Embed Size (px)
Citation preview

Universidade Federal do Rio de Janeiro
Investigação do Potencial de Aplicação de Lipases Microbianas e Vegetais
na Produção de Lipídios de Interesse Nutricional e Farmacêutico
Joab Sampaio de Sousa
RIO DE JANEIRO, RJ - BRASIL
AGOSTO DE 2014

ii
Investigação do Potencial de Aplicação de Lipases Microbianas e
Vegetais na Produção de Lipídios de Interesse Nutricional e Farmacêutico
Joab Sampaio de Sousa
RIO DE JANEIRO, RJ - BRASIL
AGOSTO DE 2014
Tese de Doutorado submetida ao corpo docente do Programa de Pós Graduação em Ciência de Alimentos do Instituto de Química da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências. Orientadores: Profa Denise Maria Guimarães Freire
Prof Alexandre Guedes Torres
Universidade Federal do Rio de Janeiro
Instituto de Química
Programa de Pós Graduação em Ciência de Alimentos

iii
Investigação do Potencial de Aplicação de Lipases Microbianas e Vegetais
na Produção de Lipídios de Interesse Nutricional e Farmacêutico
Joab Sampaio de Sousa Tese de Doutorado submetida ao corpo docente do Programa de Pós Graduação em Ciência de Alimentos do Instituto de Química da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências. Aprovada por:
______________________________________________ Profa Denise Maria Guimarães Freire, D.Sc. - UFRJ (Orientador)
______________________________________________ Prof Alexandre Guedes Torres, D.Sc. - UFRJ (Orientador)
_______________________________________________ Profa Selma Gomes Ferreira Leite, D.Sc. – UFRJ
_______________________________________________ Elisa dÀvila Cavalcanti-Oliveira, D.Sc. – UFRJ
_______________________________________________ Profa Viridiana Santana Ferreira Leitão, D.Sc. – INT
_______________________________________________ Prof Rodrigo Volcan Almeida, D.Sc. - UFRJ
Rio de Janeiro
AGOSTO de 2014
Orientadores: Profa Denise Maria Guimarães Freire
Prof Alexandre Guedes Torres

iv
RESUMO
Investigação do Potencial de Aplicação de Lipases Microbianas e Vegetais na Produção de Lipídios de Interesse Nutricional e Farmacêutico
Joab Sampaio de Sousa
Orientadores: Denise Maria Guimarães Freire e Alexandre Guedes Torres
Resumo da Tese de Doutorado submetida ao corpo docente do Programa de Pós Graduação em Ciência de Alimentos do Instituto de Química da Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutor em Ciências. O presente trabalho objetivou a investigação de 3 lipases home made e 14 lipases comerciais microbianas, na produção de 3 tipos de lipídios de interesse nutricional e farmacêutico. Dentre esses lipídios foi possível gerar óleos alimentares (óleo de coco, palma e oliva) enriquecidos em ácidos graxos poli-insaturados (AGPI) eicosapentaenóico (EPA, C20:5n3) e docosahexaenóico (DHA, C22:6n3). Com as lipases home made de Rhizomucor miehei, Candida antarctica B e de Rhizopus oryzae foi possível obter lipídios com 374, 392 e 586 mg de EPA+DHA no óleo de coco, respectivamente. Valores que atendem à necessidade de ingestão diária recomendada preconizada pela Organização Mundial da Saúde (OMS) para estes ácidos graxos. Também foi possível gerar um produto de elevado potencial farmacológico, com o enriquecimento nutricional do óleo de coco, com a aplicação da Lipozyme® TL IM (Thermomyces lanuginosus) e da Amano AK (Pseudomonas fluorescens), que pode ser utilizado na prevenção de doença coronariana (ataque cardíaco) e de acidente vascular cerebral (AVC), tendo em vista que os percentuais alcançados neste processo equivalem a 915 e 1198 mg de EPA+DHA, respectivamente. Para o óleo de palma os melhores resultados foram obtidos com as lipases comerciais de Rhizomucor miehei e de Rhizopus oryzae, com 546 e 512 mg de EPA+DHA. Já com o óleo de oliva, o melhor resultado foi obtido com a lipase de Burkholderia cepacia (Amano PS), que incorporou 455 mg de EPA+DHA.Também foi possível gerar um lipídio que pode ser utilizados como ingrediente em fórmulas infantis por ser um mimético do leite materno pela incorporação de ácido oléico (C18:1n9) na tripalmitina com as lipases de Rhizomucor miehei (Sigma-Aldrich e Lipozyme® RM IM), Novozym® 435 (Candida antarctica B), Lipozyme® TL IM (Thermomyces lanuginosus), Lipomod™ 34P (Candida rugosa) e com as lipases de Rhizopus oryzae (Sigma-Aldrich e ROL UAB). Com as quais foi possível inserir de 74,2 ± 2,14 a 41,9 ± 2,92. Também foram obtidos lipídios de menor densidade energética a partir da acidólise do óleo de peixe com o ácido octanóico (C8:0) com as lipases de Thermomyces lanuginosus (Lipozyme® TL IM e Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e de Rhizomucor miehei (Lipozyme® RM IM). Com as quais foi possível inserir de 25,38 ± 3,75 a 54,85 ± 2,34 de C8:0 no óleo de peixe, respectivamente. Não foi possível obter resultados satisfatórios com as lipases presentes nas sementes germinadas de macaúba (Acrocomia aculeata), buriti (Mauritia flexuosa L.), girassol (Helianthus annuus), gergelim (Sesamum indicum), amendoim (Arachis hypogaea L.) e abacate (Persea americana), qauando utilizadas na sob a forma de acetone powder. Palavras-chave: Lipases, lipídios estruturados, Ácidos graxos poli-insaturados, Alimentos fuoncionais, lipídios de baixo teor calórico, miméticos do leite materno.

v
ABSTRACT
Potential Application of Microbial and Vegetable Lipases Production in Lipid Nutrition and Pharmaceutical Interest
Joab Sampaio de Sousa
Orientadores: Denise Maria Guimarães Freire e Alexandre Guedes Torres
Summary of Doctoral Thesis submitted ao Programa de Pos Graduação em Ciência de Alimentos do Instituto de Química da Universidade Federal do Rio de Janeiro as part of the requirements for obtaining the degree of Doctor of Sciences. The present study aimed to investigate three home made lipases and fourteen commercial microbial lipases in order to produce lipids of nutritional and pharmaceutical interest. Among these lipids it was possible to produce cooking oils (coconut oil, palm and olive) enriched in polyunsaturated (PUFA), eicosapentaenoic (EPA, C20:5n3) and docosahexaenoic (DHA, C22: 6n3) fatty acid. Using the home made lipases from Candida antarctica B and Rhizomucor miehei B (LaBiM) and Rhizopus oryzae (UAB) it was possible to obtain lipids with 374, 392 and 586 mg of EPA+DHA in coconut oil. Values that meet the daily intake need recommended by the World Health Organization (WHO) for these fatty acids. Through the nutritional enrichment of coconut oil using the Lipozyme® TL IM (Thermomyces lanuginosus) and Amano AK (Pseudomonas fluorescens), it was also possible to generate a product of high pharmacological potential which can be used in the prevention of coronary heart disease (CHD) and stroke, given that the percentage achieved in this process is equivalent to 915 and 1198 mg of EPA+DHA, respectively. The best results with palm oil were obtained using the commercial lipases from Rhizomucor miehei and Rhizopus oryzae, both supplied by Sigma-Aldrich, with 546 and 512 mg of EPA+DHA. The best result with olive oil was obtained using lipase from Burkholderia cepacia (Amano PS), which incorporated 455 mg of EPA+DHA. It was also possible to generate a lipid which can be used as an ingredient for infant formulas because it is a mimetic for breastmilk due the incorporation of oleic acid (C18:1n9) in tripalmitin. Using lipases from Rhizomucor miehei (Sigma-Aldrich and Lipozyme® RM IM), Novozym® 435 (Candida antarctica B), Lipozyme® TL IM (Thermomyces lanuginosus), Lipomod™ 34P (Candida rugosa) and Rhizopus oryzae (ROL UAB) it was possible to achieve a performance similar to that found using the reference commercial lipase, and so to enter 41.9 ± 2.92 to 74.2 ± 2.14 of C18:1n9 in tripalmitin. Another interesting point was the obtaining of lower calorie density lipids which were acquired through the acidolysis of fish oil with octanoic acid (C8:0) using the lipases from Thermomyces lanuginosus (Lipozyme® TL IM and Lipozyme® TL 100L), Candida antarctica (Novozym® 735) and Rhizomucor miehei (Lipozyme® RM IM). With these lipases it was possible to insert 25.38 ± 3.75 to 54.85 ± 2.34 C8:0 in fish oil. Thus, it is believed that the lipid produced in this study, in addition to being functionally and nutritionally beneficial, they can be used as an important ingredient in other food formulations.
Keywords: Lipase, Structured Lipids, Polyunsaturated Fatty Acid, Functional Foods, Low-calorie Structured Lipids, Human Milk Fat Substitutes.

vi
ÍNDICE DE FIGURAS
Figura 1: Fluxograma dos objetivos específicos ............................................................................................. 1 Figura 2: Representação das reações de transesterificação, acidólise e alcoólise catalisada por lipases
na formação de diacilgliceróis, monoacilgliceróis, ácidos graxos livres e glicerol. ........................... 8 Figura 5: (a) Representação da molécula de glicerol (propan-1,2,3-triol) com a identificação dos
átomos de carbono, oxigênio e hidrogênio. (b) Estrutura geral de um triacilglicerol com as posições estereoespecíficas (sn-1, 2 ou 3) definidas. .......................................................................... 13
Figura 6: Apresentação resumida de oito etapas que compõe o processo de digestão e absorção de lipídios provenientes da alimentação. .................................................................................................. 16
Figura 7: Reação de acidólise da tripalmitina pelo ácido oléico para produção de Betapol™ ............... 26 Figura 3: Estrutura de ácidso graxos poli-insaturados de origem vegetal e animal ................................ 32 Figura 4: Via metabólica da conversão dos ácidos graxos essenciais de cadeia longa alfa-linolênico . 34 Figura 8: (a) Semente germinada de macaúba no estagio de germinação que é fornecida pela empresa
Acrotech. .................................................................................................................................................. 50 Figura 9: Comparação do perfil das famílias de ácidos graxos presentes no óleo de coco antes e após
a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Pseudomonas fluorescens (Amano AK), (ii) Thermomyces lanuginosus (Lipozyme® TL IM), (iii) Rhizomucor miehei (Lipozyme® RM IM), (iv) Rhizopus oryzae (ROL IM Sigma-Aldrich), (v) Candida antarctica B (Novozym® 435), (vi) Rhizopus oryzae (ROL UAB), (vii) Burkholderia cepacia (Amano PS), (viii) Rhizomucor miehei (RM IM LaBiM), (ix) Candida antarctica B (Lip. B LaBiM), (x) Candida antarctica A (Novozym® 735), (xi) Rhizomucor miehei (Sigma-Aldrich), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Candida rugosa (Amano AY 30) e (xiv) Candida rugosa (Lipomod™ 34P)............................................................................................................. 55
Figura 10: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de coco antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Pseudomonas fluorescens (Amano AK), (ii) Thermomyces lanuginosus (Lipozyme® TL IM), (iii) Rhizomucor miehei (Lipozyme® RM IM), (iv) Rhizopus oryzae (ROL IM Sigma-Aldrich), (v) Candida antarctica B (Novozym® 435), (vi) Rhizopus oryzae (ROL UAB), (vii) Burkholderia cepacia (Amano PS), (viii) Rhizomucor miehei (RM IM LaBiM), (ix) Candida antarctica B (Lip. B LaBiM), (x) Candida antarctica A (Novozym® 735), (xi) Rhizomucor miehei (Sigma-Aldrich), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Candida rugosa (Amano AY 30) e (xiv) Candida rugosa (Lipomod™ 34P). .................................................................. 56
Figura 11: Comparação do perfil das famílias de ácidos graxos presentes no óleo de palma antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Rhizopus oryzae (ROL IM Sigma-Aldrich), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizomucor miehei (Sigma-Aldrich), (iv) Burkholderia cepacia (Amano PS), (v) Candida antarctica B (Lip. B LaBiM), (vi) Candida antarctica B (Novozym® 435), (vii) Rhizomucor miehei (RM IM LaBiM), (viii) Rhizopus oryzae (ROL UAB), (ix) Thermomyces lanuginosus (Lipozyme® TL IM), (x) Candida rugosa (Amano AY 30), (xi) Candida rugosa (Lipomod™ 34P), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Pseudomonas fluorescens (Amano AK) e (xiv) Candida antarctica A (Novozym® 735). ................................................................................................................ 62
Figura 12: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de palma antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Rhizopus oryzae (ROL IM Sigma-Aldrich), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizomucor miehei (Sigma-Aldrich), (iv) Burkholderia cepacia (Amano PS), (v) Candida antarctica B (Lip. B LaBiM), (vi) Candida antarctica B

vii
(Novozym® 435), (vii) Rhizomucor miehei (RM IM LaBiM), (viii) Rhizopus oryzae (ROL UAB), (ix) Thermomyces lanuginosus (Lipozyme® TL IM), (x) Candida rugosa (Amano AY 30), (xi) Candida rugosa (Lipomod™ 34P), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Pseudomonas fluorescens (Amano AK) e (xiv) Candida antarctica A (Novozym® 735). ............................................. 63
Figura 13: Comparação do perfil das famílias de ácidos graxos presentes no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Burkholderia cepacia (Amano PS), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizopus oryzae (ROL IM Sigma-Aldrich), (iv) Thermomyces lanuginosus (Lipozyme® TL 100L), (v) Thermomyces lanuginosus (Lipozyme® TL IM), (vi) Rhizomucor miehei (Sigma-Aldrich), (vii) Rhizopus oryzae (ROL UAB), (viii) Candida antarctica B (Novozym® 435), (ix) Pseudomonas fluorescens (Amano AK), (x) Candida rugosa (Lipomod™ 34P), (xi) Candida antarctica B (Lip. B LaBiM), (xii) Candida rugosa (Amano AY 30), (xiii) Rhizomucor miehei (RM IM LaBiM) e (xiv) Candida antarctica A (Novozym® 735). .................................................................................................. 68
Figura 14: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Burkholderia cepacia (Amano PS), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizopus oryzae (ROL IM Sigma-Aldrich), (iv) Thermomyces lanuginosus (Lipozyme® TL 100L), (v) Thermomyces lanuginosus (Lipozyme® TL IM), (vi) Rhizomucor miehei (Sigma-Aldrich), (vii) Rhizopus oryzae (ROL UAB), (viii) Candida antarctica B (Novozym® 435), (ix) Pseudomonas fluorescens (Amano AK), (x) Candida rugosa (Lipomod™ 34P), (xi) Candida antarctica B (Lip. B LaBiM), (xii) Candida rugosa (Amano AY 30), (xiii) Rhizomucor miehei (RM IM LaBiM) e (xiv) Candida antarctica A (Novozym® 735). ............................................. 69
Figura 15: Análise do perfil em ácidos graxos da acidólise da tripalmitina com ácido oléico catalisada por pelas lipases: Rhizomucor miehei (Sigma-Aldrich), Novozym® 435 (Candida antarctica B), Lipozyme® TL IM (Thermomyces lanuginosus), Lipozyme® RM IM (Rhizomucor miehei), Lipomod™ 34P (Candida rugosa), ROL IM (Sigma-Aldrich) (Rhizopus oryzae), ROL LaBiM (Rhizopus oryzae), Amano PS IM (Burkholderia cepacia), Amano AK IM (Pseudomonas fluorescens), Lipozyme® TL 100L (Thermomyces lanuginosus), Lip. B LaBiM (Candida antarctica B), Amano AY 30 IM (Candida rugosa) e Novozym® 735 (Candida antarctica A). ............................................................................................... 76
Figura 16: Comparação do perfil das famílias de ácidos graxos presentes no óleo de peixe antes e após a reação de acidólise com o ácido octanóico (C8:0) catalisado pelas lipases de Thermomyces lanuginosus (Lipozyme® TL IM e Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e de Rhizomucor miehei (Lipozyme® RM IM). ................................................................................................ 80
Figura 17: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de peixe antes e após a reação de acidólise com o ácido octanóico (C8:0) catalisado pelas lipases de Thermomyces lanuginosus (Lipozyme® TL IM e Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e de Rhizomucor miehei (Lipozyme® RM IM). ...................... 81

ÍNDICE DE TABELAS
Tabela 3: Quantificação da atividade de esterificação dos biocatalisadores comerciais aplicados no presente estudo........................................................................................................................................ 49
Tabela 4: Quantificação da atividade de esterificação dos biocatalisadores não comerciaisaplicados no presente estudo........................................................................................................................................ 49
Tabela 5: Potencial catalítico para hidrólise do acetone powder da semente germinada de macaúba e de buriti. ......................................................................................................................................................... 51
Tabela 6: Análise do perfil das famílias de ácidos graxos presentes no óleo de coco antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases e análise da presença de EPA e DHA e da razão ω-6/ω-3. ........................ 57
Tabela 8: Análise do perfil das famílias de ácidos graxos presentes no óleo de palma antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases e análise da presença de EPA e DHA e da razão ω-6/ω-3. ........................ 64
Tabela 10: Análise do perfil das famílias de ácidos graxos presentes no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases e análise da presença de EPA e DHA e da razão ω-6/ω-3. ........................ 70
Tabela 12: Análise da incorporação de EPA+DHA nos óleos vegetais após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases. Os valores em miligramas foram estipulados com base na ingestão diária de uma colher de sopa (8,0g). ........................................................................................................................................................ 73
Tabela 13: Análise do perfil em ácidos graxos da acidólise da tripalmitina catalisada por diversas lipases. ....................................................................................................................................................... 75
Tabela 14: Análise do perfil das famílias de ácidos graxos presentes no óleo de peixe antes e após a reação de acidólise com o ácido octanóico (C8:0) catalisado por diferentes lipases e análise da presença dos ácidos octanóico, EPA e DHA. .................................................................................... 81

ix
Sumário
RESUMO iv ABSTRACT v ÍNDICE DE FIGURAS vi ÍNDICE DE TABELAS viii 1. INTRODUÇÃO E JUSTIFICATIVA 1 2. OBJETIVO 3 2.1. Objetivo Geral 3 2.2. Objetivos Específicos 3 3. REVISÃO BIBLIOGRÁFICA 2 3.1. Avaliação do Mercado de Enzimas 2 3.1.1. Mercado de Enzimas Industriais 2
3.2. Lipases (E.C.3.1.1.3) 3 3.3. Lipases Vegetais 8 3.4. Lipases Microbianas 10 3.5. Aplicação de Lipases na Indústria Alimentícia e Farmacêutica 10 3.6. Lipídios e Nutrição 12 3.7. Metabolismo e Absorção dos Lipídios 14 3.8. Lipídios Estruturados 17 3.9. Reações para Obtenção dos Lipídios Estruturados 18 3.10. Alimentos Funcionais / Nutracêuticos / Bioativos 19 3.11. Aplicações Específicas 21 3.11.1. Enriquecimento Nutricional de Óleos Vegetais Comestíveis com Ácidos Graxos Poli-insaturados da Família ω-3. 22 3.11.1.1. Mercado de Produtos Nutracêuticos 22 3.11.2. Lipídios Estruturados de Baixo Teor Calórico 23 3.11.2.1. Mercado de Produtos para Perda de Peso 24 3.11.3. Produção de Substitutos da Gordura do Leite Humano 24 3.11.3.1. Mercado de Fórmulas Infantis 26
3.12. Óleos Vegetais 27 3.12.1. Óleo de coco (Cocos nucifera L.) 28 3.12.1.1. Características Gerais 28 3.12.1.2. Benefícios Funcionais / Usos Terapêuticos 28 3.12.2. Óleo de Palma (Elaeis guineenses Jacq.) 29 3.12.2.1. Características Gerais 29 3.12.2.2. Benefícios Funcionais / Usos Terapêuticos 30 3.12.3. Óleo de Oliva (Olea europaea) 30 3.12.3.1. Características Gerais 30 3.12.3.2. Benefícios Funcionais / Usos Terapêuticos 31 3.13. Óleo de Peixe 31 3.13.1. Características Gerais 31 3.14. Dose Diária Recomendada de Ácidos Graxos Poli-insaturados da Família ω-3 36 3.15. Ingestão Diária Recomendada de Ácidos Graxos Ômega-6 / Ômega-3 na Dieta 36
4. MATERIAL E MÉTODOS 38 4.1. MATERIAIS 38 4.1.1. Sementes 38 4.1.2. Preparo dos Biocatalisadores Vegetais a partir das Sementes Germinadas 38

x
4.1.3. Lipases Microbianas 38 4.1.4. Óleos Vegetais e de Peixe 40
4.2. MÉTODOS ANALÍTICOS 40 4.2.1. Reação de Hidrólise com o Preparado Enzimático Vegetal (PEV) 40 4.2.2. Reação de Hidrólise com o Acetone Powder 40 4.2.3. Reação de Esterificação 41 4.2.4. Determinação da Acidez nos Óleos 41 4.2.5. Determinação da Atividade de Esterificação 41 4.2.6. Reação de Hidrólise do Óleo de Peixe 42 4.2.7. Concentração dos Ácidos Graxos Poli-insaturados do Óleo de Peixe Hidrolisado com Uréia 42 4.2.8. Reação de Acidólise dos Óleos Vegetais com os Ácidos Graxos Poli-insaturados 43 4.2.8. Reação de Acidólise da Tripalmitina com Ácido Oléico para Produção de Miméticos do Leite Materno 44 4.2.9. Reação de Acidólise do Óleo de Peixe com Ácido Octanóico para Produção de Lipídios de Baixo Teor Calórico 44 4.2.10. Separação do Produto por Cromatografia em Camada Delgada (TLC) 44 4.2.11. Metilação dos Ácidos Graxos para Análise por Cromatografia Gasosa 45 4.2.12. Análise da Composição em Ácidos Graxos por Cromatografia Gasosa 45
5. RESULTADOS E DISCUSSÃO 46 5.1. Análise do Perfil em Ácidos Graxos das Materias Primas Oleosas Utilizadas 46 5.2 Quantificação da Atividade de Esterificação dos Biocatalisadores 48 5.3. Avaliação do Potencial dos Biocatalisadores Vegetais 50 5.3.1. Macaúba (Acrocomia aculeata) e Buriti (Mauritia flexuosa L.) 50 5.3.1.1. Germinação das Sementes de Macaúba e Buriti 50 5.3.1.2. Avaliação do Potencial Hidrolítico e de Esterificação 50 5.3.2. Girassol (Helianthus annuus), Gergelim (Sesamum indicum) e Amendoim (Arachis hypogaea L.) 52 5.3.2.1. Germinação das Sementes de Girassol/Gergelim/Amendoim 52 5.3.2.2. Avaliação do Potencial Hidrolítico e de Esterificação 52
5.4. Enriquecimento Nutricional de Óleos Vegetais com Ácidos Graxos Poli-insaturados 53 5.4.4. Seleção de Lipases para o Enriquecimento Nutricional do Óleo de Coco (Cocos nucifera L.) 54 5.4.5. Enriquecimento Nutricional do Óleo de Palma (Elaeis guineenses Jacq.) 61 62 63 5.4.6. Enriquecimento Nutricional do Óleo de Oliva (Olea europaea) 67 5.4.7. Avaliação da ingestão da dose diária recomendada de EPA+DHA 72
5.5. Avaliação da Produção de Miméticos do Leite Materno 74 5.6. Produção de Lipídios Estruturados M-L-M de Baixo Teor Calórico a partir da Acidólise do Óleo de Peixe com Ácido Graxo de Cadeia Média 79 6. CONCLUSÕES 86 6.1. Avaliação do Potencial dos Biocatalisadores Vegetais 86 6.2. Enriquecimento Nutricional de Óleos Vegetais com Ácidos Graxos Poli-insaturados 86 6.3. Avaliação da Produção de Miméticos do Leite Materno 87 6.4. Produção de Lipídios Estruturados M-L-M de Baixo Teor Calórico. 87 7. PERSPECTIVAS / TRABALHOS FUTUROS 88 8. CONSIDERAÇÕES FINAIS 88 9. REFERENCIAS BIBLIOGRÁFICAS 90

1
1. INTRODUÇÃO e JUSTIFICATIVA
A versatilidade catalítica das lipases resulta em vantagens importantes sobre os catalisadores
químicos clássicos, tais como: especificidade pelo substrato, regiosseletividade e enanciosseletividade,
que permitem a catálise de reações com um número reduzido de coprodutos, menor geração de
efluentes e condições mais brandas de temperatura e pressão, apresentando assim um vasto campo
de aplicação tecnológica (LEE et al, 2006; WU et al, 2006; FREIRE e CASTILHO, 2008). As lipases
são glicerol éster hidrolases (E.C. 3.1.1.3) que constituem um grupo de enzimas inicialmente descritas
pela capacidade de catalisar a hidrólise de ligações éster de óleos (triacilgliceróis) gerando ácidos
graxos livres e glicerol. Geralmente, as lipases apresentam alguma seletividade particular, e a maioria
delas apresenta especificidade pelo substrato, podendo ser empregada de forma vantajosa para o
enriquecimento específico ou isolamento de um tipo ou de uma classe de lipídio.
Atualmente, as lipases respondem por cerca de 5% do mercado mundial de enzimas;
entretanto, existe uma forte tendência de crescimento por conta do seu vasto campo de aplicação. O
aumento da utilização de enzimas é completamente justificável pelo fato de serem naturais e
biodegradáveis. Além disso, atualmente busca-se não só processos industriais mais eficientes, mas
também conscientes, que não gerem resíduos e onde os catalisadores possam ser reutilizados. Com
isso, os processos de fabricação estão sendo reconstruídos em processos ambientalmente mais
amigáveis, pela substituição de produtos químicos pelo uso de enzimas. Neste contexto, a tecnologia
da catálise enzimática se insere e vem ganhando importância no cenário mundial.
Uma forma de aplicação das lipases de interesse tecnológico para a indústria de alimentos se
dá na síntese de lipídios estruturados, que são triacilgliceróis re-estruturados ou modificados por
meio da interesterificação química ou enzimática, com a finalidade de alterar sua composição em
ácidos graxos e/ou sua distribuição na molécula de glicerol (LEE e AKOH, 1998).
Com o aumento do conhecimento sobre os efeitos de ácidos graxos específicos no
metabolismo humano, principalmente dos poli-insaturados, há um crescente interesse no uso de
óleos e gorduras para o tratamento e prevenção de diversas doenças crônicas. Sob este ponto de
vista, os lipídios estruturados podem ser considerados como alimentos funcionais, que são alimentos
ou ingredientes de alimentos que podem proporcionar um efeito benéfico para a saúde, além dos
nutrientes básicos que eles contêm, visando doenças específicas ou condições metabólicas anormais
(YANKAH e AKOH, 2000).
O mercado mundial apresenta uma ampla variedade de suplementos alimentares de ácidos
graxos poli-insaturados, principalmente das famílias ômega-3 (ω-3) e ômega-6 (ω-6) e de produtos,

2
nos quais estes ácidos graxos são incorporados com a finalidade não só de complementar a ingestão
diária recomendada destes ácidos graxos (fins nutritivos); como também, para uso clínico (MU; XU;
HOY, 1998). Com isso, os lipídios estruturados podem propiciar o meio mais efetivo de fornecer
ácidos graxos desejados para fins nutritivos ou terapêuticos, visando doenças específicas ou
condições metabólicas anormais (D’AGOSTINI et al, 2001). Também podem ser sintetizados para
melhorar ou alterar as características físicas e/ou químicas dos triacilgliceróis, tais como ponto de
fusão, conteúdo de gordura sólida, viscosidade, consistência, índices de iodo e de saponificação,
sendo que alguns deles também apresentam menor valor calórico (GUNSTONE, 1998,
AUERBACH; KLEMANN; HEYDINGER, 2001).
Embora a indústria de produtos nutracêuticos seja inferior a 10% da indústria alimentar total,
o mercado oferece oportunidades significativas de crescimento e margens de lucro mais amplas, uma
vez que o mercado mundial de nutracêuticos foi estimado em cerca de US$ 151 bilhões em 2011.
Neste ramo a previsão é que se chegue a US$ 207 bilhões em 2016, com crescimento anual de 6,5%
(BCC RESEARCH, 2011a).
Na reação de interesterificação, o triacilglicerol reage com um ácido graxo livre (acidólise) ou
um álcool (alcoólise), resultando em rearranjo na distribuição dos ácidos graxos do triacilglicerol
formando um novo triacilglicerol, com propriedades químicas e físicas diversas (AKOH, 1995). Os
produtos resultantes da reação de interesterificação de uma mistura de triacilgliceróis ou entre um
triacilglicerol e ácidos graxos livres, catalisada por lipases regioespecíficas para as posições sn-1,3,
permite que o processo seja dirigido para a formação de produtos com composição e propriedades
peculiares. Portanto, o uso de lipases específicas propicia a formação de produtos que não podem ser
obtidos pelos métodos de interesterificação química, pois os catalisadores químicos não são
específicos.
Com base nisso, lipídios estruturados têm grande potencial para agregar valor a alimentos
conhecidos e possivelmente contribuir no desenvolvimento de novos produtos alimentícios com
valor nutracêutico.
O maior desafio encontrado na substituição dos catalisadores químicos pelas lipases na
indústria alimentícia reside no elevado custo de produção, imobilização e da menor estabilidade das
mesmas. Além disso, as lipases comerciais disponíveis no mercado são importadas e de elevado
custo, o que, na maioria das vezes, inviabiliza sua utilização em maiores escalas ou para produção de
comodities de menor valor agregado.
Neste contexto, o presente trabalho tem como objetivo investigar do potencial de aplicação
de lipases comerciais e home-made, de origens microbiana e vegetal na produção de lipídios de
interesse nutricional e farmacêutico.

3
2. OBJETIVO
2.1. Objetivo Geral
Aplicar lipases microbianas e vegetais para produzir lipídios funcionais e de interesse
nutracêutico, enriquecidos com ácidos graxos poli-insaturados de cadeia longa; óleo de peixe
modificado para a obtenção de lipídios com menor densidade energética e a produção de um
substituto dos lipídios do leite materno.
2.2. Objetivos Específicos
2.2.1. Investigar novas lipases vegetais obtidas a partir de sementes germinadas, avaliando o
potencial catalítico desses biocatalisadores em reações de hidrólise e esterificação;
2.2.2. Obter lipídios modificados de interesse nutricional e farmacêutico, utilizando lipases
comerciais e não comerciais;
2.2.2.1. Óleos vegetais, para fins alimentícios, enriquecidos com ácidos graxos poli-
insaturados da família ω-3;
2.2.2.2. Produzir um lipídio de menor teor calórico, tendo como matéria-prima o óleo
de peixe;
2.2.2.3 Obter um lipídio que seja mimético do leite materno;
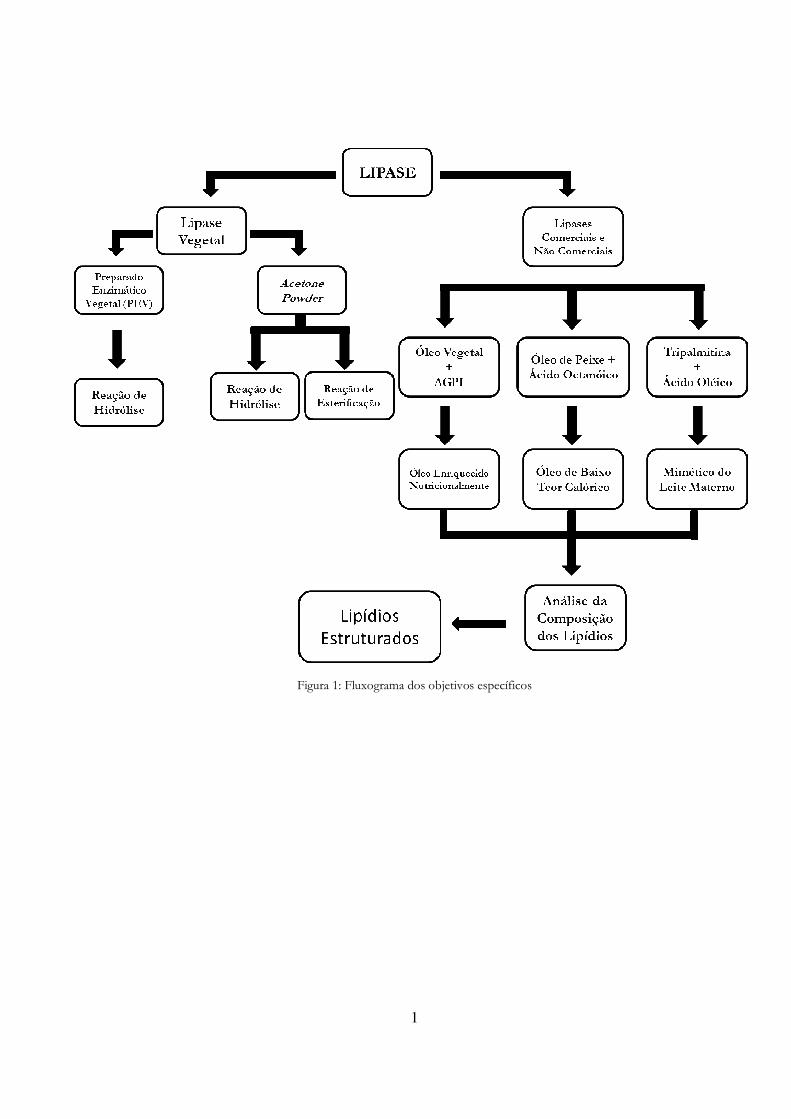
1
Figura 1: Fluxograma dos objetivos específicos

2
3. REVISÃO BIBLIOGRÁFICA
3.1. Avaliação do Mercado de Enzimas
O mercado mundial de enzimas industriais é muito competitivo, tendo a Novozymes como a
empresa líder de mercado, seguida por DSM, e DuPont (depois adquiriu uma participação
majoritária na Danisco e sua divisão Genencor), entre outras.
As empresas competem principalmente com base na qualidade do produto, seu desempenho,
pelo uso de direitos de propriedade intelectual, bem como a capacidade de inovar, entre outros
fatores. América do Norte e Europa são os maiores consumidores de enzimas industriais, embora a
região da Ásia-Pacífico esteja passando por um crescimento acentuado devido à demanda gerada pela
China, Japão e Índia, refletindo o tamanho da força das economias destes países nos dias atuais
(ADRIO e DEMAIN, 2014).
Com o aumento da conscientização sobre questões ambientais e de custos, a biotecnologia
está ganhando terreno rapidamente devido a diversas vantagens que ela oferece em relação às
tecnologias convencionais. O campo de enzimas industriais está agora a experimentar grandes
iniciativas de P & D, resultando tanto no desenvolvimento de uma série de novos produtos quanto
na melhoria do processo e desempenho de vários produtos existentes.
Com base no que foi explicitado, este tópico apresentará um ponto de vista mais arrojado e
atual, pela análise da evolução recente e futura do mercado, através da observação dos produtos
nutricionais. Este tópico descreve a indústria, sua relação com o setor de saúde, os seus participantes,
e as tendências para o futuro. Além disso, fornece uma visão geral da nutrição como um todo,
definindo produtos dentro de cada mercado, e os detalhes do tamanho do mercado.
Todos os mercados são subdivididos em tipos específicos de produtos e cada subsegmento
será mostrado mais detalhadamente, com a identificação do tamanho do mercado atual e a previsão
em curto prazo para alguns deles.
3.1.1. Mercado de Enzimas Industriais
O principal objetivo deste segmento é o de apresentar uma análise abrangente do mercado
atual e sua direção futura no mercado de enzimas como uma importante ferramenta para aumentar a
eficiência e especificidade dos produtos em que as enzimas são usadas, que poderão incluir, por
exemplo, os setores alimentícios e de bebidas, detergentes e derivados e de ração animal.

3
O aumento da utilização de enzimas é completamente justificável pelo fato de serem naturais
e biodegradáveis. Com isso, os processos de fabricação estão sendo reconstruídos em um processo
ambientalmente mais amigável, com condições menos corrosivas, devido a substituição de alguns
produtos químicos pelo uso de enzimas biodegradáveis. Esta “compatibilidade” da enzima com o
meio ambiente associada a sua capacidade de aumentar a eficiência em alguns processos, ajudam a
alavancar ainda mais o seu uso.
O mercado mundial de enzimas industriais atingiu a marca de US$ 3,6 bilhões em 2010, e em
2011, quando chegou a cerca de US$ 3,9 bilhões, foi feita uma projeção de crescimento anual de
9,1%, aonde se chegaria a US$ 6 bilhões até 2016 (BCC RESEARCH, 2012a).
Enzimas de alimentos e bebidas compõem o maior segmento da indústria de enzimas
industriais, com faturamento de cerca de US$ 1,2 bilhões em 2010. Neste ramo a previsão é que se
chegue a US$ 2,1 bilhões em 2016, com crescimento anual de 10,4% (BCC RESEARCH, 2012a).
Também devemos levar em consideração, o mercado global de nutrição ética, que foi de US$
26 bilhões em 2010. Nutrição ética é a prática de fornecer os nutrientes adequados para uma ampla
gama de indivíduos, incluindo bebês, crianças, adultos e pessoas com enfermidades e injúrias. O
mercado de nutrição ética surgiu de uma necessidade de proporcionar aos indivíduos uma nutrição
mais adequada, tanto em hospitais quanto em casa. Já em 2015, prevê-se que as receitas poderão
atingir quase US$ 44 bilhões, com previsão de expansão nos próximos cinco anos a uma taxa de
crescimento anual de 10,9% (BCC RESEARCH, 2011d).
O mercado de nutrição infantil é o maior segmento do mercado de nutrição ética, com
vendas de US$ 12 bilhões em 2010, com previsão de aumento para US$ 23 bilhões até 2015, com
uma taxa de crescimento anual de 13,2%. Os produtos de nutrição enteral, que atingiram a marca de
US $ 9,7 bilhões em 2010, tem uma previsão de mercado em torno de US$ 15,6 bilhões em 2015,
com uma taxa de crescimento anual de 9,9% (BCC RESEARCH, 2011d).
3.2. Lipases (E.C.3.1.1.3)
As enzimas são classificadas e codificadas pela NC-IUBMB (Nomenclature Committee of the
International Union of Biochemistry and Molecular Biology) de acordo com a reação catalisada. A
nomenclatura utiliza a abreviação de E.C. (Enzyme Comission) seguida de até quatro dígitos referentes
à classe e subclasses a que pertencem. Lipase é um nome genérico para um grupo de enzimas
pertencentes à classe das hidrolases (E.C.3.1) e que atuam sobre as ligações éster (E.C.3.1.1). As
lipases constituem um grupo de enzimas inicialmente descritas pela capacidade de catalisar a hidrólise
de ligações éster de óleos (triacilgliceróis) gerando ácidos graxos livres e glicerol. Por isso, foram

4
definidas como glicerol éster hidrolases (E.C. 3.1.1.3). Hoje em dia, sabe-se que são capazes de
hidrolisar também outros ésteres de ácidos graxos (FREIRE e CASTILHO, 2000).
Atualmente, as lipases respondem por cerca de 5% do mercado mundial de enzimas;
entretanto, existe uma forte tendência de crescimento por conta do seu vasto campo de aplicação.
Além disso, atualmente busca-se não só processos industriais mais eficientes, mas também
conscientes, que não gerem resíduos e onde os catalisadores possam ser reutilizados. Com isso, os
processos de fabricação estão sendo reconstruídos em processos ambientalmente mais amigáveis,
pela substituição de produtos químicos pelo uso de enzimas. Neste contexto, a tecnologia da catálise
enzimática se insere e vem ganhando importância no cenário mundial.
A utilização de enzimas como catalisadores tem uma série de vantagens em relação aos
métodos convencionais de síntese orgânica. As reações podem ocorrer a temperaturas moderadas e
dessa forma, há uma menor degradação tanto de substratos, como dos produtos formados, além da
reação ser mais específica (SONNET, 1988). Alguns produtos fabricados industrialmente a partir de
gorduras e óleos por meio de processos químicos podem ser gerados por lipases com maior rapidez e
melhor especificidade, sob condições amenas (SIH e WU, 1989). Além disso, a quimio-, régio- e
enantioespecífica de algumas enzimas desperta um enorme interesse para sua aplicação em métodos
industriais (SAXENA; SHEORAN; DAVIDSON, 2003).
O catalisador “ideal” é considerado bioquimicamente em termos de número de turnover
(kcat) ou, para um determinado processo, em termos de especificidade constante máxima
(kcat/KM). No entanto, do ponto de vista de bioprocessos, cada bioprocessos é limitado por um
conjunto de condições ditadas pelas propriedades específicas dos substratos, produtos e a reação de
bioconversão (BURTON; COWAN; WOODLEY, 2002). PRA QUE ISSO?
Muitas enzimas são ativas em interfaces de sistemas biológicos vivos, tais como nos
processos de sinalização na superfície das membranas celulares, na digestão dos lipídios na dieta,
degradação do amido e celulose, etc; mas o estudo da enzimologia permanece em grande parte
voltada para as interações entre enzimas e substratos solúveis. A caracterização bioquímica e cinética
de enzimas lipolíticas abriu novos caminhos de pesquisa na área de enzimologia interfacial
(ABDELKAFI et al, 2011).
As lipases pertencem à família de serina-hidrolases que podem catalisar tanto a reação de
hidrólise de ésteres carboxílicos em água; quanto a reação inversa, a acilação de álcoois com ácidos
carboxílicos, na presença de solventes (SCHMID e VERGER, 1998). Elas têm em comum uma α/ß
hidrolase e um sítio ativo com a tríade catalítica formada por serina-histidina-ácido
aspártico/glutâmico e a cavidade do oxiânion (OLLIS et al, 1992).

5
A água tem um efeito duplo em sistemas enzimáticos, pois ela é essencial para a formação e
manutenção da conformação cataliticamente ativa das enzimas (SCHMID e VERGER, 1998;
FICKERS; DESTAIN; THONART, 2008) e também se faz necessária na maioria dos processos de
inativação de enzimas, em particular as de inativação térmica (OLLIS et al, 1992). Várias enzimas
mostram-se eficientes em meios orgânicos lipofílicos na presença de uma pequena percentagem de
água (FISCHER e PLEISS, 2003).
Em meio aquoso, o sítio ativo das lipases é coberto por um uma estrutura móvel denominada
de tampa, que se abre quando a lipase se encontra em uma interface hidrofóbica (BRADY et al,
1990; SOUMANOU; PÉRIGNON; VILLENEUVE, 2013). Esta estrutura pode ser elucidada pela
determinação da estrutura 3D de algumas lipases. Esta mudança de estrutura proporciona acesso
livre a tríade catalítica que anteriormente estava encoberta na estrutura tridimensional da enzima e,
portanto, permite um encaixe entre o local do sítio ativo e o substrato (SOUMANOU;
PÉRIGNON; VILLENEUVE, 2013).
A atividade catalítica da lipase aumenta dramaticamente na interface. Este fenômeno é
conhecido como ativação interfacial, que corresponde à abertura da tampa, quando a lipase se liga à
interface (BROCKMAN, 1984; GILL e PARISH, 1997; SOUMANOU; PÉRIGNON;
VILLENEUVE, 2013).
Em 1958, Sarda e Desnuelle definiram as lipases em termos cinéticos, com base no
fenômeno da “ativação interfacial”. Quando esta propriedade não era observada em certas enzimas,
as mesmas eram classificadas como esterases, ou seja, aqueles que agem somente em moléculas de
ésteres carboxílicos que são solúveis em água (VERGER, 1998). Isto equivale ao fato de a atividade
das lipases serem melhores em substratos insolúveis (tais como emulsões) em comparação com as
soluções verdadeiras monoméricas. A partir destes estudos foi proposto que lipases podem ser uma
categoria especial de esterases que são altamente eficientes na hidrólise de moléculas que possuem
um grupo éster carboxílico, e que são agregados em água. Esta propriedade foi utilizada durante
muito tempo para distinguir entre as lipases e esterases (VERGER, 1998).
As enzimas que catalisam a hidrólise de triacilgliceróis têm sido estudadas há mais de 300
anos e a capacidade de as lipases para catalisar a hidrólise e também a síntese de ésteres foi
reconhecida a quase 70 anos (VAN DER WALLE, 1927).
A especificidade das lipases pelos ácidos graxos é afetada pelo comprimento da cadeia
carbônica, bem como o número e posição das insaturações (LONG, 2010). O elevado grau de
especificidade e enantiosseletividade para as reações de esterificação e transesterificação, as tornam
um excelente biocatalisador para as reações de trans e interesterificação (OKAHATA; FUJIMOTO;
IJIRO, 1995; SCHMID e VERGER, 1998).

