Embed Size (px)
Citation preview

1
Detection of IMP metallo-β-lactamase in carbapenem-non-susceptible 1
Enterobacteriaceae and glucose non-fermenting Gram-negative rods by 2
immunochromatography assay 3
4
Shigeyuki Notake1, 2, Mari Matsuda1, Kiyoko Tamai2, Hideji Yanagisawa2, 5
Keiichi Hiramatsu1 and Ken Kikuchi1* 6
7
Department of Infection Control Science, Faculty of Medicine, Juntendo University, 8
2-1-1 Hongo, Bunkyo-ku, Tokyo 113-8421, Japan1 and Miroku Medical Laboratory Inc., 9
659-2 Innai, Saku, Nagano 384-2201, Japan2. 10
11
Running title: Detection of IMP by immunochromatography 12
13
*Corresponding author. Mailing address: Ken Kikuchi, M.D., Ph.D. 14
Department of Infection Control Science, Faculty of Medicine, Juntendo University, 15
2-1-1 Hongo, Bunkyo-ku, Tokyo 113-8421, Japan 16
17
Phone: 81-3-3813-3111 ext. 3822 18
Fax: 81-3-5684-7830 19
E-mail: [email protected]
Copyright © 2013, American Society for Microbiology. All Rights Reserved.J. Clin. Microbiol. doi:10.1128/JCM.00234-13 JCM Accepts, published online ahead of print on 27 March 2013
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

2
ABSTRACT 21
Metallo-β-lactamases (MBLs) are transmissible carbapenemases of increasing 22
prevalence in Gram-negative bacteria among healthcare facilities worldwide. Further 23
spread of these carbapenem-resistant bacteria relies on clinical microbiological 24
laboratories correctly identifying and classifying the MBLs. In this study, we evaluated a 25
simple and rapid detection method of IMP, the most prevalent metallo-β-lactamase 26
(MBL) in Japan, we evaluated a simple and rapid detection method of IMP, the most 27
prevalent metallo-β-lactamase (MBL) in Japan, using an immunochromatography (IC) 28
assay for 181 carbapenem-non-susceptible (non-susceptible to imipenem or meropenem: 29
CNS) strains comprising 74 IMP-producing and 33 IMP-non-producing strains of 30
glucose non-fermenting Gram-negative rods (NFGNR), as well as 64 IMP-producing and 31
10 IMP-non-producing Enterobacteriaceae strains. The IC assay results were compared 32
to those from the double-disc synergy test (DDST), the MBL Etest, and the modified 33
Hodge test (MHT, only for Enterobacteriaceae). IMP type was confirmed by specific 34
PCR and direct sequencing. IC assay could detect all the IMP-type MBL including IMP-1, 35
2, 6, 7, 10, 11, 19, 20, 22, and 40-42 (new type) with 100% specificity and sensitivity 36
against all strains tested. Although sensitivity and specificity of the DDST and MHT was 37
equivalent to that for the IC assay, the positive MBL Etest was only 87% for NFGNR and 38
31% for Enterobacteriaceae due to the low MIC of imipenem causing an indeterminate 39
evaluation. These results indicated that the IC assay could be a useful alternative to PCR 40
for IMP-MBL detection screening. 41
42
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

3
INTRODUCTION 43
The recent worldwide emergence and dissemination of carbapenemase-producing 44
Gram-negative rods (GNR) that are resistant to carbapenems is a significant concern with 45
respect to patient care and infection control strategies (1). The transmissible 46
carbapenemases are divided into three different classes: class A (serine carbapenemases, 47
such as KPC), class B (metallo-β-lactamase: MBL, such as IMP, VIM, NDM), and class 48
D (OXA carbapenemases, such as OXA-23, OXA-48) (1, 2). Rapid and adequate 49
detection of carbapenemases is very important for appropriate antimicrobial 50
chemotherapies and infection control measures. Various phenotypic confirmation tests 51
for detecting carbapenemases have been performed including inhibition tests of 52
carbapenemase activity, the modified Hodge test (MHT), and detection of carbapenem 53
hydrolysis (1-8). However, there are no complete assays available to confirm and specify 54
carbapenemases correctly because carbapenemase-producing bacteria, notably 55
Enterobacteriaceae, show variable carbapenem MIC distribution (even under 56
breakpoint) and sometimes have carbapenemase-independent mechanisms, such as 57
reduced permeability by porin alternations, efflux pumping, and hyperproduction of class 58
C β-lactamases (e.g. AmpC) or extended-spectrum β-lactamases (ESBLs) that operate 59
with or without carbapenemase activity (1-4). Moreover, phenotypic assays cannot 60
specify types within each class of carbapenemases, such as IMP, VIM, NDM, SIM, and 61
GIM in MBL (1-4). Therefore, molecular confirmation of carbapenemases is 62
recommended for suspected carbapenemase-producing strains (1-4). However, although 63
molecular detection methods such as PCR and sequencing of carbapenemase genes are 64
reliable for confirmation of carbapenemases, it is difficult to perform such tests in routine 65
clinical microbiology laboratories because of the skill level required, higher cost, and 66
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

