Embed Size (px)
Citation preview


Jurnal Biologi Indonesia diterbitkan oleh Perhimpunan Biologi Indonesia. Jurnal ini memuat hasil penelitian ataupun kajian yang berkaitan dengan masalah biologi yang diterbitkan secara berkala dua kali setahun (Juni dan Desember).
Editor Ketua
Prof. Dr. Ibnu Maryanto Anggota
Prof. Dr. I Made Sudiana Dr. Deby Arifiani
Dr. Izu Andry Fijridiyanto
Dewan Editor Ilmiah
Dr. Abinawanto, F MIPA UI
Dr. Achmad Farajalah, FMIPA IPB
Prof. Dr. Ambariyanto, F. Perikanan dan Kelautan UNDIP
Dr. Didik Widiyatmoko, Pusat Konservasi Tumbuhan Kebun Raya-LIPI
Dr. Dwi Nugroho Wibowo, F. Biologi UNSOED
Dr. Gatot Ciptadi F. Peternakan Universitas Brawijaya
Dr. Parikesit, F. MIPA UNPAD
Dr. Faisal Anwari Khan, Universiti Malaysia Sarawak Malaysia
Assoc. Prof. Monica Suleiman, Universiti Malaysia Sabah, Malaysia
Dr. Srihadi Agungpriyono, PAVet(K), F. Kedokteran Hewan IPB
Y. Surjadi MSc, Pusat Penelitian ICABIOGRAD
Drs. Suharjono, Pusat Penelitian Biologi-LIPI
Dr. Tri Widianto, Pusat Penelitian Limnologi-LIPI
Dr. Witjaksono Pusat Penelitian Biologi-LIPI
Sekretariat Eko Sulistyadi M.Si, Dewi Citra Murniati M.Si, Hetty Irawati PU, S.Kom
Alamat d/a Pusat Penelitian Biologi - LIPI
Jl. Ir. H. Juanda No. 18, Bogor 16002 , Telp. (021) 8765056 Fax. (021) 8765068
Email : [email protected]; [email protected]; [email protected]; [email protected] Website : http://biologi.or.id
Jurnal Biologi Indonesia : Akreditasi: No. 657/AU3/P2MI-LIPI/07/2015.

JURNAL BIOLOGI INDONESIA
Diterbitkan Oleh:
Perhimpunan Biologi Indonesia
Bekerja sama dengan
PUSLIT BIOLOGI-LIPI

OBITUARI
Redaksi Jurnal Biologi Indonesia telah kehilangan seorang editor penelaah Dr. Ir Sri Sulandari, M.Sc.
yang telah berpulang kerahmat Allah SWT pada tanggal 18 Agustus 2015 Jam 16.10 di RSCM,
Jakarta. Jabatan terakhir almarhumah sebagai Peneliti Madya/IVc di Pusat Penelitian Biologi-LIPI
sebagai ahli DNA Molekuler yang menekuni kajian DNA pada ayam lokal Indonesia dan berbagai
hidupan liar khususnya pada burung. Tiga tahun terakhir sangat aktif berusaha menyelamatkan
populasi kambing Gembrong di Kabupaten Karanganyar, Bali. Almarhumah meninggalkan seorang
suami Prof. Dr. Muladno, MSA yang bekerja sebagai guru besar di Fakultas Peternakan, Institut
Pertanian bogor dan saat ini juga sebagai Direktur Jendral Peternakan dan Kesehatan Hewan,
Kementerian Pertanian, serta dua anak laki-laki Aussie Andry Vermarchnanto M. dan Endyea
Mendelian.

Jurnal Biologi Indonesia yang diterbitkan oleh PERHIMPUNAN BIOLOGI INDONESIA bekerjasama
dengan PUSLIT BIOLOGI-LIPI. Edisi volume 11 No. 2 tahun 2015 memuat 15 artikel lengkap dan
satu artikel tulisan pendek. Penulis pada edisi ini sangat beragam yaitu dari Balai Besar Penelitian
Veteriner-Deptan, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik
Pertanian, Bogor, Balai Penelitian Tanaman Sayuran Lembang, Bandung, Departemen Konservasi
Sumberdaya Hutan dan Ekowisata, Fakultas Kehutanan-IPB, Dept. Biokimia FMIPA-IPB, Institut
Sains dan Teknologi Nasional Jakarta, Pusat Penelitian dan Pengembangan Sumberdaya Pesisir &
Laut, Balitbang Kelautan & Perikanan, Kementerian Kelautan & Perikanan, Departemen Manajemen
Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, IPB. Program Studi Manajemen
Sumberdaya Perairan, Fakultas Ilmu Kelautan dan Perikanan-Universitas Maritim Raja Ali Haji-
Tual, Pusat Konservasi Tumbuhan Kebun Raya–LIPI, Puslit Biologi-LIPI, Puslit Bioteknologi-LIPI.

