuperficie y bajo ella) de una especieanécica era de 14.3 t/ha por año enla sabana nativa y de 378 t/ha poraño en un pastizal constituido porgramíneas y leguminosas mejoradas.El trabajo desarrollado en estecapítulo consistió en evaluar el efectoque causan las lombrices de laespecie Martiodrilus carimaguensis(Oligochaeta: Glossoscolecidae) en ladinámica del N en diferentessistemas de uso de la tierrapracticados en los Llanos Orientalesde Colombia.
Materiales y MétodosEl estudio se realizó en la estaciónexperimental de Carimagua (CIAT-CORPOICA) (4o37’ N, 71o19’ O,175 msnm) en los Llanos Orientalesde Colombia. La precipitación mediaanual oscila entre 2100 y 2300 mm yla temperatura media anual es de26 °C. Los suelos son Oxisoles biendrenados de tipo franco-arcilloso(fino, mezclado, isohipertérmico típicoHaplustox), con un pH (H2O) de 4.5,una saturación de Al > 80%y concentraciones bajas de loscationes de Ca, Mg, K y P (Rao et al.1993).
Se tomaron muestras en tresparcelas experimentales querepresentaban tres sistemasdiferentes de uso de la tierra:
- la sabana nativa (Sav), manejadadel modo tradicional que incluyela quema anual durante la épocaseca;
- un pastizal introducido de17 años en que se sembróBrachiaria decumbens cv. Basilisk(Bdec);
- un pastizal introducido de17 años de edad, en el que sesembró B. decumbens asociadocon la leguminosa forrajera
herbácea Pueraria phaseoloidesCIAT 9900 (“kudzú”) (Bdec/Pp).
Los pastizales que seestablecieron donde antes habíasabana nativa fueron fertilizados conlos siguientes elementos (en kg/ha):44 de P, 40 de K, 14 de Mg y 22 deS al inicio del ensayo, y con 10 de P,9 de K, 92.5 de Mg y 11 de S cadasegundo año en los siguientes 9 años(Lascano y Estrada 1989). La cargaanimal en los pastizales introducidosera de 1 animal/ha durante la épocaseca y de 2 animales/ha durante lalluviosa.
Las heces frescas deM. carimaguensis fueron marcadas yluego aisladas en el terreno bajo tresjaulas de 1 x 1 m que se colocaron alazar en cada pastizal introducido, ytambién en un área de 4 x 4 mdelimitada en la sabana nativa.Cada día se aislaban muestras deheces frescas que se mantenían bajocondiciones de campo para obtenerlos siguientes tiempos de incubacióntotal: 0.5, 1, 2, 4, 7, 15 y 20 días, enlos cuales las muestras equilibrabansu humedad con la del suelo. Lasmuestras fueron molidas ytamizadas a 2 mm de trama paradeterminar el C total, el N total(método Kjeldahl) y el N mineral(método colorimétrico). La cantidadde N total digerido anualmente, porhectárea, por las lombrices deM. carimaguensis y que se liberacomo N mineral, representa latotalidad de la tasa de mineralizacióndel N.
ResultadosEl contenido de C total del suelo fue11% y 23% más alto en Bdec(22.64 g/kg) y en Bdec/Pp (25.09 g/kg), respectivamente, que en lasabana nativa (20.4 g/kg); no hubo,en cambio, diferencias significativasrespecto al contenido de N total entre
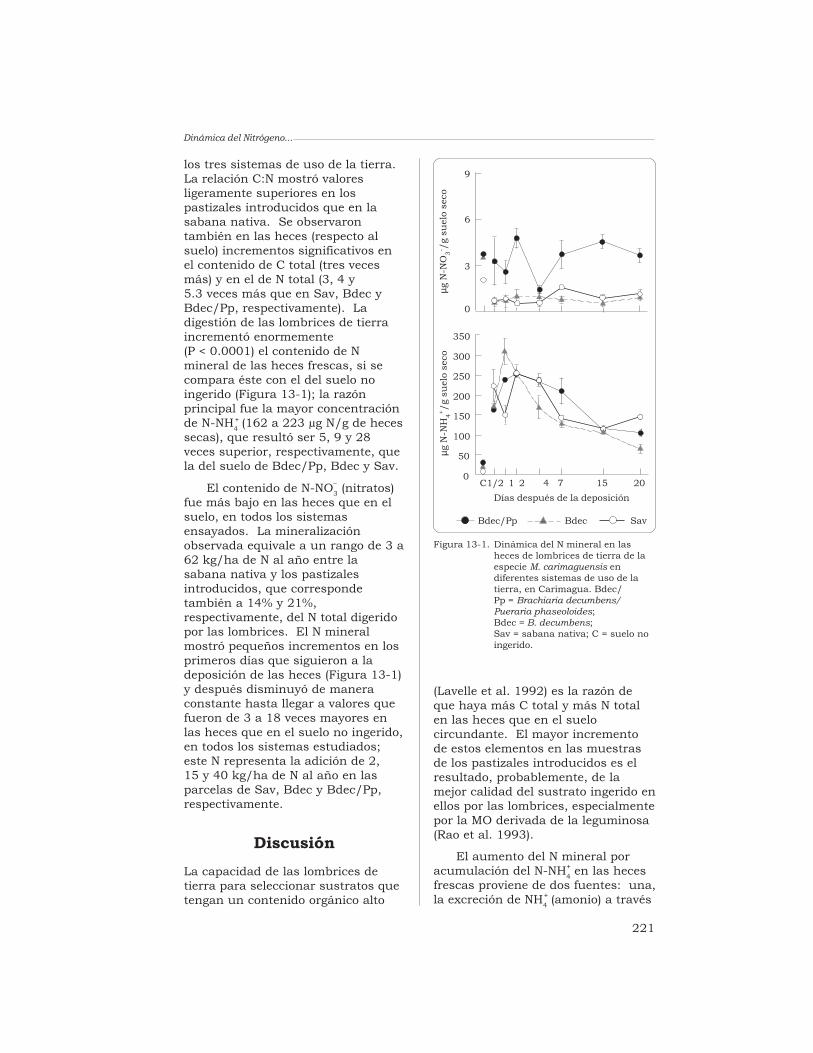
Dinámica del Nitrógeno...
221
Figura 13-1. Dinámica del N mineral en lasheces de lombrices de tierra de laespecie M. carimaguensis endiferentes sistemas de uso de latierra, en Carimagua. Bdec/Pp = Brachiaria decumbens/Pueraria phaseoloides;Bdec = B. decumbens;Sav = sabana nativa; C = suelo noingerido.
9
6
3
0
µg N
-NO
3- /g
suel
o se
co
350
300
250
200
150
100
50
0
µg N
-NH
4+ /g
suel
o se
co
C1/2 1 2 4 7 15 20Días después de la deposición
Bdec/Pp Bdec Sav
los tres sistemas de uso de la tierra.La relación C:N mostró valoresligeramente superiores en lospastizales introducidos que en lasabana nativa. Se observarontambién en las heces (respecto alsuelo) incrementos significativos enel contenido de C total (tres vecesmás) y en el de N total (3, 4 y5.3 veces más que en Sav, Bdec yBdec/Pp, respectivamente). Ladigestión de las lombrices de tierraincrementó enormemente(P < 0.0001) el contenido de Nmineral de las heces frescas, si secompara éste con el del suelo noingerido (Figura 13-1); la razónprincipal fue la mayor concentraciónde N-NH4 (162 a 223 µg N/g de hecessecas), que resultó ser 5, 9 y 28veces superior, respectivamente, quela del suelo de Bdec/Pp, Bdec y Sav.
El contenido de N-NO3 (nitratos)fue más bajo en las heces que en elsuelo, en todos los sistemasensayados. La mineralizaciónobservada equivale a un rango de 3 a62 kg/ha de N al año entre lasabana nativa y los pastizalesintroducidos, que correspondetambién a 14% y 21%,respectivamente, del N total digeridopor las lombrices. El N mineralmostró pequeños incrementos en losprimeros días que siguieron a ladeposición de las heces (Figura 13-1)y después disminuyó de maneraconstante hasta llegar a valores quefueron de 3 a 18 veces mayores enlas heces que en el suelo no ingerido,en todos los sistemas estudiados;este N representa la adición de 2,15 y 40 kg/ha de N al año en lasparcelas de Sav, Bdec y Bdec/Pp,respectivamente.
DiscusiónLa capacidad de las lombrices detierra para seleccionar sustratos quetengan un contenido orgánico alto
+
–
(Lavelle et al. 1992) es la razón deque haya más C total y más N totalen las heces que en el suelocircundante. El mayor incrementode estos elementos en las muestrasde los pastizales introducidos es elresultado, probablemente, de lamejor calidad del sustrato ingerido enellos por las lombrices, especialmentepor la MO derivada de la leguminosa(Rao et al. 1993).
El aumento del N mineral poracumulación del N-NH4 en las hecesfrescas proviene de dos fuentes: una,la excreción de NH4 (amonio) a través
+
+

222
El Arado Natural:…
de los endonefridios de la lombriz,y otra, la mineralización de la MOpor la microflora del suelo en laspartes media y posterior delintestino de la lombriz (Lavelle etal. 1992). Entre el 79% y el 96% detodo el N mineral liberado en lasheces frescas proviene de ladigestión de las lombrices. El valorbajo de N-NO3 obtenido en lasheces frescas corrobora la hipótesisde que los nitratos no son unproducto metabólico de laslombrices de tierra (Lavelle et al.1992).
El descenso del N mineral conel tiempo sugiere, probablemente,la transformación del N en biomasamicrobiana y la pérdida de Ndebida a volatilización,denitrificación o lixiviación (Lavelleet al. 1992). En los experimentosrealizados a largo plazo con lamisma especie de lombriz se haobservado que el N mineralaumenta en el suelo que circundalas heces después de la deposiciónde éstas y que hay un incrementoen el valor de N-NO3, lo que indicaque el N pasa de las heces al suelo(Decaëns et al. 1999).
Aunque en la sabana nativa sehallaron las tasas más altas demineralización (214 µg N/g sueloseco), las amplias diferencias,respecto a la sabana, en lacantidad de N mineral liberado encada hectárea de los pastizalesmejorados pueden explicarse por lagran cantidad de biomasa delombrices que hay en éstos porunidad de área y por la cantidad desuelo que ellas ingieren (Jiménez etal. 1998). Estos resultadosseñalan, finalmente, la importanciaque tiene el sistema de uso de latierra en la actividad de laslombrices.
ConclusionesLa introducción de pastizalesmejorados en áreas de sabana nativaaumenta en ésta el nivel de MO delsuelo, lo que contribuye a unincremento en la densidadpoblacional y en la actividad de lascomunidades de lombrices de tierra.El proceso de digestión de unalombriz de tierra ocasiona unaumento (respecto al suelo) de la MOen las deposiciones, así como unamayor mineralización del nitrógenoen general. La magnitud de dichoscambios depende de la MO contenidaen el suelo y de la calidad de lahojarasca ingerida por la lombriz. Lagran cantidad de N mineralencontrado en los pastizalesestudiados proviene de la altadensidad de población de laslombrices de tierra que ingierenanualmente varias toneladas desuelo por hectárea.
El manejo adecuado de lascomunidades de lombrices en lospastizales mejorados puede ser elmecanismo que incremente la MOdel suelo y el ciclo de los nutrientes,especialmente del N, lo que reduciríael riesgo de degradación del pastizal.Las investigaciones que se lleven acabo, a largo plazo, sobre los efectosde las lombrices de tierra en lacalidad del suelo deberánconcentrarse en lo que esasinvestigaciones impliquen para lasostenibilidad de un sistema de usode la tierra.
AgradecimientosEl antiguo Programa de Trópico Bajodel CIAT y el proyecto Macrofauna(Comunidad Europea) apoyaron estainvestigación. Los autores agradecenla ayuda recibida de todo el personalde Carimagua y de la sección deServicios Analíticos del CIAT.Agradecen también al Dr. Rao y al
−
−

Dinámica del Nitrógeno...
223
Dr. Friesen por sus comentariossobre el manuscrito. Se obtuvotambién financiación del Consorciopara el Manejo de Suelos Ácidos(MAS) que pertenece al programa delCGIAR sobre Manejo de Suelos, Aguay Nutrientes (SWNM, en inglés).
ReferenciasDecaëns T; Rangel AF; Asakawa N;
Thomas RJ. 1999. Carbon andnitrogen dynamics in in-situ ageingearthworm casts in natural savannaand man-made pasture of theEastern Plains of Colombia. BiolFertil Soils 30:20-28.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the EasternPlains of Colombia. Appl Soil Ecol9:153-160.
Lascano CE; Estrada J. 1989. Long-termproductivity of legume-based andpure grass pastures in the EasternPlains of Colombia. En: Proceedingsof the XVI International GrasslandCongress, Niza, Francia.p 1179-1180.
Lavelle P; Schaefer R; Zaidi Z. 1989. Soilingestion and growth in Millsoniaanomala, a tropical earthworm, asinfluenced by the quality of theorganic matter ingested.Pedobiologia 33:379-388.
Lavelle P; Meléndez G; Pashanasi B;Schaefer R. 1992. Nitrogenmineralization sand reorganizationin cast of the geophagous tropicalearthworm Pontoscolex corethrurus(Glossoscolecidae). Biol Fertil Soils14:49-53.
Rao IM; Borrero V; Ayarza MA; García R.1993. Adaptation of tropical foragespecies to acid soils: the influenceof varying phosphorus supply andsoil type on phosphorus uptake anduse. En: Barrow NJ, ed. Plantnutrition: from genetic engineeringto field practice. Kluwer AcademicPublishers, Dordrecht, Holanda.p 345-348.
Syers JK; Sharpley A; Kenney DR. 1979.Cycling of nitrogen by surface-casting earthworms in a pastureecosystem. Soil Biol Biochem11:181-185.

224
El Arado Natural:…
ResumenLas lombrices de tierra son ingenierosdel ecosistema que desempeñan unpapel importante en la dinámica de lamateria orgánica (MO) y la estructurafísica del suelo. Tanto por su tamañocomo por sus actividades, estosorganismos pueden acelerar ladescomposición y la mineralizaciónde la MO ya que reducen en tamañolos residuos orgánicos hastaconvertirlos en partículas másdisponibles para la microflora. Enconsecuencia, el impacto de susactividades en el ciclo y en ladisponibilidad de los nutrientes,tanto en ecosistemas naturales comoen agroecosistemas, llega a serconsiderable. Se realizaron tresexperimentos en una sabanaherbácea y en un pastizal deBrachiaria decumbens y Pueraria
phaseoloides (“kudzú”): un estudio decampo y dos ensayos de incubaciónen el laboratorio en la estaciónexperimental Carimagua del convenioCIAT-CORPOICA, situada en lassabanas isohipertérmicaspertenecientes a la cuenca delOrinoco en Colombia. En lasmuestras de los ensayos de campohabía más fósforo (P) en las hecesdepositadas por las lombrices que enel suelo alrededor de ellas: 50% másen las de la sabana nativa y 100%más en las del pastizal; en lasmuestras de heces producidas en ellaboratorio, esta diferencia fue nula(o muy pequeña: 10% a 20%). En elensayo de campo, casi sin excepción,todas las fracciones de P aumentaronen las heces respecto al suelo quehacía de testigo. Este incremento fuemás notorio, relativamente, en lasfracciones lábiles de P inorgánico (Pi).Excepto en la sabana (en condicionesde campo), la actividad de la fosfatasafue menor en las heces. Losresultados del estudio sugieren quelas lombrices de tierra, en condicionesnaturales, incorporan el P provenientede la hojarasca o de algún materialvegetal o de raíces no descompuestasque, normalmente, no se evalúa en elsuelo testigo. Este P proveniente de lahojarasca llega a todas las fracciones(orgánicas e inorgánicas), aunque sedistribuye más en las fraccioneslábiles de Pi. El objetivo de este
CAPÍTULO 14
Disponibilidad de Fósforo en las Hecesde una Especie Anécica de Lombriz deTierra de las Sabanas de Suelo Oxisolen Colombia1
J. J. Jiménez*, A. Cepeda*, D. K. Friesen**,T. Decaëns*** y A. Oberson†
1. Presentado por el primer autor en el XIIICongreso Internacional de Zoología delSuelo, Ceské Budejovice, República Checa,14 a 18 de agosto de 2000.
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** CIMMYT-IFDC, P.O. Box 25171, Nairobi,Kenia.
*** Laboratoire d’Ecologie, UFR Sciences etTechniques, Université de Rouen, F-76821Mont Saint Aignan Cedex, Francia.
† ETH, Institute of Plant Sciences, Eschikon,33, CH-8315, Lindau, Suiza.
> >

Disponibilidad de Fósforo en las Heces...
225
estudio fue evaluar el efecto queejerce una especie de lombriz detierra anécica, que es nativa yabundante, en la disponibilidad delP en los Oxisoles considerados debaja fertilidad.
Palabras clave: ciclo de nutrientes,fauna del suelo, fósforo,ingenieros del ecosistema,lombrices de tierra, sabanatropical
Este artículo está dedicado aPaulo César Cepeda Virviescas
in memoriam (A.C.)
IntroducciónLos invertebrados son los principalesorganismos de la fauna del suelo quedeterminan los procesos ocurridosen el suelo de los ecosistemastropicales (Lavelle et al. 1994). Enmuchos estudios se ha encontradomayor cantidad de fósforo (P)asimilable en las heces de laslombrices de tierra que en el suelotestigo (Guggenberger et al. 1996;Krishnamoorthy 1990; Lal 1974;Lunt y Jacobson 1944; Nye 1955).Estos organismos causan unimpacto notable en la mineralizacióndel P (Brossard et al. 1996; Chapuisy Brossard 1995; James 1991;López-Hernández et al. 1993;Sharpley y Syers 1976). Sharpley ySyers (1976) encontraron mayordisponibilidad de P en las heces delos individuos de la especieAllolobophora caliginosa Savigny queen el suelo adyacente a ellas, ensuelos de textura franco-arenosa deuna cuenca experimental de NuevaZelanda. López-Hernández et al.(1993) encontraron, igualmente, másP disponible en las heces dePolypheretima elongata Perrier(Megascolecidae) que en el suelotestigo: un 50% más al ser extraídocon una solución de resina y 30%más al usar una solución de
bicarbonato. El 32P intercambiableextraído en agua fue también mayoren las heces de la lombrizpantropical Pontoscolex corethrurusMüller (Glossoscolecidae) que en elsuelo no ingerido (López-Hernándezet al. 1993); además, dos suelos dediferente capacidad de adsorciónpresentaron, después de ingeridospor P. corethrurus, mayor cantidadde P isotópicamente intercambiableque los suelos empleados comotestigo (López-Hernández et al.1993).
Las lombrices de tierrapertenecen al grupo funcional de losingenieros del ecosistema (sensuJones et al. 1994, 1997; Stork yEggleton 1992) ya que modulan losrecursos disponibles, no sólotróficos sino también físicos,mediante las estructuras queproducen. Con sus actividades deremoción del suelo, de mezcla delsuelo con la hojarasca, y dedeposición de heces en el interior oen la superficie del suelo, laslombrices afectan las propiedadesfísicas del suelo, su ciclo denutrientes y el crecimiento de lasplantas (Lal 1991; Lavelle 1997;Thompson et al. 1993). Martiodriluscarimaguensis Jiménez y Moreno(Glossoscolecidae) es una especie delombriz anécica nativa (Jiménez etal. 1998a) que causa un impactocuantificable en las propiedades delsuelo (Decaëns et al. 1999a,1999b), en el crecimiento de lasplantas y en el banco de semillaspermanente del suelo (Decaëns etal. 2003), y que tiene potencial paraser utilizada en el manejo de losagroecosistemas tropicales (Mariani,información sin publicar). Elimpacto de las lombrices anécicasen el ciclo de nutrientes no ha sidoinvestigado aún plenamente, apesar de que producen grandescantidades de heces frescas sobre lasuperficie del suelo.

226
El Arado Natural:…
El P es un nutriente limitativo enlas sabanas tropicales,especialmente cuando la saturaciónde aluminio del suelo es alta, ya queentonces el P es retenido encompuestos de Al muy estables. Losestudios que se han realizado sobreel papel que los ingenieros delecosistema desempeñan en lassabanas del neotrópico son muyescasos. Los Oxisoles son suelostropicales altamente meteorizadosque se caracterizan por su bajocontenido de P total y de P disponibley, además, por una capacidad deretención de P alta debida a lasaturación de aluminio quepresentan esos suelos (Friesen et al.1997). Existe un gran interés enlograr una mayor eficiencia en ladisponibilidad de P, ya que, porejemplo, sólo el 20% del P totalaplicado como fertilizante a este tipode suelos, en estas regiones, esasimilado por las plantas (Friesen etal. 1997). Estos Oxisoles de pocafertilidad encierran una reserva de Pque no está disponible. Los autoresantes mencionados sugieren que unade las soluciones para incrementar larecuperación de P, desde las formasmenos accesibles, es la siembra devariedades de cultivos y de especiesforrajeras que extraigan el P de unmodo más eficiente. Los autores deeste capítulo creen, respecto a laanterior sugerencia, que el efecto quepuede ejercer la fauna del suelo (enespecial, los ingenieros delecosistema) en el ciclo del P sería unmecanismo importante para liberareste nutriente y lograr que seadisponible para las plantas.
Un estudio previo realizado porGuggenberger et al. (1996) indicó quelas heces de lombrices de la especieanécica M. carimaguensis estabanenriquecidas con formas lábiles de Py concluyeron que esta especie delombriz acumula una reserva de P enel suelo de los pastizales, la cual estádisponible para las plantas. El
objetivo de este estudio fuedeterminar la dinámica temporal delfraccionamiento del P en las heces deindividuos de M. carimaguensis deedades diferentes, recolectadas enlos suelos meteorizados de los LlanosOrientales de Colombia, en donde seestán sembrando cultivos ypastizales para aumentar laproducción agrícola (Thomas et al.1995). Se diseñaron, por tanto,experimentos en condiciones decampo y de laboratorio para estudiarel impacto que causa, en las reservasde P del suelo, la especie M.carimaguensis, cuyos individuosdepositan grandes cantidades deheces sobre la superficie del suelo(Decaëns et al. 1999b; Jiménez et al.1998a, 1998b).
Materiales y Métodos
Sitio de estudioLos experimentos de campo y delaboratorio se realizaron en laestación experimental Carimagua delconvenio CIAT-CORPOICA (4°30’ N,71°19’ O, y 150 msnm), una regiónque representa las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia.La precipitación media anual es de2240 mm y se distribuye desdefinales de marzo hasta principios dediciembre; la temperatura mediaanual es de 26 °C. La vegetaciónnativa varía según la topografía: enlas zonas altas (los “altos”) seencuentran sabanas herbáceasabiertas, y en las zonas bajas (los“bajos”) hay bosques de galeríaasociados con los cursos de agua ycon los ríos y sabanas inundables(o esteros). Los suelos de la estaciónexperimental son Oxisoles biendrenados que se caracterizan por suspropiedades físicas favorables, unasaturación de Al alta (> 80%), y unafertilidad química baja.

Disponibilidad de Fósforo en las Heces...
227
Biología de M. carimaguensisMartiodrilus carimaguensis Jiménez yMoreno (Glossoscolecidae) es unaespecie nativa anécica grande, esdecir, de 9.3 mm de diámetro,194 mm de longitud y 9.2 g de pesofresco (en formol al 4%), enpromedio; esta especie se encuentraen los ecosistemas naturales yperturbados de Carimagua (Jiménezet al. 1998a). La introducción de unpastizal y la acción del ganado que lopastorea generan un aumentoespectacular de las poblaciones deesta especie, que llegan amultiplicarse 10 veces con respecto alas de la sabana nativa (Jiménezet al. 1998b). Los individuos deMartiodrilus carimaguensis seencuentran activos y en la mismagalería semivertical durante, almenos, la primera semana que sigueal comienzo de la deposición de lasheces en la superficie del suelo(Decaëns et al. 1999b; Jiménez et al.1998a). Las heces superficiales deesta especie son de tipo globular(sensu Lee 1985) y estánconstituidas por una deposicióncontinuada, durante varios días, dematerial depositado junto a laentrada de la galería, lo que daorigen a una superposición de capasde diferentes edades. La obra finales una estructura en forma de torre(o sea, un turrículo) que se distinguefácilmente de otras deposicionesencontradas en la superficie delsuelo. Las heces más grandesalcanzan los 15 cm de altura y llegana pesar, en seco, hasta 400 g(Decaëns 2000; Jiménez et al.1998a).
Muestras de campoEl experimento de campo se realizóen parcelas contiguas de sabananativa, donde crecían las especiesAndropogon bicornis L. yTrachypogon vestitus Anders., y deun pastizal de 18 años conformado
por Brachiaria decumbens cv.Basilisk y Pueraria phaseoloides(Roxb.) Benth. CIAT 9900 (el kudzú),desde mayo hasta agosto de 1994.Se tomaron al azar tres áreas de4 x 4 m2 en la sabana y otras tres de1 x 1 m2 en el pastizal. Inicialmentese retiraron todas las heces quehabía sobre la superficie del suelo decada área mencionada y,transcurrido un día, se revisaron lasparcelas antes mencionadas. Seidentificaron las heces frescas reciéndepositadas de M. carimaguensis, sedesplazaron ligeramente a un ladode la entrada de la galería y seidentificaron con etiquetas blancasde plástico para establecer lostiempos de incubación a los que seiban a exponer. Se ensayaron seisperíodos de incubación: 1, 4, 8, 16,32 y 64 días; pasado este tiempo, secombinaron y mezclaron en el campolas heces de cada área que tenían lamisma edad y se introdujeron entermos con hielo para transportarlasal laboratorio. Las áreas demuestreo definidas en el pastizal deB. decumbens y kudzú se protegieroncon jaulas metálicas para evitar elpisoteo del ganado al pastorear elterreno.
Incubación en el laboratorioSe prepararon varios recipientes(potes) con suelo de las parcelasestudiadas recolectado en la capa de0 a 15 cm, secado al aire y tamizado(trama de 2 mm). En el experimentode laboratorio se incluyó untratamiento adicional que simulabalas condiciones de campo: consistíaen mezclar con el suelo de la parcela(a razón de 20 g/kg de suelo)material vegetativo verde obtenido dela misma parcela, previamentesecado al horno y trituradofinamente. Cada recipiente contenía6 kg de suelo, que se llevó a casi lacapacidad de campo (25 g/g; pF 2)5 días antes de introducir en él las

228
El Arado Natural:…
lombrices. Se recolectaron en lasparcelas estudiadas 160 adultos deM. carimaguensis. En cada pote seintrodujeron 12 individuos(suficientes para producir como hecesentre 60 y 100 g de suelo por día)durante un período deacondicionamiento, con el fin deeliminar el suelo que habían ingeridoen el campo. Pasados 6 días, laslombrices se retiraron de los potes deacondicionamiento y se transfirierondirectamente a potes que contenían2 kg de suelo preparado de idénticamanera que en los primeros potes.Transcurrido 1 día, los nuevos potesfueron revisados y las lombricestrasladadas a nuevos potes. Lasheces producidas por estas lombricesde tierra fueron retiradas del suelo ycolocadas en placas Petri sobre papelde filtro húmedo. De este modo semantenía la humedad de lasmuestras en el laboratorio, atemperatura ambiente, durante losperíodos de incubación de 1, 4, 8, 16,32 y 64 días. Todos los tratamientosse replicaron tres veces.
Análisis del PEl P del suelo testigo y el de las hecesincubadas se determinó empleandoun método modificado defraccionamiento de Hedley et al.(1982), en el que se usansucesivamente las siguientessoluciones de extracción agresiva:H2O con resina de intercambioaniónico (H2O-Res.), 0.5 M NaHCO3,0.1 M NaOH, 1 M HCl y HClconcentrado caliente (Tiessen y Moir1993).
- El P inorgánico (Pi) de todos losextractos obtenidos se determinómediante el método de ácidoascórbico-molibdato (sal demolibdeno) (Murphy y Riley1962).
- El P total de la extracciónsucesiva con H2O-Res., NaHCO3 y
NaOH se determinó, previadigestión con persulfato potásico(K2S2O8) (Bowman 1988).
- El P total (de suelo y heces, Pt) yel P residual se determinaronmediante la digestión de lamuestra con ácido perclórico(Olsen y Sommers 1982).
- El P orgánico (Po) se calculó comola diferencia entre el Pt y el Pi.
- La actividad de la fosfatasa ácidase determinó, tanto en el suelotestigo como en las heces, segúnel método de Tabatabai y Bremner(descrito en Alef et al. 1995), queestá basado en la medicióncolorimétrica del p-nitrofenolliberado por la actividadenzimática.
- El P de origen microbiano se midiósegún el método de fumigación yextracción con cloroformo.
- El P microbiano liberado se calculócomo la diferencia entre el Piextraído por Bray-II, de lasmuestras fumigadas y de las nofumigadas.
Análisis estadísticosSe ejecutó un análisis de varianza(ANOVA) de dos vías, en el que seconsideraron el sistema de uso de latierra y la edad de la deposición comoefectos fijos principales. Estos análisisse aplicaron a las fracciones de Porgánico e inorgánico extraídas consoluciones de H2O-Res., de bicarbonatosódico y de NaOH, y también al P total,al C total, al P en Bray-II, a la actividadde la fosfatasa, al P microbiano, y alpH de las muestras. Se hicieron otrascomparaciones de las mediasempleando la prueba de Tukey HSD.Todos los análisis estadísticos serealizaron utilizando el programaStatistica 5.1 bajo entorno Windows(Statsoft Inc. 1996).
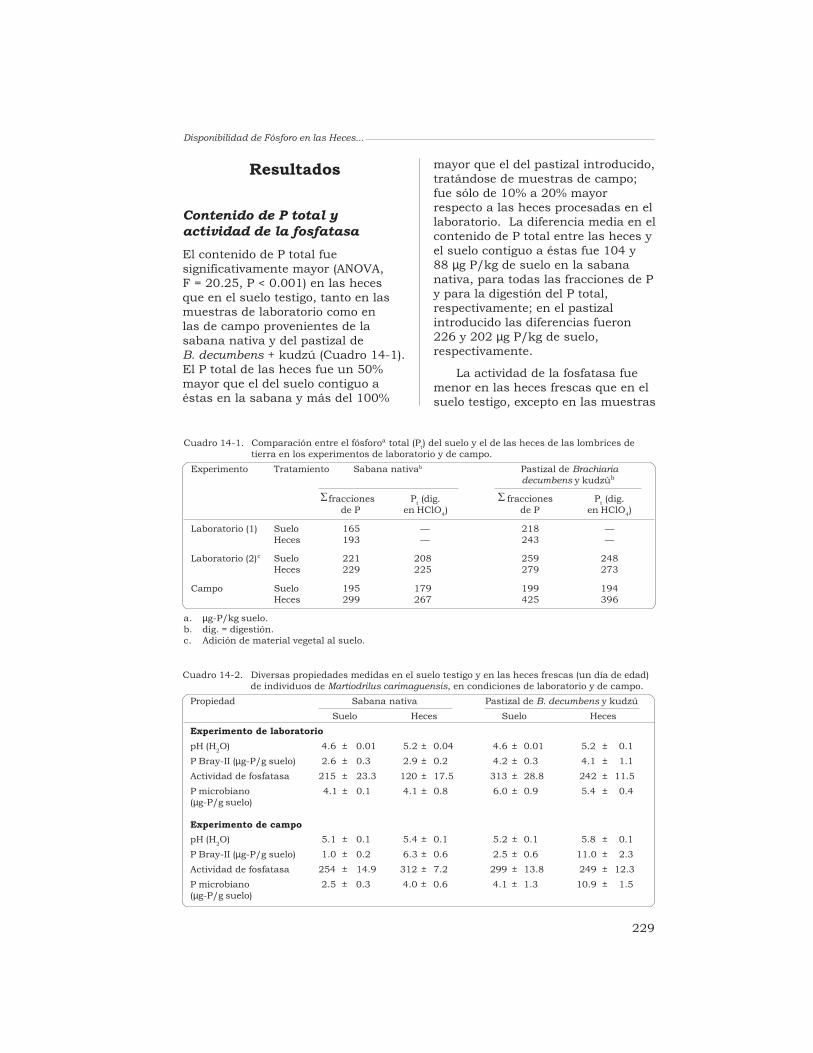
Disponibilidad de Fósforo en las Heces...
229
Cuadro 14-1. Comparación entre el fósforoa total (Pt) del suelo y el de las heces de las lombrices detierra en los experimentos de laboratorio y de campo.
Experimento Tratamiento Sabana nativab Pastizal de Brachiariadecumbens y kudzúb
fracciones Pt (dig. fracciones Pt (dig.de P en HClO4) de P en HClO4)
Laboratorio (1) Suelo 165 — 218 —Heces 193 — 243 —
Laboratorio (2)c Suelo 221 208 259 248Heces 229 225 279 273
Campo Suelo 195 179 199 194Heces 299 267 425 396
a. µg-P/kg suelo.b. dig. = digestión.c. Adición de material vegetal al suelo.
Resultados
Contenido de P total yactividad de la fosfatasaEl contenido de P total fuesignificativamente mayor (ANOVA,F = 20.25, P < 0.001) en las hecesque en el suelo testigo, tanto en lasmuestras de laboratorio como enlas de campo provenientes de lasabana nativa y del pastizal deB. decumbens + kudzú (Cuadro 14-1).El P total de las heces fue un 50%mayor que el del suelo contiguo aéstas en la sabana y más del 100%
mayor que el del pastizal introducido,tratándose de muestras de campo;fue sólo de 10% a 20% mayorrespecto a las heces procesadas en ellaboratorio. La diferencia media en elcontenido de P total entre las heces yel suelo contiguo a éstas fue 104 y88 µg P/kg de suelo en la sabananativa, para todas las fracciones de Py para la digestión del P total,respectivamente; en el pastizalintroducido las diferencias fueron226 y 202 µg P/kg de suelo,respectivamente.
La actividad de la fosfatasa fuemenor en las heces frescas que en elsuelo testigo, excepto en las muestras
Cuadro 14-2. Diversas propiedades medidas en el suelo testigo y en las heces frescas (un día de edad)de individuos de Martiodrilus carimaguensis, en condiciones de laboratorio y de campo.
Propiedad Sabana nativa Pastizal de B. decumbens y kudzúSuelo Heces Suelo Heces
Experimento de laboratoriopH (H2O) 4.6 ± 0.01 5.2 ± 0.04 4.6 ± 0.01 5.2 ± 0.1P Bray-II (µg-P/g suelo) 2.6 ± 0.3 2.9 ± 0.2 4.2 ± 0.3 4.1 ± 1.1Actividad de fosfatasa 215 ± 23.3 120 ± 17.5 313 ± 28.8 242 ± 11.5P microbiano 4.1 ± 0.1 4.1 ± 0.8 6.0 ± 0.9 5.4 ± 0.4(µg-P/g suelo)
Experimento de campopH (H2O) 5.1 ± 0.1 5.4 ± 0.1 5.2 ± 0.1 5.8 ± 0.1P Bray-II (µg-P/g suelo) 1.0 ± 0.2 6.3 ± 0.6 2.5 ± 0.6 11.0 ± 2.3Actividad de fosfatasa 254 ± 14.9 312 ± 7.2 299 ± 13.8 249 ± 12.3P microbiano 2.5 ± 0.3 4.0 ± 0.6 4.1 ± 1.3 10.9 ± 1.5(µg-P/g suelo)
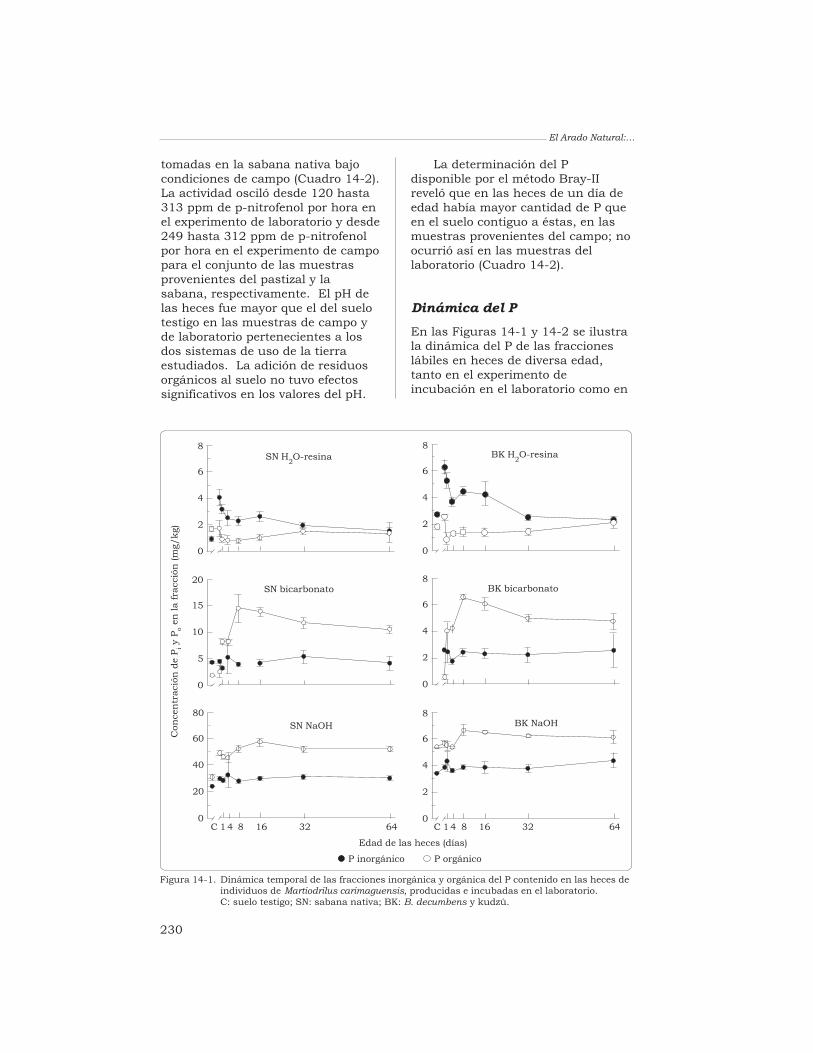
230
El Arado Natural:…
tomadas en la sabana nativa bajocondiciones de campo (Cuadro 14-2).La actividad osciló desde 120 hasta313 ppm de p-nitrofenol por hora enel experimento de laboratorio y desde249 hasta 312 ppm de p-nitrofenolpor hora en el experimento de campopara el conjunto de las muestrasprovenientes del pastizal y lasabana, respectivamente. El pH delas heces fue mayor que el del suelotestigo en las muestras de campo yde laboratorio pertenecientes a losdos sistemas de uso de la tierraestudiados. La adición de residuosorgánicos al suelo no tuvo efectossignificativos en los valores del pH.
La determinación del Pdisponible por el método Bray-IIreveló que en las heces de un día deedad había mayor cantidad de P queen el suelo contiguo a éstas, en lasmuestras provenientes del campo; noocurrió así en las muestras dellaboratorio (Cuadro 14-2).
Dinámica del PEn las Figuras 14-1 y 14-2 se ilustrala dinámica del P de las fraccioneslábiles en heces de diversa edad,tanto en el experimento deincubación en el laboratorio como en
8
6
4
2
0
20
15
10
5
0
80
60
40
20
0
8
6
4
2
0
8
6
4
2
0
8
6
4
2
0
Edad de las heces (días)
SN H2O-resina BK H2O-resina
SN NaOH BK NaOH
C 1 4 8 16 32 64
SN bicarbonato BK bicarbonato
P inorgánico P orgánico
Con
cen
trac
ión
de
P i y P
o en
la fr
acci
ón (m
g/kg
)
Figura 14-1. Dinámica temporal de las fracciones inorgánica y orgánica del P contenido en las heces deindividuos de Martiodrilus carimaguensis, producidas e incubadas en el laboratorio.C: suelo testigo; SN: sabana nativa; BK: B. decumbens y kudzú.
C 1 4 8 16 32 64
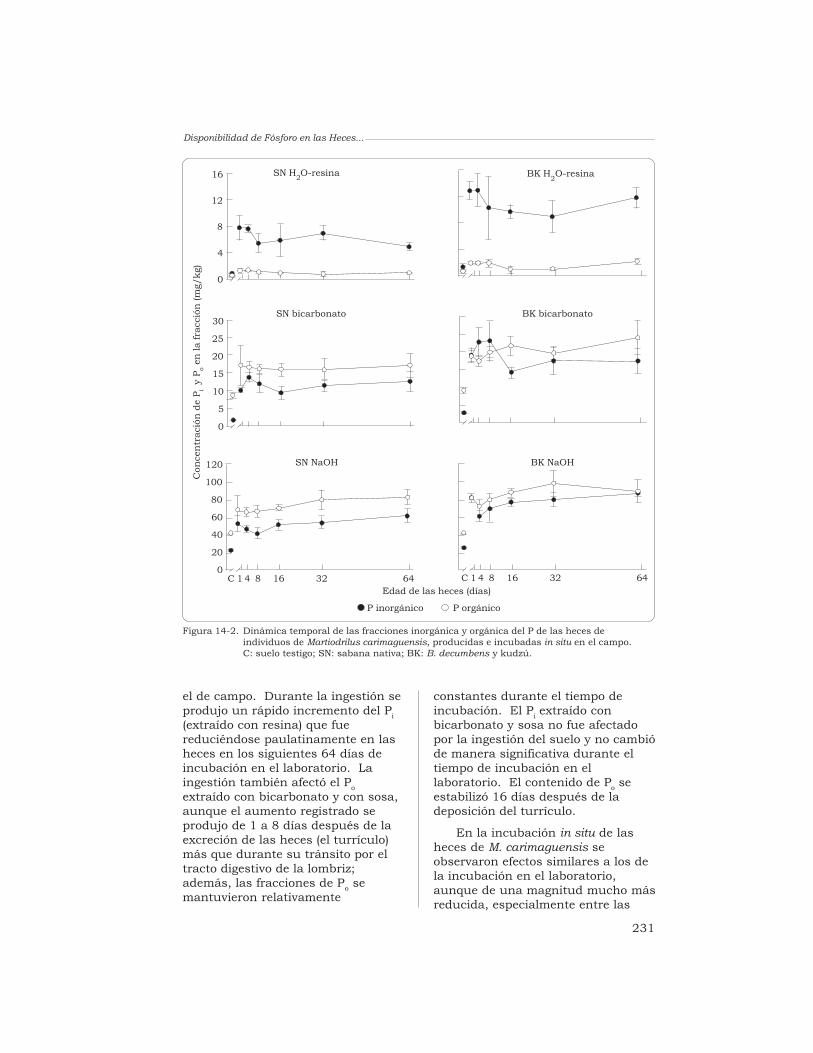
Disponibilidad de Fósforo en las Heces...
231
Figura 14-2. Dinámica temporal de las fracciones inorgánica y orgánica del P de las heces deindividuos de Martiodrilus carimaguensis, producidas e incubadas in situ en el campo.C: suelo testigo; SN: sabana nativa; BK: B. decumbens y kudzú.
el de campo. Durante la ingestión seprodujo un rápido incremento del Pi(extraído con resina) que fuereduciéndose paulatinamente en lasheces en los siguientes 64 días deincubación en el laboratorio. Laingestión también afectó el Poextraído con bicarbonato y con sosa,aunque el aumento registrado seprodujo de 1 a 8 días después de laexcreción de las heces (el turrículo)más que durante su tránsito por eltracto digestivo de la lombriz;además, las fracciones de Po semantuvieron relativamente
constantes durante el tiempo deincubación. El Pi extraído conbicarbonato y sosa no fue afectadopor la ingestión del suelo y no cambióde manera significativa durante eltiempo de incubación en ellaboratorio. El contenido de Po seestabilizó 16 días después de ladeposición del turrículo.
En la incubación in situ de lasheces de M. carimaguensis seobservaron efectos similares a los dela incubación en el laboratorio,aunque de una magnitud mucho másreducida, especialmente entre las
SN H2O-resina BK H2O-resina
SN NaOH BK NaOH
SN bicarbonato BK bicarbonato
16
12
8
4
0
Con
cen
trac
ión
de
P i y
P o en
la fr
acci
ón (m
g/kg
)
Edad de las heces (días)C 1 4 8 16 32 64
P inorgánico P orgánico
C 1 4 8 16 32 64
30
25
20
15
10
5
0
120
100
80
60
40
20
0

232
El Arado Natural:…
fracciones de Po (Figura 14-2). Tantoel Pi como el Po extraídos conbicarbonato y sosa aumentaron enlas heces de las lombrices alcomparar a éstas con el suelo; en ellaboratorio, en cambio, sólo el Poaumentó. Sin embargo, tras elaumento inicial, el Pi y el Popermanecieron relativamenteconstantes en todas las fraccionesdurante los 64 días de incubación.
En el ensayo de laboratorio, lasfracciones de P de las heces queaumentaron en comparación con lasdel suelo testigo fueron sólo lassiguientes: el Pi extraído con resinay, en menor grado, el Po extraído conH2O de la solución extractora y lasfracciones de Pi y de Po extraidas conNaOH. El incremento del P total fuetambién marginal. En términosrelativos, las heces depositadasafectaron poco las reservas de P, aexcepción del aumento ocurridoen la fracción de Pi extraído con laresina de la solución extractora(Cuadro 14-3).
Discusión
Con el fin de valorar la contribuciónde la fauna del suelo alfuncionamiento de éste y a ladinámica de los ecosistemas, esnecesario hacer una descripción delos procesos más importantes quetienen lugar en las heces (Martin yMarinissen 1993). Este estudioconfirma los resultados obtenidospor varios autores, quienes hanenfatizado la importancia de laslombrices de tierra tanto en el ciclode nutrientes como en ladisponibilidad del P en losecosistemas de pastizal (Barois yLavelle 1986; Barois et al. 1987;James 1991; López-Hernández et al.1993).
Cumplido el tránsito intestinal,se produce un aumento significativo
de P en las heces de P. corethrurus; ladiferencia más grande se observó enel P extraído con H2O, que se duplicódespués de la ingestión del suelo porlas lombrices (Barois et al. 1999). Laestabilización de los valores de P enlas heces de la lombriz endógea P.corethrurus se produce 4 díasdespués de la deposición de éstas(López-Hernández et al. 1993),mientras que en las de M.carimaguensis la estabilizaciónocurre de 16 a 64 días más tarde.
No se ha aclarado aún el origendel P incorporado en las heces deM. carimaguensis, ya que puedeprovenir o bien de la hojarasca o defragmentos de raíces y partes defollaje descompuestos o de laingestión de heces de otraslombrices, como se ha explicadorecientemente (Mariani et al. 2001).Este P, que procede de cualquierfuente orgánica, fue incluido entodas las fracciones de P (orgánico einorgánico) y en mayor medida en lasfracciones lábiles de este nutriente.No es posible concluir que la especiede lombriz aquí estudiada propicia lamineralización del P orgánico, ya quela relación proporcional del P lábil enel sustrato puede haber sido mayorque la de las formas estables de P.El hecho de que, en condiciones delaboratorio en que no se aportanresiduos orgánicos al suelo, lasreservas relativas de P no fueronafectadas, apoya la interpretacióndada. Sin embargo, el aumentorelativo del Pi extraído con resina(una reserva pequeña de P) sugierealgún tipo de mineralización del Pmenos disponible que proviene dereservas grandes, en donde loscambios relativos serían difíciles dedetectar. El modo en que fuerontratados los residuos orgánicos, esdecir, secados al horno y luegotriturados, pudo haber influido en laausencia de diferencias significativasen los tratamientos en que se
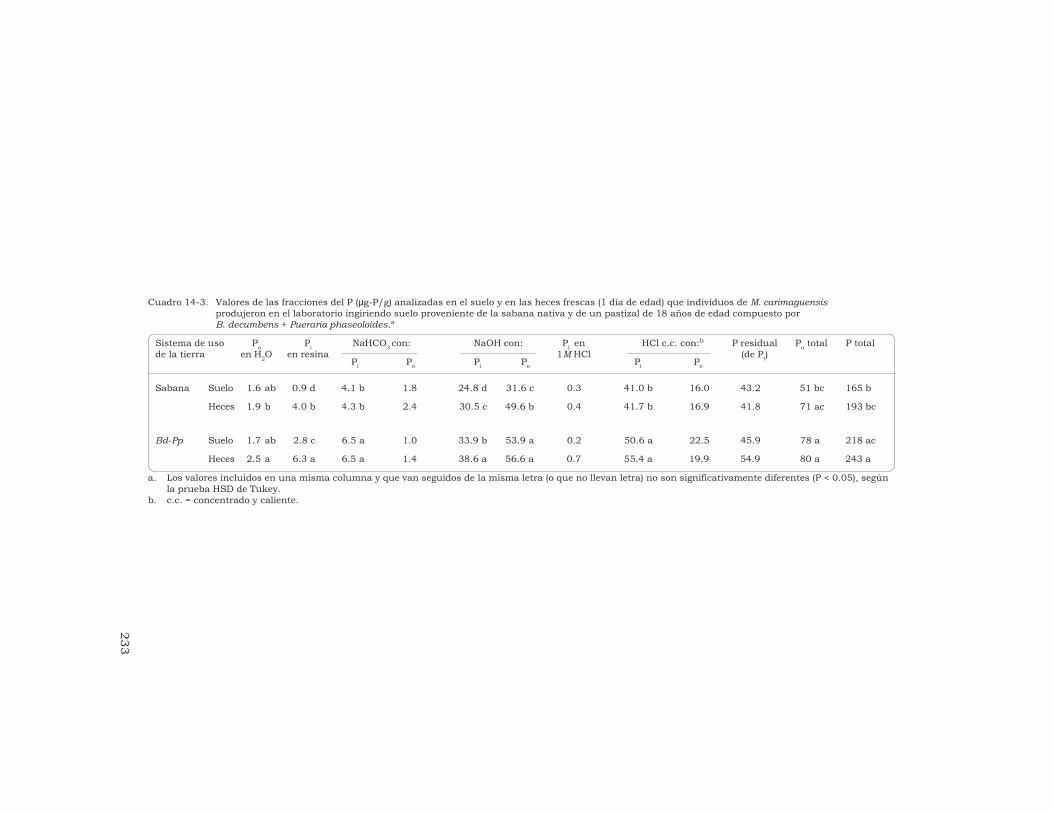
Disponibilidad de Fósforo en las H
eces...
233
Cuadro 14-3. Valores de las fracciones del P (µg-P/g) analizadas en el suelo y en las heces frescas (1 día de edad) que individuos de M. carimaguensisprodujeron en el laboratorio ingiriendo suelo proveniente de la sabana nativa y de un pastizal de 18 años de edad compuesto porB. decumbens + Pueraria phaseoloides.a
Sistema de uso Po Pi NaHCO3 con: NaOH con: Pi en HCl c.c. con:b P residual Po total P totalde la tierra en H2O en resina 1M HCl (de Pt)Pi Po Pi Po Pi Po
Sabana Suelo 1.6 ab 0.9 d 4.1 b 1.8 24.8 d 31.6 c 0.3 41.0 b 16.0 43.2 51 bc 165 b
Heces 1.9 b 4.0 b 4.3 b 2.4 30.5 c 49.6 b 0.4 41.7 b 16.9 41.8 71 ac 193 bc
Bd-Pp Suelo 1.7 ab 2.8 c 6.5 a 1.0 33.9 b 53.9 a 0.2 50.6 a 22.5 45.9 78 a 218 ac
Heces 2.5 a 6.3 a 6.5 a 1.4 38.6 a 56.6 a 0.7 55.4 a 19.9 54.9 80 a 243 a
a. Los valores incluidos en una misma columna y que van seguidos de la misma letra (o que no llevan letra) no son significativamente diferentes (P < 0.05), segúnla prueba HSD de Tukey.
b. c.c. = concentrado y caliente.

234
El Arado Natural:…
comparaba la adición de residuosorgánicos al suelo frente a la noadición de éstos. Por otro lado, elaumento de P en las fraccioneslábiles extraídas con agua y sosapuede explicarse por la selección dela MO y la reorganización otranslocación de P desde las reservasestables a formas disponibles paralas plantas.
El contenido alto de P en lasheces de M. carimaguensis se obtuvoen muestras de campo provenientesdel pastizal de B. decumbens y P.phaseoloides. Este resultado pudodeberse o bien a la presencia de laleguminosa en la asociación, la cualcontribuye a la fijación de Natmosférico y, por ende, a una mayoractividad biológica (Oberson et al.1995; Thomas 1992, 1995; Thomaset al. 1992), o bien a ladescomposición de las deposicionesdel ganado, que son introducidas enla parte mineral del suelo por algunoscoleópteros de la familia Scarabeidaey por las lombrices (Jiménez et al.1998a, 1998b) o a la fertilización delpastizal (Thomas et al. 1995).
Las heces superficiales (losturrículos) de M. carimaguensisrepresentan una fuente significativadel P disponible directamente por lasplantas, de modo similar al modelopropuesto por Decaëns et al (1999b)para la difusión del N. Este modeloconsistiría en que el P está primerodisponible en las heces frescas yenseguida se difunde rápidamente,tanto en sentido vertical comolateralmente dentro de las galerías delas lombrices y por la superficie delas heces durante el tiempo en queéstas envejecen. Este P seríaabsorbido por las raíces localizadascerca de la superficie del suelo, lo quepodría relacionarse también con elaumento de la biomasa de las raícesque se encuentran bajo las heces delas lombrices de tierra (Decaëns et al.1999b). Un nuevo estudio debería
considerar la diponibilidad del Po queestá protegido físicamente en lasheces de más edad de las lombrices deM. carimaguensis.
La actividad de los ingenieros delecosistema (sensu Jones et al. 1994,1997) influye en la cantidad y en ladistribución del P en los suelos desabana y controla esas dos variables,mediante la construcción de nidos ygalerías (López-Hernández et al.1989). La actividad de las lombricesde tierra hace más disponible el P decarácter mineral que proviene de lahojarasca o del suelo (Mansell et al.1981); Anderson y Wood (1984)encontraron que las termitas que sealimentan de suelo en los bosquestropicales de Africa actúan del mismomodo. Son escasos los estudios sobreel papel de los ingenieros delecosistema en el ciclo de nutrientes yno es posible, por ello, establecerpatrones generales dentro de lostaxones más importantes de la faunadel suelo y entre ellos. Enconsecuencia, los estudios que serealicen en el futuro deben considerarlos efectos que causa este grupofuncional en la dinámica de losnutrientes; por ejemplo, lasestructuras creadas porMicrocerotermes sp., Spinitermes sp.(Termitinae) y Velocitermes sp.(Nasutitermitinae) o por Atta laevigata,Acromyrmex sp. y Trachymyrmex sp.(Decaëns et al. 2001).
Partiendo de los resultadosobtenidos en este estudio, se puedesugerir que M. carimaguensis participaen la mineralización del P orgánicodisponible, lo que concuerda con losresultados de Guggenberger et al.(1996), quienes mostraron que laactividad de las lombrices deCarimagua causaba serios efectos enla disponibilidad del P del suelo.Estos autores sugirieron, basándoseen Lavelle y Martin (1992), que laacumulación de la reserva del Porgánico lábil se debe a que las heces

Disponibilidad de Fósforo en las Heces...
235
se estabilizan, en términos físicos, enescalas de tiempo cortas. Porconsiguiente, la actividad de losindividuos de M. carimaguensis en lasabana nativa y en el pastizalintroducido desempeña, al parecer, unpapel importante, por dos razones:incrementa la reserva activa del P, ymejora y mantiene la disponibilidaddel P en los Oxisoles de los LlanosOrientales de Colombia.
AgradecimientosLos autores agradecen al personal dela estación experimental de Carimaguapor su inestimable ayuda en el trabajode campo y por su amistad.Agradecen también a Gonzalo Borrero,del CIAT, por su ayuda en ladeterminación de las fracciones de Pen el laboratorio y a Alex Feijoo (CIAT),Patrick Lavelle (IRD, Francia) y MichelBrossard (IRD, Brasil) por suscomentarios útiles. El primer autoragradece profundamente a RichardThomas y a los miembros de la Unidadde Suelos y Nutrición de Plantas(CIAT) por sus críticas constructivas ala primera versión del manuscrito.
ReferenciasAlef K; Nannipieri P; Trazar-Cepeda C.
1995. Phosphatase activity. En: AlefK; Nannipieri P, eds. Methods inapplied soil microbiology andbiochemistry. Academic Press,Londres. p 335-344.
Anderson JM; Wood TG. 1984. Moundcomposition and soil modification bytwo soil-feeding termites (Termitinae,Termitidae) in a riparian Nigerianforest. Pedobiologia 26:77-82.
Barois I; Lavelle P. 1986. Changes inrespiration rates and somephysicochemical properties of atropical soil during transit throughPontoscolex corethrurus(Glossoscolecidae, Oligochaeta). SoilBiol Biochem 18:539-541.
Barois I; Verdier B; Kaiser P; Lavelle P;Rangel P. 1987. Influence of thetropical earthworm Pontoscolexcorethrurus (Glossoscolecidae) on thefixation and mineralization ofnitrogen. En: Bovincini Pagliai AM;Omodeo P, eds. On earthworms;vol. 2: Selected symposia andmonographs, UZI. Mucchi, Módena,Italia. p 151-158.
Barois I; Lavelle P; Brossard M; Tondoh J;Martínez MA; Rossi JP; Senapati BK;Angeles A; Fragoso C; Jiménez JJ;Decaëns T; Lattaud C; Kanyonyo J;Blanchart E; Chapuis L; Brown GG;Moreno A. 1999. Ecology ofearthworm species with largeenvironmental tolerance and/orextended distributions. En: Lavelle P;Brussaard L; Hendrix PF, eds.Earthworm management in tropicalagroecosystems. CAB International,Wallingford, Reino Unido. p 57-85.
Bowman RA. 1988. A rapid method todetermine total phosporous in soils.Soil Sci Soc Am J 52:1301-1304.
Brossard M; Lavelle P; Laurent JY. 1996.Digestion of a vertisol by the endogeicearthworm Polypheretima elongata(Megascolecidae) increases soilphosphate extractability. Eur J SoilBiol 32:107-111.
Chapuis L; Brossard M. 1995. Modificationset stabilité du phosphore échangeabled’un ferrasol ingeré par un vergéophage. C R Acad Sci Ser IIa320:587-592.
Decaëns T. 2000. Degradation dynamics ofsurface earthworm casts in grasslandsof the Eastern Plains of Colombia. BiolFertil Soils 32:149-156.
Decaëns T; Galvis JH; Amézquita E. 2001.Propriétés des structures produitespar les ingénieurs écologiques à lasurface du sol d’une savanecolombienne. C R Acad Sci Ser III, SciVie 324:465-478.
Decaëns T; Mariani L; Lavelle P. 1999a. Soilsurface macrofaunal communitiesassociated with earthworm casts ingrasslands of the Eastern Plains ofColombia. Appl Soil Ecol 13:87-100.

236
El Arado Natural:…
Decaëns T; Rangel AF; Asakawa N;Thomas RJ; Lavelle P. 1999b.Carbon and nitrogen dynamics inageing earthworm casts ingrasslands of the Eastern plains ofColombia. Biol Fertil Soils30:20-28.
Decaëns T; Mariani L; Betancourt N;Jiménez JJ. 2003. Seed dispersionby surface casting activities ofearthworms in Colombiangrasslands. Acta Oecol24(4):175-185.
Friesen DK; Rao IM; Thomas RJ; ObersonA; Sanz JI. 1997. Phosphorousacquisition and cycling in crop andpasture systems in low fertilitytropical soils. En: Ando T; Fujita K;Mae T; Matsumoto H; Mori S;Sekiya J, eds. Plant nutrition forsustainable food production andenvironment. Kluwer AcademicPublishers, Japón. p 493-498.
Guggenberger G; Haumaier L; ThomasRJ; Zech W. 1996. Assesing theorganic phosphorous status of anOxisol under tropical pasturesfollowing native savanna using 31PNMR spectroscopy. Biol Fertil Soils23:332-339.
Hedley MJ; Stewart JWB; Chauhan BS.1982. Changes in inorganic andorganic soil phosphorous fractionsinduced by cultivation practicesand by laboratory incubations. SoilSci Soc Am J 46:970-976.
James SW. 1991. Soil, nitrogen,phosphorous and organic matterprocessing by earthworms intallgrass prairie. Ecology72:2101-2109.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innative savanna and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78:1946-1957.
Krishnamoorthy RV. 1990. Mineralizationof phosphorous by faecalphosphatases of some earthwormsof Indian tropics. Proc Ind Acad SciAnim Sci 95:341-351.
Lal R. 1974. No tillage effects on soilproperties and maize (Zea mays L.)production in western Nigeria. PlantSoil 40:321-331.
Lal R. 1991. Soil conservation andbiodiversity. En: Hawksworth DL,ed. The biodiversity ofmicrorganisms and invertebrates:its role in sustainable agriculture.CAB International, Wallingford,Reino Unido. p 89-103.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humidtropics. Soil Biol Biochem24(12):1491-1498.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B; Brussard L. 1994.The relationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. Wiley, ReinoUnido. p 137-169.

Disponibilidad de Fósforo en las Heces...
237
Lee KE. 1985. Earthworms: their ecologyand relationships with soil and landuse. Academic Press, Nueva York,Estados Unidos. 411 p.
López-Hernández D; Lavelle P; FardeauJC; Niño M. 1993. Phosphoroustransformations in two P-sorptioncontrasting tropical soils duringtransit through Pontoscolexcorethrurus (Glossoscolecidae:Oligochaeta). Soil Biol Biochem25(6):789-792.
López-Hernández D; Fardeau JC; Niño M;Nannipieri P; Chacón P. 1989.Phosphorous accumulation insavanna termite mounds inVenezuela. J Soil Sci 40:635-640.
Lunt HA; Jacobson GM 1944. Thechemical composition of earthwormcasts. Soil Sci 58:367.
Mansell GP; Syers JK; Gregg PEH. 1981.Plant availability of phosphorous indead herbage ingested by surface-casting earthworms. Soil BiolBiochem 13:163-167.
Mariani L; Bernier N; Jiménez JJ; DecaënsT. 2001. Régime alimentarie d’un verde terre des savanes colombiennes:une remise en question des typesécologiques. C R Acad Sci Ser III. SciVie 324:733-742.
Martin A; Marinissen JYC. 1993.Biological and physico-chemicalprocesses in excrements of soilanimals. Geoderma 56:331-347.
Murphy J; Riley JP. 1962. A modifiedsingle solution method for thedetermination of phosphate innatural waters. Anal Chim Acta27:31-36.
Nye PH. 1955. Some soil-formingprocesses in the humid tropics; IV:The action of soil fauna. J Soil Sci6(1):73-83.
Olsen SR; Sommers LE. 1982.Phosphorous. En: Page AL; MillerRH; Keeney DR, eds. Methods of soilanalysis; Parte 2. 2a. ed. ASA,Madison, WI, Estados Unidos.p 404-430.
Oberson A; Friesen DK; Tiessen H; MoirJO; Borrero G. 1995. Phosphoroustransformations in improvedpastures. En: Tropical LowlandsProgram annual report 1994.Working document no. 148. CIAT,Cali, Colombia. p 182-187.
Sharpley AN; Syers JK. 1976. Potentialrole of earthworm casts for thephosphorous enrichment of run-offwaters. Soil Biol Biochem 8:341-346.
Stork NE; Eggleton P. 1992. Invertebratesas determinants and indicators ofsoil quality. Am J Alt Agric 7:38-47.
Thomas RJ. 1992. The role of the legumein the nitrogen cycle of productiveand sustainable pastures. GrassForage Sci 47:133-142.
Thomas RJ. 1995. Role of legumes inproviding N for sustainable tropicalpasture systems. Plant Soil174:103-118.
Thomas RJ; Fisher MJ; Ayarza MA; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA, eds.Soil management: experimentalbasis for sustainability andenvironmental quality. Advances insoil science series. Lewis Publishers,Boca Ratón, FL, Estados Unidos.p 61-83.
Thomas RJ; Lascano C; Sanz JI; Ara MA;Spain JM; Vera RR; Fisher MJ. 1992.The role of pastures in productionsystems. En: CIAT. Pastures for thetropical lowlands: CIAT’scontributions. Cali, Colombia.p 121-144.
Thompson L; Thomas CD; Radley JMA;Williamson S; Lawton JH. 1993. Theeffect of earthworms and snails in asimple plant community. Oecologia95:171-178.
Tiessen H; Moir JO 1993. Characterizationof available P by sequentialextraction. En: Carter MR, ed. Soilsampling and methods of analysis.Lewis Publishers, Boca Ratón, FL,Estados Unidos. p 75-86.

238
El Arado Natural:…
ResumenSe investigaron tanto la estabilidadestructural como la composición dela materia orgánica (MO) del sueloasociada con agregados de diferentestamaños presentes en las hecesgrandes y redondeadas quedepositan, en la superficie del suelo,las lombrices de tierra de hábitoanécico de la especie Martiodrilus sp.(familia Glossoscolecidae). Secuantificaron tanto el contenido delignina (por oxidación con CuO)como el de los carbohidratos(hidrólisis ácida) de la MO y serealizó, además, una espectroscopíade resonancia magnética nuclear(RMN) con 13C. Los datos obtenidospara las heces se compararon conlos del suelo superficial contiguo aellas, que provenía de un pastizal degramínea y leguminosa establecidoen una sabana nativa de sueloOxisol. Las heces de las lombricescontenían mayor cantidad deagregados estables de gran tamaño
(de 3.15 a 5 mm y de 5 a 8 mm) quesu suelo circundante (765 g/kg vs.390 g/kg). Esta mayor estabilidadestructural de las heces se debe a sumás alta concentración de Corgánico (56 g C/kg suelo ) encomparación con el suelo que lascircunda (26 g C/kg suelo). Esteincremento fue muy notorio para el Corgánico asociado con los agregadosde la fracción arenosa, es decir, parala MO que no participa en laformación de complejos órgano-minerales. Los indicios obtenidosrespecto a la composición de loscarbohidratos y a la alteraciónmicrobiana de la lignina, así como laespectroscopía de RMN con 13C,revelaron que la MO asociada a lafracción arenosa estaba compuesta,principalmente, por residuosvegetales ligeramentedescompuestos. Por tanto, laconcentración elevada de lignina y decarbohidratos (recursos primarios) yla relación proporcional baja de Caromático sustituido y de grupos–COOH (ambos en compuestoshúmicos) que se observaron en lasheces —al compararlas con el suelocircundante no fraccionado— sedeben, en parte, a una mezcla muyestrecha de hojarasca fresca consuelo mineral (efecto anécico). LaMO del suelo asociada a la fracciónarcillosa era más abundante en losresiduos microbianos, y los
CAPÍTULO 15
Materia Orgánica del Suelo Contenidaen las Heces que Produce unaComunidad de Lombrices Anécico-Endógea de un Pastizal Tropical, enColombia1
G. Guggenberger* , R. J. Thomas** y W. Zech*
1. Publicado en Applied Soil Ecology 3,p 263-374, por Guggenberger et al.Copyright © 1996 de Elsevier. Impreso conpermiso de la casa editorial.
* Institute of Soil Science and Soil Geography,University of Bayreuth, D-95440, Bayreuth,Alemania.
** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.

Materia Orgánica del Suelo...
239
compuestos orgánicos humificadosdominaron la MO asociada con lafracción de limos. Sin embargo, losanálisis químicos de carácterestructural indicaron que la MOunida a partículas de arcilla y delimo parecía hallarse en un estado dedescomposición más avanzado en elsuelo contiguo a las heces que enéstas.
Los datos obtenidosproporcionaron pruebas claras deque los residuos vegetales ricos encarbohidratos, junto con losmucopolisacáridos sintetizados pormicroorganismos que viven en elintestino de la lombriz y por laspropias lombrices, son losresponsables de la estabilidadestructural observada en las heces.Según Golchin et al. (1994), se hapropuesto la hipótesis de que elmetabolismo microbiano de loscarbohidratos de origen vegetalconduce a la liberación de mucílagoy de ciertos metabolitos, los cualespermean el revestimiento de laspartículas minerales y, de este modo,estabilizan las heces. Laconsecuencia de esta íntimaasociación entre los residuosvegetales ligeramente descompuestosy la fase mineral del suelo es laacumulación de una reserva de Cque, aunque físicamente protegida,está activa y se libera en el momentoen que se desintegran las heces.
Palabras clave: dinámica de lamateria orgánica del suelo, hecesde lombrices, pastizal, protecciónfísica, reserva de C
IntroducciónLos Llanos Orientales de Colombia secaracterizan por la presencia desuelos ácidos de fertilidad baja(Oxisoles). Las mayores limitantesdel crecimiento vegetal son la acidez(por niveles tóxicos de Al) y el bajo
contenido de P disponible y de Nmineral (Rao et al. 1993). Lafertilización de estos suelos y laintroducción de gramíneas yleguminosas forrajeras producen unaumento de la MO del suelo (Fisheret al. 1994), tasas más altas demineralización del N (Rao et al. 1994)y un ciclo del P mucho más eficiente(Oberson, datos sin publicar). Estosefectos benéficos que los pastizalesejercen en los parámetros químicosde los suelos mencionados serelacionan con la calidad de lahojarasca de las gramíneas y de lasleguminosas que los conforman(Thomas y Asakawa 1993), con lacalidad de la MO en cuanto a sudegradabilidad (Guggenberger et al.1995a), y con la actividad de laslombrices (Decaëns et al. 1994).
Las lombrices pueden influirmucho en la regulación de ladinámica de la MO de los suelostropicales (Anderson y Flanagan1989). Las actividades drilosféricas(la drilosfera es la parte del suelo querecibe la influencia de las lombricesde tierra) pueden ser considerables sise considera la cantidad de suelo yde MO que transita, anualmente, porel intestino de las lombrices. Lavelley Martin (1992) mostraron que lacomunidad de lombrices que habitaen las sabanas africanas (unos 350 a500 kg/ha, en peso fresco) ingeríaentre 800 y 1250 Mg/ha de sueloseco al año, que equivalen a unos14 ó 15 Mg de MO. En los pastizalestropicales se encontraron valoressimilares (Decaëns et al. 1994).Estos datos muestran, hasta ciertopunto, la influencia de las lombricesde tierra en la dinámica de la MO delos suelos tropicales; no obstante,apenas existe información sobre losefectos de la actividad de dichaslombrices en la composición químicade la MO.
El fraccionamiento del suelo enpartículas de diferente tamaño

240
El Arado Natural:…
(arenas, limos y arcillas) da origen afracciones órgano-minerales, depropiedades bien diferenciadas en loque respecta al ciclo de la MO(Anderson y Paul 1984; Tiessen yStewart 1983). La MO asociada conla fracción arenosa del suelo esimportante en el ciclo que aquéllacumple a corto plazo, la que estáunida a la fracción arcillosa dominaen el ciclo a mediano plazo, y la queestá unida a la fracción de los limosparticipa en el ciclo a largo plazo. Porconsiguiente, la separación de la MOsegún el tamaño de las partículas esuna herramienta empleada paraevaluar las fracciones lábiles ypasivas de esa MO (Christensen1992). Guggenberger et al. (1994)señalaron que el fraccionamientofísico del suelo según el tamaño desus partículas, unido a las técnicasquímicas degradativas, ofrece unpotencial extraordinario para evaluarlos cambios que ocurran en lacomposición de la MO.
Aunque las técnicas químicasdegradativas son útiles paraestablecer la naturaleza de losrecursos primarios y secundarios dela MO, fallan, generalmente, en lacaracterización de la estructuracompleja de los compuestos estableshumificados de la MO (Kögel-Knabneret al. 1988). En cambio, laespectroscopía no invasiva en estadosólido que emplea la RMN con 13C yCPMAS (“cross-polarization magic-angle spinning”), sirve para calcularla estructura química básica de la MOen las muestras de suelo completo(Kögel-Knabner et al. 1988) y en lasfracciones que de éste se obtienensegún el tamaño de sus partículas(Baldock et al. 1992). Laespectroscopía de RMN con 13C ensolución está diseñada parainvestigar la fracción extraíble de laMO del suelo (Preston 1987).
Un objetivo de este estudio fueobtener información sobre los efectos
de la actividad de las lombrices detierra en el ciclo del C en el suelo.Con el fin de estudiar la distribuciónde la MO y su composiciónestructural, tanto en las hecesproducidas por una comunidad delombrices de la sabana como en elsuelo que las circundaba, sehicieron, adoptando un enfoqueintegral, el fraccionamiento del suelosegún el tamaño de sus partículas yla caracterización química de la MOdel suelo mediante métodos bien seadestructivos (oxidación de la ligninacon CuO e hidrólisis ácida de loscarbohidratos) o bien no destructivosmediante la espectroscopía de RMNcon 13C. Se relacionaron estosresultados con la estabilidadestructural de las heces,determinada mediante el tamizadoen húmedo, y se discutieron lasposibles interacciones entre la MOdel suelo y la fase mineral de éste.
Materiales y Métodos
Descripción del sitioEl área de estudio se encuentra en elCentro Nacional de InvestigacionesAgropecuarias (CNIA) Carimagua, enColombia (4°37’ N, 71°19’ O,175 msnm), que corresponde a lassabanas isohipertérmicas biendrenadas. La temperatura mediaanual es de 26 °C y la precipitaciónmedia anual es de 2200 mm que sedistribuye, principalmente, de abril adiciembre. Los suelos son, en sumayoría, Oxisoles de baja fertilidadcaracterizados por un pH bajo (4.5,H2O), una alta saturación de Al, yniveles bajos de los cationesintercambiables de Ca, Mg y K(Lascano y Estrada 1989).
En 1978 se sembraron, enasociación, una gramínea forrajeratropical, Brachiaria decumbens cv.Basilisk y una leguminosa tropical,

Materia Orgánica del Suelo...
241
Pueraria phaseoloides CIAT 9900(kudzú), donde había antes unasabana nativa. La combinación deuna gramínea de raíces profundascon una leguminosa fijadora de Nparece incrementar el reciclaje denutrientes, la producción animal, laactividad biológica en el suelo y elalmacenamiento de C (Fisher et al.1994; Thomas et al. 1995).
En la época lluviosa seobservaron algunas heces delombrices de tierra sobre lasuperficie del suelo en la sabananativa, y una cantidad abundante deheces en el suelo del pastizal degramínea y leguminosa; esto indicaque tras el establecimiento delpastizal, la comunidad de lombricescambió y sus poblacionesaumentaron. Jiménez et al. (1994)identificaron 15 especies delombrices de tierra, algunas noconocidas hasta entonces niclasificadas a nivel ecológico; lasespecies dominantes fueron laanécica Martiodrilus n. sp., de lafamilia Glossoscolecidae y laendoanécica Andiorrhinus n. sp.Como la mayoría de las heces erande tipo globular (Lavelle y Martin1992) y de gran tamaño (2 a 8 cm dediámetro), es probable queprovinieran, en su mayoría, deMartiodrilus n. sp.
MuestreoSe recogieron aleatoriamente unas20 heces superficiales del pastizal de1 ha en julio de 1993. Para hacercomparaciones, se tomaronmanualmente, en la parcela de 1 ha,cuatro réplicas del suelo quecircundaba las heces y que no habíasido ingerido recientemente; se evitael término “no ingerido” porque, dadala intensa actividad de las lombricesde tierra, todo el suelo habrá sidoreingerido en unos pocos años. Enel laboratorio, las muestras se
mezclaron y de ese conjunto seretiraron los restos visibles de raíces.Posteriormente, las muestras sedejaron secar al aire y, excepto en ladeterminación de la estabilidad delos agregados, se tamizaron a 2 mmde trama y se secaron de nuevo a40 ºC durante 5 días.
Análisis de laboratorio
Estabilidad de losagregadosLa estabilidad de los agregados
de las heces y del suelo que lascircunda se determinó por triplicadomediante el tamizado en húmedo demuestras secadas al aire yremojadas en un aparato Yoder(Beare y Bruce 1993). Se tomaronagregados secos porque tanto lasheces superficiales como la superficiedel suelo están expuestos,normalmente, a la desecación y alrehumedecimiento. Los agregadosmenores de 8 mm se tamizaron auna frecuencia de 35 ciclos/mindurante 5 min. La trama de lostamices empleados era de 5, 3.15, 2,1, 0.5 y 0.2 mm.
Separación según eltamaño de las partículasSiguiendo el método de
Christensen (1985), la separaciónsegún el tamaño de las partículas sehizo por triplicado, en muestras desuelo secado a 40 °C durante 5 días.Se dispersaron mediante ultrasonido30 g de suelo en 150 ml de aguaempleando un desintegrador del tiposonda (Heat Systems-Ultrasonics,Model W 185-F); el total de energíadisipada fue de 1500 J/ml. Lafracción arcillosa (< 2 µm) y lafracción de limos (de 2 a 20 µm) sesepararon mediante sedimentacióngravitacional en agua; la fracción dearenas (de 20 a 2000 µm) se

242
El Arado Natural:…
recuperó como sedimento una vezobtenidas las fracciones de arcilla yde limo. Posteriormente, losagregados se secaron en frío.
Análisis químicosEl C y el N totales se
determinaron por combustión secaen un analizador de C/N (ElementarVario EL C/N Analyzer). Loscoeficientes de variación entre lasrepeticiones fueron de ±2%. A causadel bajo pH del suelo, el C totalcorrespondió al C orgánico.
La cantidad de lignina y suestado de descomposición oxidativase calcularon mediante losparámetros de lignina obtenidos dela oxidación alcalina con CuO(Hedges y Ertel 1982; Kögel-Knabneret al. 1991). Los fenoles derivadosde la lignina mediante la oxidacióncon CuO se separaron ycuantificaron empleando lacromatografía de gas (CG) de tipocapilar. La porción de lignina intactase calculó con la suma de lasunidades de vanililo (V), de siringilo(S) y de cinamilo (C) (VSC). El estadode descomposición oxidativa sedeterminó como el grado deoxidación de la cadena lateralmicrobiana de la lignina remanente,que se calculó mediante la relaciónmasal ácido-aldehído de lasunidades de vanilina [(ac/al)V].Kögel-Knabner et al. (1991)obtuvieron coeficientes de variacióndel 8% en la determinación de la
cantidad de fenol y del 6% en lasrelaciones (ac/al)V. La eficiencia dela oxidación con CuO es diferentesegún el tejido de la planta y oscilaentre el 30% y el 90% en la ligninade tipo leñoso (Ertel et al. 1984). Larelativa liberación dependerá másadelante del grado de alteración de lalignina durante su descomposición.Es posible introducir un factor derendimiento equivalente, aunque lacorrección de los datos supone unalto grado de incertidumbre (Kögel-Knabner et al. 1991). Por dicharazón, se emplearon los rendimientosbrutos con fines comparativos.
Los carbohidratos no celulósicosfueron hidrolizados con ácidotrifluoroacético 4 M (TFA) segúnGuggenberger et al. (1994). Laseparación y la cuantificación de losmonómeros liberados se realizaronmediante un análisis decromatografía de gas y deespectrometría de masa (CG/EM),después de la oximación conO-metilhidroxilamina y la sililacióncon bis(trimetilsilil)-trifluoroacetamida (Andrews 1989).Los coeficientes de variación de losanálisis de los monosacáridos fueron1% y 17% (J. Hejzlar, com. pers.1992). Las relaciones galactosa +manosa/arabinosa + xilosa [(G+M)/(A+X)] y ramnosa+fucosa/arabinosa+xilosa [(R+F)/(A+X)] seemplearon para calcular lacontribución de las hemicelulosas delas células vegetales y de losazúcares derivados de la actividad
Cuadro 15-1. Distribución de los agregados estables en agua (g/kg de suelo) en las heces y en el suelocontiguo a ellas (Oxisol del pastizal Brachiaria decumbens y Pueraria phaseoloides).Los valores se basan en varios análisis de tres repeticiones, n = 3. Las letras diferentesindican diferencias significativas a P < 0.05 según la prueba de t-Student.
Muestra Agregados según clase de tamaño Suma de las(mm) de: fracciones
8-5 5-3.15 3.15-2 2-1 1-0.5 0.5-0.2 <0.2
Heces de lombriz 494 271 188 22 3 2 3 983Suelo circundante 168 a 222 b 201 192 c 100 d 57 e 56 f 996
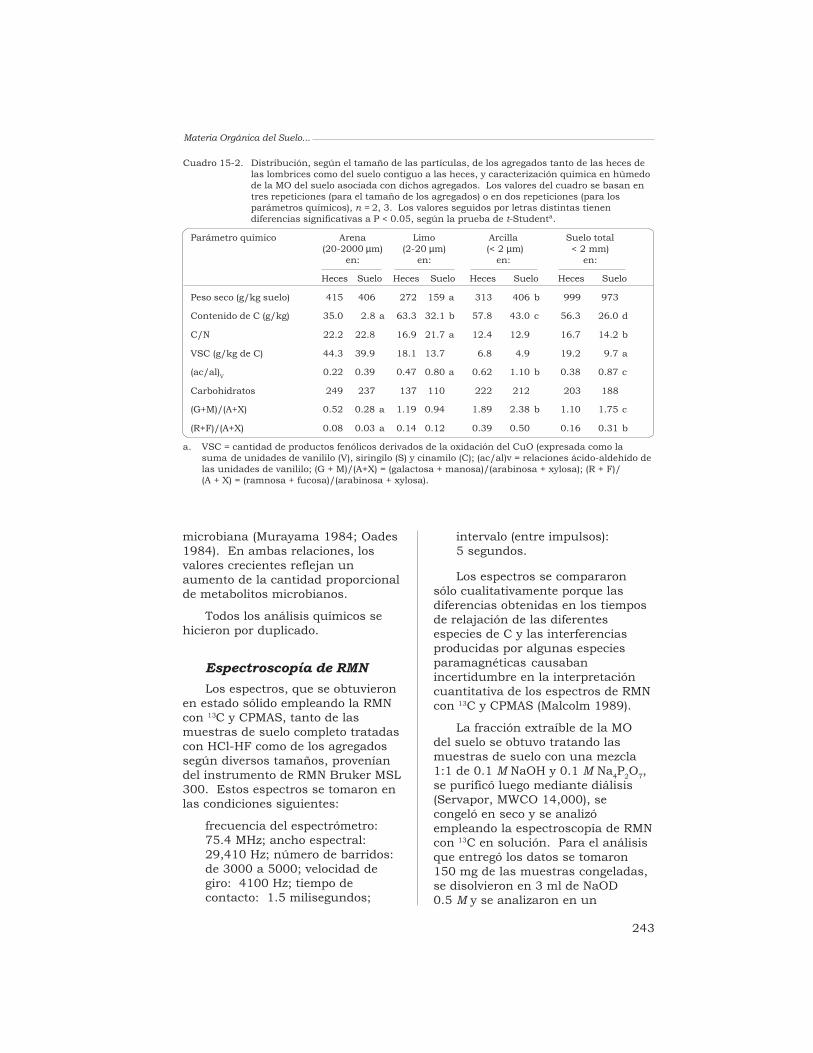
Materia Orgánica del Suelo...
243
Cuadro 15-2. Distribución, según el tamaño de las partículas, de los agregados tanto de las heces delas lombrices como del suelo contiguo a las heces, y caracterización química en húmedode la MO del suelo asociada con dichos agregados. Los valores del cuadro se basan entres repeticiones (para el tamaño de los agregados) o en dos repeticiones (para losparámetros químicos), n = 2, 3. Los valores seguidos por letras distintas tienendiferencias significativas a P < 0.05, según la prueba de t-Studenta.
Parámetro químico Arena Limo Arcilla Suelo total(20-2000 µm) (2-20 µm) (< 2 µm) < 2 mm)
en: en: en: en:
Heces Suelo Heces Suelo Heces Suelo Heces Suelo
Peso seco (g/kg suelo) 415 406 272 159 a 313 406 b 999 973
Contenido de C (g/kg) 35.0 2.8 a 63.3 32.1 b 57.8 43.0 c 56.3 26.0 d
C/N 22.2 22.8 16.9 21.7 a 12.4 12.9 16.7 14.2 b
VSC (g/kg de C) 44.3 39.9 18.1 13.7 6.8 4.9 19.2 9.7 a
(ac/al)V 0.22 0.39 0.47 0.80 a 0.62 1.10 b 0.38 0.87 c
Carbohidratos 249 237 137 110 222 212 203 188
(G+M)/(A+X) 0.52 0.28 a 1.19 0.94 1.89 2.38 b 1.10 1.75 c
(R+F)/(A+X) 0.08 0.03 a 0.14 0.12 0.39 0.50 0.16 0.31 b
a. VSC = cantidad de productos fenólicos derivados de la oxidación del CuO (expresada como lasuma de unidades de vanililo (V), siringilo (S) y cinamilo (C); (ac/al)v = relaciones ácido-aldehído delas unidades de vanililo; (G + M)/(A+X) = (galactosa + manosa)/(arabinosa + xylosa); (R + F)/(A + X) = (ramnosa + fucosa)/(arabinosa + xylosa).
microbiana (Murayama 1984; Oades1984). En ambas relaciones, losvalores crecientes reflejan unaumento de la cantidad proporcionalde metabolitos microbianos.
Todos los análisis químicos sehicieron por duplicado.
Espectroscopía de RMNLos espectros, que se obtuvieron
en estado sólido empleando la RMNcon 13C y CPMAS, tanto de lasmuestras de suelo completo tratadascon HCl-HF como de los agregadossegún diversos tamaños, proveníandel instrumento de RMN Bruker MSL300. Estos espectros se tomaron enlas condiciones siguientes:
frecuencia del espectrómetro:75.4 MHz; ancho espectral:29,410 Hz; número de barridos:de 3000 a 5000; velocidad degiro: 4100 Hz; tiempo decontacto: 1.5 milisegundos;
intervalo (entre impulsos):5 segundos.
Los espectros se compararonsólo cualitativamente porque lasdiferencias obtenidas en los tiemposde relajación de las diferentesespecies de C y las interferenciasproducidas por algunas especiesparamagnéticas causabanincertidumbre en la interpretacióncuantitativa de los espectros de RMNcon 13C y CPMAS (Malcolm 1989).
La fracción extraíble de la MOdel suelo se obtuvo tratando lasmuestras de suelo con una mezcla1:1 de 0.1 M NaOH y 0.1 M Na4P2O7,se purificó luego mediante diálisis(Servapor, MWCO 14,000), secongeló en seco y se analizóempleando la espectroscopía de RMNcon 13C en solución. Para el análisisque entregó los datos se tomaron150 mg de las muestras congeladas,se disolvieron en 3 ml de NaOD0.5 M y se analizaron en un

244
El Arado Natural:…
espectrómetro Bruker AM 500. Lasseñales del 13C fueron desacopladasrespecto a los protones mediante latécnica del desacoplamiento deacceso invertido, y se aplicó untiempo total de repetición de 2 s quepermitía la relajación completa detodos los espines atómicos. Unavez acumulados de 9,000 a27,000 barridos, se empleó unabanda ancha de 100 Hz con el fin dereducir el ruido de la señal(mejorando la relación señal:ruido).
Resultados
Estabilidad de los agregadosEn comparación con los del suelo,los agregados estables en aguamayores que 3.15 mm de las hecesfueron significativamente másabundantes (P < 0.05) y losagregados menores que 2 mm seredujeron drásticamente(Cuadro 15-1). Esta tendencia hacialos macroagregados en ladistribución por tamaño de losagregados indicó que la estabilidadestructural de las heces de laslombrices de tierra era mayor que lade los agregados del suelo quecircundaba las heces (Beare y Bruce1993).
Distribución según el tamañode las partículasLas heces de las lombrices estaban,al parecer, enriquecidas con lafracción de limos en detrimento de lafracción de arcillas, relación que nose presentó en las muestras de suelo(Cuadro 15-2). Sin embargo, dadoque las heces presentan,normalmente, mayor porcentaje deminerales de textura fina que elsuelo que las circunda (Lee 1985), elresultado en cuestión, de esteestudio, se relacionaría más con unadispersión incompleta de las heces
mediante ultrasonido (Christensen1992) que a un enriquecimiento deéstas con partículas de la fracción delimos. La formación demicroagregados estables en lafracción de limos puede estarrelacionada con la presencia demucopolisacáridos que aglutinan laspartículas primarias de arcilla (Parle1963).
C en el suelo y en losagregadosComo en los datos obtenidos porLoquet (1973) y Lee (1985), elcontenido de C orgánico de las hecesde las lombrices de este estudio fuemayor (casi el doble) que el del suelo(Cuadro 15-2). Más interesante aúnque el contenido total de C fue ladistribución de éste en cada uno delos agregados que se diferencian porel tamaño de las partículas. El sueloque no había sido ingeridorecientemente se caracterizó por unanotoria reducción de la MO del sueloasociada con la fracción arenosa ypor un incremento en el contenido dela MO unida a la fracción de arcillas,principalmente, y a la de limos. Esteesquema de distribución de la MOdel suelo es típico de la mayoría delos suelos (Christensen 1992). Enlas heces de las lombrices tambiénse observaron las concentracionesmás bajas de C en la fracciónarenosa, aunque las de C orgánicono fueron tan bajas al comparar lasheces con la misma fracción delsuelo circundante. En consecuencia,la actividad de las lombrices de tierraprodujo una redistribución de la MOdel suelo dentro de los agregados dediferente tamaño.
La relación C/N disminuyó alreducirse el tamaño de laspartículas, lo que indica que el gradode humificación iba en aumento(Baldock et al. 1992). Excepto en laspartículas del tamaño del limo, no

Materia Orgánica del Suelo...
245
hubo diferencias significativas en larelación C/N de los agregados dediferente tamaño de partículapertenecientes a las heces o al suelo.Esta relación C/N más baja de lafracción arenosa de las heces puedeatribuirse a la contribución de laarcilla a dicha fracción, por causa deuna dispersión incompleta (verantes). En cambio, la relación C/Ndel suelo fue mayor en las hecespuesto que es relativamente alto elporcentaje de MO del suelo (cuyarelación C/N es alta) asociada con lafracción de arenas.
Lignina en el suelo y en losagregados de tamaño diferenteEl rendimiento de los productosfenólicos de la oxidación con CuO(VSC) disminuyó constantementesegún el orden arena > limo > arcilla,tanto en las heces de las lombricescomo en el suelo que las circundaba(Cuadro 15-2). La creciente relación(ac/al)V encontrada sugiere unincremento en la oxidación de lacadena lateral de la ligninaremanente, en el mismo sentido.
En todos los agregadosanalizados, el VSC fue mayor en lasheces que en el suelo que lasrodeaba, aunque la diferencia no fuesignificativa. La cantidad de fenolesderivados de la lignina que sehallaron en las heces fue el doble delos hallados en el suelo. Esto sedebió, en parte, a una mayorconcentración de VSC en losagregados presentes en las heces y,principalmente, al enriquecimiento deestas heces con la MO del sueloasociada a la fracción arenosa.Simultáneamente, el suelo querodeaba las heces se caracterizó porpresentar un grado de oxidación de lacadena lateral de la ligninaremanente mayor que el de las hecesde las lombrices.
Carbohidratos en el suelo y enlos agregados de tamañodiferenteEn ambos tipos de muestras (suelo yheces), los carbohidratos nocelulósicos aumentaron en lasfracciones arenosa y arcillosa, y seredujeron en la fracción limosa(Cuadro 15-2). No se encontrarondiferencias significativas entre lasheces y el suelo circundante respectoa la concentración de carbohidratos,como revela el análisis químicorealizado en húmedo. La tendenciacreciente de las relaciones [(G+M)/(A+X)] y [(R+F)/(A+X)] sugiere que, encuanto disminuye el tamaño de losagregados, la contribución de losazúcares microbianos es cada vezmayor. La comparación entre lasheces y el suelo que las rodea revelóuna cantidad proporcionalmentemás grande de derivadosmicrobianos en la fracción arenosade las heces. Sin embargo, si seconsidera el suelo en su totalidad,las heces se caracterizaron por unacantidad proporcionalmente muyalta de carbohidratos derivados delas plantas (hemicelulosas).
Espectros de RMN con 13C yCPMAS del suelo total y de susagregadosAunque todos los espectros semostraron similares en suresonancia, se diferenciaron encuanto a su intensidad (Figura 15-1).Los espectros se dividieron encuatro intervalos químicos con el finde establecer comparacionescualitativas entre ellos: 0-50 ppm(C alquil), 50-110 ppm (C alquil-O),110-160 ppm (C aril), 160-200 ppm(C carboxil). Como se deduce de lospicos altos obtenidos a 30 ppm, lasresonancias en el intervalo del C delgrupo alquilo provenían,principalmente, de las cadenas depolimetileno (Oades et al. 1987). Las
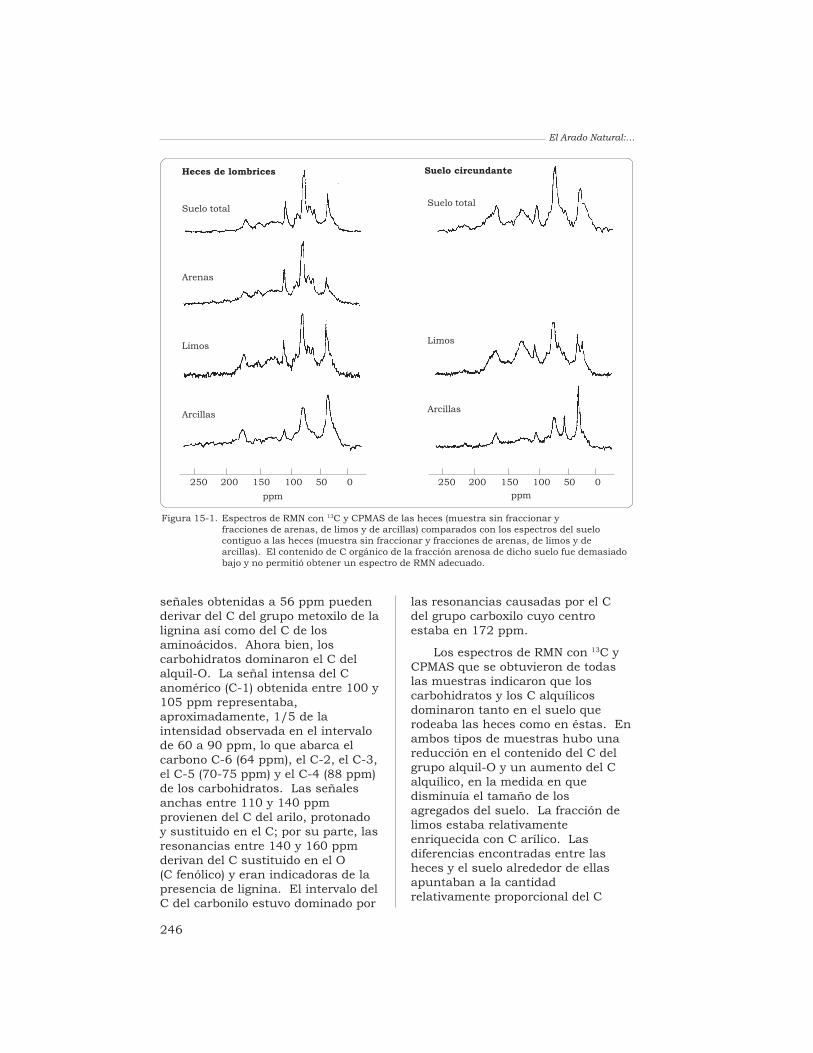
246
El Arado Natural:…
Suelo total
Limos
Arcillas
Heces de lombrices
Suelo total
Arenas
Limos
Arcillas
250 200 150 100 50 0 250 200 150 100 50 0ppm ppm
Figura 15-1. Espectros de RMN con 13C y CPMAS de las heces (muestra sin fraccionar yfracciones de arenas, de limos y de arcillas) comparados con los espectros del suelocontiguo a las heces (muestra sin fraccionar y fracciones de arenas, de limos y dearcillas). El contenido de C orgánico de la fracción arenosa de dicho suelo fue demasiadobajo y no permitió obtener un espectro de RMN adecuado.
Suelo circundante
señales obtenidas a 56 ppm puedenderivar del C del grupo metoxilo de lalignina así como del C de losaminoácidos. Ahora bien, loscarbohidratos dominaron el C delalquil-O. La señal intensa del Canomérico (C-1) obtenida entre 100 y105 ppm representaba,aproximadamente, 1/5 de laintensidad observada en el intervalode 60 a 90 ppm, lo que abarca elcarbono C-6 (64 ppm), el C-2, el C-3,el C-5 (70-75 ppm) y el C-4 (88 ppm)de los carbohidratos. Las señalesanchas entre 110 y 140 ppmprovienen del C del arilo, protonadoy sustituido en el C; por su parte, lasresonancias entre 140 y 160 ppmderivan del C sustituido en el O(C fenólico) y eran indicadoras de lapresencia de lignina. El intervalo delC del carbonilo estuvo dominado por
las resonancias causadas por el Cdel grupo carboxilo cuyo centroestaba en 172 ppm.
Los espectros de RMN con 13C yCPMAS que se obtuvieron de todaslas muestras indicaron que loscarbohidratos y los C alquílicosdominaron tanto en el suelo querodeaba las heces como en éstas. Enambos tipos de muestras hubo unareducción en el contenido del C delgrupo alquil-O y un aumento del Calquílico, en la medida en quedisminuía el tamaño de losagregados del suelo. La fracción delimos estaba relativamenteenriquecida con C arílico. Lasdiferencias encontradas entre lasheces y el suelo alrededor de ellasapuntaban a la cantidadrelativamente proporcional del C
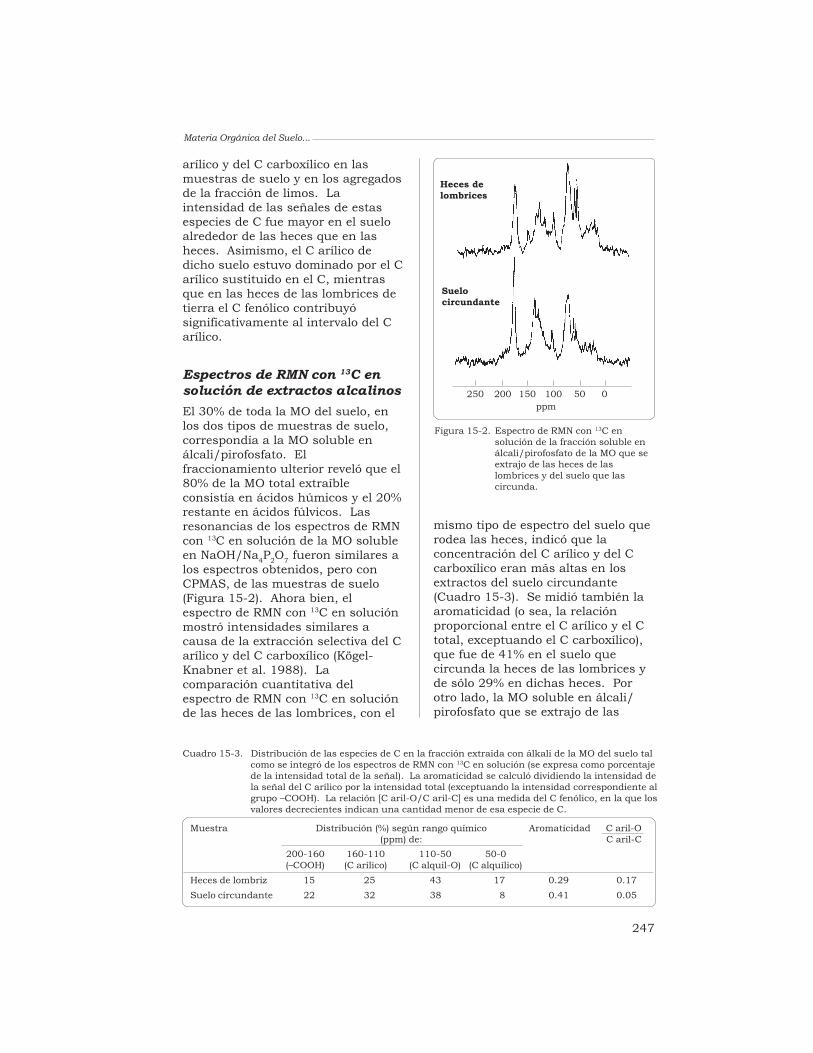
Materia Orgánica del Suelo...
247
arílico y del C carboxílico en lasmuestras de suelo y en los agregadosde la fracción de limos. Laintensidad de las señales de estasespecies de C fue mayor en el sueloalrededor de las heces que en lasheces. Asimismo, el C arílico dedicho suelo estuvo dominado por el Carílico sustituido en el C, mientrasque en las heces de las lombrices detierra el C fenólico contribuyósignificativamente al intervalo del Carílico.
Espectros de RMN con 13C ensolución de extractos alcalinosEl 30% de toda la MO del suelo, enlos dos tipos de muestras de suelo,correspondía a la MO soluble enálcali/pirofosfato. Elfraccionamiento ulterior reveló que el80% de la MO total extraíbleconsistía en ácidos húmicos y el 20%restante en ácidos fúlvicos. Lasresonancias de los espectros de RMNcon 13C en solución de la MO solubleen NaOH/Na4P2O7 fueron similares alos espectros obtenidos, pero conCPMAS, de las muestras de suelo(Figura 15-2). Ahora bien, elespectro de RMN con 13C en soluciónmostró intensidades similares acausa de la extracción selectiva del Carílico y del C carboxílico (Kögel-Knabner et al. 1988). Lacomparación cuantitativa delespectro de RMN con 13C en soluciónde las heces de las lombrices, con el
mismo tipo de espectro del suelo querodea las heces, indicó que laconcentración del C arílico y del Ccarboxílico eran más altas en losextractos del suelo circundante(Cuadro 15-3). Se midió también laaromaticidad (o sea, la relaciónproporcional entre el C arílico y el Ctotal, exceptuando el C carboxílico),que fue de 41% en el suelo quecircunda la heces de las lombrices yde sólo 29% en dichas heces. Porotro lado, la MO soluble en álcali/pirofosfato que se extrajo de las
Cuadro 15-3. Distribución de las especies de C en la fracción extraída con álkali de la MO del suelo talcomo se integró de los espectros de RMN con 13C en solución (se expresa como porcentajede la intensidad total de la señal). La aromaticidad se calculó dividiendo la intensidad dela señal del C arílico por la intensidad total (exceptuando la intensidad correspondiente algrupo –COOH). La relación [C aril-O/C aril-C] es una medida del C fenólico, en la que losvalores decrecientes indican una cantidad menor de esa especie de C.
Muestra Distribución (%) según rango químico Aromaticidad C aril-O(ppm) de: C aril-C
200-160 160-110 110-50 50-0(–COOH) (C arílico) (C alquil-O) (C alquílico)
Heces de lombriz 15 25 43 17 0.29 0.17Suelo circundante 22 32 38 8 0.41 0.05
Heces delombrices
Suelocircundante
250 200 150 100 50 0ppm
Figura 15-2. Espectro de RMN con 13C ensolución de la fracción soluble enálcali/pirofosfato de la MO que seextrajo de las heces de laslombrices y del suelo que lascircunda.