4
requirement for special equipment (1-4). A simple and rapid alternative method is thus 67
needed to confirm carbapenemase presence in bacteria. 68
In Japan, IMP-MBL is the most prevalent transmissible carbapenemase, and 69
especially members of the IMP-1 group (9, 10), while KPC is quite rare and OXA-48 has 70
not been reported (11). The first IMP-MBL was described in Pseudomonas aeruginosa in 71
Japan (12), and is now found worldwide in glucose non-fermenting Gram-negative rods 72
(NFGNR) other than P. aeruginosa and Enterobacteriaceae (1-4, 8, 13). Recently, Kitao 73
et al. (14) developed an immunochromatography (IC) assay for the production of 74
IMP-MBL in P. aeruginosa and Acinetobacter. This assay is easy to perform, rapid (≤ 20 75
min required), requires no special equipment, and detects the 24 established IMP types. In 76
addition it shows excellent correlation with PCR results. In countries like Japan, wherein 77
IMP-MBL is the most prevalent mechanism of carbapenem resistance, this assay 78
provides a useful alternative to PCR for classifying MBLs in clinical microbiology 79
laboratories. Since it is uncertain that this system can detect IMP-MBL in 80
Enterobacteriaceae, we evaluated the usefulness of this IC assay in 81
carbapenem-non-susceptible (CNS) Enterobacteriaceae and NFGNR strains with the 82
MIC of imipenem (IPM) or meropenem (MEM) > 1 μg/ml. 83
84
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

5
MATERIALS AND METHODS 85
Strains 86
A total of 181 CNS strains including Pseudomonas aeruginosa MRY 06-352 87
(producing IMP-1), and Serratia marcescens MRY 06-353 (producing IMP-19) as 88
IMP-positive controls provided by Dr. Y. Arakawa (National Infectious Disease Institute, 89
Japan) were used. These included 74 NFGNR (Pseudomonas aeruginosa, Pseudomonas 90
spp., Acinetobacter spp., and Achromobacter xylosoxidans) and 64 Enterobacteriaceae 91
(Citrobacter freundii, Enterobacter cloacae, Escherichia coli, Klebsiella spp., 92
Providencia rettgeri, and Serratia marcescens) strains that produce IMP-MBL. An 93
additional 43 CNS, but carbapenemase-negative, strains (33 NFGNR and 10 94
Enterobacteriaceae) were also used as negative controls. The 179 strains, excluding the 95
IMP-positive controls, were collected in Miroku Medical Laboratory from 2001 to 2012. 96
Each strain was species-identified by MicroScan Breakpoint Combo Panel Type 6.23J 97
(Siemens Healthcare Diagnostics, Tarrytown, NY, USA), and all the NFGNR strains were 98
re-identified using matrix-assisted laser desorption ionization-time-of-flight mass 99
spectrometry (MALDI-TOF MS, Microflex LT, Bruker Daltonik GmbH, Leipzig, 100
Germany) with MALDI Biotyper software (version 3.0, Bruker Daltonik) (15, 16). 101
Identification of Pseudomonas spp. other than P. aeruginosa and Acinetobacter spp., was 102
confirmed by 16S rRNA gene, rpoB, or gyrB sequence (17, 18). Strains used are listed in 103
Table 1, while the clinical sources and the place and date of isolation are presented in the 104
supplement material. 105
106
MIC determination 107
MICs of IPM and MEM were determined by the broth microdilution method 108
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