Jurnal Biologi Indonesia mengucapkan terima kasih dan penghargaan kepada para pakar yang telah
turut sebagai penelaah dalam Volume 11 No 2, Desember 2015:
Dr. Niken Tunjung Murti Pratiwi, Fakultas Perikanan dan Ilmu Kelautan IPB
Dr. Agus Prijono Kartono, Fakultas Kehutanan IPB
Ir. Drs. Eko Harsono MSi, Puslit Limnologi-LIPI
Dra. Donowati Tjokrokusumo M.Phil, Pusat Teknologi Bioindustri, BPPT
Ir. M. Syamsul Arifin Zein MSi, Puslit Biologi LIPI
Drh. Anang S. Achmadi MSc, Puslit Biologi LIPI
Dr. Yuyu S. Poerba, Puslit Biologi LIPI
Ir. Dwi Agustiyani MSc, Puslit Biologi LIPI
Dr. Apon Zaenal Mustopa, Puslit Bioteknologi LIPI
Dr. Yopi Puslit Bioteknologi LIPI
Dr. Joeni S. Rahajoe, Puslit Biologi LIPI
Dr. Wartka Rosa Farida, Puslit Biologi LIPI
BIOLOGI

Halaman
Efikasi Vaksin Inaktif Bivalen Avian Influenza Virus Subtipe H5N1 (Clade 2.1.3. dan Clade
2.3.2) di Indonesia
169
NLP. Indi Dharmayanti & Risa Indriani
Klon-klon Kentang Transgenik Hasil Persilangan Terseleksi Tahan terhadap Penyakit
Hawar Daun Phytophthora infestans Tanpa Penyemprotan Fungisida di Empat Lapangan
Uji Terbatas
177
Alberta Dinar Ambarwati, Kusmana, & Edy Listanto
Penambahan Inokulan Mikroba Selulolitik pada Pengomposan Jerami Padi untuk Media 187
Tanam Jamur Tiram Putih (Pleurotus ostreatus)
Iwan Saskiawan
Identifikasi Molekular dan Karakterisasi Morfo-Fisiologi Actinomycetes Penghasil Senyawa
Antimikroba
195
Arif Nurkanto & Andria Agusta
Populasi dan Kesesuaian Habitat Langkap (Arenga obtusifolia Mart.) 205
di Cagar Alam Leuweung Sancang, Jawa Barat
Didi Usmadi, Agus Hikmat, Joko Ridho Witono, & Lilik Budi Prasetyo
Optimasi Produksi Enzim Amilase dari Bakteri Laut Jakarta (Arthrobacter arilaitensis ) 215
Awan Purnawan, Y. Capriyanti, PA. Kurniatin, N. Rahmani, & Yopi
Pengaruh Antioksidan Eksopolisakarida dari Tiga Galur Bakteri Asam Laktat pada Sel
Darah Domba Terinduksi tert-Butil Hidroperoksida (t-BHP)
225
Fifi Afiati, Nina Ainul Widad, & Kusmiati
Ekosistem Lamun sebagai Bioindikator Lingkungan di P. Lembeh, Bitung, Sulawesi Utara 233
Agustin Rustam, Terry L. Kepel, Mariska A. Kusumaningtyas, Restu Nur Afi
Ati, August Daulat, Devi D. Suryono, Nasir Sudirman, Yusmiana P. Rahayu,
Peter Mangindaan, Aida Heriati, & Andreas A. Hutahaean
Identification of Bioactive Compound from Microalga BTM 11 as Hepatitis C Virus RNA 243
Helicase Inhibitor
Apon Zaenal Mustopa, Rifqiyah Nur Umami, Prabawati Hyunita Putri, Dwi
susilaningsih, & Hilda Farida
Kemampuan Cerna Protein dan Energi Metabolisme Perkici Pelangi (Trichoglossus
haematodus )
253
Rini Rachmatika & Andri Permata Sari
Optimasi Enzim α-Amilase dari Bacillus amyloliquefaciens O1 yang Diinduksi Substrat
Dedak Padi dan Karboksimetilselulosa
259
Yati Sudaryati Soeka, Maman Rahmansyah, & Sulistiani
Kajian Aspek Ekologis dan Daya Dukung Perairan Situ Cilala 267
Niken T.M. Pratiwi, Sigid Hariyadi, Inna Puspa Ayu, Aliati Iswantari,
Novita MZ, & Tri Apriadi

Halaman
Penanda Genetik Tarsius (Tarsius spp.) dengan Menggunakan Gen Cytochrome Oxidase I
(COI) DNA Mitokondria (mtDNA) Melalui Metode Sekuensing
275
Wirdateti, Sri Wijayanti Wulandari, & Paramita Cahyaningrum Kuswandi
Carboxymethyl Cellulose Hydrolyzing Yeast Isolated from South East Sulawesi, Indonesia 285
Atit Kanti
Uji Bakteri Simbiotik dan Nonsimbiotik Pelarutan Ca vs. P dan Efek Inokulasi Bakteri pada
Anakan Turi (Sesbania grandiflora L. Pers.)
295
Sri Widawati
TULISAN PENDEK 309
Mating behavior of Slow Loris (Nycticebus coucang ) at Captivity
Wartika Rosa Farida & Andri Permata Sari

Penambahan Inokulan Mikroba Selulolitik pada Pengomposan Jerami Padi
untuk Media Tanam Jamur Tiram Putih (Pleurotus ostreatus)
(The addition of Cellulolytic Microorganisms in Composting Process of Paddy Straw as
White Oyster Mushroom (Pleurotus ostreatus) Substrate)
Iwan Saskiawan
Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia,
Jl. Raya Jakarta Bogor Km. 46 Cibinong 16911. Email: [email protected]
Memasukkan: Juni 2014, Diterima: Februari 2015
ABSTRACT
Recently, the cultivation of oyster mushroom (Pleurotus ostreatus) has increased enormously because of some reasons. Mushroom growers utilize sawdust, byproduct of timber industry as main substrate in fruiting body production. Consequently, the availability of sawdust becomes an obstacle during mushroom cultivation. The aim of this study was to evaluate the effetivity of paddy rice straw as an alternative substrate in oyster mushroom cultivation. The paddy rice straw was inoculated with a cellulolytic microbs during composting process. They are Bacillus subtilis, Pseudomonas aeruginosa, Trichoderma harzianum and Aspergillus niger. The result showed that the fastest growing mycelia by fully colonizing 1.1 kg size baglog was obtained when the paddy rice straw was treated with B. subtilis (63.00 days), followed by the treatment with P. aeruginosa (63.67 days), A. niger (65.00 days), T. harzianum (67.33 days), and negative control (67.33 days) respectively. On the other hand, the treatment of P. aeruginosa gaved the highest production of fruiting body (123.33g) followed by the treatment with B. subtilis (113.33g), A. niger (90.00g), control (83.33g) and T. harzianum (78.33g) per bag log over 2 period of time harvesting.
