Embed Size (px)
Citation preview

XIII
LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE
Gli Americani sono soliti spiegare gran parte delle azioni della loro vita con il principio dell’interesse personale, compreso nel modo giusto;…..A riguardo penso che essi continuamente si contraddicano; negli Stati Uniti così come altrove, le persone dimostrano impulsi disinteressati e spontanei che sono naturali al genere umano; ma gli Americani raramente ammettono di provare emozioni di tal tipo.
Alexis de Tocqueville, Democracy in America (1830)
Sia che la lotta tra gruppi prenda la forma di una guerra o quella di una più intensa competizione per l’offerta di commercio e di cibo, il gruppo in cui la competizione interna non controllata ha prodotto un ampio proletariato senza alcun interesse concreto nello Stato, sarà il primo a collassare.
Karl Pearson, Socialism and Natural Selection (1894)
Quando quattro giovani facinorosi, in una pizzeria aperta durante la notte nella
città italiana di Rimini, iniziarono a gettare cibo e ad insultare il fornaio, un uomo
senegalese di nome Sarr Gaye Diouf intervenne per difenderlo (Meletti 2001). Uno dei
teppisti afferrò Diouf per le braccia e gli altri tre lo pugnalarono una quindicina di volte
con i coltelli per la pizza. Diouf morì immediatamente e i colpevoli vennero arrestati.
Diouf lavorava temporaneamente come uomo delle consegne, sperando di diventare
presto un tassista. Egli non conosceva il fornaio se non come cliente occasionale e i suoi
aggressori, in visita a Rimini da Napoli, non avevano mai visto Diouf prima. Eppure
Diouf diede la sua vita per difendere il fornaio e i giovani malviventi uccisero lo
straniero – Diouf – per nulla scoraggiati dalla certezza dell'arresto.
Una tale tragedia evoca orrore, ma non sorpresa. Di continuo si assiste a persone
sacrificarsi per degli sconosciuti, né é raro vedere persone ucciderne altre, anche per
piccole provocazioni, specialmente quando l'obiettivo é un “outsider”. Questi due aspetti

2 | MICROECONOMIA
del comportamento umano sono pensati, genericamente, come antitetici, ma come
vedremo, possono avere un'origine comune: la competizione tra gruppi può avere
favorito quelle nazioni, tribù, bande ed altri gruppi che hanno promosso preferenze
altruistiche nei confronti di alcuni stranieri e ostilità nei confronti di altri. Un altro
esempio empirico – la coevoluzione di cittadinanza moderna e guerra – può suggerire
alcuni dei processi sottostanti.
Figura 13.1. Citta’ autonome nel Sud della Germania (Fonte: T.A. Brady (1985),p. xvi)
Otto secoli fa, l'area intorno a Rimini, dove Diouf è stato ucciso, era governata
da piú di una dozzina di corpi sovrani. Nell'attuale Italia vi erano dalle due alle trecento
città-stato diverse. Nella Germania del Sud, mezzo millennio fa, vi erano sessantanove
città libere, oltre a numerosi episcopati, principati, ducati ed altre entità simili allo Stato
(Brady 1985). Le figure 13.1 e 13.2 illustrano questa proliferazione di sovranità nel
quindicesimo secolo. L'intera Europa, a quel tempo, era governata all'incirca da
cinquecento corpi sovrani. Ma con la Prima Guerra Mondiale, rimasero poco più di una
trentina di Stati. Questa riduzione degli Stati non solo abbassò il numero dei corpi
sovrani, ma ridusse anche l'eterogeneità delle forme di governo. Emerse una singolare
forma politica – lo Stato nazionale – dove una volta regnavano, secondo Charles Tilly
(1990:5), “imperi, città-stato, federazioni di città, gruppi di proprietari, ordini religiosi,
leghe di pirati, bande guerriere e molte altre forme di governo”. A differenza delle varie
forme di governo concorrenti che esso ha cancellato, lo Stato nazione esibiva una
struttura burocratica centralizzata e manteneva l'ordine su un territorio definito, con la
capacità di incrementare sostanzialmente il reddito attraverso la tassazione e di spiegare

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |3
forze armate permanenti1.
Figura 13.2. Entità politiche nel quindicesimo secolo in Italia. La grande maggioranza delle entità sovrane più piccole (come San Gimignano) non sono mostrate; da notare le molte entità, una volta autonome (come Verona, Bergamo, Padova, Vicenza) tutte assorbite da Venezia agli inizi del secolo. Fonte: Matthew (1992:212).
Cosa spiega il successo di questa nuova forma di organizzazione della società?
Una semplice risposta é che quando gli Stati-nazione erano in guerra con le altre forme
di governo, essi tendevano a vincere. Ma Tilly scrive: “Nessun monarca poteva condurre
una guerra senza assicurasi prima l'acquiescenza di tutta la popolazione e la cooperazione
attiva di alcuni notabili” (Tilly 1990:75). Un sistema di tassazione pagato in moneta,
insieme con la capacità di prendere a prestito somme elevate di denaro, permise agli
Stati-nazione di condurre guerre senza dover ricorrere a misure impopolari, come la
confisca diretta di cibo, armi e animali. Inoltre, l'introduzione di diritti di proprietà ben
definiti e dei mercati facilitava questa tassazione – e l'approccio, basato sul debito, alla
mobilizzazione delle risorse necessarie a vincere la guerra. L'istituzione dei mercati
1 In aggiunta a Tilly (1990), sono state utilizzate anche le seguenti referenze: Gellner (1983), Bright e
Harding (1984), Tilly (1975), Mack Smith (1959), Anderson (1974). Wallerstein (1974), e Bowles e Gintis (1984).

4 | MICROECONOMIA
favoriva, invece, la formazione dello Stato in un modo meno ovvio, introducendo
l'osservanza alla legge. Tilly continua nel suo commento:
Coloro che partecipavano ai mercati contribuivano significativamente alla necessaria sorveglianza registrando prezzi e cessioni. I cittadini propriamente civilizzati, inoltre, finirono per attribuire valore morale al pagamento delle tasse; essi si controllavano a vicenda, accusando gli evasori delle tasse di free-riding.
Gli stati europei esibivano una particolare struttura spaziale con entità politiche
grandi ma difficilmente controllabili nelle zone periferiche (per esempio, la Russia e
l'Impero Ottomano), con raggruppamenti di cittá-stato e di federazioni (le cittá-stato
italiane ed i cantoni svizzeri), e Stati più centralizzati, alla fine vittoriosi, come la Francia
e il Brandeburgo, una via di mezzo tra i due. Gli stati-nazione di successo assimilavano i
popoli che assoggettavano e con il tempo promuovevano e, a volte, imponevano un
processo di integrazione delle nuove generazioni attraverso l'istruzione scolastica2.
A causa del loro successo, forme politiche simili agli stati nazionali furono
esportate si svilupparono in tutto il mondo, soppiantando forme di organizzazione
differenti. Con lo stato-nazione e con l’emergere dell'economia capitalistica, la
popolazione Europea crebbe notevolmente – si moltiplicò di quindici volte in Gran
Bretagna nei quattro secoli successivi al 1500 - superando largamente i tassi di crescita
della popolazione in tutto il mondo (eccetto, forse, per il diciottesimo secolo in Cina).
Come risultato, la diffusione globale dello stato-nazione venne promossa non solo dalla
competizione tra stati della periferia europea, ma anche dalla massiccia emigrazione di
persone che non solo esportarono i tratti culturali europei, ma anche le capacità militari
che favorirono la costruzione dello Stato in Europa.
In breve, lo Stato nazionale si sviluppò perché sconfisse con le guerre differenti
forme di organizzazione politica. L'abilità di vincere le guerre dipendeva dalla capacità di
mobilizzare soldati e le altre risorse militari. Tale capacità dipendeva dall’ampiezza degli
scambi commerciali, dalla disponibilità del credito, dall'osservanza della normativa fiscale
e dalla volontà di servire il sovrano in guerra. Queste caratteristiche erano rafforzate dalla
diffusione delle norme riguardanti i comportamenti individuali, che, anche se all'inizio
non vantaggiose per i singoli individui, contribuivano al successo del gruppo. Tra le
ragioni vi erano la volontaria osservanza della normativa fiscale, la volontà di incorrere in
2 Weber (1976) descrive l'assimilazione di popolazioni distinte dallo Stato della Francia. Gellner (1983)
sviluppa la connessione tra la nascita del commercio, lo Stato nazionale e il sorgere di ciò che ha chiamato “exo-education”, ovvero, la socializzazione dell'infanzia portata avanti da specialisti che non siano membri della famiglia o parenti stretti del bambino.

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |5
rischi durante una guerra per il sovrano o per la nazione, ed il rispetto dei diritti di
proprietà. La norma della monogamia può aver giocato un ruolo simile anche se meno
ovvio nel salvaguardare la cooperazione popolare alla realizzazione dei progetti della
elite3. Ognuna di queste norme contribuisce, direttamente o indirettamente, alla capacità
di uno Stato di fare guerra ma richiede a coloro che rispettano le norme di rinunciare a
possibili guadagni e di tollerare eventuali perdite (incluso il minore successo
riproduttivo).
Ovviamente, gli Stati nazionali crearono anche un ambiente legale e culturale in
cui coloro che aderivano alle norme avrebbero potuto migliorare le loro capacità di fare
guerra soffrendo una perdita piccola o nulla rispetto coloro che non aderivano a queste
norme. Ma l'emergenza e la diffusione rapida degli Stati nazionali si é tipicamente basata
sulle norme vantaggiose per il gruppo e costose per il singolo individuo.
Altri casi empirici e ben documentati di conflitti ed assimilazioni tra gruppi sono
quelli della conquista della Dinka ad opera di Nuer (Kelly 1985) ed il processo
dell'evoluzione culturale nella Nuova Guinea (Soltis, Boyd, e Richerson 1995). Un altro
esempio é dato dall’ampia diffusione dell'Islam nel secolo seguente la morte di
Maometto – nel 750 esso si estendeva su un'ampia zona che andava dal fiume Indo ad
est sino al fiume Duro, in Spagna, ad ovest. Ciò fu possibile (secondo Levy 1957:3)
perchè la fede in Allah forniva “un legame ben più forte, anche se più sottile di quello di
una semplice somiglianza culturale” e facilitava un sistema inclusivo di tasse,
arruolamento militare ed alleanza”4. Quindi, il processo del conflitto di gruppo, seguito
dall'assimilazione culturale o dall'estinzione fisica, sembra essere un fenomeno molto
generale.
In questo capitolo, esploreremo il ruolo della competizione tra gruppi
nell'evoluzione delle norme altruistiche, e analizzeremo perché alcuni individui sono
disposti a rischiare la propria vita per difendere uno sconosciuto assalito da giovani
criminali, o ad andare in guerra per la gloria della nazione.
3 Herlihy e Klapische-Zuber (1985-157) scrive: “La grande conquista sociale del primo Medioevo fu
l'imposizione delle stesse regole di condotta sessuale e domestica sia per i ricchi che per i poveri.” Si veda anche MacDonald (1995). Mentre riduceva i vantaggi delle persone di potere e di successo, la norma della monogamia (così come la successiva estensione del suffragio ai lavoratori) può essere stata strumentale, come Alexander (1979) ed altri suggeriscono, nel permettere ai potenti di reclutare gli altri nei loro progetti, incluso quello della guerra.
4 Un altro caso ben documentato di selezione di gruppo spiega la pratica della condivisione dell'Ilama tra individui non legati da rapporti di parentela nell'altopiano peruviano (Flannery, Marcus e Reynold 1989 e Weinstein, Shugart e Brandt 1983).

6 | MICROECONOMIA
ALTRUISMO RECIPROCO E RECIPROCITA’ FORTE
I modelli nel capitolo 7 hanno mostrato come le strutture delle interazioni sociali
possano rendere comportamenti cooperativi come le strategie “occhio per occhio
benevolo” (“nice tit-for-tat”) una miglior risposta reciproca, anche quando gli individui
posseggono le convenzionali preferenze egoistiche sui risultati delle loro azioni. La
cooperazione convenzionale é una forma di ciò che il biologo R. Trivers (1971) ha
chiamato altruismo reciproco, ovvero, azioni che conferiscono un beneficio agli altri
attraverso un costo a se stessi nei casi in cui ci si aspetta un beneficio successivo
reciproco tale da controbilanciare il costo affrontato. L'altruismo reciproco e l’altruismo
famigliare (kin altruism) – l'azione, costosa per chi la compie, a beneficio dei membri della
famiglia o di altri parenti stretti – sono spiegazioni comuni di atti apparentemente
generosi tra individui ed altri animali.
Le interazioni ripetute e sfaccettate, permettendo la ritorsione contro le azioni
anti-sociali, contribuiscono senza dubbio al successo evoluzionistico degli atti
apparentemente generosi. Allo stesso tempo l'altruismo reciproco non costituisce una
spiegazione adeguata di quelle forme di cooperazione e mutua assistenza così diffuse tra
gli essere umani. Anzitutto, la gran parte dell'evidenza sperimentale riguardo le
preferenze altruistiche (capitolo 3) é basata su giochi con interazioni non ripetute, o sulla
fase finale di giochi ad interazione ripetuta. E' molto improbabile che in questi
esperimenti i soggetti non siano consapevoli di tale struttura basata su un’interazione
non ripetuta. Vi é un'evidenza molto forte del fatto che le persone distinguono tra
interazioni ripetute e non, e adattano i loro comportamenti di conseguenza. L'evidenza
non sperimentale é ugualmente importante: molti atteggiamenti comuni in guerra così
come nella vita di tutti i giorni non sono facilmente spiegabili dall'aspettativa di un
successivo comportamento reciproco.
In secondo luogo, le condizioni degli uomini primitivi possono aver reso il
meccanismo ripetizione-punizione uno strumento inefficace per diffondere
comportamenti altruistici. I membri delle bande nomadi potevano evitare facilmente la
punizione semplicemente spostandosi. Inoltre, in molte situazioni critiche all'evoluzione
umana, la ripetizione delle interazioni é abbastanza probabile, come nel caso di gruppi
che rischiavano l'estinzione a causa di conflitti o condizioni climatiche sfavorevoli.
In terzo luogo, il celebrato “Folk Theorem” mostra che se le interazioni ripetute
sono sufficientemente probabili ed i tassi di sconto sufficientemente bassi, l'equivalente

