Embed Size (px)
Citation preview

Isolation, characterization, and in vitro and in vivodifferentiation of putative thecal stem cellsArata Honda*, Michiko Hirose*, Kenshiro Hara†, Shogo Matoba†, Kimiko Inoue*, Himomi Miki*, Hitoshi Hiura‡,Mito Kanatsu-Shinohara§, Yoshiakira Kanai†, Tomohiro Kono‡, Takashi Shinohara§, and Atsuo Ogura*¶
*BioResource Center, RIKEN, Tsukuba, Ibaraki 305-0074, Japan; †Department of Veterinary Anatomy, University of Tokyo, Bunkyo-ku, Tokyo 113-8657,Japan; ‡Department of BioScience, Tokyo University of Agriculture, Setagaya-ku, Tokyo 156-8502, Japan; and §Department of Molecular Genetics, GraduateSchool of Medicine, Kyoto University, Kyoto 606-8501, Japan
Edited by R. Michael Roberts, University of Missouri, Columbia, MO, and approved June 7, 2007 (received for review April 26, 2007)
Although ovarian theca cells play an indispensable role in follicu-logenesis by providing follicular structural integrity and steroidsubstrates for estrogen production, little information is availableabout their recruitment, growth, and differentiation because theirimmature forms have not been identified. We have isolated puta-tive thecal stem cells with the ability to self-renew and differen-tiate in vivo and in vitro. They are similar to fibroblasts in mor-phology and proliferate in vitro as round colonies with ahomogenous cell population. They were induced to differentiateinto early precursors and steroidogenic cells in a stepwise mannerafter treatment with serum, luteinizing hormone, and paracrinefactors from granulosa cells. At each differentiation step, thesecells displayed appropriate gene expression and morphologicalmarkers and later secreted androstenedione. The fully maturemorphology was achieved by coculture with isolated granulosacells. When transplanted into the ovaries, the putative thecal stemcells colonized exclusively in the ovarian interstitium and the thecallayer of follicles as differentiated cells. Thus, thecal stem cellsappear to be present in neonatal ovaries and can be isolated,purified, and induced to differentiate in vitro. Thecal stem cellscould provide an invaluable in vitro experimental system to studyinteractions among the oocytes, granulosa cells, and theca cellsduring normal folliculogenesis and to study ovarian pathologycaused by theca cell dysfunction.
follicle � oocyte � ovary
During folliculogenesis in mammals, oocytes grow whilesurrounded by an increasing number of granulosa cell
layers. From the preantral stage onward, theca cells differentiateas the outer layer of the follicle. These elements providestructural integrity and the androgen substrate for granulosa cellestrogen production, thus playing an indispensable role in fol-licular development (1). Mature theca cells can be distinguishedby their morphology and location in situ and can be isolatedeasily in a crude suspension for in vitro analysis. Much informa-tion has been reported about the factors and mechanismsregulating theca cell steroidogenesis in several mammalian spe-cies (2). However, our knowledge about theca cell recruitmentand growth is limited because of the lack of in vitro experimentalsystems to isolate, culture, and differentiate immature thecacells. Generally, it is difficult to distinguish immature theca cellsfrom ovarian stroma cells based on their morphology (3).
In contrast to theca cells, their male counterparts, Leydig cells,have been studied in detail because their putative stem cells canbe isolated from mouse and rat testes by flow cytometric sorting(4, 5). Leydig cells can remain undifferentiated in vitro formonths and can respond to growth factors to differentiate intosteroidogenic precursor cells (5). After transplantation into thetestes of Leydig cell-deficient mice, these stem cells recolonizenormally in the host testicular interstitium and restore the serumtestosterone concentration (4). These in vitro and in vivo ap-proaches using isolated Leydig stem cells may provide further
clues about the mechanisms of formation and maintenance ofthe Leydig cell population and their biochemical characteristics.
Initially, we undertook this study to isolate putative ‘‘femalegerm-line stem (GS) cells,’’ the presence of which is still highlycontroversial among reproductive biologists and stem cell re-searchers (6–8). We initially expected that the isolation proce-dure optimized to male GS cells from neonatal testes (9, 10)might be applicable to putative female counterparts in neonatalovaries. Under our experimental conditions, we were able togenerate round spherical colonies that produced a number ofsmall primitive oocytes as if oogenesis had proceeded inside.However, detailed biochemical analysis revealed that most of thecell types comprising the colonies were somatic cells and notgerm-line cells. These cells were later identified as putativeovarian thecal stem cells.
A fundamental property of stem and progenitor cell division isthe capacity to retain the proliferative state or to generate differ-entiated daughter cells. These putative thecal stem cells werepurified easily in vitro, formed characteristic anchor-independentround colonies, and, after stimulation, started to differentiate andshow characteristic signs of steroidogenesis. They also colonized thehost ovarian tissue after transplantation in a manner similar to thatof Leydig stem cells in mice and rats (4, 5). Our study providesevidence for the presence of putative thecal stem cells in theneonatal mouse ovary. Interestingly, unlike Leydig stem cells,putative thecal stem cells can be purified in vitro simply by opti-mizing the culture conditions without any cell-sorting procedures.Thus, these isolated putative thecal stem cells may provide aninvaluable and reproducible experimental system to study theirbiochemical characteristics. This system would include gene expres-sion and signaling cascades, which are essential for normal follicu-logenesis in mammals.
ResultsIsolation of Neonatal Ovarian Cells by Selective Culture. Because theoriginal purpose of this study was to establish cell lines of femaleGS cells (6), cells were cultured from newborn mouse ovariesaccording to a protocol for male GS cells with slight modifica-tions (9, 10). The culture medium was essentially a serum-freeGS medium (GSM-K) containing growth factors [see Materials
Author contributions: A.H., M.H., T.K., and A.O. designed research; A.H., M.H., K.H., S.M.,K.I., H.M., H.H., M.K.-S., Y.K., T.K., T.S., and A.O. performed research; M.K.-S. and T.S.contributed new reagents/analytic tools; A.H., M.H., K.H., S.M., K.I., and Y.K. analyzed data;and A.H., Y.K., and A.O. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Abbreviations: GS, germ-line stem; GSM, GS medium; GSM-K, serum-free GSM; GSM-S,serum-containing GSM; ER, endoplasmic reticulum; LH, luteinizing hormone; MVH, mousevasa homologue.