6
according to the CLSI M07-A9 guideline (19) and the supplement M100-S21 (20). CNS 109
was designated as intermediate or resistant to IPM or MEM (MIC > 1 μg/ml). 110
111
Phenotypic detection of MBL 112
Double disk synergy tests (DDST) with sodium mercaptoacetate (SMA; 113
metallo-β-lactamase SMA 'Eiken'; Eiken Chemical Co., Ltd., Tokyo, Japan) were 114
performed according to the manufacturer's instructions based on the method described 115
previously (21). A McFarland 0.5 standard suspension of each test strain was inoculated 116
on Mueller-Hinton agar (Nippon Beckton-Dickinson, Fukushima, Japan). Two 117
commercial Kirby-Bauer (KB) disks (Nippon Beckton-Dickinson) containing 30 μg of 118
ceftazidime (CAZ) or 10 μg IPM were placed on the plate and an SMA disk was placed at 119
a distance of 10 mm (edge to edge). Each agar plate was incubated at 35°C overnight. The 120
presence of a synergistic inhibition zone of CAZ or IPM (≥5 mm of enlargement with the 121
SMA disk side) was interpreted as positive. The MBL Etest and MHT were performed 122
according to a previous report (22) and the CLSI guideline (20). 123
124
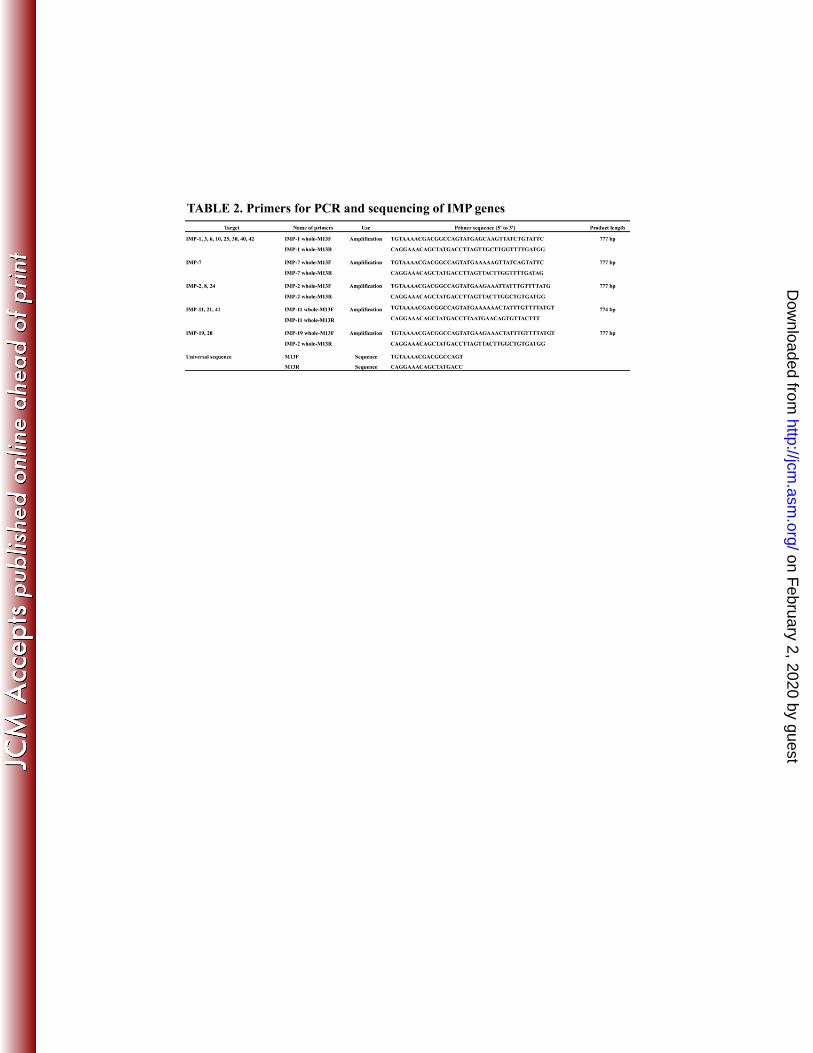
Determination of IMP-MBL genes 125
Screening of carbapenemase genes was carried out by PCR as described previously 126
(9, 23). Strains carrying transmissible carbapenemases other than IMP (VIM, SIM, GIM, 127
AIM, DIM, GIM, NDM, KPC, BIC, and OXA-48) were excluded from this study. If 128
blaIMP was positive by screening PCR, blaIMP types were determined using each 129
IMP-specific PCR to amplify the whole length of blaIMP, and direct sequencing was 130
performed. Primers used are listed in Table 2. PCR was performed in 50-μl reaction 131
mixtures that comprised 2.5 U of Ex Taq DNA polymerase (Takara Bio Inc., Shiga, 132
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

7
Japan), 0.2 mM dNTP, 25 pmol of each primer, and 2 μl of DNA template. PCR 133
conditions were as follows: initial denaturation at 95°C for 10 min, followed by 30 cycles 134
of denaturation at 95°C for 30 sec, annealing at 62°C for 1 min and DNA extension at 135
72°C for 1 min, with final extension at 72°C for 10 min. PCR products were visualized 136
under UV light exposure after 1% agarose gel electrophoresis with ethidium bromide. 137
Amplicons obtained from each PCR were sequenced using M13F and M13R primers, the 138
BigDye Terminator v3.1 Cycle Sequencing Kit (ABI, Carlsbad, CA), and an ABI 139
sequence analyzer 3730XL (ABI). Each type of IMP was determined by BLAST search 140
and data based on all the blaIMP described previously 141
(http://www.lahey.org/studies/other.asp#Table1, 24). 142
143
Detection of IMP-MBL by immunochromatography 144
The IMP-MBL IC assay kit (Quick Chaser® IMP) was kindly provided by Mizuho 145
Medy Co., Ltd. (Saga, Japan), and used according to the manufacturer’s instruction, 146
based on a previous report (14). Briefly, fresh cultured colonies were dispensed into 700 147
μl of extraction reagent solution with nonionic detergent at McFarland 4.0 standard. After 148
vigorous vortexing, 3 drops of this suspension were applied onto the sample area of the 149
IC assay test plate. These plates were incubated for 15 min at room temperature and 150
results were interpreted visually (14). 151
152
Statistical analysis 153
Receiver operating characteristic (ROC) curves and the area under the ROC curve 154
with its standard error were used to analyze the IC assay, DDST, and MHT method results, 155
using PCR results for IMP as the gold standard. Statistically significant differences were 156
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

