Embed Size (px)
Citation preview

La determinación de la síntesis de proteína microbianaen el rumen: comparación entre marcadores microbianos (Revisión)
M.D. CarroDpto. de Producción Animal I, Universidad de León, 24071 León
RESUMEN
La cuantificación de la síntesis de proteína microbiana y de la degradación de la proteína del alimento en elrumen son dos puntos clave en todos los sistemas de valoración nutritiva para los animales rumiantes. La deter-minación de la síntesis microbiana se realiza mediante la utilización de marcadores microbianos, los cuales seclasifican en internos (constituyentes de los microorganismos ruminales) y externos (sustancias administradas alos animales que son incorporadas por los microorganismos). Sin embargo, ninguno de los marcadores utiliza-dos hasta el momento cumple todos los requisitos exigidos a un «marcador ideal». En esta revisión se analizanlas ventajas y los inconvenientes de los principales marcadores microbianos internos (ácidos nucleicos, basespúricas, ácido diaminopimélico, D-alanina, ácido aminoetilfosfónico, perfiles de aminoácidos y ATP) y externos(15N, 35S y 32P), y se discuten los resultados obtenidos en algunos estudios en los que se han utilizado variosmarcadores.
PALABRAS CLAVE: Proteína microbianaRumenMarcadores microbianosPurinas, 15N
INTRODUCCIÓN
La posesión del retículo-rumen, y de la población microbiana que alberga, permite alos animales rumiantes transformar las complejas estructuras de la pared celular de los ve-getales en elementos nutritivos que pueden ser utilizados, una vez absorbidos, para sumantenimiento y funciones productivas. Sin embargo, el hecho de que los alimentos seanfermentados en el retículo-rumen antes de la digestión gástrica dificulta el conocimientode los nutrientes aportados en la ración que son utilizables por el animal. En lo referente a
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
Recibido: 5-5-00Aceptado para su publicación: 16-3-01

la proteína, los aminoácidos que llegan al duodeno pueden tener tres orígenes diferentes:la proteína microbiana sintetizada en el rumen, la proteína del alimento que no ha sido de-gradada y la proteína endógena (descamaciones celulares, jugos digestivos, etc.). La pro-teína microbiana constituye, generalmente, una proporción considerable del flujo duode-nal de nitrógeno (N) aminoacídico en los animales rumiantes. Así, se calcula que cuandolos animales reciben raciones convencionales, la proteína microbiana representa entre el40 y el 90 % de los aminoácidos que llegan al intestino delgado (Schniffen y Robinson,1987), aunque en ocasiones puede alcanzar el 100 % (AFRC, 1992; Stern et al., 1994).Debido al papel que juegan los microorganismos ruminales en la digestión de las estructu-ras de la pared celular de los vegetales y a que la proteína microbiana tiene una gran cali-dad, las raciones de los rumiantes deben ser formuladas para permitir el crecimiento ópti-mo de estos microorganismos.
La cuantificación de la síntesis de proteína microbiana y la determinación de la de-gradación de la proteína de los alimentos en el rumen son dos puntos críticos en todos lossistemas de valoración nutritiva para los animales rumiantes (Clark et al., 1992), y cobranespecial relevancia en el caso de los animales que presentan altos niveles de producción,ya que un aporte insuficiente de aminoácidos puede limitar la misma (Nocek y Russel,1988). Uno de los mayores impedimentos para obtener información fiable sobre estos dosaspectos es la falta de un método simple y preciso para determinar la síntesis de proteínamicrobiana. La cuantificación del crecimiento microbiano en el rumen se puede llevar acabo únicamente mediante la utilización de marcadores microbianos (Schönhusen et al.,1995). Generalmente se utilizan animales provistos de cánulas en el rumen y el duodeno,y se determina el flujo duodenal de N no amoniacal (NNA). Posteriormente, es necesariodiferenciar el origen del N (microbiano, alimenticio y endógeno), y para ello se determinala relación marcador/N en la digesta duodenal y en una muestra de microorganismos ais-lados del rumen o del duodeno de los animales. La relación entre ambos valores (marca-dor/N en la digesta duodenal / marcador/N en los microorganismos) representa la fracciónnitrogenada de la digesta duodenal que es de origen microbiano. El resto del N que llegaal duodeno corresponde a la fracción proteica del alimento no degradada en el rumen y alas secreciones y descamaciones endógenas. Este procedimiento in vivo es el más amplia-mente aceptado, pero presenta tres puntos conflictivos. El primero de ellos es la cuantifi-cación de la digesta duodenal. Esta cuantificación puede realizarse mediante una colec-ción total de la digesta en animales provistos de cánulas reentrantes, ya que éstas permitenrecoger la digesta, muestrearla y volverla a introducir en el duodeno. Sin embargo, estetipo de cánulas no es muy utilizado, debido a que su implantación exige un doble corte enla pared del duodeno, lo que afecta a la inervación del intestino y a su motilidad (Wenhamy Whyburn, 1980). Por ello es más frecuente el uso de cánulas simples o en T. Estas cá-nulas únicamente permiten el muestreo de la digesta duodenal en momentos puntuales, ypara cuantificar el total de la digesta es necesario utilizar marcadores de flujo, lo que aña-de nuevas imprecisiones al proceso. Cada marcador presenta una serie de ventajas e in-convenientes (Warner, 1981), pero el método más utilizado actualmente es la técnica deldoble marcador propuesta por Faichney (1975), que combina un marcador de la fase sóli-da y otro de la líquida.
Un segundo punto conflictivo es el aislamiento de una fracción microbiana represen-tativa de toda la población ruminal. En la mayoría de los estudios en los que se ha deter-minado la síntesis de proteína microbiana in vivo, la muestra de microorganismos que setoma como referencia se ha aislado de la fase líquida del rumen, debido a que éste es el
6 M.D. CARRO

procedimiento más sencillo, aunque en algunos estudios se ha aislado también a partir delcontenido duodenal (Siddons et al., 1982; Kennedy et al., 1984; Schönhusen et al., 1995).Sin embargo, prácticamente en todos los estudios en los que se han aislado bacterias aso-ciadas a la fase sólida (BAS) y a la fase líquida (BAL) del rumen se ha observado queambas difieren en su composición química (Merry y McAllan, 1983; Legay-Carmier yBauchart, 1989), y que ésta a su vez difiere de la de los protozoos (Martin et al., 1994).En cuanto a los hongos ruminales, en el momento actual no existen datos definitivos so-bre su composición química y su contribución a la proteína microbiana.
El tercer punto conflictivo es la utilización de un marcador microbiano adecuado. Se-gún Broderick y Merchen (1992) un marcador microbiano «ideal» debería cumplir los si-guientes requisitos: (1) ser fácil de determinar y cuantificar, (2) no formar parte de los ali-mentos que reciben los animales, (3) distribuirse de manera uniforme en todas las espe-cies microbianas ruminales y (4) ser biológicamente estable. Dehority (1995) añade otros3 requisitos: (5) no ser absorbido en el tracto digestivo, (6) estar en una proporción cons-tante en los microorganismos ruminales en todas las fases de su crecimiento y (7) que to-das sus formas (p.e. libre o formando parte de células intactas de microorganismos rumi-nales o de alguna de sus partes) presenten el mismo ritmo de tránsito a través del tractodigestivo. Sin embargo, ninguno de los marcadores que se utilizan hasta el momento cum-ple todos los requisitos señalados anteriormente (Broderick y Merchen, 1992; Obispo yDehority, 1999), por lo que la elección de uno u otro para realizar estudios sobre la sínte-sis de proteína microbiana en el rumen resulta especialmente complicada. El objetivo deeste trabajo es revisar la información disponible sobre los diferentes marcadores micro-bianos, haciendo especial hincapié en los más utilizados, y comparar los resultados obte-nidos en aquellos estudios en los que se ha utilizado más de uno de ellos.
Los marcadores microbianos se clasifican en dos grandes grupos: marcadores inter-nos y externos. La principal característica de los marcadores internos es que son constitu-yentes de las células microbianas y por ello no necesitan ser administrados a los animalesexperimentales. Por el contrario, los marcadores externos deben ser administrados a losanimales experimentales para que sean incorporados por los microorganismos ruminalesy formen parte de sus estructuras.
MARCADORES MICROBIANOS INTERNOS
Los principales marcadores internos son: los ácidos nucleicos (ácido ribonucleico(ARN) y ácido desoxirribonucleico (ADN)), las bases púricas (purinas), el ácido diamino-pimélico (DAPA), la D-alanina, el ácido aminoetilfosfónico (AEPA), los perfiles de ami-noácidos y el ATP.
Ácidos nucleicos (ARN y ADN) y purinas
La utilización de estas sustancias como marcadores microbianos se basa en las altasconcentraciones de ADN y, especialmente, de ARN presentes en los organismos unicelu-lares (Broderick y Merchen, 1982). Su utilización como marcadores microbianos se re-monta a la década de los sesenta, en la que diversos autores observaron que la síntesis de
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 7