6
A especificidade posicional (regiosseletividade) de lipases também é muito importante para o
processamento de matrizes oleosas. As lipases são classificadas em dois grandes grupos, onde a
primeira é uma enzima 1,3 específica, por reconhecer as ligações éster na posição sn-1,3 do
triacilglicerol (TAG); e a segunda, é uma enzima não específica, ou seja, atua sobre todas as ligações
éster do TAG.
A especificidade pelos álcoois também é importante no campo das reações oleoquímicas.
Geralmente, os álcoois primários são os melhores nucleófilos enquanto álcoois secundários são
menos reativos e por sua vez, os álcoois terciários são os menos ainda. Zaks e Klibanov (1988)
mostraram que as lipases podem ser ativas até mesmo em solventes orgânicos anidros. Além do que,
a seletividade das lipases pode ser afetada pelos parâmetros do meio de reação, isto é, a polaridade do
solvente ou log P (coeficiente hidrofóbico-hidrofílico), atividade da água (aw), a imobilização, etc
(CATONI; CERNIA; PALOCCI, 1996).
Considerando todas as vantagens anteriormente descritas, as lipases podem ser aplicadas em
uma infinidade de processos, sendo algumas delas bastante promissoras, tais como: síntese orgânica,
formulações de detergentes, síntese de biossurfactantes, indústria oleoquímica, biocombustíveis,
indústria de laticínios, indústria têxtil, indústria de agroquímicos, polímeros, fabricação de papel,
nutrição, cosméticos, farmacêutica, lipídios funcionais etc (LIESE; SEELBACH; WANDREY, 2000;
LONG, 2010; JAEGER e EGGERT, 2002). As duas últimas aplicações são as mais desafiadoras e
populares no momento, devido ao fato delas ajudarem a melhorar a saúde humana e por ser um
mercado de bilhões de dólares (LONG, 2010).
As lipases têm sido isoladas a partir de várias fontes: bacterias (45%), fungos (21%), animais
(18%), plantas (11%), e algas (3%) (PATIL; CHOPDA; MAHAJAN, 2011); e um número cada vez
maior de lipases com propriedades específicas está se tornando disponível para comercialização
(LIESE; SEELBACH; WANDREY, 2000). Somado a isso, o aumento da disponibilidade e
estabilidade das lipases comercialmente disponíveis tem resultado num aumento do interesse nas
aplicações potenciais de biocatálise de lipase (KUMAR e KANWAR, 2011). Também são
reconhecidas pela sua função fisiológica na digestão e absorção de lipídios. Estas propriedades são
agora utilizadas em várias aplicações biotecnológicas.
As lipases catalisam a hidrólise de uma ampla variedade de gorduras e óleos (triacilgliceróis)
para formar diacilgliceróis, monoacilgliceróis, ácidos graxos livres e glicerol (Figura 3). Como as
lipases atuam em meio aquoso, a reação de hidrólise dos substratos oleosos, que são insolúveis em
água, ocorre na interface lipídio/água (BORNSCHEUER et al, 2002). Este processo catalítico pode
ser descrito em termos de uma etapa de adsorção de lipase que ocorre na interface óleo/água,

7
seguindo-se a formação de um complexo enzima/substrato e a liberação de produtos de lipólise
(VERGER; MIERAS; HAAS, 1973).
A presença necessária de água em reações enzimáticas foi investigado por Zaks e Klibanov
(1986) que, a partir de seus estudos concluiu que as enzimas necessitam de uma fina camada de água
na sua superfície para reter a sua conformação cataliticamente ativa em ambientes anidros
(SOUMANOU; PÉRIGNON; VILLENEUVE, 2013). Uma das grandes vantagens da utilização de
meio anidro para biotransformações é que as reações que são termodinamicamente desfavoráveis em
água (como transesterificação e aminólise) agora podem ser realizadas com sucesso. Além disso, a
enantiosseletividade pode ser aumentada muitas vezes em meios não aquosos, em comparação com a
fase aquosa (SAKURAI et al, 1988). As desvantagens incluem a insolubilidade de certas enzimas e
substratos, a utilização de solventes não amigáveis ao meio ambiente, tais como diclorometano e uma
possível perda de estereoquímica (GILL e PARISH, 1997).
Os sistemas sem solventes são adequados para produtos alimentares, pois a ausência do
solvente facilita o processamento, reduz custos e não interfere na pureza dos produtos. No entanto,
a alta viscosidade dos substratos no meio reacional pode resultar em uma má homogeneização e uma
taxa de reação mais lenta.
Vários tipos de reações catalisadas por lipases têm sido aplicados à síntese: por exemplo, a
hidrólise do éster, esterificação, transesterificação, interesterificação, e transferência de grupos acila
entre os ésteres de outros nucleófilos tais como aminas, tióis e hidroperóxidos (GILL e PARISH,
1997).
A estabilidade das lipases também é afetada pela qualidade de lipídios. Verificou-se que os
compostos menores encontrados em óleos e gorduras, tais como hidroperóxidos, emulsificantes,
fosfolipídios, clorofila, carotenóides, polímeros, íons metálicos, tem efeitos sobre a estabilidade de
lipases (XU e HØY, 1998). No entanto, o refino de óleos e gorduras é um processo dispendioso
(XU, 2000).

8
Figura 2: Representação das reações de transesterificação, acidólise e alcoólise catalisada por lipases na formação de diacilgliceróis, monoacilgliceróis, ácidos graxos livres e glicerol.
Há muitos fatores que influenciam o interesse crescente na biocatálise, os quais incluem a
versatilidade da enzima, métodos computacionais combinados com evolução dirigida e o
desenvolvimento de tecnologias para melhorar as propriedades de enzimas para atender as
perspectivas dos processos (KUMAR e SINGH, 2013). Com isso, os próximos anos devem ser
promissores no desenvolvimento de novas enzimas área de biocatálise industrial.
3.3. Lipases Vegetais
As lipases de plantas tem atraído a atenção devido ao seu baixo custo, elevada especificidade
para alguns substratos e o fato de que elas estão amplamente disponíveis a partir de fontes naturais,
sem a necessidade de tecnologia genética molecular para produzí-las. Isso as torna uma boa
alternativa para exploração comercial como enzimas industriais (VILLENEUVE, 2003).
Estas enzimas têm sido isoladas a partir de folhas, caules, látex e de sementes de oleaginosas
e cereais (VILLENEUVE, 2003; EASTMOND, 2004; YOU et al, 2011; AVELAR et al, 2013).
Lipases de látex e de sementes de plantas oleaginosas são usados principalmente em
Acidólise
Transesterificação
Alcoólise

9
biotransformações de óleos e gorduras (VILLENEUVE, 2003). Em alguns casos, tem sido relatado
que a atividade da lipase está ausente em sementes não germinadas (ou latentes/dormentes) e
aumenta rapidamente quando começa germinação, no entanto, em alguns casos, a atividade lipolítica
foi encontrada em sementes dormentes (VILLENEUVE, 2003; EASTMOND, 2004; SOUSA et al,
2010; CAVALCANTI et al, 2007; HUANG, 1984). Como exemplo temos os casos em que a enzima
é ativa nas sementes dormentes, como nas sementes de mamona (Ricinus communis L.)
(CAVALCANTI et al, 2007; HUANG, 1984), amendoim (Arachis hypogaea L.) (SANDERS e
PATTEE, 1975) e a de pinhão manso (Jatropha curcas Lin.) (ABIGOR et al, 2002) parece ser inativa
em vivo, mas ativo in vitro. Em outras palavras, o tempo que a semente está no estado intacto, a
enzima permanece inativa, mas qualquer ligeira alteração na semente ou condições de armazenagem,
provavelmente irá iniciar a atividade lipásica.
A aplicação direta de extratos brutos de oleaginosas é economicamente atraente para a
indústria oleoquímica, devido à sua preparação fácil e de baixo custo, pois as etapas purificação e
imobilização não são necessárias porque as moléculas de enzimas são naturalmente "imobilizadas"
nos materiais sólidos provenientes de sementes, o que permite uma fácil recuperação do
biocatalisador para usos sequenciais (YOU et al, 2011; SOUSA et al, 2010; CAVALCANTI et al,
2007).
Preparações em pó, na forma de preparado acetônico (acetone powder), com atividade lipásica
obtidas a partir de semente de oleaginosas como a mamona (Ricinus communis L.) colza (Brassica napus),
pinhão manso (Jatropha curcas Lin.) e Nigella sativa foram utilizadas como biocatalisadores para
hidrólise e esterificação de lipídios e esterificação de ácido oléico com metanol e glicerol (BARON et
al, 1999; SOUSA et al, 2010; CAVALCANTI et al, 2007; MUKHERJEE, 1996; AFOLABI et al,
1991; DANDIK; ARIOGLU; AKSOY, 1993).
Lipases de oleaginosas têm grande potencial para exploração comercial, especialmente as
sementes oleaginosas que são atualmente consideradas subutilizadas.
A vantagem de utilizar lipases vegetais em indústrias alimentares, em comparação com outras
fontes de lipase é a sua aceitabilidade, em comparação com as lipases microbianas (SETH et al,
2014). Uma das lipases vegetais mais estudadas é a lipase de Carica papaya. Ela tem sido utilizada com
sucesso na síntese de lipídios de baixo teor calórico para fórmulas infantis (MANGOS; JONES;
FOGLIA, 1999), na produção de triacilgliceróis estruturados (GANDHI e MUKHERJEE, 2001), na
interesterificação de óleo de palma para a síntese de equivalente de manteiga de cacau
(PINYAPHONG e PHUTRAKUL, 2009), etc.

10
3.4. Lipases Microbianas
Os micro-organismos podem fornecer uma quantidade impressionante de catalisadores com
uma ampla gama de aplicações em diversos ramos industriais, tais como: indústria alimentícia, ração
animal, química fina e farmacêutica, etc. As propriedades singulares das enzimas, tais como a
especificidade e biodegradabilidade, permitem que se tenham processos industriais viáveis sob
condições mais brandas de temperatura e pressão, obtendo melhores rendimentos e com a redução
da geração de resíduos (KUMAR e SINGH, 2013). COLOCAR EM LIPASE GERAL
Enzimas microbianas são conhecidas por desempenhar um papel crucial como catalisadores
metabólicos, levando a sua utilização em diversas indústrias e aplicações, onde mais de 500 produtos
industriais estão sendo feitos com a utilização de enzimas microbianas (CHERRY e FIDANTSEF,
2003).
Lipases microbianas constituem um importante grupo de enzimas biotecnologicamente
valiosas, principalmente por causa da versatilidade das suas propriedades e facilidade de produção em
massa. São amplamente diversificadas em suas propriedades enzimáticas e especificidade de
substrato.
As enzimas microbianas têm algumas vantagens em relação às enzimas derivadas de plantas
ou animais por causa da grande variedade de atividades catalíticas disponíveis, facilidade de
manipulação genética, fornecimento regular devido à ausência de flutuações sazonais e rápido
crescimento de micro-organismos (WISEMAN, 1995).
O custo da produção da lipase microbiana depende basicamente dos custos da sua
fermentação e do processo de downstream (separação e purificação). Logo, o aumento da
produtividade e na estabilidade da enzima contribuirá para que se tenha um processo
economicamente mais viável (LONG, 2010).
3.5. Aplicação de Lipases na Indústria Alimentícia e Farmacêutica
Enzimas relacionadas à comidas e bebidas constituem o maior segmento de enzimas
industriais, onde se faturou cerca de US$ 1,2 bilhões em 2011, e estima-se que este mercado chegue a
US$ 1,8 bilhões até 2016 (WORLD ENZYMES, 2011).
Enzimas alimentares podem aumentar a digestibilidade dos nutrientes, levando a uma maior
eficiência na utilização dos alimentos. Além disso, podem degradar componentes que aparentemente
são indesejáveis, prejudiciais ou de pouco valor (CHOCT, 2006).

11
Lipases microbianas são utilizadas na obtenção de ácidos graxos poli-insaturados a partir de
lipídios animais e vegetais tais como óleo de savelha, atum e de borragem (Borago officinalis L.). Os
ácidos graxos poli-insaturados livres e os seus mono- e diglicerídeos são posteriormente utilizados na
produção de uma variedade de produtos farmacêuticos, incluindo hipocolesterolêmicos, anti-
inflamatórios, e trombolíticos (BELARBI; MOLINA; CHISTI, 2000). Devido aos seus efeitos
metabólicos, os ácidos graxos poli-insaturados são cada vez mais utilizados como produtos
farmacêuticos, nutracêuticos e outros aditivos alimentares (GILL e VALIVETY, 1997; BELARBI;
MOLINA; CHISTI, 2000). Muitos desses ácidos graxos poli-insaturados são essenciais na
composição de membranas lipídicas das células de todo o organismo.
Óleos vegetais modificados, como os triacilgliceróis estruturados, são importantes tanto
nutricionalmente quanto por suas propriedades físico-químicas. Isso faz com eles tenham um grande
potencial no mercado futuro. As lipases, que são régioespecíficas e as que apresentam especificidade
por ácidos graxos, são de grande importância neste tipo de modificação e devem ser exploradas cada
vez mais no design de óleos vegetais.
Óleos de baixo valor comercial são os alvos preferenciais para a síntese dos triacilgliceróis
estruturados de relevância nutricional na obtenção de substitutos da manteiga de cacau, lipídios de
baixo teor calórico, óleos enriquecidos em ácidos graxos poli-insaturados ou ricos em ácido linoleico
conjugado (CLA), etc. O valor nutricional, sensorial e as propriedades físicas de um triglicerídeo são
definidos pela sua composição em ácidos graxos, que podem ser alterados pela atuação ds lipases.
Desta forma, um lipídio que seja relativamente barato e menos desejável, pode ser modificado para
um de maior valor agregado (COLMAN e MACRAE, 1980; UNDURRAGA; MARKOVITS;
ERAZO, 2001).
As lipases também são adicionadas nos alimentos com o intuito de modificar a síntese e
degradação de ésteres de ácidos graxos de cadeia curta, uma vez que estes compostos são conhecidos
por influenciarem no sabor e aroma dos alimentos (MACEDO; LOZANO; PASTORE, 3003). Esta
característica é muito bem explorada pela indústria de laticínios pela sua atuação na hidrólise da
gordura de leite. As aplicações atuais incluem o realce de sabor dos queijos, a aceleração da
maturação de queijos e na lipólise de manteiga. Os ácidos graxos livres gerados pela ação de lipases
na gordura do leite são marcantes em alguns casos, principalmente queijos de pasta mole, onde
geram características de sabor específico em alguns produtos (HASAN; SHAH; HAMEED, 2006).
O controle preciso da concentração da lipase, pH, temperatura e na emulsão são necessários
para maximizar a produção de sabor e fragrância (MENDEZ e SALAS, 2001; OKANISHI;
SUZUKI; FURITA, 1996).

12
As lipases têm sido utilizadas como agentes terapêuticos no tratamento de perturbações
gastrointestinais, manifestações cutâneas de alergias no trato digestivo (MAUVERNAY;
LABOREUR; LABROUSSE, 1970).
Também são alvos de drogas importantes ou enzimas marcadoras no sector médico. Elas
podem ser utilizadas como ferramentas de diagnóstico e a sua presença ou os níveis crescentes
podem indicar determinada infecção ou doença (HASAN; SHAH; HAMEED, 2006).
As lipases podem ser utilizadas na resolução de misturas racêmicas e para sintetizar os blocos
de construção quirais para produtos farmacêuticos, agroquímicos e pesticidas. Podem ser utilizadas
na hidrólise de ésteres insolúveis em água, como por exemplo, na resolução de misturas racêmicas
através de hidrólise estereoespecífica (KIRCHNER; SCOLLAR; KLIBANOV, 1985; MANOEL et
al, 2012).
Dentre as principais vantagens da utilização de enzimas na síntese de compostos orgânicos,
podemos destacar a sua versatilidade, velocidade de reação elevadas, condições brandas de reação e
estereoespecificidade. Sendo que a estereoespecificidade é especialmente importante na síntese de
moléculas bioativas, pois geralmente apenas uma das formas enantioméricas manifesta bioatividade,
enquanto que a outra é muitas vezes é tóxica (IDING et al, 1998).
3.6. Lipídios e Nutrição
Não há um consenso entre as definições que são comumente aceitas pelos pesquisadores da
área. De certa forma, este problema é até compreensível, devido à complexidade e heterogeneidade
química apresentada por este grupo de moléculas. Por exemplo, Christie (1982) define lipídios como:
“uma variedade de produtos naturais, incluindo os ácidos graxos e seus derivados, esteróides,
terpenos, carotenos e os ácidos biliares, os quais têm em comum, a solubilidade em solventes
orgânicos tais como: éter dietílico, hexano, benzeno, clorofórmio ou metanol”. Para Kates (1986)
lipídios são “as substâncias que são: (i) insolúveis em água; (ii) solúveis em solventes orgânicos, tais
como clorofórmio, éter ou benzeno; (iii) contenham grupos de hidrocarbonetos de cadeia longa em
suas moléculas; e (iv) estão presentes ou são derivados de organismos vivos”. Já Gurr e James (1971)
apontam os lipídios como “um grupo quimicamente heterogêneo de substâncias, tendo em comum a
propriedade de ser insolúvel em água, mas solúvel em solventes apolares, tais como clorofórmio,
hidrocarbonetos ou álcoois”. Mais recentemente, tem sido proposta a sua origem (bio)sintética como
o fator que define os lipídios, que seriam compostos derivados química ou bioquimicamente de
ácidos graxos.

13
Algumas classificações podem ser feitas com base nas suas propriedades físicas. Dentre elas
que, em temperatura ambiente, os óleos são líquidos e gorduras são sólidas; com relação a
polaridade, os lipídios poder ser polares (por exemplo, glicerofosfolipídios e gliceroglicolipídios) ou
neutros (por exemplo, ácidos graxos, álcoois e esteróis) (AKOH e MIN, 2002).
Com base na estrutura, os lipídios podem ser classificados como: derivado, simples ou
complexo. Nos lipídios derivados estão inclusos os ácidos graxos e álcoois, que são os blocos de
construção para os lipídios simples e complexos. Já nos lipídios simples estão os acilgliceróis, éteres,
esteróis, cera, etc. Em termos gerais, os lipídios simples quando hidrolisados, geralmente liberam um
álcool e/ou ácido graxo. Os lipídios complexos englobam os fosfolipídios, glicolipídios e
esfingolipídios. Estas estruturas geram três ou mais compostos diferentes quando sofrem hidrólise
(AKOH e MIN, 2002).
Os triacilgliceróis são compostos por uma molécula de glicerol com três ácidos graxos
esterificados a ela (Figura 6a). Os ácidos graxos estão esterificados em posições distintas no esqueleto
carbônico do glicerol. Estas posições são representadas através de um sistema de numeração
estereoespecífica (sn): sn-1, sn-2 e sn-3. A sigla sn refere-se à posição dos átomos de carbono no
glicerol. O átomo de carbono que está na parte superior, pela projeção vertical de Fischer, com o
grupo hidroxila no carbono-2 orientado para a esquerda é chamado de sn-1 (posição exterior). sn-2
refere-se à posição no carbono-2 (posição intermédia) e sn-3 (posição exterior) refere-se à posição
abaixo sn-2 (Figura 6b) (SARAH e BERRY, 2009).
A posição e composição dos ácidos graxos na molécula de triacilglicerol determinam as
propriedades físicas dos óleos, que por consequência, afeta sua absorção, metabolismo e sua
Figura 3: (a) Representação da molécula de glicerol (propan-1,2,3-triol) com a identificação dos átomos de carbono, oxigênio e hidrogênio. (b) Estrutura geral de um triacilglicerol com as posições estereoespecíficas (sn-1, 2 ou 3) definidas.
Posição sn-3
Posição sn-1
Posição sn-2

14
distribuição nos tecidos. Todas essas mudanças têm implicações diretas na saúde humana. Assim
sendo, o designer de óleos obtidos por modificação posicional corrobora a importância dos lipídios
na dieta humana tanto para fins nutritivos quanto para terapêuticos.
O consumo de lipídios, apesar de essencial, pode oferecer tanto impactos positivos, quanto
negativos sobre a saúde humana. Os triacilgliceróis são uma importante fonte de energia na dieta
humana, mas o seu consumo excessivo tem sido associado a vários problemas de saúde, tais como
doenças coronarianas, obesidade, diabetes, enfarte e outras doenças. No entanto, o consumo de
determinados matrizes lipídicas com componentes bioativos, tais como vitaminas lipossolúveis,
fitoesteróis, carotenóides, antioxidantes e ácidos graxos poli-insaturados, são consideradas benéficas
para a saúde humana (CHEN; McCLEMENTS; DECKER, 2013). Nos dias atuais, há uma intensa
campanha para reduzir o consumo excessivo de óleos e gorduras, a fim de reduzir os problemas de
saúde supracitados.
É sabido que as propriedades funcionais de óleos e gorduras estão diretamente relacionadas
com o tipo de triacilgliceróis que eles contêm. As características físicas e propriedades nutricionais
das gorduras e óleos comestíveis também dependem da composição dos seus ácidos graxos (isto é,
do grau de saturação e do comprimento da cadeia carbônica e da sua distribuição estereoquímica na
molécula de glicerol), e das suas formas polimórficas (SENANAYAKE e SHAHIDI, 2005).
No organismo humano, os triacilgliceróis atuam como fonte de energia armazenada no tecido
adiposo (camada protetora térmica e mecânica que envolve órgãos importantes), fonte de ácidos
graxos essenciais, fonte e veículo de absorção de vitaminas lipossolúveis e outros compostos não
polares. Eles compõem uma parte importante da dieta humana e o desequilíbrio no seu metabolismo
pode levar a diversas desordens metabólicas, tais como: doenças cardíacas, dislipidemia, obesidade,
erros inatos do metabolismo, etc (SMALL, 1991).
Os ácidos graxos, um dos principais componentes dos triacilglicerol, podem variar no
tamanho da cadeia carbônica, no grau de saturação das suas ligações e na conformação cis ou trans
das ligações duplas. Centenas de ácidos graxos naturais foram identificados e de acordo com as
diferentes posições estereoquímicas sn-1, 2 ou 3 na molécula de glicerol ou configuração ótica (R/S)
do triacilglicerol, uma enorme variedade e complexidade de triacilgliceróis pode ser obtida (LÍSA e
HOLCAPEK, 2008).
3.7. Metabolismo e Absorção dos Lipídios
O efeito da composição dos ácidos graxos, sua posição do triacilglicerol (TAG) e suas
características físicas e bioquímicas influenciam diretamente na sua absorção no trato gastrointestinal,

15
no metabolismo dos enterócitos e quilomícrons e consequentemente, na sua distribuição nos tecidos
(GUNSTONE, 2006).
Estas características peculiares dos lipídios e seus ácidos graxos geram subsequentes
implicações na saúde humana, uma vez que eles estão diretamente relacionados às doenças
coronarianas (LEFEVRE et al, 2004), no balanço energético, sensibilidade à insulina e no
metabolismo em jejum e pós-prandial (SARAH e BERRY, 2009).
Pesquisas sobre a digestão e absorção de lipídios indicam que os ácidos graxos presentes nas
posições sn-1,3 dos TAGs são metabolizados de forma diferente daqueles presentes na posição sn-2,
devido a ação da lipase pancreática no duodeno. Por se tratar de uma lipase sn-1,3 específica, ela atua
no TAG, gerando ácidos graxos livres (das posições sn-1,3) e o 2-monoacilglicerol (2-MAG), ambos
absorvidos na mucosa intestinal. Isto influenciará diretamente na composição dos quilomícrons
formados na ressíntese dos triacilgliceróis no enterócito, consequentemente na biodisponibilidade
desses ácidos graxos (BERRY et al, 2007; YANG e KUKSIS, 1991; XU, 2000; MU e HØY, 2004;
RAMÍREZ; AMATE; GIL, 2001).
Devido à forma que os AGCM são digeridos, eles podem ser absorvidos no estômago, após a
hidrólise pela lipase gástrica (CHISTENSEN et al, 1995), e também pode ser solubilizados na fase
aquosa do conteúdo intestinal, onde se ligam à albumina, e são transportados até o fígado através da
veia porta (DECKER, 1996). Por estas razões, os AGCM são usados como fonte rápida de energia
em algumas síndromes metabólicas, como na deficiência de enzima pancreática (fibrose cística)
(JENSEN; CHRISTENSEN; HØY, 1994).
Os lipídios ingeridos na dieta, compostos por ácidos graxos essenciais ou de interesse
nutricional são mais eficientemente absorvidos quando presentes na posição sn-2 dos TAGs. Além
disso, os ácidos graxos presentes na posição sn-2 do TAG podem ser preferencialmente
transportados para o fígado, ao invés de serem transportados para o tecido adiposo (HUNTER,
2001). Além disso, os ácidos graxos livres podem interagir com cátions divalentes, como o cálcio,
tornando-se insolúveis no meio alcalino do intestino grosso e, portanto, menos biodisponíveis
(TOMARELLI et al, 1968) .
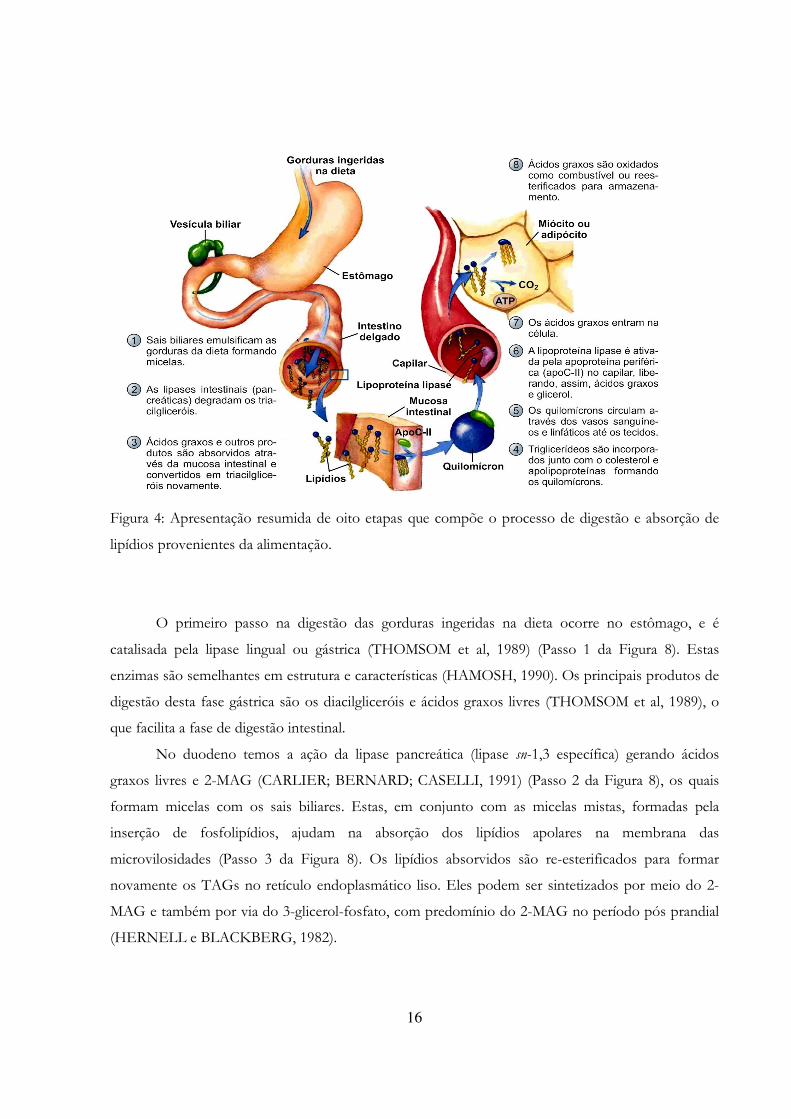
16
Figura 4: Apresentação resumida de oito etapas que compõe o processo de digestão e absorção de
lipídios provenientes da alimentação.
O primeiro passo na digestão das gorduras ingeridas na dieta ocorre no estômago, e é
catalisada pela lipase lingual ou gástrica (THOMSOM et al, 1989) (Passo 1 da Figura 8). Estas
enzimas são semelhantes em estrutura e características (HAMOSH, 1990). Os principais produtos de
digestão desta fase gástrica são os diacilgliceróis e ácidos graxos livres (THOMSOM et al, 1989), o
que facilita a fase de digestão intestinal.
No duodeno temos a ação da lipase pancreática (lipase sn-1,3 específica) gerando ácidos
graxos livres e 2-MAG (CARLIER; BERNARD; CASELLI, 1991) (Passo 2 da Figura 8), os quais
formam micelas com os sais biliares. Estas, em conjunto com as micelas mistas, formadas pela
inserção de fosfolipídios, ajudam na absorção dos lipídios apolares na membrana das
microvilosidades (Passo 3 da Figura 8). Os lipídios absorvidos são re-esterificados para formar
novamente os TAGs no retículo endoplasmático liso. Eles podem ser sintetizados por meio do 2-
MAG e também por via do 3-glicerol-fosfato, com predomínio do 2-MAG no período pós prandial
(HERNELL e BLACKBERG, 1982).

17
Os TAGs, fosfolipídios, colesterol e apoproteínas são utilizados para sintetizar os
quilomícrons (Passo 4 da Figura 8), que são secretados para a linfa e em seguida para a corrente
sanguínea, em geral através do canal torácico (RAMÍREZ; AMATE; GIL, 2001) (Passo 5 da Figura
8). Nos tecidos periféricos, eles são clivados pela lipoproteína lipase (Passo 6 da Figura 8), liberando
os TAGs, que serão metabolizados no fígado, juntamente com os outros compostos.
3.8. Lipídios Estruturados
Nos últimos anos temos assistido a um extraordinário interesse na modificação enzimática
(biotransformações) de óleos e gorduras para síntese de lipídios com propriedades nutricionais
desejáveis. Lipídios esses que foram denominados de triacilgliceróis estruturados (BORNSCHEUER;
ADAMCZAK; SOUMANOU, 2003).
O termo triacilglicerol estruturado foi introduzido primeiramente por Babayan (1987) para
descrever óleos e gorduras modificados, objetivando a alteração na composição dos ácidos graxos,
consequentemente, na estrutura do triacilglicerol após reações de hidratação, interesterificação,
esterificação, pelo uso de técnicas de engenharia genética, etc (BEDNARSKI, 2003).
Os lipídios estruturados são muitas vezes ditos como a nova geração de gorduras que podem
ser considerados como produtos nutracêuticos. Neste caso, nutracêutico refere-se a alimentos ou
partes de alimentos que proporcionam benefícios médicos ou de saúde, incluindo seu potencial para
a prevenção e/ou tratamento de doenças (AKOH, 2002; D’AGOSTINI et al, 2001; GUNSTONE,
1998). Segundo Hoy e Xu (2001), triacilgliceróis estruturados geralmente são quaisquer gorduras que
são modificados ou re-estruturados a partir de óleos e gorduras naturais, com funcionalidades ou
propriedades nutricionais para fins comestíveis ou farmacêuticos.
Uma definição mais abrangente foi descrita por Xu (2000) onde os triacilgliceróis são
modificados ou re-estruturados para atender a funções especiais, com aplicações em alimentos,
nutrição e farmacêuticas.
A sua produção só é possível com a utilização de lipases como catalisadores onde a
estereoespecificidade destas enzimas é explorada. Portanto, métodos químicos são impróprios para a
produção desse tipo de triacilglicerol por serem não estereoespecíficos. Dentre as aplicações clássicas
que são feitas por meio dos lipídios estruturados, podemos destacar a produção de: margarinas,
manteiga de cacau, substituto de gordura do leite humano, modificação de óleo de peixe, entre outras
(HAMAM e SHAHIDI, 2005; JENSEN, 2001; XU, 2000, QUINLAN e MOORE, 1993). Devemos
salientar também que as modificações na composição, estrutura e propriedades de triacilgliceróis
visam: reduzir o consumo de ácidos graxos saturados e seus isômeros trans; aumentar o teor de

18
ácidos graxos essenciais nos triacilgliceróis; diminuir o teor calórico de óleos e gorduras; melhorar as
suas propriedades físico-químicas, melhorar a estabilidade oxidativa, ponto de congelamento,
produção de óleos e gorduras caracterizados por propriedades de promoção da saúde (ricos em ácido
linoléico conjugado - CLA); etc (ADAMCZAK, 2004; OSBORN e AKOH, 2002).
De uma forma geral, o conhecimento sobre a distribuição estereoespecífica dos ácidos graxos
no triacilglicerol é de grande interesse para o entendimento das funções biológicas tanto dos próprios
ácidos graxos, como das diferentes classes de lipídios que fazem parte da dieta humana
(WANASUNDARA e SHAHIDI, 1999; SHAHIDI e WANASUNDARA, 1998; SENANAYAKE e
SHAHIDI, 2002). Ademais, há uma demanda crescente por uma melhor qualidade nutritiva nos
alimentos nos dias atuais, muito em conta da busca por uma vida mais saudável.
Entretando, devido ao elevado custo destes biocatalisadores, a sua utilização em processos
para obtenção de produtos de menor valor agregado ainda é bem limitada.
3.9. Reações para Obtenção dos Lipídios Estruturados
A posição estereoespecífica dos ácidos graxos nos lipídios pode ser estudada pelo processo
de interesterificação. Esta reação ocorre quando os ácidos graxos alteram suas posições
estereoquímica (sn-1, 2 ou 3) na molécula de glicerol (intramolecular) ou pela troca de ácidos graxos
entre duas moléculas de triacilglicerol (intermolecular) até o momento que o equilíbrio
termodinâmico é atingido (HAMAM e SHAHIDI, 2005). Estas características são exploradas pela
indústria alimentícia com intuito de alterar as propriedades físico-químicas dos lipídios, tais como:
propriedades de fusão de óleos e gorduras, forma de cristalização, textura, espalhabilidade,
plasticidade, estabilidade a oxidação, conteúdo de gordura sólida, etc (ADAMCZAK, 2004;
MARANGONI e ROUSSEAU, 1995). A Unilever Ltd. foi uma das primeiras empresas a usar
interesterificação “dirigida” catalisada por lipases 1,3 específicas em 1980 em gorduras mais baratas
que pudessem ser utilizadas no lugar da manteiga de cacau, a partir do óleo de palma com ácido
esteárico (MARANGONI e ROUSSEAU, 1995).
A reação de interesterificação oferece um grande potencial de aplicação na produção de
lipídios estruturados. Ela engloba três abordagens: acidólise, alcoólise e transesterificação. Podem ser
empregados três tipos de doadores de grupamento acila são comumente utilizados na produção de
lipídios estruturados. São eles: os triacilgliceróis, os ésteres de ácidos graxos e os ácidos graxos livres.
A escolha desses doadores de grupamento acila depende da pureza do produto, a reatividade, grau de
pureza e preço.

19
A interesterificação química é o método mais utilizado na interesterificação de óleos e
gorduras em virtude de sua simplicidade e por empregarem catalisadores de menor custo quando
comparado a interesterificação enzimática (SREENIVASAN, 1976; MARANGONI e ROUSSEAU,
1999). Porém há uma série de desvantagens, tais como: síntese aleatória (produtos randomizados)
por não oferecer especificidade com pouco ou nenhum controle sobre a distribuição posicional dos
ácidos graxos no produto final, a necessidade de um período de indução dos catalisadores antes de
iniciar a reação, perda de matéria prima por degradação ou formação de sabão, necessidade de etapas
de extras de purificação para remoção do catalisador (SREENIVASAN, 1976; MARANGONI e
ROUSSEAU, 1995; GUPTA; RATHI; BRADOO, 2003). Em geral, a interesterificação química
permite uma troca aleatória de ácidos graxos enquanto a enzimática catalisa a troca dos ácidos graxos
em posições específicas, porém é amplamente empregada, em particular na Europa, na produção de
gorduras saturadas com baixos níveis, ou totalmente isenta de gordura trans (MARANGONI e
ROUSSEAU, 1999; MARANGONI e ROUSSEAU, 1995).
3.10. Alimentos Funcionais / Nutracêuticos / Bioativos
Hoje em dia, a modificação de óleos e gorduras é uma das principais áreas da indústria de
processamento de alimentos. Mas antes de nos aprofundarmos neste ítem, algumas definições devem
ser consideradas para um melhor entendimento dos produtos abordados e das modificações
propostas neste trabalho; bem como para o efeito de comparação e melhor entendimento do que é
descrito pela literatura na classificação dessa nova geração de lipídios modificados.
Assim sendo, os alimentos funcionais podem ter um ou mais compostos com funções
bioquímicas e fisiológicas benéficas para a saúde humana. Segundo a American Dietetic Association,
alimento funcional é qualquer alimento modificado que possa trazer algum efeito benéfico além dos
nutrientes que ele originalmente os contenha. Já os nutracêuticos, foram classificados como qualquer
substância considerada como alimento, ou parte dele, que seja benéfica à saúde ou traga benefícios
médicos, incluindo a prevenção e tratamento de doenças (BLOCH e THOMSON, 1995).
Thomas e Earl (1994) definiram alimento funcional como qualquer alimento modificado ou
ingrediente alimentar que possa trazer benefícios à saúde, além dos nutrientes que ele previamente
possui. O termo nutracêutico foi proveniente da junção dos termos “nutrição” e “farmacêutico”, o
que novamente sugere um benefício de saúde mais amplo do que o obtido somente pelo nutriente,
foi proposto em 1989 por Stephen De Felice.
Uma abordagem interessante e singular foi proposta por Zeisel (1999), onde ele define
nutracêuticos como suplementos alimentares que fornecem uma elevada quantidade de um

20
composto alimentar bioativo, tendo como veículo uma matriz não alimentar, e usado com o
propósito de promover benefícios para a saúde, por meio de doses que excedam a quantidade do que
seria obtida pela ingestão normal dos alimentos.
De certa forma, os alimentos beneficiados ou processados podem ser gerados a partir de
alimentos que já contenham esses ingredientes ou enriquecidos com nutrientes conhecidos, como
por exemplo: produtos enriquecidos com cálcio são utilizados na tentativa de reduzir o risco de
osteoporose; alimentos reforçados com fibra podem reduzir o risco de câncer de cólon; etc.
Também temos como opção, a remoção de certos tipos de nutrientes, como no caso na
obtenção de gorduras com baixo teor de energia. No entanto, cada vez mais, o termo alimento
funcional está sendo aplicado a uma gama de alimentos, que incluem o óleo de peixe, em decorrência
da sua composição e benefício dos ácidos graxos poli-insaturados (GHOSH; SKINNER;
FERGUSON, 2006).
Haja vista isso, podemos observar que não há um consenso geral para a classificação de um
alimento sendo como funcional ou nutracêutico, uma vez que, dependendo da fonte consultada, uma
definição pode tanto compreender a outra, como ser totalmente díspare.
Como a fronteira entre nutracêuticos e alimentos funcionais nem sempre é clara. Uma das
formas utilizadas para esta diferenciação é o modo como o alimento será consumido. Normalmente
os nutracêuticos são consumidos na forma de cápsulas, pílulas, comprimidos, etc; enquanto os
alimentos funcionais são consumidos como alimentos in natura. Assim, quando um fitoquímico está
incluído numa formulação de alimentos é considerado um alimento funcional; já se o mesmo
fitoquímico está incluído numa cápsula, ele vai ser enquadrado como um nutracêutico (ESPÍN;
GARCÍA-CONESA; TOMÁS-BARBERÁN, 2007).
Nutracêuticos podem ser encontrados em vários produtos nos ramos alimentício e
farmacêutico, tais como: indústria de alimentos; suplementos fitoterápicos e dietéticos; indústria
farmacêutica; mais recentemente em micro-organismos engenheirados; agroprodutos; biomoléculas
ativas; produtos processados, como cereais, sopas e bebidas (SHARMA e SINGH, 2010).
Quimicamente os nutracêuticos podem ser classificados como derivados isoprenóides
(terpenóides, carotenóides, saponinas, tocotrienóis, tocoferóis, terpenos), compostos fenólicos
(curaminas, taninos, antocianinas, isoflavonas, flavononas, flavonóides), derivados de carboidratos
(ácido ascórbico, oligossacarídeos), ácidos graxos e lipídios estruturais (ácido graxo poli-insaturado ω-
3, ácido linoléico conjugado, ácidos graxos monoinsaturados, esfingolipídios, lecitinas), derivados de
aminoácidos (capsaicina, isotiocianatos, indóis, ácido fólico, colina), micro-organismos (probióticos,
prebióticos) e minerais (Ca, Zn, Cu, K, SE) (DUREJA; KAUSHIK; KUMAR, 2003).