8
evaluated by comparing 95% confidence intervals of the corresponding areas. The 157
statistical analysis was performed using STATA 11.0 (StataCorp LP, College Station, 158
TX). 159
160
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

9
RESULTS 161
Species identification of test strains and determination of IMP-type 162
Identification of all the strains of A. xylosoxidans and P. aeruginosa was identical 163
between MicroScan and MALDI-TOF MS. Both IMP-producing and 164
IMP-non-producing Pseudomonas spp. strains were identified as P. putida or P. moteilii, 165
which belonged to the Pseudomonas putida group by MALDI-TOF MS, but were 166
distinguished from any species in this group by 16S rRNA, rpoB, gyrB sequence, 167
suggesting a new species. The seventeen IMP-producing Acinetobacter strains were 168
identified as ten A. pittii, three Acinetobacter genomospecies 13, three A. soli, and one A. 169
johsonii by rpoB sequence (18), whereas the ten non-IMP-producing Acinetobacter were 170
A. baumannii (Table 1). 171
The most prevalent IMP was IMP-1, comprising 48 of 74 (64%) NFGNR and 49 of 64 172
(76%) Enterobacteriaceae strains. In this study, three new IMP-type MBLs were 173
discovered and designated as IMP-40, IMP-41, and IMP-42. IMP-40 is closely related to 174
IMP-10, with a nucleotide alteration in the T206C of blaIMP-10 causing F69S amino acid 175
substitution according to the standard numbering scheme of MBL (25). IMP-41 is similar 176
to IMP-11, with a G145T nucleotide change causing a V49F amino acid substitution. 177
IMP-42 shows a G45R amino acid substitution of IMP-1, by the G133A nucleotide 178
change. IMP-40, IMP-41, and IMP-42 were found in two P. aeruginosa, one P. 179
aeruginosa, and two A. soli strains, respectively (Table 1 and Supplemental material). 180
One A. pittii and one E. coli strain showed a C294T nucleotide change in blaIMP-1 and a 181
C306T change in blaIMP-6, respectively, but neither base change altered the amino acid 182
sequence (Table 1). 183
184
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