proteína microbiana en el rumen estaba relacionada con el flujo total de polinucleótidos(Ellis y Pfander, 1965) y con la excreción urinaria de derivados púricos (Topps y Elliot,1965). Posteriormente se han utilizado como marcadores microbianos el total de ácidosnucleicos (Zinn y Owens, 1980; Ushida et al., 1985), el ADN (Arambel et al., 1982) y elARN (Ling y Buttery, 1978; Merry y McAllan, 1983; Schönhusen et al., 1995), aunqueen los últimos años la utilización de las purinas ha adquirido un mayor protagonismo (Pé-rez et al., 1996a, 1997b; Calsamiglia et al., 1996; Carro y Miller, 1999a).
El análisis de purinas (Zinn y Owens, 1986) es simple, rápido y más barato que lamayoría de los otros procedimientos utilizados para estimar la síntesis microbiana en elrumen. Sin embargo, la utilización de las purinas como marcador microbiano presentatambién algunos inconvenientes. En primer lugar, los ingredientes que forman parte de lasraciones de los animales contienen, en mayor o menor cantidad, purinas. Algunos alimen-tos presentan contenidos muy inferiores a los existentes en los microorganismos rumina-les, pero otros (p.e. harina de pescado) presentan valores ligeramente menores a los en-contrados por algunos autores en estos microorganismos (Titgemeyer et al., 1989; Pérezet al., 1996b; Carro y Miller, 2001). McAllan y Smith (1973a) observaron que cuando seintroducían ácidos nucleicos libres (ARN y ADN) en el rumen, éstos eran degradados rá-pidamente a nucleósidos y bases, las cuales eran posteriormente catabolizadas (McAllan ySmith, 1973b). Sin embargo, la situación puede ser diferente en el caso de los ácidos nu-cleicos de los alimentos, ya que en este caso se encuentran protegidos por las estructurasde la pared celular (McAllan, 1982). La degradabilidad de las purinas de origen alimenti-cio se ha determinado en algunos estudios, pero los resultados obtenidos son contradicto-rios. Algunos autores (McAllan y Smith, 1973a; Schelling y Byers, 1984) concluyen queel flujo duodenal de purinas de origen alimenticio es insignificante, a pesar de que otrosestudios, en los que se marcaron los ácidos nucleicos microbianos con 32P (Smith et al.,1978) o con 15N (Pérez et al., 1997a), señalan que, dependiendo de la ración ingerida porlos animales, hasta un 30 % del flujo duodenal de purinas puede ser de origen alimenticio,lo que provocaría una sobreestimación de la síntesis de proteína microbiana cuando se uti-lizan las purinas como marcador. Pérez et al. (1996b) determinaron la degradación apa-rente y verdadera de las purinas de nueve alimentos (heno de alfalfa, harinas de pescado ycarne, gluten feed, cebada, maíz, tortas de soja y girasol, y paja de cebada) tras su incuba-ción en bolsas de nylon en el rumen de ovejas. Los valores de degradación verdadera delas purinas de estos alimentos tras 24 horas de incubación (valor correspondiente a un rit-mo de paso a través del rumen de 0,042 h–1) oscilaron entre 80 y 99 % (excepto la paja decebada que presentó un valor del 50 %). Por otra parte, Calsamiglia et al. (1996) determi-naron la degradación de las purinas de ocho raciones diferentes en fermentadores conti-nuos, y señalaron que la degradación microbiana del N púrico de origen alimenticio osci-ló entre 81 y 110 %.
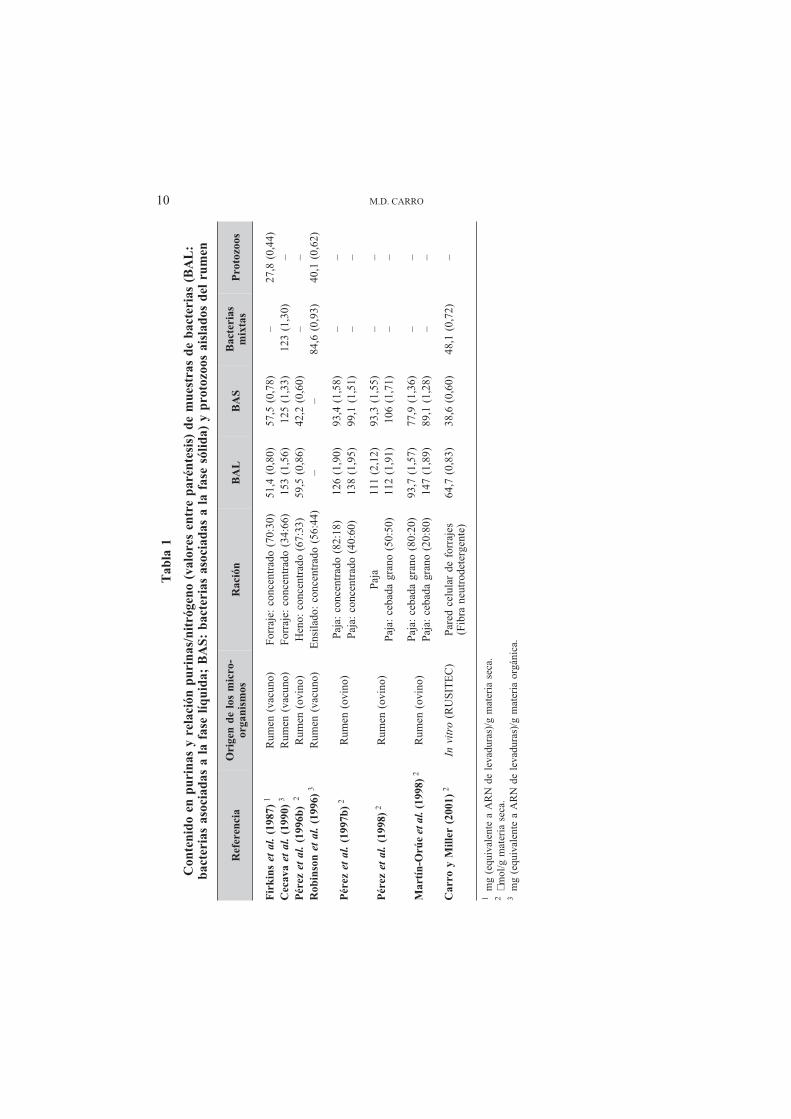
Un segundo inconveniente de las purinas como marcador microbiano es que la rela-ción purinas/N no es la misma en las bacterias y protozoos, e incluso varía en las diferen-tes especies bacterianas. Arambel et al. (1982) analizaron las relaciones ARN/N yADN/N en diecisiete especies bacterianas ruminales y observaron que éstas oscilaban en-tre 8,0 y 102 y entre 0,2 y 39, respectivamente. Cuando las bacterias se agruparon segúnlas características de su pared celular, observaron que los valores medios de la relaciónARN/N eran de 37 y 62 para las bacterias gram-positivas y gram-negativas, respectiva-mente, y de 8,8 y 19 para la relación ADN/N. Estas diferencias entre bacterias se han ob-servado también en la relación purinas/N, tal como puede observarse en la Tabla 1. En la
8 M.D. CARRO

mayoría de los trabajos en los que se han comparado distintos extractos microbianos, seha observado que las BAS presentan un menor contenido en purinas (o en ARN y ADN)y una menor relación purinas/N, tanto en estudios in vivo (Merry y McAllan, 1983; Le-gay-Carmier y Bauchart, 1989; Pérez et al., 1996b, 1997b, 1998a; Martín-Orúe et al.,1998), como in vitro (Carro y Miller, 2001). Estas diferencias en el contenido en purinasparecen deberse a las características intrínsecas de las diferentes especies que integrancada fracción bacteriana y al menor ritmo de crecimiento de las BAS en comparación conlas BAL (Bates et al., 1985; Legay-Carmier et al., 1989).
En la casi totalidad de los estudios en los que se ha determinado la síntesis de proteí-na microbiana utilizando las purinas como marcador, las BAL se han tomado como mues-tra de referencia, lo que provoca una subestimación de la síntesis de proteína microbiana,tal y como se ha observado en los escasos estudios en los que se han comparado los resul-tados obtenidos con BAL y BAS (Pérez et al., 1998a; Carro y Miller, 2001). En el trabajode Carro y Miller (2001) las raciones estaban constituidas por pared celular (fibra neu-tro-detergente) y la cantidad de proteína microbiana sintetizada fue una media de un 38 %mayor cuando se utilizaron las BAS como extracto microbiano de referencia que cuandose utilizaron las BAL. En general, el error cometido en la determinación de la cuantía dela síntesis microbiana dependerá de la proporción de BAS presente en el rumen. Esta pro-porción depende del tipo de ración que reciben los animales, pero puede llegar a ser de un90 % en animales que reciben raciones constituidas únicamente por forrajes (Merry yMcAllan, 1983). Una solución válida para paliar este problema sería obtener extractosbacterianos representativos de todas las bacterias ruminales, que incluyeran tanto las BALcomo las BAS. A pesar de que existen diversos procedimientos para aislar las BAS, sueficacia suele ser pequeña debido a que estas bacterias se encuentran firmemente adheri-das a las partículas de alimento. La mayoría de los métodos comúnmente utilizados paradesligar las BAS de las partículas de alimento permiten liberar entre un 30 y un 60 % delas mismas (Merry y McAllan, 1983; Legay-Carmier y Bauchart, 1989; Martín-Orúe etal., 1998), aunque otros autores (Whitehouse et al., 1994; Ranilla et al., 2001) han utiliza-do procedimientos que permiten desligar aproximadamente un 80 % de las BAS. Sin em-bargo, algunos de estos tratamientos pueden afectar a la integridad de las bacterias y oca-sionar pérdidas celulares que modifican relación purinas/N, tal y como han observadoMartín-Orúe et al. (1998) y Carro y Miller (2001). Por otra parte, incluso cuando se hanaplicado tratamientos que permiten un alto porcentaje de desligamiento de las BAS, la re-cuperación final de éstas en el pellet bacteriano es inferior al 60 % (Ranilla et al., 2001).
Son muy pocos los estudios en los que se ha intentado aislar un extracto bacteriano«representativo». Carro y Miller (2001) aislaron un extracto bacteriano que incluía bacte-rias asociadas a la fase sólida y líquida de la digesta obtenida de un fermentador semicon-tinuo (RUSITEC), y observaron que la relación purinas/N de dicho extracto presentó unvalor intermedio entre los encontrados en las BAL y las BAS (ver Tabla 1). Cecava et al.(1990) observaron que la relación purinas/N de un extracto microbiano «mixto» aisladode terneros fue similar a la encontrada en las BAS aisladas de los mismos animales (verTabla 1).
Por otra parte, los protozoos que pasan al abomaso y duodeno contribuyen también alflujo duodenal de purinas. Estos microorganismos suelen presentar valores para la rela-ción purinas/N en torno al 50 % de los que presentan las bacterias (ver Tabla 1; Ling yButtery, 1978; Firkins et al., 1987; Robinson et al., 1996), por lo que la síntesis de proteí-na microbiana se subestima de nuevo cuando se utilizan extractos bacterianos como
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 9
10 M.D. CARRO
Tab
la1
Con
ten
ido
enp
uri
nas
yre
laci
ónp
uri
nas
/nit
róge
no
(val
ores
entr
ep
arén
tesi
s)d
em
ues
tras
de
bac
teri
as(B
AL
:b
acte
rias
asoc
iad
asa
lafa
selí
qu
ida;
BA
S:
bac
teri
asas
ocia
das
ala
fase
sóli
da)
yp
roto
zoos
aisl
ados
del
rum
en
Ref
eren
cia
Ori
gen
de
los
mic
ro-
orga
nis
mos
Rac
ión
BA
LB
AS
Bac
teri
asm
ixta
sP
roto
zoos
Fir
kin
set
al.
(198
7)1
Rum
en(v
acun
o)F
orra
je:
conc
entr
ado
(70:
30)
51,4
(0,8
0)57
,5(0
,78)
–27
,8(0
,44)
Cec
ava
etal.
(199
0)3
Rum
en(v
acun
o)F
orra
je:
conc
entr
ado
(34:
66)
153
(1,5
6)12
5(1
,33)
123
(1,3
0)–
Pér
ezet
al.
(199
6b)
2R
umen
(ovi
no)
Hen
o:co
ncen
trad
o(6
7:33
)59
,5(0
,86)
42,2
(0,6
0)–
–R
obin
son
etal.
(199
6)3
Rum
en(v
acun
o)E
nsil
ado:
conc
entr
ado
(56:
44)
––
84,6
(0,9
3)40
,1(0
,62)
Pér
ezet
al.
(199
7b)
2R
umen
(ovi
no)
Paj
a:co
ncen
trad
o(8
2:18
)12
6(1
,90)
93,4
(1,5
8)–
–P
aja:
conc
entr
ado
(40:
60)
138
(1,9
5)99
,1(1
,51)
––
Pér
ezet
al.
(199
8)2
Rum
en(o
vino
)P
aja
111
(2,1
2)93
,3(1
,55)
––
Paj
a:ce
bada
gran
o(5
0:50
)11
2(1
,91)
106
(1,7
1)–
–
Mar
tín
-Orú
eet
al.
(199
8)2
Rum
en(o
vino
)P
aja:
ceba
dagr
ano
(80:
20)
93,7
(1,5
7)77
,9(1
,36)
––
Paj
a:ce
bada
gran
o(2
0:80
)14
7(1
,89)
89,1
(1,2
8)–
–
Car
roy
Mil
ler
(200
1)2
Invi
tro
(RU
SIT
EC
)P
ared
celu
lar
defo
rraj
es(F
ibra
neut
rode
terg
ente
)64
,7(0
,83)
38,6
(0,6
0)48
,1(0
,72)
–
1m
g(e
quiv
alen
tea
AR
Nde
leva
dura
s)/g
mat
eria
seca
.2
�mol
/gm
ater
iase
ca.
3m
g(e
quiv
alen
tea
AR
Nde
leva
dura
s)/g
mat
eria
orgá
nica
.

