Embed Size (px)
Citation preview

KARAKTERISTIK ANTI BAKTERI ISOLAT Lactobacillus
DARI TEMPOYAK
(Skripsi)
Oleh :
Pratika Viogenta
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS LAMPUNG
BANDAR LAMPUNG
2010

ABSTRAK
KARAKTERISTIK ANTIBAKTERI
ISOLAT Lactobacillus DARI TEMPOYAK
Oleh
PRATIKA VIOGENTA
Antimikroba merupakan senyawa yang dikeluarkan oleh mikroorganisrne dan
dapat menghambat atau membunuh mikroorganisme lain. Lactobacillus
merupakan salah satu genus bakteri yang mampu menghasilkan senyawa
antibakteri seperti asam organik, hidrogen peroksida (H2O2), karbon dioksida
(CO2), diacetyl dan bakteriosin. Terdapat empat jenis isolat Lactobacillus dari
tempoyak yaitu L1, L2, L3 dan L4. Keempat isolat Lactobacillus tersebut mampu
menghambat pertumbuhan Escherichia coli. Namun, karakteristik antibakteri
yang dihasilkan oleh ketiga isolat belum diketahui.
Tujuan dari penelitian ini adalah mendeteksi senyawa antibakteri yang dihasilkan
isolat Lactobacillus L1, Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4
dengan cara mengukur total asam dan berat molekul bakteriosin isolat
Lactobacillus L1, Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4.
Penelitian ini akan dilaksanakan di Laboratorium Mikrobiologi dan Laboratorium
Biomolekuler FMIPA Unila. Penentuan karakterisasi jenis antibakteri dilakukan
dua tahapan yaitu penentuan asam organik dan protein sebagai senyawa
antibakteri. Asam organik ditetapkan dengan mengukur pH media kultur dan
total asam melalui titrasi menggunakan 0,1 N NaOH. Karakterisasi protein
antibakteri dilakukan secara observasi. Karakterisasi protein antibakteri
ditentukan dengan menetralkan supernatant kemudian diuji dengan metode difusi
sumuran terhadap bakteri uji yaitu Esherchia.coli, Salmonella paratyphii ,

Bacillus substilis dan Staphilococcus aureus. Terbentuknya zona jernih
menunjukkan zat antibakteri berupa protein, dilanjutkan dengan menetukan berat
molekul protein yang dapat menghambat pertumbuhan bakteri uji dengan
menggunakan teknik Sodium Dodecyl Sulfat Polyakrilamide Gel Elektoforesis
(SDS-PAGE).
Hasil penelitian menunjukkan bahwa antibakteri yang dihasilkan dari isolat
Lactobacillus L1, Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4
berupa senyawa asam organik. Isolat Lactobacillus L1, L2, L3 dan L4
menghasilkan asam organik maksimum berturut-turut yaitu sebesar 37,15 untuk
Lactobacillus L1, 36,13 untuk Lactobacillus L2, 29,94 untuk Lactobacillus L3,
dan 7,89 untuk Lactobacillus L4. Selama fermentasi berlangsung, total asam
organik yang diproduksi oleh keempat isolat Lactobacillus meningkat hingga hari
ke 5 waktu produksi. Dari hasil SDS-Page diperoleh pada isolat L1, L2 dan L3
tidak terdapat pita protein yang terbentuk, sedangkan isolat Lactobacillus L4
menghasilkan protein dengan berat molekul 15 kDa, 24 kDa, 53 kDa dan 169 kDa
akan tetapi bukan sebagai antibakteri
Key Word : Lactobacillus, Asam organik, Bakteriosin,Total Asam, SDS-PAGE

KARAKTERISTIK ANTI BAKTERI ISOLAT Lactobacillus
DARI TEMPOYAK
Oleh
Pratika Viogenta
Skripsi
Sebagai Salah Satu Syarat untuk Mencapai Gelar
SARJANA SAINS
Pada
Jurusan Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS LAMPUNG
BANDAR LAMPUNG
2010

Judul Skripsi : KARAKTERISTIK ANTI BAKTERI ISOLAT
Lactobacillus DARI TEMPOYAK
Nama Mahasiswa : Pratika Viogenta
Nomor Pokok Mahasiswa : 0617021058
Jurusan : Biologi
Fakultas : Matematika dan Ilmu Pengetahuan Alam
MENYETUJUI
1. Komisi Pembimbing
Dr. Sumardi, M.Si Dra. Christina.N Ekowati, M.Si
NIP 196503251991031003 NIP. 195808181985032001
2. Ketua Jurusan Biologi
Dra. Nuning Nurcahyani, M. Sc
NIP. 196603051991032001

MENGESAHKAN
1. Tim Penguji
Ketua : Dr. Sumardi, M. Si ...............
Sekretaris : Dra. Christina N. Ekowati, M. Si ...............
Penguji
Bukan Pembimbing : Kusuma Handayani, M. Si ...............
2. Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Dr. Sutyarso, M. Biomed.
NIP. 195704241987031001
Tanggal Lulus Ujian Skripsi : 18 November 2010

DAFTAR RIWAYAT HIDUP
Penulis dilahirkan di Kota Bandar Lampung pada tanggal 24
Maret 1989, dari pasangan Sanyoto dan Inah Lestari, S.Pd,
sebagai anak kedua dari dua bersaudara. Penulis menyelesaikan
pendidikan Taman Kanak-kanak di TK Al-Azahar Perumnas Way Halim pada
tahun 1994, pendidikan dasar diselesaikan pada tahun 2000 di Sekolah Dasar Al-
Azahar Perumnas Way Halim, Sekolah Lanjutan Tingkat Pertama diselesaikan di
SLTP Negeri 19 Bandar Lampung pada tahun 2003, dan Sekolah Menengah Atas
di SMA Negeri 2 Bandar Lampung pada tahun 2006. Penulis tercatat sebagai
mahasiswa di Jurusan Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam
Universitas Lampung melalui jalur SPMB ( Seleksi Penerimaan Mahasiswa Baru)
pada tahun 2006.
Selama menjadi mahasiswa, penulis berkesempatan mengikuti berbagai kegiatan
keorganisasian di UKM. Rohani Islam tahun 2007-2009 sebagai anggota Biro
Akademik, di HIMBIO sebagai anggota Bidang Hubungan Masayarakat pada
tahun 2007-2008 dan sebagai seketaris Bidang Keilmuan pada tahun 2008-2009,
dan di BEM FMIPA sebagai seketaris Dinas Lingkungan Hidup pada tahun 2009.
Penulis juga pernah menjadi asisten dosen pada mata kuliah Fisiologi Tumbuhan,
Genetika, Mikrobiologi Pangan dan Industri, Mikrobiologi Umum, Mikrobiologi
Lingkungan, Fisiologi Mikrobiologi jurusan biologi FMIPA, Mikrobiologi
STIKES UMITRA, Mikrobiologi Umum guru-guru SMP tahun 2010, dan

Pengenalan SAINS kepada guru-guru SD tahun 2010. Pada tahun 2009 penulis
melaksanakan kerja praktek di Loka Penelitian dan Pengembangan Pengendalian
Penyakit Bersumber Binatang (Litbang P2B2) Banjarnegara, Jawa Tengah.

“Jika niat sudah tertuju karena Allah, tidak akan ada
halangan yang bisa menghentikan seseorang melakukan sesuatu. Niat karena Allah ialah motivator yang utama dan
seharusnya menjadi satu-satunya motivator kita.”
“Allah SWT memerintahkan kita untuk mau berpikir tentang penciptaan-Nya yang begitu menakjubkan, rumit, dan
kompleks. Namun semua itu telah Allah SWT tundukan untuk kita. Ini sebagai tanda bahwa manusia memiliki
kemampuan (dari Allah) untuk menundukan apa yang ada di langit dan di bumi.”
“Allah SWT, tidak akan pernah menjanjikan hari-hari kita berlalu tanpa sakit, berhias tawa tanpa kesedihan, senang
tanpa kesulitan, tapi Allah SWT menjajikan kekuatan kepada kita untuk dapat melewatinya”

Dengan Menyebut Nama Allah yang Maha Pengasih Dan Maha Penyayang
Dengan segala Cinta dan Kasih sayang kupersembahkan karya sederhana ini teruntuk orang-
orang yang akan selalu berharga dalam hidupku :
Ibu dan Bapak tercinta dan dicintai Allah SWT Masku : Septian D.C
Almamater yang ku banggakan

SANWACANA
Puji syukur penulis panjatkan kehadiran Allah SWT, yang telah melimpahkan
rahmat, taufik serta hidayah-Nya sehingga penulis dapat menyelelesaikan skripsi
yang berjudul “KARAKTERISTIK ANTIBAKTERI ISOLAT Lactobacillus
DARI TEMPOYAK” dengan tepat waktu. Penelitian ini merupakan rangkaian
dari penelitian “ISOLASI DAN KARAKTERISASI Lactobacillus DARI
TEMPOYAK YANG BERPOTENSI SEBAGAI PENGAWET HAYATI
BAHAN PANGAN” yang didanai oleh Dirjen Dikti Kementrian Pendidikan
Nasional melalui Hibah Penguasaan Teknologi (Strategis Unila). Shalawat serta
salam semoga selalu tercurah kepada suri taudalan terbaik umat manusia Nabi
Muhamammad SAW.
Penyelesaian ini tidak terlepas dari bantuan berbagai pihak baik berupa bantuan
pemikiran ataupun bantuan moril sehingga pada kesempatan ini, penulis
menyampaikan terima kasih dengan setulus hati kepada.
1. Bapak Dr. Sumardi, M.Si., selaku pembimbing satu atas ide, saran-saran,
motivasi, kesabarannya, waktunya serta bimbingannya kepada penulis dalam
penyelesaian hasil penelitian ini.
2. Ibu Dra. Christina. N. Ekowati, M.Si., selaku pembimbing kedua atas
kesabarannya dalam membimbing, mengkoreksi, kasih sayangnya, dan selalu
memotivasi penulis.

3. Ibu Kusuma Handayani, M. Si selaku penguji yang telah memberikan saran-
saran, kritik serta koreksinya kepada penulis.
4. Bapak dan Ibu tercinta yang senantiasa memanjatkan doa, memberikan
dukungan moril dan materil pada penulis. Semoga Allah SWT, senantiasa
memberikan kemuliaan di dunia dan akhirat.
5. Ibu Dra. Rumyati, M.Si., selaku dosen pembimbing akademik atas
bimbingannya selama penulis menjadi mahasiswa di Jurusan Biologi FMIPA
Unila.
6. Bapak Dr. Sutyarso, M.Biomed., selaku Dekan FMIPA Unila.
7. Ibu Dra. Nuning Nurcahyani, M.Sc., selaku Ketua Jurusan Biologi FMIPA
yang telah memberikan kemudahan pada penulis.
8. Seluruh dosen dan staf karyawan FMIPA khususnya jurusan Biologi
9. Kakakku yang telah memberikan perhatian serta kasih sayang yang mengalir
tiada hentinya selama ini.
10. Teman seperjuanganku Kak Robi yang selalu membantu meski sudah lulus
terlebih dahulu.
11. Penghuni lab mikrobiologi (mbak wiwin, mbak nur, kak asep, deby, ros, ria,
mahendra) terima kasih atas bantuannya, perhatiannya, nasehatnya dan canda
tawa sehingga penulis tidak merasa jenuh.
12. Teman-temanku Dora, Nensi, Gina, Rima dan semua teman-teman angkatan
2006 yang tidak bisa disebutkan satu persatu atas kebersamaan, bantuannya
dan semua kenangan yang telah kita lalui bersama.
13. Adik-adikku penerus generasi mikrobiologi Ratna, Zahra, Dwi, Diah dan
Wiwik jangan mudah menyerah dan selalu semangat.

