Embed Size (px)
DESCRIPTION
Klasszikus genetika és evolúciógenetika. Fedor Anna [email protected]. Félévi tematika. szept. 10: Müller Viktor: Életdefiníció szept. 17. Kun Ádám: Az élet eredete szept. 24. Kun Ádám: Anyagcsere okt. 1. Fedor Anna: Genetika és evolúciógenetika - PowerPoint PPT Presentation
Citation preview

Félévi tematikaszept. 10: Müller Viktor: Életdefiníciószept. 17. Kun Ádám: Az élet eredeteszept. 24. Kun Ádám: Anyagcsereokt. 1. Fedor Anna: Genetika és evolúciógenetikaokt. 8. Gedeon Gábor: Egyedfejlõdés és szabályozásokt. 15. Gedeon Gábor: Egyedfejlõdésokt. 22. Zachár István: Makrotaxonómia és replikátoroknov. 5. Garay József: Evolúciónov. 12. Müller Viktor: Immunológianov. 19. Fedor Anna: Neurobiológianov. 26. Czárán Tamás: Ökológiadec. 3. Számadó Szabolcs: Kommunikáció, kooperációdec. 10. Számadó Szabolcs: Kultúra

A mai óra témái
I. Mendel törvényei II. Eltérés a mendeli arányoktól III. A szex evolúciója IV. A nem meghatározás módjai V. Az XY rendszer következményei VI. Kvantitatív genetika VII. Evolúciógenetika

I. A genetika alaptörvényei az emlősök szőrszínének példáján
(gimnáziumi ismétlés egy új példán)

A gén: sárga sáv a szőrszálakban/sima szín
Az „A” gén két allélje: A és a.A → sárga sáv → agouti szőrszín („grízes
szürkés” kinézet, a fekete szőrszálakban sárga sáv jelenik meg),
a → sima szín → fekete szőrszín(A lovak háziasítása során az A allél
elveszett, tehát az összes ló aa.)

Fenotípusok:
Diploid geno- és fenotípusok: AA genotípus → agouti fenotípus aa genotípus → fekete fenotípus Aa genotípus → agouti fenotípust
eredményez, mivel A domináns és a recesszív.
A_-lal jelöljük a genotípust, ha lehet AA és Aa is.

Mendel I.: Uniformitás
Tiszta vonalból származó, homozigóta szülők keresztezésével nyert utódok egységes geno- és fenotípusúak.
P: AA x aaF1 : Aa

Mendel II.: Hasadás
A szülői tulajdonságokért felelős gének nem olvadnak össze az F1 generációban, a szülői tulajdonságok újra megjelennek az F2 nemzedékben.
F1 : Aa x Aa 100% agouti
F2 : ¼ AA, ½ Aa, ¼ aa ¾ agouti ¼ fekete

B gén: fekete/barna alapszín
B allél → fekete színanyagb allél → barna színanyagDomináns-recesszív öröklődés

A és B gén kombinációi
A_B_ → agouti (feketében sárga sávok, grízes szürkés)
aaB_ → fekete A_bb → fahéj (barnában sárga
sávok, grízes barna) aabb → barna (lovaknál
„gesztenye”)

Mendel III. Függetlenség
Ha egyszerre több tulajdonságban különböző tiszta vonalakat keresztezünk, akkor az F2 generációban a szülői típusoktól eltérő tulajdonság-kombinációkat is találunk, ezek a rekombináns utódok.

Mendel III. FüggetlenségP: AAbb (fahéj) x aaBB (fekete)
F1: AaBb (agouti)
F2: 9/16 A_B_ (agouti), 3/16 A_bb (fahéj),
1/16 aabb (barna) , 3/16 aaB_ (fekete)↑ ↑
rekombináns típusok szülői típusok
Ugyanezt kapnánk, ha agoutit kereszteznénk barnával:
AABB x aabb.

Mendel III. Függetlenség
Ez akkor igaz, ha a két lókusz nem kapcsolt, tehát különböző kromoszómákon helyezkednek el, vagy egymástól távol.
Ha a lókuszok szorosan kapcsoltak, akkor a rekombináns utódok aránya nagyon kicsi.

II. Eltérések a mendeli arányoktól
A) Különböző életképességű genotípusok
B) Recesszív episztázisC) Kapcsoltság

A) Letális allélek
Ritkán előfordulnak világosabb, ”sárga” színű egerek. Két sárga egér keresztezéséből származó utódok első ránézésre nem-mendeli arányokat mutatnak.
P: sárga x sárgaF1: 2/3 sárga, 1/3 normál
agouti

Letális allélek
Magyarázat: Az A lókuszon egy harmadik allél is megjelenik ritkán, az AY.
A heterozigóta AYA egerek sárgák, a homozigóta AYAY viszont életképtelen.
P: AYA x AYA
F1 zigóták: ¼ AA, ½ AYA, ¼ AYAY †
sávos sárga letális

Letális allélek: Manx macska
Hasonló: MLM farok nélküli, életképes, MLML életképtelen

Különböző életképességű genotípusok
Az előbbi példa egy szélsőséges esete annak az általános jelenségnek, hogy a különböző genotípusok túlélési esélye eltérő. Így zigótakorban a mendeli arányoknak megfelelően (1:2:1) oszlanak meg a genotípusok, de mire a kísérletező megszámolja a különböző utódokat, már megváltozhatnak az arányok.
Általában ilyen kísérleteknél olyan jó környezetet biztosítanak, hogy minden genotípus túlélése a lehető legjobb legyen, így csökkenthető a mendeli arányok torzulása.