muestra de referencia. Sin embargo, la cuantificación de la proteína protozoaria presentauna gran dificultad, ya que los protozoos suelen sufrir una retención selectiva en el rumendebido a su adhesión a las partículas de alimento (Broderick y Merchen, 1992), y son es-casos los estudios en los que se ha determinado su contribución al flujo de N duodenal(Ling y Buttery, 1978; Robinson et al., 1996).
Otro inconveniente de la utilización de las purinas como marcador microbiano es quela relación purinas/N en las bacterias cambia tras la administración de alimento a los ani-males (Craig et al., 1987; Cecava et al., 1990). Craig et al. (1987) observaron que los va-lores de la relación purinas/N en las bacterias ruminales (BAS y BAL) eran mínimos in-mediatamente antes de la administración de alimento y aumentaban hasta presentar valo-res máximos (entre 33 y 44 % superiores a los registrados anteriormente) entre 7 y 9horas tras la administración de alimento. Sin embargo, este problema puede paliarse reali-zando muestreos seriados del contenido ruminal, de tal forma que se abarquen todas lasfases del ciclo alimenticio (Cecava et al., 1990).
Como se ha comentado anteriormente, una de las principales ventajas de las purinascomo marcador microbiano es que su análisis por el método de Zinn y Owens (1986) essimple, rápido y barato. El método consiste en hidrolizar las purinas mediante su trata-miento con una solución de ácido perclórico, seguido por la precipitación de las mismascon nitrato de plata y su cuantificación mediante espectrofotometría. Sin embargo, Mak-kar y Becker (1999) observaron que la recuperación de purinas de extractos bacterianosruminales que habían sido añadidos a celulosa, almidón y fibra neutro-detergente era sola-mente del 50 % cuando se usaba este método, y propusieron la utilización de la técnicadescrita por Balcells et al. (1992) para obtener una recuperación de las purinas cercana al100 %. Dicha técnica utiliza la cromatografía líquida de alta resolución (HPLC), por loque para llevar a cabo los análisis de purinas se necesita disponer de un equipo de HPLC.Si bien es cierto que no todos los centros de investigación de nuestro país disponen de di-cho equipo, existen laboratorios que realizan los análisis de purinas mediante esta técnicaa un precio asequible (aproximadamente 2.000 pesetas por análisis).
A pesar de todos los inconvenientes anteriormente descritos, las purinas son recomen-dadas como uno de los métodos de elección para determinar la síntesis de proteína micro-biana (Broderick y Merchen, 1992; Brodiscou y Jouany, 1995).
Las purinas que llegan al duodeno pueden ser absorbidas y posteriormente reutiliza-das por el animal o metabolizadas y excretadas en la orina en forma de alantoína y otrosmetabolitos (McAllan, 1982). Debido a ello se ha propuesto que la excreción urinaria dederivados púricos (DP) puede constituir un parámetro válido para estimar el flujo duode-nal de proteína microbiana. A pesar de que los primeros estudios que mostraron la exis-tencia de una estrecha relación entre la excreción urinaria de DP y la concentración deácidos nucleicos en el rumen se llevaron a cabo hace más de treinta años (Topps y Elliot,1965), ha sido en la última década cuando se han realizado numerosos trabajos sobre estetema. Las purinas de los microorganismos ruminales que llegan al duodeno son absorbi-das rápidamente a través de la mucosa intestinal, en forma de nucleósidos o como baseslibres, si bien en el ganado vacuno sufren una cierta degradación a ácido úrico en su pasopor la mucosa (Stangassinger et al., 1995). Las purinas absorbidas se transportan hasta elhígado, donde tiene lugar su utilización y degradación. Tras su metabolismo, las purinasse excretan en la orina en forma de alantoína, ácido úrico, xantina e hipoxantina, aunqueen el ganado vacuno las cantidades de estos dos últimos compuestos son insignificantes(Verbic et al., 1990). Otras vías de excreción son la saliva y la leche, pero cuantitativa-
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 11

mente representan una pequeña proporción (Chen et al., 1990a). La estrecha relaciónexistente entre la excreción urinaria de DP y el flujo duodenal de células microbianas seha puesto de manifiesto en numerosos trabajos (Sibanda et al., 1982; Balcells et al., 1991;Chen et al., 1990b; 1991), y ha permitido el desarrollo de ecuaciones que permiten prede-cir el flujo duodenal de N microbiano a partir de la excreción urinaria de DP en el ganadovacuno (Verbic et al., 1990) y ovino (Chen et al., 1990b; Balcells et al., 1991), aunquehasta el momento no se dispone de ninguna ecuación para el ganado caprino.
La utilización de la excreción urinaria de DP para estimar el flujo duodenal de N mi-crobiano presenta varias ventajas respecto a otros métodos. La ventaja principal es que setrata de una técnica no invasiva, con lo cual los posibles trastornos en el comportamientoalimentario de los animales o en su motilidad digestiva (producidos por la implantaciónde cánulas) quedan descartados. Por otra parte, esta técnica es relativamente fácil de lle-var a cabo en la mayoría de las circunstancias, ya que únicamente se necesita realizar larecogida de la orina de los animales. En algunos estudios (Lebzien et al., 1993; Gieseckeet al., 1994) se ha puesto de manifiesto la relación existente entre la excreción de alantoí-na en leche y el flujo duodenal de N microbiano, hecho que abre nuevas posibilidades aesta técnica. La posibilidad de estimar el flujo duodenal de N microbiano a partir de laconcentración de alantoína en la leche [se ha observado que el ácido úrico excretado en laleche procede en su mayoría del metabolismo de la glándula mamaria (Giesecke et al.,1994)] es especialmente interesante, ya que en las vacas lecheras interesa lograr un altoritmo de síntesis de proteína microbiana debido a la repercusión que tiene la nutriciónproteica no sólo en la producción de leche, sino también en la función reproductora (Fer-guson y Chalupa, 1989). Si bien los resultados obtenidos son sólo aproximados, esta téc-nica resulta muy útil en aquellos casos en los que no se puede predecir la síntesis de pro-teína microbiana porque no se conoce la energía ingerida por los animales (p.e. cuandolos animales reciben forrajes), y no resulta complicado llevarla a cabo, ya que el muestreode la leche durante el ordeño es una práctica habitual (Stangassinger et al., 1995).
La excreción urinaria de DP es una técnica prometedora, pero existen ciertos aspectosque deben ser clarificados para su mejor utilización. Uno de ellos es la cuantificación dela excreción de DP de origen endógeno. A pesar de que diferentes grupos de trabajo(Chen et al., 1990b; Balcells et al., 1991) han determinado valores similares para la ex-creción urinaria endógena de DP en el ganado ovino, no se conoce bien como se ve afec-tada esta excreción cuando los animales reciben raciones de diferente calidad y/o a dife-rentes niveles de ingestión. Chen et al. (1992) señalaron que la excreción urinaria de DPde origen endógeno disminuía a medida que el animal disponía de purinas de origen exó-geno. Sin embargo, en un estudio más reciente, Pérez et al. (1998b) observaron que la ex-creción endógena de DP aumentaba al hacerlo la cantidad de concentrado ingerida por losanimales, y aumentar por ello el flujo duodenal de purinas. Aunque la contribución de laspurinas de origen endógeno a la excreción urinaria de DP es proporcionalmente pequeña(Chen et al., 1992), una cuantificación precisa de su contribución bajo diferentes circuns-tancias alimenticias permitiría obtener estimaciones más exactas del flujo duodenal de Nmicrobiano (Stangassinger et al., 1995).
Otro aspecto conflictivo es que en el cálculo del flujo duodenal de N microbiano apartir de la excreción urinaria de DP se asume que los microorganismos ruminales presen-tan una relación purinas/N constante, ya que ésta no es determinada en cada experimento.Sin embargo, esta relación varía con los factores que se han comentado anteriormente, eincluso se ve afectada por las características de la ración, como son la cantidad de proteí-
12 M.D. CARRO

na de la misma y su relación forraje:concentrado (Clark et al., 1992). Por otra parte, se haobservado que existen diferencias en el metabolismo de las purinas entre las diferentes es-pecies animales, y también entre animales jóvenes y adultos (Stangassinger et al., 1995).La identificación de estas diferencias permitiría desarrollar modelos que pudieran aplicar-se a todo tipo de animales, así como extrapolar los datos de una especie a otra.
Por último, hay que tener en cuenta que la excreción urinaria de DP es un parámetroque refleja posibles cambios en la eficiencia de digestión y absorción de los microorganis-mos, es decir, es un parámetro que refleja la biodisponibilidad del N microbiano, mientrasque el flujo duodenal de N microbiano determinado directamente mediante el uso de mar-cadores es independiente de estos procesos (Stangassinger et al., 1995). Asimismo, hayque considerar que las estimaciones del flujo duodenal de N microbiano obtenidas a partirde la excreción urinaria de DP no son valores exactos (Broderick y Merchen, 1982; Chenet al., 1992; Stangassinger et al., 1995), ya que se obtienen aceptando determinados su-puestos que pueden no ser ciertos en todos los casos (p.e. una relación constante puri-nas/N en los microorganismos ruminales), pero se acepta su validez a efectos comparati-vos. Una posible alternativa para el futuro sería analizar la excreción urinaria de DP en to-dos aquellos experimentos en los que se determine la síntesis de proteína microbianamediante la utilización de otros marcadores, de tal forma que se pudieran llegar a clarifi-car los aspectos mencionados anteriormente. Por otra parte, el análisis de la relación puri-nas/N en los microorganismos ruminales de animales que reciban raciones de diferentecalidad y administradas a diferentes niveles de ingestión permitiría obtener un rango devalores, de tal forma que en trabajos posteriores se dispusiera de los valores más apropia-dos para utilizar en cada caso.
Ácido diaminopimélico (DAPA)
El DAPA es un aminoácido que forma parte de la pared celular de numerosas bacte-rias ruminales, pero que no se encuentra en los protozoos (Ling y Buttery, 1978). Su posi-ble utilización como marcador microbiano se planteó en los años cincuenta y desde enton-ces se ha utilizado en multitud de estudios, a pesar de que presenta numerosos inconve-nientes. El hecho de que el DAPA se encuentre únicamente en la pared celular de lasbacterias hace que la relación DAPA/N en las mismas varíe con aquellas condiciones quealteran el tamaño celular. Las condiciones que favorecen el crecimiento de las bacteriasde mayor tamaño provocan una disminución de la relación DAPA/N, ya que la relaciónpared celular/protoplasma será proporcionalmente menor cuanto mayor sea el tamaño delas bacterias (Broderick y Merchen, 1992). Por el contrario, las bacterias de menor tama-ño presentan una proporción DAPA/N comparativamente mayor. Además, en algunasbacterias como los cocos gram-positivos no existe DAPA en su pared celular (Merry yMcAllan, 1983). Debido a estas diferencias entre bacterias no resulta extraño que en va-rios estudios (Merry y McAllan, 1983; Olubobokun y Craig, 1990) se haya observado queel contenido en DAPA y la relación DAPA/N es significativamente menor en las BASque en las BAL (ver Tabla 2).
Por otra parte, en algunos estudios (Ling y Buttery, 1978; Olubobokum y Craig,1990) se ha observado que la cantidad de DAPA en los microorganismos ruminales va-ría en función del tiempo transcurrido desde la ingestión de alimento. Dufva et al. (1982)señalaron que la relación DAPA/N era menor en las BAL aisladas de animales que
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 13