14. Pak Sungadi, Pak Tris, Pak Imron, Bu Endang dan seluruh saudaraku biologi
angkatan (’05-’09) serta semua pihak yang tidak bisa disebutkan satu persatu
terima kasih atas bantuan selama ini dan persahabatannya.
Semoga Allah SWT membalas segala bantuan yang telah diberikan kepada
penulis dan semoga penulisan skripsi ini dapat bermanfaat bagi yang membacanya
Bandar Lampung, November 2010
Pratika Viogenta

DAFTAR ISI
Halaman
DAFTAR TABEL .......................................................................................... i
DAFTAR GAMBAR ...................................................................................... ii
I. PENDAHULUAN ................................................................................... 1
A. Latar Belakang .................................................................................... 1
B. Tujuan Penelitian ................................................................................ 3
C. Kerangka Pemikiran ............................................................................ 3
D. Hipotesis ............................................................................................. 4
E. Manfaat Penelitian .............................................................................. 5
II. TINJAUAN PUSTAKA .......................................................................... 6
A. Lactobacillus ....................................................................................... 6
B. Antibakteri .......................................................................................... 7
C. Asam Organik ..................................................................................... 8
D. Hidrogen Peroksida ............................................................................. 11
E. Karbon Dioksida ................................................................................. 12
F. Bakteriosin .......................................................................................... 13
2.6.1. Karakteristik Bakteriosin .......................................................... 13
2.6.2. Penggolongan Bakteriosin ........................................................ 14
2.6.3. Mekanisme Kerja Bakteriosin ................................................... 18
2.6.4. Imunitas Bakteri Penghasil Bakteriosin .................................... 20
III. METODE PENELITIAN ....................................................................... 22
A. Tempat dan Waktu Penelitian ............................................................. 22
B. Alat dan Bahan .................................................................................... 22
C. Metode Penelitian ............................................................................... 23
D. Prosedur Kerja .................................................................................... 24
1. Peremajaan Bakteri ....................................................................... 24
2. Produksi Senyawa Antibakteri ...................................................... 24
3. Karakterisasi Antibakteri Berupa Asam Organik ......................... 25
4. Uji Daya Antibakteri ..................................................................... 25

5. Sodium Dodecyl Sulfat-Polyakrilamide Gel Elektoforesis
(SDS-PAGE) dan Identifikasi Aktivitas Pita Protein ................... 27
6. Diagram Alir ................................................................................. 30
IV. HASIL DAN PEMBAHASAN ............................................................... 31
A. Karakterisasi Antibakteri Senyawa Asam Organik ............................ 31
B. Karakterisasi Antibakteri Senyawa Protein ........................................ 37
V. KESIMPULAN DAN SARAN ............................................................... 42
A. Kesimpulan ......................................................................................... 42
B. Saran ................................................................................................... 43
DAFTAR PUSTAKA ..................................................................................... 44
LAMPIRAN

DAFTAR TABEL
Tabel Halaman
1. Klasifikasi Bakteriosin .............................................................................. 16
2. Penggolongan Bakteriosin Berdasarkan Kandungan Sistein .................... 17
3. Komposisi Gel Pemisah dan Gel Penahan ................................................ 27
4. Analisis Ragam Total Asam yang Dihasilkan Isolat Lactobacillus L1,
L2, L3 Dan L4 Terhadap Waktu Produksi (Hari) ..................................... 33
5. Zona Hambat dari Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3
dan L4 Sebelum dan Sesudah Dinetralkan Terhadap Bakteri Uji ........ 38
6. Hasil Pengukuran pH Media Kultur Isolat Lactobacillus ......................... 48
7. Hasil Pengukuran Total Asam Media Kultur Isolat Lactobacillus ........... 48
8. Penetuan Kurva Logaritma Berat Molekul Protein Standart Terhadap
Mobilitas Relatif (rf) ................................................................................. 54
9. Penetuan Berat Molekul Protein Terhadap Mobilitas Relatif (Rf) Pada
Isolat Lactobacillus L4

DAFTAR GAMBAR
Gambar Halaman
10. Model Pembentukan Pori pada Membran, Barrel-Stave dan Carpet
Model ....................................................................................................... 20
11. Model Mekanisme Kerja Protein Imunitas ............................................... 21
12. Bentuk Sumur Dilihat dari Bagian Bawah Cawan Petri ........................... 27
13. Pola Perubahan pH Media Isolat Lactobacillus L1, L2, L3 dan L4 yang
Diamati Tiap Hari Sampai Hari Ke 5........................................................ 31
14. Total Asam Organik Isolat Lactobacillus L1, L2, L3 dan L4 yang
Diamati Tiap Hari Sampai Hari Ke 5........................................................ 32
15. Uji Polinomial Orthogonal Isolat Lactobacillus L1 .................................. 34
16. Uji Polinomial Orthogonal Isolat Lactobacillus L2 .................................. 34
17. Uji Polinomial Orthogonal Isolat Lactobacillus L3 .................................. 35
18. Uji Polinomial Orthogonal Isolat Lactobacillus L4 .................................. 35
19. SDS-PAGE Isolat Lactobacillus L1, Lactobacillus L2, Lactobacillus
L3 dan Lactobacillus L4 ........................................................................... 40
20. Gel SDS-PAGE Ektrak Antibakteri L4 yang Diuji Terhadap St.Aureus .. 40
21. Isolat Lactobacillus L1 ............................................................................. 46
22. Isolat Lactobacillus L2 ............................................................................. 46
23. Isolat Lactobacillus L3 ............................................................................. 47
24. Isolat Lactobacillus L4 ............................................................................. 47

25. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sebelum Dinetralkan Pada Bakteri Uji Staphylococcus aureus ............... 48
26. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sesudah Dinetralkan Pada Bakteri Uji Staphylococcus aureus ................ 48
27. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sebelum Dinetralkan Pada Bakteri Uji Bacillus substilis ......................... 48
28. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sesudah Dinetralkan Pada Bakteri Uji Bacillus substilis.......................... 49
29. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sebelum Dinetralkan Pada Bakteri Uji Escherchia coli ........................... 49
30. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sesudah Dinetralkan Pada Bakteri Uji Escherchia coli ............................ 49
31. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sebelum Dinetralkan Pada Bakteri Uji Salmonella paratyphii ................ 50
32. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan L4
Sesudah Dinetralkan Pada Bakteri Uji Salmonella paratyphii ................. 50
33. Grafik Logaritma Berat Molekul Protein Standart Terhadap Mobilitas
Relatif ....................................................................................................... 52

I. PENDAHULUAN
A. Latar Belakang
Lactobacillus merupakan salah satu mikroorganisme yang aman jika
ditambahkan dalam bahan pangan karena sifatnya tidak tosik dan tidak
menghasilkan toksik. Bahkan, Lactobacillus bermanfaat bagi kesehatan dan
meningkatkan keamanan bahan pangan melalui penghambatan secara alami
terhadap pertumbuhan mikroorganisme berbahaya yang menyebabkan
pembusukan pada makanan maupun menyebabkan penyakit.
Lactobacillus berfungsi sebagai pengawet makanan karena mampu
memproduksi senyawa antibakteri seperti asam organik, hidrogen
peroksida (H2O2), karbon dioksida (CO2), diacetyl dan bakteriosin
(Kusmiati dan Malik, 2002).
Lactobacillus ditemukan pada produk-produk makanan fermantasi seperti
thongcai, lemma, tempoyak, ikan fermentasi, tauco, rebung asin dan sawi asin
(Wulandari, 2005). Tempoyak merupakan salah satu jenis makanan
tradisional yang melibatkan bakteri Lactobacillus dalam prosesnya.
Tempoyak berasal dari daging buah durian yang diolah dengan cara
fermentasi secara spontan dengan menambahkan garam 6-16 % dalam wadah
tertutup. Tempoyak dikenal di Indonesia, terutama di Sumatera dan
Kalimatan serta Malaysia (Yuliana dan Garcia, 2010). Isolat yang ditemukan

di dalam tempoyak antara lain Lactobacillus plantarum, L. brevis, L. mali, L.
fermentum L.casei, dan L. corynebacterium. Jenis bakteri asam laktat lain
yang juga ditemukan pada tempoyak yaitu Leuconostoc mesenteroides,
Pediacoccus acidilactici dan Weisella mesenteroides ( Yuliana dan Garcia,
2009).
Ekowati (2000) melaporkan bahwa dua jenis isolat Lactobacillus yang
diperoleh dari tempoyak mampu menghambat pertumbuhan Escherchia coli
dan Streptococcus haemoliticus. Konsentrasi filtrat kedua isolat
Lactobacillus tersebut mulai dari 40 % (v/v) efektif menurunkan jumlah sel
bakteri E. coli dan S. haemoliticus. Berdasarkan hasil analisis asam organik,
isolat Lactobacillus memproduksi asam laktat 0,4257 % (v/v), asam asetat
0,0717 % (v/v), asam propionat 0,0502 % (v/v), dan asam butirat 0,01 %
(v/v).
Ekowati (2009) memperoleh tiga jenis isolat Lactobacillus dari tempoyak
yaitu L1, L2 dan L3. Ketiga isolat Lactobacillus tersebut mampu
menghambat pertumbuhan Escherichia coli. Daya hambat zat antibakteri dari
ketiga isolat terhadap pertumbuhan Escherichia coli relatif sama, yang
terlihat dari diameter zona hambat berkisar 1,65 cm-2,2 cm. Hal ini
menunjukkan bahwa ketiga isolat tersebut menghasilkan suatu senyawa
antibakteri tertentu. Menurut Ogunbawo et all (2003), Lactobacillus
plantarum ST194BZ menghasilkan senyawa antibakteri dalam bentuk asam
organik dan protein. Jenis senyawa antibakteri yang dihasilkan oleh ketiga
isolat belum diketahui. Berdasarkan permasalahan tersebut maka perlu

dilakukan penelitian mengenai karakteristik antibakteri yang dihasilkan oleh
isolat Lactobacillus dari tempoyak.
B. Tujuan Penelitian
Penelitian ini bertujuan untuk mengidentifikasi senyawa antibakteri yang
dihasilkan isolat Lactobacillus dari tempoyak.
C. Kerangka Pemikiran
Lactobacillus dapat diisolasi dari makanan yang telah mengalami fermentasi
seperti tempoyak. Pada proses terbentuknya tempoyak, penambahan garam
dapur sebesar 3-7 % pada daging buah durian akan menyebabkan
Lactobacillus tumbuh dengan baik. Hasil metabolit Lactobacillus pada
tempoyak terbentuk asam-asam organik dari karbohidrat yang berasal dari
daging buah durian. Beberapa asam organik dapat bersifat toksik terhadap
mikroorganisme patogen ataupun mikroorganisme pembusuk pada makanan.
Isolat Lactobacillus yang diperoleh dari tempoyak berpotensi menghasilkan
senyawa antibakteri yang mampu menghambat pertumbuhan Escherichia
coli. Daya hambat zat antibakteri dari ketiga isolat terhadap pertumbuhan
Escherichia coli relatif sama, yang terlihat dari diameter zona hambat
berkisar 1,65 cm – 2,2 cm.
Lactobacillus yang berasal dari tempoyak mampu memproduksi jenis asam-
asam organik, yaitu asam laktat 0,4257 % (v/v), asam asetat 0,0717 % (v/v),
asam propionat 0,0502 % (v/v), dan asam butirat 0,01 % (v/v). Senyawa-
senyawa tersebut dapat menghambat pertumbuhan dari bakteri patogen dan

bakteri pembusuk makanan sehingga dapat digunakan sebagai zat antibakteri.
Pada umumnya tidak hanya asam organik yang dihasilkan Lactobacillus
mampu menghambat mikroorganisme lain tetapi terdapat senyawa lain yang
ikut berperan di dalam penghambatan pertumbuhan bakteri pembusuk dan
patogen seperti H2O2, diasetil, CO2, dan bakteriosin. Dalam metabolisme
Lactobacillus, ada kemungkinan Lactobacillus yang berasal dari tempoyak
mengubah asam-asam organik menjadi asam amino. Asam – asam amino
tersebut dapat digunakan oleh Lactobacillus untuk menyusun bakteriosin
atau protein penghambat. Bakteriosin merupakan substansi protein yang
dikode oleh plasmid, umumnya mempunyai berat molekul kecil serta
memiliki aktivitas menghambat pertumbuhan bakteri lain (bakterisidal),
terutama yang memiliki kekerabatan erat secara filogenik.
Bakteriosin dihasilkan oleh bakteri Lactobacillus antara fase logaritmik dan
awal fase stasioner. Faktor lingkungan sangat mempengaruhi pertumbuhan
bakteri Lactobacillus maupun hasil metabolitnya. Penambahan substrat
seperti ekstrak yeast (3%), NaCl (1.0-2.0%), glukosa (1.0%) dan Tween 80
(0.5%) menghasilkan bakteriosin dalam jumlah optimal. Substrat-substrat
tersebut digunakan Lactobacillus sebagai prekursor senyawa antibakteri.
Aktivitas maksimal bakteriosin dicapai pada awal pH 5.5 dan diinkubasi
antara 48 jam hingga 60 jam pada suhu 30 – 37 oC.
D. Hipotesis
Hipotesis yang diajukan yaitu isolat Lactobacillus dari tempoyak dapat
menghasilkan antibakteri berupa senyawa asam organik dan protein.

E. Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan informasi mengenai jenis
senyawa antibakteri yang dihasilkan oleh isolat Lactobacillus dari tempoyak
dalam menghambat pertumbuhan bakteri lain. Selanjutnya informasi ini
diharapkan dapat bermanfaat sebagai biopreservatif terhadap bahan pangan.

II. TINJAUAN PUSTAKA
A. Lactobacillus
Lactobacillus merupakan bakteri gram positif, tidak berspora, berbentuk
batang atau coccosbacilli. Pada umumnya mengandung guanin-citosin di
dalam DNA kurang dari 50 mol%. Lactobacillus dapat melakukan
fermentasi, bersifat aero-toleran atau anaerobik, aciduric atau acidophilic dan
memerlukan asupan nutrisi yang kompleks seperti karbohidrat, asam amino,
peptida, ester asam lemak, garam, derivat asam nukleat dan vitamin.
Lactobacillus tidak mensintesis porphyrinoids begitu juga dengan aktivitas
yang terkait dengan porphyrinoids. Namun, terdapat beberapa strain dari
Lactobacillus dapat menggunakan porphorinoid dari lingkungan dan
memperlihatkan aktivitas katalase, reduksi nitrit bahkan sitrokrom. Strain Lb.
mali mampu membentuk pseudocatalase (Wood dan Holzapfel, 1995).
Lactobacillus mampu merombak senyawa kompleks menjadi senyawa yang
lebih sederhana dengan hasil akhirnya yaitu asam laktat (Rostini, 2007).
Glukosa sebagai sumber karbon dimetabolisme membentuk 85 % asam laktat
oleh Lactobacillus yang bersifat homofermentatif atau menghasilkan asam
laktat, CO2, etanol oleh Lactobacillus yang bersifat heterofermentatif (Nur,
2005).

Apabila jumlah oksigen atau senyawa oksidan lain meningkat, sejumlah asam
asetat akan terbentuk dari asam laktat atau etanol melalui reaksi asetat kinase.
Demikian variasi metabolisme Lactobacillus membentuk metabolit. Berbagai
hasil metabolit seperti sitrat, malat, tartar, quinolat, nitrat, dan nitrit dapat
juga di metabolisme kembali dan digunakan sebagai sumber energi atau
penerima elektron terakhir (Wood dan Holzapfel, 1995)..
B. Antibakteri
Antibakteri adalah senyawa yang dihasilkan oleh mikroorganisme hidup yang
mampu menghambat pertumbuhan mikroorganisme lain terutama bakteri,
bahkan dapat membunuhnya (Irianto, 2007). Suatu antibakteri yang ideal
memiliki toksisitas selektif, berarti obat antibakteri tersebut hanya berbahaya
bagi bakteri, tetapi relatif tidak membahayakan bagi hospes. Berdasarkan
sifat toksisitas selektif. Ada bakteri yang bersifat menghambat pertumbuhan
bakteri (bakteriostatik) dan ada yang bersifat membunuh bakteri (bakterisida).
Berdasarkan mekanisme kerjanya, antibakteri dibagi menjadi 5 kelompok,
yaitu:
1) Merusak dinding sel yaitu dengan menghambat pembentukan dan
mengubahnya setelah selesai terbentuk.
2) Mengganggu permeabilitas sel yaitu dengan merusak membran sel. Fungsi
membran sel adalah mempertahankan bahan-bahan dalam sel serta
mengatur aliran keluar masuknya bahan lain. Adanya kerusakan pada
membran ini mengakibatkan terhambatnya pertumbuhan sel atau matinya.

3) Merubah molekul protein dan asam nukleat yaitu dengan
mendenaturasikan protein dan asam nukleat sehingga kerusakan sel tidak
dapat diperbaiki lagi karena hidup suatu sel tergantung pada molekul
protein dan asam nukleat dalam keadaan alamiah.
4) Menghambat kerja enzim dengan mengganggu reaksi biokimiawi.
Penghambatan ini dengan mengakibatkan terganggunya metabolisme sel.
5) Menghambat sintesis asam nukleat dan protein. Gangguan pada
pembentukan atau fungsi-fungsi DNA, RNA dan protein dapat
mengakibatkan kerusakan total pada sel, karena zat-zat tersebut
memegang peranan penting dalam proses kehidupan normal sel
(Mulyanti, 2009).
Bakteri Lactobacillus berpotensi menghasilkan antibakteri yang mampu
menghambat pertumbuhan mikroorganisme berbahaya yang
menyebabkan pembusukan pada makanan maupun menyebabkan
penyakit melalui produk metabolik seperti asam organik, hidrogen
peroksida (H2O2), karbon dioksida (CO2), diacetyl dan bakteriosin
(Ogunbanwo et all, 2003).
C. Asam Organik
Ketika proses fermentasi glukosa, Lactobacillus mampu memproduksi asam
laktat secara homofermentasi atau sebanding dengan jumlah asam laktat,
asam asetat, etanol dan karbondioksida secara heterofermentatif. Apabila
proses tersebut diamati selama seminggu, asam organik yang dihasilkan
Lactobacillus memiliki suatu aktivitas antimikroba yang lebih besar pada pH

rendah dibandingakan dengan pH netral. Mengenai kedua asam yang
terbentuk, asam asetat memiliki aktivitas penghambat yang paling kuat dan
mempunyai suatu cakupan lebih luas dalam hal aktivitas menghambat
pertumbuhan khamir, kapang dan bakteri sedangkan asam propionat
memiliki efek antimikroba yang kuat khususnya untuk khamir dan kapang
(Salminen et all, 2004).
Aktivitas antimikroba asam asetat dan asam propionat lebih kuat
dibandingkan dengan aktivitas antimikroba dari asam laktat. Hal ini dapat
dijelaskan dari tingginya nilai pKa asam asetat ( pKa = 4,87) dan asam
propionat (pKa = 4, 75) dibandingkan dengan nilai pKa asam laktat (pKa =
3,08). Sebagai contoh, pada pH 4, hanya 11 % dari asam laktat yang tidak
terdisosiasi sedangkan 85 % asam asetat dan 92 % asam propionat tidak
terdisosiasi. Namun, ketika terbentuk campuran asam-asam organik, terjadi
kerja sama dalam meningkatkan aktivitas menghambat. Sebagian besar asam
laktat menyebabkan pH menurun disaat asam asetat dan asam propionat yang
tidak terdisosiasi menjadi agen antimikroba. Gabungan dari asam laktat dan
asam asetat mampu mengurangi laju pertumbuhan Salmonella enetric ser var
Typhimurium lebih baik daripada aktivitas masing-masing dari asam laktat
atau asam asetat sendiri (Salminen et all, 2004).
Asam laktat selain menurunkan pH juga dapat menggangu permeabilitas
membran, dengan demikian dapat meningkatkan aktivitas dari substansi
antimikroba lainnya. Mekanisme antimikroba asam laktat berdasarkan teori
"chemiosmotic" dan pH homeostasis. Ketika asam laktat yang diproduksi

disekresikan ke lingkungan, beberapa molekul terdisosiasi menjadi H+ dan
anion, sementara yang lain tidak terdisosiasi. Salah satu faktor yang
berperanan terhadap terdisosiasi atau tidaknya suatu molekul adalah pH
lingkungan dan pK (tetapan keseimbangan). Hal ini menyebabkan
peningkatan proton transmembran yang pada akhirnya menyebabkan gradient
proton. Perbedaan ini menyebabkan proton lebih cepat masuk ke dalam sel
sehingga meningkatkan kebutuhan energi untuk mempertahankan pH alkali
dalam sel. (Lunggani, 2007).
Banyak orang mengasumsikan bahwa molekul asam lemah yang tidak
terdisosiasi menjadi racun, walaupun asam yang terdisosiasi telah banyak
yang mengamati juga mampu menghambat pertumbuhan mikroba. Hal ini
dapat dijelaskan bahwa asam organik yang tidak terdisosiasi (netral) dapat
berdifusi melewati membran sel karena asam organik yang tidak terdisosiasi
larut dalam lipid. Setelah memasuki sel, asam akan terdisosiasi pada pH
sitoplasma yang biasanya mendekati netral (Salminen et all, 2004).
Banyak peneliti menyatakan bahwa pelepasan proton ke dalam sitoplasma
berperan penting di dalam proses pengasaman dan menghilangnya perbedaan
pH yang berlebih pada membran sehingga menyebabkan pertumbuhan
mikroba terhambat. Namun, beberapa peneliti lainnya menyatakan bahwa
hipotesis ini harus ditinjau kembali karena melihat bahwa proton tidak dapat
ditranslokasi. Menurut mereka, akumulasi anion yang menjadi faktor utama
menyebabkan pertumbuhan mikroba terhambat jika dilihat dari jumlah anion
berkurang pada sintesis makromolekul dan anion mempengaruhi sistem

transport pada membran sel. Seperti bakteri lainnya, bakteri asam laktat
memberikan efek penetralan dari akumulasi anion dengan cara mengurangi
pH sitoplasma mikroba patogen dan mikroba pembusuk (Salminen et all,
2004).
D. Hidrogen Peroksida
Dalam kondisi adanya oksigen, bakteri asam laktat menghasilkan hydrogen
peroksida melalui oksidasi molekul yang mengandung flavoprotein, oksidasi
NADH, dan superoxide dismutase. Bakteri asam laktat tidak menghasilkan
katalase yang berfungsi mungurai hydrogen peroksida. Pada sistem lainnya
bahwa penguraian hydrogen peroksida tidak seaktif dibandingkan dengan
produksi hydrogen peroksida itu sendiri sehingga terjadi akumulasi hydrogen
peroksida. Hydrogen peroksida tidak akan terakumulasi sebab hydrogen
proksida diuraikan oleh peroksidase, flavoprotein, dan pseudcatalase.
Pengaruh bakterisidal dari hydrogen peroksida dihubungkan dengan efek
oksidasi yang kuat di dalam sel bakteri seperti kelompok sulfidril dari protein
sel dan lipid membran dapat dioksidasi. Untuk menghasilkan hidrogen
peroksida dibutuhkan oksigen sehingga menyebabkan lingkungan menjadi
anerobik. Hal ini tidak baik untuk organisme yang bersifat aerobik (Salminen
et all, 2004).
Di dalam kondisi normal, pengaruh antimikroba dari hidrogen proksida
kemungkinan ditingkatkan karena adanya lactoperoksidase dan thiocyanate
(SCN-).
SCN- +H2O2
lactoperoxidase OSCN
- + H2O

OSCN- menyebabkan kerusakan struktural dan perubahan pada membran sel
bakteri. Namun yang menjadi faktor utama hidrogen peroksida menjadi
senyawa antimikroba yaitu menghambat proses glikolisis. Hidrogen
peroksida menghambat pengangkutan glukosa, aktivitas heksokinase, dan
aktivitas glyceraldehyde-3-phosphat dehidrogenase dengan cara
mengoksidasi sulfhydryl yang terdapat didalam enzim tersebut (Salminen et
all, 2004).
E. Karbon Dioksida
Karbon dioksida dihasilkan selama proses fermentasi glukosa dan respirasi
berlangsung. Karbon dioksida memiliki pengaruh antimikroba ganda.
Karbon dioksida dapat menyebabkan lingkungan menjadi anaerob dan karbon
dioksida itu sendiri bersifat antimikroba. Mekanisme penghambatan dari
karbon dioksida belum banyak diketahui tetapi karbon dioksida mampu
menghambat aktivitas enzim dehidrogenase dan terjadinya akumulasi karbon
dioksida di dalam lipid bilayer menyebabkan tidak berfungsinya
permeabilitas membran. Pada konsentrasi karbon dioksida dalam keadaan
rendah dapat merangsang pertumbuhan beberapa organisme sedangkan pada
konsentrasi yang lebih tinggi dapat menghambat pertumbuhan (Salminen et
all, 2004).