10
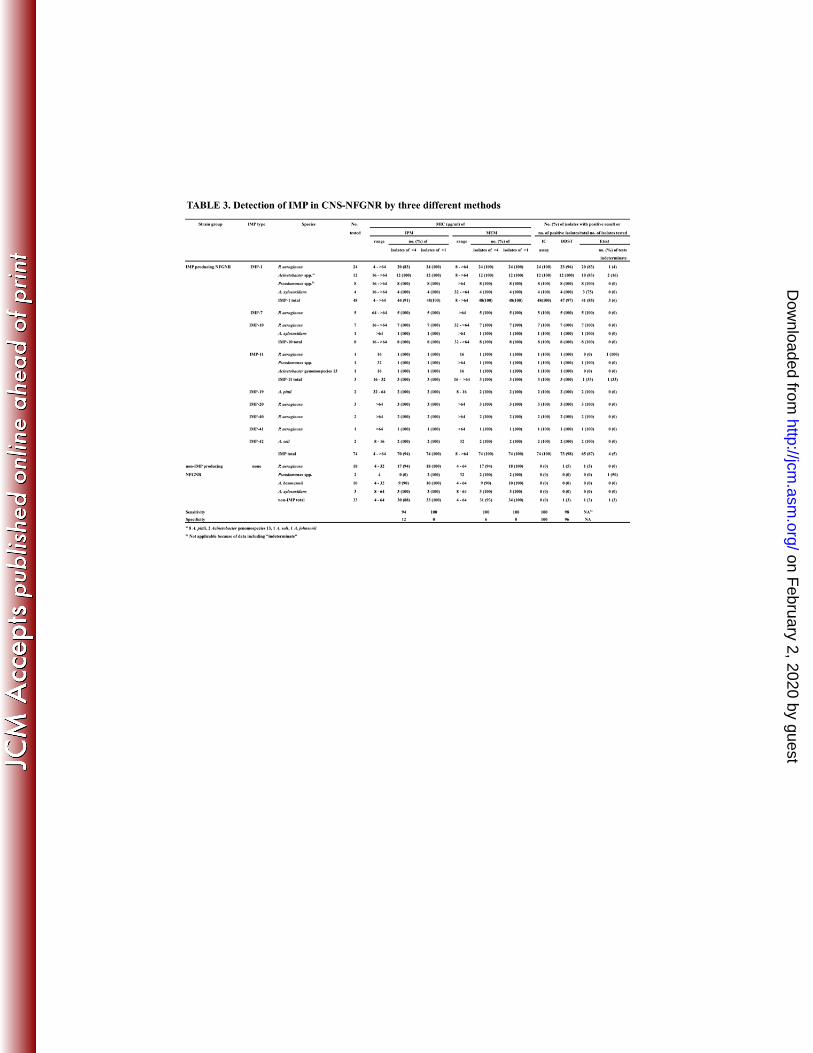
Detection of IMP in CNS-NFGNR by three different methods 185
Table 3 shows the results of three IMP detection methods in CNS-NFGNR. The IC 186
assay detected all IMP-MBL regardless of IMP type including the new IMPs, IMP-40, 187
IMP-41, and IMP-42, with 100% sensitivity and specificity (area under the curve of 188
ROC: 1.000). DDST also showed good sensitivity and specificity at 96% and 98%, 189
respectively. The areas under the curve of ROC for the IC assay and DDST were not 190
statistically different. However, there were fewer positive detections with the MBL Etest 191
(87%) compared to the IC assay and DDST because some strains showed a low MIC of 192
IPM and four tests (5%) were be evaluated as "indeterminate" due to the MIC being out of 193
range. Moreover, the MBL Etest showed negative results for three strains of 194
IMP-1-producing P. aeruginosa, one IMP-1-producing A. xylosoxidans and one 195
IMP-11-Acinetobacter genomospecies 13. The MBL Etest results were excluded for 196
statistical analysis because of data including "indeterminate". 197
198
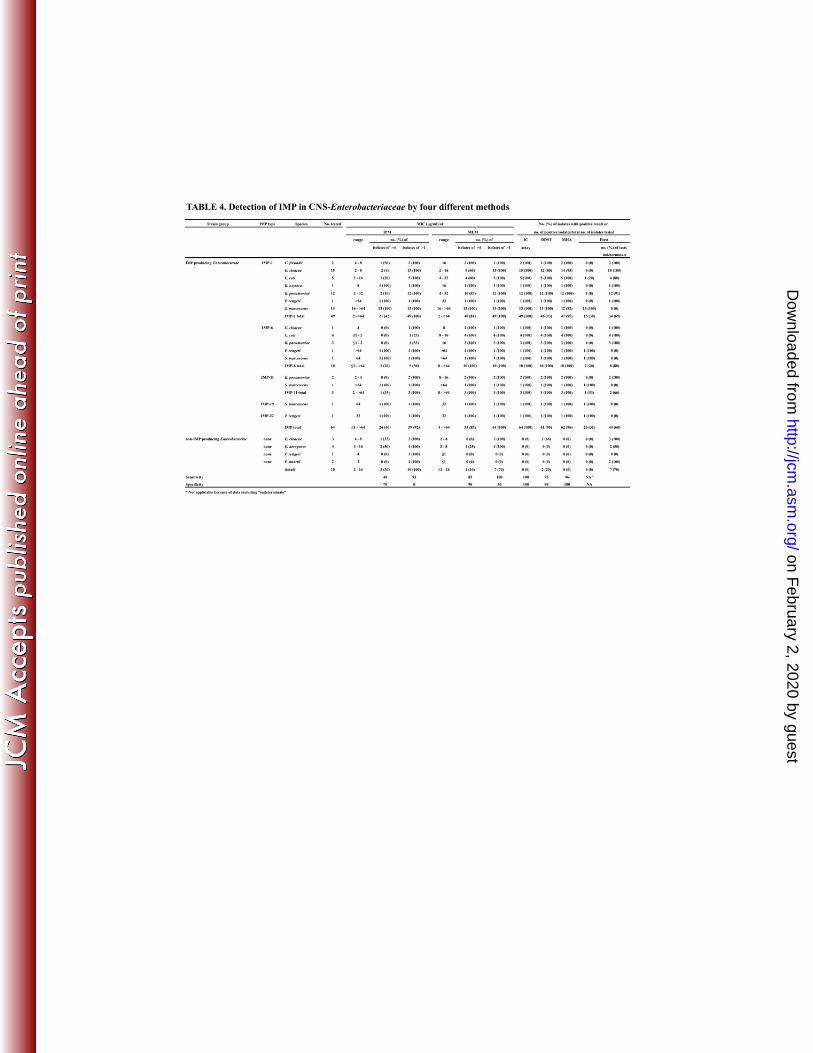
Detection of IMP in CNS-Enterobacteriaceae by four different methods 199
Table 4 details results for the four IMP detection methods in CNS-Enterobacteriaceae. 200
In these bacteria, the IC assay also showed 100% sensitivity and specificity (area under 201
the curve of ROC: 1.000), as did DDST and MHT at 95% and 96%, respectively, and the 202
areas under the curve were not statistically different among these three detection methods. 203
In comparison, the MBL Etest was inadequate for 68% of IMP-producing 204
CNS-Enterobacteriaceae, because 59% of these strains showed low susceptibility to IMP 205
within the Etest range (≤4 μg/ml). In addition, the area under the curve of ROC could not 206
be calculated for the MBL Etest results because some data were deemed "indeterminate". 207
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

