Embed Size (px)
Citation preview

153
ISSN: 0215-7950
Volume 13, Nomor 5, September 2017Halaman 153–160
DOI: 10.14692/jfi.13.5.153
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, Jalan Kamper, Kampus IPB Darmaga, Bogor 16680 Tel: 0251-8629364, Faks: 0251-8629362; surel: [email protected]
Kemampuan Mikrob Endofit dan Rizosfer Tanaman Karet dalam Mengendalikan Rigidoporus lignosus
The Ability of Endophytic and Rhizospheric Microbes of Rubber Trees to Control Rigidoporus lignosus
Siti Hardiyanti, Bonny Purnomo Wahyu Soekarno*, Titiek Siti YulianiInstitut Pertanian Bogor, Bogor 16680
ABSTRAK
Rigidoporus lignosus merupakan patogen penting pada tanaman karet yang mengakibatkan penyakit akar putih. Penggunaan mikrob antagonis merupakan salah satu teknik pengendalian yang direkomendasikan untuk patogen ini. Penelitian ini bertujuan memperoleh agens hayati dari dalam jaringan akar dan rizosfer tanaman karet yang berpotensi mengendalikan R. lignosus. Tahapan penelitian terdiri atas isolasi mikrob endofit dan rizosfer dari tanaman karet, uji patogenisitas, seleksi secara in vitro, pengujian secara in vivo, pengujian pelarut fosfat dan pengikat nitrogen, dan identifikasi. Sebanyak 99 isolat bakteri dan 18 isolat cendawan nonpatogen berhasil diisolasi dari akar dan rizosfer tanaman karet. Berdasarkan pengujian in vitro dan in vivo diperoleh 2 isolat bakteri dan 3 isolat cendawan yang mampu menghambat pertumbuhan R. lignosus, yaitu bakteri endofit ME8, bakteri rizosfer MR3, cendawan endofit CB8, CB6, dan CL3. Bakteri endofit ME8 mampu melarutkan fosfat dan memfiksasi nitrogen. Bakteri rizosfer MR3 hanya mampu memfiksasi nitrogen. Berdasarkan karakter morfologinya, isolat CB8 teridentifikasi sebagai hifa steril, CB6 sebagai Chaetomium sp., dan CL3 sebagai Penicillium sp. Berdasarkan perunutan gen 16S rRNA, bakteri ME8 memiliki kekerabatan dengan Bacillus siamensis B268, dan bakteri MR3 memiliki kekerabatan dengan B. amylolyquifaciens SXAU001. Mikrob endofit dan rizosfer yang telah diisolasi dari tanaman karet memiliki potensi sebagai agens hayati untuk mengendalikan R. lignosus.
Kata kunci: fiksasi nitrogen, mikrob antagonis, pelarut fosfat, penyakit akar putih
ABSTRACT
Rigidoporus lignosus is the most important pathogen of rubber tree which causes white root rot disease. The use of antagonistic microbe is recommended to control this pathogen. This research was conducted to isolate endophytic and rhizospheric microbes, and to study their ability to inhibit growth of R. lignosus. Research consisted of isolation of endophytic and rhizospheric microbes, pathogenicity test, in vitro and in vivo assays, growth promotion assays, and identification. There were 99 isolates of bacteria and 18 isolates of fungi isolated from the root and rhizosphere of rubber trees. In vitro and in vivo assay showed that 2 bacterial isolates, i.e. endophytic bacteria ME8, and rhizospheric bacteria MR3; and 3 fungal isolates, i.e. endophytic fungi CB8, CB6, and CL3 were able to inhibit the growth of R. lignosus. Endophytic bacteria ME8 showed the ability of solibilizing phosphate and fixing nitrogen. Rhizospheric bacteria MR3 showed the ability of solubilizing phosphate. The isolates CB6 and CL3 were very similar with Chaetomium sp. and Penicillium sp., respectively based on morphological characters; while CB8 was identified as mycelial sterile. Based on 16S rRNA sequences, endophytic bacterium ME8 and rhizospheric bacteria ME3 were identified as Bacillus siamensis B268 and B.

J Fitopatol Indones Hardiyanti et al.
154
amylolyquefaciens BCRh10, respectively. Endophytic and rhizospheric microbes isolated from rubber trees has the potency as biocontrol agents of R. lignosus.
Key words: antagonistic microbe, nitrogen fixer, phosphate solubilizer, root rot disease
PENDAHULUAN
Karet (Hevea brasiliensis) merupakan salah satu komoditas perkebunan yang memegang peranan penting bagi perekonomian Indonesia. Penyakit akar putih yang disebabkan oleh Rigidoporus lignosus merupakan salah satu faktor pembatas dalam budi daya tanaman karet. Kehilangan hasil yang disebabkan penyakit akar putih lebih besar dibandingkan dengan penyakit lain atau hama di berbagai negara penghasil karet. Pada tahun 2015 luas serangan R. lignosus di Indonesia mencapai 26 ribu ha dan menyebabkan kerugian hasil sebesar Rp75.67 miliar (Isnaini 2016).
Tanaman yang terinfeksi R. lignosus menunjukkan gejala daun menguning, daun gugur, pangkal batang membusuk dan mengakibatkan pohon mudah tumbang. Pada stadium lanjut penyakit ini menyebabkan kematian tanaman sehingga produktivitas kebun menurun. Rizomorf dari sumber infeksi akan menyebar melalui tanah dan menginfeksi tanaman sehat. Selain itu tubuh buah (basidiokarp) pada bagian pangkal batang tanaman juga dapat menyebarkan basidiosporanya (Omo-Ikerodah 2012).
Pengendalian penyakit akar putih yang telah dilakukan ialah penggunaan fungisida sintetis, kultur teknis, mekanis, sanitasi, dan pengendalian hayati. Pengendalian hayati dengan pemanfaatan agens antagonis merupakan alternatif yang saat ini banyak diteliti dan digunakan karena dinilai lebih efektif dan ramah lingkungan. Agens antagonis yang telah dimanfaatkan dalam mengendalikan R. lignosus sebagian besar adalah Trichoderma harzianum dan T. koningii (Jayasuriya dan Thennakoon 2007). Selain memiliki sifat sebagai agens biokontrol, banyak agens antagonis dapat memacu pertumbuhan tanaman karena mampu memproduksi hormon pertumbuhan, memfiksasi nitrogen, melarutkan fosfat,
dan memproduksi siderofor. Penelitian ini bertujuan mendapatkan agens hayati dari dalam jaringan akar dan rizosfer tanaman karet yang berpotensi mengendalikan R. lignosus.
BAHAN DAN METODE
Isolasi Mikrob dari Jaringan Akar Tanaman
Akar tanaman karet diperoleh dari perkebunan karet rakyat di Kabupaten Muaro Jambi dan Kabupaten Sarolangun, Provinsi Jambi. Sampel akar dicuci menggunakan air mengalir dan dipotong ukuran 1–2 cm. Sebanyak 1 g akar disterilkan permukaannya dengan merendam dalam alkohol 70% selama 1 menit dan dilanjutkan dengan merendam pada NaOCl 2.5% selama 2 menit, kemudian dibilas menggunakan akuades steril sebanyak 3 kali. Keberhasilan sterilisasi diuji dengan menggoreskan akar pada medium trypticase soya agar (TSA) 10%.
Isolasi bakteri dilakukan dengan meng-haluskan akar tersebut dan menambahkan 9 mL akuades steril, kemudian diencerkan secara berseri hingga 10-6. Sebanyak 0.1 mL suspensi disebar pada medium TSA 10% dan diinkubasi selama 2 hari. Isolasi cendawan dilakukan dengan meletakkan potongan akar yang telah disterilisasi permukaannya pada medium agar-agar dekstrosa kentang (ADK). Cawan diinkubasi selama 7 hari untukmemperoleh cendawan. Koloni yang menunjukkan morfologi berbeda masing-masing diremajakan pada medium ADK untuk mendapatkan biakan murni.
Isolasi Mikrob dari Rizosfer Tanaman Karet
Isolasi mikrob dari rizosfer tanaman karet dilakukan dengan metode pengenceran berseri. Sebanyak 1 g tanah ditambahkan akuades steril hingga 10 mL. Suspensi lalu digoyang pada kecepatan 100 rpm selama

J Fitopatol Indones Hardiyanti et al.
155
15 menit, kemudian diencerkan secara berseri hingga 10-6. Isolasi cendawan dilakukan dengan mengulturkan 0.1 mL suspensi tanah pada medium ADK dan diinkubasi selama 7 hari, sedangkan bakteri pada medium TSA 10% dan diinkubasi selama 2 hari. Isolat cendawan dan bakteri dengan morfologi berbeda dimurnikan.
Penyiapan Cendawan PatogenR. lignosus diperoleh dari Balai Penelitian
Karet Sembawa, Sumatra Selatan. Galur cendawan ini diremajakan selama 7 hari pada medium ADK untuk digunakan pada uji lanjut.
Uji Patogenisitas Uji patogenisitas bakteri dilakukan dengan
pengujian respons hipersensitif pada tanaman tembakau. Galur bakteri ditumbuhkan pada medium trypticase soya broth (TSB) dan digoyang selama 24 jam dengan kecepatan 100 rpm. Sebanyak 1 mL (108 cfu mL-1) suspensi bakteri diinfiltrasikan pada bagian bawah daun tembakau dan diinkubasi selama 48 jam. Galur bersifat patogen pada tanaman apabila terjadi nekrotik pada daun tembakau.
Uji patogenisitas cendawan dilakukan dengan mengamati gejala pada benih padi karena dapat menimbulkan gejala yang lebih cepat. Benih padi disterilkan permukaannya dengan merendam benih dalam larutan NaOCl 2% selama 2 menit, kemudian dibilas dengan akuades steril sebanyak 3 kali. Benih ditumbuhkan pada galur cendawan yang telah berumur 7 hari pada medium ADK. Pengamatan dilakukan setelah 14 hari dengan melihat gejala pada benih. Apabila terjadi nekrotik atau benih tidak tumbuh sempurna maka galur tersebut bersifat patogen pada tanaman.
Uji AntibiosisUji antibiosis dilakukan untuk menentukan
kemampuan mikrob dalam menekan per-tumbuhan R. lignosus. Pengujian menggunakan metode biakan ganda, yaitu galur bakteri digoreskan pada bagian tengah cawan petri kemudian pada sisi kanan dan kiri bakteri diinokulasi R. lignosus dengan jarak 2.5 cm.
Galur cendawan endofit dan rizosfer diinokulasikan pada cawan petri berisi medium ADK dan pada jarak 2.5 cm dari cendawan R. lignosus. Zona bening yang terbentuk di antara kedua isolat menunjukkan kemampuan mikrob dalam menghasilkan senyawa antibiosis. Persentase hambatan diukur pada 7 hari setelah inokulasi menggunakan rumus:
Persentase hambatan = R1– R2 × 100%, denganR1 R1, jari-jari koloni R. lignosus yang tumbuh menjauhi mikrob yang diuji; R2, jari-jari koloni R. lignosus yang mendekati mikrob yang diuji.
Uji HiperparasitismeUji hiperparasitisme dilakukan terhadap
3 galur cendawan terbaik pada uji antibiosis. Pengujian dilakukan dengan mengamati pertemuan antara kedua koloni pada uji biakan ganda menggunakan mikroskop. Indikator aktivitas hiperparasitisme dilihat dari adanya pelilitan pada hifa R. lignosus.
Uji Produksi Senyawa VolatilPengujian dilakukan terhadap 3 galur
terbaik pada uji antibiosis. Pengujian dilakukan menggunakan dua cawan petri berukuran sama. Isolat cendawan antagonis dikulturkan pada medium ADK, sedangkan bakteri antagonis dikulturkan pada medium TSA, kemudian biakan tersebut ditutup dengan cawan petri yang telah ditumbuhkan R. lignosus. Setelah 7 hari, diameter R. lignosus diukur dan persentase penghambatan dihitung menggunakan rumus:
ɸK– ɸP × 100%, denganɸK% penghambatan =
ɸK, diameter R. lignosus pada perlakuan kontrol; ɸP, diameter R. lignosus pada perlakuan mikrob antagonis.
Uji Antagonisme terhadap R. lignosus secara in Vivo
Seleksi in vivo dilakukan menggunakan potongan akar tanaman karet mengikuti protokol pada Jayasuriya dan Thenaakoon (2007) yang telah dimodifikasi. Akar yang digunakan berasal dari tanaman Klon GT umur 3 tahun dari kebun percobaan Pusat

J Fitopatol Indones Hardiyanti et al.
156
Penelitian Bioteknologi dan Bioindustri Indonesia di Ciampea. Galur mikrob antagonis yang digunakan ialah isolat yang memiliki daya hambat lebih dari 50% pada seleksi in vitro. Pengujian dilakukan dengan menggunakan kotak plastik dan diisi dengan tanah steril hingga 1/2 bagian. Potongan akar karet (panjang 8–10 cm, lebar 2–3 cm) disterilkan menggunakan penangas air pada suhu 50 °C selama 15 menit, kemudian direndam dalam suspensi agens antagonis (108 cfu mL-1 ) selama 6 jam. Sebagai kontrol, akar direndam dalam akuades steril. Akar tanaman yang telah diberi perlakukan diletakkan di atas permukaan tanah. Sisa agens antagonis disiramkan di atas permukaan akar. Inokulasi cendawan R. lignosus dilakukan pada jarak 1 cm dari ujung akar pada waktu yang sama. Setelah diinkubasi selama 10 haripertumbuhan miselium dan persentase penghambatannya dihitung.
Karakterisasi Bakteri Antagonis sebagai Pelarut Fosfat dan Penambat Nitrogen
Uji Pelarut Fosfat. Bakteri antagonis dikulturkan di atas kertas cakram pada medium Pikovskaya’s agar lalu diinkubasi pada suhu ambien selama 7 hari. Kemampuan bakteri dalam menambat fosfat ditunjukkan dengan terbentuknya zona bening di sekitar koloni bakteri.
Uji Penambat Nitrogen. Bakteri antagonis dikulturkan pada medium semi padat nitrogen free malat dan diinkubasi selama 48 jam. Apabila terdapat lapisan lendir maka isolat bakteri uji tersebut memiliki kemampuan dalam penambat nitrogen.
Identifikasi Agens AntagonisCendawan antagonis diidentifikasi
berdasarkan karakter morfologi menurut Watanabe (2002) dan Barnet dan Hunter (2006). Bakteri antagonis diidentifikasi menggunakan teknik polymerase chain reaction (PCR) dengan primer universal, yaitu forward primer 27F (5’-AGAGTTTGATCCTGGCTCAG-3’) dan reverse primer 1429R (5’-GGTTACC TTGTTACGACTT-3’) (Islam et al. 2012).
Proses amplifikasi yang dilakukan sebanyak 30 siklus pada kondisi pra PCR suhu 95 °C selama 5 menit, denaturasi suhu 95 °C selama 1 menit, aneling suhu 55 °C selama 5 menit, elongasi suhu 72 °C selama 1.5 menit, dan post-PCR pada suhu 72 °C selama 5 menit. Hasil amplifikasi DNA selanjutnya dirunutkan di laboratorium First Base, Malaysia. Data hasil perunutan dicocokkan dengan data GenBank National Center for Biotechnology Information (NCBI) menggunakan BLAST pada http://www.ncbi.nlm.nih.org.
Analisis DataPengujian antibiosis, uji produksi senyawa
volatil, dan pengujian secara in vivo disusun dalam rancangan acak lengkap. Masing-masing percobaan diulang 3 kali dengan menggunakan kontrol negatif. Isolat dengan persentase hambatan tertinggi pada uji antibiosis merupakan kandidat isolat unggul yang digunakan dalam seleksi selanjutnya. Pertumbuhan miselium pada pengujian in vivo dianalisis menggunakan ANOVA dengan program SAS versi 9.1. Perlakuan yang berpengaruh nyata diuji lanjut dengan duncan multiple range test pada taraf α 5%.
HASIL
Mikrob dari Jaringan Tanaman dan Rizosfer Tanaman Karet
Pada penelitian ini diperoleh 131 isolat bakteri dan 28 isolat cendawan yang terdiri atas 55 isolat bakteri dari jaringan tanaman, 76 isolat bakteri rizosfer, 15 isolat cendawan jaringan tanaman dan 13 isolat cendawan dari rizosfer. Berdasarkan pengujian patogenisitas sebanyak 99 isolat bakteri dan 18 isolat cendawan bersifat nonpatogen (Tabel 1).
Potensi Bakteri Antagonis dalam Menghambat Pertumbuhan R. lignosus
Hasil uji biakan ganda terhadap 99 isolat bakteri menunjukkan 2 isolat bakteri mampu menghambat pertumbuhan R. lignosus lebih dari 50%, yaitu bakteri endofit ME8 dan bakteri rizosfer MR3 (Gambar 1). Keduanya