F. Bakteriosin
2.6.1 Karakteristik Bakteriosin
Bakteriosin merupakan antimikrobia yang berupa protein dan disintesis
secara ribosomal (Suparjo, 2008). Bakteriosin memiliki pengaruh
bakterisidal dan bakteriostatik terhadap bakteri yang mempunyai
hubungan dekat dengan bakteri penghasilnya (Kusmiati dan Malik,
2002). Bakteri target memiliki sifat pengikatan spesifik (specific
binding site) (Usmiati,2009).
Bakteriosin biasanya tahan terhadap panas, dan aktivitasnya masih tetap
ada dalam lingkungan asam misalnya pada suhu 100˚C atau 121˚C
selama 15 menit (Ogunbawo et all, 2003), demikian pula suhu yang
sangat rendah dalam penyimpanan tidak mempengaruhi aktivitas
bakteriosin. Umumnya, bakteriosin memiliki sifat mudah didegradasi
enzim proteolitik seperti protease (Usmiati,2009).
Bakteriosin dihasilkan baik oleh bakteri gram positif maupun bakteri
gram negatif. Bakteriosin gram positif mengandung 30 sampai 60
asam amino dengan aktifitas yang bervariasi dari spektrum sempit
sampai luas delam melawan bakteri gram positif lainnya
(Suparjo,2009).
Bakteriosin disintesis selama fase eksponensial pertumbuhan sel
mengikuti pola sintesis protein. Sistem ini diatur oleh plasmid DNA.
Pada umumnya, bakteriosin non lantibiotik disintesis melalui jalur

ribosomal, sedangkan kelompok lantibiotik disintesis secara ribosomal
sebagai prepeptida kemudian mengalami modifikasi. Sekresi
prepeptida dilakukan pada fase eksponensial dan diproduksi secara
maksimal pada fase stasioner. Prinsip regulasi sintesis bakteriosin
diatur oleh adanya gen pengkode produksi dan pengkode immunitas.
Aktivitas produksi bakteriosin oleh bakteri asam laktat dipengaruhi oleh
faktor pH, suhu, sumber karbon, serta fase pertumbuhan. Jenis sumber
karbon maupun sumber nitrogen yang digunakan dalam medium
produksi mempengaruhi laju pertumbuhan sel bakteri asam laktat,
selanjutnya berpengaruh terhadap metabolisme produksi bakteriosin.
Selain itu, tingkat salinitas medium produksi seperti kandungan garam
dari media turut mempengaruhi metabolisme produksi bakteriosin.
Secara umum kondisi optimum produksi bakteriosin selain dipengaruhi
oleh fase pertumbuhan, pH media, suhu inkubasi, jenis sumber karbon
dan sumber nitrogen juga konsentrasi NaCl (Kim, 1990).
2.6.2 Penggolongan Bakteriosin
Bakteriosin berdasarkan sifat kimia, struktur dan fungsinya dibagi
menjadi 4 kelas yaitu
a. Kelas I : Lantibiotik, peptida molekul kecil dengan berat molekul
kurang 5 kDa mengandung lanthionine (Lan), β-metyl lanthionine
(MeLan), dehydroalanine dan dehydrobutyrine serta mengandung
19 sampai 50 asam amino. Kelas ini dibagi lagi menjadi menjadi 2
tipe berdasarkan struktur kimia dan aktifitas antimikroba, yaitu tipe

A dan tipe B. Tipe A memiliki bentuk ulir, bermuatan positif,
aktifitasnya berhubungan dengan pembentukan pori pada membran
sel. Tipe B memiliki bentuk globular bermuatan negatif atau netral,
aktifitas mikrobanya terkait dengan penghambatan enzim spesifik.
b. Kelas II : peptida yang stabil terhadap panas, berat molekul lebih
kecil dari 10 kDa, tidak memiliki asam amino lantionine dan tidak
terjadi perubahan asam amino. Kelas ini dibagi menjadi tiga
subkelas, yaitu bakterosin yang mempunyai efek antilisterial (IIa),
bakteriosin dengan dua peptida (IIb), dan bakteriosin yang
disekresikan melalui sec-dependent (IIc). Namun menurut van
Belkum dan Stiles dalam Supajo (2009) membagi kelas bakteriosin
ini menjadi enam subkelas antara lain IIa : cystibiotics dengan dua
ikatan disulfida yang dihasilkan dari empat asam amino sistein.
Contoh pediosin PA-1, pediosin AcH, Enterocin A dan Divercin
V41. IIb : Cyntibiotics dengan satu ikatan disulfida dari dua residu
sistein pada N-section peptida, contoh pada leucocin A. IIc :
cyntibiotics dengan satu ikatan disulfida pada N- dan C-section
peptida, contoh pada carnobacteriocin A dan enterocin B. IId :
peptida yang mengandung satu (thiolbiotics) atau tanpa sisten,
contoh pada lactococcin A dan B. IIe : bakteriosin yang memiliki
dua peptida, contoh pada thermophilin 13, lactacin F, plantaracin S,
plantaracin A, plantaracin EF, plantarain JK, lactococcin G dan
lactococcin M. Iif : bakteriosin khas, contoh entrocin 4.
c. Kelas III : protein labil terhadap panas dengan molekul lebih besar
dari 30 kDa
d. Kelas IV : suatu kelompok bacteriocin kompleks. Protein ini
mengandung lipid atau karbohidrat (glikoprotein dan lipoprotein),
Bakteriosin yang bersifat hidrofobik dan stabil terhadap panas.
(Alpay et all, 2003 ; Suparjo, 2008).
Tabel 1. Klasifikasi Bakteriosin
Kelompok Karakteristik Contoh Bakteriosin Bakteri Penghasil
I A Molekul kecil (2-5 kDa),
mengandung asam amino
lanthionine dan β-
methyllanthionine,
bermuatan positif,
berbentuk ulir
Nisin Lactococcus lactis
Pep 5 Stapylococcus epidermidis
Epidermin Stapylococcus epidermidis
Lactoccin S Lactobacillus sake
Gallidermin Stapylococcus gallinarum
Lacticin 481 Lactococcus lactis
B Molekul kecil (< 2 kDa),
bermuatan negatif atau
netral, berbentuk globular
Marsacidin Bacillus subtilis
Actagardin Actinoplasnes sp
Cinnamycin Streptomyces cinnamoneus
II a Peptida anti-listerial Pediosin PA-1/AcH Pediococcus acidilactici H /
PAC1.0
Sakacin A Lactobacillus sake LB706
Sakacin P Lactobacillus sake LTH 674
Leucocin A-
UAK187
Leuconostoc gelidium
UAL187
Carnobacteriocin B2 Carnobacterium piscicola
LV17B
Mesentrecin Y105 Leuconostoc mesenteroides
Lactococcin MMFII Lactococcus lactis
B Bakteriosin 2-peptida Lactococcin G Lactococcus lactis
Lactococcin M Lactococcus lactis
Lactacin F Lactobacillus johnsonii
Plantacirin A Lactobacillus plantarum
Plantacirin EF Lactobacillus plantarum
Plantacirin JK Lactobacillus plantarum
C Bakteriosin yang
disekresikan melalui sec-
dependent
Acidin B Lactobacillus acidophilus
Carnobacteriocin A Carnobacterium piscicola
LV17A
Divergicin A Arnobacterium divergens
LV13
Enterocin P Enterococcus faecum
Enterocin B Enterococcus faecum T136
Lactococcin A Lactococcus lactis LMG2130
Lactococcin B Lactococcus lactis WMA4
Acidocin B Lactobacillus acidophilus
M46
Cerein 7 / 8 Bacillus cereus Bc7
III Molekul besar (> 30 kDa)
sensitif terhadap panas
Helveticins J Lactobacillus helveticus
Helveticins V-1829 Lactobacillus helveticus
IV Mengandung protein dan
lipid
Lactococcin 27
Lacstrecins
(Suparjo,2008)

Bakteriosin berdasarkan kandungan sistein atau ikatan disulfida
dikelompokkan menjadi 3 kelompok yaitu
1. Cystibiotic; mengandung dua atau lebih asam amino sistein untuk
ikatan disulfida
2. Thiobiotic ; mengandung satu sistein
3. Tanpa sistein
Tabel 2. Penggolongan Bakteriosin Berdasarkan Kandungan Sistein
Bakterosin Berat Molekul
(kDa)
Asam
Amino
Bakteri Penghasil
Cystibiotics
Pediosin AcH/PA1
Leucocin A/UAL 187
Mesentericin Y 105
Sakacin A
Sakacin P
Lactacin F
Carnobacteriocin A
Carnobacteriocin BM1
Carnobacteriocin B2
Cerein 7/8
4,6
3,9
3,8
4,3
4,4
5,6
6,1
4,5
4,9
4,9
44
37
37
41
43
57
53
43
48
56
Pediococcus acidilactic H/PAC1.0
Leuconostoc gelidium UAL 187
Leuconostoc mesenteroides Y 105
Lactobacillus sake LB706
Lactobacillus sake LTH 674
Lactobacillus acidophilus 11088
Carnobacterium piscicola LV 17A
Carnobacterium piscicola LV17B
Carnobacterium piscicola LV17B
Bacillus cereus Bc7
Thiolbiotics
Lactococcin B 5,3 47 Lactococcus lactis subsp cremoris 9B4
Non Sistein
Lactococcin A
Lactococcin M
Lactococcin N
Lactococcin Gα
Lactococcin Gβ
5,8
4,3
4,4
4,3
4,1
54
48
47
39
65
Lactococcus lactis subsp cremoris 9B4,
Lac. lactis subsp cremoris LMG 2130
Lac. lactis subsp lactis bv diacetylactic
WM4
Lac. lactis subsp cremoris 9 B4
Lac. lactis subsp cremoris 9 44
Lac. lactis subsp lactis LMG 2081
Lac. lactis subsp lactis LMG 2081
(Suparjo,2008)

2.6.3 Mekanisme Kerja Bakteriosin
Usmiati (2009) menyebutkan bahwa target utama bakteriosin adalah
membran sitoplasma sel bakteri karena reaksi awal bakteriosin adalah
merusak permeabilitas membran dengan membentuk pori pada
membrane sel dan menghilangkan gaya gerak proton (proton motive
force (PMF)). Gaya gerak proton merupakan gradien elektrokimia
membran sitoplasma yang mengatur sintesis dan penimbunan ATP.
Kegagalan gaya gerak proton menyebabkan kematian sel melalui
penghentian semua reaksi yang membutuhkan energi, biosintsesis
protein atau asam nukleat (Suparjo, 2008).
Pembentukan pori pada membran sel menyebabkan destabilitas
membran sehingga dapat mengganggu kesetimbangan ADP/ATP
intraseluler akibat kebocoran pospat anorganik, mengurangi daya gerak
proton dan jumlah kation bivalensi ( Mg2+
atau Ca2+
) yang
menyebabkan penetralan muatan negatif fosfolipid dan memungkinkan
perembesan ion ( K+ dan Mg
2+ ), asam amino dan ATP (Suparjo, 2008).
Aktivitas penghambatan bakteriosin membutuhkan reseptor spesifik
permukaan sel. Lipid membran sitoplasma yang bermuatan negatif
merupakan reseptor utama bakteriosin dalam proses pembentukan pori.
Interaksi elektrostatik bakteriosin yang bermuatan positif yang bersifat
hidrofobik dengan gugus fosfat bermuatan negatif pada membran sel
target merupakan tahap awal pengikatan bakteriosin dengan membran
target. Bagian hidrofobik bakteriosin masuk ke dalam membran

membentuk pori. Konduktivitas dan stabilitas pori pada bakteriosin
lantibiotik ditingkatkan melaui pengikatan molekul (molecule docking)
sedangkan pada bakteriosi kelas II, reseptor membran target bekerja
terhadap spesifikasi tertentu (Suparjo, 2008).
Proses pembentukan pori pada membran fosfolipid oleh peptida
membran aktif umumnya terjadi melalui dua mekanisme yaitu metode
tong kayu (barrel-stave model) dan model baji atau karpet (wedge
model or carpet model) (Gambar 1). Pada model tong, peptida
menghadap hampir tegak lurus terhadap membran, kemudian masuk
dan membuat saluran ion sepanjang membran diikuti dengan
pengikatan monomer tambahan membentuk pori. Pada model karpet,
peptida berikatan dengan permukaan membran, jika konsentrasi
ambang batas monomer peptida tercapai, membran ditembus dan pori
sementara terbentuk (Zhao,2003).
Gambar 1. Model Pembentukan Pori pada Membran, Barrel-Stave dan
Carpet Model (Zhao, 2003).