¶To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0703787104/DC1.
© 2007 by The National Academy of Sciences of the USA
www.pnas.org�cgi�doi�10.1073�pnas.0703787104 PNAS � July 24, 2007 � vol. 104 � no. 30 � 12389–12394
DEV
ELO
PMEN
TAL
BIO
LOG
Y
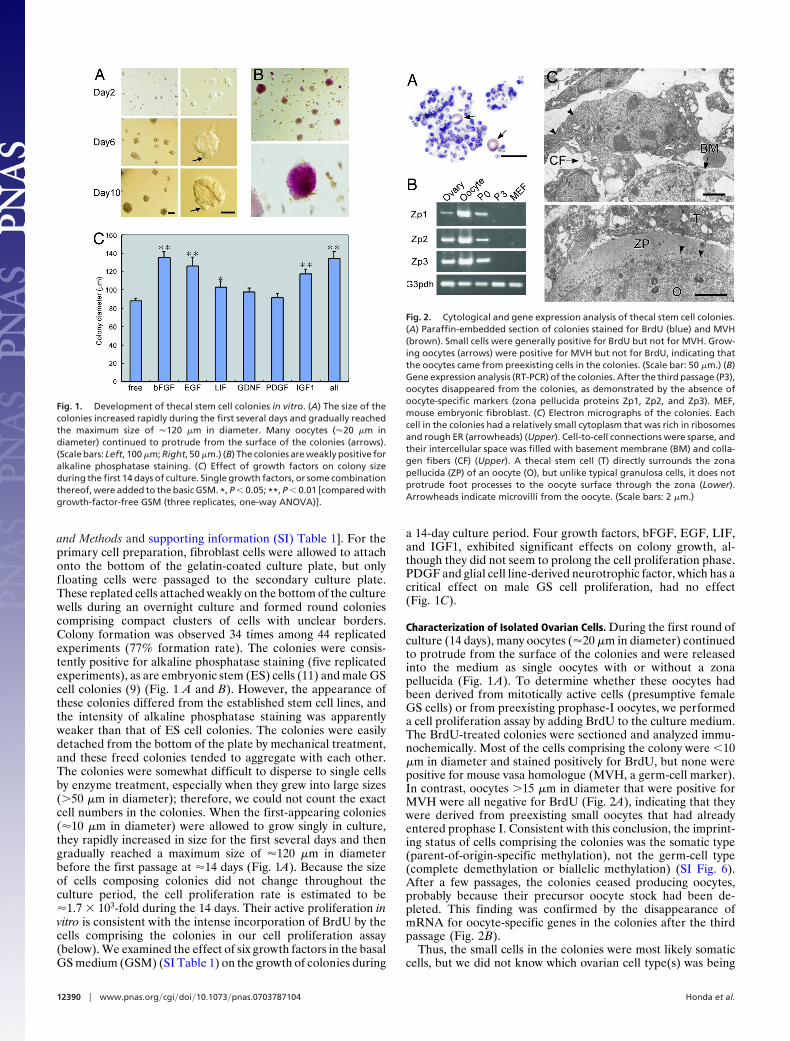
and Methods and supporting information (SI) Table 1]. For theprimary cell preparation, fibroblast cells were allowed to attachonto the bottom of the gelatin-coated culture plate, but onlyfloating cells were passaged to the secondary culture plate.These replated cells attached weakly on the bottom of the culturewells during an overnight culture and formed round coloniescomprising compact clusters of cells with unclear borders.Colony formation was observed 34 times among 44 replicatedexperiments (77% formation rate). The colonies were consis-tently positive for alkaline phosphatase staining (five replicatedexperiments), as are embryonic stem (ES) cells (11) and male GScell colonies (9) (Fig. 1 A and B). However, the appearance ofthese colonies differed from the established stem cell lines, andthe intensity of alkaline phosphatase staining was apparentlyweaker than that of ES cell colonies. The colonies were easilydetached from the bottom of the plate by mechanical treatment,and these freed colonies tended to aggregate with each other.The colonies were somewhat difficult to disperse to single cellsby enzyme treatment, especially when they grew into large sizes(�50 �m in diameter); therefore, we could not count the exactcell numbers in the colonies. When the first-appearing colonies(�10 �m in diameter) were allowed to grow singly in culture,they rapidly increased in size for the first several days and thengradually reached a maximum size of �120 �m in diameterbefore the first passage at �14 days (Fig. 1A). Because the sizeof cells composing colonies did not change throughout theculture period, the cell proliferation rate is estimated to be�1.7 � 103-fold during the 14 days. Their active proliferation invitro is consistent with the intense incorporation of BrdU by thecells comprising the colonies in our cell proliferation assay(below). We examined the effect of six growth factors in the basalGS medium (GSM) (SI Table 1) on the growth of colonies during
a 14-day culture period. Four growth factors, bFGF, EGF, LIF,and IGF1, exhibited significant effects on colony growth, al-though they did not seem to prolong the cell proliferation phase.PDGF and glial cell line-derived neurotrophic factor, which has acritical effect on male GS cell proliferation, had no effect(Fig. 1C).
