Embed Size (px)
Citation preview
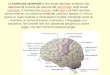
CORPO CALLOSO MESENCEFALO
TALAMO
IPOTALAMO
IPOFISI
PONTE
BULBO CERVELLETTO
La formazione della cortecciacerebrale
Lo studio del ruolo dei geni nello sviluppo embrionale di animali alivelli diversi della scala evolutiva chiarisce i processi che fecero
emergere la struttura più straordinaria e complessa nota in natura
N
el nostro cervello si distinguonosezioni diverse preposte a com-piti diversi. Il talamo, per esem-
pio, costituisce una prima stazione diraccolta e di smistamento dei segnaliche provengono dagli organi di senso; icosiddetti gangli della base regolano,tra l'altro, i dettagli fini dei nostri movi-menti; il sistema limbico controlla lenostre risposte istintive e la nostra emo-tività, oltre a sovrintendere al consoli-damento dei nostri ricordi. Al di sopradi tutto vi è la corteccia cerebrale, lamateria grigia che riveste gli emisfericerebrali e alla quale spetta, per così di-re, l'ultima parola su tutto l'operato delcervello.
Dal punto di vista evolutivo la cor-teccia cerebrale ha origine con i primimammiferi. I mammiferi inferiori pre-sentano tuttavia una corteccia ancora
di Edoardo Boncinelli
poco sviluppata e molte delle loro azio-ni possono prescindere dalla «supervi-sione» della corteccia stessa. Nella li-nea evolutiva che ha condotto allescimmie e da queste all'uomo il con-trollo delle attività nervose si èprogressivamente spostato verso la cor-teccia, di modo che oggi tutto il nostrovissuto «passa» attraverso di essa, an-che ciò che forse preferiremmo viverein modo più istintivo. Con l'evoluzio-ne, quindi, la corteccia è venuta assu-mendo un ruolo sempre più centrale,nel bene e nel male, a livello sia indivi-duale sia sociale.
La corteccia cerebrale rappresenta lavera punta di diamante dell'evoluzionebiologica e molti considerano il suostudio come l'ultima frontiera dellabiologia, anzi, della scienza tout-court.Recentemente è stata lanciata un'offen-
siva in grande stile per espugnare i se-greti del cervello e della corteccia ce-rebrale, offensiva sferrata con l'ausiliodi un'incredibile varietà di strumentiintellettuali e materiali sempre più per-fezionati e potenti. E già si profilano iprimi timidi tentativi di affrontare ilproblema dei problemi: quello relativoalla natura della coscienza. Non vi èquindi da meravigliarsi se esperimentirecenti hanno gettato un po' di luce an-che sui meccanismi che regolano la for-mazione della corteccia cerebrale du-rante lo sviluppo embrionale.
LE TAPPE DELLO SVILUPPOEMBRIONALE
Un insieme coordinato di eventi bio-logici conduce alla formazione di unindividuo maturo, costituito da migliaia
Nel cervello umano si distinguono regioni, quali il talamo, igangli della base, il sistema limbico e la corteccia, i cui diver-
si compiti sottendono le attività straordinariamente ricche earticolate della mente.
LE SCIENZE n. 346, giugno 1997 47

MIELENCEFALO
EMISFERICEREBRALI
MESENCEFALOTELENCEFALO MESENCEFALO
DIENCEFALO METENCEFALO
ROMBOENCEFALOPROSENCEFALO
Il
CORPIQUADRIGEMINI
CERVELLETTO
III
MIDOLLOEMISFERI PONTE ALLUNGATO
CEREBRALI
MESENCEFALOTALAMO EIPOTALAMO
Mutanti omeotici di Drosophila. A sinistra è mostrata unadrosofila di tipo selvatico, al centro il mutante bithorax, che
porta quattro ali, e a destra il mutante Antennapedia, che re-ca sul capo un paio di zampette al posto delle antenne.