Keywords : Pleurotus ostreatus, paddy rice straw, compost
ABSTRAK
Jamur tiram (Pleurotus ostreatus) merupakan salah satu jenis jamur pangan yang banyak dibudidayakan saat ini. Budidaya jamur tiram biasanya menggunakan serbuk gergaji sebagai media tanam. Oleh karena itu ketersediaan serbuk gergaji menjadi masalah yang dihadapi oleh pembudidaya jamur tiram. Penelitian ini bertujuan untuk mengkaji penggunaan jerami padi sebagai media tanam alternatif dalam budidaya jamur tiram dengan menggunakan mikroba selulolitik dalam proses pengomposannya. Rancangan penelitian yang digunakan adalah Rancangan Acak Lengkap (RAL) dengan empat mikroorganisme selulolitik sebagai perlakuan yaitu Bacillus subtilis (M1), Pseudomonas aeruginosa (M2), Trichoderma harzianum (M3), dan Aspergillus niger (M4). Hasil penelitian menunjukkan bahwa perlakuan M1 (B. subtilis) menunjukkan waktu pertumbuhan miselia jamur yang paling cepat yaitu 63,00 hari, diikuti dengan perlakuan M2, M4, M3 dan kontrol masing-masing selama 63,67; 65,00; 67,33 dan 67,33 hari. Sedangkan untuk produksi tubuh buah pada panen pertama, perlakuan M2 (P. aeruginosa) menunjukkan hasil yang tertinggi (123,33 g), diikuti dengan M1 (113,33 g), M4 (90,00 g) , kontrol (83,33 g) dan M3 (78,33 g).
Kata Kunci :jamur tiram, jerami padi, kompos
Jurnal Biologi Indonesia11 (2): 187-193 (2015)
PENDAHULUAN
Produksi jamur pangan atau edible mushroom
merupakan salah satu komoditas hortikultura
yang akhir-akhir ini berkembang dengan pesat.
Masyarakat luas sudah mulai mengenal jamur
pangan sebagai bahan sayuran yang dikonsumsi
setiap hari. Menurut data dari Direktorat Budidaya
dan Pascapanen Sayuran dan Tanaman Obat,
Direktorat Jenderal Hortikultura, Kementrian
Pertanian luas panen jamur di Indonesia meningkat
terus dari tahun ke tahun. Selain itu nilai ekspor
jamur pangan ternyata juga menduduki peringkat
yang cukup tinggi dibandingkan dengan komoditas
sayuran lainnya (Bahar 2012). Menurut Chang &
Miles (2004), karena nilai gizinya yang tinggi
dan mengandung beberapa senyawa aktif yang
bersifat sebagai imunomodulator yang dapat
meningkatkan ketahanan tubuh, jamur pangan
juga dapat digunakan sebagai bahan pangan

188
Iwan Saskiawan
fungsional (nutirceutical) yang bersifat sebagai
bahan nutrisi yang berefek kesehatan. Selain itu,
jamur juga merupakan salah satu komoditas
sayuran organik yang tidak menggunakan pupuk
sintetik dan pestisida sehingga sangat membantu
dalam menjaga kelestarian lingkungan. Limbah yang
berasal dari media tumbuh jamur juga dapat
dijadikan sebagai pupuk organik yang sangat baik
untuk kesuburan tanah dan tanaman (Siddhant &
Singh 2009).
Budidaya jamur pangan biasanya dilakukan
dengan menggunakan limbah pertanian sebagai
media tanam. Limbah pertanian mengandung
lignoselulosa, selulosa, dan hemiselulosa yang
sangat diperlukan oleh jamur untuk pertumbuhannya
(Mandel et al. 2005). Beberapa jenis jamur pangan
bisa dibudidayakan dengan menggunakan limbah
jerami padi, limbah kopi, tanaman eceng gondok,
serbuk gergaji dan limbah pertanian lainnya
(Saskiawan & Sudarmono 1993; Bisaria et al.
1987; Madan et al. 1987; Royse et al. 2004;
Murugesan, et al. 1995). Salah satu jenis jamur
pangan yang paling mudah dan banyak
dibudidayakan adalah jamur tiram putih (P.
ostreatus). Jamur ini memiliki kandungan
protein lebih tinggi daripada kacang-kacangan,
mengandung vitamin B1 dan B2 lebih tinggi
daripada jamur yang lain, serta memiliki asam
folat lebih tinggi dibandingkan dengan sayuran
dan daging. Asam folat sebagai komponen obat
dapat mengatasi gejala anemia, diabetes, dan
tekanan darah tinggi. Kandungan kalorinya
yang rendah baik untuk dikonsumsi orang yang
sedang menjalankan program diet (Cohen et al.
2002).
Jamur tiram biasanya dibudidayakan dengan
menggunakan media tanam serbuk gergaji. Media
tanam tersebut dengan penambahan bahan-bahan
tertentu seperti bekatul/dedak, tepung jagung, kapur
dan gypsum. Setelah melalui proses pencampuran
bahan-bahan tersebut kemudian dimasukkan
dalam kantung plastik tahan panas ukuran 18 x
25 cm dan dibentuk menyerupai botol. Media tanam
ini disebut dengan baglog. Baglog tersebut
kemudian disterilkan, didiamkan selama satu malam
untuk menurunkan suhu kemudian diinokulasi
dengan bibit jamur tiram.
Peningkatan jumlah pembudidaya jamur
menyebabkan ketersediaan serbuk gergaji
menjadi terbatas. Oleh karena itu perlu
dilalukan pencarian bahan alternatif pengganti
serbuk gergaji untuk budidaya jamur tiram.
Penelitian ini bertujuan menggunakan limbah
jerami padi sebagai media tanam alternatif
dalam budidaya jamur tiram. Dalam penelitian
ini digunakan inokulan mikroba selulolitik
dalam proses pengomposan jerami padi untuk
persiapan budidaya jamur tiram.
BAHAN DAN CARA KERJA
Kultur murni Trichoderma harzianum,
Aspergillus niger, Bacillus subtilis, dan
Pseudomonas aeruginosa merupakan koleksi
dari LIPI Microbial Culture Collection (MC),
Pusat Penelitian Biologi LIPI. Bibit jamur tiram
putih (P. ostreatus) diperoleh dari Laboratorium
Biokimia Mikroba, Bidang Mikrobiologi, Pusat
Penelitian Biologi LIPI. Jerami padi diperoleh
dari kebun percobaan yang berada di Cibinong
Science Center.