14 M.D. CARRO
Tab
la2
Con
ten
ido
enác
ido
dia
min
opim
élic
o(D
AP
A)
yre
laci
ones
(val
ores
entr
ep
arén
tesi
s)D
AP
A/N
yác
ido
amin
oeti
lfos
fón
ico-
N(A
EP
A-N
)/N
de
mu
estr
asd
eb
acte
rias
(BA
L:
bac
teri
asas
ocia
das
ala
fase
líq
uid
a;B
AS
:b
acte
rias
asoc
iad
asa
lafa
sesó
lid
a)y
pro
tozo
osai
slad
osd
elru
men
Mar
cad
orm
icro
bia
no
yre
fe-
ren
cia
Ori
gen
de
los
mic
roor
gan
is-
mos
Rac
ión
BA
LB
AS
Bac
teri
asm
ixta
sP
roto
zoos
Áci
do
dia
mim
opim
élic
o
Mer
ryy
McA
llan
(198
3)1
Rum
en(v
acun
o)C
ebad
a:pa
jace
bada
:ur
ea(4
5:48
:3)
3,81
(0,0
47)
2,20
(0,0
31)
––
Rha
nem
ay
The
urer
(198
6)1
Rum
en(v
acun
o)S
orgo
gran
o:m
elaz
as:
forr
aje
(84:
6:10
)–
–3,
28(0
,049
)1,
02(0
,017
)L
egay
-Car
mie
ryB
auch
art(
1989
)1R
umen
(vac
uno)
Hen
o:co
ncen
trad
o(5
0:50
)2,
23(0
,034
)1,
39(0
,024
)–
–O
lubo
boku
my
Cra
ig(1
990)
1R
umen
(vac
uno)
Hen
o:co
ncen
trad
o(9
4:6)
1,90
(0,0
21)
1,58
(0,0
22)
––
Mar
tin
etal.
(199
4)2
Rum
en(v
acun
o)H
eno:
ceba
da(6
5:35
)2,
52(0
,028
)1,
10(0
,013
)–
–R
obin
son
etal.
(199
6)2
Rum
en(v
acun
o)E
nsil
ado:
conc
entr
ado
(56:
44)
7,05
(0,0
78)
––
2,03
(0,0
31)
Áci
do
dia
min
oeti
lfos
fón
ico
Lin
gy
But
tery
(197
8)R
umen
(ovi
no)
Ceb
ada
gran
o:pa
ja:
mel
azas
:to
rta
deso
ja(5
8:16
:10:
16)
––
–(0,
64)
–(2,
26)
Ceb
ada
gran
o:pa
ja:
mel
azas
:ur
ea(7
2:16
:10:
2)–
––(
0,22
)–(
1,72
)
1m
gde
DA
PA
/gm
ater
iase
ca.
2m
gde
DA
PA
/gm
ater
iaor
gáni
ca.

recibían raciones forrajeras que en las aisladas de animales que recibían raciones con altasproporciones de concentrado, si bien Robinson et al. (1996) no observaron efectos signifi-cativos de la ración en la relación DAPA/N de bacterias y protozoos aislados de novillosque recibían raciones compuestas por ensilado y concentrado.
Otro problema grave es la ausencia de DAPA en los protozoos ruminales (Ling yButtery, 1978). A pesar de ello, las preparaciones de protozoos aislados del rumen presen-tan ciertas cantidades de DAPA, bien por su contaminación con bacterias durante el aisla-miento o, principalmente, debido a la presencia de restos de bacterias no digeridas en elinterior de los mismos (Coleman y Sandford, 1979). Sin embargo, la relación DAPA/N enlos protozoos es muy inferior (entre 3 y 8 veces menor) a la de las bacterias (ver Tabla 2;Czerkawski, 1974; Rahnema y Theurer, 1986; Robinson et al., 1986), por lo que no consi-derar este hecho produce una subestimación de la producción de N microbiano. Estos he-chos hacen que el DAPA no cumpla la premisa de encontrarse distribuido de forma uni-forme en todos los microorganismos ruminales.
La lisis microbiana que tiene lugar en el rumen y abomaso provoca una pérdida delcontenido celular, de tal forma que muchos restos de paredes celulares de bacterias pa-san al duodeno (Gómez et al., 1991). Dado que el DAPA solamente se encuentra en lapared celular de las bacterias, este hecho hace que la digesta duodenal presente una rela-ción DAPA/N anormalmente alta, y por ello se sobreestima la producción de N micro-biano. A lo largo de varios experimentos realizados con ovejas, Masson et al. (1991)observaron que del total de DAPA presente en la digesta duodenal sólo un 69 % forma-ba parte de células bacterianas intactas cuando las muestras se obtenían una hora antesde la ingestión de alimento, y esta cifra se reducía al 29 % si las muestras se obteníanvarias horas después de la ingestión. Estos datos indican que, a menudo, más de la mi-tad del DAPA que llega al duodeno puede encontrarse en forma libre o en restos de pa-redes celulares, de tal forma que en algunos estudios se han obtenido proporciones de Nmicrobiano en la digesta duodenal superiores al 100 % (Nikolic y Jovanovic, 1973). Enotros estudios se ha observado que el uso del DAPA provocó una sobreestimación de laproducción de N microbiano en comparación con otros marcadores, como el 35S (Sid-dons et al., 1982; Whitelaw et al., 1984), el 15N (Siddons et al., 1982; Schönhusen et al.,1995), el perfil de aminoácidos (Siddons et al., 1982; Schönhusen et al., 1995) y las pu-rinas (Illg y Stern, 1994). Por el contrario, Ling y Buttery (1978) y Walker y Nader(1975) obtuvieron menores valores de síntesis de proteína microbiana con DAPA quecon 35S. Parte de estas diferencias pueden deberse al método analítico usado para deter-minar las concentraciones de DAPA. Este análisis (Czerkawski, 1974) utiliza técnicasde cromatografía de intercambio iónico y de colorimetría, y aunque es el método utiliza-do en la mayoría de los laboratorios, muchos de ellos han introducido modificaciones enel mismo. En cualquier caso, el análisis del DAPA implica numerosos pasos y es unatécnica larga y laboriosa.
Algunos autores (Czerkawski, 1974; Rahnema y Theurer, 1986) han encontradoDAPA en algunos alimentos de uso habitual en las raciones de los animales rumiantes.Rahnema y Theurer (1986) señalaron que los valores de la relación DAPA/N en diferen-tes alimentos oscilaban entre 18 y 40 % de los encontrados en las bacterias ruminales. Sinembargo, otros estudios (Czerkawski, 1974) han determinado valores de DAPA muchomenores (inferiores a la mitad) en alimentos de características similares, y se han atribui-do los altos valores encontrados por Rahnema y Theurer (1986) a imprecisiones del méto-do analítico utilizado por estos autores (Broderick y Merchen, 1992).
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 15

Debido a los inconvenientes comentados, principalmente al hecho de que gran partedel DAPA presente en la digesta duodenal no está asociado con células bacterianas intac-tas, actualmente no se recomienda su uso como marcador microbiano (Masson et al.,1991; Broderick y Merchen, 1992; Illg y Stern, 1994), a pesar de que ha sido muy em-pleado en las últimas décadas.
D-alanina y ácido aminoetilfosfónico (AEPA)
La D-alanina es un aminoácido que forma parte de la pared celular de numerosas bac-terias ruminales, pero que no se encuentra en los protozoos (Ling y Buttery, 1978) ni enlos alimentos utilizados en las raciones de los rumiantes (Garrett et al., 1986). Sin embar-go, la D-alanina ha sido utilizada en muy pocos estudios, y en alguno de ellos se han obte-nido proporciones de N microbiano en la digesta duodenal superiores al 100 % (Quigley ySchwat, 1988), lo que teóricamente resulta imposible. Por estas razones la D-alanina nosuele utilizarse como marcador microbiano.
Horiguchi y Kandtsu (1959) aislaron un aminoácido, el ácido aminoetilfosfónico(AEPA), que formaba parte de los protozoos ciliados del rumen, pero que no estaba pre-sente en la mayoría de los alimentos usados comúnmente en las raciones de los animales,por lo que se propuso su utilización como marcador protozoario. Sin embargo, en estudiosmás recientes (Ling y Buttery, 1978; Horigani y Horiguchi, 1990) se ha observado la pre-sencia de AEPA en las bacterias ruminales (ver Tabla 2) y en algunos alimentos. Horiganiy Horiguchi (1990) señalan que las bacterias ruminales pueden incorporar AEPA y lasplantas pueden tomarlo de los microorganismos existentes en el suelo. Debido a estos he-chos se desaconseja el uso de AEPA como marcador de la síntesis microbiana de origenprotozoario (Horigani y Horiguchi, 1990).
Perfiles de aminoácidos
Este método, propuesto por Evans et al. (1975), se basa en estimar la cantidad de Nmicrobiano, N de origen alimenticio y N de origen endógeno a partir de las diferenciasexistentes en el contenido en los distintos aminoácidos de estas tres fracciones. Esta técni-ca acepta varias premisas que no son totalmente ciertas. En primer lugar, se asume que laproteína endógena presenta una composición aminoacídica constante, y por ello se sueletomar como patrón el pepsinógeno bovino, sin considerar la composición de otras proteí-nas que pueden formar parte de la misma (Siddons et al., 1982). Por otra parte, se admiteque la proteína de origen alimenticio no degradada en el rumen tiene la misma composi-ción en aminoácidos que la proteína del alimento, sin considerar que la composición ami-noacídica de la fracción proteica rápidamente degradable es diferente de la de aquellasfracciones que se degradan más lentamente (Nikolic y Jovanovic, 1972; Siddons et al.,1982). En cuanto a los microorganismos ruminales, se suele aislar una muestra de losmismos o se asume una composición constante (Stern y Hoover, 1979). A pesar de que enalgunos trabajos se ha señalado que los microorganismos ruminales presentan una compo-sición aminoacídica constante, Clark et al. (1992) revisaron los principales estudios en losque se determinó la composición aminoacídica de extractos bacterianos (441 muestras co-rrespondientes a animales que ingirieron 61 raciones diferentes) y observaron una amplia
16 M.D. CARRO