J Fitopatol Indones Hardiyanti et al.
157
menghambat pertumbuhan miselium R. lignosus masing-masing hingga 100% secara in vivo pada akar tanaman karet (Tabel 2).
Potensi Cendawan Antagonis dalam Menghambat Pertumbuhan R. lignosus
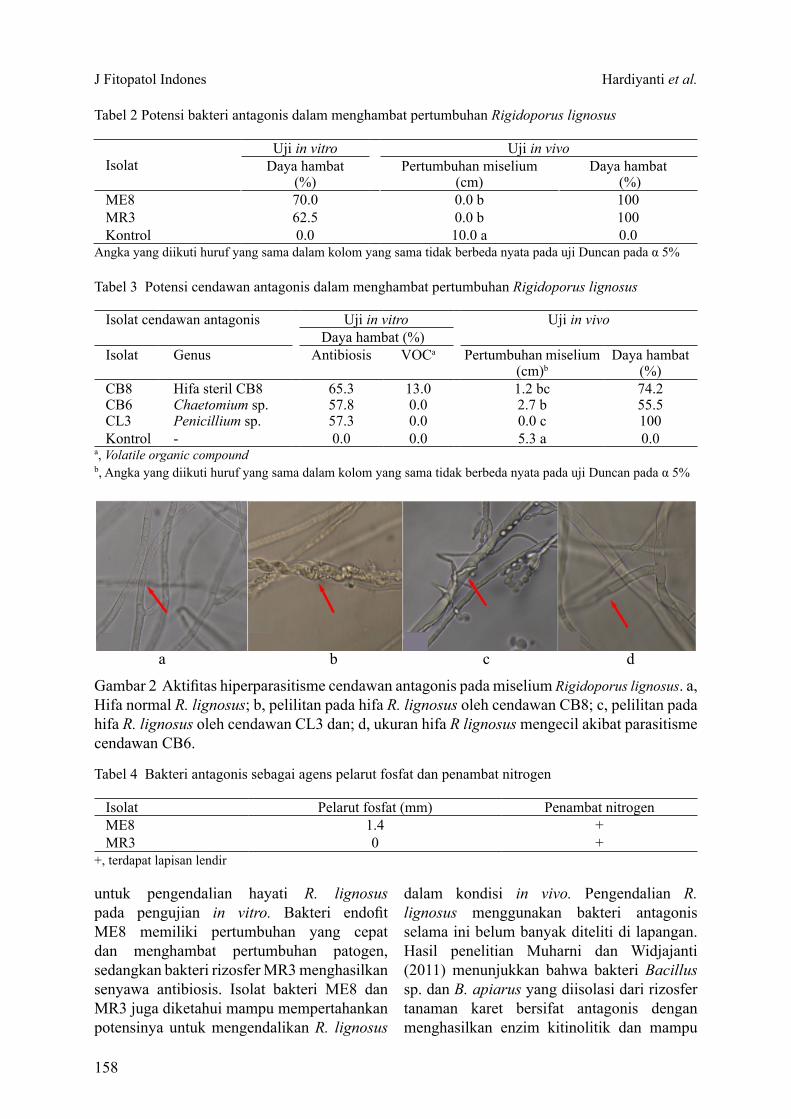
Berdasarkan uji biakan ganda diperoleh 3 cendawan endofit yang memiliki daya hambat tinggi, yaitu galur CB8, CB6, dan CL3. Ketiganya mampu menghambat pertumbuhan miselium lebih dari 50% pada pengujian in vivo. Pengujian produksi senyawa volatil menunjukkan bahwa hanya cendawan CB8 yang mampu menghambat pertumbuhan R. lignosus (Tabel 3). Cendawan CL3 dan CB8 memiliki aktivitas hiperparasitisme ditunjukkan dengan adanya pelilitan pada hifa R. lignosus, sementara cendawan CB6 menyebabkan ukuran hifa R. lignosus mengecil (Gambar 2).
Potensi Bakteri Antagonis sebagai Agens Pemacu Pertumbuhan
Bakteri ME8 mampu menambat fosfat yang ditandai oleh terbentuknya zona bening di sekitar koloni setelah 7 hari inkubasi. Hasil pengujian lainnya juga menunjukkan bakteri ME8 dan MR3 termasuk kelompok bakteri penambat nitrogen yang ditandai oleh terbentuknya lapisan lendir dan pelikel (Tabel 4).
Identifikasi Cendawan dan Bakteri Antagonis
Berdasarkan karaketristik morfologi, cendawan CB8 merupakan hifa steril, CB6 diidentifikasi sebagai Chaetomium sp., dan CL3 sebagai Penicillium sp. Berdasarkan perunutan gen 16S rRNA, isolat bakteri ME8 memiliki kekerabatan yang dekat dengan Bacillus siamensis B268 (no aksesi: KU644287), dan bakteri MR3 memiliki kekerabatan yang dekat dengan B. amylolyquifaciens SXAU001 (no aksesi: KY271752).
PEMBAHASAN
Mikrob antagonis yang telah diisolasi dari jaringan akar dan rizosfer tanaman karet memiliki potensi untuk dikembangkan sebagai agens hayati dalam mengendalikan R. lignosus. Hasil pengujian secara in vitro diperoleh isolat-isolat potensial yang memiliki beberapa mekanisme pengendalian patogen. Mekanisme bakteri antagonis meliputi kompetisi ruang dan nutrisi, memproduksi senyawa allelokimia, dan menginduksi ketahanan sistemik (ISR) tanaman inang (Compant et al. 2005). Suatu agens antagonis dapat memiliki 1 atau lebih mekanisme dalam mengendalikan patogen.
Isolat bakteri ME8 dan MR3 merupakan isolat bakteri antagonis yang potensial
Asal mikrob Mikrob total Mikrob nonpatogenikBakteri Cendawan Bakteri Cendawan
Jaringan Akar 55 15 42 10Rizosfer 76 13 57 8Total 131 28 99 18
Tabel 1 Jumlah mikrob yang diisolasi dari jaringan akar dan rizosfer tanaman karet
Gambar 1 Penghambatan pertumbuhan koloni Rigidoporus lignosus oleh mikrob antagonis. a, kontrol; b, bakteri endofit ME8; c, bakteri rizosfer MR3; d, cendawan endofit hifa steril CB8; e, cendawan endofit CB6; dan f, cendawan endofit CL3.
a b c d e f
J Fitopatol Indones Hardiyanti et al.
158
Tabel 3 Potensi cendawan antagonis dalam menghambat pertumbuhan Rigidoporus lignosus
Isolat cendawan antagonis Uji in vitro Uji in vivoDaya hambat (%)
Isolat Genus Antibiosis VOCa Pertumbuhan miselium (cm)b
Daya hambat (%)
CB8CB6CL3
Hifa steril CB8Chaetomium sp.Penicillium sp.
65.357.857.3
13.00.00.0
1.2 bc2.7 b0.0 c
74.255.5100
Kontrol - 0.0 0.0 5.3 a 0.0a, Volatile organic compoundb, Angka yang diikuti huruf yang sama dalam kolom yang sama tidak berbeda nyata pada uji Duncan pada α 5%
Gambar 2 Aktifitas hiperparasitisme cendawan antagonis pada miselium Rigidoporus lignosus. a, Hifa normal R. lignosus; b, pelilitan pada hifa R. lignosus oleh cendawan CB8; c, pelilitan pada hifa R. lignosus oleh cendawan CL3 dan; d, ukuran hifa R lignosus mengecil akibat parasitisme cendawan CB6.
a b c d
Tabel 4 Bakteri antagonis sebagai agens pelarut fosfat dan penambat nitrogen
Isolat Pelarut fosfat (mm) Penambat nitrogenME8 1.4 +MR3 0 +
+, terdapat lapisan lendir
Tabel 2 Potensi bakteri antagonis dalam menghambat pertumbuhan Rigidoporus lignosus
IsolatUji in vitro Uji in vivo
Daya hambat (%)
Pertumbuhan miselium (cm)
Daya hambat (%)
ME8 70.0 0.0 b 100MR3 62.5 0.0 b 100Kontrol 0.0 10.0 a 0.0
Angka yang diikuti huruf yang sama dalam kolom yang sama tidak berbeda nyata pada uji Duncan pada α 5%
untuk pengendalian hayati R. lignosus pada pengujian in vitro. Bakteri endofit ME8 memiliki pertumbuhan yang cepat dan menghambat pertumbuhan patogen, sedangkan bakteri rizosfer MR3 menghasilkan senyawa antibiosis. Isolat bakteri ME8 dan MR3 juga diketahui mampu mempertahankan potensinya untuk mengendalikan R. lignosus
dalam kondisi in vivo. Pengendalian R. lignosus menggunakan bakteri antagonis selama ini belum banyak diteliti di lapangan. Hasil penelitian Muharni dan Widjajanti (2011) menunjukkan bahwa bakteri Bacillus sp. dan B. apiarus yang diisolasi dari rizosfer tanaman karet bersifat antagonis dengan menghasilkan enzim kitinolitik dan mampu

J Fitopatol Indones Hardiyanti et al.
159
menghambat pertumbuhan R. lignosus sebesar 6.12 mm pada pengujian in vitro. Bakteri B. amyloliquefaciens diketahui menghasilkan senyawa surfactin, iturin, bacilomycine, azalomycin, acivicin, arthrobactin, rhodutorola acid, valinomycin, stenothricin, enterochelin, dan nocardamin (Wulff et al. 2002).
Kemampuan bakteri dalam melarutkan fosfat dan memfiksasi nitrogen akan memengaruhi kebugaran tanaman inang sehingga lebih toleran terhadap penyakit. Singh et al. (2010) menyatakan bahwa aplikasi bakteri pelarut fosfat memiliki korelasi positif terhadap performa tanaman dengan meningkatkan pertumbuhan dan meningkatkan hasil panen tanaman kedelai sebesar 23.88%. Inokulasi bakteri fiksasi nitrogen pada tanaman tomat dan cabai merah diketahui mampu meningkatkan klorofil tanaman, hara makro dan mikro tanaman, vigor, dan biomassa tanaman (Islam et al. 2012).
Selain bakteri antagonis diperoleh juga cendawan dari jaringan akar yang mampu menghambat pertumbuhan R. lignosus, yaitu cendawan endofit hifa steril CB8, cendawan endofit CB6, dan cendawan endofit CL3. Soytong et al. (2001) menyatakan bahwa Chaetomium globosum menghasilkan senyawa chaetoglobosin dan chaetoviridins yang berperan dalam menekan pertumbuhan patogen.
Cendawan endofit CL3 dan hifa steril CB8 memiliki aktivitas hiperparasitisme dan mengakibatkan hifa R. lignosus menjadi lisis, keriput, dan membengkak. Interaksi hiperparasitisme cendawan antagonis Pythium radiosum terhadap Botrytis cinerea menyebabkan koagulasi protoplasma, hifa menjadi kosong, dan kerusakan hifa ini dapat menyebabkan percabangan dan produksi struktur seksual (Paul 1999). Selain bersifat antibiosis, cendawan hifa steril CB8 juga menghasilkan senyawa volatil. Cendawan endofit dapat memproduksi asam volatil seperti alkohol, ester, keton, dan lipid yang mampu menghambat Fusarium solani, Pythium ultimun, dan Rhizoctonia solani (Strobel 2001).
Cendawan antagonis yang telah dimanfaatkan dalam mengendalikan R. lignosus sebagian besar merupakan genus Trichodema. Aplikasi T. harzianum pada potongan akar yang terinfeksi R. lignosus mengakibatkan pemulihan akar hingga 72% jika akar terinfeksi ringan, sementara jika terinfeksi berat hanya dapat memulihkan kondisi akar sebesar 23.6% (Jayasuriya dan Thenaakkon 2007). Hypocrea atroviridis yang diisolasi dari perkebunan karet di Sumatra dapat menghambat pertumbuhan patogen sebesar 77.93% pada 7 hari setelah inokulasi (Amaria et al. 2015). C. cupreum yang diformulasikan dalam bentuk tepung dan minyak mampu mengurangi insidensi penyakit akar putih pada karet sebesar 60−80% (Soytong dan Kaewchai 2014).
Hasil penelitian ini membuktikan bahwa bakteri endofit ME8, bakteri rizosfer MR3, cendawan endofit CB8, cendawan endofit CB6, dan cendawan endofit CL3 yang diisolasi dari jaringan akar dan rizosfer tanaman karet memiliki potensi sebagai agens hayati untuk mengendalikan penyakit akar putih.
DAFTAR PUSTAKA
Amaria W, Harni R, Samsudin. 2015. Evaluasi jamur antagonis dalam menghambat pertumbuhan Rigidoporus microporus penyebab penyakit jamur akar putih pada tanaman karet. JTIDP. 2(1):51−60.
Barnet HL, Hunter BB. 2006. Illustrated Genera of Imperfect Fungi. Ed ke-4. Amerika (US): APS Press.
Compant S, Duffy B, Nowak J, Clement C, Barkal EA. 2005. Use of plant growth-promoting bacteria for biocontrol of plant disease: principle, mechanisms of action, and future prospects. Appl Environ Microbiol. 71:4951−4959. DOI: https://doi.org/10.1128/AEM.71.9.4951-4959.2005.
Islam MR, Sultana T, Joe MM, Yim W, Cho JC, Sa T. 2012. Nitrogen-fixing bacteria with multiple plant growth-promoting activities enhance growth of tomato and red pepper.

J Fitopatol Indones Hardiyanti et al.
160
J Bas Microbiol. 53(1004):1−25. DOI: https://doi.org/10.1002/jobm.201200141.
Isnaini N. 2016. Pengendalian JAP pada Tanaman Karet di Indonesia dengan Dana APBN-TP 2016. www.ditjenbun.pertanian.go.id. [diakses 5 Oktober 2016].
Jayasuriya KE, Thenaakoon BI. 2007. Biological control of Rigidoporus lignosus the cause of white root disease in rubber. J Bio Sci. 36(1):9−16.
Muharni, Widjajanti H. 2011. Skrining bakteri kitinolitik antagonis terhadap pertumbuhan jamur akar putih (Rigidoporus lignosus) dari rizosfir tanaman karet. J Penel Sains. 14(1):14112−14151.
Omo-Ikerodah EE, Omorusi VI, Mokwunye MUB. 2012. Challenges and progress in the control of white root rot disease of Hevea brasiliensis in Africa. World Rural Observ. 4(1):1−2.
Paul B. 1999. Suppression of Botrytis cinera causing the grey mould disease of grape-vine by an aggressive mycoparasite, Phytium radiosum. FEMS Microbiol. 176:25−30. DOI: https://doi.org/10.1111/j.1574-6968.1999.tb13637.x.
Singh AV, Shah S, Prasad B. 2010. Effect of phosphate solubilizing bacteria on plant growth promotion and nodulation in soybeab (Glycine max (L.) Merr.). J Hill Agric. 1(1):35−39.
Soytong K, Kaewchai S. 2014. Biological control of white root of rubber trees using Chaetomium cupreum. J Agric Tech. 10(1):93−103.
Soytong K, Kanokmedhakul S, Kukongviriapa V, Isobe M. 2001. Application of Chaetomium spesies (Ketomuim®) as a new broad spectrum biological fungicide for plant disease control: Fungal Divers 7: 1−15.
Strobel GA, Dirkse E, Sears J, Markworth C. 2001. Volatile antimicrobials from Muscodor albus a novel endophytic fungus. Microbiology. 147:2943−2950. DOI: https://doi.org/10.1099/00221287-147-11-2943.
Watanabe T. 2002. Pictorial Atlas of Soil and Seed Fungi. Ed ke-2. Washington (US): CRC Pr. DOI: https://doi.org/10.1201/9781420040821.
Wulff EG, Mguni CM, Mansfeld-Giese K, Fels J, Lubeck M, Hockenhull J. 2002. Biochemical and molecular characterization of Bacillus amyloliquefaciens, B. subtilis and B. pumilus isolates with distinct antagonistic potential against Xanthomonas campestris pv. campestris. Plant Pathol. 5:574–584. DOI: https://doi.org/10.1046/j.1365-3059.2002.00753.x.

ISSN: 0215-7950
161
Volume 13, Nomor 5, September 2017Halaman 161–167
DOI: 10.14692/jfi.13.5.161
*Alamat penulis korespondensi: Program Studi Agroteknologi, Fakultas Teknologi Industri, Universitas Gunadarma. Jalan Margonda Raya 100 Pondok Cina, Depok 16424. Tel: 021-78881112, Faks: 021-78881071, Surel: [email protected]
Potensi Cendawan Endofit sebagai Pengendali Hayati Penyakit Busuk Pangkal Batang (Phytophthora capsici) pada Bibit Cabai
Potency of Endophytic Fungi as Biocontrol Agents of Stem Rot Disease (Phytophthora capsici) on Chili Pepper Seedlings
Evan Purnama Ramdan1*, Efi Toding Tondok2, Suryo Wiyono2,Sri Hendrastuti Hidayat2, Widodo2
1Universitas Gunadarma, Depok 164242Institut Pertanian Bogor, Bogor 16680
ABSTRAK
Penyakit busuk pangkal batang yang disebabkan oleh Phytophthora capsici merupakan penyakit penting pada pertanaman cabai. Sebanyak 8 cendawan endofit yang telah diisolasi dan lolos uji patogenisitas, diuji potensinya sebagai agens hayati penyakit busuk batang pada bibit cabai. Suspensi cendawan endofit diinokulasikan sebanyak 2 kali, yaitu dengan perendaman 100 butir benih dalam 100 mL suspensi dan penyiraman 10 mL suspensi per bibit cabai berumur 3 minggu setelah semai. Konsentrasi cendawan endofit yang digunakan ialah 2.8 × 106 cfu mL-1. Insidensi penyakit dan nilai area under the disease progress curve (AUDPC) dihitung selama 4 minggu setelah inokulasi P. capsici. Uji penghambatan pertumbuhan P. capsici dilakukan secara in vitro dan kemampuan kolonisasi cendawan endofit diamati pada bibit cabai umur 4 minggu. Delapan galur cendawan endofit menunjukkan penghambatan terhadap pertumbuhan P. capsici secara in vitro, di antaranya Penicillium galur MAG1 dan Penicillium galur PAB2 menunjukkan mekanisme antibiosis. Cendawan endofit memiliki kemampuan mengolonisasi pada akar (26–60%) lebih besar daripada batang (20–40%). Fusarium galur MAGR1 merupakan galur yang paling besar tingkat kolonisasinya dibandingkan dengan galur cendawan endofit lainnya, yaitu sebesar 60%. Pada pengujian secara in vivo diperoleh enam cendawan endofit (Fusarium galur MAGR1, Penicillium galur MAG1, Penicillium galur PAB2, hifa steril HAJ1, hifa steril HAJ2, dan hifa steril PBG7) yang berpotensi mengendalikan penyakit busuk pangkal batang dengan penekanan sebesar 25.5–35.5%.
Kata kunci: agens hayati, antibiosis, Capsicum annum, kolonisasi akar
ABSTRACT
Stem rot disease caused by Phytophthora capsici is an important disease on chilli. Eight endophytic fungi that had been isolated and screened based on pathogenecity test were further tested for their potential as the biological control agent of the stem rot disease of chilli. The endophytic fungi suspension was applied twice during the trial. The first application was on 100 seed lot, by soaking them in 100 mL of suspension. The second application was on the 3 weeks-old chili seedlings by drenching them with 10 mL suspension per plant. The concentration of endophytic fungi in the suspension was 2.8 × 106 cfu mL-1. The disease intensity and AUDPC value were measured for 4 weeks after the pathogen inoculation. The growth inhibition test of P. capsici was performed in vitro and the colonization abilities of endophytic fungi were observed at 4 weeks-old chilli seedlings. Eight endophytic fungi inhibited the growth of the P. capsici, and two of those isolates namely Penicillium strain MAG1 and Penicillium strain PAB2 showed antibiosis mechanism. Endophytic fungi has the ability more to colonize at the