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |7
gioco ad
€
n persone della strategia “occhio per occhio benevolo” (“nice tit-for-tat”) ed
altre strategie più complesse, possono sostenere un equilibrio di Nash con alti livelli di
cooperazione (Fudenberg e Maskin 1986). Ma il “Folk Theorem” mostra anche che
quando il meccanismo di ripetizione-punizione in effetti funziona, funziona troppo bene
nel senso che sostiene un vasto numero di risultati – alcuni di essi appena più cooperativi
della mutua defezione – mentre non fornisce spiegazioni sul perchè gli equilibri più
cooperativi e più efficienti possano essere favoriti ai risultati meno cooperativi. Alcuni
modelli recenti (Fudenberg e Maskin 1990) sono stati in grado di restringere l'insieme
degli equilibri sostenuti dalla ripetizione. Ma essi richiedono che gli attori vivano
all'infinito o (equivalentemente) che abbiano dei tassi di preferenza intertemporale pari a
zero, o altre assunzioni non realistiche.
Infine, come Boyd e Lorberbaum (1987), Joshi (1987), ed altri hanno
puntualizzato, é difficile sostenere una cooperazione attraverso la ripetizione e la
ritorsione quando le interazioni non sono diadiche – come nei mercati di cambio studiati
nel capitolo 7 – ma invece coinvolgono un gran numero di persone. Eppure, le
interazioni tra gruppi con un vasto numero di persone sono abbastanza comuni, ad
esempio la difesa o il rischio in comune o la costruzione della reputazione di un gruppo.
Per capire il problema, consideriamo un gioco del bene pubblico con individui in
cui ogni membro contribuisce se ognuno degli altri membri contribuisce e defeziona
in caso contrario. Se i membri a volte adottano un comportamento idiosincratico (non
una miglior risposta) o l'osservazione del comportamento altrui é soggetta ad errore,
eventi casuali scoraggeranno la cooperazione, in quanto vi sarà sempre qualcuno che
crederà che qualcun altro non contribuirà.
La stessa fragilità affligge anche strategie che apparentemente sono non così
stringenti. Consideriamo una strategia di cooperazione condizionale: si coopera
(contribuisce) se almeno
€
n −m tra gli altri membri cooperano nella fase finale, dove
€
m < n . Chiamiamo tale strategia
€
m -Cooperare. La sola altra strategia possibile é il
Defezionare incondizionatamente. Consideriamo tale popolazione nell'equilibrio di Nash
in cui
€
n +1−m stanno giocando la strategia
€
m -Cooperare e adottano
Defezionare (vi devono essere
€
m persone che defezionano in equilibrio poiché
altrimenti cambiare strategia da
€
m -Cooperare a Defezionare costituirebbe una miglior
risposta). Supponiamo che con una ridotta probabilità
€
ε , coloro che adottano la
strategia
€
m-Cooperare cambiano la loro strategia (o per lo meno ciò è quanto osservato
dagli altri membri). Abbiamo visto sopra come la popolazione adotterà Defezionare se

8 | MICROECONOMIA
un singolo membro tra coloro che adottavano
€
m -Cooperare defeziona (o viene
percepito tale). Quindi, un tale equilibrio di Nash sarà sostenuto in un dato periodo
solo se tutti i
€
n +1−m continueranno a giocare la strategia
€
m-Cooperare e questo
avviene con probabilità
€
(1−ε) n+1−m che diventa molto piccola per un
€
n grande. Quindi,
nei gruppi larghi, gli equilibri cooperativi sostenuti da strategie tipo
€
m-Cooperare sono
molto vulnerabili a causa eventi casuali.
Parte del problema delle strategie tipo
€
m-Cooperare deriva dal fatto che nei
gruppi grandi, non sempre ci si può basare sulla punizione inflitta a coloro che
defezionano attraverso la mancata cooperazione degli membri del gruppo.
Supponiamo che
€
m = 0 , così che tutti gli
€
n +1 membri adottano la strategia
€
m -
Cooperare e continuano a cooperare se gli altri membri non defezionano. Se un singolo
membro defeziona in qualsiasi periodo, allora tutti gli altri membri defezionano per
sempre. Supponiamo che, come forma di punizione dell'unica persona che defeziona, la
strategia
€
m-Cooperare produca un “male pubblico”: tutti i membri – gli
€
n che
€
m-
Cooperano e l'unico che defeziona – si accollano la perdita del beneficio della
cooperazione. Si può notare che un tale problema esiste, anche se in una forma molto
più attenuata nel caso diadico con la semplice strategia tit-for-tat: colui che defeziona si
accolla metà (anziché l' del costo totale derivante dalla mancata cooperazione.
Ovviamente vi é un vasto numero di strategie possibili e la dimostrazione che una di esse
– -Cooperare – non funzioni non significa che nessuna possa funzionare. Allo stesso
tempo, i problemi con la strategia -Cooperare, come modo attraverso il quale
l’egoismo può promuovere la cooperazione, sono abbastanza generali e possono
interessare la maggior parte delle strategie possibili in un tale contesto, se non tutte.
Quindi, mentre la punizione egoistica può indurre la cooperazione nelle relazioni
diadiche o di piccola scala, essa é costosa da implementare nel caso di gruppi più grandi.
Come risultato, ed anche per le ragioni descritte sopra, il tentativo di spiegare – sia in
economia che in biologia evoluzionistica – tutti o la maggior parte degli atteggiamenti
altruistici come “egoistici nel lungo periodo” non sembra persuasivo. Tocqueville aveva
ragione. A volte un atto apparentemente generoso é semplicemente tale – un
comportamento costoso che beneficia un altro membro del gruppo senza nessun
probabile comportamento reciproco verso l'individuo altruista.
Di solito, una tale forma di altruismo incondizionato é rivolta ai propri famigliari,
ma vi sono anche frequenti episodi in cui essa é rivolta a sconosciuti, come mostra
l'aneddoto raccontato all'inizio del capitolo 3. Come abbiamo visto, la reciprocitá forte – la

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |9
predisposizione a cooperare, punire o premiare gli altri, condizionata al loro
comportamento persino in una situazione one shot o in altre situazioni in cui non vi é
alcuna aspettativa di un eventuale ritorno – é anch'essa una forma comune di
comportamento osservata negli esperimenti. “Forte” sta a sottolineare che si tratta di un
comportamento altruistico da non confondere con l'altruismo reciproco di Trivers, che
alla fine non é affatto altruismo, e potrebbe essere chiamato reciprocità debole. Al
contrario dell'altruismo incondizionato, il comportamento dell'altruista forte dipende dal
tipo di persona con cui esso sta interagendo. Usando le parole del poema epico Norse
del tredicesimo secolo The Edda (Clarke 1923:55), un reciprocatore forte sarà “amico dei
suoi amici” e “sorriderà ai sorrisi e mentirà con i traditori”.
Una forma importante di reciprocità forte é la punizione altruistica, ovvero,
l'affrontare un costo per infliggere costi a coloro che violano le norme benefiche per il
gruppo. Il comportamento é altruistico se induce una maggiore aderenza alle norme,
incrementando il beneficio medio del gruppo. L'evidenza sperimentale della punizione
altruistica deriva dai giochi dei beni pubblici, passati in rassegna nel capitolo 3, ed é parte
della strategia del Punitore, descritta nel capitolo 11. La punizione altruistica permette di
avere come obiettivo coloro che violano le norme e non si basa sull'aspettativa di
guadagni futuri, evitando, quindi, alcuni degli svantaggi delle strategie come la strategia
-Cooperare nei gruppi grandi. Ma, come ogni altra forma di altruismo, essa crea un
enigma a livello di evoluzione.
Né l'altruismo incondizionato né la reciprocità forte sono facilmente spiegabili
come miglior risposte definite sull'insieme dei guadagni materiali del gioco. Ma se gli
altruisti incondizionati e i reciprocatori forti sopportano il costo di conferire dei benefici
agli altri, essi saranno sempre svantaggiati in qualsiasi processo evolutivo che favorisce
coloro che hanno un guadagno maggiore. Il fatto che le persone esibiscano
comunemente certi comportamenti necessita una spiegazione. Parte della risposta
riguarda gli effetti della competizione tra gruppi.
LA COEVOLUZIONE DEI TRATTI INDIVIDUALISTICI E DI GRUPPO
Le pratiche umane individuali altruistiche possono essere sorte ed avere persistito
perchè gli individui dei gruppi nei quali tali pratiche erano prevalenti ne hanno tratto
beneficio, anche se coloro che di fatto le mettevano in atto beneficiavano meno dei
propri compagni di gruppo. Sappiamo che i tratti del comportamento individuale

10 | MICROECONOMIA
possono proliferare in una popolazione dove gli individui copiano i vicini che sembrano
avere successo. Così pure le norme distributive, le convenzioni linguistiche o i
comportamenti individuali che sostengono le forme di governo o i sistemi di diritti di
proprietà, si diffondono o scompaiono attraverso l'emulazione delle caratteristiche dei
gruppi di successo ad opera dei membri con meno successo. Questo processo spesso ha
luogo come risultato di forme di competizione militare, economica o di altro tipo.
Charles Darwin (1873:156), nell'epigrafe del capitolo 11, si riferisce al coraggio, alla
simpatia o all'altruismo come possibili esempi: questi tratti proliferarono perchè “una
tribù che possedeva tali qualità si sarebbe diffusa e sarebbe stato vittorioso sulle altre
tribù”.
Quindi, i tratti formalmente altruistici (costosi per l'individuo che li compie ma
benefici per il gruppo), che possono proliferare sotto l'influenza della selezione del
gruppo, includono i comportamenti dannosi ai membri degli altri gruppi. In effetti, il
processo che modelliamo qui di seguito può esser meglio descritto dimostrando il
successo evolutivo dei gruppi egoistici piuttosto che degli individui generosi5. Anche se la
definizione convenzionale di altruismo si riferisce solo alle relazioni all'interno del
gruppo. Gli individui dei nostri modelli interagiscono anche con i membri di altri gruppi.
Nel modello, gli individui altruistici conferiscono benefici ai compagni del gruppo
mentre arrecano costi o perdite materiali agli estranei. I termini “benefico al gruppo” o
“egoista” si riferiscono dunque ai soli rapporti con gli altri membri dello stesso gruppo.
Come é stato da lungo tempo riconosciuto, nelle popolazioni composte da
gruppi caratterizzati da un maggior livello di cooperazione tra i membri stessi piuttosto
che con gli estranei, i processi evolutivi possono essere distinti in effetti della selezione
tra gruppi e all'interno del gruppo. Dove il grado di riproduzione dei tratti di successo
dipende dalla composizione del gruppo e dove le differenze nella composizione tra i
gruppi persistono attraverso il tempo, la selezione dei gruppi (a volte chiamata selezione a
più livelli) contribuisce a determinare la rapidità e la direzione del cambiamento
evolutivo. Il modello della prima rivoluzione dei diritti di proprietà nel capitolo 11 é un
esempio di tale processo. Il classico problema della selezione dei gruppi sorge quando gli
effetti tra i gruppi favoriscono la proliferazione di tratti altruistici, come, ad esempio,
l'altruismo che sarebbe penalizzato dalla pura selezione individuale. La selezione di
gruppo costituisce uno sbocco per l'evoluzione dell'altruismo.
5 Stiamo parafrasando Laland, Pdling-Smee e Feldman (2000:224).

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |11
Pochi studiosi delle popolazioni umane hanno messo in dubbio che le istituzioni,
le nazioni, le imprese, le bande e gli altri gruppi possano essere soggetti alla pressione
selettiva che opera a livello di gruppo piuttosto che a livello individuale. Ma, fino poco
tempo fa, la maggior parte dei modelli formali dei processi evolutivi era stata fornita da
biologi e la maggior parte di essi concluse che gli effetti a livello di gruppo non potevano
essere controbilanciati dagli effetti della selezione individuale all'interno del gruppo,
eccezion fatta per i casi in cui vi fossero delle circostanze particolari che incrementavano
le differenze tra gruppi rispetto a quelle all'interno del gruppo. La valutazione negativa
della rilevanza empirica della selezione di gruppo deriva, in primo luogo, dalla
presunzione che il tasso di selezione all'interno del gruppo sia più rapido rispetto a
quello della selezione tra i gruppi. Ciò deriva in parte dal fatto che le differenze tra
gruppi sono soprattutto dovute al caso e quindi sono insignificanti rispetto alle
differenze all'interno del gruppo. Quindi, i modelli della selezione di gruppo furono
considerati come un fallimento rispetto al loro obiettivo, ovvero, quello di spiegare il
successo evolutivo dei comportamenti altruistici. Come risultato, mentre la spiegazione
dei comportamenti benefici per il gruppo si é focalizzata su meccanismi basati su
rapporti di parentela o similarità, il notevole livello di altruismo non basato su tali
rapporti era stato interpretato come altruismo reciproco o rimaneva, per la maggior
parte, non spiegato6.
Ma gli studi successivi (vedi le letture consigliate) suggeriscono che gli
impedimenti alla selezione di gruppo possano essere meno generali di quanto affermi la
critica. Inoltre, la selezione di gruppo può essere di importanza considerevolmente
maggiore tra gli umani che non tra gli animali. Tra le caratteristiche della natura umana,
che possono favorire la selezione tra i gruppi, vi é la nostra capacità di sopprimere le
differenze fenotipiche all'interno dei gruppi attraverso la condivisione delle risorse, la
mutua assicurazione, il consenso sulle decisioni da prendere, la trasmissione culturale
conformista, le forme di differenziazione sociale che supportano alti livelli di interazioni
distribuite in gruppi, il mantenimento della frontiere dei gruppi, ed i frequenti conflitti
tra gruppi diversi. Altri animali si comportano, in parte, allo stesso modo, ma non
completamente. La selezione di gruppo può funzionare sui tratti del comportamento che
6 Ovviamente, quelle descritte non esauriscono tutte le spiegazioni offerte. Simon (1990), Caporael et al.
(1989), ed altri hanno proposto un meccanismo per mezzo del quale comportamenti costosi ma benefici per il gruppo esercitano free riding sui comportamenti che beneficiano l'individuo (la “docilità” per esempio) con cui essi sono pleitoricamente associati. Gintis, Smith e Bowles (2002) mostrano come un comportamento costoso per l'individuo che lo mette in atto ma benefico per il gruppo possa proliferare se esso costituisce un segnale veritiero del valore di uno come compagno o partner di coalizione.