2.6.4 Imunitas Bakteri Penghasil Bakteriosin
Salah satu perbedaan bakteriosin dengan antibiotik adalah adanya
mekanisme perlindungan bakteri penghasil terhadap kerja
bakteriosinnya. Perlindungan pada bekteriosin lantibiotic dapat
dimediasi melalui protein imunitas, LanI dan lanFEG. Terdapat dua
sistem yang bekerja secara sinergis untuk melindungi sel penghasil dari
bakteriosinnya sendiri. LanI, yang sebagian besar berikatan pada sisi
luar membran sitoplasma, memberikan imunitas dengan mencegah
pembentukan pori oleh bakteriosin. LanFEG bekerja melalui
pengangkutan molekul bakteriosin yang telah masuk ke dalam
membran kembali ke medium sekeliling dan menjaga konsentrasi dalam
membran di bawah tingkat kritis (Suparjo, 2008).
Protein imunitas bakteriosin non-lantibiotics disandikan oleh suatu gen
yang terdapat pada bagian hilir gen bakteriosin, kecuali gen imunitas
bakteriosin kelas IIc. Sistem imunitas bakteriosin sejauh ini belum
berhasil dijabarkan semuanya kecuali LciA, protein imunitas
Lactococcin A (Gambar 2). LciA dapat mencegah aksi Lactococcin A
dengan mengikat kemudian menetralisir bakteriosin atau dengan
berinteraksi dan merintangi reseptor bakteriosin. Melalui interaksi
Lactococcin A- reseptor dalam LciA menjangkau kedalam membran
sitoplasma. Ujung C protein imunitas berada di luar sel sedangkan
ujung N berada didalam sitoplasma. Dengan mengikat reseptor, LciA

mencegah lactococcin A masuk ke dalam membran tetapi ikatan
lactococcin A pada reseptor tetap terjadi (Suparjo, 2008).
Gambar 2. Model Mekanisme Kerja Protein Imunitas
(Suparjo, 2008)

III. METODE PENELITIAN
A. Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di Laboratorium Mikrobiologi dan Laboratorium
Molekuler Jurusan Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam
Universitas Lampung. Penelitian dilakukan pada bulan Januari 2010 hingga
Juli 2010.
B. Alat dan Bahan
Alat-alat yang digunakan dalam penelitian ini antara lain tabung reaksi,
cawan petri dengan diameter 15 cm dan 30 cm, erlenmeyer ukuran 500 mL,
250 mL, 100 mL dan 50 mL, gelas ukur, ose, spatula, bunsen, vortex mixer,
neraca analitik, kompor listrik, autoklaf, laminar air flow, inkubator dengan
suhu 37oC, inkubator shaker, piranti elektroforesis protein (Mini Protean
Tetra Cell Biorad, USA), mikrotube, tip, mikropipet, pH meter, syringe,
pipet tetes, sarung tangan dan peralatan lainnya.
Bahan-bahan yang digunakan adalah isolat Lactobacillus L1, Lactobacillus
L2, dan Lactobacillus L3 yang diperoleh dari penelitian sebelumnya serta
isolat Lactobacillus L4 yang diperoleh dari hasil isolasi terbaru dari tempoyak
dengan ciri-ciri bentuk batang, gram positif, katalase negatif dan motil,
biakan Escherchia coli, Staphylococcus aures, Bacillus substilis, dan

Salmonella paratyphii yang diperoleh dari koleksi biakan Laboratorium
Mikrobiologi FMIPA Unila, media deMan Rogosa and Sharpe (MRS) Broth,
media Nutrient Broth (NB), bacteriological agar, akuades, akubides,
alumunium foil, kasa dan kapas.
C. Metode Penelitian
Penentuan karakterisasi jenis antibakteri dilakukan dua tahapan yaitu
penentuan asam organik dan protein sebagai senyawa antibakteri. Untuk
mengkarakterisasi senyawa asam organik yaitu dengan mengukur pH media
kultur sampai hari kelima fermentasi. Terjadinya penurunan pH
menunjukkan terbentuknya asam, maka akan dilanjutkan dengan menghitung
total asam yang dihasilkan oleh isolat Lactobacillus melalui titrasi dengan
0,1 N NaOH. Perlakuan disusun dalam faktorial dengan rancangan acak
kelompok lengkap (RAKL) dengan 2 kali pengulangan. Faktor pertama
adalah jenis isolat bakteri Lactobacillus, yakni L1, L2, L3 dan L4. Faktor
kedua adalah lama produksi senyawa antibakteri dari isolat Lactobacillus,
yakni hari ke-1, ke-2, ke-3, ke-4 dan ke-5. Variabel yang diamati yaitu
perubahan pH dan total asam yang dihasilkan oleh keempat isolat
Lactobacillus. Data yang diperoleh diuji dengan analisis ragam. Adanya
perbedaan nyata (p<0.01), maka dilanjutkan dengan uji Polynomial
Ortogonal.
Karakterisasi protein antibakteri dilakukan secara observasi. Karakterisasi
protein antibakteri ditentukan dengan menetralkan supernatant kemudian diuji
dengan metode difusi sumuran terhadap bakteri uji yaitu E.coli, Sa.

paratyphii, B. substilis dan St aureus. Jika terbentuk zona jernih diduga zat
antibakteri dapat berupa protein, maka dilanjutkan dengan menetukan berat
molekul protein yang dapat menghambat pertumbuhan bakteri uji dengan
menggunakan teknik Sodium Dodecyl Sulfat Polyakrilamide Gel
Elektoforesis (SDS-PAGE). .
D. Prosedur Kerja
1. Peremajaan Bakteri
Isolat bakteri Lactobacillus L1, Lactobacillus L2, Lactobacillus L3 dan
Lactobacillus L4 dibiakkan dengan cara digores pada media MRS agar
miring, sedangkan untuk bakteri uji dibiakkan dengan cara digores pada
media NA miring.
2. Produksi Senyawa Antibakteri
Media produksi antibakteri disiapkan dengan komposisi MRS cair
ditambah 3 % glukosa (pH 6,2 ± 0,2). Masing-masing isolat Lactobacillus
L1, Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4
diinokulasikan ke dalam lima erlenmeyer yang berisi 20 mL media
produksi. Setiap erlemeyer diinkubasi di dalam inkubator pada suhu 37o C
sesuai dengan lama produksi yaitu 1 hari, 2 hari, 3 hari, 4 hari dan 5 hari.
Setiap hari kultur dideteksi karakteristik senyawa antibakteri hingga hari
ke-5 fermentasi.

3. Karakterisasi Antibakteri Berupa Asam Organik
Asam organik dapat dideteksi melalui penurunan pH media kultur dan
dilanjutkan dengan mengukur total asam yang dihasilkan Lactobacillus
L1, Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4. Media
kultur diambil sebanyak 5 mL secara aseptis dan diukur pH media dengan
menggunakan pH meter. Kemudian 5 mL kultur tersebut ditambahkan
sebanyak 10 mL aquades netral dan dititrasi dengan 0,1 N NaOH hingga
larutan menjadi netral. Total asam dapat dihitung dengan menggunakan
persamaan berikut :
Keterangan :
Jumlah NaOH = volume NaOH yang dibutuhkan untuk menetralkan
media.
Normalitas NaOH = 0,1 N
Jumlah Bahan = 5 mL
BM asam laktat = 90
4. Uji Daya Antibakteri
Setelah Lactobacillus L1, Lactobacillus L2, Lactobacillus L3 dan
Lactobacillus L4 diinkubasi selama lima hari pada media kultur, media
tersebut diambil sebanyak 2 mL. Untuk memisahkan zat antibakteri
Jumlah NaOH x Normalitas NaOH x BM asam laktat
Total Asam =
Jumlah Bahan

dengan sel penghasil maka kultur disentrifuge dengan kecepatan 11.000
rpm selama 5 menit. Supernatan yang diperoleh merupakan ekstrak
antibakteri yang diduga mengandung asam organik dan protein
penghambat. Untuk menghilangkan pengaruh aktivitas asam organik di
dalam menghambat pertumbuhan bakteri uji maka supernatan dinetralkan
dengan menggunakan 1 N NaOH. Larutan tersebut digunakan untuk uji
daya hambat terhadap bakteri uji. Bakteri uji yang digunakan, yakni E.coli,
Sa.paratyphii, B. substilis, dan St. aureus.
Bakteri uji diinokulasi sebanyak 1 ose ke dalam tabung reaksi yang berisi
5 mL aquades steril sampai diperoleh kekeruhan yang sama dengan
Standar Mac Farlan ( 3 x 108 CFU/mL ). Kemudian sebanyak 1 mL bakteri
uji dimasukkan ke dalam cawan petri steril dan ditambahkan 25 mL media
NA steril. Supaya bakteri menyebar rata pada media maka cawan petri
diputar sesuai angka 8 sebelum media NA memadat. Setelah media
memadat, di dalam media dibuat lubang membentuk sumur dengan
diameter 1 cm. Sebelum larutan antibakteri dimasukkan ke sumur, bagian
bawah cawan petri dibuat 3 garis yang saling bersilangan dan pusat sumur
sebagai titik persilangan tiga garis tersebut ( gambar 3). Setelah itu,
larutan antibakteri dimasukkan ke dalam sumur sebanyak 100 µL. Kultur
diinkubasi selama 24 jam pada suhu 37oC. Kemudian diukur diameter
zona bening yang dihasilkan sesuai dengan garis yang telah dibuat.
Gambar 3. Bentuk sumur dilihat dari bagian bawah cawan petri
5. Sodium Dodecyl Sulfat-Polyakrilamide Gel Elektoforesis (SDS-PAGE)
dan Identifikasi Aktivitas Pita Protein
Masing-masing supernatan antibakteri yang akan diindentifikasi dicampur
dengan 5 x buffer sampel (lampiran) di dalam mikrotube. Supaya
tercampur dengan rata, campuran tersebut disentrifuge dengan kecepatan
10.000 rpm selama 10 detik. Elektroforesis protein menggunakan gel
pemisah ( 8 % polyakrilamide) dan gel penahan (4 % polyakrilamide)
(tabel 3). Setelah gel dipasang pada perangkat elektroforesis, 500 mL 5 x
buffer elektroforesis pH 8,3 dituangkan ke dalam chamber. Sebanyak 20
µL campuran sampel tersebut dimasukkan ke dalam gel penahan, dan
untuk standar sebanyak 2 µL ke dalam sumur gel penahan.
Tabel 3 . Komposisi Gel Pemisah dan Gel Penahan
Komposisi 8 % Gel Pemisah
(mL)
4 % Gel Penahan
(mL)
Akuades 4,80 1,54
1,5 M buffer tris-HCl pH 8,8
+SDS
2,55 -
0,5 M buffer tris-HCL pH 6,8 - 0,65
30 % akrilamide 2,66 0,67

10 % APS 0,10 0,05
TEMED 0,01 0,01
Elektroforesis dijalankan pada tegangan konstan 100 volt 50 mA selama 1-
2 jam hingga migrasi bromo fenol biru berada dibagian bawah gel
pemisah. Gel elektroforesis dilepas dari cetakan dan jarak migrasi bromo
fenol biru diukur dari batas atas gel pemisah. Gel untuk mengetahui berat
molekul protein diwarnai dengan pewarnaan perak nitrat (AgNO3). Dalam
pewarnaan perak nitrat, mula-mula gel direndam dalam larutan fiksasi
selama 1 jam hingga semalam dengan diinkubasi bergoyang kecepatan
3.000 rpm. Kemudian gel tetap dalam keadaan bergoyang di rendam 50 %
etanol selama 20 menit. Setelah itu, gel tersebut direndam sebanyak dua
kali di larutan 30 % etanol (masing-masing selama 20 menit).
Selanjutnya gel direndam dalam larutan enhancer selama 1 menit.
Kemudian gel tersebut dicuci tiga kali dalam akuabides masing-masing 20
detik. Setelah dicuci, gel direndam dalam larutan perak nitrat selama 30
menit. Untuk memunculkan pita protein, gel direndam dengan larutan
natrium karbonat selama 3-5 menit sampai muncul pita protein. Untuk
menghentikan munculnya pita protein tersebut, gel direndam dengan
larutan fiksasi. Agar gel tidak menyusut ketika dalam penyimpanan,
maka gel direndam pada akuades selama semalam. Setelah itu, pita
protein yang muncul diukur jarak migrasinya. Berat molekul protein
diukur dengan menggunakan standar berat molekul rendah, yaitu
fosforilase b (otot kelinci) 94 kDa, albumin (serum bovin) 67 kDa,
ovalbumin (putih telur) 45 kDa, karbonat anhidrase (eritrosin bovin) 30

kDa, tripsin inhibitor (kedelai) 20,1 kDa, dan α-laktal-bumin (susu bovin)
14,4 kDa.
Gel untuk identifikasi pita protein penghambat kemudian direnaturasi
untuk menghilangkan SDS sesuai dengan Osmanagaoglu et all (1998).
Mula-mula gel direndam dalam larutan 20 % isopropanol selama 1 jam
dan dilanjutkan pada larutan 10 % asam asetat. Kemudian gel direndam
akuabides selama semalam dalam shaker dengan kecepatan 3.000 rpm
untuk menghilangkan pengaruh dari isopropanol dan asam asetat. Untuk
mengetahui daya hambat masing-masing protein penghambat, gel
diletakkan di atas media NA yang telah mengandung bakteri uji yang
paling sensitif terhadap protein penghambat (disesuaikan dari hasil uji
supernatan antibakteri yang dinetralkan). Media tersebut diinkubasi
selama 24 jam pada suhu 37 oC. Di sekitar pita protein penghambat akan
bebas dari bakteri uji.