Characterization of Isolated Ovarian Cells. During the first round ofculture (14 days), many oocytes (�20 �m in diameter) continuedto protrude from the surface of the colonies and were releasedinto the medium as single oocytes with or without a zonapellucida (Fig. 1 A). To determine whether these oocytes hadbeen derived from mitotically active cells (presumptive femaleGS cells) or from preexisting prophase-I oocytes, we performeda cell proliferation assay by adding BrdU to the culture medium.The BrdU-treated colonies were sectioned and analyzed immu-nochemically. Most of the cells comprising the colony were �10�m in diameter and stained positively for BrdU, but none werepositive for mouse vasa homologue (MVH, a germ-cell marker).In contrast, oocytes �15 �m in diameter that were positive forMVH were all negative for BrdU (Fig. 2A), indicating that theywere derived from preexisting small oocytes that had alreadyentered prophase I. Consistent with this conclusion, the imprint-ing status of cells comprising the colonies was the somatic type(parent-of-origin-specific methylation), not the germ-cell type(complete demethylation or biallelic methylation) (SI Fig. 6).After a few passages, the colonies ceased producing oocytes,probably because their precursor oocyte stock had been de-pleted. This finding was confirmed by the disappearance ofmRNA for oocyte-specific genes in the colonies after the thirdpassage (Fig. 2B).
Thus, the small cells in the colonies were most likely somaticcells, but we did not know which ovarian cell type(s) was being
Fig. 1. Development of thecal stem cell colonies in vitro. (A) The size of thecolonies increased rapidly during the first several days and gradually reachedthe maximum size of �120 �m in diameter. Many oocytes (�20 �m indiameter) continued to protrude from the surface of the colonies (arrows).(Scale bars: Left, 100 �m; Right, 50 �m.) (B) The colonies are weakly positive foralkaline phosphatase staining. (C) Effect of growth factors on colony sizeduring the first 14 days of culture. Single growth factors, or some combinationthereof, were added to the basic GSM. *, P � 0.05; **, P � 0.01 [compared withgrowth-factor-free GSM (three replicates, one-way ANOVA)].
Fig. 2. Cytological and gene expression analysis of thecal stem cell colonies.(A) Paraffin-embedded section of colonies stained for BrdU (blue) and MVH(brown). Small cells were generally positive for BrdU but not for MVH. Grow-ing oocytes (arrows) were positive for MVH but not for BrdU, indicating thatthe oocytes came from preexisting cells in the colonies. (Scale bar: 50 �m.) (B)Gene expression analysis (RT-PCR) of the colonies. After the third passage (P3),oocytes disappeared from the colonies, as demonstrated by the absence ofoocyte-specific markers (zona pellucida proteins Zp1, Zp2, and Zp3). MEF,mouse embryonic fibroblast. (C) Electron micrographs of the colonies. Eachcell in the colonies had a relatively small cytoplasm that was rich in ribosomesand rough ER (arrowheads) (Upper). Cell-to-cell connections were sparse, andtheir intercellular space was filled with basement membrane (BM) and colla-gen fibers (CF) (Upper). A thecal stem cell (T) directly surrounds the zonapellucida (ZP) of an oocyte (O), but unlike typical granulosa cells, it does notprotrude foot processes to the oocyte surface through the zona (Lower).Arrowheads indicate microvilli from the oocyte. (Scale bars: 2 �m.)
12390 � www.pnas.org�cgi�doi�10.1073�pnas.0703787104 Honda et al.
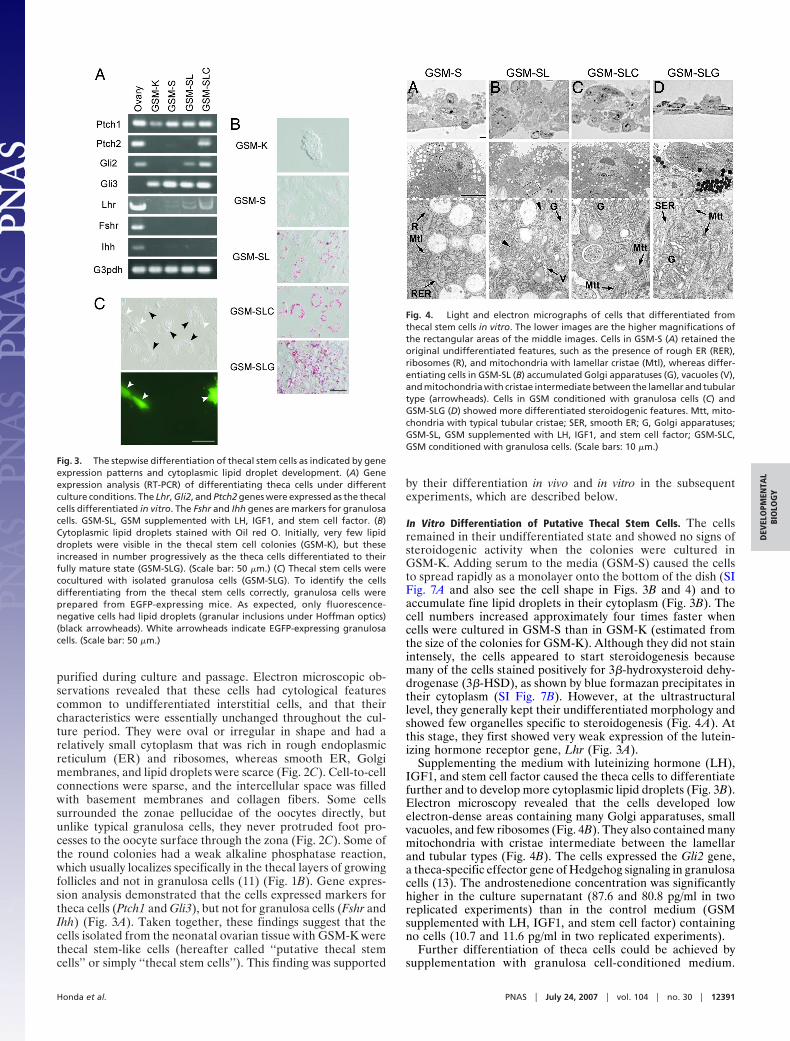
purified during culture and passage. Electron microscopic ob-servations revealed that these cells had cytological featurescommon to undifferentiated interstitial cells, and that theircharacteristics were essentially unchanged throughout the cul-ture period. They were oval or irregular in shape and had arelatively small cytoplasm that was rich in rough endoplasmicreticulum (ER) and ribosomes, whereas smooth ER, Golgimembranes, and lipid droplets were scarce (Fig. 2C). Cell-to-cellconnections were sparse, and the intercellular space was filledwith basement membranes and collagen fibers. Some cellssurrounded the zonae pellucidae of the oocytes directly, butunlike typical granulosa cells, they never protruded foot pro-cesses to the oocyte surface through the zona (Fig. 2C). Some ofthe round colonies had a weak alkaline phosphatase reaction,which usually localizes specifically in the thecal layers of growingfollicles and not in granulosa cells (11) (Fig. 1B). Gene expres-sion analysis demonstrated that the cells expressed markers fortheca cells (Ptch1 and Gli3), but not for granulosa cells (Fshr andIhh) (Fig. 3A). Taken together, these findings suggest that thecells isolated from the neonatal ovarian tissue with GSM-K werethecal stem-like cells (hereafter called ‘‘putative thecal stemcells’’ or simply ‘‘thecal stem cells’’). This finding was supported
by their differentiation in vivo and in vitro in the subsequentexperiments, which are described below.