21
Já o termo “alimento bioativo” foi definido pela primeira vez como: alimentos, ingredientes
alimentares ou suplementos alimentares que demonstrem benefícios específicos à saúde, ou
benefícios médicos que incluam a prevenção e o tratamento da doença além de funções nutricionais
básicas (GERMAN e DILLARD, 2000). Inicialmente, até 1990, os alimentos bioativos eram
considerados como alimentos naturais que pudessem fornecer a energia que atendesse as
necessidades diárias do metabolismo.
Os lipídios bioativos podem estar naturalmente presentes nos alimentos, tais como: óleos,
ovos, leite, vegetais, etc; ou podem ser adicionados como ingredientes funcionais específicos, tais
como: fitoesteróis e vitaminas. O fator que determinará se um lipídio é um alimento bioativo, será o
seu potencial para promover a saúde humana (além de uma fonte de energia simples) (CHEN;
McCLEMENTS; DECKER, 2013).
Posteriormente, na ultima década, os nutracêuticos foram ganhando importância pelos
efeitos benéficos sobre as doenças cardiovasculares; e a partir do século 21, com a utilização
crescente dos nutracêuticos em virtude dos seus benefícios, uma nova concepção de suplementos
terapêuticos esta emergindo. Esta forte corrente vai em direção a um novo conceito de medicina
nutracêutica, chamada medicina complementar alternativa (SHARMA e SINGH, 2010); e agora, os
alimentos bioativos e nutracêuticos surgiram como suplementos para alimentação que sejam
potenciais fontes naturais de prevenção de doenças cardiovasculares e do câncer (GERMAN e
DILLARD, 2000).
Eles podem ser encontrados em várias matrizes biológicas de diferentes famílias, tais como:
lipídios, vitaminas, proteínas, glicosídeos, compostos fenólicos (BERNAL et al, 2011). Uma dessas
matrizes são os lipídios, que são um grande grupo de compostos naturais que inclui gorduras, ceras,
esteróis, vitaminas solúveis em gordura (tais como as vitaminas A, D, E e K), monoglicerídeos,
diglicerídeos, fosfolipídios, os carotenóides, etc. Moléculas como os ácidos graxos e seus derivados
(incluindo tri-, di- e monoglicerídeos e fosfolipídios), metabólitos derivados dos esteróis, tais como o
colesterol, também estão agrupados na família dos lipídios.
Os lipídios desempenham importantes funções biológicas no metabolismo, dentre elas
podemos citar: armazenamento de energia, são componentes estruturais das membranas celulares,
participam de mecanismos de sinalização celular, são fonte e veículos de absorção para vitaminas
lipossolúveis (BERNAL et al, 2011).
3.11. Aplicações Específicas

22
3.11.1. Enriquecimento Nutricional de Óleos Vegetais Comestíveis com Ácidos Graxos Poli-insaturados da Família ω-3.
A incorporação desses ácidos graxos de cadeia longa em alimentos de matriz oleosa
representa um desafio considerável, devido ao aumento no risco de oxidação lipídica, que pode
acerretar no desenvolvimento de sabores e odores desagradáveis, assim como a redução da vida útil
de prateleira (ERIC et al, 2012). Logo, a oxidação lipídica tem um impacto importante sobre o sabor
dos óleos ou de qualquer alimento fabricado a partir de óleos.
Dessa maneira, a pesquisa por novos veículos, tais como os óleos vegetais, aos quais esses
ácidos graxos possam ser incorporados, os tornam alvos ideias para aumentar a ingestão de AGPI ω-
3 com o intuito de alcançar a dose de ingestão diária recomendada. Eles podem ser adicionados a
uma variedade de alimentos e demonstrar igual aceitação, em relação aos óleos vegetais originais de
uso comum, na medida em que as propriedades organolépticas não sofram alterações inesperadas.
Para tal, devemos assegurar que a fonte de AGPI ω-3 não afete o sabor do alimento e que os ácidos
graxos devem ser oxidativamente estável durante o vida de prateleira típica do produto alimentar.
Averiguado estes fatos, podemos explorar fontes como os óleos de coco, óleo de palma e azeite de
oliva, pois os mesmos têm o potencial para atender a estes critérios. Ademais, estes óleos já são
comumente utilizados na culinária.
3.11.1.1. Mercado de Produtos Nutracêuticos
Empresas tradicionais de alimentos, mesmo sob a pressão da intensa concorrência do
segmento, veêm o mercado de alimentos funcionais, bebidas e suplementos com bons olhos.
Embora a indústria de produtos nutracêuticos seja inferior a 10% da indústria alimentar total, o
mercado oferece oportunidades significativas de crescimento e margens de lucro mais amplas, ainda
que haja uma falta de categorização clara no mercado de nutracêuticos em termos de ingredientes,
aplicações, produtos e tecnologia (BCC RESEARCH, 2011a).
O mercado mundial de nutracêuticos global foi de US$ 151 bilhões em 2011 e projeções para
2016 apontam para cerca de US$ 207 bilhões, com uma taxa de crescimento anual de 6,5% entre
2011 e 2016. Já para o mercado de bebidas funcionais é esperado um maior crescimento, em torno
de 8,8% ao ano, durante o período de cinco anos. Este setor, que chegou a US$ 57 bilhões em 2011,
deverá girar ao redor de US$ 87 bilhões em 2016 (BCC RESEARCH, 2011a).
Portanto, a perspectiva de novos avanços geraram uma necessidade de redução de custos de
produção. Gastos com P & D, juntamente com o aumento da concorrência entre as empresas,

23
expirações de patentes e a implementação de novas tecnologias, estão levando este mercado a novos
rumos.
3.11.2. Lipídios Estruturados de Baixo Teor Calórico
Como a obesidade é hoje uma “epidemia” global e tem sido associada a doenças fatais, tais
como doenças cardiovasculares, diabetes e hipertensão, o desenvolvimento de óleos funcionais e
gorduras saudáveis, com o perfil lipídico (M-L-M), ou seja, que contêm ácidos graxos de cadeia longa
na posição sn-2 e ácidos graxos de cadeia média na posição sn-1,3 do triacilglicerol pode ser utilizadao
de uma forma eficiente no combate a perda de peso (FITCH-HAUMANN, 1997). Esta estruturação
com AGPI na posição sn-2 é especialmente atraente em virtude dos efeitos benéficos desses ácidos
graxos para o sistema imunológico humano e o consumo deste tipo de triacilglicerol também é
benéfico para as funções do cérebro e do fígado (GILL e VALIVETY, 1997).
Lipídios estruturados de baixa caloria são feitos sob medida são usados tanto por ser uma
fonte de energia facilmente disponível, como por ser produto de baixo teor calórico. Podem ser
aplicados na nutrição de crianças, bem como na nutrição clínica de pacientes com distúrbios de
digestão ou absorção de nutrientes. Essas triacilgliceróis também exibem propriedades físico-
químicas e estabilidade oxidativa desejáveis (BABAYAN e ROSENAU, 1991; FITCH-HAUMANN,
1997; MEGREMIS, 1991).
Atualmente o mercado oferece uma ampla variedade de produtos contendo triacilgliceróis de
cadeia média. O Caprenin® é um exemplo de produto comercial de baixo teor calórico. É um
triacilglicerol randomizado compreendendo ácido caprílico (C8:0), cáprico (C10:0) e behênico
(C22:0). O valor calórico do Caprenin® foi calculado como sendo de 4,3 - 5,0 kcal/g (LONG, 2010;
OSBORN e AKOH, 2002b). O Caprenin® apresenta propriedades semelhantes às de manteiga de
cacau e pode ser usado em doces e revestimentos de confeitaria. Os ácidos graxos caprílico e cáprico
são obtidos a partir de óleos de coco e de palmiste; já o ácido behênico, é encontrado naturalmente
no amendoim, mas para utilização na Caprenin® é produzido a partir de óleo de colza hidrogenado
(FINLEY; McDONALD; KLEMANN, 1997), tornando-o seguro para os indivíduos com alergia a
amendoim. O ácido behênico é parcialmente absorvido pelo organismo, sendo responsável pelo
valor calórico reduzido do produto desenvolvido pela Procter & Gamble Co. (OSBORN e AKOH,
2002b, AUERBACH; KLEMANN; HEYDINGER, 2001).
O Salatrim® também é uma gordura com teor calórico reduzido, desenvolvida pelo grupo
Nabisco Foods e comercializado pela Danisco Cultor sob o a marca registada Benefat™ (FINLEY;
McDONALD; KLEMANN, 1997). A palavra Salatrim® é um acrônimo derivado de triacilglicerol de

24
cadeia curta e longa. Salatrim® é uma mistura de TAG composta de pelo menos um ácido graxo de
cadeia longa (C16-C22, principalmente pelo ácido esteárico) e de ácidos graxos de cadeia curta (C2-
C4) (SMITH; FINLEY; LEVEILLE, 1994). Salatrim® é um substituto à base de gordura que pode
ser utilizado em chocolate, produtos de confeitaria e produtos lácteos. Devido à composição
molecular única, o Salatrim® fornece uma média de 5 kcal/g, ou 55% do valor calórico de gorduras
convencionais (9 kcal/g) (SMITH; FINLEY; LEVEILLE, 1994).
Em 2009, o mercado de produtos de emagrecimento e serviços foi de quase US$ 121 bilhões,
com previsão de crescimento para mais de US$ 134 bilhões em 2014. Alimentos e bebidas são as
maiores categorias neste tipo de mercado. Este segmento chegou a US$ 79 bilhões em 2009. A
tendência atual de aumento no consumo de alimentos saudáveis deve impulsionar o crescimento
deste setor em 2% ao ano, chegando a mais de US$ 86 bilhões até 2014 (BCC RESEARCH, 2011c).
3.11.2.1. Mercado de Produtos para Perda de Peso
À medida que o número de indivíduos com sobrepeso e obesidade aumenta a uma taxa
alarmante, principalmente nos Estados Unidos, numerosas oportunidades de negócio para
implementação de produtos e serviços que visam, no todo ou em parte, a perda de peso tem surgido.
Em 2009, o mercado de produtos de emagrecimento e serviços foi de quase US$ 121 bilhões,
com previsão de crescimento anual de 2,2%, resultando em um mercado de mais de US$ 134 bilhões
para o ano de 2014 (BCC Research, 2010). Alimentos e bebidas são a maior categoria neste tipo de
mercado. Este segmento foi avaliado em cerca de US$ 79 bilhões em 2009 e a tendência de aumento
no consumo de alimentos saudáveis deve impulsionar o crescimento deste setor em 2% ao ano,
chegando a mais de US$ 86 bilhões em 2014 (BCC Research, 2010).
3.11.3. Produção de Substitutos da Gordura do Leite Humano
Embora o leite humano seja a primeira escolha para recém-nascidos, substitutos do leite
materno desempenham um papel indispensável na nutrição infantil quando a amamentação não é
possível, desejável ou suficiente. Fórmulas lácteas infantis têm sido projetadas para fornecer as
crianças os nutrientes necessários para o crescimento ideal e desenvolvimento (MARTINE; PETRA;
JACQUES, 2004).
Para crianças alimentadas exclusivamente com leite materno, ele é a única fonte de nutrientes
durante os primeiros meses. Apesar de conter um percentual reduzido de lipídios, os triacilgliceróis

25
são responsáveis por fornecer 50-60% da energia da dieta e também estão envolvidos na absorção de
nutrientes (JENSEN; BUIST; WILSON, 1986; KENNEDY, 1999; JENSEN e JENSEN, 1992;
WEBER e MUKHERJEE, 2005).
A gordura do leite materno é a maior fonte de energia para as crianças (NESE; CASIMIR;
ARTEMIS, 2005; MADUCO; AKOH; PARK, 2007; XU, 2000). Composto por 3-5% de lipídios
totais, sendo mais de 90% na forma de triacilglicerol; contém de 20-25% de ácido palmítico (16:0),
sendo que a maior parte dele está esterificado na posição sn-2 do TAG; Já as posições sn-1,3
encontramos principalmente ácidos graxos insaturados tais como oléico (18:1n9) (30-35%), linoléico
(18:2n6) (7-14%) e esteárico (C18:0) (5-7%) (JENSEN, 1996; MENDEZ e SALAS, 2001;
ADLERCREUTZ, 1994; SRIVASTAVA; AKOH; CHANG, 2006; JENSEN, 1996; BRACCO,1994;
NIELSEN et al, 2006).
Essa distribuição posicional é diferente das formas que normalmente são encontradas em
óleos derivados de fontes vegetais e animais, com exceção de banha de porco e da estearina de palma
(INNIS; QUINLAN; NELSON, 1998; XU, 2000; OSBORN e AKOH, 2002; SOUMANOU;
PÉRIGNON; VILLENEUVE, 2013).
Estudos têm mostrado que a adição dessa composição em fórmulas infantis, como a fonte de
gorduras, pode desempenhar um efeito benéfico na absorção de lipídios, na sua disponibilidade
energética, na absorção de minerais e na melhora das fezes tanto em bebês prematuros, como em
neonatos, quando comparado a fórmulas comerciais que contêm baixos níveis de ácido palmítico na
posição sn-2 do triacilglicerol (LUCAS, 1997; CARNIELLI, 1996). Isso se deve ao fato de que o
predomínio de ácido palmítico na posição sn-2 evita a formação de sabões insolúveis de cálcio
causada pela interação de ácidos graxos e íons de cálcio, o que leva a perdas de energia e minerais nas
crianças (LÓPEZ-LÓPEZ, 2001; BASS e CHAN, 2006; LNNIS; DYER; NELSON, 1994). Nesta
conformação, ela irá formar micelas mistas com sais biliares e será mais eficazmente absorvida nos
enterócitos por difusão (THOMSON et al, 1989; SMALL, 1991; LONG, 2010).
Dentre os produtos que tem esta formulação como base, destacamos o Betapol™. É um
produto desenvolvido pela Loders Croklaan (Unilever Co.), utilizado em substituição da gordura do
leite materno, produzido à base dos óleos vegetais específica do leite materno, tendo como base o
1,3-dioleoil-2-palmitoilglicerol (OPO), que é o principal TAG presente no Betapol™ (XU, 2000). A
Unilever Co. depositou as primeiras patentes para a síntese desses lipídios catalisada por lipases
(KING e PADLEY, 1990).
Este processo utiliza lipases sn-1,3 específicas e ácidos graxos livres como doadores de
grupamento acila (QUINLAN e MOORE, 1993). Devido ao alto teor de ácido oléico, a estearina de
palma e o óleo de girassol são industrialmente utilizados pela Loders Croklaan.

26
Além Betapol™, outros substitutos da gordura do leite materno comerciais, como o InFat™,
também são fabricados seguindo o mesmo conceito de biocatálise (MEIRI-BENDEK, 2011).
Uma abordagem interessante é feita em única etapa usando tripalmitina e ácido oléico
catalisado por lipases 1,3-regioespecífica (Figura 9) (SRIVASTAVA et al, 2006). Recentemente,
muitos estudos têm sido feitos para produção de lipídios estruturados utilizados em fórmulas de leite
infantil ou como substituto da gordura do leite materno (SAHIN; AKOH; KARAALI; 2005,
MUKHERJEE e KIEWITT, 1998). Muitos destes estudos foram realizados utilizando a lipase de
Rhizomuco miehei (Lipozyme® RM IM) como biocatalisador para as reações de transesterificação
(THOMSON, 1989; SAHIN; AKOH; KARAALI; 2005). Além do mais, pesquisas têm sido feitas
com a utilização de EPA e DHA, devido aos benefícios para a saúde que são associados aos ácidos
graxos poli-insaturados família ω-3 (SAHIN; AKOH; KARAALI, 2006).
3.11.3.1. Mercado de Fórmulas Infantis
O número crescente de mulheres indo para o mercado de trabalho associado as crescentes
preocupações dos pais sobre nutrição e das restrições de tempo inerentes a modernidade aumentou a
demanda por alimentos mais apropriados para bebês. Estas refeições ou fórmulas imediatas e
práticas tornam-se cada vez mais atraentes no atendimento às demandas nutricionais atuais de bebês
e crianças no mundo todo (BCC RESEARCH, 2012b).
Nesse contexto, o mercado de fórmulas infantis oferece muitas oportunidades para produtos
novos e/ou inovadores. Neste item estão inclusos categorias como fórmula infantil e as refeições
para bebê.
Figura 5: Reação de acidólise da tripalmitina pelo ácido oléico para produção de Betapol™

27
O mercado global de alimentos para bebês em 2011 foi de US$ 28,2 bilhões e deve chegar a
US$ 35,2 bilhões até o final de 2016, com uma taxa de crescimento de 4,5% ao ano (BCC
RESEARCH, 2012b).
A América do Norte detém a maior participação global com 37,2% do mercado e obteve
uma receita de US$ 10,2 bilhões em 2011, com perspectiva que chegue a US$ 10,9 bilhões em 2016.
Já o mercado Europeu em 2011 foi de US$ 8,8 bilhões, deverá chegar a US$ 9,6 bilhões em 2016
(BCC RESEARCH, 2012b).
3.12. Óleos Vegetais
Os óleos vegetais são uma fonte lipídica importante e amplamente utilizada em produtos para
fins alimentares. No entanto, alguns óleos vegetais não são aconselháveis para atender a satisfação do
consumidor, em termos das suas propriedades físico-químicas ou para a textura e a estabilidade dos
alimentos (REYES-HERNÁNDEZ et al, 2007).
Os óleos vegetais são compostos de uma mistura de diferentes ácidos graxos, em diferentes
proporções. Eles também são a principal fonte de esteróis de plantas (fitoesteróis) na dieta humana e
contêm componentes menores, como esqualeno e esfingolipídios, os quais podem fornecer uma
série de benefícios para a saúde (FOSTER; WILLIAMSON; LUNN, 2009).
As propriedades dos óleos podem ser modificadas através de processos como a
hidrogenação, fracionamento e interesterificação. A hidrogenação é um processo industrial
comumente utilizado para aumentar a dureza e estabilidade das gorduras. No entanto, esta
hidrogenação, seja parcial ou total, resulta na isomerização e migração de duplas ligações que levam à
formação de ácidos graxos trans. Ácidos graxos trans são os ácidos graxos insaturados com pelo
menos uma ligação dupla na configuração trans. Isso resulta em um ângulo de ligação maior do que a
dos ácidos graxos insaturados contendo ligação dupla na conformação cis e uma cadeia de carbono
mais volumosa em relação aos graxos saturados (FILIP et al, 2010).
A ingestão de grandes quantidades de ácidos graxos trans foi positivamente correlacionada
ao aumento do risco de doença cardíaca coronariana (HU et al, 1997), processos inflamatórios
(MOZAFFARIAN et al, 2004) e câncer (ASTORG, 2005). Estudos têm mostrado que o aumento na
ingestão desses ácidos graxos também aumenta os níveis de colesterol e LDL (lipoproteínas de baixa
densidade) e diminui o HDL (lipoproteína de alta densidade) no sangue (MOZAFFARIAN E
CLARKE, 2009). Alguns fabricantes de alimentos têm usado a hidrogenação completa de óleos
seguida pelo processo de interesterificação, no lugar da hidrogenação parcial para assegurar que o

28
óleo tenha as propriedades funcionais e as qualidades organolépticas desejadas, sem a inconveniente
formação de ácidos graxos trans (FOSTER; WILLIAMSON; LUNN, 2009).
Atualmente, o processo de interesterificação é um dos mais utilizados para modificar as
propriedades físicas dos óleos e gorduras, alterando assim, suas propriedades funcionais e
nutricionais. Nele, as posições dos três ácidos graxos da molécula de triacilglicerol podem ser
rearranjados através da utilização de enzimas (lipase) ou de catalisadores químico. Ao longo dos anos,
a interesterificação tem demonstrado ser um processo seguro, embora poucos estudos tenham
investigado especificamente os benefícios para a saúde ou a inocuidade provenientes dessas gorduras
interesterificadas (FOSTER; WILLIAMSON; LUNN, 2009).
3.12.1. Óleo de coco (Cocos nucifera L.)
3.12.1.1. Características Gerais
O óleo de coco é uma fonte natural de ácidos graxos de cadeia média (AGCM), contendo
mais de 90% de ácidos graxos saturados, tende a ser sólidos à temperatura ambiente, sendo
altamente resistente a rancidez oxidativa.
Os ácidos graxos de cadeia média ao serem absorvidos nos enterócitos entram no sistema
circulatório através da veia porta e são transportados até o fígado, onde são prontamente oxidados a
cetonas. Isto porque os ácidos graxos de cadeia média não necessitam de transportadores (carnitina
palmitoil transferase) que estão presentes na membrana interna das mitocôndrias. Este transportador
é uma enzima que limita a velocidade da ß-oxidação. Assim, os AGCM induzem a termogênese e não
contribuem para o ganho de peso, uma vez que eles não são depositados nos tecidos adiposos.
3.12.1.2. Benefícios Funcionais / Usos Terapêuticos
Estudos epidemiológicos sugerem que o consumo de grandes quantidades de gordura
saturada leva a níveis elevados de colesterol no sangue (GERMAN e DILLARD, 2004). Devido a
isso, o óleo de coco tem recebido má reputação. No entanto, nos últimos anos, em estudos clínicos
conduzidos em óleo de coco, resultados positivos foram obtidos, o que podem refutar estes
argumentos (MARINA; CHE MAN; AMIN, 2009).
Os AGCM têm inúmeras aplicações funcionais, pois eles podem estimular o sistema
imunológico, são utilizados no controle de peso, como agentes antimicrobianos, como adjuvantes

29
dietéticos em casos de doença crônica degenerativa, tais como a doença cardiovascular, doenças do
fígado e câncer (GOPALA et al, 2010).
O ácido láurico, que compõe aproximadamente 55% do óleo de coco, apresenta uma forte
propriedade antimicrobiana, inibindo várias bactérias patogênicas como Listeria monocytogenes (WANG
e JOHNSON, 1992). Também foi mostrado o efeito protetor da mistura do óleo de coco juntamente
com o óleo de savelha na redução da incidência de tumores mamários (CRAIG-SCHMIDT et al,
1993). Os AGCM também são fortemente utilizados na área clínica de nutrição enteral e parenteral,
para o tratamento de pacientes que sofrem de má absorção de gordura (CHE MAN e MARINA,
2006).
Devido à absorção mais rápida, os ácidos graxos de cadeia média são úteis como uma fonte
de calorias no tratamento de pacientes hospitalizados ou debilitados. Além disso, óleos láuricos
promovem um efeito inesperado no metabolismo proteico, pois geram um saldo positivo de
nitrogênio e indiretamente preservam o catabolismo das proteínas (WO 2003033633 A1, 2003).
Uma abordagem interessante é feita com a incorporação de ácido eicosapentaenóico (EPA) e
ácido docosahexaenóico (DHA) a partir de óleo de peixe no óleo de coco. Esta reação de
interesterificação gera um lipídio estruturado com uma composição única de AGCM e AGPI, que
proporciona a soma dos efeitos benefícios para o sistema imunológico, pelo aumento na ingestão de
EPA e DHA com as vantagens nutricionais dos ácidos graxos de cadeia média de serem
hipocolesterolêmico e hipotrigliceridêmico.
3.12.2. Óleo de Palma (Elaeis guineenses Jacq.)
3.12.2.1. Características Gerais
O óleo de palma (Elaeis guineensis) é atualmente o segundo maior óleo comestível
comercializado no mundo (EDEM, 2002). O fruto da palma dá origem a dois óleos distintos. O óleo
de palma, a partir do mesocarpo do fruto; e óleo de palmiste, a partir da amêndoa. Esses óleos são
distintos em termos de sua composição de ácidos graxos.
O óleo da polpa e óleo de amêndoa de palma têm inúmeras aplicações, sendo que cerca de
80% é destinado a alimentação. Entre os usos alimentares, a oleína é usada principalmente para
preparo nas refeições, como óleo de fritura e sorvete; enquanto estearina é utilizada para a produção
de margarina e gordura vegetal (LIU e LAMPERT, 1999; PORIM, 1988; SOTERO et al., 2008).
O óleo de palma e seus derivados são comumente usados na indústria alimentícia, por duas
razões principais. Em primeiro lugar, o óleo tem uma alta estabilidade oxidativa, devido a

30
composição dos ácidos graxos, carotenóides e tocotrienóis; assim, pode ser utilizado a altas
temperaturas e suporta uma vida útil prolongada de produtos. Por outro lado, sendo, naturalmente,
semi sólido, que não exige processos de hidrogenação. Em segundo lugar, devido à alta produção de
óleo por hectare, o que a torna uma escolha relativamente econômica e facilmente disponível
(EDEM, 2002; FOSTER; WILLIAMSON; LUNN, 2009; EMBRANDIRI; IBRAHIM; SINGH,
2013).
Óleo de palma e óleo de palmiste também são ingredientes para a produção de gorduras
especiais, que incluem equivalentes à manteiga de cacau (CBE) e substitutos de manteiga de cacau
(CBS). CBE e CBS têm propriedades físicas que são semelhantes à manteiga de cacau, e são
amplamente utilizados para a produção de produtos de confeitaria de chocolate.
Geralmente, os ácidos graxos de cadeia curta ou média são absorvidos mais rapidamente do
que os ácidos graxos de cadeia longa e ácidos graxos insaturados são melhor absorvidos do que os
ácidos graxos saturados (BRACCO, 1994). O ácido palmítico é preferencialmente absorvido como 2-
moniacilglicerídeo do que na forma de ácido graxo livre (FILER; MATTSON; FOMON, 1969), pois
o ácido palmítico livre tende a complexar com sais minerais, tais como cálcio e formar sabões
insolúveis que não podem ser absorvidos no intestino delgado e, portanto, são excretados nas fezes
(JENSEN; BUIST; WILSON, 1986).
3.12.2.2. Benefícios Funcionais / Usos Terapêuticos
O óleo de palma tem baixos níveis de ácidos graxos poli-insaturados, especialmente ômega-3,
e o enriquecimento do óleo de palma com esses ácidos graxos melhora as suas propriedades
nutricionais; já as propriedades nutricionais dos componentes minoritários do óleo de palma têm
sido extensivamente estudadas, na qual os betacarotenos têm sido utilizados de forma eficaz em
estudos de intervenção dietética para combater a deficiência de vitamina A (ZAGRE et al, 2003).
3.12.3. Óleo de Oliva (Olea europaea)
3.12.3.1. Características Gerais
Este óleo desfruta de uma posição privilegiada entre os óleos comestíveis, devido ao grande
consumo de habitantes do Mediterrâneo (OH et al, 2009). A composição em ácidos graxos do óleo
de oliva (azeite de oliva) varia, dependendo do cultivar, da maturidade do fruto, altitude, clima, e

31
vários outros fatores (GALTIER et al, 2008). Os principais ácidos graxos do azeite são: (i) ácido
oléico (C18:1n9), que é um monoinsaturado da família ômega-9, que compõe 55-83%; (ii) ácido
linoléico (C18:2n6), que é um poli-insaturado da família ômega-6, que compõe 3,5-21% e (iii) ácido
palmítico (C16:0), é um ácido graxo saturado presente de 7,5-20% (OH et al, 2009)
O azeite de oliva é mais comumente usado como condimento (tempero) para saladas e para
frituras. A presença de antioxidantes naturais pode aumentar a sua estabilidade à oxidação térmica,
evitando que o óleo deteriore rapidamente a temperaturas elevadas.
3.12.3.2. Benefícios Funcionais / Usos Terapêuticos
Ao longo das últimas três décadas, estudos nutricionais e epidemiológicos têm fornecido
evidências que o consumo de azeite de oliva virgem contribui para benefícios de saúde que podem
ser atribuídos aos seus antioxidantes fenólicos, incluindo principalmente tirosol e hidroxitirosol e
seus derivados (FRANKEL, 2011; OMAR, 2010) e pela presença de uma pequena quantidade de
carotenóides (BOSKOU, 1996).
A composição química e sua fração fenólica são eficazes na diminuição do risco de doença
cardiovascular (EUROPEAN UNION, 2012). Portanto, recomenda-se o consumo diário deste óleo
(EUROPEAN FOOD SAFETY AUTHORITY, 1996).
Outros pesquisadores têm investigando os efeitos do azeite de oliva no perfil lipídico no
sangue, em fatores hemostáticos, na função imunológica e controle glicêmico, demonstrando os
efeitos de promoção da saúde que este óleo pode ter sobre a progressão de muitas doenças crônicas
(FOSTER; WILLIAMSON; LUNN, 2009).
3.13. Óleo de Peixe
3.13.1. Características Gerais
A análise dos óleos de peixe tem sido um desafio, pois geralmente eles contêm entre 30 e 60
diferentes ácidos graxos, resultando em mais de 5000 combinações possíveis TAG (BEZARD e
SEMPORE, 1995).
Os peixes de água salgada e seus óleos são a principal fonte de ácidos graxos poli-insaturados
da família ω-3 (AGPI ω-3) no Ocidente. Atualmente, seus óleos estão sendo utilizados como
suplementos (NAPIER et al, 2004), e em alguns casos, representam a única fonte natural de AGPI
ω-3, onde se tem os ácidos eicosapentaenóico (EPA, 20:5n3) e docosahexaenóico (DHA, 22:6n3),

32
como os mais biologicamente ativos desta família (FLACHS et al, 2009; HORROCKS e YEO,
1999). Os benefícios do óleo de peixe para a saúde, como a melhoria na função imunológica, redução
dos níveis de triglicerídeos e aumento da ação anti-inflamatória, estão bem documentados e são
atribuídas principalmente aos AGPI ω-3 (HORROCKS e YEO, 1999; ABBAS; MOHAMED;
JAMILAH, 2009; MILES e CALDER, 2012).
Como fontes de AGPI ω-3 de cadeia longa, temos peixes como o atum, salmão e sardinha.
Abordagens para o aumento da ingestão de AGPI ω-3 de cadeia longa incluem não só o aumento no
consumo de peixes, como também alimentos enriquecidos nesses ácidos graxos ou simplesmente
pela ingestão de cápsulas de suplementos contendo óleo de peixe (HOWE et al, 2002).
Muitos AGPIs são essenciais, uma vez que não podem ser sintetizados no organismo
humano e, portanto, tem de ser adquirido na dieta. Com base na localização da última ligação dupla
em relação à extremidade de metil terminal de um do ácido graxo, existem duas principais famílias de
AGPIs, as famílias ω-3 e ω-6 (BOURRE, 2004). Ambas as classes de ácidos graxos são importantes
em inúmeros processos relacionados à saúde humana (STILLWELL, 2006). Os potenciais benefícios
à saúde desses ácidos graxos têm sido relatados desde a descoberta de uma baixa incidência de
doenças cardiovasculares na população da Groenlândia, cuja dieta contem grande quantidade de
EPA e DHA (DYERBERG, 1989). Nas últimas três décadas, os seus benefícios têm sido
Ácido α-Linolênico ALA (18:3n3)
Figura 6: Estrutura de ácidso graxos poli-insaturados de origem vegetal e animal
Ácido Graxo Poli-insaturado da Família ω-3 de Origem Animal (peixes)
Ácido Graxo Poli-insaturado da Família ω-3 de Origem Vegetal
Ácido Eicosapentaenóico EPA (20:5n3)
Ácido Docosahexaenóico DHA (22:6n3)
Ácido Docosapentaenóico DPA (22:5n3)

33
extensivamente estudados em modelos celulares, ensaios clínicos e estudos epidemiológicos. Com
isso, inúmeros benefícios à saúde do EPA e DHA foram demonstrados em diferentes fases da vida
(MARKET RESEARCH, 2010).
Os AGPIs da família ω-3 também são muito importantes para o desenvolvimento do
cérebro, que é um órgão rico em lipídios, no qual aproximadamente dois terços do seu peso seco, são
constituídos por gorduras, particularmente AGPI de cadeia longa. Estes ácidos graxos são
incorporados principalmente nas membranas celulares e são considerados como sendo os blocos de
construção para o bom desenvolvimento do tecido cerebral. Dentre as suas funções, a principal é a
manutenção da fluidez das membranas celulares, pois isto permite que as células do cérebro possam
enviar sinais de forma mais eficiente (FOSTER; WILLIAMSON; LUNN, 2009). Durante o último
trimestre da gravidez, o cérebro passa por um período de rápido crescimento. Nele, o DHA está
presente em níveis elevados em membranas de células no cérebro e na retina, onde se acumula
rapidamente, tanto no último trimestre de gestação, quanto no primeiro ano de vida (CLANDININ
et al, 1980; SIMOPOULOS, 1991; CONNOR, 2000). Na avaliação do potencial de benefícios
cognitivos do DHA, McCann e Ames (2005) concluíram que alterações nas concentrações de DHA
cerebrais foram positivamente associadas com as mudanças no desempenho cognitivo e
comportamental.
Os AGPIs têm inúmeras funções biológicas no organismo humano, das quais se podem citar:
a redução do risco de doenças coronarianas, efeito vascular protetor, redução nos níveis de
triglicerídeos no sangue, prevenção de arritmia cardíaca, diminuição da pressão arterial, diminuição
no potencial de coagulação do sangue, melhora neurológica no desenvolvimento em crianças, efeito
estabilizador do humor, melhora em distúrbios cognitivos e disfunção imunológica, etc
(MOLENDI-COSTE; LEGRY; LECLERCQ, 2011; CONNOR, 2000; SIMOPOULOS 1991;
HORROCKS e YEO, 1999; STILLWELL, 2006; DZIEZAK, 1989).
Evidências epidemiológicas têm demonstrado que o aumento de AGPI ω-3 na dieta tem
benefícios contra o câncer (mama, cólon, próstata e pâncreas), ansiedade, nefropatia diabética,
doença inflamatória do intestino, doença de Alzheimer, bem como muitas outras (RIEDIGER et al,
2009; HUANG, 2010; ROCHA; VILARIM; NARDI, 2010; SHAPIRO et al, 2010; TURNER et al,
2010; HORROCKS e YEO, 1999; STILLWELL, 2006).
A diminuição das concentrações de AGPI ω-3 no sangue tem sido associada a várias
condições metabólicas anormais, incluindo doenças neuropsiquiátricas, esquizofrenia, Alzheimer e
depressão (ROGERS, 2001). O equilíbrio entre as famílias AGPI ω-3 e ω-6 exerce um papel
fundamental em diversos processos celulares importantes, incluindo as vias de transdução de sinal,
pelo fato de constituirem precursores de derivados biologicamente ativos, os eicosanóides (tais como

34
as prostaglandinas e leucotrienos) (Figura 5) (FLACHS et al, 2009; SIMOPOULOS, 2002). Ambas as
famílias de ácidos graxos poli-insaturados influenciam a composição da membrana celular, atuam na
sua fluidez, na mobilidade das proteínas, no potencial para a produção de hidroperóxidos, na
atividade enzimática, na ligação entre as moléculas e receptores, nas interações celulares e transporte
de nutrientes (FISCHER e BLACK, 1991; BOURRE, 2004).
Como podemos observar pela figura 5, os ácidos graxos precursores dessas famílias são
representados pelos ácidos graxo α-linolênico (C18:3n3, ALA) e ácido linoléico (C18:2n6, AL). O
ALA é convertido em ácidos graxos ω-3 de cadeia mais longa, pela ação de enzimas dessaturases
(inserem uma ligação dupla pela remoção de átomos de hidrogênio da cadeia carbônica) e alongases
(aumentam a cadeia carbônica em consequência da adição deunidades de 2 átomos de carbono). Por
outro lado, o AL é convertido a ácido araquidônico (C20:4n-6, AA). O AA é um componente de
membrana e precursor de potentes sinalizadores moleculares, tais como as prostaglandinas e os
leucotrienos.
Figura 7: Via metabólica da conversão dos ácidos graxos essenciais de cadeia longa alfa-linolênico
(série ômega-3) e linoleico (série ômega-6). Adaptado de Leonard et al, 2004.

35
O ALA pode ser metabolizado basicamente de três maneiras; sofrendo β-oxidação para
produzir energia, pode ser armazenado sob a forma de triacilglicerol e fosfolípidos das membranas
celulares; e também convertidos em ácidos graxos de cadeia mais longa do tipo ômega-3, na forma
dos ácidos eicosapentaenóico (EPA) e docosahexaenóico (DHA).
No entanto, a taxa de conversão de ALA em EPA/DHA é baixa em humanos (GOYENS et
al, 2006). Isso se deve a baixa atividade das dessaturases (∆5 e ∆6 desaturases), as quais ainda podem
ser moduladas por fatores genéticos, ambientais e co-factores dietéticos, como por exemplo: ação do
magnésio, zinco e vitamina B6 (DAS, 2005), a ingestão de outros ácidos graxos (BURDGE et al,
2003; PAWLOSKY et al, 2001; BERGER; GERSHWIN; GERMAN, 1992; LI; MANN;
SINCLAIR, 1999), de colesterol (GARG et al, 1988; LEIKIN e BRENNER, 1987) e de gordura
trans (ACKMAN e CUNNANE, 1992; HOUWELINGEN e HORNSTRA, 1994).
A variação na taxa de conversão do ALA em EPA foi estimada entre 0,2 a 6%; já para o
DHA, essa taxa não passa de 0,2% (PAWLOSKY et al, 2001, EMKEN; ADLOF; GULLEY, 1994).
Esta diferença reflete a dificuldade encontrada para a medida precisa desta conversão, evidenciando a
necessidade de pesquisas que levem a conclusões mais concretas. Esses dados levam muitos autores a
preconizar em seus trabalhos que o organismo humano não é capaz de produzir dois tipos de ácidos
graxos em quantidades suficientes.
Para efeito de comparação, os frangos podem facilmente converter o ALA em EPA e DHA.
Fato este que é amplamente explorado pelos produtores de ovos, no que tange a geração de ovos
enriquecidos em AGPI da família ω-3 (SCHEIDELER e FRONING, 1996).
Os AGPI ω-3 também desempenham um papel como precursores de mediadores lipídicos
anti-inflamatórios potentes (HUDERT et al, 2006). Estes compostos estão envolvidos no processo
de inflamação aguda, promovendo a remoção e recrutamento de fagócitos (SCHWAB et al, 2007).
Assim, os AGPI ω-3 e seus derivados são agonistas potentes essenciais no processo de inflamação
dos tecidos.
Atualmente, o mercado mundial de alimentos mostra um grande interesse nos alimentos que
fornecem não apenas os valores nutritivos, mas que também promovem benefícios à saúde, além
daqueles fornecidos pelos seus componentes inerentes. Esse segmento, que pode ser chamado de
“alimentos funcionais” está em expansão e as pesquisas científicas na área também acompanham este
crescimento.
Grande parte desse interesse se deve ao fato das tendências mundiais em relação ao uso de
aditivos naturais em alimentos. Presume-se que as substâncias (suplementos) naturais são totalmente
seguras, uma vez que a sua origem são os próprios alimentos encontrados na natureza. Logo, são
mais desejáveis do que seus homólogos sintéticos. Dentre eles, o melhor exemplo vem dos peixes e

36
óleos marinhos, que são importantes fontes de nutrientes alimentares com componentes funcionais
(TORRES-GINER et al, 2010).
Componentes bioativos eficazes que tem sido amplamente introduzido em produtos
alimentares, são os ácidos graxos poli-insaturados da família ômega-3. Isso se reflete no mercado,
onde os gastos dos consumidores globais em produtos alimentares enriquecidos em ômega-3
chegaram a US$ 13 bilhões em 2010, sendo US$ 4 bilhões somente nos Estados Unidos (Market
Research. 2010).
3.14. Dose Diária Recomendada de Ácidos Graxos Poli-insaturados da Família ω-3
Agências públicas especializadas em diversos países (França, Bélgica, Reino Unido, Holanda,
Nova Zelândia e Austrália) e organizações de saúde (FAO/OMS, American Dietetic Association e a
American Heart Association) recomendam a ingestão dietética de total de AGPI ω-3 de 1,4-2,5 g/dia,
com a mistura de EPA+DHA variando entre 140-600 mg por dia, dependendo da autoridade que
emite a diretriz. Dentre os órgãos reguladores, a FAO/OMS faz uma recomendação de 250 mg/dia,
sendo que a recomendação média, em alguns países, é de 500 mg/dia (FAO/WHO, 2008;
YASHODHARA et al, 2009; HARRIS, 2007). Isso representa menos de 2 porções de peixe (salmão,
atum, cavala e sardinha) por semana (30-40 g/dia).
À luz da literatura e da variabilidade do metabolismo, sugere-se que o mínimo de ingestão de
EPA+DHA, para os adultos saudáveis, deve ficar em torno de 0,5-1 g/dia (2-4 porções por semana
dos peixes marinhos apontados anteriormente). A ingestão diária de AGPI ω-3 de 5-6 g/dia,
apontada por Simopoulos (2006), como praticada na pré-história, foi considerada como mais
adequada ao nosso metabolismo. Tais níveis são atendidos na dieta tradicional japonesa, que contém
80-100 g/dia de peixe e marisco (YASHODHARA et al, 2009; NAGATA; TAKATSUKA;
SHIMIZU, 2002; SIMOPOULOS, 2006).
3.15. Ingestão Diária Recomendada de Ácidos Graxos Ômega-6 / Ômega-3 na Dieta
Dietas que seguem o padrão de alimentação fast food são ricas em ácidos graxos da família
ômega-6 e pobres em ácidos graxos ômega-3, resultando em uma proporção de ω-6/ω-3 em torno de
9:1 (SIMOPOULOS, 1999a; SIMOPOULOS, 1999b). Esta razão pode ser elevada para 17:1, quando
levamos em consideração a dieta ocidental de uma forma geral (SIMOPOULOS, 2001). Pessoas com
alta ingestão de carne, frituras em geral, gorduras hidrogenadas, alimentos fast food, entre outros,
fatalmente terão uma razão ω-6/ω-3 maior que a média.

37
O elevado nível atual de ácidos graxos ômega-6 fornecidos na alimentação se tornou uma
preocupação, pois as dietas ricas nesses ácidos graxos elevam os níveis do ácido araquidônico (ácido
graxo da família ω-6, C20:4n6, ARA), nos fosfolipídios de membrana, que, ao longo do tempo,
resultam em um desequilibrio na produção de eicosanoides, que por sua vez, estimulam a liberação
de citocinas inflamatórias e proteínas de fase aguda. O resultado final é a inflamação crônica de baixo
grau que contribui para problemas de saúde, tais como aterosclerose, doença de Alzheimer, câncer,
doenças cardiovasculares, síndroma metabólica, obesidade, osteoporose, diabetes tipo 2 e
periodontite (BRIGELIUS-FLOHÉ e TRABER 1999; DUQUE, 1997, RICKARD e THOMPSON,
1997; AXELSON et al, 1982). Portanto, balancear a dieta com ácidos graxos e ômega-3 e ômega-6
ajuda a diminuir o risco de desenvolvimento destas doenças crônicas.
Em vista destes fatos, serão consideradas as recomendações da Organização para a
Alimentação e Agricultura/Organização Mundial de Saúde (FAO/OMS), que preconiza uma relação
ω-6/ω-3 entre 5:1 e 10:1

38
4. MATERIAL E MÉTODOS
4.1. MATERIAIS
4.1.1. Sementes
As sementes pré-germinadas de macaúba (Acrocomia aculeata) foram compradas da empresa
Acrotech, disponível em www.acrotech.com.br. As sementes pré-germinadas de buriti (Mauritia
flexuosa L.) e não germinadas de pupunha (Bactris gasipaes Kunth) e açaí (Euterpe oleracea)
foram compradas pelo site da empresa MFRural, disponível em www.mfrural.com.br. Já as sementes
de girassol (Helianthus annuus), gergelim (Sesamum indicum), amendoim (Arachis hypogaea L.) e abacate
(Persea americana), foram adquiridas em comercio local.
As sementes pré-germinadas passaram por um período extra de germinação, em nosso
laboratório, que variou de 1 a 2 meses. Este período foi necessário para um melhor desenvolvimento
do embrião, o qual é utilizado para preparação do acetone powder. Já as sementes compradas em
mercado local foram germinadas por até 4 dias em estufa a 30 °C.
4.1.2. Preparo dos Biocatalisadores Vegetais a partir das Sementes Germinadas
Para a produção do pó bruto acetônico (acetone powder) utilizado como fonte de lipase, 100 g
de sementes (germinadas) foram trituradas em turmix com 150 mL de acetona a 4 ºC. Ao extrato
obtido foram adicionados mais 150 mL de acetona a 4 ºC, com posterior filtração em funil de
Büchner, onde o pó acetônico que fica retido no filtro é recuperado e em seguida lavado com 150
mL de acetona a 4 ºC. A ele foram adicionados mais 150 mL de acetona a 4 ºC e o mesmo foi
mantido por 24 horas sob agitação, seguindo-se de nova lavagem com 150 mL de acetona a 4 ºC. O
pó desengordurado (biocatalisador) foi mantido em recipiente aberto em capela por 24 horas para a
evaporação da acetona residual, e depois armazenado em geladeira até à utilização (Cavalcanti et al,
2007).
O Preparado Enzimático Vegetal (PEV) é obtido apenas com a maceração da semente
germinada, que sem sofrer nenhum outro tipo de tratamento, é arrmazenada em geladeira até à
utilização.
4.1.3. Lipases Microbianas

39
Para o estudo desenvolvido nesta tese, 11 lipases comerciais foram utilizadas na síntese dos
lipídios estruturados, das quais 5 são sn-1,3 regioespecífica, sendo elas: (i) Lipozyme® RM IM
(Rhizomucor miehei), (ii) Lipozyme® TL IM (Thermomyces lanuginosus) e (iii) Lipozyme® TL 100L
(Thermomyces lanuginosus) adquiridas da Novozymes; (iv) Rhizomucor miehei e (v) Rhizopus oryzae IM
obtidas da Sigma-Aldrich.
As 6 lipases não específicas utilizadas foram as seguintes: (vi) Novozym® 735 (Candida
antarctica A) e (vii) Novozym® 435 (Candida antarctica B), adquiridas Novozymes; (viii) Lipomod™ 34P
(Candida rugosa) adquirida da Biocatalysts; (ix) Amano AY 30 (Candida rugosa), (x) Amano AK
(Pseudomonas fluorescens), (xi) Amano PS (Burkholderia cepacia) adquiridas da Sigma-Aldrich.
Para o estudo das lipases próprias ou home made, foram aplicadas as lipases:
(xii) Lip. B – lipase recombinante de Candida antarctica expressa em Pichia pastoris (Patente: PI
0905122-8), denominada de Lip. B LaBiM;
(xiii) ROL – lipase recombinante de Rhizopus oryzae expressa em Pichia pastoris gentilmente
cedida pelo professor Francisco Valero do Dept. Enginyeria Química (ETSE) da Universitat
Autònoma Barcelona (Espanha) em trabalho de cooperação internacional, denominada de ROL
UAB. Esta lipase foi produzida por uma cultura de uma cepa de P. pastoris recombinante usando
metanol como indutor. A biomassa foi separada a partir do caldo de cultura por centrifugação e
microfiltração. O sobrenadante foi concentrado através de ultrafiltração com um sistema
Centrasette® Pall Filtron equipado com uma membrana Omega de 10 kDa, sendo subsequentemente
dialisadas contra tampão de Tris-HCl 10 mM a pH 7.5 e, finalmente, liofilizada. O teor médio de
proteína em pó liofilizado foi de 3,4% (p/p), Segundo metodologia descrita por Arnau et al, 2010.
(xiv) Rhizomucor miehei - lipase obtida a partir da Fermentação em Estado Sólido (FES),
adaptado de Santos (2010), aplicada na forma imobilizada, denominada de RM IM LaBiM;
O preparado enzimático sólido (PES) liofilizado de Rhizomucor miehei foi obtido a partir da
fermentação do micro-organismo em torta de babaçu moída e peneirada (partículas com tamanho
inferior a 1,18 mm). As fermentações foram conduzidas em biorreatores contendo 15 g de torta com
umidade ajustada para 65%. Cada becher foi esterelizado a 121°C por 15 minutos por autoclavação e
a torta de babaçu foi inoculada com 107 esporos por grama de sólido (peso seco) e incubada a 30°C
em estufa com injeção de ar umidificado. O tempo de fermentação foi de 72 horas. Ao final da
fermentação, o meio fermentado foi liofilizado e estocado a 4°C. Ao final da fermentação, foram
adicionados à torta fermentada 5 ml de tampão fosfato de sódio (5 mM, pH 7,0) por grama de massa
seca. A extração das enzimas foi realizada em agitador rotatório à temperatura de 35°C e velocidade
de agitação de 200 rpm por 20 minutos. Posteriormente, a torta foi prensada para a separação da fase
líquida e esta, por sua vez, foi submetida a centrifugação à 3000 rpm por 5 minutos para remoção