B) Recesszív episztázis
A C lókuszon homozigóta recesszív cc genotípusú egyedek albínó fenotípust mutatnak, bármely allélek is vannak az A ill. B lókuszon.
A C gén feljebb helyezkedik el a genetikai-biokémiai láncban:
C_ B_x → y → fekete színanyag
C_ bbx → y → barna színanyag
cc x →│ nincs színanyag

Albinizmus, cc genotípus

Albinizmus, cc genotípus

Recesszív episztázis: F2 9:3:4
P: BBcc (albínó) x bbCC (barna)
F1: BbCc (fekete)
F2: 9/16 B_C_, 3/16 bbC_, 4/16 __cc
fekete barna albínó

Recesszív episztázis: az aranyszínű Labrador kutya
fekete barna aranyszínű

Recesszív episztázis: az aranyszínű Labrador kutya
Az E gén hasonló F2 arányokat okoz, mert az ee genotípusban gátolt a pigmentnek a szőrszálban való lerakódása → fekete vagy barna szín helyett aranyszín.
P: BBee (arany) x bbEE (barna)
F1: BbEe (fekete)
F2: 9/16 B_E_, 3/16 bbE_, 4/16 __eefekete barna arany

ch: egy hőmérsékletérzékeny allél

ch: egy hőmérsékletérzékeny allél
A test melegebb részein a ch a c-hez hasonlóan albínó szőrszálakat eredményez, de a kiálló, hidegebb testrészeken működik a festékszintézisért felelős enzim, ezért azok sötétek: sziámi macskák, Himalája nyulak és egerek stb.

Tanulságok
A gének kifejeződését befolyásolja a genetikai környezet, pl. gátolhatják más gének (episztázis).
A gének kifejeződését befolyásolja a fizikai környezet (hőmérséklet).

C) Kapcsoltság
1910: Morgan 2 lókuszt vizsgált muslicán: + : vad típus, dom. (normál szárny, piros
szem)
Mutánsok:pr → bíbor szem vg → csökevényes
szárny

Kapcsoltság
P: pr+pr+ vg+vg+ x prpr vgvghomozigóta vad típusú homozigóta mutáns
F1 : pr+pr vg+vg
heterozigóta vad típusú

A testcross
Az F1 egyedeit testcross-nak vetette alá, vagyis a homozigóta recesszív genotípussal keresztezte őket:
F1: pr+pr vg+vg x prpr vgvg
tester

A testcross
A testcross célja az F1 génjeinek pontos megismerése.
Mivel a „tester” szülőtől csak recesszív alléleket kap az F2 utód, ezért a fenotípusa egyértelműen megmutatja a genotípusát, illetve hogy milyen alléleket kapott az F1 szülőtől származó ivarsejttel.
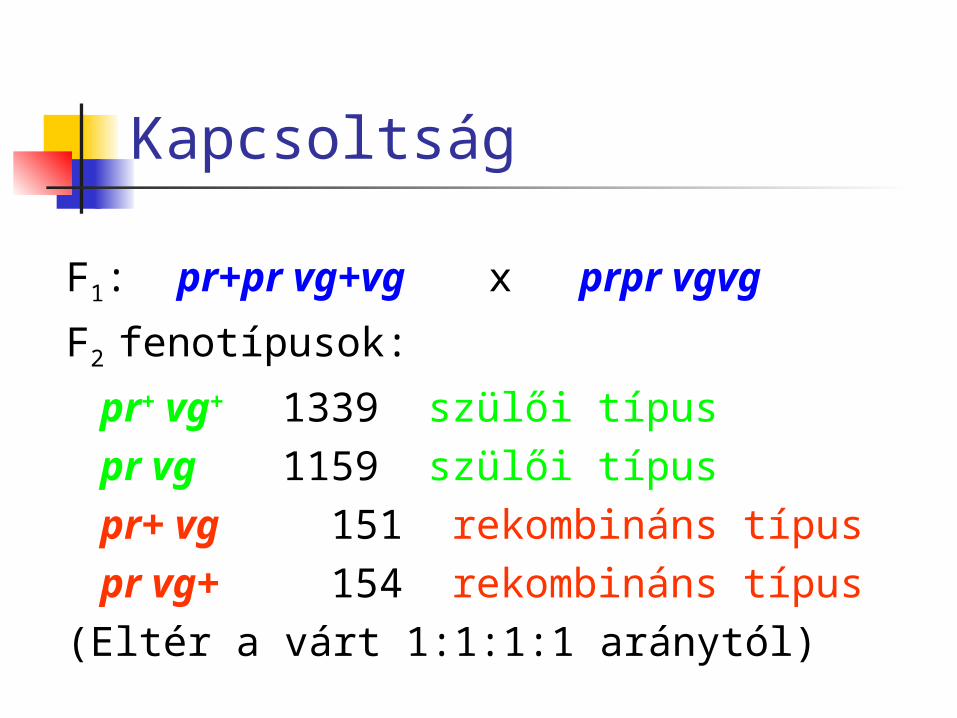
Kapcsoltság
F1: pr+pr vg+vg x prpr vgvg
F2 fenotípusok:
pr+ vg+ 1339 szülői típuspr vg 1159 szülői típuspr+ vg 151 rekombináns típuspr vg+ 154 rekombináns típus
(Eltér a várt 1:1:1:1 aránytól)