248
El Arado Natural:…
heces de las lombrices tenía másabundancia de carbohidratos (62,72 y 103 ppm) y de lignina (56 y150 ppm). El contenido de Cfenólico, calculado mediante larelación [C aril-O/C arílico] fuemucho mayor en las heces (0.17) queen el suelo alrededor de ellas (0.05).De este modo, cada uno de los dosmétodos espectroscópicos de RMNempleados corroboró los resultadosobtenidos con el otro.
DiscusiónDecaëns et al. (1994) señalaron elefecto notable que tenía laintroducción del pastizal deB. decumbens + P. phaseoloides en labiomasa de las lombrices de tierra.Esta biomasa fue de 511 kg de pesofresco por hectárea, un valor más de10 veces superior al encontrado en lasabana natural. Jiménez et al.(1994), por su parte, calcularon queestas lombrices ingerían 680 Mg desuelo seco por hectárea en el pastizalque contenía una leguminosa. Unalto porcentaje de esa cantidad sehalla como capa de heces sobre lasuperficie del suelo. Las heces enque se tomaron las muestraspresentan un gradiente de edadesque va desde las que tenían unospocos días hasta las de varios meses.Por consiguiente, no fue posibleseguir la dinámica de la MO del sueloen las heces durante elenvejecimiento de éstas. Este estudiofue planeado, en realidad, pararesaltar las diferencias principalesque presentan, en la estructuraquímica de la MO del suelo, las hecesde las lombrices y el suelo que lasrodea, y para esclarecer los posiblesmecanismos que asocian la MO delsuelo con la fase mineral de éste.
Al determinar los agregadosestables en agua, se hizo evidenteuna notoria estabilización de laspartículas secundarias en las heces
de las lombrices. A pesar desometerlos a una acción sónicaintensa, una porción de losagregados de la fracción de limosresultó estable. Una estabilidadestructural muy alta de las heces delas lombrices obtuvieron también DeVleeschauwer y Lal (1981), quienesreportaron que las heces puedensoportar de 5 a 54 veces más energíacinética proveniente de las gotas delluvia antes de desintegrarse que losagregados del suelo. Las hecesdemuestran también, generalmente,una resistencia a la tensión mayorque los agregados del suelo(McKenzie y Dexter 1987). Esteincremento en la estabilidad de susagregados refleja, en las heces, unaconcentración más elevada de Corgánico. Zhang y Schrader (1993)mostraron la relación existente entrela estabilidad de los agregados(agregación en agua, resistencia a latensión) y el C orgánico. Postulanestos autores la formación deuniones entre un humato de Ca (delhumus) o un catión polivalente deMO, de un lado, y una partícula desuelo, del otro, uniones que implicanla presencia dominante de la MO delsuelo humificada en los procesos deestabilización.
La distribución del C orgánico enlos agregados de suelo reveló, sinembargo, que la MO del sueloasociada con la fracción arenosa era13 veces más abundante en lasheces que en la fracción equivalentedel suelo situado alrededor de éstas.Según Christensen (1992), estafracción está compuesta,principalmente, por MO que no seasocia para formar complejosórgano-minerales. Se puedepostular, por tanto, una relaciónpositiva entre la alta estabilidad delos agregados de las heces y lacantidad grande de MO del suelo queno participa en dichas uniones. El Corgánico asociado con la fracciónmineral fue sólo 2 veces (en fracción

Materia Orgánica del Suelo...
249
de limos) y 1.3 veces (en fracción dearcillas) superior en las heces queen el suelo alrededor de éstas.
La caracterización analítica delas heces y del suelo que lascircundaba reveló que la MO delsuelo asociada con la fracciónarenosa consistía solamente enhojarasca ligeramentedescompuesta (un recurso primario).Los tejidos vegetales vasculares nolignificados y aún sin descomponerse caracterizan por presentarvalores de VSC que oscilan entre30 y 80 g/kg de C y un valor de0.15±0.05 en la relación (ac/al)V(Hedges y Mann 1979). Por tanto,los valores obtenidos sugieren que lacantidad de lignina presente en losagregados de la fracción arenosa delas heces es alta. Los azúcaresderivados de las plantas dominaronla MO del suelo asociada con lafracción arenosa, como lo demuestrala composición de los carbohidratos.Más aún, el espectro de RMN con13C y CPMAS de los agregados de lafracción arenosa se asemejó muchoal de la hojarasca y al de la MOpresente en los agregados libres delos suelos del pastizal de largaduración (Golchin et al. 1994;Skjemstad et al. 1986). Losespectros de RMN con 13C y CPMASde los residuos vegetales frescos secaracterizaron siempre por unaintensidad de señal alta generadapor los carbohidratos (C alquil-O) ypor la lignina (C aril-O).
En los dos tipos de muestrasanalizadas (heces y suelo) seobservó que, al reducirse el tamañode los agregados, tanto la relaciónC/N como el contenido (losparámetros) de lignina, ladistribución de los carbohidratos yla espectroscopía de RMN con 13C yCPMAS indicaban un grado más altode descomposición de la MO delsuelo. Según Kögel-Knabner et al.(1988), el descenso de los valores de
VSC y el aumento de la relación(ac/al)V se consideran indicadores dela descomposición de la lignina en elperfil del suelo. Guggenberger et al.(1994, 1995b) señalaron que, en losInceptisoles, una reducción en losproductos de oxidación del CuO y unaumento en la funcionalidadcarboxílica del remanente de lignina—siguiendo el orden arena, limo,arcilla— estaban relacionados,directamente, con la descomposiciónde la lignina. La distribución de loscarbohidratos indicó que la cantidadproporcional de azúcares derivadosde la actividad microbianaaumentaba en la medida en quedisminuía el tamaño de laspartículas en los agregados. Elenriquecimiento en carbohidratos deorigen microbiano de la fracciónarcillosa de los agregados se debe,probablemente, a un aporte mayorde metabolitos derivados de laactividad microbiana a dichafracción (Baldock et al. 1990) ytambién a la estabilización de losazúcares de origen microbianocuando se asocian íntimamente conlas arcillas (Guggenberger et al.1994). La espectroscopía de RMNcon 13C y CPMAS mostró que lafracción arcillosa de los agregadosestaba también enriquecida con Calquílico. Puesto que losmicroorganismos sintetizanbásicamente carbohidratos y Calquílico (Baldock et al. 1990), estafunción constituía una pruebaadicional de que la MO del sueloasociada con la fracción arcillosa delos agregados estaba enriquecida conmetabolitos derivados de la actividadmicrobiana (los recursossecundarios). En cambio, la fracciónde limos de los agregados secaracterizó porque presentaba lasconcentraciones más altas de Caromático y de grupos –COOH. LaMO del suelo unida a la fracción delimos representa los componentesorgánicos humificados (Oades 1990).

250
El Arado Natural:…
Se puede decir, en general, que lacomposición de la MO del sueloasociada a los agregados de diferentetamaño, en este suelo tropical, escomparable a la que se halló en lossuelos de zona templada enAlemania (Guggenberger et al. 1994,1995b).
Los análisis químicosestructurales indicaron, al parecer,que la MO del suelo asociada con lasfracciones de limos y de arcillas sehallaba en un estado más avanzadode descomposición en el suelocontiguo a las heces que en éstasmismas. Mayor interés despiertanlas diferencias observadas en laforma en que la MO se distribuyeentre las diversas fracciones delsuelo, lo que da una relaciónproporcional diferente para la MOasociada, bien sea con la fracción dearenas (dominada por los recursosprimarios) o bien con la fracción dearcillas (dominada por los recursossecundarios), o con la fracción delimos (dominada por la MOhumificada) del suelo. El espectro deRMN con 13C y CPMAS de las hecescompletas de las lombrices seasemejó bastante al de la fracciónarenosa de los agregados quegeneraba señales dominantes en laregión del C del grupo alquil-O. Encambio, el espectro de RMN con 13C yCPMAS del suelo no fraccionado fuesimilar al obtenido en la fracción delimos de los agregados. Enconsecuencia, dicho suelo era másrico en C aromático y en Ccarboxílico que las heces de laslombrices. Esta conclusión fuetambién obvia en los espectros deRMN con 13C en solución de la MOsoluble en álcali/pirofosfato que seextrajo de las muestras de suelo.Las señales de intensidad más altaprovenientes de la lignina (o sea, unarelación alta de [C aril-O/C aril-C] yde C metoxil) y de los carbohidratos(o sea, C alquil-O) de las hecesindicaron que éstas tenían una
concentración mayor de recursosprimarios que el suelo alrededor deellas. La MO soluble en NaOH/Na4P2O7, proveniente del sueloalrededor de las heces, parecía estarmás enriquecida en sustanciashúmicas cuyo contenido de Ccarboxílico y de C aromáticosustituido en el C (o sea, C arílico;baja relación [C aril-O/C aril-C]) erarelativamente alto.
Los datos obtenidos demuestranque las uniones entre el humato-Ca ola MO-catión polivalente, de un lado,y las partículas del suelo, del otro,difícilmente podrían ser lasresponsables de que haya mayorestabilidad en los agregados de lasheces de las lombrices que en los delsuelo alrededor de ellas. Esemecanismo implicaría la presencia enlas heces de una mayorconcentración de sustanciashúmicas, ricas en gruposcarboxílicos. No obstante, el resto dela MO del suelo contenida en lasheces era, principalmente, macro-MOlibre, que consistía en residuosvegetales ligeramente descompuestos.La estrategia ecológica de lasprincipales especies de lombriz detierra presentes en estos suelosexplica esta observación. Aquí, lacomunidad de lombrices estádominada por especies de hábitoanécico o endoanécico (Jiménez et al.1994), cuya estrategia adaptativa sebasa en alimentarse, al menosparcialmente, de hojarasca y en viviren el suelo (Bouché 1977). Duranteel tránsito de la MO por el intestinode las lombrices ocurre unaaceleración repentina del proceso dedescomposición de la MO,acompañado por una acción demineralización y fragmentación de laMO del suelo que resulta del fuerteefecto inicial de dicho proceso, acausa de la adición del agua y delmucus intestinal (Lavelle y Martin1992). La actividad microbianaprosigue con gran intensidad durante

Materia Orgánica del Suelo...
251
algunos días después de ladeposición de las heces frescas; enese momento, la descomposición sereduce drásticamente (Lavelle yMartin 1992).
La hojarasca fresca de lasuperficie del suelo se mezclaíntimamente con la fase mineralinorgánica durante el tránsito por elintestino de la lombriz, de acuerdocon las estrategias empleadas por lasespecies anécicas y endoanécicas (elllamado ‘efecto anécico’, segúnLavelle 1988). De este modo seproduce la estabilización física de losresiduos vegetales. Van Veen yKuikman (1990) explicaron el papelde la arquitectura del suelo comofactor que controla ladescomposición microbiana en losecosistemas terrestres. Laprotección física que brinda a la MO—fácilmente disponible en el suelo,por su naturaleza biológica— elconfinamiento proporcionado por losagregados, hace inaccesibles losresiduos orgánicos a la accióndescomponedora de la comunidadmicrobiana. Puesto que el 95% delos poros del suelo no son accesiblesa las bacterias, los microorganismosy sus sustratos están separados,generalmente, por barreras físicas(van Veen y Kuikman 1990); seacumula, por tanto, en el suelo unareserva de MO que se consideradisponible. La anerobiosisfacultativa que se establece en elinterior de los agregados retardatodavía más la descomposición de laMO del suelo (Blanchart et al. 1993;Lavelle y Martin 1992).
La estabilidad estructural de lasheces se debe al efecto estabilizadorde los mucopolisacáridos producidospor los microorganismos del tractointestinal de las lombrices (Parle1963) y por las propias lombrices.No obstante, el estudio realizado porGolchin et al. (1994) sobre laestructura del suelo y sobre el ciclo
del C ha ofrecido una explicaciónadicional que concuerda con el altocontenido de recursos primarios delas heces. La descomposición rápidade los residuos vegetales estálimitada por su interacción con lamatriz inorgánica; no obstante, sirvecomo fuente de energía de losmicroorganismos. El metabolismomicrobiano, principalmente de loscarbohidratos, conduce a laliberación de mucílago y de ciertosmetabolitos, los que se difunden enlas partículas minerales yestabilizan, por ello, los agregados.Golchin et al. (1994) mostraron quela estabilidad de los microagregadosdepende de la MO de fácildescomposición que estáincorporada en los agregados. Enconsecuencia, la estabilidadestructural de los agregadosdisminuye en la medida en que laMO se descompone permitiendo laacumulación de compuestosrefractarios. Más aún, Cambardellay Elliott (1994) aportaron pruebas deque los subproductos microbianostambién desempeñan un papel activoen la unión de microagregados quelleva a la formación demacroagregados. Finalmente,Gijsman y Thomas (1995) reportaronla existencia de una relación de tipono lineal (Mitscherlich) entre laestabilidad de los agregadoshúmedos y los carbohidratosextraíbles con agua caliente, en losOxisoles de la estación experimentalCarimagua. Estos autores sientan lahipótesis de que dichoscarbohidratos equivalen a lospolisacáridos provenientes de lasplantas o de la actividad microbiana,los cuales aglutinan a los agregados.
Puesto que las heces de laslombrices de tierra se caracterizanpor tener concentraciones altas derecursos primarios ligeramentealterados e íntimamente mezcladoscon la matriz mineral del suelo, sesugiere en este trabajo la presencia

252
El Arado Natural:…
en ellas de procesos similares deestabilización mediante mecanismosbióticos y no por formación deuniones entre un humato de Ca o unMO-catión polivalente y laspartículas del suelo. Laconsecuencia de la estrechaasociación entre los residuosvegetales ligeramente descompuestosy la fase mineral del suelo es laformación de una reserva de C quees activo pero está protegidofísicamente, y que se libera cuandola estabilidad de las hecesdisminuye. Esta hipótesis,naturalmente, debe probarsemediante el análisis de heces deedades diferentes que presentendiversos grados de desintegraciónestructural.
AgradecimientosLos autores agradecen en granmedida el trabajo analítico realizadopor Tanja Gonter y por IrisZimmermann. Expresan además unprofundo agradecimiento a LudwigHaumaier, quien registró losespectros de RMN con 13C ensolución.
ReferenciasAnderson DW; Paul EA. 1984. Organo-
mineral complexes and their studyby radiocarbon dating. Soil Sci SocAm J 48:298-301.
Anderson JM; Flanagan PW. 1989.Biological processes regulatingorganic matter dynamics in tropicalsoils. En: Coleman DC; Oades JM;Uehara G, eds. Tropical soil organicmatter. University of Hawaii Press,Honolulu, Estados Unidos.p 97-123.
Andrews MA. 1989. Capillary gas-chromatographic analysis ofmonosaccharides: improvements andcomparisons using trifluoroacetylationand trimethylsilylation of sugarO-benzyl- and O-methyl-oximes.Carbohydr Res 194:1-19.
Baldock JA; Oades JM; Vassallo AM; WilsonMA. 1990. Solid state CPMAS 13CN.M.R. analysis of particle size anddensity fractions of a soil incubatedwith uniformly labelled 13C-glucose.Aust J Soil Res 28:193-212.
Baldock JA; Oades JM; Waters AG; Peng X;Vassallo AM; Wilson MA. 1992.Aspects of the chemical structure ofsoil organic materials as revealed bysolid-state 13C NMR spectroscopy.Biogeochemistry 16:1-42.
Beare MH; Bruce RR. 1993. A comparisonof methods for measuring water-stableaggregates: implications fordetermining environmental effects onsoil structure. Geoderma 56:87-104.
Blanchart E; Bruand A; Lavelle P. 1993. Thephysical structure of casts of Millsoniaanomala (Oligochaeta, Megascolecidae)in shrub savanna soils (Côte d’Ivoire).Geoderma 56:119-132.
Bouché MB. 1977. Stratégies lombriciennes.Ecol Bull (Stockholm) 25:122-132.
Cambardella CA; Elliott ET. 1994. Carbonand nitrogen dynamics of soil organicmatter size/density fractions underdifferent management practices. SoilSci Soc Am J 57:1071-1076.
Christensen BT. 1985. Carbon and nitrogenin particle size fractions isolated fromDanish arable soils by ultrasonicdispersion and gravity-sedimentation.Acta Agric Scand 35:175-187.
Christensen BT. 1992. Physicalfractionation of soil and organicmatter in primary particle size anddensity separates. Adv Soil Sci20:1-90.
Decaëns T; Lavelle P; Jiménez JJ; EscobarG; Rippstein G. 1994. Impact of landmanagement on soil macrofauna inthe Oriental Llanos of Colombia. Eur JSoil Biol 30:157-168.

Materia Orgánica del Suelo...
253
De Vleeschauwer D; Lal R. 1981.Properties of worm casts undersecondary tropical forest regrowth.Soil Sci 132:175-181.
Ertel JR; Hedges JI; Perdue EM. 1984.Lignin signature of aquatic humicsubstances. Science 223:485-487.
Fisher MJ; Rao IM; Ayarza MA; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage by introduceddeep-rooted grasses in the SouthAmerican savannas. Nature371:236-238.
Gijsman AJ; Thomas RJ. 1995. Aggregatesize distribution and stability of anOxisol under legume-based andpure grass pastures in the EasternColombian savannas. Aust J SoilRes 33:153-165.
Golchin A; Oades JM; Skjemstad JO;Clarke P. 1994. Soil structure andcarbon cycling. Aust J Soil Res32:1043-1068.
Guggenberger G; Christensen BT; ZechW. 1994. Land-use effects on thecomposition of organic matter inparticle-size separates of soil. I.Lignin and carbohydrate signature.Eur J Soil Sci 45:449-458.
Guggenberger G; Zech W; Thomas RJ.1995a. Lignin and carbohydratealteration in particle-size separatesof an Oxisol under tropical pasturesfollowing native savanna. Soil BiolBiochem 27:1629-1638.
Guggenberger G; Zech W; Haumaier L;Christensen BT. 1995b. Land-useeffects on the composition oforganic matter in particle-sizeseparates of soils. II. CPMAS andsolution 13C NMR analysis. Eur JSoil Sci 46:147-158.
Hedges JI; Ertel JR. 1982.Characterization of lignin by gascapillary chromatography of cupricoxide oxidation products. AnnChem 54:174-178.
Hedges JI; Mann DC. 1979. Thecharacterization of plant tissues bytheir lignin oxidation products.Geochim Cosmochim Acta43:1803-1807.
Jiménez-Jaén JJ; Decaëns T; Moreno AG;Rossi J-P; Lavelle P. 1994.Dynamics and short-term effects ofearthworms in natural andmanaged savannas of the easternplains of Colombia. Informe delCIAT (sin publicar). CIAT, Cali,Colombia.
Kögel-Knabner I; Zech W; Hatcher PG.1988. Chemical composition of theorganic matter in forest soils; III:The humus layer. Z PflanzenernähBodenkd 151:331-340.
Kögel-Knabner I; Zech W; Hatcher PG.1991. Chemical structural studiesof forest soil humic acids: aromaticcarbon fraction. Soil Sci Soc Am J55:241-247.
Lascano CL; Estrada J. 1989. Long-termproductivity of legume-based andpure grass pastures in the EasternPlains of Colombia. En: Proceedingsof the XVI International GrasslandCongress. Niza, Francia.p 1179-1180.
Lavelle P. 1988. Earthworm activities andthe soil system. Biol Fertil Soils6:237-251.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humictropics. Soil Biol Biochem24:1491-1498.
Lee KE. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, Sydney.
Loquet M. 1972. Étude de l’activitémicrobiologique d’une prairiepermanente, le Haras du Pin;Comparaison avec les turricules devers. Rev Ecol Biol Sol 11:47-65.

254
El Arado Natural:…
Malcolm RL. 1989. Applications of solid-state 13C NMR spectroscopy togeochemical studies of humicsubstances. En: Hayes MHB;MacCarthy P; Malcolm RL; Swift RS,eds. Humic substances. II. In searchof structure. Wiley, Nueva York.p 339-372.
McKenzie BM; Dexter AR. 1987. Physicalproperties of casts of the earthwormAporrectodea rosea. Biol Fertil Soils5:152-157.
Murayama S. 1984. Changes in themonosaccharide composition duringthe decomposition of straws underfield conditions. Soil Sci Plant Nutr30:367-381.
Oades JM. 1984. Soil organic matter andstructural stability: mechanisms andimplications for management. PlantSoil 76:319-337.
Oades JM. 1990. Associations of colloidsin soil aggregates. En: de Boodt MF;Hayes MHB; Herbillon A, eds. Soilcolloids and their associations inaggregates NATO ASI Series B.Plenum Press, Londres. vol. 215,p 463-483.
Oades JM; Vassallo AM; Waters AG;Wilson MA. 1987. Characterizationof organic matter in particle-size anddensity fractions from a red-brownearth by solid-state 13C N.M.R. AustJ Soil Res 25:71-82.
Parle JN. 1963. Microorganisms in theintestines of earthworms. J GenMicrobiol 31:1-11.
Preston CM. 1987. Review of solutionNMR of humic substances. En:Wershaw RL; Mikita MA, eds. NMRof humic substances and coal. LewisPublishers, Chelsea, EstadosUnidos. p 3-32.
Rao IM; Zeigler RS; Vera R; Sarkarung S.1993. Selection and breeding foracid-soil tolerance in crops: uplandrice and tropical forages as casestudies. Bioscience 43:454-465.
Rao IM; Ayarza MA; Thomas RJ. 1994.The use of carbon isotope ratios toevaluate legume contribution to soilenhancement in tropical pastures.Plant Soil 162:177-182.
Skjemstad JO; Dalal RC; Barron PF.1986. Spectroscopic investigation ofcultivation effects on organic matterof Vertisols. Soil Sci Soc Am J50:354-359.
Thomas RJ; Asakawa NM 1993.Decomposition of leaf litter fromtropical forage grasses andlegumes. Soil Biol Biochem25:1351-1361.
Thomas RJ; Fisher MJ; Ayarza MA; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. Advances inSoil Science, vol. 21. CRC LewisPublishers, Boca Ratón, FL,Estados Unidos. p 61-83.
Tiessen H; Stewart JWB. 1983. Particle-size fractions and their use instudies of soil organic matter; II:Cultivation effects on organicmatter composition in size fractions.Soil Sci Soc Am J 47:509-514.
Van Veen JA; Kuikman PJ. 1990. Soilstructural aspects of decompositionof organic matter by micro-organisms. Biogeochemistry11:213-233.
Zhang H; Schrader S. 1993. Earthwormeffects on selected physical andchemical properties of soilaggregates. Biol Fertil Soils15:229-234.

Dinámica del Carbono y del Nitrógeno...
255
durante el envejecimiento de éstas(+100%) a causa, posiblemente, delos procesos de fijación de CO2, a laacumulación de raíces muertas o a laactividad de algunos grupos demacrofauna en las heces. Losniveles de NH4 fueron muy elevadosen las heces frescas (294.20 a233.98 µg/g de heces secas) encomparación con los del suelo(26.96 a 73.95 µg/g de suelo seco) acausa de la intensa mineralizaciónque el suelo y la MO experimentandurante su tránsito por el intestinode la lombriz. En la primera semanade envejecimiento de las heces, laconcentración de NH4 disminuyódrásticamente, mientras que la deNO3 presentó incrementos sucesivostanto en las heces como en el suelosubyacente (respecto a las heces) yen el suelo adyacente. Estosresultados indicaron que hubo unaproducción rápida de NO3mediante algún proceso denitrificación en las heces frescas,seguido de la difusión del NO3,primero vertical y despuéshorizontal, hacia el suelocircundante. Cumplidas 2 semanasde envejecimiento de las heces, lasconcentraciones de NH4 y NO3mostraron apenas ligerasvariaciones, probablemente porquela MO estaba protegida en las hecessecas, que son más estables. Labiomasa radicular fue más alta (de
CAPÍTULO 16
Dinámica del Carbono y del Nitrógenoen las Heces de Diferente EdadDepositadas por Lombrices en Areasde Sabana y de Pastizal en los LlanosOrientales de Colombia1
T. Decaëns* , A. F. Rangel**, N. Asakawa** yR. J. Thomas**
1. Publicado en Biology and Fertility of Soils 30,p 20-28, por Decaëns et al. Copyright ©1999 de Springer-Verlag. Impreso conpermiso de la casa editorial.
* Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.
** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
+
+
–
+
–
–
–
ResumenSe investigó el efecto causado por lalombriz anécica Martiodriluscarimaguensis Jiménez y Moreno enla dinámica del C y del N en un áreade sabana nativa y en un pastizalantrópico de los Llanos Orientales deColombia. Se compararon, a lo largode 11 meses, el C total, el N total y lacantidad de NH4 y NO3 en las heces,en el suelo subyacente y en el sueloadyacente a ellas. Se realizarontambién muestreos adicionales paracuantificar la biomasa y lamacrofauna del suelo. En los dossistemas estudiados (sabana ypastizal), la cantidad de C y de N fuesuperior en las heces (4.33% y7.50%, respectivamente) que en elsuelo (2.81% y 4.08%,respectivamente), lo que indica quelas lombrices seleccionaban unsustrato rico en materia orgánica(MO) para su alimentación. Engeneral, el contenido de C de lasheces aumentó significativamente
+ –

256
El Arado Natural:…
1.6 a 4.7 veces mayor) debajo de lasheces viejas. Se comenta, además,aquí el significado ecológico de losresultados obtenidos.
Palabras clave: dinámica de lamateria orgánica del suelo,dinámica del nitrógeno, fertilidaddel suelo, lombrices, sabana
IntroducciónLas sabanas tropicales cubren unárea de 243 x 106 ha, es decir, el43% de la superficie de AméricaLatina. Tradicionalmente, estassabanas han sido explotadasmediante el pastoreo extensivo debaja productividad (Rippstein et al.1996; Vera y Seré 1985). Sinembargo, la tierra cultivable para laproducción agrícola intensiva se haexpandido allí durante los últimos20 años. Los agroecosistemas quemás comúnmente se establecen vandesde los pastizales de gramíneas,solos o asociados con leguminosas,hasta los cultivos intensivos anuales(Thomas et al. 1995). Esta evoluciónde la tierra tiene implicacioneseconómicas importantes, aunquegran parte de su impacto ambientalse desconoce todavía.
Los invertebrados de grantamaño, principalmente laslombrices de tierra, las termitas y lashormigas, se consideran los“ingenieros del ecosistema” (sensuJones et al. 1994; consultar tambiénAnderson 1995). En el trópicohúmedo, ni las condicionesclimáticas ni los minerales arcillososdel suelo regulan, eficientemente, losprocesos de mineralización; llegan apredominar, por ello, estosorganismos como reguladores de losprocesos del suelo (Lavelle y Martin1992). Los ingenieros del ecosistemaregulan eficazmente la actividad delos microorganismos del suelo pormedio de las estructuras órgano-minerales que ellos producen (como
las heces, los termiteros y loshormigueros) y pueden causarefectos importantes en la agregacióndel suelo y en la regulación de ladinámica de la MO (Blanchart et al.1997; Bohlen y Edwards 1995;Krietzschmar y Monestiez 1992;Marinissen y de Ruiter 1993; Scheu1987).
Los efectos que las lombrices detierra ejercen en la dinámica de laMO varían según las escalas deespacio y de tiempo consideradas(Lavelle et al. 1998). A corto plazo, laactividad microbiana se estimula enel tracto intestinal y ocasiona así unincremento de la mineralización de laMO y una liberación importante denutrientes asimilables en las hecesfrescas (Blair et al. 1994). A largoplazo, las heces viejas, si suspropiedades físicas se conservan,protegen la MO de ulterioresprocesos de mineralización.Finalmente, parece que las lombricespropician el ciclo rápido de la MO enel suelo. Ahora bien, es precisoobtener más información, ya que sóloalgunas especies tropicales (especiesendógeas alóctonas, principalmente)han sido estudiadas intensamenterespecto a los diferentes procesosque ocurren en el suelo (Fragosoet al. 1997).
El objetivo del presente trabajofue la descripción de la dinámica, acorto y a largo plazo, del C y del N enlas heces de las lombrices de tierra yen el suelo superficial circundante.Se prestó atención especial a losefectos de Martiodrilus carimaguensisJiménez y Moreno (Oligochaeta,Glossoscolecidae), la lombriz quedepositaba mayor cantidad de hecessobre la superficie del suelo en elsitio del estudio. Los experimentosse realizaron a la escala espacio-temporal de las heces producidas poresta especie en una parcela desabana y en un pastizal derivado dela sabana. Se midió también la

Dinámica del Carbono y del Nitrógeno...
257
biomasa radicular con el fin deobservar la respuesta de las plantas ala deposición de las heces.
El estudio se realizó en laestación de investigación Carimaguadel convenio CIAT-CORPOICA(4°37’ N, 71°19’ O), situada en launidad fitogeográfica de las sabanasisohipertérmicas bien drenadas de losLlanos Orientales de Colombia. Elclima es de tipo subhúmedo tropical,presenta valores medios anuales detemperatura y de precipitación de26 °C y 2300 mm, respectivamente, yse caracteriza por tener una épocaseca muy definida, de noviembre amarzo (datos del CIAT). La vegetaciónnativa varía junto con la topografía:hay sabanas abiertas en las zonasaltas (los “altos” y “planos”) y selvasde galería o sabanas inundables enlas zonas bajas (los “bajos”). Lossuelos son Oxisoles (TropepticHaplustox Isohipertérmicos) en losaltos y Ultisoles (Ultic AericPlintaquox) en los bajos. Ambossuelos se caracterizan porque suspropiedades físicas son favorables(agregación, porosidad y retenciónde agua), su acidez es alta [pH (H2O)< 5], y su fertilidad química naturales muy baja (saturación de Al > 80%,CIC < 5 meq/100 g).
Materiales y Métodos
Parcelas experimentalesEl experimento se realizó en dossistemas diferentes de uso de latierra, en un Oxisol de una zona altabien drenada:
• Una parcela de sabana nativa enla que crecía el pastoTrachypogon vestitus; estaparcela estuvo protegida delpastoreo durante 4 años y fuequemada cada año durante laépoca seca, según el manejotradicional.
• Un pastizal de 3 años en que sesembraron Brachiariahumidicola, Arachis pintoi,Stylosanthes capitata yCentrosema acutifolium, que fuesometido a pastoreo con unacarga animal de 2.0 animales/ha.
Diseño experimentalDe las ocho especies de lombrices detierra encontradas en la sabananativa en el sitio del estudio(Jiménez et al. 1998a), se eligióM. carimaguensis porque es la únicacuya actividad de deposición de hecessobre la superficie del suelo essignificativa. Las heces producidaspor esta especie son grandes (hasta15 cm de altura), tienen forma detorre (turrículos) y su formación dura,normalmente, varios días. Además,esta especie se adapta muy bien aalgunos agroecosistemas, unacualidad que la haría útil en elcontexto de una agriculturasostenible (Jiménez et al. 1998b).
El experimento comenzó en mayode 1996, al inicio de la época lluviosa,cuando la actividad de las lombricesestá en un punto máximo (Jiménezet al. 1998b). En los dos sistemasestudiados, se eligieron 160 heces deM. carimaguensis y se dividieron en4 grupos de 40 heces que estuvieranpróximas, cada una de las cuales seidentificó con una chapa metálicapequeña. Sólo se escogieron hecesfrescas y aún pequeñas, es decir, enla fase temprana de la deposición; deeste modo, se tiene la certeza de quela lombriz está presente bajo cadauna de las heces marcadas y se evita,además, el efecto acumulativo quecausaría, sobre las heces, laprolongada actividad de la lombrizantes del inicio del experimento. Enel pastizal, cada grupo de heces seprotegió del pisoteo de los animalesmediante jaulas metálicas de 2 x 2 m.

258
El Arado Natural:…
Las muestras se tomaron en losdías 1, 2, 3, 7 y 14 y en los meses 1,2, 5, 6.5, 8 y 11 después de ladeposición de las heces. En cadauna de esas fechas se tomó unamuestra al azar de cada grupo de40 heces. Cada muestra consistía entres submuestras: (1) la deposición,(2) el suelo subyacente (localizadodirectamente debajo de ladeposición), y (3) el suelo adyacente(localizado a 20 cm de la deposición)(Figura 16-1). El suelo se recolectómediante un cilindro metálico de5 cm de diámetro y 5 cm deprofundidad y se refrigeró a 4 °C.
Análisis químicoEl N mineral se extrajo agitando 4 gde cada muestra fresca con 40 ml deuna solución de KCl 1M durante30 minutos. Las suspensiones sefiltraron y estos filtrados seconservaron a –15 °C antes delanálisis. Se emplearon métodoscolorimétricos para determinar lasconcentraciones de los iones amonioy nitrato (Anderson e Ingram 1989).
El C y el N total se analizaron ensubmuestras que habían sidotamizadas previamente en una tramade 2 mm. La concentración de Ctotal se midió usando un métodocolorimétrico tras una digestiónácida de las submuestras (Houba etal. 1988) y la de N total mediante elmétodo estándar de Kjeldahl (Krom1980).
Muestras de raíces y demacroinvertebradosSe muestrearon dos heces más porgrupo a las 0, 1 y 2 semanas y a los1, 2, 3.5, 5, 6.5, 8 y 11 mesesdespués de la deposición, paradeterminar la biomasa radicular y lade los macroinvertebrados (es decir,> 2 mm) presentes en las heces. Losmacroinvertebrados fueronrecolectados manualmente. Lasheces se lavaron y se tamizaron entrama de 0.053 mm para extraer deellas las raíces. Las raíces vivasfueron separadas manualmente,según su color, de las muertas y delos fragmentos de hojarasca. Losmacroinvertebrados se conservaronen alcohol al 70%. Tanto éstos comolos invertebrados se secaron luego alhorno a 75 ºC durante 48 horas y sepesaron.
Se tomaron al azar muestrasadicionales (diferentes de las hecesya identificadas) en los dos sistemasestudiados, con el fin de determinarel efecto de las heces en la biomasaradicular presente en el suelo. Estaseran muestras de suelo tomadas entres sitios: (1) un sitio libre de heces;(2) debajo de heces frescas; y(3) debajo de heces secas. En cadasitio se tomaron 8 muestras con uncilindro de 10 cm de diámetro y10 cm de profundidad. Las raícesfueron extraídas, secadas y pesadascomo se indicó anteriormente.
Figura 16-1. Descripción del procedimiento demuestreo. En cada fecha setomaron tres muestras de cadagrupo de heces: (1) de unturrículo, (2) del suelo subyacentey (3) del suelo adyacente.
Turrículo
5 cm
Sueloadyacente
Suelosubyacente
5 cm
20 cm
1
23
Gal
ería

Dinámica del Carbono y del Nitrógeno...
259
Tratamientos estadísticosLos datos se transformaron antes delanálisis para reducir la asimetría dela frecuencia de distribución. Lanormalización de los datos se obtuvocon la transformación Box-Cox(Sokal y Rohlf 1995), que es lasiguiente:
Los parámetros se calcularonaplicando el programa VerNorm 3.0del “módulo R” desarrollado porLegendre y Vaudor (1991).
Se llevó a cabo un ANOVA de tresvías en el cual los efectos fijosprincipales fueron el sistema de usode la tierra, el tipo de muestra(heces, suelo subyacente o sueloadyacente) y la edad de deposición.En estos análisis se emplearon lassiguientes variables: lasconcentraciones de NH4, de NO3 y deN inorgánico (en µg/g de suelo seco),la concentración de C total y de Ntotal (en % de suelo seco), la relaciónC:N, y el contenido de humedad (%).En total, fueron siete análisis consiete pruebas cada uno (tres deefectos principales más cuatro deinteracciones). Con el fin de evitar elerror estadístico de la significación,se aplicó la corrección de Bonferronipara pruebas encajadas (Cooper1968): los niveles ajustados designificación de 0.05, 0.01 y0.001 fueron entonces 0.001[= 0.05/(7x7)], 0.0002 [= 0.01/(7x7)]y 0.00002 [= 0.001/(7x7)],respectivamente.
Para la biomasa de las raíces y lade los macroinvertebrados (g pesoseco/g heces secas) se aplicó unANOVA de dos vías. Los principalesefectos considerados aquí fueron eltipo de sistema de uso de la tierra yla edad de las heces. Se aplicó lacorrección de Bonferroni, y losniveles de significación 0.05, 0.01 y0.001 se ajustaron así: 0.0125[= 0.05/(2x2)], 0.003 [= 0.01/(2x2)] y
0.0003 [= 0.001/(2x2)]. Se hicierontambién comparaciones entre lasmedias mediante una prueba PLSDde Fischer.
Resultados y Discusión
Comportamiento de laslombricesLa mayoría de las lombrices semantuvieron activas en sus galeríasdurante, al menos, la primerasemana después de que empezarona producir heces. Esto confirmó losresultados obtenidos en losestudios ecológicos anteriores sobreM. carimaguensis, que indicaban queesta especie tenía un hábito semi-sedentario (Jiménez et al. 1998b). Asípues, en ambos sistemas de uso de latierra las heces presentaban partesfrescas durante todo este períodosemanal y su contenido de humedadera superior al del suelo adyacente(Cuadro 16-1, Figura 16-2). Lacantidad de agua de las hecesdisminuyó drásticamente másadelante, cuando éstas tenían entre7 y 30 días de edad. Pasados30 días, la humedad de las hecesdisminuyó constantemente y su rangode variación, en comparación con lahumedad del suelo, aumentó. Eldescenso del contenido de aguaobservado 8 meses después del iniciodel experimento se correspondía conla parte media de la época seca.
Efecto de la actividad de laslombrices en la dinámica delnitrógenoSe observaron niveles altos de NH4 enlas heces de M. carimaguensisdurante su primer mes deenvejecimiento; en las heces de 1 díade edad, los valores fueron 5 y15 veces superiores (en la sabana yen el pastizal, respectivamente) que
y = (xδ −1) δδ
+ –
+
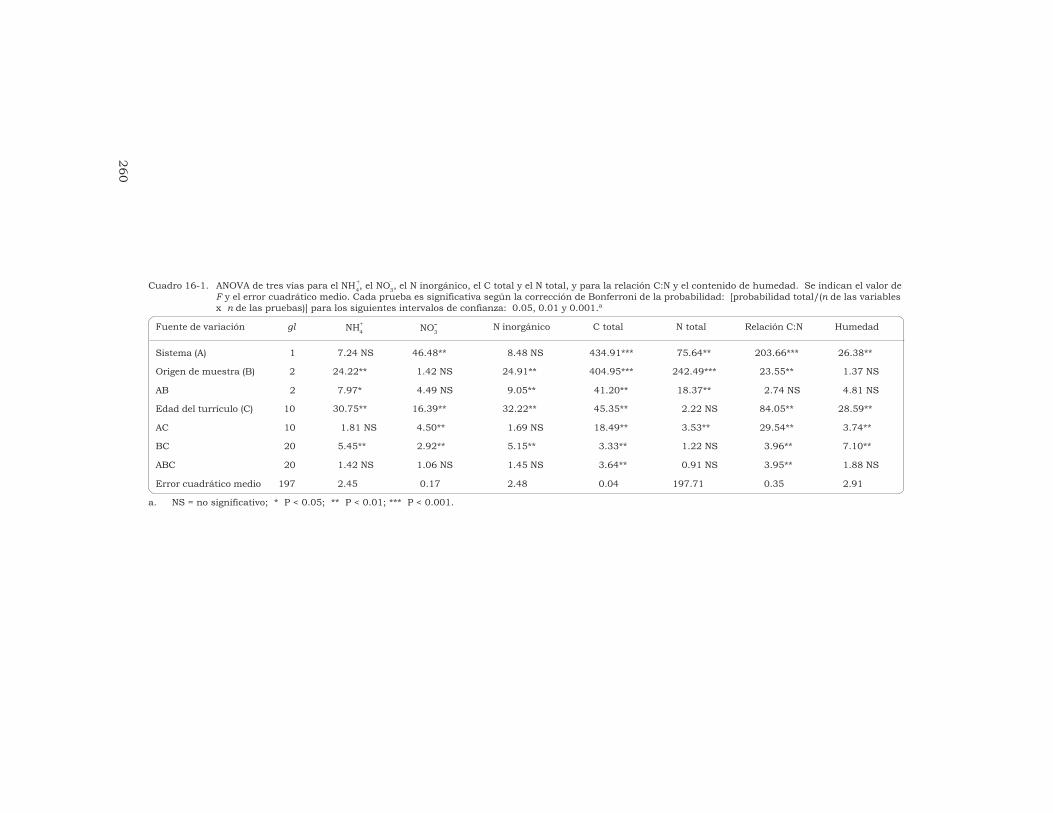
260
El A
rado Natural:…
Cuadro 16-1. ANOVA de tres vías para el NH4, el NO3, el N inorgánico, el C total y el N total, y para la relación C:N y el contenido de humedad. Se indican el valor deF y el error cuadrático medio. Cada prueba es significativa según la corrección de Bonferroni de la probabilidad: [probabilidad total/(n de las variablesx n de las pruebas)] para los siguientes intervalos de confianza: 0.05, 0.01 y 0.001.a
Fuente de variación gl N inorgánico C total N total Relación C:N Humedad
Sistema (A) 1 7.24 NS 46.48** 8.48 NS 434.91*** 75.64** 203.66*** 26.38**
Origen de muestra (B) 2 24.22** 1.42 NS 24.91** 404.95*** 242.49*** 23.55** 1.37 NS
AB 2 7.97* 4.49 NS 9.05** 41.20** 18.37** 2.74 NS 4.81 NS
Edad del turrículo (C) 10 30.75** 16.39** 32.22** 45.35** 2.22 NS 84.05** 28.59**
AC 10 1.81 NS 4.50** 1.69 NS 18.49** 3.53** 29.54** 3.74**
BC 20 5.45** 2.92** 5.15** 3.33** 1.22 NS 3.96** 7.10**
ABC 20 1.42 NS 1.06 NS 1.45 NS 3.64** 0.91 NS 3.95** 1.88 NS
Error cuadrático medio 197 2.45 0.17 2.48 0.04 197.71 0.35 2.91
a. NS = no significativo; * P < 0.05; ** P < 0.01; *** P < 0.001.
–+
NO3-NH4
+

Dinámica del Carbono y del Nitrógeno...
261
50
40
30
20
10
0
0 1 2 3 4 5 6 7 30 60 90 120 150 180 210 240 270 300 330
Hu
med
ad (
%)
50
40
30
20
10
0
Hu
med
ad (
%)
Turrículos Suelo subyacente Suelo adyacente
Días transcurridos desde la formación del turrículo
Figura 16-2. Variación del contenido de humedad del suelo y de los turrículos a lo largo del tiempo, enlos dos sistemas de uso de la tierra que se estudiaron.
Sabana nativa
Pastizal
en el suelo contiguo a ellas(Cuadro 16-1, Figura 16-3, A y B).Pasado ese tiempo, la concentraciónde NH4 bajó hasta un valor mínimo yconstante, similar al observado enlas muestras del suelo (Figura 16-3,A y B). Tanto en las heces como enel suelo subyacente y adyacente aellas, se observaron tres valoresmáximos sucesivos en laconcentración de NO3 (Cuadro 16-1,Figura 16-3, C y D). Al final delestudio, casi todo el exceso de N
inorgánico desapareció de las heces ydel suelo que las circundaba.
Los resultados obtenidosreflejaron la alta tasa demineralización que ocurre en lasheces frescas. En realidad, laactividad microbiana y lamineralización son enormementeestimuladas, primero durante eltránsito del suelo ingerido a lo largodel intestino de las lombrices y luegoen las heces frescas (Barois et al.
+
_
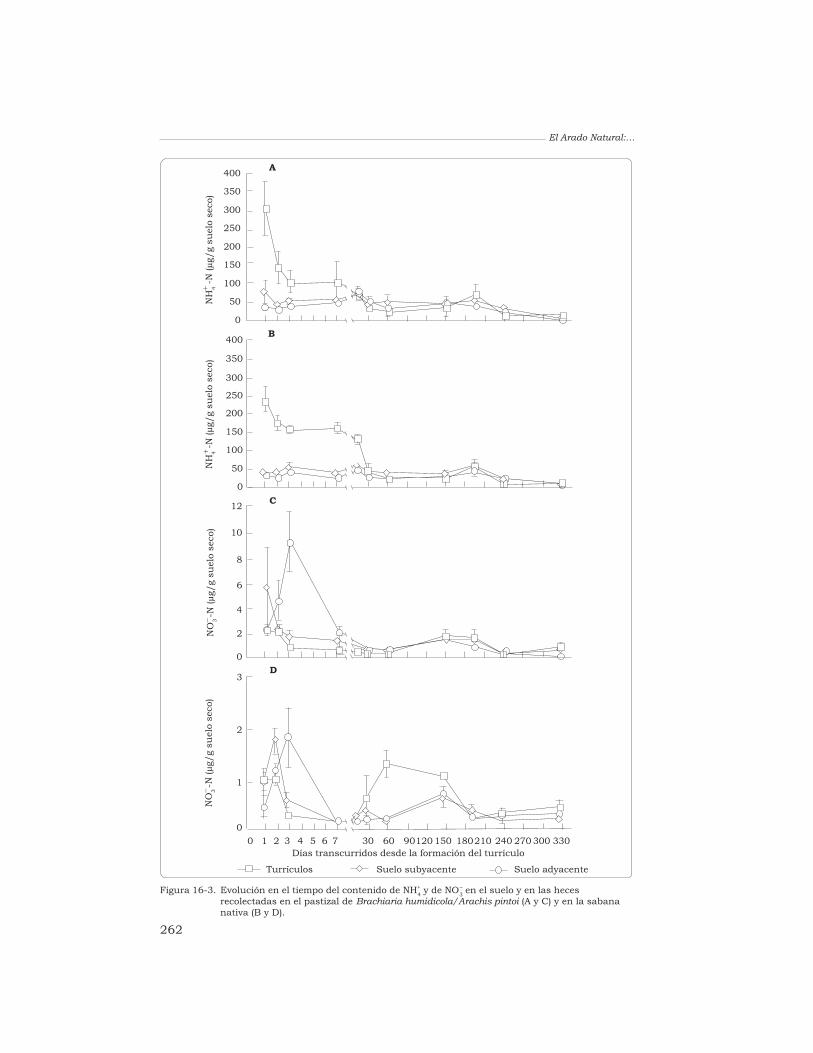
262
El Arado Natural:…
Figura 16-3. Evolución en el tiempo del contenido de NH4 y de NO3 en el suelo y en las hecesrecolectadas en el pastizal de Brachiaria humidicola/Arachis pintoi (A y C) y en la sabananativa (B y D).
+ –
C
400
350
300
250
200
150
100
50
0B400
350
300
250
200
150
100
50
0
3
2
1
0
A
D
0 1 2 3 4 5 6 7 30 60 90120 150 180210 240 270 300 330Días transcurridos desde la formación del turrículo
12
10
8
6
4
2
0
Turrículos Suelo subyacente Suelo adyacente
NH
4 -N
(µg/
g su
elo
seco
)
+
NH
4 -N
(µg/
g su
elo
seco
)
+
NO
3 -N
(µg/
g su
elo
seco
)–
NO
3 -N
(µg/
g su
elo
seco
)–

Dinámica del Carbono y del Nitrógeno...
263
Pasado este primer período enque la dinámica del N fue rápida, lasheces se secaron rápidamente en lasuperficie del suelo (Figura 16-2).Las heces secas están formadas poragregados estables, impermeables yun poco compactos (Blanchart et al.1993; Guggenberger et al. 1996;Shipitalo y Protz 1989), que protegeneficazmente la MO contenida en ellosde los procesos de descomposiciónposteriores (Lavelle y Martin 1992;Martin 1991). En este experimento,sin embargo, hubo un ligeroincremento en la concentración deNH4 y de NO3 de los 30 a los195 días después de su inicio, tantoen las heces como en el suelo
1987). Este proceso, sumado a laadición de orina en la parte final deltubo digestivo de la lombriz, explicalos altos valores de NH4 observadosen las heces frescas. Estos nivelesse mantuvieron durante el primermes por la excreción continua deheces sobre una misma deposición.Después de ese tiempo, las lombricesabandonaron las galerías y laconcentración de NH4 en las hecesdisminuyó progresivamente. Estadisminución puede atribuirse a laproducción de NO3 en las hecesfrescas mediante la nitrificación(Lavelle y Martin 1992; Scheu 1987).Una difusión rápida de NO3 hacia elsuelo sería la razón de que no sehaya observado una acumulación deeste ion en las heces. Por tanto, lostres valores máximos de NO3 sepueden interpretar del modosiguiente (Figura 16-4):
- producción rápida de NO3 en lasheces;
- difusión rápida de este NO3 en elagua que fluye por la galería enque habita la lombriz;
- difusión lateral, más lenta, con elagua de escorrentía que lava lasheces y la superficie del suelo(Figura 16-4).
La intensidad creciente de lostres picos de concentración del NO3podría reflejar la estimulación, en elsuelo alrededor de las heces, de ungrupo de bacterias (microflora) quedependen de varios nutrientes. Porúltimo, los excesos de N inorgánicodesaparecieron de las heces y delsuelo cercano a ellas por laasimilación de las raíces, por losprocesos de desnitrificación, por lareorganización de la biomasamicrobiana del suelo, o comopérdidas debidas a la lixiviación(Elliott et al. 1990; Lavelle y Martin1992; Syers et al. 1979).
Lluvia
Turrículo5
1
3
6
3
5
4
2
4
5
Gal
ería
20 cm
Figura 16-4. Dinámica del N en las heces y en elsuelo que las circunda. Lasflechas indican la dirección delflujo del agua y los númerosrepresentan un orden cronológico:(1) fuerte liberación de NH4 en lasheces frescas seguida deproducción de NO3; (2) filtraciónvertical por la galería;(3) escorrentía lateral desde elturrículo y en la superficie delsuelo; (4) lixiviación vertical dentrodel suelo; (5) desnitrificación másabsorción por raíces y pormicroorganismos;(6) inmovilización del N en lasheces secas.
+
–
+
+
_
_
_
_
_
_
+ _
_
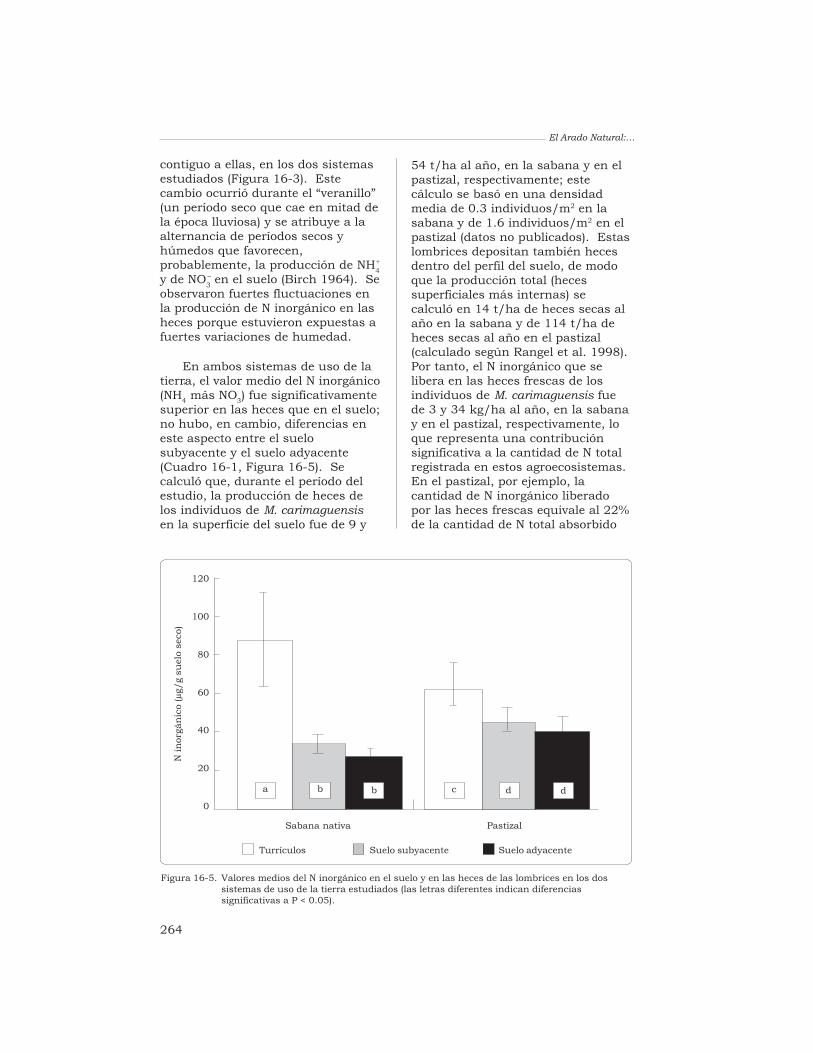
264
El Arado Natural:…
Figura 16-5. Valores medios del N inorgánico en el suelo y en las heces de las lombrices en los dossistemas de uso de la tierra estudiados (las letras diferentes indican diferenciassignificativas a P < 0.05).
120
100
80
60
40
20
0
N in
orgá
nic
o (µ
g/g
suel
o se
co)
Sabana nativa Pastizal
Turrículos Suelo subyacente Suelo adyacente
a b b c d d
contiguo a ellas, en los dos sistemasestudiados (Figura 16-3). Estecambio ocurrió durante el “veranillo”(un período seco que cae en mitad dela época lluviosa) y se atribuye a laalternancia de períodos secos yhúmedos que favorecen,probablemente, la producción de NH4y de NO3 en el suelo (Birch 1964). Seobservaron fuertes fluctuaciones enla producción de N inorgánico en lasheces porque estuvieron expuestas afuertes variaciones de humedad.
En ambos sistemas de uso de latierra, el valor medio del N inorgánico(NH4 más NO3) fue significativamentesuperior en las heces que en el suelo;no hubo, en cambio, diferencias eneste aspecto entre el suelosubyacente y el suelo adyacente(Cuadro 16-1, Figura 16-5). Secalculó que, durante el período delestudio, la producción de heces delos individuos de M. carimaguensisen la superficie del suelo fue de 9 y
54 t/ha al año, en la sabana y en elpastizal, respectivamente; estecálculo se basó en una densidadmedia de 0.3 individuos/m2 en lasabana y de 1.6 individuos/m2 en elpastizal (datos no publicados). Estaslombrices depositan también hecesdentro del perfil del suelo, de modoque la producción total (hecessuperficiales más internas) secalculó en 14 t/ha de heces secas alaño en la sabana y de 114 t/ha deheces secas al año en el pastizal(calculado según Rangel et al. 1998).Por tanto, el N inorgánico que selibera en las heces frescas de losindividuos de M. carimaguensis fuede 3 y 34 kg/ha al año, en la sabanay en el pastizal, respectivamente, loque representa una contribuciónsignificativa a la cantidad de N totalregistrada en estos agroecosistemas.En el pastizal, por ejemplo, lacantidad de N inorgánico liberadopor las heces frescas equivale al 22%de la cantidad de N total absorbido
+_
+ _

Dinámica del Carbono y del Nitrógeno...
265
por las gramíneas, es decir, unos155 kg/ha al año (Fisher y Kerridge1996) y al 48% del N total aportado alos cultivos de arroz, o sea, 70 kg/haal año (Thomas et al. 1995). Loscálculos aquí presentadosconcuerdan con otros sobre elimpacto de las lombrices de tierra enla mineralización del N (Curry et al.1995; James 1991; Martin 1991).Ahora bien, la contribución total deestas lombrices a la cantidad de Ndisponible para las plantas puede serincluso superior, dada la presenciade poblaciones abundantes de otrasespecies de lombrices (Jiménez et al.1998a) y la producción de N mineralmediante otros procesos, como laexcreción de orina o de moco cutáneopor las lombrices o la descomposiciónde sus tejidos muertos (Scheu 1994).
Acumulación de C en lasestructuras producidas por laslombricesEn los suelos que no habían tenidoun contacto reciente con laslombrices, el contenido de C total fuesignificativamente mayor en elpastizal que en la sabana, pero nohubo diferencias significativasrespecto al N total ni a la relación C:N(Figura 16-6). Estos resultadosapoyan los de otros estudios dondese demuestra que, en los pastizalesde gramíneas y leguminosas queestán sembrados en tierras desabana, aumenta generalmente,hasta cierto punto, la cantidad de Corgánico del suelo; en cambio, laconcentración de N de dichospastizales permanece más o menosigual (Thomas et al. 1995). Elaumento progresivo del C y de larelación C:N observados en el suelodel pastizal (Figura 16-7, A y E) sedeben, probablemente, a la presenciade las jaulas metálicas que protegíanla vegetación contra el pastoreo y quepermitían la acumulación en el suelo
de mayores cantidades de hojarasca yde MO fresca.
Las heces producidas en ambossistemas de uso de la tierrapresentaron contenidossignificativamente mayores de C(de 1.5 a 1.9 veces más) y de N (de 1.4a 1.6 veces más) que el sueloalrededor de ellas (Figura 16-6). Sehan reportado resultados similares enotros estudios, que se explican por lacapacidad de los individuos deM. carimaguensis para seleccionar unsustrato rico en contenido orgánico(Jiménez et al. 1998b; Rangel et al.1998). Se observaron tambiéncantidades importantes de residuosvegetales reconocibles que oscilaban,en promedio, de 1% a 3% del pesoseco total de la deposición, en lasabana y en el pastizal (datos nopresentados). Queda confirmada asíla capacidad de M. carimaguensis deingerir una mezcla de suelo y dehojarasca fresca, y la pertenencia deesta especie a la categoría ecológica delas lombrices anécicas (sensu Bouché1977). Esta característica sería laexplicación de que los niveles de Ctotal y de N total de las hecesproducidas en el pastizal, donde haymayor disponibilidad de hojarasca dela leguminosa, fueron superiores quelos observados en la sabana nativa,donde los residuos vegetales eran másescasos.
La concentración de N total fuemás o menos constante durante elenvejecimiento de las heces(Cuadro 16-1, Figura 16-7, C y D). Lomás sorprendente fue, quizás, elaumento continuo y significativo, enambos sistemas de uso de la tierra, dela cantidad de C de las heces duranteese proceso de envejecimiento(Figura 16-7, A y B). El aumento(≈ +100%) se explica por lacombinación de los siguientesfactores, cuya importancia relativa esaún desconocida:
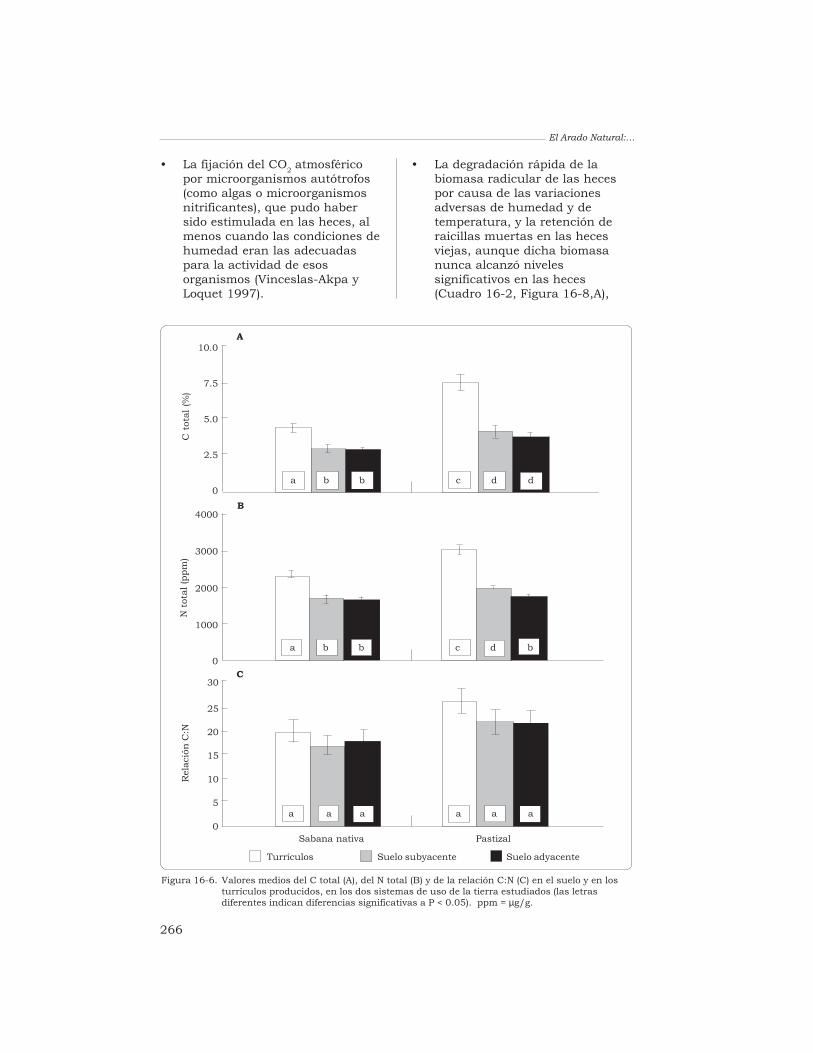
266
El Arado Natural:…
10.0
7.5
5.0
2.5
0
Rel
ació
n C
:N
Sabana nativa Pastizal
4000
3000
2000
1000
0
30
25
20
15
10
5
0
Turrículos Suelo subyacente Suelo adyacente
N t
otal
(ppm
)C
tot
al (%
)
Figura 16-6. Valores medios del C total (A), del N total (B) y de la relación C:N (C) en el suelo y en losturrículos producidos, en los dos sistemas de uso de la tierra estudiados (las letrasdiferentes indican diferencias significativas a P < 0.05). ppm = µg/g.
a b b c d d
a b b c d b
a a a a a a
A
B
C
• La fijación del CO2 atmosféricopor microorganismos autótrofos(como algas o microorganismosnitrificantes), que pudo habersido estimulada en las heces, almenos cuando las condiciones dehumedad eran las adecuadaspara la actividad de esosorganismos (Vinceslas-Akpa yLoquet 1997).
• La degradación rápida de labiomasa radicular de las hecespor causa de las variacionesadversas de humedad y detemperatura, y la retención deraicillas muertas en las hecesviejas, aunque dicha biomasanunca alcanzó nivelessignificativos en las heces(Cuadro 16-2, Figura 16-8,A),
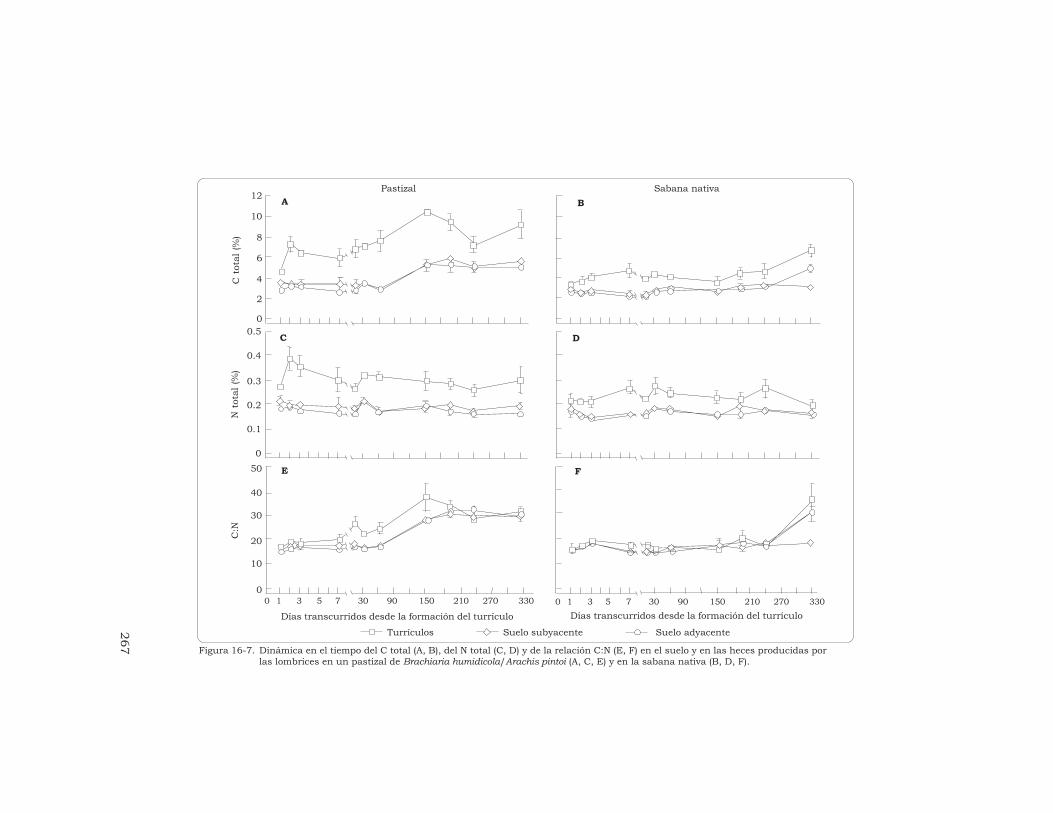
Dinám
ica del Carbono y del N
itrógeno...
267
Figura 16-7. Dinámica en el tiempo del C total (A, B), del N total (C, D) y de la relación C:N (E, F) en el suelo y en las heces producidas porlas lombrices en un pastizal de Brachiaria humidicola/Arachis pintoi (A, C, E) y en la sabana nativa (B, D, F).
Días transcurridos desde la formación del turrículo
12
10
8
6
4
2
00.5
0.4
0.3
0.2
0.1
050
40
30
20
10
0
A B
D
F
C
E
0 1 3 5 7 30 90 150 210 270 330 0 1 3 5 7 30 90 150 210 270 330Días transcurridos desde la formación del turrículo
Pastizal Sabana nativa
Turrículos Suelo subyacente Suelo adyacente
C:N
N t
otal
(%)
C t
otal
(%)
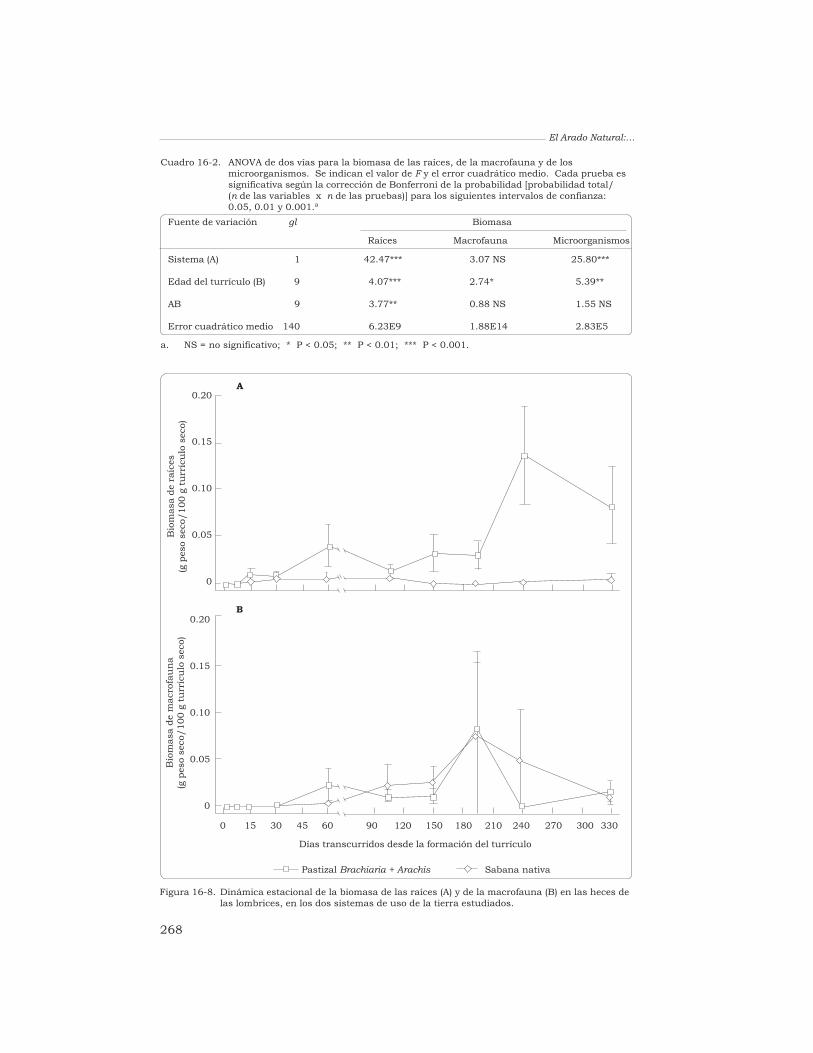
268
El Arado Natural:…
0 15 30 45 60 90 120 150 180 210 240 270 300 330
Bio
mas
a de
mac
rofa
un
a(g
pes
o se
co/1
00 g
tu
rríc
ulo
sec
o)
0.20
0.15
0.10
0.05
0
0.20
0.15
0.10
0.05
0
Bio
mas
a de
raí
ces
(g p
eso
seco
/100
g t
urr
ícu
lo s
eco)
Días transcurridos desde la formación del turrículo
B
A
Pastizal Brachiaria + Arachis Sabana nativa
Figura 16-8. Dinámica estacional de la biomasa de las raíces (A) y de la macrofauna (B) en las heces delas lombrices, en los dos sistemas de uso de la tierra estudiados.
Cuadro 16-2. ANOVA de dos vías para la biomasa de las raíces, de la macrofauna y de losmicroorganismos. Se indican el valor de F y el error cuadrático medio. Cada prueba essignificativa según la corrección de Bonferroni de la probabilidad [probabilidad total/(n de las variables x n de las pruebas)] para los siguientes intervalos de confianza:0.05, 0.01 y 0.001.a
Fuente de variación gl Biomasa
Raíces Macrofauna Microorganismos
Sistema (A) 1 42.47*** 3.07 NS 25.80***
Edad del turrículo (B) 9 4.07*** 2.74* 5.39**
AB 9 3.77** 0.88 NS 1.55 NS
Error cuadrático medio 140 6.23E9 1.88E14 2.83E5
a. NS = no significativo; * P < 0.05; ** P < 0.01; *** P < 0.001.

Dinámica del Carbono y del Nitrógeno...
269
Efecto de la deposición de lasheces en el crecimiento de lasraíces superficialesLa biomasa de las raíces de la capasuperficial del suelo respondiósignificativamente a la presencia deheces en la superficie del suelo, apesar de que se encontraroncantidades muy pequeñas de raícesen el interior de las heces(Figura 16-9). En comparación conel suelo testigo que no tenía heces,este parámetro aumentó dos vecesen el suelo situado bajo las hecesrecientes y cinco veces en el queestaba bajo las heces más viejas. Enla sabana se observó la mismatendencia, aunque no de un modosignificativo.
Varios estudios handocumentado ampliamente losefectos que causan las lombrices enel crecimiento de las plantas, enespecial los estudios de cortaduración con plantas cultivadas eninvernadero (ver revisión de Brownet al. 1999; Lavelle 1997). Estosefectos provienen de diversosmecanismos, algunos de los cualesfueron confirmados por el presentetrabajo. La deposición de heces porlos individuos de M. carimaguensisestimula primero la movilización delos nutrientes (por ejemplo, del NH4 ydel NO3 en este estudio) quegeneralmente no están disponiblesen el suelo (Rangel et al. 1998).Asimismo, estando activa estaespecie al inicio de la época lluviosa,principalmente (Jiménez et al.1998b), el período en que laslombrices inducen la mineralizaciónpuede coincidir con la época demáximo requerimiento de nutrientespor las plantas.
Como conclusión, el presenteestudio confirma la hipótesis de quehay efectos derivados de la actividadde las lombrices sobre la MO delsuelo que contrastan entre sí, según
• La acumulación de materialorgánico y la producción debolitas fecales ricas en C (de5.2% a 10.2% de C, datos nopublicados), provenientesambos de losmacroinvertebrados que, desdeel primer mes del experimento,se encontraban en las heces(Cuadro 16-2, Figura 15-8,B),pudieron contribuir al aumentoen ellas de la concentraciónde C.
Un efecto posible de laconcentración de C orgánico en lasheces y en las galerías de laslombrices de la especieM. carimaguensis es la formación deuna reserva de C relativamenteactivo, que está protegidofísicamente por las heces y se liberaa medida que éstas se desintegran(Guggenberger et al. 1996; Lavelle yMartin 1992). La gran cantidad desuelo excretado en las heces por laslombrices puede propiciar unaconsiderable acumulación de C enlos agregados estables de las heces.Esta cantidad se ha calculado en0.6 t/ha al año en la sabana y8.6 t/ha al año en el pastizal. Partede este C (83% en la sabana y 62%en el pastizal) correspondió alaumento de C observado en lasheces frescas (en comparación conel del suelo) y puede ser el resultadode la ingestión selectiva que hacenlas lombrices de sustratos ricos enmaterial orgánico. El C restanteprovenía del aumento en laconcentración de C que se presentatras la producción de las heces. Laconcentración total de C en lasheces depositadas por individuos dela especie M. carimaguensisrepresentó el 2% y el 30% (en lasabana y en el pastizal,respectivamente) de todo el Ccontenido en los primeros 10 cm delsuelo, partiendo de una densidadaparente de 1.0 g/cm3.
+_
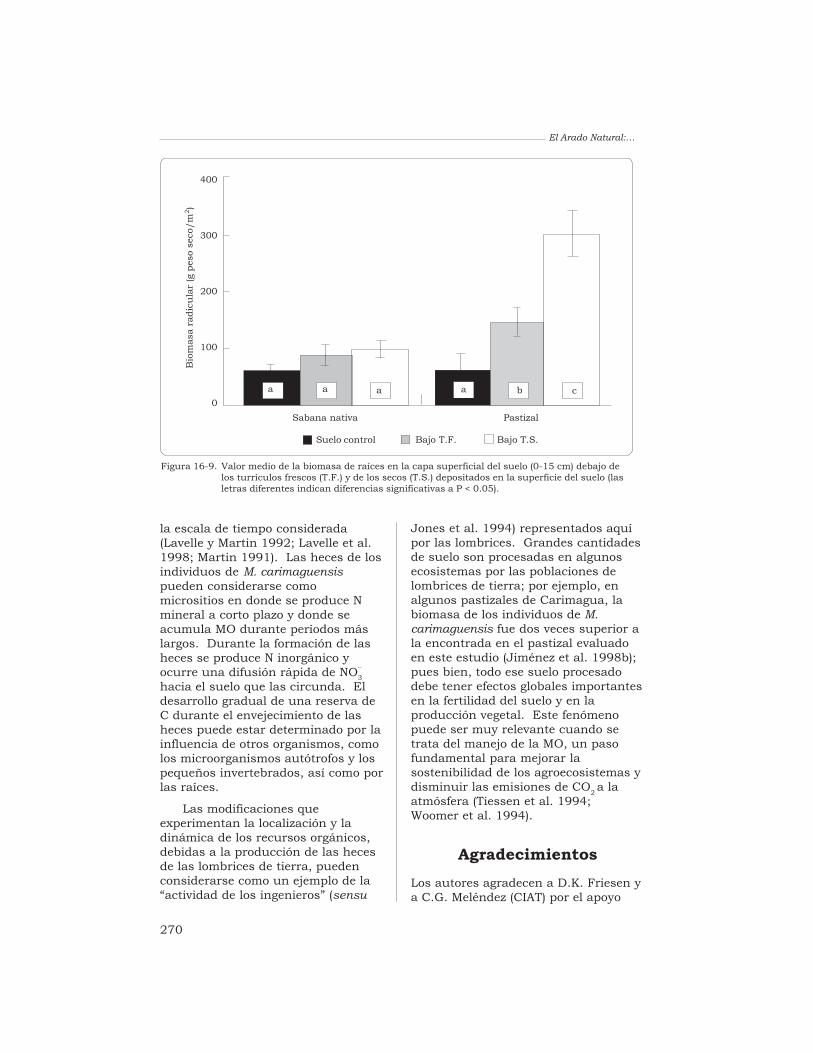
270
El Arado Natural:…
la escala de tiempo considerada(Lavelle y Martin 1992; Lavelle et al.1998; Martin 1991). Las heces de losindividuos de M. carimaguensispueden considerarse comomicrositios en donde se produce Nmineral a corto plazo y donde seacumula MO durante períodos máslargos. Durante la formación de lasheces se produce N inorgánico yocurre una difusión rápida de NO3hacia el suelo que las circunda. Eldesarrollo gradual de una reserva deC durante el envejecimiento de lasheces puede estar determinado por lainfluencia de otros organismos, comolos microorganismos autótrofos y lospequeños invertebrados, así como porlas raíces.
Las modificaciones queexperimentan la localización y ladinámica de los recursos orgánicos,debidas a la producción de las hecesde las lombrices de tierra, puedenconsiderarse como un ejemplo de la“actividad de los ingenieros” (sensu
Jones et al. 1994) representados aquípor las lombrices. Grandes cantidadesde suelo son procesadas en algunosecosistemas por las poblaciones delombrices de tierra; por ejemplo, enalgunos pastizales de Carimagua, labiomasa de los individuos de M.carimaguensis fue dos veces superior ala encontrada en el pastizal evaluadoen este estudio (Jiménez et al. 1998b);pues bien, todo ese suelo procesadodebe tener efectos globales importantesen la fertilidad del suelo y en laproducción vegetal. Este fenómenopuede ser muy relevante cuando setrata del manejo de la MO, un pasofundamental para mejorar lasostenibilidad de los agroecosistemas ydisminuir las emisiones de CO2 a laatmósfera (Tiessen et al. 1994;Woomer et al. 1994).
AgradecimientosLos autores agradecen a D.K. Friesen ya C.G. Meléndez (CIAT) por el apoyo
Bio
mas
a ra
dicu
lar
(g p
eso
seco
/m2 )
400
300
200
100
0Sabana nativa Pastizal
Suelo control Bajo T.F. Bajo T.S.
Figura 16-9. Valor medio de la biomasa de raíces en la capa superficial del suelo (0-15 cm) debajo delos turrículos frescos (T.F.) y de los secos (T.S.) depositados en la superficie del suelo (lasletras diferentes indican diferencias significativas a P < 0.05).
a a a a b c
_

Dinámica del Carbono y del Nitrógeno...
271
técnico recibido; a M. Vinceslas-Akpay a M. Loquet (de la Universidad deRouen) y a P. Lavelle, L. Mariani yJ.-P. Rossi (IRD) por sus útilessugerencias a una primera versiónde este artículo.
Referencias
Anderson JM. 1995. Soil organisms asengineers: microsite regulation ofmacroscale processes. En: JonesCG, Lawton JH, eds. Linkingspecies and ecosystems. Chapmanand Hall, Nueva York. p 94-106.
Anderson JM; Ingram JS, eds. 1989.Tropical soil biology and fertility, ahandbook of methods. 2a. ed. CABInternational, Oxford, Reino Unido.
Birch AF. 1964. Mineralization of plantnitrogen following alternance of wetand dry conditions. Plant Soil20:43-49.
Barois I; Verdier B; Kaiser P; Mariotti A;Rangel P; Lavelle P. 1987. Influenceof the tropical earthwormPontoscolex corethrurus(Glossoscolecidae) on the fixationand mineralisation of nitrogen. En:Bonvicini Pagliai AM; Omodeo P,eds. On earthworms. MuchiEditore, Modena, Italia. p 151-158.
Blair JM; Parmelee RW; Lavelle P. 1994.Influences of earthworms onbiochemistry in North Americanecosystems. En: Hendrix PH, ed.Earthworm ecology in forest,rangeland and crop ecosystems ofNorth America. Lewis Publishers,Chelsea, Estados Unidos. p 1-44.
Blanchart E. 1993. The physicalstructure of casts of Millsoniaanomala (Oligochaeta:Megascolecidae) in shrub savannasoils (Côte d’Ivoire). Geoderma56:119-132.
Blanchart E; Lavelle P; Braudeau E;Bissonnais YL; Valentin C. 1997.Regulation of soil structure bygeophagous earthworm activities inhumid savannas of Côte d’Ivoire.Soil Biol Biochem 29:431-439.
Bouché MB. 1977. Stratégieslombriciennes. En: Lohm U;Persson T, eds. Soil organisms ascomponent of ecosystems. Ecol Bull25:122-132.
Brown G; Pashanasi B; Gilot-Villenave C;Patrón JC; Senapati BK; Giri S;Barois I; Lavelle P; Blanchart E;Blakemore RJ; Spain AV; Boyer J.1999. Effects of earthworms onplant production in the tropics. En:Lavelle P; Brussaard L; Hendrix PF,eds. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido. p 87-147.
Cooper DW. 1968. The significance levelin multiple tests madesimultaneously. Heredity23:614-617.
Curry JP; Byrne D; Boyle KE. 1995. Theearthworm population of a wintercereal field and its effects on soiland nitrogen turnover. Biol FertilSoils 19:166-172.
Elliot PW; Knight D; Anderson JM. 1990.Denitrification in earthworm castsand soil from pastures underdifferent fertilizer and drainageregimes. Soil Biol Biochem22:601-605.
Fisher MJ; Kerridge PC. 1996. Theagronomy and physiology ofBrachiaria species. En: Miles JW;Mass BL; Valle CB do, eds.Brachiaria: biology, agronomy andimprovement. CIAT, Cali, Colombia.p 43-52.
Fragoso C; Brown GG; Patrón JC;Blanchart E; Lavelle P; PashanasiB; Senapati S; Kumar T. 1997.Agricultural intensification, soilbiodiversity and agroecosystemfunction in the tropics: the role ofearthworms. Appl Soil Ecol 6:17-35.