DROSOPHILA
4—Abd-B Abd-A Ubx Antp Scr Dfd Pb Lab
L_IMPT7
- HOX
EMBRIONE DI TOPO
Geni HOX dei mammiferi e loro rapporto con i geni omeotici di Drosophila. Sonomostrati schematicamente i geni appartenenti ai 13 gruppi di geni HOX del topo edell'uomo. Geni corrispondenti nel topo e nella drosofila hanno lo stesso colore. Èillustrato anche il fenomeno della colinearità, per cui i geni HOX del topo, o i corri-spondenti geni omeotici della drosofila, si susseguono sul cromosoma in un ordinedefinito che corrisponde a quello delle regioni del corpo dove essi sono attivi.
Fasi principali dello sviluppo del cervellodi un mammifero. All'inizio si distinguo-no solo tre vescicole cerebrali chiamate,dall'anteriore alla posteriore, prosencefa-lo, mesencefalo e romboencefalo. Più tar-di, all'interno del prosencefalo si distin-guono un telencefalo e un diencefalo, e ilromboencefalo si articola nel metencefalo(da cui deriverà essenzialmente il cervel-letto) e nel mielencefalo, che darà luogoalle strutture poste al confine fra il cer-vello vero e proprio e il midollo spinale.
tutta la regione dorsale dell'embrioneappena uscito dalla fase di gastrulazio-ne riceve segnali che orientano le cellu-le in direzione neurale. Si viene così aformare in questa regione un ispessi-mento, la placca neurale, che costitui-sce l'abbozzo del futuro sistema nervo-so. Successivamente la placca neuralesi espande e si ripiega a formare dap-prima una doccia e poi il tubo neurale.Durante questo periodo il primo abboz-zo di tubo neurale deve essere suddivi-so in ampie regioni che si succedonolungo il suo asse antero-posteriore e insottoregioni e domini anatomo-funzio-nali lungo lo stesso asse e lungo quellodorsoventrale. All'interno di questearee e di questi domini si deve inoltredifferenziare un certo numero di tipicellulari diversi.
La parte anteriore del tubo neurale,per esempio, si presenta costituita datre vescicole cerebrali che sono, dal-l'anteriore alla posteriore, il prosence-falo, il mesencefalo e il romboencefalo.Più o meno verso il decimo giorno del-
lo sviluppo embrionale del topo o versola quarta settimana dello sviluppo del-l'uomo, la parte anteriore della vesci-cola corrispondente al prosencefalo co-mincia a organizzarsi per dare luogo adue regioni caratterizzate da un'archi-tettura cellulare completamente diver-sa. La parte dorsale si orienta verso unastruttura stratificata a piani cellulari so-vrapposti (che darà luogo alla cortecciacerebrale) mentre la parte ventrale siorganizza in una costellazione diconglomerati nucleari di cellule nervo-se separati da tratti di sostanza bianca,che prendono collettivamente il nomedi nuclei o gangli della base.
Lo SVILUPPODELLA CORTECCIA CEREBRALE
In un uomo adulto questo tessuto ècostituito da circa cento miliardi di cel-lule nervose vere e proprie, dette neu-roni, e da quasi altrettante cellule di so-stegno dette cellule gliali. Tutta questaenorme popolazione cellulare si originada uno straterello di cellule precorritri-ci, i neuroblasti, presenti nell'abbozzodella corteccia fin dall'inizio. È chiaroquindi che il primo passo nel suo svi-luppo sarà necessariamente rappresen-tato da un rapido succedersi di cicli diproliferazione cellulare. Per qualchetempo dopo la prima definizione delterritorio embrionale destinato a dareluogo alla corteccia, le cellule che vi sitrovano non fanno altro che moltipli-carsi, a una velocità che può raggiunge-re le 250 000 nuove cellule al minuto.Mentre si dividono, esse si mantengonoin una ristretta regione immediatamenteadiacente al ventricolo, la cavità cere-brale di cui la corteccia costituisce iltetto. Per questo motivo la regione nel-la quale si moltiplicano le cellule dellafutura corteccia prende il nome di zonaventricolare, o zona germinativa ventri-colare. Solo dopo che il numero dellecellule della futura corteccia ha rag-
giunto un certo valore, alcune di essesmettono di moltiplicarsi e comincianoa differenziarsi e a maturare per rag-giungere la loro identità finale.