J Fitopatol Indones Ramdan et al.
162
root (26–60%) than in the stem (20–40%). Fusarium strain MAGR1 has the highest level of endophytic colonization i.e. 60% compared to others. Based on in vivo assay, six endophytic fungi isolates, i.e. Fusarium strain MAGR1, Penicillium strain MAG1, Penicillium strain PAB2, sterile hyphae HAJ1, sterile hyphae HAJ2, and sterile hyphae PBG7, showed the potency to control stem rot disease with inhibition level of 25.5–35.5%.
Key words: antibiosis, biocontrol agent, Capsicum annum, root colonization
PENDAHULUAN
Tanaman cabai merupakan komoditas hortikultura utama yang banyak ditanam di seluruh dunia, khususnya di Asia. Salah satu kendala budi daya tanaman ini ialah adanya penyakit busuk pangkal batang yang disebabkan Phytophthora capsici. Penyakit ini dapat menyerang tanaman dari pembibitan sampai tanaman tua. Penyakit ini mempunyai arti ekonomi penting dan menyebabkan kerusakan serius pada tanaman cabai di seluruh dunia (Lamour et al. 2012). Pada tanaman rentan, P. capsici menyebabkan kematian pada hari kesepuluh setelah inokulasi, sedangkan pada tanaman tahan menyebabkan timbulnya luka kecokelatan di bagian akar sekunder pada waktu yang sama (Dunn dan Smart 2015).
Infeksi P. capsici sulit dikendalikan, karena perkembangan awal dari penyakit ini sulit dideteksi dan penyakit cepat menyebar saat kondisi lingkungan mendukung. Oleh karena itu, strategi dan teknik pengendaliannya perlu dikembangkan, di antaranya ialah menggunakan cendawan endofit. Cendawan endofit asal tanaman cabai telah dilaporkan dapat menekan insidensi penyakit antraknosa sebesar 33–43%, dan penyakit layu bakteri lebih dari 50% (Irawati et al. 2016). Mekanisme penekanan penyakit oleh cendawan endofit, yaitu melalui produksi metabolit sekunder (Tellenbach et al. 2013), antibiotik (You et al. 2009), serta ketahanan tanaman melalui peningkatan aktivitas peroksidase dan fenilalanin amoniliase (Redman et al. 1999).
Cendawan endofit telah dilaporkan mempunyai aktivitas antagonis terhadap P. capsici secara in vitro (Paul et al. 2012), sedangkan pada pengujian in vivo mampu memperlambat laju infeksi dan menekan intensitas serangan P. capsici pada
tanaman lada (Kusumawardani et al. 2015). Sementara ini belum ditemukan laporan ilmiah penggunaan cendawan endofit untuk mengendalikan penyakit busuk P. capsici pada tanaman cabai. Penelitian ini bertujuan menguji potensi cendawan endofit dalam menekan penyakit busuk pangkal batang pada pembibitan cabai.
BAHAN DAN METODE
Uji in Vitro Cendawan Endofit Delapan cendawan endofit (Aspergillus
galur HAG1, Fusarium galur MAGR1, Penicillium galur MAG1, Penicillium galur PAB2, hifa steril isolat HAJ1, HAJ2, PBG7, dan isolat CBG5) yang digunakan merupakan kultur koleksi dari Laboratorium Mikologi Tumbuhan IPB yang telah teruji tidak patogenik pada bibit cabai serta memiliki potensi sebagai agens pemacu pertumbuhan (Ramdan et al. 2013). Isolat P. capsici berasal dari koleksi Klinik Tanaman IPB.
Aktivitas penghambatan cendawan endofit terhadap pertumbuhan P. capsici diuji menggunakan metode biakan ganda pada medium agar-agar dekstrosa kentang (ADK), mengikuti metode Li et al. (2007) yang dimodifikasi waktu inkubasinya. Waktu inkubasi diubah dari 1–3 minggu menjadi 3–4 hari pada suhu ruang, karena cendawan endofit yang diuji lebih cepat pertumbuhannya.
Pengujian disusun dalam rancangan acak lengkap, dengan ulangan 4 kali. Pada akhir inkubasi, peubah yang diamati ialah diameter koloni P. capsici dan penghambatan pertumbuhan P. capsici yang dihitung menggunakan rumus berikut:
(r1– r2) r1Persentase penghambatan = × 100%

J Fitopatol Indones Ramdan et al.
163
r1, diameter koloni P. capsici pada kontrol; dan r2, diameter koloni P. capsici yang diberi perlakuan cendawan endofit .
Penghambatan P. capsici dikelompokkan sebagai mekanisme antibiosis jika terdapat zona hambat antara pertumbuhan miselium cendawan endofit dan P. capsici, serta kompetisi jika pertumbuhan cendawan endofit mendominasi medium ADK.
Uji in Vivo Cendawan Endofit Kemampuan kolonisasi cendawan endofit
dan penekanan penyakit busuk pangkal batang pada bibit cabai merah varietas Gelora disusun dalam rancangan acak kelompok dengan 4 ulangan, tiap ulangan terdiri atas 20 bibit cabai. Sebanyak 10 mL akuades steril ditambahkan pada cendawan endofit yang ditumbuhkan pada medium ADK dan telah membentuk massa konidium. Pemanenan konidium dilakukan dengan menggoreskan jarum ose untuk memisahkan konidium dari medium. Cendawan endofit yang berupa hifa steril ditumbuhkan pada medium dekstrosa kentang cair (DKC) yang diinkubasikan selama 7 hari pada inkubator bergoyang dengan kecepatan 130 rpm. Kumpulan miselium yang tumbuh dipisahkan dari DKC dan dibilas akuades steril sebanyak 3 kali. Miselium dimasukkan ke dalam 100 mL akuades steril, kemudian dihancurkan menggunakan homogenizer Ika Ultra Turrax T18 Basic pada kecepatan 3500 sampai 24 000 rpm selama 5 menit hingga didapatkan suspensi yang relatif homogen. Konsentrasi cendawan endofit yang digunakan ialah 2.8 × 106 cfu mL-1.
Inokulasi cendawan endofit dilakukan 2 tahap, tahap ke-1 dengan perendaman benih dan tahap ke-2 penyiraman bibit. Benih terlebih dahulu disterilisasi permukaan menggunakan NaOCl 1% selama 2 menit, kemudian dibilas akuades steril sebanyak 2 kali. Setiap 100 butir benih direndam dalam 100 mL suspensi cendawan endofit selama 15 jam, kemudian disemai pada baki semai dengan medium semai komersial. Penyiraman suspensi cendawan endofit pada bibit cabai berumur 3 minggu setelah semai (MSS) sebanyak 10 mL
per tanaman. Pada kontrol digunakan akuades steril untuk perendaman dan penyiraman.
Inokulum P. capsici yang dipakai untuk pengujian berupa suspensi zoospora. Zoospora diproduksi mengikuti protokol Asian Vegetable Research and Development Center (AVRDC) seperti yang dilaporkan oleh Yunianti (2007). Sebanyak 5 × 105 cfu mL-1 suspensi zoospora dipakai untuk perlakuan. Inokulasi dilakukan pada bibit cabai berumur 4 MSS dengan cara menyiramkan 5 mL suspensi pada daerah sekitar pangkal batang bibit cabai. Perkembangan insidensi penyakit diamati tiap minggu sejak munculnya gejala. Insidensi penyakit (IP) dihitung dengan menggunakan rumus berikut:
IP = nN × 100%, dengan
n, jumlah tanaman yang terinfeksi dan N, jumlah tanaman yang diamati.
Aktivitas Kolonisasi Cendawan Endofit Pada Bibit Cabai.
Bibit cabai umur 4 MSS yang telah diinokulasi cendawan endofit diambil sebanyak 5 tanaman. Reisolasi cendawan endofit dari bagian akar dan pangkal batang mengikuti metode Rodrigues (1994) yang dimodifikasi pada sterilisasi permukaan bagian tanaman. Konsentrasi NaOCl untuk sterilisasi permukaan akar sebesar 1% dan batang sebesar 3%. Isolasi pada akar dilakukan dengan memilih secara acak bagian akar primer, akar lateral, rambut akar, dan ujung akar sebanyak 5 potongan, sedangkan isolasi pada pangkal batang diambil 5 cm dari pangkal batang. Pengamatan dilakukan terhadap koloni cendawan endofit yang keluar dari potongan sampel tanaman, kemudian dilakukan konfirmasi secara morfologi. Penghitungan kolonisasi cendawan endofit dilakukan dengan rumus:
Kolonisasi CE=∑ n∑ N × 100%, dengan
n, jumlah potongan sampel tanaman yang terinfeksi cendawan endofit dan N, jumlah potongan sampel tanaman yang diamati.
J Fitopatol Indones Ramdan et al.
164
Analisis DataData hasil pengamatan dianalisis meng-
gunakan uji analisis ragam menggunakan program SAS versi 9.1. Apabila rerata menunjukkan adanya perbedaan nyata maka dilanjutkan dengan uji Tukey pada α 5% untuk pengujian in vitro dan diuji lanjut dengan uji Duncan pada α 5% untuk pengujian in vivo.
HASIL
Penghambatan Pertumbuhan P. capsici in Vitro
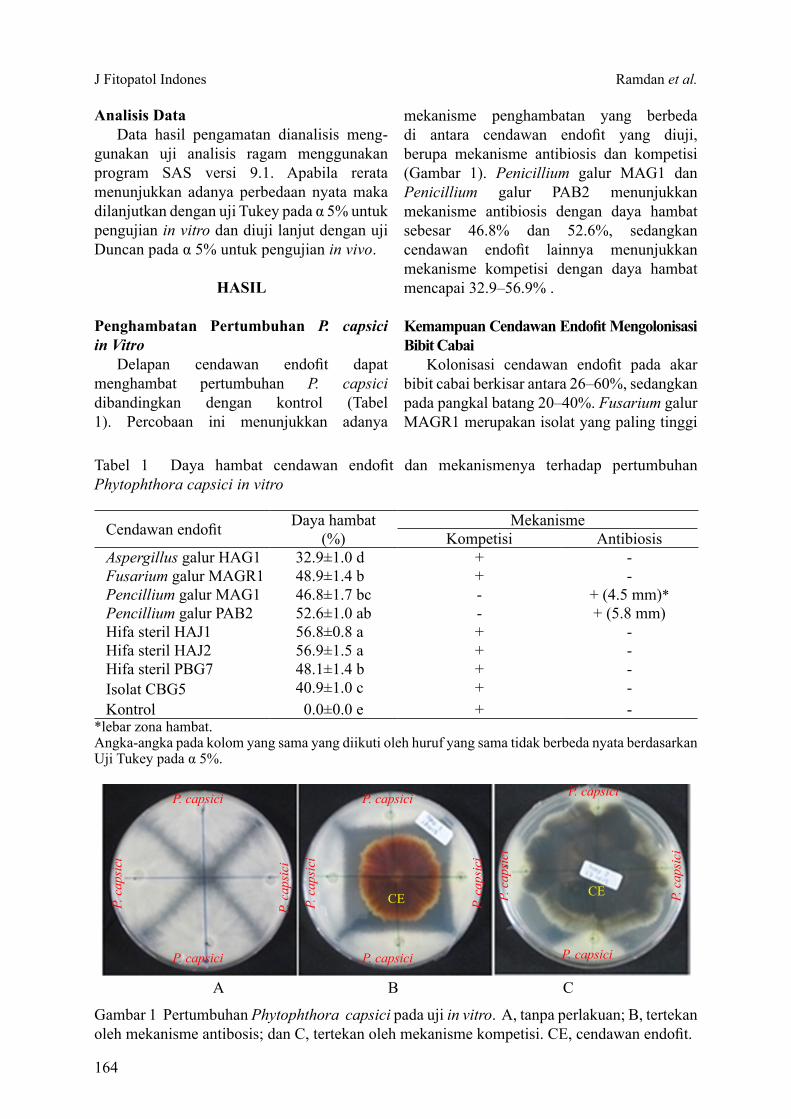
Delapan cendawan endofit dapat menghambat pertumbuhan P. capsici dibandingkan dengan kontrol (Tabel 1). Percobaan ini menunjukkan adanya
mekanisme penghambatan yang berbeda di antara cendawan endofit yang diuji, berupa mekanisme antibiosis dan kompetisi (Gambar 1). Penicillium galur MAG1 dan Penicillium galur PAB2 menunjukkan mekanisme antibiosis dengan daya hambat sebesar 46.8% dan 52.6%, sedangkan cendawan endofit lainnya menunjukkan mekanisme kompetisi dengan daya hambat mencapai 32.9–56.9% .
Kemampuan Cendawan Endofit Mengolonisasi Bibit Cabai
Kolonisasi cendawan endofit pada akar bibit cabai berkisar antara 26–60%, sedangkan pada pangkal batang 20–40%. Fusarium galur MAGR1 merupakan isolat yang paling tinggi
Tabel 1 Daya hambat cendawan endofit dan mekanismenya terhadap pertumbuhan Phytophthora capsici in vitro
Cendawan endofit Daya hambat (%)
MekanismeKompetisi Antibiosis
Aspergillus galur HAG1 32.9±1.0 d + -Fusarium galur MAGR1 48.9±1.4 b + -Pencillium galur MAG1 46.8±1.7 bc - + (4.5 mm)*Pencillium galur PAB2 52.6±1.0 ab - + (5.8 mm)Hifa steril HAJ1 56.8±0.8 a + -Hifa steril HAJ2 56.9±1.5 a + -Hifa steril PBG7 48.1±1.4 b + -Isolat CBG5 40.9±1.0 c + -Kontrol 0.0±0.0 e + -
*lebar zona hambat.Angka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan Uji Tukey pada α 5%.
Gambar 1 Pertumbuhan Phytophthora capsici pada uji in vitro. A, tanpa perlakuan; B, tertekan oleh mekanisme antibosis; dan C, tertekan oleh mekanisme kompetisi. CE, cendawan endofit.
A B C
P. capsici P. capsici P. capsici
CE CE
P. capsiciP. capsici P. capsici
P. c
apsi
ci
P. c
apsi
ci
P. c
apsi
ci
P. c
apsi
ci
P. c
apsi
ci
P. c
apsi
ci
J Fitopatol Indones Ramdan et al.
165
persentase kolonisasi di bagian akar jika dibandingkan dengan perlakuan lain, yaitu sebesar 60%. Meskipun tidak berbeda nyata antar perlakuan, Fusarium galur MAGR1 dan hifa steril isolat HAJ2 merupakan cendawan yang lebih tinggi kolonisasinya pada bagian batang, yaitu sebesar 40% (Gambar 2).
Penyakit Busuk Pangkal Batang pada Bibit Cabai
Pada akhir pengamatan in vivo, insidensi penyakit busuk pangkal batang yang diberi perlakuan cendawan endofit Fusarium galur MAGR1, Penicillium galur PAB2, hifa steril isolat HAJ1 dan PBG7 berbeda nyata dibandingkan dengan kontrol (Tabel 2). Hal tersebut berkorelasi dengan nilai AUDPC selama periode pengamatan dari
masing-masing perlakuan yang rendah dan berbeda nyata berdasarkan analisis ragam dibandingkan dengan kontrol. Pada akhir pengamatan, cendawan endofit memiliki efikasi atau kemampuan penekanan penyakit busuk pangkal sebesar 13.7–35.5%.
PEMBAHASAN
Pada penelitian ini Penicillium galur MAG1 dan Penicillium galur PAB2 menunjukkan adanya penghambatan pertumbuhan P. capsici secara antibiosis yang ditunjukkan dengan adanya zona hambat dengan lebar 4.5–4.8 mm. Hal ini sesuai dengan pendapat Wakana et al. (2016) yang menyatakan bahwa P. citirin memiliki senyawa antifungi seperti citrinin yang ditunjukkan dengan adaya zona hambat
Tabel 2 Pengaruh cendawan endofit terhadap insidensi penyakit busuk pangkal batang
Cendawan endofit Insidensi penyakit (%)*
Nilai AUDPC
Penekanan penyakit di akhir pengamatan (%)
Aspergillus galur HAG1 70.0 ± 4.0 ab 106.5 b 17.7Fusarium galur MAGR1 63.3 ± 5.7 b 90.0 b 25.5Pencillium galur MAG1 61.7 ± 5.7 b 90.8 b 27.5Pencillium galur PAB2 65.0 ± 4.5 ab 97.5 b 35.5Hifa steril HAJ1 61.7 ± 4.7 b 89.2 b 27.5Hifa steril HAJ 2 65.0 ± 6.0 ab 95.2 b 23.5Hifa steril PBG7 63.3 ± 5.7 b 90.8 b 25.5Isolat CBG5 73.3 ± 3.6 ab 103.3 b 13.7Kontrol 85.0 ± 2.7 a 144.0 a 0.0
*Angka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata berdasarkan Uji Duncan pada α 5%
Gambar 2 Kolonisasi cendawan endofit pada bagian akar ( ) dan batang ( ) bibit cabai. Huruf yang sama di atas grafik batang menunjukkan tidak berbeda nyata berdasarkan Uji Tukey pada α 5%).
Isolat Cendawan Endofit
Ting
kat K
olon
isas
i (%
)
Isolat CBG 5Aspergillus sp. HAG1
Fusarium sp. MAGR1
Pencillium sp. MAG1
Pencillium sp.PAB2
Hifa sterilHAJ1
Hifa steril HAJ2
Hifa steril PBG7
70
60
50
40
30
20
10
0
a
aab
a ab
a
ab
a
ab
ab
ab
a
ab
b
abb

J Fitopatol Indones Ramdan et al.
166
sebesar 13–17 mm pada pengujian terhadap Candida albicans dan Aspergillus fumigatus. Biakan ganda ujiantara Penicillium dan P. capsci menunjukkan adanya abnormalitas miselium P. capsici, yaitu miselium menjadi lebih tebal, ujung hifa memilin atau memutar dan bercabang (Ma et al. 2008).
Tingkat kolonisasi cendawan endofit ditemukan lebih tinggi pada bagian akar dibandingkan dengan pada bagian batang. Hal ini diduga berkaitan dengan perlakuan perendaman benih dan penyiraman cendawan endofit yang dilakukan di sekitar perakaran. Windriyati (2015) juga melaporkan bahwa cendawan endofit asal cabai memiliki tingkat kolonisasi pada akar lebih tinggi (40–52%) dibandingkan dengan pada bagian batang (20–28%). Pada pengujian ini Fusarium galur MAGR1 merupakan cendawan yang memiliki kemampuan mengolonisasi akar paling tinggi karena pertumbuhannya cepat dan mampu menyebar secara sistemik di dalam jaringan tanaman. Istikorini (2008) menyatakan bahwa Fusarium memiliki kemampuan mengolonisasi yang baik pada akar, batang, dan daun cabai.
Efikasi cendawan endofit untuk menekan penyakit busuk pangkal batang masih rendah, yaitu 13.7–35.5%. Hal ini berkaitan dengan masih rendahnya kolonisasi pada akar dan batang yaitu berkisar 20–60%. cendawan endofit yang mampu mengolonisasi di semua bagian tanaman dengan cepat akan mampu berkompetisi dengan mikroorganisme lain sehingga berpotensi sebagai agens pengendali hayati yang efektif. Selain itu, di alam cara kerja cendawan endofit tidaklah tunggal tetapi dalam komunitas untuk menghasilkan multi-mekanisme yang melindungi inangnya dari lingkungan biotik dan abiotik yang tidak mendukung (Malinowski dan Belesky 2000).
Berdasarkan pengamatan dari serangkaian pengujian in vitro dan in vivo diperoleh enam cendawan endofit (Fusarium galur MAGR1, Penicillium galur MAG1, Penicillium galur PAB2, hifa steril isolat HAJ1, HAJ2 dan PBG7) yang memiliki potensi sebagai agens pengendali penyakit busuk pangkal batang. Potensi cendawan endofit tersebut dapat
dimaksimalkan untuk menekan penyakit melalui kombinasi perlakuan, cara aplikasi yang tepat, formulasi yang mendukung dan perawatan tanaman yang tepat.
UCAPAN TERIMA KASIH
Penelitian ini didanai melalui Hibah Penelitian Unggulan Strategis Nasional, Direktorat Perguruan Tinggi, Kementerian Pendidikan dan Kebudayaan Republik Indonesia tahun 2011–2012.
DAFTAR PUSTAKA
Dunn AR, Smart CH. 2015. Interactions of Phytophthora capsici with resistant and susceptible pepper roots and stems. Mycology. 105(10):1355–1361. DOI: https://doi.org/10.1094/PHYTO-02-15-0045-R.
Irawati AFC, Sastro Y, Sulastri, Suhartono MT, Mutaqin KH, Widodo. 2016. Cendawan endofit yang potensial meningkatkan ketahanan cabai merah terhadap penyakit layu bakteri. J Fitopatol Indones.12(4):133–141.
Istikorini Y. 2008. Potensi cendawan endofit untuk mengendalikan penyakit antraknosa pada cabai (Capsicum annum L.) [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Kusumawardani Y, Sulistyowati L, Cholil A. 2015. Potensi antagonis jamur endofit pada tanaman lada (Piper nigrum L.) terhadap jamur Phytophthora capsici Leonian penyebab busuk pangkal batang. JHPT. 3(1):21–29.
Lamour KH, Stam R, Jupe J, Huitema E. 2012. The oomycete broad-host range pathogen Phytophthora capsici. Mol Plant Pathol. 13:329–337. DOI: https://doi.org/10.1111/j.1364-3703.2011.00754.x.
Li C, Gao J, Nan Z. 2007. Interactions of Neotyphodium gansuense, Achnatherum inebrians, and plant-pathogenic fungi. Mycol Res. 3:1220–1227. DOI: https://doi.org/10.1016/j.mycres.2007.08.012.