variabilidad en el contenido en todos los aminoácidos, con coeficientes de variación queoscilaron entre 4,8 y 25,6 % (valores para el ácido aspártico y la metionina, respectiva-mente). Además, la composición aminoacídica de los microorganismos ruminales cambiatras la administración de alimento (Clark et al., 1992), por lo que el aislamiento de unafracción microbiana representativa resulta especialmente difícil.
Evans et al. (1975) compararon los resultados de este método con los obtenidos conDAPA y observaron que los valores eran similares, pero inferiores a los obtenidos utili-zando ARN como marcador. De forma similar, Siddons et al. (1982) y Schönhusen et al.(1995) obtuvieron valores de síntesis de proteína microbiana mucho menores con los per-files de aminoácidos que con otros marcadores (DAPA, ARN, 15N y 35S). Debido a que laspremisas en las que se basa esta técnica no son válidas y a los resultados erráticos obteni-dos en varios estudios, este método no suele utilizarse para cuantificar la síntesis de pro-teína microbiana.
ATP
Forsberg y Lam (1977) utilizaron el ATP como marcador microbiano para cuantificarlas proporciones de BAL y BAS en el rumen, y para ello aceptaron las siguientes premi-sas: a) que el ATP está presente en todas las células vivas, pero no forma parte de las cé-lulas muertas; b) que la concentración de ATP es similar en todos los microorganismosruminales y c) que la extracción y posterior análisis de la concentración de ATP en las cé-lulas son relativamente simples y baratos (Stern y Hoover, 1979). Sin embargo, estos mis-mos autores observaron que existían diferencias en la concentración de ATP entre los di-ferentes microorganismos ruminales, y que, dependiendo del método utilizado, había unagran variabilidad en la eficiencia de la extracción de ATP de las células. Posteriormente,Wallace y West (1982) también observaron diferencias en el contenido en ATP entre bac-terias y protozoos. Estos autores señalan que la concentración de ATP refleja el nivel deactividad de los microorganismos ruminales más que la cantidad total de los mismos, porlo que no debe ser usado como marcador microbiano.
MARCADORES MICROBIANOS EXTERNOS
Los marcadores microbianos externos deben administrarse a los animales para quesean incorporados por los microorganismos ruminales y formen parte de sus estructuras.El procedimiento más empleado consiste en realizar una infusión continua del marcadoren el rumen de los animales durante el período experimental hasta que se alcanzan condi-ciones de equilibrio («steady state»). Los principales marcadores externos son 15N, 35S y32P, aunque en algunos estudios se ha determinado la incorporación de compuestos marca-dos con 3H y 14C (Chikunya y Miller, 1997; Lambert et al., 1998).
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 17

15N
El 15N se administra a los animales experimentales en forma de sales amónicas(15NH4Cl, (15NH4)2SO4) o de urea. Estas sustancias se disuelven fácilmente en agua, y trasun período de infusión de 3-4 días se consigue un enriquecimiento constante de los mi-croorganismos ruminales (Firkins et al., 1987; Beckers et al., 1995). El 15N es uno de losmarcadores externos más utilizados, tanto en estudios in vivo (Carro et al., 1992a; Leb-zien et al., 1993; Pérez et al., 1996a, 1996b) como in vitro (Carro et al., 1992b; Blümmelet al., 1997; Carro y Miller, 1999b). Este hecho es debido a algunas de las ventajas quepresenta: 1) es un isótopo no radioactivo, por lo que puede ser utilizado en cualquier oca-sión sin riesgos de contaminación medioambiental; 2) los alimentos de los animales nosuelen presentar enriquecimientos en 15N superiores a la abundancia natural (0,3663), y lainfusión de 15N únicamente enriquece a los microorganismos; 3) el 15N infundido se incor-pora a todos los microorganismos, ya que las bacterias incorporan el 15NH3 y los proto-zoos lo incorporan indirectamente cuando engullen y digieren a las bacterias, aunque elgrado de enriquecimiento de las diferentes fracciones de microorganismos suele ser dife-rente.
Otra ventaja del 15N es que marca directamente el N microbiano, es decir, el materialcuya cuantía se desea determinar. Sin embargo, lo que realmente se determina con estemétodo es la incorporación de 15NH3 por los microorganismos, y no se tiene en cuenta laincorporación directa de aminoácidos y péptidos. Este hecho cobra una especial importan-cia si se tiene en cuenta que las diferentes especies bacterianas ruminales difieren en lautilización de amoniaco y N aminoacídico para su síntesis proteica (Russell et al., 1992).En varios estudios (Komisarczuk et al., 1987; Carro y Miller, 1999b; Ranilla et al., 2000)se ha observado que las BAS incorporan una menor proporción de amoníaco que lasBAL, por lo que resulta lógico que presenten habitualmente un menor enriquecimiento en15N (ver Tabla 3). Este hecho se ha observado tanto en condiciones in vivo (Martin et al.,1994; Beckers et al., 1995; Pérez et al., 1996b), como in vitro (Komisarczuk et al., 1987;Carro y Miller, 1999b; Ranilla et al., 2000). Por ello, cuando se utilizan las BAL comoextracto microbiano de referencia se subestima la síntesis de proteína microbiana. Carro yMiller (1998) observaron que la síntesis microbiana en fermentadores semicontinuos(RUSITEC), al que se administraban cuatro raciones diferentes, era una media de un22 % menor cuando se usaban las BAL como extracto de referencia que cuando se usabanlas BAS. Tal y como se ha comentado para otros marcadores microbianos, este problemapodría solucionarse mediante el aislamiento de una fracción microbiana «representativa».
La precisión y exactitud del análisis de 15N por espectrometría de masas son excelen-tes (se pueden detectar enriquecimientos del 0,001 % por encima del enriquecimiento na-tural), pero la preparación de las muestras para el análisis es laboriosa, ya que a menudoes necesario un procesado previo de las mismas. Este procesado suele consistir en un aná-lisis kjeldahl, seguido de una destilación y evaporación del destilado. En caso de ser nece-sario este procesado, el manejo de las muestras debe ser muy cuidadoso para evitar lacontaminación de las mismas durante la destilación (Firkins et al., 1992). Por otra parte,los espectrómetros de masas son equipos muy caros y presentan un manejo complicado,por lo que no están al alcance de la mayoría de los laboratorios, y esto representa uno delos principales puntos limitantes de esta técnica. Sin embargo, en nuestro país existen cen-tros que realizan análisis de 15N a un precio aproximado de 2.500 pesetas por muestra.Una posible alternativa es el análisis de 15N mediante espectrometría de emisión, técnica
18 M.D. CARRO

Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 19T
abla
3
Val
ores
de
lare
laci
ón15
N/N
(%)
y35
S/N
enm
ues
tras
de
bac
teri
as(B
AL
:b
acte
rias
asoc
iad
asa
lafa
selí
qu
ida;
BA
S:
bac
teri
asas
ocia
das
ala
fase
sóli
da)
yp
roto
zoos
aisl
ados
del
rum
en
Mar
cad
orm
icro
bia
no
yre
fere
nci
a
Ori
gen
de
los
mi-
cro-
orga
nis
mos
Rac
ión
BA
LB
AS
Bac
teri
asm
ixta
sP
roto
zoos
15N F
irki
nset
al.
(198
7)R
umen
(vac
uno)
For
raje
:co
ncen
trad
o(7
0:30
)0,
748
0,74
9–
0,63
1
Kom
isar
czuk
etal.
(198
7)1
Invi
tro
(RU
SIT
EC
)P
aja:
pulp
ade
rem
olac
ha:
man
ioca
(50:
29:2
1)0,
932
0,61
1–
–
Mar
tin
etal.
(199
4)1
Rum
en(v
acun
o)H
eno
0,52
50,
479
–0,
456
Hen
o:ce
bada
gran
o(6
5:35
)0,
536
0,47
6–
0,46
4
Pér
ezet
al.
(199
6b)
1R
umen
(ovi
no)
Hen
o:co
ncen
trad
o(6
7:33
)0,
563
0,52
1–
–
Pér
ezet
al.
(199
7b)
Rum
en(o
vino
)P
aja
ceba
da:
conc
entr
ado
(40:
60)
0,48
00,
478
––
Mar
tín-O
rúe
etal.
(199
8)1
Rum
en(o
vino
)P
aja:
ceba
dagr
ano
(80:
20)
0,64
90,
629
––
Paj
a:ce
bada
gran
o(2
0:80
)0,
611
0,53
8–
–
Chi
kuny
ay
Mill
er(1
999)
1R
umen
(ovi
no)
Paj
a:pu
lpa
dere
mol
acha
(70:
30)
0,42
30,
420
––
Car
roy
Mil
ler
(199
9b)
1In
vitr
o(R
US
ITE
C)
Par
edce
lula
rde
forr
ajes
(Fib
rane
utro
-det
erge
nte)
0,66
80,
572
0,60
0–
Ran
illa
etal.
(200
0)1
Invi
tro
(bot
ella
sde
cult
ivo)
Par
edce
lula
rde
forr
ajes
(Fib
rane
utro
-det
erge
nte)
0,75
10,
626
0,70
2–
35S M
erry
yM
cAll
an(1
983)
Rum
en(v
acun
o)C
ebad
a:pa
jace
bada
:ur
ea(4
5:48
:3)
2,11
2,04
––
Chi
kuny
ay
Mil
ler
(199
9)R
umen
(ovi
no)
Paj
a:pu
lpa
dere
mol
acha
(70:
30)
3,16
3,63
––
1L
asdi
fere
ncia
sen
tre
las
BA
Ly
las
BA
Sen
lare
laci
ón15
N/N
son
esta
díst
icam
ente
sign
ific
ativ
as(P
<0,
05).