12 | MICROECONOMIA
sono trasmessi sia geneticamente che culturalmente. Il modello dell'evoluzione dei diritti
di proprietà del capitolo 11 considerava gli effetti della selezione di gruppo sui tratti
trasmessi culturalmente, In questo capitolo consideriamo un modello di selezione di
gruppo dei tratti trasmessi geneticamente.
Affronteremo due questioni. In primo luogo, cosa contribuisce all'evoluzione
delle forme, individualmente costose e benefiche al gruppo, di altruismo nei confronti
delle persone non legate da rapporti di parentela? E, in secondo luogo, cosa contribuisce
al successo delle strutture istituzionali comuni a livello di gruppo, che Parson (1964) ha
definito “universali evolutivi” come gli Stati, la condivisione delle risorse e la
monogamia, che sono emersi ed hanno proliferato ripetutamente in un'ampia varietà di
circostanze durante il corso della storia umana? Il processo di coevoluzione che andremo
a studiare e simulare é basato sull'idea che le due questioni di cui sopra possano essere
meglio risolte congiuntamente piuttosto che singolarmente.
Un esempio di certe caratteristiche strutturali a livello di gruppo é costituito dal
livellamento delle istituzioni, come la monogamia e la condivisione del cibo tra i non
parenti, ovvero quelle istituzioni che riducono le differenze all'interno del gruppo nella
capacità riproduttiva o nel benessere materiale. Riducendo le differenze nel successo
individuale all'interno del gruppo (in benessere, guadagni materiali o altre misure), certe
strutture possono aver attenuato le pressioni selettive all'interno del gruppo, le quali
operano contro le pratiche costose per l'individuo ma benefiche per il gruppo, dando,
quindi, ai gruppi che le mettono in atto, vantaggi in termini di competizione verso gli
altri gruppi7. In questo caso, la presenza diffusa di caratteristiche strutturali del gruppo,
come il livellamento delle istituzioni viene spiegato dal loro contributo alla proliferazione
dei tratti individuali benefici per il gruppo di appartenenza e dal contributo di questi
tratti alla sopravvivenza del gruppo.
L'idea secondo la quale la soppressione della competizione all'interno del gruppo
può influenzare, in maniera rilevante, la dinamica evolutiva é stata ampiamente
riconosciuta in alcuni insetti ed altre specie. In un articolo che esamina il caso del
fungo Dictyostelium discoideum, Steven Frank (1995:520) scrive: “La teoria dell'evoluzione
non ha spiegato come la competizione tra le unità di livello più basso sia soppressa nella
formazione di unità evolutive di livello più alto,” aggiungendo che “il mutuo vigilare e il
7 Nel modello consideriamo la “condivisione delle risorse”. Da notare che mentre essa può essere
motivata da egualitarismo, sicurezza, o altri motivi, i suoi effetti attenuano le differenze all'interno del gruppo.

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |13
rafforzamento dell’equità (fairness) evolutiva sono anche richiesti per l'evoluzione della
crescente complessità sociale”. Christopher Boehm (1999:211) si é riferito al processo
della sanzione irrogata dal gruppo agli agenti anti-sociali modellati nel capitolo 11 come
“una 'rivoluzione politica' vissuta dagli umani del Paleolitico che crearono le condizioni
sociali sotto le quali la selezione di gruppo poteva efficacemente supportare i geni
altruistici”. Similmente, Irenaus Eibl-Eibesfeldt (1982:177) si riferiva all'importanza della
“indottrinamento per rafforzare i valori, per obbedire all'autorità, e ... per condividere
principi etici” e pensava che “attraverso tali strategie, i gruppi diventano talmente
compatti da agire come unità di selezione.”
Nelle pagine che seguono, presenteremo un modello del processo di selezione di
gruppo basato su una semplificazione notevole dei processi evolutivi – l'equazione di
Price – interpretata e corretta per affrontare le peculiarità dell'evoluzione nelle
popolazioni umane. Gli effetti tra i gruppi sono basati su guerre periodiche tra i gruppi
in cui i “vincitori” sostituiscono i “perdenti” occupando i loro luoghi. Le estinzioni dei
gruppi quindi giocano un ruolo importante nel processo evolutivo.
Anzitutto spiegheremo come l'analisi dei conflitti tra i gruppi possa illuminare
l'evoluzione dei comportamenti individuali formalmente altruistici. Successivamente
svilupperemo un modello di riproduzione differenziale dei tratti individuali soggetti alla
selezione a più livelli attraverso i conflitti di gruppo, le estinzioni e le nascite. Questo
modello degli effetti della selezione di gruppo sui tratti trasmessi geneticamente può
essere usato per studiare l'evoluzione culturale8. Useremo poi una simulazione ad agenti
(agent-based) per determinare le condizioni secondo le quali un tratto costoso per
l'individuo portatore e benefico per il gruppo possa proliferare nella popolazione (i valori
dei parametri chiave riguardano la frequenza dei conflitti nel gruppo, gli aggiornamenti
degli individui, la grandezza del gruppo e la migrazione tra gruppi). La popolazione
simulata viene calibrata così da riprodurre le condizioni sociali ed ecologiche dei 50.000
anni precedenti l’avvento dell'agricoltura, un periodo abbastanza lungo da modellare i
processi di selezione a livello individuale e di gruppo in modo da avere i maggiori effetti
sull’evoluzione genetica. Le simulazioni mostrano che in assenza di istituzioni a livello di
gruppo che proteggono l'individuo altruistico dai non altruisti, le pressioni della
selezione all’interno del gruppo supportano l'evoluzione dei tratti altruistici solo quando
i conflitti tra i gruppi sono molto frequenti, i gruppi sono piccoli, ed i tassi di migrazione
bassi. Tuttavia, quando le istituzioni a livello di gruppo vengono introdotte ed
8 Vedi Bowles (2001).

14 | MICROECONOMIA
assoggettate alle pressioni della competizione insieme ai tratti individuali, l'altruismo
prolifera per un ampio insieme di parametri, che includono approssimazioni plausibili
degli ambienti dei nostri antenati.
LA LOGICA DELLA SELEZIONE MULTI-LIVELLO
Molte delle organizzazioni che rivestono un ruolo importante nello studio della
società umana sono aggregati di organizzazioni a livelli più bassi: le nazioni sono
composte da imprese, famiglie, classi sociali che, a loro volta, sono composte da persone
che a loro volta sono composte da cellule e così via. La nostra rappresentazione della
struttura sociale é semplicemente la distribuzione di queste organizzazioni ai livelli più
alti o più bassi e dei modi con cui esse interagiscono. Il processo di cambiamento può
quindi essere considerato attraverso l’evoluzione di tali entità, con alcune di esse che si
diffonderanno e diventeranno comuni, altre subiranno un declino e scompariranno con
conseguenti cambiamenti nelle interrelazioni tra le varie organizzazioni. La selezione a
più livelli rappresenta il processo tramite il quale l'evoluzione di una caratteristica a livello
individuale viene influenzata dalle pressioni competitive che operano sia a livello
individuale che ai livelli più alti.
La selezione a più livelli a volte appare come un gioco di prestigio tramite il quale
una caratteristica benefica per il gruppo ma apparentemente destinata a fallire dal punto
di vista evolutivo potrebbe non di meno proliferare nonostante abbia tassi di
riproduzione più bassi, violando le regole fondamentali dell'evoluzione. Ma, se
appropriatamente modellata, la selezione di gruppo non rappresenta un'alternativa alla
teoria evolutiva standard che tiene conto del cambiamento e della stabilità nella
distribuzione dei tratti in una popolazione con differenti caratteristiche. Piuttosto essa
rappresenta un'estensione del metodo standard che tiene conto degli effetti del gruppo
sulla riproduzione. Non vi sono conigli da tirar fuori da un cappello: la selezione di
gruppo é semplicemente una forma di interazione non casuale già introdotta nel capitolo
7 come segmentazione sociale. I tratti benefici per il gruppo evolvono sotto le pressioni
della selezione di gruppo poiché essi beneficiano di una più alta probabilità di interazione
con gli altri tratti evolutivi. Consideriamo un tratto singolo che può essere assente o
presente in ciascun individuo di una popolazione sufficientemente ampia i cui membri
appartengono ciascuno ad uno dei numerosi gruppi. Per concretezza, consideriamo un
comportamento altruistico (A) – per esempio il coraggio nella difesa del gruppo come
nell'esempio di Darwin – che comporta per l'individuo un costo e conferisce un

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |15
beneficio ad un qualsiasi membro del gruppo
Tabella 13.1. Il gioco dell'altruismo
A N A b-c -c N b 0
Nota: i payoff appartengono ai giocatori per riga.
Indichiamo con il fatto che l'individuo del gruppo possegga il tratto,
la situazione opposta (i soggetti senza il tratto sono gli N). Supponiamo che i
costi e i benefici dell'altruismo vengano misurati in termini di benessere, attraverso il
numero di riproduzioni (nel periodo successivo) dell'individuo portatore del tratto,
cosicché un membro in un gruppo composto esclusivamente da altruisti produce
€
b −c
riproduzioni in più rispetto ad un membro in un gruppo senza altruisti. Nel momento in
cui assumiamo che
€
b −c > 0, l'altruismo é benefico per il gruppo. Allo stesso tempo, se
messo a confronto con gli altri membri dello stesso gruppo, il benessere dell'altruista
risulterà più basso rispetto a quello dei non altruisti, cosicché la selezione all'interno del
gruppo lavorerà a sfavore degli altruisti.
Il punto di Darwin era che se la competizione tra gruppi influenza anche il
benessere, il tratto altruista potrebbe, non di meno, proliferare. La selezione di gruppo
funziona nel modo seguente. Lavorando nel tempo discreto, supponiamo che
€
p e
€
p' rappresentino, rispettivamente, la frazione della popolazione che possiede il tratto in un
dato periodo di tempo e nel periodo successivo, e
€
Δp = p'−p . George Price (1970) Ha
dimostrato che
€
Δp può essere suddiviso tra gli effetti di gruppo e quelli individuali.
Definiamo
€
πij il numero delle riproduzioni nel periodo successivo di un individuo di
tipo nel gruppo . Il processo di riproduzione può avvenire tramite copia culturale,
eredità genetica o qualsiasi altra causa conforme con l'equazione sotto. Il modello che
segue é basato sulla riproduzione differenziale dei tratti trasmessi geneticamente.
Supponiamo che dipenda, additivamente, dal tratto del tipo e dalla
frequenza di quel tratto nel gruppo, , nel modo seguente
(13.1) dove e rappresentano, rispettivamente, gli effetti parziali della frequenza
del tratto nel gruppo su e la presenza del tratto nell'individuo (gli indici si riferiscono
agli effetti di gruppo e dell'individuo), e denota il benessere di riferimento. Definiamo

16 | MICROECONOMIA
come l'effetto della frequenza del tratto nel gruppo sul numero medio delle
riproduzioni (la differenza nel numero delle riproduzioni ad opera di un individuo in un
gruppo composto interamente da coloro che posseggono il tratto ed un gruppo che ne é
del tutto privo é data da ). Quindi, usando le definizioni sopra, e
. Segue che, seguendo Price (1970)
(13.2) o
dove rappresenta il numero medio di riproduzione della popolazione
(normalizzate all'unità) e l'operatore indicate il valore attesa sta a significare una somma
pesata tra i gruppi (i pesi sono in relazione alla grandezza del gruppo). Il primo termine
cattura l'effetto di selezione di gruppo (positivo), mentre il secondo rappresenta l'effetto
della selezione individuale (negativo). Tralasciando casi particolari come quello di una
varianza pari a zero, ne segue che la frequenza interna di un tratto é stazionaria nel
momento in cui i due termini si uguagliano in termini di grandezza assoluta (assumendo
che sia i che le varianze non siano fonte di instabilità). Dato che il secondo termine é
negativo, la frequenza del tratto all'interno di tutti i gruppi sopravissuti tenderà a
diminuire col tempo. Ma, poiché é positivo, tale tendenza verrà controbilanciata dal
benessere medio più alto per il gruppo con una maggiore frequenza degli A.
Dunque, la condizione di stazionarietà per (eq. 13.2) mostra che
quando
(13.3)
con
Il termine a sinistra rappresenta il rapporto costi-benefici del tratto altruistico. Il
termine al lato destro descrive invece il rapporto della varianza del tratto tra i gruppi sulla
somma di quella all'interno del gruppo e di quella tra i gruppi. Si può facilmente