J Fitopatol Indones Ramdan et al.
167
Ma Y, Chang Z, Zhao J, Zhou M. 2008. Antifungal activity of Penicillium striatisporum Pst10 and its biocontrol effect on Phytophthora root rot of chilli pepper. Biol Cont. 44:24–31. DOI: https://doi.org/10.1016/j.biocontrol.2007.10.005.
Malinowski DP, Belesky DP. 2000. Review and interpretation: adaptations of endophyte-infected cool-season grasses to environmental stresses: mechanisms of drought and mineral stress tolerance. Crop Sci. 40(4):923–940. DOI: https://doi.org/10.2135/cropsci2000.404923x.
Paul NC, Deng JX, Sang HK, Choi YP, Yu SH. 2012. Distribution and antifungal activity of endophytic fungi in different growth stages of chili pepper (Capsicum annuum L.) in Korea. Plant Pathol J. 28:10–19. DOI: https://doi.org/10.5423/PPJ.OA.07.2011.0126.
Ramdan EP, Widodo, Tondok ET, Wiyono S, Hidayat SH. 2013. Cendawan endofit nonpatogen asal tanaman cabai dan potensinya sebagai agens pemacu pertumbuhan. J Fitopatol Indones. 9(5):138–144. DOI: https://doi.org/10.14692/jfi.9.5.139.
Redman RS, Freeman S, Clifton DR, Morrel J, Brown G, Rodrigues R. 1999. Biochemical analysis of plant protection afforded by a nonpathogenic endophytic mutant of Colletotrichum magna. Plant Physiol. 115:795–804. DOI: https://doi.org/10.1104/pp.119.2.795.
Rodrigues KF. 1994. The foliar fungal endophytes of the Amazonian palm. Euterpe oleracea. Mycologia. 86:376–385. DOI: https://doi.org/10.2307/3760568.
Tellenbach C, Sumarah MW, Grunig CR, Miller JD. 2013. Inhibition of Phytophthora species by secondary metabolites produced by the dark sepatate endophyte Phialocephala europaea. Fungal Ecol. 6:12–18. DOI: https://doi.org/10.1016/j.funeco.2012.10.003.
Wakana D, Hosoe T, Itabashi T, Okada K, Takiki CM, Yaguchi T, Fukusima K, Kawai K. 2006. New citrin derivates isolated from Penicillium citrinum. J Nat Med. 60:279–284. DOI: https://doi.org/10.1007/s11418-006-0001-2.
Windriyati Y. 2015. Seleksi cendawan endofit untuk pengendalian penyakit layu bakteri (Ralstonia solanacearum) pada tanaman cabai [tesis]. Bogor (ID): Institut Pertanian Bogor.
You F, Han T, Wu JZ, Huang BK, Qin LP. 2009. Antifungal secondary metabolites from endophytic Verticillium sp. Biochem Syst Ecol. 37:162–165. DOI: https://doi.org/10.1016/j.bse.2009.03.008.
Yunianti R, Sastrosumarjo S, Sutjiprihati S, Surahman M, Hidayat SH. 2007. Ketahanan 22 genotipe cabai (Capsicum spp) terhadap Phytophthora capsici Leonan dan keanekaragaman genetiknya. Bul Agron. 35:103–111.

ISSN: 0215-7950
168
Volume 13, Nomor 5, September 2017Halaman 168–174
DOI: 10.14692/jfi.13.5.174
*Alamat penulis korespondensi: Balai Penelitian Tanaman Hias, Jalan Raya Ciherang Segunung Pacet. Kotak POS No. 8 Sindaglaya Cianjur, 43253.Tel: 0263-514148, Faks: 0263-14148, surel : [email protected].
Evaluasi Resistensi Progeni Anggrek Phalaenopsis terhadap Penyakit Busuk Lunak (Pseudomonas viridiflava)
Evaluation of Phalaenopsis Progenies Againts Soft Rot Bacterial Disease (Pseudomonas viridiflava)
Hanudin*, Indijarto Budi Rahardjo, Kurniawan BudiartoBalai Penelitian Tanaman Hias, Cianjur, 43253.
ABstRAK
Penyakit busuk lunak yang disebabkan oleh Pseudomonas viridiflava merupakan salah satu permasalahan utama di pembibitan anggrek, terutama Phalaenopsis. Evaluasi progeni yang berasal dari 3 kombinasi silangan dilakukan untuk menelaah karakteristik gejala dan perkembangan infeksi penyakit tersebut, serta respons ketahanan progeni terhadap infeksi buatan P. viridiflava. Penelitian dilakukan di rumah kaca standar untuk Phalaenopsis yang melibatkan 31 populasi individu dari 3 kombinasi persilangan (P.157, E.2153, dan E.2189). Hasil penelitian menunjukkan bahwa karakteristik gejala infeksi P. viriflava pada daun ditandai oleh peluruhan (lesio) pada jaringan yang diinokulasi. Pada kebanyakan aksesi, lesio membesar dan bau busuk terdeteksi sebagai indikasi keberhasilan infeksi patogen tersebut. Tiga aksesi Phalaenopsis diklasifikasikan ke dalam genotipe resisten (P.157-12, P.157-45,dan P.157-71), sedangkan satu aksesi (P.157-32) dikategorikan sebagai rentan dan 27 aksesi lainnya tergolong genotipe yang sangat rentan.
Kata kunci: pembibitan anggrek, progeni persilangan, respons ketahanan
ABstRACt
A bacterial soft rot disease caused by P. viridiflava was one of problems in orchid nursery, especially Phalaenopsis. An evaluation on progenies derived from three crossing combinations were conducted to find out the symptom characteristic, development of infections of the disease and resistance response of the progenies against artificial infection of P. viridiflava. The research was conducted under a standard glass house conditions for Phalaenopsis involving 31 individuals from 3 progeny populations, namely P.157, E.2153 and E.2189. The results showed that symptoms were characterized by the lesion of the leaf tissues. On most accessions, the lesion enlarged and foul odor was detected as an indication of the successful establishment of bacterial infection. Based on the disease severity, three accessions (P.157-12, P.157-45, and P.157-71) had the lowest infections and were classified into resistant genotypes. One accession, i.e. P.157-32 was classified as susceptible and the rest 27 accessions were grouped into very susceptible class.
Key words: crossing progeny, orchid nursery, resistant response

J Fitopatol Indones Hanudin et al
169
PENDAHULUAN
Phalaenopsis adalah salah satu genus tanaman hias yang terdiri atas 50 spesies. Sebagai tanaman hias bernilai ekonomi tinggi, informasi mengenai keragaman genetikanya masih sedikit demikian juga resistensi terhadap hama dan penyakitnya (Chang et al. 2009).
Busuk lunak bakteri yang disebabkan oleh Pseudomonas viridiflava adalah salah satu masalah pada budi daya anggrek Oncidium, dan Phapiopedilum. Kerusakan akibat patogen ini dapat mencapai 100% (Hanudin dan Rahardjo 2011).
Perbaikan genetika dari kultivar melalui pemuliaan introgresif adalah salah satu upaya untuk mengatasi masalah penyakit ini. Handayati et al. (2004) melaporkan bahwa tingkat resistensi terhadap infeksi P. viridiflava sangat bervariasi antarprogeni. Informasi berkaitan dengan latar belakang genetika, pola pewarisan, proses pemuliaan, dan karakteristik resistensi terhadap P. viridiflava pada Phalaenopsis, masih jarang dilaporkan. Penelitian dirancang untuk mengevaluasi resistensi beberapa populasi hasil persilangan tetua Phalaenopsis terhadap P. viridiflava dan menelaah gejala karakteristik serangan patogen tersebut.
BAHAN DAN MEtODE
Bahan yang digunakan ialah 31 nomor aksesi Phalaenopsis yang berasal dari tiga populasi persilangan. Setiap populasi persilangan terdiri atas 10 atau 11 aksesi (Tabel 1). Aksesi Phalaenopsis merupakan koleksi Bagian Pemuliaan dan Plasma Nutfah, Balai Penelitian Tanaman Hias (Balithi).
Enam bulan setelah dikeluarkan dari botol, individu tanaman dengan jumlah daun sebanyak 6–7 helai ditanam dalam pot plastik berdiameter 20 cm yang berisi campuran pakis arang kayu. Tanaman dipelihara di rumah kaca dan ditutup dengan jaring polikarbonat yang menyerap cahaya 60%. Cara pemeliharaan berdasarkan standar operasional prosedur budi daya anggrek Phalaenopsis. Pemupukan menggunakan pupuk majemuk NPK 30:30:30
yang dicairkan dengan konsentrasi 0.2% yang disemprotkan ke daun dengan interval 2 kali dalam seminggu. Perbanyakan inokulum P. viridiflava dan metode inokulasinya
Galur bakteri P. viridiflava Ph-7 merupakan koleksi Laboratorium Bakteriologi Balithi, Cianjur. Bakteri digoreskan pada medium King ‘B (20 g proteose peptone No. 3, 10 g gliserol, 1.5 g K2HPO4, 1.5 g MgSO4-7H2O, 15.0 g agar-agar dan 100 mL-1 akuades) dan diinkubasikan dalam inkubator bersuhu 32 ± 2 °C selama 24 jam. Biakan murni dengan warna koloni hijau berpendar disuspensikan dalam akuades steril sebagai sumber inokulum dengan kerapatan 5.2 × 1010 cfu mL-1.
Inokulasi P. viridiflava pada AnggrekSetiap aksesi anggrek diinokulasi
berdasarkan metode pin pricking (Hanudin dan Rahardjo 2011). Sebanyak dua helai daun (sebagai ulangan), yaitu pada daun ke-3 dan ke-4 dari pucuk dilukai menggunakan jarum semprit hipodermik (berdiameter 0.6 mM) kemudian diinokulasikan P. viridiflava dengan menempelkan kapas steril yang sebelumnya dicelupkan ke dalam suspensi inokulum. Sebagai kontrol digunakan Phalaenopsis aksesi rentan (M22-235) yang dilukai dan ditempeli kapas basah steril tanpa P. viridiflava.
Peubah yang diamati meliputi gejala penyakit, waktu inkubasi (WI) dan keparahan penyakit (KP). Keparahan penyakit dihitung menggunakan rumus sebagai berikut:
N × VKP = × 100%, dengan(ni × vi)∑
i
i=0
ni, jumlah tanaman dengan skor ke-i; vi, nilai skala dari tiap kategori serangan (Tabel 1); N, jumlah tanaman yang diamati; dan V, skor tertinggi.
Gejala busuk lunak dinilai berdasarkan skala deskriptif ordinal mulai dari 1–9: 1, tidakada lesio di lokasi inokulasi; 2, lesio lebih kecil dari 5 mm; 3, lesio antara 5–10 mm; 4, lesio yang lebih besar dari 10 mm tetapi tidak mencapai batang; 5, lesio mencapai batang utama; 6, infeksi mencapai batang

J Fitopatol Indones Hanudin et al
170
tapi tidak pada daun yang tidak diinokulasi; 7, infeksi mencapai batang dan daun yang tidak diinokulasi; 8, seluruh batang dan daun terinfeksi tetapi belum mati; dan 9, tanaman mati (Ren et al. 2001).
Waktu inkubasi diamati setiap hari mulai satu hari setelah inokulasi (HSI) sampai seluruh daun yang diinokulasi terinfeksi busuk lunak. Kriteria resistensi ditetapkan menggunakan metode da Silva Felix et al. (2014) yang dimodifikasi pada kriteria ketahanan menjadi, yaitu imun, KP = 0%; resisten, 1.0 < KP < 10%; rentan, 11% < KP <60%; dan sangat rentan, KP ≥ 61%.
HAsIL
Perkembangan gejala penyakit oleh P. viridiflava
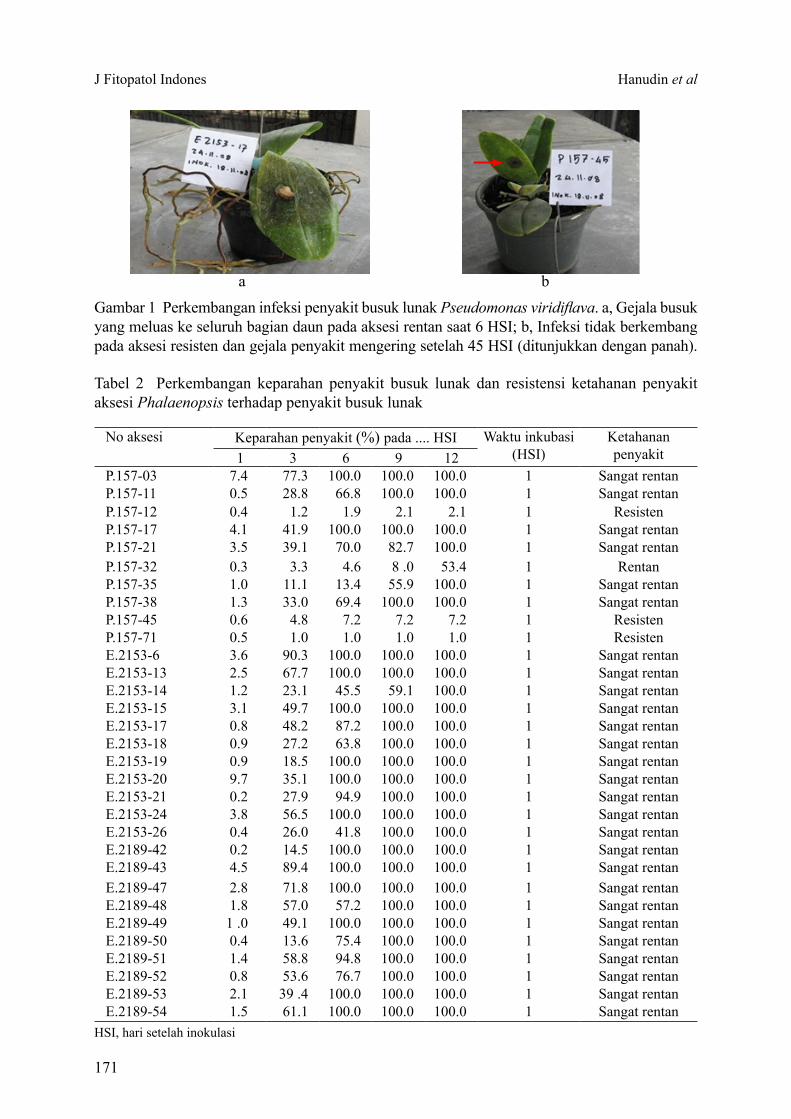
Secara umum awal serangan P. viridiflava pada daun Phalaenopsis ditandai dengan bercak hijau kecokelatan yang berkembang menjadi pembusukan jaringan (Gambar 1a). Pada individu tanaman populasi aksesi Phalaenopsis E.2153, gejala pembusukan yang meluas ke seluruh bagian daun dan bagian tanaman lain terdeteksi lebih cepat daripada populasi aksesi Phalaenopsis P.157 dan E.2189 (Tabel 2). Gejala busuk ini menimbulkan aroma yang menyengat. Tanaman dengan gejala seperti ini umumnya
mati pada 12 HSI. Inokulasi yang tidak berkembang tampak pada aksesi Phalaenopsis P.157 yang tahan (Gambar 1b).
Keparahan Penyakit dan Klasifikasi Resistensi
Perkembangan gejala infeksi P. viridiflava pada setiap aksesi anggrek yang diuji bervariasi. Pada seluruh progeni pada populasi aksesi Phalaenopsis E.2153, E.2189 dan sebagian besar P.157 yang diuji menunjukkan tingkat perkembangan penyakit dan keparahan yang tinggi. Peningkatan KP paling tajam terlihat pada aksesi E.2153-6, yaitu 3.6, 90.3 dan 100% pada 1, 3, dan 6 HSI. Pada aksesi P.157-32, tingkat keparahan terdeteksi lebih rendah, yaitu 0.3, 3.3, 4.6, 8.0 dan 53.4% pada 1, 3, 6, 9, dan 12 HSI. Tiga aksesi mempunyai tingkat perkembangan infeksi P. viridiflava paling rendah dari seluruh tanaman yang diuji, yaitu P.157-12, P.157-45 dan P.157-71 (Tabel 2). Gejala infeksi yang timbul tidak berkembang pada tiga aksesi tersebut hingga 45 HSI, bahkan luka bekas infeksi berubah menjadi kering (Gambar 1b).
Perkembangan dan tingkat keparahan penyakit busuk diklasifikasikan resisten pada tiga aksesi, yaitu P.157-12, P.157-45, dan P.157-71. Aksesi P.157-32 dengan tingkat keparahan tinggi diklasifikasikan rentan dan aksesi lainnya tergolong sangat rentan.
Kombinasi tetua silangan Phalaenopsis “Vulcan Harbour” x (“Golden
Amboin” x “Meller Gold”) (E.2189, nomor kode populasi)
‘Kenanga’ x (‘Golden Wish’ x ‘Brother Lawrence’
(E.2153, nomor kode populasi)
‘Barbara Moller’ x (‘Robert Chick’ x ‘Golden Amboin’)
(P.157, nomor kode populasi)42 6 343 13 1147 14 1248 15 1749 17 2150 18 3251 19 3552 20 3853 21 4554 24 71
26
Tabel 1 Daftar tiga populasi aksesi Phalaenopsis hasil persilangan yang digunakan pada penelitian
J Fitopatol Indones Hanudin et al
171
Gambar 1 Perkembangan infeksi penyakit busuk lunak Pseudomonas viridiflava. a, Gejala busuk yang meluas ke seluruh bagian daun pada aksesi rentan saat 6 HSI; b, Infeksi tidak berkembang pada aksesi resisten dan gejala penyakit mengering setelah 45 HSI (ditunjukkan dengan panah).
a b
Tabel 2 Perkembangan keparahan penyakit busuk lunak dan resistensi ketahanan penyakit aksesi Phalaenopsis terhadap penyakit busuk lunak
No aksesi Keparahan penyakit (%) pada .... HSI Waktu inkubasi (HSI)
Ketahanan penyakit1 3 6 9 12
P.157-03 7.4 77.3 100.0 100.0 100.0 1 Sangat rentanP.157-11 0.5 28.8 66.8 100.0 100.0 1 Sangat rentanP.157-12 0.4 1.2 1.9 2.1 2.1 1 ResistenP.157-17 4.1 41.9 100.0 100.0 100.0 1 Sangat rentanP.157-21 3.5 39.1 70.0 82.7 100.0 1 Sangat rentanP.157-32 0.3 3.3 4.6 8 .0 53.4 1 RentanP.157-35 1.0 11.1 13.4 55.9 100.0 1 Sangat rentanP.157-38 1.3 33.0 69.4 100.0 100.0 1 Sangat rentanP.157-45 0.6 4.8 7.2 7.2 7.2 1 ResistenP.157-71 0.5 1.0 1.0 1.0 1.0 1 ResistenE.2153-6 3.6 90.3 100.0 100.0 100.0 1 Sangat rentanE.2153-13 2.5 67.7 100.0 100.0 100.0 1 Sangat rentanE.2153-14 1.2 23.1 45.5 59.1 100.0 1 Sangat rentanE.2153-15 3.1 49.7 100.0 100.0 100.0 1 Sangat rentanE.2153-17 0.8 48.2 87.2 100.0 100.0 1 Sangat rentanE.2153-18 0.9 27.2 63.8 100.0 100.0 1 Sangat rentanE.2153-19 0.9 18.5 100.0 100.0 100.0 1 Sangat rentanE.2153-20 9.7 35.1 100.0 100.0 100.0 1 Sangat rentanE.2153-21 0.2 27.9 94.9 100.0 100.0 1 Sangat rentanE.2153-24 3.8 56.5 100.0 100.0 100.0 1 Sangat rentanE.2153-26 0.4 26.0 41.8 100.0 100.0 1 Sangat rentanE.2189-42 0.2 14.5 100.0 100.0 100.0 1 Sangat rentanE.2189-43 4.5 89.4 100.0 100.0 100.0 1 Sangat rentanE.2189-47 2.8 71.8 100.0 100.0 100.0 1 Sangat rentanE.2189-48 1.8 57.0 57.2 100.0 100.0 1 Sangat rentanE.2189-49 1 .0 49.1 100.0 100.0 100.0 1 Sangat rentanE.2189-50 0.4 13.6 75.4 100.0 100.0 1 Sangat rentanE.2189-51 1.4 58.8 94.8 100.0 100.0 1 Sangat rentanE.2189-52 0.8 53.6 76.7 100.0 100.0 1 Sangat rentanE.2189-53 2.1 39 .4 100.0 100.0 100.0 1 Sangat rentanE.2189-54 1.5 61.1 100.0 100.0 100.0 1 Sangat rentan
HSI, hari setelah inokulasi