40
dos sólidos mais finos. O sobrenadante, chamado de extrato bruto, foi utilizado para medir a
atividade lipásica. O extrato enzimático, livre de células, produzidos por FES foi tratado com sulfato
de amônio em saturação (40% e 80%). Ao extrato bruto foi adicionado à quantidade de sal para
obtenção de concentração a 20% da saturação e mantido sob agitação lenta/ constante e banho de
gelo. Logo após esta solução foi centrifugada à 4oC por 20 minutos a 10000 rpm. O precipitado
obtido foi ressupenso num volume menor de tampão fosfato de sódio 50mM pH 7,0 de forma a
concentrar a enzima. O sobrenadante foi utilizado para nova precipitação com saturação de 40% e
80% de sulfato de amônio e todo o protocolo repetido para estas faixas. Na imobilização, o
tratamento do suporte foi feito da seguinte forma: A 1g de suporte (ACCUREL) adicionar 10 ml de
etanol sob agitação por 30 min à temperatura ambiente. Depois da retirada do etanol com auxílio de
uma pipeta, adicionar 10 ml de água destilada. Esperar 10 minutos e lavar exaustivamente o suporte
com água destilada até retirar todo o etanol. Para a imobilização, a 1g de suporte adicionar 10 ml de
solução enzimática à temperatura ambiente overnight, sob agitação. Filtrar e lavar o ACCUREL
MP1000 com Tampão fosfato 5mM pH 7 para retirar a enzima que não foi imobilizada. Secar o
ACCUREL MP1000 em dessecador sob vácuo até o IME ficar completamente seco (+ 3 dias).
4.1.4. Óleos Vegetais e de Peixe
Os óleos vegetais de coco (COPRA), palma (Hemmer) e de oliva (Azeite GALLO) foram
adquiridos em mercado local e o óleo de peixe foi gentilmente cedido pelo Dr.Ricardo Martino da
Fundação Instituto de Pesca do Estado do Rio de Janeiro (FIPERJ).
4.2. MÉTODOS ANALÍTICOS
4.2.1. Reação de Hidrólise com o Preparado Enzimático Vegetal (PEV)
As reações de hidrólise foram conduzidas com 5% (p/v) de PEV (com diferentes dias de
germinação), 30% de substrato (óleo de soja) em tampão 0.1 M fosfato de sódio pH 7,0 em reator
termostatizado sob agitação magnética, à 35 ºC por 24 horas.
4.2.2. Reação de Hidrólise com o Acetone Powder
As reações de hidrólise foram conduzidas com 2,5% (p/v) de acetone powder das sementes
germinadas, 25% de substrato (óleo de soja) em meio tamponado (acetato de sódio pH 4,0; fosfato
de sódio pH 7,0 e bicarbonato de sódio pH 10,0) na concentração 0.1 M em reator termostatizado

41
sob agitação magnética à 35 °C por até 48 horas. Em ensaios específicos de reações de hidrólise foi
utilizado 5,0 % (p/v) dos emulsificantes Triton X-100 e Goma arábica de apenas com a utilização de
tampão fosfato de sódio pH 7,0.
4.2.3. Reação de Esterificação
As reações de esterificação foram conduzidas com 2,5% (p/v) de acetone powder das sementes
germinadas tendo com substratos o glicerol (C3H8O3) e o ácidos oléico (C18:1n9) ou ácido caprílico
(C8:0) na razão molar de 1:3 em reator termostatizado sob agitação magnética, à 35 ºC por até 48
horas.
4.2.4. Determinação da Acidez nos Óleos
A aicdez foi determinada retirando-se alíquotas do meio reacional (100 µL, em triplicata) para
a medida de acidez do meio reacional. As reações foram interrompidas pela adição de 20,0 mL uma
solução de acetona-etanol (1:1 v/v). O teor de ácidos graxos livres foi determinado por titulometria
de neutralização empregando NaOH 0.04 mol.L-1, em titulador automático Mettler Toledo modelo
DG20 até um valor de pH final de 11.0. Os brancos reacionais, neste caso, são obtidos no tempo
zero da reação, adicionando-se o acetone powder após a solução acetona-etanol. A Equação (1) descreve
o cálculo empregado para determinação da acidez do meio reacional.
m
MMNVbVaAcidez
×
××−=
10
)((%) (Equação 1)
Onde:
Va = volume de solução de NaOH gasto para a titulação da amostra (mL); Vb = volume de solução
de NaOH gasto para a titula ção do branco (mL); N = normalidade da solução de NaOH (mol.L-1);
MM = massa molar do ácido graxo predominante no óleo; m = massa de amostra (g).
4.2.5. Determinação da Atividade de Esterificação
Com relação à avaliação da atividade de esterificação do acetone powder (AP) das sementes
germinadas, as reações foram conduzidas com 2,5 % (p/v) de AP, tendo como substrato o ácido
oléico (C18:1n9) e o álcool etílco (C2H60) na razão molar (1:1) em reator termostatizado sob agitação
magnética, à 35 ºC por 15 min. O progresso da reação foi acompanhado retirando-se alíquotas do

42
meio reacional (100 µL, em triplicata) e analisadas por titulometria de neutralização. As amostras
foram dissolvidas em 40 mL de acetona/etanol (1:1, v/v) e tituladas contra uma solução de NaOH a
0.04 mol.L-1, empregando titulador automático Mettler Toledo modelo DG20. Uma unidade de
atividade corresponde a quantidade de biocatalisador que leva ao consumo de 1 µmol de ácido graxo
livre por minuto nas condições de ensaio, segundo metodologia descrita por Langone & Sant’anna
(1999) . A Equação (2) descreve o cálculo empregado para determinação da atividade enzimática de
esterificação.
Va
Vm
mt
MVfVigUçãoesterificadeAtividade ××
×
×−= 1000
)()/( (Equação 2)
Onde:
Vi = volume de NaOH consumido na titulação da amostra retirada no tempo zero de reação (mL);
Vf = volume de NaOH consumido na titulação da amostra retirada no tempo t de reação (mL); M =
concentração da solução de NaOH (mol.L-1); t = tempo de reação (min); m = massa de preparação
enzimática utilizada na reação (g); Vm = volume de meio reacional (mL); Va = volume de amostra
(mL).
4.2.6. Reação de Hidrólise do Óleo de Peixe
As reações de hidrólise do óleo de peixe foram conduzidas com a lipase de Candida rugosa
(Lipomod 34P) 5% (p/v), 50% de substrato em tampão 0.1 M fosfato de sódio pH 7.0 em reator
termostatizado sob agitação mecânica, à 40 ºC por 48 horas. A reação foi paralisada mediante a
extração da fase orgânica com hexano, em funil de separação. Foi adicionado sulfato de sódio anidro
para retirar possíveis vestígios de água do produto, que foi em seguida filtrado e concentrado em
rotaevaporador. Para evaporação de qualquer vestígio do solvente no produto, o mesmo foi
incubado overnigth em estufa a 70 ºC.
4.2.7. Concentração dos Ácidos Graxos Poli-insaturados do Óleo de Peixe Hidrolisado com Uréia
O método de fracionamento com complexação (inclusão) com uréia baseia-se na separação
pelo grau de insaturação, pois os ácidos graxos insaturados tendem a não se complexar com a uréia e
não formar precipitados. Quando a uréia é cristalizada na presença de compostos alifáticos de cadeia
longa, ela forma cristais hexagonais com um canal, no qual os compostos alifáticos podem se alojar

43
desde que não contenham grupos funcionais que aumentam a sua massa molar. Tais cristais são
conhecidos como complexos de inclusão de uréia (CHRISTIE, 1989).
A técnica de complexação com uréia é frequentemente utilizada para a concentração de
ácidos graxos poli-insaturados, em especial, da família ômega-3, a partir de uma variedade de óleos
de origem marinha, tais como: cavala, savelha, arenque, salmão, gordura de foca, lobo marinho, etc
(WANASUNDARA e SHAHIDI, 1999; SENANAYAKE e SHAHIDI, 2000). Esta técnica foi
baseada na metodologia descrita por Gámez-Meza et al, 2003 (apud WANASUNDARA e
SHAHIDI, 1999), na qual os ácidos graxos livres (15 g) provenientes da hidrólise do óleo de peixe
são misturados com uréia (25 g) em etanol 95 % (100 mL) e aquecidos com agitação até que a
mistura torne-se uma solução límpida e homogênea. Esta solução foi transferida para tubos de
centrífuga e rapidamente resfriada por imersão em água gelada, depois mantida refrigerada a 5 ºC por
8 horas. Os cristais foram removidos por centrifugação (6.000 x g) por 20 min a 5 ºC. O
sobrenadante, contendo os ácidos graxos poli-insaturados não complexados, foi mantido a -5 ºC por
12 horas, em seguida, centrifugado novamente (6.000 x g) a -5 ºC por 20 min, sendo posteriormente
acidificado a pH 4,0 com HCl 0.05 M. Em seguida, 100 mL de solução água-hexano (1:1) a 65 ºC foi
adicionada e mantida sob agitação por 30 min. Posteriormente, as fases foram separadas e o solvente
evaporado. O concentrado de ácidos graxos poli-insaturados foi armazenado em freezer a -20 ºC.
4.2.8. Reação de Acidólise dos Óleos Vegetais com os Ácidos Graxos Poli-insaturados
As reações de enriquecimento nutricional foram realizadas pela acidólise dos óleos vegetais
(coco, palma e oliva) com os ácidos graxos provenientes do óleo de peixe previamente hidrolisado e
concentrado segundo o item 4.2.7. As reações de acidólise foram conduzidas com 2,5% (p/v) de
biocatalisador* (lipase), tendo como substrato o óleo vegetal e os ácidos graxos livres na razão molar
1:3, em reator termostatizado sob agitação magnética à 40 °C por 24 horas. Ao final deste tempo, as
reações foram centrifugadas a 10.000 rpm por 5 min para a remoção do biocatalisador e congeladas
em freezer para posterior análise. As reções foram realizadas em duplicata e os ensaios em brancos
conduzidos sem a presença do biocatalisador.
* A atividade específica de esterificação de cada biocatalisador utilizado foi mensurada conforme o
ítem 4.2.5. Devido à disparidade na atividade entre as lipases, a quantidade de biocatalisador utilizado
nas reações não pode ser padronizada por atividade especifica, pois a reação se tornaria, na prática,
inviável devido à massa de biocatalisador que deveria ser utilizada para equiparar a atividade de

44
esterificação em todas as reações de acidólise. Por isso a quantidade de biocatalisador utilizado foi
expressa em percentual em relação ao substrato utilizado.
4.2.8. Reação de Acidólise da Tripalmitina com Ácido Oléico para Produção de Miméticos
do Leite Materno
As reações para produção de miméticos do leite materno foram conduzidas com 5,0% (p/p)
de biocatalisador (lipase), tendo como substrato a tripalmitina (C51H98O6) e o ácido oléico (C18:1n9)
na razão molar 1:6, em reator termostatizado sob agitação magnética à 60 °C por 24 horas. As reções
foram realizadas em duplicata e os ensaios em brancos conduzidos sem a presença do biocatalisador.
4.2.9. Reação de Acidólise do Óleo de Peixe com Ácido Octanóico para Produção de
Lipídios de Baixo Teor Calórico
As reações para produção de lipídios de menor densidade energética foram conduzidas com
2,5% (p/v) de biocatalisador (lipase), tendo como substrato o óleo de peixe e o ácido octanóico
(C8:0) na razão molar 1:2, em reator termostatizado sob agitação magnética à 40 °C por 24 horas. Ao
final deste tempo, as reações foram centrifugadas a 10.000 rpm por 5 min para a remoção do
biocatalisador e congeladas em freezer para posterior análise. As reções foram realizadas em
duplicata e os ensaios em brancos conduzidos sem a presença do biocatalisador.
4.2.10. Separação do Produto por Cromatografia em Camada Delgada (TLC)
O produto das reações de interesterificação (acidólise) foi aplicado em placas cromatográficas
de camada delgada (TLC) revestidas com sílica gel 60 Å com identificador de fluorescência F254 (20
x 20 cm, 1 mm de espessura de camada). Alíquotas de 4 a 10 µL foram aplicadas por centímetro de
placa e a mistura de solventes de desenvolvimento (fase móvel) foi composta por hexano/éter
etílico/ácido acético (70:30:1, v/v/v). Após a eluição, as placas foram pulverizadas com solução 0,2
% de 2,7-diclorofluoresceína em metanol e visualizados sob luz ultravioleta (UV). A banda referente
aos triacilgliceróis, devidamente identificada utilizando-se a trioleína como padrão, foi raspada para
análise da composição em ácidos graxos. As demais bandas, referentes aos diacilgliceróis,
monoacilgliceróis e ácidos graxos também puderam ser identificadas com a utilização de padrão
específico para cada família.

45
4.2.11. Metilação dos Ácidos Graxos para Análise por Cromatografia Gasosa
As amostras foram descongeladas em banho maria a 37 ºC e homogeneizadas. 8,0 µL das
amostras oleosas e/ou 100 a 200 mg de sílica (proveniente da TLC), foram transferidas para tubos de
hidrólise, no qual também foram adicionados 3,0 mL da solução de metanol:hexano (4:1 v/v) e em
seguida 300 µL de cloreto de acetila (catalisador) sob agitação suave. Os tubos foram fechados em
atmosfera de N2, seguindo-se o aquecimento em banho com agitação a 100 ºC por 1 hora; em
seguida, os tubos foram resfriados em água. Adicionou-se lentamente 3,0 mL de KHCO3 (10 % p/v)
para paralisar a reação e neutralizar os solventes seguindo-se a agitação em vórtex por 30 segundos e
centrifugação a 2.000 rpm por 10 min. Após centrifugação, a fase superior (orgânica) foi então
transferida cuidadosamente para frascos de armazenagem de 2,0 mL e as amostras foram analisadas
em cromatógrafo a gás. As análises foram feitas em triplicata e o ensaio em branco foi realizado em
paralelo com água deionizada (LEPAGE e ROY, 1986).
4.2.12. Análise da Composição em Ácidos Graxos por Cromatografia Gasosa
Os ácidos graxos metilados, segundo metodologia descrita no ítem 4.9, foram analisados em
cromatógrafo gasoso Shimadzu GCMS-QP2010 em uma coluna Omegawax 320 Supelco (30 m x
0,32 mm x 0,25 µm) com detector de ionização de chama (FID). As temperaturas do injetor e do
detector foram de 260 e 280 °C, respectivamente. As condições utilizadas durante a cromatografia
foram as seguintes: a temperatura inicial da coluna foi de 150 ºC, chegando a 210 ºC no final da
corrida. A temperatura do forno foi mantida a 150 ºC por 5 min, seguida por um aumento até 210
ºC, com uma rampa de 2 ºC/min. A corrida foi mantida isotermicamente por 30 min. Foi aplicada a
razão de split: 1:20, o volume de injeção foi de 1 µL, e o gás de arraste foi o hélio (1,1 ml / min). Os
resultados médios dos ácidos graxos foram calculados em mol%.
A identificação dos ácidos graxos foi determinado pela comparação com a solução padrão
comercial contendo 37 ésteres metílicos de ácidos graxos (Supelco® 37 FAME Mix) e com a solução
PUFA No.3 (menhaden oil), usadas como padrão externo, ambas adquiridas da Sigma-Aldrich.
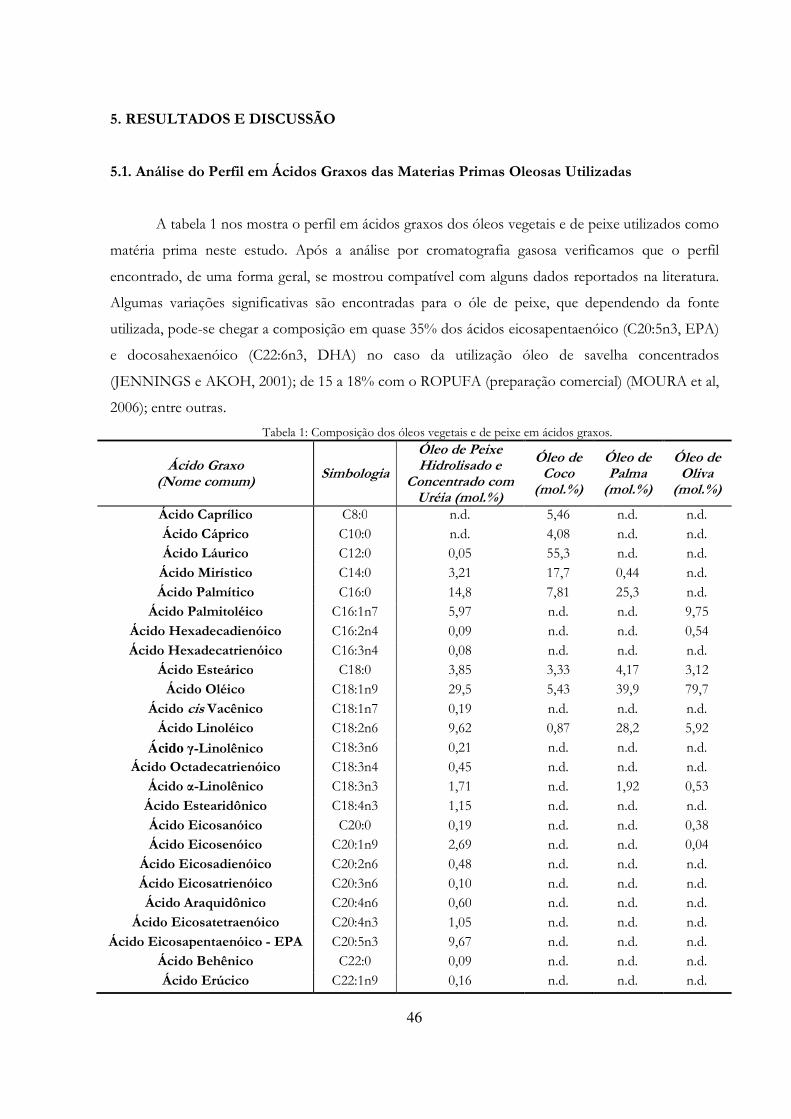
46
5. RESULTADOS E DISCUSSÃO
5.1. Análise do Perfil em Ácidos Graxos das Materias Primas Oleosas Utilizadas
A tabela 1 nos mostra o perfil em ácidos graxos dos óleos vegetais e de peixe utilizados como
matéria prima neste estudo. Após a análise por cromatografia gasosa verificamos que o perfil
encontrado, de uma forma geral, se mostrou compatível com alguns dados reportados na literatura.
Algumas variações significativas são encontradas para o óle de peixe, que dependendo da fonte
utilizada, pode-se chegar a composição em quase 35% dos ácidos eicosapentaenóico (C20:5n3, EPA)
e docosahexaenóico (C22:6n3, DHA) no caso da utilização óleo de savelha concentrados
(JENNINGS e AKOH, 2001); de 15 a 18% com o ROPUFA (preparação comercial) (MOURA et al,
2006); entre outras.
Tabela 1: Composição dos óleos vegetais e de peixe em ácidos graxos.
Ácido Graxo (Nome comum) Simbologia
Óleo de Peixe Hidrolisado e
Concentrado com Uréia (mol.%)
Óleo de Coco (mol.%)
Óleo de Palma (mol.%)
Óleo de Oliva (mol.%)
Ácido Caprílico C8:0 n.d. 5,46 n.d. n.d. Ácido Cáprico C10:0 n.d. 4,08 n.d. n.d. Ácido Láurico C12:0 0,05 55,3 n.d. n.d. Ácido Mirístico C14:0 3,21 17,7 0,44 n.d. Ácido Palmítico C16:0 14,8 7,81 25,3 n.d. Ácido Palmitoléico C16:1n7 5,97 n.d. n.d. 9,75
Ácido Hexadecadienóico C16:2n4 0,09 n.d. n.d. 0,54 Ácido Hexadecatrienóico C16:3n4 0,08 n.d. n.d. n.d.
Ácido Esteárico C18:0 3,85 3,33 4,17 3,12 Ácido Oléico C18:1n9 29,5 5,43 39,9 79,7
Ácido cis Vacênico C18:1n7 0,19 n.d. n.d. n.d. Ácido Linoléico C18:2n6 9,62 0,87 28,2 5,92
Ácido γ-Linolênico C18:3n6 0,21 n.d. n.d. n.d. Ácido Octadecatrienóico C18:3n4 0,45 n.d. n.d. n.d. Ácido α-Linolênico C18:3n3 1,71 n.d. 1,92 0,53 Ácido Estearidônico C18:4n3 1,15 n.d. n.d. n.d. Ácido Eicosanóico C20:0 0,19 n.d. n.d. 0,38 Ácido Eicosenóico C20:1n9 2,69 n.d. n.d. 0,04 Ácido Eicosadienóico C20:2n6 0,48 n.d. n.d. n.d. Ácido Eicosatrienóico C20:3n6 0,10 n.d. n.d. n.d. Ácido Araquidônico C20:4n6 0,60 n.d. n.d. n.d.
Ácido Eicosatetraenóico C20:4n3 1,05 n.d. n.d. n.d. Ácido Eicosapentaenóico - EPA C20:5n3 9,67 n.d. n.d. n.d.
Ácido Behênico C22:0 0,09 n.d. n.d. n.d. Ácido Erúcico C22:1n9 0,16 n.d. n.d. n.d.
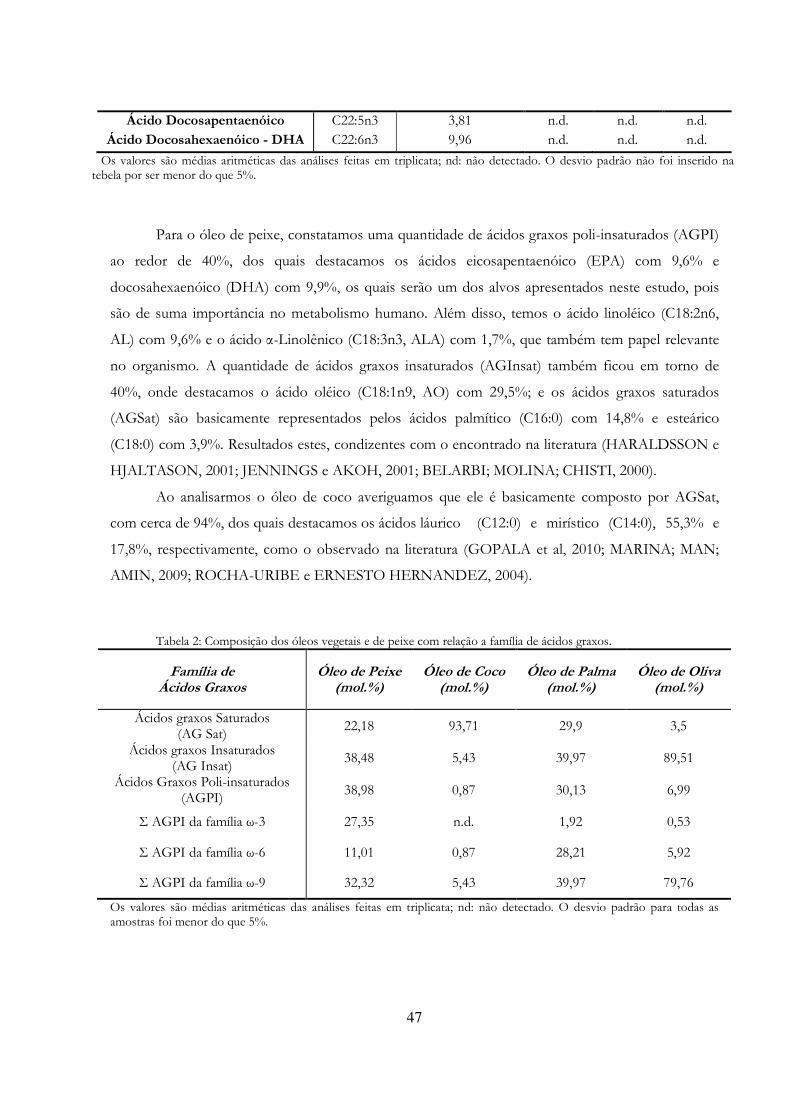
47
Ácido Docosapentaenóico C22:5n3 3,81 n.d. n.d. n.d. Ácido Docosahexaenóico - DHA C22:6n3 9,96 n.d. n.d. n.d.
Os valores são médias aritméticas das análises feitas em triplicata; nd: não detectado. O desvio padrão não foi inserido na tebela por ser menor do que 5%.
Para o óleo de peixe, constatamos uma quantidade de ácidos graxos poli-insaturados (AGPI)
ao redor de 40%, dos quais destacamos os ácidos eicosapentaenóico (EPA) com 9,6% e
docosahexaenóico (DHA) com 9,9%, os quais serão um dos alvos apresentados neste estudo, pois
são de suma importância no metabolismo humano. Além disso, temos o ácido linoléico (C18:2n6,
AL) com 9,6% e o ácido α-Linolênico (C18:3n3, ALA) com 1,7%, que também tem papel relevante
no organismo. A quantidade de ácidos graxos insaturados (AGInsat) também ficou em torno de
40%, onde destacamos o ácido oléico (C18:1n9, AO) com 29,5%; e os ácidos graxos saturados
(AGSat) são basicamente representados pelos ácidos palmítico (C16:0) com 14,8% e esteárico
(C18:0) com 3,9%. Resultados estes, condizentes com o encontrado na literatura (HARALDSSON e
HJALTASON, 2001; JENNINGS e AKOH, 2001; BELARBI; MOLINA; CHISTI, 2000).
Ao analisarmos o óleo de coco averiguamos que ele é basicamente composto por AGSat,
com cerca de 94%, dos quais destacamos os ácidos láurico (C12:0) e mirístico (C14:0), 55,3% e
17,8%, respectivamente, como o observado na literatura (GOPALA et al, 2010; MARINA; MAN;
AMIN, 2009; ROCHA-URIBE e ERNESTO HERNANDEZ, 2004).
Tabela 2: Composição dos óleos vegetais e de peixe com relação a família de ácidos graxos.
Família de Ácidos Graxos
Óleo de Peixe (mol.%)
Óleo de Coco (mol.%)
Óleo de Palma (mol.%)
Óleo de Oliva (mol.%)
Ácidos graxos Saturados (AG Sat) 22,18 93,71 29,9 3,5
Ácidos graxos Insaturados (AG Insat)
38,48 5,43 39,97 89,51
Ácidos Graxos Poli-insaturados (AGPI) 38,98 0,87 30,13 6,99
Σ AGPI da família ω-3 27,35 n.d. 1,92 0,53
Σ AGPI da família ω-6 11,01 0,87 28,21 5,92
Σ AGPI da família ω-9 32,32 5,43 39,97 79,76
Os valores são médias aritméticas das análises feitas em triplicata; nd: não detectado. O desvio padrão para todas as amostras foi menor do que 5%.

48
Para o óleo de palma detectamos um equilíbrio entre as famílias de ácidos graxos (Tabela 2).
Dos 30% de AGSat presentes no óleo, 25% correspondem ao ácido palmítico (C16:0); todo o
percentual de AGInsat (40%) corresponde ao ácido oléico (C18:1n9); e para os 30% de AGPI, temos
28% só de ácido linoléico (C18:2n6), como o observado na literatura (UNDURRAGA;
MARKOVITS; ERAZO, 2001; EMBRANDIRI; IBRAHIM; SINGH, 2013; FAJARDO; AKOH;
LAI, 2003).
Com relação ao óleo de oliva, nota-se que ele é essencialmente formado por AGInsat. Eles
correspondem a 90% da composição do óleo, sendo 80% de ácido oléico (C18:1n9) e 9,8% de ácido
palmitoléico (C16:1n7), como descrito por (BOSKOU, 1996; PHAM e PHAM, 2012; GALTIER et
al, 2008).
Com esta análise foi possível confirmar a composição distinta dos diferentes óleos no que diz
respeito à composição em ácidos graxos. A caracterização do substrato é essencial para prever ou
explicar a atuação dos biocatalisadores (lipases) testados neste trabalho. Pois a especificidade
apresentada por algumas lipases frente a diferentes substratos pode explicar os resultados obtidos
para um tipo de óleo ou reação específica.
5.2 Quantificação da Atividade de Esterificação dos Biocatalisadores
Nas tabelas 3 e 4, estão apresentados os dados de atividade de esterificação utilizando como
substrato o ácido oléico (C18:1n9) e o álcool etílico (C2H6O) para as lipases comerciais e próprias,
respectivamente.
Observa-se uma disparidade entre as atividades dos biocatalisadores comerciais
comparativamente aos próprios. Os valores muito inferiores observados nos últimos (Tabela 4) já
eram esperados visto que essas lipases próprias não sofreram melhoramento genético comumente
realizado para obtenção de biocatalisadores comerciais.
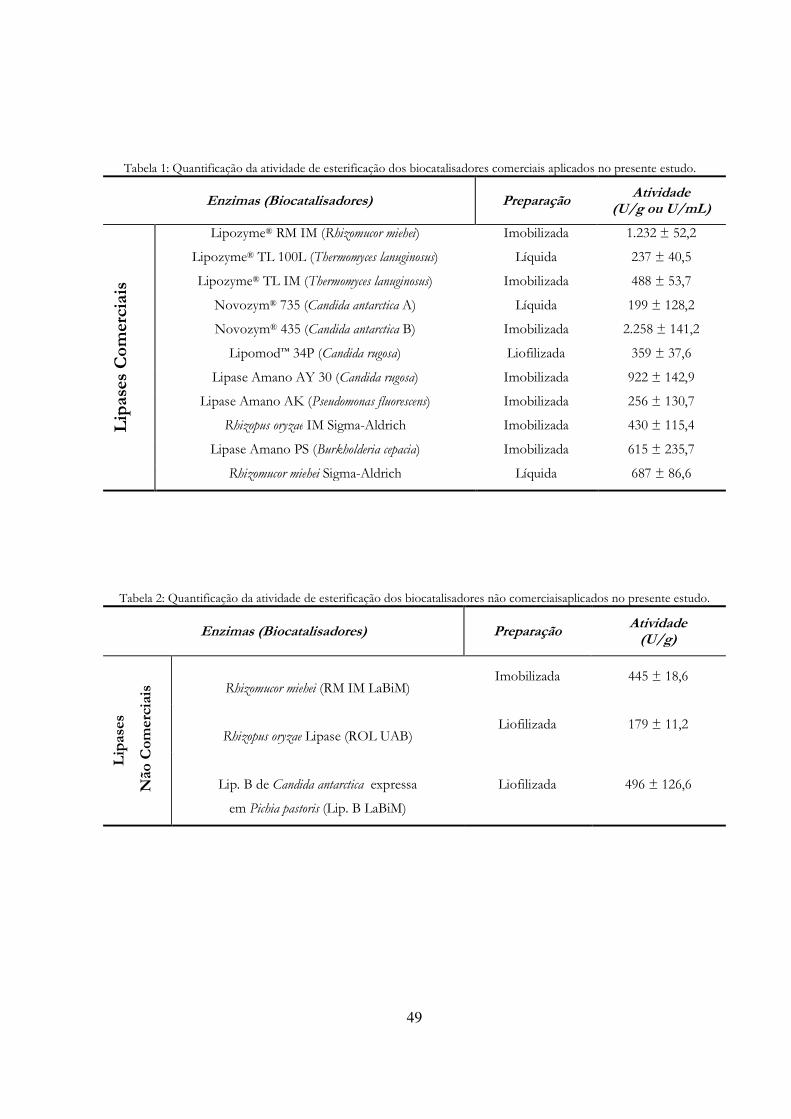
49
Tabela 1: Quantificação da atividade de esterificação dos biocatalisadores comerciais aplicados no presente estudo.
Enzimas (Biocatalisadores) Preparação Atividade (U/g ou U/mL)
Lipases Comerciais
Lipozyme® RM IM (Rhizomucor miehei) Imobilizada 1.232 ± 52,2
Lipozyme® TL 100L (Thermomyces lanuginosus) Líquida 237 ± 40,5
Lipozyme® TL IM (Thermomyces lanuginosus) Imobilizada 488 ± 53,7
Novozym® 735 (Candida antarctica A) Líquida 199 ± 128,2
Novozym® 435 (Candida antarctica B) Imobilizada 2.258 ± 141,2
Lipomod™ 34P (Candida rugosa) Liofilizada 359 ± 37,6
Lipase Amano AY 30 (Candida rugosa) Imobilizada 922 ± 142,9
Lipase Amano AK (Pseudomonas fluorescens) Imobilizada 256 ± 130,7
Rhizopus oryzae IM Sigma-Aldrich Imobilizada 430 ± 115,4
Lipase Amano PS (Burkholderia cepacia) Imobilizada 615 ± 235,7
Rhizomucor miehei Sigma-Aldrich Líquida 687 ± 86,6
Tabela 2: Quantificação da atividade de esterificação dos biocatalisadores não comerciaisaplicados no presente estudo.
Enzimas (Biocatalisadores)
Preparação Atividade (U/g)
Lipases
Não Comerciais
Rhizomucor miehei (RM IM LaBiM) Imobilizada 445 ± 18,6
Rhizopus oryzae Lipase (ROL UAB) Liofilizada 179 ± 11,2
Lip. B de Candida antarctica expressa
em Pichia pastoris (Lip. B LaBiM)
Liofilizada 496 ± 126,6

50
5.3. Avaliação do Potencial dos Biocatalisadores Vegetais
5.3.1. Macaúba (Acrocomia aculeata) e Buriti (Mauritia flexuosa L.)
5.3.1.1. Germinação das Sementes de Macaúba e Buriti
As sementes de macaúba foram adquiridas na forma pré-germinadas com empresa Acrotech;
já as de buriti, com empresa MFRural. Ao chegarem ao nosso laboratório (Figura 8a), passaram por
um período extra de germinação, que variou de 1 a 2 meses, dependendo do estágio de
desenvolvimento do embrião ou broto, o qual é utilizado como matéria prima para obtenção do
acetone powder. Para um melhor acompanhamento deste estágio, as sementes foram cortadas
longitudinalmente. Pela figura 8b podemos verificar o estágio final de crescimento do broto após 2
meses de germinação em laboratório, apartir do qual o acetone powder foi preparado, conforme
metodologia descrita no ítem 4.1.2. O mesmo procedimento foi adotado com as sementes de buriti.
5.3.1.2. Avaliação do Potencial Hidrolítico e de Esterificação
Os testes para a avaliação do potencial hidrolítico foram feitos aplicando o acetone powder das
sementes germinadas no óleo de soja (substrato) em meio tamponado. Como não se tinha nenhuma
informação prévia descrita na literatura sobre uma possível lipase presente na semente da
macaúba/buriti, o teste inicial foi realizado com três diferentes tampões: acetato de sódio (pH 4),
fosfato de sódio (pH 7) e bicarbonato de sódio (pH 10). Em nenhum dos três casos foi obtido um
Figura 8: (a) Semente germinada de macaúba no estagio de germinação que é fornecida pela empresa Acrotech. (b) Corte transversal da semente após um período extra de 1 a 2 meses de germinação em laboratório.
Figura (a) Figura (b)
1 cm 2 cm

51
resultado satisfatório, tendo em vista que a acidez, ao final de 24 horas de reação não ultrapassou 2%
(Tabela 5).
Apesar destes resultados, ensaios de hidrólise com a utilização de emulsificantes, tais como o
Triton X-100 (detergente não iônico) e a goma arábica (resina natural), foram realizados em meio
tamponado no pH 7 (fosfato de sódio). Esses compostos, ao funcionarem como emulsificantes,
atuam facilitando a ação das lipases no meio reacional, uma vez que aumentam a solubilidade do
substrato, tornando-os mais acessíveis na interface lipídio-água. Eles também podem atuar
dispersando a possível lipase que se encontra naturalmente imobilizada no acetone powder. Com este
teste, também não foi possível obter nenhum resultado satisfatório, tendo em vista que no melhor
resultado de hidrólise, a acidez do óleo de soja não passou de 2,5% (Tabela 5).
Tabela 3: Potencial catalítico para hidrólise do acetone powder da semente germinada de macaúba e de buriti.
Reações de Hidrólise Acetone powder Macaúba
Acidez (%) Acetone powder Buriti
Acidez (%) pH 4 (Acetato de sódio 0.1M) 1,4 ± 0,02 0,8 ± 0,09 pH 7 (Fosfato de sódio 0.1M) 2,0 ± 0,09 1,0 ± 0,11
pH 10 (Bicarbonato de sódio 0.1M) 2,0 ± 0,20 0,8 ± 0,17 pH 7 com Triton X-100 2,5 ± 0,14 - pH 7 com Goma Arábica 1,5 ± 0,07 -
As reações de hidrólise foram conduzidas com 2,5% de acetone powder (p/v) da semente germinada de macaúba/buriti, 25% de substrato (óleo de soja, com acidez de 0,8 ± 0,05%) nos tampões (acetato de sódio pH 4.0; fosfato de sódio pH 7.0 e bicarbonato de sódio pH 10.0) na concentração 0.1 M, incubados a 35 °C em reator termostatizado sob agitação magnética por 24 horas. Os ensaios com a utilização dos emulsificantes Triton X-100 e Goma arábica foram realizados, ambos na concentração de 5 % (p/v) apenas com a utilização de tampão fosfato de sódio pH 7.0, 0.1M.
Para os ensaios de avaliação do potencial de esterificação do acetone powder das sementes
germinadas de macaúba e do burit foram utilizados como substrato, glicerol (C3H8O3), o ácido oléico
(C18:1n9), ácido graxo insaturado de cadeia longa comumente encontrado em óleos vegetais e o
ácido caprílico (C8:0), ácido graxo saturado de cadeia média. Neste caso também não foram obtidos
resultados satisfatórios tendo em vista uma redução bem discreta na acidez ao final da reação. Foi
possível reduzir a acidez inicial de 98 ± 1,25% para 94,7 ± 0,20 e 96,0 ± 0,84, com a utilização do
ácido oléico e caprílico, respectivamente, para a macaúba. Utilizando o buriti, esta redução para 65,5
± 3,53 e 84,0 ± 1,41, com a utilização do ácido oléico e caprílico, respectivamente. Estes valores,
aparentemente, foram considerados animadores, por se tratar de um teste inicial sem qualquer tipo
de otimização. Porém, quando se tentou quantificar a atividade de esterificação do ácido oléico com
o álcool etílico, os resultados encontrados não foram conclusivos e foram muito dispersos, indicando

52
que a metodologia para a obtenção do acetone powder pode não ter sido efetiva ou apresentado
reprodutibilidade.
Deste modo pode-se supor que este método de preparação do biocatalisador, de alguma
forma, inativou a lipase que deve ter sido produzida durante a etapa de germinação da semente, ou
que a atividade lipásica pode ser muito baixa, tendo em vista o longo tempo necessário para a
germinação de palmáceas. Haja visto que para o desenvolvimento e crescimento do embrião (Figura
8b) observou-se simultaneamente a redução do tecido lipídico que deve ter sido utilizado como fonte
de energia. Não se deve descartar também a hipótese da possível lipase ser solúvel em acetona, como
é observado no caso da bromelina.
5.3.2. Girassol (Helianthus annuus), Gergelim (Sesamum indicum) e Amendoim (Arachis
hypogaea L.)
5.3.2.1. Germinação das Sementes de Girassol/Gergelim/Amendoim
As sementes de girassol, gergelim e amendoim foram adquiridas em mercado local e sua
germinação foi feita em laboratório em estufa a 30 °C, sobre algodão umedecido, por até 4 dias.
5.3.2.2. Avaliação do Potencial Hidrolítico e de Esterificação
A avaliação do potencial catalítico de hidrólise foi iniciada com a utilização do Preparado
Enzimático Vegetal (PEV) para cada dia de germinação das sementes.
Os melhores resultados para as reações de hidrólise foram obtidos com o 4° dias de
germinação, 10,57 ± 0,07% (girassol), 9,44 ± 0,81% (gergelim) e 36,7 ± 1,56% (amendoim) de acidez
final. Tendo em vista estes resultados positivos, o acetone powder foi preparado para uma melhor
caracterização desta lipase, tanto para as reações de hidrólise, quanto para as de esterificação.
Contudo, ao utilizarmos o acetone powder nas reações de hidrólise, constatamos o mesmo
comportamento que foi observado com a macaúba e o butiti, ou seja, o acetone powder não foi capaz
de catalisar as reações de hidrólise, tendo em vista que a acidez final nestas reações não foi superior a
3%. Para os ensaios de avaliação do potencial de esterificação deste acetone powder, foi observado uma
pequena redução da acidez inicial de 98 ± 1,25% para 89,35 ± 4,39 (girassol), 93,8 ± 3,56%
(gergelim) e 83,9 ± 2,64% (amendoim) em 24 horas de reação.