272
El Arado Natural:…
Guggenberger G; Thomas RJ; Zech W.1996. Soil organic matter withinearthworm casts of an anecic-endogeic tropical pasturecommunity. Appl Soil Ecol3:263-274.
Houba VJC; Lee van der JJ; NovozamskyI; Walinga J. 1988. Soil and plantanalysis. En: Department of SoilScience and Plant Nutrition, ed. Soilanalysis procedures. 4a. ed.Agricultural University,Wageningen, Holanda. p 4-14.
James SW. 1991. Soil, nitrogen,phosphorus, and organic matterprocessing by earthworms intallgrass prairies. Ecology72:2101-2109.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher M; Thomas RJ.1998a. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998b. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the EasternPlains of Colombia. Appl Soil Ecol9:153-160.
Jones CJ; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Krom M. 1980. Spectrophotometricdetermination of ammonia: a studyof modified Bertheloy reaction usingsalicylate and dichloroisocyanurate.The Analyst 105:305-316.
Lavelle P. 1979. Relations entre typesécologiques et profilsdémographiques chez les vers deterre de la savane de Lamto (Côted’Ivoire). Rev Ecol Biol Sol16:85-101.
Lavelle P. 1997. Faunal activities and soilprocesses: Adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P; Martin A. 1992. Small-scale andlarge-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humidtropics. Soil Biol Biochem24:1491-1498.
Lavelle P; Pashanasi B; Charpentier F;Gilot C; Rossi J-P; Derouard L;Andre J; Ponge JP; Bernier N. 1998.Large-scale effects of earthworms onsoil organic matter and nutrientdynamics. En: Edwards CA, ed.Earthworm ecology. St. Lucies Press,Boca Ratón, Estados Unidos.p 103-122.
Legendre P; Vaudor A. 1991. The Rpackage: multidimensional analysis,spatial analysis. Université deMontréal, Département des SciencesBiologiques, Montreal, Canadá.
Martin A. 1991. Short- and long-termeffects of the endogeic earthwormMilsonia anomala (Omodeo)(Megascolecidae, Oligochaeta) oftropical savannas, on soil organicmatter. Biol Fertil Soils 11:234-238.
Rangel AF; Thomas RJ; Madero E; LavelleP; Friesen DK; Jiménez JJ; DecaënsT; Amézquita E. 1997. Efecto de lalombriz de tierra Martiodriluscarimaguensis sp. nov. en ladinámica del nitrógeno de un Oxisolde los Llanos Orientales deColombia. Suelos Ecuatoriales27:235-241.
Rippstein G; Lascano C; Decaëns T. 1996.La production fourragère dans lessavanes d’Amérique du Sudintertropicale. Fourrages 145:33-52.
Scheu S. 1987. The influence ofearthworms (Lumbricidae) on thenitrogen dynamics in the soil littersystem of a deciduous forest.Oecologia 72:197-201.
Scheu S. 1994. There is an earthwormmobilizable pool in soil. Pedobiologia38:243-249.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research, 3a. ed. WHFreeman, Nueva York, EstadosUnidos.

Dinámica del Carbono y del Nitrógeno...
273
Syers JK; Sharpley AN; Keeney DR. 1979.Cycling of nitrogen by surface-casting earthworms in a pastureecosystem. Soil Biol Biochem11:181-185.
Thomas RJ; Fisher MJ; Ayarza MA; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. LewisPublishers, Boca Ratón, EstadosUnidos. p 61-83.
Tiessen H; Cuevas E; Chacón N. 1994.The role of organic matter insustaining soil fertility. Nature371:783-785.
Vera RR; Seré CO, eds. 1985. Sistemas deproducción pecuaria extensiva:Brasil, Colombia, Venezuela.Informe final del Proyecto ETES,1978-1982. CIAT, Cali, Colombia.
Vinceslas-Akpa M; Loquet M. 1997.Organic matter transformations inlignocellulosic waste productscomposted or vermicomposted(Eisenia fetida andrei): chemicalanalysis and 13C CPMAS NMRspectroscopy. Soil Biol Biochem29:751-758.
Woomer PL; Martin A; Albrecht A; ResckDVS; Scharpenseel HW. 1994. Theimportance and management of soilorganic matter in the tropics. En:Woomer PL; Swift MJ, eds. Thebiological management of soilfertility. Wiley-Sayce Publication.p 47-80.

274
El Arado Natural:…
ResumenEn los suelos tropicales predominanlos minerales de carga variable. Elpunto de carga cero (PCC) es funciónde los diferentes constituyentesminerales y orgánicos del suelo, y elgrado en que éstos interactúandetermina la expresión de la cargavariable. La presencia de la materiaorgánica (MO) afecta el PCC porquemodifica la carga total del suelo y,por consiguiente, la capacidad deéste para retener iones encompetencia con la lixiviación. Engeneral, la cantidad de MO en lasheces de las lombrices es mayor queen el suelo que las circunda. Estacantidad alta de MO puede inducirun cambio del PCC hacia un pH másbajo, lo que mejora la capacidad decarga y la adsorción de cationes y deaniones y reduce, por ende, el riesgode lixiviación. En este estudio seanaliza la influencia de la lombriz
Martiodrilus carimaguensis Jiménez yMoreno en el cambio del PCC, enuna parcela de sabana nativa y endos pastizales mejorados de losLlanos Orientales de Colombia. ElPCC se determinó, mediantetitulación protónica, en las heces yen el suelo que las circundaba, conel fin de evaluar la cantidad de cargapresente en la superficie de laspartículas coloidales a diferentesvalores tanto de pH como deconcentración de electrolitos. En losdos sistemas de uso de la tierra, elPCC de las heces se desplazó haciavalores más ácidos, y esto aumentóla generación y la retención de lacarga a niveles naturales de pH(pH en H2O). El nivel de Al+3
intercambiable de las heces fueinferior al encontrado en el suelo noingerido, mientras que los niveles deCa+2 y de Mg+2 aumentaron en ellas.En los pastizales introducidos seincrementó la diferencia entre el PCCy el pH (de +42% a +56%), encomparación con la sabana. Losresultados obtenidos se atribuyen ala capacidad de las lombrices deconcentrar la MO en las hecescuando ingieren sustratos ricos enMO. La modificación del PCC en lasheces implica una mejora de lacapacidad de intercambio catiónico,que podría aumentar ladisponibilidad y la retención de losnutrientes.
CAPÍTULO 17
Características del Intercambio Iónicoen las Heces de la Lombriz de TierraAnécica Martiodrilus carimaguensisJiménez y Moreno, en un Oxisol deuna Sabana de Colombia1
A. F. Rangel* , E. Madero**, R. J. Thomas*,D. K. Friesen* y T. Decaëns***
1. Publicado en Pedobiologia 43, p 795-801, porRangel et al. Copyright © 1999 de Urban &Fischer Verlag. Impreso con permiso de lacasa editorial.
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
** Universidad Nacional de Colombia, SedePalmira, Unidad de Suelos, AA 237, Palmira,Colombia.
*** Laboratoire d’Ecologie des Sols Tropicaux,IRD-Bondy, 32 Av. Henri Varagnat, F-93143Bondy Cedex, Francia.

Características del Intercambio Iónico...
275
Palabras clave: lombrices,Martiodrilus carimaguensis, puntode carga cero (PCC), suelosácidos
IntroducciónLos coloides del suelo estánconstituidos por mezclas depolímeros orgánicos e inorgánicosderivados de las alteracionesproducidas por los agentesatmosféricos (Sposito y Schindler1986). La meteorización origina unagran cantidad de óxidos de hierro yde aluminio (óxidos y oxi-hidróxidos);estos minerales presentan una cargaeléctrica superficial debida a laprotonación y a la desprotonación delos grupos hidroxílicos de susuperficie. Las arcillas que poseeneste tipo de carga reciben el nombrede arcillas de carga variable. Losgrupos funcionales, como lospolímeros orgánicos (carboxílicos,fenólicos y enólicos), la materiaorgánica del suelo (MO) y los alófanoscontribuyen también a crear unacarga variable (Oades et al. 1989).
Las condiciones ambientalesafectan mucho la carga variable de lasuperficie del suelo, que será positivao negativa según el pH y la fuerzaiónica de los electrolitos de lasolución que estén en contacto conlos polímeros. Habrá entonces unvalor determinado de pH para el cualla carga neta es cero (PCC), puntoque representará un número igual delugares protonados y desprotonadosen el suelo (Van Raij y Peech 1972).La MO del suelo, material de cargavariable por sí misma, puede afectarsensiblemente el PCC y la carga totaldel suelo y, por tanto, la capacidadde retener iones en oposición a lalixiviación (Oades et al. 1989).
Los minerales de carga variableson abundantes en los suelostropicales. Si se mantienen losvalores de la MO tan altos como sea
posible, los valores del PCC seránmínimos; por su parte, la fertilizaciónpuede mantener el pH del suelo alnivel más alto que convenga en lapráctica. Estas prácticas de manejopueden contribuir ampliamente a lacapacidad que tienen estos suelospara retener los cationes que sonnutrientes para las plantas (Oades etal. 1989).
Los invertebrados grandes delsuelo, principalmente las lombrices,las termitas y las hormigas, seconsideran “ingenieros delecosistema del suelo” (Lavelle 1997).En el trópico húmedo, donde ni losprocesos de meteorización ni lavegetación son constantes y dondelas arcillas desempeñan un papel demenor importancia en la regulaciónde la mineralización, la fauna delsuelo se convierte en el principalregulador de la dinámica de la MOdel suelo (Lavelle y Martin 1992).Estos ingenieros del suelo regulan laactividad de los microorganismos enlas estructuras órgano-minerales queproducen (por ejemplo, en las hecesde las lombrices, en los termiteros yen los hormigueros), lo cual afecta laagregación y la dinámica de la MOdel suelo (Decaëns et al. 1999;Lavelle y Martin 1992).
La amplitud de tales efectospuede ser considerable en los suelosen que la actividad de las lombriceses alta. La cantidad total de sueloingerido por una comunidad delombrices se ha calculado entre600 y 1250 t/ha de suelo seco al añoo, lo que es lo mismo, de 14 a15 t/ha de MO al año en las sabanas(Lavelle 1978). En los LlanosOrientales de Colombia, Jiménez etal. (1998) calcularon que laproducción de heces (dentro delsuelo y en su superficie) de unalombriz anécica era de 14.3 t/ha deheces secas al año en la sabananativa, y de 378 t/ha de heces secasal año en un pastizal introducido.

276
El Arado Natural:…
Por tanto, las lombrices de tierrapueden afectar el PCC y laspropiedades de carga del suelo pormedio de las estructuras órgano-minerales que producen (las heces).El objetivo de este estudio fueevaluar el efecto que ejercía unaespecie de lombriz de tierra anécicade gran tamaño (Martiodriluscarimaguensis Jiménez y Moreno) enel PCC de sus heces, en una sabananativa y en dos pastizalesintroducidos, en un Oxisol deColombia. Los efectos que tendría laactividad de las lombrices en elcontenido de Al, Ca y Mg del suelotambién se determinaron.
Materiales y Métodos
Sitio del estudioEl estudio se llevó a cabo en laestación experimental Carimagua delconvenio CIAT-CORPOICA (4o37’ N,71o19’ O, 175 msnm), situada en losLlanos Orientales de Colombia. Elvalor anual de la precipitación estáentre 2100 y 2300 mm y el de latemperatura alrededor de 26 °C. Elsuelo es un Oxisol franco-arcillosobien drenado (fino, mezclado,Haplustox típico isohipertérmico),con un pH (H2O) de 4.5, unasaturación de Al > 80%, y con bajasconcentraciones de los elementosCa+2, Mg+2 y K+, y de P (Rao et al.1993).
Los muestreos se realizaron entres parcelas experimentales querepresentaban tres sistemasdiferentes de uso de la tierra: (1) lasabana nativa (Sav), explotada segúnel manejo tradicional de quemaanual durante la época seca; (2) unpastizal Brachiaria decumbens cv.Basilisk (Bdec) que tenía 17 años;(3) un pastizal de B. decumbensasociado con la leguminosa herbáceaforrajera Pueraria phaseoloides CIAT
9900 (“kudzú”) (Bdec/Pp), tambiénde 17 años. Los pastizalesintroducidos se establecieron enparcelas de sabana y se fertilizaroncon los siguientes elementos (enkg/ha): 44 P, 40 K, 14 Mg y 22 S, almomento de establecer el pastizal; seagregaron (en kg/ha) 10 P, 9 K,92.5 Mg y 11 S cada 2 años durantelos siguientes 9 años (Lascano yEstrada 1989). La carga animal deestos pastizales fue de 1 animal/hadurante la época seca y de2 animales/ha en la época lluviosa.
Muestreo del suelo y de lashecesDespués de la introducción de lospastizales, las parcelas de la sabanade Carimagua mantuvieron labiodiversidad original de laslombrices de tierra, aunque labiomasa de éstas aumentó más de10 veces (se midieron 4.8 g/m2 depeso fresco, en la sabana nativa)(Decaëns et al. 1994). Se eligió unaespecie anécica de individuosgrandes, Martiodrilus carimaguensis,porque domina en los pastizalesintroducidos, en los que sueleaumentar la población y la biomasade otras especies de lombrices.Estas lombrices anécicas producengrandes cantidades de heces(algunas de 15 cm de altura) sobre lasuperficie del suelo, las cuales sedistinguen fácilmente de otrasdeposiciones (Jiménez et al. 1998).
Las heces frescas de M.carimaguensis se señalaron así en elcampo: en cada pastizal introducidose aislaron en tres jaulas metálicasde 1 x 1 m, dispuestas al azar, y enla sabana nativa se incluyeron en unárea de 4 x 4 m. Las heces frescasproducidas diariamente se aislaban yse mantenían en estas condicionesdurante 20 días, hasta que llegabana un equilibrio con la humedad delsuelo. Las muestras fueron luego
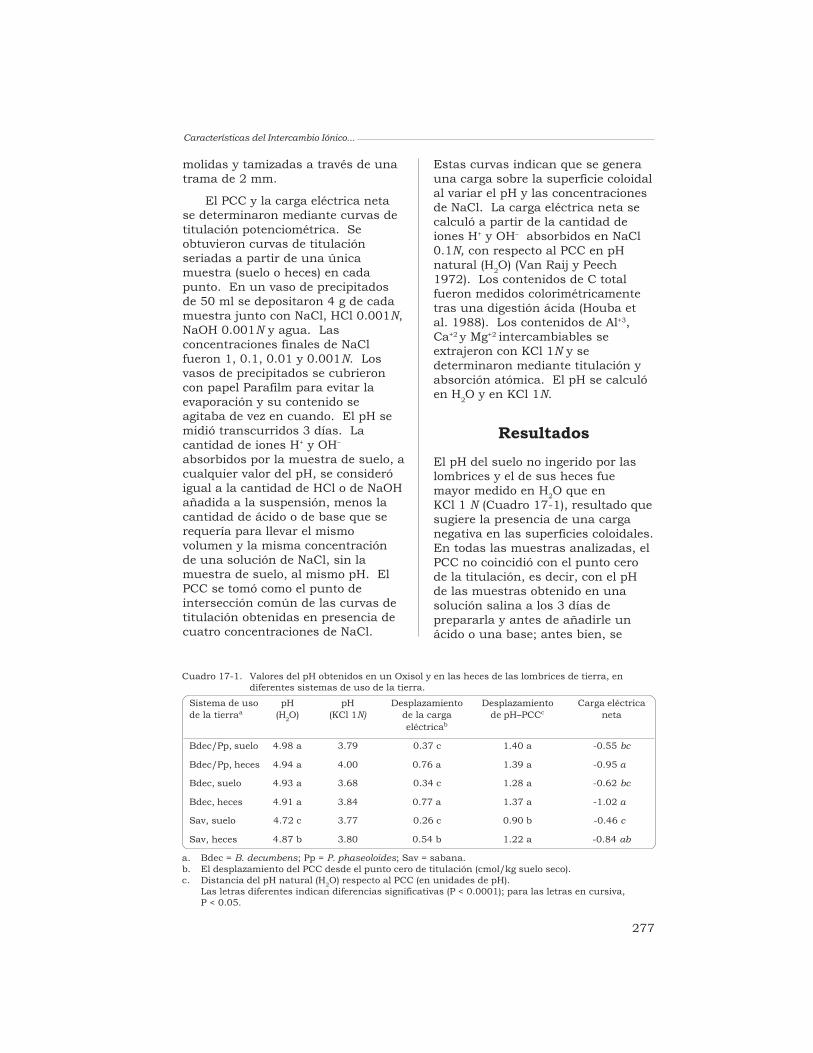
Características del Intercambio Iónico...
277
molidas y tamizadas a través de unatrama de 2 mm.
El PCC y la carga eléctrica netase determinaron mediante curvas detitulación potenciométrica. Seobtuvieron curvas de titulaciónseriadas a partir de una únicamuestra (suelo o heces) en cadapunto. En un vaso de precipitadosde 50 ml se depositaron 4 g de cadamuestra junto con NaCl, HCl 0.001N,NaOH 0.001N y agua. Lasconcentraciones finales de NaClfueron 1, 0.1, 0.01 y 0.001N. Losvasos de precipitados se cubrieroncon papel Parafilm para evitar laevaporación y su contenido seagitaba de vez en cuando. El pH semidió transcurridos 3 días. Lacantidad de iones H+ y OH–
absorbidos por la muestra de suelo, acualquier valor del pH, se consideróigual a la cantidad de HCl o de NaOHañadida a la suspensión, menos lacantidad de ácido o de base que serequería para llevar el mismovolumen y la misma concentraciónde una solución de NaCl, sin lamuestra de suelo, al mismo pH. ElPCC se tomó como el punto deintersección común de las curvas detitulación obtenidas en presencia decuatro concentraciones de NaCl.
Estas curvas indican que se generauna carga sobre la superficie coloidalal variar el pH y las concentracionesde NaCl. La carga eléctrica neta secalculó a partir de la cantidad deiones H+ y OH– absorbidos en NaCl0.1N, con respecto al PCC en pHnatural (H2O) (Van Raij y Peech1972). Los contenidos de C totalfueron medidos colorimétricamentetras una digestión ácida (Houba etal. 1988). Los contenidos de Al+3,Ca+2 y Mg+2 intercambiables seextrajeron con KCl 1N y sedeterminaron mediante titulación yabsorción atómica. El pH se calculóen H2O y en KCl 1N.
ResultadosEl pH del suelo no ingerido por laslombrices y el de sus heces fuemayor medido en H2O que enKCl 1 N (Cuadro 17-1), resultado quesugiere la presencia de una carganegativa en las superficies coloidales.En todas las muestras analizadas, elPCC no coincidió con el punto cerode la titulación, es decir, con el pHde las muestras obtenido en unasolución salina a los 3 días deprepararla y antes de añadirle unácido o una base; antes bien, se
Cuadro 17-1. Valores del pH obtenidos en un Oxisol y en las heces de las lombrices de tierra, endiferentes sistemas de uso de la tierra.
Sistema de uso pH pH Desplazamiento Desplazamiento Carga eléctricade la tierraa (H2O) (KCl 1N) de la carga de pH–PCCc neta
eléctricab
Bdec/Pp, suelo 4.98 a 3.79 0.37 c 1.40 a -0.55 bc
Bdec/Pp, heces 4.94 a 4.00 0.76 a 1.39 a -0.95 a
Bdec, suelo 4.93 a 3.68 0.34 c 1.28 a -0.62 bc
Bdec, heces 4.91 a 3.84 0.77 a 1.37 a -1.02 a
Sav, suelo 4.72 c 3.77 0.26 c 0.90 b -0.46 c
Sav, heces 4.87 b 3.80 0.54 b 1.22 a -0.84 ab
a. Bdec = B. decumbens; Pp = P. phaseoloides; Sav = sabana.b. El desplazamiento del PCC desde el punto cero de titulación (cmol/kg suelo seco).c. Distancia del pH natural (H2O) respecto al PCC (en unidades de pH).
Las letras diferentes indican diferencias significativas (P < 0.0001); para las letras en cursiva,P < 0.05.
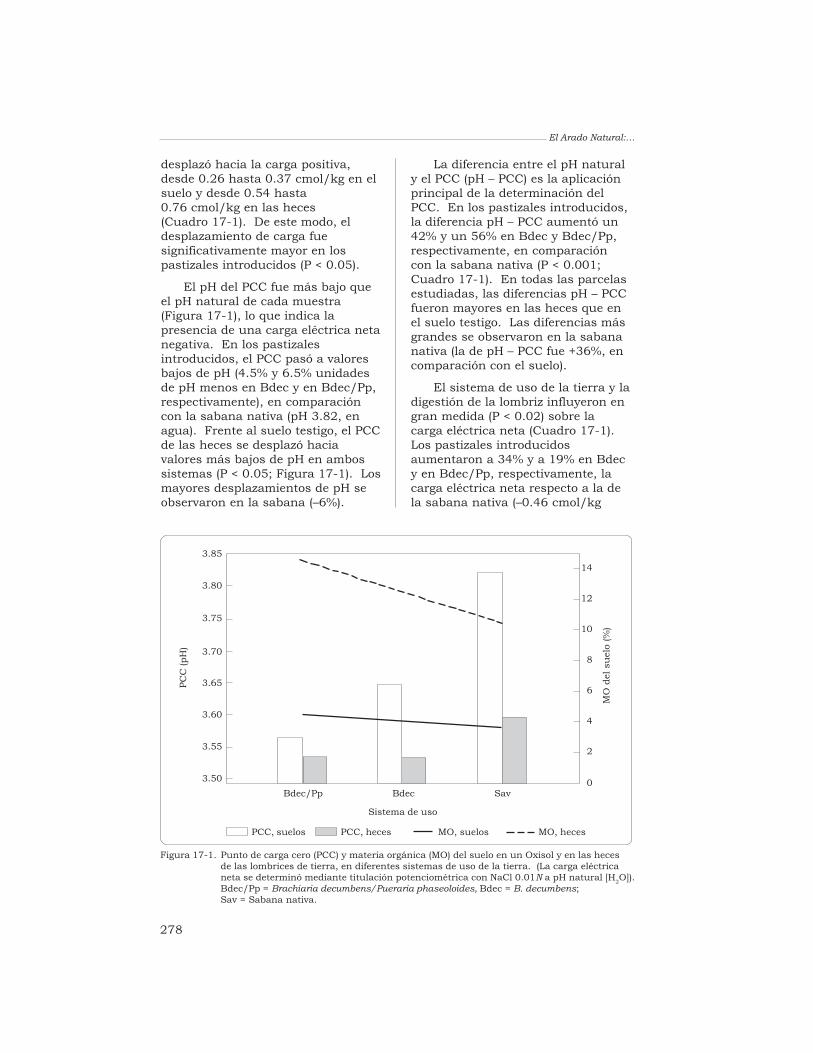
278
El Arado Natural:…
desplazó hacia la carga positiva,desde 0.26 hasta 0.37 cmol/kg en elsuelo y desde 0.54 hasta0.76 cmol/kg en las heces(Cuadro 17-1). De este modo, eldesplazamiento de carga fuesignificativamente mayor en lospastizales introducidos (P < 0.05).
El pH del PCC fue más bajo queel pH natural de cada muestra(Figura 17-1), lo que indica lapresencia de una carga eléctrica netanegativa. En los pastizalesintroducidos, el PCC pasó a valoresbajos de pH (4.5% y 6.5% unidadesde pH menos en Bdec y en Bdec/Pp,respectivamente), en comparacióncon la sabana nativa (pH 3.82, enagua). Frente al suelo testigo, el PCCde las heces se desplazó haciavalores más bajos de pH en ambossistemas (P < 0.05; Figura 17-1). Losmayores desplazamientos de pH seobservaron en la sabana (–6%).
La diferencia entre el pH naturaly el PCC (pH – PCC) es la aplicaciónprincipal de la determinación delPCC. En los pastizales introducidos,la diferencia pH – PCC aumentó un42% y un 56% en Bdec y Bdec/Pp,respectivamente, en comparacióncon la sabana nativa (P < 0.001;Cuadro 17-1). En todas las parcelasestudiadas, las diferencias pH – PCCfueron mayores en las heces que enel suelo testigo. Las diferencias másgrandes se observaron en la sabananativa (la de pH – PCC fue +36%, encomparación con el suelo).
El sistema de uso de la tierra y ladigestión de la lombriz influyeron engran medida (P < 0.02) sobre lacarga eléctrica neta (Cuadro 17-1).Los pastizales introducidosaumentaron a 34% y a 19% en Bdecy en Bdec/Pp, respectivamente, lacarga eléctrica neta respecto a la dela sabana nativa (–0.46 cmol/kg
Figura 17-1. Punto de carga cero (PCC) y materia orgánica (MO) del suelo en un Oxisol y en las hecesde las lombrices de tierra, en diferentes sistemas de uso de la tierra. (La carga eléctricaneta se determinó mediante titulación potenciométrica con NaCl 0.01N a pH natural [H2O]).Bdec/Pp = Brachiaria decumbens/Pueraria phaseoloides, Bdec = B. decumbens;Sav = Sabana nativa.
PCC
(pH
)
3.85
3.80
3.75
3.70
3.65
3.60
3.55
3.50Bdec/Pp Bdec Sav
Sistema de uso
14
12
10
8
6
4
2
0
PCC, suelos PCC, heces MO, suelos MO, heces
MO
del
su
elo
(%)
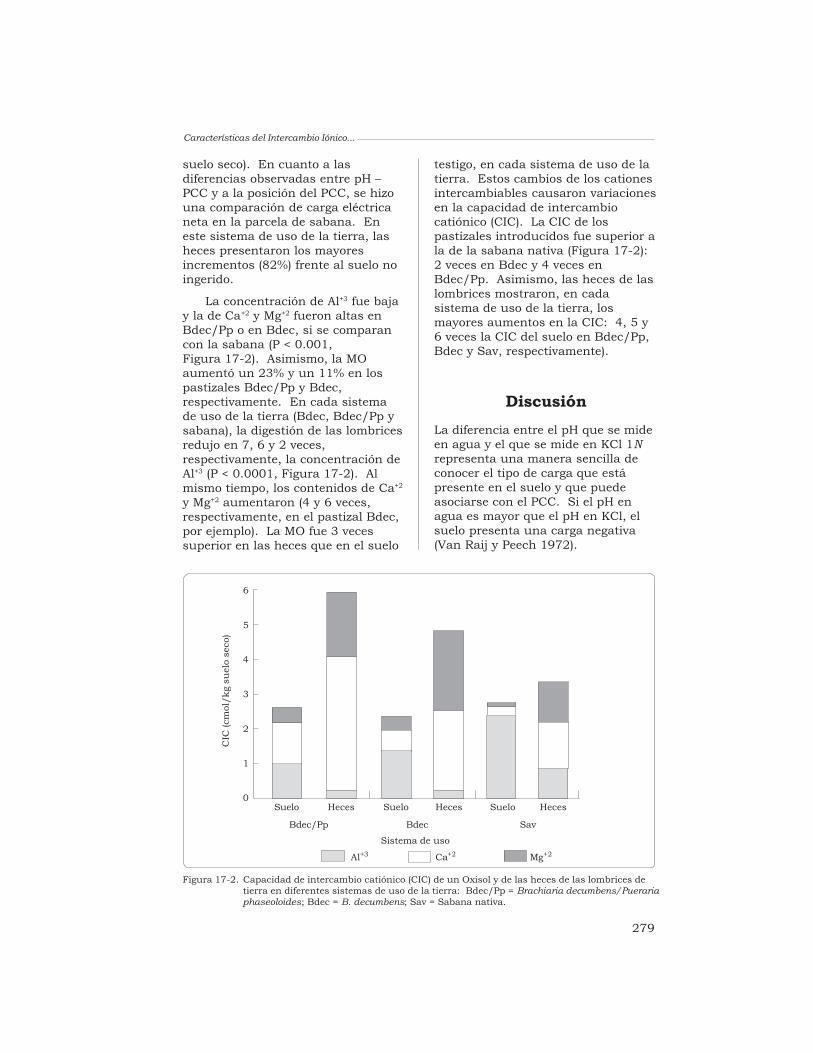
Características del Intercambio Iónico...
279
suelo seco). En cuanto a lasdiferencias observadas entre pH –PCC y a la posición del PCC, se hizouna comparación de carga eléctricaneta en la parcela de sabana. Eneste sistema de uso de la tierra, lasheces presentaron los mayoresincrementos (82%) frente al suelo noingerido.
La concentración de Al+3 fue bajay la de Ca+2 y Mg+2 fueron altas enBdec/Pp o en Bdec, si se comparancon la sabana (P < 0.001,Figura 17-2). Asimismo, la MOaumentó un 23% y un 11% en lospastizales Bdec/Pp y Bdec,respectivamente. En cada sistemade uso de la tierra (Bdec, Bdec/Pp ysabana), la digestión de las lombricesredujo en 7, 6 y 2 veces,respectivamente, la concentración deAl+3 (P < 0.0001, Figura 17-2). Almismo tiempo, los contenidos de Ca+2
y Mg+2 aumentaron (4 y 6 veces,respectivamente, en el pastizal Bdec,por ejemplo). La MO fue 3 vecessuperior en las heces que en el suelo
testigo, en cada sistema de uso de latierra. Estos cambios de los cationesintercambiables causaron variacionesen la capacidad de intercambiocatiónico (CIC). La CIC de lospastizales introducidos fue superior ala de la sabana nativa (Figura 17-2):2 veces en Bdec y 4 veces enBdec/Pp. Asimismo, las heces de laslombrices mostraron, en cadasistema de uso de la tierra, losmayores aumentos en la CIC: 4, 5 y6 veces la CIC del suelo en Bdec/Pp,Bdec y Sav, respectivamente).
DiscusiónLa diferencia entre el pH que se mideen agua y el que se mide en KCl 1Nrepresenta una manera sencilla deconocer el tipo de carga que estápresente en el suelo y que puedeasociarse con el PCC. Si el pH enagua es mayor que el pH en KCl, elsuelo presenta una carga negativa(Van Raij y Peech 1972).
Figura 17-2. Capacidad de intercambio catiónico (CIC) de un Oxisol y de las heces de las lombrices detierra en diferentes sistemas de uso de la tierra: Bdec/Pp = Brachiaria decumbens/Puerariaphaseoloides; Bdec = B. decumbens; Sav = Sabana nativa.
Suelo Heces Suelo Heces Suelo Heces
6
5
4
3
2
1
0
Al+3 Ca+2 Mg+2
Bdec/Pp Bdec Sav
Sistema de uso
CIC
(cm
ol/k
g su
elo
seco
)

280
El Arado Natural:…
Los iones del suelo (cationes yaniones) se llaman intercambiablescuando pueden ser retirados odesplazados por la lixiviación debidaa otro tipo de solución electrolítica.Los iones pueden ser adsorbidos, condiversos grados de afinidad, hacia lasuperficie del suelo; la remoción queexperimenten después dependerá delion que se emplee para remplazarlos(Oades et al. 1989). Eldesplazamiento del PCC desde elpunto de titulación cero se debe auna adsorción específica de iones H+
y OH– en sitios de oxidación que estánocupados por cationes, iones H+ osólidos diluidos; este desplazamientoequivale a la cantidad de Al+3
intercambiable (Van Raij y Peech1972). Oades et al. (1989)encontraron que, si se identifica elPCC, cualquier adsorción excesiva decationes o aniones intercambiables, aeste pH, podría atribuirse a unacarga permanente.
Los cambios ocurridos en lafertilidad del suelo (cantidadesbastante altas de Ca+2, Mg+2 y MO,reducción del Al+3 intercambiable) y elcomportamiento general de la carga(valores bajos de PCC, diferenciasentre el pH natural y el PCC, CIC ycarga eléctrica neta relativamentealtas), que provienen de laintroducción y del manejo depastizales de especies forrajerasmejoradas en suelos cubiertos hastaentonces por la sabana nativa, sontambién una consecuencia dediversos factores. Entre ellos están lafertilización y el encalado durante15 años, desde el establecimiento delpastizal; el empleo de especiesforrajeras que producen grandescantidades de biomasa en el interiordel suelo; la contribución de una MOde alta calidad proveniente de laleguminosa forrajera; y laestabilización de los procesos físicos yquímicos por la actividad de la faunadel suelo, especialmente de las
lombrices de tierra (Gijsman y Thomas1995; Rao et al. 1993).
Guggenberger et al. (1995)encontraron que, en los suelos de lasabana de Carimagua, la introducciónde B. decumbens sola o asociada conP. phaseoloides incrementó el C totaldel suelo en un 18%, especialmente enla fracción arcillosa. Otra explicación,dada por los mismos autores, es lasiguiente: los cambios en las prácticasagrícolas con que se manejan lossuelos fuertemente meteorizadospueden influir en la cantidad de MOque hay en el suelo, en su tasa dereciclaje, y en su distribución entre laparte lábil y la parte estable de losdiferentes agregados del suelo. Estosefectos podrían explicar el virajeobservado del PCC hacia valores de pHrelativamente bajos.
La importancia que tienen losefectos causados por las lombrices detierra en la fertilidad del suelo y en sucomportamiento electroquímicodependen de las escalas de espacio yde tiempo que se consideren (Lavelle yMartin 1992). Si las escalas de tiempoy de espacio son estrechas, laactividad de las poblacionesbacterianas se estimula en el tractodigestivo de las lombrices, lo queaumenta la mineralización de la MO yconduce a la acumulación denutrientes asimilables en las hecesfrescas. En una escala espacio-temporal más extensa, las heces secasprotegen la MO de una mineralizaciónulterior, estableciendo así una reservade MO que persiste en el suelo(Decaëns et al. 1999). El impactoglobal de M. carimaguensis en ladinámica de la MO puede serconsiderable dada su capacidad deingerir grandes cantidades de suelo yMO fresca (hasta 378 t/ha al año enBdec/Pp).
Las diferencias que presentan lasheces observadas en los sistemas deuso de la tierra estudiados reflejan,

Características del Intercambio Iónico...
281
probablemente, la calidad de lahojarasca y de la MO del sueloingeridas por las lombrices. Elanálisis de la hojarasca indica que,en los pastizales Bdec y Bdec/Pp,ésta presenta mayor cantidad de Ca+2
(2 y 4 veces más, respectivamente)que en la parcela de sabana nativa(Ca+2 = 0.16 cmol/kg). Asimismo, lacantidad de Mg+2 es mayor en lahojarasca de los pastizales que en lade la sabana nativa (Thomas yAsakawa 1993).
En todos los sistemas de uso dela tierra estudiados, se observarongrandes cambios en el PCC del suelodespués del tránsito de éste por elintestino de las lombrices; estoocurrió especialmente en la parcelade sabana, donde tanto la MO delsuelo como la calidad de la hojarascapresentaron los niveles más bajos.Los anteriores resultados señalan laimportancia de canalizar la actividadde las lombrices de tierra paraestabilizar la fertilidad del suelo y elcomportamiento electroquímico deéste en los pastizales; equivalen,además, esos resultados a unmétodo para mantener la calidad delsuelo. Oades et al. (1989)demostraron que, si se mantiene elcontenido de la MO en su nivel másalto posible para alcanzar valoresmínimos del PCC, se haría un granaporte a la capacidad que poseen lossuelos de carga variable de retenerlos cationes que nutren las plantas yde incrementar la CIC. Las grandesdiferencias observadas entre el pHnatural y el PCC podrían tambiénreducir el riesgo de lixiviación decationes y mejorar, de este modo, lasostenibilidad de los suelos.
ConclusionesLa introducción de especiesforrajeras mejoradas en pastizalesestablecidos en suelos que han
pertenecido anteriormente a lasabana nativa, desencadena cambiosimportantes en la fertilidad y en elcomportamiento electroquímico deesos suelos. Estos cambios en loscationes intercambiables y en laspropiedades electroquímicas semejoran notablemente cuando elsuelo ha sido sometido a la digestiónde las lombrices (M. carimaguensis),ya que en las heces de éstas hay unincremento de la MO del suelo. Deigual modo, la magnitud de talescambios depende de la calidad de laMO del suelo consumida por laslombrices (por ejemplo, en lospastizales asociados con leguminosasherbáceas). El descenso del PCC enlas heces se debe al aumento de laMO del suelo y al desplazamiento delAl+3 intercambiable. La deposición delas heces ocasiona cambios en laspropiedades del suelo, que puedenllevar a una mejora considerable dela capacidad de intercambio de ionesen un Oxisol infértil. Estos efectospueden estar relacionados con unamejoría en la retención y el reciclajede nutrientes en un sistema demanejo de la tierra en que seapliquen pocos insumos.
AgradecimientosEl antiguo Programa de Trópico Bajodel CIAT y el Proyecto Macrofauna(Comunidad Europea) aportaron losfondos para realizar estainvestigación. Los autores agradecenla ayuda del personal de Carimaguay de la sección de servicios analíticosdel CIAT. Agradecen también alDr. Carlos Iglesias y al Dr. EdgarAmézquita por sus comentarios. Serecibió también apoyo financiero delconsorcio para el Manejo de SuelosÁcidos (MAS) perteneciente alprograma del sistema CGIAR para elManejo de Suelos, Agua y Nutrientes(SWNM, en inglés).

282
El Arado Natural:…
ReferenciasDecaëns T; Rangel AF; Asakawa N;
Thomas RJ. 1999. Carbon andnitrogen dynamics in in situ ageingearthworm casts in natural savannaand man-made pasture of theEastern Plains of Colombia. BiolFertil Soils 30:20-28.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30(4):157-168.
Gijsman AJ; Thomas RJ. 1995. Aggregatesize distribution and analysis of anOxisol under legume-based andpure grass pasture in the EasternPlains of Colombia. Aust J Soil Res33:153-165.
Guggenberger G; Zech W; Thomas RJ.1995. Lignin and carbohydratealteration in particle-size separatesof an Oxisol under tropical pasturesfollowing native savanna. Soil BiolBiochem 27:1629-1638.
Houba VJC; Lee van der JJ; Novozamky I;Walinga J. 1988. Soil and plantanalysis. En: Department of SoilScience and Plant Nutrition, ed. Soilanalysis procedures. 4a. ed.Agricultural University,Wageningen, Holanda. p 4-14.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the EasternPlains of Colombia. Appl Soil Ecol9:153-160.
Lascano CE; Estrada J. 1989. Long-termproductivity of legume-based andpure grass pastures in the EasternPlains of Colombia. En: Memoriasdel XVI International GrasslandCongress, Niza, Francia.p 1179-1180.
Lavelle P. 1978. Les vers de terre de lasavane de Lamto (Côte d’Ivoire):peuplements, populations etfontions dans l’écosystème. Tesis(Doctorado). Paris VI. Publ Lab ZoolENS, 12. 301 p.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in a soil of the humidtropics. Soil Biol Biochem24:1491-1498.
Oades JM; Gillman GP; Uehara G. 1989.Interactions of soil organic matterand variable-charge clays. En:Coleman DC; Oades JM; Uehara G,eds. Dynamics of soil organicmatter in tropical ecosystems. NiftalProject, University of Hawaii,Honolulu, Estados Unidos. p 69-86.
Rao IM; Borrero V; Ayarza MA; García R.1993. Adaptation of tropical foragespecies to acid soils: the influenceof varying phosphorus supply andsoil type on phosphorus uptake anduse. En: Barrow NJ, ed. Plantnutrition: From genetic engineeringto field practice. Kluwer AcademicPublishers, Dordrecht, Holanda.p 345-348.
Sposito G; Schindler P W. 1986. Reactionat the soil colloid-soil solutioninterface. Transactions of the 13th
International Congress of SoilScience (Hamburg) 6:683-699.
Thomas RJ; Asakawa NM. 1993.Decomposition of leaf litter fromtropical forage grasses andlegumes. Soil Biol Biochem25:1351-1361.
Van Raij B; Peech M. 1972.Electrochemical properties of someOxisols and Alfisols of the tropics.Soil Sci Soc Am J 36:587-593.

Las Comunidades de la Macrofauna de la Superficie...
283
PARTE V
Efectos Producidos por los Ingenierosdel Ecosistema en la Biodiversidad
del Suelo, a Escala de lasEstructuras Biogénicas

Las Comunidades de la Macrofauna de la Superficie...
285
ResumenLas lombrices de tierra modifican,mediante sus actividadesdrilosféricas, las condiciones de vidade otros organismos. Seinvestigaron los efectos que causauna especie anécica de gran tamaño,Martiodrilus carimaguensis Jiménez yMoreno, en las comunidades demacrofauna del suelo de una sabananatural y de un pastizal introducidoen los Llanos Orientales deColombia. Se tomaron muestras delos invertebrados empleando unmétodo manual estándar, adiferentes escalas espaciales: (1) a laescala de unos pocos centímetros, secompararon durante un año ladinámica de la densidad, la biomasa,la diversidad y los pesos individualesde los macroinvertebradosencontrados en las hecesdepositadas en la superficie del suelopor la especie de lombrizmencionada, en dos lugares: en elsuelo situado debajo de las heces yen un suelo testigo libre de heces
superficiales; (2) a la escala de unospocos decímetros y en un tiempodeterminado, se tomaron muestrasde los macroinvertebrados enmonolitos de suelo cuya superficiepresentaba diferente densidad deheces. Los macroinvertebradoscolonizaron las heces 4 y 6 semanasdespués de su formación, en elpastizal y en la sabana,respectivamente. En estos dossistemas de uso de la tierra y a laescala espacial del tamaño de ladeposición, la densidad demacroinvertebrados fuesignificativamente mayor debajode las heces (de 1660 a5310 individuos/m2) que en el suelotestigo (de 400 a 1700 individuos/m2); no se hallaron aquí diferenciassignificativas en cuanto a labiomasa, la riqueza de especies, ladiversidad y la equitatividad. Lapresencia de las heces superficialesincrementó la dominancia relativa delas poblaciones epígeas (+185% a+250%) en detrimento de las especiesendógeas grandes (–39% a –76%). Elpeso individual de los invertebradosfue inferior en las heces (de 1.2 a5.7 mg/individuo) y en el suelodebajo de ellas (de 8.2 a 11.1 mg/individuo) que en el suelo testigo(25.0 a 39.6 mg/individuo). Estosresultados fueron confirmados a unaescala mayor. A la escala de algunosdecímetros, la riqueza taxonómica se
CAPÍTULO 18
Las Comunidades de la Macrofauna dela Superficie del Suelo Asociadas conlas Heces de las Lombrices de Tierraen los Llanos Orientales de Colombia1
T. Decaëns, L. Mariani y P. Lavelle*
1. Publicado en Applied Soil Ecology 13,p 87-100, por Decaëns et al. Copyright ©1999 de Elsevier. Impreso con permiso de lacasa editorial.
* Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia/Unidad de Suelos yNutrición Plantas, CIAT, Cali, Colombia.

286
El Arado Natural:…
correlacionó positivamente con elnúmero de heces en la superficie delsuelo, mientras que la diversidad nocambió y la equitatividad disminuyó.Estos resultados se atribuyen,principalmente, a la formación denuevos hábitat (macroporos, galerías)por las lombrices y a laconcentración de hojarasca y demateria orgánica (MO) en las heces.Apoyan, además, la “hipótesis de lasbiodiversidades encajadas”, según lacual los ingenieros del suelo puedencausar efectos determinantes en laabundancia y en la diversidad deotros organismos del suelo.
Palabras clave: biodiversidad delsuelo, ingenieros del ecosistema,macrofauna del suelo,Martiodrilus carimaguensis,pastizal de gramínea yleguminosa, sabana tropical
IntroducciónDe los numerosos organismos quehabitan en los suelos, unos pocosinvertebrados grandes (lombrices,termitas y hormigas, principalmente)causan un impacto considerable enmuchos procesos que determinan lafertilidad del suelo. Estos gruposfuncionales, definidos como“ingenieros del ecosistema” (sensuJones et al. 1994), elaboran unagran variedad de macroporos (porejemplo, galerías o cámaras) y deestructuras órgano-minerales (porejemplo, las heces de las lombrices,los termiteros y los hormigueros) queinfluyen en las propiedadeshidráulicas, en la macroagregación, yen la dinámica de la MO del suelo(Anderson 1995; Lavelle 1996, 1997).
Los ingenieros del ecosistema,mediante sus actividadesalimentarias y mecánicas, modificanlas condiciones de vida de otrosorganismos más pequeños y menosmóviles e influyen, por tanto, en laabundancia y en la diversidad de
éstos (hipótesis de lasbiodiversidades encajadas, Lavelle1996). Los efectos que causan laslombrices en la actividad de losmicroorganismos del suelo han sidoinvestigados en muchas partes(Barois y Lavelle 1986; Daniel yAnderson 1992; Scheu 1987, 1993),y se han conocido como la “paradojade la bella durmiente” (Lavelle 1996).Sin embargo, la información sobre elimpacto de las lombrices en ladiversidad y en la estructura de lascomunidades de microorganismos esescasa (Parkinson y McLean 1998).Algunos estudios han consideradolos efectos positivos o negativos quelas lombrices de tierra pueden teneren las comunidades demicroinvertebrados y demesoinvertebrados (ver revisión deBrown 1995; también Loranger et al.1998; Marinissen y Bok 1988), perosu impacto en las comunidades de lamacrofauna se ha investigado poco.Sólo unos pocos estudios hanhallado respuestas positivas de partede algunos grupos de macrofauna ala actividad de las lombrices de tierra(Kirk 1981; Szlavecz 1985;Thompson et al. 1993), aunquetodavía queda mucho por investigar.
Es fundamental comprendertanto el papel de la biodiversidad enlos suelos como las condicionesnecesarias para su mantenimiento,porque las diferentes especies deinvertebrados causarán un impactodiferente y, por ende, tendrándiferentes funciones en el suelo(Lavelle 1996). Con este propósito,es esencial lograr dos objetivos:(1) identificar claramente lasrelaciones existentes entre lasespecies; (2) averiguar hasta quépunto la presencia de una especiedada influye en la presencia de otras.
El objetivo de este estudio fueevaluar los efectos que ejercen lasestructuras creadas por losindividuos de Martiodrilus

Las Comunidades de la Macrofauna de la Superficie...
287
carimaguensis Jiménez y Moreno(Oligochaeta: Glossoscolecidae), unaespecie grande anécica (sensuBouché 1977) de lombriz de tierra delos Llanos Orientales de Colombia,en las comunidades de lamacrofauna del suelo. Esta especiees la única del sitio de estudio quedeposita una gran cantidad de heceso turrículos sobre la superficie delsuelo (Jiménez et al. 1998b). Losexperimentos se realizaron adiferentes escalas de tiempo y deespacio, en una parcela de la sabananatural y en un pastizal introducidopor la acción humana en un área dela sabana.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua delconvenio CIAT-CORPOICA (4°37’ N,71°19’ O), localizada en la unidadfitogeográfica de las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia.El clima es subhúmedo tropical,donde el valor medio anual de latemperatura y de la precipitación esde 26 °C y de 2300 mm,respectivamente; hay una época secaque transcurre de noviembre amarzo. La vegetación nativa estádeterminada por la topografía: haysabanas abiertas en las zonas altas(“altos” y “planos”) y selvas de galeríao sabanas inundables en las zonasbajas (“bajos”). Los suelos sonOxisoles (Tropeptic HaplustoxIsohyperthermic) en los altos yUltisoles (Ultic Aeric Plintaquox) enlos bajos. Los dos tipos de suelosson muy agregados y se caracterizanpor su baja fertilidad química (pH[H2O] < 5, saturación de Al > 80%,CIC < 5 meq/100 g).
Parcelas experimentalesLa macrofauna del suelo se muestreóen dos sistemas diferentes de uso dela tierra que pertenecían a un Oxisolbien drenado ubicado en una zonaalta:
• Una parcela de sabana nativacubierta por Trachypogon vestitusAnders., protegida del pastoreodurante 4 años, y manejadatradicionalmente con la quemaanual durante la época seca.
• Un pastizal de 3 años sembradocon Brachiaria humidicola(Rendle), Arachis pintoi Krap. yGreg., Stylosanthes capitata Vog.y Centrosema acutifolium Benth.,sometido a pastoreo a razón de2.0 animales/ha.
Diseño experimentalLos ingenieros del ecosistemainfluyen en algunos procesos delsuelo a escalas espaciales ytemporales más grandes porqueproducen estructuras físicas (porejemplo, las heces de las lombrices,los termiteros y las galerías)(Anderson 1995; Lavelle 1996, 1997).Por tanto, sus efectos puedendefinirse como una regulación, apequeña escala, de procesos queocurren a una escala mayor(Anderson 1995). Por tal razón, serealizaron dos experimentos con elfin de describir los efectos que teníael envejecimiento de las heces de losindividuos de M. carimaguensis enlas comunidades de invertebrados, ados escalas:
- a la escala de la estructuraque construyen (unospocos cm);
- a la escala de la parcelaexperimental.

288
El Arado Natural:…
suelo se revisaron manualmente ycon cuidado, en bandejas plásticasde 40 x 60 cm; losmacroinvertebrados que seencontraron allí se conservaron enalcohol al 70%.
Con el fin de describir conprecisión la estructura de lacomunidad de invertebrados, éstosse identificaron hasta Orden oFamilia; después se separaron segúnel morfotipo (sensu Oliver y Beattie1995), es decir, en taxonesmorfológicamente distintos(Cuadro 18-1). Se calculó luego ladensidad (individuos/m2) y labiomasa (g/m2 de materia fresca) de11 grandes unidades taxonómicas yfuncionales (Cuadro 18-2). Se siguióla clasificación funcional empleadanormalmente para las lombrices detierra (Bouché 1977), porque incluyelas características funcionales de lamayor parte de la macrofauna delsuelo:
- especies endógeas, que vivenexclusivamente dentro delsuelo y son, en su mayoría,lombrices y larvas rizófagasde los coleópteros;
- especies anécicas, que vivenen el suelo y se alimentan enla superficie, y son lombrices,termitas y hormigas;
- especies epígeas, que vivenentre la hojarasca, y sonlombrices, termitas, hormigasy artrópodos de la hojarasca.
La biomasa se corrigió respecto alas pérdidas ocasionadas por laconservación en alcohol, así: un19% para las lombrices y lastermitas, un 9% para las hormigas,un 11% para los coleópteros, un 6%para arácnidos y miriápodos, y un13% para otros grupos de artrópodos(Decaëns et al. 1994).
Estudio a la escala de ladeposiciónEl experimento se inició al
comienzo de la época lluviosa (mayode 1996), durante el punto máximode actividad de las lombrices. Enambos sistemas se marcaron160 heces de individuos deM. carimaguensis con pequeñasláminas metálicas, y se dividieron en8 grupos de 20 heces contiguas cadauno. Durante esta operación seprestó atención especial a las hecesdepositadas recientemente (es decir,a las heces pequeñas y frescas). Deeste modo se aseguró la presencia deuna lombriz en cada una de lasgalerías marcadas y se evitó el efectoacumulativo, que resultaría de lapresencia de una deposición muygrande sobre la superficie del sueloantes del inicio del experimento. Laslombrices no fueron eliminadas delas galerías, y permanecieron en ellasañadiendo material sobre ladeposición inicial durante losprimeros días del experimento.Pasados varios días, las heces oturrículos pueden alcanzar 15 cm dediámetro y 15 cm de altura. Lasheces se secaban en la superficie delsuelo cuando las lombricesabandonaban sus galerías.
La macrofauna del suelo (esdecir, los invertebrados mayores de2 mm) se muestreó en las siguientesfechas: 0, 7 y 14 días, y 1, 2, 3.5, 5,6.5, 8, 9.5 y 11 meses después delinicio de la producción de las heces.En cada fecha de muestreo seescogió al azar un turrículo en cadauno de los 8 grupos de 20 heces. Lamuestra constaba de los elementossiguientes: (a) el turrículo, (b) elsuelo situado bajo el turrículo; y(c) el suelo testigo, localizado a 20 cmdel turrículo. Las muestras de suelose tomaron con cilindros de aluminiode 10 cm de diámetro y 10 cm dealtura. Tanto las heces como el
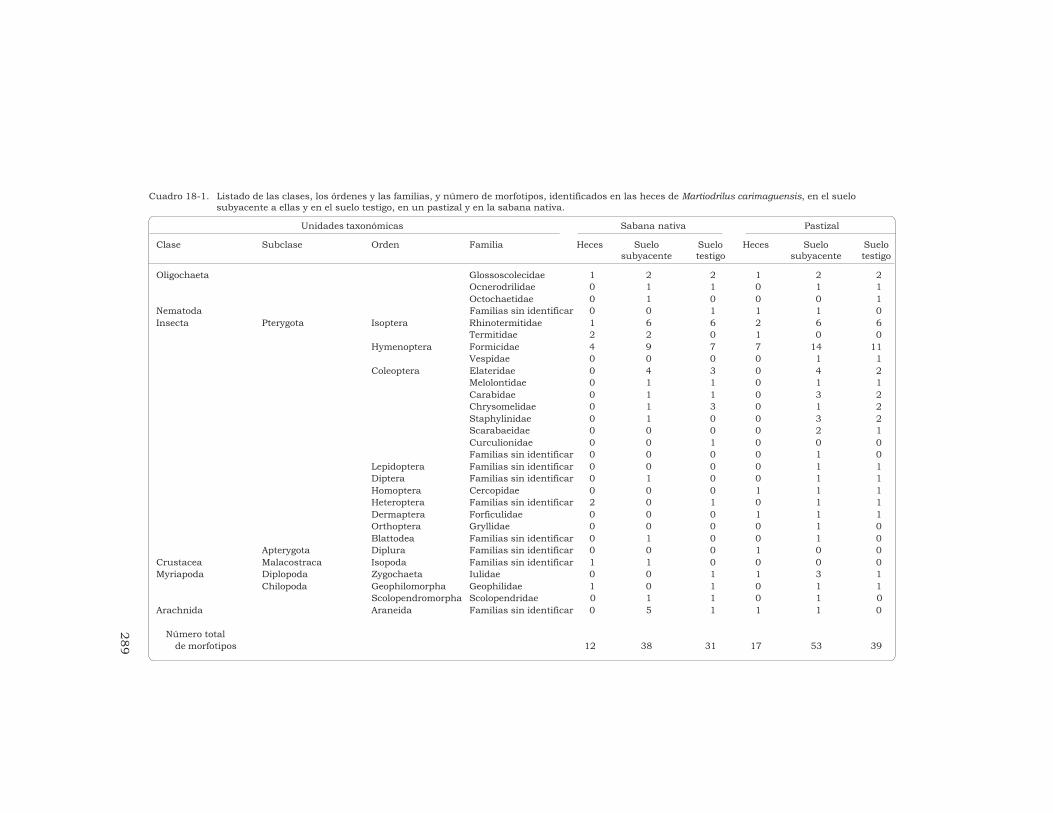
Las Com
unidades de la Macrofauna de la Superficie...
289
Cuadro 18-1. Listado de las clases, los órdenes y las familias, y número de morfotipos, identificados en las heces de Martiodrilus carimaguensis, en el suelosubyacente a ellas y en el suelo testigo, en un pastizal y en la sabana nativa.
Unidades taxonómicas Sabana nativa Pastizal
Clase Subclase Orden Familia Heces Suelo Suelo Heces Suelo Suelosubyacente testigo subyacente testigo
Oligochaeta Glossoscolecidae 1 2 2 1 2 2Ocnerodrilidae 0 1 1 0 1 1Octochaetidae 0 1 0 0 0 1
Nematoda Familias sin identificar 0 0 1 1 1 0Insecta Pterygota Isoptera Rhinotermitidae 1 6 6 2 6 6
Termitidae 2 2 0 1 0 0Hymenoptera Formicidae 4 9 7 7 14 11
Vespidae 0 0 0 0 1 1Coleoptera Elateridae 0 4 3 0 4 2
Melolontidae 0 1 1 0 1 1Carabidae 0 1 1 0 3 2Chrysomelidae 0 1 3 0 1 2Staphylinidae 0 1 0 0 3 2Scarabaeidae 0 0 0 0 2 1Curculionidae 0 0 1 0 0 0Familias sin identificar 0 0 0 0 1 0
Lepidoptera Familias sin identificar 0 0 0 0 1 1Diptera Familias sin identificar 0 1 0 0 1 1Homoptera Cercopidae 0 0 0 1 1 1Heteroptera Familias sin identificar 2 0 1 0 1 1Dermaptera Forficulidae 0 0 0 1 1 1Orthoptera Gryllidae 0 0 0 0 1 0Blattodea Familias sin identificar 0 1 0 0 1 0
Apterygota Diplura Familias sin identificar 0 0 0 1 0 0Crustacea Malacostraca Isopoda Familias sin identificar 1 1 0 0 0 0Myriapoda Diplopoda Zygochaeta Iulidae 0 0 1 1 3 1
Chilopoda Geophilomorpha Geophilidae 1 0 1 0 1 1Scolopendromorpha Scolopendridae 0 1 1 0 1 0
Arachnida Araneida Familias sin identificar 0 5 1 1 1 0
Número totalde morfotipos 12 38 31 17 53 39
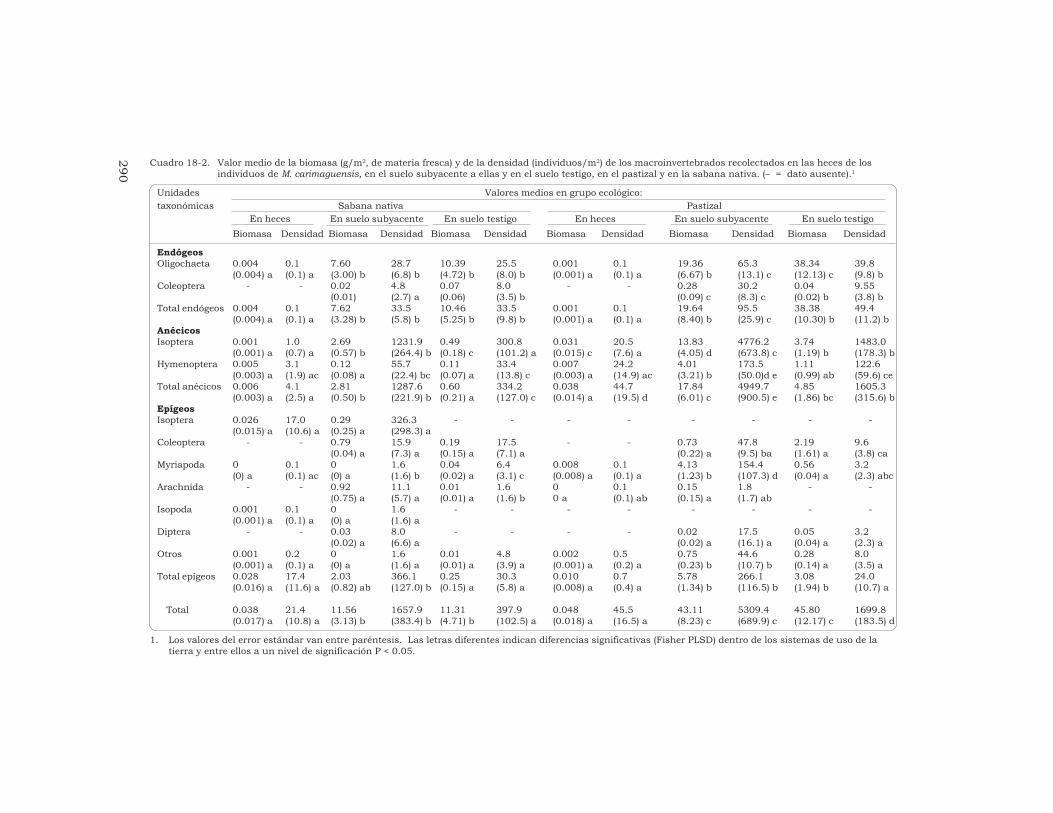
290
El A
rado Natural:…
Cuadro 18-2. Valor medio de la biomasa (g/m2, de materia fresca) y de la densidad (individuos/m2) de los macroinvertebrados recolectados en las heces de losindividuos de M. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en el pastizal y en la sabana nativa. (– = dato ausente).1
Unidades Valores medios en grupo ecológico:taxonómicas Sabana nativa Pastizal
En heces En suelo subyacente En suelo testigo En heces En suelo subyacente En suelo testigoBiomasa Densidad Biomasa Densidad Biomasa Densidad Biomasa Densidad Biomasa Densidad Biomasa Densidad
EndógeosOligochaeta 0.004 0.1 7.60 28.7 10.39 25.5 0.001 0.1 19.36 65.3 38.34 39.8
(0.004) a (0.1) a (3.00) b (6.8) b (4.72) b (8.0) b (0.001) a (0.1) a (6.67) b (13.1) c (12.13) c (9.8) bColeoptera - - 0.02 4.8 0.07 8.0 - - 0.28 30.2 0.04 9.55
(0.01) (2.7) a (0.06) (3.5) b (0.09) c (8.3) c (0.02) b (3.8) bTotal endógeos 0.004 0.1 7.62 33.5 10.46 33.5 0.001 0.1 19.64 95.5 38.38 49.4
(0.004) a (0.1) a (3.28) b (5.8) b (5.25) b (9.8) b (0.001) a (0.1) a (8.40) b (25.9) c (10.30) b (11.2) bAnécicosIsoptera 0.001 1.0 2.69 1231.9 0.49 300.8 0.031 20.5 13.83 4776.2 3.74 1483.0
(0.001) a (0.7) a (0.57) b (264.4) b (0.18) c (101.2) a (0.015) c (7.6) a (4.05) d (673.8) c (1.19) b (178.3) bHymenoptera 0.005 3.1 0.12 55.7 0.11 33.4 0.007 24.2 4.01 173.5 1.11 122.6
(0.003) a (1.9) ac (0.08) a (22.4) bc (0.07) a (13.8) c (0.003) a (14.9) ac (3.21) b (50.0)d e (0.99) ab (59.6) ceTotal anécicos 0.006 4.1 2.81 1287.6 0.60 334.2 0.038 44.7 17.84 4949.7 4.85 1605.3
(0.003) a (2.5) a (0.50) b (221.9) b (0.21) a (127.0) c (0.014) a (19.5) d (6.01) c (900.5) e (1.86) bc (315.6) bEpígeosIsoptera 0.026 17.0 0.29 326.3 - - - - - - - -
(0.015) a (10.6) a (0.25) a (298.3) aColeoptera - - 0.79 15.9 0.19 17.5 - - 0.73 47.8 2.19 9.6
(0.04) a (7.3) a (0.15) a (7.1) a (0.22) a (9.5) ba (1.61) a (3.8) caMyriapoda 0 0.1 0 1.6 0.04 6.4 0.008 0.1 4.13 154.4 0.56 3.2
(0) a (0.1) ac (0) a (1.6) b (0.02) a (3.1) c (0.008) a (0.1) a (1.23) b (107.3) d (0.04) a (2.3) abcArachnida - - 0.92 11.1 0.01 1.6 0 0.1 0.15 1.8 - -
(0.75) a (5.7) a (0.01) a (1.6) b 0 a (0.1) ab (0.15) a (1.7) abIsopoda 0.001 0.1 0 1.6 - - - - - - - -
(0.001) a (0.1) a (0) a (1.6) aDiptera - - 0.03 8.0 - - - - 0.02 17.5 0.05 3.2
(0.02) a (6.6) a (0.02) a (16.1) a (0.04) a (2.3) aOtros 0.001 0.2 0 1.6 0.01 4.8 0.002 0.5 0.75 44.6 0.28 8.0
(0.001) a (0.1) a (0) a (1.6) a (0.01) a (3.9) a (0.001) a (0.2) a (0.23) b (10.7) b (0.14) a (3.5) aTotal epígeos 0.028 17.4 2.03 366.1 0.25 30.3 0.010 0.7 5.78 266.1 3.08 24.0
(0.016) a (11.6) a (0.82) ab (127.0) b (0.15) a (5.8) a (0.008) a (0.4) a (1.34) b (116.5) b (1.94) b (10.7) a
Total 0.038 21.4 11.56 1657.9 11.31 397.9 0.048 45.5 43.11 5309.4 45.80 1699.8(0.017) a (10.8) a (3.13) b (383.4) b (4.71) b (102.5) a (0.018) a (16.5) a (8.23) c (689.9) c (12.17) c (183.5) d
1. Los valores del error estándar van entre paréntesis. Las letras diferentes indican diferencias significativas (Fisher PLSD) dentro de los sistemas de uso de latierra y entre ellos a un nivel de significación P < 0.05.

Las Comunidades de la Macrofauna de la Superficie...
291
Estudio a la escala de laparcelaEn los primeros meses de la
época lluviosa (junio-julio de 1997),se tomaron al azar 20 monolitos de25 x 25 x 30 cm en cada parcela deestudio. Se contaron las heces de losindividuos de M. carimaguensispresentes en la superficie de losmonolitos demarcados y, acontinuación, éstos fueron extraídosdel suelo y divididos en 4 capassucesivas (hojarasca, 0-10 cm,10-20 cm y 20-30 cm). Cada capa oestrato se revisó manualmentesiguiendo el método antes descrito(recomendado por el programa deFertilidad y Biología del SueloTropical, Anderson e Ingram 1993).
Procesamiento de los datosLa estructura de las comunidades demacroinvertebrados se describióempleando los índices disponibles enla literatura científica. Estos índicesse calcularon en cada submuestra(turrículo, suelo subyacente y suelotestigo) de cada parcelaexperimental, a partir del total deindividuos recogidos durante todo elexperimento. Las comparacionesentre estos índices requieren, paraque tengan validez, que todas lasmuestras sean del mismo tamaño(Hurlbert 1971); por consiguiente, nose compararon las muestras desuelo, cuyo tamaño podía calcularse,con las muestras de turrículos(heces) cuyo tamaño no se podíadeterminar fácilmente con precisión.
El índice de similitud deSørensen (Cs) se empleó paracomparar la composición taxonómica(de morfotipos) de las comunidadesde macrofauna en el suelosubyacente y en el suelo testigo,dentro de cada parcela experimentaly entre una y otra parcela (Legendrey Legendre 1979; Sørensen 1948):
donde j es el número de morfotiposcomunes a las dos muestrascomparadas, y los términos a y brepresentan el número total demorfotipos recolectados en unamuestra y en la otra,respectivamente.
La riqueza taxonómica (S) sedefinió como el número total demorfotipos. La diversidad (H) y laequitatividad (H’) se calcularon apartir de los índices de diversidad deShannon-Wiener (Pielou 1966):
donde pi es la frecuencia delmorfotipo i.
Análisis estadísticos
Datos cuantitativosLa normalidad de la frecuencia
de distribución de los datos se probócon la prueba de Kolmogorov-Smirnov (Lilliefors 1967). Los datosse transformaban antes del análisissi era necesario reducir la asimetríade la frecuencia de distribución. Lanormalización de los datos se obtuvomediante la transformación Box-Cox(Sokal y Rohlf 1995) que es:
La prueba de normalidad y elcálculo de los parámetros δ serealizaron empleando el programaVerNorm 3.0 del “paquete R”desarrollado por Legendre y Vaudor(1991).
Se hizo también un análisis de lavarianza (ANOVA) de tres vías, en elque se consideraron, como efectosprincipales, el sistema, el origen dela muestra (turrículo, suelosubyacente o suelo testigo) y la edad
Cs = 2 j a + b( )
∑=
−=n
1i2 ii plogpH
y = (xδ −1) δ
H' = H/log2S

292
El Arado Natural:…
del turrículo. Se analizaron labiomasa y la densidad de los tresgrupos funcionales y de lamacrofauna total. Se realizaronentonces ocho análisis y cadaanálisis consistió en siete pruebas(tres efectos principales y cuatrointeracciones). Se empleó lacorrección de Bonferroni (Cooper1968) para pruebas ‘encajadas’, conla cual se evita el error estadístico:los niveles de significación ajustadosdel 0.05, 0.01 y 0.001 fueron,respectivamente, 0.001 [= 0.05/(8x7)], 0.0002 [= 0.01/(8x7)] y0.00002 [= 0.001/(8x7)]. Mediante laprueba PLSD de Fisher, se hicieronlas comparaciones adicionales de ladensidad y la biomasa medias.
Se emplearon regresiones linealessimples para probar la relación entreel número de turrículos halladossobre la superficie del monolito y lacomunidad de macrofaunaencontrada dentro del monolito. Lanormalidad de los datos se probóaplicando la prueba de Kolmogorov-Smirnov antes descrita. Esta pruebase realizó para cada situación queaportara un número suficiente demuestras (al menos cuatro). Losdatos empleados para la ejecución deestas pruebas pertenecían a lassiguientes variables: la densidad, labiomasa, la riqueza taxonómica (S) yla diversidad (H) de la macrofauna, elporcentaje de individuos encontradosen la capa superficial (0 a 10 cm) delmonolito, y la densidad de lombrices,termitas y otros grupos demacroinvertebrados. Se hicieron loscálculos de tipo “jackknife” descritosmás adelante, para los índices dediversidad. La normalidad se probóa un nivel de significación del 5% entodas las distribuciones, excepto paralos pseudovalores de H en la sabanaque tenía cinco turrículos sobre lasuperficie de la muestra. Lasignificación de todos los coeficientes
de regresión r se comprobó porseparado en los siguientes niveles designificación: 0.05, 0.01 y 0.001.
Indices de diversidadCon el fin de mejorar el cálculo de lariqueza taxonómica (S) y de ladiversidad (H) y de establecerintervalos de confianza paracomparar las comunidades demacroinvertebrados, se aplicó unprocedimiento “jackknife”generalizado y de primer orden(Tukey 1958). No se encontraronreferencias que mencionaran el usode dichas pruebas en la equitatividad(H’) y en el índice de Sørensen’s (Cs);por tanto, fue posible comparar estosúltimos índices con menos rigor.Sólo se consideraron las diferenciasmás grandes (> 0.20) al compararestos índices.
El procedimiento “jackknife” sebasa en la computación de n valoresdel estadístico deseado junto con laexclusión sucesiva de una de las nmuestras. La media de estoscálculos se usa para reducir ladesviación del estadístico, y lavariabilidad entre dichos valorescuando se calcula el error estándar(Sokal y Rohlf 1995). Tukey (1958)llegó a la conclusión de que ladistribución de los pseudovalores (φ)obtenidos mediante el procedimiento“jackknife” es asintótica,independiente y normal. Enconsecuencia, se calculó aquí unintervalo de confianza mediante latabla de distribución de la t.
En el caso del índice dediversidad de Shannon-Wiener, losvalores calculados de H ( ) y los delerror estándar (σ ) se obtuvieronempleando las siguientes ecuaciones(Adams y McCune 1979; Heltshe yForrester 1985):
,ˆ H n = φ ii =1
n
∑ n =φ
ˆ H nˆ H n
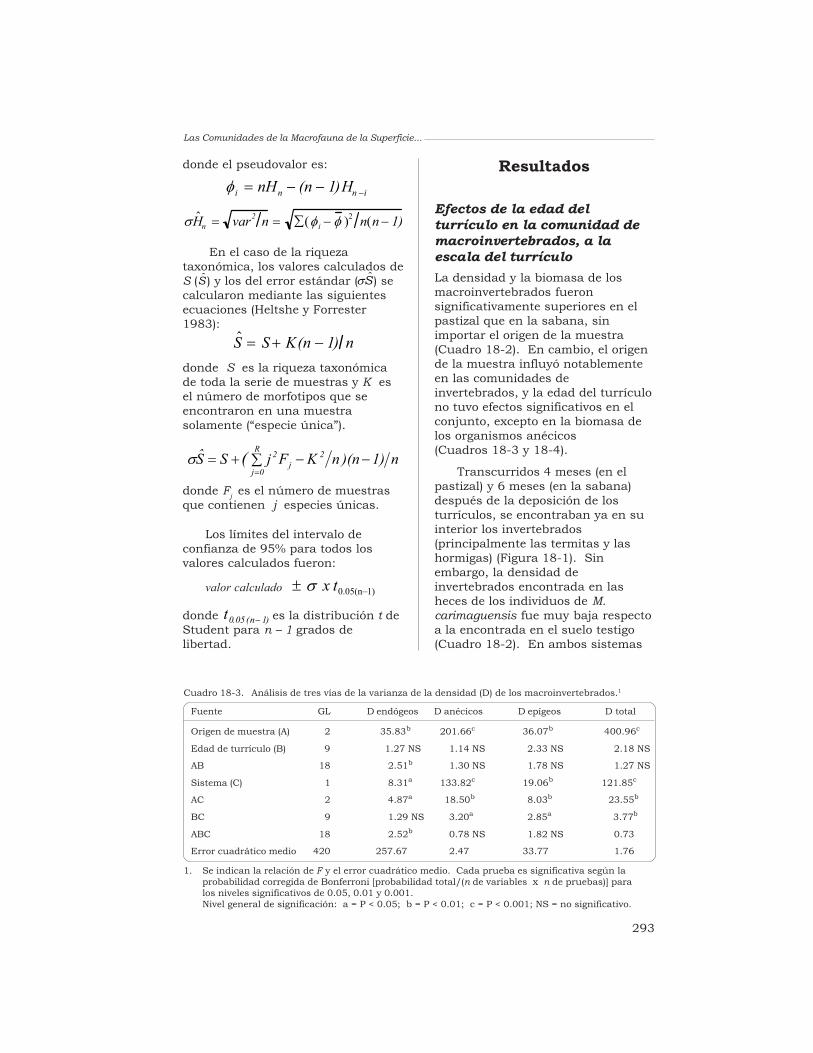
Las Comunidades de la Macrofauna de la Superficie...
293
donde el pseudovalor es:
En el caso de la riquezataxonómica, los valores calculados deS ( ) y los del error estándar ( ) secalcularon mediante las siguientesecuaciones (Heltshe y Forrester1983):
donde S es la riqueza taxonómicade toda la serie de muestras y K esel número de morfotipos que seencontraron en una muestrasolamente (“especie única”).
donde Fj es el número de muestrasque contienen j especies únicas.
Los límites del intervalo deconfianza de 95% para todos losvalores calculados fueron:
valor calculado 1)0.05(n−± txσ
donde es la distribución t deStudent para n – 1 grados delibertad.
Cuadro 18-3. Análisis de tres vías de la varianza de la densidad (D) de los macroinvertebrados.1
Fuente GL D endógeos D anécicos D epígeos D total
Origen de muestra (A) 2 35.83b 201.66c 36.07b 400.96c
Edad de turrículo (B) 9 1.27 NS 1.14 NS 2.33 NS 2.18 NS
AB 18 2.51b 1.30 NS 1.78 NS 1.27 NS
Sistema (C) 1 8.31a 133.82c 19.06b 121.85c
AC 2 4.87a 18.50b 8.03b 23.55b
BC 9 1.29 NS 3.20a 2.85a 3.77b
ABC 18 2.52b 0.78 NS 1.82 NS 0.73
Error cuadrático medio 420 257.67 2.47 33.77 1.76
1. Se indican la relación de F y el error cuadrático medio. Cada prueba es significativa según laprobabilidad corregida de Bonferroni [probabilidad total/(n de variables x n de pruebas)] paralos niveles significativos de 0.05, 0.01 y 0.001.Nivel general de significación: a = P < 0.05; b = P < 0.01; c = P < 0.001; NS = no significativo.
Resultados
Efectos de la edad delturrículo en la comunidad demacroinvertebrados, a laescala del turrículoLa densidad y la biomasa de losmacroinvertebrados fueronsignificativamente superiores en elpastizal que en la sabana, sinimportar el origen de la muestra(Cuadro 18-2). En cambio, el origende la muestra influyó notablementeen las comunidades deinvertebrados, y la edad del turrículono tuvo efectos significativos en elconjunto, excepto en la biomasa delos organismos anécicos(Cuadros 18-3 y 18-4).
Transcurridos 4 meses (en elpastizal) y 6 meses (en la sabana)después de la deposición de losturrículos, se encontraban ya en suinterior los invertebrados(principalmente las termitas y lashormigas) (Figura 18-1). Sinembargo, la densidad deinvertebrados encontrada en lasheces de los individuos de M.carimaguensis fue muy baja respectoa la encontrada en el suelo testigo(Cuadro 18-2). En ambos sistemas
φ i = nHn − (n − 1)Hn −i
σ ˆ H n = var2 n = (φ i − φ )2∑ n(n − 1)
ˆ S = S + K(n − 1) n
(R
0j
2j
2 n1)(n)nKFjSS ∑ −−+==
σ
t0.05 (n− 1)
S Sσ
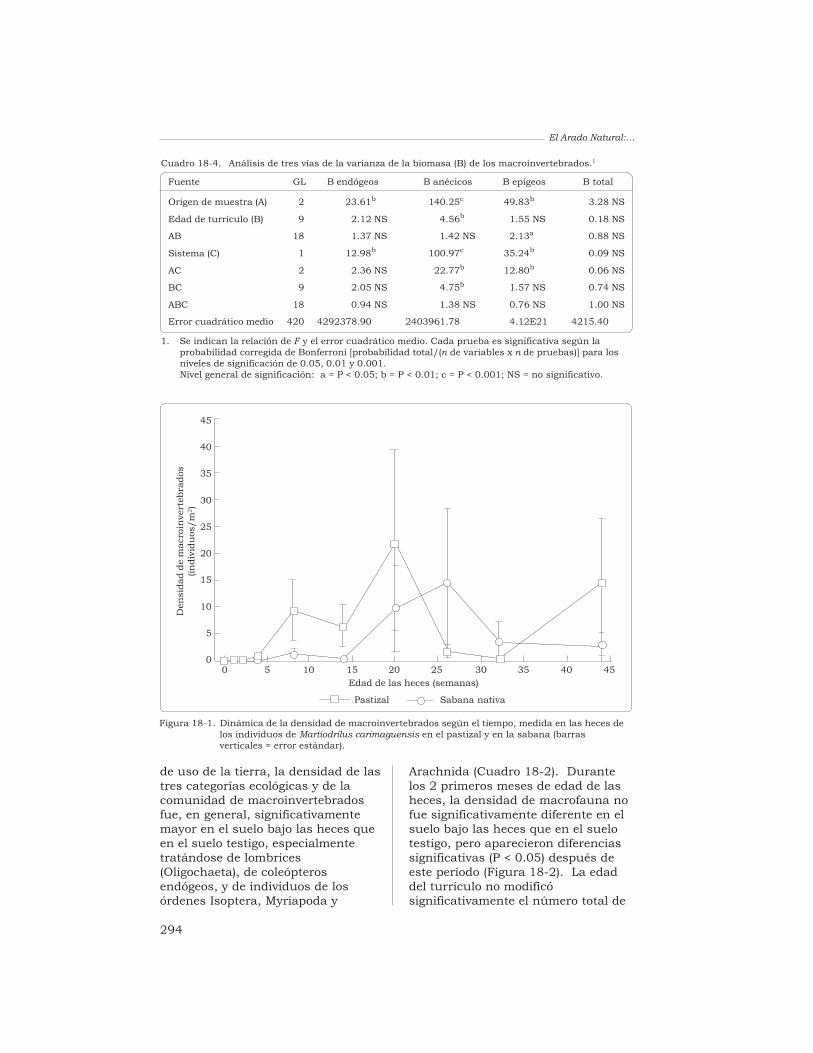
294
El Arado Natural:…
Arachnida (Cuadro 18-2). Durantelos 2 primeros meses de edad de lasheces, la densidad de macrofauna nofue significativamente diferente en elsuelo bajo las heces que en el suelotestigo, pero aparecieron diferenciassignificativas (P < 0.05) después deeste período (Figura 18-2). La edaddel turrículo no modificósignificativamente el número total de
de uso de la tierra, la densidad de lastres categorías ecológicas y de lacomunidad de macroinvertebradosfue, en general, significativamentemayor en el suelo bajo las heces queen el suelo testigo, especialmentetratándose de lombrices(Oligochaeta), de coleópterosendógeos, y de individuos de losórdenes Isoptera, Myriapoda y
Cuadro 18-4. Análisis de tres vías de la varianza de la biomasa (B) de los macroinvertebrados.1
Fuente GL B endógeos B anécicos B epígeos B total
Origen de muestra (A) 2 23.61b 140.25c 49.83b 3.28 NS
Edad de turrículo (B) 9 2.12 NS 4.56b 1.55 NS 0.18 NS
AB 18 1.37 NS 1.42 NS 2.13a 0.88 NS
Sistema (C) 1 12.98b 100.97c 35.24b 0.09 NS
AC 2 2.36 NS 22.77b 12.80b 0.06 NS
BC 9 2.05 NS 4.75b 1.57 NS 0.74 NS
ABC 18 0.94 NS 1.38 NS 0.76 NS 1.00 NS
Error cuadrático medio 420 4292378.90 2403961.78 4.12E21 4215.40
1. Se indican la relación de F y el error cuadrático medio. Cada prueba es significativa según laprobabilidad corregida de Bonferroni [probabilidad total/(n de variables x n de pruebas)] para losniveles de significación de 0.05, 0.01 y 0.001.Nivel general de significación: a = P < 0.05; b = P < 0.01; c = P < 0.001; NS = no significativo.
45
40
35
30
25
20
15
10
5
00 5 10 15 20 25 30 35 40 45
Edad de las heces (semanas)
Den
sida
d de
mac
roin
vert
ebra
dos
(indi
vidu
os/m
2 )
Pastizal Sabana nativa
Figura 18-1. Dinámica de la densidad de macroinvertebrados según el tiempo, medida en las heces delos individuos de Martiodrilus carimaguensis en el pastizal y en la sabana (barrasverticales = error estándar).
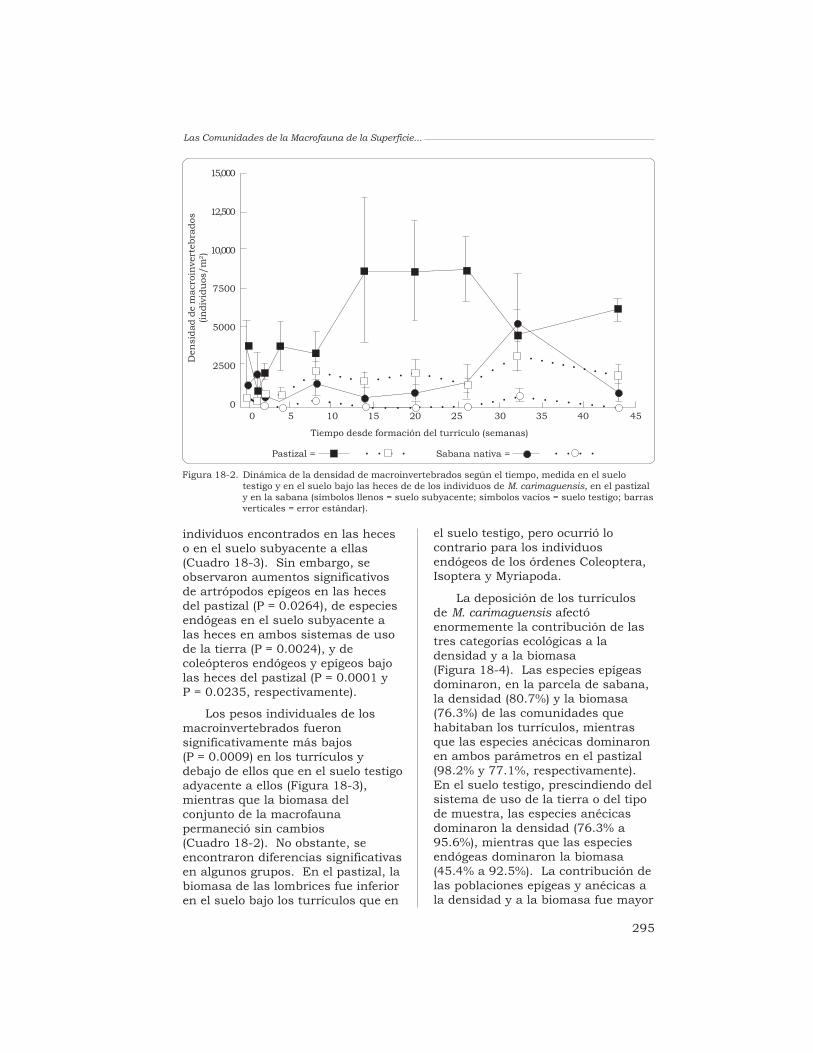
Las Comunidades de la Macrofauna de la Superficie...
295
el suelo testigo, pero ocurrió locontrario para los individuosendógeos de los órdenes Coleoptera,Isoptera y Myriapoda.
La deposición de los turrículosde M. carimaguensis afectóenormemente la contribución de lastres categorías ecológicas a ladensidad y a la biomasa(Figura 18-4). Las especies epígeasdominaron, en la parcela de sabana,la densidad (80.7%) y la biomasa(76.3%) de las comunidades quehabitaban los turrículos, mientrasque las especies anécicas dominaronen ambos parámetros en el pastizal(98.2% y 77.1%, respectivamente).En el suelo testigo, prescindiendo delsistema de uso de la tierra o del tipode muestra, las especies anécicasdominaron la densidad (76.3% a95.6%), mientras que las especiesendógeas dominaron la biomasa(45.4% a 92.5%). La contribución delas poblaciones epígeas y anécicas ala densidad y a la biomasa fue mayor
individuos encontrados en las heceso en el suelo subyacente a ellas(Cuadro 18-3). Sin embargo, seobservaron aumentos significativosde artrópodos epígeos en las hecesdel pastizal (P = 0.0264), de especiesendógeas en el suelo subyacente alas heces en ambos sistemas de usode la tierra (P = 0.0024), y decoleópteros endógeos y epígeos bajolas heces del pastizal (P = 0.0001 yP = 0.0235, respectivamente).
Los pesos individuales de losmacroinvertebrados fueronsignificativamente más bajos(P = 0.0009) en los turrículos ydebajo de ellos que en el suelo testigoadyacente a ellos (Figura 18-3),mientras que la biomasa delconjunto de la macrofaunapermaneció sin cambios(Cuadro 18-2). No obstante, seencontraron diferencias significativasen algunos grupos. En el pastizal, labiomasa de las lombrices fue inferioren el suelo bajo los turrículos que en
○○
○○
○○
○○
○○
○
○
○
○
○
○
○
○
○
○
○○
○○
○○
○○
○○
○
○
○
○
○○
15,000
12,500
10,000
7500
5000
2500
00 5 10 15 20 25 30 35 40 45
Tiempo desde formación del turrículo (semanas)
Den
sida
d de
mac
roin
vert
ebra
dos
(indi
vidu
os/m
2 )
Pastizal = Sabana nativa =
Figura 18-2. Dinámica de la densidad de macroinvertebrados según el tiempo, medida en el suelotestigo y en el suelo bajo las heces de de los individuos de M. carimaguensis, en el pastizaly en la sabana (símbolos llenos = suelo subyacente; símbolos vacíos = suelo testigo; barrasverticales = error estándar).
○○
○○
○○
○○
○○
○○
○○
○ ○ ○ ○ ○
○○
○○
○○
○
○
○○
○ ○ ○ ○
○ ○ ○ ○ ○ ○ ○ ○
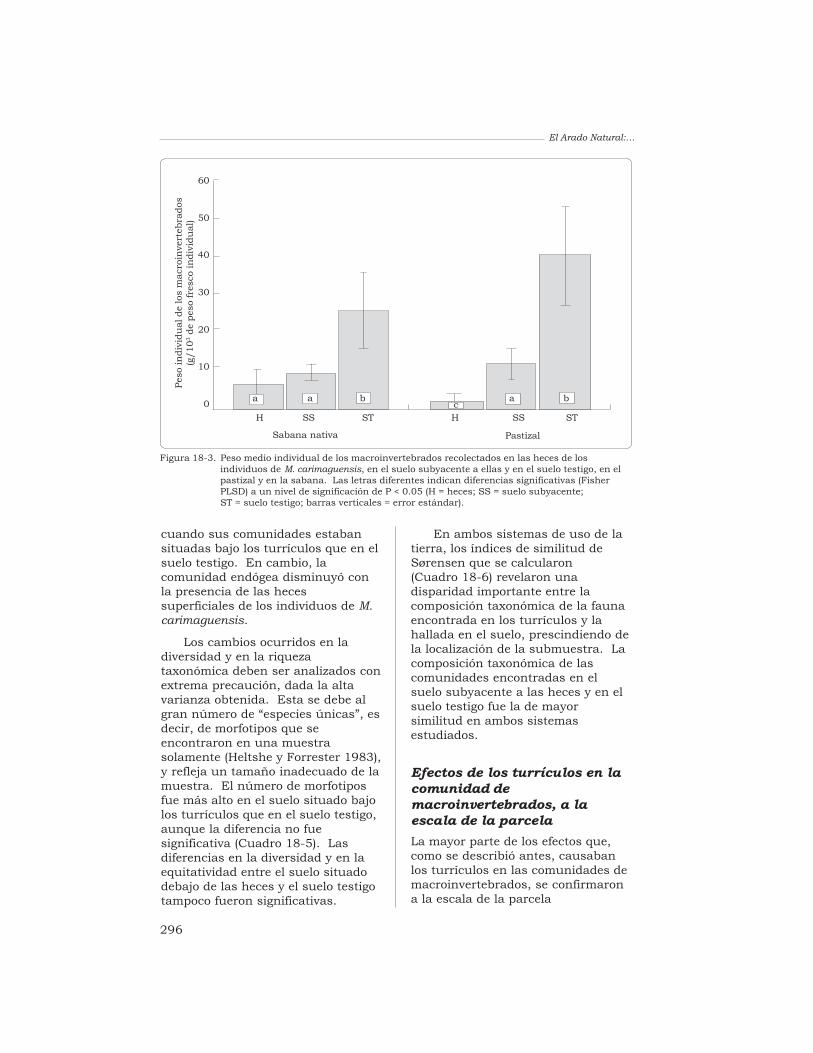
296
El Arado Natural:…
60
50
40
30
20
10
0H SS ST H SS ST
Pastizal
Peso
indi
vidu
al d
e lo
s m
acro
inve
rteb
rado
s(g
/103
de
peso
fres
co in
divi
dual
)
Figura 18-3. Peso medio individual de los macroinvertebrados recolectados en las heces de losindividuos de M. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en elpastizal y en la sabana. Las letras diferentes indican diferencias significativas (FisherPLSD) a un nivel de significación de P < 0.05 (H = heces; SS = suelo subyacente;ST = suelo testigo; barras verticales = error estándar).
ca a b a b
Sabana nativa
cuando sus comunidades estabansituadas bajo los turrículos que en elsuelo testigo. En cambio, lacomunidad endógea disminuyó conla presencia de las hecessuperficiales de los individuos de M.carimaguensis.
Los cambios ocurridos en ladiversidad y en la riquezataxonómica deben ser analizados conextrema precaución, dada la altavarianza obtenida. Esta se debe algran número de “especies únicas”, esdecir, de morfotipos que seencontraron en una muestrasolamente (Heltshe y Forrester 1983),y refleja un tamaño inadecuado de lamuestra. El número de morfotiposfue más alto en el suelo situado bajolos turrículos que en el suelo testigo,aunque la diferencia no fuesignificativa (Cuadro 18-5). Lasdiferencias en la diversidad y en laequitatividad entre el suelo situadodebajo de las heces y el suelo testigotampoco fueron significativas.
En ambos sistemas de uso de latierra, los índices de similitud deSørensen que se calcularon(Cuadro 18-6) revelaron unadisparidad importante entre lacomposición taxonómica de la faunaencontrada en los turrículos y lahallada en el suelo, prescindiendo dela localización de la submuestra. Lacomposición taxonómica de lascomunidades encontradas en elsuelo subyacente a las heces y en elsuelo testigo fue la de mayorsimilitud en ambos sistemasestudiados.
Efectos de los turrículos en lacomunidad demacroinvertebrados, a laescala de la parcelaLa mayor parte de los efectos que,como se describió antes, causabanlos turrículos en las comunidades demacroinvertebrados, se confirmarona la escala de la parcela
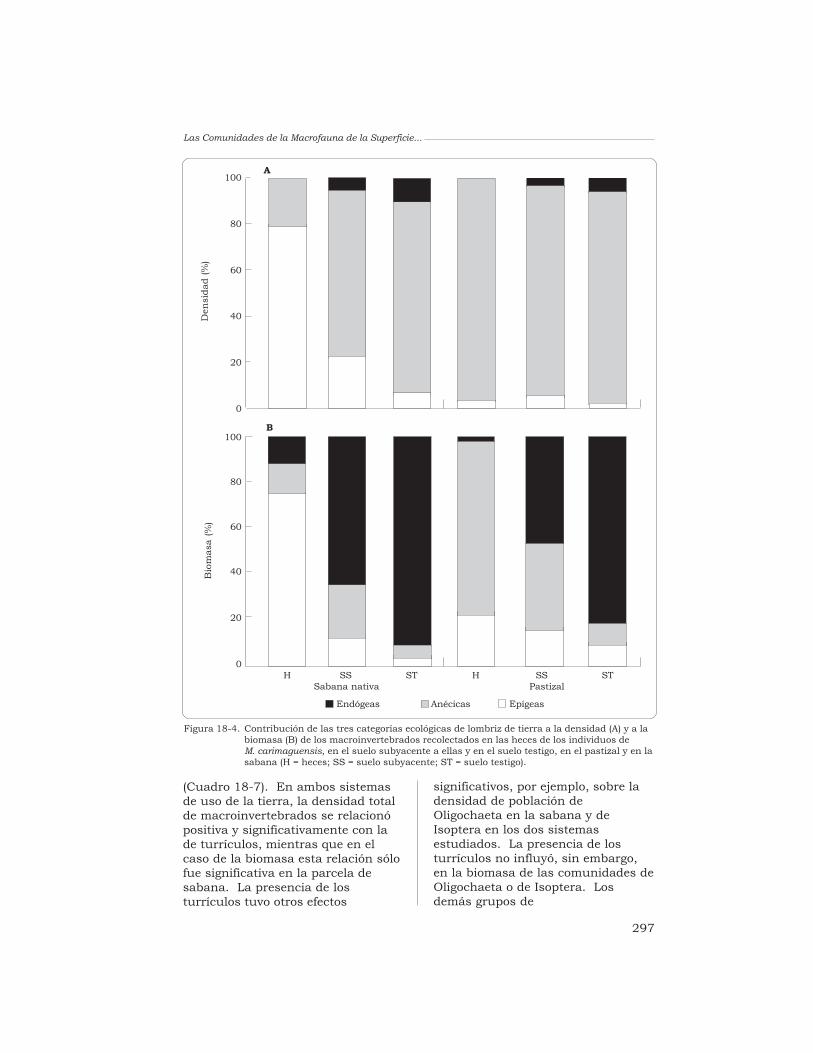
Las Comunidades de la Macrofauna de la Superficie...
297
100
80
60
40
20
0
H SS ST H SS ST
Den
sida
d (%
)
100
80
60
40
20
0
Bio
mas
a (%
)
B
A
Sabana nativa Pastizal
Endógeas Anécicas Epígeas
Figura 18-4. Contribución de las tres categorías ecológicas de lombriz de tierra a la densidad (A) y a labiomasa (B) de los macroinvertebrados recolectados en las heces de los individuos deM. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en el pastizal y en lasabana (H = heces; SS = suelo subyacente; ST = suelo testigo).
(Cuadro 18-7). En ambos sistemasde uso de la tierra, la densidad totalde macroinvertebrados se relacionópositiva y significativamente con lade turrículos, mientras que en elcaso de la biomasa esta relación sólofue significativa en la parcela desabana. La presencia de losturrículos tuvo otros efectos
significativos, por ejemplo, sobre ladensidad de población deOligochaeta en la sabana y deIsoptera en los dos sistemasestudiados. La presencia de losturrículos no influyó, sin embargo,en la biomasa de las comunidades deOligochaeta o de Isoptera. Losdemás grupos de
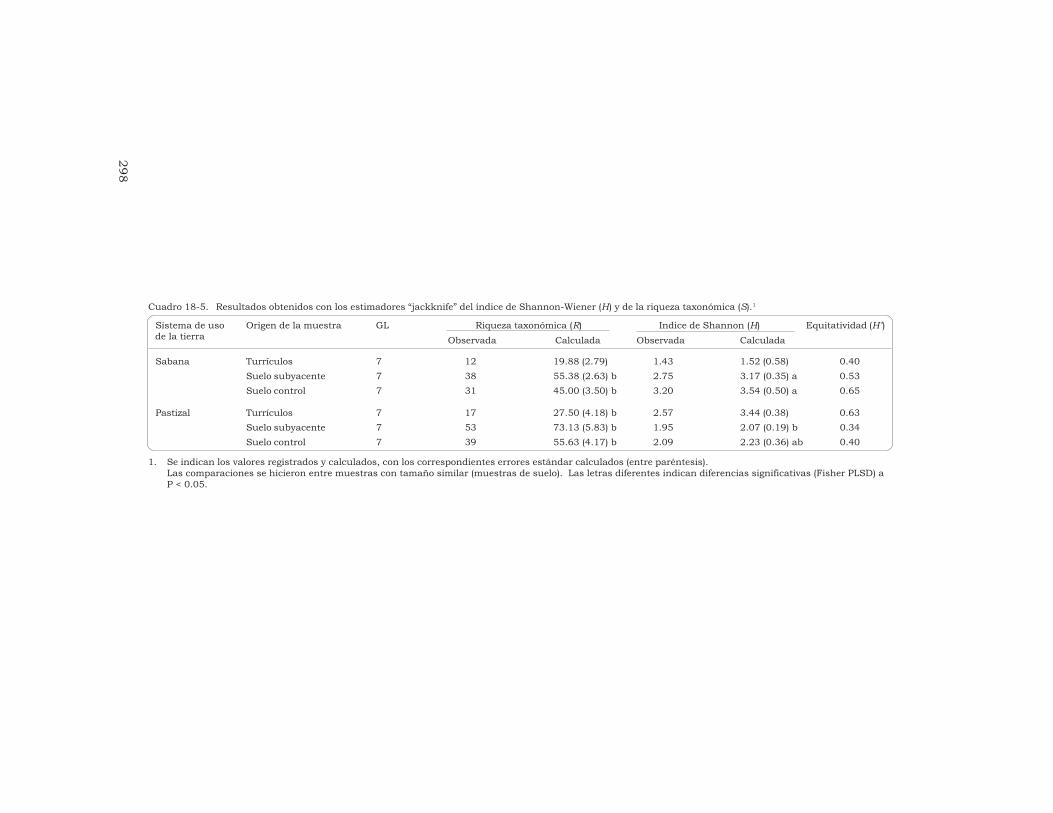
298
El A
rado Natural:…
Cuadro 18-5. Resultados obtenidos con los estimadores “jackknife” del índice de Shannon-Wiener (H) y de la riqueza taxonómica (S).1
Sistema de uso Origen de la muestra GL Riqueza taxonómica (R) Indice de Shannon (H) Equitatividad (H’) Observada Calculada Observada Calculada
Sabana Turrículos 7 12 19.88 (2.79) 1.43 1.52 (0.58) 0.40Suelo subyacente 7 38 55.38 (2.63) b 2.75 3.17 (0.35) a 0.53Suelo control 7 31 45.00 (3.50) b 3.20 3.54 (0.50) a 0.65
Pastizal Turrículos 7 17 27.50 (4.18) b 2.57 3.44 (0.38) 0.63Suelo subyacente 7 53 73.13 (5.83) b 1.95 2.07 (0.19) b 0.34Suelo control 7 39 55.63 (4.17) b 2.09 2.23 (0.36) ab 0.40
1. Se indican los valores registrados y calculados, con los correspondientes errores estándar calculados (entre paréntesis).Las comparaciones se hicieron entre muestras con tamaño similar (muestras de suelo). Las letras diferentes indican diferencias significativas (Fisher PLSD) aP < 0.05.
de la tierra
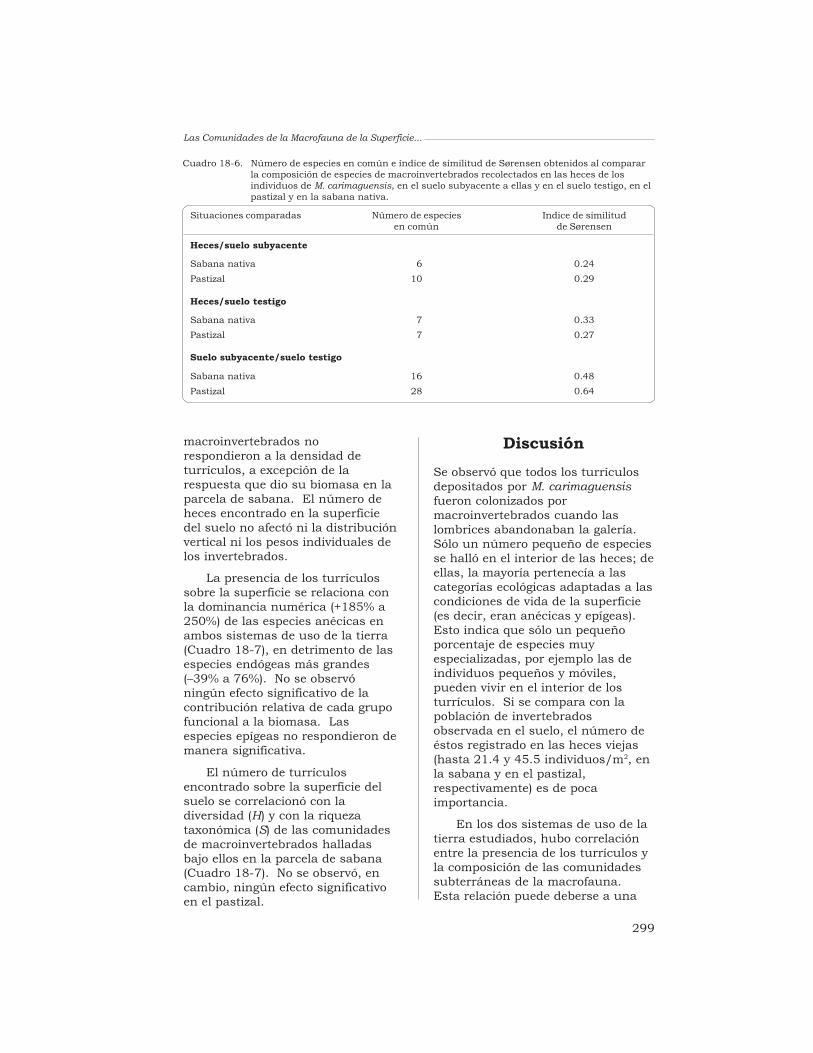
Las Comunidades de la Macrofauna de la Superficie...
299
Cuadro 18-6. Número de especies en común e índice de similitud de Sørensen obtenidos al compararla composición de especies de macroinvertebrados recolectados en las heces de losindividuos de M. carimaguensis, en el suelo subyacente a ellas y en el suelo testigo, en elpastizal y en la sabana nativa.
Situaciones comparadas Número de especies Indice de similituden común de Sørensen
Heces/suelo subyacente
Sabana nativa 6 0.24Pastizal 10 0.29
Heces/suelo testigo
Sabana nativa 7 0.33Pastizal 7 0.27
Suelo subyacente/suelo testigo
Sabana nativa 16 0.48Pastizal 28 0.64
macroinvertebrados norespondieron a la densidad deturrículos, a excepción de larespuesta que dio su biomasa en laparcela de sabana. El número deheces encontrado en la superficiedel suelo no afectó ni la distribuciónvertical ni los pesos individuales delos invertebrados.
La presencia de los turrículossobre la superficie se relaciona conla dominancia numérica (+185% a250%) de las especies anécicas enambos sistemas de uso de la tierra(Cuadro 18-7), en detrimento de lasespecies endógeas más grandes(–39% a 76%). No se observóningún efecto significativo de lacontribución relativa de cada grupofuncional a la biomasa. Lasespecies epígeas no respondieron demanera significativa.
El número de turrículosencontrado sobre la superficie delsuelo se correlacionó con ladiversidad (H) y con la riquezataxonómica (S) de las comunidadesde macroinvertebrados halladasbajo ellos en la parcela de sabana(Cuadro 18-7). No se observó, encambio, ningún efecto significativoen el pastizal.
DiscusiónSe observó que todos los turrículosdepositados por M. carimaguensisfueron colonizados pormacroinvertebrados cuando laslombrices abandonaban la galería.Sólo un número pequeño de especiesse halló en el interior de las heces; deellas, la mayoría pertenecía a lascategorías ecológicas adaptadas a lascondiciones de vida de la superficie(es decir, eran anécicas y epígeas).Esto indica que sólo un pequeñoporcentaje de especies muyespecializadas, por ejemplo las deindividuos pequeños y móviles,pueden vivir en el interior de losturrículos. Si se compara con lapoblación de invertebradosobservada en el suelo, el número deéstos registrado en las heces viejas(hasta 21.4 y 45.5 individuos/m2, enla sabana y en el pastizal,respectivamente) es de pocaimportancia.
En los dos sistemas de uso de latierra estudiados, hubo correlaciónentre la presencia de los turrículos yla composición de las comunidadessubterráneas de la macrofauna.Esta relación puede deberse a una
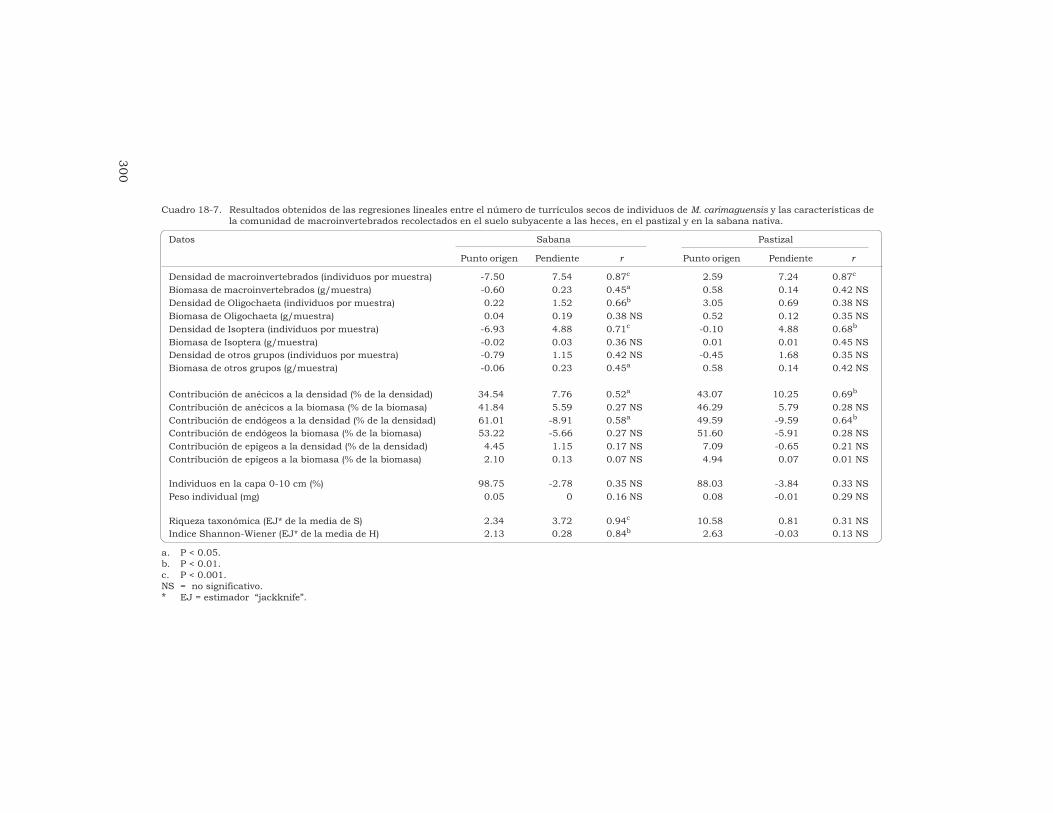
300
El A
rado Natural:…
Cuadro 18-7. Resultados obtenidos de las regresiones lineales entre el número de turrículos secos de individuos de M. carimaguensis y las características dela comunidad de macroinvertebrados recolectados en el suelo subyacente a las heces, en el pastizal y en la sabana nativa.
Datos Sabana Pastizal
Punto origen Pendiente r Punto origen Pendiente r
Densidad de macroinvertebrados (individuos por muestra) -7.50 7.54 0.87c 2.59 7.24 0.87c
Biomasa de macroinvertebrados (g/muestra) -0.60 0.23 0.45a 0.58 0.14 0.42 NSDensidad de Oligochaeta (individuos por muestra) 0.22 1.52 0.66b 3.05 0.69 0.38 NSBiomasa de Oligochaeta (g/muestra) 0.04 0.19 0.38 NS 0.52 0.12 0.35 NSDensidad de Isoptera (individuos por muestra) -6.93 4.88 0.71c -0.10 4.88 0.68b
Biomasa de Isoptera (g/muestra) -0.02 0.03 0.36 NS 0.01 0.01 0.45 NSDensidad de otros grupos (individuos por muestra) -0.79 1.15 0.42 NS -0.45 1.68 0.35 NSBiomasa de otros grupos (g/muestra) -0.06 0.23 0.45a 0.58 0.14 0.42 NS
Contribución de anécicos a la densidad (% de la densidad) 34.54 7.76 0.52a 43.07 10.25 0.69b
Contribución de anécicos a la biomasa (% de la biomasa) 41.84 5.59 0.27 NS 46.29 5.79 0.28 NSContribución de endógeos a la densidad (% de la densidad) 61.01 -8.91 0.58a 49.59 -9.59 0.64b
Contribución de endógeos la biomasa (% de la biomasa) 53.22 -5.66 0.27 NS 51.60 -5.91 0.28 NSContribución de epígeos a la densidad (% de la densidad) 4.45 1.15 0.17 NS 7.09 -0.65 0.21 NSContribución de epígeos a la biomasa (% de la biomasa) 2.10 0.13 0.07 NS 4.94 0.07 0.01 NS
Individuos en la capa 0-10 cm (%) 98.75 -2.78 0.35 NS 88.03 -3.84 0.33 NSPeso individual (mg) 0.05 0 0.16 NS 0.08 -0.01 0.29 NS
Riqueza taxonómica (EJ* de la media de S) 2.34 3.72 0.94c 10.58 0.81 0.31 NSIndice Shannon-Wiener (EJ* de la media de H) 2.13 0.28 0.84b 2.63 -0.03 0.13 NS
a. P < 0.05.b. P < 0.01.c. P < 0.001.NS = no significativo.* EJ = estimador “jackknife”.