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |17
mostrare (vedi Bowles 2001 e l'esempio illustrato sotto) che tale rapporto misura la
differenza tra la probabilità che un altruista venga accoppiato con un altro altruista
P(A|A) e quella secondo cui un non altruista venga a contatto con un altruista P(A|N).
Quindi
Il rapporto tra le varianze rappresenta, conseguentemente, una misura del grado di
segmentazione derivante non dall'accoppiamento non casuale, ma dal fatto che la
popolazione é strutturata a gruppi. L'eq. 13.3 mostra che affinché un tratto altruista
proliferi, maggiore é il costo del tratto (in relazione al beneficio), maggiore deve essere la
varianza tra i gruppi (rispetto alla varianza all'interno del gruppo). Quando la varianza tra
i valori medi del gruppo é zero, la probabilità di incontrare un altruista diventa
indipendente dal tipo di ciascun individuo. La selezione di gruppo sarebbe, dunque, non
operativa e solo forme non costose di benefici per il gruppo prolifererebbero.
Similmente, quando , tutti i gruppi sono omogenei e un individuo
incontra solo persone del suo stesso tipo, indipendentemente dalla composizione della
popolazione totale. In tal caso, la selezione all'interno del gruppo é assente e la selezione
tra gruppi é l'unica forza al lavoro. In questo caso (estremo), si può dire che il gruppo
rappresenti la sola unità di selezione. Quindi, la forza della selezione di gruppo dipende
dalla grandezza del beneficio (per il gruppo) rispetto al costo individuale (
€
b e
€
c
nell'esempio) e dalla misura in cui i gruppi differiscono nella frequenza media del tratto
rispetto la varianza all'interno del tratto nel gruppo.
Coloro che hanno familiarità con la biologia della popolazione riconosceranno
l'eq. 13.3, espressa come , come una versione della regola di Hamilton, che
spiega come un tratto altruistico, anche se raro, possa proliferare. Da questo punto di
vista, la selezione a più livelli non é distinguibile dai processi evolutivi basati su altre
forme di distribuzione (ad esempio, selezione sulla base dei rapporti di parentela, o altre
forme di segmentazione all'interno del gruppo o raggruppamenti).
Un esempio chiarirà il processo. Supponiamo che una popolazione sia composta
da due gruppi di uguali dimensioni, con le frazioni di individui altruistici in ciascuno pari
a e , cosicché . Dalla matrice utilizzata precedentemente
sappiamo che il payoff degli altruisti é minore del payoff dei non altruisti. Quindi, gli
18 | MICROECONOMIA
altruisti sono svantaggiati nel processo di riproduzione. Ciò può esser visto dalle
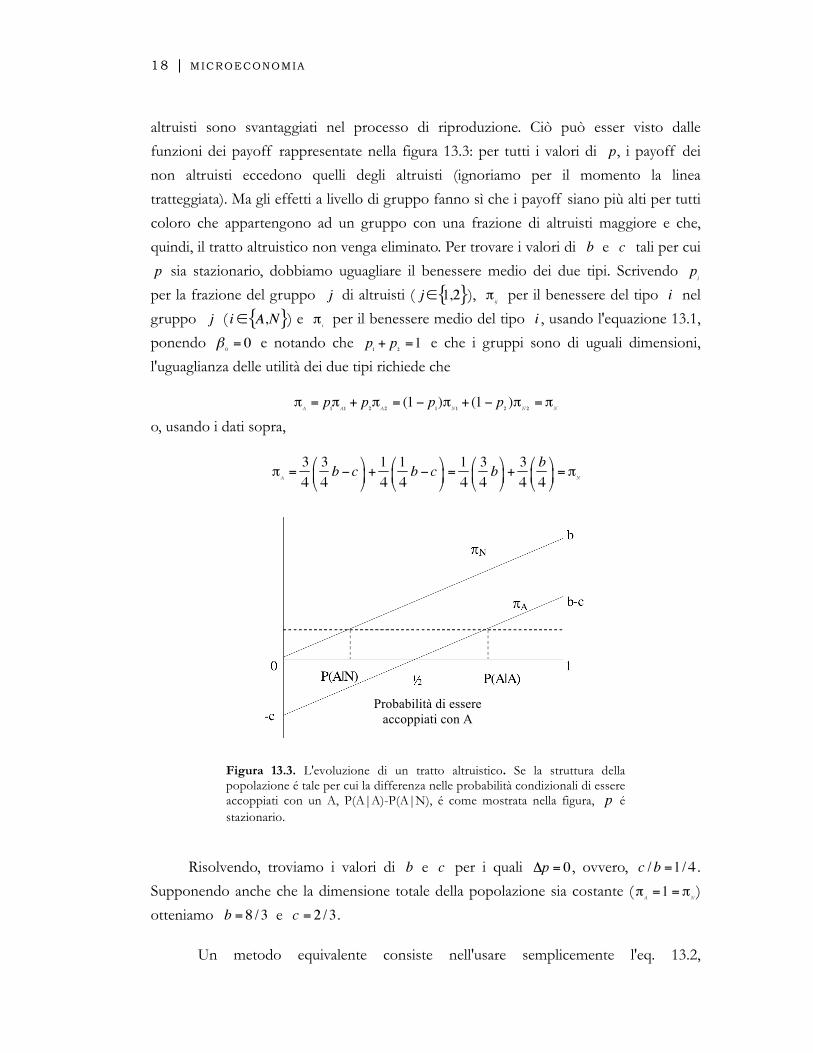
funzioni dei payoff rappresentate nella figura 13.3: per tutti i valori di , i payoff dei
non altruisti eccedono quelli degli altruisti (ignoriamo per il momento la linea
tratteggiata). Ma gli effetti a livello di gruppo fanno sì che i payoff siano più alti per tutti
coloro che appartengono ad un gruppo con una frazione di altruisti maggiore e che,
quindi, il tratto altruistico non venga eliminato. Per trovare i valori di e tali per cui
sia stazionario, dobbiamo uguagliare il benessere medio dei due tipi. Scrivendo
per la frazione del gruppo di altruisti ( ), per il benessere del tipo nel
gruppo ( ) e per il benessere medio del tipo , usando l'equazione 13.1,
ponendo e notando che e che i gruppi sono di uguali dimensioni,
l'uguaglianza delle utilità dei due tipi richiede che
o, usando i dati sopra,
Figura 13.3. L'evoluzione di un tratto altruistico. Se la struttura della popolazione é tale per cui la differenza nelle probabilità condizionali di essere accoppiati con un A, P(A|A)-P(A|N), é come mostrata nella figura, é stazionario.
Risolvendo, troviamo i valori di e per i quali , ovvero, .
Supponendo anche che la dimensione totale della popolazione sia costante ( )
otteniamo e .
Un metodo equivalente consiste nell'usare semplicemente l'eq. 13.2,
Probabilità di essere accoppiati con A

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |19
considerando che per e che
cosicché, usando l'eq.13.1 abbiamo
da cui, per otteniamo come condizione per ,
riproducendo i risultati di cui sopra. Inoltre, riutilizzando l'eq. 13.3 ed usando i valori
empirici dell'esempio abbiamo
come ci si poteva aspettare.
Quindi, per i valori , la frequenza del tratto altruistico crescerà, eccedendo
un mezzo nel periodo successivo. Ciò avviene perché la grandezza relativa del gruppo
più altruistico cresce, controbilanciando il declino della frazione degli altruisti in ogni
gruppo. La proliferazione del tratto costoso per il singolo individuo ma benefico per il
gruppo viene spiegata attraverso la struttura a gruppi della popolazione, che tiene conto
del fatto che gli altruisti tendono ad accoppiarsi con altri altruisti più di frequente
rispetto alla media della popolazione (nonostante l'accoppiamento casuale all'interno dei
gruppi). Quindi, la probabilità di incontrare un altro altruista, condizionata al fatto di
essere altruisti, é data da
mentre un non altruista incontra un altruista con probabilità
La differenza tra le due probabilità condizionali - ¼ – é data dal vantaggio atteso a
beneficio del tratto altruista attraverso la distribuzione favorevole tra i gruppi, risultando
in un modo equivalente di rappresentare l'eq. 13.3
La figura 13.3 mostra come la struttura di gruppo della popolazione superi gli
svantaggi dell'affrontare i costi derivanti dal comportamento altruistico. Mentre i payoff
dei non altruisti eccedono sempre quelli degli altruisti, data una certa probabilità di incontrare

20 | MICROECONOMIA
gli altruisti, la differenza della probabilità di incontrare un altruista, dato un certo tipo, (
) controbilancia tale svantaggio.
L'esempio mostra come la selezione di gruppo possa permettere la proliferazione
di un tratto altrimenti non possibile. L'analisi é comunque incompleta. L'equazione di
Price fornisce una descrizione sommaria dell'equilibrio piuttosto che una descrizione
completa del sistema dinamico. Da essa possiamo derivare la condizione di stazionarietà
per
€
p, ma non tiene conto dei movimenti delle varianze su cui é basato il movimento di
€
p. Nella maggior parte dei modelli biologici, i meccanismi che incrementano la varianza
tra i gruppi (ad esempio la mutazione genetica) sono deboli e tendono ad essere
sommersi dagli effetti omogeneizzanti della selezione stessa, insieme alla migrazione tra i
gruppi. Questa é la ragione per cui le pressioni della selezione di gruppo tra gli animali
non umani vengono considerate deboli. Ma, tra gli umani, dove la dimensione effettiva
del gruppo é piccola (ad esempio i membri di un gruppo nomade) e dove i gruppi di
frequente si dividono sia in risposta ad un aumento di dimensione, che a causa di
tensioni interpersonali all'interno del gruppo, gli errori di accoppiamento aumenteranno
la varianza tra i gruppi. Per ogni modello minimamente fedele alle circostanze empiriche
della natura umana, l'unico modo pratico per determinare se gli effetti che incrementano
la varianza sono forti abbastanza da rendere la selezione di gruppo un'influenza
importante sull'evoluzione, é quello di simulare la popolazione strutturata a gruppi con
valori dei parametri ragionevoli.
UN MODELLO DI SIMULAZIONE AD AGENTI DELLA SELEZIONE A
PIU’ LIVELLI
In assenza delle due istituzioni – distribuzione delle risorse e segmentazione
all'interno del gruppo – introdotte a breve, il processo di selezione all'interno del gruppo
é modellato (per l'individuo ) attraverso l'equazione dinamica di riproduzione standard
(13.4)
Ora, immaginiamo che il gruppo abbia adottato la pratica, comune tra i selvaggi
in cerca di pascoli ed altre comunità, della condivisione delle risorse all'interno del
gruppo. Una certa frazione della risorse che acquisisce un individuo – magari un tipo
specifico di bene come avviene tra gli Achè (Kaplan e Hill 1985) – viene depositata in
una pentola comune che viene suddivisa equamente tra tutti i membri del gruppo. Una
tale istituzione di condivisione può essere modellata come una tassa lineare,
€
tj∈ 0,1[ ) ,
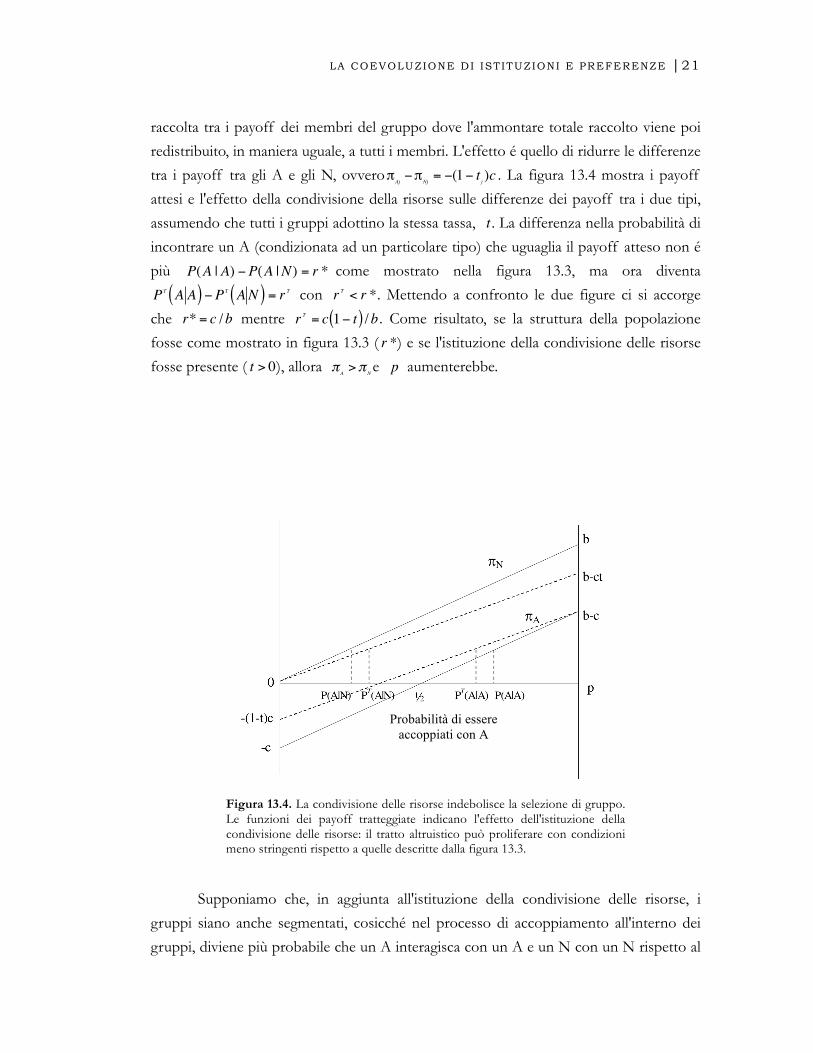
LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |21
raccolta tra i payoff dei membri del gruppo dove l'ammontare totale raccolto viene poi
redistribuito, in maniera uguale, a tutti i membri. L'effetto é quello di ridurre le differenze
tra i payoff tra gli A e gli N, ovvero
€
πAj−π
Nj= −(1− t
j)c . La figura 13.4 mostra i payoff
attesi e l'effetto della condivisione della risorse sulle differenze dei payoff tra i due tipi,
assumendo che tutti i gruppi adottino la stessa tassa,
€
t . La differenza nella probabilità di
incontrare un A (condizionata ad un particolare tipo) che uguaglia il payoff atteso non é
più
€
P(A | A) −P(A |N) = r * come mostrato nella figura 13.3, ma ora diventa
€
PT A A( ) −PT A N( ) = r T con
€
r T < r *. Mettendo a confronto le due figure ci si accorge
che
€
r* = c /b mentre
€
r T = c 1− t( ) /b . Come risultato, se la struttura della popolazione
fosse come mostrato in figura 13.3 (
€
r *) e se l'istituzione della condivisione delle risorse
fosse presente (
€
t > 0), allora
€
πA
>πNe
€
p aumenterebbe.
Figura 13.4. La condivisione delle risorse indebolisce la selezione di gruppo. Le funzioni dei payoff tratteggiate indicano l'effetto dell'istituzione della condivisione delle risorse: il tratto altruistico può proliferare con condizioni meno stringenti rispetto a quelle descritte dalla figura 13.3.
Supponiamo che, in aggiunta all'istituzione della condivisione delle risorse, i
gruppi siano anche segmentati, cosicché nel processo di accoppiamento all'interno dei
gruppi, diviene più probabile che un A interagisca con un A e un N con un N rispetto al
Probabilità di essere accoppiati con A