Le cellule che via via escono dal ci-clo proliferativo migrano per qualchefrazione di millimetro verso la superfi-cie esterna della testa e vanno a collo-carsi in una cosiddetta zona intermedia,nella quale restano per qualche tempo.Successivamente, alcune di queste ri-prendono a migrare verso una regioneancora più esterna, la cosiddetta placcacorticale, dalla quale deriverà poi lamateria grigia della corteccia adulta.
Intorno alla metà della gestazione sipossono quindi distinguere nella cortec-cia cerebrale in via di sviluppo tre zoneprincipali e cioè, dalla più interna allapiù esterna, la zona germinativa ventri-colare, la zona intermedia e la placcacorticale. All'inizio della formazionedella corteccia esiste soltanto la zonaventricolare; dopo qualche tempo com-pare anche quella intermedia; più tardiinfine si forma la placca corticale. Con ilpassare del tempo la placca corticale siispessisce sempre più a spese della zonaintermedia e di quella ventricolare. Que-st'ultima diviene sempre più tenue fin-ché, intorno al diciassettesimo giorno neltopo e alla sedicesima settimana nell'uo-mo, scompare quasi del tutto. È noto chei neuroni corticali, una volta usciti dal ci-clo proliferativo non si replicano più. Lamaggior parte di essi è già al proprio po-sto molto prima della nascita; questineuroni non possono più venire sostitui-ti, né altri possono esservi aggiunti.
Di recente si è chiarita la modalità concui si formano i vari strati della placcacorticale prima e della corteccia poi. Ineuroni migrano dalla zona intermediaverso le loro diverse collocazioni nellaplacca corticale secondo una precisa se-quenza temporale basata sulla loro datadi nascita. I primi nati, cioè i primi auscire dal ciclo proliferativo, vanno a po-sizionarsi negli strati più interni dellaplacca che col tempo daranno luogo aglistrati più profondi della corteccia adulta,cioè il 5 e il 6, mentre i neuroni che so-praggiungono successivamente migranooltre i primi arrivati, fino a una regionepiù esterna, per andare a formare gli stra-ti superiori della corteccia, cioè gli strati2, 3 e 4. Esistono particolari ceppi di to-po nei quali la struttura della corteccia sipresenta profondamente alterata perchéal momento di migrare i neuroni corticalinon hanno ricevuto i segnali giusti oppu-re non-li hanno saputi riconoscere.
Una volta formati i tipici sei strati dicellule nervose, si passa a poco a pocodalla placca corticale alla corteccia ce-rebrale vera e propria. I neuroni giun-ti in sede cominciano a differenziarsi ead acquisire i caratteri definitivi. I corpi
cellulari dei neuroni che costituisconola materia grigia emettono i tipici pro-lungamenti, o processi neuronali, equesti si allacciano a loro volta in unaselva di connessioni che costituisce ilsubstrato materiale della mente.
Ogni neurone presenta due tipi diprocessi: un assone e un certo numerodi dendriti. L'assone è un prolunga-mento della cellula nervosa che connet-te il neurone con un suo bersaglio, chepuò essere o un altro neurone, oppureun muscolo, e che si può trovare anchea distanza di decine di centimetri dalcorpo cellulare del neurone. Una buonaparte dei neuroni corticali invia gli as-soni fuori dalla corteccia. Questi posso-no collegare tra loro due diverse regio-ni della corteccia - che si trovino peresempio in due emisferi diversi - omettere in contatto una specifica regio-ne corticale con strutture nervose diver-
se dalla corteccia (dette collettivamentesubcorticali). L'insieme degli assoni af-ferenti ed efferenti costituisce la massi-ma parte della sostanza bianca degliemisferi, che si forma alla fine della ge-stazione e subito dopo la nascita sotto isei strati della materia grigia, occupan-do in pratica lo spazio lasciato liberodalla zona intermedia.