11
DISCUSSIONS 208
Many types of carbapenemase-producing Gram-negative rods have emerged 209
worldwide with various carbapenemases types disseminated among countries and 210
continents. IMP-MBL is one of the most prevalent carbapenemases in Asia, Europe, and 211
some areas of the North and South America, and Australia (13). Currently, IMP-MBL has 212
spread through most Enterobacteriaceae including E. coli and K. pneumoniae, both of 213
which are prevalent in community-acquired and healthcare-associated infections (1-5, 214
13). Since some IMP-producing Enterobacteriaceae strains, especially, E. coli and K. 215
pneumoniae, show low-level resistance or even sensitivity to carbapenems, the CLSI 216
breakpoints of carbapenems in Enterobacteriaceae have changed since June 2010, as 217
follows: IPM (S ≤ 4 μg/ml, I = 8 μg/ml, R ≥ 16 μg/ml,) and MEM (S ≤ 4 μg/ml, I = 8 218
μg/ml, R ≥ 16 μg/ml) have moved to (S ≤ 1 μg/ml, I = 2 μg/ml, R ≥ 4 μg/ml), and (S ≤ 1 219
μg/ml, I = 2 μg/ml, R ≥ 4 μg/ml) (3, 19). Moreover, EUCAST has established 220
epidemiological cut-off values (ECOFFs) that discriminate wild-type isolates lacking any 221
carbapenem-resistance mechanisms from those possessing resistance as IPM (1-4 μg/ml) 222
and MEM (0.125-0.25 μg/ml), in addition to carbapenem clinical breakpoints (2). If a 223
strain of Enterobacteriaceae for which carbapenem MICs are below ECOFFs is detected 224
and becomes prevalent, clinical microbiology laboratory staff should consider 225
surveillance of carbapenemase-producing strains (2). Such surveillance by routine 226
clinical microbiological laboratories ideally requires a rapid, reliable, inexpensive, and 227
simple detection method of IMP. The IC assay evaluated in this study showed excellent 228
sensitivity (100%) and specificity (100%) across MIC ranges (area under the curve of 229
ROC: 1.000). Current phenotypic detection of MBL in clinical laboratories uses testing 230
by DDST or MHT, and the IC assay results in this study are statistically comparable with 231
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

12
those acquired using these two tests by ROC analysis. However, both DDST and MHT 232
require overnight culture (> 16 h), compared with only 20 min for the IC assay. The 233
DDST is also not recommended by the Dutch Working Party on the Detection of Highly 234
Resistant Microorganisms because the sensitivity depends on the optimal distance 235
between the disks, and this cannot be predicted (4). In our results, the MBL Etest was 236
inappropriate for detecting IMP, especially in Enterobacteriaceae, mainly because of the 237
low MIC of IPM. These results confirm similar studies previously described (26-28). 238
Moreover, the MBL Etest had five false-negative results involving four IMP-1 and one 239
IMP-11-producing strains. In addition, Laraki (29) described that even 10 mM EDTA 240
could not inhibit the enzyme activity of IMP-1 isolated from one Japanese strain. Poor 241
inactivation of IMP by EDTA might account for such false-negative results of the MBL 242
Etest. The IC assay detected all IMP types including 3 new types as IMP-40, 41, and 42, 243
and did not react with VIM-2 in two P. aeruginosa strains, NDM-1 in one K. pneumoniae 244
isolate, OXA-23 in one A. baumannii strain and KPC-2 in K. pneumoniae ATCC 245
BAA-1705 (data not shown). Two monoclonal antibodies designated as 4C9-C/F6 and 246
4E7-C/F6 are used in this IC assay system (14). Since 4C9-C/F6 and 4E7-C/F6 recognize 247
the highly conserved amino acids 124-130 (H2 region) and 134-140 (S6 region) in IMP 248
(14, 24), and since the H2 and S6 regions of the new IMP types, IMP-40, 41, and 42, are 249
well conserved, the IC assay can detect such IMP types. Thus, the IC assay might also be 250
used for detecting emerging types of IMP that retain the original H2 and S6 regions. The 251
cost of an IC assay is also comparable to other conventional or molecular detection 252
methods of MBL at ¥1,000 per test. 253
In conclusion, the IC assay is considered an alternative and suitable method to detect 254
IMP in routine clinical microbiology testing including IMP surveillance with advantages 255
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

13
in cost, processing time, reliability, convenience, and no requirement for special 256
equipment. In countries like Japan where IMP-MBL is highly prevalent, and in areas of 257
newly developed MBL endemics, IC assay testing would be quite useful for the early 258
detection and control of nosocomial and community spreads of bacteria with IMP-MBL. 259
260
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