Kapcsoltság
A szülői típusok a várt ¼-es aránynál sokkal nagyobb arányban vannak jelen.
Morgan következtetése: a két lókusz egy kromoszómán helyezkedik el, egymáshoz közel.
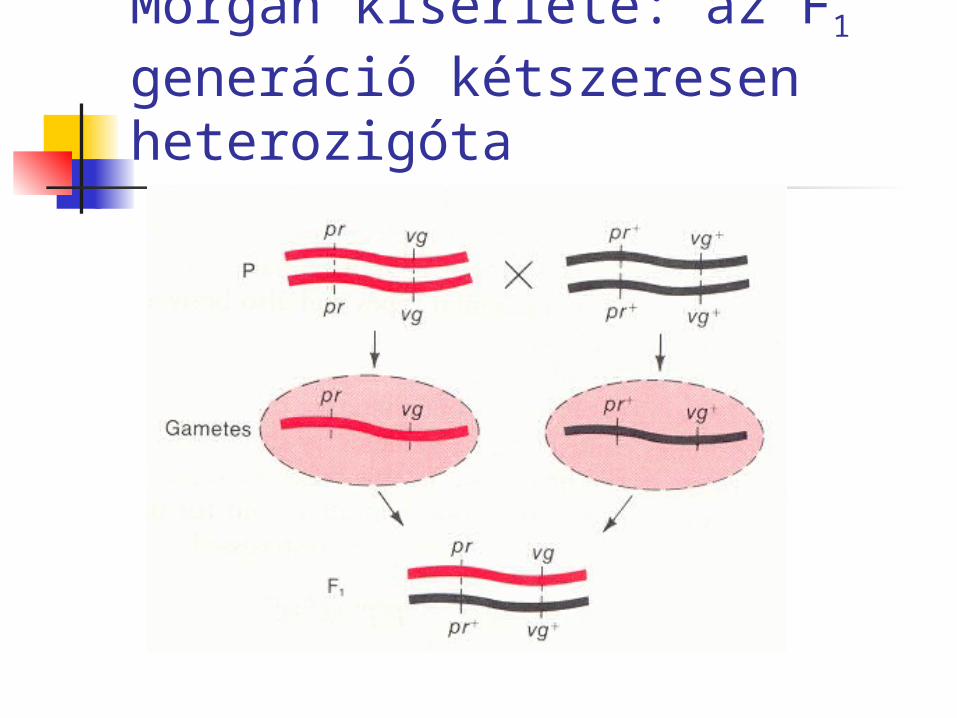
Morgan kísérlete: az F1 generáció kétszeresen heterozigóta
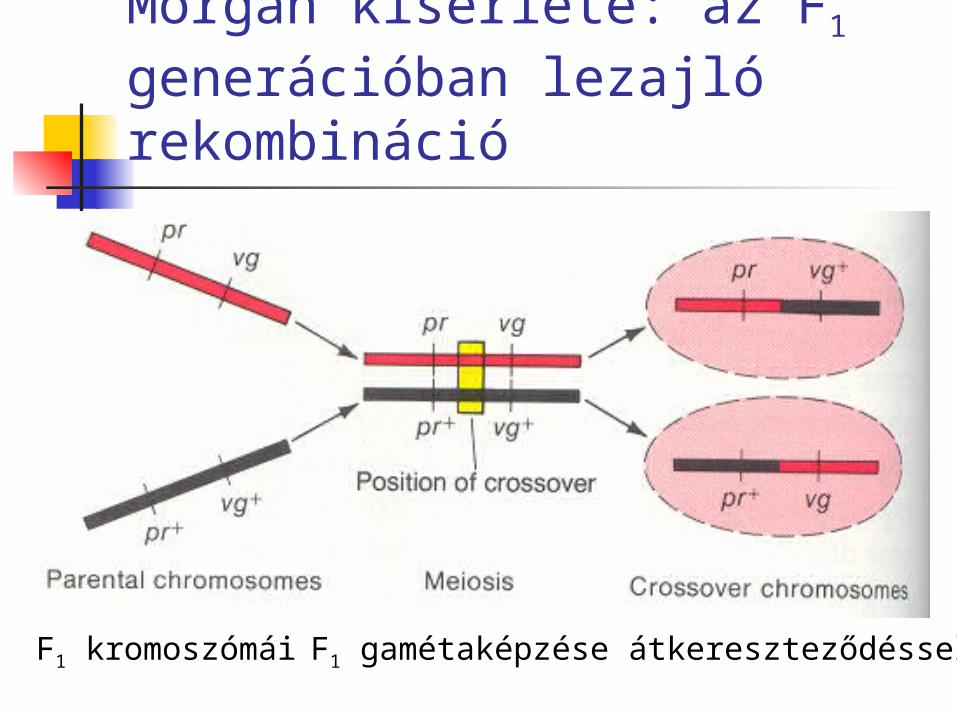
Morgan kísérlete: az F1 generációban lezajló rekombináció
F1 kromoszómái F1 gamétaképzése átkereszteződéssel
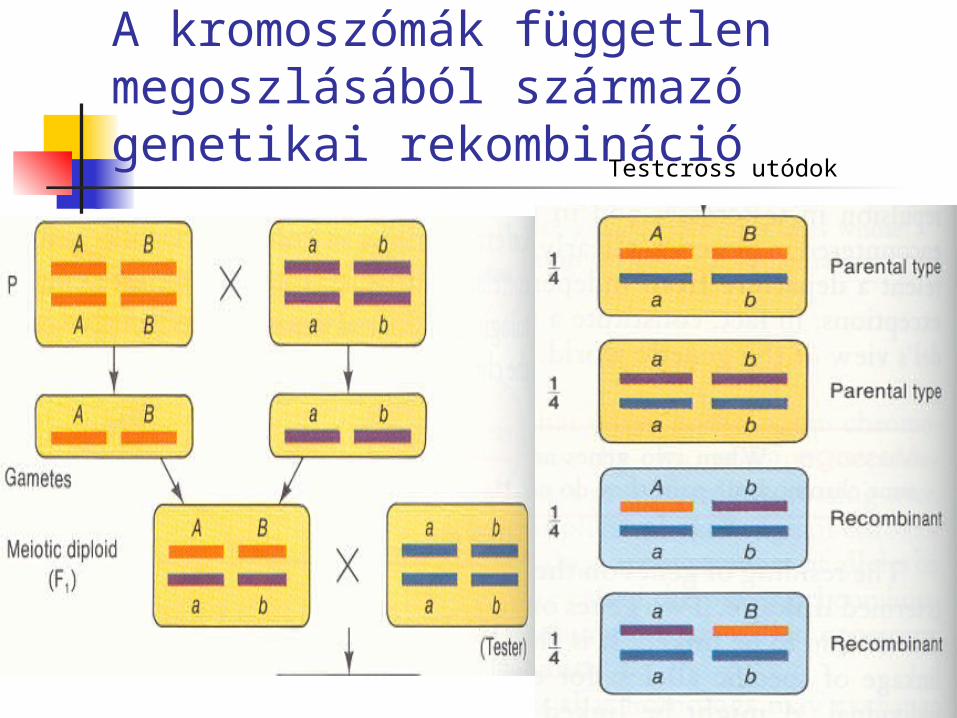
A kromoszómák független megoszlásából származó genetikai rekombináció
Testcross utódok
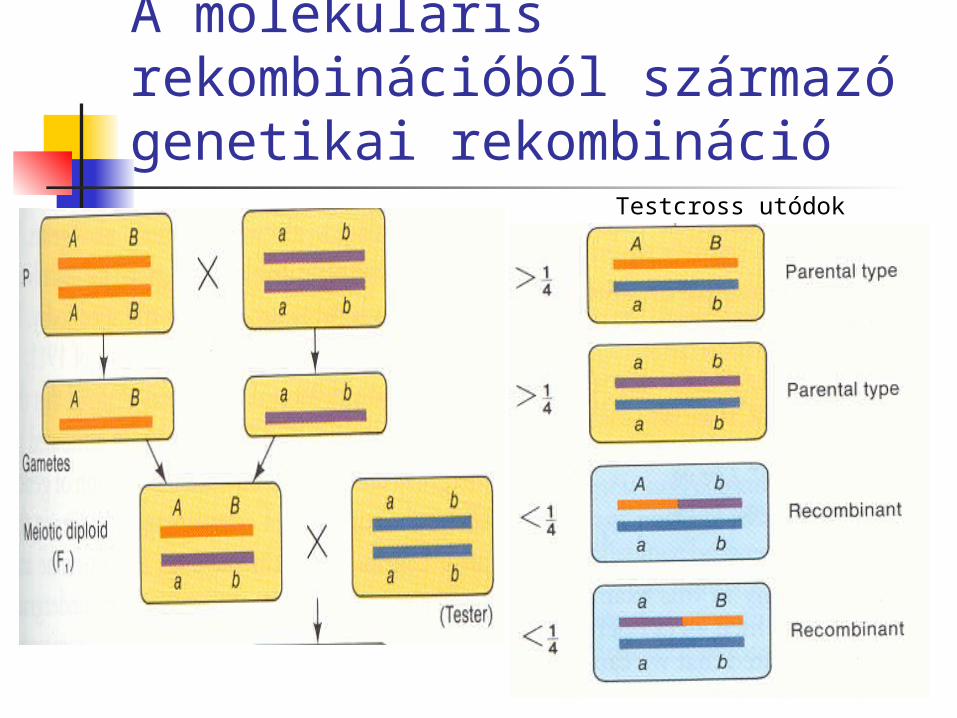
A molekuláris rekombinációból származó genetikai rekombináció
Testcross utódok