In Vitro Differentiation of Putative Thecal Stem Cells. The cellsremained in their undifferentiated state and showed no signs ofsteroidogenic activity when the colonies were cultured inGSM-K. Adding serum to the media (GSM-S) caused the cellsto spread rapidly as a monolayer onto the bottom of the dish (SIFig. 7A and also see the cell shape in Figs. 3B and 4) and toaccumulate fine lipid droplets in their cytoplasm (Fig. 3B). Thecell numbers increased approximately four times faster whencells were cultured in GSM-S than in GSM-K (estimated fromthe size of the colonies for GSM-K). Although they did not stainintensely, the cells appeared to start steroidogenesis becausemany of the cells stained positively for 3�-hydroxysteroid dehy-drogenase (3�-HSD), as shown by blue formazan precipitates intheir cytoplasm (SI Fig. 7B). However, at the ultrastructurallevel, they generally kept their undifferentiated morphology andshowed few organelles specific to steroidogenesis (Fig. 4A). Atthis stage, they first showed very weak expression of the lutein-izing hormone receptor gene, Lhr (Fig. 3A).
Supplementing the medium with luteinizing hormone (LH),IGF1, and stem cell factor caused the theca cells to differentiatefurther and to develop more cytoplasmic lipid droplets (Fig. 3B).Electron microscopy revealed that the cells developed lowelectron-dense areas containing many Golgi apparatuses, smallvacuoles, and few ribosomes (Fig. 4B). They also contained manymitochondria with cristae intermediate between the lamellarand tubular types (Fig. 4B). The cells expressed the Gli2 gene,a theca-specific effector gene of Hedgehog signaling in granulosacells (13). The androstenedione concentration was significantlyhigher in the culture supernatant (87.6 and 80.8 pg/ml in tworeplicated experiments) than in the control medium (GSMsupplemented with LH, IGF1, and stem cell factor) containingno cells (10.7 and 11.6 pg/ml in two replicated experiments).
Further differentiation of theca cells could be achieved bysupplementation with granulosa cell-conditioned medium.
Fig. 3. The stepwise differentiation of thecal stem cells as indicated by geneexpression patterns and cytoplasmic lipid droplet development. (A) Geneexpression analysis (RT-PCR) of differentiating theca cells under differentculture conditions. The Lhr, Gli2, and Ptch2 genes were expressed as the thecalcells differentiated in vitro. The Fshr and Ihh genes are markers for granulosacells. GSM-SL, GSM supplemented with LH, IGF1, and stem cell factor. (B)Cytoplasmic lipid droplets stained with Oil red O. Initially, very few lipiddroplets were visible in the thecal stem cell colonies (GSM-K), but theseincreased in number progressively as the theca cells differentiated to theirfully mature state (GSM-SLG). (Scale bar: 50 �m.) (C) Thecal stem cells werecocultured with isolated granulosa cells (GSM-SLG). To identify the cellsdifferentiating from the thecal stem cells correctly, granulosa cells wereprepared from EGFP-expressing mice. As expected, only fluorescence-negative cells had lipid droplets (granular inclusions under Hoffman optics)(black arrowheads). White arrowheads indicate EGFP-expressing granulosacells. (Scale bar: 50 �m.)
Fig. 4. Light and electron micrographs of cells that differentiated fromthecal stem cells in vitro. The lower images are the higher magnifications ofthe rectangular areas of the middle images. Cells in GSM-S (A) retained theoriginal undifferentiated features, such as the presence of rough ER (RER),ribosomes (R), and mitochondria with lamellar cristae (Mtl), whereas differ-entiating cells in GSM-SL (B) accumulated Golgi apparatuses (G), vacuoles (V),and mitochondria with cristae intermediate between the lamellar and tubulartype (arrowheads). Cells in GSM conditioned with granulosa cells (C) andGSM-SLG (D) showed more differentiated steroidogenic features. Mtt, mito-chondria with typical tubular cristae; SER, smooth ER; G, Golgi apparatuses;GSM-SL, GSM supplemented with LH, IGF1, and stem cell factor; GSM-SLC,GSM conditioned with granulosa cells. (Scale bars: 10 �m.)