53
Levando-se em consideração o tempo de reação, podemos concluir que não há diferença
entre eles e que estes valores são muito baixos para justificar a continuidade do trabalho com este
tipo de preparação enzimática.
Ao fim desta etapa, podemos concluir a partir dos resultados explicitados a cima, que a
obtenção dos biocatalisadores na forma de acetone powder foi extremamente prejudicial ao potencial
catalítico das possíveis lipases presentes nas sementes germinadas. Isso pode ser evidenciado pela
presença de atividade lipásica na formulação do PEV e ausência quando este mesmo PEV foi
convertido em acetone powder, ou seja, esta metodologia de preparo de certa forma, inativou a lipase
presente na germinação da semente.
Diferente do que foi observado com as lipases presentes na semente germinada de pinhão
manso (Jatropha curcas L.) e na semente dormente (não germinada) de mamona (Ricinus communis L.),
as quais já foram alvo de pesquisa em nosso grupo de trabalho. Com estas lipases foi possível a
realização de reações tanto de hidrólise como de síntese, com excelentes resultados (SOUSA et al,
2010; CAVALCANTI et al, 2007). Estes precedentes nos motivaram a investigar possíveis novas
fontes de lipase vegetal que ainda não foram descritas na literatura, tais como as apresentadas nesta
tese.
Vale salientar também a tentativa de estudo com as sementes de pupunha (Bactris gasipaes
Kunth), açaí (Euterpe oleracea), que não produziu bons resultados graças à incapacidade de fazer com
que suas sementes germinassem; e com a semente germinada de abacate (Persea americana), com a qual
também não foi obtido nenhum resultado satisfatório.
5.4. Enriquecimento Nutricional de Óleos Vegetais com Ácidos Graxos Poli-insaturados
Na produção de óleos e gorduras modificados, o grau de reatividade dos diferentes ácidos
graxos pode ser influenciado pela composição dos triglicerídeos, atividade de água, natureza dos
solventes e também pelo tipo/fonte de lipase. Logo, a capacidade que esses biocatalisadores têm em
discriminar os diferentes tipos de ácidos graxos, pode ser utilizada como ponto positivo em favor de
novas aplicações na indústria alimentícia, que sejam interessantes não só do ponto de vista
econômico, como também benéficas à saúde humana.
Por este ponto de vista, a incorporação dos AGPI da família ω-3 em óleos vegetais mostra-se
uma aplicação bastante interessante, podendo ser alcançada por meio da transesterificação, quando
os AGPI da família ω-3 são empregados na forma de éster metílico ou por interesterificação
(acidólise), quando se utiliza esses ácidos graxos na forma livre (LI e WARD, 1993). Atualmente, o
processo de interesterificação é um dos mais utilizados para modificar as propriedades físicas dos

54
óleos e gorduras, alterando, assim, suas propriedades funcionais e nutricionais (MARANGONI e
ROUSSEAU, 1995; SREENIVASAN, 1976; WILLIS e MARANGONI, 1998; CHRISTENSEN et
al, 2001).
No presente trabalho utilizou-se diversas lipases comerciais e próprias com o objetivo de
incorporar ácidos graxos poli-insaturados em óleos vegetais comestíveis, tendo em vista que os óleos
vegetais são uma fonte lipídica importante e amplamente utilizada em produtos para fins alimentares.
Para isso comparou-se o perfil lipídico dos produtos obtidos, avaliando a incorporação de
AGPI ω-3 específicos, como os ácidos eicosapentaenóico (20:5n3, EPA) e docosahexaenóico
(22:6n3, DHA), que oferecem inúmeros benefícios a saúde, nos óleos de coco, palma e oliva.
5.4.4. Seleção de Lipases para o Enriquecimento Nutricional do Óleo de Coco (Cocos
nucifera L.)
O óleo de coco é uma fonte natural de ácidos graxos de cadeia média (AGCM), contendo
mais de 90% de ácidos graxos saturados (AGSat.). Têm inúmeras aplicações funcionais, pois eles
podem estimular o sistema imunológico, são utilizados no controle de peso, como adjuvantes
dietéticos, entre outras (GOPALA et al, 2010). Devido à absorção mais rápida, os AGCM são úteis
como uma fonte de calorias no tratamento de pacientes hospitalizados ou debilitados (WO
2003033633 A1, 2003).
A seguir são mostrados os resultados obtidos com as lipases comerciais e próprias. Em
seguida, uma comparação mais específica foi feita com os resultados obtidos a partir das mesmas
fontes de lipases microbianas.
A Figura 9 apresenta o perfil das famílias de ácidos graxos presentes no óleo de coco antes e
após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe
catalisado por 14 lipases, apresentadas por ordem decrescente de incorporação de AGPI. A Figura
10 expõe o percentual de incorporação dos ácidos graxos eicosapentaenóico (EPA) e
docosahexaenóico (DHA) no óleo de coco antes e após a reação de acidólise com ácidos graxos poli-
insaturados provenientes do óleo de peixe catalisado por diferentes lipases. Na Tabela 6 estão
explicitados os percentuais das famílias de ácidos graxos no óleo de coco antes e após a reação de
acidólise, assim como a quantidade de EPA e DHA incorporado aos óleos e a razão ω-6/ω-3
encontrada em cada produto ao final da reação catalisada pelas diferentes lipases.
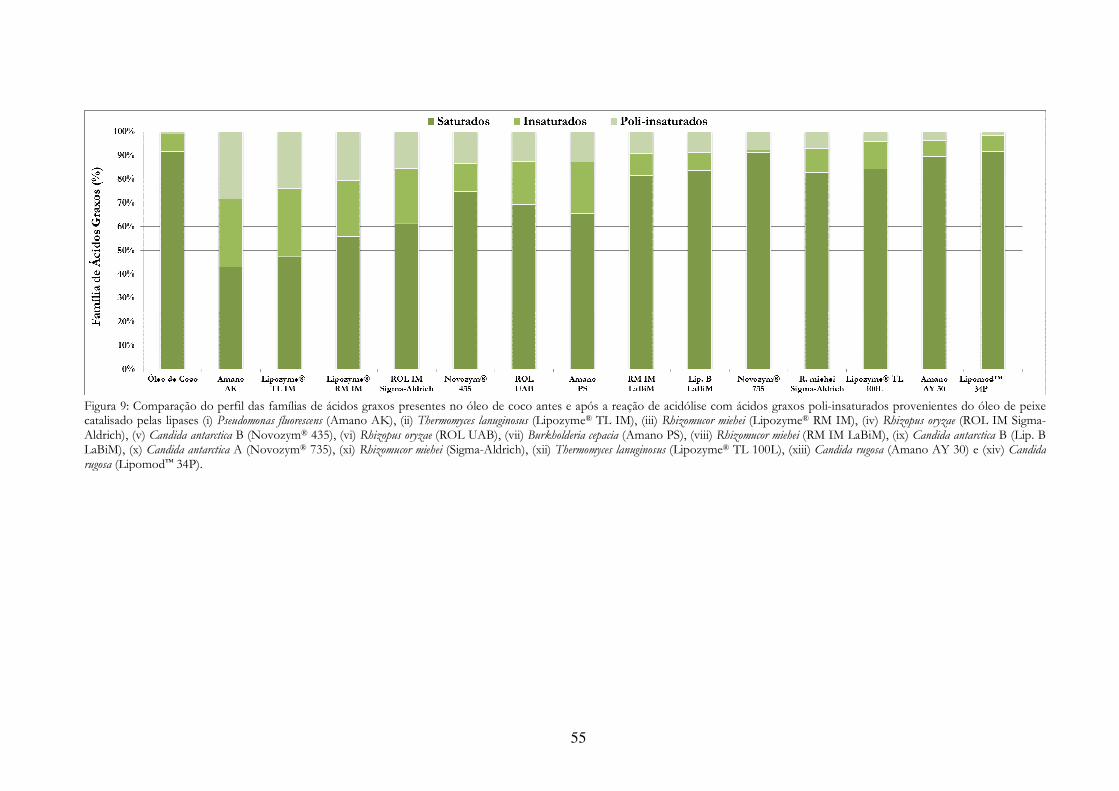
55
.
Figura 9: Comparação do perfil das famílias de ácidos graxos presentes no óleo de coco antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Pseudomonas fluorescens (Amano AK), (ii) Thermomyces lanuginosus (Lipozyme® TL IM), (iii) Rhizomucor miehei (Lipozyme® RM IM), (iv) Rhizopus oryzae (ROL IM Sigma-Aldrich), (v) Candida antarctica B (Novozym® 435), (vi) Rhizopus oryzae (ROL UAB), (vii) Burkholderia cepacia (Amano PS), (viii) Rhizomucor miehei (RM IM LaBiM), (ix) Candida antarctica B (Lip. B LaBiM), (x) Candida antarctica A (Novozym® 735), (xi) Rhizomucor miehei (Sigma-Aldrich), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Candida rugosa (Amano AY 30) e (xiv) Candida rugosa (Lipomod™ 34P).
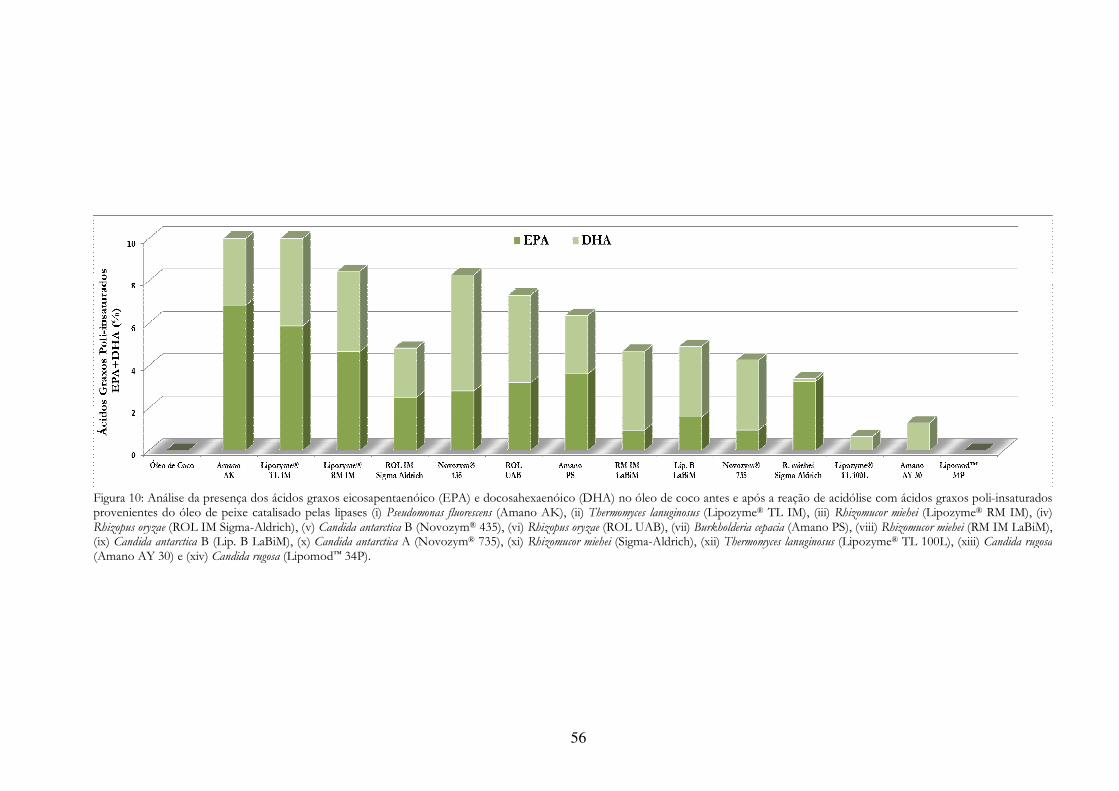
56
Figura 10: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de coco antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Pseudomonas fluorescens (Amano AK), (ii) Thermomyces lanuginosus (Lipozyme® TL IM), (iii) Rhizomucor miehei (Lipozyme® RM IM), (iv) Rhizopus oryzae (ROL IM Sigma-Aldrich), (v) Candida antarctica B (Novozym® 435), (vi) Rhizopus oryzae (ROL UAB), (vii) Burkholderia cepacia (Amano PS), (viii) Rhizomucor miehei (RM IM LaBiM), (ix) Candida antarctica B (Lip. B LaBiM), (x) Candida antarctica A (Novozym® 735), (xi) Rhizomucor miehei (Sigma-Aldrich), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Candida rugosa(Amano AY 30) e (xiv) Candida rugosa (Lipomod™ 34P).
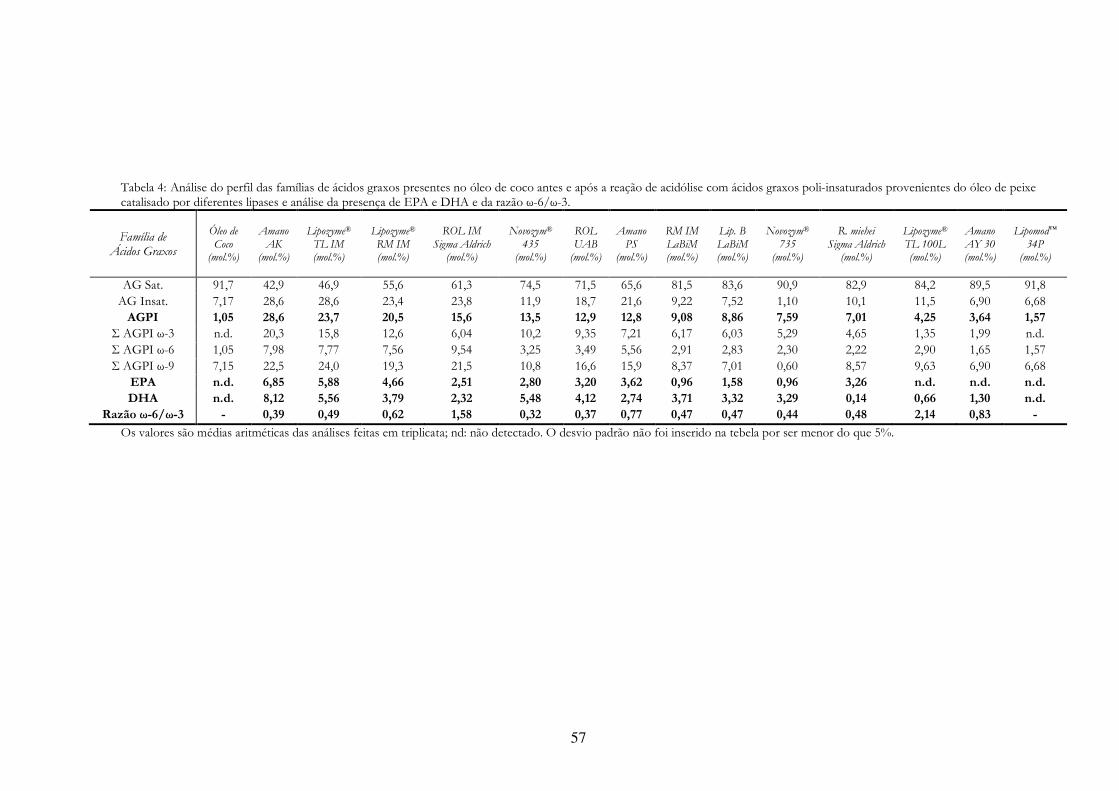
57
Tabela 4: Análise do perfil das famílias de ácidos graxos presentes no óleo de coco antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases e análise da presença de EPA e DHA e da razão ω-6/ω-3.
Família de Ácidos Graxos
Óleo de Coco
(mol.%)
Amano AK
(mol.%)
Lipozyme® TL IM (mol.%)
Lipozyme® RM IM (mol.%)
ROL IM Sigma Aldrich
(mol.%)
Novozym® 435
(mol.%)
ROL UAB (mol.%)
Amano PS
(mol.%)
RM IM LaBiM (mol.%)
Lip. B LaBiM (mol.%)
Novozym® 735
(mol.%)
R. miehei Sigma Aldrich
(mol.%)
Lipozyme® TL 100L (mol.%)
Amano AY 30 (mol.%)
Lipomod™
34P (mol.%)
AG Sat. 91,7 42,9 46,9 55,6 61,3 74,5 71,5 65,6 81,5 83,6 90,9 82,9 84,2 89,5 91,8 AG Insat. 7,17 28,6 28,6 23,4 23,8 11,9 18,7 21,6 9,22 7,52 1,10 10,1 11,5 6,90 6,68 AGPI 1,05 28,6 23,7 20,5 15,6 13,5 12,9 12,8 9,08 8,86 7,59 7,01 4,25 3,64 1,57
Σ AGPI ω-3 n.d. 20,3 15,8 12,6 6,04 10,2 9,35 7,21 6,17 6,03 5,29 4,65 1,35 1,99 n.d. Σ AGPI ω-6 1,05 7,98 7,77 7,56 9,54 3,25 3,49 5,56 2,91 2,83 2,30 2,22 2,90 1,65 1,57 Σ AGPI ω-9 7,15 22,5 24,0 19,3 21,5 10,8 16,6 15,9 8,37 7,01 0,60 8,57 9,63 6,90 6,68
EPA n.d. 6,85 5,88 4,66 2,51 2,80 3,20 3,62 0,96 1,58 0,96 3,26 n.d. n.d. n.d. DHA n.d. 8,12 5,56 3,79 2,32 5,48 4,12 2,74 3,71 3,32 3,29 0,14 0,66 1,30 n.d.
Razão ω-6/ω-3 - 0,39 0,49 0,62 1,58 0,32 0,37 0,77 0,47 0,47 0,44 0,48 2,14 0,83 -
Os valores são médias aritméticas das análises feitas em triplicata; nd: não detectado. O desvio padrão não foi inserido na tebela por ser menor do que 5%.

58
Para esta seleção, as Burkholderia cepacia (Amano PS) e Candida antarctica A (Novozym® 735)
(Figura 9), nas quais o percentual de AGPI totais passou de 1% para 29, 24, 13 e 8%,
respectivamente (Tabela 6).
A redução dos AGSat também foi acentuada com a utilização da Amano AK, Lipozyme® TL
IM e da Amano PS, tendo em vista a diminuição de 92% para 43, 47 e 66%, respectivamente. Além
disso, o incremento não foi satisfatório apenas para os AGPI, mas também para a família de
AGInsat, que aumentou de 7% para 29, 29 e 22% com a utilização das mesmas lipases (Tabela 6).
Para as demais lipases, os percentuais de incorporação AGPI variaram de 4 a 2% com as
lipases de Thermomyces lanuginosus (Lipozyme® TL 100L), Candida rugosa (Amano AY 30) e de Candida
rugosa (Lipomod™ 34P) (Tabela 6).
Na incorporação de AGInsat há um aumento com a utilização da lipase de Thermomyces
lanuginosus (Lipozyme® TL 100L) de 7 para 12%. Já com as lipases de Candida rugosa (Amano AY 30 e
Lipomod™ 34P) praticamente não há alteração; contudo, há uma acentuada redução para 1% com a
utilização da lipase de Candida antarctica A (Novozym® 735). Aparentemente a Novozym® 735
realizou uma troca dos AGInsat pelos AGPI, sem alteração nos saturados.
Com relação aos ácidos ecisapentaenóico (EPA) e docosahexaenóico (DHA), alguns pontos
devem ser destacados, dentre eles: a incorporação unicamente de DHA com as lipases Thermomyces
lanuginosus (Lipozyme® TL 100L) e de Candida rugosa (Amano AY 30), com 0,7 e 1,3% (Tabela 6).
Fato interessante que pode ser explorado para síntese de formulações que objetivem a
suplementação com este tipo específico de ácido graxo. Já a lipase de Candida rugosa (Lipomod™ 34P)
não foi capaz de inserir esses ácidos graxos no óleo de coco.
As maiores incorporações da mistura (EPA+DHA) foram alcançadas com as lipases de
Pseudomonas fluorescens (Amano AK), com 14,97%, Thermomyces lanuginosus, com 11,44% na forma
imobilizada (Lipozyme® TL IM); e com a de Burkholderia cepacia (Amano PS), aonde se chegou 6,36%
(Tabela 6). Estes valores suprem a necessidade de ingestão diária para esses ácidos graxos ao se levar
em consideração a recomendação dietética típica preconizada pela OMS (Organização Mundial da
Saúde), que é de 300 a 500 mg por dia de EPA+DHA (Institute of Medicine, 2005). Os percentuais
alcançados neste processo equivalem a 1198, 915 e 508 mg da mistura de EPA+DHA,
respectivamente. Estes cálculos foram feitos com base no consumo de apenas uma colher de sopa
(8,0 g) por dia deste óleo enriquecido.
Os produtos obtidos com a aplicação da Amano AK e da Lipozyme® TL IM fornecem um
produto cuja composição é recomendada para indivíduos com doença coronariana, tendo em vista a
recomendação da ingestão de 900 mg da mistura EPA+DHA, segundo a American Heart Association

59
em suas Diretrizes oficiais dietéticas (2000), além de também diminuir o risco de acidente vascular
cerebral (AVC) (FAO/WHO, 2008).
Baseado nestes fatos, esta reação de interesterificação gera um lipídio com uma composição
singular de AGCM e AGPI, que proporciona a soma dos efeitos benefícios para o sistema
imunológico, pelo aumento na ingestão de EPA e DHA com as vantagens nutricionais dos AGCM
de serem hipocolesterolêmico e hipotrigliceridêmico.
Além do mais, podemos concluir que as lipases mais indicadas para o enriquecimento
nutricional do óleo de coco são as de Pseudomonas fluorescens (Amano AK) e de Thermomyces lanuginosus
na forma imobilizada (Lipozyme® TL IM) e que foi possível obter um produto de grande interesse
farmacológico que pode ser utilizado na prevenção de doença coronariana (infarto do miocárdio) e
de AVC.
A lipase de Rhizomucor miehei foi descrita pela primeira vez em 1973 e é uma enzima
amplamente utilizada em diversas reações da indústria oleoquímica (RODRIGUES e
FERNANDEZ-LAFUENTE, 2010). A sua forma comercial mais utilizada é disponibilizada pela
Novozymes na forma imobilizada com o nome de Lipozyme® RM IM. Outra fonte de lipase
utilizada no enriquecimento dos óleos vegetais que se fez a comparação de desempenho, foi a lipase
de Candida antarctica B. A sua forma comercial mais utilizada é disponibilizada pela Novozymes na
forma imobilizada com o nome de Novozym® 435. Para finalizar esta parte de comparação no
desempenho do enriquecimento nutricional de óleos vegetais utilizamos a lipase de Rhizopus oryzae. A
sua forma comercial utilizada é disponibilizada pela Sigma-Aldrich com o nome de Rhizopus oryzae
lipase.
As lipases de Rhizomucor miehei e de Candida antarctica B foram obtidas no Laboratório de
Biotecnologia Microbriana (LaBiM) segundo metodologia descrita no item 4.2 em material e
métodos. Já a lipase de Rhizopus oryzae foi cedida ao LaBiM pelo professor Francisco Valero do Dept.
Enginyeria Química da Universitat Autònoma Barcelona (UAB - Espanha) em trabalho de
cooperação internacional.
A Figura 13 apresenta o perfil das famílias de ácidos graxos presentes no óleo de coco antes e
após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe
catalisado pelas lipases de Rhizomucor miehei na forma comercial (Lipozyme® RM IM e Sigma-Aldrich)
e própria (RM IM LaBiM); lipase de Candida antarctica B comercial (Novozym® 435) e própria (Lip. B
LaBiM) e por último, a lipase de Rhizopus oryzae comercial (Rhizopus oryzae lipase Sigma-Aldrich) e
própria (ROL UAB).

60
Ao avaliarmos o enriquecineto nutricional do óleo de coco (Figura 13), observamos uma
elevada incorporação de AGPI, devido ao aumento de 1%, para 21,0% com a utilização da
Lipozyme® RM IM, 9,2% com a RM IM LaBiM e 7,0% com Rhizomucor miehei da Sigma-Aldrich (RM
SA). Já a quantidade de ácidos graxos insaturados (AGInsat) foi triplicada com o uso da Lipozyme® e
aumentou em cerca de 30% com a home made e RM SA (Figura 13).
Os ácidos EPA e DHA, que não foram detectados no óleo de coco inicial, passaram a fazer
parte do produto final, com 4,7% de EPA e 3,8% de DHA com a utilização da Lipozyme®, 0,96% de
EPA e 3,7% de DHA com a utilização da RM IM LaBiM e 3,3% de EPA e 0,14% de DHA (Tabela
7). Os valores encontrados para a Lipozyme® e a RM IM LaBiM atendem a necessidade de ingestão
diária recomendada; já a RM SA não. Os percentuais alcançados neste processo equivalem a 676, 374
e 272 mg da mistura de EPA+DHA.
Como podemos observar, a incorporação de DHA foi semelhante para a Lipozyme® RM IM
e para a RM IM LaBiM. Com a RM SA observa-se incorporação de EPA quase que exclusivamente.
Uma formulação comercial que apresente a relação DHA/EPA (4:1) obtida com a RM IM LaBiM é
ideal para mulheres gestantes ou que estejam amamentando, para compensar a perda de DHA tanto
para o feto em formação como para o o desenvolvimento do recém nascido.
Com a utilização da lipase de Candida antarctica B, foi obtido um aumento de 1% para 13,63%
com a Novozym® 435 e de 1% para 8,92% com o emprego da Lip. B LaBiM. Para os AGInsat,
houve um aumento em 60% com a comercial; já com a home made não houve modificação desta
família. Na inserção de EPA+DHA, que não foram detectados neste óleo, passaram a fazer parte do
produto final com 2,8% de EPA e 5,48% de DHA com a utilização da Novozym® 435 e com 1,58%
de EPA e 3,32% de DHA com a utilização da Lip. B LaBiM (Tabela 7). Valores estes que também
suprem a necessidade de ingestão diária para esses ácidos graxos tendo em vista que os percentuais
alcançados neste processo equivalem a 662 e 392 mg da mistura de EPA+DHA, alcançados com a
Novozym® 435 e Lip. B LaBiM, respectivamente. Com a utilização desta lipase de Candida antarctica,
temos a mesma proporção de EPA e DHA para as duas preparações.
Para finalizar esta parte de comparação no desempenho do enriquecimento nutricional do
óleo de coco, utilizamos a lipase de Rhizopus oryzae. Com ela, foi possível observar uma elevada
incorporação de AGPI, devido ao aumento de 1% para 15,57% para a lipase comercial de 1% para
12,91% com a ROL UAB (Tabela 7). Com relação aos AGInsat também há um aumento
considerável, em torno de 330 e 260%, para a ROL comercial e UAB, respectivamente.

61
Os ácidos EPA e DHA, que não foram detectados no óleo de coco inicial, passaram a fazer
parte do produto final, com 2,51% de EPA e 2,32% de DHA com a utilização da ROL comercial e
com 3,20% de EPA e 4,12% de DHA com a utilização da ROL UAB (Tabela 7). Valores estes que
também suprem a necessidade de ingestão diária para esses ácidos graxos tendo em vista que os
percentuais alcançados neste processo equivalem a 386 e 586 mg da mistura de EPA+DHA, com a
Rhizopus oryzae comercial e ROL UAB, respectivamente. Neste caso a lipase própria apresentou um
desempenho superior ao apresentado pela comercial.
5.4.5. Enriquecimento Nutricional do Óleo de Palma (Elaeis guineenses Jacq.)
O óleo de palma (Elaeis guineenses Jacq.) atualmente é o segundo maior óleo comestível
comercializado no mundo (EDEM, 2002). Ele e seus derivados são comumente usados na indústria
alimentícia por terem elevada estabilidade oxidativa, devido a sua composição em ácidos graxos e a
presença de carotenóides e tocotrienóis. Com isto, ele pode ser utilizado sob altas temperaturas e
suporta uma vida útil de prateleira prolongada (FOSTER; WILLIAMSON; LUNN, 2009).
Este óleo tem baixos níveis de ácidos graxos poli-insaturados, especialmente ômega-3, e o
seu enriquecimento com esses ácidos graxos pode melhorar suas propriedades nutricionais. Por isso,
o mesmo princípio utilizado no óleo de coco foi aplicado para o óleo de palma.
A Figura 15 apresenta o perfil das famílias de ácidos graxos presentes no óleo de palma antes
e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe
catalisado por 14 lipases, apresentadas por ordem decrescente de incorporação de AGPI. A Figura
16 expõe o percentual de incorporação dos ácidos graxos eicosapentaenóico (EPA) e
docosahexaenóico (DHA) no óleo de palma antes e após a reação de acidólise com ácidos graxos
poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases. Na Tabela 7 estão
explicitados os percentuais das famílias de ácidos graxos no óleo de palma antes e após a reação de
acidólise, assim como a quantidade de EPA e DHA incorporado aos óleos e a razão ω-6/ω-3
encontrada em cada produto ao final da reação catalisada pelas diferentes lipases.
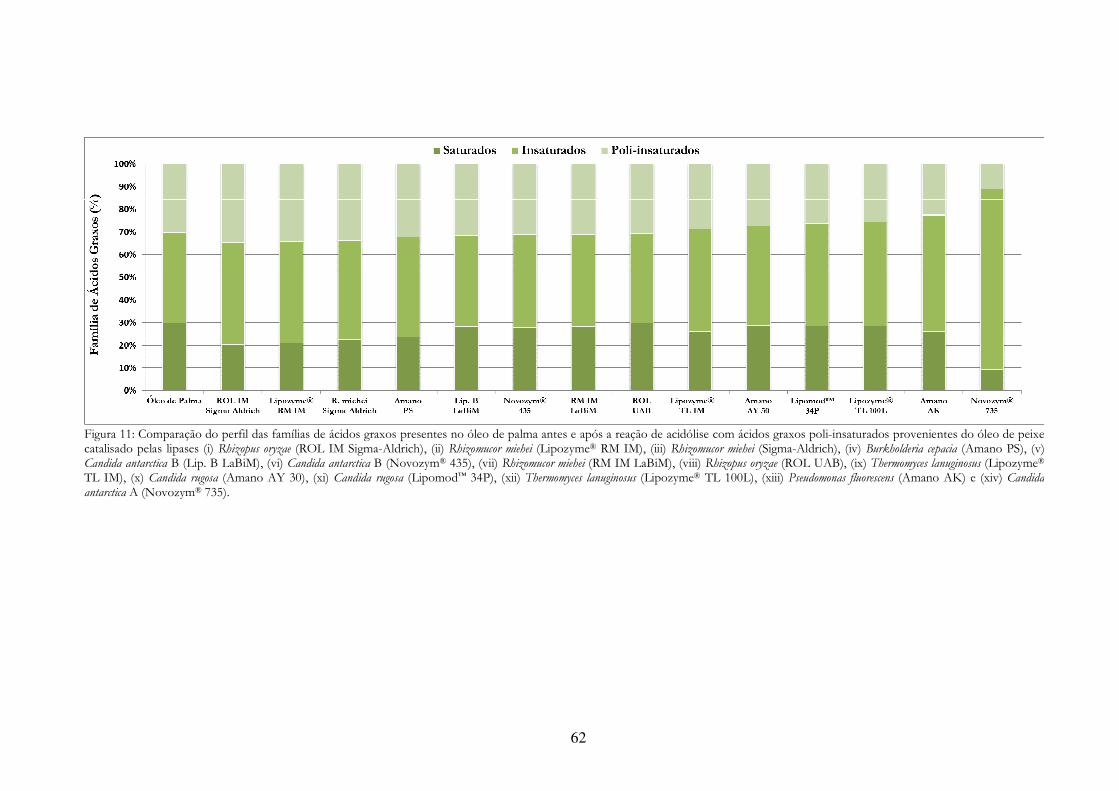
62
Figura 11: Comparação do perfil das famílias de ácidos graxos presentes no óleo de palma antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Rhizopus oryzae (ROL IM Sigma-Aldrich), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizomucor miehei (Sigma-Aldrich), (iv) Burkholderia cepacia (Amano PS), (v)Candida antarctica B (Lip. B LaBiM), (vi) Candida antarctica B (Novozym® 435), (vii) Rhizomucor miehei (RM IM LaBiM), (viii) Rhizopus oryzae (ROL UAB), (ix) Thermomyces lanuginosus (Lipozyme®TL IM), (x) Candida rugosa (Amano AY 30), (xi) Candida rugosa (Lipomod™ 34P), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Pseudomonas fluorescens (Amano AK) e (xiv) Candida antarctica A (Novozym® 735).
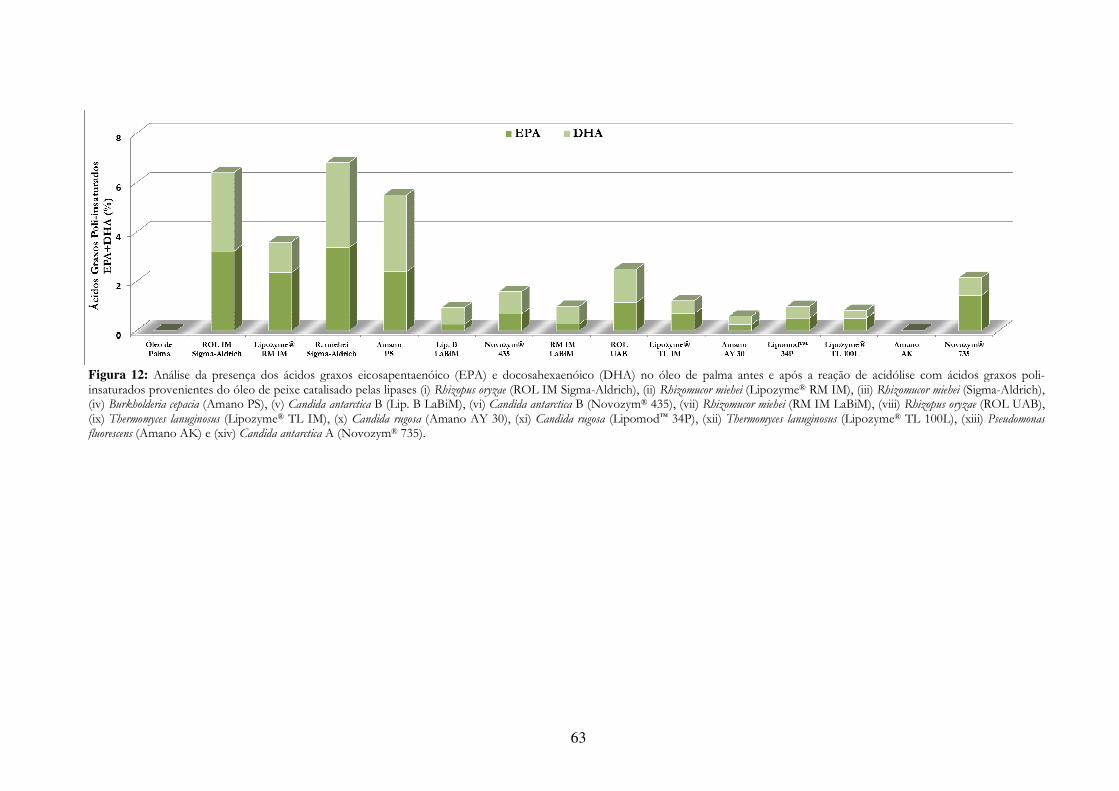
63
Figura 12: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de palma antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Rhizopus oryzae (ROL IM Sigma-Aldrich), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizomucor miehei (Sigma-Aldrich), (iv) Burkholderia cepacia (Amano PS), (v) Candida antarctica B (Lip. B LaBiM), (vi) Candida antarctica B (Novozym® 435), (vii) Rhizomucor miehei (RM IM LaBiM), (viii) Rhizopus oryzae (ROL UAB), (ix) Thermomyces lanuginosus (Lipozyme® TL IM), (x) Candida rugosa (Amano AY 30), (xi) Candida rugosa (Lipomod™ 34P), (xii) Thermomyces lanuginosus (Lipozyme® TL 100L), (xiii) Pseudomonas fluorescens (Amano AK) e (xiv) Candida antarctica A (Novozym® 735).
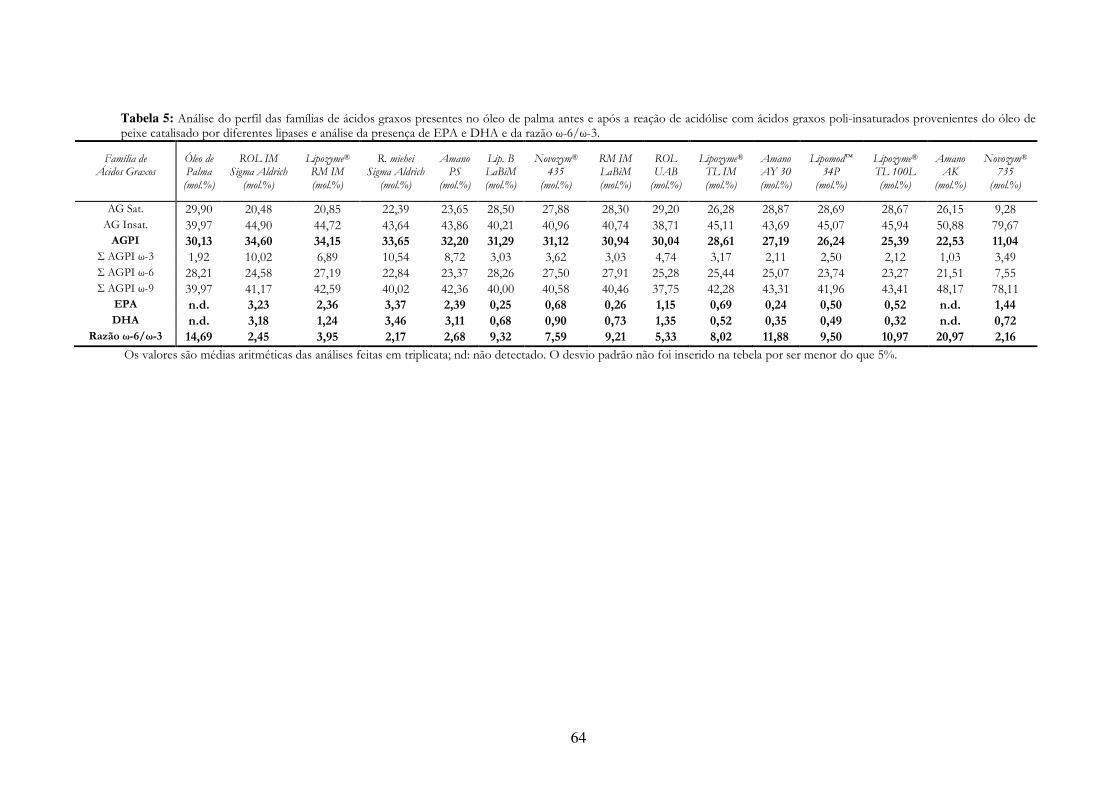
64
Tabela 5: Análise do perfil das famílias de ácidos graxos presentes no óleo de palma antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases e análise da presença de EPA e DHA e da razão ω-6/ω-3.
Família de Ácidos Graxos
Óleo de Palma (mol.%)
ROL IM Sigma Aldrich
(mol.%)
Lipozyme® RM IM (mol.%)
R. miehei Sigma Aldrich
(mol.%)
Amano PS
(mol.%)
Lip. B LaBiM (mol.%)
Novozym® 435
(mol.%)
RM IM LaBiM (mol.%)
ROL UAB (mol.%)
Lipozyme® TL IM (mol.%)
Amano AY 30 (mol.%)
Lipomod™ 34P
(mol.%)
Lipozyme® TL 100L (mol.%)
Amano AK
(mol.%)
Novozym® 735
(mol.%)
AG Sat. 29,90 20,48 20,85 22,39 23,65 28,50 27,88 28,30 29,20 26,28 28,87 28,69 28,67 26,15 9,28 AG Insat. 39,97 44,90 44,72 43,64 43,86 40,21 40,96 40,74 38,71 45,11 43,69 45,07 45,94 50,88 79,67 AGPI 30,13 34,60 34,15 33,65 32,20 31,29 31,12 30,94 30,04 28,61 27,19 26,24 25,39 22,53 11,04
Σ AGPI ω-3 1,92 10,02 6,89 10,54 8,72 3,03 3,62 3,03 4,74 3,17 2,11 2,50 2,12 1,03 3,49 Σ AGPI ω-6 28,21 24,58 27,19 22,84 23,37 28,26 27,50 27,91 25,28 25,44 25,07 23,74 23,27 21,51 7,55 Σ AGPI ω-9 39,97 41,17 42,59 40,02 42,36 40,00 40,58 40,46 37,75 42,28 43,31 41,96 43,41 48,17 78,11
EPA n.d. 3,23 2,36 3,37 2,39 0,25 0,68 0,26 1,15 0,69 0,24 0,50 0,52 n.d. 1,44 DHA n.d. 3,18 1,24 3,46 3,11 0,68 0,90 0,73 1,35 0,52 0,35 0,49 0,32 n.d. 0,72
Razão ω-6/ω-3 14,69 2,45 3,95 2,17 2,68 9,32 7,59 9,21 5,33 8,02 11,88 9,50 10,97 20,97 2,16
Os valores são médias aritméticas das análises feitas em triplicata; nd: não detectado. O desvio padrão não foi inserido na tebela por ser menor do que 5%.

65
Para o óleo de palma observamos uma diminuição no percentual de AGPI totais que
compunham o óleo antes da reação de acidólise. A sua composição inicial, que era de 30% sofreu
uma redução para: 28,6% (Lipozyme® TL IM), 27,2% (Amano AY 30), 26,2% (Lipomod™ 34P),
25,4% (Lipozyme® TL 100L) e 22,5% (Amano AK) e 11,0% (Novozym® 735). Já para as lipases
Amano PS, há um pequeno acréscimo em torno de 10% (Tabela 8). Apesar dessa redução, há
incorporação tanto em EPA quanto em DHA, mostrando que houve uma melhora na qualidade dos
AGPI. Exceto para a lipase de Pseudomonas fluorescens (Amano AK), na qual esses ácidos graxos não
foram detectados (Figura 27). Estes ácidos graxos passaram a fazer parte do produto final, com
2,39% de EPA e 3,11% de DHA com a utilização da Amano PS (Tabela 11). Estes valores suprem a
necessidade de ingestão diária para esses ácidos graxos, pois os percentuais alcançados neste
processo equivalem a 440 mg da mistura de EPA+DHA.
Há um ganho no percentual de AGInsat com todas as lipases testadas, com destaque para a
de Candida antarctica A (Novozym® 735), com a qual foi possível dobrar o seu percentual no produto
final. Esta característica poderá ser explorada em outras matrizes oleosas, como as empregadas na
síntese de similares da manteiga de cacau.
Ao final das reações de acidólise, chegou-se a formulações com as mais variadas razões ω-
6/ω-3. A partir do óleo de palma inicial que apresentava uma relação de 14:1, chegou-se não só a
mínima de 2:1 com a lipase de Candida antarctica A (Novozym® 735), como também foi possível
aumentear esta relação para 21:1 com a lipase de Pseudomonas fluorescens (Amano AK). Como visto
anteriormente (Ítem 3.13.4.3), este fato não é desejável do ponto de vista nutricional, uma vez que
elevados níveis de ácidos graxos ômega-6, geram um excesso de ácido araquidônico no organismo.
Com isso podemos concluir que a Amano AK de Pseudomonas fluorescens não deve ser utilizada
para este tipo de preparação e que a lipase mais indicada para o enriquecimento do óleo de palma é a
Amano PS de Burkholderia cepacia.
A Figura 17 apresenta o perfil das famílias de ácidos graxos presentes no óleo de palma antes
e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe
catalisado pelas lipases de Rhizomucor miehei na forma comercial (Lipozyme® RM IM e Sigma-Aldrich)
e própria (RM IM LaBiM); lipase de Candida antarctica B comercial (Novozym® 435) e própria (Lip. B
LaBiM) e por último, a lipase de Rhizopus oryzae comercial (Rhizopus oryzae lipase Sigma-Aldrich) e
própria (ROL UAB).

66
Para o óleo de palma houve uma pequena mudança, em torno de 10%, nos percentuais de
AGPI e AGInsat, com a utilização da Lipozyme® e da RM SA; já com a RM IM LaBiM,
praticamente não houve mudança. Todavia, foi possível incorporar EPA e DHA com as três lipases
de Rhizomucor miehei (Figura 18). Com as incorporações de 3,37% de EPA e 3,46% de DHA com a
utilização da RM SA foi possível alcançar a necessidade de ingestão diária para esses ácidos graxos,
pois os percentuais alcançados neste processo equivalem a 546 mg da mistura de EPA+DHA.
Também houve uma redução na razão ω-6/ω-3 de 15:1 do óleo de palma original, para 2:1,
4:1 e 9:1, com a utilização da RM SA, Lipozyme® e da RM IM LaBiM, respectivamente. Esta relação
atingida fica dentro da faixa de 5:1 a 10:1 recomendada pela FAO/OMS, que também é fortemente
recomendada para gestantes pela Health and Welfare Canada (1990). Com a RM SA chega-se a razão
2:1, que atende aos padrões do Instituto de Medicina (EUA), que ao ser mais rigoroso, aconselha
uma relação de no máximo 5:1 (Institute of Medicine, 2002).
No estudo de FAJARDO; AKOH; LAI (2003) com a acidólise do óleo de palma com AGPI
ω-3 há uma incorporação média de 20,8% de EPA e 15,6% DHA no óleo (razão molar óleo de
palma/ EPAX EE de 1:3), utilizando 10% (p/p) da lipase IM60 (Rhizomucor miehei) em meio orgânico
(hexano) a 55 °C em 24 horas de reação. Esses valores são superiores ao encontrado em nossa
investigação por utilizar uma matéria prima comercial (EPAX EE) concentrada em AGPI ω-3.
Com o autilização da lipase de Candida antarctica B, o mesmo perfil de produto foi observado,
uma vez que, praticamente não há diferença entre as composições do óleo original e os óleos que
sofreram acidólise (Figura 17). Há apenas uma pequena incorporação de EPA+DHA, que não chega
a 1%. Contudo, há uma melhora na razão ω-6/ω-3 de 14:1 do óleo de palma original, para 7:1 e 9:1,
alcançados com a Novozym® 435 e Lip. B LaBiM, respectivamente, ficando dentro da faixa de 5:1 a
10:1 recomendada pela FAO/OMS.
Com a lipase de Rhizopus oryzae, foi possível apenas uma pequena mudança com a lipase
comercial, cerca de 10%; já com a lipase ROL UAB, não houve modificação no percentual final de
AGPI. Com relação a incorporação de EPA e DHA, a ROL comercial conseguiu inserir duas vezes
mais que a ROL LaBiM (Tabela 9) e com isso alcançou a dose diária recomendada, tendo em vista
que o percentual de 6,41% equivale a 512 mg da mistura EPA+DHA . Neste caso vale salientar que
a preparação comercial além de imbolizada apresenta uma atividade de esterificação duas vezes maior
que a ROL UAB.
Também houve uma redução considerável na razão ω-6/ω-3 de 15:1 do óleo de palma
original, para 2:1 e 5:1, respectivamente. Como já foi visto, essa relação atende aos padrões do
Instituto de Medicina (EUA).