I dendriti sono generalmente più cor-ti, esili e ramificati degli assoni epermettono ai corpi cellulari dei varineuroni di connettersi ai corpi cellularidi altri neuroni limitrofi. Moltissimineuroni corticali sono incredibilmentericchi di dendriti e possono anche man-care di un vero e proprio assone. I neu-roni di questo tipo costituiscono lo spe-cifico della corteccia cerebrale: da essidipende la sua incredibile connettività.La corteccia è infatti essenzialmenteuna gigantesca rete di cellule nervose
di miliardi di cellule appartenenti acentinaia di tessuti diversi, a partire dauna singola cellula uovo fecondata. Viè da rimanere incantati a osservare letappe di questo meraviglioso processo,ma fino a poco tempo fa di tutto ciò sicomprendeva ben poco. Oggi si intrav-vede finalmente qualche barlume, e al-cuni eventi macroscopici sono statimessi in relazione con altrettanti pro-cessi microscopici. I vari passaggi checonducono alla formazione del sistemanervoso centrale dei vertebrati sonostati descritti da tempo nelle loro lineeessenziali, anche se sono tutt'oggi og-getto di studio negli embrioni dei siste-mi biologici più diversi.
Le basi per la costruzione di questocomplesso sistema vengono gettate su-bito dopo la gastrulazione, il processoche trasforma una sfera cava costitui-ta da poche cellule embrionali primi-tive in una serie di tubi paralleli cherappresentano una copia miniaturizzatama già riconoscibile del futuro organi-smo. L'epidermide che si trova lungo