14
ACKNOWLEDGMENTS 261
We thank Dr. Y. Arakawa for providing the IMP-positive control strains. We also thank 262
Mr. T. Mizutani for assistance with the statistical analysis. This study was supported in 263
part by a Grant-in-Aide (S0991013) from Ministry of Education, Culture, Sport, Science, 264
and Technology, Japan (MEXT) for the Foundation of Strategic Research Projects in 265
Private Universities. 266
267
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

15
REFERENCES 268
1. Queenan AM, Bush K. 2007. Carbapenemases: the versatile β-lactamases. Clin. 269
Microbiol. Rev. 20: 440-458. 270
2. Miriagou V, Cornaglia G, Edelstein M, Galani I, Giske CG, Gniadkowski M, 271
Malamou-Lada E, Martinez-Martinez L, Navarro F, Nordmann P, Peixe L, 272
Pournaras S, Rossolini GM, Tsakris A, Vatopoulos A, Canton R. 2010. Acquired 273
carbapenemases in Gram-negative bacterial pathogens: detection and surveillance 274
issues. Clin. Microbiol. Infect. 16: 112-122. 275
3. Nordmann P, Gniadkowski M, Giske CG, Poirel L, Woodford N, Miriagou V, The 276
European Network on Carbapenemases. 2012. Identification and screening of 277
carbapenemase-producing Enterobacteriaceae. Clin. Microb. Infect. 18: 432-438. 278
4. Stuart JC, Leverstein-van Hall MA, on behalf of members of the Dutch Working 279
Party on the Detection of Highly Resistant Microorganisms. 2010. Guideline for 280
phenotypic screening and confirmation of carbapenemases in Enterobacteriaceae. Int. 281
J. Antimicrob. Agents. 36: 205-210. 282
5. Bernabeu S, Poirel L, Nordmann P. Spectrophotometry-based detection of 283
carbapenemase producers among Enterobacteriaceae. 2012. Diagn. Microbiol. Infect. 284
Dis. 74: 88-90. 285
6. Dortet L, Poirel L, Nordmann P. Rapid identification of carbapenemase types in 286
Enterobacteriaceae and Pseudomonas spp. by using a biochemical test. 2012. 287
Antimicrob. Agents Chemother. 56: 6437-6440. 288
7. Hrabák J, Walková R, Študentová V, Chudáčková E, Bergerová T. 289
Carbapenemase activity detection by matrix-associated laser desorption 290
ionization-time of flight mass spectrometry. 2011. J. Clin. Microbiol. 49: 3222-3227. 291
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

16
8. Walsh, TR, Toleman MA, Poirel L, Nordmann P, 2005. Metallo-β-lactamases: the 292
quiet before the storm? Clin. Microbiol. Rev. 18: 306-325. 293
9. Shibata N, Doi Y, Yamane K, Yagi T, Kurokawa H, Shibayama K, Kato H, Kai K, 294
Arakawa Y. 2003. PCR Typing of genetic determinants for metallo-β-lactamases and 295
integrases carried by Gram-negative bacteria isolated in Japan, with focus on the Class 296
3 integron. J. Clin. Microbiol. 41: 5407–5413. 297
10. Nishio H, Komatsu M, Shibata N, Shimakawa K, Sueyoshi N, Ura T, Satoh K, 298
Toyokawa M, Nakamura T, Wada Y, Orita T, Kofuku T, Yamasaki K, Sakamoto 299
M, Kinoshita S, Aihara M, Arakawa Y. 2004. Metallo-β-lactamase-producing 300
Gram-negative bacilli: laboratory-based surveillance in cooperation with 13 clinical 301
laboratories in the Kinki region of Japan. J. Clin. Microbiol. 42, 5256–5263. 302
11. Suzuki S, Yamane K, Wachino J, Matsui M, Konda T, Arakawa Y. 2012. Three 303
months survey of multidrug-resistant Enterobacteriaceae in Japan. Nihon Rinsho. 70, 304
187-191 (in Japanese). 305
12. Watanabe M, Iyobe S, Inoue M, Mitsuhashi S. 1991. Transferable imipenem 306
resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 35: 147-151. 307
13. Cornglia G, Giamarellou H, Rossolini GM. 2011. Metallo-β-lactamases: a last 308
frontier for β-lactams? Lancet Infect. Dis. 11: 381-393. 309
14. Kitao T, Miyoshi-Akiyama T, Tanaka M, Narahara K, Shimojima M, Kirikae T. 310
2011. Development of an immunochromatographic assay for diagnosing the 311
production of IMP-type metallo-β-lactamases that mediate carbapenem resistance in 312
Pseudomonas. J. Microbiol. Methods 87: 330-337. 313
15. Saffert RT, Cunningham SA, Ihde SM, Jobe KEM, Mandrekar J, Patel R. 2011. 314
Comparison of Bruker Biotyper matrix-assisted laser desorption ionization-time of 315
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