22 | MICROECONOMIA
caso dell'accoppiamento casuale. Supponiamo che la probabilità che un membro A del
gruppo venga accoppiato con un altro A non sia ma e che la
probabilità che un membro N del gruppo sia accoppiato con un A sia .
Come nel capitolo 7, definiamo il grado di segmentazione del gruppo j, o la
differenza nelle probabilità condizionali che un A incontri un A e un N un A negli
accoppiamenti all'interno del gruppo. Astraendo per il momento dalla tassa (
€
tj= 0):
€
πAj−π
Nj= s
jb −c . La segmentazione riduce lo svantaggio, in termini di payoff atteso,
dell'altruista poiché, all'interno di un dato gruppo, diviene molto più probabile incontrare
un altruista, così come diviene molto più probabile per un N incontrare un N. Se
€
sj> c /b, ∀j gli A, nella media, avranno un payoff più alto degli N all'interno di ogni
gruppo, e gli A prolifereranno come risultato della selezione sia tra i gruppi che
all'interno dei gruppi. Quindi, entrambi i membri nell'equazione di Price saranno positivi.
Per affrontare il classico problema di selezione di gruppo assumiamo che
€
s < c /b cosicché gli A proliferano se le pressioni della selezione di gruppo sono
abbastanza forti. Come la condivisione delle risorse, la segmentazione é una convenzione
e viene diffusa culturalmente.
Tenendo conto sia della segmentazione che della condivisione delle risorse, le
differenze tra i payoff attesi ricevuti dagli N e dagli A all'interno di un gruppo diventano
€
1− tj( ) sj
b −c( ) cosí che si ha
€
Δpj= p
j(1− p
j)(1− t
j)(s
jb −c) (13.5)
da cui (mettendo a confronto le eq. 13.4 e 13.5) risulta chiaro che entrambe le
istituzioni ritardano la selezione all'interno dei gruppi contro gli A. Ciò può esser visto
notando che
€
∂Δpj
∂t= −p
j(1− p
j)(s
jb −c)
∂Δpj
∂s= p
j(1− p
j)(1− t
j)b
(13.6)
Per
€
pj∈ 0,1( ) entrambe le espressioni sono positive, il che implica che sia la
segmentazione che la condivisione delle risorse attenuano la selezione negativa contro gli
A. Da notare come l'effetto di ciascuna istituzione sia maggiore quando
€
pj é prossimo
ad un mezzo e quando l'altra istituzione è ad un livello basso. Quindi, in termini dei
benefici nel ritardare la selezione contro gli A, le istituzioni sono sostitute, non
complementi: i loro effetti benefici sono tanto maggiori quanto minore é la presenza

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |23
dell'altra istituzione.
La struttura del processo di aggiornamento é descritta nella figura 13.4. La
riproduzione individuale é soggetta a mutazioni cosicché, con una piccola probabilità,
€
e,
i discendenti possono essere sia A che N con uguale probabilità. Le istituzioni
rappresentate da
€
s e
€
t differiscono tra i gruppi e anch'esse evolvono. Quando vi sono
dei conflitti tra i gruppi, vince il gruppo con il payoff più alto. I membri del gruppo che
perde muoiono e il gruppo vincente va a popolare il sito occupato dai perdenti insieme ai
loro discendenti. I nuovi abitanti del sito adottano le istituzioni del gruppo vincente da
cui discendono. Anche le istituzioni sono soggette a una variazione stocastica, che
aumenta o diminuisce
€
s e
€
t casualmente in ogni periodo. Sia la condivisione delle
risorse che la segmentazione impongono dei costi al gruppo che le adotta. I gruppi più
segmentati potrebbero catturare con più difficoltà i benefici della diversità o delle
economie di scala mentre la condivisione delle risorse potrebbe ridurre gli incentivi ad
acquisire le risorse da condividere. Questi costi non vengono modellati formalmente, ma,
per catturare il loro impatto, i benefici medi del gruppo vengono ridotti di un
ammontare crescente e convesso rispetto ad e .
Tabella 13.1. Parametri chiave per la Simulazione
Valori di riferimento Intervalli considerati
Dimensione media del gruppo(n/g) 20 da 7 a 47
Tasso di Migrazione (m) 0.2 da 0.1 a 0.3
Probabilitá di conflitto (k) 0.25 da 0.18 a 0.4
Tasso di Mutazione (e) 0.001 da 0.01 a 0.000001
Nota: La dimensione totale della popolazione é n e vi sono g gruppi; m, k ed e sono indicati per ogni generazione. Gli altri parametri: beneficio, b:2, costo, c:1; payoff di riferimento: 10. Abbiamo variato la dimensione del gruppo variando n. Per le ragioni spiegate nel testo, restringiamo s in modo che non ecceda ½ mentre
€
t∈ 0,1[ ] . Il costo imposto al gruppo dalle due istituzioni é
pari a
€
1/2(s 2 + t 2 ) .
Insieme con Jung-Kyoo Choi e Astrid Hopfensitz ho simulato una popolazione
artificiale strutturata in venti gruppi. I valori di riferimento dei parametri esplorati della
popolazione sono descritti nella tabella 13.1. I valori chiave riguardano il tasso di
migrazione tra i gruppi, la dimensione dei gruppi e la frequenza dei conflitti tra i gruppi.
Dato che ogni conflitto é letale per i perdenti abbiamo deciso di utilizzare, come

24 | MICROECONOMIA
riferimento, una singola guerra ogni quattro anni. I valori di riferimento sono stati scelti
sulla base di una plausibilità empirica, la cui evidenza verrà discussa nella penultima
sezione.
Ogni simulazione viene iniziata senza altruisti né istituzioni presenti al tempo
zero, per vedere come essi possano proliferare se inizialmente rari (il processo di
mutazione individuale e di gruppo introduce una certa variabilità nella popolazione). Il
benessere di riferimento
€
βoé posto pari a 10 e i discendenti vengono prodotti in
proporzione alla quota del benessere individuale su quello totale del gruppo, cosicché, in
assenza di segmentazione e condivisione delle risorse, la differenza attesa nei payoff é
pari a
€
c =1 per cui gli N producono il dieci per cento in più di discendenti rispetto agli
A. Abbiamo considerato 10 simulazioni di 50.000 generazioni utilizzando diversi valori
dei parametri.
Una simulazione tipica appare nella figura 13.6. L'aumento di
€
p viene sostenuto
dall'aumento casuale sia di
€
s che di
€
t (tra il periodo 100 e 150). Quando
€
p raggiunge
livelli alti (dal periodo 532 a 588 per esempio) sia
€
s che
€
t diminuiscono, causando
tipicamente una diminuzione forte in
€
p. I successivi aumenti di
€
t ed
€
s avvengono
casualmente.
La dinamica che ne risulta emerge per le seguenti ragioni: quando la popolazione
é divisa equamente tra gli A e gli N, molti gruppi sono approssimativamente suddivisi
equamente. Come risultato (dall'eq. 13.6) gli effetti benefici della ritardata selezione
all'interno dei gruppi, che derivano dagli alti livelli di
€
s e
€
t , vengono massimizzati in
questa regione. Ma, se é ben superiore a 0.5, i benefici della protezione degli A
offerta dalle istituzioni diventano meno importanti. Allo stesso tempo le istituzioni sono
costose, per cui, quando é alto, é probabile che i gruppi con un sostanziale livello di
segmentazione o di condivisione delle risorse perdano i conflitti con gli altri gruppi e i
siti da loro occupati vengano popolati dai discendenti dei vincitori, che tipicamente
posseggono un livello di variabili istituzionali minore. Come risultato, quando le
istituzioni dei vincitori vengono imposte nei siti ripopolati, sia che diminuiscono.
Tabella 13.3. Le istituzioni ritardano la selezione all'interno del gruppo contro gli altruisti.
Istituzione
€
βi t Test
Nessuna -0.102 8.5

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |25
Condivisione delle risorse -0.08 16.6 Segmentazione -0.063 13.4 Entrambe -0.055 11.2
Nota: La colonna
€
βi fornisce la stima dei minimi quadrati del coefficiente
del valore medio del gruppo di
€
pj1− p
j( ) come valore per predire
(l'altro regressore é la varianza tra i gruppi, var ( )). L'ultima colonna fornisce il test t della stima.
Per esplorare ulteriormente l'impatto delle istituzioni sui processi di
aggiornamento, stimiamo l'equazione di Price, esplorando l'effetto di ciascuna istituzione
separatamente (ovvero, restringendo
€
s,t ad essere entrambe o nessuna pari a zero).
Usando i dati da 10.000 generazioni, regrediamo i valori osservati di
€
Δp sui valori del
periodo precedente relativi a var(
€
pj)e a
€
E var pj( ){ }, dove il secondo termine rappresenta
la media tra i gruppi delle varianze all'interno degli stessi. I coefficienti di queste variabili
rappresentano delle stime di
€
βGe
€
βi tratte dall'equazione 13.2. Come mostra la tabella
13.3, l'effetto combinato della condivisione delle risorse e della segmentazione é quello di
ridurre della metà l'effetto della selezione all'interno dei gruppi contro gli altruisti. Da
notare come, in assenza di istituzioni, la stima di
€
βi (0.102) sia molto vicina al valore