67
Em vista dos resultados, podemos indicar as lipases comerciais de Rhizomucor miehei e de
Rhizopus oryzae, ambas fornecidas pela Sigma-Aldrich, como as mais indicadas para o enriquecimento
do óleo de palma neste estudo comparativo.
5.4.6. Enriquecimento Nutricional do Óleo de Oliva (Olea europaea)
As modificações em óleos e gorduras para melhoria da sua funcionalidade têm ocorrido com
vários tipos de óleos, incluindo o óleo de oliva. Este óleo desfruta de uma posição privilegiada entre
os óleos comestíveis, devido ao grande consumo pelos habitantes do Mediterrâneo (OH et al, 2009).
Ao longo das últimas décadas estudos nutricionais e epidemiológicos têm fornecido
evidências de que o consumo de azeite de oliva virgem contribui para os benefícios de saúde, em
virtude da presença de compostos antioxidantes e seus derivados (FRANKEL, 2011). Porém, em
muitas culturas o azeite de oliva é utilizado mais comumente como condimento (tempero) para
saladas. Com isso, a prerrogativa nesta investigação seria a de deixá-lo ainda mais atraente do ponto
de vista nutricional pela inserção de AGPI e consequentemente aumentar os efeitos benéficos para o
sistema imunológico, pelo aumento na ingestão de EPA e DHA.
A Figura 19 apresenta o perfil das famílias de ácidos graxos presentes no óleo de oliva antes e
após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe
catalisado por 14 lipases, apresentadas por ordem decrescente de incorporação de AGPI. A Figura
20 expõe o percentual de incorporação dos ácidos graxos eicosapentaenóico (EPA) e
docosahexaenóico (DHA) no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-
insaturados provenientes do óleo de peixe catalisado por diferentes lipases. Na Tabela 7 estão
explicitados os percentuais das famílias de ácidos graxos no óleo de oliva antes e após a reação de
acidólise, assim como a quantidade de EPA e DHA incorporado aos óleos e a razão ω-6/ω-3
encontrada em cada produto ao final da reação catalisada pelas diferentes lipases.
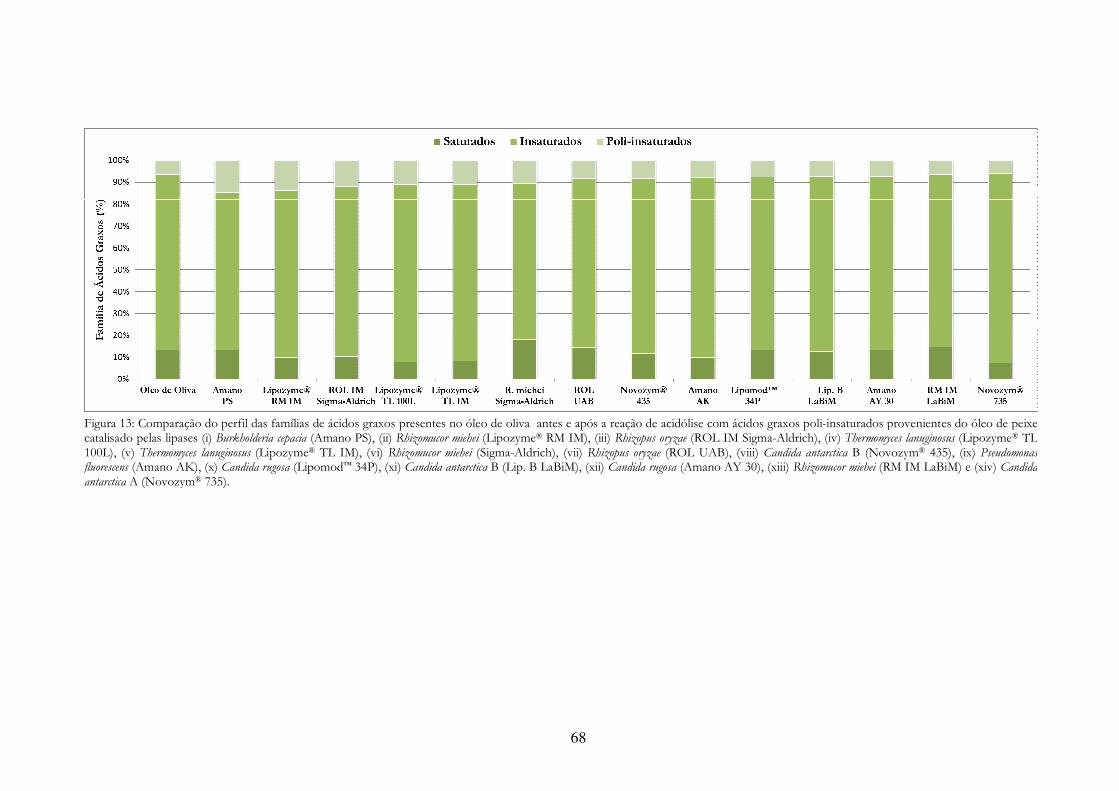
68
Figura 13: Comparação do perfil das famílias de ácidos graxos presentes no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Burkholderia cepacia (Amano PS), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizopus oryzae (ROL IM Sigma-Aldrich), (iv) Thermomyces lanuginosus (Lipozyme® TL 100L), (v) Thermomyces lanuginosus (Lipozyme® TL IM), (vi) Rhizomucor miehei (Sigma-Aldrich), (vii) Rhizopus oryzae (ROL UAB), (viii) Candida antarctica B (Novozym® 435), (ix) Pseudomonas fluorescens (Amano AK), (x) Candida rugosa (Lipomod™ 34P), (xi) Candida antarctica B (Lip. B LaBiM), (xii) Candida rugosa (Amano AY 30), (xiii) Rhizomucor miehei (RM IM LaBiM) e (xiv) Candida antarctica A (Novozym® 735).
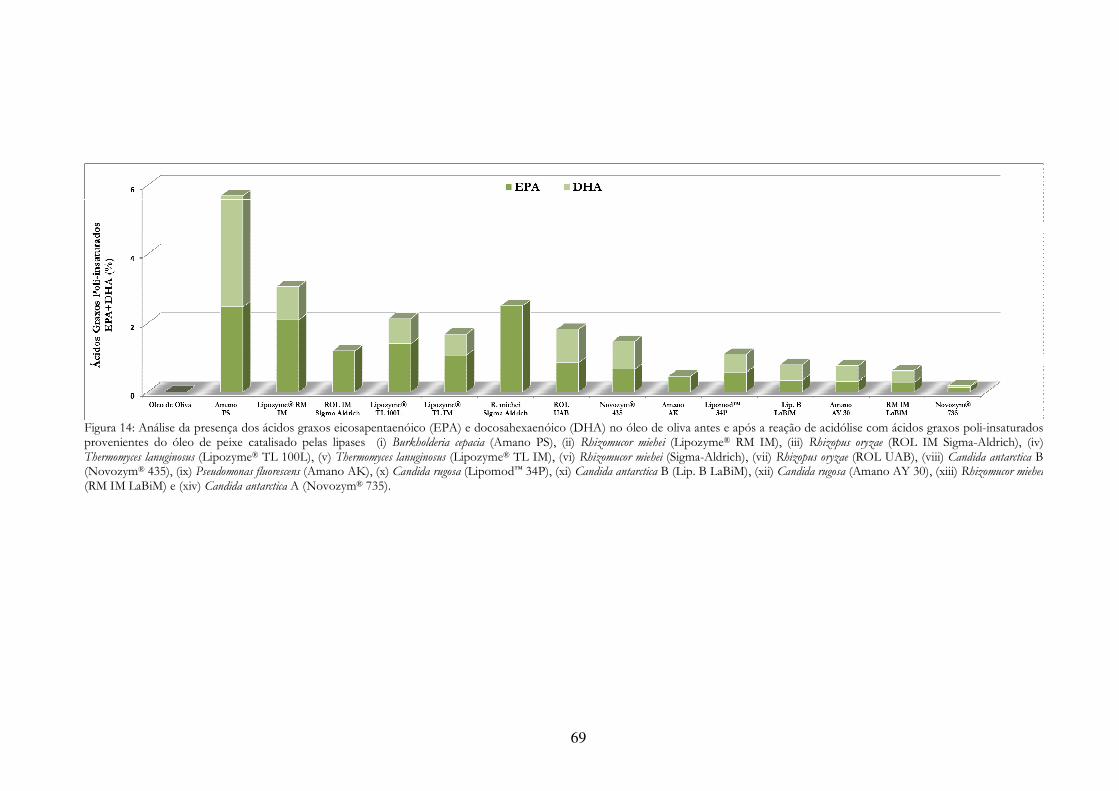
69
Figura 14: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado pelas lipases (i) Burkholderia cepacia (Amano PS), (ii) Rhizomucor miehei (Lipozyme® RM IM), (iii) Rhizopus oryzae (ROL IM Sigma-Aldrich), (iv) Thermomyces lanuginosus (Lipozyme® TL 100L), (v) Thermomyces lanuginosus (Lipozyme® TL IM), (vi) Rhizomucor miehei (Sigma-Aldrich), (vii) Rhizopus oryzae (ROL UAB), (viii) Candida antarctica B (Novozym® 435), (ix) Pseudomonas fluorescens (Amano AK), (x) Candida rugosa (Lipomod™ 34P), (xi) Candida antarctica B (Lip. B LaBiM), (xii) Candida rugosa (Amano AY 30), (xiii) Rhizomucor miehei(RM IM LaBiM) e (xiv) Candida antarctica A (Novozym® 735).
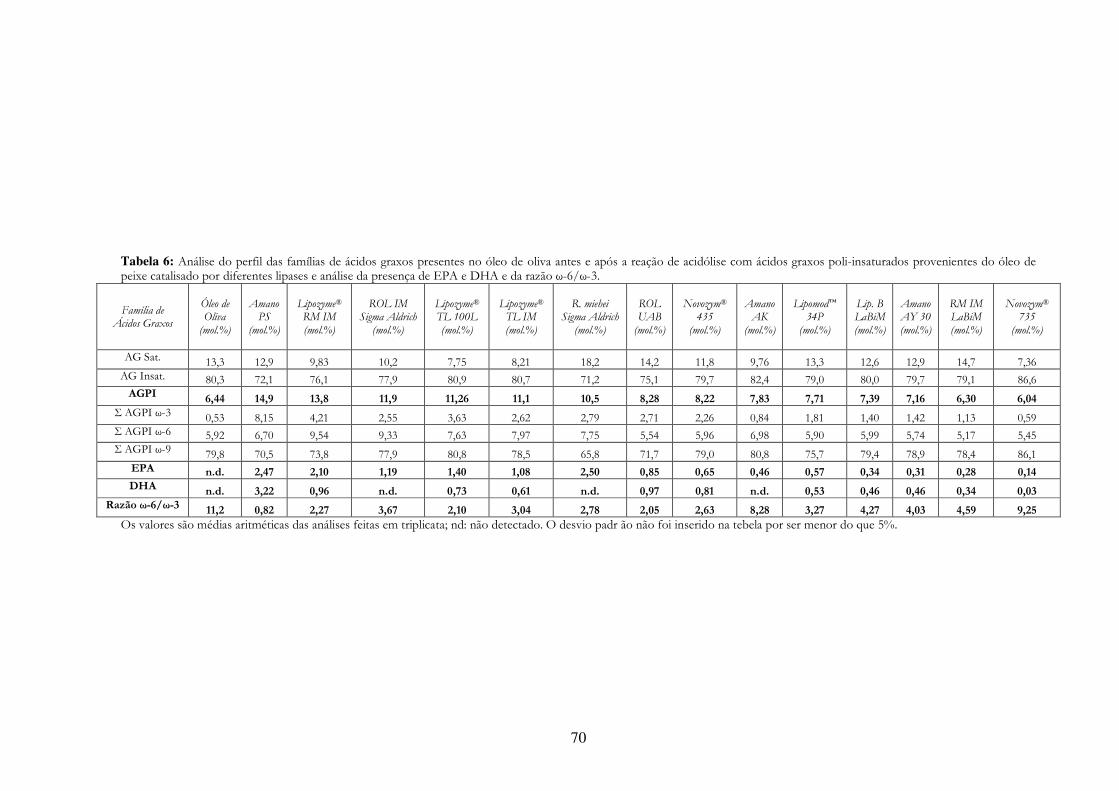
70
Tabela 6: Análise do perfil das famílias de ácidos graxos presentes no óleo de oliva antes e após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases e análise da presença de EPA e DHA e da razão ω-6/ω-3.
Família de Ácidos Graxos
Óleo de Oliva (mol.%)
Amano PS
(mol.%)
Lipozyme® RM IM (mol.%)
ROL IM Sigma Aldrich
(mol.%)
Lipozyme® TL 100L (mol.%)
Lipozyme® TL IM (mol.%)
R. miehei Sigma Aldrich
(mol.%)
ROL UAB (mol.%)
Novozym® 435
(mol.%)
Amano AK
(mol.%)
Lipomod™ 34P
(mol.%)
Lip. B LaBiM (mol.%)
Amano AY 30 (mol.%)
RM IM LaBiM (mol.%)
Novozym® 735
(mol.%)
AG Sat. 13,3 12,9 9,83 10,2 7,75 8,21 18,2 14,2 11,8 9,76 13,3 12,6 12,9 14,7 7,36 AG Insat. 80,3 72,1 76,1 77,9 80,9 80,7 71,2 75,1 79,7 82,4 79,0 80,0 79,7 79,1 86,6 AGPI 6,44 14,9 13,8 11,9 11,26 11,1 10,5 8,28 8,22 7,83 7,71 7,39 7,16 6,30 6,04
Σ AGPI ω-3 0,53 8,15 4,21 2,55 3,63 2,62 2,79 2,71 2,26 0,84 1,81 1,40 1,42 1,13 0,59 Σ AGPI ω-6 5,92 6,70 9,54 9,33 7,63 7,97 7,75 5,54 5,96 6,98 5,90 5,99 5,74 5,17 5,45 Σ AGPI ω-9 79,8 70,5 73,8 77,9 80,8 78,5 65,8 71,7 79,0 80,8 75,7 79,4 78,9 78,4 86,1
EPA n.d. 2,47 2,10 1,19 1,40 1,08 2,50 0,85 0,65 0,46 0,57 0,34 0,31 0,28 0,14 DHA n.d. 3,22 0,96 n.d. 0,73 0,61 n.d. 0,97 0,81 n.d. 0,53 0,46 0,46 0,34 0,03
Razão ω-6/ω-3 11,2 0,82 2,27 3,67 2,10 3,04 2,78 2,05 2,63 8,28 3,27 4,27 4,03 4,59 9,25 Os valores são médias aritméticas das análises feitas em triplicata; nd: não detectado. O desvio padr ão não foi inserido na tebela por ser menor do que 5%.

71
Ao avaliarmos o enriquecimento nutricional do óleo de oliva, percebemos que foi possível
quase dobrar a quantidade de AGPI, com as lipasaes Amano PS de Burkholderia cepacia e com a de
Thermomyces lanuginosus, tanto na forma imobilizada (Lipozyme® TL IM), como na forma livre
(Lipozyme® TL 100L).
Para o EPA e DHA, a incorporação unicamente de EPA com as lipases Amano AK de
Pseudomonas fluorescens e de Rhizomucor miehei (Sigma-Aldrich) (Figura 20), atingindo 0,46 e 2,50%,
respectivamente, é um fato interessante ainda não reportado e que pode ser alvo de estudos. A maior
incorporação se deu com a lipase de Burkholderia cepacia (Amano PS IM), chegando-se a 2,47% de
EPA e 3,22% de DHA. Estes valores correspondem a uma ingestão de 455 mg da mistura de
EPA+DHA e suprem a necessidade de ingestão diária para estes ácidos graxos.
Ao final das reações de acidólise do óleo de oliva, também chegou-se a formulações com as
mais variadas razões ω-6/ω-3. A partir do óleo de oliva inicial que apresentava uma relação de 11:1,
chegou-se a 3:1 e 2:1 (Tabela 12), que são consideradas nutricionalmente mais saudáveis.
Em vista dos resultados apresentados, as lipase Amano PS de Burkholderia cepacia e de
Thermomyces lanuginosus, tanto na forma imobilizada (Lipozyme® TL IM), como na forma livre
(Lipozyme® TL 100L) são as mais indicadas para a realização do enriquecimento nutricional do óleo
de oliva; sendo a primeira oferece maior vantagem devido a elevada incorporação de EPA e DHA.
No óleo de oliva houve um aumento em 120 e 60% nos AGPI com a utilização da
Lipozyme® e da RM SA, respectivamente. Com a RM IM LaBiM, não houve aumento nos AGPI.
A incorporação de DHA chegou ao máximo de 1% com a Lipozyme®, sendo que esse ácido
graxo não foi detectado com a utilização da RM SA. Para o EPA, foi visto uma incorporação de 2,1 e
2,5% para a Lipozyme® e RM SA, respectivamente; já com a RM IM LaBiM foi apenas de 0,28%
(Tabela 11). Com estes resultados não foi possível alcançar a dose de ingestão diária recomendada
para estes ácidos graxos.
Já para a relação razão ω-6/ω-3, há uma diminuição de 11:1 do óleo de oliva original, para 2:1
e 5:1, respectivamente. Relação que atende aos padrões do Instituto de Medicina (EUA).
Com a lipase de Candida antarctica B, houve uma pequena incorporação de AGPI, ao redor de
25 e 15%, para a Novozym® 435 e Lip. B LaBiM, respectivamente. A incorporação de EPA+DHA
também não foi satisfatória para estas duas preparações, tendo em vista os pequenos percentuais de
incorporação (Tabela 11). Já para a razão ω-6/ω-3, há uma diminuição de 11:1 do óleo de oliva
original, para 2:1 e 4:1, respectivamente (Tabela 11). Como vimos anteriormente, são excelentes
valores preconizados pelo Instituto de Medicina (EUA). Resultado similar foi observado no estudo
de Nagachinta e Akoh (2012) no qual realizaram a acidólise de oleína de palma com uma mistura dos
ácidos docosahexaenóico (DHA) e araquiônico (ARA), onde chegaram a 23% de incorporação

72
utilizando uma razão molar DHA+ARA /oleína de palma de 1:12, catalisada pela Novozym® 435 em
meio orgânico (hexano) a 60 °C em 24 horas de reação.
Na utilização da lipase de Rhizopus oryzae, foi possível dobrar a quantidade de AGPI com a ROL
comercial e elevar em 30% com a ROL UAB (Tabela 11).
Com a utilização da lipase comercial não foi detectado DHA no óelo de oliva, somente o
EPA; já com ROL UAB foi detectado a presença dos dois ácidos graxos, embora em pequena
quantidade.
Para este caso também foi notada uma redução na relação razão ω-6/ω-3, que passou de 11:1
do óleo de palma original, para 4:1 e 2:1, respectivamente (Tabela 11). Para as duas lipases de
Rhizopus oryzae empregadas, todos os valores de razão ω-6/ω-3 encontram-se dentro da faixa indicada
pelo Instituto de Medicina (EUA).
5.4.7. Avaliação da ingestão da dose diária recomendada de EPA+DHA
No que diz respeito às doenças cardiovasculares, estudos epidemiológicos e de intervenção
dietética indicam que o consumo de óleo de peixe ou de suplementos de AGPI da família ω-3 que
tenham entre 250 a 500 mg de EPA+DHA, diminuem o risco de mortalidade por doença arterial
coronariana e morte cardíaca súbita. Sendo que a ingestão de 250 mg/dia de EPA e DHA parece ser
suficiente para a prevenção primária em indivíduos saudáveis. Além disso, o consumo de óleos ricos
em AGPI da família ω-3 de cadeia longa durante a gravidez favorece o ganho de peso do bebê e
reduz o risco de parto prematuro (HORVATH, KOLETZKO, SZAJEWSKA, 2007; MAKRIDES;
DULEY; OLSEN, 2006; SZAJEWSKA; HORVATH; KOLETZKO, 2006).
Para o caso de gestantes ou nutrizes, doses extras de 100 a 200 mg de DHA devem ser
adicionadas na dieta para compensar as perdas pelo acúmulo deste ácido graxo em tecidos do feto
em desenvolvimento e em bebês durante o primeiro ano de vida, pois grande parte do DHA é
depositado nos fosfolipídios de membrana do tecido cerebral e da retina (MARTINEZ, 1992;
MARTINEZ, 1994). Além disso, há indícios de que um maior consumo de fontes alimentares de
EPA e DHA ocasiona uma menor incidência de depressão pós-parto (HIBBELN, 2002).
Já em crianças, a ingestão de DHA em níveis de 50 a 100 mg/dia têm sido eficaz para a
função visual e cognitiva, sendo considerados adequados para esse período, tendo em vista que o
DHA também é um componente estrutural importante dos lipídios da retina (STILLWELL e
WASSALL, 2003).
A avaliação da dose diária recomendada para os AGPI da família ω-3 empregada neste estudo
levou em consideração a recomendação dietética típica preconizada pela OMS (Organização Mundial

73
da Saúde), que é de 300 a 500 mg por dia de EPA+DHA. Estes valores foram publicados no Dietary
Reference Intakes para energia e macronutrientes (Institute of Medicine, 2005).
A Tabela 13 apresenta a quantidade, em mg, da mistura de EPA+DHA incorporada nos
óleos vegetais de coco, palma e oliva, após a reação de acidólise catalisada por diferentes lipases.
Para se chegar a estes valores em miligramas, foi estipulado como padrão, a medida de uma
colher sopa (8,0g) e com base na ingestão de apenas uma colher por dia dos produtos obtidos,
chegou-se aos valores explicitados na tabela 13.
Tabela 7: Análise da incorporação de EPA+DHA nos óleos vegetais após a reação de acidólise com ácidos graxos poli-insaturados provenientes do óleo de peixe catalisado por diferentes lipases. Os valores em miligramas foram estipulados com base na ingestão diária de uma colher de sopa (8,0g).
Biocatalisador Óleo de Coco
enriquecido em
EPA+DHA (mg)
Óleo de Palma
enriquecido em
EPA+DHA (mg)
Óleo de Oliva
enriquecido em
EPA+DHA (mg)
Rhizomucor miehei (Lipozyme® RM IM) 676 288 245
Rhizomucor miehei (RM IM LaBiM) 374 79 50
Rhizomucor miehei (Sigma-Aldrich) 272 546 200
Candida antarctica B (Novozym® 435) 662 126 116
Candida antarctica B (Lip. B LaBiM) 392 74 64
Rhizopus oryzae (Sigma-Aldrich) 386 512 95
Rhizopus oryzae (ROL UAB) 586 200 146
Thermomyces lanuginosus (Lipozyme® TL IM) 915 97 135
Thermomyces lanuginosus (Lipozyme® TL 100L) 52 67 170
Candida antarctica A (Novozym® 735) 340 173 14
Candida rugosa (Lipomod™ 34P) n.d. 79 88
Candida rugosa (Amano AY 30) 104 47 62
Pseudomonas fluorescens (Amano AK) 1198 n.d. 37
Burkholderia cepacia (Amano PS) 509 440 455
Os valores são médias aritméticas das análises feitas em duplicata; nd: não detectado. Faixa de 0-100 mg de EPA+DHA; Faixa de 101-300 mg de EPA+DHA e Faixa superior a 300 mg
Ao analisarmos a tabela 13, observamos que o óleo mais suscetível a reação proposta de
enriquecido em EPA e DHA foi o óleo de coco e a lipase com o melhor desempenho para tal reação

74
foi a Amano PS de Burkholderia cepacia, pois foi capaz de promover o enriquecimento nutricional nos
três óleos utilizados como substrato.
Para o óleo de coco as lipases mais indicadas são: (i) Amano AK de Pseudomonas fluorescens; (ii)
Lipozyme® TL IM de Thermomyces lanuginosus; (iii) Lipozyme® RM IM de Rhizomucor miehei; (iv)
Novozym® 435 de Candida antarctica B; (v) Rhizopus oryzae (UAB); (vi) Amano PS de Burkholderia
cepacia. Além do mais, vimos que é possível obter um produto de grande intesse farmacológico que
pode ser utilizado na prevenção de doença coronariana (infarto do miocardio) e de AVC com a
lipase Amano AK de Pseudomonas fluorescens.
Para o óleo de palma, foi possível chegar a dose diária recomendada com a utilização das
lipases de Rhizomucor miehei (Sigma-Aldrich), Rhizopus oryzae (Sigma-Aldrich) e com a Amano PS
Burkholderia cepacia, que foi única a incorporar esses ácidos graxos de forma eficaz no óleo de oliva.
Podemos supor que o enriquecimento parece ser mais dependente do substrato do que das enzimas
empregadas.
De uma forma geral, com a metodologia proposta neste trabalho, foi possível o
desenvolvimento de óloes enriquecidos nutricionalmente que atendessem a necessidade de ingetão
diária preconizada pela OMS (Organização Mundial da Saúde) para os ácidos graxos
eicosapentaenóico e docosahexaenóico.
5.5. Avaliação da Produção de Miméticos do Leite Materno
O objetivo desta parte do trabalho foi sintetizar um composto que apresentasse a
composição em ácidos graxos semelhantes as que são encontradas nos lipídios do leite humano. Para
tal, temos a tripalmitina, que é o substrato mais comum empregado na produção de miméticos do
leite materno, que pode ser substituído por óleos vegetais, por serem economicamente mais viáveis,
tais como a estearina de palma. Além disso, os óleos de cártamo e de soja são duas fontes de ácidos
graxos poli-insaturados que, em conjunto com o óleo de coco, produz uma combinação aceitável de
gordura que pode ser empregada na formulação de gordura de leite infantil (PACKARD, 1982).
A reação proposta nesta investigação foi a acidólise da tripalmitina com o ácido oléico
(C18:1n9) em meio livre de solvente, catalisado por diversas lipases (Figura 23).
Para tal foram utilizadas as seguintes lipases: Rhizomucor miehei (Sigma-Aldrich), Novozym®
435 (Candida antarctica B), Lipozyme® TL IM (Thermomyces lanuginosus), Lipozyme® RM IM (Rhizomucor
miehei), Lipomod™ 34P (Candida rugosa), Rhizopus oryzae (Sigma-Aldrich), ROL UAB (Rhizopus oryzae),
Amano PS (Burkholderia cepacia), Amano AK (Pseudomonas fluorescens), Lipozyme® TL 100L
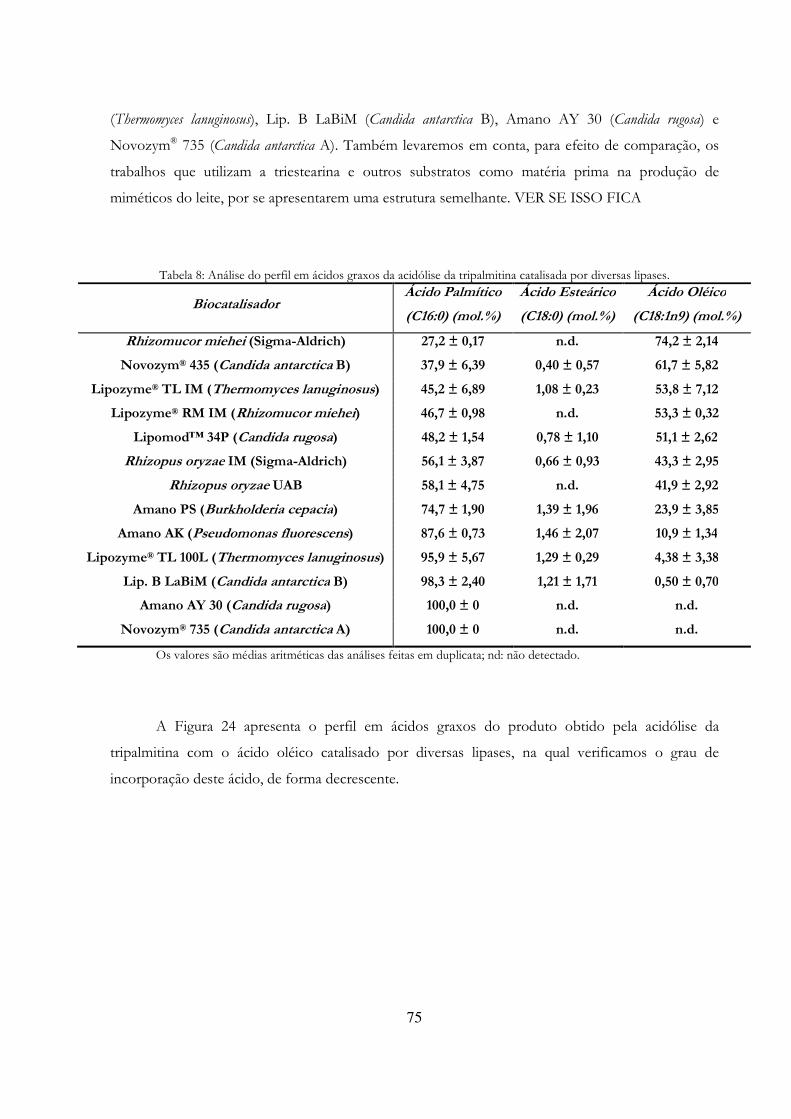
75
(Thermomyces lanuginosus), Lip. B LaBiM (Candida antarctica B), Amano AY 30 (Candida rugosa) e
Novozym® 735 (Candida antarctica A). Também levaremos em conta, para efeito de comparação, os
trabalhos que utilizam a triestearina e outros substratos como matéria prima na produção de
miméticos do leite, por se apresentarem uma estrutura semelhante. VER SE ISSO FICA
Tabela 8: Análise do perfil em ácidos graxos da acidólise da tripalmitina catalisada por diversas lipases.
Biocatalisador Ácido Palmítico
(C16:0) (mol.%)
Ácido Esteárico
(C18:0) (mol.%)
Ácido Oléico
(C18:1n9) (mol.%)
Rhizomucor miehei (Sigma-Aldrich) 27,2 ± 0,17 n.d. 74,2 ± 2,14
Novozym® 435 (Candida antarctica B) 37,9 ± 6,39 0,40 ± 0,57 61,7 ± 5,82
Lipozyme® TL IM (Thermomyces lanuginosus) 45,2 ± 6,89 1,08 ± 0,23 53,8 ± 7,12
Lipozyme® RM IM (Rhizomucor miehei) 46,7 ± 0,98 n.d. 53,3 ± 0,32
Lipomod™ 34P (Candida rugosa) 48,2 ± 1,54 0,78 ± 1,10 51,1 ± 2,62
Rhizopus oryzae IM (Sigma-Aldrich) 56,1 ± 3,87 0,66 ± 0,93 43,3 ± 2,95
Rhizopus oryzae UAB 58,1 ± 4,75 n.d. 41,9 ± 2,92
Amano PS (Burkholderia cepacia) 74,7 ± 1,90 1,39 ± 1,96 23,9 ± 3,85
Amano AK (Pseudomonas fluorescens) 87,6 ± 0,73 1,46 ± 2,07 10,9 ± 1,34
Lipozyme® TL 100L (Thermomyces lanuginosus) 95,9 ± 5,67 1,29 ± 0,29 4,38 ± 3,38
Lip. B LaBiM (Candida antarctica B) 98,3 ± 2,40 1,21 ± 1,71 0,50 ± 0,70
Amano AY 30 (Candida rugosa) 100,0 ± 0 n.d. n.d.
Novozym® 735 (Candida antarctica A) 100,0 ± 0 n.d. n.d.
Os valores são médias aritméticas das análises feitas em duplicata; nd: não detectado.
A Figura 24 apresenta o perfil em ácidos graxos do produto obtido pela acidólise da
tripalmitina com o ácido oléico catalisado por diversas lipases, na qual verificamos o grau de
incorporação deste ácido, de forma decrescente.
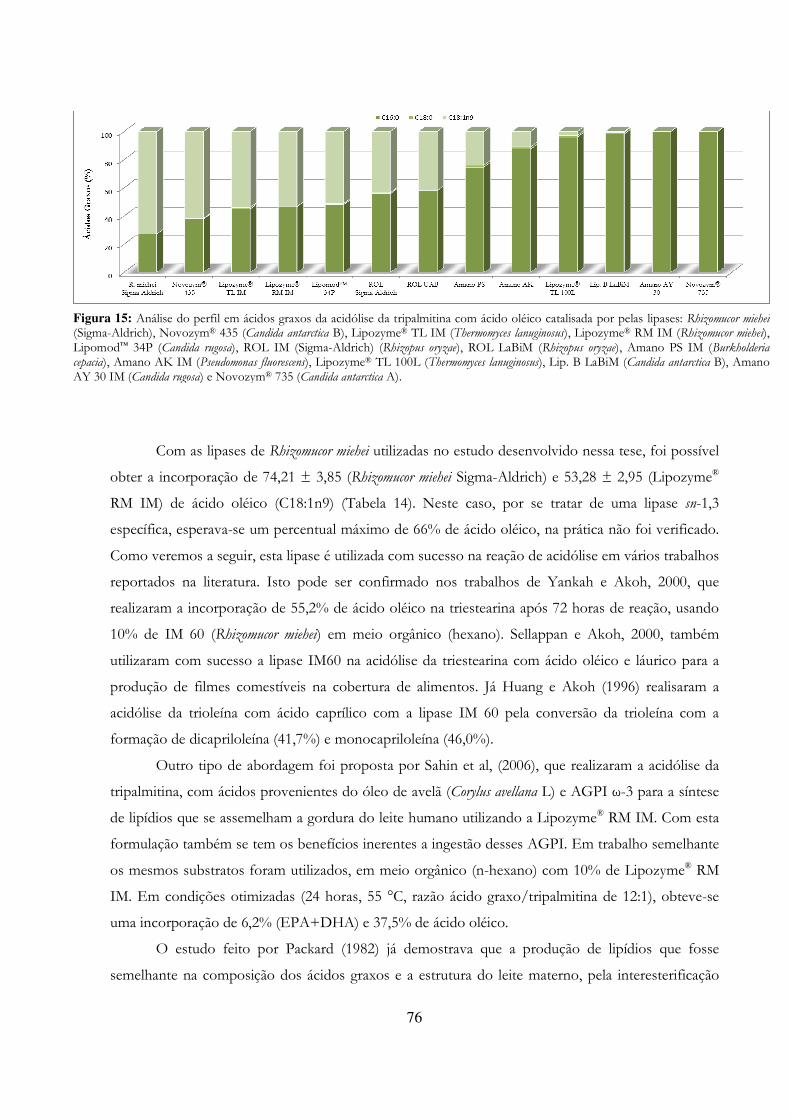
76
Com as lipases de Rhizomucor miehei utilizadas no estudo desenvolvido nessa tese, foi possível
obter a incorporação de 74,21 ± 3,85 (Rhizomucor miehei Sigma-Aldrich) e 53,28 ± 2,95 (Lipozyme®
RM IM) de ácido oléico (C18:1n9) (Tabela 14). Neste caso, por se tratar de uma lipase sn-1,3
específica, esperava-se um percentual máximo de 66% de ácido oléico, na prática não foi verificado.
Como veremos a seguir, esta lipase é utilizada com sucesso na reação de acidólise em vários trabalhos
reportados na literatura. Isto pode ser confirmado nos trabalhos de Yankah e Akoh, 2000, que
realizaram a incorporação de 55,2% de ácido oléico na triestearina após 72 horas de reação, usando
10% de IM 60 (Rhizomucor miehei) em meio orgânico (hexano). Sellappan e Akoh, 2000, também
utilizaram com sucesso a lipase IM60 na acidólise da triestearina com ácido oléico e láurico para a
produção de filmes comestíveis na cobertura de alimentos. Já Huang e Akoh (1996) realisaram a
acidólise da trioleína com ácido caprílico com a lipase IM 60 pela conversão da trioleína com a
formação de dicapriloleína (41,7%) e monocapriloleína (46,0%).
Outro tipo de abordagem foi proposta por Sahin et al, (2006), que realizaram a acidólise da
tripalmitina, com ácidos provenientes do óleo de avelã (Corylus avellana L) e AGPI ω-3 para a síntese
de lipídios que se assemelham a gordura do leite humano utilizando a Lipozyme® RM IM. Com esta
formulação também se tem os benefícios inerentes a ingestão desses AGPI. Em trabalho semelhante
os mesmos substratos foram utilizados, em meio orgânico (n-hexano) com 10% de Lipozyme® RM
IM. Em condições otimizadas (24 horas, 55 °C, razão ácido graxo/tripalmitina de 12:1), obteve-se
uma incorporação de 6,2% (EPA+DHA) e 37,5% de ácido oléico.
O estudo feito por Packard (1982) já demostrava que a produção de lipídios que fosse
semelhante na composição dos ácidos graxos e a estrutura do leite materno, pela interesterificação
Figura 15: Análise do perfil em ácidos graxos da acidólise da tripalmitina com ácido oléico catalisada por pelas lipases: Rhizomucor miehei(Sigma-Aldrich), Novozym® 435 (Candida antarctica B), Lipozyme® TL IM (Thermomyces lanuginosus), Lipozyme® RM IM (Rhizomucor miehei), Lipomod™ 34P (Candida rugosa), ROL IM (Sigma-Aldrich) (Rhizopus oryzae), ROL LaBiM (Rhizopus oryzae), Amano PS IM (Burkholderia cepacia), Amano AK IM (Pseudomonas fluorescens), Lipozyme® TL 100L (Thermomyces lanuginosus), Lip. B LaBiM (Candida antarctica B), Amano AY 30 IM (Candida rugosa) e Novozym® 735 (Candida antarctica A).

77
enzimática de tripalmitina com uma mistura de óleos vegetais, como os de coco, de soja e de
cártamo, utilizando a Lipozyme® RM IM como biocatalisador, poderia ser bem sucedida.
A lipase de Thermomyces lanuginosus, comercialmente conhecida como Lipozyme® TL, tem
recebido atenção atualmente nas pesquisas envolvendo lipídios. A Lipozyme® TL IM é uma
preparação granulada de sílica de grau alimentar de uma lipase microbiana sn-1,3 específica de
Thermomyces lanuginosus produzida por fermentação submersa a partir do micro-organismo Aspergillus
oryzae geneticamente modificado (DEREWENDA, 1994). Com as lipases de Thermomyces lanuginosus
utilizadas no estudo desenvolvido nessa tese, foi possível obter a incorporação de 53,77 ± 7,12 e 4,38
± 1,95 para as formas imobilizada e livre, respectivamente (Tabela 13).
Em trabalhos descritos na literatura, como no estudo feito por Yang; Fruekilde; Xu (2003) da
acidólise do óleo de girassol, foi observado uma menor incorporação de ácido caprílico para a lipase
de Thermomyces lanuginosus em relação a Lipozyme® RM IM, com 10 e 40%, respectivamente. Essas
reações foram conduzidas com uma razão molar óleo de girassol/ácido caprílico de 1:1, 10% (p/p)
de lipase em 5 horas de reação. No entanto, para a reação de transesterificação da tripalmitina (PPP)
com éster etílico (EE) de EPA e DHA, essa diferença foi menor. Em reações otimizadas com razão
molar de substrato de 5:1 (EE/PPP), 20% (p/p) de lipase em 20 horas de reação, houve um
aumento para 40 e 45% de incorporação para Thermomyces lanuginosus e Lipozyme® RM IM,
respectivamente.
Já Nagachinta e Akoh (2013) produziram lipídios enriquecidos com os ácidos graxos
araquidônico (ARA) e docosahexaenóico (DHA) pela acidólise da tripalmitina (PPP) usando a
Lipozyme® TL IM. Chegou-se a incorporação de 18% de ARA e 15% de DHA com uma razão
molar de 1:9 (PPP/AGL), 10% (p/p) de biocatalisador em meio orgânico (hexano), a 60 °C durante
24 horas.
No trabalho desenvolvido por Rocha-Uribe e Hernandez (2004), foi possível realizar a
acidólise da tripalmitina e do óleo de coco (fonte de ácidos graxos de cadeia média) com uma mistura
de ácidos graxos livres contendo 73% de CLA (ácido linoleico conjugado oriundos do óleo de
cártamo) como doador de grupos acila, catalisado pela Lipozyme® IM, com sucesso.
Com a utilização da lipase Novozym® 435 (Candida antarctica B) a incorporação de ácido
oléico foi de 61,68 ± 5,82 e de 0,50 ± 0,70 para Lip. B LaBiM (Tabela 13). Podemos concluir que
para este caso a lipase home made não foi capaz de catalisar a reação desejada. Resultado semelhante
ao encontrado com a lipase comercial também foi observados por Hamam e Shahidi (2007) no
estudo da acidólise da triestearina (SSS) com diferentes tipos de ácidos graxos (AG), aonde se chegou
a incorporação de 58,2% de ácido oléico e 55,9% de ácido linolênico, com uma razão molar AG/SSS
3:1, 10% (p/p) de enzima, 2% de água em meio orgânico (hexano), a 45 °C em 24 horas de reação.

78
Senanayake e Shahid, 1999 relataram que a Novozym® 435 exibiu um alto grau de
incorporação de 25,8 a 28,7%, para o ácido docosahexaenóico (DHA) no óleo de prímola.
No estudo da acidólise da tripalmitina com a estearina de palma, Quin et al, (2012) chegaram
a uma incorporação de de 31% de ácidos graxos com a utilização da Lipozyme® RM IM e da
Lipozyme® TL IM; Já com a Novozym 435®, praticamente não há incorporação. Diferente do
observado no estudo de Karabulut; Durmaz; Hayaloglu (2010), que relataram bons resultados na a
incorporação de ácidos graxos na seguinte ordem: C18:3n3> C18:2n6> C18:1n9 na acidólise da
tripalmitina, catalisada pela Lipozyme® TL IM, Lipozyme® RM IM e Novozym® 435. Em outro
estudo de Karabulut; Durmaz; Hayaloglu (2009) realizaram a acidólise de triacilmonoácidos de
diferentes tamanhos de cadeia carbônica (C6:0 a C18:0) com acido oléico catalisado pelas lipases
Lipozyme® TL IM, Lipozyme® RM IM e Novozym® 435. Com relação à acidólise da tripalmitina,
houve a incorporação de 12, 18 e 15%, respectivamente, obtida com uma razão molar de 1:1
(PPP/AGL), 10% (p/p) de biocatalisador a 60 °C durante 6 horas de reação.
Na avaliação da acidólise catalisada pela Novozym® 735 (Candida antarctica A), não foi
possível obter incorporação do ácido oléico na tripalmitina nas condições testadas e também não
foram encontrados trabalhos relacionados a essa pesquisa que pudessem ajudar a compreender, ou
fazer qualquer tipo de comparação, com a reação proposta.
Com o emprego da lipase de Candida rugosa na forma comercial Lipomod™ 34P e Amano AY
30 foi possível obter uma incorporação de 51,07 ± 2,62 com a primeira; já com a segunda, não foi
detectado o ácido oléico (Tabela 13).
Como parâmetro temos o trabalho de Hamam and Shahidi (2007) mostrando que a lipase de
Candida rugosa foi capaz de inserir uma excepcional quantidade de ácido linoleico (75,8%), ácido α-
linolênico (74,8%), e ácido linoleico conjugado (53,5%) na triestearina com uma razão molar ácido
graxo/triestearina 3:1, 10% de enzima, 2% de água em meio orgânico (hexano), a 45 °C por 24
horas.
Com as lipases Amano PS de Burkholderia cepacia e Amano AK de Pseudomonas fluorescens, foi
possível obter a incorporação de 23,92 ± 3,85 e 10,91± 1,34, respectivamente. Em trabalho
semelhante, Qin et al (2012) conseguiram a incorporação de 30,39% na acidólise da tripalmitina
utilizando a lipase PS de Burkholderia cepacia. Para o caso da Pseudomonas fluorescens, não foram
encontrados trabalhos na literatura para que uma melhor discussão pudesse ser feita.
Para as lipases de Rhizopus oryzae, as formas comercial (ROL IM Sigma-Aldrich) e ROL
LaBiM, obtiveram resultado semelhante, com cerca de 42% de incorporação do ácido oléico na
tripalmitina (Tabela 13). Em estudo semelhante, Tecelão et al, (2012), utilizando o mesmo