Kultur B. subtilis dan P. aeruginosa
diremajakan pada media NA miring, diinkubasi
pada suhu 37oC selama 3 hari. Sel bakteri dipanen
dengan menambahkan campuran akuades steril
sebanyak 3ml, selanjutnya diukur kerapatan optiknya
(OD) sampai target nilai 0,5 dengan panjang
gelombang 600 nm.
Kultur A. niger (LIPI-MC 713) dan T.
harzianum (LIPI-MC 732) diremajakan pada
media PDA miring, diinkubasi pada suhu kamar
sampai sporulasi merata (4 hari). Spora dipanen
dengan menambahkan campuran akuades steril
dan tween-80 0,1% sebanyak 3 ml, selanjutnya
diukur kerapatan optiknya (OD) sampai target
nilai 0,5 dengan panjang gelombang 600 nm.
Inokulum bakteri B. subtilis dan P.
aeruginosa yang memiliki nilai OD 0,5 masing-
masing dimasukkan ke dalam media kaldu
nurien sebanyak 4 ml (2% dari volume media)
secara aseptis, sedangkan inokulum jamur A.
niger dan T. harzianum yang memiliki nilai OD
0,5 masing-masing diinokulasikan ke dalam
media ekstrak tauge cair, diinkubasi dalam
rotary shaker dengan kecepatan 120 rpm
dengan suhu 30oC selama 4 hari.
Jerami padi di jemur di bawah sinar
matahari langsung hingga kering dan warnanya
berubah menjadi kecoklatan, kemudian dipotong-
potong dengan ukuran kurang lebih 10 cm.

189
Penambahan Inokulan Mikroba Selulolitik pada Pengomposan Jerami Padi
Potongan jerami ditimbang dengan berat kering
masing-masing 2 kg dan direndam dalam ember
yang berisi air bersih selama 3 jam, kemudian
ditiriskan selama 1 jam sampai tidak ada air yang
menetes (kadar air 60%) (Parlindungan 2001). Jerami
tersebut dimasukkan ke dalam kantong plastik
berwana hitam dan masing-masing diinokulasi
dengan suspensi bakteri B. subtilis (M1), P.
aeruginosa (M2), dan suspensi spora jamur T.
harzianum (M3), A. niger (M4) masing-masing
sebanyak 10% (b/v) dan kontrol tanpa inokulasi
mikroorganisme lignoselulolitik (K). Campuran
substrat diaduk hingga merata, kemudian
dimasukkan ke dalam kantong plastik masing-
masing dengan berat 2 kg berat kering, mulut
kantong plastik diikat, dikomposkan selama 6
hari. Perlakuan pengomposan masing-masing
diulang 3 kali.
Sebanyak 10 gram sampel kompos ditimbang
dan ditambah 50 ml akuades steril. Sampel tersebut
kemudian dibuat homogen dengan magnetic stirrer
selama 30 menit kemudian disentrifugasi selama
20 menit dengan kecepatan 9000 rpm. Supernatan
hasil sentrifugasi digunakan sebagai sampel
untuk mengukur kadar glukosa dan xilosa.
Perhitungan kadar glukosa dan xilosa
dilakukan menurut metode Bernfeld (1955)
dengan beberapa modifikasi. Sebanyak 0,25 ml
sampel ditambah dengan 0,50 ml pereaksi Dinitro
Salisilic Acid (DNS), kemudian divortex dan
dipanaskan pada waterbath dengan suhu 90oC selama
5 menit. Setelah dingin, sampel ditambah 5 ml
akuades dan diukur gula reduksinya dengan
spektrofotometer pada panjang gelombang 540
nm. Standar yang digunakan adalah larutan gula
dan xilosa.
Jerami padi yang telah dikomposkan perlakuan
K, M1, M2, M3, dan M4 masing-masing ditambah
dengan 1% gipsum (w/w), 1% CaCO3 (w/w) dan
10% dedak (w/w). Kadar air diatur hingga
kelembaban 60%. Campuran media selanjutnya
diaduk hingga rata dan dimasukkan dalam
plastik tahan panas dengan ukuran 18x25cm,
masing-masing diisi sebanyak 1kg. Baglog
kemudian dipasangi cincin plastik dengan
diameter ±2,5cm dan disumbat dengan kapas, dilapisi
kertas dan plastik selanjutnya disterilkan dalam
autoklaf dengan suhu 121oC tekanan 1 atm
selama 1 jam. Baglog yang telah disterilkan
didiamkan selama 1 malam. Baglog diinokulasi
dengan bibit jamur tiram putih sebanyak 3
sendok teh secara aseptis. Baglog disimpan di dalam
kubung inkubasi dan diamati pertumbuhan
miseliumnya sampai waktu full grown yaitu kondisi
miselium jamur tiram yang berwarna putih
memenuhi seluruh permumkaan media tanam.
Baglog yang telah full grown dipindahkan ke kubung
produksi, dibuka sumbat kapas dan cincin bambu,
diinkubasi dan diamati waktu terjadi pembentukan
tubuh buah pertama.
Pemanenan tubuh buah dilakukan secara
manual dengan cara mencabut seluruh bagian
tubuh buah jamur tiram ketika ukuran diameter
tudung mencapai 5 cm. Hal tersebut biasanya
tecapai pada umur 4-5 hari terhitung sejak
pembentukan calon tubuh buah.
Parameter utama yang diamati dalam penelitian
ini adalah waktu full grown, waktu awal
pembentukan tubuh buah, dan berat basah tubuh
buah. Suhu dan pH substrat selama pengomposan
diamati sebagai parameter pendukung.
Rancangan penelitian menggunakan Rancangan
Acak Lengkap satu faktor dengan 5 perlakuan (4
jenis mikroorganisme dan 1 kontrol) dengan 3 kali
ulangan. Data yang diperoleh dianalisis dengan
analysis of varian (ANOVA) yaitu dengan uji F pada
taraf signifikasi 95% dan 99%, kemudian dilanjutkan
dengan uji Duncan.