III. A szex evolúciója

Mi a közös?

Az ivaros folyamat (szex) nem feltétele a szaporodásnak
Valójában annak ellentettje! Partenogenezis = szűznemzés
Cnemidophorus uniparens (ostoros farkú gyík)
Pitypang, szeder, palástfű araszoló kerekesférgek (Cl. Rotifera,
O. Bdelloidea)

A szex kétszeres ára Ha a szűznemző és az ivaros változat
ugyan annyi utódot képes létrehozni, akkor a szűznemzők száma minden generációban meg fog kétszereződni.
Csak akkor igaz, ha a hímek nem járulnak hozzá az utódneveléshez. nincs utódgondozás anizogámia
A szex kialakulására nincs hatással, mert az első ivaros eukarióták biztos izogámok voltak.

Az ivarosság egyéb költségei
Összeolvadáshoz szükséges partner keresése
Gaméták előállítása meiózissal

Az ivarosság előnyei a populációk számára gyorsabb evolúció és alkalmazkodás a
környezethez csökkentheti a káros mutációk gyakoriságát:
„motor-váltó” elmélet, avagy G. B. Shaw és a színésznő véges pop.: Muller kilincskereke (az ivartalan
pop.kban káros mutációk halmozódhatnak fel) végtelen pop.: ha a káros mutációk
szinergisztikusan hatnak Ezek hosszú távú előnyök, melyek az ivarosság
fenntartásában játszhatnak szerepet.
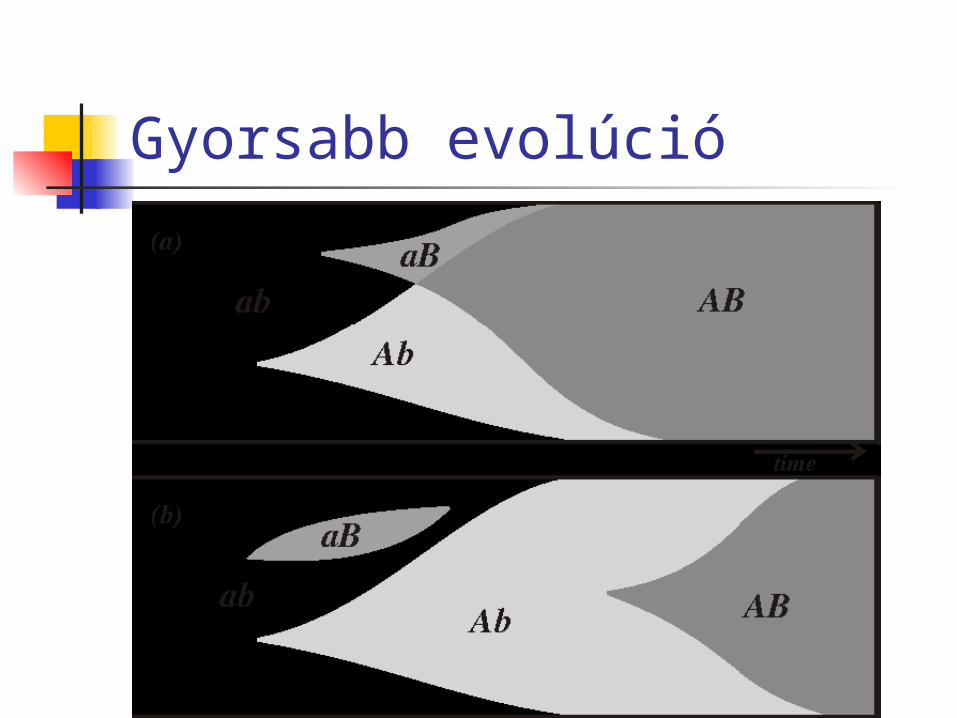
Gyorsabb evolúció