26 | MICROECONOMIA
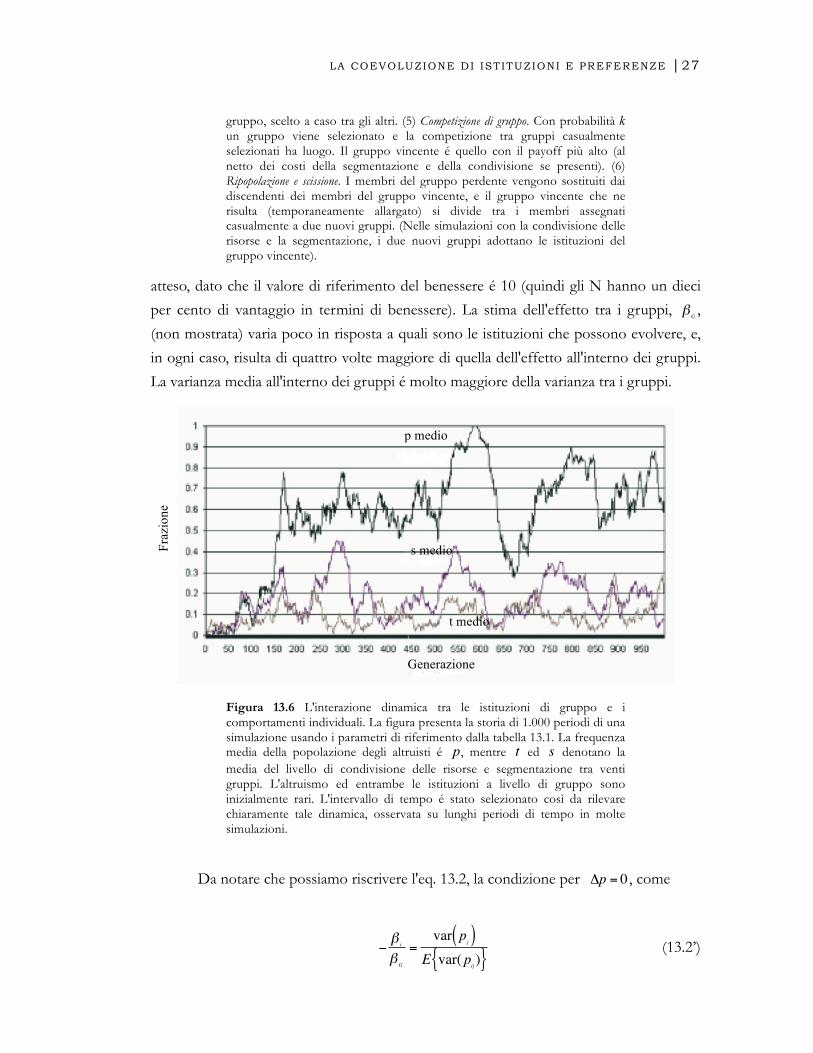
Figura 13.5. Le interazioni individuali e di gruppo. Assegniamo n individui ai g gruppi. Al tempo t=0 tutti sono N. (1) Accoppiamento. In ogni periodo, ogni membro del gruppo é accoppiato casualmente con un altro membro i cui payoff sono dati nel testo (in alcune simulazioni vengono modificati dalla regola di condivisione delle risorse). Con l'introduzione della segmentazione, i membri interagiscono con un tipo simile con probabilità s e vengono accoppiati casualmente con probabilità 1-s. (2) Riproduzione. Le riproduzioni delle generazioni correnti costituiscono la generazione successiva. Esse vengono prodotte estraendo (con sostituzione) alcuni individui dal gruppo di appartenenza dove la probabilità che si venga estratti é uguale alla quota del payoff dell'individuo rispetto a quella totale del gruppo. (3) Mutazione. Con probabilità e, un membro della generazione successiva non é la riproduzione dei suoi genitori, ma può essere A o N con uguale probabilità. (4) Migrazione. Con probabilità m ogni membro della nuova generazione si stabilisce in un
1) Accoppiare e Interagire
2) I Payoff determinano il numero di figli per ogni giocatore (fra parentesi)
3) Nuova generazione e mutazione
4) Migrazione
5) Competizione tra gruppi
6) Il gruppo vincente ripopola il sito del gruppo perdente e si divide in due nuovi gruppi
7) Nuovi gruppi
gruppo i
gruppo i
gruppo i
gruppo i
gruppo vincente i
gruppo vincente i Allargamento temporaneo
gruppo perdente j
Immigrazione dal gruppo y
Emigrazione verso il gruppo x
Agenti che giocano N
Agenti che giocano A
Agenti che cambiano per caso
Torna al Passo (1) Torna al Passo (1)
gruppo i gruppo j
LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |27
gruppo, scelto a caso tra gli altri. (5) Competizione di gruppo. Con probabilità k un gruppo viene selezionato e la competizione tra gruppi casualmente selezionati ha luogo. Il gruppo vincente é quello con il payoff più alto (al netto dei costi della segmentazione e della condivisione se presenti). (6) Ripopolazione e scissione. I membri del gruppo perdente vengono sostituiti dai discendenti dei membri del gruppo vincente, e il gruppo vincente che ne risulta (temporaneamente allargato) si divide tra i membri assegnati casualmente a due nuovi gruppi. (Nelle simulazioni con la condivisione delle risorse e la segmentazione, i due nuovi gruppi adottano le istituzioni del gruppo vincente).
atteso, dato che il valore di riferimento del benessere é 10 (quindi gli N hanno un dieci
per cento di vantaggio in termini di benessere). La stima dell'effetto tra i gruppi,
€
βG,
(non mostrata) varia poco in risposta a quali sono le istituzioni che possono evolvere, e,
in ogni caso, risulta di quattro volte maggiore di quella dell'effetto all'interno dei gruppi.
La varianza media all'interno dei gruppi é molto maggiore della varianza tra i gruppi.
Figura 13.6 L'interazione dinamica tra le istituzioni di gruppo e i comportamenti individuali. La figura presenta la storia di 1.000 periodi di una simulazione usando i parametri di riferimento dalla tabella 13.1. La frequenza media della popolazione degli altruisti é , mentre ed denotano la media del livello di condivisione delle risorse e segmentazione tra venti gruppi. L'altruismo ed entrambe le istituzioni a livello di gruppo sono inizialmente rari. L'intervallo di tempo é stato selezionato così da rilevare chiaramente tale dinamica, osservata su lunghi periodi di tempo in molte simulazioni.
Da notare che possiamo riscrivere l'eq. 13.2, la condizione per
€
Δp = 0 , come
€
−β
i
βG
=var p
j( )E var( p
ij){ }
(13.2’)
p medio
s medio
t medio
Generazione
Fraz
ione

28 | MICROECONOMIA
con
€
Δp < 0se il rapporto tra le varianze eccede il rapporto tra gli effetti all'interno
dei gruppi e quelli tra i gruppi, e viceversa. Nella nostra simulazione osserviamo una tale
relazione teorica? Usando le stime econometriche degli effetti tra i gruppi e all'interno
dei gruppi descritti nella tabella 13.2, così come i rapporti tra le varianze medie
osservate nelle stesse simulazioni, otteniamo i risultati della tabella 13.3. In assenza delle
due istituzioni, il rapporto tra l'effetto di selezione all'interno dei gruppi e quello della
selezione tra i gruppi,
€
−βi/β
G, é quasi il doppio del rapporto delle varianze tra e
all'interno dei gruppi. Quindi, nel caso di una popolazione con certi valori medi e stimati,
€
Δp sarebbe negativo. Non ci sorprende, infatti, trovare che il valore medio di
€
p é 0.06
nelle simulazioni su cui sono basate tali stime. Nel caso in cui entrambe le istituzioni
siano presenti, invece, il rapporto delle varianze uguaglia il rapporto tra gli effetti, il che
implica che gli effetti all'interno del gruppo che operano contro gli A sono esattamente
controbilanciati dagli effetti tra i gruppi che ne sostengono, invece, la proliferazione.
Nella simulazione su cui sono basate tali stime, il valore medio di
€
p é 0.51.
I conflitti tra i gruppi giocano un ruolo chiave nel sostenere sia le istituzioni a
livello di gruppo che l'altruismo a livello individuale. Nelle simulazioni riportate, la
frequenza attesa del conflitto é data da
€
1/k dove
€
k rappresenta la probabilità che un
gruppo venga estratto per un conflitto in ogni generazione. Sembra probabile che, in un
lungo periodo storico, la frequenza dei conflitti vari considerevolmente, forse in risposta
al bisogno di emigrare in tempi di variabilità climatica. Per esplorare la sensitività della
simulazione al variare della frequenza dei conflitti, abbiamo variato stocasticamente
utilizzando il sistema autoregressivo descritto nelle note della figura 13.7. Durante i
periodi in cui il conflitto era frequente (intorno alla 21.000-esima generazione) venivano
sostenuti alti livelli di altruismo, ma gli intervalli periodici di pace relativa tra i gruppi
(intorno alla 25.300-esima, 27.000-esima e 29.600-esima generazione) causavano delle
riduzioni drastiche della frazione degli A nella popolazione.
Abbiamo cercato di rispondere anche ad altre due domande. Nel caso in cui le
istituzioni a livello di gruppo non siano coevolute con l'altruismo a livello individuale,
quest'ultimo avrebbe potuto proliferare? E ancora, quanto sono sensibili le nostre
simulazioni alle variazioni dei parametri chiave? Per rispondere ad entrambe le domande,
abbiamo variato la dimensione del gruppo da 7 a 47, e, per ciascun valore della
dimensione, sono state simulate 50.000 generazioni, lasciando gli altri parametri ai loro
valori di riferimento. Lo studio é stato fatto lasciando prima coevolvere entrambe le

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |29
istituzioni, poi ciascuna singolarmente, ed infine nessuna delle due. Abbiamo seguito lo
stesso procedimento variando il tasso di migrazione da 0.1 a 0.3 e la probabilità del
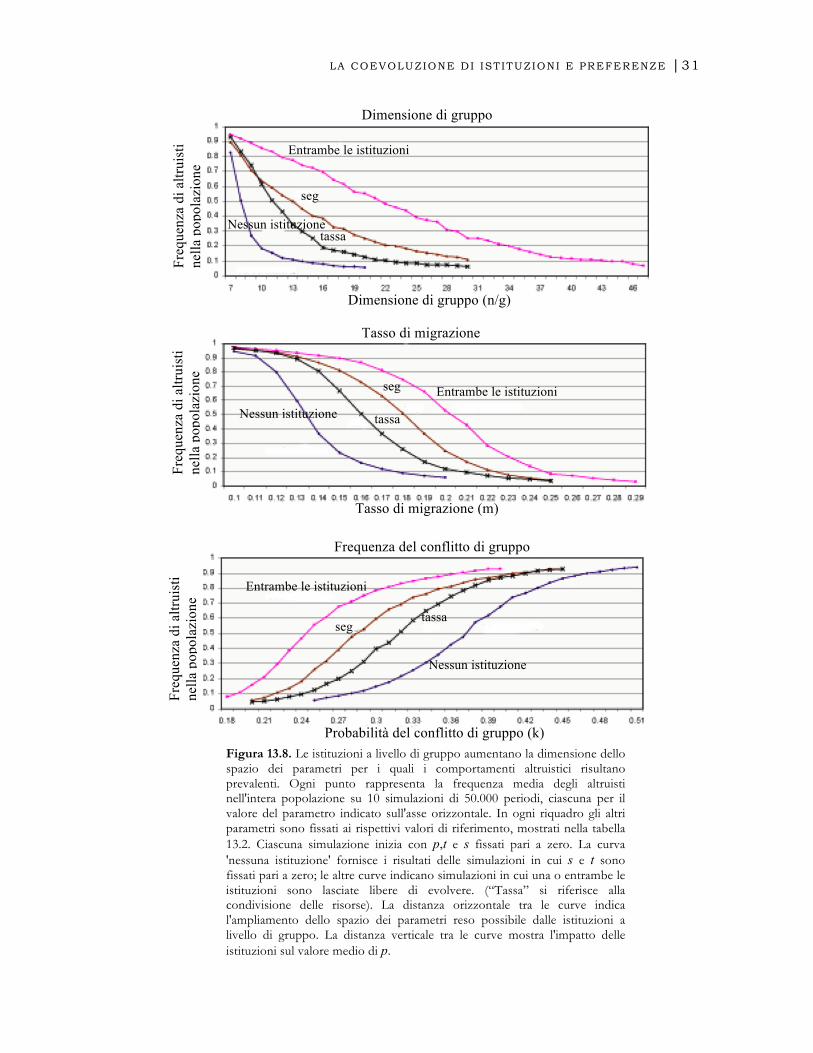
conflitto ( ) da 0.18 a 0.51. I risultati appaiono nella figura 13.8.
Tabella 13.3. Una stima dell'equazione di Price
Istituzioni Rapporto tra gli effetti Rapporto tra le varianze
Nessuna 0.25 0.13 0.06 Entrambe 0.13 0.13 0.51
Nota: Il rapporto tra le varianze rappresenta la media di
su 10.000 generazioni simulate, mentre il rapporto tra
gli effetti é dato da
€
−βi/β
G, stimato come descritto nella tabella 13.2. La
frazione degli A nella popolazione é .
Il riquadro superiore mostra che, quando entrambe le istituzioni non vengono fatte
evolvere, un gruppo di dimensione 7 sostiene alti livelli di altruismo, ma nei gruppi con
dimensione maggiore di 8, la frequenza degli altruisti diviene minore di 0.3.
Considerando come riferimento la dimensione del gruppo nel quale , vediamo
come, in assenza di istituzioni la dimensione critica sia 8, mentre quando entrambe sono
considerate,
€
p > 0.5 si manifesta nei gruppi di dimensioni minori di 22. I risultati per il
tasso di migrazione sono simili. In assenza delle istituzioni, sostenere un
€
p > 0.5
richiede un tasso di migrazione (per generazione) di 0.13, ma se entrambe le istituzioni
sono libere di evolvere, il tasso di migrazione critico é pari a 0.21. Il riquadro in fondo
mostra come le istituzioni permettano anche l'evoluzione di elevati livelli di altruismo
con un numero molto minore di conflitti. Anche una lettura “verticale” della figura può
essere illuminante: per esempio, il riquadro in fondo mostra che per , é
minore di 0.2 senza istituzioni, ma maggiore di 0.8 se entrambe le istituzioni sono libere
di evolvere9.
9 Le figura 13.8 e la tabella 13.2 suggeriscono che la segregazione ha un'influenza maggiore rispetto alla
condivisione delle risorse: la segmentazione, se considerata isolatamente, ha un effetto maggiore della condivisione quando anch'essa viene considerata da sola sia in ritardare la selezione all'interno del gruppo contro gli A sia in ampliare lo spazio dei parametri per i quali gli A costituiscono una notevole frazione della popolazione. Ciò é un artefatto delle nostre scelte del modello. Le funzioni di costo per e sono identiche, ma s ha un impatto maggiore nell'aggiornamento all'interno del gruppo, come può esser visto dall'eq. 13.6. Usando queste equazioni per mettere a confronto l'effetto di quando con quello di quando , vediamo come il primo sia volte il secondo, dove poiché l'azione altruistica é di beneficio al gruppo (Nella nostra simulazione e , cosicché che l'effetto di s è il doppio di quello di ). Inoltre, dall'eq.13.5 possiamo notare come, se ,
30 | MICROECONOMIA
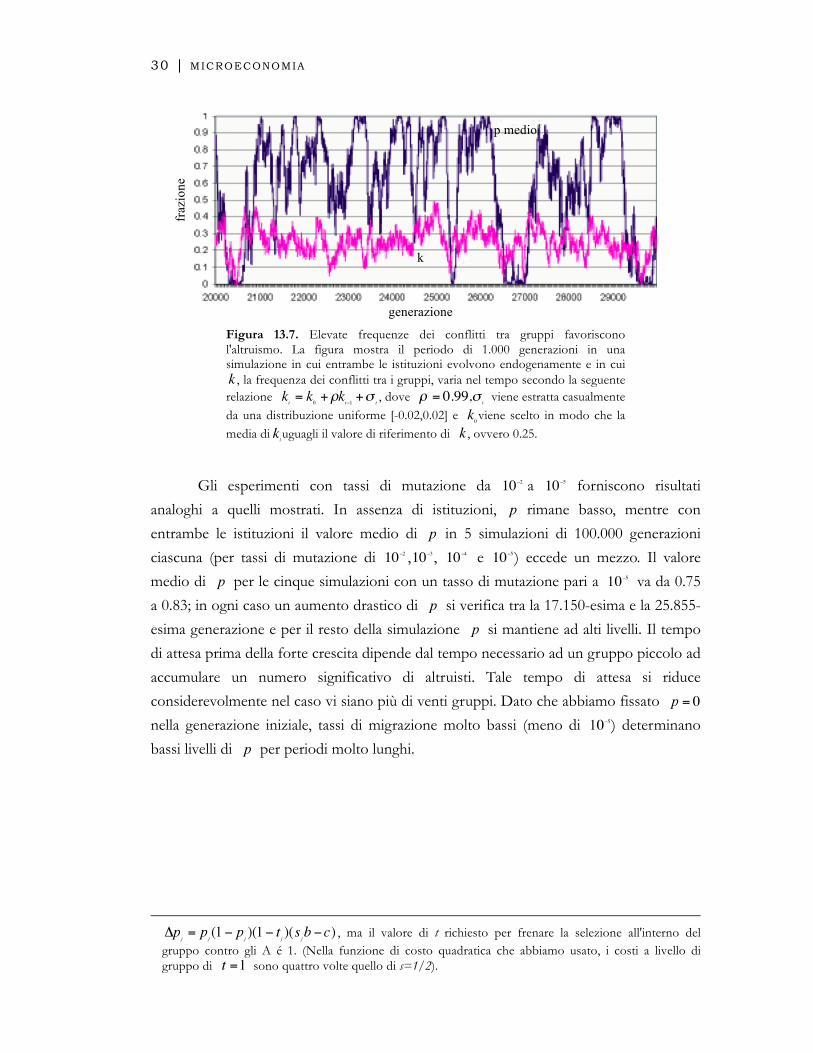
Figura 13.7. Elevate frequenze dei conflitti tra gruppi favoriscono l'altruismo. La figura mostra il periodo di 1.000 generazioni in una simulazione in cui entrambe le istituzioni evolvono endogenamente e in cui
, la frequenza dei conflitti tra i gruppi, varia nel tempo secondo la seguente relazione
€
kt= k
0+ρk
t−1+σ
t, dove
€
ρ = 0.99,σt viene estratta casualmente
da una distribuzione uniforme [-0.02,0.02] e viene scelto in modo che la media di uguagli il valore di riferimento di , ovvero 0.25.
Gli esperimenti con tassi di mutazione da
€
10−2 a
€
10−5 forniscono risultati
analoghi a quelli mostrati. In assenza di istituzioni, rimane basso, mentre con
entrambe le istituzioni il valore medio di in 5 simulazioni di 100.000 generazioni
ciascuna (per tassi di mutazione di
€
10−2 ,
€
10−3 ,
€
10−4 e
€
10−5) eccede un mezzo. Il valore
medio di per le cinque simulazioni con un tasso di mutazione pari a va da 0.75
a 0.83; in ogni caso un aumento drastico di si verifica tra la 17.150-esima e la 25.855-
esima generazione e per il resto della simulazione si mantiene ad alti livelli. Il tempo
di attesa prima della forte crescita dipende dal tempo necessario ad un gruppo piccolo ad
accumulare un numero significativo di altruisti. Tale tempo di attesa si riduce
considerevolmente nel caso vi siano più di venti gruppi. Dato che abbiamo fissato
nella generazione iniziale, tassi di migrazione molto bassi (meno di ) determinano
bassi livelli di per periodi molto lunghi.
, ma il valore di t richiesto per frenare la selezione all'interno del
gruppo contro gli A é 1. (Nella funzione di costo quadratica che abbiamo usato, i costi a livello di gruppo di sono quattro volte quello di s=1/2).
p medio
k
generazione
fraz
ione
LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |31
Figura 13.8. Le istituzioni a livello di gruppo aumentano la dimensione dello spazio dei parametri per i quali i comportamenti altruistici risultano prevalenti. Ogni punto rappresenta la frequenza media degli altruisti nell'intera popolazione su 10 simulazioni di 50.000 periodi, ciascuna per il valore del parametro indicato sull'asse orizzontale. In ogni riquadro gli altri parametri sono fissati ai rispettivi valori di riferimento, mostrati nella tabella 13.2. Ciascuna simulazione inizia con p,t e s fissati pari a zero. La curva 'nessuna istituzione' fornisce i risultati delle simulazioni in cui s e t sono fissati pari a zero; le altre curve indicano simulazioni in cui una o entrambe le istituzioni sono lasciate libere di evolvere. (“Tassa” si riferisce alla condivisione delle risorse). La distanza orizzontale tra le curve indica l'ampliamento dello spazio dei parametri reso possibile dalle istituzioni a livello di gruppo. La distanza verticale tra le curve mostra l'impatto delle istituzioni sul valore medio di p.
Dimensione di gruppo
Dimensione di gruppo (n/g)
Freq
uenz
a di
altr
uist
i ne
lla p
opol
azio
ne
Freq
uenz
a di
altr
uist
i ne
lla p
opol
azio
ne
Freq
uenz
a di
altr
uist
i ne
lla p
opol
azio
ne
Tasso di migrazione
Tasso di migrazione (m)
Frequenza del conflitto di gruppo
Probabilità del conflitto di gruppo (k)
Entrambe le istituzioni
Entrambe le istituzioni
Entrambe le istituzioni
seg
seg
seg
tassa
tassa
Nessun istituzione
tassa
Nessun istituzione
Nessun istituzione