que presenta una menor precisión, pero que requiere un equipo menos costoso y más fácilde manejar (Proksch, 1972). Otro inconveniente del 15N es el alto precio del isótopo, elcual varía en función del enriquecimiento de la fuente. Por ejemplo, 1 g de 15NH4 Cl (Sig-ma-Alldrich Química, S.A.) con un enriquecimiento del 98 y del 10 % cuesta aproxima-damente 40.000 y 1.800 pesetas, respectivamente. La cantidad mínima necesaria paramarcar los microorganismos ruminales suele ser de 80 mg de 15N por cada 100 g de N enla ración, por lo que el uso del 15N en estudios in vivo se ve limitado en muchos casos porrazones económicas. Sin embargo, estas razones no limitan su uso en estudios in vitro, enlos que la cantidad necesaria es mucho menor (1 g de 15NH4 Cl con un enriquecimientodel 98 % permite realizar varios experimentos in vitro, dependiendo del número de trata-mientos y de réplicas). En cualquier caso, el 15N es uno de los métodos más seguros y fia-bles, y es recomendado como uno de los métodos de elección para determinar la síntesisde proteína microbiana (Broderick y Merchen, 1992).
Los resultados obtenidos en los diferentes estudios en los que se han comparado el 15Ny las purinas son contradictorios. Pérez et al. (1996a) observaron que los valores de síntesismicrobiana en ovejas que recibían heno de alfalfa suplementado con diferentes cantidadesde cebada (0, 220, 400 y 550 g/día) eran entre un 14 y un 22 % menores cuando se utiliza-ban las purinas como marcador que cuando se utilizaba 15N. De acuerdo con estos resulta-dos, Calsamiglia et al. (1996) observaron que las purinas subestimaban la síntesis microbia-na entre un 1 y un 18 %, en comparación con el 15N, en fermentadores continuos que reci-bían una ración basal suplementada con diferentes concentrados proteicos. Por el contrario,Carro y Miller (1999a) obtuvieron valores entre un 15 y un 17 % mayores con purinas quecon 15N al determinar la síntesis microbiana en un fermentador semicontinuo al que se leadministraba una ración basal con un contenido mínimo de purinas (pared celular suple-mentada con cuatro fuentes nitrogenadas diferentes). De forma similar, Tejido et al. (2001)observaron que las purinas sobreestimaban la síntesis microbiana entre un 19 y un 47 %comparadas con el 15N cuando ambos marcadores se utilizaron para cuantificar el creci-miento microbiano in vitro sobre sustratos que no contenían purinas (almidón y celulosa).Carro y Miller (1999a) y Tejido et al. (2001) atribuyeron los mayores valores obtenidos conlas purinas a una pérdida de purinas en el proceso de aislamiento de los extractos bacteria-nos, tal y como ya ha sido comentado anteriormente. Otros autores (Cecava et al., 1991)han observado también una sobreestimación de la sínteis microbiana con las bases púricasen estudios in vivo, y la han atribuido al efecto de las purinas del alimento que no son de-gradadas en el rumen. A pesar de las diferencias obtenidas en los valores absolutos de sínte-sis microbiana en los diferentes estudios, en los trabajos de Pérez et al. (1996a) y Carro yMiller (1999a) se observó una correlación significativa entre los valores obtenidos paracada ración con los dos marcadores, si bien esta relación no fue significativa en los trabajosde Calsamiglia et al. (1996) y Tejido et al. (2001).
35S
De forma similar a la descrita para el 15N, el 35S se infunde en el rumen en forma desales (principalmente Na2
35SO4) y es incorporado a la proteína de los microorganismos através de la síntesis de novo de aminoácidos azufrados (cistina y metionina), aunque tam-bién puede incorporarse a otros compuestos azufrados como la coenzima A (Broderick yMerchen, 1992). Los protozoos son marcados indirectamente cuando engullen y digieren
20 M.D. CARRO

a las bacterias ruminales. El método requiere el aislamiento de una fracción microbianarepresentativa y la determinación de la relación 35S/N en la misma y en la digesta duode-nal. En los primeros estudios realizados era necesario determinar la actividad específicadel 35S en la cistina, la metionina o en el total de aminoácidos azufrados (Beever et al.,1974), lo que complicaba la parte analítica de la determinación de la síntesis microbiana.Sin embargo, Mathers y Miller (1980) propusieron una modificación del método que re-duce el número de análisis y aumenta su precisión (Siddons et al., 1982), y que ha sidousada en numerosos estudios (Kennedy et al., 1984; Chikunya y Miller, 1998, 1999).
Una limitación del 35S como marcador microbiano es el error introducido debido aque una cantidad considerable de S microbiano puede proceder de la incorporación direc-ta de péptidos y/o aminoácidos azufrados a la proteína microbiana. Por otra parte, el 35S esun isótopo radiactivo, aunque su radiación � es débil y su vida media es relativamentecorta (noventa días). Sin embargo, la utilización de este isótopo exige disponer de instala-ciones y laboratorios aptos para trabajar con material radiactivo, así como de personalcualificado para ello.
Kennedy et al. (1984) realizaron un experimento para comparar el 15N y el 35S comomarcadores microbianos y observaron que el 15N produjo valores un 24 % menores que el35S. Por el contrario, Siddons et al. (1982) señalaron que la síntesis de proteína microbia-na en ovejas alimentadas con ensilado o con heno de hierba fue hasta un 25 % menor conel 35S que con el 15N. De forma similar, Chikunya y Miller (1999) observaron que el 35Sprodujo valores un 12 % inferiores a los obtenidos con el 15N cuando ambos se utilizaronpara determinar la síntesis de proteína microbiana en ovejas alimentadas con una racióncompuesta por paja y pulpa de remolacha (70:30). Dado que las diferencias obtenidas enlos distintos estudios comparativos no son consistentes, se piensa que las condiciones ali-menticias pueden afectar de forma diferente a las estimaciones de la síntesis de proteínamicrobiana realizadas con estos dos marcadores (Siddons et al., 1982).
32P
Bucholtz y Bergen (1973) observaron que la captación de fósforo y su incorporacióna los fosfolípidos microbianos presentaban una estrecha correlación con la síntesis de pro-teína microbiana en el rumen, por lo que propusieron el uso de 32P como marcador. Poste-riormente, Smith et al. (1978) propusieron una técnica basada en la incorporación de 32P alos ácidos nucleicos microbianos. Estos autores observaron que los valores obtenidos con32P eran aproximadamente un 85 % de los estimados utilizando ARN como marcador, yatribuyeron esta diferencia a que el ARN sobreestimaba la síntesis microbiana debido a lapresencia de pequeñas cantidades de ARN de origen alimenticio. A pesar de estos resulta-dos, la alta energía de este isótopo hace que su manejo sea peligroso y por ello se ha utili-zado mayoritariamente en estudios in vitro (Van Nevel y Demeyer, 1977; Merry et al.,1990). Por otra parte, su vida media es corta (catorce días) y esto hace que se disponga deun tiempo limitado para analizar las muestras de digesta y extractos microbianos.
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 21

CONCLUSIONES
En el momento actual los dos marcadores microbianos más utilizados son las purinasy el 15N, y ambos han sido propuestos como marcadores de elección por varios autores(Broderick y Merchen, 1982; Broudiscou y Jouany, 1995). A pesar de que son pocos losestudios en los que se han comparado estos dos marcadores, en algunos de ellos se ha ob-servado una correlación significativa entre los resultados obtenidos por los dos métodos.Las purinas presentan la ventaja de ser constituyentes de los microorganismos, hecho queevita la necesidad de realizar infusiones en el rumen para administrar el marcador, peroun inconveniente es que también forman parte de los alimentos que reciben los animales.El 15N presenta principalmente problemas económicos, dado el alto precio del isótopo yde la analítica que conlleva la técnica. Un problema grave que presentan ambos marcado-res es el aislamiento de una fracción bacteriana representativa, ya que las relaciones puri-nas/N y 15N/N difieren en las diferentes poblaciones microbianas (BAS, BAL y proto-zoos).
En la medida de lo posible sería recomendable la utilización conjunta de purinas y15N como marcadores en los estudios en los que se determine la síntesis de proteína mi-crobiana. Además, sería deseable recoger la orina producida por los animales y analizarsu concentración en DP, lo que probablemente permitiría validar de forma definitiva laexcreción urinaria de DP como indicador del flujo duodenal de N microbiano.
AGRADECIMIENTOS
Este trabajo forma parte de los Proyectos LE 29/98 y LE 38/01 financiados por la Junta de Castilla y León.Deseo manifestar mi agradecimiento al Dr. Balcells por las interesantes conversaciones que hemos mantenidosobre la utilización de las bases púricas y el 15N como marcadores microbianos.
SUMMARY
Determination of microbial protein synthesis in the rumen: a comparison ofmicrobial markers (Review)
Measurement of microbial protein synthesis and protein degradation in the rumen is critical in all the pro-tein evaluation systems for ruminants. Microbial protein synthesis is determined by means of microbial markers,which are classified as internal (inherently present in rumen microorganisms) and external (compounds added tothe rumen to label the microorganisms) markers. However, no marker has proven to be completely satisfactory.This review examines the advantages and disadvantages of the internal (nucleic acids, purine bases,diaminopimelic acid, D-alanine, aminoethyl-phosphonic acid, amino acid profiles and ATP) and external (15N,35S and 32P) microbial markers and presents the results obtained from studies in which several microbial markerswere compared.
KEY WORDS: Microbial proteinRumenMicrobial markersPurines15N
22 M.D. CARRO