E. Diagram Alir
lam supernatan di
Produksi senyawa antibakteri dari isolat
Lactobacillus L1, L2, L3 dan L4 dengan lama
produksi, yakni hari ke-1, ke-2, ke-3, ke-4 dan
ke-5
Mengukur pH media
kultur
Memisahkan senyawa antibakteri
dari sel bakteri dengan cara
disentrifuge 11.000 rpm selama 5
menit
Mentitrasi 5 mL
media kultur dengan
0,1 N NaOH
Menghitung total
asam yang
dihasilkan isolat
Lactobacillus L1,
L2, L3, dan L4
Supernatan dinetralkan
dengan 1 N NaOH.
Larutan tersebut diujikan
terhadap bakteri E.coli,
Sa.paratyphii, B.
substilis, dan St. aureus
dengan metode difusi
sumur
Protein penghambat yang
terdapat di supernatant di
deteksi berat molekulnya
dengan teknik SDS-PAGE
Pewarnaan
perak nitrat
Uji daya
hambat pita
protein Menghitung
berat
molekul
protein
Menghitung
berat
molekul
protein
IV. HASIL DAN PEMBAHASAN
A. Karakterisasi Antibakteri Senyawa Asam Organik
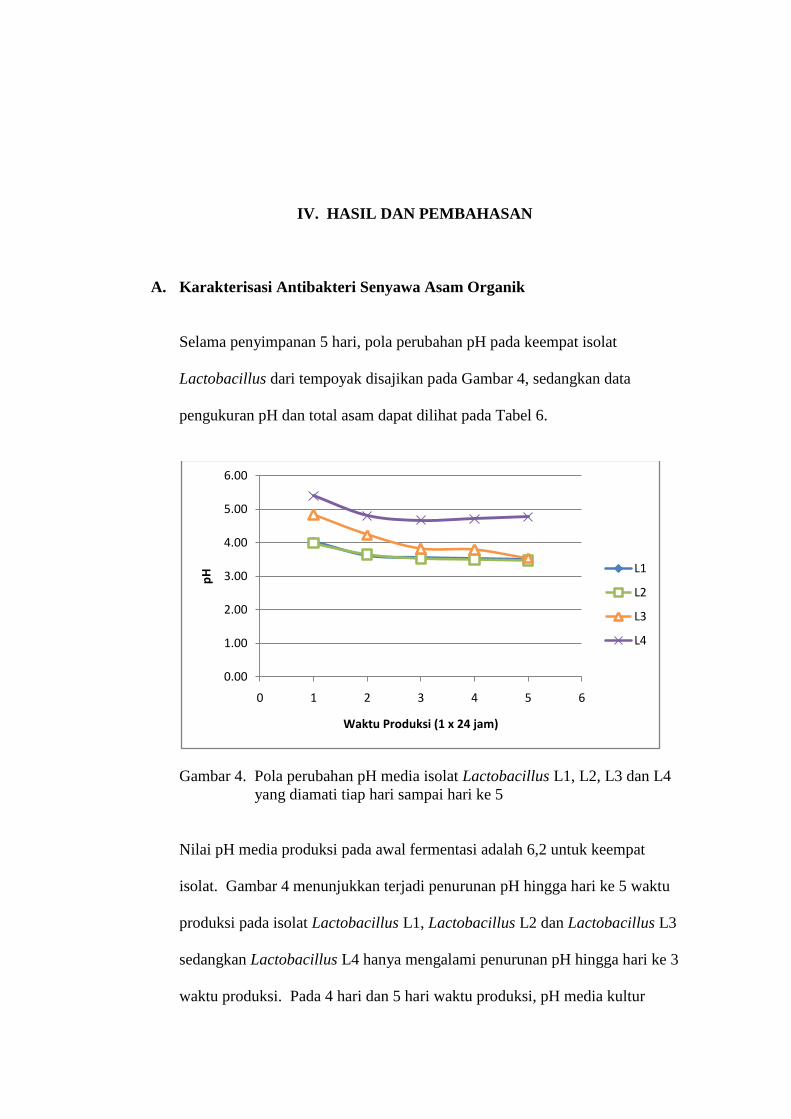
Selama penyimpanan 5 hari, pola perubahan pH pada keempat isolat
Lactobacillus dari tempoyak disajikan pada Gambar 4, sedangkan data
pengukuran pH dan total asam dapat dilihat pada Tabel 6.
Gambar 4. Pola perubahan pH media isolat Lactobacillus L1, L2, L3 dan L4
yang diamati tiap hari sampai hari ke 5
Nilai pH media produksi pada awal fermentasi adalah 6,2 untuk keempat
isolat. Gambar 4 menunjukkan terjadi penurunan pH hingga hari ke 5 waktu
produksi pada isolat Lactobacillus L1, Lactobacillus L2 dan Lactobacillus L3
sedangkan Lactobacillus L4 hanya mengalami penurunan pH hingga hari ke 3
waktu produksi. Pada 4 hari dan 5 hari waktu produksi, pH media kultur
0.00
1.00
2.00
3.00
4.00
5.00
6.00
0 1 2 3 4 5 6
pH
Waktu Produksi (1 x 24 jam)
L1
L2
L3
L4

isolat Lactobacillus L4 mengalami kenaikan. pH terendah dari masing-
masing isolat Lactobacillus L1, Lactobacillus L2, Lactobacillus L3 dan
Lactobacillus L4 yaitu 3,50, 3,47, 3,54 dan 4,67. Penurunan pH yang
terbentuk berkisar 5,40 pada isolat Lactobacillus L4 dengan waktu produksi 1
hari hingga 3,47 untuk isolat Lactobacillus L2 dengan waktu produksi 5 hari.
Terjadinya penurunan pH media menunjukkan bahwa keempat isolat
Lactobacillus dari tempoyak menghasilkan asam-asam organik yang dapat
digunakan oleh isolat tersebut untuk menghambat pertumbuhan bakteri lain.
Grafik perubahan total asam yang dihasilkan oleh isolat Lactobacillus L1,
Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4 dalam membentuk
asam organik pada media MRS disajikan pada Gambar 5. Data yang
diperoleh dianalisis dengan analisis ragam yang dapat dilihat pada Tabel 4.
. Gambar 5. Total asam organik isolat Lactobacillus L1, L2, L3 dan L4 yang
diamati tiap hari sampai hari ke 5
0.00
5.00
10.00
15.00
20.00
25.00
30.00
35.00
40.00
0 1 2 3 4 5 6
Tota
l Asa
m (
%)
Waktu Produksi (1 x 24 jam)
L1
L2
L3
L4

Tabel 4. Analisis ragam total asam yang dihasilkan isolat Lactobacillus L1,
L2, L3 dan L4 terhadap waktu produksi (hari).
Sumber
Keragaman db jk kt F Sign
Ftb
1%
Ftb
5 %
Perlakuan 19 4.808,45 253,08 17,46 Sign * 2,97 2,14
isolat 3 3.256,64 1.085,55 74,90 Sign * 4,94 3,10
waktu 4 1.255,72 313,93 21,66 Sign * 4,43 2,87
iso x wak 12 296,08 24,67 1,70 tn 3,23 2,28
Galat 20 289,86 14,49
Total 39 5.098,30
Keterangan : * = berbeda nyata pada taraf 1% dan 5%
tn = tidak berbeda nyata
Hasil analisis ragam data total asam menunjukkan hubungan antara setiap
jenis isolat Lactobacillus yaitu berbeda nyata. Hal ini menjelaskan bahwa
setiap isolat Lactobacillus yang ditemukan dari tempoyak memiliki
kemampuan yang berbeda dalam hal menghasilkan asam-asam organik.
Perbedaan kadar asam organik tidak terlepas dari karakter dari masing-
masing isolat Lactobacillus. Setiap isolat memiliki kondisi optimum yang
berbeda dalam pertumbuhannya, antara lain pH, suhu dan kebutuhan nutrisi.
Tidak semua isolat Lactobacillus pertumbuhannya optimum berada pada pH
6,2 sehingga akan mempengaruhi fase pertumbuhan. Isolat Lactobacillus L1
mampu menghasilkan total asam sebesar 37,15 dan isolat Lactobacillus L2
sebesar 36,13 kemungkinan memiliki pH optimum pada pH 6,2 sehingga
lebih mudah untuk beradapatasi dan mencapai fase logaritmik secara optimal
dibandingkan dengan isolat Lactobacillus L3 sebesar 29,94 dan isolat
Lactobacillus L4 sebesar 7,98. Pada fase logaritmik, metabolit primer yang
diantaranya yaitu asam-asam organik banyak dihasilkan pada fase ini.
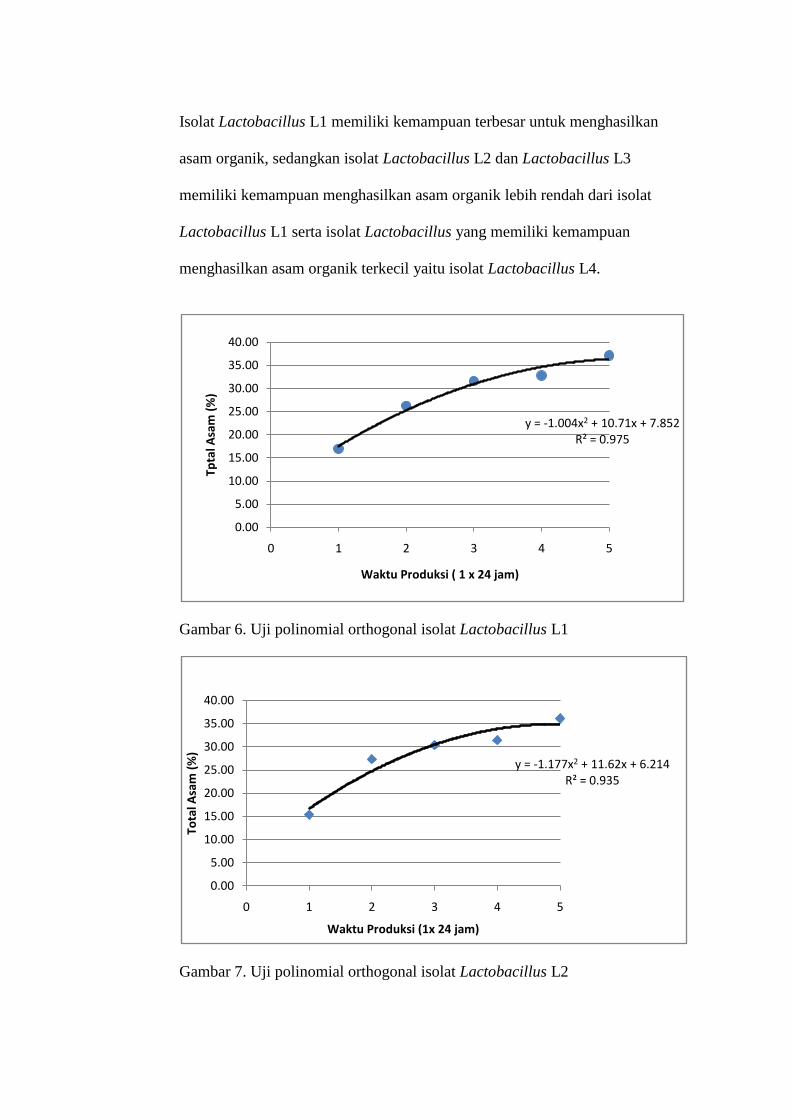
Isolat Lactobacillus L1 memiliki kemampuan terbesar untuk menghasilkan
asam organik, sedangkan isolat Lactobacillus L2 dan Lactobacillus L3
memiliki kemampuan menghasilkan asam organik lebih rendah dari isolat
Lactobacillus L1 serta isolat Lactobacillus yang memiliki kemampuan
menghasilkan asam organik terkecil yaitu isolat Lactobacillus L4.
Gambar 6. Uji polinomial orthogonal isolat Lactobacillus L1
Gambar 7. Uji polinomial orthogonal isolat Lactobacillus L2
y = -1.004x2 + 10.71x + 7.852R² = 0.975
0.00
5.00
10.00
15.00
20.00
25.00
30.00
35.00
40.00
0 1 2 3 4 5
Tpta
l Asa
m (
%)
Waktu Produksi ( 1 x 24 jam)
y = -1.177x2 + 11.62x + 6.214R² = 0.935
0.00
5.00
10.00
15.00
20.00
25.00
30.00
35.00
40.00
0 1 2 3 4 5
Tota
l Asa
m (
%)
Waktu Produksi (1x 24 jam)