Az ivarosság előnyei az egyedek számára
„tombola modell” erős és kiszámíthatatlan szelekció esetén
kétszálú DNS javítás feltétele a diploídia (bár a diploídiának nem feltétele az ivarosság)
az előbb említett előnyök itt is érvényesek a populáción belül

Az ivarosság előnyei az önző gének számára
transzpozonok (ugráló genetikai elemek) terjedése
baktérium plazmidok elérik a baktériumsejtek összekapcsolódását és ilyenkor átjutnak egyik sejtből a másikba

Párosodási típusok kialakulása
hibrid vigor organellumok egyszülős
öröklődése anizogámia kialakulása másodlagos nemi jellegek

IV. A nem meghatározás módjai
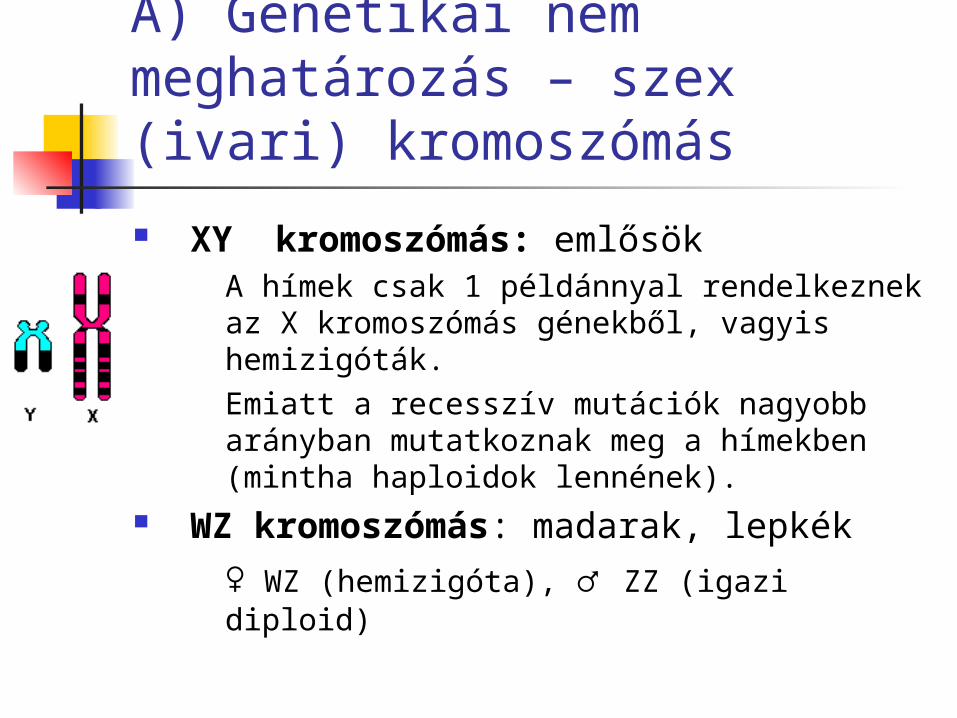
A) Genetikai nem meghatározás – szex (ivari) kromoszómás
XY kromoszómás: emlősök A hímek csak 1 példánnyal rendelkeznek az X kromoszómás génekből, vagyis hemizigóták. Emiatt a recesszív mutációk nagyobb arányban mutatkoznak meg a hímekben (mintha haploidok lennének).
WZ kromoszómás: madarak, lepkék
♀ WZ (hemizigóta), ♂ ZZ (igazi diploid)

B) Genetikai nem meghatározás – autoszómás
Autoszóma: nem ivari kromoszóma (emberben 22 pár van)
Haplodiploid rendszerek: hártyásszárnyúak, sáskák, levéltetvek, stb.
♀ (2n), ♂ (n) megtermékenyítetlen petéből X:A arány: muslica, fonálféreg
♀ : 2:2
♂ : 1:2; kétszer annyi autoszóma készlet mint X
Autoszómás domináns: házilégy

C) Környezeti nem meghatározás
Gammarus duebeninél (rák) a napszakok hossza alapján a szezon elején főleg hím, később inkább nőstény
Trachemys scripta teknősnél a hőmérséklet szerint
Alligátorok: tojás inkubálási hőmérséklete alapján

D) Egyéb nem meghatározási módok
Életkor alapján (nem váltás) pl. egyes csigák fiatalon hímek,
idősebben nőstények halaknál fordítottra is van példa
Fertőzés által egyes rovarokban: Wolbachia bakt. ZZ hímeket nősténnyé alakítja

E) Nincs nem meghatározás
Hermafroditák: egyszerre ♀ és ♂ pl. egyes csigák, földigiliszták
Partenogenetikus fajok: nincsenek hímek, csak nőstények pl. egyes gyíkok, halak, rovarok

A nem meghatározás evolúciója: miért alakul ki gyakran az XY rendszer?
Az XY rendszer lényege, hogy az Y kromoszóma génekben elszegényedik, emberben csupán kb. 20 gént tartalmaz, főleg a hím nemi funkciókért, pl. spermatogenezisért felelős géneket.

A nem meghatározás evolúciója: miért alakul ki gyakran az XY rendszer?
A kiindulás egy autoszómás hím-meghatározó allél, M; így Mm → hím, mm → nőstény.
Ha ennek a közelében, kapcsoltan megjelenik egy olyan F allél, amely csak hímekben előnyös, viszont nőstényekben káros, az elterjed a populációban.

A nem meghatározás evolúciója: miért alakul ki gyakran az XY rendszer?
Az ilyen csak hímekben előnyös allélek felgyűlnek az M allél közelében és a kromoszóma egyre nagyobb részén gátlódik a rekombináció (így ezek az allélek nem kerülhetnek nőstényekbe).
A rekombináció nélküli kromoszómát viszont sújtják az ivartalan szaporodás hátrányai, felgyűlnek a káros mutációk.