17
flight mass spectrometer to BD Phoenix automated microbiology system for 316
identification of Gram-negative bacilli. J. Clin. Microbiol. 49: 887-892. 317
16. Espinal P, Seifert H, Dijkshoorn L, Vila J, Roca I. 2012. Rapid and accurate 318
identification of genomic species from the Acinetobacter baumannii (Ab) group by 319
MALDI-TOF MS. Clin. Microbiol. Infect. 18: 1097-1103. 320
17. Mullet M, Lalucat J, Garcia-Valdes E. 2010. DNA sequence-based analysis of the 321
Pseudomonas species. Environm. Microbiol. 12: 1513-1530. 322
18. Turton JF, Shah J, Ozongwu C, Pike R. 2010. Incidence of Acinetobacter species 323
other than A. baumannii among clinical isolates of Acinetobacter. evidence for 324
emerging species. J. Clin. Microbiol. 48: 1445-1449. 325
19. CLSI 2012. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria 326
That Grow Aerobically; Approved Standard-Ninth Edition. M07-A09. Clinical and 327
Laboratory Standards Institute, Wayne, PA. 328
20. CLSI 2011. Performance Standards for Antimicrobial Susceptibility Testing; 21st 329
Informational Supplement M100-S21. Clinical and Laboratory Standards Institute, 330
Wayne, PA. 331
21. Arakawa Y, Shibata N, Shibayama K, Kurokawa H, Yagi T, Fujiwara H, and 332
Goto M. 2000. Convenient test for screening metallo-β-lactamase-producing 333
Gram-negative bacteria by using thiol compounds. J. Clin. Microbiol. 38: 40–43. 334
22. Walsh TR, Bolmström A, Qwärnström A, Gales A. 2002. Evaluation of a new 335
Etest for detecting metallo-β-lactamases in routine clinical testing. J. Clin. Microbiol. 336
40: 2755-2759. 337
23. Poirel L, Walsh TR, Cuvillier V, Nordmann P. 2011. Multiplex PCR for detection 338
of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 70: 119-123. 339
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from

18
24. Widmann M, Pleiss J, Oelschlaeger P. 2012. Systematic analysis of metallo- 340
β-lactamases using an automated database. Antimicrob. Agents Chemother. 56: 341
3481-3491. 342
25. Garau G, García-Sáez I, Bebrone C, Anne C, Mercuri P, Galleni M, Frère J-M, 343
Dideberg O. 2004. Update of the standard numbering scheme for class B β-lactamase. 344
Antimicrob. Agents Chemother. 48: 2347-2349. 345
26. Doyle D, Peirano G, Lascols C, Lloyd T, Church DL, Pitout JDD. 2012. 346
Laboratory detection of Enterobacteriaceae that produce carbapenemases. J. Clin. 347
Microbiol. 50: 3877-3880. 348
27. Espedido BA, Thomas LC, Iredell JR. 2007. Metallo-β-lactamase or 349
extended-spectrum β-lactamase: a wolf in sheep's clothing. J. Clin. Microbiol. 45: 350
2034-2036. 351
28. Galani I, Rekastina PD, Hatzaki D, Plachouras D, Souli M, Giamarellou H. 2008. 352
Evaluation of different laboratory tests for the detection of metallo-β-lactamase 353
production in Enterobacteriaceae. J. Antimicrob. Chemother. 61: 548-553. 354
29. Laraki N, Franceschini N, Rossolini GM, Santucci P, Meunier C, de Pauw E, 355
Amicosante G, Frère JM, Galleni M. 1999. Biochemical characterization of the 356
Pseudomonas aeruginosa 101/1477 metallo-β-lactamase IMP-1 produced by 357
Escherichia coli. Antimicrob. Agents Chemother. 43: 902-906. 358
on February 2, 2020 by guest
http://jcm.asm
.org/D
ownloaded from





![產生 Carbapenemase · 是 blaKPC (Klebsiella pneumoniae carbapenemase, KPC) 及blaNDM-1 (New Delhi metallo-β-lactamase 1, NDM-1) [1-4]。這些研究同時也顯示出抗藥性](https://img.pdfslide.tips/doc/110x75/5f528987e1fbfa1f2a6f78bc/cc-carbapenemase-blakpc-klebsiella-pneumoniae-carbapenemase-kpc-blandm-1.jpg)




![Carbapenemase Producerende Enterobacteriaceaeusers.skynet.be/fa686441/GAB/GAB-2013/CPE-VanVaerenbergh...Microsoft PowerPoint - Ppt0000015 [Alleen-lezen] Author gdepourcq Created Date](https://img.pdfslide.tips/doc/110x75/5ff09d71fbb91a2fb9442a71/carbapenemase-producerende-ent-microsoft-powerpoint-ppt0000015-alleen-lezen.jpg)