Las Comunidades de la Macrofauna de la Superficie...
301
respuesta común de esta especie delombriz y de otros invertebrados a lapresencia de puntos críticos (“hotspots”), por ejemplo áreas ricas ennutrientes, en el campo. Ahora bien,la densidad de invertebradosaumentó gradualmente en el suelosubyacente a los turrículos duranteel envejecimiento de éstos, unresultado que los presenta como losverdaderos responsables de loscambios observados. Unaexplicación alterna sería que laslombrices encuentran estos puntoscríticos mucho más rápidamente quelos demás invertebrados, aunqueparece improbable si se consideraque la movilidad de las lombrices,comparada con la de otrosinvertebrados forrajeros activos comolas termitas o los artrópodostransformadores de hojarasca, esrelativamente baja.
La presencia de los turrículos enla superficie del suelo ocasiona unaumento en la densidad de poblaciónde los invertebrados pequeños en elsuelo subyacente (+317% y 212%, enla sabana y en el pastizal,respectivamente). La deposición deheces superficiales puede favorecerlas condiciones de vida de un grannúmero de especies del suelo, comose deduce de la respuesta positiva dela riqueza y la diversidadtaxonómicas al aumento del númerode turrículos. Ahora bien, este efectofue significativo solamente en lasabana, lo que sugiere que laactividad de las lombrices puedecausar un impacto muy grandecuando existen condicionesdesfavorables sobre la superficie delsuelo —por ejemplo, ausencia dehojarasca en una sabana sometida ala quema— y ningún impacto, o muypequeño, cuando las condiciones delentorno son favorables —como en elcaso del pastizal.
Con el fin de explicar la forma enque las lombrices pueden influir en
las comunidades demacroinvertebrados del suelo, se hanformulado dos hipótesis que explicanlas modificaciones estructurales ytróficas del suelo a partir de laformación de las estructurasbiogénicas de las lombrices:
1. Primera, algunos invertebradosprefieren vivir en el interior de losturrículos o en el suelosubyacente a ellos, dado que haycambios en la cantidad y en lacalidad del recurso energético deéstos. Los turrículos deM. carimaguensis tienen un altocontenido de MO (Decaëns et al.1999; Guggenberger et al. 1996;Rangel et al. 1999) y representanun sustrato alimenticio valiosopara ciertas lombrices pequeñaspolihúmicas y para las termitashumívoras. La biomasaradicular aumenta localmentebajo los turrículos (Decaëns et al.1999), lo que puede serbeneficioso para las larvas decoleópteros rizófagos. Algunasespecies que viven entre lahojarasca, como laspertenecientes a los gruposIsopoda y Diplopoda, pueden seratraídas por los cambios queocurren en la composición de lahojarasca que ha sido procesadapor las lombrices (Szlavecz1985). Los depredadorespequeños (Chilopoda, Arachnida)pueden encontrarse con una altadensidad de pequeñas presas demicrofauna y de mesofauna, queresultaron favorecidas, a su vez,por mejores condiciones de vidaderivadas de la acción de laslombrices (Brown 1995; Lorangeret al. 1998).
2. Segunda, hay otras especies quepueden responder a los cambiosproducidos en la estructura físicadel suelo y a la formación denuevos microhábitat específicos acausa de la actividad de las

302
El Arado Natural:…
lombrices. Los macroporos queresultan de tal actividad sonconsiderados como hábitat paraalgunos microinvertebrados yotros mesoinvertebrados (Haukka1991; Loranger et al. 1998). Kirk(1981) encontró un gran númerode huevos de individuos delgénero Diabrotica (Coleoptera) enlas galerías de las lombrices.Durante este estudio se encontróque las hormigas y las termitasusaban las galerías como mediode comunicación. En el ambienteconstituido por una sabanadespués de la quema, losinvertebrados que viven en lasuperficie, es decir, entre lahojarasca y en el medio herbáceo,quedan totalmente desprotegidos;las estructuras producidas porlas lombrices pueden serempleadas entonces por esosartrópodos de la superficie comorefugios específicos, que losayudarían a conservar suspoblaciones o a recolonizarrápidamente la superficie delsuelo después del paso del fuego.
Casi todos los resultados aquíobtenidos indican que la presencia delos turrículos de M. carimaguensissobre la superficie del sueloaumentan, de preferencia, laspoblaciones de los invertebradosanécicos y epígeos en detrimento delos endógeos. Pueden darse dosrazones:
- la concentración de recursosorgánicos cerca de lasuperficie, en las hecessuperficiales;
- la formación de nuevoshábitat específicos sobre lasuperficie del suelo.
Las estructuras creadas en elinterior del suelo (galerías y heces)pueden causar efectos diferentes en
otros grupos de invertebrados, comoocurre con la pequeña lombrizpolihúmica de la familiaOcnerodrilidae, que se encuentrageneralmente alimentándose de lasheces de los individuos deM. carimaguensis hasta unaprofundidad de 70 cm (Jiménez et al.1998a).
Conclusiones GeneralesLos resultados presentados en esteestudio apoyan la hipótesis de Lavelle(1996) sobre las biodiversidadesencajadas, en la que sostiene que losingenieros del ecosistema (sensuJones et al. 1994) determinan laestructura de las comunidades deotros organismos más pequeños.
Se ha demostrado también eneste estudio que las heces de unalombriz anécica pueden ejercerefectos significativos en lascomunidades de macrofauna quehabitan el suelo. Cuando estasheces son superficiales (losturrículos), se produce un aumentoglobal de la densidad de laspoblaciones de macroinvertebrados yse favorecen las especies epígeas yanécicas en detrimento de lasendógeas. Es posible lanzar lahipótesis de que estos resultados sedeben a modificaciones en lossiguientes aspectos:
- el tamaño y la abundancia demicrohábitat específicos paralos invertebrados máspequeños (fisuras en lasheces o galerías);
- la localización y la dinámicade los recursos orgánicos queemplea, normalmente, unagran parte de la comunidadde organismos del suelo(carbono en las heces y en elcórtex de la galería).

Las Comunidades de la Macrofauna de la Superficie...
303
Las actividades de las lombricesde tierra, sin embargo, no estánrestringidas a la superficie del suelo.Una gran cantidad de sus estructurasse crean en el perfil del suelo ypueden actuar, por tanto, sobre otrasespecies a diferentes profundidades.La investigación futura deberíacentrarse en este aspecto y permitirasí el desarrollo de una buenaperspectiva del impacto que hacen laslombrices en la biodiversidad delsuelo.
La producción de estructurasbiogénicas por especies de lombricesgrandes parece ser un factorimportante en la determinación de laestructura de las comunidades demacroinvertebrados del suelo.¿Causan ellas el mismo efecto enotros grupos funcionales deorganismos del suelo? ¿Quéimplicaciones tendrían en talesprocesos otras especies de ingenierosdel ecosistema? Los termiteros, porejemplo, albergan normalmente ungran número de invertebrados (Woody Sands 1978), aunque faltanestudios que describan estefenómeno. Elaborar una tipología delas estructuras biogénicas, junto conlos efectos que ellas producen en losorganismos del suelo, es un pasonecesario para poder describir losnexos existentes entre los diferentesgrupos funcionales aquímencionados.
La colonización de las heces detamaño grande por otrosinvertebrados es un aspecto crítico dela conservación y la dinámica de laMO y de la regulación de laspropiedades físicas del suelo. Cuandolos invertebrados pequeños ingierenlas heces grandes y compactas de laslombrices, evitan su excesivaacumulación en la superficie del suelola cual, de presentarse, conduciría auna compactación superficial delsuelo y afectaría negativamente elcrecimiento vegetal (Chauvel et al.
1999; Rose y Wood 1980). Por otrolado, dichos invertebrados puedenreactivar la dinámica de la MOhaciendo que ciertos recursosorgánicos, que estaban inmovilizadosen las heces secas, quedendisponibles para los microorganismos(Lavelle 1996). La prueba de estahipótesis se considera crítica parapoder identificar las funciones de losinvertebrados en relación con elimpacto que causan en laspropiedades del suelo, y paraestablecer la importancia funcionalque tiene realmente la biodiversidaden el suelo.
AgradecimientosLos autores quieren agradecer aR.J. Thomas y D.K. Friesen (CIAT) porel apoyo técnico y económico recibidode ellos. Agradecen también aA.V. Spain (CSIRO), I. Poudevigne(Universidad de Rouen) y J.-P. Rossi(ORSTOM) por sus comentarios útilessobre una primera versión de esteartículo. Finalmente, a E. Baudry,X. Graves y E. Mariani por la granayuda que prestaron en el cálculo delos estimadores del procedimiento“jackknife”.
ReferenciasAdams JE; McCune ED. 1979. Application
of the generalized jackknife toShannon’s measure of informationused as an index of diversity. En:Grassle JF; Patil GP; Smith WK;Taillie C, eds. Ecological diversity intheory and practice. InternationalCo-operative Publisher House,Fairland, MD, Estados Unidos.p 117-132.
Anderson JM. 1995. Soil organisms asengineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linking speciesand ecosystems. Chapman and Hall,Londres. p 94-106.

304
El Arado Natural:…
Anderson JM; Ingram J, eds. 1993.Tropical soil biology and fertility: ahandbook of methods. 2a. ed. CAB,Oxford, Reino Unido. 221 p.
Barois I; Lavelle P. 1986. Changes inrespiration rate and some physico-chemical properties of a tropical soilduring transit through Pontoscolexcorethrurus. Soil Biol Biochem18:539-541.
Bouché MB. 1977. Statégieslombriciennes. En: Lohm U;Persson T, eds. Soil organisms ascomponent of ecosystems. Ecol Bull25:122-132.
Brown G. 1995. How do earthwormsaffect microfloral and faunalcommunity diversity? Plant Soil170:209-231.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm.Nature 398:32-33.
Cooper DW. 1968. The significance levelin multiple tests madesimultaneously. Heredity23:614-617.
Daniel O; Anderson JM. 1992. Microbialbiomass and activity in contrastingsoil materials after passage throughthe gut of the earthworm Lumbricusrubellus Hoffmeister. Soil BiolBiochem 24:465-470.
Decaëns T; Rangel AF; Asakawa N;Thomas R.J. 1999. Carbon andnitrogen dynamics in ageing castsin grasslands of the eastern plainsof Colombia. Biol Fertil Soils30:20-28.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30:157-168.
Guggenberger G; Thomas RJ; Zech W.1996. Soil organic matter withinearthworm casts of an anecic-endogeic tropical pasturecommunity, Colombia. Appl SoilEcol 3:263-274.
Haukka J. 1991. Spatial distribution andformation of earthworm burrows.Pedobiologia 35:175-178.
Heltshe JF; Forrester NE. 1983.Estimating species richness usingthe jackknife procedure. Biometrics39:1-11.
Heltshe JF; Forrester NE. 1985.Statistical evaluation of thejackknife estimate of diversity whenusing quadrant sample. Ecology66:107-111.
Hurlbert SH. 1971. The nonconcept ofspecies diversity: a critique andalternative parameter. Ecology52:577-586.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher M; Thomas RJ.1998a. Earthworm communities innative savannas and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998b. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Kirk VM. 1981. Earthworm burrows asoviposition sites for western andnorthern corn rootworms(Diabrotica : Coleoptera). J KansasEntomol Soc 54:68-74.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.

Las Comunidades de la Macrofauna de la Superficie...
305
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Legendre L; Legendre P. 1979. Écologienumérique; Tome 2: La structure desDonnées Écologiques. Masson, París,Francia, y Presses de l’Université duQuébec, Montreal, Canadá. 197 p.
Legendre P; Vaudor A. 1991. The R-package: multidimentional analysis,spatial analysis. Université deMontréal, Département des SciencesBiologiques, Montreal, Canadá.
Lilliefors HW. 1967. The Kolmogorov-Smirnov test for normality with meanand variance unknown. J Am StatAssoc 62:399-402.
Loranger G; Ponge JF; Blanchart E; LavelleP. 1998. Impact of earthworms onthe diversity of microarthropods in avertisol (Martinique). Biol Fertil Soils27:21-27.
Marinissen JCY; Bok J. 1988. Earthworm-amended soil structure: its influenceon Collembola populations ingrasslands. Pedobiologia32:243-252.
Oliver I; Beattie AJ. 1995. Invertebratemorphospecies as surrogates forspecies: a case of study. ConservBiol 10:99-109.
Parkinson D; McLean MA. 1998. Impactsof earthworms on the communitystructure of other biota in forestsoils. En: Edwards CA, ed.Earthworm ecology. St Lucie Press,Columbus, OH, Estados Unidos.p 213-226.
Pielou EC. 1966. The measurement ofdiversity in different types ofbiological collections. J Theor Biol13:131-144.
Rangel AF; Madero E; Thomas RJ; FriesenDK; Decaëns T. 1999. Ion exchangeproperties of casts of the anecicearthworm (Martiodriluscarimaguensis Jiménez and Moreno)in a Colombian savanna Oxisol.Pedobiologia 43:795-801.
Rose CJ; Wood AW. 1980. Someenvironmental factors affectingearthworms populations and sweetpotato production in the Tari Basin,Papua New Guinea Highlands.Papua New Guinea Agric J 31:1-10.
Scheu S. 1987. Microbial activity andnutrient dynamics in earthwormcasts (Lumbricidae). Biol Fertil Soils5:230-234.
Scheu S. 1993. Analysis of the microbialnutrient status in soilmicrocompartments: earthwormfaeces from a basalt-limestonegradient. Geoderma 56:575-586.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed. WHFreeman and Company, NuevaYork, Estados Unidos.
Sørensen T. 1948. A method ofestablishing groups of equalamplitude in plant sociology basedon similarity of species content andits application to analysis of thevegetation on Danish commons.Biol Srk 5:1-34.
Szlavecz K. 1985. The effect ofmicrohabitats on the leaf litterdecomposition and on thedistribution of soil animals. HolarctEcol 8:33-38.
Thompson L; Thomas CD; Radley JMA;Williamson S; Lawton JH. 1993.The effects of earthworms andsnails in a simple plant community.Oecologia 95:171-178.
Tukey JW. 1958. Bias and confidence innot quite large samples. Ann MathStat 29:614.
Wood TG; Sands WA. 1978. The role oftermites in ecosystems. En: BrianMV, ed. Production ecology of antsand termites. Cambridge UniversityPress, Cambridge, Reino Unido.p 245-292.

306
El Arado Natural:…
ResumenSe investigaron los efectos causadospor la especie de lombriz de tierraMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae) en elbanco de semillas del suelo de unaparcela de sabana y de dos pastizalesintroducidos, en Colombia. Mediantetécnicas de germinación y aplicandoel método de lavado seguido detamizado, se comparó la densidad, ladiversidad y las tasas de germinaciónde las semillas en las hecesproducidas por las lombrices y en elsuelo que las circundaba. Seencontraron grandes cantidades desemillas en las heces (desde 58 hasta163 semillas/100 g de heces secas),cuyas tasas de germinación fueron de3 a 40 veces menores que en lasencontradas en el suelo a causa,posiblemente, del daño que sufrieron
durante su tránsito por el intestinode las lombrices. La composición deespecies de las semillas viableshalladas en las heces fue muydiferente de la que se observó en lasmuestras del suelo, tal vez a causade la ingestión selectiva de semillasque hicieron las lombrices. Cadaaño se deposita, en las hecessuperficiales, del 1% al 13% delbanco total de semillas del suelo.Tanto en la sabana como en uno delos pastizales estudiados, lacomposición de especies de lassemillas que germinaron se asemejómás a la vegetación epígea presentecuando las semillas provenían de lasheces que cuando provenían delsuelo. Estos resultados indican quelas semillas que, después de seringeridas por las lombrices,sobreviven al tránsito intestinal,presentan mayores oportunidades degerminar que las que forman partedel banco de semillas del suelo. Enconsecuencia, las heces seconsideran nichos de regeneraciónde plantas, y la actividad de laslombrices de tierra es, por ello, unfactor que favorece la expresión, enla vegetación epígea, del banco desemillas del suelo.
Palabras clave: banco de semillasdel suelo, heces de lombrices,pastizales tropicales, sabanas
CAPÍTULO 19
Efectos Causados por las Lombricesen el Banco de Semillas Permanentedel Suelo en Sistemas de Sabana y dePastizal en Colombia1
T. Decaëns*, L. Mariani**, N. Betancourt*** yJ. J. Jiménez***
1. Una versión diferente de este trabajo ha sidopublicado en Acta Oecologica 24, p 175-185,por Decaëns et al. Copyright © 2003 deElsevier. Impreso con permiso de la casaeditorial.
* Laboratoire d’Ecologie, UPRES EA 1293,UFR Sciences, Université de Rouen, F-76821Mont Saint Aignan Cedex, Francia.
** Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.
*** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
307
IntroducciónLos bancos de semillas del suelo seoriginan en la acumulación desemillas en el suelo y en supersistencia en él. Thompson yGrime (1979) los definieron como“todas las semillas viables de unaespecie en un tiempo dado,incluyendo en ellas las que están enla superficie del suelo y las que sehallan debajo de ésta”. Harper(1977) considera que las semillasconstituyen la “memoria” de lasplantas porque, durante décadas,conservan los genotipos que no sehan manifestado aún en lavegetación epígea.
Además del papel quedesempeñan en la dinámica de lasformaciones vegetales naturales(Dutoit y Alard 1995; McIvor yGardener 1994), los bancos desemillas que están en el suelo de losagroecosistemas actúan, a menudo,como factor de permanencia de lasmalezas (Benoit et al. 1992; Cavers1995). La evaluación del potencialde germinación de las semillas queestán en el suelo se ha utilizado,generalmente, como un indicador delestado de infestación con malezasque presenta un cultivo. Ahora bien,pese a la importancia económica delos bancos de semillas, se sabe muypoco sobre aquéllos que seencuentran en suelos dedicados a laproducción agrícola en el trópico(García 1995).
Los animales, así como lavariación espacio-temporal de lascondiciones ambientales, puedeninfluir en la supervivencia de lassemillas, en la duración de sulatencia y en su tasa de germinación(Mayer y Poljakoff-Mayber 1989). Seconsidera, además, que la actividadde los animales puede devolver lassemillas a la superficie del suelocontribuyendo así, posiblemente, a lacomposición de la vegetación epígea
(Willems y Huijsmans 1994). Laperturbación del suelo(pedoturbación) debida a losmacroinvertebrados grandes que lohabitan y se definen como“ingenieros del ecosistema” (sensuJones et al. 1994) —las termitas, lashormigas y las lombrices,principalmente— puede tener unimpacto significativo en la dinámicadel banco de semillas del suelo.
Las especies de lombricescatalogadas como anécicas (sensuBouché 1977) horadan el suelo,viven en galerías verticales osubverticales y se alimentan de lahojarasca en la superficie del suelo ode una mezcla de hojarasca y suelo.Los individuos de estas especiespueden ingerir también semillas, demanera selectiva, cuando consumensuelo u hojarasca. Por consiguiente,estas semillas pueden serdispersadas verticalmente por laacción de las lombrices, ya que sonexcretadas en sus heces, bien sea enel interior del suelo (por ejemplo,dentro de las galerías) o en lasuperficie del suelo (a la entrada dela galería) (Grant 1983; Reest yRogaar 1988; Thompson et al. 1994;Willems y Huijsmans 1994). Laslombrices de estas especiesparticipan, al enterrar esas semillas,en la formación de grandes bancosde semillas en los ecosistemas depastizal (Grant 1983). Durante eltránsito por el intestino de laslombrices, algunas semillas quedaninservibles, pero en las quesobreviven se estimula su potencialde germinación puesto quemejoraron para ellas las condicionesquímicas que encontraron en lasheces (McRill y Sagar 1973; vanTooren y During 1988).
El objetivo del presente estudiofue cuantificar el efecto que unaespecie de lombriz anécica grande(Martiodrilus carimaguensis Jiménezy Moreno, Glossoscolecidae,

308
El Arado Natural:…
Oligochaeta) ejercía en la dinámicadel banco de semillas del suelo en lasabana natural y en la sabanadedicada a la explotación agrícolaen los Llanos Orientales Colombia.Los individuos de la especieM. carimaguensis son los másgrandes recolectados hasta elmomento en el sitio del estudio y losúnicos que depositan cantidadessignificativas de heces en las galeríasy en la superficie del suelo. Seprestó atención especialmente altransporte vertical de las semillas, alos cambios producidos en elpotencial de germinación de éstas, ya las semejanzas entre el banco desemillas y la vegetación epígea. Sediscute, también, en este trabajo elsignificado ecológico de dichosprocesos respecto a la dispersiónvertical de las semillas, a lapermanencia de los bancos desemillas en el suelo y al ciclo querecorren, a la dinámica de lagerminación de las especiesconsideradas como malezas, y a laconservación de la diversidad vegetalen la sabana nativa.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua (4°37’ N,71°19’ O) del convenio CIAT-CORPOICA, perteneciente a launidad fitogeográfica de las sabanasisohipertérmicas bien drenadas delos Llanos Orientales de Colombia.El clima es de tipo subhúmedotropical, con un valor medio anual de26 °C para la temperatura y de2300 mm para la precipitación, y conuna época seca que va desdenoviembre hasta marzo (datos deCIAT). La topografía determina lavegetación nativa: hay sabanasabiertas en las zonas altas (“altos” y“planos”) y bosques de galería o
sabanas inundables en las zonasbajas (“bajos”). Los suelospertenecen al grupo de los Oxisoles(Tropeptic HaplustoxIsohipertérmico) en los altos y al delos Ultisoles (Ultic Aeric Plintaquox)en los bajos. Ambos grupos desuelos se caracterizan por suspropiedades físicas agronómicasfavorables (agregación, porosidad,retención de agua), por su acidezalta (pH (H2O) < 5), y por sufertilidad química baja (saturaciónde Al > 80%, CIC < 5 meq/100 g).
Parcelas de muestreoLas parcelas de muestreo estabanubicadas en un alto, sobre un Oxisolbien drenado. Las parcelas medían7200 m2 y estaban separadas unos100 m unas de otras; se asumió, portanto, que no había diferencia entreuna y otra respecto al potencial delluvia de semillas. Se eligieron trestratamientos que permitíancomparar la sabana nativa condiferentes sistemas agropastorilesque se hallaban en estudio en laestación experimental (Friesen et al.1995; Tergas et al. 1984):
1. Una parcela de sabana nativadonde crecían las especiesPaspalum pectinatum Nees,Axonopus purpusii (Mez.) Chase yTrachypogon vestitus Anders.; nofue pastoreada durante 4 años yse quemaba anualmente durantela época seca.
2. Un pastizal de 18 años de edaden que se asociaban las especiesBrachiaria decumbens Stapf. yPueraria phaseoloides Benth.(pastizal A); se hallaba bajopastoreo con una carga animalde 1 animal/ha durante la épocaseca y 2 animales/ha durante laépoca lluviosa.
3. Un pastizal de 3 años en que seasociaban las especies Brachiaria

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
309
humidicola (Rendle), Arachispintoi Krap. y Greg., Stylosanthescapitata Vog. y Centrosemaacutifolium Benth. (pastizal B);se hallaba bajo pastoreo conuna carga animal media de2 animales/ha.
Toma de muestras en el bancode semillasEl muestreo se realizó en la mitad dela época de lluvias (agosto de 1996).Las muestras correspondían, portanto, al banco de semillaspermanente del suelo, porque esprobable que la mayor parte delbanco de semillas temporal hayagerminado justo al inicio de la épocalluviosa. Se supuso que este método,tomado de la literatura actual,permitía detectar la mayoría de lasespecies en los ecosistemas depastizales tropicales, y tambiénevaluar la densidad de las semillasde las especies más comunes, por lomenos. En cada una de las tresparcelas experimentales se tomaronal azar 20 muestras de suelo, con uncilindro de 8 cm de diámetro y 6 cmde altura. Cuando se toma unnúmero de muestras tan pequeño,disminuye la precisión en ladeterminación de la densidad de lassemillas recolectadas (Bigwood yInouye 1988; Thompson 1986); sinembargo, las restricciones logísticas yde espacio y tiempo del invernaderofavorecieron la elección de esteprocedimiento. No obstante, lasuperficie y el volumen totalmuestreados (1005 cm2 y 6032 cm3,respectivamente) superaron losvalores mínimos recomendados parala recolección de la mayoría de lasespecies del banco de semillas de unpastizal (1000 cm2 y 1200 cm3,respectivamente) (Forcella 1984;Roberts 1981). En cada parcela serecolectaron, también al azar, 300heces frescas de M. carimaguensis,que se separaron en 20 grupos de
15 heces “contiguas”. El volumentotal de las heces recolectadas varióde 4000 cm3 a 10,000 cm3, según laparcela experimental estudiada.
Tanto las muestras de suelocomo las de heces se secaron al airedurante 15 días y después sepesaron. Cada muestra de suelo ode heces se depositó en una bandejaplástica (26 x 27 x 6 cm) quecontenía una capa de arena de río de2 cm de altura. Para que pudieragerminar la mayoría de las semillas,la capa de suelo colocada en lasbandejas era inferior a 5 mm (Dallinget al. 1994) y las heces se ubicaronrespetando la posición quepresentaban en el campo. Seutilizaron 40 bandejas, 20 para elsuelo y otras 20 para las heces; en10 bandejas más se incluyó arenasola que sirvió de testigo paraevaluar la posible contaminación desemillas de este sustrato. Lasbandejas de germinación secolocaron aleatoriamente en elinvernadero, se mantuvieronhúmedas y se expusieron acondiciones naturales de luz y detemperatura (aproximadamente,12 h/12 h de oscuridad/luz y 26 °C,respectivamente). Una vez porsemana, las plántulas germinadas seidentificaban hasta el nivel deespecie y se retiraban de la bandeja.Después de 3 meses, las hecesfueron reducidas a pequeñosfragmentos (< 5 mm de diámetro) yse colocaron de nuevo en lasbandejas para continuar laevaluación.
La evaluación y la comparaciónde la cantidad total de semillascontenidas en el suelo y en las hecesse realizó mediante un métododerivado de Malone (1967) (Benoit etal. 1989). Las tasas de germinaciónde las semillas provenientes tanto delsuelo como de las heces secalcularon a partir del número desemillas que germinaron en la

310
El Arado Natural:…
primera parte del experimento. Decada muestra se tomaron al azar 40 gde suelo o de heces secas al comienzodel experimento, que equivalían a20 submuestras de suelo o de hecesen cada sistema estudiado yrepresentaban del 5% al 10% de lamuestra total inicial. Cadasubmuestra se agitó durante unmínimo de 30 minutos en unasolución de hexametafosfato de sodio(50 g/l) y de bicarbonato de sodio(25 g/l) para dispersar todos losagregados del suelo. La suspensiónse tamizó después (en trama de0.125 mm) y los restos orgánicos selavaron con una fina aspersión deagua y se secaron durante 48 horas a70 °C. Una vez secas, las semillasdañadas y las que no recibieron daño(es decir, las que visualmenteparecían intactas y resistían unapresión leve) se separaron y secontaron bajo una lupa binocular. Nofue posible identificarlas porque lasreferencias respecto a las especies aque pertenecen las semillasencontradas en esta región del mundoes muy escasa.
Vegetación epígeaLa composición de la vegetaciónepígea se determinó en cada parcela apartir de 100 datos puntualestomados cada 20 cm a lo largo de untransecto de 20 m, cuyo origen ydirección se escogieron al azar. Laidentificación de las plantas se realizósegún los estudios de Escobar et al.(1993) y de Mejía (1984). Estos datosse emplearon para establecer lafrecuencia relativa de cada especiedentro de la vegetación. En total, seestudiaron cuatro transectos porparcela.
Producción de hecessuperficialesSe evaluó la producción individualdiaria de heces superficiales
depositadas por individuos deM. carimaguensis señalando, entodas la parcelas experimentales,40 galerías en las que se presumíaque la lombriz estaba presente, esdecir, cuando había una deposiciónfresca a la entrada de la galería. Lasheces superficiales se recolectarondiariamente durante 8 días y, antesde pesarlas, se secaron a 70 °C. Semidió la producción total de hecessuperficiales en g/m2 por año,partiendo de los datos existentes dedensidad poblacional deM. carimaguensis (Decaëns, datos sinpublicar; Jiménez et al. 1998a) yconsiderando que la época de plenaactividad de esta especie va desdemayo hasta octubre de cada año, esdecir, durante 180 días del año(Jiménez et al. 1998a); se obtuvo asíel siguiente resultado:
PA = N x PID x 180
donde PA = producción anual;PID = producción individual diaria;N = número de individuos por m2,que equivale al número de hecesfrescas por m2 (Jiménez et al. 1998a).
Se calculó la producción total deheces (superficiales y del interior delsuelo) en la sabana y en el pastizal Ausando la misma ecuación y losdatos existentes de producción deheces obtenidos por Rangel (1996) enun experimento de microcosmos.
Procesamiento de los datosSe usó el índice de similitud deSørensen (Cs) para comparar lacomposición del banco de semillasdel suelo, del banco de semillas delas heces y de la vegetación epígea,dentro de cada parcela experimentaly entre las parcelas (Sørensen 1948):
donde j es el número de especiescomunes a las dos muestras, y a yb representan el número total de
Cs = 2j(a + b)

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
311
especies registradas en cada muestraal comparar, respectivamente, lasituación a con la b.
La riqueza específica se definecomo el número total de especiesencontradas en una situación dada(por ejemplo, en las parcelasexperimentales, en el suelo o en lasheces). La diversidad (H) y laequitatividad (H’) se calcularon apartir del índice de diversidad deShannon-Wiener (Pielou 1966):
donde pi es la frecuencia de laespecie i
Estadística
Datos cuantitativosCon el fin de reducir la asimetría
de la distribución de los datos, éstosse transformaron mediante latransformación Box-Cox (Sokal yRohlf 1995). Para ello se empleó elprograma VerNorm 3.0 contenido enel paquete estadístico R package quedesarrollaron Legendre y Vaudor(1991).
Los análisis de varianza (ANOVA)de dos vías se realizaronconsiderando el “sistema” (de uso dela tierra) y el “tipo de muestra” (porejemplo, suelo o heces) como efectosfijos principales. El análisis incluíael número total de semillas (semillas/100 g de suelo o de heces), elnúmero total de semillas viables(semillas/100 g de suelo o de heces),la relación proporcional de semillasdañadas, la tasa de germinación y ladensidad del banco de semillasviables (semillas/m2). Se realizaroncinco análisis, cada uno de los cualescomprendía tres pruebas (dos efectosprincipales y una interacción). Se usóla corrección Bonferroni para pruebas
encajadas (Cooper 1968) con el finde controlar el error estadístico; losniveles de probabilidad 0.05, 0.01 y0.001 quedaron ajustados,respectivamente, así: 0.0033[= 0.05/(3x5)], 0.0007 [= 0.01/(3x5)]y 0.00007 [= 0.001/(3x5)]. Lasmedias se compararon aplicando laprueba de Fisher PLSD.
Se hicieron también análisis devarianza simples tomando el“sistema” como efecto principal ofuente de variación en las siguientessituaciones:
a. la densidad de heces frescas,la densidad total de las heces(heces/m2), la producciónindividual de heces(g/individuo por día) y laproducción global de heces(g/m2 por año);
b. la densidad de semillasviables respecto a las heces,el número de semillas viablesexcretadas en las heces(n/m2 por año) y elporcentaje del banco desemillas del suelo que esexcretado en las heces cadaaño.
Se usó la corrección deBonferroni para pruebas encajadas;los niveles de probabilidad 0.05,0.01 y 0.001 quedaron ajustados,respectivamente, así: en a., 0.0125,0.0250 y 0.00025, y en b., 0.0167,0.0033 y 0.0003.
Indices de diversidadEl procedimiento general
“jackknife” de primer orden (Tukey1958) se usó para calcular mejor lariqueza taxonómica (S) y ladiversidad (H) y generar asíintervalos de confianza que sirvieranpara comparar, respecto a esos dosparámetros, las comunidades deplántulas. No se hallaron
H = − pi log 2 pi
i = 1
n
∑
H'= H log 2S

312
El Arado Natural:…
referencias explícitas sobre el uso deesta prueba en la equitatividad (H’) yen el índice de Sørensen (Cs); porconsiguiente, no fue posible hacercomparaciones muy rigurosas. Alcomparar después estos índices, sólose interpretaron las diferencias másgrandes (por ejemplo > +0.20 entre elvalor más bajo y el más altocomparados).
El procedimiento “jackknife” sebasa en el cómputo de n valores delestadístico deseado mediante laexclusión sucesiva de una de las nmuestras. La media de estasvaloraciones se usa en la reduccióndel sesgo del estadístico y lavariabilidad existente entre estosvalores se emplea para estimar elerror estándar (Sokal y Rohlf 1995).Tukey (1958) conjeturó que lospseudo-valores (f ) obtenidos a partirdel procedimiento “jackknife” sonasintóticamente independientes ysiguen la distribución normal. Enconsecuencia, computamos unintervalo de confianza usando unatabla de distribución de t.
En el caso del índice dediversidad de Shannon-Wiener, lavaloración de y del errorestándar se calculó a partir delas siguientes ecuaciones (Adams yMcCune 1979; Heltshe y Forrester1985):
donde el pseudo-valor es:
Para la riqueza específica, lavaloración de y la del errorestándar se calcularon segúnlas siguientes ecuaciones (Heltshe yForrester 1983):
donde S es la riqueza específica detodo el conjunto de las muestras yK es el número correspondiente alas especies que se encontraronsolamente en una bandeja degerminación (“especie única”).
donde Fj es el número de muestrasen que estaba la especie única j.
Los cálculos se realizaron con elprograma informático “EstimateS 5”(Colwell 1997). En todas lasvaloraciones realizadas, los límitesde confianza al 95% son:
Valores calculados
donde t0.05(n-1) es la distribución de lat de Student para n – 1 grados delibertad.
ResultadosLa germinación de las semillas duró,en total, 4½ meses. Al final delexperimento no se observó ningúntipo de germinación en las bandejastestigo. La densidad de semillasviables por m2 fue significativamentemenor en el suelo de la sabananativa y en el pastizal B que enel pastizal A (Cuadro 19-1,Figura 19-1,A).
La producción individual deheces de M. carimaguensis fue muyconstante cuando se compararon lossistemas de uso de la tierra, aunqueel efecto del sistema en la densidadpoblacional afectó la producciónglobal de heces de toda la poblaciónde estas lombrices de tierra(Cuadros 19-2 y 19-3). La densidadde heces frescas y, en consecuencia,de individuos de M. carimaguensisfue mínima en la sabana y en elpastizal B, pero aumentósignificativamente en el pastizal A.
)ˆ( nHH)ˆ( nHσ
ˆ H n = φ ii =1
n
∑ n =φ
)ˆ(SS)ˆ( Sσ
nnKSS )1(ˆ −+=
(R
0j
2j
2 n1)(n)nKFjSS ∑ −−+==
σ
1)0.05(n−± txσ
φ i = nHn − (n − 1)Hn −i
σ ˆ H n = var2 n = (φ i − φ )2∑ n(n − 1)

Efectos C
ausados por las Lombrices de Tierra en el B
anco de Semillas...
313
Cuadro 19-1. Promedio de la densidad (semillas/m2) y de la aparición (%) de las semillas (según el taxón a que pertenecen) germinadas en el suelo y en las hecesde las lombrices, y porcentaje de aparición de las especies de vegetación epígea registradas en los tres tratamientos del estudio (Sem. = semillas;germ. = germinadas).
Especies Sabana nativa Pastizal A Pastizal BEn En En veget. En En En veget. En En En veget.
suelo: heces: epígea: suelo: heces: epígea: suelo: heces: epígea:Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparicióngerm. total germ. total (%) germ. total germ. total (%) germ. total germ. total (%)(no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%)
Amaranthus dubius - - 0.15 4.3 - 0.30 0.8 1.50 5.0 - 0.20 2.1 0.85 10.4 -Andropogon bicornis - - - - 4.5 - - - - 0.2 - - - - -Andropogon gayanus - - - - - 0.20 0.5 0.15 0.5 - - - - - -Andropogon - - - - 3.0 - - - - - - - - -
leucostachyusAndropogon selloanus - - - - 1.9 - - - - - - - - - -Arachis pintoi - - - - - - - - - - 0.10 1.1 - - 13.0Aristida capillacea - - 0.05 1.4 2.5 - - 0.10 0.3 - - - 0.05 0.6 -Aristida riparia - - - - 0.1 - - - - - - - - - -Aristida tincta - - - - 0.5 - - - - - - - - - -Axonopus aureus - - - - 0.7 - - - - - - - - - -Axonopus - - - - 0.1 - - - - - - - - - -
chrysoblepharisAxonopus compressus - - 0.45 12.9 - 0.05 0.1 1.15 3.8 - 0.30 3.2 1.65 20.1 0.2Axonopus purpusii - - - - 17.3 - - - - - - - - - 2.9Borreria sp. 1.25 9.6 0.15 4.3 2.1 3.95 10.3 0.70 2.3 0.5 0.75 8.0 0.15 1.8 -Brachiaria decumbens - - - - - 0.15 0.4 1.05 3.5 40.2 - - - - -Brachiaria humidicola - - - - - 0.05 0.1 0.10 0.3 - 0.35 3.7 0.20 2.4 74.7Cassearia ulmifolia - - - - 0.2 - - - - - - - - - -Chamacrista diphila - - - - - - - - - - - - - - 0.8Clidemia rubra - - - - 0.2 - - - - - - - - - -Clitoria guianensis - - - - 0.4 - - - - - - - - - -Croton trinitatis - - 0.15 4.3 - 0.10 0.3 1.90 6.3 1.6 - - 0.10 1.2 -Cyperus haspan - - - - - - - - - - - - - - 0.3Cypura paludosa - - - - 0.1 - - - - - - - - - -Declieuxia fruticosa - - - - 0.1 - - 0.15 0.5 - - - - - -Desmodium barbatum - - - - 0.1 - - - - - - - - - -Desmodium - - - - 0.2 0.25 0.7 0.65 2.2 11.3 - - - - 0.2
ovalifoliumDichromena ciliata 0.25 1.9 0.30 8.6 0.1 - - 0.20 0.7 - 0.10 1.1 - - 0.2Digitaria neesiana 1.00 7.7 0.15 4.3 - - - 0.90 3.0 - 0.20 2.1 0.20 2.4 -Elyonorus candidus - - - - 0.1 - - - - - - - - - -Emilia sonchifolia 0.60 4.6 - - 0.1 0.05 0.1 - - 0.1 0.20 2.1 0.05 0.6 -Fimbristylis complanata - - - - - - - - - - 0.05 0.5 - - -Galactia glaucescens - - - - - - - - - 0.2 - - - - -Gimnopogon foliosus 0.25 1.9 - - 1.0 - - 0.05 0.2 - - - - - -
(Continúa)
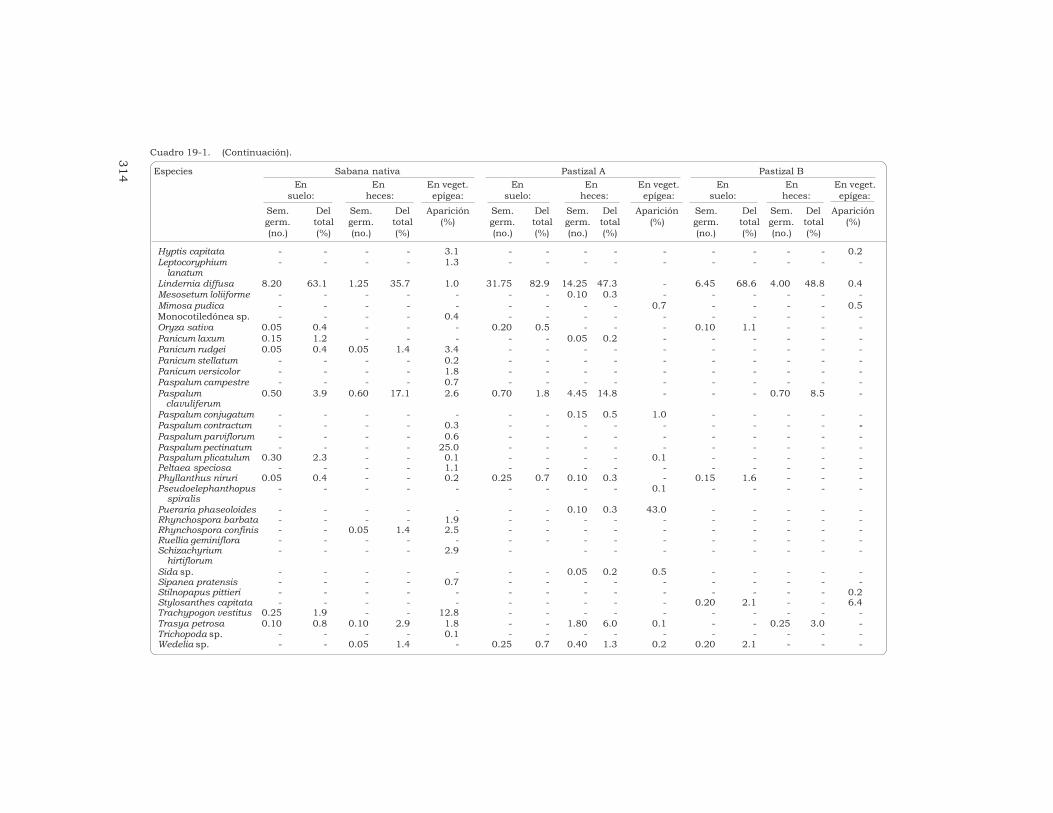
314
El A
rado Natural:…
Cuadro 19-1. (Continuación).
Especies Sabana nativa Pastizal A Pastizal BEn En En veget. En En En veget. En En En veget.
suelo: heces: epígea: suelo: heces: epígea: suelo: heces: epígea:Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparición Sem. Del Sem. Del Aparicióngerm. total germ. total (%) germ. total germ. total (%) germ. total germ. total (%)(no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%) (no.) (%)
Hyptis capitata - - - - 3.1 - - - - - - - - - 0.2Leptocoryphium - - - - 1.3 - - - - - - - - - -
lanatumLindernia diffusa 8.20 63.1 1.25 35.7 1.0 31.75 82.9 14.25 47.3 - 6.45 68.6 4.00 48.8 0.4Mesosetum loliiforme - - - - - - - 0.10 0.3 - - - - - -Mimosa pudica - - - - - - - - - 0.7 - - - - 0.5Monocotiledónea sp. - - - - 0.4 - - - - - - - - - -Oryza sativa 0.05 0.4 - - - 0.20 0.5 - - - 0.10 1.1 - - -Panicum laxum 0.15 1.2 - - - - - 0.05 0.2 - - - - - -Panicum rudgei 0.05 0.4 0.05 1.4 3.4 - - - - - - - - - -Panicum stellatum - - - - 0.2 - - - - - - - - - -Panicum versicolor - - - - 1.8 - - - - - - - - - -Paspalum campestre - - - - 0.7 - - - - - - - - - -Paspalum 0.50 3.9 0.60 17.1 2.6 0.70 1.8 4.45 14.8 - - - 0.70 8.5 - clavuliferumPaspalum conjugatum - - - - - - - 0.15 0.5 1.0 - - - - -Paspalum contractum - - - - 0.3 - - - - - - - - - -Paspalum parviflorum - - - - 0.6 - - - - - - - - - -Paspalum pectinatum - - - - 25.0 - - - - - - - - - -Paspalum plicatulum 0.30 2.3 - - 0.1 - - - - 0.1 - - - - -Peltaea speciosa - - - - 1.1 - - - - - - - - - -Phyllanthus niruri 0.05 0.4 - - 0.2 0.25 0.7 0.10 0.3 - 0.15 1.6 - - -Pseudoelephanthopus - - - - - - - - - 0.1 - - - - -
spiralisPueraria phaseoloides - - - - - - - 0.10 0.3 43.0 - - - - -Rhynchospora barbata - - - - 1.9 - - - - - - - - - -Rhynchospora confinis - - 0.05 1.4 2.5 - - - - - - - - - -Ruellia geminiflora - - - - - - - - - - - - - - -Schizachyrium - - - - 2.9 - - - - - - - - -
hirtiflorumSida sp. - - - - - - - 0.05 0.2 0.5 - - - - -Sipanea pratensis - - - - 0.7 - - - - - - - - - -Stilnopapus pittieri - - - - - - - - - - - - - - 0.2Stylosanthes capitata - - - - - - - - - - 0.20 2.1 - - 6.4Trachypogon vestitus 0.25 1.9 - - 12.8 - - - - - - - - - -Trasya petrosa 0.10 0.8 0.10 2.9 1.8 - - 1.80 6.0 0.1 - - 0.25 3.0 -Trichopoda sp. - - - - 0.1 - - - - - - - - - -Wedelia sp. - - 0.05 1.4 - 0.25 0.7 0.40 1.3 0.2 0.20 2.1 - - -
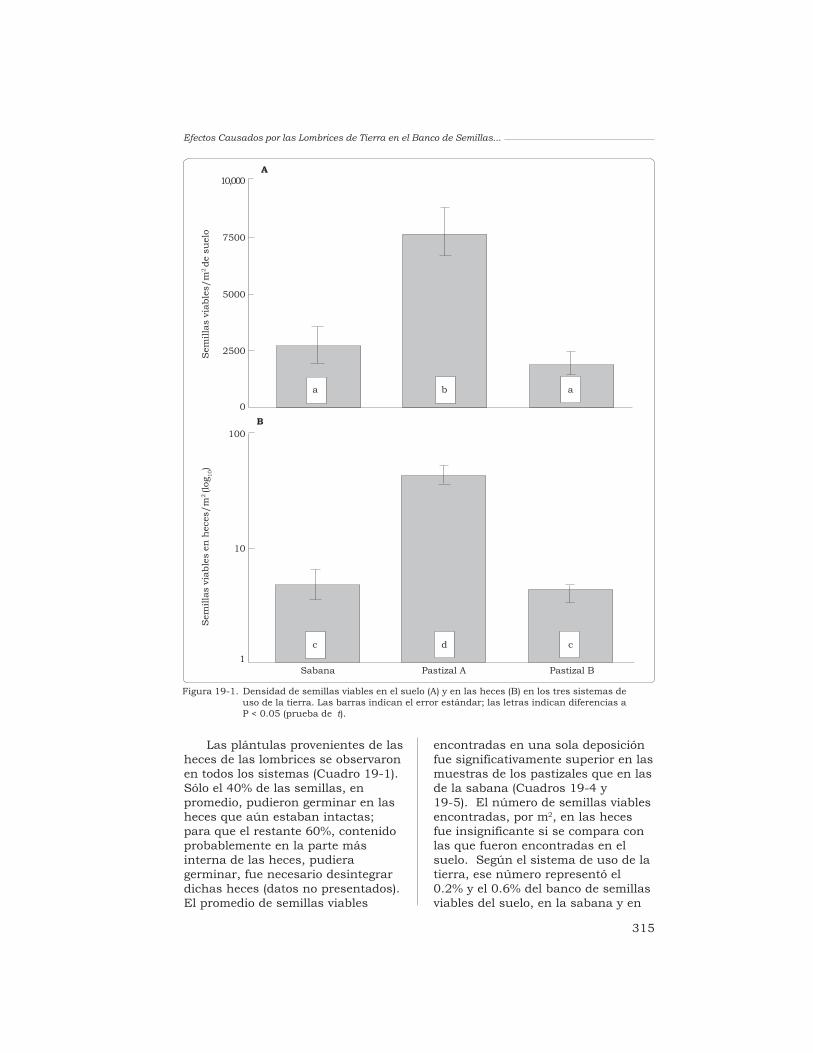
Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
315
Las plántulas provenientes de lasheces de las lombrices se observaronen todos los sistemas (Cuadro 19-1).Sólo el 40% de las semillas, enpromedio, pudieron germinar en lasheces que aún estaban intactas;para que el restante 60%, contenidoprobablemente en la parte másinterna de las heces, pudieragerminar, fue necesario desintegrardichas heces (datos no presentados).El promedio de semillas viables
encontradas en una sola deposiciónfue significativamente superior en lasmuestras de los pastizales que en lasde la sabana (Cuadros 19-4 y19-5). El número de semillas viablesencontradas, por m2, en las hecesfue insignificante si se compara conlas que fueron encontradas en elsuelo. Según el sistema de uso de latierra, ese número representó el0.2% y el 0.6% del banco de semillasviables del suelo, en la sabana y en
10,000
7500
5000
2500
0
Sem
illas
via
bles
en
hec
es/m
2 (lo
g 10)
Sem
illas
via
bles
/m2 de
su
elo
100
10
1Sabana Pastizal A Pastizal B
a b a
c d c
A
B
Figura 19-1. Densidad de semillas viables en el suelo (A) y en las heces (B) en los tres sistemas deuso de la tierra. Las barras indican el error estándar; las letras indican diferencias aP < 0.05 (prueba de t).
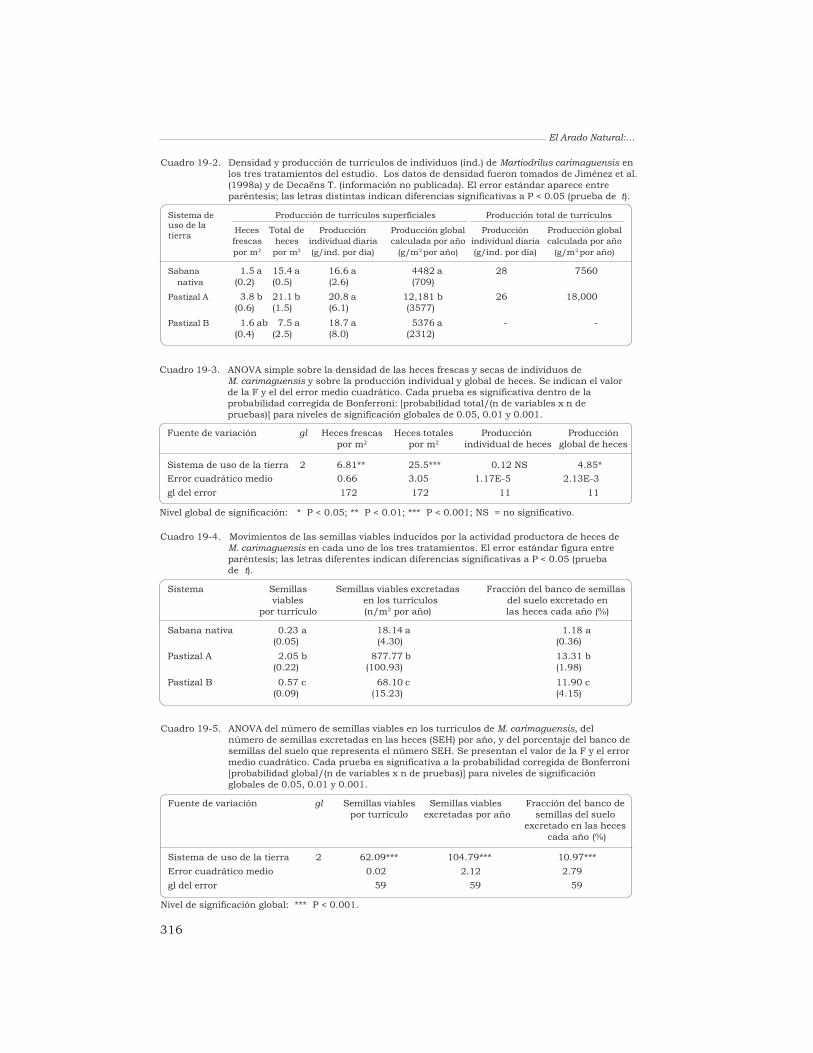
316
El Arado Natural:…
Cuadro 19-3. ANOVA simple sobre la densidad de las heces frescas y secas de individuos deM. carimaguensis y sobre la producción individual y global de heces. Se indican el valorde la F y el del error medio cuadrático. Cada prueba es significativa dentro de laprobabilidad corregida de Bonferroni: [probabilidad total/(n de variables x n depruebas)] para niveles de significación globales de 0.05, 0.01 y 0.001.
Fuente de variación gl Heces frescas Heces totales Producción Producciónpor m2 por m2 individual de heces global de heces
Sistema de uso de la tierra 2 6.81** 25.5*** 0.12 NS 4.85*Error cuadrático medio 0.66 3.05 1.17E-5 2.13E-3gl del error 172 172 11 11
Nivel global de significación: * P < 0.05; ** P < 0.01; *** P < 0.001; NS = no significativo.
Cuadro 19-4. Movimientos de las semillas viables inducidos por la actividad productora de heces deM. carimaguensis en cada uno de los tres tratamientos. El error estándar figura entreparéntesis; las letras diferentes indican diferencias significativas a P < 0.05 (pruebade t).
Sistema Semillas Semillas viables excretadas Fracción del banco de semillasviables en los turrículos del suelo excretado en
por turrículo (n/m2 por año) las heces cada año (%)
Sabana nativa 0.23 a 18.14 a 1.18 a(0.05) (4.30) (0.36)
Pastizal A 2.05 b 877.77 b 13.31 b(0.22) (100.93) (1.98)
Pastizal B 0.57 c 68.10 c 11.90 c(0.09) (15.23) (4.15)
Cuadro 19-5. ANOVA del número de semillas viables en los turrículos de M. carimaguensis, delnúmero de semillas excretadas en las heces (SEH) por año, y del porcentaje del banco desemillas del suelo que representa el número SEH. Se presentan el valor de la F y el errormedio cuadrático. Cada prueba es significativa a la probabilidad corregida de Bonferroni[probabilidad global/(n de variables x n de pruebas)] para niveles de significaciónglobales de 0.05, 0.01 y 0.001.
Fuente de variación gl Semillas viables Semillas viables Fracción del banco depor turrículo excretadas por año semillas del suelo
excretado en las heces cada año (%)
Sistema de uso de la tierra 2 62.09*** 104.79*** 10.97***Error cuadrático medio 0.02 2.12 2.79gl del error 59 59 59
Nivel de significación global: *** P < 0.001.
Cuadro 19-2. Densidad y producción de turrículos de individuos (ind.) de Martiodrilus carimaguensis enlos tres tratamientos del estudio. Los datos de densidad fueron tomados de Jiménez et al.(1998a) y de Decaëns T. (información no publicada). El error estándar aparece entreparéntesis; las letras distintas indican diferencias significativas a P < 0.05 (prueba de t).
Sistema de Producción de turrículos superficiales Producción total de turrículos
Heces Total de Producción Producción global Producción Producción globalfrescas heces individual diaria calculada por año individual diaria calculada por añopor m2 por m2 (g/ind. por día) (g/m2 por año) (g/ind. por día) (g/m2 por año)
Sabana 1.5 a 15.4 a 16.6 a 4482 a 28 7560nativa (0.2) (0.5) (2.6) (709)
Pastizal A 3.8 b 21.1 b 20.8 a 12,181 b 26 18,000(0.6) (1.5) (6.1) (3577)
Pastizal B 1.6 ab 7.5 a 18.7 a 5376 a - -(0.4) (2.5) (8.0) (2312)
uso de latierra
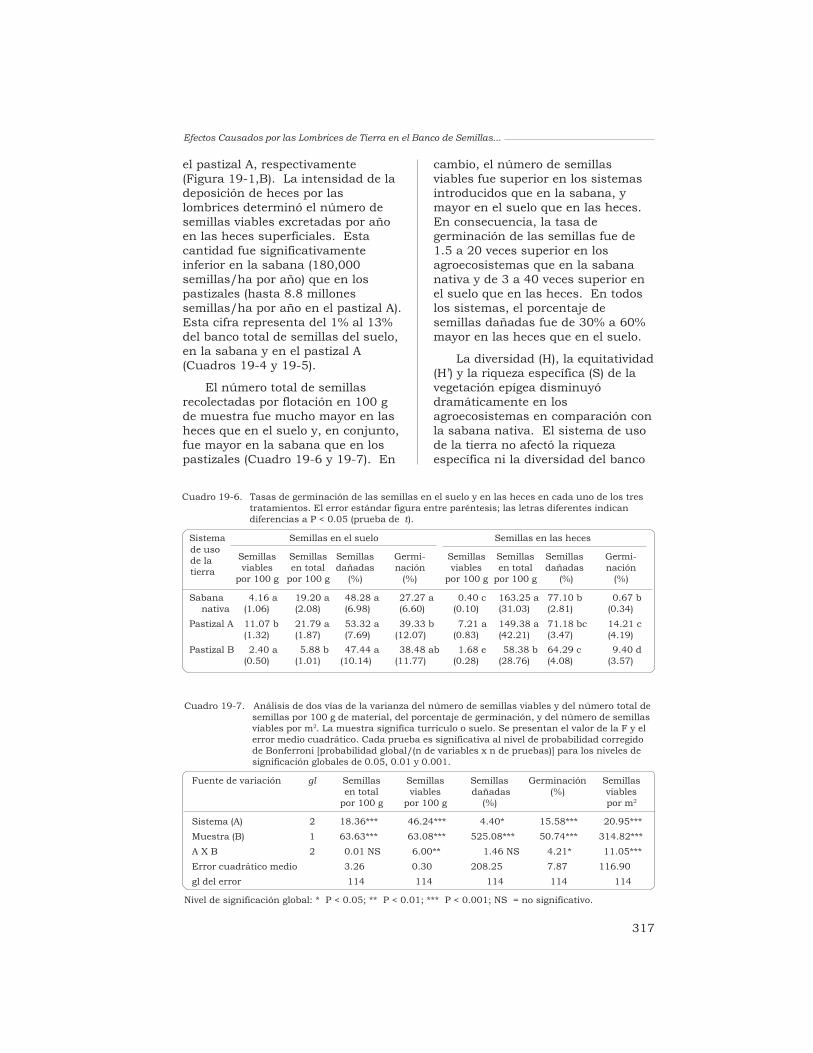
Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
317
el pastizal A, respectivamente(Figura 19-1,B). La intensidad de ladeposición de heces por laslombrices determinó el número desemillas viables excretadas por añoen las heces superficiales. Estacantidad fue significativamenteinferior en la sabana (180,000semillas/ha por año) que en lospastizales (hasta 8.8 millonessemillas/ha por año en el pastizal A).Esta cifra representa del 1% al 13%del banco total de semillas del suelo,en la sabana y en el pastizal A(Cuadros 19-4 y 19-5).
El número total de semillasrecolectadas por flotación en 100 gde muestra fue mucho mayor en lasheces que en el suelo y, en conjunto,fue mayor en la sabana que en lospastizales (Cuadro 19-6 y 19-7). En
cambio, el número de semillasviables fue superior en los sistemasintroducidos que en la sabana, ymayor en el suelo que en las heces.En consecuencia, la tasa degerminación de las semillas fue de1.5 a 20 veces superior en losagroecosistemas que en la sabananativa y de 3 a 40 veces superior enel suelo que en las heces. En todoslos sistemas, el porcentaje desemillas dañadas fue de 30% a 60%mayor en las heces que en el suelo.
La diversidad (H), la equitatividad(H’) y la riqueza específica (S) de lavegetación epígea disminuyódramáticamente en losagroecosistemas en comparación conla sabana nativa. El sistema de usode la tierra no afectó la riquezaespecífica ni la diversidad del banco
Cuadro 19-7. Análisis de dos vías de la varianza del número de semillas viables y del número total desemillas por 100 g de material, del porcentaje de germinación, y del número de semillasviables por m2. La muestra significa turrículo o suelo. Se presentan el valor de la F y elerror medio cuadrático. Cada prueba es significativa al nivel de probabilidad corregidode Bonferroni [probabilidad global/(n de variables x n de pruebas)] para los niveles designificación globales de 0.05, 0.01 y 0.001.
Fuente de variación gl Semillas Semillas Semillas Germinación Semillasen total viables dañadas (%) viables
por 100 g por 100 g (%) por m2
Sistema (A) 2 18.36*** 46.24*** 4.40* 15.58*** 20.95***Muestra (B) 1 63.63*** 63.08*** 525.08*** 50.74*** 314.82***A X B 2 0.01 NS 6.00** 1.46 NS 4.21* 11.05***Error cuadrático medio 3.26 0.30 208.25 7.87 116.90gl del error 114 114 114 114 114
Nivel de significación global: * P < 0.05; ** P < 0.01; *** P < 0.001; NS = no significativo.
Cuadro 19-6. Tasas de germinación de las semillas en el suelo y en las heces en cada uno de los trestratamientos. El error estándar figura entre paréntesis; las letras diferentes indicandiferencias a P < 0.05 (prueba de t).
Sistema Semillas en el suelo Semillas en las heces
Semillas Semillas Semillas Germi- Semillas Semillas Semillas Germi-viables en total dañadas nación viables en total dañadas nación
por 100 g por 100 g (%) (%) por 100 g por 100 g (%) (%)
Sabana 4.16 a 19.20 a 48.28 a 27.27 a 0.40 c 163.25 a 77.10 b 0.67 b nativa (1.06) (2.08) (6.98) (6.60) (0.10) (31.03) (2.81) (0.34)Pastizal A 11.07 b 21.79 a 53.32 a 39.33 b 7.21 a 149.38 a 71.18 bc 14.21 c
(1.32) (1.87) (7.69) (12.07) (0.83) (42.21) (3.47) (4.19)Pastizal B 2.40 a 5.88 b 47.44 a 38.48 ab 1.68 e 58.38 b 64.29 c 9.40 d
(0.50) (1.01) (10.14) (11.77) (0.28) (28.76) (4.08) (3.57)
de usode latierra
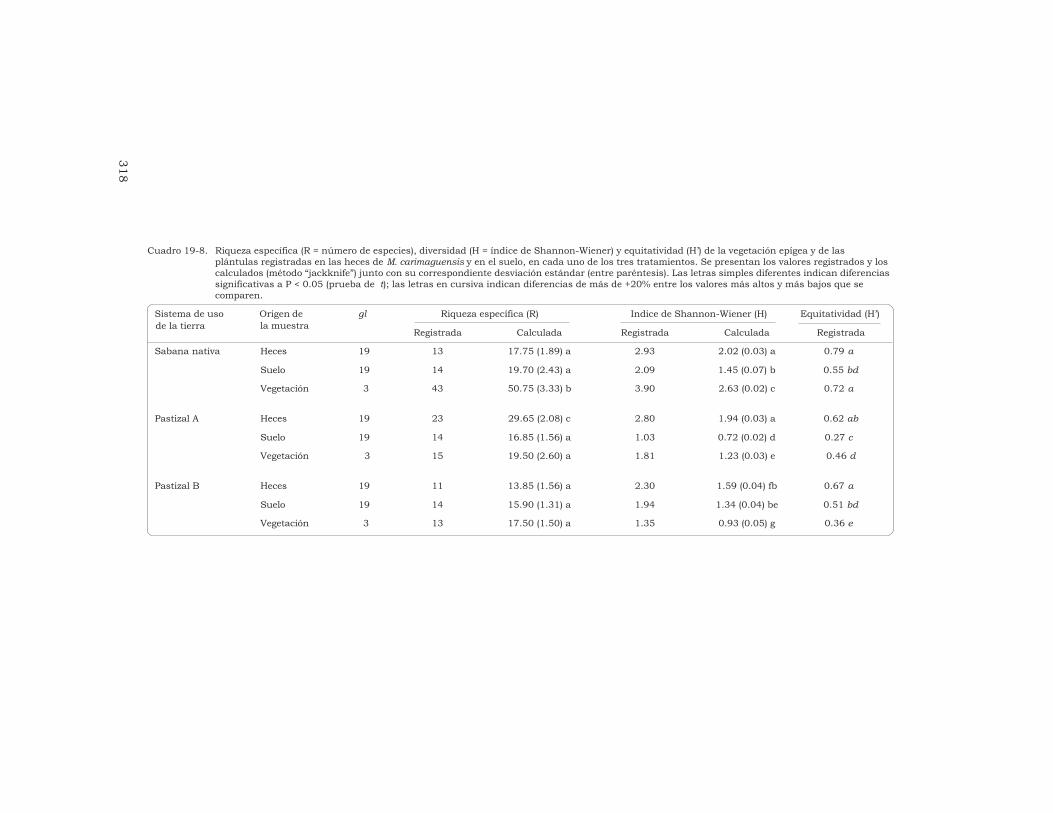
318
El A
rado Natural:…
Cuadro 19-8. Riqueza específica (R = número de especies), diversidad (H = índice de Shannon-Wiener) y equitatividad (H’) de la vegetación epígea y de lasplántulas registradas en las heces de M. carimaguensis y en el suelo, en cada uno de los tres tratamientos. Se presentan los valores registrados y loscalculados (método “jackknife”) junto con su correspondiente desviación estándar (entre paréntesis). Las letras simples diferentes indican diferenciassignificativas a P < 0.05 (prueba de t); las letras en cursiva indican diferencias de más de +20% entre los valores más altos y más bajos que secomparen.
Sistema de uso Origen de gl Riqueza específica (R) Indice de Shannon-Wiener (H) Equitatividad (H’)
Registrada Calculada Registrada Calculada Registrada
Sabana nativa Heces 19 13 17.75 (1.89) a 2.93 2.02 (0.03) a 0.79 a
Suelo 19 14 19.70 (2.43) a 2.09 1.45 (0.07) b 0.55 bd
Vegetación 3 43 50.75 (3.33) b 3.90 2.63 (0.02) c 0.72 a
Pastizal A Heces 19 23 29.65 (2.08) c 2.80 1.94 (0.03) a 0.62 ab
Suelo 19 14 16.85 (1.56) a 1.03 0.72 (0.02) d 0.27 c
Vegetación 3 15 19.50 (2.60) a 1.81 1.23 (0.03) e 0.46 d
Pastizal B Heces 19 11 13.85 (1.56) a 2.30 1.59 (0.04) fb 0.67 a
Suelo 19 14 15.90 (1.31) a 1.94 1.34 (0.04) be 0.51 bd
Vegetación 3 13 17.50 (1.50) a 1.35 0.93 (0.05) g 0.36 e
de la tierra la muestra
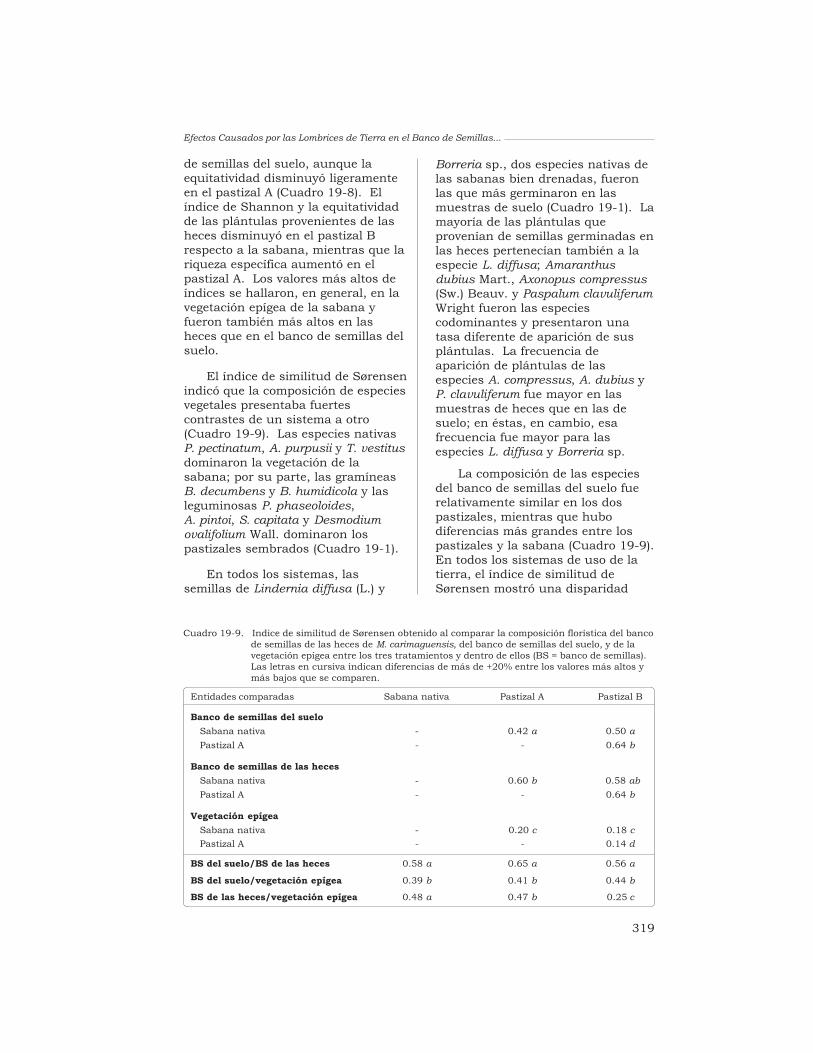
Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
319
de semillas del suelo, aunque laequitatividad disminuyó ligeramenteen el pastizal A (Cuadro 19-8). Elíndice de Shannon y la equitatividadde las plántulas provenientes de lasheces disminuyó en el pastizal Brespecto a la sabana, mientras que lariqueza específica aumentó en elpastizal A. Los valores más altos deíndices se hallaron, en general, en lavegetación epígea de la sabana yfueron también más altos en lasheces que en el banco de semillas delsuelo.
El índice de similitud de Sørensenindicó que la composición de especiesvegetales presentaba fuertescontrastes de un sistema a otro(Cuadro 19-9). Las especies nativasP. pectinatum, A. purpusii y T. vestitusdominaron la vegetación de lasabana; por su parte, las gramíneasB. decumbens y B. humidicola y lasleguminosas P. phaseoloides,A. pintoi, S. capitata y Desmodiumovalifolium Wall. dominaron lospastizales sembrados (Cuadro 19-1).
En todos los sistemas, lassemillas de Lindernia diffusa (L.) y
Borreria sp., dos especies nativas delas sabanas bien drenadas, fueronlas que más germinaron en lasmuestras de suelo (Cuadro 19-1). Lamayoría de las plántulas queprovenían de semillas germinadas enlas heces pertenecían también a laespecie L. diffusa; Amaranthusdubius Mart., Axonopus compressus(Sw.) Beauv. y Paspalum clavuliferumWright fueron las especiescodominantes y presentaron unatasa diferente de aparición de susplántulas. La frecuencia deaparición de plántulas de lasespecies A. compressus, A. dubius yP. clavuliferum fue mayor en lasmuestras de heces que en las desuelo; en éstas, en cambio, esafrecuencia fue mayor para lasespecies L. diffusa y Borreria sp.
La composición de las especiesdel banco de semillas del suelo fuerelativamente similar en los dospastizales, mientras que hubodiferencias más grandes entre lospastizales y la sabana (Cuadro 19-9).En todos los sistemas de uso de latierra, el índice de similitud deSørensen mostró una disparidad
Cuadro 19-9. Indice de similitud de Sørensen obtenido al comparar la composición florística del bancode semillas de las heces de M. carimaguensis, del banco de semillas del suelo, y de lavegetación epígea entre los tres tratamientos y dentro de ellos (BS = banco de semillas).Las letras en cursiva indican diferencias de más de +20% entre los valores más altos ymás bajos que se comparen.
Entidades comparadas Sabana nativa Pastizal A Pastizal B
Banco de semillas del sueloSabana nativa - 0.42 a 0.50 aPastizal A - - 0.64 b
Banco de semillas de las hecesSabana nativa - 0.60 b 0.58 abPastizal A - - 0.64 b
Vegetación epígeaSabana nativa - 0.20 c 0.18 cPastizal A - - 0.14 d
BS del suelo/BS de las heces 0.58 a 0.65 a 0.56 a
BS del suelo/vegetación epígea 0.39 b 0.41 b 0.44 b
BS de las heces/vegetación epígea 0.48 a 0.47 b 0.25 c

320
El Arado Natural:…
importante entre la composición delas especies del banco de semillas delsuelo y la vegetación epígea. Lasprincipales especies dominantes deesa vegetación no se encontraron en elexperimento realizado en elinvernadero y, en cambio, las especiesdominantes entre las plántulasprovenientes de semillas germinadasen el suelo estaban ausentes o eranraras en la comunidad vegetal(Cuadro 19-1). La composición deespecies entre las plántulas desemillas germinadas en las heces fuemuy constante en todos los sistemasde uso de la tierra, y más o menossimilar a la observada en las semillasprovenientes del suelo (Cuadro 19-9).En la sabana nativa, la composicióndel banco de semillas de las hecesestuvo ligeramente más próxima a lade la vegetación epígea que a la delbanco de semillas del suelo, mientrasque en el pastizal B se observó locontrario.
La lista completa de familias,géneros y especies y de su presencia(+) o ausencia (–) en las heces, en elsuelo y en la vegetación epígea,considerando los cinco sistemas comoun todo, se presenta en el Apéndice(p. 329).
Discusión
Efectos del sistema de uso de latierra en el banco de semillasdel sueloEl número de semillas germinadas enlos agroecosistemas (de 1860 hasta7600 semillas/m2) obtenido en esteestudio está entre los más bajos quese han registrado en los pastizalestropicales (de 11,000 hasta 36,000semillas/m2) (Kellman 1974, 1978;McIvor 1987). De los resultados deeste estudio se desprende que, si seusan los pastizales durante muchotiempo, aumenta considerablemente
la densidad de semillas en el suelo.Se sabe además que las condicionesambientales que caracterizan elestablecimiento de los pastizales sonidóneas para muchas especies demalezas, las cuales producengrandes cantidades de semillas delarga viabilidad y contribuyen así alestablecimiento de bancos desemillas importantes en el suelo(Thompson y Grime 1979). Lapersistencia de estas semillas en elsuelo, incluso tras un largo períodode uso del pastizal, confirma lalongevidad sorprendente de estospropágulos y su habilidad parapermanecer en los pastizales duranteperíodos largos de tiempo(Chancellor 1986; Dutoit y Alard1995; Thompson et al. 1994). Másaún, las malezas que se presenten,aun de modo esporádico, en elpastizal que esté en uso, puedenhaber participado en la formación deun banco de semillas permanente, deimportancia relativa, en el pastizalmás antiguo. Las diferenciasencontradas entre los bancos desemillas viables de los dos pastizalesse pueden interpretar como unresultado tanto de la edad de estossistemas pastoriles como del manejode las malezas durante elestablecimiento del pastizal odespués de él.
El gran aumento queexperimentó el banco de semillas delsuelo tras la aplicación de unsistema de uso de la tierra coincidecon el descubrimiento de Thompsony Grime (1979), quienes demostraronque la importancia de los bancos desemillas se relaciona positivamentecon su perturbación (por ejemplo,con la destrucción de la vegetaciónnativa) y negativamente con el estrésambiental (por ejemplo, con laslimitantes del crecimiento). En elcaso presente, la acidez del suelo, latoxicidad debida al aluminio, y lossuelos con un contenido bajo de

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
321
nutrientes son los tipos másfrecuentes de estrés ambiental. Enlos pastizales, la fertilización, elencalado o los efectos positivos de lapresencia de una leguminosa en laspropiedades químicas del suelo,reducen estas limitantes, mientrasque las perturbaciones las aumentan(al incrementar la carga animal ydestruir la vegetación nativa). Entales casos, si las condicionesambientales son aptas para lagerminación de las semillas —porejemplo, cuando el sobrepastoreoincrementa el área de suelodesprovisto de vegetación— el riesgode una invasión rápida de malezases más grande.
La ausencia de una correlaciónsignificativa entre la composiciónespecífica del banco de semillas delsuelo y de la vegetación epígeaconcuerda con los resultadosobtenidos en otros estudios (Dutoit yAlard 1995; Kellman 1974, 1978;Roberts 1986). Algunos factoresecológicos que actúan sobre lavegetación pueden explicar estosresultados (Harper 1977). Porejemplo, la reproducción vegetativa,la quema de la sabana, la dispersiónde las semillas por el viento o por losanimales, pueden afectar los bancosde semillas y la flora epígea de modoespecífico (Dutoit y Alard 1995;Mayer y Poljakoff-Mayber 1989;Thompson y Grime 1979). Laactividad humana relacionada conlos agroecosistemas añade nuevosfactores, como la presión delpastoreo sobre la vegetación o elimpacto de las prácticas agrícolas enlos ciclos de vida de las especies demalezas (Dutoit y Alard 1995; García1995; Milberg 1992). La similitudentre el banco de semillas del suelo yla vegetación epígea fue mayor en elpastizal más antiguo que en los otrossistemas; esto se debe,probablemente, a una mayorpresencia de suelo desnudo que
crea, a su vez, condiciones idóneaspara la germinación de malezasoportunistas (García 1995).
Efectos de las lombrices detierra en los bancos desemillasLos resultados obtenidos en esteestudio confirman el complejo papeldesempeñado por las lombricestanto en el proceso de formación delbanco de semillas como en lacontribución a la vegetación epígea(Willems y Huijsmans 1994). En lasabana nativa, la densidadpoblacional de M. carimaguensis esrelativamente baja y representa sóloel 15% de la biomasa total delombrices. Los efectos de laslombrices en el banco de semillas sehan subestimado porque la actividadde las dos especies dominantes:Andiodrilus sp. (24% de la biomasa)y Glossodrilus sp. (54% de labiomasa) (Jiménez et al. 1998b) nose consideraron en este estudio. Encambio, los cálculos hechos en elpastizal, donde la población de M.carimaguensis representa el 86% dela biomasa total de lombrices(Jiménez et al. 1998a, 1998b), seacercan bastante a la realidad. Elnúmero de semillas viablesexcretadas anualmente en las hecessuperficiales de M. carimaguensis, enestos sistemas de uso de la tierra, esrealmente considerable (hasta8.7 millones de semillas porhectárea al año), si lo comparamoscon el obtenido en la especieLumbricus terrestris L. en latitudestempladas (890 mil semillas porhectárea al año, en promedio)(Willems y Huijsmans 1994).
Muchas especies animales(conejos, vacas, etc.) ingieren lassemillas y modifican por ello su tasade germinación (Jones et al. 1991;Mayer y Poljakoff-Mayer 1989). Eneste estudio, las semillas incluidas

322
El Arado Natural:…
dentro de las heces perdían de un64% a un 97% de su viabilidad, loque puede atribuirse al daño sufridopor ellas durante su tránsito por elintestino de M. carimaguensis; así losugiere el porcentaje relativamentealto de semillas dañadas que seregistró en las heces. Otros estudioshan demostrado el efecto negativoque causa en la germinación de lassemillas su tránsito por el intestinode las lombrices, a causa de ladigestión, la asimilación y laescarificación de parte de lassemillas ingeridas (McRill y Sagar1973; van Tooren y During 1988).Se sabe, además, que las hecessuperficiales producidas por lasespecies de individuos grandes estánformadas por agregados estables,relativamente ricos en amonio ybastante impermeables al agua y alos gases (Blanchart et al. 1993;Rangel et al. 1999), condiciones quefavorecen el mantenimiento de lalatencia de la semilla o el retraso desu germinación (Grant 1983).
Otros estudios han señalado larelación positiva existente entre unapoblación de lombrices y la densidaddel banco de semillas del suelo en lospastizales (Grant 1983; Reest yRogaar 1988). Se cree que laslombrices de la especie M.carimaguensis son anécicas, es decir,que ingieren una mezcla de suelo yde hojarasca, sustratos ambos quecontienen cantidades importantes desemillas viables. Esta especie delombriz deposita heces sobre lasuperficie del suelo, aunque tambiénexcreta cantidades considerables deheces bajo esa superficie. Si secomparan los datos obtenidosanteriormente sobre la produccióntotal de heces de los individuos deM. carimaguensis (Rangel 1996) conlos datos de la producción de hecessuperficiales obtenidos en esteestudio (Cuadro 19-3), se estima queel 34% y el 68% de la cantidad totalde heces se producen en el interior
del suelo de la sabana y del pastizalA, respectivamente. Estos datosindican que una gran parte de lassemillas que sobreviven al tránsitointestinal es excretada en el interiorde las galerías de las lombrices,hasta los primeros 20 cm deprofundidad, principalmente,aunque a veces se encuentran hastalos 80 cm (Jiménez et al. 1998a). Laacción que desarrollan las lombricesenterrando las semillas puede tenerconsecuencias importantes en ladinámica de la comunidad vegetal,porque las semillas de muchasespecies vegetales sobreviven mejorsi están enterradas que si estánsobre la superficie del suelo; además,hay pocos mecanismos naturalesque expliquen la forma en que lassemillas se entierran en el suelo(Grant 1983). Por otra parte, lasheces producidas por las lombricespueden proteger las semillas de unagerminación temprana y participan,así, en la formación de bancos desemillas persistentes en el suelo,cuya presencia es importante.
Otra parte de las semillas esexcretada por las lombrices en lasuperficie del suelo. Cuando lassemillas enterradas sontransportadas hacia la superficie delsuelo, aumentan sus oportunidadesde germinación (Grant 1983; Reest yRogaar 1988; Willems y Huijsmans1994). Se ha demostrado, sinembargo, que gran parte de estassemillas (59%) se encuentran en elinterior de las heces y éstas debenfragmentarse primero para que lassemillas germinen. La fragmentaciónde las heces en la superficie del sueloes el resultado de una combinaciónde factores —por ejemplo, lasprecipitaciones fuertes, el fuego, elpisoteo del ganado— y ocurreprobablemente a diferentes tasas quedependen del sistema de uso de latierra y de la cubierta vegetal delsuelo (Decaëns 2000). El 41%restante puede germinar en las heces

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
323
completas y su supervivenciadependerá de la calidad de estesoporte físico. Las condicionesfavorables para la germinación deestas semillas están relacionadas,por tanto, con los factores externosque intervienen en la durabilidad delas heces (por ejemplo, el fuego, lalluvia, la escorrentía y el pisoteo delganado).
El comportamiento selectivo yactivo que manifiestan los individuosde M. carimaguensis al ingerir lassemillas puede explicar lasdiferencias encontradas entre lacomposición específica de plántulasprovenientes del suelo o de las heces.Esta característica se ha observadotambién en lombrices de latitudestempladas y está ligada al tamaño dela semilla y a su ‘gustosidad’, esdecir, a la percepción agradable(estímulo positivo enquimiorreceptores de la lombriz) quetienen las semillas para las lombrices(Grant 1983; Willems y Huijsmans1994). Las semillas pequeñas y‘gustosas’ (o ‘palatables’) soningeridas primero que las demás ytienen, por ello, mayor oportunidadde estar presentes en las heces. Eneste estudio fue imposible verificaresta relación porque no se disponeen la literatura de descripciones delas semillas de las especies vegetalesque crecen en las sabanascolombianas. Otra consecuencia deesta actitud selectiva sería lahomogeneidad relativa del banco desemillas de las heces en todos lossistemas de uso de la tierra, a pesarde que éstos modificanprofundamente la comunidad vegetaly el banco de semillas del suelo.Puesto que la mayoría de lasplántulas provenientes de las hecespertenecían a especies de la sabana,la actividad que ejercen las lombricesde tierra en el banco de semillaspuede considerarse como un factorde inercia que actúa frente a lasperturbaciones antrópicas. Para
probar esta hipótesis, hay queinvestigar los efectos que ejercen lascomunidades de lombrices en lasucesión vegetal secundaria, tras elcese de la actividad humana.
Las diferencias de riquezaespecífica y de diversidadencontradas entre las plántulasprovenientes de las heces y las delsuelo que circunda las heces es,ciertamente, uno de los rasgos másrelevantes de este estudio; es difícil,sin embargo, explicar esasdiferencias. Parece contradictorioque haya en la sabana y en elpastizal A un número mayor detaxones en las heces, ya que laingestión selectiva de semillas por laslombrices debería favorecer laausencia de especies desechadas, porejemplo semillas poco ‘ gustosas’ o detamaño grande (o de ambascaracterísticas) (Grant 1983; Willemsy Huijsmans 1994). Estecomportamiento debería, además,reducir la diversidad, ya que laslombrices seleccionarían las especiesmás atractivas que serían, por tanto,las dominantes; los datosexperimentales muestran, en cambio,el efecto opuesto. Ahora bien, lainterpretación que puede darse sebasa en las mejores condiciones queofrecen las heces para que lassemillas germinen, lo que aumenta laequitatividad de las especies ypermite su germinación por igual.Esta hipótesis podría probarse si seobserva que las especies faltantesestán en el suelo pero no germinaron.La prueba no pudo realizarse duranteeste estudio por falta de literaturarelevante para identificar las semillashasta nivel de especie. Lagerminación de algunas semillas, porsu parte, dependerá de queabandonen su período de latencia, loque ocurre tras la escarificación queles producen ya sea las contraccioneso ya la actividad enzimática del tractodigestivo de las lombrices (Grant1983).