Honda et al. PNAS � July 24, 2007 � vol. 104 � no. 30 � 12391
DEV
ELO
PMEN
TAL
BIO
LOG
Y
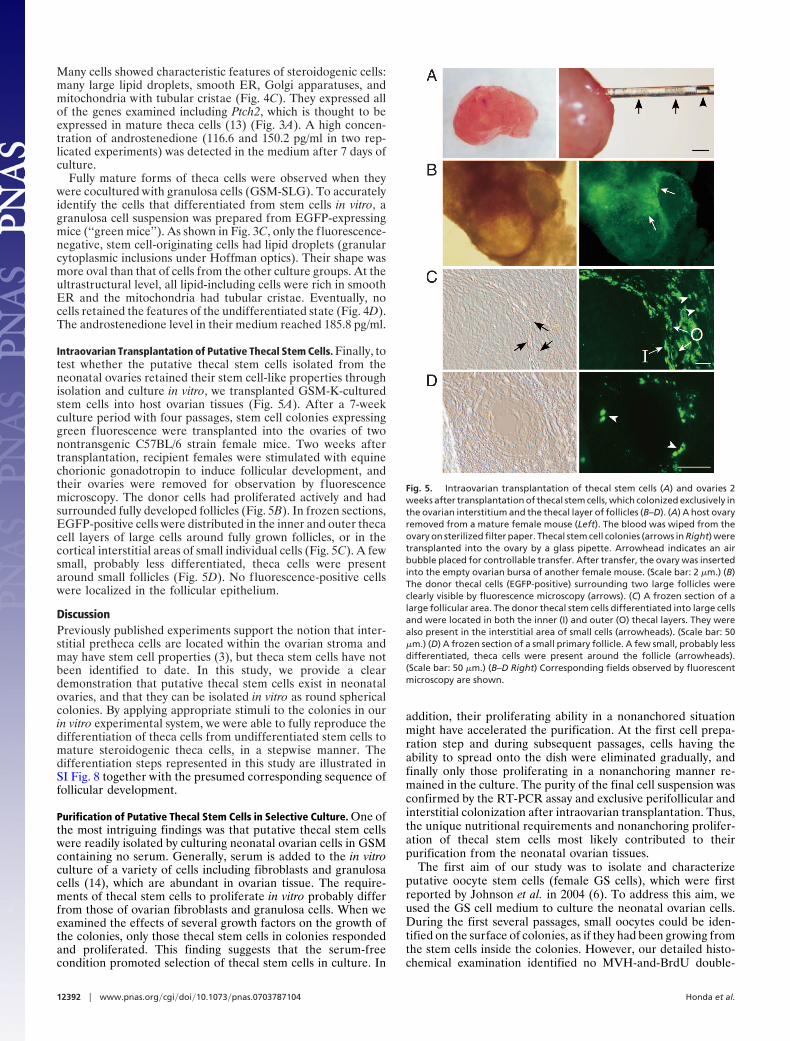
Many cells showed characteristic features of steroidogenic cells:many large lipid droplets, smooth ER, Golgi apparatuses, andmitochondria with tubular cristae (Fig. 4C). They expressed allof the genes examined including Ptch2, which is thought to beexpressed in mature theca cells (13) (Fig. 3A). A high concen-tration of androstenedione (116.6 and 150.2 pg/ml in two rep-licated experiments) was detected in the medium after 7 days ofculture.
Fully mature forms of theca cells were observed when theywere cocultured with granulosa cells (GSM-SLG). To accuratelyidentify the cells that differentiated from stem cells in vitro, agranulosa cell suspension was prepared from EGFP-expressingmice (‘‘green mice’’). As shown in Fig. 3C, only the fluorescence-negative, stem cell-originating cells had lipid droplets (granularcytoplasmic inclusions under Hoffman optics). Their shape wasmore oval than that of cells from the other culture groups. At theultrastructural level, all lipid-including cells were rich in smoothER and the mitochondria had tubular cristae. Eventually, nocells retained the features of the undifferentiated state (Fig. 4D).The androstenedione level in their medium reached 185.8 pg/ml.
Intraovarian Transplantation of Putative Thecal Stem Cells. Finally, totest whether the putative thecal stem cells isolated from theneonatal ovaries retained their stem cell-like properties throughisolation and culture in vitro, we transplanted GSM-K-culturedstem cells into host ovarian tissues (Fig. 5A). After a 7-weekculture period with four passages, stem cell colonies expressinggreen fluorescence were transplanted into the ovaries of twonontransgenic C57BL/6 strain female mice. Two weeks aftertransplantation, recipient females were stimulated with equinechorionic gonadotropin to induce follicular development, andtheir ovaries were removed for observation by fluorescencemicroscopy. The donor cells had proliferated actively and hadsurrounded fully developed follicles (Fig. 5B). In frozen sections,EGFP-positive cells were distributed in the inner and outer thecacell layers of large cells around fully grown follicles, or in thecortical interstitial areas of small individual cells (Fig. 5C). A fewsmall, probably less differentiated, theca cells were presentaround small follicles (Fig. 5D). No fluorescence-positive cellswere localized in the follicular epithelium.
DiscussionPreviously published experiments support the notion that inter-stitial pretheca cells are located within the ovarian stroma andmay have stem cell properties (3), but theca stem cells have notbeen identified to date. In this study, we provide a cleardemonstration that putative thecal stem cells exist in neonatalovaries, and that they can be isolated in vitro as round sphericalcolonies. By applying appropriate stimuli to the colonies in ourin vitro experimental system, we were able to fully reproduce thedifferentiation of theca cells from undifferentiated stem cells tomature steroidogenic theca cells, in a stepwise manner. Thedifferentiation steps represented in this study are illustrated inSI Fig. 8 together with the presumed corresponding sequence offollicular development.
Purification of Putative Thecal Stem Cells in Selective Culture. One ofthe most intriguing findings was that putative thecal stem cellswere readily isolated by culturing neonatal ovarian cells in GSMcontaining no serum. Generally, serum is added to the in vitroculture of a variety of cells including fibroblasts and granulosacells (14), which are abundant in ovarian tissue. The require-ments of thecal stem cells to proliferate in vitro probably differfrom those of ovarian fibroblasts and granulosa cells. When weexamined the effects of several growth factors on the growth ofthe colonies, only those thecal stem cells in colonies respondedand proliferated. This finding suggests that the serum-freecondition promoted selection of thecal stem cells in culture. In
addition, their proliferating ability in a nonanchored situationmight have accelerated the purification. At the first cell prepa-ration step and during subsequent passages, cells having theability to spread onto the dish were eliminated gradually, andfinally only those proliferating in a nonanchoring manner re-mained in the culture. The purity of the final cell suspension wasconfirmed by the RT-PCR assay and exclusive perifollicular andinterstitial colonization after intraovarian transplantation. Thus,the unique nutritional requirements and nonanchoring prolifer-ation of thecal stem cells most likely contributed to theirpurification from the neonatal ovarian tissues.