A nem meghatározás evolúciója: miért alakul ki gyakran az XY rendszer?
Az Y kromoszóma végül elveszti azokat a géneket, amelyek az X kromoszómán is megtalálhatóak, hiszen az X kromoszóma a nőstényekben rekombinálódik, nem gyűlhetnek fel rajta a káros mutációk.
Az emberi Y kromoszómán csak egy kis rész maradt néhány génnel, amely még rekombinálódik az X-szel, a többi rész néhány hím funkcióért felelős génen kívül „üres”, vagyis a mutáció által erodált funkcióképtelen géneket tartalmaz.

V. Az XY rendszer következményei
A nemhez kötött öröklődésDóziskompenzáció
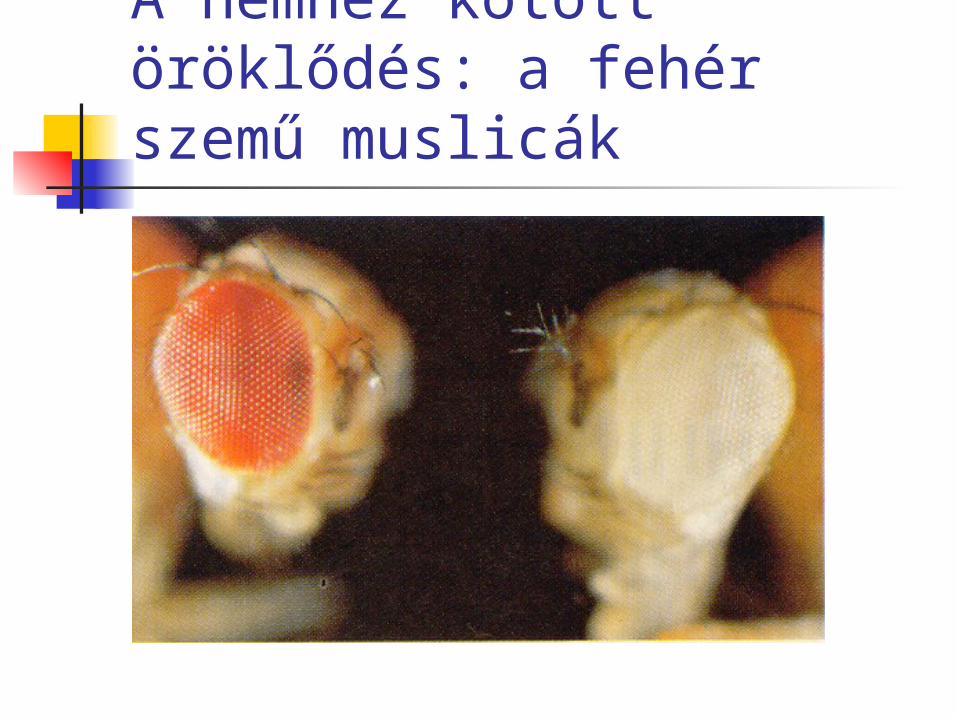
A nemhez kötött öröklődés: a fehér szemű muslicák

A nemhez kötött öröklődés felfedezése: a fehér szemű muslicák (Morgan, 1910 körül)
piros szemű ♀ X fehér szemű ♂: Első pillantásra mendeli 3:1 arányokat találunk az F2 generációban, de furcsa, hogy csak a hímek fehér szeműek!
Az ún. reciprok keresztezés méginkább meghökkentő eredményt ad: mind az F1-ben, mind az F2 -ben 1:1 arányt találunk!
Csak úgy magyarázható, ha feltételezük, hogy a fehér szemszínt okozó allél az X kromoszómán helyezkedik el.

piros szemű ♀ X fehér szemű
♂:
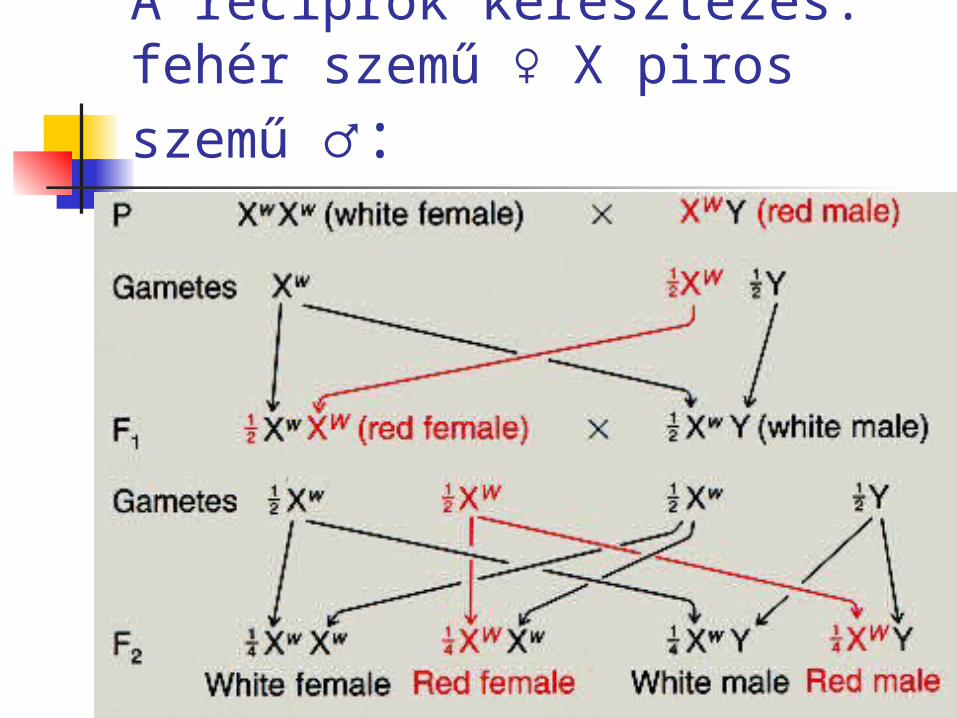
A reciprok keresztezés:fehér szemű ♀ X piros szemű
♂:

A nemhez (X-hez) kötött öröklődés az emberben A pedigrében a következő jelek
utalnak recesszív nemhez kötött öröklődésre: több férfi érintett (beteg) mint nő az érintett férfi fiai egészségesek, de
lányai hordozók, így a lányai által szült fiú-unokák fele érintett
az érintett férfi fiainak utódai mind egészségesek, mert a fiúk nem is hordozzák a beteg allélt.