32 | MICROECONOMIA
AMBIENTI EVOLUTIVI
Abbiamo descritto un processo secondo cui le istituzioni come la condivisione e
la segmentazione forniscono un ambiente all'interno del quale i tratti altruistici evolvono
e nel quale tali istituzioni proliferano nella popolazione grazie al loro contributo al
processo di evoluzione dei tratti benefici al gruppo. Questo modello fornisce anche
indicazioni sul processo di evoluzione dei comportamenti altruistici e delle istituzioni a
livello di gruppo? Dipende se lo spazio dei parametri in cui avviene tale processo
coevolutivo approssima l’ambiente rilevante nelle nostre simulazioni, ovvero i primi
50.000 o 100.000 anni dell’esistenza umana moderna, precedente alla drammatica
trasformazione della struttura sociale avvenuta con l’avvento dell’agricoltura intorno ad
11.000 anni fa.
Non si sa molto del tardo Pleistocene, ed è ben conosciuta la difficoltà di fare
inferenze riguardo all’organizzazione sociale dei gruppi umani durante tale periodo sulla
base delle semplici società contemporanee (Foley 1987, Kelly 1995). Con una certa
sicurezza possiamo dire, comunque, che le condizioni climatiche erano estremamente
variabili (Richerson, Boyd e Bettinger 2001), e che bande nomadi composte da soggetti
sia legati che non legati da vincoli di parentela e la mancanza di organizzazioni politiche
complesse costituivano forme comuni di organizzazione sociale.
La nostra dimensione di riferimento del gruppo, 20, è basata su
un’approssimazione della mediana dei 235 gruppi di cacciatori descritti da Binford
(2001), ovvero 19. La nostra scelta della grandezza del gruppo non è, comunque, del
tutto realistica. Occorre ricordare che le piccole dimensioni contribuiscono alle pressioni
della selezione aumentando la varianza tra i gruppi che aumenta quando quelli di
successo raddoppiano la loro dimensione e si dividono. In realtà, la scissione del gruppo
non è semplicemente l’esito di un’estrazione casuale ma piuttosto appare come un
processo di risoluzione dei conflitti politici in cui è probabile che soggetti legati da
vincoli di parentela e coalizioni rimangano insieme. Di conseguenza, è probabile che la
scissione abbia contributo alla varianza tra i gruppi in modi che il nostro modello non
può catturare. Uno studio sulla scissione tra le popolazioni amazzoni (Neves 1995:198)
riporta che:
La dimensione massima del villaggio è limitata dall’ammontare di relazioni di parentela e dalla solidarietà tra gli individui che deriva da tre fonti: relazioni di parentela, legami coniugali e influenza dei leader politici…La scissione nel villaggio è favorita, quindi, dall’indebolimento delle relazioni di parentela che deriva dalla crescita della popolazione; e quando avviene mantiene uniti i parenti stretti ma tende a separare quelli meno vicini…La linea potenziale della scissione è data dalla

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |33
divisione nella patrilinearitá10.
Dato che è probabile che i portatori dei tratti benefici al gruppo siano
numericamente e socialmente dominanti nei gruppi vincitori, essi possono aver messo in
atto ció che Hamilton (1975:137) ha chiamato assortative division, segregando i portatori
degli altri tratti, fintanto che il riconoscimento dei tratti o delle caratteristiche correlate
con i tratti lo permetta. In tal caso, i gruppi di dimensioni molto maggiori sosterrebbero i
processi evolutivi sopra descritti.
Si sa poco del conflitto di gruppo nel corso della storia umana primitiva. Sappiamo
però che le morti a causa di guerre costituiscono una frazione sostanziale delle morti
avvenute in molte delle societá antecedenti la nascita dello Stato delle quali si ha traccia
nei registri etnografici ed archeologici. La media riportata da Keeley (1996) per gli studi
etnografici delle societá pre-statali è 0.19 e per quelli archeologici è 0.16. Tali dati
possono essere messi a confronto con le stime molto più basse per l’Europa e per gli
Stati Uniti nel ventesimo secolo (molto inferiori a 0.1), con lo 0.03 per la Francia del
diciannovesimo secolo e con lo 0.02 per l’Europa Occidentale del diciassettesimo secolo.
Un registro di 200 guerre in 50 anni nel Mae-Enga nella Nuova Guinea, per esempio, ha
considerato 800 persone da una popolazione di 5000 e ne è risultato un tasso di mortalitá
annuale da guerre (0.0032 morti procapite per anno) che costituisce quasi il doppio di
quello della Germania e della Russia nel ventesimo secolo, ma che è molto inferiore a
quello medio delle società pre-statali di cui si ha testimonianza (Keeley 1996:195). È
difficile dire se tali episodi straordinariamente letali fossero comuni durante il tardo
Pleistocene. Ma possiamo avvalerci di alcune speculazioni basate su quanto sappiamo
riguardo al cambiamento climatico e ai probabili tassi di crescita della popolazione.
Christopher Boehm (2000a:19) scrive:
Sembra sensato supporre che, in ambienti prosperi e stabili, la densità della popolazione cresca e che le bande inizino a competere per le risorse, e che, alla fine, questo abbia creato problemi politici letali anche se, inizialmente, le risorse erano più che adeguate….Le varie vicende dei conflitti potrebbero aver aumentato la forza della selezione naturale che operava a livello di gruppi, in quanto alcune bande vennero decimate mentre altre si riprodussero e alla fine si scissero. [In risposta alle drammatiche oscillazioni climatiche nel periodo interglaciale del tardo Pleistocene], le bande nomadi erano costrette a cambiamenti molto frequenti, che consistevano sia nell’adattamento alle caratteristiche dei gruppi limitrofi che nei cambiamenti dell’ambiente naturale.
La sua conclusione è la seguente:
Verso la fine del Pleistocene, quando l’essere umano moderno cominciò ad affermarsi, il tasso di 10 Chagnon (1983:141-3) ha studiato il villaggio di Yanomamo che si suddivise e scoprí che la parentela
genetica media nel villaggioprima della scissione era piú bassa di ciascuna delle nuove unitá formatisi a seguito della divisione.

34 | MICROECONOMIA
estinzione dei gruppi avrebbe potuto crescere drammaticamente in quanto bande bisognose di cacciatori ben armati, stranieri privi di interazioni politiche ben consolidate si scontrarono di frequente, sia localmente che nel corso di lunghe migrazioni.
Carol Ember (1978) ha raccolto dati sulla frequenza dei conflitti tra una
cinquantina di bande nomadi nel presente o nel recente passato. Escludendo i pastori e
coloro che praticavano agricoltura sedentaria, il 64% dei gruppi affrontava dei conflitti
ogni due anni o piú frequentemente. Persino escludendo coloro che allevavano cavalli o
pescavano (tra i quali i conflitti erano più comuni), le guerre venivano considerate “rare”
solo nel 12% dei gruppi.
Mentre il movimento tra le unità etno-linguistiche era probabilmente poco
comune, sembra probabile che notevoli tassi di migrazione si verificavano, invece, tra le
bande che componevano tali unità. Il valore medio dei tassi di migrazione per le tredici
società studiate da Rogers (1990) è del 22 percento per generazione con il massimo (i
!Kung) minore di un mezzo. Dato che i dati di Rogers si riferiscono a gruppi di
dimensioni più ampie rispetto alle bande, essi potrebbero sottostimare in qualche modo i
tassi di migrazione.
Nonostante la natura di queste inferenze sia fortemente speculativa, sembra
possibile che gli ambienti sociali e fisici del tardo Pleistocene possano ricadere tra lo
spazio dei parametri delle traiettorie coevoluzionistiche illustrate nella figura 13.7. In tal
caso, il modello della selezione a più livelli con le istituzioni endogene può fornire per lo
meno una parziale descrizione di questo periodo cruciale dell’evoluzione umana.
CONCLUSIONE
Sembra dunque probabile che la predisposizione umana verso le attività benefiche
per il gruppo possa essere coevoluta con le comuni istituzioni che implementano la
condivisione delle risorse e la segmentazione sociale. In tal caso, questo approccio può
aiutare a capire perché gli umani siano stati portati a condividere e cooperare per il
raggiungimento di obiettivi comuni e allo stesso tempo siano stati pronti ad uccidere o
essere uccisi in nome di entità astratte come la nazione o la razza. Il modello fornisce
anche alcuni suggerimenti per capire perché certi atteggiamenti siano meno comuni tra
gli animali: la maggior parte degli animali sono incapaci di creare gli ambienti che
facilitano certi atteggiamenti, ovvero quelli costituiti da gruppi ben definiti composti
soggetti non legati da relazioni di parentela ma in cui vi siano dei codici di
comportamento comunemente compresi. E, come abbiamo visto, senza queste peculiari