The first aim of our study was to isolate and characterizeputative oocyte stem cells (female GS cells), which were firstreported by Johnson et al. in 2004 (6). To address this aim, weused the GS cell medium to culture the neonatal ovarian cells.During the first several passages, small oocytes could be iden-tified on the surface of colonies, as if they had been growing fromthe stem cells inside the colonies. However, our detailed histo-chemical examination identified no MVH-and-BrdU double-
Fig. 5. Intraovarian transplantation of thecal stem cells (A) and ovaries 2weeks after transplantation of thecal stem cells, which colonized exclusively inthe ovarian interstitium and the thecal layer of follicles (B–D). (A) A host ovaryremoved from a mature female mouse (Left). The blood was wiped from theovary on sterilized filter paper. Thecal stem cell colonies (arrows in Right) weretransplanted into the ovary by a glass pipette. Arrowhead indicates an airbubble placed for controllable transfer. After transfer, the ovary was insertedinto the empty ovarian bursa of another female mouse. (Scale bar: 2 �m.) (B)The donor thecal cells (EGFP-positive) surrounding two large follicles wereclearly visible by fluorescence microscopy (arrows). (C) A frozen section of alarge follicular area. The donor thecal stem cells differentiated into large cellsand were located in both the inner (I) and outer (O) thecal layers. They werealso present in the interstitial area of small cells (arrowheads). (Scale bar: 50�m.) (D) A frozen section of a small primary follicle. A few small, probably lessdifferentiated, theca cells were present around the follicle (arrowheads).(Scale bar: 50 �m.) (B–D Right) Corresponding fields observed by fluorescentmicroscopy are shown.
12392 � www.pnas.org�cgi�doi�10.1073�pnas.0703787104 Honda et al.

positive oocytes, indicating that only preexisting postmeioticoocytes grew out from the colonies. This result by itself does notnegate the presence of oocyte stem cells in the neonatal ovary,but these stem cells, if any, did not seem to respond to GS cellmedium, which has been used in experiments on their malecounterparts.
Proliferation Ability of Putative Thecal Stem Cells in Vitro. bFGF,EGF, LIF, and IGF1 significantly promoted the growth of stemcell colonies, but the culture conditions seemed to be suboptimalbecause they did not proliferate indefinitely. Although the cellscontinued to divide for �2 months, as shown by the increases incolony size, the proliferation rate decreased after �10 days, andthe cell number seemed to reach a plateau after 1 month ofculture. However, we cannot exclude the possibility that thecalstem cells have an inherently limited proliferation potential andthat this feature was correctly reflected in vitro. Unlike malegerm cells, which produce spermatozoa continuously throughoutlife (15), the size of the primordial oocyte reserve appears to befixed during the prenatal period. Consistent with the nondividingcharacter of oocytes, it is possible that thecal stem cells, whichlocalize in the ovarian interstitium, do not divide actively butgradually acquire a proliferation ability as they differentiate tosupport the growing follicle. Generally, the tissue-specific stemcells studied so far are thought to depend on specialized cellularmicroenvironments and the ex vivo expansion of pure popula-tions of tissue stem cells has proven elusive (16). We alsoobserved that induction of differentiation by serum simulta-neously promoted proliferation of differentiating theca cells invitro.
Differentiation of Putative Thecal Stem Cells in Vitro and in Vivo.Thecal stem cells in round colonies had morphological charac-teristics common to undifferentiated interstitial cells such assmall fibroblasts: they were oval and contained many organellesincluding mitochondria with laminar cristae, rough ER, andribosomes. These thecal stem cells maintained these undiffer-entiated features for as long as they proliferated within roundcolonies. Stimulation with serum caused these cells to spreadonto the bottom of the dish and to start to accumulate small lipiddroplets in their cytoplasm. Although they seemed to stay in arelatively undifferentiated state, they expressed a small amountof Lhr mRNA. Treatment with LH, IGF1, and stem cell factoror paracrine factors from granulosa cells caused further signif-icant differentiation of theca cells, leading to accumulation ofmany lipid droplets, formation of smooth ER, and the emer-gence of mitochondria with tubular cristae. All these cytologicalfeatures are indicative of steroidogenic ability (17), a conclusionsupported by the detection of androstenedione at a high con-centration in the supernatant. These differentiation patterns oftheca cells are consistent with previous findings showing thattheir proliferation and differentiation are independent of LHreceptors in the early stages, but become LH receptor-dependent at a later stage (18), and that IGF1 synergisticallyaugments the LH stimulation of androstenedione production(19). The stepwise emergence of several theca cell markers isalso consistent with a previous in situ hybridization study thatsuggested that granulosa-derived factors, Ihh and Dhh, arecandidate initiators of early thecal differentiation (13). Weconclude that our culture system induced the differentiation oftheca cells from their stem cells in a stepwise manner similar tothat occurring in vivo, without the presence of other cell types.However, their full differentiation as shown by cells in the innerthecal layer of follicles required the presence of granulosa cells.Granulosa cells from adult bovine ovaries also stimulate andro-stenedione production by stromal cells (20). It is unlikely thatdirect cell-to-cell contact with granulosa cells was required forsuch terminal differentiation because of the intervening base-
ment membrane between the granulosa and the inner thecallayers in normal follicles. Instead, close proximity probablypromoted a paracrine effect of the granulosa cells. The presenceof such paracrine mechanisms has been shown experimentally inan in vivo assay in hypophysectomized rats (21).
A fundamental question that remains unanswered about thedifferentiation of the theca cell lineage is the mechanisms bywhich the inner and outer thecal layers are formed from thecommon stem cells (3). The cells comprising the inner layer aresteroidogenic cells and blood vessel cells, whereas cells compris-ing the outer layer are unidentified fibroblast-like cells andsmooth muscle cells (22, 23). Our transplantation experimentsrevealed that the isolated thecal stem cells had the capacity tocontribute to the formation of both inner and outer thecal layercells, although the latter seemed to be more heterogeneous incomposition. It is possible that the difference between cells in thetwo thecal layers represents the different differentiation status ofthese theca cells’ lineage. This assumption agrees with the resultsof our in vitro differentiation experiments, in which coculturewith granulosa cells led to the terminal thecal differentiationfrom the relatively undifferentiated, fibroblast-like cells. It is alsointeresting that, only 2 weeks after transplantation, the donortheca cells were observed in fully grown follicles. Becausefollicular development in mature rodents is estimated to take �6weeks (24), it is probable that the transplanted thecal cells wereable to invade partially grown preexisting follicles.