J Fitopatol Indones Hanudin et al
172
PEMBAHAsAN
P. viridiflava ialah patogen yang memiliki kisaran inang luas. Patogen tersebut dapat menginfeksi tanaman kacang merah (Phaseolus vulgaris), kiwi (Actinidia deliciosa), selada (Lactuca sativa) (Gonzales et al. 2003), wortel (Almeida et al. 2013), kubis (Bhat et al. 2012) dan alfafa (Heydari et. al. 2012) dari bagian daun, batang hingga akar tanaman (Sarris et al. 2012).
Gejala infeksi P. viridiflava berupa bercak hijau kecokelatan diikuti pembusukan jaringan. P. viridiflava mampu mensekresikan satu set enzim maupun isoenzim yang mendegradasi polimer dinding sel Phalaenopsis (Joko et al. 2014). Bakteri ini dilaporkan mempunyai karakteristik yang sama dengan bakteri penyebab busuk lunak Erwinia dalam mensekresikan 5 isoenzim utama (PelA, PelB, PelC, PelD, PelE) dan 4 isoenzim sekunder (PelI, PelL, PelZ, PelX) pektat liase (Lee et al. 2006). Enzim pektinase merupakan faktor utama patogenisitas bakteri busuk lunak yang digunakan untuk menghancurkan pengikat di antara sel tanaman sehingga menyebabkan kematian jaringan (Mikicinski et al. 2010). Bakteri busuk lunak juga menghasilkan enzim selulase yang berfungsi memecah selulosa yang terdapat pada dinding sel primer dan sekunder tanaman dan enzim protease yang berfungsi untuk merusak protein pada dinding sel sehingga bakteri dapat masuk ke dalam sel tanaman inang (Ismail et al. 2012).
Perkembangan penyakit mencapai 100% pada 12 HSI pada sebagian besar tanaman yang diuji, walaupun beberapa aksesi menunjukkan respons yang berbeda. Hal ini menunjukkan bahwa galur P. viridiflava yang digunakan pada pengujian ini sangat virulen. Perbedaan respons yang ditunjukkan oleh aksesi P.157-12, P.157-45, dan P.157-71 berupa keparahan penyakit yang rendah (1.0–7.2% pada 12 HSI) dan gejala infeksi yang mengering hingga 45 HSI mengindikasikan bahwa terdapat mekanisme spesifik yang bersifat endogen pada ketiga aksesi ini yang menyebabkan infeksi P. viridiflava tidak dapat berkembang lebih lanjut. Karakteristik
ini diduga berhubungan dengan sifat yang diwariskan oleh tetuanya yaitu Phalaenopsis ”Barbara Moler”, Phalaenopsis ”Robert Chick”, atau Phalaenopsis ”Golden Amboin”. Davidsson et al. (2013) melaporkan bahwa resistensi tanaman terhadap penyakit bersifat kompleks dan diatur oleh banyak gen yang dapat diturunkan pada generasi berikutnya.
Secara umum resistensi tanaman terhadap patogen dapat berbentuk resistensi morfologi, anatomi, dan kimia (Henry et al. 2013). Pada resistensi morfologi yang berperan menghambat penetrasi patogen ke dalam jaringan tanaman ialah lapisan epidermis, stomata, dan ketebalan kutikula (Bessire et al. 2007). Lapisan epidermis tebal dilaporkan dapat menghambat infeksi bercak hitam pada mawar (Suhardi dan Winarto 2002); embun tepung pada jeruk (Mahfud 1991); antraknosa pada mangga (Mahfud et al. 1997); dan bercak daun pada pisang (Aliah et al. 2015). Daun yang mempunyai stomata kecil akan menyulitkan penetrasi patogen karat putih pada krisan (Budiarto et al. 2008).
Pada penyakit busuk lunak bakteri, beberapa laporan menyebutkan bahwa mekanisme ketahanan disebabkan oleh karakteristik genotipe tanaman yang mampu memproduksi fitoaleksin yang dapat menghambat atau membunuh patogen (Jeandet et al. 2013). Mekanisme ketahanan tanaman terhadap P. viridiflava hingga saat ini belum terungkap sempurna. Marina et al. (2013) melaporkan tingkat resistensi tanaman terhadap infeksi P. viridiflava berhubungan dengan meningkatnya konsentrasi poliamine dalam tubuh tanaman. Rossi et al. (2015) juga mengemukakan aktivitas enzim arginine dekarbosilase dapat menghambat patogen P. viridiflava.
Berdasarkan perkembangan dan tingkat keparahan penyakit maka aksesi P.157–32 dikategorikan rentan, sedangkan aksesi P.157-12, P.157-45, dan P.157-71 di-klasifikasikan resisten. Sebanyak 27 aksesi lainnya yang mempunyai tingkat keparahan penyakit tinggi hingga 12 HSI dikategorikan sangat rentan. Informasi penelitian ini dapat digunakan oleh pemulia dalam studi

J Fitopatol Indones Hanudin et al
173
pewarisan sifat ketahanan dan seleksi tetua persilangan Phalaenopsis yang ditujukan untuk mendapatkan progeni resisten terhadap busuk lunak bakteri.
UCAPAN tERIMA KAsIH
Penulis mengucapkan terima kasih kepada Balitbangtan, Puslitbang Hortikultura dan Balai Penelitian Tanaman Hias yang telah membiayai pelaksanaan penelitian. Penulis mengucapkan terima kasih juga kepada Saepullah, Ridwan Daelani, Asep Syamsudin, dan Iman Taufiq yang telah membantu dalam persiapan penelitian dan pengumpulan data selama penelitian berlangsung.
DAFtAR PUstAKA
Aliah NU, Sulistyowati L, Muhibbudin A. 2015. Hubungan ketebalan lapisan epidermis daun terhadap serangan jamur (Mycosphaerella musicola) penyebab penyakit bercak daun sigatoka pada sepuluh kultivar pisang. JHPT. 3(1):35–43.
Almeida IMG, Maciel KW, Neto JR, Beriam L. 2013. Pseudomonas viridiflva in imported carrot seeds. Aus Plant Dis. 8:17–19. DOI: https://doi.org/10.1007/s13314-012-0086-2.
Bessire M, Chassot C, Jacquat AC, Humphry M, Borel S, Petetot JMC, Metraux JP, Nawrath C. 2007. A permeable cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea. The EMBO Journal. 26(8):2158–2168. DOI: https://doi.org/10.1038/sj.emboj.7601658.
Budiarto K, Rahardjo IB, Suhardi. 2008. Seleksi ketahanan klon-klon harapan krisan terhadap penyakit karat. J Horti. 18(3):249–254.
Bhat KA, Bhat NA, Mohiddin FA, Sheikh PA, Wani AH. 2012. Studies on pectinase activities of isolates of Erwinia carotovora and Rhizopus sp. causing soft rot in cabbage (Brassica oleracea var. capitata L.). Afric J Agric Res. 7(45):6062–6067. DOI: https://doi.org/10.5897/AJAR12.1215.
Chang YK, Veilleux RE, Iqbal MJ. 2009. Analysis of genetic variability among
Phalaenopsis species and hybrids using amplified fragment length polymorphism. J Am Soc Hort Sci. 134 (1):58–66.
Davidsson PR, Kariola T, Niemi O, Palva ET. 2013. Pathogenicity of and plant immunity to soft rot pectobacteria. Front Plant Sci. 4(191):1–13. DOI: https://doi.org/10.3389/fpls.2013.00191.
Da Silva Felix KC, de Oliveira WJ, Mariano RLR, de Souza EB. 2014. Lettuce genotype resistance to ‘soft rot’ caused by Pectobacterium carotovorum subsp. carotovorum. Sci Agri. 71(4):287–291. DOI: https://doi.org/10.1590/0103-9016=2013-0301.
Gonzales AJ, Rodicio MR, Mendoza MC. 2003. Identification of an emergent and atypical Pseudomonas viridiflava lineage causing bacteriosis in plants of agronomic importance in a Spanish Region. App Environ Microbiol. 69(5):2936–2941. DOI: https://doi.org/10.1128/AEM.69.5.2936-2941.2003.
Hanudin, Rahardjo IB. 2011. Karakteristik Pseudomonas viridiflava: penyebab penyakit busuk lunak dan evaluasi virulensinya pada klon anggrek Phalaenopsis. JHPT Tropika. 11(2):185–193. DOI: http://dx.doi.org/10.23960/j.hptt.211185-193.
Handayati W, Hanudin, Soedjono S. 2004. Resistensi genotip anggrek Phalaenosis terhadap penyakit busuk lunak. J Hort. 14(Edisi khusus):398–402.
Henry E, Yadeta KA, Coaker G. 2013. Recognition of bacterial plant pathogens: local, systemic and transgenerational immunity. New Phytologist. 199:908–915. DOI: https://doi.org/10.1111/nph.12214.
Heydari A, Khodakaramian G, Zafari D. 2012. Characterization of Pseudomonas viridiflava causing alfalfa root rot disease in Hamedan province of Iran. J Plant Pathol Microbiol. 3:135. DOI : https://doi.org/:10.4172/2157-7471.1000135.
Ismail ME, Abdel_Monaim MF, Mostafa YM. 2012. Identification and pathogenicity of phytopathogenic bacteria associated with soft rot disease of girasole tuber in Egypt.

J Fitopatol Indones Hanudin et al
174
J. Bacteriol. Res. 4(1):1–8. DOI: https://doi.org/10.5897/JBR11.015.
Jeandet P, Clément C, Courot E, Cordelier S. 2013. Modulation of phytoalexin biosynthesis in engineered plants for disease resistance. Int J Mol Sci. 14:14136–14170. DOI: https://doi.org/10.3390/ijms140714136.
Joko T, Subandi A, Kusumandani W, Wibowo A, Priatmodjo A. 2014. Activities of plant cell wall-degradating enzymes by bacterial soft rot of orchids. Archived Phytopathol Plant Protec. 47(10):1239–1250. DOI: https://doi.org/ 080/03235408.2013.838374.
Lee YA, Chen KP, Hsu YW. 2006. Characterization of Erwinia chrysanthemi, the soft-rot pathogen of white-flowered calla lily, based on pathogenicity and PCR-RFLP and PFGE analyses. Plant Pathol. 55:530–536. DOI: https://doi.org/10.1111/j.1365-3059.2006.01392.x.
Marina M, Sirera FV, Rambla JL, Gonzalez ME, Blazquez MA, Carbonell J, Pieckenstain FL, Ruiz OA. 2013. Thermospermine catabolism increases Arabidopsis thaliana resistance to Pseudomonas viridiflava. J Experimen Bot. 64(5):1393–1402. DOI: https://doi.org/10.1093/jxb/ert012.
Mikicinski A, Sobiczewski P, Sulikowska M, Pulawska J, Treder J. 2010. Pectolytic bacteria associated with soft rot of calla lily (Zantedeschia spp.)
tubers. Phytopathology. 158(4):201–209. DOI: https://doi.org/10.1111/j.1439-0434.2009.01597.x.
Mahfud MC. 1991. Ketahanan beberapa jenis jeruk terhadap penyakit embun tepung. J Hort. 1(2):54–57.
Mahfud MC, Purnomo S, Handoko, Tegopati B, Sugiyarto M. 1997. Perbedaan ketahanan beberapa jenis mangga terhadap penyakit antraknosa. J Hort. 7(1):561–565.
Ren JR, Petzoldt R, Dickson MH. 2001. Genetics and population improvement of resistance to bacterial soft rot in Chinese cabbage. Euphytica. 117(3):197–207. DOI: https://doi.org/10.1023/A:1026541724001.
Rossi FR, Marina M, Pieckenstain FL. 2015. Role of arginine decarboxylase (ADC) in Arabidopsis thaliana defence against the pathogenic bacterium Pseudomonas viridiflava. Plant Biol (Stuttg). 17(4):831–839. DOI: https://doi.org/10.1111/plb.12289.
Sarris PF, Trantas EA, Mpalantinaki E, Ververidis F. 2012. Pseudomonas viridiflava, a multi host plant pathogen with significant genetic variation at the molecular level. Plos One. 7(4):e36090. DOI: https://doi.org/10.1371/journal.pone.0036090.
Suhardi, Winarto B. 2002. Resistensi varietas mawar (Rosa hybrida) terhadap penyakit bercak hitam. J Hort. 12(1): 56–63.

ISSN: 0215-7950
175
Volume 13, Nomor 5, September 2017Halaman 175–183
DOI: 10.14692/jfi.13.5.175
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, Jalan Kamper, Kampus IPB Darmaga, Bogor 16680Tel: 0251-8629364, Faks: 0251-8629362, Surel: [email protected]
Identifikasi Colletotrichum spp. Asal Tanaman Pepaya
Identification of Colletotrichum spp. Originated from Papaya Plant
Eryna Elfasari Rangkuti, Suryo Wiyono, Widodo*
Institut Pertanian Bogor, Bogor 16680
ABSTRAK
Colletotrichum spp. sebagai penyebab penyakit antraknosa merupakan salah satu faktor pembatas produksi pepaya (Carica papaya). Tujuan penelitian ini ialah mengidentifikasi spesies Colletotrichum spp. dari berbagai bagian tanaman pepaya yang menunjukkan gejala antraknosa. Sebanyak 20 galur Colletotrichum spp. berhasil diisolasi, selanjutnya dilakukan pengelompokkan menjadi 3 tipe morfologi berdasarkan kemiripan koloni dan bentuk konidia. Sebanyak 12 galur dari kelompok I dan 3 galur dari kelompok II diidentifikasi sebagai C. gloeosporioides sensu lato berdasarkan morfologi koloni dan bentuk konidia silinder yang kedua ujungnya membulat. Sementara itu 1 galur dari kelompok II secara molekuler diidentifikasi sebagai C. magnum. Empat galur dari kelompok III dengan bentuk konidia melengkung dan berujung runcing, serta membentuk setae pada medium buatan diidentifikasi secara molekuler sebagai C. truncatum. C. gloeosporioides ditemukan pada batang, tangkai daun, dan buah. C. magnum dan C. truncatum masing-masing ditemukan pada tangkai daun dan buah. Selain karakter morfologi dan molekuler, perbedaan respons pertumbuhan koloni terhadap suhu dapat digunakan untuk mengonfirmasi spesies Colletotrichum tersebut. Keberadaan Colletotrichum yang menginfeksi pada bagian batang dan tangkai daun pepaya di pertanaman dan temuan spesies C. magnum dan C. truncatum pada tanaman ini merupakan laporan pertama di Indonesia.
Kata kunci: C. gloeosporioides, C. magnum, C. truncatum, morfologi, molekuler
ABSTRACT
Colletotrichum spp. is known as the causal agent of anthracnose and considered as an important limiting factors on papaya production. The objective of this study was to determine the species of Colletotrichum spp. from various plant parts of papaya (Carica papaya) showing anthracnose symptom. Twenty isolates of Colletotrichum spp. was isolated and were grouped into 3 morphological groups based on colony similarity, conidial morphology, and setae formation. A total of 12 isolates of group I and 3 isolates of group II were identified morphologically as C. gloeosporioides sensu lato based on cylindrical conidia with rounded on both ends and colony morphology. Meanwhile, 1 isolate of group II was molecularly identified as C. magnum. Four isolates of group III with a curved and pointed-end conidia, and produced setae on artificial medium were identified molecularly as C. truncatum. C. gloeosporioides was obtained on stem, leaf petiole, and fruit. C. magnum and C. truncatum were only obtained on leaf petiole and fruit, respectively. In addition to morphological and molecular characters, differences in colony growth responses to temperature can be used to distinguish the species of Colletotrichum. To our knowledge the existence of anthracnose symptom on the stems and leaf petioles of papaya in the fields and the discovery of C. magnum and C. truncatum isolated from papaya was the first report in Indonesia.
Key words: C. gloeosporioides, C.magnum, C. truncatum, molecular, morphology