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |35
strutture umane di interazione, l’evoluzione dei comportamenti umani altruistici sarebbe
stata molto improbabile. Un’implicazione ulteriore, sottolineata dalla figura 13.6, consiste
nel fatto che anche l’altruismo e la guerra possano essere coevolute. I comportamenti di
gruppo che rendono la cooperazione per i mutui benefici possibile tra gli umani,
rendono possibili anche le guerre su larga scala. E frequenti conflitti, come abbiamo
visto, possono aver costituito un contributo essenziale all’evoluzione dei tratti altruistici
che li facilitano. Per esplorare tale dinamica, abbiamo considerato endogena la probabilità
dei conflitti letali tra i gruppi; abbiamo assunto che
€
k vari congiuntamente alla
frequenza degli A nella popolazione. In queste simulazioni (non mostrate), la
popolazione spende virtualmente tutto il tempo in uno dei due stati: alte frequenze di
altruismo e conflitti molto frequenti o pochi altruisti e conflitti infrequenti (Bowles e
Choi 2003).
I modelli di selezione a piú livelli, simili a quello simulato in questo capitolo,
hanno anche dimostrato che la punizione altruistica dei violatori delle norme può
proliferare anche se rara e rimanere comune anche in gruppi abbastanza numerosi. In
uno di questi modelli (Boyd, Gintis e Richerson 2003) è stato sfruttato il fatto che il
costo della punizione dei violatori delle norme è piccolo quando coloro che aderiscono
costituiscono la maggioranza del gruppo. (Ciò é simile alla dinamica del modello del
Punitore-l’Usurpatore-Condivisore, nelle vicinanze dell’equilibrio di Rousseau). In questo
caso, le pressioni relativamente deboli della selezione di gruppo che derivano dai conflitti
tra i gruppi (come nel modello sopra) sono sufficienti per mantenere alte frequenze dei
punitori altruistici e per sostenere alti livelli di cooperazione. Un secondo modello, che
riguarda la reciprocitá forte (Bowles e Gintis 2003), considera una forma di punizione
dei violatori delle norme comune tra le bande nomadi di cacciatori: l’esercizio
dell’ostracismo da parte del gruppo. I conflitti di gruppo o le estinzioni di un gruppo
non giocano alcun ruolo in questo modello: coloro che vengono puniti soffrono una
perdita di benessere durante il periodo in cui essi non fanno parte del gruppo. I free rider
che aderiscono alla norma ma non puniscono non cacciano i reciprocatori
(contraccambisti) forti perché se i free rider divengono comuni in un gruppo, i violatori
della norme proliferano, riducendo il benessere medio dei gruppi cui appartengono.
Nell’introdurre i capitoli dal 10 al 13, ci siamo chiesti come le istituzioni possano
cambiare e come le persone e le regole che governano la loro vita possano coevolvere.
Abbiamo modellato tre fonti principali di cambiamento: trend secolari esogeni
(soprattutto, cambiamenti tecnologici e ambientali, come nel capitolo 11), gli effetti

36 | MICROECONOMIA
congiunti del caso e dell’azione collettiva (capitolo 12) e gli effetti congiunti del caso e
dei conflitti tra i gruppi (in questo capitolo). I modelli introdotti suggeriscono due modi
in cui il cambiamento può essere avvenuto endogenamente, rispettivamente, con l’azione
collettiva o con la competizione di gruppo. Considerati congiuntamente, i modelli
catturano almeno parte dei desiderata descritti all’inizio del capitolo 11, ovvero,
l’importante ruolo giocato da conflitti di interesse, caso, azione collettiva, persistenza
delle istituzioni inefficienti e traiettorie altamente irregolari del cambiamento sintetizzate
dal termine “equilibri punteggiati”. I modelli forniscono anche delle buone ragioni per
aspettarsi che le istituzioni inefficienti, pur se persistenti per periodi lunghi, nel lungo
periodo non se la passino bene quanto le altre istituzioni, identiche ma più efficienti.
Abbiamo anche visto come, per ben due ragioni differenti (fornite nel capitolo 12 e 13),
le istituzioni egualitarie possano essere favorite nel processo di evoluzione. Nei modelli
basati sulla teoria dei giochi stocastici evolutivi, il bacino di attrazione delle istituzioni
egualitarie è più ampio e, nei modelli di selezione a più livelli, le istituzioni egualitarie
ritardano la selezione all’interno del gruppo contro gli altruisti e ciò aumenta la capacità
del gruppo di sopravvivere nei conflitti con gli altri gruppi. Siamo stati in grado, dunque,
di spiegare, in parte, non solo i meccanismi causali dell’evoluzione delle istituzioni e
individuale, ma anche il concetto di Parson degli “universali evolutivi”, ovvero, le
istituzioni che ci si può aspettare siano molto diffuse in ambienti differenti, emergenti in
numerose occasioni e proliferanti anche se inizialmente rare.
La strategia concettuale alla base di tutti i modelli presentati è stata quella di
estendere un numero di approcci astratti, ispirati alla biologia – la teoria dei giochi
stocastici evolutivi e la decomposizione dei processi di selezione all’interno dei-tra i
gruppi dell’equazione di Price –modellando i processi distintivi delle interazioni sociali
umane. Quindi, l’approccio stocastico evolutivo è stato esteso tenendo conto del
perseguimento intenzionale degli interessi attraverso l’azione collettiva e il modello della
selezione a più livelli è stato corretto per dare una spiegazione della condivisione delle
risorse e della segmentazione sociale all’interno dei gruppi, così come delle guerre e delle
altre forme di conflitto tra i gruppi.
I risultati delle simulazioni suggeriscono l’utilità dell’approccio. L’orizzonte
temporale in cui la storia si spiega nei più semplici dei modelli stocastici evolutivi è
troppo lungo per risultare rilevante per spiegare l’evoluzione sociale, ma l’introduzione
delle interazioni locali e la correlazione delle azioni idiosincratiche, attraverso le tendenze
conformistiche, aumentano molto il passo del cambiamento. Similmente, in assenza di

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |37
istituzioni all’interno del gruppo quali la condivisione delle risorse e la segmentazione, le
condizioni per l’evoluzione dei tratti individuali benefici al gruppo sono molto stringenti.
Ma quando si permette a questi due importanti aspetti dell’interazione umana di
coevolvere con i tratti individuali, il processo di coevoluzione che ne risulta appare una
possibile spiegazione alla storia umana.

38 | MICROECONOMIA
Riferimenti bibliografici Alexander, R. D. (1979). Darwinism and Human Affairs. Seattle, University of
Washington Press. Anderson, P. (1974). Passages from Antiquity to Feudalism. London, NLB. Boehm, C. (1999). "The Natural Selection of Altruistic Traits." Human Nature 10(3):
205-252. Boehm, C. (2000). Hierachy in the Forest. Cambridge, Harvard University Press. Bowles, S. (2001). Individual Interactions, Group Conflicts, and the Evolution of
Preferences. Social Dynamics. S. Durlauf and P. Young. Cambridge, MA, MIT Press: 155-190.
Bowles, S. and H. Gintis (1984). State and Class in European Feudalism. Statemaking and Social Movements: Essays in History and Theory. C. B. a. S. Harding. Ann Arbor, University of Michigan Press.
Bowles, S. and H. Gintis (2003). Strong reciprocity: The origin of hunman cooperation. The Cultural and Genetic Evolution of Cooperation. P. Hammerstein. Cambrige, MIT Press.
Bowles, S., J.-K. Choi, et al. (2003). "The coevolution of individual behaviors and group level institutions." Journal of Theoretical Biology 223(2): 135-147.
Boyd, R. and J. Lorberbaum (1987). "No Pure Strategy Is Evolutionarily Stable in the Repeated Prisoner's Dilemma Game." Nature 327: 58-59.
Boyd, R., H. Gintis, et al. (2003). "The evolution of altruistic punishment." Proceedings of the National Academy of Science (USA) 20: 123-143.
Brady, T. A. (1985). Turning Swiss: Cities and Empire, 1450-1550. Cambridge, Cambridge University Press.
Bright, C. and S. F. Harding (1984). Statemaking and social movements : essays in history and theory. Ann Arbor, University of Michigan Press.
Caporael, L. R., R. M. Dawes, et al. (1989). "Selfishness examined: Cooperation in the Absence of Egoistic Incentives." Behavioral and Brain Sciences 12: 683-738.
Chagnon, N. A. (1983). Yanomamo: The Fierce People. New York, Holt, Rhinehart and Winston.
Clarke, D. E. M., Ed. (1923). The Havamal With Selections From Other Poems of the Edda, Illustrating the Wisdom of the North in Heathen Times. Cambridge, Cambridge University Press.
Darwin, C. (1873). The Descent of Man. New York, D. Appleton and Company. de Tocqueville, A. (1958 [1830]). Democracy in America, Volume II. New York NY,
Vintage. Eibl-Eibesfeldt, I. (1982). "Warfare, Man's Indoctrinability and Group Selection."
Journal of Comparative Ethology 60(3): 177- 198. Ember, C. (1978). "Myths About Hunter-Gatherers." Ethology 17(4): 439-449. Flannery, K., J. Marcus, et al. (1989). The Flocks of the Wamani: A Study of Llama
Herders on the Puntas of Ayacucho, Peru. San Diego, Academic Press. Foley, R. (1987). Another Unique Species: Patterns in Human Evolutionary Ecology.
New York, John Wiley and Sons. Frank, S. A. (1995). "Mutual Policing and Repression of Competition in the Evolution of
Cooperative Groups." Nature 377: 520-522.

LA COEVOLUZIONE DI ISTITUZIONI E PREFERENZE |39
Fudenberg, D. and E. Maskin (1986). "The Folk Theorem in Repeated Games with Discounting or with Incomplete Information." Econometrica 54(3): 533-554.
Fudenberg, D. and E. Maskin (1990). "Evolution and Cooperation in Noisy Repeated Games." American Economic Review 80(2): 275-279.
Gellner, E. (1983). Nations and nationalism. Ithaca, Cornell University Press. Gintis, H., E. A. Smith, et al. (2002). "Costly signaling and cooperation." Journal of
Theoretical Biology 213(1): 103-119. Hamilton, W. D. (1975). Innate Social Aptitudes of Man: an Approach from
Evolutionary Genetics. Biosocial Anthropology. R. Fox. New York, John Wiley and Sons: 133-155.
Herlihy, D. and C. Klapische-Zuber (1985). Tuscans and their Families. New Haven, Yale University Press.
Joshi, N. V. (1987). "Evolution of cooperation by reciprocation within structured demes." Journal of Genetics 66(1): 69-84`.
Kaplan, H. and K. Hill (1985). "Food Sharing among Ache Foragers: Tests of Explanatory Hypotheses." Current Anthropology 26(2): 223-246.
Keeley, L. (1996). War Before Civilization. New York, Oxford University Press. Kelly, R. C. (1985). The Nuer Conquest: The Structure and Development of an
Expansionist System. Ann Arbor, University of Michigan Press. Kelly, R. L. (1995). The foraging spectrum : diversity in hunter-gatherer lifeways.
Washington, Smithsonian Institution Press. Laland, K. N., F. J. Odling-Smee, et al. (2000). "Group Selection: A Niche Construction
Perspective." Journal of Conciousness Studies 7(1/2): 221-224. Levy, R. (1957). The Social Structure of Islam. Cambridge, Cambridge University Press. MacDonald, K. (1995). "The Establishment and Maintenance of Socially Imposed
Monogamy in Western Europe." Politics and the Life Sciences 14(1): 3-23. Mack Smith, D. (1959). Italy: A Modern History. Ann Arbor, University of Michigan
Press. Matthew, D. (1992). Atlas of Medieval Europe. Oxford, Andromeda, Ltd. Meletti, J. (2001). Difende un fornaio, assassinato. La Republica: (June 28)13. Neves, E. G. (1995). "Village Fissioning in Amazonia." Rev. do Museo de Arqueologica
e Ethnologia (S. Paulo) 5: 195-209. Parsons, T. (1964). "Evolutionary Universals in Society." American Sociological Review
29(3): 339-357. Pearson, K. (1894). "Socialism and Natural Selection." Fortnightly Review 56: 1-21. Price, G. R. (1970). "Selection and Covariance." Nature 227: 520-521. Richerson, P., R. Boyd, et al. (2001). "Was Agriculture Impossible During the
Pleistocene but Mandatory During the Holocene? A Climate Change Hypothesis." American Antiquity 66(3): 387-411.
Rogers, A. R. (1990). "Group Selection by Selective Emigration: The Effects of Migration and Kin Structure." American Naturalist 135(3): 398-413.
Simon, H. A. (1990). "A Mechanism for Social Selection and Successful Altruism." Science 250: 1665-1667.
Soltis, J., R. Boyd, et al. (1995). "Can Group-Functional Behaviors Evolve by Cultural Group Selection: An Empirical Test." Current Anthropology 36(3): 473-483.
Tilly, C. (1975). The Formation of National States in Western Europe. Princeton, Princeton University Press.
Tilly, C. (1990). Coercion, capital, and European states, AD 990-1990. Cambridge, Mass.,

40 | MICROECONOMIA
USA, B. Blackwell. Trivers, R. L. (1971). "The Evolution of Reciprocal Altruism." Quarterly Review of
Biology 46: 35-57. Wallerstein, I. (1974). The Modern World-System: Capitalist Agriculture and the Origins
of the European World-Economy in the Sixteenth Century. New York, Academic Press.
Weber, E. (1976). Peasants into Frenchmen: The Modernization of Rural France, 1870-1914. Stanford, Stanford University Press.
Weinstein, D. A., H. H. Shugart, et al. (1983). "Energy Flow and the Persistence of a Human Population: A Simulation Analysis." Human Ecology 11(2): 201-223.





