As far as we know, this is the first demonstration of successfulintraovarian transplantation of cells isolated in vitro. We foundthat direct transplantation of cells into intact ovaries in situcaused heavy bleeding, and these transplanted cells never sur-vived. In combination with an ovary transfer (removal andreplacement) technique, the transplantation was very successful,and we could show that the putative thecal cells we purified weretransplantable, as are other tissue-specific stem cells (25). Ourintraovarian transplantation technique should enable us to as-sess the ability of a variety of cells to differentiate and functionin the ovarian tissue.
Theca Cell Culture as an in Vitro Model for Soma–Germ and Soma–Soma Interactions. In mice, assembly of oocytes and somatic cellsinto follicles occurs within the first week after birth, and fullyexpanded antral follicles appear shortly before puberty (26). Theovaries of 2-day-old mice are devoid of distinct follicular struc-tures, and when the follicles grow to enclose two to three layersof granulosa cells, theca cells start to surround the follicles andacquire the steroidogenic ability regulated primarily by LH.Androstenedione, a steroid hormone produced by theca cells, isconverted into estradiol by granulosa cells (1, 2). Thus, follicu-logenesis is a complex process in which germ cells (oocytes) andtwo types of somatic cells (theca cells and granulosa cells)interact through paracrine or direct mechanisms. In this study,we first demonstrated that putative thecal stem cells can beisolated and purified in vitro and induced to differentiate toacquire active steroidogenic ability. These isolated theca cellsmay provide invaluable experimental systems to study theca–granulosa and theca–oocyte interactions; e.g., kit–kit ligand (27)and Hedgehog signaling systems (13). We have also recentlyfound that theca cells can promote the growth of granulosa-freeoocytes in a paracrine fashion, indicating the presence of theca-derived oocyte trophic factors (unpublished data). Other thanthe factors involved in normal folliculogenesis, the mechanismsunderlying ovarian pathology caused by theca cell dysfunction,including hyperandrogenism and polycystic ovary syndrome (1),could be unraveled by studying thecal stem cells in culture.
Materials and MethodsCell Preparation and Culture. To prepare thecal stem cells, ovarieswere collected from newborn (2–4 days after birth) ICR or
Honda et al. PNAS � July 24, 2007 � vol. 104 � no. 30 � 12393
DEV
ELO
PMEN
TAL
BIO
LOG
Y

C57BL/6-Tg(CAG-EGFP)C14-Y01-FM131Osb female mice.The latter are the so-called ‘‘green mice’’ that show greenfluorescence in the whole body (28). Cell culture was performedaccording to the protocol for establishment of male GS cells withslight modifications (9, 10). In brief, isolated ovaries were treatedwith 1 mg/ml collagenase in Hanks’s solution at 37°C for 15 minand then treated with 0.2% trypsin and 1.4 mg/ml DNase for 10min. Loosened ovarian tissues were dissected into single cells bygentle pipetting and were allocated into wells of a 0.1% (wt/vol)gelatin-coated 24-well tissue-culture plate (2.0–2.5 � 105 cellsper 2.0-cm2 well). After incubation for 4 h, f loating cells wererecovered and placed into another culture well containingGSM-K (SI Table 1). The cells were maintained at 37°C under5% CO2 in air. A half-volume of the medium was changed every2–3 days. After 14 days of culture, colonies were dispersed bytrypsin treatment and replated into culture wells containingfresh medium (2� dilution). Colonies grew to the original sizein �10–14 days, and passages were repeated until the cellsstopped proliferating (at four to five passages).
Granulosa cells were prepared from follicles in the ovariesfrom 7.5 units of equine chorionic gonadotropin-stimulatedTgX(act-EGFP)Osb#50 female mice (another green mousestrain). To remove the thecal layers from the isolated follicles,they were incubated in StemPro-34 serum-free medium contain-ing 0.25% trypsin and 0.02% EDTA for 10 min at 37°C. Oocyteswere washed and stripped of the enclosing follicular cells bypipetting. Groups of granulosa cells from 50 follicles weretransferred to a four-well dish (Nunc, Roskilde, Denmark).Cultures were carried out in GSM supplemented with 15% FBS(GSM-S) (SI Table 1) at 37°C with 5% CO2 in air. Two days later,granulosa cell clumps were dispersed by trypsin treatment andreplated to a fresh culture plate (5� dilution). The lack ofcontamination by thecal cells in the granulosa cell suspensionwas confirmed by RT-PCR with specific primers (data notshown). Granulosa-conditioned medium was prepared fromwells after 24 h of incubation of 80–100% confluent granulosacells.
In Vitro Differentiation. Putative theca stem cells (1.1 � 105 cells)were cultured for 4–5 weeks with three passages and were thenreplated and cultured under one of the five conditions indicatedin SI Table 1. After 6 days of culture, these cells were subjectedto RT-PCR, Oil red O staining, or electron microscopic obser-vation. In some experiments, the androstenedione concentration
in the supernatant was measured by liquid chromatography-tandem mass spectrometric analysis (29). This analysis wasperformed by Teikoku Hormone (Kawasaki, Japan).