J Fitopatol Indones Rangkuti et al.
176
PENDAHULUAN
Pepaya (Carica papaya) merupakan salah satu buah yang banyak dikonsumsi di Indonesia. Salah satu faktor pembatas dalam peningkatan produksi, baik kualitas maupun kuantitas buah pepaya ialah infeksi Colletotrichum sp. penyebab antraknosa ketika buah masih di pertanaman maupun saat pascapanen.
Pada umumnya antraknosa pada pepaya lebih dikenal sebagai penyakit pascapanen (Rahman et al. 2008), akan tetapi Haggag dan Singer (2013) melaporkan C. capsici dapat menyebabkan antraknosa sebelum dan sesudah pascapanen. Antraknosa dapat menyerang batang, tangkai, dan buah papaya di pertanaman berdasarkan pengamatan di berbagai daerah di Indonesia. Bahkan dari hasil pengamatan di lahan, buah yang masih hijau juga sudah mulai terserang, terutama pada kultivar pepaya tipe ukuran medium(1–2 kg). Oleh karena itu penelitian ini dilakukan untuk mengoleksi spesies Colletotrichum pada berbagai bagian tanaman pepaya dan mengidentifikasinya berdasarkan pada ciri morfologi, molekuler, dan respons suhu. Pengetahuan dasar ini diharapkan berguna dalam menyusun strategi pengendalian penyakit antraknosa pada pepaya.
BAHAN DAN METODE
Isolasi Colletotrichum spp.Cendawan diisolasi dari batang, tangkai,
dan buah pepaya yang menunjukkan gejala khas antraknosa. Buah pepaya tersebut berasal dari kebun percobaan Pusat Kajian Hortikultura Tropika Pasir Kuda, Bogor dan kebun pepaya milik petani di Desa Tambak Mulyo, Kabupaten Kebumen, Jawa Tengah. Isolasi cendawan dilakukan dengan menanam jaringan tanaman pada medium agar-agar dekstrosa kentang (ADK) dan diinkubasi pada suhu 28 °C. Cendawan yang tumbuh dengan ciri-ciri sebagai cendawan Colletotrichum dimurnikan mengikuti metode yang dikemukakan oleh Choi et al. (1999).
Identifikasi MorfologiColletotrichum diidentifikasi mengikuti
Watanabe (2002), serta pustaka acuan lainnya, yaitu Smith dan Black (1990), Zakaria dan Bailey (2000), Photita et al. (2005), dan Weir et al. (2012). Pengamatan dilakukan terhadap warna koloni, pembentukan seta, dan bentuk konidium.
Identifikasi Molekuler Colletotrichum spp.Isolasi DNA Total. Sebanyak 3 potong
(diameter ± 5 mm) biakan Colletotrichum spp. berumur 10 hari dibiakkan ke dalam erlenmeyer berisi 100 mL potato dextrose broth (PDB) dan diinkubasikan selama 7 hari pada suhu kamar. Miselium cendawan dipanen dengan filtrasi vakum dan dicuci menggunakan akuades steril, selanjutnya dikeringkan. DNA diekstraksi dan dimurnikan menurut metode Sambrook et al. (1989) yang dimodifikasi pada tahap ekstraksi DNA, yaitu suhu dan waktu pemanasannya. Sebanyak 0.2 gmiselium cendawan digerus dengan nitrogen cair hingga menjadi serbuk lalu dimasukkan ke dalam tabung eppendorf, selanjutnya ditambahi 1 mL bufer ekstrak dan 10 μL β-mercapto ethanol dan dicampur hingga homogen menggunakan vorteks. Pemanasan dilakukan untuk menghancurkan dinding sel di dalam penangas air dengan suhu 65 °C selama 30 menit dan didinginkan sampai mencapai suhu ruang. Sebanyak 750 μL kloroform isoamilalkohol (24:1) ditambahkan ke dalam tabung dan dicampur hingga homogen lalu disentrifugasi pada kecepatan 11 000 rpm selama 10 menit. Supernatan yang diperoleh dipindahkan ke tabung eppendorf baru dengan menambahkan 1000 μL kloroform dicampur hingga homogen dengan vorteks dan disentrifugasi kembali pada 11 000 rpm selama 10 menit.
Supernatan dipindahkan ke tabung eppendof yang baru dan ditambahkan 1000 μLisopropanol dingin. Tabung dikocok perlahan untuk mengikat DNA dan diinkubasi pada suhu -20 °C selama 30 menit. Benang DNA yang diperoleh disentrifugasi selama 10 menit hingga mengendap. Supernatan dibuang

J Fitopatol Indones Rangkuti et al.
177
dan pelet dicuci dengan etanol 70% lalu disentrifugasi pada 11 000 rpm selama 10 menit. Etanol dibuang dan pelet dikeringvakumkan. Pelet diresuspensi dalam 100 μL bufer TE dan disimpan pada suhu -20 °C untuk digunakan dalam proses amplifikasi DNA.
Amplifikasi DNA. Proses amplifikasi dilakukan pada mesin Thermo Cycle PCR Gene Amp PCR System 9700 versi 3.12. Amplifikasi menggunakan primer universal untuk spesies Colletotrichum, yaitu forward primer ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) dan reverse primer ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) dengan ukuran target hasil amplifikasi ialah 580 pb. Reaksi amplifikasi DNA dengan volume total 25 μL terdiri atas 2 μL DNA, 2.5 μL bufer 10x dan Mg2+, 0.5 μLdNTP 10 mM, 1 μL masing-masing primer, 0.2 μL Taq DNA (5 unit μL-1), dan 17.8 μL H2O. Kondisi amplifikasi dibagi menjadi beberapa tahap, yaitu pradenaturasi suhu 95 °C selama 1 menit, diikuti 30 siklus amplifikasi yang masing-masing siklus terdiri atas pemisahan utas DNA pada suhu 94 °C selama 1 menit, penempelan primer pada suhu 58 °C selama 1 menit, sintesis DNA pada suhu 72 °C selama 1 menit dan penyambungan DNA pada suhu 72 °C selama 7 menit (White et al. 1990).
Elektroforesis DNA. Produk hasil amplifikasi dianalisis menggunakan gel agarosa 1% (0.5x Tris-Borate EDTA/TBE). Elektroforesis dilakukan pada 50 volt selama 50 menit dan selanjutnya gel agarosa direndam dalam larutan pewarna yang berisi etidium bromida (1%) selama 15 menit, lalu dicuci dengan H2O selama 10 menit. Hasil elektroforesis divisualisasikan dengan transilluminator ultraviolet (APOLLO portable light box: model LB101, 1 lamp unit, 110 volts 60 hz,). Fragmen DNA yang terbentuk pada hasil elektroforesis didokumentasi.
Analisis Runutan DNA Colletotrichum spp. Produk amplifikasi dikirim ke First Base (Malaysia) untuk perunutan nukleotida. Hasil perunutan dianalisis menggunakan program basic local alignment search tool (BLAST) dengan program optimasi untuk memperoleh urutan basa DNA yang memiliki homologi dengan sikuen DNA yang terdapat dalam situs
national center for biotechnology information (NCBI). Runutan nukleotida yang diperoleh dianalisis menggunakan penyejajaran berganda ClustalW pada perangkat lunak Bioedit sequence alignment editor versi 7.1.3. Hubungan kekerabatan antarisolat dikonstruksi menggunakan perangkat lunak molecular evolutionery genetic analysis versi 6.06 (MEGA6) dengan bootstrap 1000 kali ulangan.
Amplifikasi dengan Primer Spesifik. Sebanyak 8 isolat Colletotrichum yang mewakili warna koloni dan bagian tanaman yang diisolasi dilakukan amplifikasi menggunakan primer spesifik CaInt2/ITS4 dan CcInt/ITS4, masing-masing untuk mengidentifikasi C. acutatum dan C. capsici. Penggunaan primer spesifik CaInt2/ITS4 dilakukan untuk memastikan beberapa isolat dari beberapa tipe morfologi yang ditemukan dalam penelitian ini dan diduga sebagai C. gloeosporioides berdasarkan karakter morfologi, tetapi bentuk konidianya sering mirip dengan C. acutatum. Sementara itu penggunaan primer spesifik CcInt/ITS4 untuk memastikan bentuk konidia isolat yang berbentuk melengkung sebagai C. capsici.
Respons Suhu Colletotrichum spp. asal Berbagai Bagian Tanaman
Percobaan ini dilakukan mengikuti metode Smith dan Black (1990) untuk membedakan beberapa spesies Colletotrichum berdasar respons suhu terhadap pertumbuhan radialnya pada medium buatan. Biakan Colletotrichum spp. dengan diameter 5 mm diletakkan pada medium ADK dan diinkubasi pada suhu 16, 24, dan 32 °C. Percobaan disusun dalam rancangan acak lengkap faktorial dengan 2 faktor, yaitu biakan Colletotrichum spp. dan suhu. Diameter koloni pertumbuhan cendawan diamati pada hari ke-10 setelah inkubasi.
HASIL
Identifikasi secara MorfologiSebanyak 20 isolat diperoleh dari
beberapa bagian tanaman yang menunjukkan gejala antraknosa. Seluruhnya merupakan

J Fitopatol Indones Rangkuti et al.
178
Colletotrichum spp. Berdasarkan pada karakter morfologi, Colletotrichum yang ditemukan dalam penelitian ini secara umum dibagi menjadi 3 kelompok tipe morfologi (Gambar 1). Kelompok I, warna koloni tampak atas yang bervariasi mulai abu-abu, krem, atau hijau tua dengan koloni tampak bawah abu-abu; aservulus berwarna oranye yang berisi massa konidia terlihat jelas pada pusat koloni; konidium silinder dengan kedua ujungnya membulat, membentuk 2 gelembung minyak; dan seta tidak ada, baik pada medium maupun pada jaringan tanaman. Kelompok II, warna koloni seragam, yaitu tampak atas berwarna abu-abu dan tampak bawah berwarna abu-abu kehitaman; konidium silinder dengan ujung membulat, tidak membentuk gelembung minyak; seta tidak ada, baik pada medium maupun jaringan tanaman inang. Tipe morfologi kelompok III ialah warna koloni tampak atas krem atau abu-abu, dengan warna koloni tampak bawah cokelat dengan cincin konsentris, konidium melengkung dengan kedua ujung meruncing dan membentuk 2 gelembung minyak; seta dihasilkan hanya pada medium ADK (Gambar 2).
Colletotrichum tipe morfologi kelompok I ditemukan pada semua bagian tanaman (batang: BT1, BT14, BT1, BT16; buah: B17, B24, B24, B50; dan tangkai daun: TP, T2, T3, T20). Tipe morfologi kelompok II ditemukan pada bagian batang dan tangkai daun (batang: B17; tangkai daun: T15, T21, T26), dan kelompok III hanya ditemukan pada bagian buah (B4, B7, B8, dan B19). Jika hanya didasarkan pada warna koloni dan bentuk konidium, kelompok I dan II (16 galur) diidentifikasi sebagai C. gloeosporioides sensu lato. Meskipun bentuk konidia pada kelompok I dan II sama, akan tetapi pada kelompok II konidianya tidak membentuk gelembung minyak. Dengan adanya perbedaan ini, salah satu galur yang mewakili tipe morfologi kelompok II (T15) diuji lebih lanjut secara molekuler dengan primer universal untuk memastikan spesiesnya. Isolat-isolat dari tipe morfologi III dengan bentuk konidia melengkung dengan kedua ujung meruncing yang oleh beberapa peneliti (Haggag dan
Singer 2013; Rahman et al. 2008; Sato et al. 2015) diidentifikasi secara morfologi sebagai C. capsici, diidentifikasi lebih lanjut secara molekuler untuk lebih memastikan spesiesnya seperti yang disarankan oleh Cannon et al. (2012).
Identifikasi Molekuler Colletotrichum spp. Amplifikasi DNA. Sebanyak 8 galur yang
mewakili bagian tanaman dan tipe morfologi, yaitu BT 15 dan BT16 (tipe I dari batang), T15 dan T26 (tipe II dari tangkai daun), serta B4, B7, B8, dan B9 (tipe III dari buah) tidak teramplifikasi fragmen DNA-nya, baik dengan primer spesifik C. acutatum maupun C. capsici. Dengan hasil tersebut, ketiga kelompok tipe morfologi Colletotrichum yang ditemukan bukan spesies C. acutatum maupun C. capsici. Akan tetapi 5 dari 8 galur yang diuji, yaitu T15 (tipe morfologi II), B4, B7, B8, dan B19 (tipe morfologi III) berhasil diamplifikasi dengan ukuran ± 600 pb menggunakan primer universal ITS1/ITS4 (Gambar 3). Berdasarkan hasil sikuensing, 4 galur (B4, B7, B8 dan B9) dinyatakan sebagai C. truncatum, dan 1 galur (T15) dinyatakan sebagai C. magnum.
Filogenetika Colletotrichum dari bagian pepaya. Berdasarkan analisis filogenetika yang disejajarkan dengan database di NCBI galur C. truncatum dari hasil penelitian ini memiliki kekerabatan yang lebih dekat dengan galur dari Malaysia dibandingkan dengan galur yang berasal dari negara Asia lainnya (China, Taiwan dan Thailand). Sementara itu C. magnum yang diisolasi dari tangkai daun (T15) berkerabat dekat dengan galur yang berasal dari Brazil (Gambar 4).
Respons Pertumbuhan Colletotrichum spp. terhadap Suhu. Pertumbuhan koloni (diameter) Colletotrichum yang diisolasi dari tanaman pepaya dipengaruhi oleh suhu dan spesiesnya. Dari uji statistik, baik spesies maupun suhu secara tunggal dan interaksinya menunjukkan perbedaan yang nyata. Ketiga spesies yang diuji, C. gloesporioides, C. magnum, dan C. truncatum, secara umum tumbuh optimum pada suhu 24 °C. Pada suhu 16 oC, C. gloeosporioides cenderung tumbuh lebih cepat dibandingkan 2 spesies lainnya,
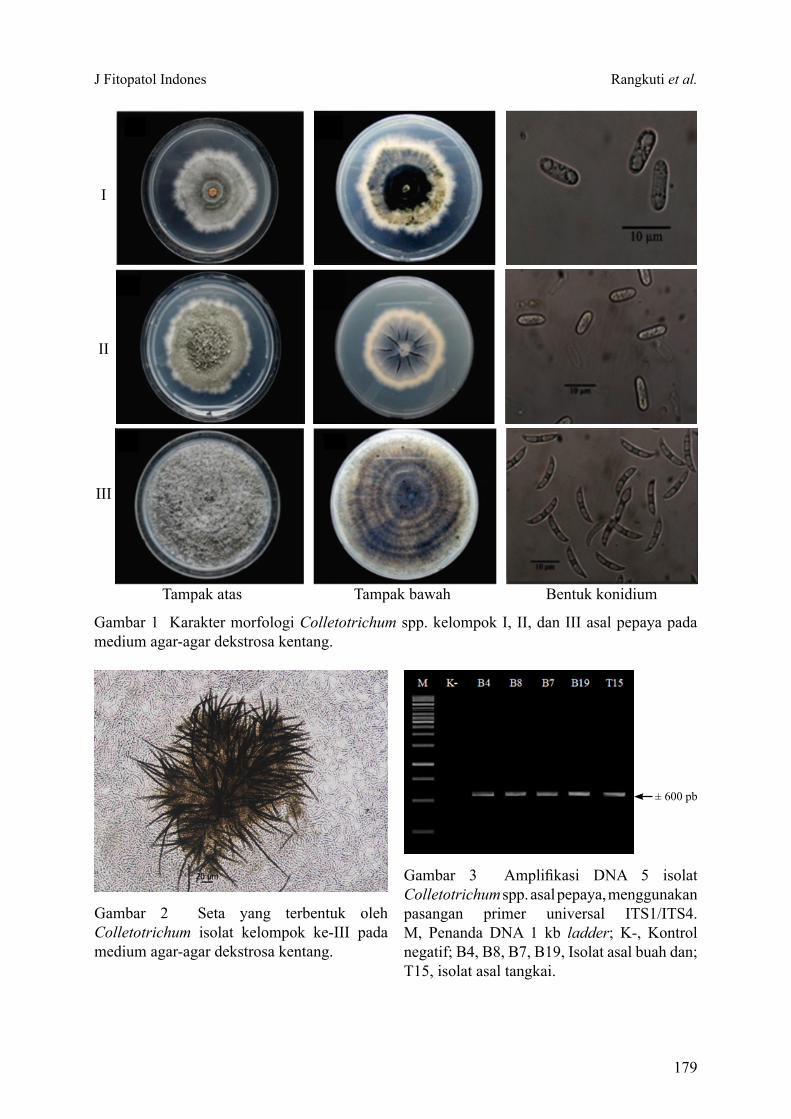
J Fitopatol Indones Rangkuti et al.
179
Gambar 2 Seta yang terbentuk oleh Colletotrichum isolat kelompok ke-III pada medium agar-agar dekstrosa kentang.
± 600 pb
Gambar 3 Amplifikasi DNA 5 isolat Colletotrichum spp. asal pepaya, menggunakan pasangan primer universal ITS1/ITS4.M, Penanda DNA 1 kb ladder; K-, Kontrol negatif; B4, B8, B7, B19, Isolat asal buah dan; T15, isolat asal tangkai.
Gambar 1 Karakter morfologi Colletotrichum spp. kelompok I, II, dan III asal pepaya pada medium agar-agar dekstrosa kentang.
I
II
III
Tampak atas Tampak bawah Bentuk konidium