324
El Arado Natural:…
En la sabana protegida de laquema, las heces permanecen aveces en la superficie del suelodurante más de un año; la quema,en cambio, lleva súbitamente a sudestrucción porque elimina el efectoprotector de la cubierta vegetal y dela hojarasca (Decaëns 2000). Esteproceso conduce a la formación deuna reserva de semillaspotencialmente listas para germinar,que pueden dispersarse por el fuegoen la superficie del suelo cercano, endonde se beneficiarán de lascondiciones idóneas para germinar,es decir, de un suelo desnudo, demayor intensidad lumínica y denutrientes disponibles en las cenizas.Las limitaciones químicasimportantes de la germinación de lassemillas y de la supervivencia de lasplántulas (por ejemplo, bajocontenido de nutrientes, saturaciónde aluminio alta) puedendesaparecer en las heces. Enconsecuencia, las semillasexcretadas en la superficie del suelotendrán más oportunidades degerminar y de participar en lavegetación epígea. Esta situaciónexplicaría la composición del bancode semillas de las heces, la cual separece más a la de la vegetaciónepígea de la sabana que la del bancode semillas del suelo. Las hecesproducidas por las lombrices puedenconsiderarse, finalmente, como unnicho importante de regeneración(sensu Grubb 1977) de la comunidadvegetal, y la actividad de esaslombrices como un factorindispensable para la sostenibilidady la diversidad del ecosistema(Willems y Huijsmans 1994).
De los sistemas agropastorilesestudiados, el pastizal B tiene unadensa cubierta herbácea que evita,probablemente, la germinación de lassemillas que llegan a su superficie;en el pastizal A ocurre el efectocontrario debido a sus extensasáreas de suelo desnudo. Estas
características pueden explicar lasdiferencias observadas, respecto a lasimilitud de composición deespecies, entre el banco de semillasde las heces o el del suelo y lavegetación epígea, en estos dossistemas de uso de la tierra. Losmecanismos descritos tienen unaimportancia especial, dado que lamayoría de las especies encontradasen las heces son consideradasmalezas por la mayor parte de losgranjeros, tanto en los pastizalescomo en los demás cultivos. Lacomprensión de tales procesos es degran importancia en el diseño desistemas de producción agrícolasostenibles, si en ellos se espera unaintensa actividad de las lombrices detierra.
Estudios recientes handemostrado que la distribuciónespacial de las lombrices (Rossi yLavelle 1998; Rossi et al. 1997) y delas semillas en el suelo (Benoit et al.1992; Bigwood e Inouye 1988;Dessaint et al. 1991; Thompson etal. 1994) es de tipo agregado.Aunque se sabe que el número deplántulas que aparecen en el suelodesnudo es mucho mayor enpresencia de las heces superficialesdepositadas por las lombrices (Grant1983), faltan más estudios sobre larelación entre la distribuciónespacial en sentido horizontal de laslombrices, de las semillas y de lasplántulas. Es necesario aplicar undiseño de muestreo espacialmenteexplícito y herramientasgeoestadísticas para abordar estetema en los estudios que se realicenmás adelante.
Es posible que la limitación másimportante de este estudio estéligada a la ausencia actual dereferencias que describan lassemillas, aparte de las que versansobre métodos de germinación. Estarestricción impidió responder aquíuna pregunta interesante: ¿en qué

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
325
difieren las semillas ingeridas por laslombrices de las que ellas noingieren? Fue imposible tambiéndeterminar la especie de las semillasque son ingeridas y no sobreviven altránsito por el intestino de laslombrices. Para subsanar estaausencia, se requeriríandescripciones detalladas de lassemillas que pertenecen a lasespecies más comunes, por lomenos, en esta parte del mundo.
El presente estudio apoya yespecifica más la “hipótesis de lasbiodiversidades encajadas” de Lavelle(1996), que propone lo siguiente:
- la diversidad de lacomunidad vegetal epígeadetermina la diversidad de lafauna del suelo; y
- los ingenieros ecológicos delsuelo determinan laestructura de lascomunidades de losorganismos más pequeños.
Las estructuras biogénicasproducidas por los ingenieros delecosistema (por ejemplo, las heces delas lombrices, los termiteros)originan cambios en el hábitat y enlas condiciones tróficas de losorganismos más pequeños o menosmóviles. La hipótesis de Lavelleconsideró, inicialmente, los efectosque ejercen los ingenieros delecosistema en diferentes grupos de lafauna del suelo, como losmesoartrópodos y losmicroartrópodos, los enquitreidos,los nematodos y algunosmicroorganismos (bacterias). Elimpacto que causan las lombrices detierra en los movimientos verticales,en la viabilidad y la diversidad de losbancos de semillas del suelodemuestran que estas interaccionesno son de una sola vía sino de doblevía, y que los ingenieros delecosistema pueden, por su parte,causar efectos indirectos en la
dinámica y en la diversidad de lavegetación mediante las estructurasbiogénicas que producen.
AgradecimientosLos autores agradecen a R.J.Thomas (CIAT) y a D.K. Friesen(CIAT) por su apoyo técnico yeconómico. Asimismo, a P. Lavelle(IRD), A. Smith (CIAT), D. Alard(Universidad de Rouen, Francia),G. Rippstein (CIRAD) y J.P. Rossi(IRD) por sus muy útiles comentariosa una primera versión de estecapítulo. También a E. Baudry,X. Graves y E. Mariani por su ayudaen el cómputo de estimadoresaplicando el procedimiento“jackknife”.
ReferenciasAdams JE; McCune ED. 1979.
Application of the generalizedjackknife to Shannon’s measure ofinformation used as an index ofdiversity. En: Grassle JF; Patil GP;Smith W; Taillie C, eds. Ecologicaldiversity in theory and practice.International Co-operativePublishing House, Fairland, MD,Estados Unidos. p 117-131.
Benoit DL; Derksen DA; Panneton B.1992. Innovative approaches toseedbank studies. Weed Sci40:660-669.
Benoit DL; Kenkel NC; Cavers PB. 1989.Factors influencing the precision ofsoil seed bank estimates. Can J Bot67:2833-2840.
Bigwood DW; Inouye DW. 1988. Spatialpattern analysis of seed banks: animproved method and optimizedsampling. Ecology 69:497-507.
Blanchart E; Bruand A; Lavelle P. 1993.The physical structure of casts ofMillsonia anomala (Oligochaeta:Megascolecidae) in shrub savannasoils (Côte d’Ivoire). Geoderma56:119-132.

326
El Arado Natural:…
Bouché MB. 1977. Stratégieslombriciennes. En: Lohm U;Persson T, eds. Soil organisms as acomponent of ecosystems. Ecol Bull25:122-132.
Cavers PB. 1995. Seed banks: memory insoil. Can J Soil Sci 75:11-13.
Chancellor RJ. 1986. Decline of arableweed seeds during 20 years in soilunder grass and the periodicity ofseedling emergence aftercultivation. J Appl Ecol 23:631-637.
Colwell RK. 1997. EstimateS: statisticalestimations of species richness andshared species from samples.Version 5. User’s guide andapplication. Published at: http://viceroy.eeb.uconn.edu/estimates
Cooper DW. 1968. The significance levelin multiple tests madesimultaneously. Heredity23:614-617.
Dalling JW; Swaine MD; Garwood NC.1994. Effect of soil depth onseedling emergence in tropical soilseed-bank investigations. FunctEcol 9:119-121.
Decaëns T. 2000. Degradation dynamicsof surface earthworm casts ingrasslands of the eastern plains ofColombia. Biol Fertil Soils32:146-156.
Dessaint F; Chadoeuf R; Barralis G.1991. Spatial pattern analysis ofweed seeds in the cultivated soilseed bank. J Appl Ecol 28:721-730.
Dutoit T; Alard D. 1995. Permanent seedbanks in chalk grassland undervarious management regimes: theirrole in the restoration of species-rich plant communities. BiodiversConserv 4:939-950.
Escobar E; Belalcázar J; Rippstein G.1993. Claves de las principalesplantas de sabana de la altillanurade los Llanos Orientales enCarimagua, Meta, Colombia. CIAT,Cali, Colombia.
Forcella F. 1984. A species-area curve forburied viable seeds. Aust J Agric Res35:645-652.
Friesen DK; Rao IM; Thomas RJ; ObersonA; Sanz JI. 1997. Phosphorusacquisition and cycling in crop andpasture systems in low fertilitytropical soils. Plant Soil196:289-294.
García MA. 1995. Relationships betweenweed community and soil seed bankin a tropical agroecosystem. AgricEcosys Environ 55:139-146.
Grant JD. 1983. The activities ofearthworms and the fates of seeds.En: Satchell JE, ed. Earthwormecology: from Darwin tovermiculture. Chapman and Hall,Londres. p 107-122.
Grubb PJ. 1977. The maintenance ofspecies-richness in plantcommunities: the importance of theregeneration niche. Biol Rev52:107-145.
Harper JL. 1977. Population biology ofplants. 1a. ed. Academic Press,Nueva York.
Heltshe JF; Forrester NE. 1983.Estimating species richness usingthe jackknife procedure. Biometrics32:1-11.
Heltshe JF; Forrester NE. 1985. Statisticalevaluation of the jackknife estimateof diversity when using quadratsamples. Ecology 66:107-111.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998a. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta: Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl Soil Ecol9:153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998b. Earthworm communities innative savannas and man-madepastures of the eastern plains ofColombia. Biol Fertil Soils28:101-110.

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
327
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones RM; Noguchi M; Bunch GA. 1991.Levels of germinable seed in topsoiland cattle faeces in legume-grassand nitrogen-fertilized pasture inSouth-East Queensland. Aust JAgric Res 42:953-968.
Kellman M. 1974. The viable weed seedcontent of some tropicalagricultural soils. J Appl Ecol2:669-677.
Kellman M. 1978. Microdistribution ofviable weed seeds in two tropicalsoils. J Biogeogr 5:291-300.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Legendre P; Vaudor A. 1991. The Rpackage: multidimentional analysis,spatial analysis. Université deMontréal, Département desSciences Biologiques, Montreal.
Malone CR. 1967. A rapid method forenumeration of viable seeds in soil.Weeds 15:381-382.
Mayer AM; Poljakoff-Mayber A. 1989. Thegermination of seeds. 4a. ed.Pergamon Press, Oxford.
McIvor JG. 1987. Changes in thegerminable seed levels in the soilbeneath pastures in the seasonallydry tropics of north Queensland.Aust J Exp Agric 27:283-289.
McIvor JG; Gardener CJ. 1994.Germinable soil seed banks innative pastures in north-easternAustralia. Aust J Exp Agric34:1113-1119.
McRill M; Sagar GR. 1973. Earthwormsand seeds. Nature 243:482.
Mejía MM. 1984. Nombres científicos yvulgares de especies forrajerastropicales. CIAT, Cali, Colombia.
Milberg P. 1992. Seed bank in a 35-year-old experiment with differentsystems of a semi-naturalgrassland. Acta Oecol 13:743-752.
Pielou EC. 1966. The measurement ofdiversity in different types ofbiological collections. J Theor Biol13:131-144.
Rangel AF. 1996. Efecto de la lombriz detierra Martiodrilus carimaguensissobre la dinámica del nitrógeno y elpunto de carga cero en un Oxisol delos Llanos Orientales de Colombia.Tesis. Universidad Nacional deColombia, Palmira.
Rangel AF; Thomas RJ; Jiménez JJ;Decaëns T. 1999. Nitrogendynamics associated withearthworm casts of Martiodriluscarimaguensis Jiménez and Morenoin a Colombian savanna Oxisol.Pedobiologia 43:557-560.
Reest PJvd; Rogaar H. 1988. The effectsof earthworm activity on the verticaldistribution of plant seeds in newlyreclaimed polder soils in theNetherlands. Pedobiologia31:211-218.
Roberts HA. 1981. Seed banks in soils.Adv Appl Biol 6:1-55.
Roberts HA. 1986. Seed persistence insoil and seasonal emergence inplant species from differenthabitats. J Appl Ecol 23:639-656.
Rossi J-P; Lavelle P. 1998. Earthwormaggregation in the savanna of Lamto(Côte d’Ivoire). Appl Soil Ecol7:195-199.
Rossi J-P; Lavelle P; Albrecht A. 1997.Relationships between spatialpattern of the endogeic earthwormPolypheretima elongata and soilheterogeneity. Soil Biol Biochem29:485-488.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed.Freeman and Company, NuevaYork.

328
El Arado Natural:…
Sørensen T. 1948. A method ofestablishing groups of equalamplitude in plant sociology basedon similarity of species content andits application to analysis of thevegetation on Danish commons.Biol Srk 5:1-34.
Tergas LE; Paladines O; KleinheisterkampI; Velásquez J. 1984. Productividadanimal de Brachiaria decumbenssola y con pastoreo complementarioen Pueraria phaseoloides en losLlanos Orientales de Colombia.Produc Anim Trop 9:1-13.
Thompson K. 1986. Small-scaleheterogeneity in the seed bank ofan acidic grassland. J Ecol74:733-738.
Thompson K; Grime JP. 1979. Seasonalvariation in the seed banks ofherbaceous species in tencontrasting habitats. J Ecol67:893-921.
Thompson K; Green A; Jewels M. 1994.Seeds in soil and worm casts from aneutral grassland. Funct Ecol8:29-35.
Tooren BF van; During HJ. 1988. Viablediaspores in the gut of earthworms.Acta Bot Neerl 37:181-185.
Tukey JW. 1958. Bias and confidence innot quite large samples. Ann MathStat 29:614.
Willems JH; Huijsmans KGA. 1994.Vertical seed dispersal byearthworms: a quantitativeapproach. Ecography 17:124-130.

Efectos Causados por las Lombrices de Tierra en el Banco de Semillas...
329
Familia Género, especie Descriptor En En bancos vegetación de semilla: epígea
De Deheces suelo
Amaranthaceae Amaranthus dubius Mart. + + +
Compositae Emilia sonchifolia (L.) D.C. + + +(= Asteraceae) Pseudoelephanthopus spiralis (Lees.) Cronq. - - +
Stilnopapus pittieri Gleason - - +Wedelia sp. — + + +
Cyperaceae Cyperus haspan L. - - +Fimbristylis complanata (Retz.) Vahl - + -Rhynchospora barbata (Vahl) Kunth - - +Rhynchospora blepharophora (J. and C. Presl) H. Pfriff + + +(= Dichromena ciliata)
Rhynchospora confinis (Nees) C.B. Clarke - + +
Euphorbiaceae Croton trinitatis Millsp. + + +Phyllanthus niruri L. + + +
Fabaceae Arachis pintoi Krap. and Greg. - + +(= Caesalpinioideae) Chamaecrista diphila (L.) Greene - - +(= Papilionoideae) Clitoria guianensis (Aublet) Bentham - - +
Desmodium barbatum (L.) Benth. - - +Desmodium ovalifolium Wall. + + +Galactia glaucescens Kunth - - +Pueraria phaseoloides Benth. + - -Stylosanthes capitata Vog. - + +
Flacourtiaceae Cassearia ulmifolia Vahl ex Benth. - - +
Iridaceae Cypura paludosa Aubl. - - +
Lamiaceae Hyptis capitata Jacq. - - +(= Labiaceae)
Malvaceae Peltaea speciosa (H.B.K.) Stand. - - +Sida sp. — + - -
Melastomataceae Clidemia rubra (Aublet) Mart. - - +
Mimosaceae Mimosa pudica L. - - +
Poaceae Andropogon bicornis L. - - +(= Gramineae) Andropogon gayanus Kunth + + +
Andropogon leucostachyus H.B.K. - - +Andropogon selloanus (Hack.) Hack. - - +Aristida capillacea Lam. + - +Aristida riparia Trin. - - +Aristida tincta Trin. and Rupr. - - +Axonopus aureus Beauv. - - +Axonopus chrysoblepharis (Lam.) Chase - - +Axonopus compressus (Sw.) Beauv. + + +Axonopus purpusii (Mez.) Chase - - +
Apéndice. Familia, género y especie a que pertenecenlas semillas presentes (+) o ausentes (-) en lasheces de la lombriz de tierra estudiada, en elsuelo que las circunda y en la vegetaciónepígea (como expresión de las semillas),considerando como un todo los tres sistemasde uso de la tierra ensayados.a
(Continúa)

330
El Arado Natural:…
Familia Género, especie Descriptor En En bancos vegetaciónde semilla: epígea
De Deheces suelo
Apéndice. (Continuación).
Poaceae Brachiaria decumbens Stapf. + + +(= Gramineae) Brachiaria humidicola (Rendle) Schweickt + + +
Digitaria neesiana Henrard + + -Elyonorus candidus (Trinius) Hackel - - +Gimnopogon foliosus (Willd.) Nees + + +Leptocoryphium lanatum (H.B.K.) Nees - - +Mesosetum loliiforme (Steud.) Chase + - -Oryza sativa L. - + +Panicum laxum Swartz + + -Panicum rudgei Roem. and Schult. + + +Panicum stellatum Humb. and Bonpl. - - +Panicum versicolor (Doell) Henrard - - +Paspalum campestre Nees - - +Paspalum clavuliferum Wright + + +Paspalum conjugatum Berg. + - -Paspalum contractum Flügge - - +Paspalum parviflorum Rohde - - +Paspalum pectinatum Nees - - +Paspalum plicatulum Michx. - + +Schizachyrium hirtiflorum (Retzius Alston) - - +Thrasya petrosa (Trin.) Chase + + +Trachypogon vestitus Anders. + - +
Rubiaceae Borreria sp. — + + +Declieuxia fruticosa (R. & S.) Kuntze + - +Sipanea pratensis Aublet - - +
Scrophulariaceae Lindernia diffusa (L.) Wettst. + + +
Trichopodaceae Trichopodium sp. — - - +(= Trichopus sp.)
Monocotiledónea sp.1 — - - +
a. La clasificación se basa en Mejía (1984) y Escobar et al. (1993).

PARTE VI
Efectos Producidos por los Ingenierosdel Ecosistema desde la Dimensión
de la Muestra Unitaria hasta laParcela Experimental

Determinación del Nivel de Fósforo Orgánico...
333
ResumenSe evaluaron —mediante laespectroscopía de la resonanciamagnética nuclear (RMN) con 31P, elfraccionamiento del P y la absorciónde P— los cambios que experimentael contenido de P en el horizonte A(0-10 cm) de los suelos ácidos desabana en Colombia, que fueronfertilizados con aplicaciones bajas deP y en los cuales habían crecidocontinuamente durante 15 años unpastizal de una sola gramínea(Brachiaria decumbens) y otro en quese asociaban la gramínea y unaleguminosa forrajera (B. decumbens+ Pueraria phaseoloides). El Pdisponible, analizado mediante elmétodo Bray, fue bajo en la sabananativa (1.3 mg/kg suelo) y dio unarespuesta moderada alestablecimiento del pastizal. Almismo tiempo, la afinidad del suelopor el P inorgánico disminuyóligeramente después de dichoestablecimiento. La espectroscopíade RMN con 31P mostró que el P
asociado con los ácidos húmicosestaba dominado por un P-monoéster y, en segundo lugar, porun P-diéster. Se observaroncantidades proporcionalmente másbajas de fosfonatos, de ácido teicóico,de ortofosfato y de pirofosfato. El Pasociado con los ácidos fúlvicospresentó una relación proporcionalmás bajas del P-diéster y uncontenido mayor de ortofosfato. Enla sabana nativa, las reservas de lasespecies de P orgánico inestable(fosfonatos y P-diéster, además delácido teicóico) asociadas con losácidos húmico y fúlvico fueron de12.4 y 1.1 kg/ha, respectivamente;estas asociaciones aumentaron,respectivamente, hasta 18.1 y1.8 kg/ha en el pastizal sinleguminosa y hasta 19.5 y 2.3 kg/haen el pastizal con leguminosa. Losdatos muestran la importancia de lasespecies de P orgánico inestable parael suministro de P a las plantas enlos pastizales mejorados de lostrópicos; indican, además, que el Pdel ácido húmico, en particular,responde a los cambios que presentael uso de la tierrra dentro de unaescala de tiempo relativamente corta.Las heces de las lombrices de tierrafueron muy abundantes en elpastizal de B. decumbens +P. phaseoloides y estabanenriquecidas con especies de Porgánico inestable. La conclusión
CAPÍTULO 20
Determinación, mediante laEspectroscopía de RMN con 31P, delNivel de Fósforo Orgánico de unOxisol, en Pastizales del TrópicoDerivados de la Sabana Nativa1
G. Guggenberger*, L. Haumaier*, R. J. Thomas** yW. Zech*
1. Publicado en Biology and Fertility of Soils 23,p 332-339, por Guggenberger et al.Copyright © 1996 de Springer. Impreso conpermiso de la casa editorial.
* Institute of Soil Science and Soil Geography,University of Bayreuth, D-95440 Bayreuth,Alemania.
** CIAT, AA 6713, Cali, Colombia.

334
El Arado Natural:…
extraída de este estudio es que laactividad de las lombrices mejora elaporte de P en los suelos de estospastizales, porque genera una reservade P orgánico fácilmente asimilable.
Palabras clave: especies orgánicas deP, espectroscopía de RMN con 31P,fósforo del suelo, heces de lombrizde tierra, pastizales tropicalesmejorados, suelos de sabanaácidos
Este trabajo está dedicado alProfesor J.C.G. Ottow con motivo de
su 60º cumpleaños (G.G.)
IntroducciónHay una firme convicción de que lossuelos del trópico húmedo estánsometidos a deficiencias de P, N, K,Ca, Mg, S, Zn y Mo, la cual revisteespecial importancia para losUltisoles y Oxisoles del neotrópico,que son viejos y están intensamentemeteorizados. En los suelos ácidos delas sabanas de América Latina, el Pconstituye un problema particularporque es fijado por los óxidos ehidróxidos de Fe y de Al, y seconvierte así en una limitante químicapara la producción agrícola en esossuelos.
Ball-Coelho et al. (1993)demostraron que el P orgánico delsuelo contribuye a la nutrición de lasplantas en los suelos fuertementemeteorizados. El conocimientodetallado de la composición del Porgánico puede revelar sucontribución potencial en la cadenaque va desde la descomposiciónmicrobiana de la materia orgánica(MO) hasta el P inorgánicosuministrado a las plantas. Newmany Tate (1980) y Tate y Newman (1982)fueron pioneros en el uso de laespectroscopía de resonanciamagnética nuclear (RMN) con 31P paracuantificar las especies de P orgánico
e inorgánico de los extractos alcalinosdel suelo. En condiciones tropicales,la fracción soluble de NaOH puedeformar una reserva relativamenteactiva de P y puede reaccionar mássensiblemente a los cambios en elsistema de uso de la tierra dentro deuna escala de tiempo relativamentecorta, representando así una fracciónmás lábil de P (Oberson et al. 1995;Tiessen et al. 1992). De los diferentestipos de compuestos de Pidentificados por el análisis de RMNcon 31P, los diésteres de ortofosfato,incluyendo en ellos los fosfolípidos ylos ácidos nucleicos, se conviertenmuy fácilmente en P inorgánico, queestá disponible para las plantas,cuando el suelo ofrece un ambientefavorable (Hinedi et al. 1988; Tate yNewman 1982).
Los pastizales se han convertidoen un sistema de uso de la tierra muyextendido, tras la deforestación o eldesarrollo de cultivos en los suelos dela sabana tropical. Hay pruebas muyclaras de que el establecimiento depastizales mejorados en los suelosácidos de fertilidad baja de unasabana, especialmente cuando sesiembran en él especies vegetales queproduzcan una gran cantidad debiomasa en el interior del suelo,puede incrementar realmente elcontenido de la MO y la fertilidad deese suelo (Thomas et al. 1995).
En el presente estudio se empleóla RMN con 31P en los ácidos fúlvico yhúmico para estudiar la composiciónquímica del P moderadamenteresistente en el horizonte A de unOxisol. Los datos obtenidos se hanusado para establecer posiblesdiferencias entre estas fracciones deP, tanto en la sabana nativa como enun pastizal de gramínea sola, en otrode gramínea asociada conleguminosa, y en las heces de laslombrices, muy abundantes en elsistema en que se sembraron lagramínea y la leguminosa. Se discute

Determinación del Nivel de Fósforo Orgánico...
335
la importancia que tienen para ladisponibilidad del P las especies de Pidentificadas, partiendo de los datosdel fraccionamiento de P y de unexperimento de sorción (o sea, deabsorción o adsorción) de P.
Materiales y Métodos
Descripción del sitio delensayoEl sitio de estudio representa lassabanas isohipertérmicas biendrenadas y se encuentra en el CentroNacional de InvestigacionesAgropecuarias (CNIA) Carimagua, enColombia (4º37’ N, 71º19’ O,175 msnm). Los valores mediosanuales de la temperatura y laprecipitación fueron 26 ºC y2200 mm, respectivamente,distribuidos desde abril hastadiciembre. Los suelos son, en sumayoría, Oxisoles de baja fertilidadcaracterizados por un pH bajo(< 5, en agua), una saturación de Alalta (> 75%) y niveles bajos de loscationes intercambiables de Ca, Mg yK. Los factores que contribuyen a lainfertilidad de estos suelos soncomplejos y entre ellos están latoxicidad debida al Al, y los bajoscontenidos de cationes básicosintercambiables y de P disponible.En 1978 se sembró la especieforrajera tropical de raíces profundasBrachiaria decumbens cv. Basiliskjunto con la leguminosa Puerariaphaseoloides CIAT 9900 (kudzú) enun área ocupada antes por la sabananativa. Ambas especies forrajerasestán muy bien adaptadas a lascondiciones de estos suelos ácidos.La aplicación de fertilizante (kg/ha)durante 15 años (1978-1993) a lospastizales introducidos fue de 0 N,103 P, 63 K, 648 Ca, 56 Mg y 77 S ala gramínea y de 0 N, 106 P, 80 K,648 Ca, 63 Mg y 90 S a las franjasde leguminosa.
El establecimiento del pastizalcausó un impacto significativo en laabundancia de la macrofauna delsuelo. La biomasa de lombrices en lasabana, en el pastizal de gramíneasola y en el de gramínea conleguminosa fue, respectivamente, de4.78, 22.95 y 51.09 g peso fresco/m2
(Decaëns et al. 1994). La ingestiónanual de suelo por las lombricesen el sistema B. decumbens +P. phaseoloides se calculó en 680 Mgsuelo/ha (Jiménez et al. 1994).
Muestreo del sueloEn septiembre de 1993 se tomaron alazar, manualmente, cuatro muestrasde suelo en cada uno de lostratamientos, los cuales habían sidosometidos a 15 años de pastoreo. Lasmuestras se tomaron en el horizontemineral A (0-10 cm) solamente, yaque la MO del suelo del pastizal esmás abundante cerca de su superficiey decrece al descender en el perfil delsuelo (Rao et al. 1994). Se recogieronademás al azar unas 20 heces delombriz de tierra en el pastizal de1 ha de B. decumbens + P.phaseoloides. En el laboratorio, lasmuestras fueron homogeneizadas y seextrajeron de ellas los restos visiblesde raíces. Todas las muestras desuelo se almacenaron secas y sutemperatura de secado, 105 °C, setomó como base de los resultados.
ExtracciónLos extractos alcalinos de los suelosse obtuvieron agregando, a 50 g desuelo fino (< 2 mm) secado al aire,200 ml de NaOH 0.1 M durante 24 hbajo una atmósfera de N2. Una vezcentrifugada la mezcla, elsobrenadante se acidificó hasta pH 2con HCl 2 M, y luego se centrifugópara separar las fracciones húmica yfúlvica ácidas. Los residuos sedispersaron mediante ultrasonidoempleando 200 ml de NaOH 0.5 M

336
El Arado Natural:…
durante 10 minutos, y undesintegrador de tipo sonda (HeatSystems-Ultrasonics, ModeloW 185-F). La cantidad de energíatotal disipada fue de 270 J/ml. Lassuspensiones se centrifugaron y lossobrenadantes se acidificaron hastapH 2 con HCl 2 M, y se centrifugaronde nuevo para separar por segundavez las fracciones húmica y fúlvica. Atodas las fracciones se les practicóuna diálisis extensa. Terminadaésta, las dos fracciones húmicasácidas se combinaron en una sola,así como las dos fracciones fúlvicasácidas, se secaron ambas en frío y seanalizaron respecto a su contenido deC en un analizador “EL CNSElementar Vario”.
El P contenido en los ácidoshúmico y fúlvico correspondió a lasuma de las fracciones de Pobtenidas con NaOH, es decir, por elmétodo del fraccionamientosecuencial de P según Hedley et al.(1982). El P orgánico soluble en aguay el P inorgánico soluble en agua y enNaHCO3 (las formas inestables delprocedimiento de Hedley et al. 1982)no pudieron ser extraídos,probablemente, con el método aquíindicado, ya que las moléculas muypequeñas extraídas con NaOHatraviesan el tubo de la diálisis.
Análisis químico del PEl P total del suelo se extrajomediante la adición en secuencia deH2SO4 concentrado, H2O2 y ácidofluorhídrico (HF) (Bowman 1988); el Pinorgánico se determinó a partir de laextracción en secuencia con H2SO4 yHF solamente, y el P orgánico secalculó como la diferencia entre esosdos valores. Los valores del Porgánico son cantidades mínimas, yaque parte de ese P puede serhidrolizado por el H2SO4. Paracalcular el P disponible para lasplantas se empleó el método Bray II(Olsen y Sommers 1982). El P total
de las fracciones húmica y fúlvica sedeterminó tras la ignición (550 ºC) yla disolución de los residuos en H2SO40.1 M; el P inorgánico se midiódirectamente en los extractos; y el Porgánico de estas fracciones secalculó como la diferencia entre losvalores anteriores. En todos estoscasos, el P se analizó mediante unmétodo modificado que emplea azulde molibdeno.
Sorción de P inorgánicoLa sorción (absorción o adsorción) porel suelo del P inorgánico se determinóagitando 10 g de suelo con 100 ml deagua deionizada que contenían,alternativamente, 4.96, 9.92, 14.88,19.84 y 24.80 mg/L de P comoKH2PO4, durante 24 h a temperaturaambiente constante (25±1 ºC) y sinajustar el pH. Tras la centrifugacióny la filtración, el P inorgánico sedeterminó colorimétricamente en elsobrenadante (Olsen y Sommers1982). La diferencia entre el Pinorgánico en solución antes delexperimento y después delexperimento dio la cantidad de Pinorgánico extraído de la solución.
Mediciones de RMN con 31PUna muestra de 150 mg secadaen frío se disolvió en 3 cm3 deNaOD 0.5 M en tubos de RMN de10 mm y se analizó a los 2 días. Seusó un espectrómetro Bruker AM 500que funcionaba a una frecuencia de202 MHz, para obtener, una vezrecolectadas entre 8,000 y 20,000imágenes de barrido, los espectros nodesacoplados de 1H. Los espectros seregistraron a una temperatura de20 ºC, con un ángulo de impulso de90º, con un retraso del impulso de0.2 s, y con un tiempo de adquisiciónde 0.1 s. En estudios preliminares enque el retraso del impulso fue de 2 sse obtuvo una distribución de lasdiferentes especies de P que se

Determinación del Nivel de Fósforo Orgánico...
337
equipara a la anterior, aunque laresolución de los espectros obtenidano fue mejor que cuando se trabajócon un retraso de impulso de 0.2 s.
Las mediciones se hicieron conrespecto al ácido ortofosfórico al 85%.Los picos fueron asignados según lainformación obtenida en la literatura(Condron et al. 1990; Newman y Tate1980) y las áreas bajo los picos semidieron por integración electrónica.Dado que el ortofosfato orgánicoasociado a especies paramagnéticas,como el hierro, no sería detectado porel análisis RMN con 31P (Bedrock et al.1995), los resultados de este trabajono son cuantitativos, pero puedenusarse para hacer comparaciones;en efecto, el contenido de hierro de lafracción húmica fue comparableen las cuatro muestras de sueloempleadas (ácido húmico:56-81 g/kg, ácido fúlvico:1.4-1.9 g/kg).
Resultados
Distribución del CLa cantidad de C orgánico del sueloaumentó desde 22 g/kg en la sabanahasta 26 g/kg en el pastizal, sinleguminosa o con ella (Guggenbergeret al. 1995). El contenido de Corgánico de las heces de las lombricesfue de 56 g/kg.
De las cuatro muestras de suelose extrajeron 330 g/kg de C orgánico
mediante el procedimiento deextracción en secuencia(Cuadro 20-1). La mayor parte deeste C fue separado como ácidohúmico. Oberson et al. (1995)hicieron extracciones en los mismossuelos mediante el esquema defraccionamiento de Hedley et al.(1982). La suma de las fracciones deC orgánico soluble, extraídassecuencialmente en agua, en NaHCO3y en NaOH, fue de 470 g/kg. Sinembargo, el rendimiento del Corgánico de los extractos alcalinos deOberson et al. (1995) pudocompararse a la suma del C orgánicode las fracciones húmica y fúlvica delas muestras de este estudio. Estosresultados indicaron que la fracciónlábil de P, soluble en NaHCO3 y enagua, se perdió durante la diálisis.
Concentraciones de PLos niveles de P total (Cuadro 20-2)aumentaron ligeramente en el ordensiguiente:
sabana pastizal de pastizal denativa gramínea gramínea y
leguminosa
(332 mg/kg) (358 mg/kg) <(382 mg/kg)
Estos valores fueron 1.6 vecessuperiores a los obtenidos mediantela digestión con ácido perclórico(Oberson et al. 1995). Según Olsen ySommers (1982), la digestión conácido perclórico puede subestimar elcontenido de P en los suelosaltamente meteorizados. Esta
Cuadro 20-1. Distribución del carbono entre las fracciones del ácido húmico (AH), del ácido fúlvico(AF) y de la humina en los suelos de Carimagua. La suma de C-AH, C-AF y C-huminaes < 1000 g/kg C debido a la pérdida de C durante la centrifugación y, principalmente,a la diálisis de los compuestos de peso molecular bajo.
Muestra de: Carbono (g/kg C) en la fracción:C-AH C-AF C-humina
Sabana nativa 226 87 325Brachiaria decumbens 248 82 271B. decumbens + Pueraria phaseoloides (B+P) 239 85 277Heces de lombrices en B+P 236 80 395

338
El Arado Natural:…
discrepancia refleja, sin duda, losproblemas que surgen cuando sedetermina el P en los suelostropicales. La cantidad de Pinorgánico fluctuó entre 144 y188 mg/kg de suelo, mientras que lade P orgánico fue aproximadamentede 190 mg/kg suelo. El métodoBray arrojó una cantidad pequeñade P en el suelo de la sabana nativa(1.3 mg/kg suelo) y una cantidadapenas un poco mayor que ésta enel pastizal de la asociación degramínea y leguminosa.
La mayor parte del P extraíbleestaba asociado a los ácidoshúmicos. Al establecer los pastizalesmejorados, aumentó la cantidad deP total de los ácidos húmico y fúlvicoen más de un 40%, si se comparacon la obtenida para ellos en lasabana nativa (Cuadro 20-2). Estosvalores fueron de dos a cuatro vecessuperiores en las heces de laslombrices que en el suelo que lascircundaba, lo que indica unenriquecimiento notable del Pextraíble.
El P orgánico dominóinvariablemente al P de los ácidoshúmicos y la relación proporcionaldel P inorgánico en dichos ácidos fuemuy alta en las muestras de lasheces de las lombrices. En general,la relación proporcional del Pinorgánico fue mayor respecto al Pdel ácido fúlvico que respecto al P
del ácido húmico; en las heces, el Pcontenido en el ácido fúlvico fuetotalmente inorgánico cuando sedeterminó por los procedimientoshúmedos de la química.
Sorción de PEl fraccionamiento del P inorgánicoentre la solución y las fases desorción (Figura 20-1) mostró que elsuelo de Carimagua tenía unaafinidad por el P inorgánico entre bajae intermedia, según los criterios de
Cuadro 20-2. Concentración de P (mg/kg de suelo) y su distribución en los ácidos húmicos y fúlvicosde los suelos de Carimagua. Los valores se basan en dos repeticiones, n = 2; elcoeficiente de variación del P en las fracciones individuales fue < 5%; PT = P total en elsuelo; Pi = P inorgánico; Po= P orgánico; P Bray = P extraíble en NH4F-HCl.
Muestra de: PT Pi Po P Bray P de ácido húmico P de ácido fúlvico
PT Po Pi PT Po Pi
Sabana nativa 332 144 188 1.3 23.1 18.9 4.9 4.0 2.3 1.7B. decumbens 358 163 195 1.4 32.9 25.7 7.2 5.7 3.6 2.1B. decumbens + 382 188 194 2.2 30.8 24.2 6.6 6.5 4.0 2.5 P. phaseoloides (B+P)
Heces de lombrices 592 392 200 n.a.a 69.9 60.1 9.8 31.4 0 31.4 en B+P
a. No analizado.
250
200
150
100
50
00 1 10
Sabana nativaB. decumbensB. decumbens + P. phaseoloidesB. decumbens + P. phaseoloides(heces de lombrices)
P en la solución
Figura 20-1. Isotermas de sorción de P en lossuelos de Carimagua.
P de
sor
ción
(mg/
kg)

Determinación del Nivel de Fósforo Orgánico...
339
Juo y Fox (1977). La MO del suelosuperficial pudo haber contribuido ala sorción relativamente baja del Ppor el suelo. Juo y Fox (1977)reportaron isotermas de sorciónsimilares en la superficie de Ultisolesque se desarrollaron sobre rocasácidas. La afinidad del suelo por el Pinorgánico disminuyó un poco tras elestablecimiento de los pastizales.Las heces de las lombricespresentaron una concentración de Pinorgánico en equilibrio muchomayor que el suelo que lascircundaba, cuando se añadieron ala solución cantidades pequeñas de Pinorgánico. Sin embargo, cuando seaplicó la cantidad más alta posiblede P, los valores fueron similares.
Espectroscopía de RMN con 31PLos espectros de RMN con 31P de losácidos húmico y fúlvico que seextrajeron de las muestras deCarimagua (Figuras 20-2 y 20-3)presentaron señales intensas enδ = [4.3-5.6] ppm, que provenían demonoésteres de ortofosfato, unaforma diversa de P que comprendefosfatos de inositol, fosfatos deazúcares y mononucleótidos. Laseñal en δ = 4.3 ppm puede serasignada a la fosfocolina. Losdiésteres de ortofosfato, por ejemplolos fosfolípidos y el ADN (Newman yTate 1980), dieron resonancia enδ = [0.0-0.2] ppm. Las señalesseparadas de los diésteres obtenidasen δ = [1.0-2.0] ppm pueden provenirdel ácido teicóico, un diéster deortofosfato que consta de unidadesde un azúcar ligadas a grupos fosfato(Condron et al. 1990). Puesto que elácido teicóico no es un compuestosimple sino uno complejo, semanifiesta en un rango de δ ppm.El pico de señal obtenido enδ = 19.2 ppm procedía de losfosfonatos. Las señalescorrespondientes al P inorgánicohicieron picos en δ = 6.5 ppm
(ortofosfato) y en δ = -4.4 ppm(pirofosfatos). En el Cuadro 20-3 semuestra la distribución de lasespecies de P mencionadas.
Los datos obtenidos con laquímica desarrollada en mediohúmedo concuerdan con laespectroscopía de RMN con 31P en losiguiente: las especies de P orgánicodominan los ácidos húmicos (> 94%de toda la intensidad de la señal). ElP monoéster constituyó unporcentaje significativo del P de losácidos húmicos (44% a 59% de la
Figura 20-2. Espectros de resonancia magnéticanuclear (RMN) con 31P de los ácidoshúmicos extraídos de los suelos deCarimagua. El contenido de P enlos ácidos húmicos fue el siguente:sabana nativa, 1146 mg/kg;B. decumbens, 1294 mg/kg;B. decumbens + P. phaseoloides,1320 mg/kg; heces de laslombrices en B. decumbens +P. phaseoloides, 2138 mg/kg.
Sabana nativa
5.0 0.14.5
B. decumbens
-4.4
B. decumbens +P. phaseoloides
B. decumbens +P. phaseoloides(heces delombrices)
1.5
ppm20 10 0 -10 -20
19.2
6.5
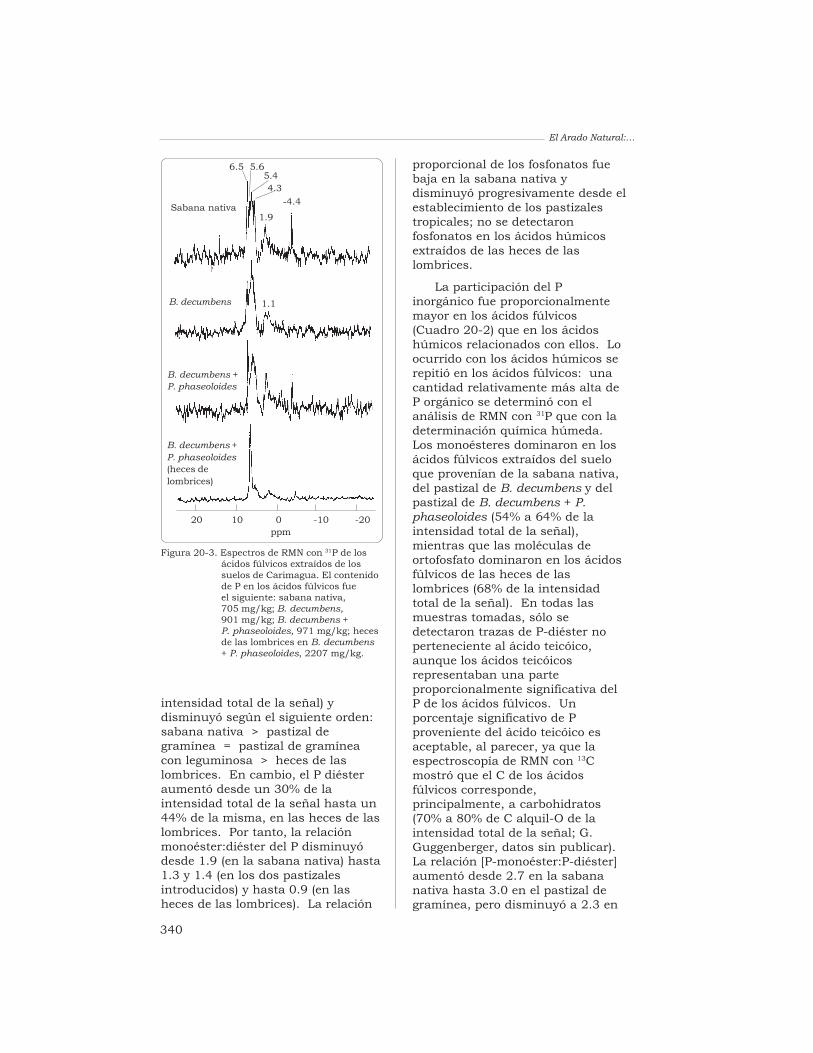
340
El Arado Natural:…
Figura 20-3. Espectros de RMN con 31P de losácidos fúlvicos extraídos de lossuelos de Carimagua. El contenidode P en los ácidos fúlvicos fueel siguiente: sabana nativa,705 mg/kg; B. decumbens,901 mg/kg; B. decumbens +P. phaseoloides, 971 mg/kg; hecesde las lombrices en B. decumbens+ P. phaseoloides, 2207 mg/kg.
intensidad total de la señal) ydisminuyó según el siguiente orden:sabana nativa > pastizal degramínea = pastizal de gramíneacon leguminosa > heces de laslombrices. En cambio, el P diésteraumentó desde un 30% de laintensidad total de la señal hasta un44% de la misma, en las heces de laslombrices. Por tanto, la relaciónmonoéster:diéster del P disminuyódesde 1.9 (en la sabana nativa) hasta1.3 y 1.4 (en los dos pastizalesintroducidos) y hasta 0.9 (en lasheces de las lombrices). La relación
proporcional de los fosfonatos fuebaja en la sabana nativa ydisminuyó progresivamente desde elestablecimiento de los pastizalestropicales; no se detectaronfosfonatos en los ácidos húmicosextraídos de las heces de laslombrices.
La participación del Pinorgánico fue proporcionalmentemayor en los ácidos fúlvicos(Cuadro 20-2) que en los ácidoshúmicos relacionados con ellos. Loocurrido con los ácidos húmicos serepitió en los ácidos fúlvicos: unacantidad relativamente más alta deP orgánico se determinó con elanálisis de RMN con 31P que con ladeterminación química húmeda.Los monoésteres dominaron en losácidos fúlvicos extraídos del sueloque provenían de la sabana nativa,del pastizal de B. decumbens y delpastizal de B. decumbens + P.phaseoloides (54% a 64% de laintensidad total de la señal),mientras que las moléculas deortofosfato dominaron en los ácidosfúlvicos de las heces de laslombrices (68% de la intensidadtotal de la señal). En todas lasmuestras tomadas, sólo sedetectaron trazas de P-diéster noperteneciente al ácido teicóico,aunque los ácidos teicóicosrepresentaban una parteproporcionalmente significativa delP de los ácidos fúlvicos. Unporcentaje significativo de Pproveniente del ácido teicóico esaceptable, al parecer, ya que laespectroscopía de RMN con 13Cmostró que el C de los ácidosfúlvicos corresponde,principalmente, a carbohidratos(70% a 80% de C alquil-O de laintensidad total de la señal; G.Guggenberger, datos sin publicar).La relación [P-monoéster:P-diéster]aumentó desde 2.7 en la sabananativa hasta 3.0 en el pastizal degramínea, pero disminuyó a 2.3 en
Sabana nativa
6.5 5.6
B. decumbens 1.1
B. decumbens +P. phaseoloides
B. decumbens +P. phaseoloides(heces delombrices)
ppm20 10 0 -10 -20
5.44.3
-4.4
1.9
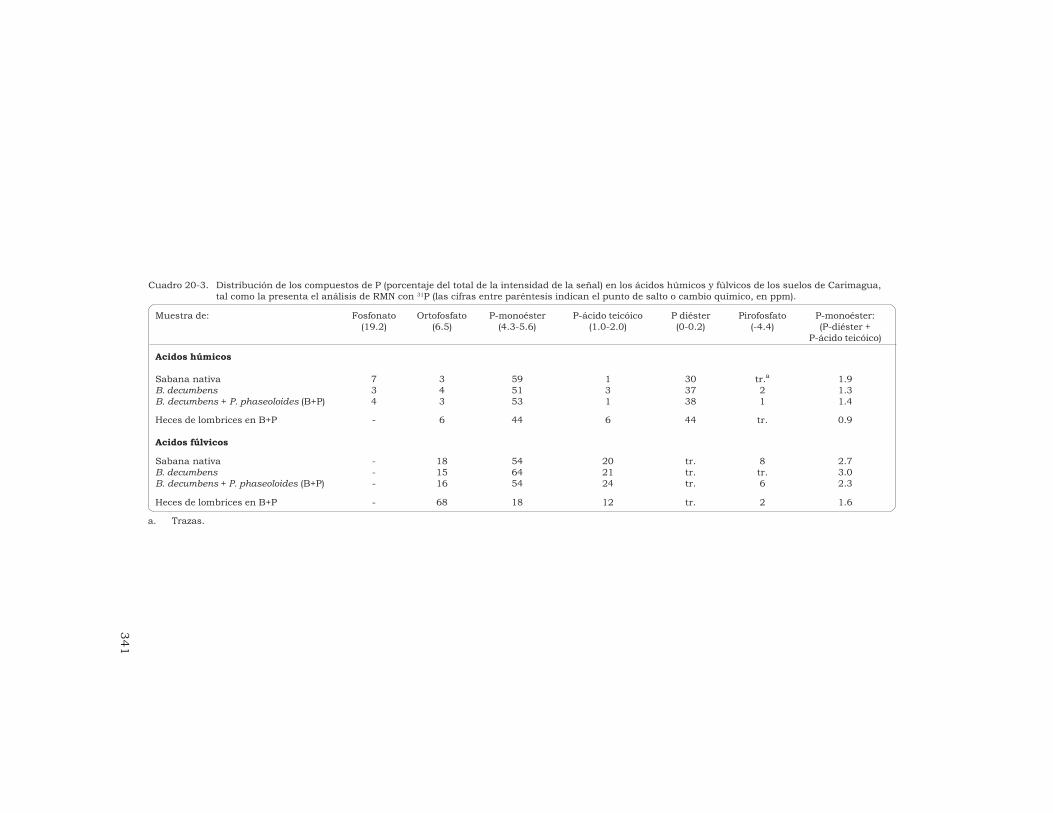
Determ
inación del Nivel de Fósforo O
rgánico...
341
Muestra de: Fosfonato Ortofosfato P-monoéster P-ácido teicóico P diéster Pirofosfato P-monoéster:(19.2) (6.5) (4.3-5.6) (1.0-2.0) (0-0.2) (-4.4) (P-diéster +
P-ácido teicóico)
Acidos húmicos
Sabana nativa 7 3 59 1 30 tr.a 1.9B. decumbens 3 4 51 3 37 2 1.3B. decumbens + P. phaseoloides (B+P) 4 3 53 1 38 1 1.4
Heces de lombrices en B+P - 6 44 6 44 tr. 0.9
Acidos fúlvicos
Sabana nativa - 18 54 20 tr. 8 2.7B. decumbens - 15 64 21 tr. tr. 3.0B. decumbens + P. phaseoloides (B+P) - 16 54 24 tr. 6 2.3
Heces de lombrices en B+P - 68 18 12 tr. 2 1.6
a. Trazas.
Cuadro 20-3. Distribución de los compuestos de P (porcentaje del total de la intensidad de la señal) en los ácidos húmicos y fúlvicos de los suelos de Carimagua,tal como la presenta el análisis de RMN con 31P (las cifras entre paréntesis indican el punto de salto o cambio químico, en ppm).

342
El Arado Natural:…
Cuadro 20-4. Reservas calculadas (kg/ha) de P total (PT), de P Bray y de las diferentes especies de Pasociadas con los ácidos húmicos y fúlvicos en la capa superficial del suelo (0-10 cm) deCarimagua. El P inorgánico incluye a los ortofosfatos y al pirofosfato; el P-diéster incluyeal P del ácido teicóico.
Muestra de: PT P Bray P de ácido húmico P de ácido fúlvico Inorgánico Mono- Diéster + Inorgánico Mono- Diéster +
éster fosfonato éster fosfonato
Sabana nativa 461 1.8 0.8 19.1 12.4 1.4 3.0 1.1B. decumbens 501 1.9 2.5 23.6 20.0 0.9 5.4 1.8B. decumbens + 534 3.0 2.1 22.5 18.1 1.5 5.2 2.3
P. phaseoloides
el pastizal de gramínea conleguminosa. Las heces de laslombrices presentaron los valoresmás bajos de esta relación del P.
DiscusiónLa cantidad de P calculada a partirde los resultados obtenidos indicóque el incremento en las reservas delP total, una vez establecido elpastizal (Cuadro 20-4) fue menor queel P total añadido en forma defertilizante (103 kg/ha de P en elpastizal de gramínea y 106 kg/ha deP en el pastizal de gramínea asociadacon leguminosa). El P que no sepudo contabilizar ascendió a63 kg/ha en el pastizal deB. decumbens y a 30 kg/ha en elpastizal de B. decumbens + P.phaseoloides. Las pérdidas de P enel suelo del pastizal de gramínea conleguminosa se atribuyeron alpastoreo (36 kg/ha, Oberson et al.1995), aunque en el suelo delpastizal de gramínea sola lacantidad de P que se pierde comoconsecuencia del pastoreo(24 kg/ha, Oberson et al. 1995) fuemuy pequeña como para responderpor todo el P que no pudo sercuantificado. En concordancia conOberson et al. (1995), el movimientodel P orgánico soluble y constituidoen partículas desde la superficie delsuelo hasta los horizontes másprofundos fue, probablemente, elresponsable del balance negativo del
P en la superficie del suelo de laparcela de B. decumbens en elpresente estudio. De este modo, ladiferencia entre el pastizal degramínea sola y el de gramíneaasociada con leguminosa se atribuyóa la mayor eficiencia de laleguminosa respecto a la gramíneapara extraer el P desde el subsuelohasta la superficie.
La fuerte interacción entre el ionfosfato y los hidróxidos de Fe y de Ales típica de los Oxisoles y fueresponsable de las bajasconcentraciones del P disponiblepara las plantas obtenidas con elmétodo Bray. Como lo reportaronOberson et al. (1995), elestablecimiento de los pastizalesmejorados y la concurrenteaplicación de P en forma defertilizante aumentaron la cantidadde P inorgánico fácilmentedisponible.
Los resultados obtenidos en esteestudio mediante el análisis de RMNcon 31P indicaron que las especies deP orgánico solubles en álcali estabanunidas, principalmente, por enlacesestéricos a la MO del suelo, de talmodo que quedaban integradas aesas moléculas orgánicas (Stewart yTiessen 1987). La mayor parte del Porgánico estaba asociada con losácidos húmicos (Cuadro 20-2). Sinembargo, tanto los datos obtenidosmediante los análisis químicos y elanálisis de RMN con 31P indicaron

Determinación del Nivel de Fósforo Orgánico...
343
que los ácidos húmicosintensamente dializados y los ácidosfúlvicos, en especial, presentabancontenidos significativos de Pinorgánico. Este efecto no es,probablemente, un resultado de lahidrólisis alcalina del P-monoéster ydel P-diéster en la solución de NaOHpreparada para la espectroscopía deRMN. Newman y Tate (1980)encontraron que los extractosalcalinos pueden almacenarsedurante algunos días sin queaparezcan cambios significativos enlos espectros; los resultados delpresente estudio indicaron solamentecambios menores en la distribuciónde las especies de P tras una semanade almacenamiento (L. Haumaier,datos sin publicar). Las diferenciasdetectadas entre los resultados delmétodo químico en medio húmedo yla espectroscopía se debieron, enparte, a las incertidumbres quegenera el método químico indirectoen la determinación del P orgánico.Sin embargo, la cantidad de Pinorgánico de los extractosanalizados mediante RMN con 31P fuesiempre menor, proporcionalmente,que la obtenida mediante la químicaen húmedo, lo que indica un errorsistemático. Bedrock et al. (1995)reportaron que las moléculas deortofosfato inorgánico asociadas conespecies paramagnéticas, como el Fe,no son detectadas por la prueba deRMN con 31P.
Bowman y Cole (1978)demostraron que la MO extraíble conNaOH contenía cantidadesproporcionalmente altas de Pmineral. En varios estudios se hamostrado que el P inorgánico estáligado a los ácidos fúlvico y húmicomediante puentes catiónicos, comolos complejos P-Ca-humato(o -fulvato) (Dormaar 1972) o elP-Fe(Al)-humato (o -fulvato) (Gerke yHermann 1992). Estos complejospueden ser lo suficientemente
grandes como para tolerar la diálisis.La degradación y la mineralizaciónde la MO hace disponible el Pmineral que se encuentra unido a lasmoléculas orgánicas (Bowman y Cole1978).
Se acepta que el P orgánicosoluble en NaOH fluctúe, en general,de moderadamente lábil amoderadamente estable (Bowman yCole 1978). Numerosasinvestigaciones han mostrado que laespectroscopía de RMN con 31P esmuy apropiada para distinguir loscompuestos de P que presentendiferentes grados de estabilidadfrente a la acción microbiana.Hinedi et al. (1988) encontraron quela fracción de P-diéster puedeconvertirse con relativa rapidez en Pinorgánico en suelos favorables,mientras que la de P-monoéster esmucho más estable. De este modo,el descenso en la tasa P-monoéster:P-diéster encontrada en los ácidoshúmicos sugiere que los niveles de Porgánico lábil aumentan tras elestablecimiento de un pastizaltropical. Hay evidencias queprueban que los productos derivadosde la acción microbiana contribuyen,de modo importante, a la cantidad deP diéster, incluyendo los ácidosteicóicos (Condron et al. 1990). Tatey Newman (1982) encontraron unafuerte correlación entre el P diéster yvarios índices obtenidos a partir dela biomasa microbiana. Según estosmismos autores, los fosfonatospueden contribuir también a lasconcentraciones del P orgánico lábilderivado de la actividad microbiana.
Las relaciones [P diéster:Pmonoéster] encontradas en losácidos húmicos (1-3:1) fueron másbajas que las referenciadas en laliteratura para los extractos alcalinos(4-20:1) (Condron et al. 1990;Hawkes et al. 1984; Zech et al.1987); esto indica que las especiesde P orgánico lábil enriquecían los

344
El Arado Natural:…
ácidos húmicos de los suelos deCarimagua. Estas elevadasconcentraciones de especies de Porgánico lábil asociadas a ácidoshúmicos entran en conflicto con lamayor parte de los hallazgosanteriores, en los que el P orgánicode los ácidos húmicos se consideró elreflejo de la larga y lentatrasformación que experimenta el P(Bowman y Cole 1978). Ahora bien,según Ball-Coelho et al. (1983),cualquier asignación que se hagaordinariamente a la estabilidad del Pno refleja, necesariamente, losprocesos que se desarrollan en elcampo; además, Tiessen et al. (1992)mostraron, en los suelos cercanos aBrasilia, que la fracción de Porgánico soluble en NaOH reflejó loscambios globales experimentadospor la MO del suelo y por los nivelesde P orgánico cuando se modificó eluso dado a la tierra. La fracción deP orgánico encontrada en los ácidoshúmicos era representativa de lafracción de P orgánico alcalino-soluble determinada por Oberson etal. (1995), ya que los ácidos húmicosde los suelos de Carimaguacontenían 90% del total del Porgánico alcalino-soluble(Cuadro 20-2). Los mismos autoresconcluyeron que se debe ajustar lainterpretación actual del P orgánicoalcalino-soluble a las condicionestropicales, porque esta fracciónreacciona, en períodos cortos detiempo, ante los cambios queocurran en el uso de la tierra.
A diferencia de lo ocurrido en larelación [P monoéster:P diéster] delos ácidos húmicos, esta clase derelación no mostró, en los ácidosfúlvicos, una tendencia definidacuando cambió el uso de la tierra.Es más, la relación [P monoéster:P diéster] de los ácidos fúlvicos fuemayor que la correspondiente de losácidos húmicos, al menos en lasfracciones extraídas del suelo
cubierto con pastizales mejorados.Estos resultados sugieren que el Porgánico de los ácidos fúlvicos esmás estable, lo que concuerda, a suvez, con la medida más alta de larelación [C:P orgánico] que se hallóen los ácidos fúlvicos. En los ácidoshúmicos, la relación [C:P orgánico]fue de 225:1 (sabana nativa), 200:1(B. decumbens) y 220:1 (B.decumbens + P. phaseoloides), y elvalor de esta relacióncorrespondiente a los ácidos fúlvicosfue de 620:1 (sabana nativa), 450:1(B. decumbens) y 430:1 (B.decumbens + P. phaseoloides). El Pse mineraliza cuando la relación [C:Porgánico] es igual a 22 o menor queeste valor, según Dalal (1977), y lainmovilización del P inorgánicoocurre cuando esa relación es igual a300 o es mayor que este valor.Ahora bien, Oberson et al. (1995)pusieron en duda las tasas críticasde la relación [C:P orgánico]empleadas y sugieren su revisiónpara adaptarlas a las condicionesencontradas en los suelos tropicales.
El P que se encuentra en losgrupos fosfonato y diéster, enparticular (incluyendo aquí el P delácido teicóico) podría suministrarrápidamente, si se mineraliza, el Pinorgánico disponible para lasplantas (Tate y Newman 1982). Elcálculo de la reserva de las diferentesfracciones de P en el suelo superficial(0 a 10 cm) demostró que lasreservas de P-diéster lábil y de Pfosfonado que se asocian con losácidos húmicos excedieron concreces la reserva del P disponiblepara las plantas (Bray P;Cuadro 20-4). Las reservas de Porgánico lábil asociadas con losácidos fúlvicos fueron similares a lasobtenidas por el método Bray. Elmanejo del pastizal de gramínea solao asociada con leguminosa, juntocon una aplicación de fertilizante,incrementó ligeramente la cantidad

Determinación del Nivel de Fósforo Orgánico...
345
del P determinado por el métodoBray y causó un incrementósignificativo en las reservas de lasespecies lábiles de P orgánico. Esteresultado se debió, principalmente, auna relación proporcional más altadel P-diéster en los extractos.
Es difícil establecer, empleandosolamente los datos de la RMN con31P, si una reserva grande de Porgánico fácilmente disponible puedaatribuirse al aumento de la actividadmicrobiana y a tasas de reciclajealtas o, en cambio, a unaacumulación de P orgánico lábildebida a una actividad microbianalimitada. Condron et al. (1990) yHawkes et al. (1984), por ejemplo,informaron que el descenso de la MOasociado con el cultivo de pastizalesen los suelos de Europa y Canadácoincidía con una pérdida constantede P-diéster. Ahora bien, lasrelaciones proporcionalmente másaltas de P-diéster y de fosfonatos sehan encontrado en sueloshidromórficos, los cuales presentanuna actividad microbiana reducida(Condron et al. 1990; Zech et al.1987). Por consiguiente, losresultados obtenidos mediante elanálisis de RMN con 31P en elpresente estudio deberíanrelacionarse con otros estudiosrealizados en los suelos deCarimagua.
Oberson et al. (1995) aportaronpruebas evidentes de que,combinando una aplicación baja defertilizantes con el establecimiento deun pastizal mejorado, se modificabanlos parámetros químicos del suelo yse favorecía su actividad biológica.Estos autores concluyeron que el Porgánico de los residuos vegetales yde las deposiciones del ganado fuereciclado con más eficiencia en lospastizales mejorados que en lasabana nativa, habida cuenta de laactividad relativamente alta de lafosfatasa y del alto valor de las
reservas de P encontradas.Asimismo, la concentración y ladistribución de los carbohidratos yde la lignina en los agregados dediferente tamaño indicaron que laMO estaba más disponible para lasbacterias, como fuente de C y deenergía, en los pastizales que en lasabana nativa. Guggenberger et al.(1995) y Rao et al. (1994)encontraron tasas potencialescrecientes de mineralización del N enel pastizal cubierto con gramínea yleguminosa. Se sugiere, por tanto,que la concentración más alta, conrespecto a la de la sabana nativa, delas especies de P orgánico lábilencontradas en los pastizalesmejorados en el presente estudioindica que la situación nutricional delas poblaciones microbianas y de lamacrofauna del suelo han mejoradoy que se intensificaron, por ello, elciclo de los nutrientes y el de la MOdel suelo.
Los resultados de este estudiomostraron también que la actividadde las lombrices de tierra tuvo unfuerte efecto en la disponibilidad delP en los Oxisoles estudiados, y pudohaber causado un efecto mayor alaumentar las reservas del P activo.La biomasa debida a las lombricesen el pastizal de gramínea conleguminosa fue dos veces superior ala encontrada en el pastizal degramínea sola, la cual fue, por suparte, cinco veces mayor que laregistrada en la sabana nativa(Decaëns et al. 1994). En las hecesde las lombrices se detectó menorafinidad por el P inorgánico(experimento de sorción) que en elsuelo que las circundaba, lo quesugiere una disponibilidad más altade P en las heces. Asimismo, laconcentración del P extraíble conálcali fue mucho mayor en las hecesque en el suelo testigo, destacándosemás en esto las especies de Porgánico lábil de los ácidos húmicos.Lavelle y Martin (1992) reportaron

346
El Arado Natural:…
también un aumento en ladisponibilidad de P a corto plazo,gracias a la actividad de laslombrices.
Las heces de las lombrices quecomponen las comunidades de lasabana neotropical son estructurasresistentes (Lavelle y Martin 1992).La acumulación de las reservas de Porgánico lábil se debió a suestabilización física en el interior delas heces, puesto que la inclusiónfísica de un material orgánico enmicroagregados, a los cuales losmicroorganismos no tienen acceso, esel principal control ejercido sobre losprocesos de descomposición en losque intervienen las bacterias (vanVeen y Kuikman 1990). Lavelle yMartin (1992) señalaron que estaestabilización física de la MO delsuelo en las estructuras compactasde las heces de las lombrices es unafase intermedia porque, a largo plazo,la actividad de las lombrices tiende aacelerar el ciclo de la MO y de losnutrientes asociados con ella, procesoen que gran parte de esta MO se hallaen forma activa más que en formapasiva.
En las heces de las lombrices, losfulvatos son elementos constitutivosimportantes para la sorción de P,dado que se han halladoconcentraciones altas de P inorgánicoen los ácidos fúlvicos. Lavelle yMartin (1992) encontraron que el Pinorgánico intercambiable era másabundante en los suelos ingeridos porlas lombrices que en aquéllos que nolo fueron. Probablemente, el Pinorgánico unido a los ácidos fúlvicosestá más disponible que el que hasido fijado por los óxidos e hidróxidosde Fe y Al.
AgradecimientosLos autores agradecen a Tanja Gonterpor los análisis químicos en húmedo
del P y a Iris Zimmermann por lapreparación de los extractosalcalinos. Están también altamenteagradecidos con Astrid Oberson porsus valiosos comentarios sobre elmanuscrito.
ReferenciasBall-Coelho B; Salcedo H; Tiessen H;
Stewart JWB. 1993. Short- andlong-term phosphorus dynamics ina fertilized ultisol under sugarcane.Soil Sci Soc Am J 57:1027-1034.
Bedrock CN; Cheshire MV; Chudek JA;Fraser AR; Goodman BA; ShandCA. 1995. Effect of pH onprecipitation of humic acid frompeat and mineral soils on thedistribution of phosphorus forms inhumic and fulvic acid fractions.Commun Soil Sci Plant Anal26:1411-1425.
Bowman RA. 1988. A rapid method todetermine total phosphorus insoils. Soil Sci Soc Am J52:1301-1304.
Bowman RA; Cole CV. 1978. Anexploratory method forfractionation of organic phosphorusfrom grassland soils. Soil Sci125:95-101.
Condron LM; Frossard E; Tiessen H;Newman RH; Stewart JWB. 1990.Chemical nature of organicphosphorus in cultivated anduncultivated soils under differentenvironmental conditions. J Soil Sci41:41-50.
Dalal RC. 1977. Soil organic phosphorus.Adv Agron 29:83-117.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein, G. 1994.Inpact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30:157-168.
Dormaar JF. 1972. Seasonal patterns ofsoil organic phosphorus. Can J SoilSci 52:107-112.

Determinación del Nivel de Fósforo Orgánico...
347
Gerke J; Hermann R. 1992. Adsorption oforthophosphate to humic-Fe-complexes and to amorphous Fe-oxides. Z Pflanzenernaerh Bodenkd155:233-236.
Guggenberger G; Zech W; Thomas RJ.1995. Lignin and carbohydratealteration in particle-size separatesof an Oxisol under tropical pasturesfollowing native savanna. Soil BiolBiochem 27:1629-1638.
Guggenberger G; Haumaier L; ThomasRJ; Zech W. 1996. Assessing theorganic phosphorus status of anOxisol under tropical pasturesfollowing native savanna using 31PNMR spectroscopy. Biol Fertil Soils23:332-339.
Hawkes GE; Powlson DS; Randall EW;Tate KR. 1984. A 31P nuclearmagnetic resonance study of thephosphorus species in soils fromlong continued field experiments.J Soil Sci 35:35-45.
Hedley MJ; Stewart JWB; Chauhan BS.1982. Changes in inorganic andorganic soil phosphorus fractionsinduced by cultivation practicesand by laboratory incubations. SoilSci Soc Am J 46:970-976.
Hinedi ZR; Chang AC; Lee RWK. 1988.Mineralization of phosphorus insludge-amended soils monitored byphosphorus-31-nuclear magneticresonance spectroscopy. Soil SciSoc Am J 52:1593-1596.
Jiménez JJ; Decaëns T; Moreno AG; RossiJ.-P.; Lavelle P. 1994. Dynamicsand short-term effects ofearthworms in natural andmanaged savannas of the easternplains of Colombia. CIAT Report,Cali, Colombia.
Juo ASR; Fox RL. 1977. Phosphatesorption characteristics of somebench-mark soils of West Africa.Soil Sci 124:370-376.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humidtropics. Soil Biol Biochem24:1491-1498.
Newman RH; Tate KR. 1980. Soilphosphorus characterization by 31Pnuclear magnetic resonance.Commun Soil Sci Plant Anal11:835-842.
Oberson A; Friesen DK; Tiessen H; MoirJO; Borrero G. 1995. Phosphorustransformations in improvedpastures. En: CIAT, ed. Tropicallowlands program annual report1994. Documento de trabajono. 148. CIAT, Cali, Colombia.p 182-187.
Olsen SR; Sommers LE. 1982.Phosphorus. En: Page AL; MillerRH; Keeney DR, eds. Methods ofsoil analysis, Part 2. 2a. ed. ASA-SSSA, Madison, WI, EstadosUnidos. p 404-430.
Rao IM; Ayarza MA; Thomas RJ. 1994.The use of carbon isotope ratios toevaluate legume contribution to soilenhancement in tropical pastures.Plant Soil 162:177-182.
Stewart JWB; Tiessen H. 1987. Dynamicsof soil organic phosphorus.Biogeochemistry 4:41-60.
Tate KR; Newman RH. 1982. Phosphorusfractions of a climosequence of soilsin New Zealand tussock grassland.Soil Biol Biochem 14:191-196.
Tiessen H; Salcedo IH; Sampaio EVSB.1992. Nutrient and soil organicmatter dynamics under shiftingcultivation in semi-aridnortheastern Brazil. Agric EcosystEnviron 38:139-151.

348
El Arado Natural:…
Thomas RJ; Fisher MJ; Ayarza MA; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. LewisPublishers, Boca Ratón, FL,Estados Unidos. p 61-83.
Van Veen JA; Kuikman PJ. 1990. Soilstructural aspects of decompositionof organic matter by micro-organisms. Biogeochemistry11:213-233.
Zech W; Alt HG; Haumaier L; Blasek R.1987. Characterization ofphosphorus fractions in mountainsoils of the Bavarian Alps by 31PNMR spectroscopy. ZPflanzenernaerh Bodenkd150:119-123.

Alteración de la Lignina y de los Carbohidratos...
349
ResumenLos pastizales mejoradosestablecidos en los suelos ácidos delas sabanas tropicales puedenincrementar tanto el contenido de lamateria orgánica (MO) como lafertilidad de un suelo. El objetivo deeste estudio fue seguir los cambioscuantitativos y cualitativos de la MOasociada a las fracciones de arenas,de limos y de arcillas en los suelos delas sabanas isohipertérmicas deColombia, después de establecer enellos pastizales de gramínea sola(Brachiaria decumbens) y pastizalesde gramínea con leguminosa(B. decumbens + Puerariaphaseoloides). Se investigaron,asimismo, las heces de las lombricesanécicas y endo-anécicasprovenientes del pastizal quecontenía la leguminosa. Sedeterminaron el C, el N, los fenolesderivados de la lignina (oxidaciónalcalina con CuO) y loscarbohidratos (hidrólisis ácida con
ácido trifluoracético 4 M), en el suelototal y en sus agregados.
El contenido de C se incrementó,desde la sabana hasta el pastizal conleguminosa, en el siguiente orden (eng/kg de fracción de suelo): enarenas (1.6 a 3.3) < en limos (25.8 a32.1) < en arcillas (38.8 a 43.0);además, la relación C:N disminuyóen el mismo orden. Después de15 años contados desde laintroducción del pastizal, la cantidadde C se incrementó, respecto a losvalores obtenidos en la sabananativa, en los siguientes factores:2 (fracción de arenas), 1.3 (fracciónde limos) y 1.1 (fracción de arcillas).El descenso observado en laproducción de compuestos deoxidación de CuO de tipo fenólico y elincremento en la oxidación de lacadena lateral de la ligninaremanente indicaron que hay unaalteración progresiva de la ligninarelacionada con la disminución en eltamaño de las partículas del suelo.En el suelo de la sabana se encontróuna cantidad menor de fragmentosintactos de lignina que en el suelo delos dos tipos de pastizal. Losfactores de enriquecimiento, E =(mg/kg de suelo del pastizal / mg/kgde suelo de la sabana), de los fenolesderivados de la lignina tras elestablecimiento de los pastizales deB. decumbens y B. decumbens +
CAPÍTULO 21
Alteración de la Lignina y de losCarbohidratos en los Agregados de unOxisol, en Pastizales TropicalesDerivados de la Sabana Nativa1
G. Guggenberger* , W. Zech* y R. J. Thomas**
1. Publicado en Soil Biology and Biochemistry27, p 1629-1638, por Guggenberger et al.Copyright © 1995 de Elsevier. Impreso conpermiso de la casa editorial.
* Institute of Soil Science and Soil Geography,University of Bayreuth, D-95440 Bayreuth,Alemania.
** Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.