HASIL
Pengamatan secara visual menunjukkan
bahwa jerami padi kering yang digunakan
berwarna kecoklatan. Setelah proses pengomposan
dengan perlakuan penambahan mikroba selulolitik
selama 6 hari, warna jerami berubah menjadi
coklat kehitaman, memiliki aroma seperti tanah,
dengan tekstur jerami sedikit lebih lembek atau
remah. Gambar 1 menunjukkan bahwa
pengomposan jerami padi dengan inokulan B.
subtilis (M1) menghasilkan kadar monosakarida
glukosa yang paling tinggi yaitu 4,90 mmol per 10
gram sampel dibandingkan dengan Kontrol (K) 0,62
mmol, 4,21 mmol (M2), 2,96 mmol (M3) dan 2,37
mmol (M4). Sedangkan monosakarida xilosa
tertinggi dihasilkan dari pengomposan jerami dengan
perlakuan inokulan P. aeruginosa (M2) yaitu 7,24
mmol per 10 gram sampel dibandingkan dengan
Kontrol (K) 0,96 mmol, 6,09 mmol (M1), 4,08
mmol (M3) dan 4,28 mmol (M4).

190
Iwan Saskiawan
Perubahan suhu selama proses pengomposan
jerami dengan penambahan mikroba selulolitik dapat
dilihat pada Gambar 2. Perubahan suhu pada
seluruh perlakuan penambahan mikroba menunjukan
pola yang sama. Kenaikan suhu terlihat pada proses
pengomposan dari hari ke 1 sampai ke 3. Suhu
tertinggi dicapai ketika pengomposan memasuki hari
ke 3 dan dilanjutkan dengan penurunan suhu sampai
hari ke 6. Perlakuan P. aeruginosa (M2)
menunjukkan suhu tertinggi pada hari ke 3
dibandingkan dengan perlakuan yang lain yaitu
49,6oC.
Selain suhu, parameter lain yang diukur
selama proses pengomposan adalah pH.
Gambar 3. menunjukkan perubahan pH substrat
pada 5 perlakuan dalam masa inkubasi 6 hari.
Grafik penurunan pH pada seluruh perlakuan
menunjukkan pola perubahan pH yang hampir
sama. Penurunan pH yang cukup signifikan
terjadi pada hari ke 3 proses pengomposan.
Sedangkan pada hari ke 6, perlakuan T.
harzianum (M3) menunjukkan pH yang paling
rendah yaitu 6,65.
Pengaruh pengomposan dengan penambahan
inokulan mikroba terhadap pertumbuhan jamur
tiram, waktu panen pertama dan berat basah tubuh
buah pada pemanenan pertama ditampilkan pada
Tabel 1. Pertumbuhan jamur tiram ditetapkan
dengan mengukur waktu yang diperlukan oleh
miselium untuk memenuhi seluruh permukaan
media tanam (full grown). Dari 5 perlakuan
yang dicobakan, perlakuan B. subtilis (M1)
menunjukkan waktu full grown yang lebih cepat
yaitu 63 hari. Waktu full grown tersebut berbeda
nyata dengan perlakuan T. harzianum (M3), A.
niger (M4), dan kontrol pada pada taraf 5%.
Sedangkan waktu produksi tubuh buah jamur
tiram yang pertama diantara 4 perlakuan dan
kontrol menunjukkan bahwa perlakuan B.
subtilis (M1) menghasilkan tubuh buah pertama
paling cepat yaitu pada hari ke 82,3. Waktu
pertama produksi tubuh buah tersebut berbeda
nyata pada taraf 5% dengan perlakuan T.
Gambar 1. Produksi gula reduksi glukosa dan xilosa selama proses pengomposan ( □ = glukosa,
■ = xilosa, K = Kontrol; M1 = B. subtilis; M2 = P. aeruginosa; M3 = T. harzianum; dan M4 = A. niger)
Gambar 2. Perubahan suhu selama proses pengom-posan (○ = K, Kontrol; □ = M1, B. subtilis; Δ = M2, P. aeruginosa; ◊ = M3, T. harzi-
anum; dan × = M4, A. niger)
Gambar 3. Perubahan pH selama proses pengompo-san (○ = K, Kontrol; □ = M1, B. subtilis; Δ = M2, P. aeruginosa; ◊ = M3, T. harzianum; dan × = M4, A. niger)
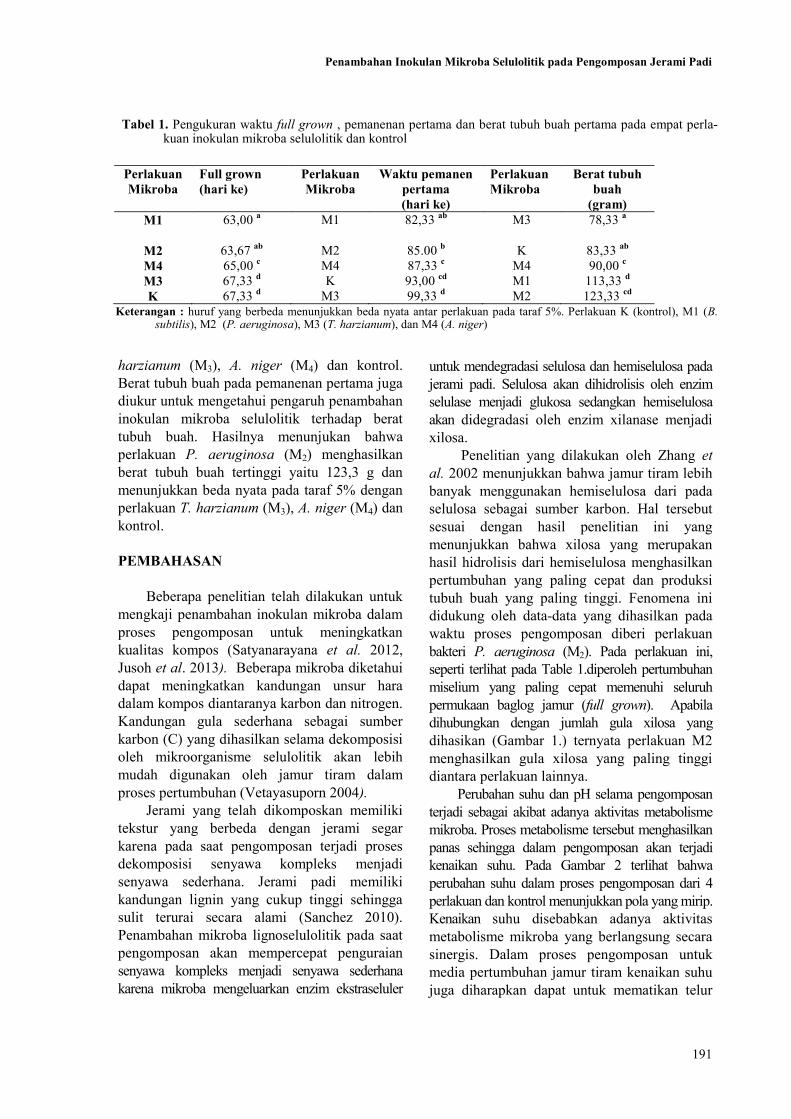
191
Penambahan Inokulan Mikroba Selulolitik pada Pengomposan Jerami Padi
harzianum (M3), A. niger (M4) dan kontrol.