J Fitopatol Indones Rangkuti et al.
180
sementara itu pada suhu 32 °C C. truncatum tumbuh paling cepat dibandingkan dengan lainnya (Gambar 5).
PEMBAHASAN
Gejala awal antraknosa pada buah yang disebabkan oleh C. gloeosporioides ditandai dengan bercak oval, sedikit berair, membentuk lesio cekung pada permukaan buah yang akan berkembang menjadi nekrosis, dan kematian pada jaringan (Patel et al. 2005). Selama ini Colletotrichum sebagai penyebab penyakit antraknosa pada pepaya lebih dikenal sebagai penyakit pascapanen, akan tetapi proses infeksinya sudah dimulai sejak di pertanaman dan bersifat laten (Maeda dan Nelson 2014). Hasil survei lapangan menunjukkan bahwa akhir-akhir ini mulai banyak muncul gejala penyakit di pertanaman pepaya di sebagian wilayah Indonesia. Gejala penyakit antraknosa tersebut ditemukan selain pada bagian buah, juga terjadi pada tangkai daun dan batang. Berdasakan pustaka standar (Semangun 2007) serangan patogen ini pada bagian buah hanya terjadi ketika buah menjelang matang di lapangan dan diteruskan sampai pascapanen, akan tetapi dari hasil penelitian ini menunjukkan bahwa cendawan tersebut juga ditemukan menyerang pada buah yang masih hijau. Selain di Indonesia, peneliti lain sudah pernah melaporkan adanya
infeksi antraknosa di lapangan, tetapi hanya melaporkan kejadiannya pada buah (Haggag dan Singer 2013).
Hasil identifikasi secara morfologi dan/atau molekuler ditentukan tiga spesies Colletotrichum pada pepaya di pertanaman, yaitu C. gloeosporioides, C. magnum, dan C. truncatum. Berdasarkan warna koloni yang bervariasi, Colletotrichum dalam kelompok I ini diidentifikasi sebagai C. gloesporioides seperti yang dikemukakan oleh Rampersad (2011). Tipe morfologi kelompok I yang dicirikan dengan koloni berwarna abu-abu sampai abu-abu kehijauan (olive) dan terbentuknya aservulus dengan massa konidia berwarna oranye juga dikuatkan oleh peneliti lain sebagai C. gloeosporioides (Molina-Chaves et al. 2017; Torres-Calzada et al. 2013). Mengingat C. gloeosporioides oleh Weir et al. (2012) diketahui sebagai kompleks spesies (species complex), maka Colletotrichum tipe morfologi I yang dalam penelitian ini baru diidentifikasi secara morfologi masih perlu diklarifikasi lebih lanjut secara molekuler. Kelompok morfologi III dengan konidium melengkung (kurva) dan ujung meruncing pernah dilaporkan di Mesir (Haggag dan Singer 2013) dan Malaysia (Rahman et al. 2008) pada pepaya pascapanen. Kedua laporan tersebut berhasil mengidentifikasi Colletotrichum dengan tipe konidia ini sebagai C. capsici. Akan tetapi di dalam penelitian
Gambar 4 Pohon filogenetika spesies Colletotrichum spp. yang berasal dari buah dan tangkai pepaya di Kebumen, Indonesia. C. acutatum_THA_FJ972591 digunakan sebagai pembanding di luar group.
C. trucantum _B4
C. trucantum _B7C. trucantum _B8
C. magnum_MYS_JX971159C. trucantum_CHN_KC460308
C. trucantum_TWN_KX197397
C. acutatum_THA_FJ972591C. magnum_BRZ_HM163177
C. trucantum _B19
C. magnum_T15C. truncatum_THA_AY266372
51
51
100
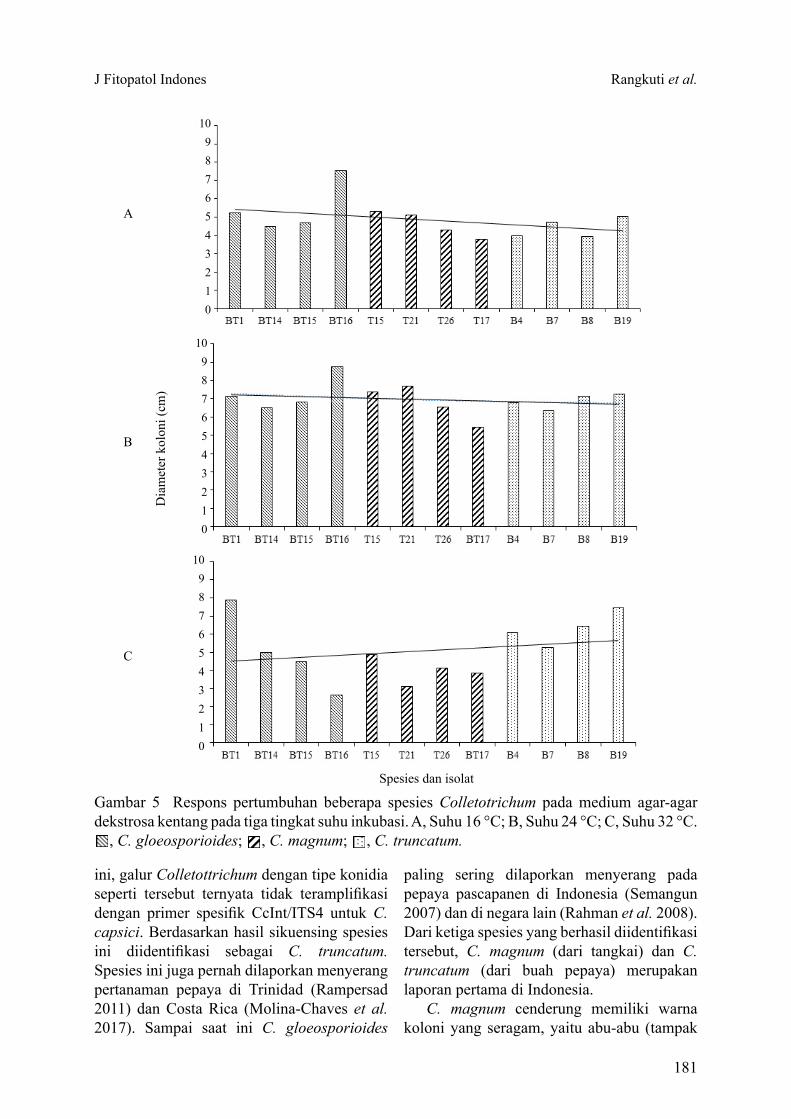
J Fitopatol Indones Rangkuti et al.
181
Gambar 5 Respons pertumbuhan beberapa spesies Colletotrichum pada medium agar-agar dekstrosa kentang pada tiga tingkat suhu inkubasi. A, Suhu 16 °C; B, Suhu 24 °C; C, Suhu 32 °C.
, C. gloeosporioides; , C. magnum; , C. truncatum.
A
B
C
Spesies dan isolat
Dia
met
er k
olon
i (cm
)
109876543210
109876543210
109876543210
ini, galur Colletottrichum dengan tipe konidia seperti tersebut ternyata tidak teramplifikasi dengan primer spesifik CcInt/ITS4 untuk C. capsici. Berdasarkan hasil sikuensing spesies ini diidentifikasi sebagai C. truncatum. Spesies ini juga pernah dilaporkan menyerang pertanaman pepaya di Trinidad (Rampersad 2011) dan Costa Rica (Molina-Chaves et al. 2017). Sampai saat ini C. gloeosporioides
paling sering dilaporkan menyerang pada pepaya pascapanen di Indonesia (Semangun 2007) dan di negara lain (Rahman et al. 2008). Dari ketiga spesies yang berhasil diidentifikasi tersebut, C. magnum (dari tangkai) dan C. truncatum (dari buah pepaya) merupakan laporan pertama di Indonesia.
C. magnum cenderung memiliki warna koloni yang seragam, yaitu abu-abu (tampak

J Fitopatol Indones Rangkuti et al.
182
atas) dan abu-abu kehitamaan (tampak bawah). Dari bentuk konidium, C. gloeosporioides dan C. magnum memiliki bentuk yang sama, yaitu silinder dengan kedua ujung membulat. Keduanya dapat dibedakan dari ada tidaknya gelembung minyak pada konidium. C. truncatum yang ditemukan dalam penelitian ini mudah dibedakan dari dua spesies lainnya. Koloni spesies ini membentuk cincin konsentris, konidium melengkung dan membentuk seta pada medium ADK.
Colletotrichum yang diisolasi dari buah, tangkai daun dan batang di lapangan ialah C. truncatum, C. magnum dan C. gloeosporioides. Dari penelitian ini, C. truncatum hanya ditemukan di lapangan pada bagian buah, sedangkan C. magnum sebagian besar (3 dari 4 isolat) ditemukan pada tangkai daun. Sementara itu C. gloeosporioides merupakan spesies yang dominan dan ditemukan pada semua bagian tanaman. Berdasarkan hasil identifikasi molekuler, isolat asal buah dengan konidium melengkung (isolat kelompok III) menunjukkan hubungan kekerabatan yang paling dekat dengan spesies C. truncatum asal Malaysia, sedangkan salah satu galur (T15) asal tangkai dengan konidium silindris dan tanpa gelembung minyak (tipe morfologi II) yang diperoleh dalam penelitian ini memiliki hubungan kekerabatan dengan C. magnum asal Brazil. Keberadaan C. magnum sebagai patogen antraknosa pada tanaman pepaya juga pernah dilaporkan oleh peneliti lain di Meksiko (Tapia-Tussell et al. 2016) dan Costarica (Molina-Chaves et al. 2017).
Dari percobaan pengaruh suhu, per-tumbuhan Colletotrichum asal pepaya dipengaruhi oleh spesies dan suhu. Secara umum, ketiga spesies Colletotrichum tumbuh optimum pada suhu 24 °C dan relatif sama, tetapi menunjukkan perbedaan ketika ditumbuhkan pada suhu yang lebih rendah (16 °C)dan suhu lebih tinggi (32 °C). Pada suhu yang lebih rendah (16 °C), C. gloesporioides cenderung tumbuh lebih cepat dibandingkan dua spesies lainnya. Sementara itu C. truncatum tumbuh lebih cepat pada suhu yang lebih tinggi (32 °C) dibandingkan dua speseis lainnya. Sangeetha dan Rawal (2010) melaporkan
bahwa konidium C. gloeosporioides belum mampu berkecambah pada suhu 15–20 °Cdan 30 °C. Penelitian Smith dan Black (1990) juga menunjukkan adanya perbedaan respons pertumbuhan koloni tiga spesies Colletotrichum pada stroberi terhadap suhu, dimana salah satu spesies tumbuh lebih lambat dibandingkan dua spesies lainnya pada semua kisaran suhu yang diujikan. Dengan demikian, perbedaan respons pertumbuhan koloni terhadap suhu dapat digunakan sebagai salah satu faktor dalam membedakan tiga spesies Colletotrichum pada tanaman pepaya.
UCAPAN TERIMA KASIH
Penghargaan dan terima kasih disampaikan kepada Lembaga Pengelola Dana Pendidikan (LPDP), Kementerian Keuangan Republik Indonesia melalui Beasiswa Tesis Batch II Tahun 2016.
DAFTAR PUSTAKA
Cannon PF, Damm U, Johnston PR, Weir BS. 2012. Colletotrichum-current status and future directions. Stud Mycol. 73:181–213. DOI: https://doi.org/10.3114/sim0014.
Choi YW, Hyde KD, Ho WH. 1999. Single spore isolation of fungi. Fungal Divers. 3:29–38.
Haggag WM, Singer S. 2013. First report of Colletotrichum capsici causing pre and postharvest anthracnose on papaya in Egypt. Int J Engineer Innov Technol. 3(6):151–152.
Maeda C, Nelson S. 2014. Anthracnose of papaya in Hawai’i. Mãnoa (US): College of Tropical Agriculture and Human Resources, University of Hawai’i.
Molina-Chaves A, Gómeⱬ-Alpἰⱬar L, Umaṅa-Rojas G. 2017. Identificación de especies del género Colletotrichum asociadas a la antracnosis en papaya (Carica papaya L.) en Costa Rica. Agronomía Costarricense 41(1): 69-80. DOI: https://doi.org/10.15517/rac.v41i1.29752.
Patel RV, Joshi KR, Solanky KU, Sabalpara AN. 2005. Colletotrichum gloeosporioides:

J Fitopatol Indones Rangkuti et al.
183
a new leaf spot pathogen of turmeric in Gujarat. Ind J Phytopathol. 58 (1):125.
Photita W, Taylor PWJ, Ford R, Hyde KD, Lumyong S. 2005. Morphological and molecular characterization of Colletotrichum species from herbaceous plants in Thailand. Fungal Divers. 18:117–133.
Rahman MA, Mahmud TMM, Kadir J, Abdul Rahman R, Begum MM. 2008. Major postharvest fungal diseases of papaya cv. Sekaki in Selangor, Malaysia. Pertanika J Trop Agric Sci. 31(1):27–34.
Rampersad SN. 2011. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease of papaya in Trinidad. Plant Dis. (95): 1244–1254. DOI: https://doi.org/10.1094/PDIS-02-11-0080.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning: a Laboratory Manual. Ed ke-2. New York (US): Cold Spring Harbor.
Sangeetha CG, Rawal RD. 2010. Temperature requirement of different isolates of Colletotrichum gloeosporioides isolated from mango. Afr J Biotechnol. 9(21):3086−3090.
Sato T, Moriwaki I, Kaneko S. 2015. Anthracnose fungi with curved conidia Colletotrichum spp. belonging to ribosomal groups 9-13, and their host ranges in Japan. JARQ. 49(4):351–362. DOI: https://doi.org/10.6090/jarq.49.351.
Semangun H. 2007. Penyakit-penyakit Tanaman Hortikultura di Indonesia. Yogyakarta (ID): Gadjah Mada University Press.
Smith BJ, Black LL. 1990. Morphological, cultural, and pathogenic variation among
Colletotrichum species isolated from strawberry. Plant Dis. 74:69–76. DOI: https://doi.org/10.1094/PD-74-0069.
Tapia-Tussell R, Cortes-Velazquez A, Valencia-Yah T. 2016. First report of Colletotrichum magnum causing anthracnose in papaya in Mexico. Plant Dis. 100(11):2323. DOI: https://doi.org/10.1094/PDIS-03-16-0324-PDN.
Torres-Calzada C, Tapia-Tussel R, Higuera-Ciapara I, Perez-Brito D. 2013. Morphological, pathological and genetic diversity of Colletotrichum species responsible for anthracnose in papaya (Carica papaya L). Eur J Plant Pathol. 135(1):67-79. DOI: https://doi.org/10.1007/s10658-012-0065-7.
Watanabe T. 2002. Pictorial Atlas of Soil and Seed Fungi Morphologies of Cultured Fungi and Key to Species. Ed ke-2. Boca Raton (US). CRC Pr. DOI: https://doi.org/10.1201/9781420040821.
Weir BS, Johnston PR, Damm U. 2012. The Colletotrichum gloeosporioides species complex. Stud Mycol. 73:115–180. DOI: https://doi.org/10.3114/sim0011.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Di dalam: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editor. PCR Protocols: A Guide to Methods and Applications. San Diego (US): Academic Pr. hlm 315–322. DOI: https://doi.org/10.1016/B978-0-12-372180-8.50042-1.
Zakaria M, Bailey JA. 2000. Morphology and cultural variation among Colletotrichum isolates obtained from tropical forest nurseries. J Trop For Sci. 12(1):1–20.

184
ISSN: 0215-7950
Volume 13, Nomor 5, September 2017Halaman 184–190
DOI: 10.14692/jfi.13.5.184
*Alamat penulis korespondensi: Fakultas Pertanian, Universitas Udayana, Jalan PB Sudirman, Denpasar, Bali 80225. Tel: 0361-255346, Faks: 0361-255346; surel: [email protected]
Penyakit Layu Bakteri Stewart pada Jagung di Bali
Stewart’s Wilt Disease of Maize in Bali
I Gede Rai Maya Temaja*, G.N. Alit Susanta Wirya, Ni Made Puspawati, Khairun Nisak Syahdu
Universitas Udayana, Denpasar 80225
ABSTRAK
Penyakit layu stewart merupakan penyakit serius pada tanaman jagung manis (Zea mays). Gejala khas penyakit ini ialah garis berwarna hijau pucat sampai kekuningan yang sejajar dengan tulang daun. Gejala seperti ini ditemukan pada daun jagung manis di daerah Denpasar, Tabanan, Gianyar dan Karangasem pada tahun 2015. Deteksi patogen dengan metode polymerase chain reaction dilakukan menggunakan DNA total yang diisolasi dari daun tanaman bergejala dan pasangan primer CPSL1/CPSR2c. Amplikon dengan ukuran ~1100 pb berhasil diperoleh dari sampel asal Denpasar. Analisis sikuen nukleotida memastikan bahwa gejala penyakit layu stewart tersebut disebabkan oleh bakteri Pantoea stewartii subsp. stewartii. Hasil analisis sikuen menunjukkan bahwa P. stewartii subsp. stewartii isolat Bali memiliki tingkat homologi yang tinggi (98.97–99.08%) dan berada dalam satu clade dengan isolat asal Kanada, Amerika Serikat, dan Jepang. Ini merupakan laporan pertama mengenai P. stewartii subsp. stewartii yang menginfeksi tanaman jagung di Provinsi Bali.
Kata kunci: analisis sikuen, nukleotida, Pantoea, Zea mays
ABSTRACT
Stewart’s wilt is a serious disease of sweet corn (Zea mays). The typical symptoms of the disease are pale-green to yellow linear streaks parallel to the veins. The symptoms were observed on sweet corn in Denpasar, Tabanan, Gianyar, and Karangasem areas during a survey in 2015. Pathogen detection based on a polymerase chain reaction was carried out using total DNA obtained from symptomatic leaf samples and the pairs of primers, CPSL1/CPSR2c. The expected sized (~1100 bp) amplicon was detected in samples from Denpasar. Sequence analysis confirmed that Stewart’s wilt disease symptoms are caused by Pantoea stewartii subsp. stewartii. Nucleotide sequence and phylogenetic analysis showed that P. stewartii subsp. stewartii from Bali has high homology (98.97-99.08 %) and placed in the same clade with isolates from Canada, USA and Japan. This is the first report of P. stewarti subsp. stewartii on corn in Bali.
Key words: nucleotide, sequence analysis, Pantoea, Zea mays

J Fitopatol Indones Temaja et al.
185
PENDAHULUAN
Jagung (Zea mays) adalah salah satu bahan pangan pokok di Indonesia dan juga merupakan komponen utama pangan dan pakan ternak dan unggas. Kebutuhan jagung di Indonesia belum bisa dipenuhi dari produksi dalam negeri, sehingga masih harus dilakukan impor. Pada tahun 2013 impor jagung sebesar 3.19 juta ton, volume impor ini naik 76.94 % dibandingkan dengan tahun 2012. Pada tahun 2014 volume impor jagung belum berubah dari tahun sebelumnya yaitu sebesar 3.17 juta ton (Pusat Data dan Sistem Informasi Pertanian Kementan 2015).
Tingginya impor jagung ke Indonesia, meningkatkan kemungkinan masuk dan tersebarnya OPTK A1 yang dapat terbawa melalu biji jagung, salah satunya ialah Pantoea stewartii subsp. stewartii. Bakteri tersebut merupakan penyebab penyakit layu stewart yang endemik di Amerika dan telah tersebar di beberapa negara di dunia (OEPP/EPPO 2006). Bakteri ini menyebabkan layu vaskular dan hawar pada daun (Coplin et al. 2002). Penyakit layu stewart telah menimbulkan masalah besar bagi negara produsen jagung, yaitu mengakibatkan kehilangan hasil. Pada jagung manis, kehilangan hasil berhubungan dengan infeksi sistemik, yaitu pengurangan hasil 0.8% untuk setiap insidensi penyakit 1% dari bibit tanaman terinfeksi sistemik (Freeman dan Pataky 2001).
P. stewartii subsp. stewartii pertama kali dilaporkan menginfeksi tanaman jagung di Bogor, Jawa Barat pada tahun 2014 (Rahma et al. 2014). Gejala penyakit layu stewart ditandai dengan perubahan warna daun dari hijau pucat ke kuning, disertai garis linear teratur atau tepinya bergelombang sejajar dengan tulang daun. Survei pada tahun 2015 menemukan penyakit yang menunjukkan gejala menyerupai infeksi penyakit layu stewart pada daun jagung manis di Desa Kesiman Kertalangu, Kota Denpasar. Penyakit seperti ini belum pernah dijumpai pada tanaman jagung di Bali. Oleh karena itu, penelitian ini bertujuan mengidentifikasi penyebab gejala layu stewart pada tanaman jagung di Bali.
BAHAN DAN METODE
Survei PenyakitSurvei untuk mendapatkan status
penyebaran penyakit layu stewart dilakukan di semua kabupaten dan kota di Bali. Pengamatan meliputi tanaman jagung yang menunjukkan gejala layu stewart. Pengambilan sampel di lapangan dilakukan pada tanaman yang bergejala secara acak. Sampel tanaman jagung dicabut dan dimasukkan ke dalam plastik untuk selanjutnya dideteksi patogen penyebabnya dengan metode polymerase chain reaction (PCR).
Insidensi penyakit (IP) dan keparahan penyakit (KP) diukur menggunakan rumus:
IP = nN × 100%, dengan
n, jumlah tanaman yang terserang; dan N, jumlah tanaman yang diamati.
N × VKP = × 100%, dengan(ni × vi)∑
i
i=0
ni, jumlah tanaman dengan skor ke-i; vi, nilai skala dari tiap kategori serangan (Tabel 1); N, jumlah tanaman yang diamati; dan V, skor tertinggi.
Deteksi Patogen dengan PCREkstraksi DNA dan Amplifikasi Gen cps.
DNA total diekstrak dari daun jagung bergejala menggunakan kit ekstraksi DNeasy Plant Mini Kit (Qiagen). Amplifikasi gen capsule
Tabel 1 Skala dan tingkat serangan Pantoea stewartii subsp. stewartii pada tanaman jagung
Skala Tingkat serangan
(%)
Kriteria serangan
Tingkat virulensi
1 < 1 Sehat Tidak virulen2 1 Sangat ringan Kurang virulen3 2 Sangat ringan Kurang virulen4 3 Sangat ringan Kurang virulen5 4 Sangat ringan Kurang virulen6 5–25 Ringan Virulen7 25–50 Sedang Virulen8 50–90 Berat Sangat virulen9 > 90 Sangat berat Sangat virulen
Sumber: Rahma et al. (2010)