350
El Arado Natural:…
P. phaseoloides fueron,respectivamente, 3.9 y 2.7, 4.3 y 2.4y 2.6 y 2.6 para las respectivasfracciones de arenas, de limos y dearcillas. La MO asociada con lasarcillas tenía azúcares derivados dela actividad microbiana, mientrasque los azúcares encontrados en laMO asociada con las arenasprovenían de las plantas. La MOasociada con la fracción de limospresentó, por otro lado,concentraciones bajas de ambasclases de azúcar. El establecimientodel pastizal de gramínea y del quetiene leguminosa incrementó,respectivamente, la cantidad deazúcares fácilmente degradables enlos siguientes factores: 3.0 y 3.5 (enla fracción de arenas), 1.9 y 1.3 (enla fracción de limos) y 1.1 y 1.1 (en lafracción de arcillas). La distribuciónde la MO en los agregados del suelosugiere que hubo, tras laintroducción de los pastizales, unaumento en la cantidad de la macro-MO no protegida. Al mismo tiempo,las trazas de lignina y decarbohidratos indicaron nivelesbastante altos de MO vegetal sindescomponer, en especial en lafracción de arenas. Se concluyó queel aumento de esta fracción de la MOtiene efectos benéficos en el ciclointerno del N, en lo que concierne ala mineralización e inmovilización deéste. En comparación con el sueloque las circunda, las heces de laslombrices se caracterizaron por unenriquecimiento muy grande, en lafracción arenosa, en C, en lignina yen azúcares derivados de las plantas.Estos resultados sugieren que lahojarasca fresca se incorpora, demanera notoria, a la parte mineraldel suelo (efecto anécico), que losprocesos de descomposición adoptanun ritmo lento en las heces, y que sedesarrolla una reserva de MO en elsuelo que, aunque relativamentelábil, está protegida físicamente enlas heces.
Palabras clave: agregados, heces,lombrices, materia orgánica delsuelo, nitrógeno, pastizales,suelos de sabana
IntroducciónLa materia orgánica (MO) del suelose compone de un variado conjuntode residuos vegetales, animales ymicrobianos, en todos los estadosposibles de descomposición debido ala compleja interacción de losprocesos biológicos, químicos yfísicos que se desarrollan en el suelo(Swift et al. 1979). Por cuestiones deanálisis funcional, en la MO delsuelo se clasifican unas pocasentidades, y las diferentes tasas dedescomposición de dicha MO seinterpretan conceptualmente comomodelos matemáticos queconsideran la cinética de losdiferentes compartimentos(Jenkinson y Rayner 1977; Partonet al. 1987).
Los cambios en las prácticasagrícolas de manejo de los cultivosinfluyen en la cantidad de la MOpresente en el suelo y en su tasa dedescomposición. En las revisionesde literatura realizadas, se halló lasugerencia de que la pérdida de MOque sigue a la tala del bosquetropical y a su conversión en pastizalfluctúa entre un 20% y un 50%(Bouwman 1990; Sombroek et al.1993). Las prácticas relacionadascon el uso dado a la tierra afectantambién la distribución de la MO delsuelo entre su fracción lábil y sufracción estable (Cambardella yElliott 1992; Christensen 1992;Tiessen y Stewart 1983). Las laboresdel cultivo disminuyen, de maneramucho más notoria, la cantidad deMO en el suelo, en especial en lafracción lábil. La eliminación de estafracción lábil y el enriquecimientorelativo en materiales biológicamenteresistentes, conduce a un suministro

Alteración de la Lignina y de los Carbohidratos...
351
menor de nutrientes de tipoorgánico, en especial los de N y losde P. La reducción en la fertilidaddel suelo asociada con la situaciónanterior es, probablemente, mayorque la hallada cuando se compara laMO total de un suelo virgen y de otrocultivado (Tiessen y Stewart 1983).
Los pastizales son un sistema deuso de la tierra que se ha extendidoenormemente tras la deforestaciónde los ecosistemas tropicales y lasiembra en sus suelos de cultivosintroducidos (Sombroek et al. 1993).Hay indicios de que elestablecimiento de pastizales ensuelos de fertilidad baja, como los delas sabanas ácidas, y en especialcuando se emplean especies queproducen niveles altos de biomasaradicular, pueden aumentar lacantidad de MO y mejorar lafertilidad del suelo (Serrão et al.1979; Sombroek et al. 1993; Thomaset al. 1995). Estos pastizalestropicales se basan, a menudo, enmezclas de gramíneas y leguminosasforrajeras. Rao et al. (1994)emplearon la relación de isótopos delC para evaluar la contribución de lasleguminosas al mejoramiento de lacalidad del suelo. Estos autoresdemostraron que la crecientecontribución de MO derivada de lasleguminosas está ligada a unincremento de la fertilidad del suelo,como lo indica el aumento de lastasas potenciales de mineralizacióndel N.
El fraccionamiento del suelosegún el tamaño de sus agregados(arenas, limos y arcillas) generafracciones órgano-minerales cuyaspropiedades se diferencian mucho delas que caracterizan el ciclo de la MO(Anderson y Paul 1984; Tiessen yStewart 1983). Algunos estudioshan demostrado que la MO del sueloasociada a la fracción de arenas esimportante en los ciclos a corto plazode la MO, mientras que la fracción de
arcillas domina en los de plazointermedio, y la de limos participa enlos de largo plazo (Christensen1992). Guggenberger et al. (1994)demostraron la utilidad de laoxidación de CuO de tipo alcalino yde la hidrólisis ácida paradeterminar, respectivamente, laalteración de la lignina y de loscarbohidratos, y esto desde losagregados más gruesos hasta losmás finos y después de un cambioen el sistema de uso de la tierra.
El objetivo de este estudio fueseguir los cambios cuantitativos ycualitativos de la MO del suelo en elhorizonte A de un Oxisol, despuésdel establecimiento de un pastizal degramíneas introducidas en un áreade sabana nativa. Se empleó laseparación de los agregados segúnsu tamaño para calcular la cantidadde MO libre en el suelo en partículas(en la fracción de arenas) y la de MOestabilizada por la asociación con lasfracciones de limos y de arcillas. Seemplearon la oxidación del CuO detipo alcalino y la hidrólisis de loscarbohidratos con ácidotrifluoroacético (ATF) para detectar laalteración de la lignina y de loscarbohidratos. Los datos sediscutieron respecto a ladisponibilidad del C y a los efectosque puede ejercer el fenómenoestudiado en el ciclo del N del suelo.
Materiales y Métodos
Descripción del sitioEl área del estudio se encuentra enel Centro Nacional de InvestigacionesAgropecuarias (CNIA) Carimagua(4°37’ N, 71°19’ O, 175 msnm), enColombia. La temperatura mediaanual es de 26 °C y la precipitaciónmedia anual es de unos 2200 mm,que se distribuyen desde abril hastadiciembre. El área es representativa

352
El Arado Natural:…
de las sabanas isohipertérmicas biendrenadas, donde predominan losOxisoles de fertilidad baja. Estossuelos se caracterizan por un pHbajo (4.5, en agua), 96% desaturación de Al, y cantidadespequeñas de cationesintercambiables, como los de Ca, Mgy K (Lascano y Estrada 1989). Losfactores que contribuyen a la escasafertilidad de estos suelos ácidos y losefectos que causan en el crecimientode las plantas son complejos; entreellos figuran la toxicidad debida alAl, los niveles bajos de P disponible,y las tasas bajas de mineralizacióndel N (Rao et al. 1993).
En 1978 se sembraron tanto lagramínea tropical Brachiariadecumbens cv. Basilisk como laleguminosa Pueraria phaseoloidesCIAT 9900 (kudzú), sola o enasociación, en un terreno dondeantes había sólo vegetación desabana. Ambas especies están bienadaptadas a las condiciones de lossuelos ácidos. Se aplicaron lassiguientes dosis de fertilizante(kg/ha) durante 15 años (1978-1993)en los pastizales establecidos: 0 N,103 P, 63 K, 648 Ca, 56 Mg y 77 Sen el de gramínea sola, y 0 N, 106 P,80 K, 648 Ca, 63 Mg y 90 S en el degramínea asociada con laleguminosa. La caracterización delas variables físicas y químicas (porejemplo, la textura, los óxidos ehidróxidos de Al o de Fe) del sueloconfirmaron que el origen de éste erasimilar en todos los tratamientos.
El establecimiento del pastizalintroducido ejerció un efectosignificativo en la abundancia de lamacrofauna del suelo. La biomasade lombrices en la sabana nativa, enel pastizal de gramínea sola y en elpastizal en que se asocian gramíneay leguminosa fueron,respectivamente, 4.78, 22.95 y51.09 g PF/m2, respectivamente(Decaëns et al. 1994). J.J. Jiménez
estimó una ingestión de 680 Mgsuelo/ha en el pastizal deB. decumbens + P. phaseoloides. Lasespecies de lombriz de tierra quepredominaron fueron Martiodrilussp., de tipo anécico, y Andiorrhinussp., que es endo-anécica, ambaspertenecientes a la familiaGlossoscolecidae.
Muestreo del sueloEn 1993 y tras 15 años de pastoreo,se tomaron muestras del suelo en lasabana nativa, en el pastizal degramínea y en el de gramínea conleguminosa. En cada tratamiento seextrajeron, manualmente, cuatromuestras al azar. Se tomaronmuestras solamente del horizontemineral A, puesto que el contenidode C del pastizal era muy alto en lasuperficie del suelo y descendíagradualmente según la profundidad.Rao et al. (1994) informaron que el Cderivado del pastizal introducidoconstituía el 29% del C orgánico enel intervalo de 0 a 2 cm deprofundidad y menos del 7% en loshorizontes inferiores. Por tanto, seesperaba obtener diferenciassignificativas en la distribución y enla composición del C orgánicosolamente en la superficie del suelo.Los cambios que se puedanpresentar a través del perfil delsuelo, especialmente los más sutilesen los horizontes profundos, puedentardar décadas y aun siglos endesarrollarse totalmente.
Se tomaron también muestras delas heces depositadas por laslombrices en la superficie del pastizalde B. decumbens + P. phaseoloides,en el que había gran cantidad deheces de tamaño considerable (2 a8 cm ∅ ). Se recogieron al azar20 heces superficiales en la parcelade 1 ha. Las muestras se llevaron allaboratorio en donde se secaron alaire, se tamizaron a una trama de

Alteración de la Lignina y de los Carbohidratos...
353
2 mm y, finalmente, se volvieron asecar a 40 °C durante 5 días. Losresultados obtenidos en los análisisquímicos se calcularon a partir demuestras que se habían secado a105 °C.
Aislamiento de las partículasdel suelo según su tamañoLa separación de las partículas desuelo por tamaños se realizó portriplicado en un material < 2 mm,según el método de Christensen(1985); las raíces visibles se retiraronantes del fraccionamiento.Empleando un desintegrador (“HeatSystems-Ultrasonics”, ModeloW 185-F) se dispersaron medianteultrasonido 30 g de suelo en 150 mlde agua. La energía total disipadafue de 1500 J/ml. Las fracciones dearcillas (< 2 µm) y de limos (2-20 µm)se separaron por sedimentacióngravitacional en agua y el sedimentoobtenido, tras el aislamiento de lasarcillas y los limos, constituyó lafracción de arenas (20-2000 µm).Los resultados del estudio seexpresan en materiales secados a40 °C. Antes del análisis, la fracciónde arenas se molió (a < 20 µm).
Análisis químicosLos contenidos totales de C y de Ndel suelo y de cada una de lasfracciones se determinaron con unanalizador de combustión seca(“Elementary Vario EL C:NAnalyzer”). Los coeficientes devariación entre las repeticiones de losanálisis fueron de ± 2%.
La cantidad y el estado dedescomposición oxidativa de lalignina en el suelo y en las fraccionesdel suelo se calcularon mediante elsistema de parámetros de la ligninaobtenidos de la oxidación alcalina deCuO (Ertel y Hedges 1984; Kögel-Knabner et al. 1991). El CuO se
empleó para oxidar muestras de[100(arcilla) – 1000(arena)] mg juntocon NaOH 2 M durante 2 h, a 170 °Cbajo una atmósfera de N2 (Hedges yErtel 1982). Los productos fenólicosde la oxidación de CuO fueronanalizados mediante cromatografíade gas capilar (a lo largo de unacolumna de sílice fundido OV-1)obteniendo derivados del trimetil-silil, en un aparato HP 5890equipado con un detector FID(“Flame Ionization Detector”). Elácido fenilacético se empleó comoestándar de control interno.
La suma de las unidades devanililo (V), siringilo (S) y cinamilo (C)(V+S+C) se tomó como un indicadorde la cantidad de residuos intactosde lignina. No se calculó la cantidadde lignina dentro de las muestrasporque la cantidad de fenoles querinde la lignina difiere según lasespecies vegetales (Sarkanen yLudwig 1971) y porque no existe,además, una forma independiente decalcular la cantidad de ligninacontenida en la MO del suelo (Kögel-Knabner et al. 1991). En cambio,fue más oportuno usar losrendimientos brutos de modocomparativo. Se consideraron lasrelaciones ácido-aldehído de lasunidades de vanililo (ac/al)V ysiringilo (ac/al)S para establecer elgrado de oxidación de la cadenarestante de lignina debido a lasbacterias. El coeficiente de variaciónobtenido en la determinación de unfenol a nivel individual fue de 8% yde 6% para las tasas de (ac/al)V y(ac/al)S que se calcularon (Kögel-Knabner et al. 1991).
La concentración y lacomposición de los carbohidratos sedeterminaron mediante el análisis delos monómeros de azúcar liberadosen la hidrólisis ácida. Sehidrolizaron muestras de 100 mg(arcillas) – 1000 mg (arenas) conácido trifluoroacético (ATF) a 100 °C

354
El Arado Natural:…
durante 4 h bajo una atmósferaenriquecida con N2. Losmonosacáridos liberados se separaronde los residuos húmicos y de loscationes con C activo (Charcoal) y conresina Dowex 50, respectivamente(Boas 1953). La derivación de losazúcares incluye, también, laoximación con O-metilhidroxylaminay la sililación con bis-trimetilsilil-trifluoroacetamida (Andrews 1989).Los monómeros de los azúcares seanalizaron en un cromatógrafo de gasHP 5890 equipado con una columnade sílice fundida HP-5 y un detectorselectivo de masa HP 5971A. Lacuantificación se logró mediante laintegración del área bajo los picos dela respuesta de los fragmentos demasa de características selectas,empleando el 1,3-difenoxibencenocomo elemento estándar de controlinterno. El coeficiente de variación enlos análisis de los monosacáridososciló entre el 1% y el 17% (J. Hejzlar,com. pers.).
Puesto que los carbohidratoscristalinos (la celulosa) no sondigeridos por el ATF, los azúcaresmonoméricos liberados por lahidrólisis ácida provinieron de lahemicelulosa de las plantas y de losmetabolitos de origen microbiano.Los polisacáridos encontrados en lascélulas vegetales se caracterizan porpresentar una relación alta,proporcionalmente, de las pentosas(azúcares con cinco átomos de C)arabinosa y xylosa, mientras que laspoblaciones microbianas sintetizan,principalmente, las hexosas (seisátomos de C) galactosa y manosa, losoxiazúcares ramnosa y fucosa y otrosazúcares menores (Moers et al. 1990).La cantidad proporcional decarbohidratos derivados de lasplantas y de la actividad microbianapueden calcularse a partir de larelación molar simple de losmonómeros del azúcar. Oades (1984)estableció que la relación
galactosa+manosa/arabinosa+xylosa [(G+M/A+X)] esbaja (< 0.5) en el caso de loscarbohidratos derivados de lasplantas y alta (> 2.0) en el caso delos polisacáridos bacterianos.Murayama (1984) usó la relacióncreciente ramnosa+fucosa/arabinosa+xylosa [(R+F)/(A+X)] parainvestigar la síntesis de loscarbohidratos bacterianos durantela descomposición de la paja.
Resultados
Carbono y nitrógenoLa cantidad de sólidos del sueloobtenidos por dispersiónultrasónica y sedimentacióngravitacional varió desde 973hasta 999 g/kg de suelo total(Cuadro 21-1). No hubo diferenciassignificativas entre los tratamientosen la fracción de arenas; por suparte, el suelo de los dos pastizalesintroducidos tenía un contenidomenor de limos y mayor de arcillasque el de la sabana nativa. Latextura global de los suelos indicó,en términos de peso, que éstos erancomparables. Las heces de laslombrices presentaban mayorcontenido de partículas de lafracción de limos que de la dearcillas.
La cantidad de C total del suelode sabana fue de 22.6 g/kg desuelo. El establecimiento delpastizal introducido de gramíneasola aumentó esta cantidad de Chasta 26.6 g/kg de suelo y el degramínea con leguminosa hasta26.0 g/kg de suelo. La mayorcantidad de C se registró, sinembargo, en las heces de laslombrices (56.3 g/kg) recolectadasen el pastizal de gramínea conleguminosa. La cantidad de C en lafracción de arenas fue
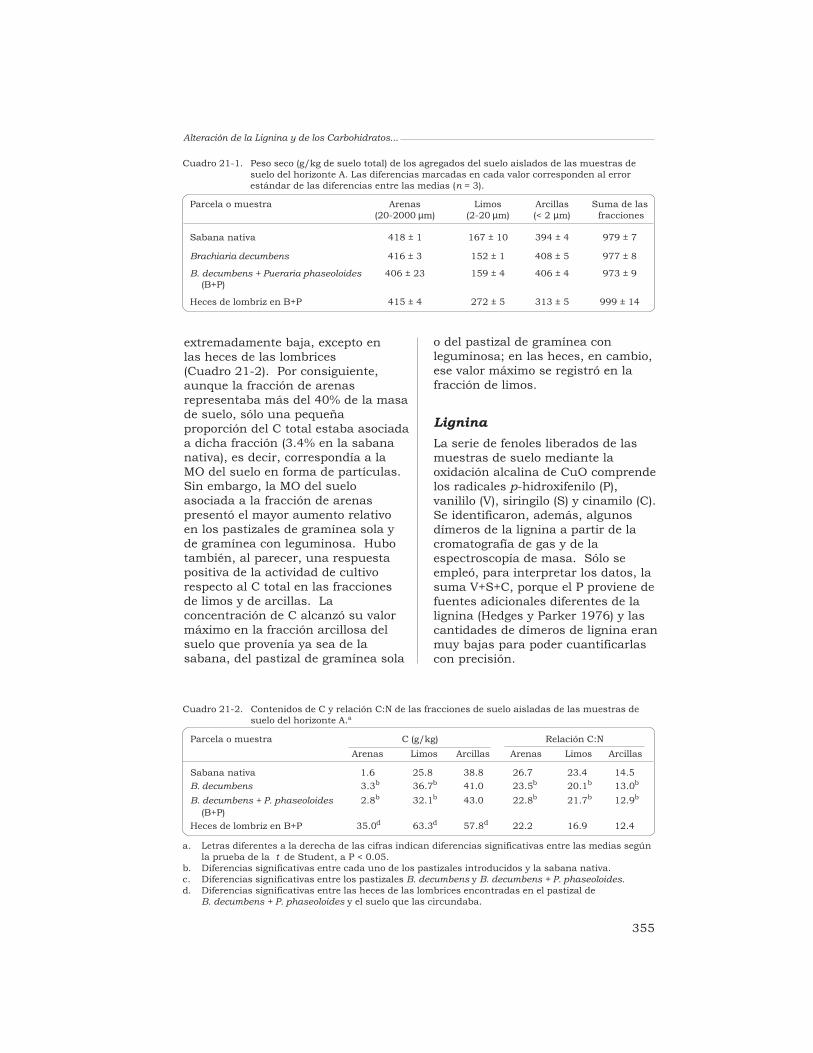
Alteración de la Lignina y de los Carbohidratos...
355
extremadamente baja, excepto enlas heces de las lombrices(Cuadro 21-2). Por consiguiente,aunque la fracción de arenasrepresentaba más del 40% de la masade suelo, sólo una pequeñaproporción del C total estaba asociadaa dicha fracción (3.4% en la sabananativa), es decir, correspondía a laMO del suelo en forma de partículas.Sin embargo, la MO del sueloasociada a la fracción de arenaspresentó el mayor aumento relativoen los pastizales de gramínea sola yde gramínea con leguminosa. Hubotambién, al parecer, una respuestapositiva de la actividad de cultivorespecto al C total en las fraccionesde limos y de arcillas. Laconcentración de C alcanzó su valormáximo en la fracción arcillosa delsuelo que provenía ya sea de lasabana, del pastizal de gramínea sola
o del pastizal de gramínea conleguminosa; en las heces, en cambio,ese valor máximo se registró en lafracción de limos.
LigninaLa serie de fenoles liberados de lasmuestras de suelo mediante laoxidación alcalina de CuO comprendelos radicales p-hidroxifenilo (P),vanililo (V), siringilo (S) y cinamilo (C).Se identificaron, además, algunosdímeros de la lignina a partir de lacromatografía de gas y de laespectroscopía de masa. Sólo seempleó, para interpretar los datos, lasuma V+S+C, porque el P proviene defuentes adicionales diferentes de lalignina (Hedges y Parker 1976) y lascantidades de dímeros de lignina eranmuy bajas para poder cuantificarlascon precisión.
Cuadro 21-2. Contenidos de C y relación C:N de las fracciones de suelo aisladas de las muestras desuelo del horizonte A.a
Parcela o muestra C (g/kg) Relación C:NArenas Limos Arcillas Arenas Limos Arcillas
Sabana nativa 1.6 25.8 38.8 26.7 23.4 14.5B. decumbens 3.3b 36.7b 41.0 23.5b 20.1b 13.0b
B. decumbens + P. phaseoloides 2.8b 32.1b 43.0 22.8b 21.7b 12.9b
(B+P)Heces de lombriz en B+P 35.0d 63.3d 57.8d 22.2 16.9 12.4
a. Letras diferentes a la derecha de las cifras indican diferencias significativas entre las medias segúnla prueba de la t de Student, a P < 0.05.
b. Diferencias significativas entre cada uno de los pastizales introducidos y la sabana nativa.c. Diferencias significativas entre los pastizales B. decumbens y B. decumbens + P. phaseoloides.d. Diferencias significativas entre las heces de las lombrices encontradas en el pastizal de
B. decumbens + P. phaseoloides y el suelo que las circundaba.
Cuadro 21-1. Peso seco (g/kg de suelo total) de los agregados del suelo aislados de las muestras desuelo del horizonte A. Las diferencias marcadas en cada valor corresponden al errorestándar de las diferencias entre las medias (n = 3).
Parcela o muestra Arenas Limos Arcillas Suma de las(20-2000 µm) (2-20 µm) (< 2 µm) fracciones
Sabana nativa 418 ± 1 167 ± 10 394 ± 4 979 ± 7
Brachiaria decumbens 416 ± 3 152 ± 1 408 ± 5 977 ± 8
B. decumbens + Pueraria phaseoloides 406 ± 23 159 ± 4 406 ± 4 973 ± 9(B+P)
Heces de lombriz en B+P 415 ± 4 272 ± 5 313 ± 5 999 ± 14

356
El Arado Natural:…
En el suelo de la sabana nativase registró la menor cantidad deunidades de V+S+C (3.5 g/kg C),mientras que en los pastizales losvalores observados fueron 11.5 g/kgC (en B. decumbens) y 9.7 g/kg C (enB. decumbens + P. phaseoloides)(Cuadro 21-3). Las heces de lombrizdepositadas en el pastizal degramínea con leguminosapresentaron los valores más altos deV+S+C (19.2 g/kg C). La suma delos productos de oxidación de CuOdisminuyó en el orden siguiente:arenas > limos > arcillas. Como elsuelo total, también los agregadosdel suelo de la sabana nativapresentaron la cantidad más bajaobservada de V+S+C. Una vezestablecido un pastizal, el mayorincremento en los productos de laoxidación fenólica de CuO se produjoen la fracción de arenas. Aunque lasuma V+S+C en los agregadoscontenidos en las heces no fuesignificativamente mayor que en losrespectivos agregados del suelo quelas circundaba, las mediciones en lasmuestras de suelo total arrojaronvalores mucho más altos que losanteriores en los productos de laoxidación fenólica de CuO de lasheces. Estos resultados se explicanporque la MO asociada a la fracciónde arenas contribuye, en gran parte,
al total de MO del suelo presente enlas heces (Cuadro 21-2).
Las relaciones ácido:aldehído delos radicales vanililo y siringilo,[(ac/al)V, (ac/al)S] muestran el gradode alteración microbiana de lalignina, y el aumento en la relación(ac/al)V refleja directamente unincremento en la oxidación de lacadena lateral del radicalfenilpropano (Hedges et al. 1988).En todos los suelos analizados, ladisminución del tamaño de laspartículas del suelo trajo consigo unaumento de las relaciones (ac/al)V y(ac/al)S (Cuadro 21-4), lo que indicaun incremento en la degradación dela lignina según el orden siguiente:arenas > limos > arcillas. En todaslas fracciones de suelo analizadas,las relaciones (ac/al)V y (ac/al)S másaltas se observaron en la sabananativa. En los pastizales, los valoresde estas relaciones fueron menoresque los anteriores y fueron los másbajos en las heces analizadas.
En la Figura 21-1 se muestranlos cambios de la lignina respecto ala tasa C:N. Según la Figura 21-1A,el valor generado por la suma V+S+Cdisminuía cuando la relación C:N sereducía, es decir, cuando aumentabala descomposición de la MO delsuelo. Al mismo tiempo, la oxidación
Cuadro 21-3. Rendimiento (expresado como la suma de los radicales V, S y C) de los productos deoxidación de CuO, en la MO del suelo asociada con las fracciones aisladas de muestrastomadas en el horizonte A del suelo.
Parcela o muestra V + S + C (g/kg C) en:Arenas Limos Arcillas Suma de las Suelo
fracciones total
Sabana nativa 24.8 5.8 2.8 3.9 3.5B. decumbens 48.2b 19.1b 6.6b 10.9b 11.5b
B. decumbens + P. phaseoloides 39.9b 13.7b 4.9b,c 7.6b 9.7b
(B+P)Heces de lombriz en B+P 44.3 18.1 6.8 19.1d 19.2d
a. Letras diferentes a la derecha de las cifras indican diferencias significativas entre las medias segúnla prueba de la t de Student, a P < 0.05.
b. Diferencias significativas entre cada uno de los pastizales introducidos y la sabana nativa.c. Diferencias significativas entre los pastizales B. decumbens y B. decumbens + P. phaseoloides.d. Diferencias significativas entre las heces de las lombrices encontradas en el pastizal de
B. decumbens + P. phaseoloides y el suelo que las circundaba.

Alteración de la Lignina y de los Carbohidratos...
357
de la cadena lateral de la ligninaremanente, es decir, la relación(ac/al)V, aumentó al disminuir larelación C:N (Figura 21-1B). Ahorabien, al comparar los diferentessistemas de uso de la tierra, losparámetros de la lignina sugierenque la descomposición de éstaalcanza un estado más avanzadocuando aumenta la relación C:N, enel siguiente orden:
heces de lombrices en el pastizalde B. decumbens + P.phaseoloides <
Cuadro 21-4. Relaciones (ac/al)v y (ac/al)s de la MO del suelo asociada con las fracciones de sueloaisladas de las muestras recolectadas en el horizonte A.a
Parcela o muestra (ac/al)v (ac/al)sArenas Limos Arcillas Arenas Limos Arcillas
Sabana nativa 0.77 1.33 1.54 1.27 1.56 1.95 B. decumbens 0.45 1.16 1.21 0.66b 1.00b 1.42b
B. decumbens + P. phaseoloides 0.39b 0.80b 1.10b 0.58b 0.84b 0.88b,c
(B+P)Heces de lombriz en B+P 0.22 0.47d 0.62d 0.33d 0.62 0.84
a. Letras diferentes a la derecha de las cifras indican diferencias significativas entre las medias segúnla prueba de la t de Student, a P < 0.05.
b. Diferencias significativas entre cada uno de los pastizales introducidos y la sabana nativa.c. Diferencias significativas entre los pastizales B. decumbens y B. decumbens + P. phaseoloides.d. Diferencias significativas entre las heces de las lombrices encontradas en el pastizal de
B. decumbens + P. phaseoloides y el suelo que las circundaba.
0 10 20 30 40 50 0 0.4 0.8 1.2 1.6
30
25
20
15
10
5
0
V+S+C (g/kg)
A B
(ac/al)v
Figura 21-1. (A) Cambios en el rendimiento de los productos de oxidación de CuO (V+S+C) y(B) cambios en las relaciones (ac/al)V al aumentar la descomposición, desde la fracción dearenas hasta la de arcillas, en muestras de suelo tomadas en el horizonte A. = sabananativa; = Brachiaria decumbens; = B. decumbens + Pueraria phaseoloides. = hecesde lombrices en B. decumbens + P. phaseoloides.
C/N
Arenas
Limos
Arcillas
Arenas
Limos
Arcillas
< pastizal de B. decumbens+ P. phaseoloides B. <
< pastizal de B. decumbens< sabana nativa.
CarbohidratosLa concentración de loscarbohidratos del suelo recolectadoen la sabana nativa, en el pastizal degramínea y en el pastizal de gramíneaasociada con leguminosa fueronsimilares (190 g/kg C,aproximadamente), y fue un poco
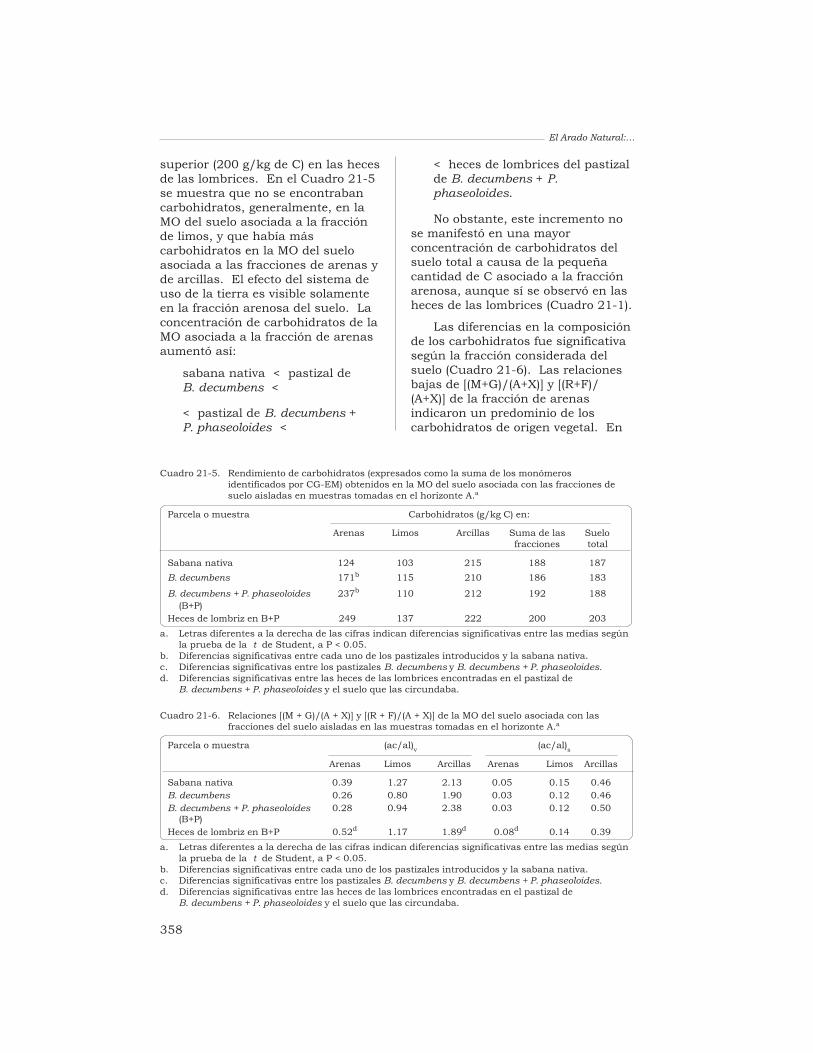
358
El Arado Natural:…
superior (200 g/kg de C) en las hecesde las lombrices. En el Cuadro 21-5se muestra que no se encontrabancarbohidratos, generalmente, en laMO del suelo asociada a la fracciónde limos, y que había máscarbohidratos en la MO del sueloasociada a las fracciones de arenas yde arcillas. El efecto del sistema deuso de la tierra es visible solamenteen la fracción arenosa del suelo. Laconcentración de carbohidratos de laMO asociada a la fracción de arenasaumentó así:
sabana nativa < pastizal deB. decumbens <
< pastizal de B. decumbens +P. phaseoloides <
< heces de lombrices del pastizalde B. decumbens + P.phaseoloides.
No obstante, este incremento nose manifestó en una mayorconcentración de carbohidratos delsuelo total a causa de la pequeñacantidad de C asociado a la fracciónarenosa, aunque sí se observó en lasheces de las lombrices (Cuadro 21-1).
Las diferencias en la composiciónde los carbohidratos fue significativasegún la fracción considerada delsuelo (Cuadro 21-6). Las relacionesbajas de [(M+G)/(A+X)] y [(R+F)/(A+X)] de la fracción de arenasindicaron un predominio de loscarbohidratos de origen vegetal. En
Cuadro 21-5. Rendimiento de carbohidratos (expresados como la suma de los monómerosidentificados por CG-EM) obtenidos en la MO del suelo asociada con las fracciones desuelo aisladas en muestras tomadas en el horizonte A.a
Parcela o muestra Carbohidratos (g/kg C) en:
Arenas Limos Arcillas Suma de las Suelofracciones total
Sabana nativa 124 103 215 188 187B. decumbens 171b 115 210 186 183
B. decumbens + P. phaseoloides 237b 110 212 192 188(B+P)
Heces de lombriz en B+P 249 137 222 200 203a. Letras diferentes a la derecha de las cifras indican diferencias significativas entre las medias según
la prueba de la t de Student, a P < 0.05.b. Diferencias significativas entre cada uno de los pastizales introducidos y la sabana nativa.c. Diferencias significativas entre los pastizales B. decumbens y B. decumbens + P. phaseoloides.d. Diferencias significativas entre las heces de las lombrices encontradas en el pastizal de
B. decumbens + P. phaseoloides y el suelo que las circundaba.
Cuadro 21-6. Relaciones [(M + G)/(A + X)] y [(R + F)/(A + X)] de la MO del suelo asociada con lasfracciones del suelo aisladas en las muestras tomadas en el horizonte A.a
Parcela o muestra (ac/al)v (ac/al)s
Arenas Limos Arcillas Arenas Limos Arcillas
Sabana nativa 0.39 1.27 2.13 0.05 0.15 0.46B. decumbens 0.26 0.80 1.90 0.03 0.12 0.46B. decumbens + P. phaseoloides 0.28 0.94 2.38 0.03 0.12 0.50
(B+P)Heces de lombriz en B+P 0.52d 1.17 1.89d 0.08d 0.14 0.39
a. Letras diferentes a la derecha de las cifras indican diferencias significativas entre las medias segúnla prueba de la t de Student, a P < 0.05.
b. Diferencias significativas entre cada uno de los pastizales introducidos y la sabana nativa.c. Diferencias significativas entre los pastizales B. decumbens y B. decumbens + P. phaseoloides.d. Diferencias significativas entre las heces de las lombrices encontradas en el pastizal de
B. decumbens + P. phaseoloides y el suelo que las circundaba.

Alteración de la Lignina y de los Carbohidratos...
359
cambio, estas relaciones fueronaltas en las fracciones de limos y dearcillas, e indicaron así la presenciade carbohidratos sintetizados porlas bacterias. Las relaciones[(M+G)/(A+X)] y [(R+F)/(A+X)] fueronalgo menores en las fracciones dearenas y de limos de los dospastizales introducidos que en lasfracciones correspondientes de lasabana nativa, lo que ilustra lagran importancia de las plantas enla presencia de los carbohidratosdetectados en las señales. En lafracción de arenas de las heces delas lombrices se encontró mayorcantidad de azúcares microbianosque en la respectiva fracción de lasdemás muestras.
DiscusiónEl objetivo de este estudio fueinvestigar las posibles diferenciasexistentes entre la composición delos carbohidratos y de la lignina delsuelo tras la introducción depastizales tropicales mejorados.Puesto que el ciclo de la MO delsuelo está muy influido por suasociación con las partículasminerales primarias (Anderson yPaul 1984), se trató de distinguir,ante todo, entre la MO del suelo queno había sido estabilizada por lasinteracciones con la matriz mineraly la MO que se hallaba dentro delos complejos órgano-minerales.Christensen (1992) señaló que laseparación de las partículas delsuelo por medio de la dispersiónultrasónica es un mecanismo útilpara separar la MO del suelo enforma de complejos órgano-minerales de arcillas y de limos, ycomo MO libre y reducida apartículas que se fraccionabanjunto con los minerales de lafracción de arenas. Este mismo
autor recomienda que la energíaadecuada para la dispersión delsuelo sea de 1500 J/ml; sinembargo, se desconoce aún lapotencia de sonda efectiva.
En la literatura científicaconsultada se discuten aún laslimitaciones de la dispersiónultrasónica del suelo. Una de lasprincipales limitantes asociadas aluso del ultrasonido (sonicación)para dispersar el suelo es la posibleredistribución, entre las fraccionesdel suelo, de la fracción ligera de laMO del suelo (Elliott y Cambardella1991). La MO libre y la fracciónligera de la MO puede permaneceren las fracciones de limos y dearcillas después de un tratamientoultrasónico. Ahora bien, los datosobtenidos sobre la composiciónquímica de la MO del sueloasociada a la fracción de limos y dearcillas no confirmaron laexistencia de una considerableredistribución de la fracción ligerade la MO. La MO asociada a lasfracciones del suelo de menortamaño aparece mucho másdegradada que la fracción ligera dela MO asociada a la fracción dearenas (Baldock et al. 1992;Guggenberger et al. 1994). Otradesventaja de la aplicación delultrasonido se relaciona con laredistribución del C microbiano.Los tratamientos ultrasónicos, aúnlos de acción limitada, liberan elcontenido de muchas célulasbacterianas (Ahmed y Oades 1984),que es adsorbido, probablemente,por las fracciones de limos y dearcillas. La biomasa microbiana noexcede, normalmente, el 5% de laMO del suelo (Jenkinson y Powlson1976); por consiguiente, laredistribución de la MO se limita auna parte proporcionalmentepequeña de la MO total.

360
El Arado Natural:…
Alteración de la cantidad y ladistribución de la MO totalLa reducción de la MO asociada a lafracción de arenas del suelo y elaumento de la MO asociada a lasfracciones de limos y de arcillasconforman un proceso que seobserva a menudo en el suelo(Christensen 1992). Enconsonancia con Christensen(1986), la MO del suelo asociada a lafracción arenosa consiste en macro-MO que no forma complejos órgano-minerales (= la fracción liviana de laMO) y, por tanto, es mucho mássusceptible a la mineralización quela MO unida a los limos y a lasarcillas. En comparación con lafracción arenosa de los suelos de laspraderas norteamericanas(Anderson et al. 1981; Tiessen yStewart 1983), la arena aislada delsuelo de las sabanasisohipertérmicas tenía una cantidadde MO mucho menor, lo quesubraya la importancia de lasasociaciones órgano-minerales en laestabilización de la MO en los suelostropicales. De acuerdo con Duxburyet al. (1989), las condicionesclimáticas cálidas y húmedasconducen a una mayor y máscompleta descomposición de losmateriales orgánicos que no esténprotegidos. En cambio, la MO delsuelo estabilizada en los complejosórgano-minerales fue menosafectada por los cambios dehumedad y de temperatura(Duxbury et al. 1989).
Es probable que la introducciónde B. decumbens y de B. decumbens+ P. phaseoloides en los pastizalesincremente el contenido de C en elsuelo, principalmente porque enellos se genera un volumenconsiderable de desechos de raíces(Serrão et al. 1979; Sombroek et al.1993). Muertas las raíces, entranen primer lugar en la fracciónarenosa porque son partículas
discretas. El cálculo delenriquecimiento en C (EC) tras laintroducción de los pastizales, esdecir:
indicó que ese contenido de C seduplicaba en la fracción de arenas(Cuadro 21-7). Durante los procesosque siguen a la descomposición yhumificación de la hojarasca, haypartes de la MO del suelo quepueden formar complejos con losminerales de la fracción de arcillas yde limos (Anderson y Flanagan 1989;Baldock et al. 1992). No se produjoun aumento significativo en elcontenido de C del complejo dearcillas (EC = 1.1) 15 años despuésde la introducción del pastizal,puesto que la cantidad de C en lafracción de arcillas era relativamentealta y la transformación de losresiduos vegetales frescos encomplejos órgano-minerales eralenta.
La actividad de las lombrices detierra tuvo un efecto considerable enel proceso de redistribución de la MOen el suelo; la fracción de arenas delas heces, especialmente, era másabundante en C que el suelo que lascircundaba (EC = 12.8). Puesto quelas especies de lombrices quepredominaban en el pastizalpertenecían a las categorías anécicay endo-anécica —las cuales sealimentan, al menos en parte, dehojarasca aunque vivan en galeríasconstruidas en el suelo (Lee 1985)—esta hojarasca se mezclóíntimamente con la fracción mineraldel suelo en el tracto digestivo de laslombrices. De acuerdo con E.Blanchart [tesis doctoral sinpublicar, Universidad de Rennes I,1990, citado en Lavelle y Martin(1992)], las heces grandesdepositadas por las lombrices en las
EC = g C/kg de fracción del sueloo de suelo total de la sabana
g C/kg de fracción del sueloo de suelo total del pastizal
(Ecuación 1)
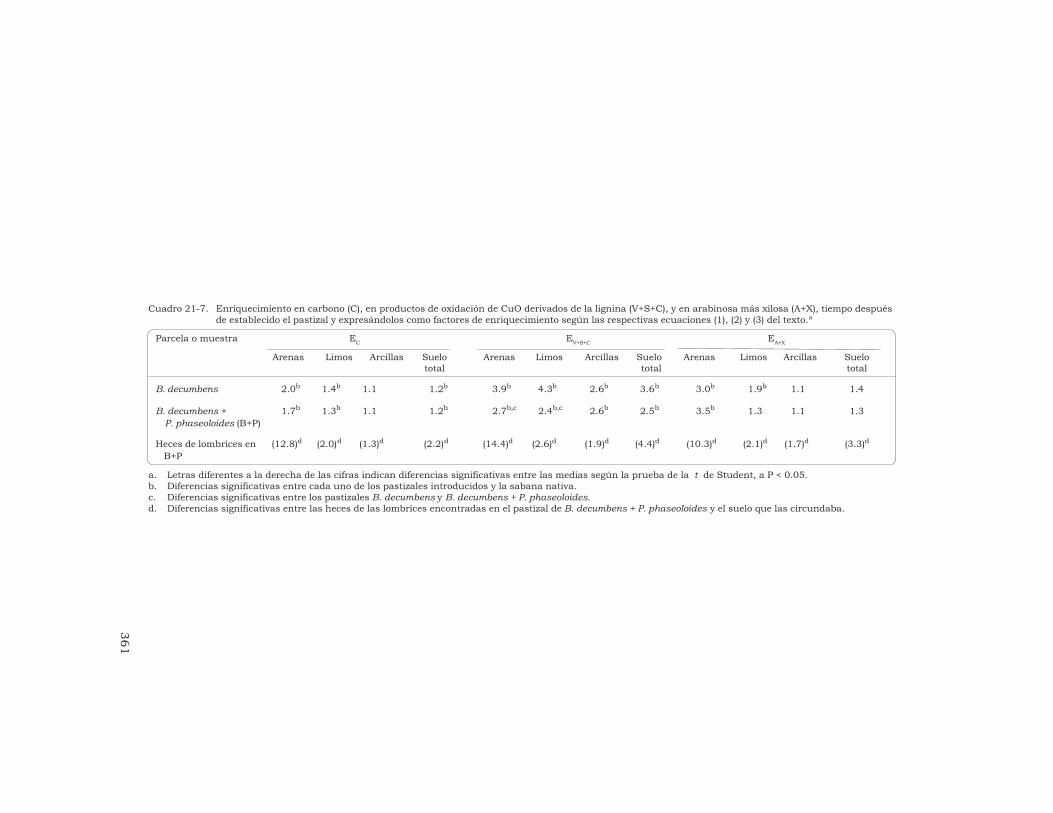
Alteración de la Lignina y de los C
arbohidratos...
361
Cuadro 21-7. Enriquecimiento en carbono (C), en productos de oxidación de CuO derivados de la lignina (V+S+C), y en arabinosa más xilosa (A+X), tiempo despuésde establecido el pastizal y expresándolos como factores de enriquecimiento según las respectivas ecuaciones (1), (2) y (3) del texto.a
Parcela o muestra EC EV+S+C EA+X
Arenas Limos Arcillas Suelo Arenas Limos Arcillas Suelo Arenas Limos Arcillas Suelototal total total
B. decumbens 2.0b 1.4b 1.1 1.2b 3.9b 4.3b 2.6b 3.6b 3.0b 1.9b 1.1 1.4
B. decumbens + 1.7b 1.3b 1.1 1.2b 2.7b,c 2.4b,c 2.6b 2.5b 3.5b 1.3 1.1 1.3P. phaseoloides (B+P)
Heces de lombrices en (12.8)d (2.0)d (1.3)d (2.2)d (14.4)d (2.6)d (1.9)d (4.4)d (10.3)d (2.1)d (1.7)d (3.3)d
B+P
a. Letras diferentes a la derecha de las cifras indican diferencias significativas entre las medias según la prueba de la t de Student, a P < 0.05.b. Diferencias significativas entre cada uno de los pastizales introducidos y la sabana nativa.c. Diferencias significativas entre los pastizales B. decumbens y B. decumbens + P. phaseoloides.d. Diferencias significativas entre las heces de las lombrices encontradas en el pastizal de B. decumbens + P. phaseoloides y el suelo que las circundaba.

362
El Arado Natural:…
sabanas tropicales son estructurasresistentes. Los estudiospreliminares sobre estabilidad de losagregados demostraron que habíauna cantidad mucho mayor deagregados estables en agua en lasheces analizadas que en el sueloalrededor de ellas (Guggenberger,información sin publicar). Lavelle yMartin (1992) mostraron que losprocesos de descomposición sonconsiderablemente lentos en lasheces viejas, especialmente a causade la protección física que recibe laMO en las estructuras compactas delas heces. Esta MO del sueloprotegida físicamente deja de serloen la fracción arenosa después de laaplicación del ultrasonido(Christensen 1992).
El aumento de la fracción delimos en detrimento de la de arcillas(Cuadro 21-1) se debió,probablemente, a la unión fuerte delas partículas de arcilla con lafracción de limos mediante elmucílago segregado por laslombrices, fracción que no fuedestruida por el ultrasonido. La MOdel suelo presente en losmicroagregados puede ser muyestable frente a la acción de lasbacterias puesto que está protegidafísicamente en las heces (Skjemstadet al. 1993).
Alteración de la lignina delsueloTanto los rendimientos totalesobtenidos de la suma V+S+C comolas relaciones (ac/al)V y (ac/al)S de laMO del suelo asociada a la fracciónarenosa fueron muy parecidos a losde la MO ligeramente alterada en losestratos orgánicos del suelo (Kögel etal. 1988; Ziegler et al. 1986); estehecho apoya la idea de que la MO delsuelo distribuida en la fracción dearenas presenta las característicasbioquímicas de aquellos residuos
vegetales que no han sido alterados(Baldock et al. 1992; Christensen1992).
La biodegradación de la ligninalleva a una modificación de lospolímeros de ésta y aparecen,entonces, contenidos más bajos deresiduos intactos de lignina que sedeben a la descomposición de losanillos y a un mayor grado deoxidación de las cadenas laterales(Kögel-Knabner et al. 1988; Zech yKögel-Knabner 1994). Por lo tanto,la disminución del rendimiento enproductos de la oxidación de CuO,junto con el aumento de lasrelaciones (ac/al)V y (ac/al)S, sonindicadores del proceso dedescomposición de la lignina que seobserva cuando aumenta laprofundidad del suelo (Kögel-Knabner et al. 1988; Zech y Kögel-Knabner 1994) y cuando disminuyeel tamaño de las partículas del suelo(Guggenberger et al. 1994).
Los datos indicaron también quela lignina de la sabana nativa seencontraba en un estado dedescomposición muy avanzado. Lacantidad de lignina que rindió elsuelo total fue bastante baja y lasrelaciones (ac/al)V y (ac/al)Scorrespondientes fueronconsiderablemente más altas que lasencontradas habitualmente en lossuelos de bosque de las zonastempladas (V+S+C = 8 a 20 mg/g C;(ac/al)V = 0.4 a 1.0 mg/g C; Zech etal. 1992). Este resultado seexplicaría por el régimen detemperaturas altas de la sabanatropical. Los procesos dedescomposición de la MO en el sueloocurren, generalmente, a unavelocidad varias veces mayor en losclimas tropicales que en lostemplados (Coleman et al. 1989). Noobstante, la descomposición de lalignina se hallaba en un punto muybajo tras el establecimiento de lospastizales.

Alteración de la Lignina y de los Carbohidratos...
363
El notable incremento enrendimiento de los productos deoxidación del CuO, junto con eldescenso del valor de las relaciones(ac/al)V y (ac/al)S, en los pastizalesproductivos de gramínea sola o degramínea con leguminosa ocasionó,probablemente, en comparación conla sabana nativa, un nivelrelativamente alto de incorporaciónde la hojarasca (Serrão et al. 1979).Los tejidos vasculares sindescomponer de los vegetales noleñosos se caracterizan porque lasuma de V+S+C rinde entre 30 y80 g/kg C y porque presentan unarelación (ac/al)V de 0.15 ± 0.05(Hedges y Mann 1979). Porconsiguiente, un aporte mayor deresiduos vegetales frescos no sóloaumenta la cantidad de macro-MOdel suelo (ver más arriba) sinotambién la cantidad de ligninaintacta del suelo (y mucho más quela anterior). Hay pruebas evidentesde que la lignina se descompone másen un sistema en que hayalimitaciones de N (Fog 1988; Haider1986). De ahí que el contenido másalto de N que se registró en lospastizales pudo aminorar lavelocidad de la lignólisis, lo que noocurrió en el suelo más pobre en Nde la sabana nativa.
Los factores de enriquecimientorelacionados con la suma V+S+C,que se expresan como:
indican que, después delestablecimiento de un pastizal, hayun enriquecimiento en productos deoxidación del CuO mayor que en Ctotal, en cada fracción por tamañoanalizada (Cuadro 21-7). Puesto queun valor alto de la suma de V+S+C esun indicador de la presencia dematerial vegetal sin descomponer
(Kögel-Knabner et al. 1988; Ziegler etal. 1986), los resultados obtenidosindicarían que la MO adicionalconsiste, principalmente, en recursosprimarios (de origen vegetal).
El valor de la suma V+S+Cobtenido en las heces de laslombrices fue el doble del obtenidoen el suelo que las circundaba, en elpastizal de B. decumbens + P.phaseoloides (Cuadro 21-3); además,el 60% de los productos de laoxidación fenólica de CuO estabaasociado a la fracción arenosa. En elcálculo de estos datos se ponderó lasuma V+S+C con el contenido de C ycon la masa de cada fracción. Enrelación con esto, el valor 0.22 de larelación (ac/al)V es sólo ligeramentesuperior al del tejido vascular vegetalque no se ha degradado (Hedges yMann 1979). Estos resultadossugieren que, considerada laestrategia ecológica de las lombricesde hábito anécico o endo-anécico, lahojarasca se incorporó fuertemente ala fracción mineral del suelo [efectoanécico según Lavelle (1988)] y lalignina quedó bien preservada dentrode las heces de las lombrices.
Alteración de loscarbohidratos del sueloLa concentración del total decarbohidratos hidrolizables seregistró, en cada caso, en todo elrango de 50 a 250 mg de C (derivadode carbohidratos)/g de C, tal como loreportó Cheshire (1979). Asimismo,la distribución de los carbohidratosen las diferentes fracciones resultócomparable con los datosencontrados en suelos de latitudestempladas (Angers y Mehuys 1990;Guggenberger et al. 1994).
El enriquecimiento relativo enazúcares (derivados de plantas) de lafracción de arenas confirmó lahipótesis de que la MO del sueloasociada a esta fracción estaba
g (V+S+C)/kg de fraccióndel suelo o de suelo total
de la sabana
g (V+S+C)/kg de fraccióndel suelo o de suelo total
del pastizal(Ecuación 2)EV+S+C =

364
El Arado Natural:…
compuesta, en gran parte, porresiduos vegetales. Por su parte, losvalores altos de [(M+G)/(A+X)] y[(R+F)/(A+X)] de la fracción dearcillas indicaron ya sea tasaselevadas de síntesis de carbohidratospor las bacterias o ya estabilizaciónde los metabolitos de origenbacteriano. De acuerdo conChristensen (1992), la colonizaciónbacteriana se concentra dentro delas partículas de la fracción dearcillas y hay pruebas claras de quelos carbohidratos provenientes demicroorganismos se estabilizanmediante una asociación íntima concomponentes inorgánicos, entre ellosel Fe, en la fracción de arcillas(Oades et al. 1987; Oades 1989). LaMO del suelo asociada a la fracciónde limos estuvo ausente en ambostipos de azúcares (de origen vegetal obacteriano). Oades (1989) señalóque la fracción de limos estáenriquecida en ácidos húmicos, queson químicamente complejos yamorfos, y presentan un contenidobajo de carbohidratos (Kögel-Knabner et al. 1988).
En el Cuadro 21-7 se consideróque la cantidad de carbohidratos deorigen vegetal estaba representadapor la suma de la arabinosa y laxilosa (A+X), y de ahí se calculó elenriquecimiento de A+X tras laintroducción del pastizal:
El sistema de uso de la tierraafectó principalmente la cantidad decarbohidratos de la fracción arenosa.Esta fracción se caracterizó, despuésde establecido el pastizal, por unenriquecimiento proporcionalmentemayor en carbohidratos de origenvegetal que en C total. Losresultados indicaron un incremento
en el componente lábil de la MO delsuelo, ya que las hemicelulosas sedegradan fácilmente. La diversidadde la vegetación afectó menos lacantidad y la composición de loscarbohidratos asociados a lasfracciones de limos y de arcillas.
Las señales detectadas paracarbohidratos en los análisis de lasheces indicaron que el efectoprincipal de la actividad de laslombrices no fue una alteración de laconcentración del C de loscarbohidratos asociado a cada unade las fracciones, sino el notableenriquecimiento en carbohidratos deorigen vegetal debido a laconcentración alta de C en lafracción de arenas.
Consecuencias para el ciclo delC y del NRao et al. (1994) demostraron,empleando los isótopos del C, que11 años después de la introduccióndel pastizal, la MO del suelo derivadade éste contribuyó significativamentea la MO total; por ejemplo, en losprimeros 2 cm del suelo, el 29% delC orgánico del suelo del pastizal quecontenía gramínea y leguminosaprovenía de residuos de laleguminosa. En el presente estudio,la mayor parte de la MO adicionalencontrada en el pastizal se habíaseparado como fracción arenosa.Las señales detectadas en el análisispara la lignina y los carbohidratosrevelaron, además, que estasituación correspondía a unaumento en la cantidad desustancias de origen vegetalligeramente alteradas.
En algunos estudios (Schulten yHempfling 1992; Schulten yLeinweber 1993; Preston et al. 1994)se ha señalado que la MO de unsuelo que no esté degradadopresenta, generalmente,concentraciones más altas de
(Ecuación 3)EA+X = g (A+X)/kg de fraccióndel suelo o de suelo total
de la sabana
g (A+X)/kg de fraccióndel suelo o de suelo total
del pastizal

Alteración de la Lignina y de los Carbohidratos...
365
recursos primarios que se puedendescomponer, que la MO de un suelodegradado adyacente al anterior;éste se caracteriza por una cantidadconsiderable de sustanciascompletamente humificadas. Seobservan, en general, cantidadesdecrecientes de sustancias lábilestras el cultivo de un suelo nativo enque haya un débil retorno deresiduos vegetales al suelo(Christensen 1992).
A diferencia de la anteriorsituación, el Oxisol investigado eneste estudio se caracterizó por tenerconcentraciones bajas de sustanciasfácilmente degradables que noestaban protegidas dentro decomplejos órgano-minerales. Lamineralización del N se reducetambién cuando hay cantidadesbajas de MO lábil (Duxbury et al.1989). Aunque no fue posibledistinguir entre los efectos de la solafertilización y del establecimiento dela gramínea y la leguminosa, lasinergia de ambas acciones produjoconcentraciones mayores derecursos primarios. Rao et al. (1994)encontraron que los residuosprovenientes de la leguminosacontribuían a la fertilidad del sueloal aumentar las tasas potenciales demineralización del N. Avanzando enesta línea, Follett y Schimel (1989) ySchulten y Hempfling (1992)hallaron que la baja disponibilidad ocalidad (en cuanto a la capacidad dedegradación) de las fuentes de Cpropicia un crecimiento bacterianolimitado, que reduce a su vez lacapacidad de inmovilizar y conservarel N mineral. El incremento de lareserva de fuentes de C disponibleque se presenta tras elestablecimiento de un pastizalpermite, por tanto, estimular elcrecimiento bacteriano y evitar laspérdidas de N del suelo porlixiviación (puesto que aumenta la
capacidad del suelo para almacenarN) (Follett y Schimel 1989).
Las señales detectadas en elanálisis para la lignina y loscarbohidratos sugieren que en lasheces de las lombrices seincrementan los residuos vegetalesligeramente alterados en la fracciónde arenas. Estos resultados apoyanla hipótesis de que los procesos dedescomposición adquieren,temporalmente, un ritmo lento enlas heces de las lombrices, donde secrea una reserva lábil de MO queestá protegida físicamente (Lavelle yMartin 1992). La actividad de laslombrices aumenta también laincorporación de MO en el horizontemineral puesto que, al excavar susgalerías, las lombrices incorporan lahojarasca fresca en el suelo. Dadoque los suelos de la sabanapresentan, a menudo, una intensaactividad de lombrices de tierra(Lavelle 1988) cuyas heces cubrencompletamente, a veces, lasuperficie del suelo (como en el casode la parcela de B. decumbens + P.phaseoloides), el papel de estaslombrices en la transformación de laMO del suelo y en la liberación delos nutrientes unidos a complejosorgánicos (N, P, S) merece serinvestigado con más detenimiento.
AgradecimientosLos autores agradecen a Bent T.Christensen por su apoyo en laintroducción del método deseparación de fracciones del sueloen el laboratorio de que ellosdisponían. Asimismo, a LudwigHaumaier por sus comentariosútiles sobre el manuscrito. Graciastambién a Tanja Gonter y a IrisZimmermann por su apoyo técnicodurante la realización de estetrabajo.

366
El Arado Natural:…
ReferenciasAhmed M; Oades JM. 1984. Distribution
of organic matter and adenosinetriphosphate after fractionation ofsoils by physical procedures. SoilBiol Biochem 16:465-470.
Anderson DW; Paul EA. 1984. Organo-mineral complexes and their studyby radiocarbon dating. Soil Sci SocAm J 48:298-301.
Anderson DW; Saggar S; Bettany JR;Stewart JWB. 1981. Particle sizefractions and their use in studies ofsoil organic matter: I. The natureand distribution of forms of carbon,nitrogen and sulfur. Soil Sci Soc AmJ 45:767-772.
Anderson JM; Flanagan PW. 1989.Biological processes regulatingorganic matter dynamics in tropicalsoils. En: Coleman DC; Oades JM;Uehara G, eds. Tropical soil organicmatter. University of Hawaii Press,Honolulu, Estados Unidos.p 97-123.
Andrews MA. 1989. Capillary gas-chromatographic analysis ofmonosaccharides: improvementsand comparisons usingtrifluoroacetylation andtrimethylsilylation of sugarO-benzyl- and O-methyl-oximes.Carbohydr Res 194:1-19.
Angers DA; Mehuys GR. 1990. Barley andalfalfa cropping effects oncarbohydrate contents of a clay soiland its size fractions. Soil BiolBiochem 22:285-288.
Baldock JA; Oades JM; Waters AG; PengX; Vassallo AM; Wilson MA. 1992.Aspects of the chemical structure ofsoil organic materials as revealed bysolid-state 13C NMR spectroscopy.Biogeochemistry 16:1-42.
Boas NF. 1953. Method for thedetermination of hexosamines intissues. J Biol Chem 204:553-563.
Bouwman AF, ed. 1990. Soils and thegreenhouse effect. John Wiley,Nueva York.
Cambardella CA; Elliott ET. 1992.Particulate soil organic-matterchanges across a grasslandcultivation sequence. Soil Sci SocAm J 56:777-783.
Cheshire MV. 1979. Nature and origin ofcarbohydrates in soils. AcademicPress, Londres.
Christensen BT. 1985. Carbon andnitrogen in particle size fractionsisolated from Danish arable soils byultrasonic dispersion and gravity-sedimentation. Acta Agr Scand35:175-187.
Christensen BT. 1986. Strawincorporation and soil organicmatter in macro-aggregates andparticle size separates. J Soil Sci37:125-135.
Christensen BT. 1992. Physicalfractionation of soil and organicmatter in primary particle size anddensity separates. En: Stewart BA,ed. Advances in soil science.Springer-Verlag, Nueva York.Vol. 20, p 1-90.
Coleman DC; Oades JM; Uehara G. 1989.Introduction: soil organic matter:tropical vs. temperate. En: ColemanDC; Oades JM; Uehara G, eds.Tropical soil organic matter.University of Hawaii Press,Honolulu, Estados Unidos. p 1-4.
Duxbury JM; Smith MS; Doran JW. 1989.Soil organic matter as a source anda sink of plant nutrients. En:Coleman DC; Oades JM; Uehara G.eds. Tropical soil organic matter.University of Hawaii Press,Honolulu, Estados Unidos. p 33-67.
Elliott ET; Cambadella CA. 1991. Physicalseparation of soil organic matter.Agric Ecosyst Environ 34:407-419.
Ertel JR; Hedges JI. 1984. The lignincomponent of humic substances:distribution among soil sedimentaryhumic, fulvic, and base-insolublefractions. Geochim Cosmochim Acta48:2065-2074.

Alteración de la Lignina y de los Carbohidratos...
367
Fog K. 1988. The effect of added nitrogenon the rate of decomposition oforganic matter. Biol Rev 63:533-462.
Follett RF; Schimel DS. 1989. Effect oftillage practices on microbialbiomass dynamics. Soil Sci Soc AmJ 53:1091-1096.
Guggenberger G; Christensen BT; Zech W.1994. Land use effects on thecomposition of organic matter inparticle-size separates of soils; I:Lignin and carbohydrate signature.Eur J Soil Sci 45:449-458.
Guggenberger G; Zech W; Thomas RJ.1995. Lignin and carbohydratealteration in particle-size separatesof an Oxisol under tropical pasturesfollowing native savanna. Soil Biol.Biochem 27:11629-1638.
Haider K. 1986. The synthesis anddegradation of humic substances insoil. En: Transaction of the XIIICongress International Society ofSoil Science, Hamburgo, Alemania.p 644-656.
Hedges JI; Ertel JR. 1982.Characterization of lignin by gascapillary chromatography of cupricoxide oxidation products. Ann Chem54:174-178.
Hedges JI; Mann DC. 1979. Thecharacterization of plant tissues bytheir lignin oxidation products.Geochim Cosmochim Acta43:1803-1807.
Hedges JI; Parker PL. 1976. Land-derivedorganic matter in surface sedimentsfrom the Gulf of Mexico. GeochimCosmochim Acta 40:1803-1807.
Hedges JI; Blanchette RA; Weliky K; DevolAH. 1988. Effects of fungaldegradation on the CuO oxidationproducts of lignin: a controlledlaboratory study. GeochimCosmochim Acta 52:2717-2726.
Jenkinson DS; Powlson DS. 1976. Theeffect of biocidal treatments onmetabolism in soil; V: A method formeasuring soil microbial biomass.Soil Biol Biochem 8:209-214.
Jenkinson DS; Rayner JH. 1977. Theturnover of soil organic matter insome of the Rothamsted classicalexperiments. Soil Sci 123:298-305.
Kögel I; Hempfling R; Zech W; HatcherPG; Schulten H-R. 1988. Chemicalcomposition of the organic matter inforest soils; I: Forest litter. Soil Sci146:124-136.
Kögel-Knabner I; Zech W; Hatcher PG.1988. Chemical compositionof the organic matter in forestsoils; III: The humus layer. ZPflanzenernaerh Bodenkd151:331-340.
Kögel-Knabner I; Zech W; Hatcher PG.1991. Chemical structural studiesof forest soil humic acids: aromaticcarbon fraction. Soil Sci Soc Am J55:241-247.
Lascano CL; Estrada J. 1989. Long-termproductivity of legume-based andpure grass pastures in the easternplains of Colombia. En: Proceedingsof the XVI International GrasslandCongress, Niza, Francia.p 1179-1180.
Lavelle P. 1988. Earthworm activities andthe soil system. Biol Fertil Soils6:237-251.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humidtropics. Soil Biol Biochem24:1491-1498.
Lee KE. 1985. Earthworms: their ecologyand relationships with soils andland use. Academic Press, Londres.
Moers MEC; Baas M; de Leeuw JW; BoonJJ; Schenk PA. 1990. Occurrenceand origin of carbohydrates in peatsamples from a red mangroveenvironment as reflected byabundances of neutralmonosaccharides. GeochimCosmochim Acta 54:367-381.

368
El Arado Natural:…
Murayama S. 1984. Changes in themonosaccharide compositon duringthe decomposition of straws underfield conditions. Soil Sci Plant Nutr30:367-381.
Oades JM. 1984. Soil organic matter andstructural stability: mechanismsand implications for management.Plant Soil 76:319-337.
Oades JM. 1989. An introduction toorganic matter in mineral soils. En:Dixon JB; Weed SB, eds. Mineralsin soil environments. Book SeriesNo. 1. Soil Science Society ofAmerica, Madison, Estados Unidos.p 89-159.
Oades JM; Vassallo AM; Waters AG;Wilson MA. 1987. Characterizationof organic matter in particle-sizeand density fractions from a red-brown earth by solid-state 13C NMR.Aust J Soil Res 25:71-82.
Parton WJ; Schimel DS; Cole CV; OjimaDS. 1987. Analysis of factorscontrolling soil organic matter levelsin Great Plains grasslands. Soil SciSoc Am J 51:1173-1179.
Preston CM; Newman RH; Rother P. 1994.Using 13C CPMAS NMR to assesseffects of cultivation on the organicmatter of particle size fractions in agrassland soil. Soil Sci 157:26-35.
Rao IM; Zeigler RS; Vera R; Sarkarung S.1993. Selection and breeding foracid-soil tolerance in crops: uplandrice and tropical forages as casestudies. Bioscience 43:454-465.
Rao IM; Ayarza MA; Thomas RJ. 1994.The use of carbon isotope ratios toevaluate legume contribution to soilenhancement in tropical pastures.Plant Soil 162:177-182.
Sarkanen KV; Ludwig CH. 1971. Lignins.Wiley-Interscience, Nueva York.
Schulten H-R; Hempfling R. 1992.Influence of agricultural soilmanagement on humuscomposition and dynamics:classical and modern analyticaltechniques. Plant Soil 142:259-271.
Schulten H-R; Leinweber P. 1993.Pyrolysis-field ionization massspectrometry of agricultural soilsand humic substances: effect ofcropping systems and influence ofthe mineral matrix. Plant Soil151:77-90.
Serrão EAS; Fales IC; da Vega JB;Teixeira Neto JF. 1979.Productividad de los pastoscultivados en suelos de bajafertilidad de la región amazónica deBrasil. En: Sánchez PA; Tergas LE,eds. Producción de pastos en suelosácidos de los trópicos. CIAT, Cali,Colombia. p 199-229.
Skjemstad JO; Janik LJ; Head MJ;McClure SG. 1993. High energyultraviolet photo-oxidation: a noveltechnique for studying physicallyprotected organic matter in clay-and silt-sized aggregates. J Soil Sci44:485-499.
Sombroek WG; Nachtergaele FO; Hebel A.1993. Amounts, dynamics andsequestering of carbon in tropicaland subtropical soils. Ambio22:417-426.
Swift MJ; Heal OW; Anderson JM. 1979.Decomposition in terrestrialecosystems. Blackwell Scientific,Oxford.
Thomas RJ; Fisher MJ; Ayarza MA; SanzJI. 1995. The role of forage grassesand legumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. Advances inSoil Science Series, Lewis Pub.,Boca Ratón, Estados Unidos.p 61-83.
Tiessen H; Stewart JWB. 1983. Particle-size fractions and their use instudies of soil organic matter; II:Cultivation effects on organicmatter composition in size fractions.Soil Sci Soc Am J 47:509-514.

Alteración de la Lignina y de los Carbohidratos...
369
Zech W; Kögel-Knabner I. 1994. Patternsand regulation of organic mattertransformation in soils: litterdecomposition and humification.En: Schulze ED, ed. Flux control inbiological systems. Academic Press,Nueva York. p 303-334
Zech W; Ziegler F; Kögel-Knabner I;Haumaier L. 1992. Humicsubstances distribution andtransformation in forest soils. SciTotal Environ 117/118:155-174.
Ziegler F; Kögel I; Zech W. 1986.Alteration of gymnosperm andangiosperm lignin duringdecomposition in forest humuslayers. Z Pflanzenernaerh Bodenkd149:323-331.

370
El Arado Natural:…
ResumenSe evaluaron los efectos que unaespecie de lombriz anécica quehorada galerías profundas en elsuelo, Martiodrilus carimaguensis,causa en la fertilidad del suelo y enel crecimiento de las plantas, endos pastizales de los LlanosOrientales de Colombia. Estaespecie permanece inactiva (enestivación) a cierta profundidaddurante la época seca. Paraimpedir que estas lombricescolonicen la capa superficial delsuelo al retornar a ella al inicio dela época lluviosa, se usó una red defibra de vidrio. Mediante unanálisis de componentesprincipales, se estudiaron losefectos de la ausencia de estaespecie en las propiedades del
suelo y en la biomasa de lasplantas. La manipulaciónexperimental logró impedir laactividad de M. carimaguensis en lacapa superficial del suelo. Lamayor parte de la varianzaregistrada en este estudio (29.35%)se debió al efecto del tipo y la edaddel pastizal en la estructura delsuelo y en la materia orgánica.Una menor parte de la varianza(10.81%) se atribuyó al efecto de laleguminosa en las propiedadesfísicas del suelo. El efecto causadopor la ausencia de las lombrices enlas propiedades del suelo y en elcrecimiento de las plantas explicó el16.50% de la varianza total. Labiomasa de los individuos deM. carimaguensis se relacionó conlos siguientes factores: menorcompactación del suelo, mayorcontenido de C, menor saturaciónde Al, mayor biomasa herbácea ymenor biomasa de malezas. Estosresultados apoyan la creenciageneral de que las lombricesafectan la fertilidad del suelo y elcrecimiento de las plantas.
Palabras clave: ecología del suelo,fertilidad del suelo, lombricesanécicas, manipulación delombrices, pastizales tropicales
CAPÍTULO 22
Efecto de la Exclusión de la LombrizAnécica Martiodrilus carimaguensisJiménez y Moreno en las Propiedadesdel Suelo y en el Crecimiento de lasPlantas, en Pastizales de los LlanosOrientales de Colombia1
T. Decaëns* , J. J. Jiménez** y P. Lavelle*
1. Publicado en Pedobiologia 43, p 835-841, porDecaëns et al. Copyright © 1999 de Urban &Fischer Verlag. Impreso con permiso de lacasa editorial.
* Laboratoire d’Ecologie des Sols Tropicaux,IRD/Unidad de Suelos y Nutrición dePlantas, CIAT, 32 Av. Henri Varagnat,F-93143 Bondy Cx, Francia.
** Departamento de Biología Animal I, Facultadde Biología, Universidad Complutense,28040 Madrid, España.

Efecto de la Exclusión de la Lombriz Anécica...
371
IntroducciónLa mayoría de los estudios sobre elimpacto que hace en el suelo laactividad de las lombrices de tierrahan sido realizados enexperimentos de laboratorio decorta duración (Derouard et al.1997; Pashanasi et al. 1992; Spainet al. 1992). A pesar de que esteenfoque ha permitido un avancesignificativo en la comprensión delpapel que desempeñan laslombrices en los procesosdesarrollados en el suelo, son aúnincompletos los conocimientosactuales sobre la forma en queestos organismos influyen en losprocesos relacionados con unecosistema, en condiciones decampo in situ (Bohlen et al. 1995).
El objetivo de este estudio fue lavaloración de un método innovador(Baker 1998) que excluyeraselectivamente, de pequeñasunidades experimentales, laslombrices anécicas (sensu Bouché1977). Se evaluó la eficiencia delmétodo respecto a su capacidad deeliminar las lombrices anécicasgrandes, pertenecientes a la especieMartiodrilus carimaguensis Jiménezy Moreno, del suelo superficial dedos pastizales de gramínea yleguminosa en los Llanos Orientalesde Colombia. Allí, en el CNICarimagua, las poblaciones de estaespecie han sido favorecidas por lospastizales intensivos, pero se hanreducido dramáticamente en loscultivos anuales (Jiménez et al.1998a). El estudio determina losefectos que produce la desapariciónde esta especie en los procesos delsuelo y en el crecimiento de lasplantas, y los compara con losefectos debidos a otros factoresambientales.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua delconvenio CIAT-CORPOICA (4°37’ N,71°19’ O), en los Llanos Orientales(Meta), en Colombia. El clima estropical húmedo con unatemperatura y una precipitación de26 °C y 2300 mm, respectivamente,como valor medio anual (datos delCIAT). La vegetación nativa estácompuesta por parches de sabanaabierta y de bosques de galería. Lossuelos son Oxisoles bien agregados,de acidez alta y de fertilidad químicabaja.
Diseño y parcelasexperimentalesEl experimento se realizó en dospastizales, cada uno de ellossometido a pastoreo con2.0 animales/ha: un pastizal A, de3 años, que contiene Brachiariahumidicola, Arachis pintoi,Stylosanthes capitata y Centrosemaacutifolium; un pastizal B, de 1 año,con Panicum maximum y A. pintoi.
La ecología de las lombrices deCarimagua puede consultarse enJiménez et al. (1998a, 1998b).Todas las especies están inactivas enla época seca. Las lombrices de laespecie anécica M. carimaguensishacen galerías profundas en el suelo(60 a 100 cm) y entran en diapausahasta la siguiente época lluviosa(Jiménez et al. 1998b). La idea delestudio fue impedir el retorno de losindividuos hacia las capassuperficiales del suelo cuandocomenzaran las lluvias. Dado queotras especies estivan en esas capassuperficiales, se esperaba que de esemodo el experimento tendría efectosespecíficos en los individuos de M.carimaguensis.

372
El Arado Natural:…
Al final de la época seca (finalesde marzo de 1996) se establecieron12 unidades experimentalespequeñas en cada pastizalmencionado, que fueron sometidas alpastoreo habitual. El protocolo deestablecimiento se resume en laFigura 22-1:
1. Se extrajo, con una pala, unmonolito de suelo de40 x 40 x 30 cm;
2. Cada uno de los lados delmonolito (excepto la partesuperior) se envolvió en unared de fibra de vidrio de0.5 mm de luz;
3. El monolito se introdujo denuevo en el hueco que dejó laextracción y, para evitar
cualquier movimiento de laslombrices, se enterraronplanchas metálicas alrededorde las unidadesexperimentales;
4. Se delimitó un área dondelos individuos deM. carimaguensis, que selocalizaban por la presenciade las heces frescas (Jiménezet al. 1998b), fueronsistemáticamente controladoscon la inyección de 0.01 g decarbofuran activo en cadagalería.
Se establecieron también12 unidades sin red como testigo.
Se tomaron muestras de lavegetación y del suelo antes del
4
31
15 cm15 cm
15 c
m
Lámina demetal
Malla de fibrade vidrio,completo
30 c
m
40 cm
Superficie de muestreo
Superficie de controlquímico
2
Figura 22-1. Descripción del procedimiento de muestreo. Los números corresponden al ordencronológico de los diferentes pasos del procedimiento empleado y se refieren adescripciones completas incluidas en el texto.
70 cm

Efecto de la Exclusión de la Lombriz Anécica...
373
experimento, 6 meses después de suinicio y 18 meses después del mismo(en octubre de 1995, en agosto de1996 y en junio de 1997,respectivamente). En cada fecha setomaron cuatro unidadesexperimentales al azar en cadapastizal. Las muestras se usaronpara caracterizar la vegetación(biomasas radicular y aérea), laspropiedades del suelo (densidadaparente, densidad del tamañomedio de los agregados, resistencia ala penetración, sorción [capacidad deabsorción o adsorción], infiltración ypropiedades químicas), laspoblaciones de lombrices (densidad ybiomasa), las de hormigas y las determitas (número de cámaras). Seemplearon los métodosrecomendados por el programa TSBF(Programa de Biología y Fertilidad delos Suelos Tropicales) (Anderson eIngram 1989). De cada muestra setomaron cuatro repeticiones. Lossitios en que se establecían lasunidades experimentales en elcampo se ubicaron al azar, evitandosiempre el efecto de borde delpastizal.
EstadísticaMediante la prueba PLSD de Fisherse probó el efecto de la manipulaciónexperimental, tanto en la densidad yen la biomasa de las lombrices, comoen la abundancia de las cámaras delas hormigas y de las termitas. Laasimetría de los datos se redujo alaplicar una transformación Box-Cox(Sokal y Rohlf 1995) empleando elprograma estadístico “Vernorm”(Legendre y Vaudor 1991). Se aplicóla corrección de Bonferroni y losniveles ajustados de significación al0.05, 0.01 y 0.001 fueron 0.008,0.002 y 0.0002, respectivamente.
El Análisis de ComponentesPrincipales (ACP) se realizó con elprograma (software) ADE (Thioulouseet al. 1997), con el fin de extraer la
principal fuente de variación de losdatos ambientales. Se diseñó unamatriz de 40 líneas (muestras) y23 columnas (variables del suelo y dela vegetación). La biomasa de cadaespecie de lombriz y la abundanciade las cámaras de las hormigas y delas termitas se proyectaron sobre losejes del ACP como columnasadicionales. Se hizo un análisis deregresión simple para comprobar lascorrelaciones entre los parámetros dela fauna del suelo y los ejes del ACP.
ResultadosAl final del experimento sólo seencontró un individuo de M.carimaguensis en el conjunto de lasunidades experimentales. El grantamaño de los individuos deM. carimaguensis fue la causa de queuna ligera disminución de ladensidad de las lombricesdeterminara una reducción bruscade su biomasa total (desde -89%hasta -100%, P < 0.01) (Figura 22-2).Las diferencias observadas en lasotras especies (Figura 22-2) y en losartrópodos sociales (no se presentanaquí) no fueron significativas.
El eje 1 extraído del ACP explicóel 29.33% de la inercia total de losdatos (Cuadro 22-1). La correlaciónasociada con las variables dio valoresnegativos entre: (a) de un lado,biomasa alta de las gramíneas, delas raíces y del componenteherbáceo; valores altos de densidadaparente en el estrato de 5 a 10 cmdel suelo; alta resistencia a lapenetración; pH alto; y contenido denutrientes disponibles; y (b) del otro,saturación de Al alta (Figura 22-3,A,Cuadro 22-1). El ordenamiento delas muestras mostró una separaciónentre: (a) de un lado, las tomadas enlas dos primeras fechas y lastomadas posteriormente(Figura 22-3,B); y (b) del otro, lastomadas en el pastizal A y las del
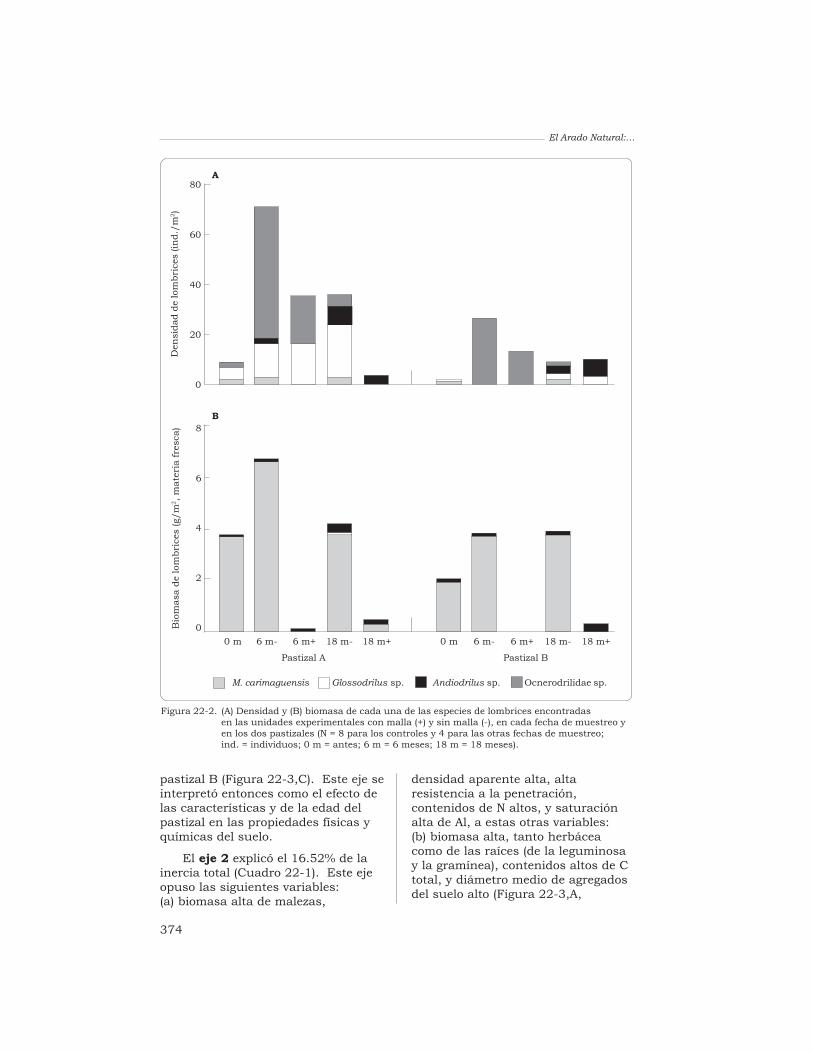
374
El Arado Natural:…
pastizal B (Figura 22-3,C). Este eje seinterpretó entonces como el efecto delas características y de la edad delpastizal en las propiedades físicas yquímicas del suelo.
El eje 2 explicó el 16.52% de lainercia total (Cuadro 22-1). Este ejeopuso las siguientes variables:(a) biomasa alta de malezas,
densidad aparente alta, altaresistencia a la penetración,contenidos de N altos, y saturaciónalta de Al, a estas otras variables:(b) biomasa alta, tanto herbáceacomo de las raíces (de la leguminosay la gramínea), contenidos altos de Ctotal, y diámetro medio de agregadosdel suelo alto (Figura 22-3,A,
80
60
40
20
0
8
6
4
2
00 m 6 m- 6 m+ 18 m- 18 m+ 0 m 6 m- 6 m+ 18 m- 18 m+
M. carimaguensis Glossodrilus sp. Andiodrilus sp. Ocnerodrilidae sp.
Pastizal B
Bio
mas
a de
lom
bric
es (g
/m2 ,
mat
eria
fres
ca)
Den
sida
d de
lom
bric
es (i
nd.
/m2 )
Figura 22-2. (A) Densidad y (B) biomasa de cada una de las especies de lombrices encontradasen las unidades experimentales con malla (+) y sin malla (-), en cada fecha de muestreo yen los dos pastizales (N = 8 para los controles y 4 para las otras fechas de muestreo;ind. = individuos; 0 m = antes; 6 m = 6 meses; 18 m = 18 meses).
A
B
Pastizal A
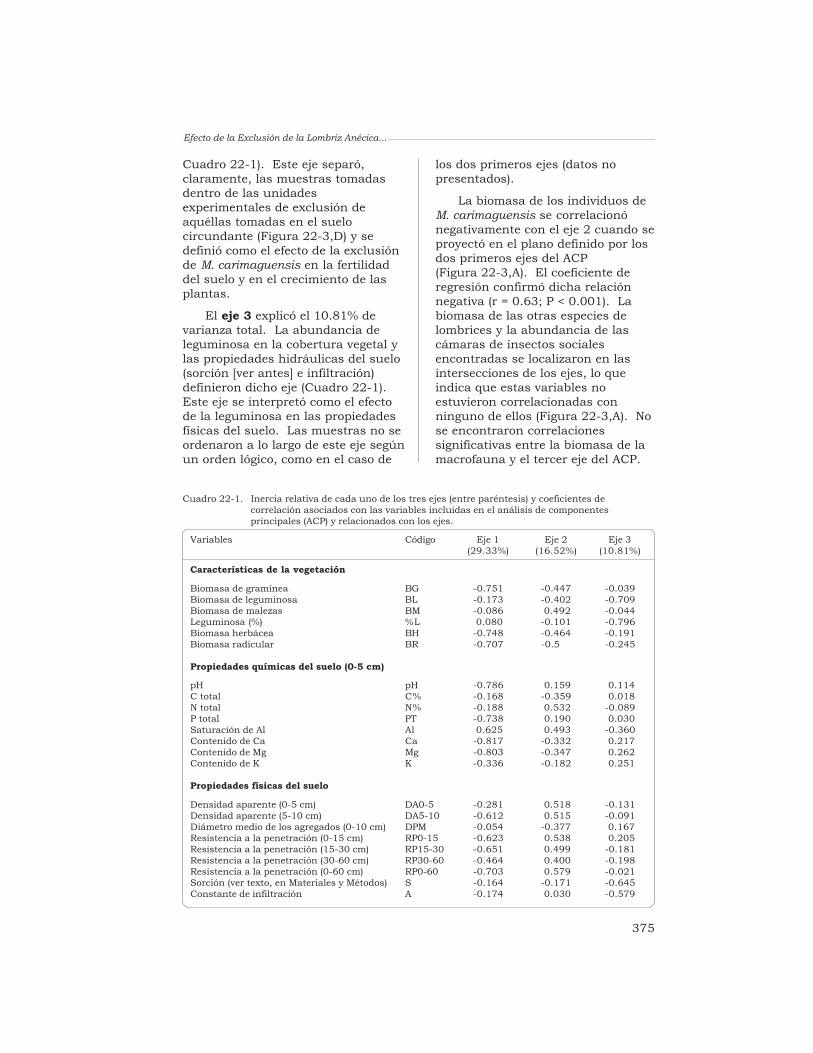
Efecto de la Exclusión de la Lombriz Anécica...
375
Cuadro 22-1). Este eje separó,claramente, las muestras tomadasdentro de las unidadesexperimentales de exclusión deaquéllas tomadas en el suelocircundante (Figura 22-3,D) y sedefinió como el efecto de la exclusiónde M. carimaguensis en la fertilidaddel suelo y en el crecimiento de lasplantas.
El eje 3 explicó el 10.81% devarianza total. La abundancia deleguminosa en la cobertura vegetal ylas propiedades hidráulicas del suelo(sorción [ver antes] e infiltración)definieron dicho eje (Cuadro 22-1).Este eje se interpretó como el efectode la leguminosa en las propiedadesfísicas del suelo. Las muestras no seordenaron a lo largo de este eje segúnun orden lógico, como en el caso de
los dos primeros ejes (datos nopresentados).
La biomasa de los individuos deM. carimaguensis se correlacionónegativamente con el eje 2 cuando seproyectó en el plano definido por losdos primeros ejes del ACP(Figura 22-3,A). El coeficiente deregresión confirmó dicha relaciónnegativa (r = 0.63; P < 0.001). Labiomasa de las otras especies delombrices y la abundancia de lascámaras de insectos socialesencontradas se localizaron en lasintersecciones de los ejes, lo queindica que estas variables noestuvieron correlacionadas conninguno de ellos (Figura 22-3,A). Nose encontraron correlacionessignificativas entre la biomasa de lamacrofauna y el tercer eje del ACP.
Cuadro 22-1. Inercia relativa de cada uno de los tres ejes (entre paréntesis) y coeficientes decorrelación asociados con las variables incluidas en el análisis de componentesprincipales (ACP) y relacionados con los ejes.
Variables Código Eje 1 Eje 2 Eje 3(29.33%) (16.52%) (10.81%)
Características de la vegetación
Biomasa de gramínea BG -0.751 -0.447 -0.039Biomasa de leguminosa BL -0.173 -0.402 -0.709Biomasa de malezas BM -0.086 0.492 -0.044Leguminosa (%) %L 0.080 -0.101 -0.796Biomasa herbácea BH -0.748 -0.464 -0.191Biomasa radicular BR -0.707 -0.5 -0.245
Propiedades químicas del suelo (0-5 cm)
pH pH -0.786 0.159 0.114C total C% -0.168 -0.359 0.018N total N% -0.188 0.532 -0.089P total PT -0.738 0.190 0.030Saturación de Al Al 0.625 0.493 -0.360Contenido de Ca Ca -0.817 -0.332 0.217Contenido de Mg Mg -0.803 -0.347 0.262Contenido de K K -0.336 -0.182 0.251
Propiedades físicas del suelo
Densidad aparente (0-5 cm) DA0-5 -0.281 0.518 -0.131Densidad aparente (5-10 cm) DA5-10 -0.612 0.515 -0.091Diámetro medio de los agregados (0-10 cm) DPM -0.054 -0.377 0.167Resistencia a la penetración (0-15 cm) RP0-15 -0.623 0.538 0.205Resistencia a la penetración (15-30 cm) RP15-30 -0.651 0.499 -0.181Resistencia a la penetración (30-60 cm) RP30-60 -0.464 0.400 -0.198Resistencia a la penetración (0-60 cm) RP0-60 -0.703 0.579 -0.021Sorción (ver texto, en Materiales y Métodos) S -0.164 -0.171 -0.645Constante de infiltración A -0.174 0.030 -0.579

376
El Arado Natural:…
DiscusiónLos experimentos que se realizancon lombrices de tierra encondiciones de campo sirven paraevaluar los efectos causados porellas en los parámetros del suelo yen el crecimiento de las plantas.Este enfoque puede ser muy útil
RP0-15
RP
RP15-30
pH
PT
DA5-10
AyRP30-60
BMDA0-5 N%
A
K
SCaMgBGBH BR
Al
Gs
C%BL
DPM
Mc
Oc
HorTerm
%L
Eje 2
Eje 1
A Eje 2
Eje 1
Fecha de muestreo
4.63.9-5.7
-4.4
18m
0m
Eje 2
Eje 1
Tipo de pastizalEje 2
Eje 1
Presencia deM. carimaguensis
B
C D
B
A
+
–
Figura 22-3. Resultados del análisis de componentes principales (ACP): (A): círculo de correlación y devalores propios o ‘eigenvalues’ (variables en letra sencilla; variables adicionales en negrita;códigos igual que en el Cuadro 22-1. Mc = biomasa de M. carimaguensis; Ay = biomasa deAndiodrilus n. sp.; Gs = biomasa de Glossodrilus n. sp.; Oc = biomasa de Ocnerodrilidaesp.; Ant = abundancia de hormigas; Term = abundancia de termitas); (B) (C) y(D): ordenación de las muestras en el plano definido por los ejes 1 y 2 del ACP (el cuadrointerno de (B) representa la escala de las figuras 3B, 3C y 3D; 0m = antes; 6m = 6 meses;18m = 18 meses; A = pastizal de B. humidicola; B = pastizal de P. maximum; + = conmalla; – = sin malla).
6m
cuando se trata de establecer elpapel de las lombrices en laproducción primaria de losecosistemas naturales y de los queson explotados por el hombre(Bohlen et al. 1995). En nuestroestudio, el bloqueo del movimientovertical de las lombrices, en laépoca y a la profundidad

Efecto de la Exclusión de la Lombriz Anécica...
377
adecuadas, resultó ser un métodoeficiente para eliminar la especieanécica deseada.
Los principales factores queinfluyeron en las propiedades delsuelo fueron el tipo y la edad delpastizal. Este resultado puedeatribuirse a los efectos específicos delas plantas, a la protección que da alsuelo la cubierta vegetal, a la calidady a la producción de la hojarasca, yal efecto acumulativo del pisoteo delganado que puede variar según lahistoria y la naturaleza de lospastizales (Gijsman 1996; Gijsman yThomas 1996; Sánchez et al. 1991).El efecto de las leguminosas en laspropiedades físicas del suelo ocupóun tercer lugar en importancia; laexplicación de este resultado estaríaen el efecto de los canales que abrenlas raíces y aumentan lamacroporosidad y la permeabilidaddel suelo (Mytton et al. 1993).
La desaparición de M.carimaguensis produjo cambiossignificativos en las propiedades delsuelo y en la biomasa de las plantas,aunque este impacto fue deimportancia secundaria encomparación con el efecto delpastizal. La disminución de labiomasa de las lombrices se asociócon la degradación del suelo(aumentó la compactación del sueloy la saturación de Al y disminuyó elcontenido de C) y de la biomasaherbácea. A su vez, la disminuciónde la biomasa vegetal favoreció lapresencia de especies de malezasoportunistas. Estos resultadosconcuerdan con el conocimientogeneral de que las lombrices puedeninfluir en las propiedades del suelo yen el crecimiento de las plantas (verrevisión de Edwards y Bohlen 1996;Lal 1988; Lavelle 1997; Lee 1985).Se demuestra también que lapérdida de una especie en unacomunidad de lombrices de tierra,cuando aquella especie está asociada
a un importante descenso de labiomasa total, puede originarpérdidas significativas en lasfunciones del ecosistema. Ahorabien, la rapidez e intensidad de losefectos observados en este estudiofueron acentuadas, probablemente,por el pastoreo intensivo. Encondiciones naturales, sinperturbación del suelo, el impactoque hacen las lombrices de tierra enlos procesos del suelo persiste a unaescala grande de tiempo (de variosmeses o años) (Blanchart 1992).
Las poblaciones de lombrices sonmuy sensibles a las prácticas demanejo del suelo (ver, por ejemplo,Lavelle y Pashanasi 1989; Reddy etal. 1995). En Carimagua, la biomasade las especies nativas aumentaenormemente en los pastizalesintroducidos por el hombre;disminuye, en cambio,dramáticamente en los cultivosanuales (Decaëns et al. 1994;Jiménez et al. 1998a). Tales efectospueden tener, a su vez,consecuencias decisivas para lafunción del suelo y su producciónprimaria (Lavelle 1996, 1997). Sedebe centrar la atención ahora en elmanejo de las poblaciones delombrices en los agroecosistemastropicales, con el fin de beneficiarsede su actividad y del impacto quecausan en la fertilidad del suelo,para mejorar de este modo lasostenibilidad de la producciónagrícola.
AgradecimientosLos autores agradecen a R.J.Thomas, D.K. Friesen, E. Amézquita,C.G. Meléndez y N. Asakawa (CIAT)por su apoyo técnico y financiero. AJ.-P. Rossi (IRD) y a dos revisoresanónimos por la revisión útil quehicieron a una versión anterior deeste artículo.

378
El Arado Natural:…
ReferenciasAnderson JM; Ingram JS, eds. 1989.
Tropical soil biology and fertility: ahandbook of methods. 2a. ed. CABInternational, Oxford, Reino Unido.221 p.
Baker GH. 1998. The ecology,management, and benefits ofearthworms in agricultural soils,with particular reference toSouthern Australia. En: EdwardsCA. ed. Earthworm ecology. St.Lucies Press, Boca Ratón, EstadosUnidos. p 229-257.
Blanchart E. 1992. Restoration byearthworms (Megascolecidae) of themacroaggregate structure of adestructured savanna soil underfield conditions. Soil Biol Biochem24:1587-1594.
Bohlen PJ; Parmelee RW; Blair JM;Edwards CA; Stinner BR. 1995.Efficacy of methods formanipulating earthwormpopulations in large-scale fieldexperiments in agroecosystems. SoilBiol Biochem 27:993-999.
Bouché MB. 1977. Stratégieslombriciennes. En: Lohm U;Persson T, ed. Soil organisms ascomponent of ecosystems.Ecological Bulletin 25:122-132.
Decaëns T; Lavelle P; Jiménez JJ;Escobar G; Rippstein G. 1994.Impact of land management on soilmacrofauna in the Oriental Llanosof Colombia. Eur J Soil Biol30:157-168.
Derouard L; Tondoh J; Vilcosqui L;Lavelle P. 1997. Effects ofearthworm introduction on soilprocesses and plant growth. SoilBiol Biochem 29:541-545.
Edwards CA; Bohlen PJ. 1996. Biologyand ecology of earthworms. 3a. ed.Chapman and Hall, Londres, ReinoUnido. 426 p.
Gijsman AJ. 1996. Soil aggregate stabilityand soil organic matter fractionsunder agropastoral systemsestablished in native savanna. AustJ Soil Res 34:891-907.
Gijsman AJ; Thomas RJ. 1996.Evaluation of some physicalproperties of an Oxisol afterconversion of native savanna intolegume-based or pure grasspasture. Trop Grassl 30:237-248.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher M; Thomas RJ.1998a. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998b. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Lal R. 1988. Effects of macrofauna on soilproperties in tropical systems. AgricEcosyst Environ 24:101-116.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lee KE. 1985. Earthworms. Their ecologyand relationships with soils andland use. Academic Press, Orlando,Estados Unidos. 411 p.
Legendre P; Vaudor A. 1991. The Rpackage: multidimentional analysis,spatial analysis. Université deMontréal, Département desSciences Biologiques, Montreal,Canadá.

Efecto de la Exclusión de la Lombriz Anécica...
379
Mytton LR; Cresswell A; Colbourn P.1993. Improvement in soil structureassociated with white clover. GrassForage Sci 48:84-90.
Pashanasi B; Meléndez G; Szott L; LavelleP. 1992. Effect of inoculation withthe endogeic earthworm Pontoscolexcorethrurus (Glossoscolecidae) on Navailability, soil microbial biomassand the growth of three tropicalfruit tree seedlings in potexperiment. Soil Biol Biochem24:1655-1659.
Reddy MV; Kumar VPK; Reddy VR;Balashouri P; Yule DF; Cogle AL;Jangawad LS. 1995. Earthwormbiomass response to soilmanagement in a semi-arid tropicalAlfisol agroecosystem. Biol FertilSoils 19:317-321.
Sánchez PA; Castilla CE; Alegre JC. 1991.Grazing pressure effects on thepasture degradation process. En:Tropical Soils Technical Report,1988-1989. p 182-186.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed. W.H.Freeman and Company, NuevaYork, Estados Unidos.
Spain AV; Lavelle P; Mariotti A. 1992.Stimulation of plant growth bytropical earthworms. Soil BiolBiochem 24:1629-1633.
Thioulouse J; Chessel D; Doledec S;Olivier JM. 1997. ADE 4: amultivariate analysis and graphicaldisplay software. Stat Comput7:75-83.

380
El Arado Natural:…
CAPÍTULO 23
Estructura Espacio-Temporal de unaComunidad de Lombrices de Tierra yHeterogeneidad del Suelo en unPastizal Tropical en Carimagua,Colombia1
T. Decaëns* y J.-P. Rossi**
ResumenSe estableció la estructura espacio-temporal de una comunidad delombrices de tierra en un pastizaltropical en Colombia y se comparódicha estructura con laheterogeneidad espacial del suelo.Se tomaron muestras de laslombrices en seis fechas distintasdurante 1 año; las propiedades delsuelo, en cambio, se establecieron enuna sola muestra tomada al final dedicho período. El diseño delmuestreo, que fue espacialmenteexplícito, consistió en una redregular de puntos separados entre sí5 m. En cada fecha se tomaron120 muestras de lombrices paradeterminar su densidad, pero setomaron 60 puntos para establecerlas propiedades del suelo. Con el finde describir la estructura espacial delos datos recolectados se hicieron
análisis multivariables (variablemúltiple) y análisis geoestadísticos.La prueba de Mantel y la pruebaparcial de Mantel se usaron paraestablecer las relaciones entre lospatrones de distribución espacio-temporal de las lombrices y laheterogeneidad del suelo. Losresultados obtenidos demostraroncon claridad la existencia de unconjunto estructurado en lacomunidad de lombrices. Parte deestos patrones fueron estables en eltiempo, al menos en la escalatemporal correspondiente al períodode estudio, y respondían a undominio alterno de parches depoblación de determinados conjuntosde especies. A partir de estosresultados se discute el significadoecológico de estos grupos deespecies.
Las propiedades del suelomanifestaron también unaorganización espacial significativa ysólo una parte pequeña de laestructura espacio-temporal de ladistribución de las lombrices serelacionó con esas propiedades. Lasespecies pequeñas fueron másabundantes en áreas (parches) enque el suelo era más compacto,mientras que las especies grandes seasociaban con áreas en que labiomasa de las raíces y el nivel de Ceran altos. Se discute también en
1. Publicado en Ecography 24(6), p 671-682,por Decaëns y Rossi. Copyright © 2001 deEcography. Impreso con permiso de la casaeditorial.
* Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia/Unidad de Suelos yNutrición de Plantas, CIAT, AA 6713, Cali,Colombia.
** Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.