Berat tubuh buah pada pemanenan pertama juga
diukur untuk mengetahui pengaruh penambahan
inokulan mikroba selulolitik terhadap berat
tubuh buah. Hasilnya menunjukan bahwa
perlakuan P. aeruginosa (M2) menghasilkan
berat tubuh buah tertinggi yaitu 123,3 g dan
menunjukkan beda nyata pada taraf 5% dengan
perlakuan T. harzianum (M3), A. niger (M4) dan
kontrol.
PEMBAHASAN
Beberapa penelitian telah dilakukan untuk
mengkaji penambahan inokulan mikroba dalam
proses pengomposan untuk meningkatkan
kualitas kompos (Satyanarayana et al. 2012,
Jusoh et al. 2013). Beberapa mikroba diketahui
dapat meningkatkan kandungan unsur hara
dalam kompos diantaranya karbon dan nitrogen.
Kandungan gula sederhana sebagai sumber
karbon (C) yang dihasilkan selama dekomposisi
oleh mikroorganisme selulolitik akan lebih
mudah digunakan oleh jamur tiram dalam
proses pertumbuhan (Vetayasuporn 2004).
Jerami yang telah dikomposkan memiliki
tekstur yang berbeda dengan jerami segar
karena pada saat pengomposan terjadi proses
dekomposisi senyawa kompleks menjadi
senyawa sederhana. Jerami padi memiliki
kandungan lignin yang cukup tinggi sehingga
sulit terurai secara alami (Sanchez 2010).
Penambahan mikroba lignoselulolitik pada saat
pengomposan akan mempercepat penguraian
senyawa kompleks menjadi senyawa sederhana
karena mikroba mengeluarkan enzim ekstraseluler
untuk mendegradasi selulosa dan hemiselulosa pada
jerami padi. Selulosa akan dihidrolisis oleh enzim
selulase menjadi glukosa sedangkan hemiselulosa
akan didegradasi oleh enzim xilanase menjadi
xilosa.
Penelitian yang dilakukan oleh Zhang et
al. 2002 menunjukkan bahwa jamur tiram lebih
banyak menggunakan hemiselulosa dari pada
selulosa sebagai sumber karbon. Hal tersebut
sesuai dengan hasil penelitian ini yang
menunjukkan bahwa xilosa yang merupakan
hasil hidrolisis dari hemiselulosa menghasilkan
pertumbuhan yang paling cepat dan produksi
tubuh buah yang paling tinggi. Fenomena ini
didukung oleh data-data yang dihasilkan pada
waktu proses pengomposan diberi perlakuan
bakteri P. aeruginosa (M2). Pada perlakuan ini,
seperti terlihat pada Table 1.diperoleh pertumbuhan
miselium yang paling cepat memenuhi seluruh
permukaan baglog jamur (full grown). Apabila
dihubungkan dengan jumlah gula xilosa yang
dihasikan (Gambar 1.) ternyata perlakuan M2
menghasilkan gula xilosa yang paling tinggi
diantara perlakuan lainnya.
Perubahan suhu dan pH selama pengomposan
terjadi sebagai akibat adanya aktivitas metabolisme
mikroba. Proses metabolisme tersebut menghasilkan
panas sehingga dalam pengomposan akan terjadi
kenaikan suhu. Pada Gambar 2 terlihat bahwa
perubahan suhu dalam proses pengomposan dari 4
perlakuan dan kontrol menunjukkan pola yang mirip.
Kenaikan suhu disebabkan adanya aktivitas
metabolisme mikroba yang berlangsung secara
sinergis. Dalam proses pengomposan untuk
media pertumbuhan jamur tiram kenaikan suhu
juga diharapkan dapat untuk mematikan telur
Perlakuan
Mikroba
Full grown
(hari ke)
Perlakuan
Mikroba
Waktu pemanen
pertama
(hari ke)
Perlakuan
Mikroba
Berat tubuh
buah
(gram)
M1 63,00 a M1 82,33
ab M3 78,33
a
M2 63,67 ab
M2 85.00 b K 83,33
ab
M4 65,00 c M4 87,33
c M4 90,00
c
M3 67,33 d K 93,00
cd M1 113,33
d
K 67,33 d M3 99,33
d M2 123,33
cd
Tabel 1. Pengukuran waktu full grown , pemanenan pertama dan berat tubuh buah pertama pada empat perla-kuan inokulan mikroba selulolitik dan kontrol
Keterangan : huruf yang berbeda menunjukkan beda nyata antar perlakuan pada taraf 5%. Perlakuan K (kontrol), M1 (B. subtilis), M2 (P. aeruginosa), M3 (T. harzianum), dan M4 (A. niger)

192
Iwan Saskiawan
atau larva serangga yang akan menjadi hama
dalam budidaya jamur tiram. Dalam penelitian
suhu paling tinggi yang dicapai relatif lebih
rendah dibandingkan dengan penelitian dari
Jusoh et al. 2013 hal ini disebabkan karena
dalam proses pengomposan dipengaruhi oleh
volume material bahan kompos. Dalam penelitian ini
pengomposan dilakukan dalam karung ukuran 20 kg,
sehingga panas yang dihasilkan tidak setinggi
apabila dilakukan proses pengomposan dalam
jumlah banyak .