J Fitopatol Indones Temaja et al.
186
biosynthesis (cps) dilakukan dengan teknik PCR menggunakan pasangan primer CPSL1 (5'-CCTGTCAGTCTCGAACC-3'); dan CPSR2c (5'-ATCTCGAACCGGTAACC-3')dengan produk PCR 1100 pb. Primer CPSL1 dan CPSR2c yang dirancang dari sikuen daerah cpsD mengode sintesis kapsul polisakarida stewartan yang spesifik untuk P. stewartii (Coplin et al. 2002). DNA diamplifikasi pada volume reaksi 25 µL yang mengandung PCR Bead, 1 µL ( 0.4 µM )primer CPSL1, 1 µL (0.4 µM ) primer CPSR2c, 22 µL nuclease free water, dan 1 µL template DNA. Amplifikasi dilakukan pada Gene Amp PCR System 9700 thermocycler, dengan program: predenaturasi pada 94 °C selama 5 menit, dilanjutkan dengan 25 siklus denaturasi 94 °C selama 1 menit; aneling 54 °Cselama 1 menit dan ekstensi 72 °C selama 30 detik. Siklus terakhir ditambah tahapan sintesis 72 °C selama 10 menit, kemudian siklus berakhir dengan suhu 4 °C. Produk PCR dielektroforesis pada gel agarosa 1.2% yang dijalankan pada 80 V selama 1 jam. Gel diwarnai dalam 2 µg mL-1 etidium bromida dalam bufer elektroforesis (40 mM Tris, 20 mM natrium asetat, dan 1 mM EDTA pH 7.0) dan divisualisasi pada transluminator UV (Bio-Rad Translluminator 2000).
Sikuensing dan Analisis Filogenetika. Analisis sikuen dilakukan oleh PT. Genetika Science Indonesia. Sikuen DNA selanjutnya diedit menggunakan piranti lunak ChromasPro versi 1.5. Analisis sikuen untuk mengetahui tingkat homologinya dengan sikuen yang telah didepositkan pada GenBank dilakukan dengan piranti lunak basic local alignment search tool (BLAST) melalui situs: http://blast.ncbi.nlm.nih.gov/Blast.cgi (Thompson et al. 1997). Data sikuen nukleotida yang terpilih kemudian dianalisis kespesifikan nukleotidanya dengan program multiple alignment (ClustalW) yang terintegrasi pada program piranti lunak molecular evolutionary genetics analysis (MEGA) versi 6.0 (Tamura et al. 2013). Pohon filogenetika dikonstruksi menggunakan piranti lunak PAUP 4.0b10 dengan metode neighbor joining (NJ) berdasarkan pendekatan maximum parsimony
(MP) dan bootstrap 1000 ulangan (Brady et al. 2012). Selanjutnya, pohon filogenetika ditampilkan menggunakan piranti lunak TreeGraph 2.0 (Stover dan Muller 2010).
HASIL
Keberadaan pertanaman jagung, terutama jagung manis sangat berkaitan erat dengan munculnya gejala layu stewart yang disebabkan oleh bakteri P. stewartii subsp. stewartii. Pada penelitian ini gejala tanaman jagung yang terinfeksi layu stewart terdapat di Desa Kesiman Kertalangu, Kota Denpasar; Desa Kediri, Kabupaten Tabanan; Desa Blahbatuh, Kabupaten Gianyar; dan Desa Manggis, Kabupaten Karangasem. Gejala layu stewart yang menginfeksi tanaman jagung di daerah tersebut ialah adanya garis lurus berwarna hijau pucat sampai kekuningan sejajar dengan tulang daun pada daun (Gambar 1). Insidensi penyakit tertinggi (23%) terjadi di Desa Kesiman Kertalangu, Denpasar, dan paling rendah (11%) di Desa Manggis, Karangasem. Sedangkan keparahan penyakit tertinggi terdapat di kota Denpasar dan Kabupaten Gianyar sebesar 10% dan terendah terdapat di Kabupaten Karangasem yaitu sebesar 3% (Tabel 2). Semua tanaman yang menunjukkan gejala merupakan varietas jagung manis.
Deteksi dengan PCR hanya berhasil mengamplifikasi fragmen DNA dari sampel asal Desa Kesiman Kertalangu, Kota Denpasar. Fragmen DNA berukuran ~1100 pb didapatkan dari amplifikasi menggunakan pasangan primer CPSL dan CPSR2c yang mengamplifikasi gen cps (Gambar 2). Fragmen DNA yang dihasilkan selanjutnya dipurifikasi dan disikuensing untuk mengidentifikasi bakteri berdasarkan kemiripan sikuen nukleotida dengan spesies bakteri yang telah teridentifikasi di GenBank. Berdasarkan hasil penjajaran sikuen, bakteri isolat Denpasar (yang selanjutnya dinamakan isolat Bali) memiliki kekerabatan yang dekat dengan P. stewartii subsp. stewartii DOAB 021 (EU215383.1) asal Kanada, P. stewartii subsp. stewartii DC283 (AF077292.2) asal USA, P. stewartii subsp. stewartii cpsD

J Fitopatol Indones Temaja et al.
187
Lokasi Insidensi penyakit(%)
Keparahan penyakit(%)
Gejala penyakit
Denpasar 23 10 Garis kuning membujur sejajar tulang daunTabanan 16 5 Garis kuning membujur sejajar tulang daunGianyar 21 10 Garis kuning tak beraturan sejajar tulang daunKarangasem 11 3 Bercak kuning pada daun dan layu
Tabel 2 Insidensi dan keparahan penyakit layu stewart di Provinsi Bali
Gambar 1 Gejala layu stewart di pertanaman jagung di Desa Kesiman Kertalangu, Denpasar. Garis lurus berwarna hijau pucat sampai kekuningan yang sejajar dengan tulang daun.
Gambar 2 Hasil amplifikasi gen cps P. stewartii subsp. stewartii menggunakan primer CPSL1 dan CPSR2c Sampel berasal dari tanaman jagung di Provinsi Bali. 1 dan 2, Denpasar; 3 dan 4, Tabanan; 5 dan 6, Gianyar; 7 dan 8, Karangasem; M, Penanda DNA 100 pb (Fermentas).
1 2 3 4M 5 6 7 8
1100 pb
(AB894429.1) asal Jepang, dan P. stewartii subsp. stewartii DOAB 022 (EU215384.1) asal Kanada (Tabel 3). Hasil analisis pohon filogenetika menggunakan metode NJ dengan 1000 kali ulangan bootstrap dan pendekatan MP menunjukkan bahwa isolat Bali ialah P. stewartii subsp. stewartii karena berada pada satu kelompok dengan sikuen-sikuen bakteri P. stewartii subsp. stewartii dengan dukungan 100% bootstrap support (Gambar 3).
PEMBAHASAN
Gejala penyakit layu stewart ditemukan pada tanaman jagung di Bali. Gejala layu stewart yang terdapat pada pertanaman jagung ini sesuai dengan yang diungkapkan oleh Rahma et al. (2010). Keparahan penyakit menunjukkan bahwa tingkat infeksi penyakit di lapangan tergolong ringan; belum ditemukan adanya vektor kumbang Chaetocnema pulicaria di lapangan.
Amplifikasi berhasil mendapatkan fragmenberukuran ~1100 pb hanya dari sampel daun
asal Denpasar, sesuai dengan target yang diharapkan. Sampel dari daerah lain tidak teramplifikasi dikarenakan sampel tersebut tidak terinfeksi bakteri P. stewartii subsp. stewartii. Hal ini sesuai dengan hasil uji morfologi koloni bakteri yaitu hanya sampel asal Denpasar yang dapat dikulturkan. Diagnosis menggunakan primer spesifik cukup baik digunakan untuk P. stewartii subsp. stewartii apalagi bila dikombinasikan dengan analisis matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) (Wensing et al.2010).
Kedekatan jarak genetika antara isolat P. stewartii subsp. stewartii isolat Bali dan isolat P. stewartii subsp. stewartii dari Kanada, USA, dan Jepang dimungkinkan karena adanya lalu lintas komoditas pertanian antarnegara dan antararea sehingga bakteri tersebut dapat terbawa melalui benih yang masuk ke wilayah Provinsi Bali. Penyakit layu stewart yang disebabkan oleh bakteri P. stewartii subsp. stewartii muncul pertama kali di Indonesia pada tahun 2010 (Rahma et al. 2010). Hasil

J Fitopatol Indones Temaja et al.
188
Isol
atN
o. A
kses
i G
ene
Ban
kSi
mila
ritas
(%)
12
34
56
78
910
1112
1314
1516
1718
1920
21ET
ET1
99
CU
4681
35.1
ID
EP D
SM 1
2163
FN
3922
35.1
82.0
1 ID
EP
Ep1
96
FP
2368
42.1
82.0
3 10
0ID
PA
PA
13C
P003
085.
170
.10
67.1
7 67
.28
ID
PA L
MG
534
2
HE6
1716
0.1
69.9
3 67
.17
67.2
8 99
.90
ID
Isol
at B
ali
na67
.33
66.0
5 66
.16
87.1
5 87
.05
ID
PSS
DO
AB
021
EU
2153
83.1
67.1
6 65
.89
65.8
3 86
.95
86.8
4 98
.97
ID
PSS
DC
283
A
F077
292.
267
.33
66.0
5 66
.00
86.9
5 86
.84
98.9
7 99
.79
ID
PSS
cpsD
AB
8944
29.1
67.3
3 66
.05
66.0
0 87
.05
86.9
5 99
.08
99.9
0 99
.90
ID
PSS
DO
AB
022
EU
2153
84.1
67.3
3 66
.05
66.0
0 87
.05
86.9
5 99
.08
99.9
0 99
.90
100
ID
PR N
D04
C
P009
454.
162
.64
62.9
9 63
.05
76.4
1 76
.58
74.0
3 73
.71
73.8
7 73
.87
73.8
7 ID
C
N N
D02
C
P011
427.
160
.57
61.0
9 62
.22
76.0
9 76
.25
74.1
7 73
.83
74.0
0 74
.00
74.0
0 85
.20
ID
SL A
TCC
275
92
CP0
0625
2.1
42.5
2 44
.40
44.4
4 45
.74
45.6
0 44
.72
45.2
7 45
.27
45.2
7 45
.27
45.7
8 46
.11
ID
PA A
J133
55 D
NA
AP0
1203
2.2
40.7
2 38
.70
38.6
3 44
.43
44.2
5 44
.96
45.3
0 45
.47
45.4
7 45
.47
41.1
0 40
.62
38.5
1 ID
PA
R10
0
CP0
1420
7.1
40.9
6 38
.70
38.6
3 44
.25
44.0
8 44
.96
45.3
0 45
.47
45.4
7 45
.47
41.1
0 40
.38
38.7
8 99
.07
ID
PA L
MG
201
03C
P001
875.
240
.48
38.4
6 38
.39
44.0
8 44
.08
44.4
4 44
.79
44.9
6 44
.96
44.9
6 40
.98
40.2
6 38
.66
99.0
7 99
.28
ID
PAG
FD
AA
R-
GO
S 16
0
CP0
1412
9.1
41.4
7 40
.38
40.4
3 44
.86
44.6
9 45
.05
45.0
5 45
.22
45.2
2 45
.22
42.9
7 42
.16
38.7
6 81
.98
82.0
581
.61
ID
PV C
9 1
CP0
0220
6.1
42.2
3 41
.03
41.0
8 46
.18
46.0
1 46
.54
46.5
4 46
.71
46.7
1 46
.71
44.1
1 43
.14
39.0
9 81
.09
81.0
8 80
.63
96.5
4ID
EB
Eb6
61
FP23
6843
.137
.91
37.5
3 37
.57
43.4
6 43
.29
44.6
7 43
.82
43.9
9 43
.99
43.9
9 39
.56
38.7
1 36
.55
57.9
7 57
.85
57.8
558
.65
58.9
4 ID
EA
ATC
C B
AA
21
58
FR71
9191
.141
.70
40.1
2 40
.06
41.4
4 41
.27
41.4
6 41
.46
41.4
6 41
.46
41.4
6 40
.31
40.0
2 39
.81
60.6
9 61
.10
60.7
7 59
.53
58.4
5 72
.84
ID
EA C
FBP1
430
FN43
4113
.140
.59
39.4
1 39
.80
41.5
2 41
.36
41.3
8 41
.38
41.3
8 41
.38
41.3
8 39
.93
38.3
2 39
.44
60.2
6 60
.68
60.3
5 59
.35
58.2
7 73
.42
99.7
5ID
Tabe
l 3 M
atrik
sim
ilarit
as P
. ste
warti
i sub
sp. s
tew
artii
isol
at B
ali d
an is
olat
hom
olog
inya
yan
g te
rdaf
tar d
i Gen
eBan
k be
rdas
arka
n sik
uen
gen
cps (
%)
*na:
bel
um d
idaf
tar d
i Gen
eban
k; ID
: sik
uen
iden
tik; E
T: E
rwin
ia ta
sman
iens
is ;
EP: E
rwin
ia p
yrifo
liae;
PA
: Pan
toea
ana
natis
; PSS
: Pan
toea
stew
artii
Sub
sp. s
tew
artii
; PR
: Pan
toea
rwan
dens
is; C
N: C
edec
ea
nete
ri; S
L: S
erra
tia li
quef
acie
ns; P
AG
: Pan
toea
agg
lom
eran
s; P
V: P
anto
ea v
agan
s; E
B: E
rwin
ia b
illin
giae
; EA
:Erw
inia
am
ylov
ora

J Fitopatol Indones Temaja et al.
189
Gambar 3 Filogenetika kekerabatan P. stewartii subsp. stewartii isolat Bali terhadap galur-galur homologinya yang terdaftar di GenBank berdasarkan gen cps. Hasil analisis menggunakan metode neighbor joining dengan menggunakan 1000 kali ulangan bootstrap dan pendekatan maximum parsimony. Isolat Bali adalah P. stewartii subsp. stewartii dengan dukungan 100 % bootstrap support.
diagnosis P. stewartii subsp. stewartii di Provinsi Bali menunjukkan penyebaran yang lebih luas di Indonesia, padahal sesuai dengan Peraturan Menteri Pertanian No.51 tahun 2015 P. stewartii subsp. stewartii merupakan organisme pengganggu tumbuhan karantina (OPTK) golongan A1.
Berdasarkan hasil identifikasi molekuler dapat disimpulkan bahwa isolat bakteri yang berasal dari Desa Kesiman Kertalangu, Denpasar merupakan bakteri P. stewartii subsp. stewartii yang menyebabkan penyakit layu stewart pada tanaman jagung di Denpasar. Penyakit ini baru ditemukan hanya di kota Denpasar dan belum pernah dilaporkan di Provinsi Bali. Penelitian dasar untuk mengetahui karakter molekuler dan biologi P. stewartii subsp. stewartii perlu dilakukan sebagai landasan menyusun strategi pengendalian.
UCAPAN TERIMA KASIH
Penelitian ini dibiayai oleh Direktorat Riset dan Pengabdian Masyarakat, Direktorat Jenderal Penguatan Riset dan Pengembangan, Kementerian Riset, Teknologi, dan Pendidikan Tinggi melalui Penelitian Fundamental dengan Surat Perjanjian Penugasan Pelaksanaan Penelitian No. 486.132/UN14.2/PNL.01.03.00/2016.
DAFTAR PUSTAKA
Brady CL, Cleenwerck I, Westhuizen LVD, Venter SN, Coutinho TA, Vos PD. 2012. Pantoea rodasii sp. nov., Pantoea rwandensis sp. nov. and Pantoea wallisii sp. nov., isolated from eucalyptus. Int J Syst Evol Microbiol. 62:1457–1464. DOI: https://doi.org/10.1099/ijs.0.032615-0.
Isolat BaliEU215384.1 Pantoea stewartii subs stewartii DOAB 22EU215383.1 Pantoea stewartii subs stewartii DOAB 2AF077292.2 Pantoea stewartii subs stewartii DC283AB894429.1 Pantoea stewartii subs stewartii cpsDHE617160.1 Pantoea ananatis LMG 5342CP003085.1 Pantoea ananatis PA 13CP011427.1 Cadecea neteri ND02CP009454.1 Pantoea rwandensis ND04FP236842.1 Erwinia pyrifoliae Ep1 96FN392235.1 Erwinia pyrifoliae DSM 12163CU468135.1 Erwinia tasmaniensis ET1 99FR719191.1 Erwinia amylovora ATCC BAA 2158FN434113.1 Erwinia amylovora CFBP1430FP236843.1 Erwinia billingiae Eb661CP014207.1 Pantoea ananatis R100CP001875.2 Pantoea ananatis LMG 20103AP012032.2 Pantoea ananatis AJ13355 DNACP014129.1 Pantoea Agglomerans FDAARGOS 160CP002206.1 Pantoea vagans C9 1CP006252.1 Serratia liquefaciens ATCC 27592EU898724 Escherichia coli TW14359 16S
100
100
100
100100
100
100
100
99.9
99.9
99.9
98.9
54.8
75.8
85.1
85.7
83.9
91.5
0.0 100.0

J Fitopatol Indones Temaja et al.
190
Coplin DL, Majerczak DR, Zhang Y, Kim WS, Jock S, Geider K. 2002. Identification of Pantoea stewartii subsp. stewartii by PCR and strain differentiation by PFGE. Plant Dis. 86:304–311. DOI: https://doi.org/10.1094/PDIS.2002.86.3.30.
Dharmayanti NLPI. 2011. Filogenetika molekuler: metode taksonomi organisme berdasarkan sejarah evolusi. Wartazoa. 21(1):15–18.
Freeman ND, Pataky JK. 2001. Levels of Stewart’s wilt resistance necessary to prevent reductions in yield of sweet corn hybrids. Plant Dis. 85:1278–1284. DOI: https://doi.org/10.1094/PDIS.2001.85.12.1278.
OEPP/EPPO. 2006. Pantoea stewartii Subsp. stewartii. Bulletin OEPP/EPPO. 36: 111–115. DOI: https://doi.org/10.1111/j.1365-2338.2006.00920.x.
[PDSIPKM] Pusat Data dan Sistem Informasi Pertanian Kementerian Pertanian. 2015. Outlook Komodiitas Pertanian Tanaman Pangan. Jagung. PDSIPKM.
Rahma H, Nurbailis, Liswarni Y, Puspita Y. 2010. Uji virulensi beberapa isolat Pantoea stewartii penyebab layu stewart pada bibit jagung (Zea mays L). Manggaro. 11(1):12–17.
Rahma H, Sinaga MS, Surahman M, Giyanto. 2014. First report of Stewart’s wilt of maize caused by Pantoea stewartii subsp. stewartii in Bogor district, Indonesia. J. ISSAAS 20(2):131–141.
Stover BC, Muller KF. 2010. TreeGraph 2: combining and visualizing evidence from different phylogenetic analysis. BMC Bioinformatics 11(7):1–9. DOI: https://doi.org/10.1186/1471-2105-11-7.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 30(12):2725–2729. DOI: https://doi.org/10.1093/molbev/mst197.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. The clustal X windows interfase: flexible strategies for multiple sequence alignment aided by quality analysis tool. Nucleic Acids Res. 25(24):4876–4882. DOI: https://doi.org/10.1093/nar/25.24.4876.
Wensing A, Zimmermann S, Geider K. 2010. Identification of the corn pathogen Pantoea stewartii by mass spectrometry of whole-cell extracts and its detection with novel PCR primers. Appl. Environ. Microbiol. 76(18):6248–6256. DOI: https://doi.org/10.1128/AEM.01032-10.





