48 LE SCIENZE n. 346, giugno 1997LE SCIENZE n. 346, giugno 1997 49
HOXb 3
HOXb 4
HOXb 5
rl
b7 b6 b5 b4 b3
HOXb
b2 b1
Schema della regione romboencefalica di un embrio-ne di topo, in cui è mostrata la localizzazione dell'at-tività di alcuni geni HOX; il codice di colori è ugualea quello dell'illustrazione a pagina 49. Sono indicatele radici di alcuni nervi cranici e la vescicola otica(VO), che segna la posizione del futuro orecchio.
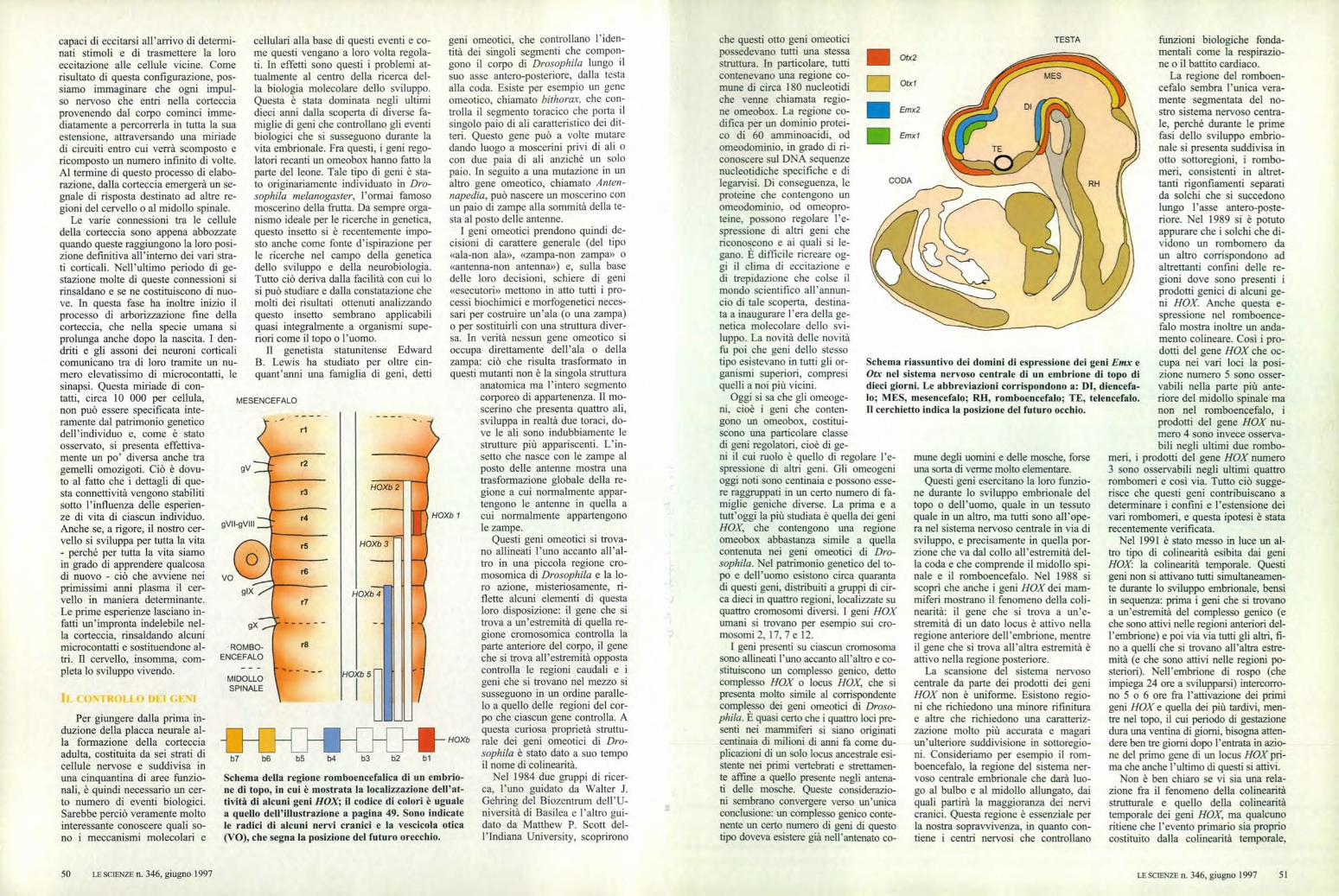
Otx2
Otxl
Emx2
Emxl
Schema riassuntivo dei domini di espressione dei geni Emx eOtx nel sistema nervoso centrale di un embrione di topo didieci giorni. Le abbreviazioni corrispondono a: DI, diencefa-lo; MES, mesencefalo; RH, romboencefalo; TE, telencefalo.Il cerchietto indica la posizione del futuro occhio.
gV
glX
gVll-gVlIl
gX
ROMBO-ENCEFALO
- - -
MIDOLLOSPINALE
capaci di eccitarsi all'arrivo di determi-nati stimoli e di trasmettere la loroeccitazione alle cellule vicine. Comerisultato di questa configurazione, pos-siamo immaginare che ogni impul-so nervoso che entri nella cortecciaprovenendo dal corpo cominci imme-diatamente a percorrerla in tutta la suaestensione, attraversando una miriadedi circuiti entro cui verrà scomposto ericomposto un numero infinito di volte.Al termine di questo processo di elabo-razione, dalla corteccia emergerà un se-gnale di risposta destinato ad altre re-gioni del cervello o al midollo spinale.