Perubahan pH terjadi karena adanya
aktivitas mikroorganisme selama proses
fermentasi yang menghasilkan asam organik
(Stofella & Brian 2001). Dalam penelitian ini
grafik penurunan pH pada semua perlakuan
menunjukkan pola yang mirip. Analisis statistik
menunjukkan pH pada akhir pengomposan tidak
berbeda nyata pada setiap perlakuan. Perubahan pH
pada proses fermentasi ini juga menyediakan
kondisi derajat keasaman yang sesuai untuk
pertumbuhan jamur tiram.
Tubuh buah jamur merupakan salah satu
fase generatif dalam siklus hidup jamur. Secara
umum pembentukan tubuh buah pada jamur
pangan dipengaruhi oleh faktor lingkungan dan
faktor fisiologis jamur pangan tersebut. Faktor
lingkungan tersebut adalah temperatur, kelembaban,
intensitas cahaya, dan konsentrasi CO2. Kebutuhan
temperatur pada pertumbuhan miselium biasanya
lebih tinggi daripada fase pembentukan tubuh
buah. Masing-masing jenis jamur memerlukan
temperatur dan kelembaban yang berbeda untuk
pembentukan tubuh buahnya. Pada fase
pertumbuhan vegetatif atau pertumbuhan miselium,
jamur tiram putih memerlukan suhu udara berkisar
antara 27-29oC, dengan kelembaban udara 75-85
%. Sedangkan pada fase pembentukan tubuh
buah jamur tiram memerlukan suhu udara
berkisar antara 25-27oC dan kelembaban 80-
90%. Dalam penelitian ini produksi tubuh buah
tertinggi yang dihasilkan pada pemanenan
pertama diperoleh pada perlakuan pemberian P.
aeruginosa (M2). Sedangkan waktu full grown
dan pemanenan hari pertama tercepat dicapai
pada perlakuan M1 (B. subtilis) meskipun
analisa statistik antara perlakuan M1 dan M2
untuk parameter ini tidak menghasilkan
perbedaan yang signifikan pada taraf 5%. Hasil
penelitian Liestianty & Nurhasanah (2011)
menunjukkan bahwa beberapa bakteri dari
genus Bacillus mempunyai aktivitas enzim
hydrolase yang dapat menghidrolisis gula
rantrai panjang.
Hasil yang diperoleh dari panen pertama
jamur tiram yang dibudidayakan dengan media
jerami padi dengan perlakuan M2 yaitu 123,33
gr masih relatif lebih rendah apabila dibandingkan
dengan hasil yang diperoleh apabila media tanam
yang digunakan adalah serbuk kayu yaitu 140 gr
(Moonmoon et al. 2010). Meskipun demikian
dalam penelitian ini jerami padi dapat
digunakan sebagai media tanam alternatif ketika
ketersediaan serbuk kayu mulai sulit untuk
didapatkan.
KESIMPULAN
Penggunaan mikroorganisme selulolitik
dalam pengomposan substrat jerami padi
sebagai media tanam alternatif jamur tiram
mempengaruhi waktu full grown dan berat hasil
panenan jamur tiram. Bakteri B. subtilis
merupakan mikroorganisme yang menghasilkan
kompos dengan waktu full grown terbaik dan
berat hasil panenan pertama jamur tiram
terbanyak yaitu 63,00 hari dan 113,33 gram.
UCAPAN TERIMAKASIH
Ucapan terima kasih disampaikan kepada
Diar Anggraini Savira Dewi dari Fakultas Sains
dan Matematika, Universitas Diponegoro,
Semarang yang telah membantu pelaksanaan
kegiatan penelitian ini. Penelitian ini dibiayai
dari dana penelitian DIPA Tematik Pusat
Penelitian Biologi, Lembaga Ilmu Pengetahuan
Indonesia (LIPI) tahun anggaran 2012.
DAFTAR PUSTAKA
Bahar, YH. 2012. Kebijakan dan Dukungan
Pengembangan Agribisnis Jamur. Direktorat
Budidaya dan Pascapanen Sayuran dan
Tanaman Obat, Direktorat Jenderal
Hortikultura. Disampaikan pada Pendampi-
ngan Kelembagaan Jamur di Surakarta.
Bernfeld. 1955. Amylase a and b, In Methods in

193
Penambahan Inokulan Mikroba Selulolitik pada Pengomposan Jerami Padi
Enzymology (Colowick SP, Kaplan NO,
ed.) Academic Press Inc, New York 1: 149
-158.
Bisaria R, M. Madan, & VS. Bisaria. 1987.
Biological efficiency and nutritive value of
Pleurotus sajor caju cultivated on different
agro-wastes. Biological Waste 19: 239-255.
Chang, ST. & PG. Miles 2004. Mushrooms
Cultivation, Nutritional Value, Medicinal
Effect, and Environmental Impact. Second
Edition. CRC Press LLC. Boca Raton,
Florida.
Cohen, R., L. Persky, & Y. Hadar. 2002.
Biotechnological applications and potential
of wood-degrading mushrooms of the
genus Pleurotus. Applied Microbiology
Biotechnology. 58: 582-594.
Jusoh, MLC, LA. Manaf, & PA. Latiff. 2013.
Composting of rice straw with effective
microorganisms (EM) and its influence on
compost quality. Iranian Journal of
Environmental Health Sciences and
Engineering. 10: 17-26.
Liestianty, D. & Nurhasanah. 2011. Penapisan
dan Isolasi Bacillus Penghasil Amilase
Dari Limbah Sagu (Metroxylon sagu
Rottb). Jurnal Biologi Indonesia. 7 (2):317
-327.
Madan, M, P. Vasudevan, & S. Sharma. 1987.
Cultivation of Pleurotus sajor-caju on
different wastes. Biological Waste. 22: 241
-250.
Mandel QA, AA. Al-Laith & SA Mohamed. 2005.
Cultivation of Oyster Mushrooms (Pleurotus
spp.) on Various Lignocellulosic Wastes.
World Journal of Microbiology & Biotech-
nology. 21:601–607.
Moonmoon, M., MN. Uddin, S. Ahmed, NJ.