REFERENCIAS BIBLIOGRÁFICAS
AFRC, 1992. Nutritive Requirements of Ruminant Animals: Protein. AFRC Technical Committee on Responsesto Nutrients. Report No. 9. Nutr. Abstr. Rev. (Series B), 62, 787-835.
ARAMBEL M.J., BARTLEY E.E., DUFVA G.S., NAGARAJA T.G., DAYTON A.D., 1982. Effect of diet onamino and nucleic acids of rumen bacteria and protozoa. J. Dairy Sci., 65, 2095-2101.
BALCELLS J., GUADA J.A., CASTRILLO C., GASA J., 1991. Urinary excretion of allantoin and allantionprecursors by sheep after different rates of purine infusion into the doudenum. J. Agric. Sci., Camb., 116,309-317.
BALCELLS J., GUADA J.A., PEIRÓ J.M., PARKER D.S., 1992. Simultaneous determination of allantoin andoxipurines in biological fluids by high-performance liquid chromatography. J. Chromat., 575, 153-157.
BATES D.B., GILLETT J.A., BARAO S.A., BERGER W.G., 1985. The effect of specific growth rate and stageof growth on nucleic acid-protein values of pure cultures and mixed ruminal bacteria. J. Anim. Sci., 61,713-724.
BECKERS Y., THÉWIS A., MAUDOUX B., FRANCOIS E., 1995. Studies on the in situ nitrogen degradabilitycorrected for bacterial contamination of concentrate feeds in steers. J. Anim. Sci., 73, 220-227.
BEEVER D.E., HARRISON D.G., THOMSON D.J., CAMMELL S.B., OSBOURN D.F., 1974. A method forthe estimation of dietary and microbial protein in duodenal digesta of ruminants. Br. J. Nutr., 32, 99-112.
BLÜMMEL M., STEINGAb H., BECKER K., 1997. The relationship between in vitro gas production, in vitromicrobial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed in-take of roughages. Br. J. Nutr., 77, 911-921.
BRODERICK G.A., MERCHEN N.R., 1992. Markers for quantifying microbial protein synthesis in the rumen.J. Dairy Sci., 75, 2618-2632.
BROUDISCOU L., JOUANY J.P., 1995. Reassessing the manipulation of protein synthesis by rumen microbes.Reprod. Nutr. Develop., 35, 517-535.
BUCHOLTZ H.F., BERGEN W.G., 1973. Microbial phospholipid synthesis as a marker for microbial proteinsynthesis in the rumen. Appl. Microbiol., 25, 504-509.
CALSAMIGLIA S., STERN M.D., FIRKINS J.L., 1996. Comparison of nitrogen-15 and purines as microbialmarkers in continuous culture. J. Anim. Sci., 74, 1375-1381.
CARRO M.D., LEBZIEN P., ROHR K., 1992a. Effects of yeast culture on rumen fermentation, digestibility andduodenal flow in dairy cows fed a silage based diet. Livest. Prod. Sci., 32, 219-229.
CARRO M.D., LEBZIEN P., ROHR K., 1992b. Influence of yeast culture on the «in vitro» fermentation (Rusi-tec) of diets containing variable portions of concentrates. Anim. Feed Sci. Technol., 37, 209-220.
CARRO M.D., MILLER E.L., 1998. Effect of microbial isolates on microbial yield estimation in RUSITECsystem. In: In vitro Techniques for Measuring Nutrient Supply to Ruminants. ed. BSAS, Midlothian, pp.306-308, Reino Unido.
CARRO M.D., MILLER E.L., 1999a. Comparison of 15N and purines as microbial markers in a RUSITECsystem. In: Proceedings of the BSAS Winter Meeting 1999, BSAS, pp. 214.
CARRO M.D., MILLER E.L., 1999b. Effect of supplementing a fibre basal diet with different nitrogen formson ruminal fermentation and microbial growth in an in vitro semi-continuous culture system (RUSITEC).Br. J. Nutr., 82, 149-157.
CARRO M.D., MILLER E.L., 2001. Comparison of bacterial isolates and of microbial markers (15N and purinebases) for the determination of microbial protein synthesis in semicontinuous fermenters. J. Anim. Sci. (Enfase de evaluación).
CECAVA M.J., MERCHEN N.R., GAY L.C., BERGER L.L., 1990. Composition of ruminal bacteria harvestedfrom steers as influenced by dietary energy level, feeding frequency, and isolation techniques. J. Dairy Sci.73, 2480-2488.
CECAVA, M.J., MERCHEN, N.R., BERGER, L.L., MACKIE, R.I. AND FAHEY JR., G.C., 1991. Effects ofdietary energy level and protein source on nutrient digestion and ruminal nitrogen metabolism in steers. J.Anim. Sci. 69, 2230-2243.
CHEN X.B., HOVELL F.D.DEB., ØRSKOV E.R., 1990a. Excretion of purine derivatives in ruminants: recy-cling of allantion into the rumen via saliva and its fate in the gut. Br. J. Nutr., 63, 197-205.
CHEN X.B., HOVELL F.D.DEB., ØRSKOV E.R., BROWN D.S., 1990b. Excretion of purine derivatives by ru-minants: effect of exogenous nucleic acid supply on purine derivative excretion by sheep. Br. J. Nutr., 63,131-142.
CHEN X.B., ØRSKOV E.R., HOVELL F.D.DEB., 1991. The use of intragastric infusion in studies on excretionof purine derivatives as a measure of microbial protein supply in ruminants. In: Protein Metabolism andNutrition, Vol. 2. Eggum, B.O., Boisen, S., Børsting, C., Danfaer, A. and Hvelplund, R., eds. NationalInstitute of Animal Science, Foulum, pp. 67-70. Dinamarca.
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 23

CHEN X.B., CHEN Y.K., FRANKLIN M.F., ØRSKOV E.R., SHAND W.J., 1992. The effect of feed intakeand body weight on purine derivative excretion and microbial protein supply in sheep. J. Anim. Sci., 70,1534-1542.
CLARK J.H., KLUSMEYER T.H., CAMERON M.R., 1992. Microbial protein synthesis and flows of nitrogenfractions to the doudenum of dairy cows. J. Dairy Sci., 75, 2304-2323.
CHIKUNYA S., MILLER E.L., 1998. The influence of supplementing a rapidly degraded fibre basal diet withdifferent forms of nitrogen on microbial activity in continuous fermenters. In: Proceedings of the BSASWinter Meeting 1998, BSAS, pp. 169.
CHIKUNYA S., MILLER E.L., 1999. Effects of source of bacterial isolate and microbial marker on the magni-tude of absolute values of microbial nitrogen yield in sheep. In: Proceedings of the BSAS Winter Meeting1999, BSAS, pp. 29.
COLEMAN G.S., SANDFORD D.C., 1979. The engulfment and digestion of mixed rumen bacteria and indivi-dual bacterial species of rumen ciliate protozoa grown in vivo. J. Agric .Sci, Camb., 92, 729-742.
CRAIG W.M., BROWN D.R., BRODERICK G.A., RICKER D.B., 1987. Post-pandrial composition changes offluid- and particle-associated ruminal microorganisms. J. Anim. Sci., 65, 1042-1053.
CZERKAWSKI J.W., 1974. Methods for determining 2-6-diaminopimelic and 2-aminoethylphosphonic acid ingut contents. J. Sci. Food Agric., 25, 45-55.
DEHORITY B.A., 1995. Methodology for measuring microbial growth in the rumen. In: Proceedings of theInternational Symposium on the Nutrition Requirements of Ruminants, Universidad Federal de Vicosa, Vi-cosa-MG-Brasil. pp 121-137.
DUFVA G.S., BARTLEY E.E., ARAMBEL M.J., NAGARAJE T.G., DENNIS S.M., GALITZER S.G.,DAYTON A.D., 1982. Diaminopimelic acid content of feeds and rumen bacteria and its usefulness as a ru-men bacterial marker. J. Dairy Sci., 65, 1754-1759.
EVANS R.A., AXFORD F.E., OFFER N.W., 1975. A method for estimating the quantities of microbial and die-tary proteins flowing in the duodenal digesta of ruminants. Proc. Nutr. Soc., 34, 65A.
ELLIS, W.C, PFANDER W.H., 1965. Rumen microbial polynucleotide synthesis and its possible role in rumi-nant nitrogen synthesis. Nature (Lond.), 205, 974-975.
FAICHNEY G.J., 1975. The use of markers to partition digestion within the gastrointestinal tract of ruminants.In: Digestion and Metabolism in the Ruminant. McDonald I.W., Warner A.C.I. eds. Armidale Universityof New England Publishing Unit, pp. 277-291.
FERGUSON J.D., CHALUPA W., 1989. Impact of protein nutrition on reproduction in dairy cows. J. DairySci., 72, 746-766.
FIRKINS J.L., BERGER L.L., MERCHEN N.R., FAHEY G.C.JR., MULVANEY R.L., 1987. Ruminal nitrogenmetabolism in steers as affected by feed intake and dietary urea concentration. J. Dairy Sci., 70,2302-2311.
FIRKINS J.L., WEISS W.P., PIWONKA E.J., 1992. Quantification of intraruminal recycling of microbial nitro-gen using nitrogen-15. J. Dairy Sci., 70, 3223-3233.
FORSBERG C.W., LAM K., 1977. Use of adenosine 5’-triphosphate as an indicator of the microbiota biomassin rumen contents. Appl. Environ. Microbiol., 33, 528-537.
GARRET, J.E., GOODRICH, R.D., MEISKE, J.C., STERN, M.D., 1986. Influence of supplemental nitrogensource on digestion of nitrogen, dry matter and organic matter and on in vivo rate of ruminal protein degra-dation. J. Anim. Sci., 64, 1801-1810.
GIESECKE D., EHRENTREICH L., STANGASSINGER M., AHRENS F., 1994. Mammary and renal excre-tion of purine metabolites in relation to energy intake and milk yield in dairy cows. J. Dairy Sci., 77,2376-2380.
GOMEZ L., BOGAERT C., JOUANY J.P., LASSALAS B., 1991. The influence of lasalocid and cationmycinon nitrogen digestion in sheep: comparison of ethods for estimating microbial nitrogen. Can. J. Anim. Sci.,71, 389-399.
HORIGANE, A., HORIGUCHI M., 1990. Nutritional aspects and metabolism of aminophosphonic acids in ru-minants. In: The Rumen Ecosystem. The Microbial Metabolism and Its Regulation. Hoshino, S., Onodera,R., Minato, H. and Itabashi, H., eds. Springer Verlag, , pp. 51-72. Nueva York.
HORIGUCHI M., KANDATSU M., 1959. Isolation of 2-aminoethanephosphonic acid from rumen protozoa.Nature (Lond.), 184, 901-902.
ILLG D.J., STERN M.D., 1994. In vitro and in vivo comparisons of diaminopimelic acid and purines for esti-mating protein synthesis in the rumen. Anim. Feed Sci. Technol., 48, 49-55.
KENNEDY P.M., HAZLEWOOD G.P., MILLIGAN L.P., 1984. A comparison of methods for the estimation ofthe proportion of microbial nitrogen in duodenal digesta, and of correction for microbial contamination innylon bags incubated in the rumen of sheep. Br. J. Nutr., 52, 403-417.
24 M.D. CARRO