Gambar 8. Uji polinomial orthogonal isolat Lactobacillus L3
Gambar 9. Uji polinomial orthogonal isolat Lactobacillus L4
Keragaman waktu produksi juga menunjukkan adanya perbedaan yang nyata
terhadap kadar asam organik yang diproduksi oleh keempat isolat
Lactobacillus. Selama fermentasi berlangsung, total asam organik yang
diproduksi oleh keempat isolat Lactobacillus meningkat hingga hari ke 5
waktu produksi. Waktu fermentasi menunjukkan beda nyata dengan
y = 0.075x2 + 4.859x + 3.276R² = 0.992
0.00
5.00
10.00
15.00
20.00
25.00
30.00
35.00
0 1 2 3 4 5
Tota
l Asa
m (
%)
Waktu Produksi (1x 24 jam)
y = -0.25x2 + 2.32x + 2.408R² = 0.930
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
8.00
9.00
0 1 2 3 4 5
Tota
l Asa
m (
%)
Waktu Produksi (1 x 24 jam)

membentuk garis lengkung (kuadratik) dengan persamaan regresi
y = -1,004x2 + 10,71x + 7,852, R² = 0,975 untuk isolat Lactobacillus L1,
y = -1,177x2 + 11,62x + 6,214,R² = 0,935 untuk isolat Lactobacillus L2 dan
y = -0,25x2 + 2,32x + 2,408, R² = 0,930 untuk isolat Lactobacillus L4. Hal
ini berarti bahwa dengan semakin lama waktu fermentasi, total asam organik
memiliki kecendrungan untuk meningkat dan menurun pada hari tertentu.
Penurunan total asam disebabkan karena asam organik yang dihasilkan
keempat isolat Lactobacillus ada yang bersifat volatil. Menurut Ekowati
(2000), hasil analisis asam organik isolat Lactobacillus dari tempoyak
terdapat asam-asam organik antara lain asam laktat, asam asetat, asam
propionat dan asam butirat. Asam butirat merupakan asam organik yang
bersifat volatil sehingga tidak terjadi akumulasi asam butirat. Selain itu, asam
organik yang tidak volatil kemungkinan juga dimanfaatkan oleh bakteri untuk
metabolisme selanjutnya (Mayasari, 2007).
Berdasarkan persamaan regresi, isolat Lactobacillus L1 menghasilkan asam
organik maksimum pada hari ke 5 waktu produksi yaitu sebesar 37,15.
Begitu juga dengan isolat Lactobacillus L2 dan Lactobacillus L4
menghasilkan asam organik maksimum pada hari ke 5 waktu produksi yaitu
36,13 untuk Lactobacillus L2 dan 7,89 untuk Lactobacillus L4.
Peningkatan produksi asam organik terbesar pada hari ke-5 waktu produksi
menunjukkan bahwa aktivitas ketiga isolat bakteri Lactobacillus mencapai
fase pertumbuhan logaritmik. Hal ini sesuai dengan hasil penelitian Kartika

(2000) bahwa fase logaritmik isolat Lactobacillus yang berasal dari tempoyak
mulai pada hari ke-0 sampai hari ke-7.
Total asam yang dihasilkan isolat Lactobacillus L3 terhadap waktu produksi
(hari) menunjukkan pola linier dengan persamaan regresi y = 0,075x2 +
4,859x + 3,276, R² = 0,992. Hal ini berarti produksi asam organik oleh isolat
Lactobacillus L3 selama 5 hari fermentasi mengalami peningkatan.
Interaksi antara isolat dengan waktu produksi tidak berbeda nyata. Hal ini
berarti bahwa interaksi setiap isolat dengan keragaman waktu adalah sama.
Data total asam terbesar dihasilkan oleh isolat Lactobacillus L1 pada hari
kelima sedangkan total asam terkecil dihasilkan oleh isolat Lactobacillus L4
pada hari pertama.
B. Karakterisasi Antibakteri Senyawa Protein
Ekstrak antibakteri isolat Lactobacillus L1, Lactobacillus L2, Lactobacillus
L3 dan Lactobacillus L4 sebelum dinetralkan mampu menghambat
pertumbuhan dari bakteri Stapylococcus aureus, Bacillus substilis,
Escherchia coli, dan Salmonella paratyphii dengan terbentuknya zona hambat
disekitar sumur. Namun, ketika ekstrak antibakteri keempat isolat tersebut
dinetralkan dengan NaOH hanya ekstrak antibakteri isolat Lactobacillus L4
terhadap St. aureus yang masih membentuk zona hambat sedangkan yang
lainnya kemampuan ekstrak antibakteri isolat Lactobacillus L1, Lactobacillus
L2 dan Lactobacillus L3 terhadap St aureus, B. substilis, E. coli dan Sa.
paratyphii dan ektrak antibakteri L4 terhadap B. substilis, E. coli dan

Sa. paratyphii hilang atau tidak menghasilkan zona hambat ( Tabel 5 ).
Fungsi penetralan bertujuan untuk menghilangkan pengaruh dari asam
organik yang digunakan oleh keempat isolat Lactobacillus untuk
menghambat pertumbuhan bakteri-bakteri uji.
Tabel 5. Zona hambat dari ekstrak antibakteri isolat Lactobacillus L1, L2, L3
dan L4 sebelum dan sesudah dinetralkan terhadap bakteri uji
Bakteri Uji
Ekstrak antibakteri
sebelum dinetralkan
Ekstrak antibakteri
sesudah dinetralkan
L1 L2 L3 L4 L1 L2 L3 L4
Staphylococcus aureus + + + + - - - +
Bacillus substilis + + + + - - - -
Escherchia coli + + + + - - - -
Salmonella paratyphii + + + + - - - -
Keterangan : + = terbentuk zona hambat ( ekstrak antibakteri menghambat
pertumbuhan bakteri uji) ,
- = tidak terbentuk zona hambat (ekstrak antibakteri tidak
menghambat pertumbuhan bakteri uji)
Hasil dari elektroforesis gel polyakrilamid, SDS PAGE, dapat dilihat bahwa
pada isolat Lactobacillus L1, Lactobacillus L2 dan Lactobacillus L3 tidak
terdapat pita protein yang terbentuk, sedangkan isolat Lactobacillus L4
terdapat empat pita protein dengan berat molekul 15 kDa, 24 kDa, 53 kDa
dan 169 kDa akan tetapi protein tersebut tidak mampu menghambat
pertumbuhan St. aureus ( Gambar 6 dan 7). Tidak terbentuknya zona hambat
pada pita protein Lactobacillus L4 kemungkinan disebabkan karena kadar
protein yang terlalu sedikit untuk menghambat pertumbuhan St. aureus yang
tumbuh pada gel.

Keempat isolat Lactobacillus yang ditemukan pada tempoyak tidak memiliki
protein penghambat yang dapat menghambat pertumbuhan St. aureus,
B. substilis, E. coli dan Sa. paratyphii.. Menurut Kusmiati dan Malik (2002),
protein penghambat memiliki pengaruh bakterisidal dan bakteriostatik
terhadap bakteri yang mempunyai hubungan dekat dengan bakteri
penghasilnya karena bakteri target memiliki sifat pengikatan spesifik (specific
binding site) sehingga kemungkinan isolat Lactobacillus L4 memiliki protein
penghambat yang menghambat pertumbuhan bakteri selain bakteri St aureus,
B. substilis, E. coli dan Sa. paratyphii.
Ekstrak antibakteri Lactobacillus L4 yang telah dinetralkan membentuk zona
hambat disebabkan karena ada senyawa lain selain protein dan asam organik
yang berperan dalam menghambat pertumbuhan Sa aureus pada isolat
Lactobacillus L4, seperti hydrogen peroksida, etanol, diasetyl dan karbon
dioksida.

169 kDa
53 kDa
24 kDa
15 kDa
M a b c d
Gambar 6. SDS-PAGE isolat Lactobacillus L1, L2, L3 dan L4
Keterangan : M = protein standart
a = protein dari isolat Lactobacillus L1
b = protein dari isolat Lactobacillus L2
c = protein dari isolat Lactobacillus L3
d = protein dari isolat Lactobacillus L4
Gambar 7. Gel SDS-PAGE ektrak antibakteri L4 yang diuji terhadap
St.aureus.

Jenis – jenis Lactobacillus yang mampu menghasilkan protein penghambat
dalam bentuk bakteriosin antara lain L. plantarum ST194BZ menghasilkan 2
bakteriosin dengan berat molekul 3,3 kDa dan 14 kDa (Todorov dan Dick,
2005), L. amylovorus LMPGP-13139 menghasilkan bakteriosin dengan berat
molekul 4,5 dan 6 kDa (Contreras et all, 1996) , L. sake menghasilkan
sakacin B dengan berat molekul 6,3 kDa (Samelis et all, 1994), L.
acidophilus menghasilkan acidophilin 801 dengan berat molekul 6,5 kDa
(Zamfir et all, 2001), L. bavaricus M1401 menghasilkan bavaricin A dengan
berat molekul 3,5-4 kDa (Larsen et all, 1993), L. gaseri KT7 menghasilkan
gassericin KT7 dengan berat molekul 4,5-5 kDa (Zhu et all, 2000). Namun,
ada juga jenis Lactobacillus yang tidak menghasilkan protein penghambat
berupa bakteriosin seperti L. fermentum, L. delbrueckii dan L. reuteri (Sanni
et all,1999)
Bakteriosin disintesis mengikuti pola sintesis protein. Sistem ini diatur oleh
plasmid DNA. Pada umumnya, bakteriosin non lantibiotik disintesis melalui
jalur ribosomal, sedangkan kelompok lantibiotik disintesis secara ribosomal
sebagai prepeptida kemudian mengalami modifikasi. Sekresi prepeptida
dilakukan pada fase eksponensial dan diproduksi secara maksimal pada fase
stasioner. Prinsip regulasi sintesis bakteriosin diatur oleh adanya gen
pengkode produksi dan pengkode immunitas (Kim, 1990).

V. KESIMPULAN DAN SARAN
A. Kesimpulan
Dari hasil penelitian yang telah dilakukan, maka didapatkan kesimpulan
sebagai berikut :
1. Senyawa antibakteri yang dihasilkan oleh keempat isolat Lactobacillus
yaitu berupa asam organik.
2. Isolat Lactobacillus L1 paling berpotensi menghasilkan asam organik
sebesar 37,15 dibandingkan dengan isolat Lactobacillus L2 sebesar 36,13,
isolat Lactobacillus L3 sebesar 29,94 dan isolat Lactobacillus L4 sebesar
7,98.
3. Waktu dalam mencapai total asam maksimum isolat Lactobacillus L1,
Lactobacillus L2, Lactobacillus L3 dan Lactobacillus L4 pada hari ke-5
waktu produksi.
4. Isolat Lactobacillus L1, Lactobacillus L2 dan Lactobacillus L3 tidak
menghasilkan protein sedangkan isolat Lactobacillus L4 menghasilkan
protein dengan berat molekul 15 kDa, 24 kDa, 53 kDa dan 169 kDa akan
tetapi bukan sebagai antibakteri.

B. Saran
Perlu diadakan penelitian lanjutan untuk mengetahui kadar potein penghambat
dan nilai optimum dari asam organik yang dihasilkan isolat Lactobacillus dari
tempoyak dengan memperbanyak waktu produksi dan mengetahui kondisi
optimum pertumbuhan dari keempat isolat Lactobacillus seperti suhu dan pH.