Le varie connessioni tra le celluledella corteccia sono appena abbozzatequando queste raggiungono la loro posi-zione definitiva all'interno dei vari stra-ti corticali. Nell'ultimo periodo di ge-stazione molte di queste connessioni sirinsaldano e se ne costituiscono di nuo-ve. In questa fase ha inoltre inizio ilprocesso di arborizzazione fine dellacorteccia, che nella specie umana siprolunga anche dopo la nascita. I den-driti e gli assoni dei neuroni corticalicomunicano tra di loro tramite un nu-mero elevatissimo di microcontatti, lesinapsi. Questa miriade di con-tatti, circa 10 000 per cellula,non può essere specificata inte-ramente dal patrimonio geneticodell'individuo e, come è statoosservato, si presenta effettiva-mente un po' diversa anche tragemelli omozigoti. Ciò è dovu-to al fatto che i dettagli di que-sta connettività vengono stabilitisotto l'influenza delle esperien-ze di vita di ciascun individuo.Anche se, a rigore, il nostro cer-vello si sviluppa per tutta la vita- perché per tutta la vita siamoin grado di apprendere qualcosadi nuovo - ciò che avviene nei VO
primissimi anni plasma il cer-vello in maniera determinante.Le prime esperienze lasciano in-fatti un'impronta indelebile nel-la corteccia, rinsaldando alcunimicrocontatti e sostituendone al-tri. Il cervello, insomma, com-pleta lo sviluppo vivendo.
CONTROLLO D = FNI
Per giungere dalla prima in-duzione della placca neurale al-la formazione della cortecciaadulta, costituita da sei strati dicellule nervose e suddivisa inuna cinquantina di aree funzio-nali, è quindi necessario un cer-to numero di eventi biologici.Sarebbe perciò veramente moltointeressante conoscere quali so-no i meccanismi molecolari e
cellulari alla base di questi eventi e co-me questi vengano a loro volta regola-ti. In effetti sono questi i problemi at-tualmente al centro della ricerca del-la biologia molecolare dello sviluppo.Questa è stata dominata negli ultimidieci anni dalla scoperta di diverse fa-miglie di geni che controllano gli eventibiologici che si susseguono durante lavita embrionale. Fra questi, i geni rego-latori recanti un omeobox hanno fatto laparte del leone. Tale tipo di geni è sta-to originariamente individuato in Dro-sophila melanogaster, l'ormai famosomoscerino della frutta. Da sempre orga-nismo ideale per le ricerche in genetica,questo insetto si è recentemente impo-sto anche come fonte d'ispirazione perle ricerche nel campo della geneticadello sviluppo e della neurobiologia.Tutto ciò deriva dalla facilità con cui losi può studiare e dalla constatazione chemolti dei risultati ottenuti analizzandoquesto insetto sembrano applicabiliquasi integralmente a organismi supe-riori come il topo o l'uomo.
Il genetista statunitense EdwardB. Lewis ha studiato per oltre cin-quant'anni una famiglia di geni, detti
geni omeotici, che controllano l'iden-tità dei singoli segmenti che compon-gono il corpo di Drosophila lungo ilsuo asse antero-posteriore, dalla testaalla coda. Esiste per esempio un geneomeotico, chiamato bithorax, che con-trolla il segmento toracico che porta ilsingolo paio di ali caratteristico dei dit-teri. Questo gene può a volte mutaredando luogo a moscerini privi di ali ocon due paia di ali anziché un solopaio. In seguito a una mutazione in unaltro gene omeotico, chiamato Anten-napedia, può nascere un moscerino conun paio di zampe alla sommità della te-sta al posto delle antenne.
I geni omeotici prendono quindi de-cisioni di carattere generale (del tipo«ala-non ala», «zampa-non zampa» o«antenna-non antenna») e, sulla basedelle loro decisioni, schiere di geni«esecutori» mettono in atto tutti i pro-cessi biochimici e morfogenetici neces-sari per costruire un'ala (o una zampa)o per sostituirli con una struttura diver-sa. In verità nessun gene omeotico sioccupa direttamente dell'ala o dellazampa: ciò che risulta trasformato inquesti mutanti non è la singola struttura
anatomica ma l'intero segmentocorporeo di appartenenza. Il mo-scerino che presenta quattro ali,sviluppa in realtà due toraci, do-ve le ali sono indubbiamente lestrutture più appariscenti. L'in-setto che nasce con le zampe alposto delle antenne mostra unatrasformazione globale della re-gione a cui normalmente appar-tengono le antenne in quella acui normalmente appartengonole zampe.