79
biocatalisador, conseguiram a incorporação de 30 e 22% de ácido oléico com a imobilização desta
lipase em Lewatit® VP OC 1600 e Accurel® MP 1000, respectivamente.
Acredita-se que lipídios, como o proposto neste estudo, podem ser utilizados como
ingredientes em fórmulas infantis com intuito de proporcionar vantagens semelhantes às encontradas
em miméticos do leito materno. Ademais, a fortificação de leite para lactentes com ácidos graxos de
cadeia longa poli-insaturados pela adição de óleo de peixe está se tornando cada vez mais popular
para os benefícios adicionais de saúde. No entanto, o sucesso da produção desses lipídios para a
formulação de leite infantil pode ser impedida pela sua alta susceptibilidade à deterioração oxidativa
(AKOH e MOUSSATA, 2001).
De uma forma geral, o tipo de enzima usada para a catálise no processo de transesterificação
é um fator determinante na produção e composição do produto, pois a conformação da enzima é
afetada por vários fatores no microambiente do meio reacional, isto inclui o solvente, a solubilidade
dos substratos, quantidade de água livre, pH do meio e temperatura; e quaisquer outras mudanças
conformacionais que possam afetar a atividade catalítica da enzima.
5.6. Produção de Lipídios Estruturados M-L-M de Baixo Teor Calórico a partir da Acidólise
do Óleo de Peixe com Ácido Graxo de Cadeia Média
Lipídios que contêm ácidos graxos de cadeia média nas posições sn-1,3 e ácidos graxos de
cadeia longa na posição sn-2 do triacilglicerol, fornecem menos calorias em comparação com os
lipídios regulares e são denominados lipídios estruturados de “baixas calorias ou reduzido teor
calórico”. São projetados principalmente para aplicações nutricionais especiais, para atender à
crescente necessidade de alimentos mais saudáveis e prevenir a obesidade. Assim, os ácidos graxos de
cadeia média são usados tanto por ser uma fonte de energia facilmente disponível, como por ser
produto de baixo teor calórico. Somado a isso, temos os benefícios de saúde do óleo de peixe, como
a melhoria na função imunológica, redução dos níveis de triglicerídeos e aumento da ação anti-
inflamatória, estão bem documentados e são atribuídas principalmente aos ácidos graxos poli-
insaturados da família ômega-3, destacando-se os ácidos eicosapentaenóico (EPA - 20:5n3) e
docosahexaenóico (DHA - 22:6n3) (ABBAS; MOHAMED; JAMILAH, 2009; MILES E CALDER,
2012; HORROCKS E YEO, 1999).
A reação proposta nesta investigação foi a acidólise do óleo de peixe com o ácido octanóico
em meio livre de solvente, catalisado por diversas lipases.
A síntese desses lipídios de menor densidade energética proposta neste trabalho utilizou
como biocatalisadores as lipases: Lipozyme® RM IM (Rhizomucor miehei); Lipozyme® TL 100L
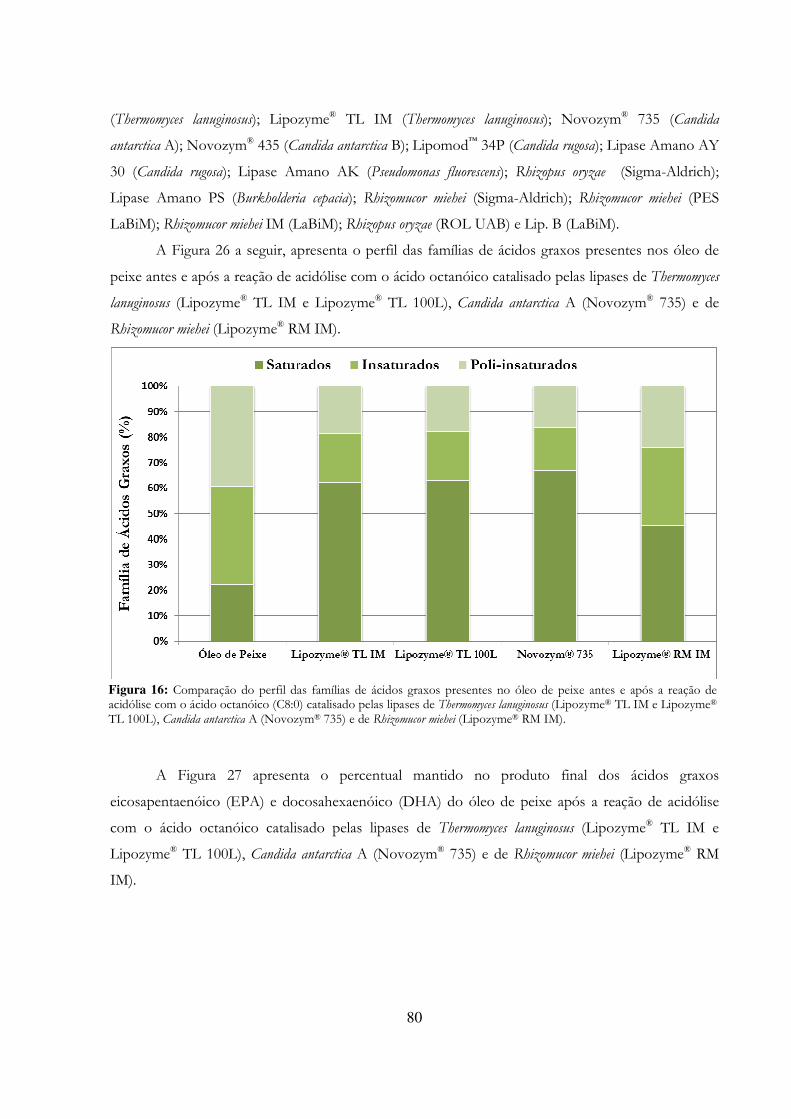
80
(Thermomyces lanuginosus); Lipozyme® TL IM (Thermomyces lanuginosus); Novozym® 735 (Candida
antarctica A); Novozym® 435 (Candida antarctica B); Lipomod™ 34P (Candida rugosa); Lipase Amano AY
30 (Candida rugosa); Lipase Amano AK (Pseudomonas fluorescens); Rhizopus oryzae (Sigma-Aldrich);
Lipase Amano PS (Burkholderia cepacia); Rhizomucor miehei (Sigma-Aldrich); Rhizomucor miehei (PES
LaBiM); Rhizomucor miehei IM (LaBiM); Rhizopus oryzae (ROL UAB) e Lip. B (LaBiM).
A Figura 26 a seguir, apresenta o perfil das famílias de ácidos graxos presentes nos óleo de
peixe antes e após a reação de acidólise com o ácido octanóico catalisado pelas lipases de Thermomyces
lanuginosus (Lipozyme® TL IM e Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e de
Rhizomucor miehei (Lipozyme® RM IM).
A Figura 27 apresenta o percentual mantido no produto final dos ácidos graxos
eicosapentaenóico (EPA) e docosahexaenóico (DHA) do óleo de peixe após a reação de acidólise
com o ácido octanóico catalisado pelas lipases de Thermomyces lanuginosus (Lipozyme® TL IM e
Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e de Rhizomucor miehei (Lipozyme® RM
IM).
Figura 16: Comparação do perfil das famílias de ácidos graxos presentes no óleo de peixe antes e após a reação de acidólise com o ácido octanóico (C8:0) catalisado pelas lipases de Thermomyces lanuginosus (Lipozyme® TL IM e Lipozyme®TL 100L), Candida antarctica A (Novozym® 735) e de Rhizomucor miehei (Lipozyme® RM IM).
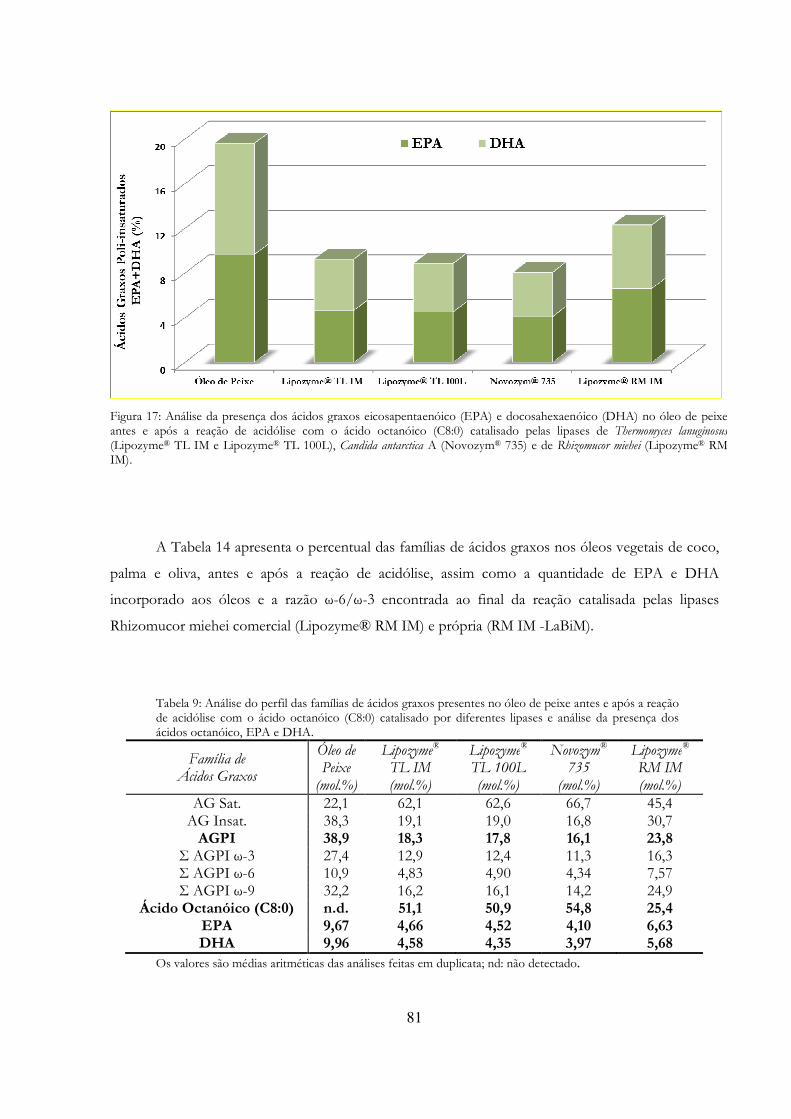
81
A Tabela 14 apresenta o percentual das famílias de ácidos graxos nos óleos vegetais de coco,
palma e oliva, antes e após a reação de acidólise, assim como a quantidade de EPA e DHA
incorporado aos óleos e a razão ω-6/ω-3 encontrada ao final da reação catalisada pelas lipases
Rhizomucor miehei comercial (Lipozyme® RM IM) e própria (RM IM -LaBiM).
Tabela 9: Análise do perfil das famílias de ácidos graxos presentes no óleo de peixe antes e após a reação de acidólise com o ácido octanóico (C8:0) catalisado por diferentes lipases e análise da presença dos ácidos octanóico, EPA e DHA.
Família de Ácidos Graxos
Óleo de Peixe (mol.%)
Lipozyme® TL IM (mol.%)
Lipozyme® TL 100L (mol.%)
Novozym® 735
(mol.%)
Lipozyme® RM IM (mol.%)
AG Sat. 22,1 62,1 62,6 66,7 45,4 AG Insat. 38,3 19,1 19,0 16,8 30,7 AGPI 38,9 18,3 17,8 16,1 23,8
Σ AGPI ω-3 27,4 12,9 12,4 11,3 16,3 Σ AGPI ω-6 10,9 4,83 4,90 4,34 7,57 Σ AGPI ω-9 32,2 16,2 16,1 14,2 24,9
Ácido Octanóico (C8:0) n.d. 51,1 50,9 54,8 25,4 EPA 9,67 4,66 4,52 4,10 6,63 DHA 9,96 4,58 4,35 3,97 5,68
Os valores são médias aritméticas das análises feitas em duplicata; nd: não detectado.
Figura 17: Análise da presença dos ácidos graxos eicosapentaenóico (EPA) e docosahexaenóico (DHA) no óleo de peixe antes e após a reação de acidólise com o ácido octanóico (C8:0) catalisado pelas lipases de Thermomyces lanuginosus (Lipozyme® TL IM e Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e de Rhizomucor miehei (Lipozyme® RM IM).

82
Com a metodologia empregada nesta pesquisa, os melhores resultados de incorporação do
ácido octanóico no óleo de peixe foram obtidos com as lipases de Thermomyces lanuginosus, com 50,99
± 1,98 e 51,06 ± 0,18, para as formas livre (Lipozyme® TL 100L) e imobilizada (Lipozyme® TL IM),
respectivamente; e com a de Candida antarctica A (Novozym® 735) chegou-se a 54,85 ± 2,34. Apesar
da menor incorporação, mas não menos importante, foi possível chegar a 25,38 ± 3,75 com a lipase
de Rhizomucor miehei (Lipozyme® RM IM) (Tabela.14).
Os resultados obtidos estão de acordo com trabalhos descritos na literatura, tanto para a
acidólise do óleo de peixe, quanto à de substratos semelhantes. Dentre eles podemos citar o trabalho
de Akoh e Moussata (2001), que realizaram a acidólise dos óleos de peixe (savelha) e de canola pelo
ácido caprílico (C8:0) catalisada pela Lipozyme® RM IM, onde foi possível adicionar 29,5% e 40,1%
de C8:0, respectivamente.
Em trabalho semelhante, Jennings e Akoh (2001) realizaram a incorporação 31,1 ± 4,6l% de
ácido cáprico (C10:0) no óleo de peixe (savelha) catalisado pela IM 60 de Rhizomucor miehei em meio
orgânico (n-hexano) e de 28,8 ± 4,7l% de C10:0 na ausência de solvente, obtidos com a razão molar
óleo/C10 de 1:2, 10% (p/p) de IM 60, a 55 °C em 24 horas de reação. Os mesmos autores já haviam
conseguido anteriormente Jennings e Akoh (1999) nas mesmas condições 43,0 ± 1,6%, em meio
orgânico (n-hexano) e 31,8 ± 8,5l%, na ausência de solvente; porém, após otimização dos parâmetros
chegou-se a uma incorporação de 65% em meio orgânico, com uma razão molar óleo/C10:0 de 1:8;
já em meio livre de solvente, 56% foi alcançado com uma razão molar óleo/C10 de 1:6. Ambas com
10% (p/p) de IM60 a 55 °C por 24 horas de reação.
Com o uso de óleos vegetais, resultados similares também foram observados. No trabalho
desenvolvido por Lee e Akoh (1998) com a acidólise do óleo de amendoim pelo com ácido caprílico
(C8:0) catalisado pela lipase IM60 (Rhizomucor miehei), onde chegou-se a 30% C8:0, com uma razão
molar óleo/C8 de 1:2, 2% de IM60 a 50 °C por 72 h de reação. Jennings e Akoh (2000) utilizando a
mesma lipase, conseguiram incorporações de ácido cáprico (C10:0) no óleo de gergelim na faixa de
28,3 ± 3,5% e 25,7 ± 4,3 mol% de C10:0 com a razão molar óleo/C10 de 1:2, 10% (p/p) de IM 60, a
55 °C, em meio orgânico (n-hexano) e na sua ausência, respectivamente, em 24 horas de reação.
Após otimização, chegaram a 62% em meio orgânico com uma razão molar óleo/C10 de 1:7, com
15% (p/p) de IM60 a 55 °C por 24 horas de reação.
No trabalho desenvolvido por Gokçe; Yesilçubuk; Ustun (2013) foi feita a acidólise do óleo
da semente de Echium (Echium vulgare) com ácido láurico (C12:0) catalisado pela Lipozyme® RM IM.
Em condições otimizadas (razão molar C12:0/óleo de 5:1, 5% de RM IM, em meio orgânico
(hexano) por 4 horas de reação, obteve-se uma incorporação de 43% de C12:0.

83
No trabalho desenvolvido por Nunes et al, 2012 com a otimização da acidólise do óleo de
oliva com os ácidos caprílico (C8:0) e cáprico (C10:0) catalisado pela lipase heterologa de Rhizopus
oryzae (r-ROL) imobilizada em Eupergit® C, em meio livre de solvente por 24 horas de reação, foi
possível obter a incorporação de 21,6 e 34,3 mol.% dos ácidos caprílico e cáprico, respectivamente.
Dados estes que não foram observados em nosso estudo, usando a mesma lipase, porém a mesma
não estava imobilizada.
Em um estudo interessante, Khodadadi et al., 2013 (318) avaliaram a interesterificação do
óleo de linhaça com tricaprilina em meio orgânico utilizando as lipases Lipozyme® TL IM (2% p/v
a 50 °C por 4h), Novozym® 435 (1% p/v a 50 °C por 24h) e Lipozyme® RM IM (2% p/v a 30 °C
por 8 horas) e apontaram um rendimento, em condições otimizadas, na seguinte ordem, Novozym®
435 > Lipozyme® TL IM > Lipozyme® RM IM.
Com a utilização de substratos “sintéticos” de diferentes tamanhos de cadeia, tais como a
trioleína, triestearina e tributirina, observamos alguns resultados interessantes, como os encontrados
por Fomuso e Akoh (1997) na acidólise da trioleína (OOO) com os ácidos capróico (C6:0) e butírico
(C4:0) catalisado por várias lipases, sendo a IM60 de Rhizomucor miehei a mais eficaz. Com ela foi
possível gerar 13% de trioleína (não modificada) e triacilgliceróis dissubstituído (49%) e
monossubstituído (38%), contendo os ácidos graxos de cadeia curta. Isto foi obtido com a razão
molar OOO/C6/C4 de 1:4:4, 10% (p/p) de IM 60, a 55 °C, em meio orgânico (n-hexano) em 24
horas de reação.
Já Yankah e Akoh (2000) conseguiram 37% de incorporação de ácido caprílico (C8:0) na
triestearina (SSS) catalisado pela lipase IM60 (Rhizomucor miehei). Este resultado foi obtido com uma
razão molar SSS/C8 de 1:6, 5% de IM60, em meio orgânico (n-hexano) a 50 °C por 12 horas de
reação.
Bektas et al, 2008 estudaram acidólise da tripalmitina (PPP) pelo ácido cáprico (C10:0)
catalisado pela Lipozyme® TL IM em meio orgânico (hexano). Em condições otimizadas (razão
molar C10/PPP de 6,5:1, 11.6% de TL IM, em 9 horas de reação) obteve-se 45% de incorporação de
C10:0. Resultado semelhante foi encontrado por Zhao et al, 2007, onde chegaram incorporação de
50% de C10:0 na banha de porco, com a utilização da Lipozyme® TL IM (10% p/p) com uma razão
molar C10:0/banha de porco de 2:1, a 55 °C por 24 horas de reação.
Em uma abordagem diferente, Han et al, (2011) realizaram interesterificação da tributirina
(BBB) com o estearato de metila (St-ME), catalisado pela Lipozyme® RM IM. Em condições
otimizadas chegou-se a inserção 78% de ácido esteárico com uma razão molar St-ME:BBB de 2:1,
10% (p/p) de RM IM a 65 °C em 7 horas de reação.

84
Para as lipases de Candida antarctica B, tanto a comercial (Novozym® 435) quanto a Lip. B
LaBiM a incorporação não foi satisfatória, pois foi obtido apenas 1,8 e 0,52% de incorporação,
respecticamente; assim como as de Rhizomucor miehei (RM PES LaBiM e RM IM LaBiM) com 0,11 e
0,13% de incorporação, respectivamente. Com a utilização da lipase de Rhizopus oryzae, tanto a forma
comercial (ROL Sigma-Aldrich), quanto a ROL LaBiM, não foi detectado a presença do ácido
octanóico. O mesmo foi observado para as lipases de Candida rugosa (Lipomod™ 34P e Amano AY
30), Pseudomonas fluorescens (Amano AK), Burkholderia cepacia (Amano PS) e para a de Rhizomucor miehei
(Sigma-Aldrich).
Alguns trabalhos descritos na literatura demostrarem que a lipase de Pseudomonas fluorescens
tem uma boa utilização em reações de hidrólise, como por exemplo, nos óleos de sardinha
(CHAKRABORTY e RAJ, 2009), atum e concentrados de ácidos graxos poli-insaturados,
principalmente de EPA e DHA (RAKSHIT; VASUHI; KOSUGI, 2000) e outras fontes de óleos
(KOJIMA e SHIMIZU, 2003).
Com relação a lipase Burkholderia cepacia alguns trabalhos citam a sua utilização para a
produção de biodiesel, utilizando como matéria prima o óleo de pinhão manso e metanol (YOU et
al, 2013); na incorporação de ácidos graxos de diferentes tamanhos de cadeia carbônica (C4:0 a
C18:1n9) na molécula de glicerol (FU e PARKIN, 2004); na etanólise do óleo de rícino (BARON et
al, 2004), entre outras matrizes oleosas, mas para o estudo nesta reação específica com o óleo de
peixe ela também não foi eficaz.
A diferença nos resultados pode ser atribuída às diferenças de acidez e propriedades físicas
do ácido graxo no meio reacional. A perda de atividade lipásica na presença de concentrações
elevadas de ácidos graxos livres tem sido associada a acidificação da fase aquosa e o aumento da
solubilidade dos ácidos graxos de cadeia curta (WILLIS e MARANGONI, 1998).
Sabe-se que o sítio ativo de muitas lipases é recoberto por uma alça helicoidal anfipática ou
tampa, tornando o mesmo inacessível ao substrato (DEREWENDA, 1994). Na reação de acidólise, a
abertura da tampa pode ser impedida pela presença de uma grande quantidade de ácidos graxos livres
(YANG; FRUEKILDE; XU, 2003). Isto poderia explicar uma menor atividade catalítica na reação
de acidólise para algumas lipases.
Como já foi explicitado anteriormente, altas concentrações de ácidos graxos livres podem
levar a níveis elevados de grupamentos de ácidos carboxílicos livres ou ionizados, que podem
acidificar a fase micro aquosa circundante da lipase ou causar a dessorção da água da interface. Além
disso, os ácidos graxos de cadeia curta e média poderiam se particionar longe da interface aquosa que
circunda a enzima, devido à sua maior solubilidade em água, de modo que esse particionamento
limite o acesso do substrato a essa interface (YANKAH e AKOH, 2000).

85
Além disso, a mudança na quantidade de água em torno (superfície) influencia na polaridade,
flexibilidade e na acessibilidade do sítio ativo para o substrato. Além disso, essa acessibilidade pode
ser influenciada pela variação de comprimento de cadeia, a saturação, e a solubilidade do substrato
(FLORES et al, 2000).

86
6. CONCLUSÕES
Alguns resultados obtidos com esta tese mostram que é factível a geração de produtos (óleos
alimentícios) com o desenvolvimento de tecnologia enzimática nacional, reduzindo no futuro a
dependência por parte das multinacionais que dominam o mercado de enzimas.
Os processos desenvolvidos neste estudo podem se beneficiar da crescente demanda por
lipídios estruturados para aditivos nas indústrias de nutracêuticos, explorando a potencialidade do
beneficiamento de algumas matrizes lipídicas de baixo custo e principalmente do desenvolvimento
biocatalisadores produzidos em território nacional.
6.1. Avaliação do Potencial dos Biocatalisadores Vegetais
� A metodologia aplicada para obtenção de lipases a partir de sementes germinadas por meio
da produção de acetone powder não se mostrou eficaz para os casos analisados, tendo em vista
que essa preparação, de certa forma, pode ter inativado as possíveis lipases presentes no
estágio de germinação.
6.2. Enriquecimento Nutricional de Óleos Vegetais com Ácidos Graxos Poli-insaturados
� Foi possível gerar óleos vegetais enriquecidos nutricionalmente pela inserção de ácidos graxos
poli-insaturados que atendessem à ingestão diária recomendada preconizada pela OMS
(Organização Mundial da Saúde) para os ácidos graxos eicosapentaenóico (EPA) e
docosahexaenóico (DHA). Óleos que podem ser comercializados como suplementos
alimentares de maior valor agregado;
� A lipase de Rhizomucor miehei produzida em laboratório (LaBiM) se mostrou eficaz no
enriquecimento dos óleos vegetais, principalmente com relação ao óleo de coco, onde foi
possível gerar um produto que além de suprir as necessidades diárias de EPA+DHA,
também pode ser recomendado para a suplementação dietética de mulheres gestantes ou que
estejam no período de amamentação; Além disso, esta lipase foi capaz de melhorar a razão ω-
6/ω-3 dos óleos;

87
� Com a Lip. B (LaBiM) também foi possível obter excelentes resultados com o óleo de coco,
haja vista que também atingiu-se a recomendação para a ingestão de EPA/DHA no produto
gerado. Com ela também foi possível melhorar a razão ω-6/ω-3 dos óleos;
� Para a lipase de Rhizopus oryzae (ROL UAB), o destaque vai para o enriquecimento do óleo de
coco, onde o resultado final de incorporação foi melhor do que o encontrado com a lipase
comercial; também houve melhora na razão ω-6/ω-3 dos óleos;
� Foi possível gerar um produto de elevado potencial farmacológico, no enriquecimento
nutricional do óleo de coco com a aplicação da Lipozyme® TL IM e da Amano AK, que
pode ser utilizado na prevenção de doença coronariana (infarto do miocárdio) e de acidente
vascular cerebral (AVC).
� Acredita-se que os lipídios produzidos neste estudo, além de serem funcional e
nutricionalmente benéficos, que eles possam ser utilizados como importantes ingredientes
em variadas formulações alimentares;
6.3. Avaliação da Produção de Miméticos do Leite Materno
� Acredita-se que lipídios, como os obtidos neste estudo, podem ser utilizados como
ingredientes em fórmulas infantis com intuito de proporcionar vantagens semelhantes às
encontradas em miméticos comerciais do leito materno.
� Com a lipase de Rhizopus oryzae (ROL UAB) foi possível obter um desempenho semelhante
ao encontrado com a utilização da lipase comercial de referência.
6.4. Produção de Lipídios Estruturados M-L-M de Baixo Teor Calórico.
� Foi possível gerar lipídios de menor densidade energética com as lipases de Thermomyces
lanuginosus (Lipozyme® TL IM e Lipozyme® TL 100L), Candida antarctica A (Novozym® 735) e
de Rhizomucor miehei (Lipozyme® RM IM).

88
� Os óleos gerados neste processo obtidos como resultado deste estudo têm, o potencial de
oferecer inúmeros benefícios a saúde associados aos ácidos graxos poli-insaturados ômega-3
e ômega-6;
7. PERSPECTIVAS / TRABALHOS FUTUROS
� Aprimoramento de metodologias para análise estereoespecífica de triacilgliceróis;
� Estudo cinético das reações de esterificação para as lipases mais promissoras;
� Otimização das condições de reação para melhorar a produtividade;
� Busca/descoberta de novas fontes de lipases a partir de micro-organismos e/ou vegetais;
� Análise da viabilidade econômica para o emprego de biocatalisadores home made em processos
de modificação de óleos e gorduras;
� Design de alimentos com lipídios bioativos para a melhoria na promoção da saúde.
� Estabilidade dos produtos gerados
8. CONSIDERAÇÕES FINAIS
A produção de lipídios estruturados catalisada por lipases ainda não está amplamente
implementada industrialmente, tendo em vista que a tecnologia enzimática ainda está em franco
desenvolvimento. No entanto, com o seu avanço, não há dúvida de que a aplicação desses
biocatalisadores para a modificação de óleos e gorduras vai disparar em um futuro próximo.
Apesar desse cenário favorável, a utilização das lipases esbarra na politica de preços
proibitivo praticado pelas multinacionais que dominam o mercado. O que acaba levando alguns
processos a serem economicamente inviáveis.
Portanto, o mercado nacional de enzimas só poderá se solidificar se houver oferta de
preparações enzimáticas de qualidade com custos relativamente baixos. Nesse contexto, a
fermentação em meio sólido apresenta-se como uma tecnologia extremamente apropriada, uma vez
que a produção de enzimas pode ser feita por uma técnica simples e de baixo custo. Assim como o
uso de enzimas imobilizadas, que oferece importantes vantagens em processos industriais e tem sido
utilizado extensivamente por facilitar o desenvolvimento de processos em escala comercial. Somado
a isso, o desenvolvimento de um sistema mais eficiente de expressão em combinação com a melhoria

89
das tecnologias de engenharia de proteína devem permitir que a indústria seja impulsionada para a
produção de lipases em larga escala, tornando o seu custo de aplicação mais competitivo.
O conhecimento profundo destes processos pode ajudar no desenvolvimento de lipases
customizadas para aplicações específicas e em médio/longo prazo, abrir novas perspectivas das mais
diversas aplicações, tornando as lipases tão importantes industrialmente no futuro, como as proteases
são atualmente.

90
9. REFERENCIAS BIBLIOGRÁFICAS
ABBAS, K. A.; MOHAMED, A.; JAMILAH, B. Fatty acids in fish and beef and their nutritional values: A review. Journal of Food, Agriculture and Environment, v. 7, p. 37-42, 2009. ABDELKAFI, S.; BAROUH, N.; FOUQUET, B.; FENDRI, I.; PINA, M.; SCHEIRLINCKX, F., VILLENEUVE, P.; CARRIÈRE, F. Carica papaya Lipase: A Naturally Immobilized Enzyme with Interesting Biochemical Properties. Plant Foods Human Nutrition, v. 66, p. 34-40, 2011. ABIGOR, R. D.; UADIA, P. O.; FOGLIA, T. A.; HAAS, M. J.; SCOTT, K.; SAVARY, B. J. (2002). Partial purification and properties of lipase from germinating seeds of Jatropha curcas L. Journal of American Oil Chemists Society, v. 79 (11), p. 1123–1126, 2002. ACKMAN, R. G.; CUNNANE, S. C. Long-chain polyunsaturated fatty acids: Sources, biochemistry, and nutritional/clinical applications. Advances in Applied Lipid Research, v. 1, p. 161-215, 1992. ADAMCZAK, M. The application of lipases in modifying the composition, structure and properties of lipids – a review. Polish Journal Of Food And Nutrition Sciences, v. 13, No 1, p. 3-10, 2004. ADLERCREUTZ P. Enzyme-catalyzed lipid modification. Biotechnology & Genetic Engineering Reviews, v. 12, p. 231-254, 1994. ADRIO, J. L.; ARNOLD L. Demain. Microbial Enzymes: Tools for Biotechnological Processes. Biomolecules, v.4, p. 117-139, 2014. AFOLABI, O. A.; OLOGUNDE, M. A.; ANDERSON, W. A.; READ, J. S.; DACOSTA, M. D.; EPPS, F. A.; AYORINDE, F. O. The Use of Lipase (Acetone Powder) from Vernonia galemensis in Fatty Acids, Analysis of Seed Oils. Journal of Chemical Technology and Biotechnology, v. 51, p. 41-46, 1991. AKOH, C. C. Structured lipids – enzymatic approach. INFORM, v. 6, p. 1055-1061, 1995. AKOH, C. C. Structured lipids. In Food Lipids Chemistry, Nutrition, and Biology, 2nd ed.; Akoh, C. C., Min, D. B., Eds.; Dekker: New York, p, 877-908, 2002. AKOH, C. C.; MIN, D. B. Food Lipids: Chemistry, Nutrition, and Biotechnology. Published by Marcel Dekker, Inc, New York, NY, USA, p, 21, 2002. AKOH, C. C.; MOUSSATA, C. O. Characterization and oxidative stability of enzymatically produced fish and canola oil-based structured lipids. Journal of the American Oil Chemists Society, v. 78, p. 25-30, 2001. ARAUJO, R.; CASAL, M.; CAVACO-PAULO, A. Application of enzymes for textile fibres processing. Biocatalysis and Biotransformation, v 26(5), p. 332-349, 2008. ARNAU, C; RAMÓN, R; CASAS, C; VALERO, F. Optimization of the heterologous production of a Rhizopus oryzae lipase in Pichia pastoris system using mixed sub-strates on controlled fed-batch bioprocess, Enzyme and Microbial Technology, v. 46, p. 494-500, 2010.

91
ASTORG, P. Dietary fatty acids and colorectal and prostate cancer: epidemiological studies. Bulletin du Cancer, v. 92, p. 670-684, 2005. ATOMI, H.; SATO, T.; KANAI, T. Application of hyperthermophiles and their enzymes. Current Opinion in Biotechnology, v. 22, p. 618-626, 2011. AUERBACH, M. H.; KLEMANN, L. P E HEYDINGER, J. A. Reduced-energy lipids, in Structured and Modified Lipids. Gunstone, F.D., Ed., Marcel Dekker, New York, p. 485-510, 2001. AU-KBC Research Center. Applications of Lipases., Life Sciences, Anna University, Chennai, India. Disponível em: www.au-kbc.org/beta/bioproj2/introduction.html. AVELAR, M .H. M.; CASSIMIRO, D. M. J.; SANTOS, K. C.; DOMINGUES, R. C. C.; CASTRO, H. F.; MENDES, A. A. Hydrolysis of vegetable oils catalyzed by lipase extract powder from dormant castor bean seeds. Industrial Crops and Products, v. 44, p. 452-458, 2013. AXELSON, M.; SJÖVALL, J.; GUSTAFSSON, B. E.; SETCHELL, K. D. R. Origin of lignans in mammals and identification of a precursor from plants. Nature, v. 298, p. 659-660, 1982. BABAYAN, V. K. Medium chain triglycerides and structure lipids. Lipids, v. 22, p. 417-420, 1987. BARON, M. H.; REVAULT, M.; SERVAGEANT-NOINVILLE, S.; ABADIE, J.; QUIQUAMPOIX, H. Chymotrypsin adsorption on Montmorillonite: enzymatic activity and kinetic FTIR structural analysis, Journal of Colloid and Interface Science, v. 214, p. 319-332, 1999. BARON, A. M.; BAROUH, N.; BAREA, B.; VILLENEUVE, P.; MITCHELL, D. A.; KRIEGER, N. Transesterification of castor oil in a solvent-free medium using the lipase from Burkholderia cepacia LTEB11 immobilized on a hydrophobic support. Fuel, v. 117, p. 458-462, 2014. BASS, J. K.; CHAN, G. M. Calcium nutrition and metabolism during infancy. Nutrition, 22, 1057-1066, 2006. BCC Research (Business Communications Company, Inc). Report code: FOD027C. Weight Loss Markets for Products and Services; BCC Research: Wellesley, MA, USA, 2010. BCC Research (Business Communications Company, Inc). Report Code: FOD013D. Nutraceuticals: Global Markets and Processing Technologies, BCC Research, Wellesley, MA, USA, 2011a. BCC Research (Business Communications Company, Inc). Report code: FOD027C. Weight Loss Markets for Products and Services; BCC Research: Wellesley, MA, USA, 2011c. BCC Research (Business Communications Company, Inc). Report code: FOD029D. Global Markets for Ethical Nutrition in Healthcare; BCC Research: Wellesley, MA, USA, 2011d. BCC Research (Business Communications Company, Inc). Report code: BIO082A. Medical Enzymes: Technologies and Global Markets; BCC Research: Wellesley, MA, USA, 2011e.

92
BCC Research (Business Communications Company, Inc). Report code: BIO030G. Global Markets for Enzymes in Industrial Applications; BCC Research: Wellesley, MA, USA, 2012a. BCC Research (Business Communications Company, Inc). Report code: FOD046A. Global Markets for Baby Foods; BCC Research: Wellesley, MA, USA, 2012b. BEDNARSKI, W.; ADAMCZAK, M. Modified lipids and fat mimetics, In: Chemical and Functional Properties of Food Lipids (Z.E. Sikorski, A. Kolakowska). CRC Press, Boca Raton, p. 309–324, 2003. BELARBI, E. H.; MOLINA, E.; CHISTI, Y. A process for high yield and scaleable recovery of high purity eicosapentaenoic acid esters from microalgae and fish oil. Enzyme and Microbial Technology, v. 26, p 516-529, 2000. BEKTAS¸ I.; YUCEL, S.; USTUN, G.; AKSOY, H. A. Production of reduced calorie structured lipid by acidolysis of tripalmitin with capric acid: optimisation by response surface methodology. Journal of the Science of Food and Agriculture, v. 88, p. 1927-1931, 2008. BERGER, A.; GERSHWIN, M. E.; GERMAN, J. B. Effects of various dietary fats on cardiolipin acyl composition during ontogeny of mice. Lipids, v. 27, p. 605-612, 1992. BERNAL, J.; MENDIOLA, J. A.; IBÁÑEZ, E.; CIFUENTES, A. Advanced analysis of nutraceuticals. Journal of Pharmaceutical and Biomedical Analysis, v. 55, p. 758-774, 2011. BERRY, S. E.; WOODWARD, R.; YEOH, C.; MILLER, G. J.; SANDERS, T. A. Effect of interesterification of palmitic acid-rich triacylglycerol on postprandial lipid and factor VII response. Lipids, v. 42, p. 315-323, 2007. BEZARD, J. A.; SEMPORE, B. G. In J.-L. Sebedio & E. G. Perkins (Eds.), New trends in lipid and lipoprotein analyses. Illinois: AOAC Press, p. 106, 1995. BLOCH, A.; THOMSON, C. A. Position of the American dietetic association: phytochemicals and functional foods. Journal of the American Dietetic Association, v. 95, p. 493-96, 1995. BORNSCHEUER, U. T.; ADMACZAK, M.; SOUMANOU, M. M. in: Gunstone, F. D. (Ed.), Lipids as Constituents of Functional Foods, P.J. Barnes & Associates, Bridgwater, p. 149-182, 2002. BORNSCHEUER, U. T.; ADAMCZAK, M.; SOUMANOU, M. M. Lipase-catalysed synthesis of modified lipids, In: Lipids for Functional Foods and Nutraceuticals. (F.D. Gunstone). The Oil Press, Bridgwater, p. 149–182, 2003. BOSKOU, D. Olive Oil Composition. In Olive Oil Chemistry and Technology; Boskou, D., Ed.; AOC Press: Champaign, IL, USA, p. 52-83, 1996. BOURRE, J. M. Roles of unsaturated fatty acids (especially omega-3 fatty Acids) in the brain at various ages and during ageing. The Journal of Nutrition, Health & Aging, v. 8 (3), p. 163-174, 2004.

93
BRACCO, U. Effect of triglyceride structure on fat absorption. The American Journal of Clinical Nutrition, v. 60 (6), p. 1002-1009, 1994. BRADY, L.; BRZOZOWSKI, A. M.; DEREWENDA, Z. S.; DODSON, E.; DODSON, G.; TOLLEY, S.; TURKENBURG, J. P.; CHRISTIANSEN, L.; HUGE-JENSEN, B.; NORSKOV, L. . A serine protease triad forms the catalytic center of a triacylglycerol lipase. Nature, v. 343, p. 767-770, 1990. BRIGELIUS-FLOHÉ, R.; TRABER, M. G. Vitamin E: Function and metabolism. Federation of American Societies for Experimental Biology Journal, v. 13, p. 1145-1155, 1999. BROCKMAN, H. L. General features of lipolysis. In Lipases. B. Borgstrom and H. L. Brockman, editors. Elsevier Science Publishers, Amsterdam, p 1-46, 1984. BURDGE, G. C.; FINNEGAN, Y. E.; MINIHANE, A. M.; WILLIAMS, C. M.; WOOTTON, S. A. Effect of altered dietary n-3 fatty acid intake upon plasma lipid fatty acid composition, conversion of α-linolenic acid to longer-chain fatty acids and partitioning towards beta-oxidation in older men. British Journal of Nutrition, v. 90, p. 311–321, 2003. CAMMAROTA, M. C.; FREIRE, D. M. G. A review on hydrolytic enzymes in the treatment of wastewater with high oil and grease content. Bioresource Technology, v.97 (17), p. 2195-2210, 2006. CARLIER, H.; BERNARD, A.; CASELLI, C. Digestion and absorption of polyunsaturated fatty acids. Reproduction Nutrition Development, v. 31, p. 475-500, 1991. CATONI, E.; CERNIA, E.; PALOCCI, C. Different aspects of “solvent engineering” in lipase biocatalysed esterification. Journal of Molecular Catalysis B: Enzymatic, v. 105, p. 79-86, 1996. CAVALCANTI, E. D. C.; MACIEL, F. M.; VILLENEUVE, P.; LAGO, R. C. A.; MACHADO, O. L. T.; FREIRE, D. M. G. Acetone power from dormant seeds of Ricinus communis L. Applied Biochemistry and Biotechnology, v. 137, p. 57-65, 2007. CHAKRABORTY, K.; RAJ, R. P. Selective enrichment of n3 polyunsaturated fatty acids with C18–C20 acyl chain length from sardine oil using Pseudomonas fluorescens MTCC 2421 lipase. Food Chemistry, v. 114 p. 142-150, 2009. CHANDLER, I. C.; QUINLAN, P. T.; MCNEILL, G. P. Lipase-Catalyzed Synthesis of Chiral Triglycerides, Journal of the American Oil Chemists' Society, v. 75, p. 1513-1518, 1998. CHE MAN, Y. B.; MARINA, A. M. Medium chain triacylglycerol. In F. Shahidi (Ed.), Nutraceutical and specialty lipids and their coproducts, Boca Raton: Taylor & Francis Group, p. 27-56, 2006. CHEN, B.; McCLEMENTS, D. J.; DECKER, E. A. Design of Foods with Bioactive Lipids for Improved Health. Annual Review of Food Science Technology, v. 13 (4), p. 35-56, 2013. CHRISTENSEN, M. W.; ANDERSEN, L.; KIRK, O.; HOLM, H. C. Enzymatic Interesterification of Commodity Oils and Fats: Approaching the Tonne Scale, Lipid Technology, v. 7, p. 33-37, 2001.

94
CHISTENSEN, M. S.; HOY, C. E.; BECKER, C. C.; REDGRAVE, T. G. Intestinal absorption and lymphatic transport of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), and decanoic acids: dependence on intramolecular triacylglycerol structure. The American Journal of Clinical Nutrition, v. 61, p, 56-61, 1995. CHRISTIE, W. W. Silver ion chromatography using solid-phase extraction columns packed with a bonded-sulfonic acid phase. Journal of Lipid Research, v. 30, p. 1471-1473, 1989. CHRISTIE, W. W. Lipid Analysis. Pergamon Press, New York, p. 1, 1982. CHOCT, M. Enzymes for the feed industry: Past, present and future. World's Poultry Science Journal, v. 62, p. 5-15, 2006. CLANDININ, M. T.; CHAPPELL, J. E.; LEONG, S.; HEIM, T.; SWYER, P. R.; CHANCE, G. W. Extrauterine fatty acid accretion in infant brain: implications for fatty acid requirements. Early Human Development, v. 4, p. 131-138, 1980. COLMAN, M. H.; MACRAE, A. R. UK Patent No. 1577933, 1980. CONNOR, W. E. Importance of n-3 fatty acids in health and disease. American Journal of Clinical Nutrition, v. 71(1), p. 171-175, 2000. CRAIG-SCHMIDT, M.; WHITE, M. T.; TEER, P.; JOHNSON, J.; LANE, H. W. Menhaden, coconut and corn oils and mammary tumor incidence in BALB/c virgin female mice treated with DMBA. Nutrition and Cancer, v. 20, p. 99-106, 1993. D’AGOSTINI, D.; FERRAZ, R. C.; GIOIELLI, L. A.; SOTERO V. E. Lípidos estructurados obtenidos por interesterificación de las mezclas binarias y ternarias de las grasas de palma, semilla de palma y triglicéridos de cadena media. Grasas y Aceites, v. 52 (3), p. 214-221 (2001). DANDIK, L.; ARIOGLU, G.; AKSOY, H. A. The Hydrolysis of Used Frying Oil by Native Lipase, Applied Biochemistry and Biotechnology, v. 42, p. 119-126, 1993. DAS, U. N., “A defect in the activity of ∆6 and ∆5 desaturases may be a factor predisposing to the development of insulin resistance syndrome,” Prostaglandins Leukotrienes and Essential Fatty Acids, Lipids in Health and Disease, v. 72 (5), p. 343-350, 2005. DECKER, E. A. The role of stereospecific saturated fatty acid position on lipid nutrition. Nutrition Reviews, v. 54, p. 108-110, 1996. DEREWENDA, Z. S. Structure and Function of Lipases, Advances in Protein Chemistry, v. 45, p. 1-52, 1994. DUQUE, A. G. The role of lipids in fetal brain development. Perinat. Nutr. Rep. Spring, p. 4-5, 1997. Dureja D, Kaushik D, Kumar V. Developments in nutraceuticals. Indian J Pharmacol 2003; 35: 363-72.

95
Dyerberg J. 1989. Coronary heart disease in Greenland Inuit: a paradox. Implications for Western diet patterns. Arct. Med. Res. 48:47–54. Dziezak, J.D. (1989). Fats, oils and fats substitutes. Food Technol. 43: 663–688. Eastmond, P.J., 2004. Cloning and characterization of the acid lipase from castorbeans. J. Biol. Chem. 279, 45540–45545. Edem DO. 2002. Palm oil: Biochemical, physiological, nutritional, hematological and toxicological aspects: A review. Plant Foods for Human Nutrition (Formerly Qualitas Plantarum) 57, 319-341. Asha Embrandiri, Mahamad H. Ibrahim, Rajeev P. Singh. Palm Oil Mill Wastes Utilization; Sustainability in the Malaysian Context. International Journal of Scientific and Research Publications, Volume 3, Issue 3, 1-7. 2013. Emken EA, Adlof RO, Gulley RM. 1994. Dietary linoleic acid influences desaturation and acylation of deuterium-labeled linoleic and linolenic acids in young adult males. Biochim. Biophys. Acta 1213: 277-288. Eric A. Decker, Casimir C. Akoh, and Richard S. Wilkes. Incorporation of (n-3) Fatty Acids in Foods:Challenges and Opportunities. The Journal of Nutrition. Supplement: Heart Healthy Omega-3s for Food—Stearidonic Acid (SDA) as a Sustainable Choice. 1-4 (2012). European Union. Commision Regulation (EU) No 432/2012 of 16 May 2012 establishing a list of permitted health claims made on foods, other than those referring to the reduction of disease risk and to children’s development and health. Off. J. Eur. Union 2012, L136, 1–40. A. Ramírez Fajardo, Casimir C. Akoh and O.M. Lai. Lipase-Catalyzed Incorporation of n-3 PUFA into Palm Oil. JAOCS 80, 1197-1200 (2003). FAO/WHO. Interim Summary of Conclusions and Dietary Recommendations on Total Fat & Fatty Acids—From the Joint FAO/WHO Expert Consultation on Fats and Fatty Acids in Human Nutrition, WHO HQ, Geneva, Switzerland, 2008. Feed Enzymes: The Global Scenario; Frost & Sullivan: London, UK, 2007. Fickers, P. J., Destain, J., Thonart, P., Lipases are atypical hydrolases: Principal characteristics and applications. Biotechnol. Agron. Soc. Environ. 2008, 12, 119–130. Fischer MA, Black HS. Modification of membrane composition, eicosanoid metabolism, and immunoresponsiveness by dietary omega-3 and omega-6 fatty acid sources, modulators of ultraviolet-carcinogenesis. Photochem Photobiol. 1991;54:381-387. Fischer M, Pleiss J. The lipase engineering database: a navigation and analysis tool for protein families. Nucleic Acids Res. 2003;31:319–321. Filer LJ Jr, Mattson FH, Fomon SJ (1969) Triglyceride configuration and fat absorption by the human infant. J Nutr 99(3) 293-298.