Estructura Espacio-Temporal de una Comunidad de Lombrices...
381
este trabajo si la distribuciónespacio-temporal de las lombricesinduce patrones similares dedistribución en las propiedadesfísico-químicas del suelo o si, por elcontrario, son éstas las quedeterminan la distribución de laslombrices. Los resultados de estetrabajo sugieren que la organizaciónde la comunidad de lombricescorresponde, en gran parte, a unsistema que no está en equilibrio,donde las poblaciones móviles dealgunas especies de lombricesinfluyen en las propiedades delsuelo. Es difícil establecer el efectoglobal de la comunidad de lombricesen el suelo porque en él predominanfactores que se expresan,principalmente, a una escala mayoren la jerarquía ecológica.
IntroducciónEn los ecosistemas terrestres, losprocesos que acontecen en el sueloejercen una función reguladora enla mayoría de los ciclosbiogeoquímicos (Anderson yFlanagan 1989; Swift et al. 1979). Elpapel que desempeñan en estosciclos los macroinvertebradosexcavadores de tamaño grande (porejemplo, las lombrices, las termitas ylas hormigas) se ha reconocidoampliamente (Lavelle et al. 1993), ydichos organismos han sidodefinidos como los ingenieros delecosistema (Jones et al. 1994). Estosorganismos producen, en efecto,estructuras físicas (heces, domos ygalerías) que constituyen lugaresespecíficos de regulación dondeocurren procesos importantes delsuelo a una escala mayor (Anderson1995; Lavelle 1996), que puedeninfluir o bien en la proliferación deotros organismos del suelo o bien enla estructura de las comunidades deéstos. Jones et al. (1994, 1997)señalaron la necesidad de establecer
los patrones de distribución espacialde las poblaciones de ingenierosecológicos con el fin de comprendermejor la importancia que tienen lasfunciones que ellos cumplen.
Se sabe, desde hace algúntiempo, que las lombrices de tierrainfluyen en los procesos y en laspropiedades del suelo a diferentesescalas de tiempo y de espacio(Lavelle et al. 1992; Syers y Springett1984). Las heces que producenestán compuestas por agregadosórgano-minerales cuya estabilidadestructural es, generalmente, mayorque la de los agregados del suelo(Blanchart et al. 1997; Lal yAkinremi 1983; Lee y Foster 1991).Los procesos de mineralización de lamateria orgánica (MO) se intensificanen las heces frescas y se reducen odesaparecen en las heces secas(Lavelle et al. 1998; Martin 1991).Se ha demostrado también, enexperimentos realizados a nivel demicrocosmos, que una comunidadsencilla de lombrices puede causarefectos significativos en laspropiedades del suelo y en elcrecimiento de las plantas (Bohlen yEdwards 1995; Derouard et al. 1997;Haimi y Huhta 1990). Por otro lado,falta información sobre el posibleimpacto in situ de conjuntoscomplejos de especies a escalasespacio-temporales más grandes.
La distribución de la fauna delsuelo es, con frecuencia, agregada, almenos en una escala espacial devarias decenas de metros (Robertsony Freckman 1995; Rossi et al. 1995,1997; Webster y Boag 1992). Lospatrones de distribución espacial,incluso cuando varían de unaespecie a otra, son principalmenteoposiciones o alternancias entreparches de alta densidad deindividuos y parches de bajadensidad. Esta heterogeneidadocurre a una escala generalmentemenor que 100 m y se extiende

382
El Arado Natural:…
desde 20 hasta 40 m para laslombrices, aunque se han observadoestructuras más reducidas de 2 a10 m (Rossi 1998). Se desconocen,en gran parte, los factores quepropician esta ordenación relativa dela estructura espacial. Pueden sermuy complejos, probablemente, ypueden implicar el influjo que causanla heterogeneidad ambiental y losparámetros demográficos de laspoblaciones (Rossi et al. 1997).
Se demostró, hace poco tiempo,que la distribución espacial de lascomunidades de lombrices presentauna estabilidad temporal relativa, almenos durante períodos de 1 ó2 años (Jiménez 1999; Rossi 1998).En algunos casos, las estructurasespacio-temporales consisten en unaalternancia de áreas dondepredominan determinados conjuntosde especies o algunos gruposfuncionales. A consecuencia de esto,diferentes parches de poblacióncorresponderían a efectos diferentesen las propiedades del suelo y en elcrecimiento de las plantas (Rossi1998). Por tanto, el papel funcionalde la organización espacio-temporalde la comunidad de lombrices puedetener un efecto regulador en algunosaspectos del funcionamiento delsuelo, al menos a cierta escalaespacio-temporal de observación(Rossi 1998).
El objetivo de este estudio fueestablecer la posible relación entre laorganización espacio-temporal de lacomunidad de lombrices y laheterogeneidad espacial de algunaspropiedades del suelo, en un pastizaltropical. Numerosos estudiosrealizados en el sitio experimental deeste estudio y en otros lugares handemostrado que algunos procesosque acontecen en el suelo estánregulados por la actividad de laslombrices, al menos a la escala deobservación de la bioestructura queellas producen (heces de algunos cm)
o en el entorno cercano (unasdecenas de cm). La meta del trabajofue verificar si el impacto causado porlas lombrices en el suelo puedeobservarse a una escala mayor (unaparcela de algunas decenas demetros). Se probó la hipótesis de quela estructura espacio-temporal de losconjuntos de especies de lombricesinduce patrones similares deheterogeneidad espacial en el suelo(al mismo tamaño de resolución) y deque el fenómeno inverso, o sea, quedicha heterogeneidad influya en laagrupación de especies de la fauna desuelo, no se presenta.
Materiales y Métodos
Sitio del estudioEl estudio se realizó en la estaciónexperimental Carimagua del convenioCORPOICA-CIAT (4°37’ N, 71°19’ O),localizada en la unidad fitogeográficade las sabanas isohipertérmicas biendrenadas de los Llanos Orientales deColombia. El clima presenta unrégimen tropical subhúmedo con unamedia anual de 26 °C de temperaturay 2300 mm de precipitación: la épocaseca está muy bien definida y vadesde noviembre hasta marzo (datosdel CIAT). La vegetación nativa varíacon la topografía: hay sabanasabiertas en las zonas altas (los“altos”) y bosques de galería osabanas inundables en las zonasbajas (los “bajos”). Los suelos sonOxisoles (Tropeptic HaplustoxIsohipertérmicos) en los altos yUltisoles (Ultic Aeric Plintaquox) enlos bajos. Ambos tipos de suelos secaracterizan por sus propiedadesfísicas favorables (agregación,porosidad, retención de agua), suacidez alta (pH (H2O) < 5), y unafertilidad química baja (saturación deAl > 80%, CIC < 5 meq/100 g desuelo seco) (datos del CIAT).

Estructura Espacio-Temporal de una Comunidad de Lombrices...
383
Las muestras se tomaron en unpastizal previamente arado deBrachiaria humidicola (Rendle) (CIAT679), Arachis pintoi Krap. y Greg.(CIAT 17434), Stylosanthes capitataVog. (cv. Capica) y Centrosemaacutifolium Benth. (cv. Vichada) quetenía 3 años. La parcela experimentalestaba situada en un alto biendrenado y el ganado pastoreaba enella con una carga animal de2 animales/ha. Las muestras setomaron en un área de 35 x 70 m quese delimitó en el centro de la parcela.
Muestreo de lombricesLas fechas de muestreo se escogieronpara cubrir, de manera precisa, unciclo estacional completo. Laslombrices se muestrearon desdeoctubre de 1995 hasta octubre de1996 en las siguientes fechas: (a) enoctubre de 1995 y (b) cada 2 meses,entre febrero y octubre de 1996.
El método empleado pararecolectar las lombrices se adaptó delaplicado en el programa de Biología yFertilidad de los Suelos Tropicales(TSBF, acrónimo en inglés) deAnderson e Ingram (1993) y consistíaen la extracción rápida de unmonolito de suelo de 25 x 25 x 30 m.Este suelo se revisó manualmente enel campo sobre una lámina deplástico y las lombrices recolectadasse identificaron hasta el nivel deespecie (según Jiménez 1999), secontaron, y se devolvieron al lugar enque se hallaba el monolito junto contodo el suelo ya revisado.
Antes de la extracción delmonolito, se calculó la densidad de laespecie anécica grande, Martiodriluscarimaguensis Jiménez y Moreno, apartir del número de heces frescasdepositadas sobre una superficie de1 m2, cuyo centro se hallaba en elárea ocupada por el monolito. Larelación existente entre el número deindividuos y el de heces frescas
presentes sobre la superficie delsuelo se ha considerado como elmétodo más eficiente paradeterminar la densidad poblacionalde esta especie (Jiménez et al.1998a). En realidad, M.carimaguensis escapa fácilmentehacia los horizontes profundos delsuelo cuando se emplea el métodoclásico de extracción manual, acausa de su gran tamaño y de lasgalerías profundas que excava.
Los sitios de muestreo selocalizaron regularmente cada 5 men la red diseñada de 35 x 70 m.Este procedimiento permitía ubicarlos puntos en el espacio mediantecoordenadas (x, y), y facilitaba laaplicación de técnicas de estadísticaespacial. Para evitar que semuestrearan dos monolitosexactamente en el mismo punto, lasmuestras se desplazaban, de unafecha a otra, a lo largo de una espiralcuyo origen correspondía al puntomuestreado en la primera fecha.Esta diferencia de localización (unos30 cm) se consideró despreciable alcompararla con el tamaño total de laparcela estudiada; las coordenadasde los puntos, por tanto, fueronidénticas de una fecha a la otra. Entotal, se muestrearon 120 monolitosen cada fecha, número que excedeen mucho el tamaño mínimo de lamuestra, el cual se consideranecesario para realizar la mayoría delos análisis espaciales (Legendre yFortin 1989; Rossi 1996).
Propiedades del suelo ybiomasa radicularSe tomaron muestras de suelo alterminar el período en que se hizo elmuestreo de las lombrices(septiembre de 1996). La dinámicatemporal de la heterogeneidad delsuelo no se tuvo en cuenta porque esdifícil establecer vínculos entre ésta yla dinámica temporal reciente de la

384
El Arado Natural:…
distribución de las lombrices. Esposible, en realidad, que tanto laslombrices como las propiedades delsuelo se influencien mutuamente conun determinado intervalo temporal,que es el tiempo necesario para quelas estructuras influyan en elambiente cercano o para que losindividuos ocupen áreas favorables.En consecuencia, es posible queestos dos compartimentos (laslombrices y las propiedades del suelo)no manifiesten simultáneamentepatrones espacio-temporales. Esteestudio partió, por tanto, delsupuesto de que el diseño delmuestreo arrojaría dos tipos posiblesde resultados, que corresponderían adistintos tipos de interacciones entreel suelo y la fauna contenida en él.
- En primer lugar, la distribuciónde las lombrices es estable en eltiempo y muestra una correlaciónmuy fuerte con los parámetrosdel suelo, lo que indica que laheterogeneidad del suelopresenta una inercia temporalalta e influye en la distribuciónde los organismos durante todo elperíodo de muestreo (hipótesisdel equilibrio).
- En segundo lugar, laspoblaciones de lombrices sonmóviles y su distribución no serelaciona con los parámetros delsuelo o, si existe, es muy débil;esto significa que los patrones dedistribución espacial de lacomunidad y del suelo puedeninfluirse mutuamente en unsistema que no está en equilibrio.
La biomasa de las raíces secalculó también en esta misma fechadel muestreo del suelo con el fin deconocer el efecto que causan laslombrices en el crecimiento de lasplantas. En ningún caso se tomaronmuestras de la biomasa de la parteaérea de las plantas, porque elpastoreo intensivo influye más en ella
que la actividad de las lombrices o lascaracterísticas del suelo. Lasmuestras que se tomaron del suelo yde las raíces provenían solamente dela capa de 0 a 10 cm, porque durantela época lluviosa la mayoría de laslombrices están activas a esaprofundidad (Jiménez y Decaëns2000).
La resistencia a la penetración semidió en cuatro profundidades (0,2.5, 5 y 10 cm) con un penetrómetroDAIKI 5520 (diámetro del cono de1 cm, ángulo de 26°). La densidadaparente se determinó en las capasde 0 a 5 cm y de 5 a 10 cm, partiendodel peso de un cilindro de suelo (5 cmde diámetro) que se había secadopreviamente durante una semana a75 °C. El C total se calculó en lacapa de 0 a 10 cm mediante elmétodo de digestión ácida y posteriordeterminación colorimétrica, enmuestras de 2 g de suelo secado alaire (Houba et al. 1988). Las raícesse recolectaron mediante el lavado yel tamizado (en malla de 0.053 mm)de muestras de suelo que habían sidotomadas con cilindros de 10 cm dediámetro hasta 10 cm deprofundidad, y que se habían secadoa 75 °C durante 48 horas.
Los puntos de muestreo selocalizaron en la misma red diseñadapara el muestreo de las lombrices.Aunque se tomaron muestrassolamente en la mitad de estos120 puntos, el total de 60 puntossigue siendo superior al que seconsidera suficiente para aplicar elanálisis estadístico espacial (Legendrey Fortin 1989; Rossi 1996).
Análisis estadísticos
Análisis triádico parcialEl análisis triádico parcial (ATP)
es un método específico paraconjuntos o matrices cúbicas de

Estructura Espacio-Temporal de una Comunidad de Lombrices...
385
datos. Este método es muy útilcuando se analiza una serie de tablasrelacionadas con t fechas demuestreo que tuvieron lugar en lamisma localidad (o estación) s. Encada fecha se establecen p variablesen cada estación o cada punto demuestreo y el ATP permite extraer laestructura de inercia que es común alas diferentes fechas. El métodoimplica la realización de tres pasossucesivos (Centofanti et al. 1989;Thioulouse y Chessel 1987), de loscuales se hicieron dos en esteestudio:
• El análisis de lainterestructura: este análisisproporciona una descripción delas estaciones en función de latipología de las fechas demuestreo y extrae la estructuraque es común a las diferentesfechas, para cada una de lasvariables p.
• El análisis de compromiso: esteanálisis permite describir lasestaciones en función de latipología de las variables, ytambién identificar las variablesque explican la estructura comúna las diferentes fechas demuestreo.
En este estudio, la tabla oconjunto original de datosrepresentaba seis tablas iniciales(correspondientes a las seis fechas demuestreo), cada una de ellascompuesta de 120 líneas (estación =punto de muestreo) y 5 columnas(variable = especie de lombriz). Lascoordenadas factoriales de los puntosde muestreo, extraídas en el análisisde la interestructura, serepresentaron en un mapa con el finde describir los patrones dedistribución espacio-temporal decada especie de lombriz. Asimismo,los patrones espaciales, comunes alas diferentes fechas, de los distintosconjuntos de especies se ubicaron en
un mapa con las coordenadasfactoriales suministradas por elanálisis de compromiso. Los análisisy sus cifras correspondientes fueronprocesados en el módulo STATIS y enlos módulos gráficos asociados delprograma informático (software)ADE-4 (Thioulouse et al. 1997).
Análisis de componentesprincipalesPara analizar los parámetros del
suelo y la biomasa de las raíces sellevó a cabo un análisis decomponentes principales (ACP). Latabla de datos comprendía 60 líneas(los puntos de muestreo) y8 columnas (los parámetros del sueloy la biomasa radicular). Se trazaronmapas con las coordenadasfactoriales de los puntos de muestreocon el propósito de identificar lospatrones de variación espacial, tantode las variables del suelo como de labiomasa de las raíces, siguiendo elmismo procedimiento descritoanteriormente para el ATP. Esteanálisis se realizó con el módulo ACPdel programa informático ADE-4(Thioulouse et al. 1997).
Cálculo de los valoresfaltantes mediante“kriging”Se calcularon las coordenadas
factoriales sobre el eje del ACP paralos 60 puntos en que no se tomaronmuestras de suelo ni de raíces, conel fin de comparar, posteriormente,los diferentes patrones espacialesrevelados en el análisis factorial.Este cálculo se hizo empleando latécnica de interpolación mediante el“kriging” puntual (Burgess y Webster1980; Yost et al. 1982a), unprocedimiento que tiene en cuentalos valores de la variable en lospuntos cercanos y su autocorrelacióntal como se describe en su

386
El Arado Natural:…
semi-variograma (es decir, en larepresentación de su semivarianza enfunción de la distancia entre lospares de puntos muestreados). Tantolos semi-variogramas como losvalores calculados con “kriging” seobtuvieron con el programainformático (software) VAR5, que fuedesarrollado por la Universidad deHawaii (Yost et al. 1989).
Autocorrelogramas deMoranLas estructuras espacio-
temporales extraídas del análisis devariable múltiple (multivariado)fueron probadas y descritas mediantelos autocorrelogramas de Moran(Moran 1950). El índice de Moran secalculó para las coordenadasfactoriales de los puntos de muestreoen los dos primeros ejes tanto delACP como del análisis decompromiso. Los datos se asignarona 14 clases de distancia (verCuadro 23-1, donde se muestran loslímites y el número de pares de
puntos de cada clase). El nivel designificación estadística en que seprobó cada coeficiente fue deP = 0.05. El correlograma entero seconsideró significativo cuando almenos uno de los coeficientes lo fueal nivel de probabilidad corregidode Bonferroni: 0.003 = 0.05/14(14 = número de clases de distancia)(Cooper 1968). La prueba estadísticade Kolmogorov-Smirnov se utilizópara comprobar la normalidad de ladistribución de la frecuencia de losdatos, antes de hacer el cómputo delos índices. Cuando fue necesarioreducir la asimetría de la distribuciónde los datos, se aplicó latransformación Box-Cox (Sokal yRohlf 1995). Los diversos módulosdel paquete estadístico “R Package” seusaron en todos estos pasos(Legendre y Vaudor 1991).
Prueba de MantelSe empleó la r estadística de la
prueba sencilla de Mantel (Mantel1967) para subrayar los patronescomunes tanto a la distribuciónespacio-temporal de las lombricescomo a la distribución espacial de laspropiedades del suelo y de la biomasaradicular. Esta prueba consiste en elcálculo y la comprobación de lacorrelación que existe entre dosmatrices de distancia que reflejan laestructura espacial de dos variablesdadas (Legendre y Fortin 1989;Legendre y Trousselier 1988). Secrearon cuatro matrices en este caso,que son las siguientes:
- EAW1 y EAW2, quecontienen las coordenadasfactoriales de las muestrassituadas en los dos primerosejes del análisis decompromiso; y
- SOIL1 y SOIL2, que contienenlas coordenadas factoriales delas muestras situadas en losdos primeros ejes del ACP
Cuadro 23-1. Límites inferior y superior ynúmero de pares de puntos encada una de las 14 clases dedistancia empleadas en elcómputo del correlograma.
Clase de Límite Límite Pares dedistancia inferior superior puntos
(m) (m) (no.)
1 0 5.6 2172 5.6 11.2 7403 11.2 16.8 6354 16.8 22.4 10445 22.4 28.0 7536 28.0 33.5 9887 33.5 39.1 7158 39.1 44.7 5549 44.7 50.3 558
10 50.3 55.9 39011 55.9 61.5 24012 61.5 67.1 19613 67.1 72.7 8614 72.7 78.3 24

Estructura Espacio-Temporal de una Comunidad de Lombrices...
387
(obtenidas una vez aplicadoel “kriging”).
De este modo, la prueba deMantel se llevó a cabo entre cadauna de las matrices EAW y SOIL.
Se puede lograr una estadísticasignificativa en la prueba de Mantel apartir de un factor inesperado queejerza un efecto similar y simultáneoen ambas variables estudiadas(Legendre y Fortin 1989; Rossi yQuénéhervé 1998). Se empleó, portanto, en lo sucesivo la estadística dela prueba parcial de Mantel (Smouseet al. 1986) para comprobar estaeventualidad y resaltar lasverdaderas relaciones entre lasmatrices EAW y SOIL. Esta pruebaconsiste en probar cada matriz EAWrespecto a cada matriz SOIL,condicionando la prueba a unamatriz de distancia SPACE que seconstituye a partir de lascoordenadas geográficas de lospuntos de muestreo. Se presume, deeste modo, que la matriz SPACErepresenta los patrones espacialescomunes a todas las variablesanalizadas (Legendre y Troussellier1988; Rossi y Quénéhervé 1998).
El módulo “Mantel 3.0” delpaquete estadístico “R Package” deLegendre y Vaudor (1991) se utilizópara llevar a cabo la prueba simplede Mantel y la prueba parcial deMantel. La significación estadísticase verificó mediante una prueba depermutación (1000 permutaciones) aun nivel de P = 0.05 (Legendre yFortin 1989).
Resultados
Estructura espacio-temporalde la comunidad de lombricesLa comunidad de lombricesestudiada estaba constituida, en sumayor parte, por especies geófagas
(endógeas, sensu Bouché 1977) ycontaba con la presencia de unaespecie polihúmica (sensu Lavelle1979) que ingiere suelo rico en MO(Ocnerodrilidae sp.), de una especiemesohúmica (sensu Lavelle 1979)que ingiere suelo con un contenidobajo de MO (Andiodrilus sp.), y deuna especie intermedia entre lasanteriores (Glossodrilus sp.). Laespecie de tamaño grandeM. carimaguensis, que ingiere tantohojarasca fresca como suelo yconstruye galerías subverticales,representaba la especie anécica(sensu Bouché 1977), mientras queuna sola especie, Aymara sp., seconsideró como transformadora de lahojarasca (epígea, sensu Bouché,1977) (Jiménez et al. 1998b).
La densidad media de laslombrices fue de 4.8 y95.4 individuos/m2 durante febrero yjunio, respectivamente. Las especiesendógeas predominaron en ladensidad poblacional:Ocnerodrilidae sp. (≅ 35% de ladensidad total de lombrices),Andiodrilus sp. (≅ 35%) yGlossodrilus sp. (≅ 20%). Sólo el 2%de la densidad total de las lombricesestaba representada porM. carimaguensis, pero esta especiedominó en la estructura de lacomunidad en cuanto a la biomasa,dado su gran tamaño encomparación con el de las demásespecies.
Análisis de lainterestructuraEl valor propio (“eigenvalue”) del
primer eje sobresalió entre lossiguientes, en tanto que eldecrecimiento global fue suave(Figura 23-1,A); este comportamientosugirió que solamente el primer ejepodía escogerse para el análisis decompromiso.

388
El Arado Natural:…
123123123
12341234123412341234
Figura 23-1. Resultados obtenidos en el análisis de la tabla de interestructura: (A) Diagrama de losvalores propios o “eigenvalues”. (B) Círculo de correlación con las fechas de muestreoproyectadas en el plano factorial definido por los dos primeros ejes del ACP de la matriz deinterestructura. (C) Mapas de las coordenadas factoriales de los 120 puntos de muestreosobre el primer componente del análisis para cada una de las cinco especies de lombriz(los círculos y los cuadrados representan, respectivamente, las coordenadas positivas ynegativas; la superficie del símbolo es proporcional al valor correspondiente obtenido).
1212
12
123412341234
11
11
1234123412341234123
123123
121212
1212
1212
11
1212
1234123412341234
1212
11
12
1212
12
1212
1234123412341234
123123123
123123123
121211
1
123456123456123456123456123456123456
1234512345123451234512345
1234567123456712345671234567123456712345671234567
1234512345123451234512345
121212345
123451234512345123451212
123451234512345123451234512345
123123123
1212
12345123451234512345
12341234123412341234
1234123412341234
1234123412341234
123451234512345123451234512345
1231231231234
1234123412341234
123412341234
12345123451234512345
123123123
123412341234123412341234
12341234
123123123
1234123412341234
123456123456123456123456123456
123412341234
123123123
1234123412341234
123123123123
123412341234
1234123412341234
1234123412341234
1234123412341234
123123123
123123
1234123412341234123123123
1234512345123451234512345
123123123123123123
123123123123
123123123
123123123
123123123
1231231231234
123412341234
1234567123456712345671234567123456712345671234567
1234123412341234
123456123456123456123456123456123456123456
1231231231234
123412341234
1234123412341234
123412341234
123123123
121212
123123123
123412341234
123412341234
1234123412341234
123123123
123456123456123456123456123456123456
1212
1212
123412341234123412345
123451234512345
123123
123123123 1234
123412341234
123123123
123123123
12 1
1212
123123
1212
1234512345123451234512345 123
123123
123456123456123456123456123456123456
1212
1234123412341234
1234567123456712345671234567123456712345671234567
12341234123412341234
123456123456123456123456123456123456
1212
123456712345671234567123456712345671234567
123123123
1212
1212
123123123
123123123123
1234123412341234
123123123123
1234512345123451234512345
123412341234
123412341234
123412341234
123123123
11
123123123
123123123
123123123
123123123
123123123
1234123412341234
1234512345123451234512345
12341234123412341234
123412341234 1234
123412341234
1234512345123451234512345
12345123451234512345
1234123412341234123412341234
121212
12121212121212
1212
123123123
1212
1212
1234123412341234
123412341234
123123
123123123
123123123
123123123
123123123
123123123
123123123123123123
123412341234
123123123
1212
1212
1212
121212
121212
12341234123412341234
123412341234 1234
1234123412341234
1234512345123451234512345
1234512345123451234512345
12345123451234512345
123451234512345123451234512345
1234512345123451234512345
123456123456123456123456123456123456
123123123
123456712345671234567123456712345671234567
123123123123
1212
11
1212
1234123412341234123412341234
123451234512345123451234512345
1234123412341234
1234123412341234
1234512345123451234512345
123123123
1212
1234123412341234
1234123412341234 123
123123
1234512345123451234512345
Eje 2 (17.8%)
Eje 1 (20.2%)
II/96
VI/96
IV/96
X/96
X/95
VIII/96
B
A
Glossodrilus sp.
Aymara sp.
Andiodrilus sp.
Martiodrilus carimaguensis
Ocnerodrilidae sp.
C
N
123123
11
1
1212
12
1212
1212
12
1212
1212
1212
1212
1212
12
1212
1212
1
1212

Estructura Espacio-Temporal de una Comunidad de Lombrices...
389
El primer eje (20.2% de la inerciatotal) correspondió claramente alllamado “efecto de tamaño” entre lastablas (todas las variables tuvieron elmismo signo en el eje 1)(Figura 23-1,B), y esto significa quelas correlaciones entre las tablasfueron todas positivas. En otraspalabras, no hubo inversión de laestructura espacial de una fecha aotra y los patrones observados en ladistribución espacial de las lombricesfueron, al menos en parte, comunes atodas las fechas de muestreo. Lavariabilidad de los datos restantes nofue afectada por las estructurasespacio-temporales, lo que refleja laalta variabilidad espacio-temporalglobal de los conjuntos de especies.
Los mapas trazados con lascoordenadas factoriales de los puntosde muestreo (Figura 23-1,C) revelaronla existencia de patrones espacialesagregados que eran estables en eltiempo, al menos en la escalatemporal considerada en este estudio.La estructura agregada, enparticular, fue muy clara respecto ala distribución espacio-temporal deGlossodrilus sp., especie que presentóun área o parche de densidad altamuy bien definida en la parte inferiorizquierda de la red diseñada en laparcela. En cambio, la densidad deAndiodrilus sp. fue alta en la mitadderecha de dicha red y baja en sumitad izquierda. Las estructurasespaciales no fueron muy claras en elcaso de M. carimaguensis, aunque seobservó un área más densa en lamitad izquierda de la zonamuestreada. Los parches dedensidad alta que presentaron lasespecies Glossodrilus sp. yM. carimaguensis eran, en parte,superponibles y se oponíanclaramente al parche de altadensidad de Andiodrilus sp. De lasdemás especies se observaronpequeños parches distribuidos portoda la parcela (como paraOcnerodrilidae sp.) o agrupados en la
parte superior de ésta (como paraAymara sp.)
Primer análisis decompromisoEl descenso fuerte de los valores
propios de los ejes justificó laelección de sólo los dos primeros ejespara la interpretación de los datos(Figura 23-2,A). Este análisis decompromiso se hizo con el primer ejedel análisis de la interestructura, quesólo representaba el 20.2% de lainercia total de la matriz de datos.
El primer eje (34.2% de la inerciatotal) estuvo caracterizado, en granparte, por una oposición entre dosconjuntos de especies: por un lado,Andiodrilus sp. y Aymara sp. y, por elotro, Glossodrilus sp. junto conOcnerodrilidae sp. y M.carimaguensis (Figura 23-2,B). Elmapa de las coordenadas de lospuntos de muestreo sobre el primereje del análisis reveló la presencia dedos zonas diferenciadas: la primera,situada en la parte inferior izquierdade la parcela, estaba dominada porel conjunto de las especiesGlossodrilus sp.-Ocnerodrilidae sp.-M. carimaguensis; la segunda, en laparte superior derecha, estabaocupada, en su mayor parte, porAndiodrilus sp. y Aymara sp.(Figura 23-2,C). El correlograma deMoran, calculado sobre lascoordenadas factoriales de lospuntos de muestreo, fue globalmentesignificativo (P < 0.001, Figura 23-3),donde las cuatro primeras clases dedistancia tuvieron valoresindividuales positivos, que fueronnegativos desde la sexta clase enadelante. Estos resultados indicanque los conjuntos de especiesdescritos se localizaban en parchesagregados que medían unos 20 m.
El segundo eje (23.0% de lainercia total) mostró la separaciónentre las especies pequeñas
390
El Arado Natural:…
12345671234567123456712345671234567123456712345671234567
1234512345123451234512345
Figura 23-2. Resultados de la tabla del análisis de compromiso: (A) Diagrama de los valores propios o“eigenvalues”. (B) Círculo de correlación con las variables (especies) proyectadas en elplano factorial definido por los dos primeros ejes del ACP de la matriz de compromiso.(C) Mapas de las coordenadas factoriales de los 120 puntos de muestreo sobre losprimeros dos componentes del análisis (los círculos y los cuadrados representan,respectivamente, las coordenadas positivas y negativas; la superficie del símbolo esproporcional al valor correspondiente obtenido).
123123123123
1234123412341234
123412341234
123412341234
123412341234
123123123
1234123412341234
123412341234123412345
12345123451234512345
1234123412341234
123123
1212
123123123
1234512345123451234512345
123456123456123456123456123456123456
121212
1234123412341234
1231231231231234
1234123412341234
121212
1212
1212
1234123412341234
123456123456123456123456123456123456
123456123456123456123456123456123456
123456712345671234567123456712345671234567
121212341234123412341234
123451234512345123451234
1234123412341234
123123123
123123123
1234123412341234
1234512345123451234512345
123456123456123456123456123456123456
1234123412341234
12345678123456781234567812345678123456781234567812345678
12341234123412341234
123123123
1212
123123123
123123123
123123123
123123123
123456123456123456123456123456123456
1234567123456712345671234567123456712345671234567
12341234123412341234
12341234123412341234
1234567123456712345671234567123456712345671234567
1234123412341234
1234567812345678123456781234567812345678123456781234567812345678
123123123
1234512345123451234512345
123123123
123456123456123456123456123456123456
1234512345123451234512345
123412341234
1212
1212
123123
123451234512345123451234512345
123123123
12
1212
123456712345671234567123456712345671234567
123412341234
123123123123
1234123412341234
123456123456123456123456123456123456
123456123456123456123456123456123456
123123
1234123412341234
123412341234
1212
123412341234
123412341234
1234123412341234
123123123
123123123
123123123
12341234123412341234
123456123456123456123456123456123456
1234512345123451234512345
123451234512345123451234512345
123456781234567812345678123456781234567812345678123456781234567812345678
123456123456123456123456123456123456123456
1234512345123451234512345
1212
1212
1212
1234512345123451234512345123456123456123456123456123456123456
1212
123123123
1234512345123451234512345
123456123456123456123456123456
123123
123456123456123456123456123456123456
1234567123456712345671234567123456712345671234567
123456123456123456123456123456
123412341234
12345678123456781234567812345678123456781234567812345678
1212
1212
Eje 2 (23.0%)
Eje 1 (34.2%)A
B
C
Glossodrilus sp.
Aymara sp.
Andiodrilus sp.
M. carimaguensis
Ocnerodrilidae sp.
Eje 1
Eje 2
N123456789123456789123456789123456789123456789123456789123456789123456789123456789
123456123456123456123456123456123456
12345123451234512345
1234512345123451234512345
1234512345123451234512345
123456123456123456123456123456123456
12
1212
11
123456712345671234567123456712345671234567
123456123456123456123456123456123456
123456123456123456123456123456123456
11
1212
1212
1212
12

Estructura Espacio-Temporal de una Comunidad de Lombrices...
391
(Aymara sp. y Ocnerodrilidae sp.) ylas especies de mayor tamaño(Glossodrilus sp., Andiodrilus sp. yM. carimaguensis) (Figura 23-2,B). Adiferencia del patrón descrito para elprimer eje, el mapa trazado con lascoordenadas factoriales sobre esteeje no reflejó una estructura espacialclara (Figura 23-2,C). Sin embargo,se observaron pequeños parches decoordenadas positivas o negativas,las primeras en la parte inferiorderecha y en la superior derecha dela parcela, y las segundas en elcentro y en la parte inferior izquierdade ésta. El correlograma de Moranconfirmó la presencia de estasestructuras espaciales (P < 0.003,
Figura 23-3), aunque sólo aparecióuna autocorrelación positiva en lasegunda clase de distancia. De ahíque este eje reflejara la oposición quemanifiestan en el espacio lasespecies de diferente tamaño que selocalizaban en parches pequeños deunos 10 m.
Heterogeneidad del sueloLos valores mínimos y máximos quese obtuvieron para las propiedadesdel suelo y para la biomasa radicularse muestran en el Cuadro 23-2. Eldescenso de los valores propiosasociados con los ejes del ACP fuebrusco, lo que indicó que sólo los dosprimeros ejes debían tomarse parainterpretar los datos (Figura 23-4,A).
El primer eje (44.3% de la inerciatotal) presentó la oposición entre el Ctotal y la biomasa de las raíces, porun lado y, por el otro, entre algunasvariables que describen lacompactación del suelo (por ejemplo,la resistencia a la penetración y ladensidad aparente) (Figura 23-4,B).El mapa trazado con las coordenadasfactoriales reveló la presencia de dosáreas, fácilmente identificables, en
Figura 23-3. Correlogramas de Moran obtenidosa partir de las coordenadasfactoriales de los 120 puntos demuestreo sobre los dos primerosejes del ACP de la matriz decompromiso ( = valoressignificativos individuales al nivelde P < 0.05; = valores nosignificativos; el valor de P serefiere al nivel de significaciónglobal del correlograma).
0.50
0.25
0
-0.25
-0.50
0.50
0.25
0
-0.25
-0.501 2 3 4 5 6 7 8 9 10 1112 13 14
Eje 1 del compromiso
Eje 2 del compromiso
Clases de distancia
P < 0.003
P < 0.001
Indi
ce d
e M
oran
Indi
ce d
e M
oran
Cuadro 23-2. Valor medio de las propiedadesdel suelo y de la biomasaradicular (ss = suelo seco,rs = raíces secas).
Propiedades Capa Mín. Máx.(cm)
C orgánico total (%) 0-10 2.2 3.6
Densidad aparente 0-5 0.82 1.43(g/cm3 de ss) 5-10 1.07 1.77
Resistencia a la 0 0 4.3penetración 2.5 0 6.0
(kg/cm2) 6 0.7 8.310 4.0 16.820 8.8 17.830 11.3 23.040 15.2 25.050 16.3 25.0
Biomasa radicular 0-10 21.4 141.0(g/m2 de rs)
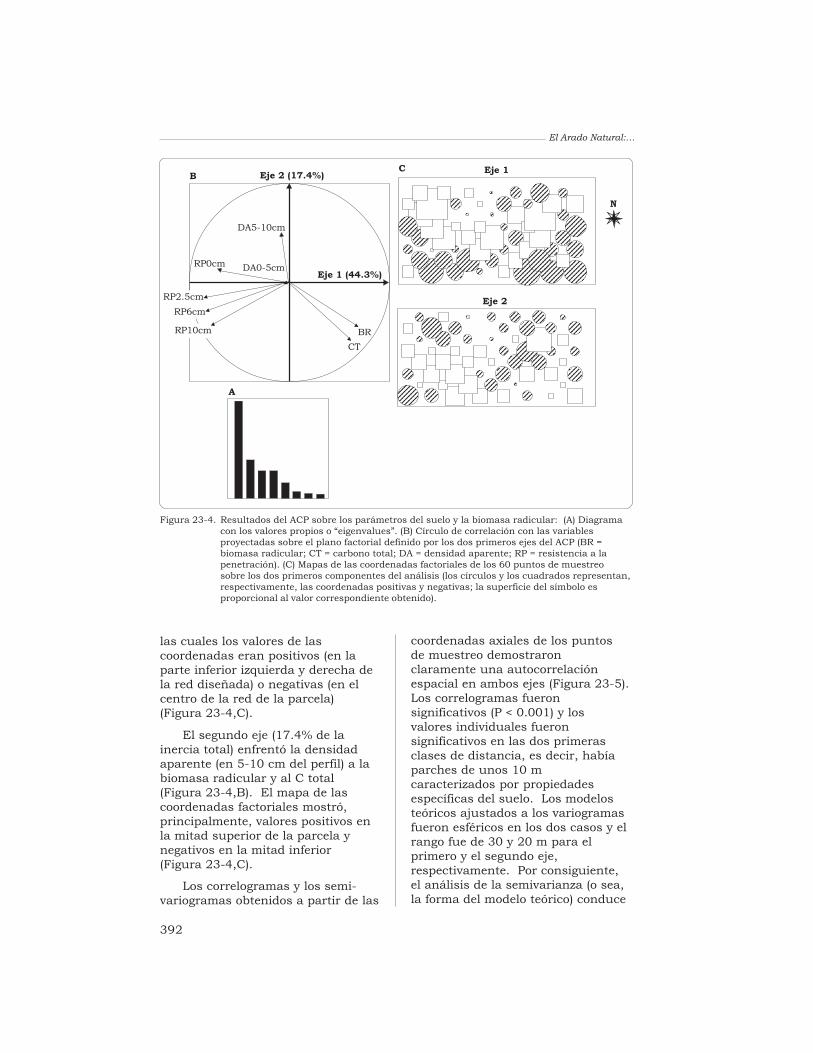
392
El Arado Natural:…
las cuales los valores de lascoordenadas eran positivos (en laparte inferior izquierda y derecha dela red diseñada) o negativas (en elcentro de la red de la parcela)(Figura 23-4,C).
El segundo eje (17.4% de lainercia total) enfrentó la densidadaparente (en 5-10 cm del perfil) a labiomasa radicular y al C total(Figura 23-4,B). El mapa de lascoordenadas factoriales mostró,principalmente, valores positivos enla mitad superior de la parcela ynegativos en la mitad inferior(Figura 23-4,C).
Los correlogramas y los semi-variogramas obtenidos a partir de las
coordenadas axiales de los puntosde muestreo demostraronclaramente una autocorrelaciónespacial en ambos ejes (Figura 23-5).Los correlogramas fueronsignificativos (P < 0.001) y losvalores individuales fueronsignificativos en las dos primerasclases de distancia, es decir, habíaparches de unos 10 mcaracterizados por propiedadesespecíficas del suelo. Los modelosteóricos ajustados a los variogramasfueron esféricos en los dos casos y elrango fue de 30 y 20 m para elprimero y el segundo eje,respectivamente. Por consiguiente,el análisis de la semivarianza (o sea,la forma del modelo teórico) conduce
Figura 23-4. Resultados del ACP sobre los parámetros del suelo y la biomasa radicular: (A) Diagramacon los valores propios o “eigenvalues”. (B) Círculo de correlación con las variablesproyectadas sobre el plano factorial definido por los dos primeros ejes del ACP (BR =biomasa radicular; CT = carbono total; DA = densidad aparente; RP = resistencia a lapenetración). (C) Mapas de las coordenadas factoriales de los 60 puntos de muestreosobre los dos primeros componentes del análisis (los círculos y los cuadrados representan,respectivamente, las coordenadas positivas y negativas; la superficie del símbolo esproporcional al valor correspondiente obtenido).
1234123412341234
123123123
12341234123412341234
123456712345671234567123456712345671234567
1234512345123451234512345
123456123456123456123456123456123456
1212
123123
1234123412341234
123123123
12
123123123
1234512345123451234512345
123412341234
123412341234123412341234512345123451234512345123456123456123456123456123456123456
1234123412341234
1234567123456712345671234567123456712345671234567
123456712345671234567123456712345671234567
123123123
1212
1234123412341234
1234123412341234
123123123123
123456123456123456123456123456123456
1212
123123123
123123123123
123123
1212
1234123412341234
1231231234
123412341234
12345123451234512345
123123123
1234123412341234
123123123
1212
1234123412341234
1234123412341234
123456123456123456123456123456123456
123123123
123412341234
12345123451234512345
1234512345123451234512345
123123123
Eje 1 (44.3%)
A
C Eje 1
N123451234512345123451234512345
12341234123412341234
1234512345123451234512345
123123
1234123412341234
123456123456123456123456123456123456
123456123456123456123456123456123456
123123123
1234512345123451234512345
123412341234
123451234512345123451234512345
1234123412341234
123123123
Eje 2 (17.4%)B
DA5-10cm
DA0-5cmRP0cm
BRCT
Eje 2RP2.5cmRP6cm
RP10cm
12
1
11
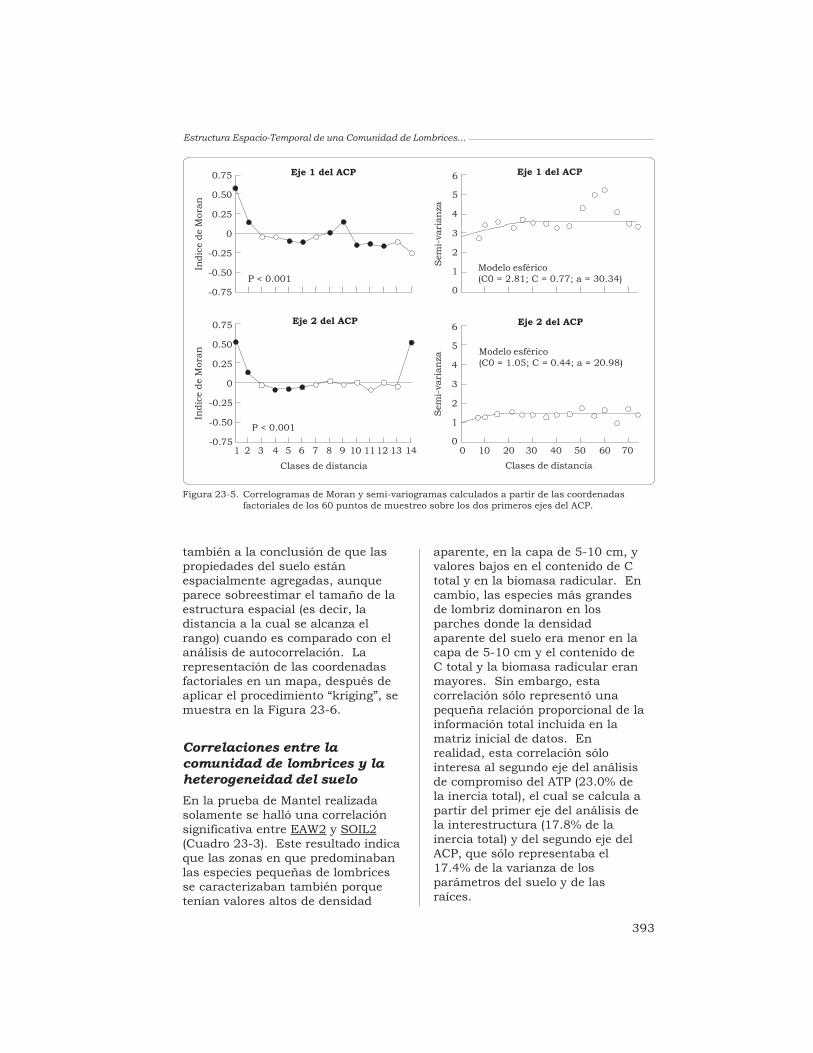
Estructura Espacio-Temporal de una Comunidad de Lombrices...
393
también a la conclusión de que laspropiedades del suelo estánespacialmente agregadas, aunqueparece sobreestimar el tamaño de laestructura espacial (es decir, ladistancia a la cual se alcanza elrango) cuando es comparado con elanálisis de autocorrelación. Larepresentación de las coordenadasfactoriales en un mapa, después deaplicar el procedimiento “kriging”, semuestra en la Figura 23-6.
Correlaciones entre lacomunidad de lombrices y laheterogeneidad del sueloEn la prueba de Mantel realizadasolamente se halló una correlaciónsignificativa entre EAW2 y SOIL2(Cuadro 23-3). Este resultado indicaque las zonas en que predominabanlas especies pequeñas de lombricesse caracterizaban también porquetenían valores altos de densidad
aparente, en la capa de 5-10 cm, yvalores bajos en el contenido de Ctotal y en la biomasa radicular. Encambio, las especies más grandesde lombriz dominaron en losparches donde la densidadaparente del suelo era menor en lacapa de 5-10 cm y el contenido deC total y la biomasa radicular eranmayores. Sin embargo, estacorrelación sólo representó unapequeña relación proporcional de lainformación total incluida en lamatriz inicial de datos. Enrealidad, esta correlación sólointeresa al segundo eje del análisisde compromiso del ATP (23.0% dela inercia total), el cual se calcula apartir del primer eje del análisis dela interestructura (17.8% de lainercia total) y del segundo eje delACP, que sólo representaba el17.4% de la varianza de losparámetros del suelo y de lasraíces.
Figura 23-5. Correlogramas de Moran y semi-variogramas calculados a partir de las coordenadasfactoriales de los 60 puntos de muestreo sobre los dos primeros ejes del ACP.
0.75
0.50
0.25
0
-0.25
-0.50
-0.75
1 2 3 4 5 6 7 8 9 10 1112 13 14
Eje 1 del ACP Eje 1 del ACP
Clases de distancia
P < 0.001
P < 0.001
Indi
ce d
e M
oran
Indi
ce d
e M
oran
0.75
0.50
0.25
0
-0.25
-0.50
-0.750 10 20 30 40 50 60 70
Clases de distancia
6
5
4
3
2
1
0
6
5
4
3
2
1
0
Sem
i-va
rian
zaS
emi-
vari
anza
Modelo esférico(C0 = 2.81; C = 0.77; a = 30.34)
Modelo esférico(C0 = 1.05; C = 0.44; a = 20.98)
Eje 2 del ACP Eje 2 del ACP

394
El Arado Natural:…
La significación de la estadísticade Mantel se examinó, una vezconsiderado el efecto de la matrizSPACE, para establecer si lacorrelación entre EAW2 y SOIL2 eraverdadera o dudosa. La pruebaparcial de Mantel [EAW2.SOIL2].SPACE (que es una prueba parcialentre EAW2 y SOIL2 condicionada aSPACE) se llevó a cabo para verificarla validez de la correlaciónobservada. El coeficiente r obtenidofue 0.42 (P = 0.039), cuyo significadoes que la similitud observada entrelos dos patrones de organizaciónespacial correspondía a unacorrelación fuerte.
Figura 23-6. Mapas de las coordenadas factoriales de los 120 puntos de muestreo sobre los dos primerosejes del ACP después de la interpolación puntual de “kriging” de los puntos no muestreados(valores ausentes). (Los círculos y los cuadrados representan, respectivamente, lascoordenadas positivas y negativas; la superficie del símbolo es proporcional al valorcorrespondiente obtenido).
123412341234
1234567812345678123456781234567812345678123456781234567812345678
123412341234
123412341234
123123123
123123123
123123123
123456123456123456123456123456123456
1212
123123123123
1212
1212
123123
12345123451234512345
1234123412341234
123456123456123456123456123456123456123456123456123456123456123456123456
123456123456123456123456123456123456
1234123412341234
12345123451234512345
123123123
123456123456123456123456123456123456
123456123456123456123456123456123456
123456123456123456123456123456123456123456
123456123456123456123456123456123456
1234567812345678123456781234567812345678123456781234567812345678
123123123
123123123
1234512345123451234512345
12345123451234512345
1234123412341234
123456123456123456123456123456123456
123456789123456789123456789123456789123456789123456789123456789123456789
1234567812345678123456781234567812345678123456781234567812345678
123456123456123456123456123456123456
123456123456123456123456123456123456
123123123
1234512345123451234512345123451234512345123451234512345
123456712345671234567123456712345671234567123456712345
123451234512345
1234123412341234
1234123412341234
123123123
123123123
1234123412341234
123456789123456789123456789123456789123456789123456789123456789123456789123456789
123456123456123456123456123456123456123456
1234512345123451234512345
123456123456123456123456123456123456123456123456123456123456123456
1234567812345678123456781234567812345678123456781234567812345678
123456123456123456123456123456123456
12345671234567123456712345671234567123456712345671234567
1234567812345678123456781234567812345678123456781234567812345678
1234123412341234
1234123412341234
1234123412341234
12341234123412341234
1234123412341234
12341234123412341234123412341234
123123123
1234123412341234
1234123412341234
1234123412341234
1234123412341234
123412341234
1234123412341234
1234123412341234
1234123412341234
1234123412341234
1234123412341234
12345612345612345612345612345612345
1234512345123451234512345
12345123451234512345
1234567812345678123456781234567812345678123456781234567812345678
123451234512345123451234512345
12345123451234512345
1234567812345678123456781234567812345678123456781234567812345678
123456123456123456123456123456
1234512345123451234512345
1212
123456123456123456123456123456
1234512345123451234512345
123123123
123123
1212
1212
123123123
12341234123412341234
1234512345123451234512345
1234512345123451234512345123123123
12345123451234512345123451234512345123451234512345
12345123451234512345
12341234123412341234512345123451234512345
123456712345671234567123456712345671234567123456712341234123412341234
123456123456123456123456123456
123123
121212
121212
12341234123412341234123456123456123456123456123456123456
1212
1212
1212
12
1
1212
11
12
Eje 1N
Eje 2
1
123456123456123456123456123456123456
12345678123456781234567812345678123456781234567812345678
Cuadro 23-3. Coeficientes r estandarizados deMantel y probabilidad asociada.(EAW1 y EAW2 = matrices dedistancia para las coordenadasfactoriales de los puntos demuestreo sobre el primero y elsegundo ejes del análisis decompromiso, respectivamente;SOIL1 y SOIL2 = matrices dedistancia para las coordenadasfactoriales de los puntos demuestreo sobre el primero y elsegundo ejes del ACP,respectivamente.)
r estándar Probabilidada
de Mantel
EAW1.SOIL1 0.079 0.332 NSEAW1.SOIL2 0.170 0.257 NSEAW2.SOIL1 -0.266 0.276 NSEAW2.SOIL2 0.424 0.039*
a. NS = no significativo; * significativo aP < 0.05.

Estructura Espacio-Temporal de una Comunidad de Lombrices...
395
DiscusiónEste estudio demuestra, en formaexplícita, la existencia de un conjuntoestructurado en las comunidades delombrices, al menos en la escalatemporal referida al período delestudio. Esta estructura corresponde,en gran parte, a la alternancia deparches donde predominan conjuntosde especies particulares. El patrónglobal se puede describir en diferentesniveles de organización espacial, quese encajan unos dentro de otros enun momento dado y quecorresponden a distintoscomponentes del análisis decompromiso. Estas estructurasespacio-temporales han sido yadescritas para algunas comunidadesde lombrices de las sabanas de Costade Marfil (Rossi 1998; Tondoh 1998) yde pastizales de Colombia (Jiménez1999). La escala espacial a la que seexpresan estas estructuras (unos20 m) concuerda con la quepresentan otros autores respecto a ladistribución espacial de las lombricesen estudios diseñados para una solafecha de muestreo (Nuutinen et al.1998; Rossi et al. 1995, 1997). Esimportante recalcar que lasestructuras descritas sólo representanun porcentaje bajo de la variabilidadtotal de la matriz de datos (57.0% del20.2% que fue explicado por el primereje de la interestructura). Esosresultados indican que los conjuntosde especies son relativamente móviles,aunque se observa una estabilidadespacial relativa en ciertas áreas de laparcela experimental. Esta falta deestabilidad en el patrón espacio-temporal de las lombrices reflejaríauna situación en la que el sistema noestá en equilibrio. De este modo, laoposición entre los conjuntos deespecies sería el efecto de unfenómeno de exclusión competitiva, otambién de una segregación espacialen el caso de que las lombricesrespondan a factores ambientales que
no son estables ni en el tiempo ni enel espacio (es decir, cuando unacaracterística de estos mismosfactores es que no están en equilibrio).
El primer eje del análisis decompromiso expresó, principalmente,la segregación espacial entreAndiodrilus sp. y Glossodrilus sp.Este resultado fue observado ya enuna sabana nativa y en un pastizal demuchos años de edad sembrado congramínea y leguminosa en Carimagua(Jiménez 1999). Ambas especies sonendógeas, presentan característicasbiométricas similares y, por tanto,muestran un alto índice desuperposición de nichos, que fuedesarrollado (Jiménez 1999) a partirde los datos de biometría y dedistribución vertical. Porconsiguiente, se puede interpretar laoposición espacial entre estas dosespecies como el resultado de unfenómeno de exclusión competitiva.De ahí que, en los parches en quepredomina una u otra especie, lacoexistencia con otras especies debeser un efecto de los mecanismos dedistribución del nicho ecológico.Parece que esta hipótesis es bastantereal, ya que las especies que coexistentienen tamaños diferentes, distintoshábitos alimenticios y diversasestrategias adaptativas. No obstante,se necesitan aún más datos ecológicosdetallados (p.ej., el régimenalimenticio) para obtener lossiguientes resultados:
- establecer el grado deseparación de nichos quepresenta realmente estacomunidad de lombrices;
- identificar las especies quecompiten entre sí, losmecanismos de exclusión quese observan, el mejorcompetidor encontrado, y losfactores ambientales quefacilitan la presencia de otrasespecies.

396
El Arado Natural:…
El segundo eje manifestó laoposición entre las especies más omenos pequeñas (Ocnerodrilidae sp.y Aymara sp.) y las grandes(Glossodrilus sp., Andiodrilus sp. yM. carimaguensis). Se supone quelos hábitos de las especies grandesson diferentes de los que tienen laspequeñas y que probablemente poresto no compiten entre sí. De estemodo, la alternancia de parches quedescribe este eje se puede interpretarcomo la respuesta de la comunidadde lombrices a la heterogeneidadambiental (o sea, segregaciónespacial). El estiércol del ganado,por ejemplo, puede tener efectospositivos en las especies pequeñas,las cuales ingieren, de preferencia,sustratos ricos en MO y puedenpresentar hábitos coprófagosoportunistas; las especies grandes,en cambio, se alimentangeneralmente de hojarasca, deraicillas muertas o de MOhumificada.
Los parches en quepredominaban o bien las especiesgrandes o las pequeñas seencontraban en áreas que secaracterizaban por tener propiedadesdel suelo y valores de biomasaradical específicos. Sin embargo,este resultado no permitía establecersi la distribución espacial de laslombrices determinaba laheterogeneidad del suelo o si ocurríalo contrario. Se pueden postular, porconsiguiente, dos hipótesis:
• Hipótesis A. Las lombricespueden influir en las propiedadesdel suelo, de modo que sudistribución espacial presentapatrones similares a los de laheterogeneidad del suelo. Estahipótesis supone que lasdiferentes especies de lombricesproducen heces de tamaño ypropiedades distintas y tienen,por tanto, efectos diferentes enlas propiedades del suelo
(Blanchart 1992; Derouard et al.1997). En Carimagua,Ocnerodrilidae sp. produceagregados pequeños y se creeque fragmenta las heces grandesdepositadas por otras especies(Jiménez et al. 1998b), mientrasque Aymara sp. produce,probablemente, pequeñosgránulos holorgánicos cuyoefecto en la estructura del sueloes reducido. Las especies detamaño grande producen hecesestables, compactas y ricas enMO que se degradan en pocosmeses por la actividad de otrosinvertebrados, los cualeshoradan en ellas galerías y lasfragmentan en agregados máspequeños y menos compactos(Decaëns 2000; Decaëns et al.1999b, 1999c). Puedepresentarse una regulacióneficiente de la estructura delsuelo en aquellos parchesdominados por las especiesgrandes, la cual propiciaría unamacroagregación más intensa ypermitiría, probablemente, quese acumule una reserva de Cdado que se protege físicamentela MO contra los procesos demineralización. Estos efectos enla MO y en las propiedadesfísicas del suelo tendrían,lógicamente, consecuencias en elcrecimiento de las plantas, lascuales se han observado en esteestudio como aumento de labiomasa de las raíces. Encambio, el predominio de lasespecies pequeñas, queproducen grandes cantidades deagregados poco estables ypequeños, puede hacer que elsuelo desarrolle una estructuradebilitada y sensible al pisoteodel ganado.
• Hipótesis B. La heterogeneidadespacial del suelo puede influiren el conjunto de las especies de

Estructura Espacio-Temporal de una Comunidad de Lombrices...
397
lombrices. Esta hipótesispostula que la segregaciónespacial es el mecanismo queconduce a la distribución delconjunto de especies que se haobservado. Las especiesendógeas grandes (Andiodrilussp. y Glossodrilus sp.) selocalizan en zonas donde ladisponibilidad del recurso tróficoes grande, por ejemplo dondehay un alto contenido de C. Losvalores altos de C estánasociados a una alta biomasa deraíces y a una agregación delsuelo también alta (Fisher et al.1994; Gijsman 1996; Gijsman yThomas 1995; Rao 1998).Cuando el contenido de MO deun parche es relativamente bajo,el suelo presentará unacapacidad de carga menor, lo quelleva a una reducción en elnúmero de especies endógeasgrandes y a un aumentoproporcional de las especiespequeñas. Además, el dominiode una especie polihúmica puedeacentuarse por su aptitud paravivir en un suelo compacto. Estahabilidad ya ha sido demostradapor una especie pequeñaafricana de la familia Eudrilidae(Blanchart et al. 1997; Rossi1998), que presenta similitudesmorfológicas y ecológicas conOcnerodrilidae sp.
Para elegir la hipótesis que seaproxima más a la realidad, convieneconsiderar que la correlación sóloafectó una pequeña parte de lavariabilidad inicial de los datos. Enrealidad, se podría suponer que lavariabilidad temporal de laspropiedades del suelo es muchomenor que la variabilidad de ladistribución espacial de laslombrices. Por tanto, si lascomunidades de lombricesrespondieran a la heterogeneidadambiental —es decir, si las especies
se localizan en parches quepresenten condiciones óptimas parala vida— se observaría unavariabilidad temporal baja de suspatrones de organización espacial,así como una correlación alta con laspropiedades del suelo. Ahora bien,la baja correlación observada indicaque las lombrices influyen en elsuelo, aunque estos efectos sondifíciles de observar por la altavariabilidad espacio-temporal delconjunto de las especies. Así pues,la correlación entre la comunidad yel ambiente sólo es perceptible dondeaparece una estabilidad temporalrelativa en el conjunto de lasespecies (es decir, las estructurasobservadas en el ATP).
Otros experimentos y variasobservaciones empíricas sobre elimpacto que hace M. carimaguensisen el funcionamiento del sueloapoyan la primera hipótesis. Losefectos producidos por esta especieen el suelo pueden serdesproporcionados si losrelacionamos con su densidad (dadosu peso individual tan alto) auncuando la especie no participesignificativamente en los patrones dedistribución aquí descritos. Lasheces superficiales producidas poresta especie tienen tambiénimportancia en los procesosrelacionados con la estructura delsuelo, la dinámica de la MO, laliberación de nutrientes, e inclusocon los puntos del suelo en que seincrementa la biomasa radicular yque están situados justo debajo deesas bioestructuras (Decaëns 2000;Decaëns et al. 1999c). En un trabajoreciente, Decaëns et al. (1999a)demostraron que la desaparición deM. carimaguensis conducía a unacompactación superficial rápida y aun descenso de los niveles de C en elsuelo y de la biomasa vegetal, tantode la parte aérea como de las raíces.Estos resultados concuerdan con la

398
El Arado Natural:…
idea general de que las lombricesregulan la estructura del suelo, ladinámica de la MO del suelo y elcrecimiento de las plantas, a laescala de las heces y del suelo quelas circunda (Edwards y Lofty 1978,1980; Lavelle y Martin 1992; Lavelleet al. 1998; Martin 1991). Sedescribió recientemente, a la escalamás grande de la parcela en elpastizal, el impacto que hace en laestructura del suelo de una sabanade Costa de Marfil la distribuciónespacial de las lombrices, cuandoestá caracterizada por una oposiciónde parches dominados por especiesde diferente tamaño (Rossi 1998).
Un aspecto sorprendente de esteestudio es que la mayor parte de lavariabilidad de las propiedades delsuelo y de la biomasa de las raícesno estaba ligada a la distribuciónespacio-temporal de las lombrices.Esto significa que son otros losfactores que desempeñan un papelpredominante en la regulación delfuncionamiento del suelo o que elefecto debido a las lombrices seexpresa en una escala espacio-temporal muy pequeña (por ejemplo,la escala de las bioestructuras queellas producen) que difícilmentepodía ser percibida por la reddiseñada para el muestreo en elpresente estudio. La biomasa de lasraíces, por ejemplo, parece tener unpapel predominante; el índice decorrelación r obtenido en unaprueba parcial de Mantel que se hizoadicionalmente, en la que serelacionó la biomasa radicular con elC total, fue de 0.62 (P < 0.001). Esteresultado apoya las conclusionesobtenidas en estudios recientesrealizados en los suelos de la sabanacolombiana, donde se observó queuna biomasa radicular alta conducea la acumulación de C en el suelo(Fisher et al. 1994; Rao 1998),contribuye así al mejoramiento de laestructura del suelo (Gijsman 1996;
Gijsman y Thomas 1995), y mantienefinalmente las propiedades físicas deéste (Gijsman y Thomas 1996).Tanto las raíces como las lombricesse consideran ingenieros delecosistema y la función de ambos enel suelo exhibe, al parecer,características convergentes, almenos para ciertos procesosespecíficos, por ejemplo laacumulación de C o la agregación delsuelo. Es necesario emprendernuevos estudios para establecer laactividad funcional de un amplioespectro de organismos del suelo:este trabajo permitirá definir losgrupos funcionales de los ingenierosdel ecosistema.
Los atributos funcionales de unaespecie o de un grupo funcionalpueden considerarse a diferentesniveles de observación, dentro deuna jerarquía de sistemas queinteractúan y se presentan encajadosunos dentro de otros. Cuando seasciende en esta jerarquía, losniveles sucesivos describirán losmismos procesos, pero con unadinámica cada vez más lenta y enescalas espaciales cada vez másgrandes (Allen y Starr 1982). Unmétodo muy utilizado para definir lafunción de las lombrices de tierra esla descripción del impacto que hacecada especie individualmente. Esteprimer paso es necesario, aunqueimplica el riesgo de ejecutar ciertasextrapolaciones que permitiríantrasladar los resultados obtenidos ala escala del ecosistema (el enfoque“bottom-up”: de abajo a arriba o delo inferior a lo superior, Anderson1995). En este trabajo se aplicó, sinembargo, el enfoque “top-down” (dearriba a abajo o de lo superior a loinferior) para deducir los atributosfuncionales de una especie delombriz partiendo de un estudio agran escala. Se identificaron lascorrelaciones, aunque no fue tareafácil llegar hasta ellas. Esta

Estructura Espacio-Temporal de una Comunidad de Lombrices...
399
situación es, probablemente,inevitable en el diseño experimentalempleado, en razón de los efectospredominantes de los factoresambientales que regulan tanto losprocesos del suelo como la dinámicade las comunidades en niveles muyaltos de la escala jerárquica(Anderson 1995). Ahora bien, losestudios de tipo “top-down” sobre laactividad funcional de losorganismos del suelo son necesariospara determinar el alcance a quepuede llegar la extrapolación, aescalas espacio-temporales másgrandes, de un estudio realizado anivel de los individuos.
En conclusión, pocos indiciospermiten afirmar que las lombricesinfluyen en las propiedades del sueloo que ocurre lo contrario. Algunosresultados indican, sin embargo, queel sistema se encontraba en unestado no equilibrado y que laspoblaciones móviles de lombricesejercían un influjo global en el suelo,el cual fue, por su parte, difícil dedetectar. Probar esta hipótesisimplica la realización de estudioscomparativos de la dinámicaespacio-temporal tanto de losconjuntos de lombrices como de laspropiedades del suelo, con el fin deidentificar los intervalos de tiempo yde establecer los enlaces de tipocausal que relacionan estos doscompartimentos.
AgradecimientosLos autores agradecen a R.J.Thomas, D.K. Friesen, E. Amézquitay C.G. Meléndez (CIAT) y aD. Cárdenas (CORPOICA) por suayuda científica y técnica. Elprograma global de los centros delGCIAI para el Manejo de Suelos,Agua y Nutrientes (SWNM, en inglés),perteneciente al consorcio del Manejo
de Suelos Acidos (MAS, en inglés)financió, en parte, este trabajo.Expresan además su más sinceroagradecimiento a L. Mariani yP. Lavelle (IRD), a D. Alard yP. Margerie (Universidad de Rouen) ya R. Thomas, I. Rao y J.J. Jiménez(CIAT) por sus comentarios útiles auna versión preliminar delmanuscrito.
ReferenciasAllen TFH; Starr TB. 1982. Hierarchy,
perspectives for ecologicalcomplexity. The University ofChicago Press, Chicago, IL, EstadosUnidos.
Anderson JM. 1995. Soil organisms asengineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linking speciesand ecosystems. Chapman and Hall,Londres. p 94-106.
Anderson JM; Flanagan P. 1989.Biological processes regulatingorganic matter dynamics in tropicalsoils. En: Coleman DC; Oades JM;Uheara G, eds. Dynamics of soilorganic matter in tropicalecosystems. University of HawaiiPress, Honolulu, Estados Unidos.p 97-123.
Anderson JM; Ingram J, eds. 1993.Tropical soil biology and fertility: ahandbook of methods. CAB, Oxford,Reino Unido.
Blanchart E. 1992. Restoration byearthworms (Megascolecidae) of themacroaggregate structure of adestructured savanna soil underfield conditions. Soil Biol Biochem24:1587-1594.
Blanchart E; Lavelle P; Braudeau E;Bissonnais YL; Valentin C. 1997.Regulation of soil structure bygeophagous earthworm activities inhumid savannas of Côte d’Ivoire.Soil Biol Biochem 29:431-439.

400
El Arado Natural:…
Bohlen PJ; Edwards CA. 1995. Earthwormeffects on N dynamics and soilrespiration in microcosms receivingorganic and inorganic nutrients. SoilBiol Biochem 27:341-348.
Bouché MB. 1977. Stratégieslombriciennes. En: Lohm U; PerssonT, eds. Soil organisms as componentof ecosystems. Ecol Bull25:122-132.
Burgess TM; Webster R. 1980. Optimalinterpolation and isarithmicmapping of soil properties; I: Thesemi-variogram and punctualkriging. J Soil Sci 31:315-331.
Centofanti M; Chessel D; Doledec S. 1989.Stabilité d’une structure spatiale etcompromis d’une analyse statistiquemulti-tableaux: Application à laphysico-chimie d’un lac réservoir.Rev Sci Eau 2:71-93.
Cooper DW. 1968. The significance level inmultiple tests made simultaneously.Heredity 23:614-617.
Decaëns T. 1999. Rôle fonctionnel etréponses aux pratiques agricoles desvers de terre et autres ingénieursécologiques dans les savanescolombiennes. Tesis (Doctorado).Université Pierre et Marie Curie,París.
Decaëns T. 2000. Degradation dynamicsof surface earthworm casts ingrasslands of the Eastern Plains ofColombia. Biol Fertil Soils32:149-156.
Decaëns T; Jiménez JJ; Lavelle P. 1999a.Effects of exclusion of the anecicearthworm Martiodriluscarimaguensis Jiménez and Morenoon soil properties and plant growthin grasslands of the Eastern Plainsof Colombia. Pedobiologia43:835-841.
Decaëns T; Mariani L; Lavelle P. 1999b.Soil surface macrofaunalcommunities associated withearthworm casts in grasslands of theEastern Plains of Colombia. ApplSoil Ecol 13:87-100.
Decaëns T; Rangel AF; Asakawa N;Thomas RJ. 1999c. Carbon andnitrogen dynamics in ageingearthworm casts in grasslands ofthe Eastern Plains of Colombia. BiolFertil Soils 30:20-28.
Derouard L; Tondoh J; Vilcosqui L;Lavelle P. 1997. Effects ofearthworm introduction on soilprocesses and plant growth. SoilBiol Biochem 29:541-545.
Edwards CA; Lofty JR. 1978. Theinfluence of arthropods andearthworms upon root growth ofdirect drilled cereals. J Appl Ecol15:789-795.
Fisher MJ; Rao IM; Ayarza MA; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage by introduceddeep-rooted grasses in the SouthAmerican savannas. Nature371:236-238.
Gijsman AJ. 1996. Soil aggregate stabilityand soil organic matter fractionsunder agropastoral systemsestablished in native savanna. AustJ Soil Res 34:891-907.
Gijsman AJ; Thomas RJ. 1995. Aggregatesize distribution and stability of anOxisol under legume-based andpure grass pasture in the easternColombian savannas. Aust J SoilRes 33:153-165.
Gijsman AJ; Thomas RJ. 1996.Evaluation of some physicalproperties of an Oxisol afterconversion of native savanna intolegume-based or pure grasspasture. Trop Grassl 30:237-248.
Haimi J; Huhta V. 1990. Effects ofearthworms on decompositionprocesses in raw humus forest soil:a microcosm study. Biol Fertil Soils10:178-183.
Houba VJC; van der Lee JJ; NovozamskyI; Walinga J. 1988. Soil and plantanalysis. En: Department of SoilScience and Plant Nutrition, ed. Soilanalysis procedures. 4a. ed.Agricultural University,Wageningen, Holanda. p 4-14.

Estructura Espacio-Temporal de una Comunidad de Lombrices...
401
Jiménez JJ. 1999. Estructura de lascomunidades y dinámica de laspoblaciones de lombrices de tierraen las sabanas naturales yperturbadas de Carimagua(Colombia). Tesis (Doctorado).Universidad Complutense deMadrid. 311 p.
Jiménez JJ; Decaëns T. 2000. Verticaldistribution of earthworms ingrassland soils of the Colombian“Llanos”. Biol Fertil Soils32:463-473.
Jiménez JJ; Moreno AG; Lavelle P;Decaëns T. 1998b. Populationdynamics and adaptive strategies ofMartiodrilus carimaguensis(Oligochaeta, Glossoscolecidae), anative species from the well-drainedsavannas of Colombia. Appl SoilEcol 9:153-160.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998a. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78:1946-1957.
Lal R; Akinremi OO. 1983. Physicalproperties of earthworm casts andsurface soil as influenced bymanagement. Soil Sci 135:114-123.
Lavelle P. 1979. Relations entre typesécologiques et profilsdémographiques chez les vers deterre de la savane de Lamto (Côted’Ivoire). Rev Ecol Biol Sol16:85-101.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P; Martin A. 1992. Small-scaleand large-scale effects of endogeicearthworms on soil organic matterdynamics in soils of the humidtropics. Soil Biol Biochem24:1491-1498.
Lavelle P; Blanchart E; Martin A; MartinS. 1993. A hierarchical model fordecomposition in terrestrialecosystems: application to soils ofthe humid tropics. Biotropica25:130-150.
Lavelle P; Blanchart E; Martin A; SpainAV; Martin S. 1992. Impact of soilfauna on the properties of soils inthe humid tropics. En: Segoe S, ed.Myths and science of soils of thetropics. Special Publication. Soil SciSoc Am, Madison, Estados Unidos.p 157-185.
Lavelle P; Pashanasi B; Charpentier F;Gilot C; Rossi J-P; Derouard L;André J; Ponge JP; Bernier N. 1998.Large-scale effects of earthwormson soil organic matter and nutrientdynamics. En: Edwards CA, ed.Earthworm ecology. St. LuciesPress, Boca Ratón, Estados Unidos.p 103-122.
Lee KE; Foster RC. 1991. Soil fauna andsoil structure. Aust J Soil Res29:745-775.
Legendre P; Fortin MJ. 1989. Spatialpattern and ecological analysis.Vegetatio 80:107-138.
Legendre P; Troussellier M. 1988. Aquaticheterotrophic bacteria: modeling inthe presence of spatialautocorrelation. Limnol Oceanogr33:1055-1067.
Legendre P; Vaudor A. 1991. The Rpackage: multidimensionalanalysis, spatial analysis.Université de Montréal,Département des SciencesBiologiques, Montreal.
Mantel N. 1967. The detection of diseaseclustering and a generalizedregression approach. Cancer Res27:209-220.

402
El Arado Natural:…
Martin A. 1991. Short- and long-termeffects of the endogeic earthwormMillsonia anomala (Omodeo)(Megascolecidae, Oligochaeta) oftropical savannas, on soil organicmatter. Biol Fertil Soils 11:134-138.
Moran PAP. 1950. Notes on continuousstochastic phenomena. Biometrika37:17-23.
Nuutinen V; Pitkänen J; Kuusela E;Wildbom T; Lohilahti H. 1998.Spatial variation of earthwormcommunity in relation to soilproperties and yield on a grass-clover field. Appl Soil Ecol 8:85-94.
Rangel AF; Thomas RJ; Jiménez JJ;Decaëns T. 1999. Nitrogen dynamicassociated with earthworm casts(Martiodrilus carimaguensis Jiménezand Moreno) in a Colombianlowland Oxisol. Pedobiologia43:557-560.
Rao IM. 1998. Root distribution andproduction in native and introducedpastures in the South Americansavannas. En: Box JE, ed. Rootdemography and their efficiencies insustainable agriculture, grasslandsand forest ecosystems. KluwerAcademic Publishers, Dordrecht,Holanda. p 19-42.
Robertson GP; Freckman D. 1995. Thespatial distribution of nematodetrophic groups across a cultivatedecosystem. Ecology 76:1425-1432.
Rossi J-P. 1996. Statistical tool for soilbiology; XI: Autocorrelogram andMantel test. Eur J Soil Biol32:195-203.
Rossi J-P. 1998. Rôle fonctionnel de ladistribution spatiale des vers deterre dans une savane humide deCôte d’Ivoire. Tesis (Doctorado).Université Pierre et Marie Curie,París. 191 p + anexos.
Rossi J-P; Lavelle P. 1998. Earthwormaggregation in the savanna of Lamto(Côte d’Ivoire). Appl Soil Ecol7:195-199.
Rossi J-P; Delaville L; Quénéhervé P.1996. Microspatial structure of aplant-parasitic nematodecommunity in a sugarcane field inMartinique. Appl Soil Ecol 3:17-26.
Rossi J-P; Lavelle P; Albrecht A. 1997.Relationships between spatialpattern of the endogeic earthwormPolypheretima elongata and soilheterogeneity. Soil Biol Biochem29:485-488.
Rossi J-P; Lavelle P; Tondoh JE. 1995.Statistical tool for soil biology; X:Geostatistical analysis. Eur J SoilBiol 31:173-181.
Smouse PE; Long JC; Sokal RR. 1986.Multiple regression and correlationextensions of the Mantel test ofmatrix correspondence. Syst Zool35:627-632.
Sokal RR; Rohlf FJ. 1995. Biometry: theprinciples and practice of statisticsin biological research. 3a. ed. W.H.Freeman and Company, NuevaYork.
Swift MJ; Heal OW; Anderson JM. 1979.Decomposition in terrestrialecosystems. Blackwell ScientificPublications, Oxford, Reino Unido.
Syers JK; Springett JA. 1984.Earthworms and soil fertility. PlantSoil 76:93-104.
Thioulouse J; Chessel D. 1987. Lesanalyses multitableaux en écologiefactorielle; I: De la théorie d’état à latypologie de fonctionnement parl’analyse triadique. Acta Oecol8:463-480.
Thioulouse J; Chessel D; Dolédec S;Olivier JM. 1997. ADE-4: amultivariate analysis and graphicaldisplay software. Stat Comput7:75-83.
Tondoh JE. 1998. Dynamique despopulations du vers de terreHyperodrilus africanus dans unesavane protégée du feu de Lamto(Côte d’Ivoire). Tesis (Doctorado).Université Pierre et Marie Curie,París.

Estructura Espacio-Temporal de una Comunidad de Lombrices...
403
Yost RS; Trangmar BB; Ndiaye JP;Yoshida NS. 1989 Geostatisticalsoftware for PC-DOS and MS-DOS.Department of Agronomy and SoilScience, University of Hawaii,Honolulu, Estados Unidos.
Yost RS; Uehara G; Fox RL. 1982a.Geostatistical analysis of soilchemical properties of large landareas; I: Semi-variograms. Soil SciSoc Am J 46:1028-1032.
Yost RS; Uehara G; Fox RL. 1982b.Geostatistical analysis of soilchemical properties of large landareas; II: Kriging. Soil Sci Soc Am J46:1033-1037.

PARTE VII
Conclusiones Generales yPrioridades para la
Investigación Futura

Conclusiones Generales...
407
CAPÍTULO 24
Conclusiones Generales, ResultadosImportantes de la Investigación yNecesidades Futuras
J. J. Jiménez*, T. Decaëns**, R. J. Thomas*,L. Mariani*** y P. Lavelle***
Esta obra recopila los conocimientosrecientes sobre losmacroinvertebrados del suelo,haciendo énfasis especialmente enlas lombrices de tierra de lassabanas situadas sobre los suelosácidos de los Llanos Orientales deColombia. La mayor parte de estainvestigación se realizó en el CentroNacional de Investigaciones (CNI)Carimagua. La hipótesis básica queha servido de hilo conductor en estelibro es que las lombrices de tierrason un recurso natural que puedemanejarse para ayudar a lasostenibilidad de aquellosagroecosistemas cuyos recursosbiológicos se consideraríanindicadores potenciales de fertilidady de calidad del suelo.
En Carimagua se hizo uninventario de los recursos biológicos,es decir, de la abundancia y ladiversidad de las especies y de suecología particular, así como unaevaluación del impacto que causandichos recursos en el suelo y en las
plantas. En este libro se describe elimpacto de los ecosistemas, tantonaturales como antrópicos, sobre lascomunidades de la macrofauna delsuelo, y se estudian la biologíabásica, la dinámica de laspoblaciones, las estrategiasadaptativas y el ciclo de vida de laslombrices de tierra; asimismo, sedetallan los efectos que un grupofuncional, el de los ingenieros delecosistema, causa en diferentesaspectos físicos, químicos ybiológicos de estos suelos. Sepresentan, también, opciones paraconservar dichos recursos biológicosy para aprovecharlos en sistemas demanejo que integren diversasprácticas agrícolas. Finalmente, seresumen los resultados másrelevantes de la investigaciónobtenidos hasta la fecha y sediscuten las necesidades queenfrenta esta investigación en elfuturo.
La Fauna del Suelo comoRecurso Natural Valioso
La macrofauna del suelo en lasabana natural de Carimagua secaracteriza por su riquezataxonómica y su alta densidad depoblación. Las termitas (Isoptera)son el grupo más abundante (47% dela biomasa total) y lo siguen las
* Unidad de Suelos y Nutrición de Plantas,CIAT, AA 6713, Cali, Colombia.
* Laboratoire d’Ecologie, UFR Sciences,Université de Rouen, F-76821 Mt SaintAignan Cedex, Francia.
*** Laboratoire d’Ecologie des Sols Tropicaux,IRD, 32 Av. Henri Varagnat, F-93143 BondyCedex, Francia.