A nemhez (X-hez) kötött öröklődés az emberben
Példák (mind recesszív betegségek): vérzékenység vörös-zöld színtévesztés tesztikuláris feminizáció:
érzéketlenség a férfi nemi hormonokra, XY kromoszómákkal nőies (de steril) fenotípus alakul ki.

A dóziskompenzáció lehetőségei:
emlősök: a nőstényekben random X kromoszóma-inaktiváció történik (heterokromatinizálódik→Barr test)
muslica: a hímek X kromoszómájáról gyorsabb az átírás (up-regulation, hipertranszkripció)
fonalféreg: a nőstények X kromoszómájáról lassabb az átírás
Az X kromoszómán lévő "nem nemi" tulajdonságok kialakulásáért felelős gének esetében zavarokat okozhatna, ha a nőben mindíg két, a férfiban csak egy gén működne.

Az X kromoszóma-inaktiváció
Az emlős nőstények egyedfejlődésének korai szakaszában véletlenszerűen inaktiválódik minden sejtben az egyik X.
Emiatt az emlős nőstények mind genetikai mozaikok, de mivel ez a mozaikosság elég finom-szemcséjű, így „nem látszik”.
Pl. heterozigóta (hordozó) nőstényekben vannak működöképes sejtek a test egészét ill. a funkció egészét nézve, pl. vannak véralvadási faktort termelő sejtek, piros és zöld csapok a retinában.

Az X kromoszóma-inaktiváció
Egyes nemhez-kötött betegségek vagy jellegek azonban a kültakarón mozaikosan megfigyelhető rendellenességekben, foltokban mutatkoznak meg (durva-szemcsés).

Az X kromoszóma-inaktiváció
Ilyen a verejtékmirigyek hiánya. Az X kromoszómás A gén szükséges a verejtékmirigyek megjelenéséhez.
XAXA: egészséges nő XAY: egészséges férfi XaY: beteg férfi, nincs
verejtékmirigye XAXa: mozaikos nő, foltokban nincs
verejtékmirigye
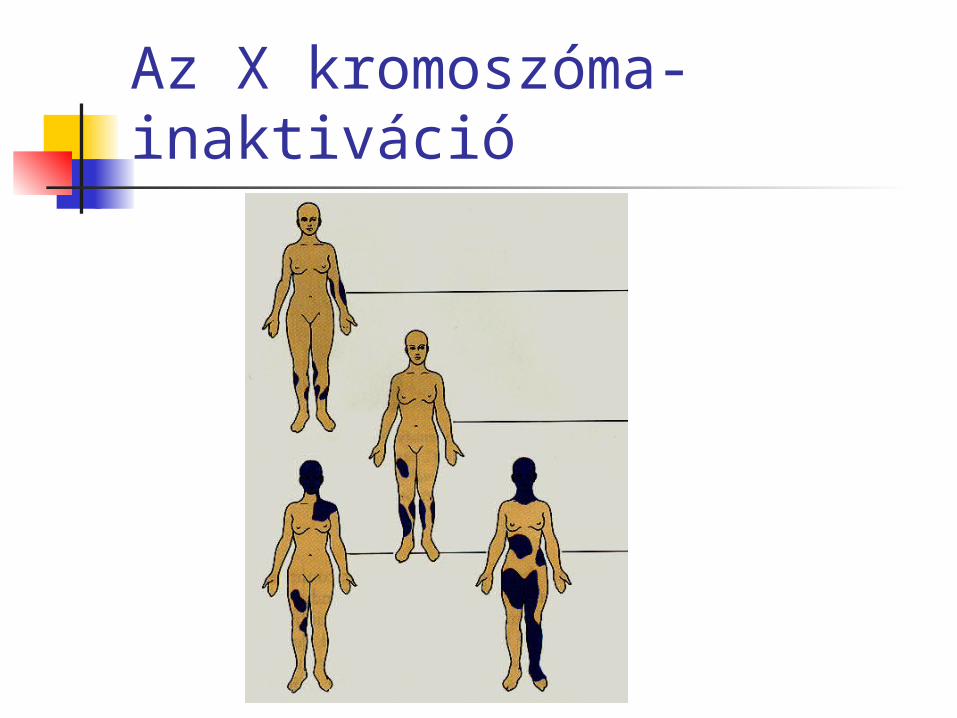
Az X kromoszóma-inaktiváció

Az X kromoszóma-inaktiváció
Az un. calico macskák heterozigóta nőstények, Oo genotípussal. Azokban a foltokban ahol az O allélt tartalmazó X kromoszóma marad aktív narancssárga lesz, ahol az o allél aktív ott fekete. (A fehér foltokat egy másik gén okozza, ez nem nemhez kötött.)

VI. Kvantitatív genetika

Mennyiségi jellegek genetikája(kvantitatív genetika)
Kvantitatív (mennyiségi) jellegek: azok a tulajdonságok, amelyek valamilyen mérőszámmal jellemezhetők.
Pl.: testmagasság, testsúly, tejtermelés, utódok száma, magvak olajtartalma, IQ stb.
Általában normális eloszlást követnek a populációban.

Mennyiségi jellegek genetikája(kvantitatív genetika)
Sok (10-100), egyenként kis hatású mendelező gén befolyásolja a fenotípust.
Egy-egy allél befolyása általában a környezet hatásával összemérhetően kicsi, ezért a jelleg nem „digitálisan”, hanem folytonosan változik, nehéz meghatározni a genotípust a fenotípusból.

http://falco.elte.hu/kvanti/
A pop. fenotípusos eloszlása
fenotípus
gyako
risá
g

Mennyiségi jellegek genetikája(kvantitatív genetika) Minél nagyobb a genetikai meghatározottság,
annál nagyobb az öröklékenység (heritabilitás). A nagy heritabilitás azt jelenti, hogy a populáció egyedei főleg azért térnek el egymástól, mert különbözik a genotípusuk és csak kisebb arányban okozza a különbségeket a környezet.
Az öröklékenység mindig csak egy adott populációra jellemző, más populációban más (és tulajdonságonként is változik).
Minél nagyobb a heritabilitása egy tulajdonságnak annál nagyobb előrehaladást lehet elérni a mesterséges szelekcióban (pl. tenyésztésben).