96
Fitch-Haumann B., Structured lipids allow fat. Inform, 1997, 8, 1004–1011. P. Flachs, M. Rossmeisl, M. Bryhn, and J. Kopecky, “Cellular and molecular effects of n-3 polyunsaturated fatty acids on adipose tissue biology and metabolism,” Clinical Science, vol. 116, no. 1, pp. 1–16, 2009. Flores, M.V., J.J.W. Sewalt, A.E.M. Janssen, and A. van der Padt, The Nature of Fatty Acid Modifies the Equilibrium Position in the Esterification Catalyzed by Lipase, Biotechnol. Bioeng. 67:364–371 (2000). Filip, S., Fink, R., Hribar, J., & Vidrih, R. (2010). Trans fatty acids in food and their influence on human health. Food Technology and Biotechnology, 48, 135-142. Lydia B. Fomuso and Casimir C. Akoh. Enzymatic Modification of Triolein: Incorporation of Caproic and Butyric Acids to Produce Reduced-Calorie Structured Lipids. JAOCS 74, 269–272 (1997). R. Foster, C.S. Williamson and J. Lunn. Culinary oils and their health effects. British Nutrition Foundation Nutrition Bulletin, 34, 4–47. 2009. Freire, D. M. G. e Castilho, L. C. (2008). “Lipases em Biocatálise” em: Enzimas em Biotecnologia: Produção, aplicação e mercado, 1ª edição Rio de Janeiro, RJ, Editora interciência., Cap. 16 pp: 369-385. Edwin N. Frankel. Nutritional and Biological Properties of Extra Virgin Olive Oil. J. Agric. Food Chem. 2011, 59, 785–792. X. Fu e K.L. Parkin. Selectivity of fatty acid incorporation into acylglycerols in esterification reactions using Rhizomucor miehei and Burkholderia cepacia lipases. Food Research International 37 (2004) 651–657. Galtier, O.; LeDreau, Y.; Ollivier, D.; Krister, J.; Artaud, J & Dupuy, N. 2008. Lipid Compositions and French Registered Designations of Origins of Virgin Olive Oils Predicted by Chemometric Analysis of Mid Infrared Spectra. Applied Spectroscopy. Vol. 6, No.5 Hoy, C.E. & Xu. X. (2001). Structured Triacylglycerols , In:Structured and Modified Llipids, Gunstone, F. (Ed), pp.209-239, Marcel Dekker, Inc.ISBN:0-8247-0253-0, New York. GÁMEZ-MEZA, N.; NORIEGA-RODRÍGUEZ, J. A.; MEDINA-JUÁREZ, L. A.; ORTEGA-GARCÍA, J.; MONROY-RIVERA, J.; TORO-VÁZQUEZ, F. J.; GARCÍA, H. S. e ANGULO-GUERRERO, O. Concentration of eicosapentaenoic acid and docosahexaenoic acid from fish oil by hydrolysis and urea complexation. Food Research International. 36, 721–727. (2003). N.N. Gandhi, K.D. Mukherjee, Reactivity of medium-chain substrates in the interesterification of tripalmitin catalyzed by papaya lipase, J. Am. Oil Chem. Soc. 78 (2001) 965–968. Garg ML, Wierzbicki AA, Thomson ABR, Clandinin MT. 1988. Dietary cholesterol and/or n-3 fatty acid modulate ∆9-desaturase activity in rat liver microsomes. Biochim. Biophys. Acta 962: 330-336. German JB, Dillard CJ. Phytochemicals: nutraceuticals and human health. J Sci Food Agric 2000; 80: 1744-56.

97
German, G. B., & Dillard, C. J. (2004). Saturated fats: what dietary intake? American Journal of Clinical Nutrition, 80, 550-559. J GILL and J H PARISH. Minireview: Lipases--Enzymes at an Interface. BIOCHEMICAL EDUCATION 25(1) 2-5. 1997. Ghosh D, Skinner M, Ferguson LR. The role of the Therapeutic Goods Administration and the Medicine and Medical Devices Safety Authority in evaluating complementary and alternative medicines in Australia and New Zealand. Toxicology. 2006;221:88-94. Gill I, Valivety R. Polyunsaturated fatty acids: Part 1. Occurrence, biological activities and applications. Trends Biotechnol 1997;15:401–9. Godfrey T, Reichelt J. Industrial applications. In: Industrial enzymology - applications of enzymes in industry. London: The Nature Press; 1983. pp. 170–465. Julide Gokçe, Nese Sahin Yesilçubuk, and Guldem Ustun. Enzymatic production of low-calorie structured lipid from Echium seed oil and lauric acid: optimisation by response surface methodology. International Journal of Food Science and Technology 2013, 48, 1383–1389. Gopala Krishna A.G., Gaurav Raj, Ajit Singh Bhatnagar, Prasanth Kumar P.K. and Preeti Chandrashekar. Coconut Oil: Chemistry, Production and Its Applications - A Review. Indian Coconut Journal. 15-27. 2010. Goyens PLL, Spilker ME, Zock PL, KatanMB, Mensink RP. 2006. Conversion ofα-linolenic acid in humans is influenced by the absolute amounts of α-linolenic acid and linoleic acid in the diet and not by their ratio. Am. J. Clin. Nutr. 84:44–53. Gunstone F. D. (1998). Movements towards tailor-made fats. Progress in Lipid Research. Vol. 37, Issue 5, November, P. 277-305. F.D. Gunstone, Modifying Lipids for Use in Food, Published by Woodhead Publishing Limited, Cambridge, UK, 2006. Disponível em: www.woodheadpublishing.com Gupta R., Rathi P., Bradoo S., “Lipase mediated up-gradation of dietary fats and oils”, Critical Reviews in Food Science and Nutrition, 2003; Vol. 43: 635-644. M. I. Gurr and A. T. James. Lipid Biochemistry and Introduction. Cornell University Press, Ithaca, NY, 1971, p. 1. Hamam, F.; Shahidi, F. Enzymatic incorporation of capric acid into a single cell oil rich in docosahexaenoic acid and docosapentaenoic acid and oxidative stability of the resultant structured lipid. Food Chem. 2005, 91, 583-591. Han, L., Xu, Z., Huang, J., Meng, Z., Liu, Y. & Wang, X. (2011). Enzymatically catalyzed synthesis of low-calorie structured lipid in a solvent-free system: Optimization by response surface methodology. Journal of Agricultural a nd Food Chemistry, 59(23), 12635–12642.

98
Fayez Hamam and Fereidoon Shahidi. Acidolysis of Tristearin with Selected Long-Chain Fatty Acids. J. Agric. Food Chem. 2007, 55, 1955-1960. M. Hamosh, Lingual and Gastric Lipases: Their Role in Fat Digestion, Boca Raton, FL: CRC Press; 1990, p. 128– 32. Haraldsson, G. G.; Hjaltason, B. Fish oils as sources of important polyunsaturated fatty acids. In Structured and Modified Lipids; Gunstone, F. D., Ed.; Dekker: New York, 2001; pp 313-350. W. S. Harris, “International recommendations for consumption of long-chain omega-3 fatty acids,” Journal of Cardiovascular Medicine, vol. 8, supplement 1, pp. S50–S52, 2007. Fariha Hasan, Aamer Ali Shah, Abdul Hameed. Industrial applications of microbial lipases. Enzyme and Microbial Technology 39 (2006) 235–251. Health and Welfare Canada. 1990. Nutrition Recommendations. The Report of the Scientific Review Committee. Department of Supply and Services, Cat. No. H49-42/1990E, Ottawa, ON. Hernell O, Blackberg L. Digestion of human milk lipids: physiological significance of sn-2 monoacylglycerol hydrolysis by salt-stimulated lipase. Pediatr Res 1982; 16: 882-885. Hibbeln JR. Seafood consumption, the DHA content of mothers’ milk and prevalence rates of postpartum depression: a cross-national, ecological analysis. J Affect Disord. 2002;69:15–29. Houwelingen ACv, Hornstra G. 1994. Trans fatty acids in early human development. World Rev. Nutr. Diet. 75: 175-178. Hu, F. B., Stampfer, M. J., Manson, J. E., Rimm, E., Colditz, G. A., Rosner, B. A. (1997). Dietary fat intake and the risk of coronary heart disease in women. New England Journal of Medicine, 337, 1491e1499. Huang TL. 2010. Omega-3 fatty acids, cognitive decline, and Alzheimer’s disease: a critical review and evaluation of the literature. J. Alzheimer’s Dis. 21:673–90. Kuan-Hsiang Huang and Casimir C. Akoh. Enzymatic Synthesis of Structured Lipids: Transesterification of Triolein and Caprylic Acid Ethyl Ester. JAOCS 73, 245-250 (1996). Hudert CA, Weylandt KH, Lu Y, Wang J, Hong S, Dignass A, Serhan CN, Kang JX. Transgenic mice rich in endogenous omega-3 fatty acids are protected from colitis. Proc Natl Acad Sci U S A. 2006;103:11276-11281. Hunter JE. 2001. Studies on effects of dietary fatty acids as related to their position on triglycerides. Lipids 36:655–68. Horrocks LA, Yeo YK. 1999. Health benefits of docosahexaenoic acid (DHA). Pharmacol Res 40:211–25. Horvath A, Koletzko B, Szajewska H. Effect of supplementation of women in high-risk pregnancies with long-chain polyunsaturated fatty acids on pregnancy outcomes and growth measures at birth: a meta-analysis of randomized controlled trials. Br J Nutr. 2007;98:253–9.

99
Howe PR, Downing JA, Grenyer BF, Grigonis-Deane EM, Bryden WL. Tuna fish meal as a source of DHA for n-3 PUFA enrichment of pork, chicken, and eggs. Lipids. 2002;37:1067-1076. Hoy, C.E. & Xu. X. (2001). Structured Triacylglycerols , In:Structured and Modified Llipids, Gunstone, F. (Ed), pp.209-239, Marcel Dekker, Inc.ISBN:0-8247-0253-0, New York. Huang, A.H.C. (1984). Plant lipases. In: B. Borgstrom, & H.L. Brockman (Eds.), Lipase (pp. 419–422). Amsterdam: Elsevier Publishers. H. Iding, P. Siegert, K. Mesch, M. Pohl. Application of alpha-keto acid decarboxylases in biotransformations. Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology. Volume 1385, Issue 2, 1998, Pages 307–322. Innis, S. M., Quinlan, P. T., & Nelson, C. M. (1998). Structured triacylglycerols in infant nutrition and metabolism. In S. Y-Huang, & A. J. Sinclair (Eds.), Lipids in infant nutrition (pp. 268e281). Champaign, IL: American Oil Chemists’ Society. Institute of Medicine. 2002. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids, National Academies Press, Washington, DC, pp. 7-1— 7-69 (dietary fiber), 8-1— 8-97 (fat and fatty acids). Institute of Medicine, Dietary Reference Intakes for Energy and Macronutrients. Washington, DC: National Academy Press, 2005. Jaqueline Scholz Issa, Jayme Diament, Neusa Forti. Lipemia Pós-Prandial. Influência do Envelhecimento. Arquivos Brasileiros de Cardiologia - Volume 85, Nº 1, Julho 2005. Jäckel, C.; Hilvert, D. Biocatalysts by evolution. Curr. Opin. Biotechnol. 2010, 21, 753–759. Jaeger KE, Eggert T. Lipases for biotechnology. Curr Opin Biotechnol 2002;13(4):390–7. Brenda H. Jennings and Casimir C. Akoh. Enzymatic Modification of Triacylglycerols of High Eicosapentaenoic and Docosahexaenoic Acids Content to Produce Structured Lipids. JAOCS 76, 1133–1137 (1999). Brenda H. Jennings and Casimir C. Akoh. Lipase-catalyzed modification of sesame oil to incorporate capric acid. Journal of Food Lipids 7 (2000) 2 1-30. Brenda H. Jennings, Casimir C. Akoh. Lipase catalyzed modi®cation of ®sh oil to incorporate capric acid. Food Chemistry 72 (2001) 273±278. Jensen C, Buist NR, Wilson T (1986). Absorption of individual fatty acids from long chain or medium chain triglycerides in very small infants. Am J Clin Nutr 43(5) 745-751. R. G. Jensen, G. L. Jensen: Specialty lipids for infant nutrition. I. milks and formulas. J. Pediatr. Gastroenterol. Nutr. 15 (1992) 232–245. Jensen MM, Christensen MS, Hoy CE. Intestinal absorption of octadecanoic, decanoic and linoleic acid: effect of triglyceride structure. Ann Nutr Metab 1994; 38:104-16.

100
Jensen, R. G. (1996). The lipids in human milk. Progress in Lipid Research, 35(1), 53–92. Jensen, R. G. Infant formulas. In Structured and Modified Lipids; Gunstone, F. D., Ed.; Dekker: New York, 2001; pp 455-464. Kamariah Long. Unlocking the Miracle of Lipases. Malaysian Agricultural Research and Development Institute (MARDI). Kuala Lumpur, Malaysia. 1-46. 2010. Karabulut, I.; Durmaz, G.; Hayaloglu, A. A. C18 unsaturated fatty acid selectivity of lipases during the acidolysis reaction between tripalmitin and oleic, linoleic, and linolenic acids. J. Am. Oil Chem. Soc. 2010, 87, 1301−1307. Ihsan Karabulut, Gokhan Durmaz and Ali Adnan Hayaloglu. Fatty Acid Selectivity of Lipases during Acidolysis Reaction between Oleic Acid and Monoacid Triacylglycerols. J. Agric. Food Chem. 2009, 57, 10466–10470. M. Kates. Techniques of Lipidology: Isolation, Analysis and Identification of Lipids. Elsevier, New York, 1986, p. 1. Kennedy, K.; Fewtrell, M. S.; Morley, R.; Abbott, R.; Quinlan, P. T.; Wells, J. C. K.; Bindels, J. G.; Lucas, A. Double-blind, randomized trial of a synthetic triacylgylcerol in formula-fed term infants: effects on stool biochemistry, stool characteristics, and bone mineralization. Am. J. Clin. Nutr. 1999, 70, 920-927. King, D. M., Padley, F. B., European patent No 0 209 327 (Unilever plc), 1990. Kirchner G, Scollar MP, Klibanov AM. Resolution of racemic mixtures via lipase catalysis in organic solvents. Biosci Biotechnol Biochem 1985;107:7072–251. Klibanov, A.M. Improving enzymes by using them in organic solvents. Nature 2001, 409, 241–246. Kojima, Y., & Shimizu, S. (2003). Purification and characterization of the lipase from Pseudomonas fluorescens HU380. Journal of Bioscience and Bioengineering, 96, 219–226. Kumar A., Kanwar SS., “Synthesis of isopropyl ferulate using silica-immobilized lipase in an organic medium. Enzyme Research, 2011; Vol. 2011: 1- 8. Kumar, A.; Singh, S. Directed evolution: Tailoring biocatalysis for industrial application. Crit. Rev. Biotechnol. 2013, 33, 365–378. Langone, M. A. P. e Sant’anna, G. L. Jr. (1999). Enzymatic synthesis of medium-chain triglycerides in a solvent-free system. Applied Biochemistry and Biotechnology. 77-79, p. 759-770. Lee, D. H., Kim J. M., Shin, H. Y., Kang, S. W. e Kim, S. W. (2006). Biodiesel production using a mixture of immobilized Rhizopus oryzae and Candida rugosa lipases. Biotechnology and Bioprocess Engineering. 11: 522-525.

101
Lee, K. T. e Akoh, C. C. (1998). Characterization of Enzymatically Synthesized Structured Lipids Containing Eicosapentaenoic, Docosahexaenoic and Caprylic acids. Journal of American Oil Chemist’s Society. 75:495-499. Ki-Teak Lee and Casimir C. Akoh. Solvent-Free Enzymatic Synthesis of Structured Lipids from Peanut Oil and Caprylic Acid in a Stirred Tank Batch Reactor. JAOCS 75, 1533–1537 (1998). Leikin AI, Brenner RR. 1987. Cholesterol-induced microsomal changes modulate desaturase activities. Biochim. Biophys. Acta 922: 294-303. Leonard AE, Pereira SL, Sprecher H, Huang YS. Elongation of long-chain fatty acids. Prog Lipid Res. 2004; 43(1):36-54. Lefevre M, Kris-Etherton PM, Zhao G, RUSSELL P. TRACY (2004) Dietary fatty acids, hemostasis, and cardiovascular disease risk. J Am Diet Assoc 104, 410–419. LEPAGE, G. e ROY, C. C. “Direct transesterification of all classes of lipids in a one-step reaction”. Journal of Lipid Research. v. 27, pp. 114-120. (1986). Li D, Mann NJ, Sinclair AJ. 1999. Comparison of n-3 polyunsaturated fatty acids from vegetable oils, meat, and fish in raising platelet eicosapentaenoic acid levels in humans. Lipids 34: S309. Liese A, Seelbach K, Wandrey C, editors. Industrial biotransformationsWeinheim: Wiley-VCH, 2000. LIU, L. e LAMPERT, D. Monitoring chemical interesterification. Journal of American Oil Chemist’s Society. 76, 783–787. (1999). Miroslav Lísa, Michal Holcapek. Triacylglycerols profiling in plant oils important in food industry, dietetics and cosmetics using high-performance liquid chromatography–atmospheric pressure chemical ionization mass spectrometry. Journal of Chromatography A, 1198–1199 (2008) 115–130. Lnnis S M, Dyer R, Nelson C M. Evidence that palmitic acid is absorbed as Sn-2 monoacylycerol from human milk by breast-fed infant. Lipids, 1994, 29: 541 Kamariah Long. Unlocking the Miracle of Lipases. Malaysian Agricultural Research and Development Institute (MARDI). Kuala Lumpur, Malaysia. 1-46. 2010. López-López, A., Castellote-Bargalló, A. I., Campoy-Folgoso, C., Rivero-Urgel, M., Tormo-Carnicé, R., Infante-Pina, D., et al. (2001). The influence of dietary palmitic acid triacylglyceride position on the fatty acid, calcium and magnesium contents of at term newborn faeces. Early Human Development, 65, 83-94 Lucas A, Quinlan P, Abrams S, Ryan S, Meah S, Lucas PJ (1997) Randomised controlled trial of a synthetic triglyceride milk formula for preterm infants Arch Dis Child Fetal Neonatal Ed 77(3) 178-184. Macedo GA, Lozano MMS, Pastore GM. Enzymatic synthesis of short chain citronellyl esters by a new lipase from Rhizopus sp. Electronic Journal of Biotechnology ISSN: 0717-3458 2003 Vol.6 No.1, 72-75.

102
Maduco C O, Akoh C C, Park Y W. Enzymatic interesterification of tripalmitin with vegetable oil blends for formulation of caprine milk infant formula analogs. J Dairy Science, 2007, 90: 594-601. Makrides M, Duley L, Olsen SF.Marine oil, and other prostaglandin precursor, supplementationfor pregnancy uncomplicated by pre-eclampsia or intrauterine growth restriction. Cochrane Database of Systematic Reviews 2006, Issue 3. Art. No.: CD003402. DOI: 10.1002/14651858.CD003402.pub2. MANOEL, E. A. ; Pais, Karla C. ; Cunha, Aline G. ; Coelho, Maria Alice Z. ; Freire, Denise M.G. ; Simas, Alessandro B.C. . On the kinetic resolution of sterically hindered myo-inositol derivatives in organic media by lipases. Tetrahedron: Asymmetry (Print), v. 23, p. 47-52, 2012 Marangoni AG., Rousseau D., “Engineering Triacylglycerols: The Role of Interesterification”, Trends in Food Science Technoogy, 1995; Vol. 6: 329-335. Marangoni, A.G. and D. Rousseau. 1999. Chemical and enzymatic modification of butterfat and butterfat-canola oil blends. J. Food Res. Int. 31:595-599. A.M. Marina, Y.B. Che Man, and I. Amin. Virgin coconut oil: emerging functional food oil. Trends in Food Science & Technology 20 (2009) 481-487. Market Research. 2010. Omega 3 foods and beverages in theU.S. Mark. Res. Rep.,Market Research, Rockville, MD. 3rd ed. Martine S A, Petra A M, Jacques G B. Current trends in the composition of infant milk formulas. Current Paediatrics, 2004, 14: 51-63. Martinez M. Tissue levels of polyunsaturated fatty acids during early human development. J Pediatr. 1992;120: S129. Martinez M. Polyunsaturated fatty acids in the developing human brain, red cells and plasma: influence of nutrition and peroxisomal disease. J World Rev Nutr Diet. 1994; 75:70. Mauvernay RY, Laboreur P, Labrousse M. Composition and its products, United States Patent 3,513,073 (1970). McCann JC, Ames BN. 2005. Is docosahexaenoic acid, an n-3 long-chain polyunsaturated fatty acid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. Am. J. Clin. Nutr. 82:281–95. Mendez, C.; Salas, J.A. Altering the glycosylation pattern of bioactive compounds. Trends Biotechnol. 2001, 19, 449–456. Meiri-Bendek, I., Ben Dror, G., Laouz, H., Yaakobi, D. et al., Human breast milk lipid mimetic as dietary supplement, European Patent EP 1 681 945 B1, 2011. Miles, E. A., & Calder, P. C. (2012). Influence of marine n-3 polyunsaturated fatty acids on immune function and a systematic review of their effects on clinical outcomes in rheumatoid arthritis. British Journal of Nutrition, 107, 171-184.

103
Olivier Molendi-Coste, Vanessa Legry, and Isabelle A. Leclercq. Why and HowMeet n-3 PUFA Dietary Recommendations?. Gastroenterology Research and Practice. Volume 2011, Article ID 364040, 11 pages. Kiro Mojsov. APPLICATION OF ENZYMES IN THE TEXTILE INDUSTRY : A REVIEW. II International Congress “Engineering, Ecology and Materials in the Processing Industry” 230-239, 2011. Juliana Maria Leite Nóbrega de Moura*, Lireny Aparecida Guaraldo Gonçalves, Renato Grimaldi, Marinalda da Silva Soares e Ana Paula Badan Ribeiro. Otimização das condições de produção de ésteres etílicos a partir de óleo de peixe com elevado teor de ácidos graxos ω-3. Quim. Nova, Vol. 29, No. 5, 956-959, 2006. A.M. Marina, Y.B. Che Man, and I. Amin. Virgin coconut oil: emerging functional food oil. Trends in Food Science & Technology 20 (2009) 481-487. Mozaffarian, D., Pischon, T., Hankinson, S. E., Rifai, N., Joshipura, K., Willett, W. C. (2004). Dietary intake of trans fatty acids and systemic inflammation in women. American Journal of Clinical Nutrition, 79, 606e612. Mozaffarian, D., & Clarke, R. (2009). Quantitative effects on cardiovascular risk factors and coronary heart disease risk of replacing partially hydrogenated vegetable oils with other fats and oils. European Journal of Clinical Nutrition, 63, S22eS33. Mu, H., Xu, X. e Hoy C. E. (1998). Production of specific structured triacylglycerols by lipasecatalyzed interesterification in a laboratory scale continuous reactor. Journal of American Oil Chemist’s Society. 75, 1187-1193. Mu H, Høy CE. 2004. The digestion of dietary triacylglycerols. Prog. Lipid Res. 43:105–33 Nesaretnam K, Yew WW, Wahid MB. 2007. Tocotrienols and cancer: beyond antioxidant activity. Eur. J. Lipid Sci. Technol. 109:445–52. Mukherjee, K.D., Plant Lipases in Lipid Biotransformations, in Engineering of/with Lipases, edited by F.X. Malcata, Kluver Academic, Dordrecht, 1996, pp. 391–401. Mukherjee, K. D.; Kiewitt, I. Structured triacylglycerols resembling human milk fat by transesterification catalyzed by papaya (Carcia papaya) latex. Biotechnol. Lett. 1998, 20, 613-616. S. Nagachinta, C.C. Akoh. Enrichment of palm olein with long chain polyunsaturated fatty acids by enzymatic acidolysis . LWT - Food Science and Technology 46 (2012) 29-35. Supakana Nagachinta and Casimir C. Akoh. Synthesis of Structured Lipid Enriched with Omega Fatty Acids and sn-2 Palmitic Acid by Enzymatic Esterification and Its Incorporation in Powdered Infant Formula. J. Agric. Food Chem. 2013, 61, 4455-4463. Nagata, N. Takatsuka, and H. Shimizu, “Soy and fish oil intake and mortality in a Japanese community,” American Journal of Epidemiology, vol. 156, no. 9, pp. 824–831, 2002. J. A. Napier, O. Sayanova, B.Qi, and C.M. Lazarus, “Progress toward the production of long-chain polyunsaturated fatty acids in transgenic plants,” Lipids, vol. 39, no. 11, pp. 1067-1075, 2004.

104
Nese S, Casimir C A, Artemis K. Enzymatic production of human milk fat substitutes containing γ-linolenic acid: optimization of reactions by response surface methodology. J Am Oil Chem Soc, 2005, 82: 549-557. Nielsen, N. S.; Yang, T.; Xu, X.; Jacobsen, C. Production and oxidative stability of a human milk fat substitute produced from lard by enzyme technology in a pilot packed-bed reactor. Food Chem. 2006, 94, 53-60. P. A. Nunes, P. Pires-Cabral, M. Guillén, F. Valero, D. Luna, S. Ferreira-Dias. Production of MLM-Type Structured Lipids Catalyzed by Immobilized Heterologous Rhizopus oryzae Lipase. J Am Oil Chem Soc (2011) 88:473-480. OH, J. E.; Lee, K.; Park, W.; H.K., Kim, J.Y.; Kwang, I.K.; Kim, J.W., Kim, H.R. & Kim, I.H. (2009). Lipase Catalyzed Acidolysis of Olive oil with Capric Acid: Effect of Water Activity on incorporation and Acyl Migration. J. Agric Food Chem., Vol 57, No.19, pp9280-9283 Okahata Y., Fujimoto Y., Ijiro K., “A lipid coated lipase as enantioselective ester synthesis catalyst in homogeneous organic solvents”, Journal of Organic Chemistry, 1995; Vol. 60: 2244-2250. Okanishi, M.; Suzuki, N.; Furita, T. Variety of hybrid characters among recombinants obtained by interspecific protoplast fusion in streptomycetes. Biosci. Biotechnol. Biochem. 1996, 6, 1233–1238. Ollis DL, Cheah E, Cygler M, Dijkstra B, Frolow F, Franken SM, Harel M, Remington SJ, Silman I, Schrag J, Sussman JL, Verschueren KHG, Goldman A. The alpha/beta hydrolase fold. Protein Eng. 1992;5:197–211. Omar, S.H. Oleuropein in olive and its pharmacological effects. Sci. Pharm. 2010, 78, 133–154. H.T. Osborn, C.C Akoh, Comp. Rev. Food Sci. Food Saf. 1 (2002) 93–103. H.T. Osborn and C.C. Akoh. Structured Lipids–Novel Fats with Medical, Nutraceutical, and Food Applications. Comprehensive reviews in food science and food safety, Vol. 3, 110-120, 2002b. Packard VS (1982) Infant formula, composition, formulation and processing. In: Stewart GF, Schweigert SS, Hawthorn J (eds) Food science and technology, a series of monographs. Academic Press, New York, pp 141–175. Panke, S.; Wubbolts, M. Advances in biocatalytic synthesis of pharmaceutical intermediates. Curr. Opin. Chem. Biol. 2005, 9, 188–194. K.J. Patil, M.Z. Chopda, R.T. Mahajan, Lipase biodiversity, Indian J. Sci. Technol. 4 (2011) 971–982. Pawlosky RJ, Hibbeln JR, Novotny JA, Salem N Jr. Physiological compartmental analysis of α-linolenic acid metabolism in adult humans. J Lipid Res 2001;42:1257–65. Laura J. Pham and Patrisha J. Pham. (2012). Biocatalyzed Production of Structured Olive Oil Triacylglycerols. Olive Oil – Constituents, Quality, Health Properties 448 and Bioconversions, Dr. Dimitrios Boskou (Ed.), pp. 447-456, ISBN: 978-953-307-921-9, InTech, Available from:

105
http://www.intechopen.com/books/olive-oil-constituents-quality-healthproperties-and-bioconversions/biocatalyzed-production-of-structured-olive-oil-triacylglycerols Pinyaphong, P. & Phutrakul, S. (2009). Modification of palm oil structure to cocoa butter equivalent by carica papaya lipase- catalyzed interesterification. World Academy of Science, Engineering and Technology, 54, Sanders, T. H., & Pattee, H. E. (1975). Peanut alkaline lipase. Lipids, 10(1), 50–54. PORIM (Palm Oil Research Institute of Malaysia). Minilibro de los usos del aceite de palma, Ed., Percetekan, Malindo, Malasia. (1988). Xiao-Li Qin, Bo Yang, Hui-Hua Huang, and Yong-Hua Wang. Lipase-Catalyzed Incorporation of Different Fatty Acids into Tripalmitin Enriched Triacylglycerols: Effect of Reaction Parameters. J. Agric. Food Chem. 2012, 60, 2377-2384. P. Quinlan, S. Moore: Modification of triglycerides by lipases: Process technology and its application to the production of nutritionally improved fats. INFORM 4 (1993) 580–585. S. K. Rakshit, R. Vasuhi, Y. Kosugi. Enrichment of polyunsaturated fatty acid from tuna oil using immobilized Pseudomonas fluorescens lipase. Bioprocess Engineering 23 (2000), 251-255. M. Ramírez, L. Amate, A. Gil. Absorption and distribution of dietary fatty acids from different sources. Early Human Development. 65 Suppl. (2001) 95-101. Rickard SE, Thompson LU. 1997. Phytoestrogens and lignans: Effects on reproduction and chronic disease. In: Antinutrients and Phytochemicals in Foods, ed Shahidi F, Oxford University Press, New York, pp. 273-293. Riediger ND, Othman RA, Suh M, Moghadasian MH. 2009. A systemic review of the roles of n-3 fatty acids in health and disease. J. Am. Diet. Assoc. 109:668–79. Rogers P (2001) A healthy body, a healthy mind: long-term impact of diet on mood and cognitive function. Proceedings of the Nutrition Society 60: 135-143. Rocha ADM, Vilarim MM, Nardi AE. 2010. What is the effectiveness of the use of polyunsaturated fatty acid omega-3 in the treatment of depression? Expert Rev. Neurother. 10:1117–29. Alejandro Rocha-Uribe and Ernesto Hernandez. Solvent-Free Enzymatic Synthesis of Structured Lipids Containing CLA from Coconut Oil and Tricaprylin. JAOCS 81, 685–689 (2004). Rodrigues, R. C.; Fernandez-Lafuente, R. Lipase from Rhizomucor miehei as a biocatalyst in fats and oils modification. J. Mol. Catal. B: Enzym. 2010, 66, 15−32. Sahin, N.; Akoh, C. C.; Karaali, A. Lipase-catalyzed acidolysis of tripalmitin with hazelnut oil fatty acids and stearic acid to produce human milk fat subsititute. J. Agric. Food Chem. 2005, 53, 5779-5783. Sahin, N., Akoh, C. C., Karaali, A., Human milk fat substitutes containing omega-3 fatty acids. J. Agric. Food Chem. 2006, 54, 3717–3722.

106
SANTOS, B. C. P. (2010). Purificação e caracterização de lipases de Penicillium simplicissimum produzidas por Fermentação no Estado Sólido e Submersa, Dissertação de M.Sc., Instituto de Química, UFRJ, Rio de Janeiro, RJ, Brasil. Sarah E. E. Berry. Triacylglycerol structure and interesterification of palmitic and stearic acid-rich fats: an overview and implications for cardiovascular disease. Nutrition Research Reviews (2009), pp. 1-15. Saxena RK, Sheoran A, Giri B, Davidson WS. Purification strategies for microbial lipases. J Microbiol Meth 2003;52:1–18. Schmid RD, Verger R. Lipases: interfacial enzymes with attractive applications. Angew Chem Int Ed. 1998;37:1609–1633. Senanayake, S. P. J. N.; Shahidi, F. Enzyme-assisted acidolysis of borage (Borage officinalis L.) and evening primrose (Oenothera biennis L.) oils: incorporation of omega-3 polyunsaturated fatty acids. J. Agric. Food Chem. 1999, 47, 3105-3112. Senanayake, S. P. J. N., Shahidi, F., in: Shahidi, F. (Ed.), Bailey’s Industrial Oil and Fat Product, John Wiley & Sons, New York, USA 2005, pp. 555–584. Senanayake, N.; Shahidi, F. Concentration of docosahexaenóico acid (DHA) from algal oil via urea complexation. J. Food Lipids 2000, 7, 51-61. Sih CJ, Wu SH. Resolution of enantiomers via biocatalysts. Topics Stereochem 1989;19:63–125. SILVA, J. N. (2014), produção de lipase por fermentação em estado sólido utilizando o fungo Rhizomucor miehei, Dissertação de M.Sc., Escola de Química, UFRJ, Rio de Janeiro, RJ, Brasil. Simopoulos AP. Omega-3 fatty acids in inflammation and autoimmune diseases. J Am Coll Nutr. 2002;21:495-505 Shahidi, F. and Wanasundara, U.N. 1998. Omega-3 fatty acid concentrates: nutritional aspects and production technologies. Trends Food Sci. Tech. 9, 230-240. Shapiro H, Theilla M, Attal-Singer J, Singer P. 2010. Effects of polyunsaturated fatty acid consumption in diabetic nephropathy. Nat. Rev. Nephrol. 7:110–21. R. E. Smith, J. W. Finley, and G. A. Leveille, J. Agric. Food. Chem., 42, 432 (1994). Rakesh Sharma and Ram B. Singh. Bioactive Foods and Nutraceutical Supplementation Criteria in Cardiovascular Protection. The Open Nutraceuticals Journal, 2010, 3, 141-153. Sonali Seth, Debamitra Chakravorty, Vikash Kumar Dubey, Sanjukta Patra. An insight into plant lipase research – challenges encountered. Protein Expression and Purification 95 (2014) 13–21. Sonnet, PE., 1988. Lipase selectivities. J. Am. Oil Chem. Sot., 65: 900-904.

107
T.J. Mangos, K.C. Jones, T.A. Foglia, Lipase-catalyzed synthesis of structured low-calorie triacylglycerols, J. Am. Oil Chem. Soc. 76 (1999) 1127-1132. Reyes-Hernández J, Dibildox-Alvarado E, Charó-Alonso M, Toro-Vazquez J. 2007. Physicochemical and Rheological Properties of Crystallized Blends Containing trans-free and Partially Hydrogenated Soybean Oil. J. Am. Oil Chem. Soc. 84, 1081-1093. Mohamed M. Soumanou, Marlène Pérignon and Pierre Villeneuve. Lipase-catalyzed interesterification reactions for human milk fat substitutes production: A review. Eur. J. Lipid Sci. Technol. 2013, 115, 270–285. Sousa, Joab Sampaio ; Cavalcanti-Oliveira, Elisa d Avila ; Aranda, Donato Alexandre Gomes; Freire, Denise Maria Guimarães . Application of lipase from the physic nut (Jatropha curcas L.) to a new hybrid (enzyme/chemical) hydroesterification process for biodiesel production. Journal of Molecular Catalysis. B, Enzymatic (Print) , v. 65, p. 133-137, 2010. Sakurai, T, Margolin, A L, Russell, A J and Klibanov, A M (1988)Jam Chem Soc 110, 7236-7237. Sarda, L. and Desnuelle, P. (1958) Biochim. Biophys. Acta 30, 513-521. Senanayake, N.S.P.J. and Shahidi, F. 2002. Enzyme-catalyzed synthesis of structured lipids via acidolysis of seal (Phoca groenlandica) blubber oil with capric acid. Food Res. Int. 35, 745–752. Scheideler SE, Froning GW. 1996. The combined influence of dietary flaxseed variety, level, form, and storage conditions on egg production and composition among vitamin E-supplemented hens. Poult. Sci. 75: 1221-1226. Schmid, R. D., Verger, R., Lipases: Interfacial enzymes with attractive applications. Angew. Chem. Int. Ed. Engl. 1998, 37, 1608–1633. Schwab JM, Chiang N, Arita M, Serhan CN. Resolvin E1 and protectin D1 activate inflammation-resolution programmes. Nature. 2007; 447:869-874. Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. Maurizio Servili, Beatrice Sordini, Sonia Esposto, Stefania Urbani, Gianluca Veneziani, Ilona Di Maio, Roberto Selvaggini and Agnese Taticchi. Biological Activities of Phenolic Compounds of Extra Virgin Olive Oil. Antioxidants 2014, 3, 1-23. Simopoulos, A. P. (1991). Omega-3 fatty acids in health and disease and in growth and development. American Journal of Clinical Nutrition, 54(3), 438–463. Simopoulos AP. 1999a. New products from the agri-food industry: The return of n-3 fatty acids into the food supply. Lipids 34: S297-S301. Simopoulos AP. 1999b. Evolutionary aspects of omega-3 fatty acids in the food supply. Prostaglandins Leuko. Essent. Fatty Acids 60: 421-429.

108
Simopoulos AP. 2001. n-3 Fatty acids and human health: Defining strategies for public policy. Lipids 36: S83-S89. A. P. Simopoulos, “Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: nutritional implications for chronic diseases,” Biomedicine and Pharmacotherapy, vol. 60, no. 9, pp. 502–507, 2006. Small, D. M., The effect of triglyceride structure on absorption and metabolism. Ann. Rev. Nutr. 1991, 11, 413–434. SOTERO, D. G.; ÁGUILA, J. S.; RAMÍREZ, R. S.; REÁTEGUI, G. C.; RIOS, J. A. S.; SOLÍS, V. S.; TORRES, R. P.; e FILHO, J. M. Fraccionamiento e interesterificacion del aceite de palma (Elaeis guineensis) cultivado en la amazonia peruana. Grasas y Aceites. 59 (2), Abril-Junio, 104-109. (2008). Sreenivasan, B. 1976. Interesterification of fats. J. Amer. Oil Chem. Soc. 55:796-805. Srivastava A, Akoh C C, Chang S W. Candida rugosa lipase LIP1-catalyzed transesterification to produce human milk fat substitute. J Agric Food Chem, 2006, 54: 5175-5181. Stephen De Felice1989. Disponível em: http://www.nutraceuticalsworld.com/contents/view_health-e-insights/2011-10-28/an-interview-with-dr-stephen-defelice/#sthash.zMRcJ5vc.dpuf Stephanie G Burton, Don A Cowan, and John M Woodley. The search for the ideal biocatalyst. http://biotech.nature.com • JANUARY 2002 • VOLUME 20 • nature biotechnology 37-45. William Stillwell. The role of polyunsaturated lipids in membrane raft function. Scandinavian Journal of Food and Nutrition 2006; 50 (2): 107-113. Stillwell W, Wassall SR. Docosahexaenoic acid: membrane properties of a unique fatty acid. Chem and Phys of Lipids. 2003;126:1. Subramani Sellappan and Casimir C. Akoh. Enzymatic Acidolysis of Tristearin with Lauric and Oleic Acids to Produce Coating Lipids. JAOCS 77, 1127–1133 (2000). TAHIN. Q.S. Importancia fisiológica e patológica dos ácidos graxos Arq. BioI. Tecnol., 28 (3) 335-361.1985. Carla Tecelão, Marina Guillén, Francisco Valero, Suzana Ferreira-Dias. Immobilized heterologous Rhizopus oryzae lipase: A feasible biocatalyst for the production of human milk fat substitutes. Biochemical Engineering Journal 67 (2012) 104– 110. Thomas PR, Earl R. Opportunities in the Nutrition and Food Sciences: Research Challenges and the Next Generation of Investigators. Washington, DC: National Academies Press; 1994:109. Thomson, A. B.; Keelan, M.; Garg, M. L.; Clandinin, M. T. Intestinal aspects of lipid absorption: a review. Can. J. Physiol. Pharmacol. 1989, 67, 179-191.

109
Tomarelli RM, Meyer BJ, Weaber JR, Bernhart FW. Effect of positional distribution on the absorption of the fatty acids of human milk and infant formulas. J Nutr 1968;95:583– 90. Sergio Torres-Giner, Antonio Martinez-Abad, Maria J. Ocio, and Jose M. Lagaron. Stabilization of a Nutraceutical Omega-3 Fatty Acid by Encapsulation in Ultrathin Electrosprayed Zein Prolamine. Vol. 75, Nr. 6, 2010 Journal of Food Science N69-79. Tschocke C. Enzymatic treatment of fats in wastewater treatment plants. Eau Ind, Nuisances 1990;138:63–4. Tufvesson, P.; Lima-Ramos, J.; Nordblad, M.; Woodley, J.M. Guidelines and cost analysis for catalyst production in biocatalytic processes. Org. Process Res. Dev. 2011, 15, 266–274. Turner D, Shah PS, Steinhart AH, Zlotkin S, Griffiths AM. 2011. Maintenance of remission in inflammatory bowel disease using omega-3 fatty acids (fish oil): a systematic review and meta-analyses. Inflamm. Bowel Dis. 17:336–45. Undurraga D, Markovits A, Erazo S. Cocoa butter equivalent through enzymic interesterification of palm oil midfraction. Process Biochem 2001;36:933–9. Van Der Walle N. U¨ ber synthetische Wirkung bakterieller Lipasen. Cbl Bakt Parasitenk Infektionskr 1927;70:369–73. Verger, R., Mieras, M. C. E., and de Haas, G. H. (1973) Action of phospholipase A at interfaces, J. Biol. Chem. 248, 4023-4034. Villeneuve, P., 2003. Plant lipases and their applications in oils and fats modification.Eur. J. Lipid Sci. Technol. 105, 308–317. Vivienne V. Yankah and Casimir C. Akoh. Lipase-Catalyzed Acidolysis of Tristearin with Oleic or Caprylic Acids to Produce Structured Lipids. JAOCS 77, 495–500 (May 2000). Wanasundara, U.N. and Shahidi, F. 1999. Concentration of omega 3-polyunsaturated fatty acids of seal blubber oil by urea complexation: Optimization of reaction conditions. Food Chem. 65, 41-49. Wang, L. L., & Johnson, E. A. (1992). Inibition of Listeria monocytogenes by fatty acids and monoglycerides. Applied and Environmental Microbiology, 58, 624-629. Weber, N., & Mukherjee, K. D. (2005). Lipids in infant formulas and human milk fat substitutes. In C. C. Akoh, & O. Lai (Eds.), Healthful lipids (pp. 607e641). Champaign, IL: American Oil Chemists’ Society. Willis, W.M., and A.G. Marangoni, Enzymatic Interesterification, in Food Lipids: Chemistry, Nutrition, and Biotechnology, edited by C.C. Akoh and D.B. Min, Marcel Dekker, New York, 1998, pp. 665–698. Wiseman A. Introduction to principles. In: Wiseman A, editor. Handbook of enzyme biotechnology. 3rd ed. Padstow, Cornwall, UK: Ellis Horwood Ltd. T.J. Press Ltd.; 1995. p. 3–8. WHO and FAO Joint Consultation. 1995. Fats and oils in human nutrition. Nutr. Rev. 53: 202-205.

110
WO 2003033633 A1. Cholesterol lowering structured lipids with omega 3 PUFA, 2003. World Enzymes. Freedonia Group: Cleveland, OH, USA, 2011. Wu, J. C., Low, H. R., Leng, Y., Chow, Y., Li, R., Talukder, M. e Choi, W. J. (2006). Ketoprofen resolution by enzymatic esterification and hydrolysis of the ester product. Biotechnology and Bioprocess Engineering. 11: 211-214. X. Xu, C.-E. Høy, J. Adler-Nissen: Effects of lipid-borne compounds on the activity and stability of lipases in microaqueous systems for lipase-catalyzed reaction. In: Stability and Stabilization of Biocatalysts. Eds. A. Ballesteros, F. J. Plou, J. L. Iborra, P. Halling, Elsevier Science, Amsterdam 1998, pp. 441–446. Xuebing Xu. Production of specific-structured triacylglycerols by lipase-catalyzed reactions: a review. Eur. J. Lipid Sci. Technol. 2000, 287–303. Yang LY & Kuksis A (1991) Apparent convergence (at 2-monoacylglycerol level) of phosphatidic acid and 2-monoacylglycerol pathways of synthesis of chylomicron triacylglycerols. J Lipid Res 32, 1173–1186. Tiankui Yang, Maj-Britt Fruekilde, and Xuebing Xu. Applications of Immobilized Thermomyces lanuginose Lipase in Interesterification. JAOCS 80, 881–887 (2003). Yankah, V. V. e Akoh, C. C. (2000). Lipase catalyzed acidolysis of tristearin with oleic or caprylic acids to produce structured lipids. Journal of American Oil Chemist’s Society. Champaign, v.77, n.5, p.495-500. B. M. Yashodhara, S. Umakanth, J. M. Pappachan, S. K. Bhat, R. Kamath, and B. H. Choo, “Omega-3 fatty acids: a comprehensive review of their role in health and disease,” Postgraduate Medical Journal, vol. 85, no. 1000, pp. 84–90, 2009. Qing hong You, Xiulian Yin, Yuping Zhao, Yan Zhang. Biodiesel production from jatropha oil catalyzed by immobilized Burkholderia cepacia lipase on modified attapulgite. Bioresource Technology 148 (2013) 202-207. You, P., Su, E., Yang, X., Mao, D., Wei, D., 2011. Carica papaya lipase-catalyzed syn-thesis of terpene esters. J. Mol. Catal. B: Enzym. 71, 152–158. N.M. Zagre et al., Public Health Nutr., 6, pp. 733-42 (2003). Zaks, A and Klibanov, A M (1986)JAm Chem Soc 108, 2767-2768. Zaks, A. and Klibanou, A.M. (1988). The effect of water on enzyme action in organic media. J. Biol. Chem. 263: 8017–8021. Zhao H, Lu Z, Bie X, Lu F and Liu Z, Lipase catalyzed acidolysis of lard with capric acid in organic solvent. J Food Eng 78:41–46 (2007). S.H. Zeisel, Regulation of ‘nutraceuticals’, Science 285 (1999) 1853–1855.

111





