408
El Arado Natural:…
lombrices (Oligochaeta) con un 31%del total. Entre los demás gruposrepresentados están las hormigas ylas avispas (Hymenoptera), losescarabajos (Coleoptera), las arañas(Arachnida), los miriápodos(Myriapoda), los nematodos(Nematoda), las cigarras(Homoptera), las chinches(Hemiptera), los isópodos (Isopoda) ylas moscas y mosquitos (Diptera).
Los dos grupos funcionales másimportantes de losmacroinvertebrados del suelo estánrepresentados en la sabana deCarimagua: son los“transformadores de la hojarasca”,p.ej., algunos coleópteros y laslombrices de hábito epígeo, y los“ingenieros del ecosistema”, como lastermitas, las hormigas, las lombricesendógeas y las anécicas. De modosimilar, y unida a la abundanteriqueza específica de la sabana, unagran diversidad funcional semanifiesta en cuatro tipos deestructuras biogénicas superficialesproducidas por 14 especies deingenieros del ecosistema; son lassiguientes: termiteros (3 tipos),placas de termitas (1), hormigueros(8) y heces de lombrices (2) (Decaënset al. 2001). Este descubrimientorefuerza la idea de que lacaracterización de la macrofauna encualquier ecosistema debe iracompañada de un inventario de lasestructuras biogénicas, las cualesson, además, el factor que une labiodiversidad de los organismos conla función que éstos desempeñan enel ecosistema (Lavelle 1997).
Cuando los ecosistemasnaturales son sustituidos poragroecosistemas, la intensidad de loscambios que experimenta lacomunidad de lombrices dependerá,en su mayor parte, de la amplitud delas modificaciones ambientalesinducidas por el uso agrícola delsuelo. Por ejemplo, la riqueza
específica disminuye drásticamentecuando la selva tropical o la sabanase convierten en monocultivosanuales (Dangerfield 1990; Decaënset al. 1994; Fragoso et al. 1997;Lavelle 1988; Lavelle y Pashanasi1989). De otra parte, pocos cambiosse observan cuando elagroecosistema es funcionalmentesimilar al ecosistema original, comoen el caso de un pastizal provenientede una sabana (Decaëns et al. 1994;Jiménez et al. 1998; Schneidmadl yDecaëns 1995) o el de un sistemaagroforestal que deriva del bosquetropical (Fragoso et al. 1997; Gilot-Villenave et al. 1995; Lavelle yPashanasi 1989).
La pérdida de la diversidad de lascomunidades de lombrices causauna alteración en el funcionamientodel ecosistema (Fragoso et al. 1997;Giller et al. 1997). En India, porejemplo, surgen problemas en loscultivos de té tras la desaparición delas lombrices de hábito anécicoporque la hojarasca, que sin ellas nopuede ser introducida rápidamenteen el suelo, se acumulaexcesivamente sobre éste (Senapatiet al. 1994). En América del Sur, laconversión del bosque amazónico enpastizales intensivos conduce a laeliminación de las especies nativasde lombrices en beneficio de otraespecie alóctona; ahora bien, laintensa actividad de ésta, unida a laausencia de la diversidad original,degradan el ecosistema (Barros et al.1998; Blanchart et al. 1999; Chauvelet al. 1999; Duboisset 1995).
La IntensificaciónAgrícola y los Ingenieros
del EcosistemaLas prácticas agrícolas puedenclasificarse, siguiendo un gradientecontinuo, desde las extensivas(utilización del fuego, baja carga

Conclusiones Generales...
409
animal) hasta las intensivas (usonormal de maquinaria agrícola,aplicación de grandes cantidades deinsumos y pastoreo intenso). Laintensificación agrícola y lasprácticas asociadas con ella, como laeliminación de la vegetación nativa,la mecanización del suelo y el uso depesticidas, conducen a ciertasmodificaciones ambientales(especialmente en el microclima y enel recurso trófico) que disminuyen lamayor parte de la biodiversidad delsuelo (Swift et al. 1996).
La biodiversidad de unagroecosistema se reduce no sólo enrelación con la macrofauna del suelosino también con las comunidadesvegetales y animales, encomparación con los ecosistemasnaturales (Giller et al. 1997; Swift yAnderson 1994; Swift et al. 1996).El concepto de intensificaciónagrícola incluye también cambios enla estructura de los agroecosistemas,por ejemplo cuando se pasa de unaagricultura tradicional a una másintensiva (Swift et al. 1996). Laintensificación agrícola se caracterizapor tres acciones (Giller et al. 1997;Swift y Anderson 1994):
• El uso más frecuente de lamisma parcela de suelo, es decir,la intensificación en el uso delrecurso.
• La especialización de las especiesproductivas, tanto animalescomo vegetales, es decir, elcomienzo de la pérdida de labiodiversidad de la cubiertavegetal inicial.
• La utilización de insumos, comofertilizantes y pesticidas, juntocon la mecanización del suelo.
Los diferentes agroecosistemasestudiados en Carimagua puedenclasificarse en función del grado deintensificación agrícola (Swift et al.1996), el cual puede expresarse
mediante un índice (adaptado deGiller et al. 1997) que permitevisualizar de manera precisa dichaintensificación en los LlanosOrientales (Decaëns y Jiménez2002). El orden ascendente deintensificación del gradiente sería elsiguiente (Figura 24-1):
Sabana nativa < Cultivos demarañón < Pastizalestradicionales <
Pastizales introducidos antiguos< Pastizales introducidosrecientes <
Monocultivos < Cultivos anuales
La riqueza taxonómica de lamacrofauna del suelo varía a lo largode este gradiente según dos factores:(a) la intensidad y la frecuencia de laperturbación y (b) la productividaddel ecosistema (cantidad y calidad delos recursos) (Huston 1996). Engeneral, se pueden distinguir dosgrandes agroecosistemas quecontrastan según los efectoscausados en las prácticas agrícolasempleadas: los pastizales y losmonocultivos anuales.
Los pastizales no sólo conservanla riqueza de especies de lombricessino que aumentan sus poblaciones(Jiménez et al. 1998), es decir, tienenuna capacidad de carga mayor. Larazón es la siguiente: los pastizales,que incorporan C al suelo mediantela fotosíntesis y las raíces,suministran además con losdesechos animales, en particular lasdeposiciones del ganado, recursosorgánicos de mayor calidad tantopara las lombrices como para lostransformadores de la hojarasca. Elestiércol del ganado es fragmentadoe incorporado al suelo rápidamente,junto con la hojarasca, por losingenieros del ecosistema, es decir,uno y otra son ingeridos por ellos yse mezclan con la parte mineral delsuelo en el tracto digestivo de laslombrices anécicas, o son mezclados
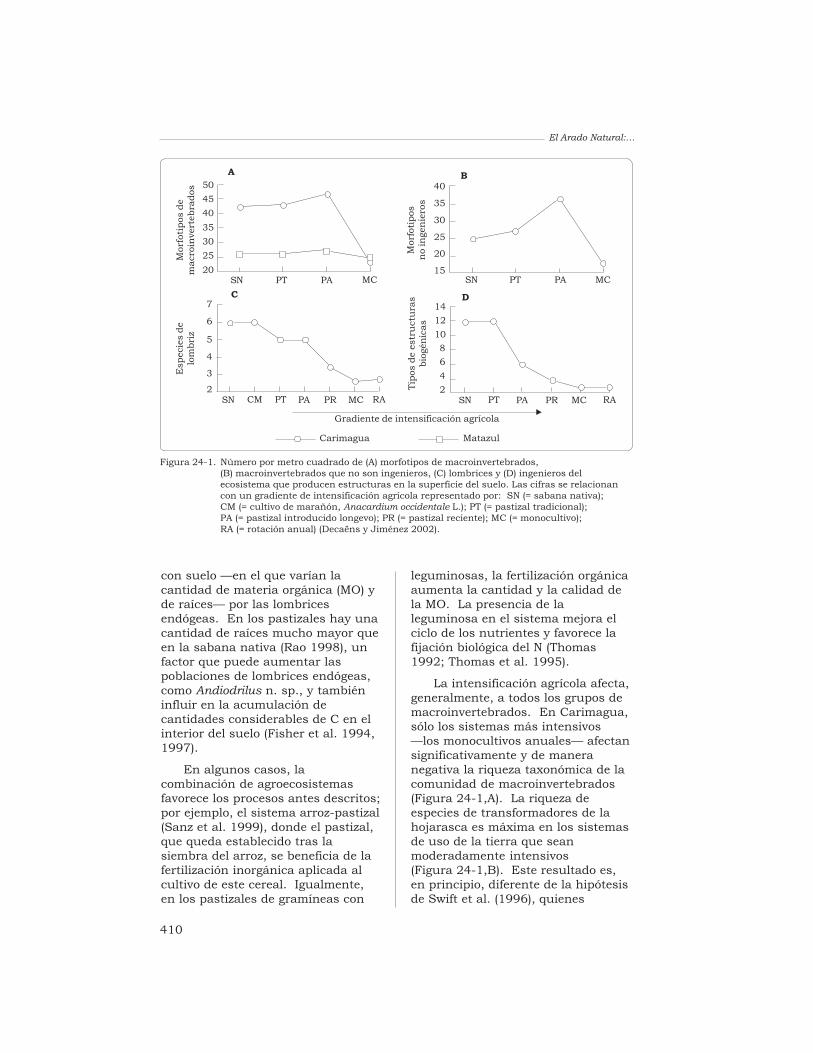
410
El Arado Natural:…
con suelo —en el que varían lacantidad de materia orgánica (MO) yde raíces— por las lombricesendógeas. En los pastizales hay unacantidad de raíces mucho mayor queen la sabana nativa (Rao 1998), unfactor que puede aumentar laspoblaciones de lombrices endógeas,como Andiodrilus n. sp., y tambiéninfluir en la acumulación decantidades considerables de C en elinterior del suelo (Fisher et al. 1994,1997).
En algunos casos, lacombinación de agroecosistemasfavorece los procesos antes descritos;por ejemplo, el sistema arroz-pastizal(Sanz et al. 1999), donde el pastizal,que queda establecido tras lasiembra del arroz, se beneficia de lafertilización inorgánica aplicada alcultivo de este cereal. Igualmente,en los pastizales de gramíneas con
leguminosas, la fertilización orgánicaaumenta la cantidad y la calidad dela MO. La presencia de laleguminosa en el sistema mejora elciclo de los nutrientes y favorece lafijación biológica del N (Thomas1992; Thomas et al. 1995).
La intensificación agrícola afecta,generalmente, a todos los grupos demacroinvertebrados. En Carimagua,sólo los sistemas más intensivos—los monocultivos anuales— afectansignificativamente y de maneranegativa la riqueza taxonómica de lacomunidad de macroinvertebrados(Figura 24-1,A). La riqueza deespecies de transformadores de lahojarasca es máxima en los sistemasde uso de la tierra que seanmoderadamente intensivos(Figura 24-1,B). Este resultado es,en principio, diferente de la hipótesisde Swift et al. (1996), quienes
Figura 24-1. Número por metro cuadrado de (A) morfotipos de macroinvertebrados,(B) macroinvertebrados que no son ingenieros, (C) lombrices y (D) ingenieros delecosistema que producen estructuras en la superficie del suelo. Las cifras se relacionancon un gradiente de intensificación agrícola representado por: SN (= sabana nativa);CM (= cultivo de marañón, Anacardium occidentale L.); PT (= pastizal tradicional);PA (= pastizal introducido longevo); PR (= pastizal reciente); MC (= monocultivo);RA (= rotación anual) (Decaëns y Jiménez 2002).
A50454035302520
Carimagua Matazul
SN PT PA MC
7
6
5
4
3
2
C
SN PT PACM PR MC RA
40
35
30
25
20
15
1412108642
SN PT PA MC
SN PT PA PR MC RA
B
D
Mor
foti
pos
no
inge
nie
ros
Tipo
s de
est
ruct
ura
sbi
ogén
icas
Mor
foti
pos
dem
acro
inve
rteb
rado
sE
spec
ies
delo
mbr
iz
Gradiente de intensificación agrícola
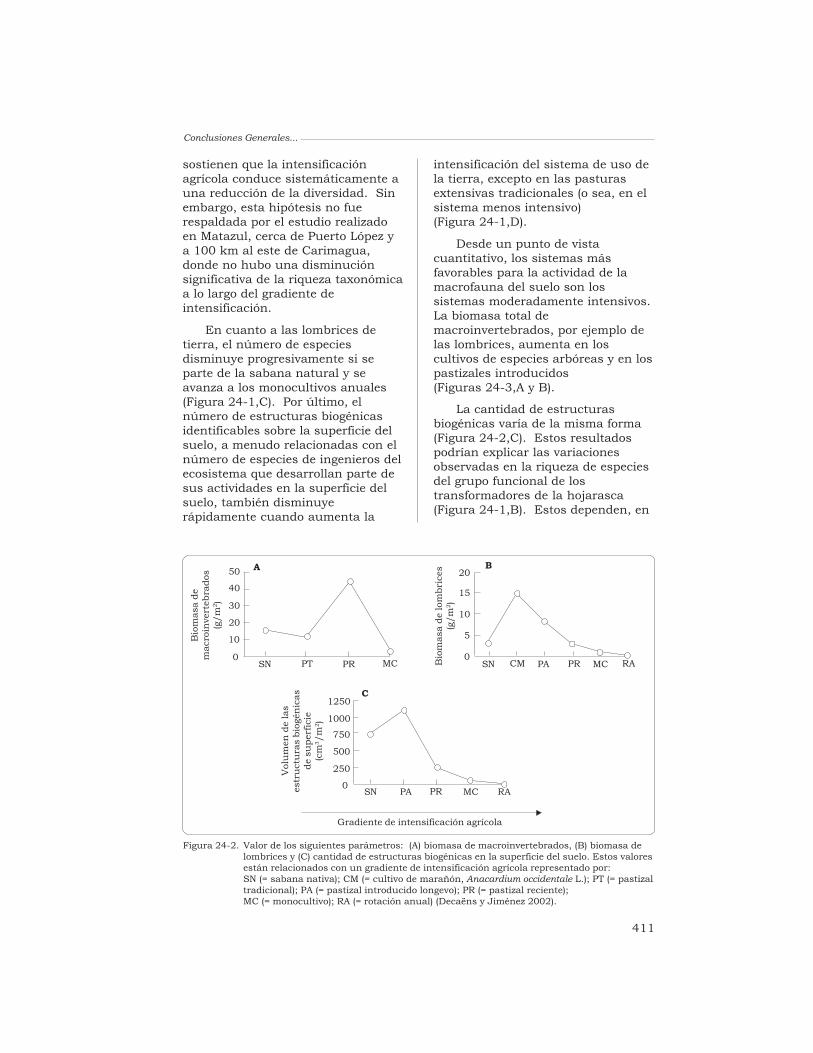
Conclusiones Generales...
411
sostienen que la intensificaciónagrícola conduce sistemáticamente auna reducción de la diversidad. Sinembargo, esta hipótesis no fuerespaldada por el estudio realizadoen Matazul, cerca de Puerto López ya 100 km al este de Carimagua,donde no hubo una disminuciónsignificativa de la riqueza taxonómicaa lo largo del gradiente deintensificación.
En cuanto a las lombrices detierra, el número de especiesdisminuye progresivamente si separte de la sabana natural y seavanza a los monocultivos anuales(Figura 24-1,C). Por último, elnúmero de estructuras biogénicasidentificables sobre la superficie delsuelo, a menudo relacionadas con elnúmero de especies de ingenieros delecosistema que desarrollan parte desus actividades en la superficie delsuelo, también disminuyerápidamente cuando aumenta la
intensificación del sistema de uso dela tierra, excepto en las pasturasextensivas tradicionales (o sea, en elsistema menos intensivo)(Figura 24-1,D).
Desde un punto de vistacuantitativo, los sistemas másfavorables para la actividad de lamacrofauna del suelo son lossistemas moderadamente intensivos.La biomasa total demacroinvertebrados, por ejemplo delas lombrices, aumenta en loscultivos de especies arbóreas y en lospastizales introducidos(Figuras 24-3,A y B).
La cantidad de estructurasbiogénicas varía de la misma forma(Figura 24-2,C). Estos resultadospodrían explicar las variacionesobservadas en la riqueza de especiesdel grupo funcional de lostransformadores de la hojarasca(Figura 24-1,B). Estos dependen, en
Figura 24-2. Valor de los siguientes parámetros: (A) biomasa de macroinvertebrados, (B) biomasa delombrices y (C) cantidad de estructuras biogénicas en la superficie del suelo. Estos valoresestán relacionados con un gradiente de intensificación agrícola representado por:SN (= sabana nativa); CM (= cultivo de marañón, Anacardium occidentale L.); PT (= pastizaltradicional); PA (= pastizal introducido longevo); PR (= pastizal reciente);MC (= monocultivo); RA (= rotación anual) (Decaëns y Jiménez 2002).
Bio
mas
a de
mac
roin
vert
ebra
dos
(g/m
2 )
Bio
mas
a de
lom
bric
es(g
/m2 )
50
40
30
20
10
0
Gradiente de intensificación agrícola
20
15
10
5
0
1250
1000
750
500
250
0
SN CM PA PR MC RASN PT PR MC
A B
SN PA MC RA
C
Vol
um
en d
e la
ses
tru
ctu
ras
biog
énic
asde
su
perf
icie
(cm
3 /m
2 )
PR
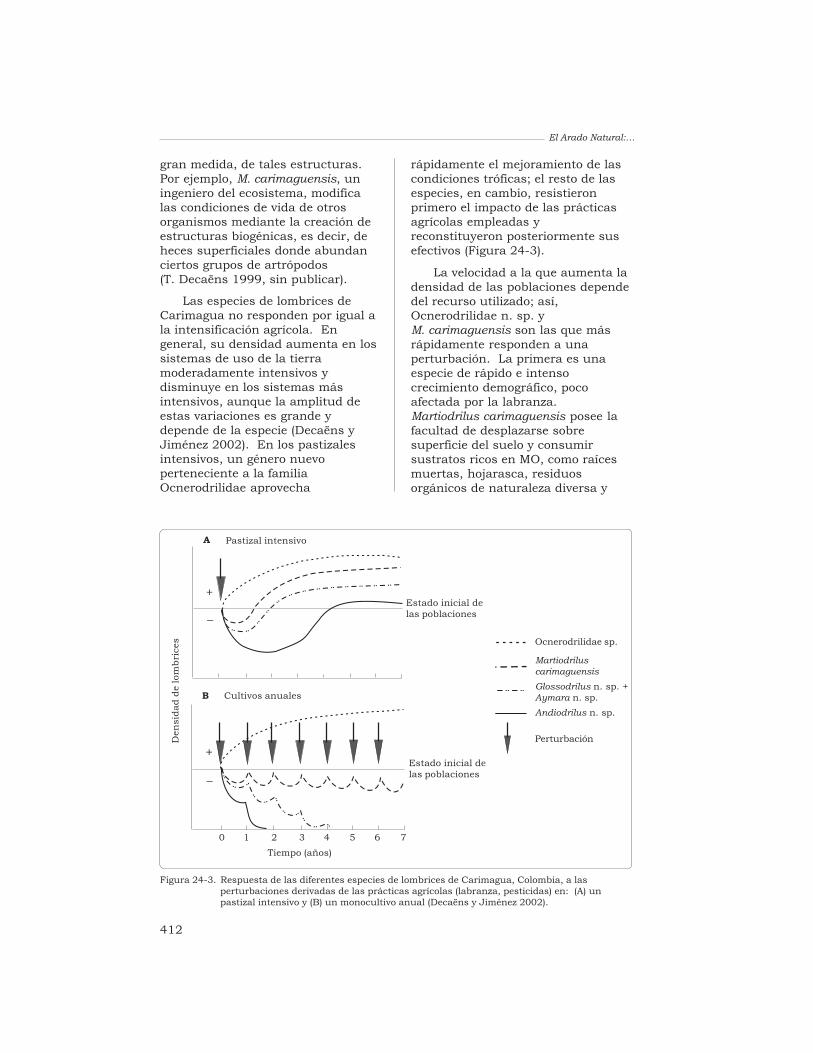
412
El Arado Natural:…
gran medida, de tales estructuras.Por ejemplo, M. carimaguensis, uningeniero del ecosistema, modificalas condiciones de vida de otrosorganismos mediante la creación deestructuras biogénicas, es decir, deheces superficiales donde abundanciertos grupos de artrópodos(T. Decaëns 1999, sin publicar).
Las especies de lombrices deCarimagua no responden por igual ala intensificación agrícola. Engeneral, su densidad aumenta en lossistemas de uso de la tierramoderadamente intensivos ydisminuye en los sistemas másintensivos, aunque la amplitud deestas variaciones es grande ydepende de la especie (Decaëns yJiménez 2002). En los pastizalesintensivos, un género nuevoperteneciente a la familiaOcnerodrilidae aprovecha
rápidamente el mejoramiento de lascondiciones tróficas; el resto de lasespecies, en cambio, resistieronprimero el impacto de las prácticasagrícolas empleadas yreconstituyeron posteriormente susefectivos (Figura 24-3).
La velocidad a la que aumenta ladensidad de las poblaciones dependedel recurso utilizado; así,Ocnerodrilidae n. sp. yM. carimaguensis son las que másrápidamente responden a unaperturbación. La primera es unaespecie de rápido e intensocrecimiento demográfico, pocoafectada por la labranza.Martiodrilus carimaguensis posee lafacultad de desplazarse sobresuperficie del suelo y consumirsustratos ricos en MO, como raícesmuertas, hojarasca, residuosorgánicos de naturaleza diversa y
Figura 24-3. Respuesta de las diferentes especies de lombrices de Carimagua, Colombia, a lasperturbaciones derivadas de las prácticas agrícolas (labranza, pesticidas) en: (A) unpastizal intensivo y (B) un monocultivo anual (Decaëns y Jiménez 2002).
A
B
Pastizal intensivo
Cultivos anuales
Estado inicial delas poblaciones
Estado inicial delas poblaciones
+
_
+
_
0 1 2 3 4 5 6 7
Den
sida
d de
lom
bric
es
Tiempo (años)
Ocnerodrilidae sp.
MartiodriluscarimaguensisGlossodrilus n. sp. +Aymara n. sp.Andiodrilus n. sp.
Perturbación

Conclusiones Generales...
413
heces de otras lombrices (Mariani etal. 2001). En los monocultivosanuales, las especies no tienen eltiempo suficiente para recuperar ladensidad que tenían antes de lasperturbaciones y la mayoría de ellasdesaparece de estos sistemas.
Ciclo de Vida, Estructurade la Comunidad y
PerturbaciónEl estudio de la diversidad y de losprocesos ecológicos asociados con esabiodiversidad y su función en elecosistema constituye la base de lacomprensión y del manejo de losecosistemas naturales y perturbados(Giller 1996). En general, lascomunidades de lombrices sonsensibles a los factores climáticos yedáficos, los cuales determinan ladisponibilidad de los recursosalimenticios y las condicionesmicroclimáticas. Existe, por tanto, elriesgo de que las alteraciones que elhombre causa en los ecosistemasnaturales alteren las comunidades demacroinvertebrados. Ahora bien, lacapacidad de respuesta de dichafauna puede emplearse para manejarsus actividades (Lavelle et al. 1994,1999).
En relación con la biología y laecología de las especies de la sabana,las hipótesis que se han probado sonlas siguientes (Jiménez 1999):
• La sustitución de la sabana nativapor el pastizal conduce a unareducción drástica de la riquezaespecífica y de la diversidad de laslombrices de tierra, que puedeconducir a la eliminación de lasespecies nativas.
• Aunque las especies nativasconsigan adaptarse al nuevosistema de uso de la tierra, laestructura de su comunidadcambiará.
• El manejo de la producciónprimaria en el pastizal y suaumento influyen en laabundancia y en la biomasa delas especies, bien incrementandoo bien reduciendo el número deindividuos.
El cambio que experimenta elecosistema natural, que es la sabananativa, influye en la estructurafuncional de la comunidad delombrices, pero poco o nada afecta lariqueza en especies de éstas. En lospastizales de Carimagua seencuentran las mismas especiesnativas que en la sabana natural yno se observa ninguna invasión deespecies exóticas. Sin embargo, laestructura de la comunidad hacambiado, es decir, la contribuciónrelativa de las diferentes categoríasecológicas y sus efectos en losagroecosistemas fueron alterados.
Dos pastizales introducidosconservan la fauna nativa enCarimagua: (a) Andropogon gayanusy Brachiaria decumbens, solos oasociados con la leguminosa Puerariaphaseoloides; (b) Brachiariahumidicola, sola o asociada conArachis pintoi u otras leguminosas.Estos resultados son excepcionalesporque, según la teoría generalmenteaceptada, las perturbaciones queexperimentan los ecosistemasnaturales traen consigo ladisminución o la desaparición de lasespecies nativas (Lee 1991). Lo máscomún es la aparición de especies dedistribución pantropical con unrango de tolerancia a las propiedadesfísico-químicas del medio muyamplio, como Pontoscolex corethrurusMüller (Cuadro 24-1) y Polypheretimaelongata Perrier (Barois et al. 1988;Barros et al. 1998; Fragoso et al.1999; Lavelle y Pashanasi 1989).
Es probable que el principalfactor que interviene en elmantenimiento de la fauna nativa
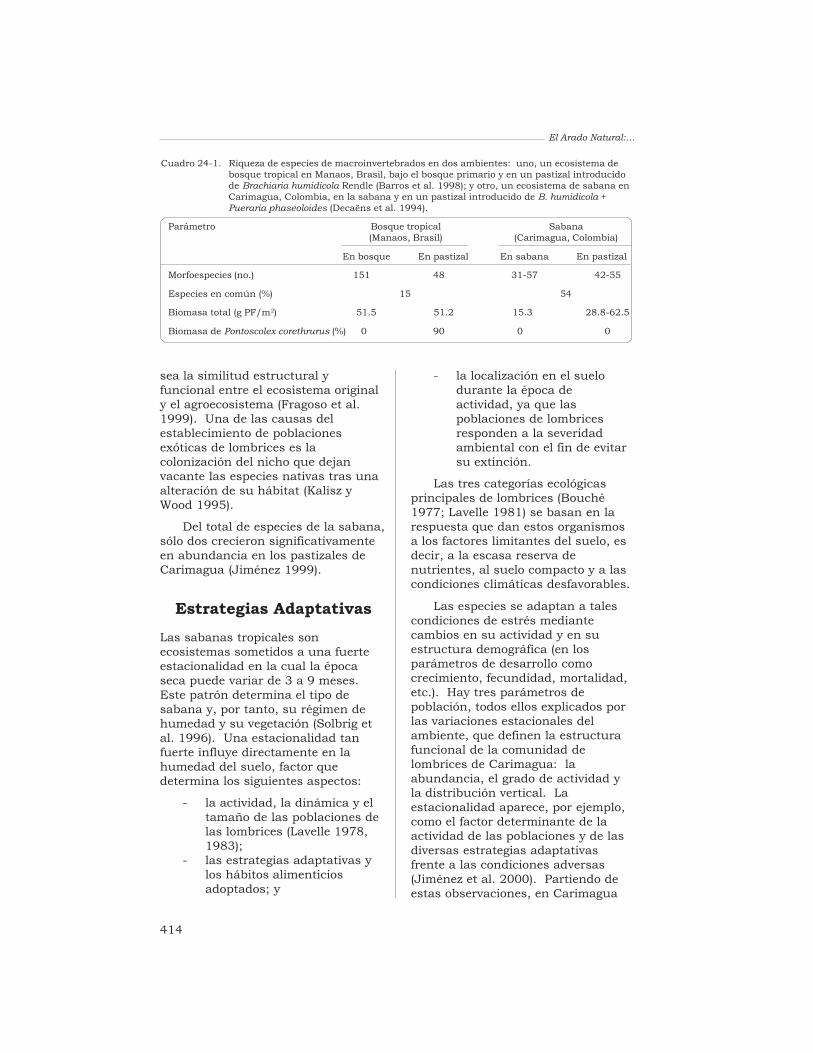
414
El Arado Natural:…
sea la similitud estructural yfuncional entre el ecosistema originaly el agroecosistema (Fragoso et al.1999). Una de las causas delestablecimiento de poblacionesexóticas de lombrices es lacolonización del nicho que dejanvacante las especies nativas tras unaalteración de su hábitat (Kalisz yWood 1995).
Del total de especies de la sabana,sólo dos crecieron significativamenteen abundancia en los pastizales deCarimagua (Jiménez 1999).
Estrategias AdaptativasLas sabanas tropicales sonecosistemas sometidos a una fuerteestacionalidad en la cual la épocaseca puede variar de 3 a 9 meses.Este patrón determina el tipo desabana y, por tanto, su régimen dehumedad y su vegetación (Solbrig etal. 1996). Una estacionalidad tanfuerte influye directamente en lahumedad del suelo, factor quedetermina los siguientes aspectos:
- la actividad, la dinámica y eltamaño de las poblaciones delas lombrices (Lavelle 1978,1983);
- las estrategias adaptativas ylos hábitos alimenticiosadoptados; y
- la localización en el suelodurante la época deactividad, ya que laspoblaciones de lombricesresponden a la severidadambiental con el fin de evitarsu extinción.
Las tres categorías ecológicasprincipales de lombrices (Bouché1977; Lavelle 1981) se basan en larespuesta que dan estos organismosa los factores limitantes del suelo, esdecir, a la escasa reserva denutrientes, al suelo compacto y a lascondiciones climáticas desfavorables.
Las especies se adaptan a talescondiciones de estrés mediantecambios en su actividad y en suestructura demográfica (en losparámetros de desarrollo comocrecimiento, fecundidad, mortalidad,etc.). Hay tres parámetros depoblación, todos ellos explicados porlas variaciones estacionales delambiente, que definen la estructurafuncional de la comunidad delombrices de Carimagua: laabundancia, el grado de actividad yla distribución vertical. Laestacionalidad aparece, por ejemplo,como el factor determinante de laactividad de las poblaciones y de lasdiversas estrategias adaptativasfrente a las condiciones adversas(Jiménez et al. 2000). Partiendo deestas observaciones, en Carimagua
Cuadro 24-1. Riqueza de especies de macroinvertebrados en dos ambientes: uno, un ecosistema debosque tropical en Manaos, Brasil, bajo el bosque primario y en un pastizal introducidode Brachiaria humidicola Rendle (Barros et al. 1998); y otro, un ecosistema de sabana enCarimagua, Colombia, en la sabana y en un pastizal introducido de B. humidicola +Pueraria phaseoloides (Decaëns et al. 1994).
Parámetro Bosque tropical Sabana(Manaos, Brasil) (Carimagua, Colombia)
En bosque En pastizal En sabana En pastizal
Morfoespecies (no.) 151 48 31-57 42-55
Especies en común (%) 15 54
Biomasa total (g PF/m2) 51.5 51.2 15.3 28.8-62.5
Biomasa de Pontoscolex corethrurus (%) 0 90 0 0

Conclusiones Generales...
415
se distinguen tres grupos deespecies, según la respuesta que denal estrés ambiental:
1. Especies de tamaño pequeño,localizadas superficialmente en elsuelo (en la época activa), condistribución vertical profundadurante la fase de quiescencia.Por ejemplo, Ocnerodrilidae n.sp., así como Aymara n. sp.(epígea) y Glossodrilus n. sp.(endógea), aunque la distribuciónvertical de las dos últimas no esmuy profunda.
2. Especies de tamaño intermedio,de localización superficialdurante la época lluviosa, y sinadaptación especial a la época desequía; por ejemplo, Andiodrilusn. sp. (endógea).
3. Especies de tamaño grande, condiapausa verdadera en las capasprofundas del suelo; por ejemplo,Andiorrhinus n. sp. (endo-anécica, aunque sin diapausa) yM. carimaguensis (anécica).
Ingenieros del Ecosistemadel Suelo
Los Capítulos del 11 al 23 estándedicados al estudio de los efectosproducidos por los ingenieros delecosistema en algunos procesosfísicos y químicos del suelo, y giranen torno a tres hipótesis (Decaëns1999):
• Los ingenieros del ecosistemaproducen estructuras biogénicasque son características de suespecie.
• Los ingenieros del ecosistemacausan, mediante las estructurasbiogénicas producidas, unimpacto significativo en losprocesos del suelo y en losorganismos que viven en él.
• La intensificación agrícola tieneefectos contrastantes en lascomunidades de ingenierosecológicos del suelo y altera, portanto, el impacto que éstosharían, ya que la composición dela comunidad de ingenieros y sufunción pueden variar.
La comprobación empírica yexperimental de estas hipótesis hapermitido establecer y describir elpapel funcional de los ingenieros delecosistema en los suelos de lassabanas colombianas, así comoevaluar el impacto que tienen lasprácticas agrícolas en laspoblaciones de tales ingenieros.
Martiodrilus carimaguensiscomo ingeniero del ecosistemaLos ingenieros ecológicos sonorganismos que tienen la facultad demodular la disponibilidad de losrecursos que serán usados por otrosorganismos vivos mediante lasestructuras biogénicas que producen(Jones et al. 1994, 1997; Lawton yJones 1995). Numerososmacroinvertebrados responden aesta definición (Anderson 1995;Jones et al. 1994; Lavelle 1997),incluyendo a M. carimaguensis. Estaespecie, en efecto, modifica suambiente haciendo pasar losmateriales (hojarasca, humus ysuelo) de un estado físico a otro, esdecir, abriendo macroporos en elsuelo, incorporando hojarasca alsuelo y formando agregados estables.Las estructuras producidas—galerías y turrículos— constituyenun recurso (microhábitat y recursotrófico, respectivamente) que losmacroinvertebrados y las raícesaprovechan directamente. Demanera relativamente indirecta,M. carimaguensis transforma elsuelo, llevándolo de un estado físicoinicial a uno bioturbado a través deuna serie de modificaciones en sus

416
El Arado Natural:…
propiedades físicas, en la dinámicade la MO y en la disponibilidad de losnutrientes, todo lo cual tiene unefecto positivo en el crecimiento delas plantas (Decaëns et al. 1999a,1999b).
En resumen, M. carimaguensisdesarrolla las siguientes acciones pormedio de las estructuras queproduce (Decaëns 1999):
• Suministra un recurso trófico, yaque Andiodrilus n. sp. yOcnerodrilidae n. sp. ingierenesas estructuras, y uno espacial,porque proporciona un espacio aciertos macroinvertebrados en elinterior de las galerías o bajo losturrículos.
• Modifica indirectamente ladisponibilidad y la calidad delrecurso trófico y del recursoespacial; es decir, al modificar laestructura del suelo y ladinámica de su MO, favorece ladisponibilidad de los nutrientesque requieren las plantas.
• Facilita el transporte de algunosorganismos, aproximándolos alos recursos.
Los atributos funcionales de unaespecie o de un grupo funcional seconsideran a diferentes niveles deorganización dentro de los sistemasjerárquicamente conformados. Encuanto aumenta la escala deobservación, de lo más próximo a lomás alejado, los niveles describen losmismos procesos (Allen y Starr1982). Una primera aproximaciónascendente (“bottom-up”, sensuAnderson 1995) ha mostrado que lasestructuras biogénicas producidaspor M. carimaguensis influyen en laestructura del suelo, en la dinámicade la MO del suelo, en la diversidadde la macrofauna y en el crecimientoradicular de las plantas. En una
aproximación descendente(“top-down”, sensu Anderson 1995),el impacto de M. carimaguensis en elecosistema sigue siendo apreciable,aunque está marcadoprogresivamente por los efectosproducidos por otras comunidadesde organismos y por los factoresambientales que regulan los procesosdel suelo dentro de la escalajerárquica (Decaëns et al. 1999a).
Tipología de las estructurasbiogénicasLos ingenieros del suelo delecosistema en Carimagua producenestructuras biogénicas decaracterísticas físicoquímicasparticulares de cada especie y,generalmente, diferentes del sueloque las circunda. Esto abre laposibilidad, mas allá de los criteriosclásicos de clasificación ecológica delas lombrices de tierra, de estableceruna clasificación nueva de losingenieros del ecosistema basada enlos atributos funcionales de susestructuras biogénicas y sinconsiderar necesariamente losaspectos taxonómicos (Decaëns et al.2001); por ejemplo, la ingeniería quehace M. carimaguensis en elecosistema disminuye lacompactación del suelo —un efectoque equilibra, en cierta forma, lacompactación proveniente de lasheces grandes y compactas queproduce la lombriz— y nos permitiríallamarla una especiedescompactadora. Hay queconsiderar también el efecto de laespecie perteneciente a la familiaOcnerodrilidae, pues su contribucióna la estabilidad estructural del suelopuede ser importante, aun cuandolos efectos que tenga en elcrecimiento de las plantas seanpequeños.

Conclusiones Generales...
417
Guía para el Manejo de laFauna Benéfica del Suelo
Tanto la investigación presentada enesta obra como las referenciasbibliográficas consultadas nos llevana ser cautos respecto a laformulación de una guía generalpara el manejo de la macrofauna delsuelo. Algunas prácticas, como lospastizales mejorados, puedengenerar, sin duda, un aumentosustancial de las poblaciones demacrofauna. Ahora bien, esteaumento puede ser benéfico, comoen el caso de M. carimaguensis en lassabanas colombianas, o perjudicial,como el efecto compactador deP. corethrurus en los pastizales de laAmazonia que, aunado a la pérdidade la fauna nativa del suelo, llevaríaa la degradación de los pastizales(Chauvel et al. 1999). La similitudentre el ecosistema original y elagroecosistema que de él provienesería el principal determinante de lasupervivencia, de la adaptación, dela capacidad de resistencia y de laestabilidad de las especies nativas delombrices, dentro de los límites delmanejo de los ecosistemas.
Antes de pensar en estableceruna de las guías indicadas, esnecesario conocer la biología y laecología básicas de la macrofauna decualquier ecosistema. Este procesoexige un activo trabajo deinvestigación, lo que explica laescasa presencia de estudiossimilares en la zona tropical. Elconocimiento mencionado debeenfocarse a los principales ingenierosdel ecosistema que se hanencontrado, puesto que los efectoscausados por estas especies tiendena dominar los que provienen de lasdemás poblaciones de la macrofauna(Lavelle 1997).
El caso de M. carimaguensis enlos Llanos Orientales de Colombia es,hasta ahora, una excepción, ya que
los efectos de esta especie en lospastizales mejorados tienden a serbeneficiosos cuando aumentan suspoblaciones (Parte III y Parte VI deesta obra). Estas lombrices son unejemplo excepcional de que lasactividades de la macrofauna nativacontribuyen a la conservación, a laproductividad y a la sostenibilidad deun agroecosistema, y logran mejorarestos parámetros.
La disposición espacial de unpastizal junto a un cultivo puedeacelerar la recuperación de laspoblaciones de macrofauna de esteúltimo. Esta práctica de manejonecesita ser investigada aún enprofundidad, especialmente si laespecie benéfica, que puedeestablecerse más rápidamente,ayuda a revertir los problemas dedegradación de la estructura delsuelo debidos a los cultivos, y haceasí innecesaria la solución de dichosproblemas mediante el empleo deestrategias intensivas y el uso demaquinaria costosa. Las lombricesde tierra se convierten entonces enun recurso que se podría utilizarpara mejorar la calidad delecosistema.
La disposición espacial deparcelas que conservan laspoblaciones de lombrices junto aaquéllas que tienden a reducirlas esun ejemplo de manejo indirecto(Figura 24-4). Lavelle et al. (1999)suministran mayor informaciónsobre este tema y lo discuten endetalle.
Tipología de una comunidadde macrofauna benéficaAunque es grande el número deespecies que pueden considerarseingenieros del ecosistema porquecausan cambios físicos en elambiente, no todos esos cambiostendrán consecuencias ecológicassignificativas (Jones et al. 1997). Sin

418
El Arado Natural:…
embargo, algunos ingenieros puedenmultiplicar estos efectos mediante laacción desarrollada por su dominiofuncional. Estos efectos dependende una serie de factores, entre losque se encuentran la densidadpoblacional, la distribución espaciallocal y regional, y el tipo deestructuras físicas producidas (Joneset al. 1997). Por tanto, es precisoidentificar las especies presentes enel suelo y las estructuras que ellasproducen, y describir con precisiónlos mecanismos mediante los cualeslos ingenieros del ecosistemainfluyen en el ambiente del suelo.Las interacciones tróficas eindirectas (interacciones deingeniería, sensu Jones et al. 1994)pueden incluirse en modelospredictivos del impacto quecausarían los ingenieros delecosistema. Estos modelos seránparticularmente útiles cuando seintente mejorar la sostenibilidad de
los agroecosistemas mediante unaumento de las actividades o de ladiversidad de la macrofauna delsuelo (o de ambos factores) (Giller etal. 1997). Los modelos seemplearían para probar las hipótesissobre la función de la biodiversidaden los agroecosistemas (Swift yAnderson 1994; Swift et al. 1996).
El estudio aquí presentadopermite sugerir que el manejo de ladiversidad de las lombrices de tierraen los agroecosistemas debe iniciarseestableciendo una comunidad quecontenga una o más especies epígeasy una o más especies anécicas —queconstruyan galerías verticales,produzcan estructuras biogénicas enla superficie y mezclen residuosvegetales con el sustrato mineral— yuna o más especies endógeas —quese alimenten de MO y de exudadosde raíces y horaden galeríashorizontales. La comunidad de
Figura 24-4. Alternativas de manejo en las tecnologías con lombrices (Senapati et al. 1999). Losrectángulos indican actividades humanas.
Vermicompost
Inoculación sola1. Potes2. Parcelas
Inoculación +manejo de
residuos orgánicos
Manejoorgánico
Rotación decultivos
Yurimaguas(Perú)
Tamil Nadu(India)
Carimagua(Colombia)
Yierpadi(India)
Producción masiva
Yurimaguas (Perú)Yierpadi (India)
Manejo directo(inoculación)
Manejo indirecto
En el suelo
Tecnologías con lombrices
Fuera del suelo

Conclusiones Generales...
419
lombrices de Carimagua respondeexactamente a este modelo, y haceposible la propuesta de un manejodeterminado de las comunidades delombrices en los agroecosistemasestudiados.
Las opciones de manejo de laactividad de las lombrices tendríanen cuenta, principalmente, lossiguientes aspectos relevantes:
• La comunidad de ingenieros delecosistema del suelo en elagroecosistema debería ser lamisma que la del ecosistemaoriginal, que es el natural.
• Los organismos ingenieros delsuelo y, en particular,M. carimaguensis, influyen endiferentes procesos relacionadoscon la fertilidad del suelo.
• Las actividades de los ingenierosdel suelo se han desarrolladobastante en los pastizales, perose reducen mucho en los cultivosanuales.
• Los efectos de los ingenieros delecosistema permanecen durantealgún tiempo porque lasestructuras biogénicasproducidas persisten en elentorno, aun después de ladesaparición del organismo quelas produjo.
La especie perteneciente a lafamilia Ocnerodrilidae yM. carimaguensis son las únicasespecies de lombriz que soportan laintensificación continua de losmonocultivos. Ocnerodrilidae n. sp.es una especie endógea polihúmicaque tolera la labranza porque susindividuos son de tamaño muypequeño. Los individuos deM. carimaguensis, aparte de la buenaestrategia con que, en la épocadesfavorable, ocupan los estratosprofundos del suelo (Jiménez yDecaëns 2000), migran también por
la superficie del suelo. Lacolonización que haríaM. carimaguensis de una parcelapobre en lombrices podría facilitarsesi se distribuyen adecuadamente lasparcelas. Además, las rotaciones enque un monocultivo sigue a unpastizal de varios años de serviciopueden ser favorables para eldesarrollo de poblacionesimportantes de lombrices.
Un sistema de producciónagrícola que preserve las lombricesdesarrollaría las siguientes prácticas:
• Mantener los pastizales nativos(la sabana) como reservorios debiodiversidad.
• Establecer cultivos forrajeroscomo bancos de proteína(Rippstein et al. 1996) quepermitan el desarrollo de unabiomasa de lombricesimportante.
• Sembrar cultivos anuales queroten con pastizales de variosaños (bancos de proteína) o concultivos forrajeros, de maneraque los fenómenos de migraciónse maximicen. En este sistema,las estructuras biogénicas queaparezcan serían indicadorescualitativos y cuantitativos delestado de las poblaciones de losingenieros del ecosistema.
Un desafío importante para elfuturo cercano es la identificación delos agroecosistemas sostenibles quemantienen niveles aceptables dediversidad y de biomasa de lamacrofauna del suelo, para optimizarlas actividades de ésta. Lainvestigación presentada en estaobra es un ejemplo de la decisión deabrir la “caja negra” de la biología delsuelo y del consiguiente desarrollo deuna guía de manejo de la faunaencontrada. Ahora bien, para quelos productores acepten y adoptenesta guía, los estudios mencionados

420
El Arado Natural:…
deben combinarse con un enfoquemultidisciplinario que incluya losconocimientos locales, laexperimentación en la finca delproductor y algunos estudiossocioeconómicos. De este modo, elsuelo se vería como un lugar másdiverso, productivo y saludable,gracias a un proceso que aumente elnúmero de opciones viables para elproductor.
Aceptación por losProductores
Cualquier práctica nueva de manejode los organismos del suelo debe seraceptada por el agricultor y requiere,a menudo, que éste la desarrolle y laadapte para que pueda incorporarlacon éxito a su sistema. Elconocimiento actual de la percepciónque tienen los agricultores de losorganismos del suelo es todavía muylimitado. Algunos estudios recientessobre esta visión del agricultor hanrevelado, por ejemplo, una granvariación tanto en los conocimientoslocales sobre las lombrices como enel uso que pueden hacer de ellas ensus sistemas agrícolas (Ortiz et al.1999). Los agricultores de Méxicoque fueron consultados estabanrelativamente bien informados sobrela importancia y los beneficios de laslombrices de tierra en la fertilidad delsuelo; los agricultores del Congo, encambio, tenían muy escasainformación al respecto. Sepercibieron también diferencias entrehombres y mujeres sobre este temaen el estudio de México (Ortiz et al.1999). Los conocimientos que tienenlos agricultores de una localidadsobre los organismos microscópicosson incluso menores que losrelacionados con los organismos másgrandes y visibles; los síntomas deenfermedades en las plantas serían,quizás, el único indicador obvio paraellos de la presencia de organismos
perjudiciales. No obstante, resultaestimulante anotar que losagricultores de México podíandistinguir entre tres o cuatro tipos delombrices según su tamaño, su colory el lugar donde se encontraban.Ortiz et al. (1999) publicaron elprimer informe, probablemente,sobre la percepción que tienen losagricultores de la macrofauna delsuelo.
Los agricultores y los científicosnecesitan desarrollar conjuntamenteun lenguaje común sobre la faunadel suelo, ya que así se aceptaríanmejor los conceptos sobre losorganismos del suelo y sobre lasprácticas de manejo de éstos, y seaumentaría, por ello,significativamente las poblaciones deestos organismos en los sistemas deproducción. Es necesario, también,desarrollar una guía del usuario,sencilla y amigable, para manejar lasactividades de los organismosbenéficos. Los agricultores necesitandistinguir, por ejemplo, entre lasespecies de lombrices que compactanel suelo y las que no lo compactan.Necesitan también métodos demuestreo simples y rápidos paraobtener, al menos, un cálculosemicuantitativo de la abundanciade la macrofauna. Los científicos,por su parte, necesitan establecer overificar los umbrales de abundanciade organismos o de poblaciones,desde los que se obtengan impactossignificativos en los procesos delsuelo y en la producción vegetal.
Estas herramientas y guías paratomar decisiones deben desarrollarseconjuntamente entre los agricultoresy los científicos, incorporando elconocimiento local existente y,cuando éste no exista,suministrando material educativo ydidáctico para los que puedan leer ypara los que no sepan hacerlo.Existen ejemplos de dicho material:uno de ellos es la serie de guías de

Conclusiones Generales...
421
manejo de los recursos naturales queha sido publicada para usarla en elmanejo de las cuencas (CIAT 1999).Estas guías indicarán cómoidentificar y desarrollar un enfoquesobre el manejo de los recursosnaturales y cómo desarrollar unmaterial atractivo para losagricultores y otros usuarios, comolos extensionistas y los integrantes deorganizaciones no gubernamentales(ONG). Actualmente se estánelaborando módulos de capacitaciónsobre biología del suelo.
Resultados Importantesde la Investigación
Capítulo 2Impacto del uso de la tierraen la macrofauna del suelode los Llanos Orientales deColombia1. Las comunidades de
macroinvertebradosresponden claramente a lasalteraciones derivadas delsistema de uso del suelo. Elpastoreo y la quemafavorecen la actividad de laslombrices, aunque suimportancia decrece cuandohay sobrepastoreo.
2. La asociación deB. decumbens y P.phaseoloides durante untiempo largo (15 años)conserva la calidad y labiodiversidad del suelo ymantiene también la riquezataxonómica de la sabana.
3. Los cultivos anuales tienenun impacto negativodramático en las poblacionesde lombrices y artrópodosporque reducen su biomasa,la densidad de suspoblaciones y su riquezataxonómica.
Capítulo 3Cuantificación de lamacrofauna del suelo enuna cuenca hidrográfica deColombia1. La diversidad, la abundancia
y la estructura funcional delas comunidades de losmacroinvertebrados del suelovariaban según el sistema deuso de la tierra; el bosquesecundario fue el ecosistemade mayor diversidad.
2. Varios organismos puedenser utilizados comoindicadores de calidad delsuelo en los ambientes deladera alterados de los Andescolombianos.
Capítulo 4Las comunidades delombrices en las sabanasnativas y en los pastizalesintroducidos de los LlanosOrientales de Colombia1. La riqueza específica de la
sabana nativa se mantuvo enlos pastizales, ya sean degramínea sola o asociada conleguminosas, y ningunaespecie exótica de lombrizcolonizó estosagroecosistemas.
2. La comunidad de lombricesde Carimagua estácompuesta por especiespertenecientes a lasprincipales categoríasecológicas. Estas especiesson de diverso tamaño yemplean estrategiasadaptativas diferentes que lespermiten sobrevivir a severascondiciones ambientales.
3. La conversión de la sabanaen pastizal produjodiferencias significativas entérminos de densidad y de

422
El Arado Natural:…
biomasa de las lombrices(10 veces mayores en elpastizal introducido), que sedebieron, principalmente, alespectacular aumento de laspoblaciones de la especieanécica M. carimaguensis.
4. En la sabana nativa, laslombrices endógeascontribuyeron con más del80% a la biomasa total delombrices, mientras que en elpastizal, la especie M.carimaguensis aportó el 88%de esa biomasa y lasendógeas sólo el 12%.
Capítulo 5Dinámica de poblaciones yestrategias adaptativas deMartiodrilus carimaguensis(Oligochaeta,Glossoscolecidae), unaespecie nativa de lassabanas bien drenadas deColombia1. La producción de heces
frescas o turrículos en lasuperficie del suelo fue 11veces superior en el pastizalintroducido que en la sabananativa, lo que indica que haydiferencias entre ambossistemas respecto a laabundancia de M.carimaguensis.
2. Martiodrilus carimaguensispresentó una verdaderadiapausa, que teníadiferencias si se trataba deun adulto o de un individuoinmaduro. Estos últimos sonactivos solamente 4 meses alaño, hasta la mitad delperíodo de lluvias. Estaslombrices construyencámaras de estivación al finalde sus galerías, y en ellashacen varios septos con lasheces.
3. La capacidad de entrar endiapausa antes de la llegadade la época seca y laconstrucción de galeríassemipermanentes son dosfactores que explican por quéla sequía afecta tan poco alos individuos de esta especiey por qué pueden mantenersu densidad poblacional.
4. Las diferencias en la calidadde la MO y el aporte que haceel estiércol del ganado, quees rápidamente incorporadoal suelo por especies dehábito anécico (M.carimaguensis y algunoscoleópteros) pueden ser losfactores que explican elaumento de la actividad deestas lombrices en lospastizales.
Capítulo 6Estrategias reproductorasde tres especies delombrices nativas de lassabanas de Carimagua(Colombia)1. La fuerte estacionalidad del
sitio restringe la deposiciónde los capullos a sólo8 meses del año. Seobservan dos estrategiasclaras: o bien deposicióncontinua, como enAndiodrilus n. sp., odeposición y diapausa, comoen M. carimaguensis; o bienmuerte de los adultos, comoen Glossodrilus n. sp.
2. Cuanto mayor es el tamañodel adulto, mayor es el delcapullo; M. carimaguensisinvierte el 16% del peso deladulto en la formación delcapullo, el porcentaje másalto registrado hasta la fecha.

Conclusiones Generales...
423
Capítulo 7Distribución vertical de laslombrices de tierra en lossuelos de los “Llanos” deColombia cubiertos porespecies forrajeras1. La sustitución de la sabana
por el pastizal no produceningún efecto en ladistribución vertical de laslombrices en el suelo.
2. Las especies más grandes nose encuentran a muchaprofundidad, ya que no huborelación entre el tamaño delas lombrices y laprofundidad a la que fueronencontradas. Sin embargo,una especie endógeapolihúmica muy pequeña,que se encuentrahabitualmente en las hecesde M. carimaguensis en elinterior del suelo, suelehallarse a una profundidadrelativa.
3. El principal factor en ladistribución vertical de laslombrices es la humedad delsuelo, aunque las diferenciasencontradas en este factorentre los adultos y losindividuos inmaduros de M.carimaguensis pueden sertambién de origen biótico. Elinicio temprano de ladiapausa de los individuosinmaduros estáfisiológicamente inducido; laseñal que desencadena elproceso necesita másinvestigación.
4. Durante la diapausa, tantolos adultos como lossubadultos de M.carimaguensis (los individuosmaduros) se encontraron amayor profundidad que losindividuos inmaduros.
Capítulo 8Distribución espacial de laslombrices en las sabanas desuelos ácidos de los LlanosOrientales de Colombia1. Las lombrices de la sabana y
de los pastizales, quepertenecían a categoríasecológicas diferentes y eran detamaño diverso, presentaronuna distribución espacialfuertemente agregada enparches de varias decenas demetros, en cualquiera de losdos sistemas de uso de latierra ensayados.
2. Existe una relación entre losíndices de agregación y eltamaño de las especies (larelación longitud/diámetropreclitelar, L/D). El análisisfactorial discrimina lasespecies grandes que viven acierta profundidad de las quese encuentran cerca de lasuperficie del suelo, y tambiénlas especies de agregaciónespacial fuerte pero conrelación L/D baja de lasespecies con valores altos deL/D, como en el caso deGlossodrilus sp., la cual tieneademás un índice deagregación cercano a 1. Elpatrón de distribuciónespacial de las lombrices seexplica por variablesbiológicas y no por el sistemade uso de la tierra.
Capítulo 9Diferencias en laprogramación de ladiapausa y en los patronesde estivación de algunasespecies de lombriz de tierradel trópico1. La actividad de las lombrices
estuvo determinada por la

424
El Arado Natural:…
fuerte estacionalidad delecosistema de las sabanas ycasi todas las poblaciones delombrices descendían a losestratos profundos del suelodurante la época seca.
2. Las especies que afrontan laépoca seca muestrandiferentes estrategiasadaptativas, por ejemplo ladiapausa facultativa uobligatoria, para evitar quesus tegumentos se sequen enel interior de las cámaras deestivación localizadas a ciertaprofundidad del suelo.
3. Los individuos inmaduros deM. carimaguensis comienzanla diapausa en la mitad de laépoca lluviosa; la fuerteestacionalidad de las sabanassería tal vez el factor queregula la densidadpoblacional de esta especie.
Capítulo 10La dieta de una lombrizanécica de las sabanascolombianas: Una cuestiónrelacionada con lascategorías ecológicas1. Se observaron diferencias
significativas mínimas alcomparar el contenido de lamolleja de los individuosprovenientes ya sea de lasabana o del pastizalintroducido. Los de lasabana tenían en la mollejauna cantidad relativamentegrande de complejos órgano-minerales. Se encontrótambién una gran cantidadde fragmentos vegetales< 500 µm (44.2% V/V) en losagregados órgano-minerales,de los cuales las raíces eranel componente principal.
2. El contenido de las mollejasanalizadas era el siguiente:
un 33% de fracción liviana deMO, un 6.6% de MO amorfay un 53% de partículasminerales.
3. Los individuos recolectadosen las dos parcelasensayadas compartían unrégimen alimenticio basadoen la necrofagia y en lacoprofagia, y a vecesadoptaban una dietatípicamente anécica, es decir,ingerían una mezcla dehojarasca y suelo.
4. Los estudios que seemprendan más tardedeberán considerar lahipótesis de la ingestiónselectiva de una masaórgano-mineral en lasuperficie del suelo, es decir,un comportamiento anécicotípico aunque con unafunción ecológica modificada.Este comportamiento sería laconsecuencia de unaadaptación de esta especie ala ausencia de hojarasca enla sabana que es sometida ala quema periódica.
Capítulo 11Propiedades de lasestructuras construidas porlos ingenieros delecosistema en la superficiedel suelo de una sabanacolombiana1. En las sabanas ácidas de
Carimagua, 14 especiesdiferentes demacroinvertebradosproducen estructurasbiogénicas sobre la superficiedel suelo, que son fácilmentedistinguibles entre sí:hormigueros (8 tipos),termiteros (3), placas determitas (1) y heces delombrices (2).

Conclusiones Generales...
425
2. Los turrículos producidos porAndiodrilus sp. (endógea) yM. carimaguensis (anécica)están formados por agregadosde gran tamaño que, en elcaso de esta última especie,son más estables que losagregados del suelo de tamañocomparable.
3. En conjunto, se hanidentificado tres tipos deestructuras biogénicas:a. compactas y ricas en MO
(turrículos);b. ligeramente compactas y
ricas en MO (termiteros); yc. granulares, no
compactadas y con bajoscontenidos de MO(hormigueros y placas determitas)
4. A partir de la tipología de lasestructuras biogénicas, sepuede establecer unaclasificación funcional de lafauna del suelo y valorar susefectos en el funcionamientodel ecosistema.
Capítulo 12Dinámica de la degradaciónde las heces superficiales dela lombriz de tierra en áreasde sabana y de pastizal enlos Llanos Orientales deColombia1. La formación de cámaras y
galerías en el interior de losturrículos se debió a lacolonización de éstos poralgunos macroorganismos(macroinvertebrados y raíces)y principalmente por laactividad de las termitaspertenecientes al géneroNasutitermes.
2. La persistencia de losturrículos deM. carimaguensis sobre lasuperficie del suelo parece
estar determinada por dosfactores antagónicos:a. la densidad de la
vegetación y la cantidadde hojarasca presenteque protege los turrículosdel impacto de la lluvia, y
b. el pisoteo del ganado queacelera su destrucción.
3. El papel que desempeñan losmacroinvertebradospequeños en lafragmentación de lasestructuras biogénicasproducidas por organismosde tamaño mayor apoya lahipótesis de que laestabilidad de losecosistemas depende de unadiversidad funcional alta.
4. Los individuos deM. carimaguensis soningenieros del ecosistema, yaque los turrículos queproducen son micrositiosdonde se regulan importantesprocesos físicos y químicosdel suelo. La producción degrandes cantidades deturrículos produce efectossignificativos en la estructuradel suelo, en sus propiedadesfísicas y en la dinámica de laMO en el perfil del suelo.
Capítulo 13Dinámica del nitrógeno enlas heces de las lombricesde tierra de la especieMartiodrilus carimaguensisJiménez y Moreno, en unOxisol de las sabanas deColombia1. La introducción de un
pastizal en un área desabana conduce al aumentode las reservas de MO, delcual dependería elincremento de la densidad delas lombrices de tierra.

426
El Arado Natural:…
2. Las lombrices de lospastizales, cuya densidad esalta, ingieren variastoneladas de suelo por año, yson por ello responsables delas grandes cantidades de Nmineralizado.
3. El manejo de lascomunidades de lombrices enlos pastizales introducidospuede convertirse en unmecanismo que aumente enel suelo la MO y el ciclo denutrientes, especialmente delN, reduciendo así el riesgo dedegradación del pastizal.
Capítulo 14Disponibilidad de fósforoen las heces de una especieanécica de lombriz de tierrade las sabanas de sueloOxisol en Colombia1. Martiodrilus carimaguensis
juega un papel importante enmejorar y en mantener ladisponibilidad del P en losOxisoles de baja fertilidad.
2. Todas las fracciones de Pencontradas en las heceseran mayores que las delsuelo que circundaba lasheces (aumento quecorrespondía al del P total),principalmente en lasfracciones inorgánicas lábilesde P.
3. Los individuos deMartiodrilus carimaguensis,en condiciones de campo,incorporan el P de lahojarasca o del materialvegetativo aéreo y radiculardescompuesto. Este P seincorpora en todas lasfracciones (orgánicas einorgánicas) de P, peroespecialmente en susfracciones inorgánicaslábiles.
Capítulo 15Materia orgánica del suelocontenida en las heces queproduce una comunidad delombrices anécico-endógeade un pastizal tropical, enColombia1. En las heces de lombriz se
encontraron contenidos deagregados estables más altosque en el suelo circundante,que correspondían, a su vez,a concentraciones altas de Corgánico.
2. La mezcla estrecha dematerial fresco con suelo(efecto anécico) llevó a mayorconcentración de recursosprimarios (carbohidratos ylignina) y a menorconcentración de compuestoshúmicos (C aromáticos ygrupos –COOH sustituidos enel C).
3. La actividad de las lombricestrajo consigo unaredistribución muy notoria dela MO en agregados detamaños diferentes.
4. La descomposición de la MOdel suelo asociada conpartículas de limo y arcilla sehalla, al parecer, en unestado más avanzado en elsuelo circundante que en lasheces.
Capítulo 16Dinámica del carbono y delnitrógeno en las heces dediferente edad depositadaspor lombrices en áreas desabana y de pastizal en losLlanos Orientales deColombia1. Las lombrices seleccionan
sustratos alimenticios cuyovalor de C orgánico se

Conclusiones Generales...
427
incrementa durante elenvejecimiento de losturrículos, lo cual se debe, enparte, a los procesos defijación del CO2, a lasraicillas muertas o a laactividad en las heces dealgunos grupos demacrofauna.
2. Durante el primer mes seobservan contenidos altos deNH4 en las heces de losindividuos deM. carimaguensis a causa,principalmente, de ladeposición continua dematerial fresco sobre elturrículo. Una vez que lalombriz abandona la galería,ese nivel desciende y elcontenido de N inorgánicoque queda provendría de laproducción de NO3
en hecesfrescas mediante procesos denitrificación.
3. En las heces frescas seobserva una rápidaproducción de NO3
que, mástarde, se difunde hacia elsuelo circundante, primeroen sentido vertical y despuésen sentido horizontal.
4. Entre 3 y 34 kg/ha al año deN inorgánico se liberan en lasheces frescas de losindividuos de M.carimaguensis en la sabana yen el pastizal,respectivamente, lo queequivale a casi un 22% del Ntotal anual asimilado por esteúltimo.
5. La gran cantidad de sueloprocesado por las lombriceshace extensivos,especialmente en el manejode la MO del suelo, losefectos que causaglobalmente dicho proceso enla fertilidad del suelo y en laproducción vegetal; éste esun paso fundamental en el
mejoramiento de lasostenibilidad de losagroecosistemas y en ladisminución de las emisionesde CO2 a la atmósfera.
Capítulo 17Características delintercambio iónico en lasheces de la lombriz anécicaMartiodrilus carimaguensisJiménez y Moreno, en unOxisol de una sabana deColombia1. La introducción de pastizales
en los suelos de sabana y elmanejo de éstos conducen acambios importantes en dosparámetros: la fertilidad delsuelo y el comportamientoelectroquímico de susnutrientes.
2. El tránsito del suelo por elintestino de la lombrizocasiona grandes cambios enel punto de carga cero,especialmente en la sabana,donde la calidad de la MO delsuelo y de la hojarasca esmuy baja.
3. Los cambios de laspropiedades del suelo quederivan de la actividad de laslombrices pueden llevar auna mejora en la capacidadde intercambio catiónico delos Oxisoles infértiles.
Capítulo 18Las comunidades de lamacrofauna de la superficiedel suelo asociadas con lasheces de las lombrices detierra en los LlanosOrientales de Colombia1. La densidad y la biomasa de
las tres categorías ecológicasde lombrices fueronfuertemente afectadas por la
-
-
+

428
El Arado Natural:…
cantidad de turrículosdepositados por M.carimaguensis sobre lasuperficie del suelo; además,las comunidades deorganismos encontradas enlos turrículos estabanrepresentadasprincipalmente (en sudensidad y en su biomasa)por especies epígeas en lasabana y por especiesanécicas en el pastizal.
2. Las heces de las lombricesparecen mejorar lascondiciones de vida de otrasespecies, puesto que lariqueza taxonómica y ladiversidad de los organismosaumentó cuando seincrementó el número dedeposiciones superficiales.
3. Los resultados obtenidosapoyan la hipótesis de lasbiodiversidades encajadas,según la cual los ingenierosdel ecosistema determinan laabundancia y la estructurade las comunidades deorganismos más pequeños.
Capítulo 19Efectos causados por laslombrices en el banco desemillas permanente delsuelo en sistemas desabana y de pastizal enColombia1. El porcentaje de germinación
de las semillas que sehallaban en las heces de laslombrices fue de 3 a 40 vecesmenor que en lasencontradas en el suelo, aconsecuencia de los dañosque sufren las semillas
cuando son ingeridas por laslombrices. En las hecesdepositadas sobre lasuperficie del suelo hay de18 a 878 semillasgerminables por m2 por año(hasta 8.7 millones desemillas por hectárea poraño), cantidad querepresenta del 1% al 13% deltotal de semillas del bancopermanente del suelo.
2. La diversidad, la riqueza deespecies y la equitatividad dela vegetación disminuyeronconsiderablemente en losagroecosistemas, si secomparan éstos con lasabana nativa.
3. Es débil la relación entre lacomposición específica delbanco de semillas del suelo yla vegetación que crece en lasuperficie del suelo,resultado que se explica porfactores ecológicos, como lareproducción vegetativa, laquema de la sabana, ladispersión de las semillas porel viento o los animales, y elsobrepastoreo.
4. La composición de lavegetación superficial estuvomás cerca de la que seobtuvo en el banco desemillas de las heces que dela obtenida en el banco desemillas del suelo.
5. Es posible considerar que losturrículos de M.carimaguensis son nichosimportantes de regeneraciónde la comunidad vegetal, yque la actividad de laslombrices es un factorrelevante de la diversidad y lasostenibilidad de losecosistemas.

Conclusiones Generales...
429
Capítulo 20Determinación, mediante laespectroscopía de RMN con31P, del nivel de fósforoorgánico de un Oxisol, enpastizales del trópicoderivados de la sabananativa1. El establecimiento de los
pastizales mejoradosaumentó, en más del 40%, elcontenido total de P en losácidos húmicos y fúlvicos, yestos valores fueron de 2 a4 veces superiores en lasheces que en el suelo que lascircundaba, lo que indica unenriquecimiento notable del Pextraíble.
2. El P del ácido húmico estuvosiempre dominado por lasespecies que contenían Porgánico, y la cantidad de Porgánico en el ácido húmicofue proporcionalmente másalta en las heces que en elsuelo.
3. La actividad de las lombricesmejora la disponibilidad del Pen los pastizales porque creauna reserva de P orgánicoque proviene de laestabilización física de ésteen el interior de las heces.
Capítulo 21Alteración de la lignina yde los carbohidratos en losagregados de un Oxisol, enpastizales tropicalesderivados de la sabananativa1. Los turrículos de las
lombrices se caracterizan porun notable enriquecimientoen C, en lignina y enazúcares de la fracción dearena, si se comparan conlos contenidos del suelo
alrededor de ellos, lo quesugiere una importanteincorporación de hojarasca ala parte mineral del suelo(efecto anécico).
2. La concentración decarbohidratos en la MOasociada a la fracción dearenas aumentó según elorden siguiente: sabananativa < B. decumbens < B.decumbens + P. phaseoloides< turrículos de lombrices deB. decumbens + P.phaseoloides.
3. La actividad de las lombricestiene un efecto importante enla redistribución de la MO delsuelo. En la fracción dearenas de los turrículos seencontraron valores altos deC, en comparación con elsuelo que los circundaba. Lahojarasca fresca fueincorporada a la partemineral del suelo, mientrasque la lignina se preservabaen los turrículos.
4. El enriquecimiento de lafracción de limos endetrimento de la de arcillasse produjo, probablemente,por la unión fuerte que existeentre las partículas de arcillay las de limos mediante elmucílago secretado por laslombrices.
Capítulo 22Efecto de la exclusión de lalombriz anécicaMartiodrilus carimaguensisJiménez y Moreno en laspropiedades del suelo y enel crecimiento de lasplantas, en pastizales delos Llanos Orientales deColombia1. Los efectos que causa la
ausencia de M. carimaguensis

430
El Arado Natural:…
en las propiedades del sueloy en el crecimiento de lasplantas están asociados auna compactación grande delsuelo, a un contenido bajo deC, a la saturación alta de Al,a una baja biomasa vegetal ya una alta biomasa demalezas; sin embargo, losprincipales factores quecontrolan las propiedades delsuelo fueron el tipo depastizal y la edad de éste.
2. La pérdida de alguna especie,principalmente si estáasociada a una reducciónimportante de la biomasa,conduce a pérdidassignificativas en elfuncionamiento delecosistema. Porconsiguiente, hay que prestaratención al manejo de lascomunidades de lombrices detierra en los agroecosistemas,con el fin de obtener, graciasa sus actividades, un efectofavorable en la fertilidad delsuelo.
Capítulo 23Estructura espacio-temporal de una comunidadde lombrices de tierra yheterogeneidad del suelo enun pastizal tropical enCarimagua, Colombia1. Mediante un estudio de
análisis espacial y unenfoque del nivel más alto almás bajo (“top-down”) sedemostró la existencia deuna estructura espacial en lacomunidad de lombrices delas sabanas de Carimagua.
2. Existe una relaciónsignificativa entre laestructura espacio-temporalde la comunidad delombrices y la heterogeneidad
espacial del suelo y de labiomasa radicular.
3. Los patrones de distribuciónespacial son estables ymuestran la oposiciónexistente entre Andiodrilussp. y Glossodrilus sp.,resultado que indicaría lapresencia de un fenómeno deexclusión competitiva.
4. Existe una oposición espacialentre las especies pequeñas ylas de mayor tamaño. Losparches de poblacióndominados por las especiesgrandes se oponen a otros enque las especies pequeñasson más abundantes, ycorresponden a áreascaracterizadas porpropiedades del suelo yvalores de biomasa radicularque son específicos.
Necesidades de laInvestigación Futura
Todo el conocimiento recopilado eneste estudio se está empleando en eldesarrollo de un modelo que permitael manejo indirecto de las actividadesde las lombrices de tierra, con el finde optimizar sus efectos benéficos enlos agroecosistemas (Figura 24-5;L. Mariani sin publicar, citado enLavelle 2000). El objetivo del modeloes establecer una acción permanentede la fauna del suelo en losagroecosistemas, con el fin de deobtener efectos favorables en laestructura física del suelo y en ladinámica de la MO del suelo. Elmodelo explica las interacciones queocurren entre tres entidades: elagroecosistema (o sea, las opcionesde manejo elegidas), las propiedadesdel suelo (la dinámica de la MO delsuelo y la estructura física de éste) yla macrofauna del suelo,especialmente las actividades de losingenieros invertebrados del suelo y

Conclusiones Generales...
431
los efectos que causan lasestructuras biogénicas producidaspor ellos (Lavelle 2000).
El seguimiento de las estructurasbiogénicas producidas por losingenieros del ecosistema del sueloes un primer paso para comprenderlas relaciones existentes entre labiodiversidad y la función que éstadesempeña. Además de este enfoquepreliminar, es necesario fortalecer eltrabajo orientado a entender losaspectos funcionales de estasestructuras, mediante el desarrollode estudios a diferentes escalasespacio-temporales y desde la escalade la estructura hasta la de laparcela o la población. Debenconsiderarse también la dinámica delas estructuras biogénicas durantesu envejecimiento y sus efectos
concomitantes en las diferentespropiedades del suelo. Esta es unalabor importante ya que, cuandodesaparecen de la parcela algunosgrupos de la fauna del suelo (porejemplo, las lombrices de tierra) susefectos pueden persistir dentro de laescala de su dominio funcional, esdecir, la drilosfera. Por lo tanto,gracias a los procesos bien sea físicoso químicos, como se ha mostrado enlos capítulos previos, que ocurren enlas estructuras biogénicas, lasostenibilidad del agroecosistemapuede mantenerse durante algúntiempo. Una prioridad de lainvestigación debería ser ladeterminación del período de vida deestas estructuras y de laspropiedades relacionadas con ellas,por ejemplo, la infiltración del agua.
Figura 24-5. Modelo propuesto para el manejo de las poblaciones de lombrices y de otros ingenieros delecosistema (L. Mariani, sin publicar; citado en Lavelle 2000).
Materia orgánica
Estructura
Disposición espacialde las parcelas
Prácticasagrícolas
AGROECOSISTEMA
SUELO
MigraciónTratamiento del suelo
Otros ingenierosdel ecosistema
Lombrices
Estructurasbiogénicas de
superficie
MACROFAUNA
Valor alimenticiodel suelo
Recursotrófico
Estimulación delcrecimiento vegetal
AgregaciónPorosidad
Indicador

432
El Arado Natural:…
Los ingenieros del ecosistema delsuelo deberían clasificarse tambiénsegún los efectos que ejerzan en laestructura física del suelo, en ladinámica de la MO del suelo, en labiodiversidad de otros organismos yen la producción primaria. Senecesitan más estudiospormenorizados para desarrollaropciones alternas de manejo, si sequiere prevenir o revertir ladegradación del suelo mediante lamanipulación de las actividades delos organismos ingenieros delecosistema que hagan impactopositivo y significativo en laestructura y en la fertilidad del suelo.Si se conserva una faunadiversificada en que haya especiesepígeas, endógeas y anécicas, sefavorecerán también aquellasespecies que no son ingenieras, esdecir, las epígeas o las quetransforman la hojarasca (Lavelle1996). Se puede, por tanto,optimizar e incrementar lasactividades de los organismosbenéficos del suelo a escala de laparcela o del agroecosistema, sindejar de atender los requisitos delproductor en términos deproductividad y de beneficios.
A escala de la parcela se podríaincrementar la capacidad de cargade algunas especies selectas de estosorganismos; sin embargo, y enprimer lugar, se debe conocer lasensibilidad de las diferentesespecies de la comunidad demacroinvertebrados a las diversasprácticas agrícolas empleadas, comola labranza, la fertilización y elcontrol de plagas, así como adiferentes regímenes alimenticios.
Es posible lograr un manejoindirecto de las actividades de lamacrofauna del suelo manteniendodiferentes sistemas de uso de latierra, naturales y no naturales, a laescala del paisaje. Los ecosistemasnaturales, en este caso la sabana
nativa, actuarían como el reservoriooriginal de la fauna del suelo, que esfuncionalmente diverso. Esnecesario estudiar, a la escala delagroecosistema o de la práctica demanejo, la capacidad que tienen lasespecies clave para migrar desde elpastizal mejorado y desde la sabananativa donde los niveles,respectivamente, de biomasa y dediversidad son altos, hacia sistemasde manejo de la tierra, como loscultivos, donde la biodiversidad esbaja.
Las estructuras biogénicas sepueden emplear como índice de lasactividades de los ingenieros delecosistema en los cultivos, yservirían de indicadores ensustitución de las mediciones muylaboriosas que requieren laspoblaciones de dichos ingenieros. Sise logra entender la dinámica (o sea,la producción y la degradación) delas estructuras biogénicas, seráposible establecer las densidadescríticas que sirvan como advertenciaoportuna de los efectos que podríacausar un cambio ocurrido en lasprácticas agrícolas.
Es difícil estudiar este tema auna escala que supere el tamaño delas estructuras biogénicas; por tanto,los trabajos descritos en la mayorparte de los capítulos de la obrasobre los efectos de los ingenieros delecosistema se han realizado en dichaescala. Se requieren aún estudiosque establezcan el papel funcional delos organismos del suelo y lasopciones de que se dispone paraoptimizar sus actividades medianteun modelo de manejo integrado enlos agroecosistemas. Esos estudioscaracterizarían primero lasestructuras biogénicas producidaspor otras especies de lombrices paraprobar la hipótesis de que los efectospercibidos en los procesos del sueloestán, de hecho, jerárquicamenteregulados. La dinámica de las

Conclusiones Generales...
433
malezas y la actividad de laslombrices son también un temadigno de un estudio futuro, lo mismoque el papel de las lombrices en ladispersión de los propágulos demicorriza y en la estimulación de lamicroflora.
Es necesario investigar tambiénel papel de la macrofauna en ladegradación de los pastizalestropicales, especialmente en elCerrado brasileño donde, a diferenciade lo que ocurre en Carimagua, sehan encontrado pruebas evidentes dedegradación física, química ybiológica. Las causas de ladegradación de los pastizales deltrópico húmedo no se hanesclarecido todavía (Fisher et al.1995). Hay que considerar la posiblecontribución de las especiesendémicas de lombriz de tierra —quese adaptan a los cambios ocurridosen el ecosistema original— al diseñode prácticas agrícolas cuyos efectosen el ecosistema sean menosdañinos. Este conocimientopermitirá mejorar la sostenibilidad,ya que el uso calculado de losrecursos biológicos podría mantenerla fertilidad de los ecosistemasnaturales y lograr un nivel alto deproductividad primaria en los suelospobres.
Los bioindicadores de ladiversidad y la calidad del suelo sontemas importantes que deberíanconsiderarse como una prioridad enla investigación futura. A partir de lapresencia y la abundancia de losgrupos de macrofauna mássensibles, se computa un índice y serelaciona con las prácticas agrícolas,para evaluar el valor de labiodiversidad en el mantenimiento delas propiedades físicas del suelo, enla conservación de su MO y en laproducción vegetal primaria. Esposible calcular un índice indirectomediante un análisis multivariableque establezca la relación que existe
entre la biodiversidad, la funciónestructural y las propiedades físicasy químicas del suelo. Estos estudios,tomados en conjunto, darían unmejor manejo del agroecosistemaporque promoverán la integración dela productividad, la sostenibilidad yla conservación de los recursosnaturales.
Finalmente, la adopción de lasprácticas de manejo que incrementeno estimulen, ya sea elestablecimiento o ya elmantenimiento de poblacionessignificativas de macrofauna,requiere de una labor adicional decapacitación y de motivación de losagricultores. Este proceso variará deun país a otro y de una región a otra.Es necesario establecer un lenguajecomún entre los agricultores y loscientíficos, de modo que losprincipios y los conceptos relativos almanejo de los organismos benéficosdel suelo ganen mayor aceptación ysean desarrollados más tarde por lospropios agricultores.
ReferenciasAllen TFH; Starr TB. 1982. Hierarchy:
perspectives for ecologicalcomplexity. University of ChicagoPress, Chicago, IL, Estados Unidos.
Anderson JM. 1995. Soil organisms asengineers: microsite modulation ofmacroscale processes. En: JonesCG; Lawton JH, eds. Linkingspecies and ecosystems. Chapmanand Hall, Londres. p 94-106.
Barois I; Cadet P; Albrecht A; Lavelle P.1988. Systèmes de culture et faunades sols: Quelques donnés. En:Feller C, ed. Fertilité des sols dansles agricultures paysannescaribéenes: effet des restitutionsorganiques. Institut Français deRecherche Scientifique pour leDéveloppement en Coopération(ORSTOM)–Martinique, París,Francia. p 85-96.

434
El Arado Natural:…
Barros E; Grimaldi M; Desjardins T;Sarrazin M; Chauvel A; Lavelle P.1998. Conversion of forest intopastures in Amazonia: effects on soilmacrofaunal diversity and soil waterdynamics. En: Proceedings of the16th World Congress of Soil Science,agosto 1998, Montpellier, Francia.ISSS, AISS, IBG, SICS and AFES,Montpellier, Francia. vol. 1, p 203(también en CD-ROM).
Blanchart E; Albrecht A; Alegre J;Duboisset A; Gilot-Villenave C;Pashanasi B; Lavelle P; Brussaard L.1999. Effects of earthworms on soilstructure and physical properties.En: Lavelle P; Brussaard L; HendrixPF, eds. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido. p 149-172.
Bouché MB. 1977. Strategieslombriciennes. En: Lohm U; PerssonT, eds. Soil organisms as componentof ecosystems. Ecol Bull 25:122-132.
Chauvel A; Grimaldi M; Barros E;Blanchart E; Desjardins T; SarrazinM; Lavelle P. 1999. Pasture damageby an Amazonian earthworm. Nature398:32-33.
CIAT. 1999. Decision making forsustainable natural resourcemanagement: nine tools that help.Cali, Colombia.
Dangerfield JM. 1990. Abundance,biomass and diversity of soilmacrofauna in savanna woodlandand associated managed habitats.Pedobiologia 34:141-150.
Decaëns T. 1999. Rôle fonctionnel etréponses aux pratiques agricoles desvers de terre et d’autres ingénieursécologiques dans les savanescolombiennes. Tesis (Ph.D.).Université Paris VI, París, Francia.412 p.
Decaëns T; Jiménez JJ. 2002. Earthwormcommunities under an agriculturalintensification gradient in Colombia.Plant Soil 240(1):133-143.
Decaëns T; Lavelle P; Jiménez JJ; EscobarG; Rippstein G. 1994. Impact of landmanagement on soil macrofauna inthe Oriental Llanos of Colombia. EurJ Soil Biol 30(4):157-168.
Decaëns T; Galvis JH; Amézquita E. 2001.Properties of some structurescreated by soil ecosystem engineersin a Colombian savanna. C R AcadSci, Ser III. 324(5):465-478.
Decaëns T; Jiménez JJ; Lavelle P. 1999a.Effects of exclusion of the anecicearthworm Martiodriluscarimaguensis Jiménez and Morenoon soil properties and plant growthin grasslands of the Eastern Plains ofColombia. Pedobiologia 43:835-841.
Decaëns T; Rangel AF; Asakawa N;Thomas R. 1999b. Carbon andnitrogen dynamics in earthwormcasts in grasslands of the EasternPlains of Colombia. Biol Fertil Soils30:20-28.
Duboisset A. 1995. Caractérisation etquantification par analyse d’imagedes modifications structuralesengendrées par Pontoscolexcorethrurus (Glossoscolecidae,Oligochaeta) sur un sol ferrallitiquecultivé (Pérou). DEA Pédologia,Université Nancy I, Francia.
Fisher MJ; Rao IM; Ayarza MA; LascanoCE; Sanz JI; Thomas RJ; Vera RR.1994. Carbon storage by introduceddeep-rooted grasses in the SouthAmerican savannas. Nature371:236-238.
Fisher MJ; Rao IM; Thomas RJ; LascanoCE. 1995. Grasslands in the well-watered tropical lowlands. En:Hodgson J; Illius AW, eds. Theecology and management of grazingsystems. CAB International,Wallingford, Reino Unido.p 393-425.

Conclusiones Generales...
435
Fisher MJ; Thomas RJ; Rao IM. 1997.Management of tropical pastures inacid-soil savannas of SouthAmerica for carbon sequestration inthe soil. En: Lal R; Kimble JM;Follet RF; Stewart BA, eds.Management of carbonsequestration in the soil. CRCPress, Boca Ratón, FL, EstadosUnidos. p 405-420.
Fragoso C; Brown GG; Patrón JC;Blanchart E; Lavelle P; PashanasiB; Senapati BK; Kumar T. 1997.Agricultural intensification, soilbiodiversity and ecosystem functionin the tropics: the role ofearthworms. Appl Soil Ecol 6:17-35.
Fragoso C; Lavelle P; Blanchart E;Senapati BK; Jiménez JJ; MartínezMA; Decaëns T; Tondoh J. 1999.Earthworm communities of tropicalagroecosystems: origin, structureand influence of managementpractices. En: Lavelle P; BrussaardL; Hendrix PF, eds. Earthwormmanagement in tropicalagroecosystems. CAB International,Wallingford, Reino Unido. p 27-55.
Giller PS. 1996. The diversity of soilcommunities, the “poor man’stropical rainforest”. BiodiversConserv 5:135-168.
Giller KE; Beare MH; Lavelle P; Izac A-MN; Swift MJ. 1997. Agriculturalintensification, soil biodiversity andagroecosystem function. Appl SoilEcol 6:3-16.
Gilot-Villenave C; Lavelle P; Keli J;Kouassi P; Guillaume G. 1995.Biological activity of soil underrubber plantations in Côte d’Ivoire.Acta Zool Fenn 196:186-189.
Huston MA. 1996. Biological diversity:the coexistence of species inchanging landscape. CambridgeUniversity Press, Reino Unido.
Jiménez JJ. 1999. Estructura de lascomunidades y dinámica de laspoblaciones de lombrices de tierraen sabanas naturales y perturbadasde Carimagua (Colombia). Tesis(Doctorado). UniversidadComplutense de Madrid, España.311 p.
Jiménez JJ; Decaëns T. 2000. Verticaldistribution of earthworms ingrassland soils from the ColombianLlanos. Biol Fertil Soils 32:463-473.
Jiménez JJ; Brown GG; Decaëns T; FeijooT; Lavelle P. 2000. Differences inthe timing of diapause and patternsof aestivation in some tropicalearthworms. Pedobiologia44:677-694.
Jiménez JJ; Moreno AG; Decaëns T;Lavelle P; Fisher MJ; Thomas RJ.1998. Earthworm communities innative savannas and man-madepastures of the Eastern Plains ofColombia. Biol Fertil Soils28:101-110.
Jones CG; Lawton JH; Shachak M. 1994.Organisms as ecosystem engineers.Oikos 69:373-386.
Jones CG; Lawton JH; Shachak M. 1997.Positive and negative effects oforganisms as physical ecosystemengineers. Ecology 78(7):1946-1957.
Kalisz PJ; Wood HB. 1995. Native andexotic earthworms in wildlandecosystems. En: Hendrix PF, ed.Earthworm ecology andbiogeography in North America.Lewis Publishers, Boca Raton, FL,Estados Unidos. p 117-126.
Lavelle P. 1978. Les vers de terre de lasavanne de Lamto (Côte d’Ivoire):Peuplements, populations etfonctions dans l’ecosystème. Tesis(Doctorado). Université Paris VI,Francia.
Lavelle P. 1981. Stratégies dereproduction chez les vers de terre.Acta Oecol 2(2):117-133.

436
El Arado Natural:…
Lavelle P. 1983. The soil fauna of tropicalsavannas; II: The earthworms. En:Bourlière F, ed. Tropical savannas.Elsevier Scientific Publishing,Amsterdam, Holanda. p 485-504.
Lavelle P. 1988. Earthworm activities andthe soil system. Biol Fertil Soils6:237-251.
Lavelle P. 1996. Diversity of soil faunaand ecosystem function. Biol Int33:3-16.
Lavelle P. 1997. Faunal activities and soilprocesses: adaptive strategies thatdetermine ecosystem function. AdvEcol Res 27:93-132.
Lavelle P. 2000. Ecological challenges forsoil science. Soil Sci 165:73-86.
Lavelle P; Pashanasi B. 1989. Soilmacrofauna and land managementin Peruvian Amazonia (Yurimaguas,Loreto). Pedobiologia 33:283-291.
Lavelle P; Dangerfield M; Fragoso C;Eschenbrenner V; López-HernándezD; Pashanasi B. 1994. Therelationship between soilmacrofauna and tropical soilfertility. En: Woomer PL; Swift MJ,eds. The biological management oftropical soil fertility. Wiley andSons, Chichester, Reino Unido.p 137-169.
Lavelle P; Brussaard L; Hendrix PF, eds.1999. Earthworm management intropical agroecosystems. CABInternational, Wallingford, ReinoUnido.
Lawton JH; Jones CG. 1995. Linkingspecies and ecosystems: organismsas ecosystem engineers. En: JonesCG; Lawton JH, eds. Linkingspecies and ecosystems. Chapmanand Hall, Londres. p 141-150.
Lee KE. 1991. The diversity of soilorganisms. En: Hawksworth DL, ed.The biodiversity of microorganismsand invertebrates: its role insustainable agriculture. CABInternational, Wallingford, ReinoUnido. p 73-87.
Mariani L; Bernier N; Jiménez JJ; DecaënsT. 2001. Régime alimentaire d’un verde terre des savanes colombiennes:une remise en question des typesécologiques. C R Acad Sci, Ser III.324(8):733-742.
Ortiz B; Fragoso C; M’Boukou I; PashanasiB; Senapati BK; Contreras A. 1999.Perception and use of earthworms intropical farming systems. En: LavelleP; Brussaard L; Hendrix PF, eds.Earthworm management in tropicalagroecosystems. CAB International,Wallingford, Reino Unido.p 239-252.
Rao IM. 1998. Root distribution andproduction in native and introducedpastures in the South Americansavannas. En: Box, Jr. JE, ed. Rootdemographics and their efficienciesin sustainable agriculture,grasslands and ecosystems. KluwerAcademic Publishers, Amsterdam,Holanda. p 19-41.
Rippstein G; Lascano CE; Decaëns T.1996. La production fourragère dansles savanes d’Amerique de Sudintertropicale. Fourrages 145:33-52.
Sanz JI; Zeigler RS; Sarkarung S; MolinaDL; Rivera M. 1999. Sistemasmejorados arroz-pasturas parasabana nativa y pasturasdegradadas en suelos ácidos deAmérica del Sur. En: Guimarães EP;Sanz JI; Rao IM; Amézquita MC;Amézquita E, eds. Sistemasagropastoriles en sabanas tropicalesde América Latina. CIAT, Cali,Colombia. p 232-244.
Schneidmadl J; Decaëns T. 1995. Soilmacrofauna in managed savannas atMatazul farm, Eastern Plains,Colombia. En: CIAT annual report,1995. CIAT, Cali, Colombia.
Senapati BK; Panigrahi PK; Lavelle P.1994. Macrofaunal status andrestoration strategy in degraded soilunder tea cultivation in India. En:Etchevers JD, ed. Proceedings of the15th World Congress of Soil Science,Acapulco, México. p 64-75.

Conclusiones Generales...
437
Senapati BK; Lavelle P; Giri S; PashanasiB; Alegre J; Decaëns T; Jiménez JJ;Albrecht A; Blanchart E; MahieuxM; Rousseaux L; Thomas RJ;Panigrahi PK; Venkatachalam M.1999. In-soil earthwormtechnologies for tropicalagroecosystems. En: Lavelle P;Brussaard L; Hendrix PF, eds.Earthworm management in tropicalagroecosystems. CAB International,Wallingford, Reino Unido.p 199-237.
Solbrig OT; Medina E; Silva JF. 1996.Determinants of tropical savannas.En: Solbrig OT; Medina E; Silva JF,eds. Biodiversity and savannaecosystem processes. Springer-Verlag, Berlín, Alemania. p 31-41.
Swift MJ; Anderson JM. 1994.Biodiversity and ecosystem functionin agricultural systems. En: SchulzeED; Mooney HA, eds. Biodiversityand ecosystem function. Springer-Verlag, Berlín, Alemania. p 15-41.
Swift MJ; Vandermeer J; RamakrishnanPS; Anderson JM; Ong CK; HawkinsBA. 1996. Biodiversity andagroecosystem function. En:Mooney HA; Cushman JH; MedinaE; Sala OE; Schulze ED, eds.Functional roles of biodiversity: aglobal perspective. John Wiley andSons, Nueva York, Estados Unidos.p 261-298.
Thomas RJ. 1992. The role of legume inthe nitrogen cycle of productive andsustainable pastures. Grass ForageSci 47:133-142.
Thomas RJ; Lascano CE; Sanz JI; AraMA; Spain JM; Vera RR; Fisher MJ.1995. The role of forage grasses andlegumes in maintaining theproductivity of acid soils in LatinAmerica. En: Lal R; Stewart BA,eds. Soil management: experimentalbasis for sustainability andenvironmental quality. Advances inSoil Science Series. LewisPublishers, Boca Ratón, FL,Estados Unidos. p 61-83.
Acrónimos y Abreviaturas Usados en el Texto
439
Acrónimos y Abreviaturas Usados enel Texto
AcrónimosAECI Agencia Española de
CooperaciónInternacional
CE Comunidad Europea
CIAT Centro Internacional deAgricultura Tropical
CIRAD Centre de CoopérationInternationale enRecherche Agronomiquepour le Développement(Montpellier, Francia)
CNIA Centro Nacional deInvestigacionesAgropecuarias
CORPOICA Corporación Colombianade InvestigaciónAgropecuaria
CPMAS Cross Polarization Magic-Angle Spinning (giros deángulo mágico en lapolarización cruzada,técnica del análisis RMNcon 13C)
CSIRO Organización de laCommonwealth para laInvestigación Industrial yCientífica (Australia);(CommonwealthScientific and IndustrialResearch Organisation(Australia)
CIMMYT Centro Internacional deMejoramiento de Maíz yTrigo
ETH EidgenössischeTechnische HochschuleZürich (Instituto deTecnología Federal Suizo)
GCIAI Grupo Consultivo para laInvestigación AgrícolaInternacional
IBP Programa BiológicoInternacional
IFDC International Fertilizerand Development Center
IFS Fundación Internacionalpara la Ciencia
IGAC Instituto GeográficoAgustín Codazzi (enBogotá)
IRD Institut de Recherchepour le Développement(ex-ORSTOM)
IUBS Unión Internacional deCiencias Biológicas
LEST Laboratoire d’Ecologiedes Sols Tropicaux (enIRD)
MAS Manejo de Suelos Acidos(del programa SWNM delCGIAR)
ORSTOM Institut Français deRecherche Scientifiquepour le Développementen Coopération
RMN (Espectroscopía de)Resonancia MagnéticaNuclear
SAS© Sistema de AnálisisEstadístico

440
El Arado Natural:…
SPSS© Paquete Estadístico paraCiencias Sociales. (Serefiere al SPSS original)
STD Programa de DesarrolloCientífico y Técnico de laUnión Europea
SWNM (Consorcio Ejecutor delPrograma de Todos losCentros del CGIAR sobreel Manejo de Suelos,Agua y Nutrientes
TSBF (Programa de) Biología yFertilidad de los SuelosTropicales
UE Unión Europea
USDA Departamento deAgricultura de losEstados Unidos deAmérica
Abreviaturas[(ac/al)C] Relación ácido/aldehído
en un grupo (unidad) decinamil
[(ac/al)S] Relación ácido/aldehídoen un grupo (unidad) desiringil
[(ac/al)V] Relación ácido/aldehídoen un grupo (unidad) devanilil
ACP Análisis de componentesprincipales
Ad Adulto
ADE-4 Análisis de variablemúltiple (multivariado) yprograma (software) deexpresión gráfica
ADN Acidodesoxirribonucléico
AF Acido fúlvico
AH Acido húmico
ANOVA Análisis de varianza (enestadística paramétrica)
ATF Acido trifluoroacético
ATP Análisis triádico parcial
B Biomasa
b Indice de agregaciónespacial de Taylor
BA Barbecho de malezas ohelechos arbustivos (deporte bajo; BU en inglés)
Bdec Pastizal de Brachiariadecumbens
Bdec/Pp Pastizal de Brachiariadecumbens + Puerariaphaseoloides
BH Pastizal de Brachiariahumidicola
BK Pastizal de B. decumbensy Kudzú (P. phaseoloides)
BL Biomasa de lombrices detierra
BR Biomasa de las raíces
BS Bosque secundario(Cap. 3); banco desemillas (en el suelo)(Cap. 19)
c.c. Concentrado y caliente13C Carbono 13 (isótopo)
C/N Relación carbono/nitrógeno
°C Grados centígrados
© Copyright
CC Cultivo tradicional decafé
CG Cromatografía de gases
CG/EM Relación entre lacromatografía de gases yla espectrometría demasa
CIC Capacidad deintercambio catiónico
cm Centímetros
CM Cultivo de marañón(Anacardium occidentaleL.)
cmol centimol
CO2 Dióxido de carbono
COM Complejos órgano-minerales

Acrónimos y Abreviaturas Usados en el Texto
441
COOH¯ Grupo carboxílico
Cs Indice de similitud deSørensen
CT, CT Carbono total
CuO Oxido de cobre (cúprico)
CV Coeficiente de variación(en estadísticas)
cv. Cultivar, variedad
D Densidad
DA Densidad aparente
DDF Días después del fuego
DEH Distribución espacialhorizontal
DPM Diámetro ponderal medio(de los agregados delsuelo)
dt Desviación típica (enestadística)
DV Distribución vertical (delas lombrices)
EAA Estabilidad de losagregados (del suelo) enagua
EC Enriquecimiento en C
EJ Estimador “jackknife”
EM Extracción manual
ES Error estándar
Estrategia K Estrategia usada poralgunos organismos, porejemplo las lombrices detierra, con el fin de hacerun aporte de individuosdurante la fasereproductora y mantenerla población cerca de sucapacidad de carga.Estos organismos son detamaño grande, de ciclode vida largo (variosaños) y producen pocoscapullos pero de grantamaño (hasta un 20%del peso del adulto).
Estrategia r Estrategia usada poralgunos organismos, porejemplo las lombrices detierra, con el fin de hacer
un aporte muy grande deindividuos durante lafase reproductora,aunque con una elevadamortalidad. Estosorganismos son engeneral de tamañopequeño, de ciclo de vidacorto (varios meses) yproducen muchoscapullos.
g Gramos
g/g Relación entre el peso engramos del elemento quese considera y el pesototal en gramos de lamuestra
gl Grados de libertad (enestadística)
[(G+M)/(A+X)] Relación entre la suma(galactosa + manosa) y lasuma (arabinosa +xilosa), cuatro azúcaressintetizados pormicroorganismos
H Heces
H Indice de diversidadbiológica de Shannon
H’ Equitatividad (unamedida de la diversidadbiológica)
H+ Protones (Iones deHidrógeno)
H2O Fórmula química delagua
H2SO4 Acido sulfúrico
ha Hectárea
HCl Acido clorhídrico
HClO4 Acido perclórico
HF Acido fluorhídrico
HS Humedad del suelo
HSD Diferencia honestamentesignificativa (en inglés)(en estadísticaparamétrica)
Hz Hertz (unidad defrecuencia de ondas deradio)
442
El Arado Natural:…
Id Indice de Morisita(agregación espacial)
ind. Individuos
Inm Inmaduros
J Julio (unidad de potenciao trabajo)
K2S2O8 Persulfato potásico
KCl Cloruro potásico
KH2PO4 Fosfato dihidrogenado depotasio (Bifosfato depotasio)
L/D Relación entre lalongitud y el diámetromáximo de la lombriz
LA Longitud del adulto
LB Lombrices
LI Longitud del inmaduro
ln Logaritmo neperiano
LT Lavado y tamizado
m Metros
Macroinv. Macroinvertebrados
Macroorg. Macroorganismos
Max. Máximo
MC Macrofauna (Cap. 2);monocultivo (Cap. 24)
meq Miliequivalente gramo
Mg Mega-gramos (=tonelada)
mg miligramos (10-3 g)
MHz Megahertz (mil hertz)
Min. Mínimo
ml mililitro (10-3 l)
mm milímetros (10-3 m)
MO Materia orgánica
MOS Materia orgánica delsuelo
msnm Metros sobre el nivel delmar
N Inm Número inmaduros
n Número deobservaciones (enestadística)
n.sp. Nueva especie
Na4O2P7 Ortofosfato de sodio
NaCl Cloruro de sodio (sal)
NaHCO3 Bicarbonato de sodio(carbonato ácido desodio)
NaOH Hidróxido de sodio
NH4 Amonio (radical positivo)
NH4F Fluoruro de amonio
NO3 Nitrato (radical negativo)
NS No significativo (enestadística)
Ø Diámetro (símbolo)
OH¯ Iones hidróxido
ONG Organización nogubernamental
p.ej. Por ejemplo
P/D Relación entre el peso yel diámetro máximo de lalombriz
p/p Relación entre el peso delelemento que seconsidera y el peso totalde la muestra
PA Peso del adulto (Cap. 7);producción anual(Cap. 19); pastizalintroducido longevo(Cap. 24)
PC Peso del capullo
PCC Punto de carga cero
PF Peso fresco
pF Símbolo de la expresiónlogarítmica para elpotencial de succión deagua del suelo
pH Símbolo de la expresiónlogarítmica para el gradode acidez o alcalinidadde una solución (indicaconcentración de ionesde hidrógeno)
Pi Fósforo inorgánico
PI Peso del individuo (oestado) inmaduro
+
¯

Acrónimos y Abreviaturas Usados en el Texto
443
PID Producción individualdiaria
PK Pastizal de Pennisetumclandestinum (“kikuyo”)
PLSD Probabilidad de que hayauna diferencia demínimos cuadrados (unaprueba estadística)
PM Pastizal mejorado
Po Fósforo orgánico
PP Plantaciones de pino
ppm Partes por millón
PR Pastizal reciente
Pt Fósforo total
PT Pastizal tradicional
PY Pastizal de Melinisminutiflora (“yaraguá”)
R Riqueza específica
r Coeficiente de regresión(estadística)
[(R+F)/(A+X)] Relación entre la suma(ramnosa + fucosa) y lasuma (arabinosa +xilosa), cuatro azúcaressintetizados por losmicroorganismos
RA Rotación anual
RP Resistencia a lapenetración
rpm Revoluciones por minuto
rs Raíces secas
S40 Bosque de 40 años deedad (S40+ = de más de40 años)
Sav Sabana nativa
SD Sin determinar
SN Sabana nativa
SS Selva secundaria(Cap. 3); suelosubyacente (Cap. 18)
ss Suelo seco
ST Suelo testigo
Su Subadulto
t Tonelada
TF Turrículo fresco
TS Turrículo seco
UA Unidad animal (medidaempleada en ganaderíaque representa 250 kg depeso)
UT Unidad taxonómica
V/V Relación entre elvolumen del elementoque se considera y elvolumen total de lamuestra
VAR5 Programa (software)geoestadísticodesarrollado por laUniversidad de Hawai
vs. Versus (frente)
VSC La suma de las unidades(grupos químicos) devanilil (V), siringil (S) ycinamil (C)
YA Yuca asociada (con frijoly maíz)
µg Microgramo (10-6 g)
µm Micrómetro (10-6 m)
∆ Delta (letra griega que seusa para indicarvariación)
∑ Ypsilon (letra griega), enmatemáticas representala suma de un conjuntode datos
χ2 Prueba de Chi-cuadrado(en estadística)
γ (h) Semi-varianza; engeoestadística es el valorde la varianza de unavariable dada en relacióncon la distancia quesepara los puntos demuestreo

444
El Arado Natural:…
Elementos químicos:Lista de los símbolos
usados en el textoAl Aluminio
C Carbono
Ca Calcio
Cl Cloro
Fe Hierro
H Hidrógeno
K Potasio
Mg Magnesio
Mo Molibdeno
N Nitrógeno
Na Sodio
O Oxígeno (también comoO2)
P Fósforo
S Azufre
Zn Cinc

Publicación CIAT No. 336
Proyecto Recuperación de Suelos Degradadosy
Unidad de Comunicaciones
Edición: Francisco MottaGladys Rodríguez (asistente editorial)
Traducción al español: Juan José Jiménez (21 capítulos)
Producción: Artes Gráficas, CIATOscar Idárraga (diagramación)Julio C. Martínez (diseño de carátula)
Impresión: Feriva S.A., Cali, Colombia





