Questi geni omeotici si trova-no allineati l'uno accanto all'al-tro in una piccola regione cro-mosomica di Drosophila e la lo-ro azione, misteriosamente, ri-flette alcuni elementi di questaloro disposizione: il gene che sitrova a un'estremità di quella re-gione cromosomica controlla laparte anteriore del corpo, il geneche si trova all'estremità oppostacontrolla le regioni caudali e igeni che si trovano nel mezzo sisusseguono in un ordine paralle-lo a quello delle regioni del cor-po che ciascun gene controlla. Aquesta curiosa proprietà struttu-rale dei geni omeotici di Dro-sophila è stato dato a suo tempoil nome di colinearità.
Nel 1984 due gruppi di ricer-ca, l'uno guidato da Walter J.Gehring del Biozentrum dell'U-niversità di Basilea e l'altro gui-dato da Matthew P. Scoli del-l'Indiana University, scoprirono
che questi otto geni omeoticipossedevano tutti una stessastruttura. In particolare, tutticontenevano una regione co-mune di circa 180 nucleotidiche venne chiamata regio-ne omeobox. La regione co-difica per un dominio protei-co di 60 amminoacidi, odomeodominio, in grado di ri-conoscere sul DNA sequenzenucleotidiche specifiche e dilegarvisi. Di conseguenza, leproteine che contengono unomeodominio, od omeopro-teine, possono regolare l'e-spressione di altri geni chericonoscono e ai quali si le-gano. È difficile ricreare og-gi il clima di eccitazione edi trepidazione che colse ilmondo scientifico all'annun-cio di tale scoperta, destina-ta a inaugurare l'era della ge-netica molecolare dello svi-luppo. La novità delle novitàfu poi che geni dello stessotipo esistevano in tutti gli or-ganismi superiori, compresiquelli a noi più vicini.
Oggi si sa che gli omeoge-ni, cioè i geni che conten-gono un omeobox, costitui-scono una particolare classedi geni regolatori, cioè di ge-ni il cui ruolo è quello di regolare l'e-spressione di altri geni. Gli omeogenioggi noti sono centinaia e possono esse-re raggruppati in un certo numero di fa-miglie geniche diverse. La prima e atutt'oggi la più studiata è quella dei geniHOX, che contengono una regioneomeobox abbastanza simile a quellacontenuta nei geni omeotici di Dro-sophila. Nel patrimonio genetico del to-po e dell'uomo esistono circa quarantadi questi geni, distribuiti a gruppi di cir-ca dieci in quattro regioni, localizzate suquattro cromosomi diversi. I geni HOXumani si trovano per esempio sui cro-mosomi 2, 17, 7 e 12.
I geni presenti su ciascun cromosomasono allineati l'uno accanto all'altro e co-stituiscono un complesso genico, dettocomplesso HOX o locus HOX, che sipresenta molto simile al corrispondentecomplesso dei geni omeotici di Droso-phda. È quasi certo che i quattro loci pre-senti nei mammiferi si siano originaticentinaia di milioni di anni fa come du-plicazioni di un solo locus ancestrale esi-stente nei primi vertebrati e strettamen-te affine a quello presente negli antena-ti delle mosche. Queste considerazio-ni sembrano convergere verso un'unicaconclusione: un complesso genico conte-nente un certo numero di geni di questotipo doveva esistere già nell'antenato co-
TESTA
mune degli uomini e delle mosche, forseuna sorta di verme molto elementare.