KOMISARCZUK S., DURAND M., BEAUMATIN PH., HANNEQUART G., 1987. Utilisation de l’azote 15pour la mesure de la protéosynthèse microbienne dans les phases solide et liquide d’un fermenteursemi-continu (Rusitec). Reprod. Nutr. Develop., 27 (1B), 261-262.
LAMBERT A., LUCAS F., BLANCHART G., 1998. Dégradation et prélèvement de peptides de caséines mar-qués au 14C par des bactéries mixtes du rumen. Reprod. Nutr. Dev., 38, 69-79.
LEBZIEN P., GIESECKE D., WIESMAYR S., ROHR K., 1993. Messung der mikrobiellen Proteinsynthese miPansen von Kühen mittels 15N-Bestimmung mi Doudenalchymus und Allantoinausscheidung in der Milch.J. Anim. Physiol. Anim. Nutr., 70, 82-882.
LEGAY-CARMIER F., BAUCHART D., 1989. Distribution of bacteria in the rumen contents of dairy cows gi-ven a diet supplemented with soya-bean oil. Br. J. Nutr., 61, 725-740.
LING J.R., BUTTERY P.J., 1978. The simultaneous use of ribonucleic acid, 35S, 2,6-diaminopimelic acid and2-aminoethylphosphonic acid as markers of microbial nitrogen entering the duodenum of sheep. Br. J.Nutr., 39, 165-179.
MAKKAR H.P.S., BECKER K., 1999. Purine quantification in digesta from ruminants by spectrophotometricand HPLC methods. Br. J. Nutr., 81, 107-112.
MARTIN C., WILLIAMS A.G., MICHALET-DOREAU B., 1994. Isolation and characteristics of the protozoaland bacterial fractions from bovine ruminal contents. J. Anim. Sci., 72, 2962-2968.
MARTÍN-ORÚE S.M., BALCELLS J., ZAKRAOUI F., CASTRILLO C., 1998. Quantification and chemicalcomposition of mixed bacteria harvested from solid fractions of rumen digesta: effect of detachment proce-dure. Anim. Feed Sci. Technol., 71, 269-282.
MASSON H.A., DENHOLM A.M., LING J.R., 1991. In vivo metabolism of 2,2’-diaminopimelic acid fromgram-positive and gram-negative bacterial cells by ruminal microorganims and ruminants and its use as amarker of bacterial biomass. Appl. Environ. Microbiol., 57, 1714-1720.
MATHERS J.C., MILLER E.L., 1980. A simple procedure using 35S incorporation for the measurement of mi-crobial and undegraded food protein in ruminant digesta. Br. J. Nutr., 43, 503-514.
MERRY R.J., MCALLAN A.B., SMITH R.H., 1990. In vitro continuous culture studies on the effect of nitro-gen source on rumen microbial growth and fibre digestion. Anim. Feed Sci. Technol., 31, 55-64.
MCALLAN A.B., 1982. The fate of nucleic acids in ruminants. Proc. Nutr. Soc., 41, 309-317.MCALLAN A.B., SMITH R.H., 1973a. Degradation of nucleic acids in the rumen. Br. J. Nutr., 29, 331-345.MCALLAN A.B., SMITH R.H., 1973b. Degradation of nucleic acid derivatives by rumen bacteria in vitro. Br.
J. Nutr., 29, 467-474.MERRY R.J., MCALLAN A.B., 1983. A comparison of the chemical composition of mixed bacteria harvested
from the liquid and solid fractions of rumen digesta. Br. J. Nutr., 50, 701-709.NIKOLIC J.A., JOVANOVIC M., 1973. Preliminary studies on the use of different methods for determining the
proportions of bacterial nitrogen in the total nitrogen of rumen contents. J. Agric. Sci., Camb., 81, 1-7.NOCEK J.E., RUSSELL J.B., 1988. Protein and carbohydrate as an integrated system. Relationship of ruminal
availability to microbial contribution and milk producion. J. Dairy Sci., 71, 2070-2082.OLUBOBOKUM J.A., CRAIG W.M., 1990. Quantity and characteristics of microorganisms associated with ru-
minal fluid or particles. J. Anim. Sci., 68, 3360-3370.OBISPO N.E., DEHORITY B.A., 1999. Feasibility of using total purines as a marker for ruminal bacteria. J.
Anim. Sci., 77, 3084-3095.PÉREZ J.F., BALCELLS J., GUADA J.A., CASTRILLO C., 1996a. Determination of rumen microbial-nitro-
gen production in sheep: a comparison of urinary purine excretion with methods using 15N and purine ba-ses as markers of microbial-nitrogen entering the duodenum. Br. J. Nutr., 75, 699-709.
PÉREZ J.F., RODRÍGUEZ C.A., GONZÁLEZ J., BALCELLS J., GUADA J.A., 1996b. Contribution of dietarypurine bases to duodenal digesta in sheep. In situ studies of purine degradability corrected for microbialcontamination. Anim. Feed Sci. Technol., 62, 251-262.
PÉREZ J.F., BALCELLS J., GUADA J.A., CASTRILLO C., 1997a. Contribution of dietary nitrogen and purinebases to the duodenal digesta: comparison of duodenal and polyester-bag measurements. Anim. Sci., 65,237-245.
PÉREZ J.F., BALCELLS J., GUADA J.A., CASTRILLO C., 1997b. Rumen microbial production estimated eit-her from urinary purine derivative excretion or from direct measurements of 15N and purine bases as micro-bial markers: effect of protein source and rumen bacteria isolates. Anim. Sci., 65, 225-236.
PÉREZ J.F., BALCELLS J., FONDEVILLA M., GUADA J.A., 1998a. Composition of liquid- and particle-as-sociated bacteria and their contribution to the rumen outflow. Aust. J. Agric. Res., 49, 907-914.
PÉREZ J.F., BALCELLS J., CEBRIÁN J.A., MARTÍN-ORÚE S.M., 1998b. Excretion of endogenous and exo-genous purine derivatives in sheep: effect of increased concentrate intake. Br. J. Nutr., 79, 237-240.
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 25

PROKSCH G., 1972. 14N/15N ratio determination by emission spectrometry and its present possibilities. In: Tra-cer Studies on Non-protein Nitrogen for Ruminants. International Atomic Energy Agency, Vienna, pp.163-169.
QUIGLEY J.D., SCHWAB C.G., 1988. Comparison of D-alanine and diaminopimelic acid as bacterial markersin young calves. J. Anim. Sci., 66, 758-767.
RAHNEMA S.H., THEURER B., 1986. Comparison of various amino acids for estimation of microbial nitrogendigesta. J. Anim. Sci., 63, 603-612.
RANILLA M.J., CARRO M.D., LÓPEZ S., VALDÉS C., NEWBOLD J.C., WALLACE R.J. 2000. 15N-ammo-nia incorporation by ruminal bacteria growing on different N sources. Repr. Nutr. Develop., 40:2, 199.
RANILLA M.J., TEJIDO, M.L., CARRO M.D., 2001. Comparación de diversos métodos de desligamiento debacterias ruminales asociadas a la fase sólida de la digesta en un sistema in vitro (Rusitec). ITEA (Vol. ex-tra), 22, pp. 382-384.
ROBINSON P.H., FADEL J.G., IVAN M., 1996. Critical evaluation of diaminopimelic and robonucleic acid asmarkers to estimate rumen pools and duodenal flows of bacterial and protozoal nitrogen. Can. J. Anim.Sci., 76, 587-597.
RUSSELL J.B., O’CONNOR J.D., FOX D.G., VAN SOEST P.J., SNIFFEN C.J., 1992. A net carbohydrate andprotein system for evaluating cattle diets: I. Ruminal fermentation. J. Anim. Sci., 70, 3551-3561.
SCHELLING G.T., BRYERS F.M., 1984. Cytosine as a marker for microbial nitrogen leaving the rumen. Can.J. Anim. Sci., 64 (Suppl.), 62-71.
SCHNIFFEN C.J., ROBINSON P.H., 1987. Microbial growth and flow as influenced by dietary manipulation.J. Dairy Sci., 70, 425-441.
SCHÖNHUSEN ULRIKE, VOIGT J., KREIENBRING F., TEUSCHER F., 1995. Bewertung verschiedenerMarker für die Messung der Mikrobiellen Stickstoffpassage am Duodenum der Milchkuh. Arch. Anim.Nutr., 48, 147-158.
SIBANDA S., TOPPS J.H., STORM E., ØRSKOV E.R., 1982. The excretion of allantoin by ruminants in rela-tion to protein entering the abomasum. Proc. Nutr. Soc., 41, 75A.
SIDDONS R.C., BEEVER D.E., NOLAN J.V., 1982. A comparison of methods for the estimation of microbialnitrogen in duodenal digesta of sheep. Br. J. Nutr., 48, 377-389.
SMITH R.H., MCALLAN A.B., HEWITT P., LEWIS P.E., 1978. Estimation of amounts of microbial and die-tary nitrogen compounds entering the duodenum of cattle. J. Agric. Sci., Camb., 90, 557-568.
STANGASSINGER M., CHEN X.B., LINDBERG J.E., GIESECKE D., 1995. Metabolism of purines in rela-tion to microbial production. In: Ruminant Physiology: Digestion, Metabolism, Growth and Reproduction.Engelhardt, W.w., Leonhard-Marek, S., Breves, G., Giesecke, D., eds. Ferdinand Enke Verlag, Stuttgart,pp. 387-406.
STERN M.D., HOOVER W.H., 1979. Methods for determining and factors affecting rumen microbial proteinsynthesis: a review. J. Anim. Sci., 49, 1590-1603.
STERN M.D., VARGA G.A., CLARK J.H., FIRKINS J.L., HUBER J.T., PALMQUIST D.L., 1994. Evaluationof chemical and physical properties of feeds that affect protein metabolism in the rumen. J. Dairy Sci., 77,2762-2786.
TEJIDO M.L., CARRO M.D., RANILLA M.J., ARÍN M.J., DÍEZ M.T., LÓPEZ S., 2001. Utilización de 15N ybases púricas como marcadores para estimar la síntesis microbiana in vitro. ITEA (Vol. extra), 22, pp.385-387.
TITGEMEYER E.C., MERCHEN N.R., BERGER L.L., 1989. Evaluation of soybean meal, corn gluten meal,blood meal and fish meal as sources of nitrogen and amino acids disappearing from the small intestine ofsteers. J. Anim. Sci., 67, 262-275.
TOPPS J.H., ELLIOTT R.C., 1965. Relationship between concentrations of ruminal nucleic acids and excretionof purine derivatives by sheep. Nature (Lond.), 205, 498-499.
USHIDA K., LASSALAS B., JOUANY J.P., 1985. Determination of assay parameters for RNA analysis in bac-terial and duodenal samples by spectrophotometry. Influence of sample treatment and preservation. Re-prod. Nutr. Develop., 25, 1037-1046.
VAN NEVEL C.J., DEMEYER D.I., 1977. Determination of rumen microbial growth in vitro from 32P-labelledphosphate incorporation. Br. J. Nutr., 38, 101-114.
VERBIC J., CHEN X.B., MACLEOD N.A., ØRSKOV E.R., 1990. Excretion of purine derivatives by rumi-nants: effect of microbial nucleic acid infusion on purine derivative excretion by steers. J. Agric. Sci.,Camb., 114, 243-248.
WALKER D.J., NADER C.J., 1975. Measurement in vivo of rumen microbial protein synthesis. Aust. J. agric.Res., 26, 689-698.
WALLACE, R.J., WEST, A.A., 1982. Adenosine triphosphate and deoxyribonucleic acidin the alimentary tractof cattle fed different N sources. J. Appl. Bacteriol., 45, 49-57.
26 M.D. CARRO

WARNER A.C.I., 1981. Rate of passage of digesta through the gut of mammals and birds. Nutr. Abstr. Rev.,Series B, 51, 789-820.
WENHAM G., WHYBURN R.S., 1980. A radiological investigation of the effects of cannulation on intestinalmotility and digesta flow in sheep. J. Agric. Sci., Camb., 95, 539-546.
WHITEHOUSE N.L., OLSON V.M., SCHWAB C.G., CHESBRO W.R., CUNNINGHAM K.D., LYKOS T.,1994. Improved techniques for dissociating particle-associated mixed ruminal microorganims from ruminaldigesta solids. J. Anim. Sci., 72, 1335-1343.
ZINN R.A., OWENS F.N., 1986. A rapid procedure for purine measurement and its use for estimating net rumi-nal protein synthesis. Can. J. Anim. Sci., 66, 157-166.
Invest. Agr.: Prod. Sanid. Anim. Vol. 16 (1), 2001
SÍNTESIS DE PROTEÍNA MICROBIANA Y MARCADORES MICROBIANOS 27





