Shelly, & MA. Khan. 2010. Cultivation of
different strains of king oyster mushroom
(Pleurotus eryngii) on saw dust and rice
straw in Bangladesh. Saudi Journal of
Biological Sciences. King Saud University.
17, 341-345.
Murugesan, AG., GS. Vijayalakshmi, N. Sukumaran,
& Mariappan. 1995. Utilization of water
hyacinth for oyster mushroom cultivation.
Bioresource Technology. 51: 97-98.
Parlindungan, A.K. 2001. Karakteristik
Pertumbuhan dan Produksi Jamur Kuping
Merah (Auricularia yudae) pada Baglog
Alang-alang. Jurnal Natur Indonesia. 3(2):
113-120.
Royse, DJ., TW. Rhodes, S. Ohga, & JE. Sanchez.
2004. Yield, mushroom size and time to
production of Pleurotus cornucopiae
(oyster mushroom) grown on switch grass
substrate spawned and supplemented at
various rates. Bioresource Technology. 91:
85-91.
Sanchez, M. 2010. Cultivation of Pleurotus
ostreatus and Other Edible Mushrooms.
Appl Microbiol Biotechnol. 85:1321-1337.
Saskiawan, I & Sudarmono. 1993. Alternatif
penggunaan sekam padi pada budidaya
jamur tiram (Pleurotus). Prosiding Seminar
Hasil Litbang SDH. Puslitbang Biologi
LIPI Bogor.
Satyanarayana T, NJ. Bhavdish, & P. Anil. 2012.
Microorganism in Sustainable Agriculture
and Biotechnology. Springer.
Siddhant & CS. Singh. 2009. Recycling of Spent
Oyster Mushroom Substrate to Recover
Additional Value. Kathmandu University
Journal of Science, Engineering and
Technology. 5 (1): 66-71.
Stofella, PJ. & AK. Brian. 2001. Compost Utilization
in Horticultural Cropping System. Lewis
Publisher.
Vetayasuporn S. 2004. Effective Microorganisms
for Enhancing Pleurotus ostreatus (Fr.)
Kummer Production. Journal of Biological
Science 4 (6): 706-710. Department of
Biotechnology, Faculty of Technology
Mahasarakham University. Thailand
Zhang R, X Li, & JG. Fade. 2002. Oyster
mushroom cultivation with rice and wheat
straw. Bioresources Technology. 82: 277-
284.

PANDUAN PENULIS
Naskah dapat ditulis dalam bahasa Indonesia atau bahasa Inggris. Naskah disusun dengan urutan: JUDUL (bahasa Indonesia dan Inggris), NAMA PENULIS (yang disertai dengan alamat Lembaga/Instansi), ABSTRAK (bahasa Inggris, dan Indonesia maksimal 250 kata), KATA KUNCI (maksimal 6 kata), PENDAHULUAN, BAHAN DAN CARA KERJA, HASIL, PEMBAHASAN, UCAPAN TERIMA KASIH (jika diperlukan) dan DAFTAR PUSTAKA. Penulisan Tabel dan Gambar ditulis di lembar terpisah dari teks.
Naskah diketik dengan spasi ganda pada kertas HVS A4 maksimum 15 halaman termasuk gambar, foto, dan tabel disertai CD. Batas dari tepi kiri 3 cm, kanan, atas, dan bawah masing-masing 2,5 cm dengan program pengolah kata Microsoft Word dan tipe huruf Times New Roman berukuran 12 point. Setiap halaman diberi nomor halaman secara berurutan. Gambar dalam bentuk grafik/diagram harus asli (bukan fotokopi) dan foto (dicetak di kertas licin atau di scan). Gambar dan Tabel di tulis dan ditempatkan di halaman terpisah di akhir naskah. Penulisan simbol a, b, c, dan lain-lain dimasukkan melalui fasilitas insert, tanpa mengubah jenis huruf. Kata dalam bahasa asing dicetak miring. Naskah dikirimkan ke alamat Redaksi sebanyak 3 eksemplar (2 eksemplar tanpa nama dan lembaga penulis).
Penggunaan nama suatu tumbuhan atau hewan dalam bahasa Indonesia/Daerah harus diikuti nama ilmiahnya (cetak miring) beserta Authornya pada pengungkapan pertama kali.
Pustaka didalam teks ditulis secara abjad.
Contoh penulisan Daftar Pustaka sebagai berikut :
Jurnal : Achmadi, AS., JA. Esselstyn, KC. Rowe, I. Maryanto & MT. Abdullah. 2013. Phylogeny, divesity , and
biogeography of Southeast Asian Spiny rats (Maxomys). Journal of mammalogy 94 (6):1412-123. Buku : Chaplin, MF. & C. Bucke. 1990. Enzyme Technology. Cambridge University Press. Cambridge. Bab dalam Buku : Gerhart, P. & SW. Drew. 1994. Liquid culture. Dalam : Gerhart, P., R.G.E. Murray, W.A. Wood, & N.R.
Krieg (eds.). Methods for General and Molecular Bacteriology. ASM., Washington. 248-277. Abstrak : Suryajaya, D. 1982. Perkembangan tanaman polong-polongan utama di Indonesia. Abstrak Pertemuan
Ilmiah Mikrobiologi. Jakarta . 15 –18 Oktober 1982. 42. Prosiding : Mubarik, NR., A. Suwanto, & MT. Suhartono. 2000. Isolasi dan karakterisasi protease ekstrasellular dari
bakteri isolat termofilik ekstrim. Prosiding Seminar nasional Industri Enzim dan Bioteknologi II. Jakarta, 15-16 Februari 2000. 151-158.
Skripsi, Tesis, Disertasi : Kemala, S. 1987. Pola Pertanian, Industri Perdagangan Kelapa dan Kelapa Sawit di Indonesia.[Disertasi].
Bogor : Institut Pertanian Bogor. Informasi dari Internet : Schulze, H. 1999. Detection and Identification of Lories and Pottos in The Wild; Information for surveys/
Estimated of population density. http//www.species.net/primates/loris/lorCp.1.html.






