Intraovarian Transplantation. Thecal stem cells (3.5 � 105) derivedfrom C57BL/6-Tg(CAG-EGFP)C14-Y01-FM131Osb were cul-tured in GSM-K for 7 weeks with four passages. In a preliminarystudy, we found that direct transplantation of cells into intactovaries in situ caused heavy bleeding, and no cells survived afterthe operation. Therefore, we first removed the host ovaries frommature C57BL/6 females, and the blood was wiped from theovaries on sterilized filter paper (Fig. 5A). Approximately 20colonies were picked up by a glass pipette (�100 �m innerdiameter) together with a minimal amount of medium. The tipof the pipette was inserted into the host ovary along the longeraxis (Fig. 5A). The cell colonies were transplanted in theopposite side of the ovarian cortex by applying gentle positivepressure, and the pipette was slowly pulled out of the ovary. Eachovary that received donor cells was inserted into the emptyovarian bursa of another host C57BL/6 female mouse underanesthesia as described (30). Two weeks after transplantation,the host females were administered 5 units of equine chorionicgonadotropin, and the ovaries were retrieved 48 h later. Thesetransplantation experiments were replicated twice.
Cytological, Histological, and Gene Expression Analyses. Cytological,histological, and gene expression analyses were performed ac-cording to established protocols (see SI Materials and Methods).
Animals. All animals were maintained and used for experimentsin accordance with the guidelines of the RIKEN Institute.
EGFP-expressing strains of mice (green mice) used in this study belongto Dr. M. Okabe (Osaka University, Osaka, Japan) and were providedby RIKEN BioResource Center with the support of the NationalBioResources Project of the Ministry of Education, Culture, Sports,Science, and Technology of Japan. The anti-MVH antibody was kindlyprovided by Dr. T. Noce (Mitsubishi Kagaku Institute of Life Science,Tokyo, Japan). This research was supported by grants from the Ministryof Education, Culture, Sports, Science, and Technology (to A.H. andA.O.); the Ministry of Health, Labour, and Welfare (A.O.); and CoreResearch for Evolutional Science and Technology (A.O.). A.H. is therecipient of a research fellowship from the RIKEN Special PostdoctoralResearchers Program.
1. Magoffin DA (2005) Int J Biochem Cell Biol 37:1344–1349.2. Demeestere I, Centner J, Gervy C, Englert Y, Delbaere A (2005) Reproduction
(Bristol, UK) 130:147–156.3. Hirshfield AN (1991) Biol Reprod 44:1157–1162.4. Lo KC, Lei Z, Rao Ch V, Beck J, Lamb DJ (2004) Endocrinology 145:4011–
4015.5. Ge RS, Dong Q, Sottas CM, Papadopoulos V, Zirkin BR, Hardy MP (2006)
Proc Natl Acad Sci USA 103:2719–2724.6. Johnson J, Canning J, Kaneko T, Pru JK, Tilly JL (2004) Nature 428:145–150.7. Eggan K, Jurga S, Gosden R, Min IM, Wagers AJ (2006) Nature 441:1109–
1114.8. Telfer EE, Gosden RG, Byskov AG, Spears N, Albertini D, Andersen CY,
Anderson R, Braw-Tal R, Clarke H, Gougeon A, et al. (2005) Cell 122:821–822.9. Kanatsu-Shinohara M, Miki H, Inoue K, Ogonuki N, Toyokuni S, Ogura A,
Shinohara T (2005) Biol Reprod 72:985–991.10. Kanatsu-Shinohara M, Ogonuki N, Inoue K, Miki H, Ogura A, Toyokuni S,
Shinohara T (2003) Biol Reprod 69:612–616.11. Kawase E, Suemori H, Takahashi N, Okazaki K, Hashimoto K, Nakatsuji N
(1994) Int J Dev Biol 38:385–390.12. Cortvrindt R, Hu Y, Smitz J (1998) Hum Reprod 13:1292–1302.13. Wijgerde M, Ooms M, Hoogerbrugge JW, Grootegoed JA (2005) Endocrinol-
ogy 146:3558–3566.14. Zhao J, Dorland M, Taverne MA, van der Weijden GC, Bevers MM, van den
Hurk R (2000) Mol Reprod Dev 55:65–74.15. Meistrich ML, van Beek MEAB (1993) in Cell and Molecular Biology of the
Testis, eds Desjardins C, Ewing LL (Oxford Univ Press, Oxford), pp 266–295.
16. Conti L, Pollard SM, Gorba T, Reitano E, Toselli M, Biella G, Sun Y, SanzoneS, Ying QL, Cattaneo E, et al. (2005) PLoS Biol 3:1594–1606.
17. Fawcett DW (1981) The Cell (Saunders, Philadelphia), 2nd Ed.18. Gelety TJ, Magoffin DA (1997) Biol Reprod 56:938–945.19. Huang CT, Weitsman SR, Dykes BN, Magoffin DA (2001) Biol Reprod
64:451–456.20. Orisaka M, Tajima K, Mizutani T, Miyamoto K, Tsang BK, Fukuda S, Yoshida
Y, Kotsuji F (2006) Biol Reprod 75:734–740.21. Smyth CD, Miro F, Whitelaw PF, Howles CM, Hillier SG (1993) Endocrinology
133:1532–1538.22. Sato E, Kimura N, Yokoo M, Miyake Y, Ikeda JE (2006) Microsc Res Tech
69:427–435.23. Motta PM, Nottola SA, Familiari G, Makabe S, Stallone T, Macchiarelli G
(2003) Int Rev Cytol 223:177–288.24. Hirshfield AN (1991) Int Rev Cytol 124:43–101.25. Mayhall EA, Paffett-Lugassy N, Zon LI (2004) Curr Opin Cell Biol 16:713–
720.26. van den Hurk R, Zhao J (2005) Theriogenology 63:1717–1751.27. Hutt KJ, McLaughlin EA, Holland MK (2006) Mol Hum Reprod 12:61–69.28. Okabe M, Ikawa M, Kominami K, Nakanishi T, Nishimune Y (1997) FEBS Lett
407:313–319.29. Kashiwagi B, Shibata Y, Ono Y, Suzuki R, Honma S, Suzuki K (2005) J Androl
26:586–591.30. Migishima F, Suzuki Migishima R, Song SY, Kuramochi T, Azuma S, Nishijima
M, Yokoyama M (2003) Biol Reprod 68:881–887.
12394 � www.pnas.org�cgi�doi�10.1073�pnas.0703787104 Honda et al.





