VII. Evolúciógenetika

A mutáció A mutáció a DNS valamilyen megváltozása. Általában a mutáció során káros allélok
keletkeznek. Ritkán előnyös mutációk bukkannak fel: az
evolúció nyersanyagát jelentő változatosság forrása.
A eukariótákban a mutációk többsége (majdnem) semleges: nem-kódoló szakszokat, intronokat érint a kodonok 3. nukleotidjának megváltozása
gyakran nem okoz változást a kódolt fehérjében

A mutáció típusai Pont-mutációk:
egy bázispár megváltozása egy bázispár beékelődése vagy elvesztése →
„frame-shift” (leolvasási keret) mutációk Nagyobb DNS szakaszokat érintő
változások: beékelődés (inszerció) elvesztése (deléció) megfordulása (inverzió) megkettőződése (duplikáció) → fontos az
evolúcióban, nyersanyag, gén-családok

A mutációk gyakorisága
Báziscsere valószínűsége: 10-9 / nukleotid / DNS másolás.
RNS vírusoknál ez sokkal nagyobb: 10-4 / nukleotid / DNS másolás → HIV változékonysága
Lókuszonkénti mutációs ráta: 10 -5/ lókusz / generáció
Az evolúció feltételei Szaporodás, öröklődés, változékonyság

Az genetikai változatosságot befolyásoló tényezők
A genetikai változatosságot növelik: mutáció rekombináció (allélgyakoriságot nem bef.!) migráció
A genetikai változatosságot csökkentik: szelekció sodródás (véletlen allél-gyakoriság-
változások)
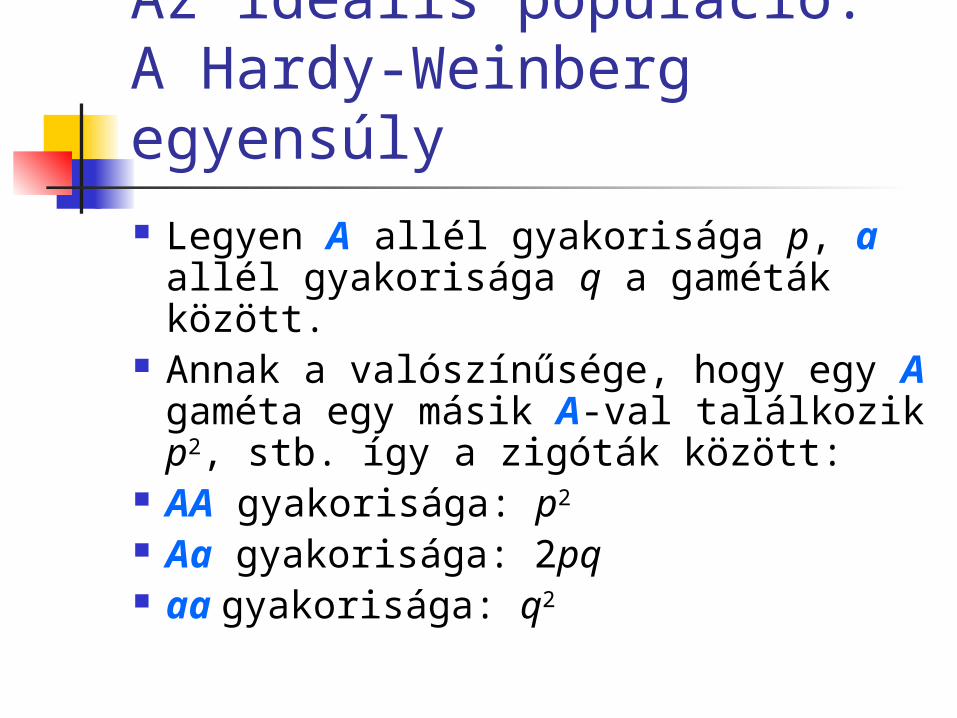
Az ideális populáció: A Hardy-Weinberg egyensúly Legyen A allél gyakorisága p, a allél
gyakorisága q a gaméták között. Annak a valószínűsége, hogy egy A
gaméta egy másik A-val találkozik p2, stb. így a zigóták között:
AA gyakorisága: p2
Aa gyakorisága: 2pq aa gyakorisága: q2

Mikor nem áll fenn a A Hardy-Weinberg egyensúly?
Ha nem ideális a populáció: Ha a mutáció megváltoztatta az arányokat Ha a migráció megváltoztatta az arányokat Ha a szelekció megváltoztatta az
arányokat Ha a populáció kicsi → sodródás Ha a párosodás nem véletlenszerű
(beltenyésztés) Ha a szaporodás ivartalan

A rekombináció szerepe az evolúcióban
A genetikai változatosság növelése, prokariótákban ritkább a rekombináció, ivaros eukariótákban gyakori.
A rekombináció a 2 vagy több lókuszos genotípusokat tekintve növeli a változatosságot, ha csak egy lókuszt vizsgálunk, a rekombináció nem játszik szerepet (ld. H-W egyensúly).

A sodródás szerepe az evolúcióban
A genetikai sodródás azt jelenti, hogy az allél-gyakoriságok változását a véletlen is befolyásolja a szelekció mellett.
Kis populációkban a véletlen nagyobb szerepet kap a szelekcióhoz képest.

A sodródás szerepe az evolúcióban Átlagosan igaz, hogy a legrátermettebbnek
van a legtöbb utódja, de az utódok száma ingadozik, kis populációkban előfordul, hogy a legrátermettebbeknek nem lesz egy utódja sem.
Nagy populációban többen vannak jelen a legrátermettebb genotípussal, kicsi az esélye, hogy mindegyik utód nélkül pusztul el valamilyen szerencsétlen véletlen miatt.






