Tabel 6. Hasil Pengukuran pH Media Kultur Isolat Lactobacillus
Lactobacillus L1 L2 L3 L4
Hari
1 4,03 3,99 4,84 5,40
2 3,63 3,65 4,24 4,80
3 3,55 3,53 3,83 4,67
4 3,53 3,50 3,80 4,71
5 3,50 3,47 3,54 4,77
Tabel 7 . Hasil Pengukuran Total Asam Media Kultur Isolat Lactobacillus
Perlakuan Ulangan
X X2 X
1 2
L1H1 16,65 17,28 33,93 575,82 16,97
L1H2 29,07 23,40 52,47 1.392,62 26,24
L1H3 32,94 30,31 63,25 2.003,74 31,63
L1H4 34,02 31,57 65,59 2.154,03 32,80
L1H5 37,31 36,99 74,30 2.760,30 37,15
L2H1 17,46 13,32 30,78 482,27 15,39
L2H2 31,14 23,49 54,63 1.521,48 27,32
L2H3 32,38 28,42 60,80 1.856,16 30,40
L2H4 32,94 29,88 62,82 1.977,86 31,41
L2H5 36,16 36,09 72,25 2.610,03 36,13
L3H1 9,86 6,12 15,98 134,67 7,99
L3H2 16,56 10,44 27,00 383,23 13,50
L3H3 21,78 16,74 38,52 754,60 19,26
L3H4 28,26 17,26 45,52 1.096,54 22,76
L3H5 37,44 22,43 59,87 1.904,86 29,94
L4H1 4,25 4,21 8,46 35,79 4,23
L4H2 5,94 7,20 13,14 87,12 6,57
L4H3 5,76 8,32 14,08 102,40 7,04
L4H4 6,66 7,87 14,53 106,29 7,27
L4H5 7,56 8,39 15,95 127,55 7,98
Total 823,87 22.067,35 411,94
Gambar 9. Isolat Lactobacillus L1
Gambar 10. Isolat Lactobacillus L2

Gambar 11. Isolat Lactobacillus L3
Gambar 12. Isolat Lactobacillus L4

L1 L2
L3 L4
Gambar 13. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sebelum Dinetralkan Pada Bakteri Uji Staphylococcus aureus
L1 L2
L3 L4
Gambar 14. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sesudah Dinetralkan Pada Bakteri Uji Staphylococcus aureus
L1 L2
L3 L4
Gambar 15. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sebelum Dinetralkan Pada Bakteri Uji Bacillus substilis

L1 L2
L3 L4
Gambar 16. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sesudah Dinetralkan Pada Bakteri Uji Bacillus substilis
L1 L2
L3 L4
Gambar 17. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sebelum Dinetralkan Pada Bakteri Uji Escherchia coli
L1 L2
L3 L4
Gambar 18. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sesudah Dinetralkan Pada Bakteri Uji Escherchia coli

L1 L2
L3 L4
Gambar 19. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sebelum Dinetralkan Pada Bakteri Uji Salmonella paratyphii
L1 L2
L3 L4
Gambar 20. Uji Sumur Ekstrak Antibakteri Isolat Lactobacillus L1, L2, L3 dan
L4 Sesudah Dinetralkan Pada Bakteri Uji Salmonella paratyphii

Tabel 8. Penetuan kurva logaritma berat molekul protein standart terhadap
mobilitas relatif (rf)
Standart Berat Molekul
(Da) (Y) Log (Y) Mobilitas
Relatif (rf) (X)
α-lactal-bumin 14.400 4,16 0,956
Trypsin inhibitor 20.100 4,30 0,911
Carbonic an-hydrase 30.000 4,48 0,856
Ovalbumin 45.000 4,65 0,822
Albumin 66.000 4,82 0,778
Fosforilase b 97.000 4,99 0,722
Keterangan :
1. Jarak migrasi bromofenol biru untuk standart protein diukur dari batas atas
gel pemisah yaitu 4,5 cm
2. a. Jarak dari batas atas gel pemisah ke Fosforilase b yaitu 3,25 cm
b. Jarak dari batas atas gel pemisah ke Albumin yaitu 3,5 cm
c. Jarak dari batas atas gel pemisah ke Ovalbumin yaitu 3,7 cm
d. Jarak dari batas atas gel pemisah ke Carbonic an-hydrase yaitu 3,85 cm
e. Jarak dari batas atas gel pemisah ke Trypsin inhibitor yaitu 4,1 cm
f. Jarak dari batas atas gel pemisah ke α-lactal-bumin yaitu 4,3 cm
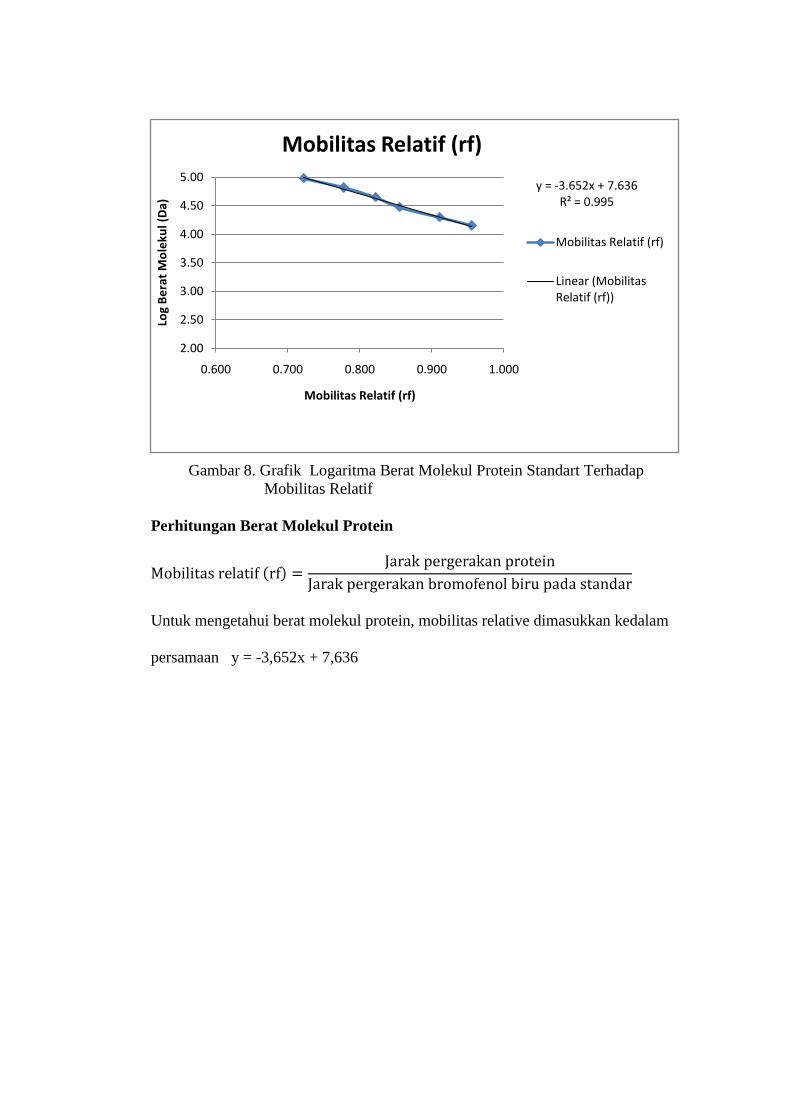
y = -3.652x + 7.636R² = 0.995
2.00
2.50
3.00
3.50
4.00
4.50
5.00
0.600 0.700 0.800 0.900 1.000
Log
Be
rat
Mo
leku
l (D
a)
Mobilitas Relatif (rf)
Mobilitas Relatif (rf)
Mobilitas Relatif (rf)
Linear (Mobilitas Relatif (rf))
Gambar 8. Grafik Logaritma Berat Molekul Protein Standart Terhadap
Mobilitas Relatif
Perhitungan Berat Molekul Protein
Untuk mengetahui berat molekul protein, mobilitas relative dimasukkan kedalam
persamaan y = -3,652x + 7,636

Tabel 9. Penetuan Berat Molekul Protein Terhadap Mobilitas Relatif (Rf) Pada
Isolat Lactobacillus L4
Mobilitas Relatif
(x) Log (y)
Berat Molekul
(Da) (y)
0,947 4,178 15.066,07
0,894 4,373 23.604,78
0,798 4,722 52.722,98
0,660 5,227 168.655,30
Keterangan :
1. Jarak migrasi bromofenol biru untuk sampel L4 diukur dari batas atas gel
pemisah yaitu 4,7 cm.
2. Jarak dari batas atas gel pemisah ke protein pada sampel L4 adalah 3,1
cm, 3,75 cm, 4,2 cm, dan 4,45 cm
Reagent yang digunakan
1. Larutan Standar Mac Farlan
Larutan 1 % asam sulfat (H2SO4) sebanyak 9,9 mL dicampur dengan 1 %
barium klorida (BaCl2.2H2O) sebanyak 0,1 mL. Kocok terlebih dahulu sebelum
digunakan.
2. 0,1 N NaOH
Sebanyak 40 g NaOH dilarutkan dalam1000 mL akuades. Larutan disimpan
dalam suhu ruang.

3. 1 % phenophetylen
Sebanyak 1 gram phenophetylen dilarutkan dalam 50 mL alkohol 96 % .
Setelah larut ditambahkan 50 mL akuades. Larutan disimpan dalam suhu
ruang.
4. 5 x buffer sampel
Larutan ini terdiri dari 0,12 mL 1 M buffer tris-HCl pH 6,8 (konsentrasi akhir
60 mM), 1 mL 50 % gliserol (konsentrasi akhir 25 %), 0,4 mL 10 % SDS
(2%), 0,1 mL β-merkaptoetanol (14,4 mM), 0,2 mL 1 % bromofenol biru
(0,1%) dan 0,18 mL akuabides. Sampel protein diencerkan dengan perbadingan
1:4 dengan 5x larutan stok. Larutan disimpan pada suhu 4oC.
5. 5 x buffer elektroforesis pH 8,3
Larutan terdiri atas 9 g tris basa, 43,2 g glisin, 3 g SDS yang dilarutkan hingga
600 mL akuades. Untuk membuat 300 mL larutan, 60 mL 5 x larutan stok
diencerkan dengan akuades sebanyak 240 mL. Larutan disimpan pada suhu
4oC.
6. Elektroforesis gel polyakrilamide 30 % ( 29% : 1 % )
Sebanyak 29,2 g akrilamida dicampur dengan 0,8 g N’N’-bis-akrilamida dan
dilarutkan dengan akuades hingga 100 mL. Campuran disaring dengan kertas
saring, dan disimpan dalam botol gelap pada suhu 4oC. Sebelum digunakan
campuran tersebut divakum selama 15 menit pada suhu kamar.

7. 1,5 M buffer tris-HCl pH 8,8
Ke dalam 50 mL akuades dimasukkan 18,15 g tris (hidroksimetil) aminometan
(tris basa), pH diatur 8,8 dengan 1 N HCl. Akuades ditambah hingga 100 mL.
Larutan disimpan pada suhu 4oC.
8. 0,5 M buffer tris-HCl pH 6,8
Ke dalam 60 mL akuades dimasukkan 6 g tris basa, pH diatur 8,8 dengan 1 N
HCl. Akuades ditambah hingga 100 mL. Larutan disimpan pada suhu 4oC.
9. 10 % SDS
Sebanyak 0,1 g SDS dilarutkan dalam 1 mL akuades. Larutan disimpan pada
suhu 4oC dan dibuat segar setiap minggu.
10. 10 % Amonium Persulfat (APS)
Satu mL akuades mengandung 1 g APS. Larutan ini tersedia harus dalam
keadaan segar.
11. Larutan fiksasi
Larutan ini terdiri atas 25 % metanol, 12 % asam asetat, dan 63 % akuades.
Larutan ini disimpan dalam botol gelap pada suhu kamar.
12. Larutan enhancer
Sebanyak 0,02 g Na2S2O3.5H2O dilarutkan dalam 100 mL akuabides.
Larutan ini disimpan dalam botol gelap pada suhu kamar.

13. Larutan perak nitrat
Larutan ini terdiri atas 0,2 g AgNO3, 35 µL formaldehida, dan dilarutkan
dalam 100 mL akuabides. Larutan ini disimpan dalam botol gelap pada suhu
kamar.
14. Larutan natrium karbonat
Larutan ini terdiri dari 6 g NaCO3 dan 48 µL formaldehida dalam 100 mL
akuabides. Larutan ini tersedia dalam keadaan segar.





