Questi geni esercitano la loro funzio-ne durante lo sviluppo embrionale deltopo o dell'uomo, quale in un tessutoquale in un altro, ma tutti sono all'ope-ra nel sistema nervoso centrale in via disviluppo, e precisamente in quella por-zione che va dal collo all'estremità del-la coda e che comprende il midollo spi-nale e il romboencefalo. Nel 1988 siscoprì che anche i geni HOX dei mam-miferi mostrano il fenomeno della coli-nearità: il gene che si trova a un'e-stremità di un dato locus è attivo nellaregione anteriore dell'embrione, mentreil gene che si trova all'altra estremità èattivo nella regione posteriore.
La scansione del sistema nervosocentrale da parte dei prodotti dei geniHOX non è uniforme. Esistono regio-ni che richiedono una minore rifiniturae altre che richiedono una caratteriz-zazione molto più accurata e magariun'ulteriore suddivisione in sottoregio-ni. Consideriamo per esempio il rom-boencefalo, la regione del sistema ner-voso centrale embrionale che darà luo-go al bulbo e al midollo allungato, daiquali partirà la maggioranza dei nervicranici. Questa regione è essenziale perla nostra sopravvivenza, in quanto con-tiene i centri nervosi che controllano
funzioni biologiche fonda-mentali come la respirazio-ne o il battito cardiaco.
La regione del romboen-cefalo sembra l'unica vera-mente segmentata del no-stro sistema nervoso centra-le, perché durante le primefasi dello sviluppo embrio-nale si presenta suddivisa inotto sottoregioni, i rombo-meri, consistenti in altret-tanti rigonfiamenti separatida solchi che si succedonolungo l'asse antero-poste-riore. Nel 1989 si è potutoappurare che i solchi che di-vidono un rombomero daun altro corrispondono adaltrettanti confini delle re-gioni dove sono presenti iprodotti genici di alcuni ge-ni HOX Anche questa e-spressione nel romboence-falo mostra inoltre un anda-mento colineare. Così i pro-dotti del gene HOX che oc-cupa nei vari loci la posi-zione numero 5 sono osser-vabili nella parte più ante-riore del midollo spinale manon nel romboencefalo, iprodotti del gene HOX nu-mero 4 sono invece osserva-bili negli ultimi due rombo-
meri, i prodotti del gene HOX numero3 sono osservabili negli ultimi quattrorombomeri e così via. Tutto ciò sugge-risce che questi geni contribuiscano adeterminare i confini e l'estensione deivari rombomeri, e questa ipotesi è statarecentemente verificata.
Nel 1991 è stato messo in luce un al-tro tipo di colinearità esibita dai geniHOX: la colinearità temporale. Questigeni non si attivano tutti simultaneamen-te durante lo sviluppo embrionale, bensìin sequenza: prima i geni che si trovanoa un'estremità del complesso genico (eche sono attivi nelle regioni anteriori del-l'embrione) e poi via via tutti gli altri, fi-no a quelli che si trovano all'altra estre-mità (e che sono attivi nelle regioni po-steriori). Nell'embrione di rospo (cheimpiega 24 ore a svilupparsi) intercorro-no 5 o 6 ore fra l'attivazione dei primigeni HOX e quella dei più tardivi, men-tre nel topo, il cui periodo di gestazionedura una ventina di giorni, bisogna atten-dere ben tre giorni dopo l'entrata in azio-ne del primo gene di un locus HOX pri-ma che anche l'ultimo di questi si attivi.
Non è ben chiaro se vi sia una rela-zione fra il fenomeno della colinearitàstrutturale e quello della colinearitàtemporale dei geni HOX, ma qualcunoritiene che l'evento primario sia propriocostituito dalla colinearità temporale,
MESENCEFALO
50 LE SCIENZE n. 346, giugno 1997
LE SCIENZE n. 346, giugno 1997 51
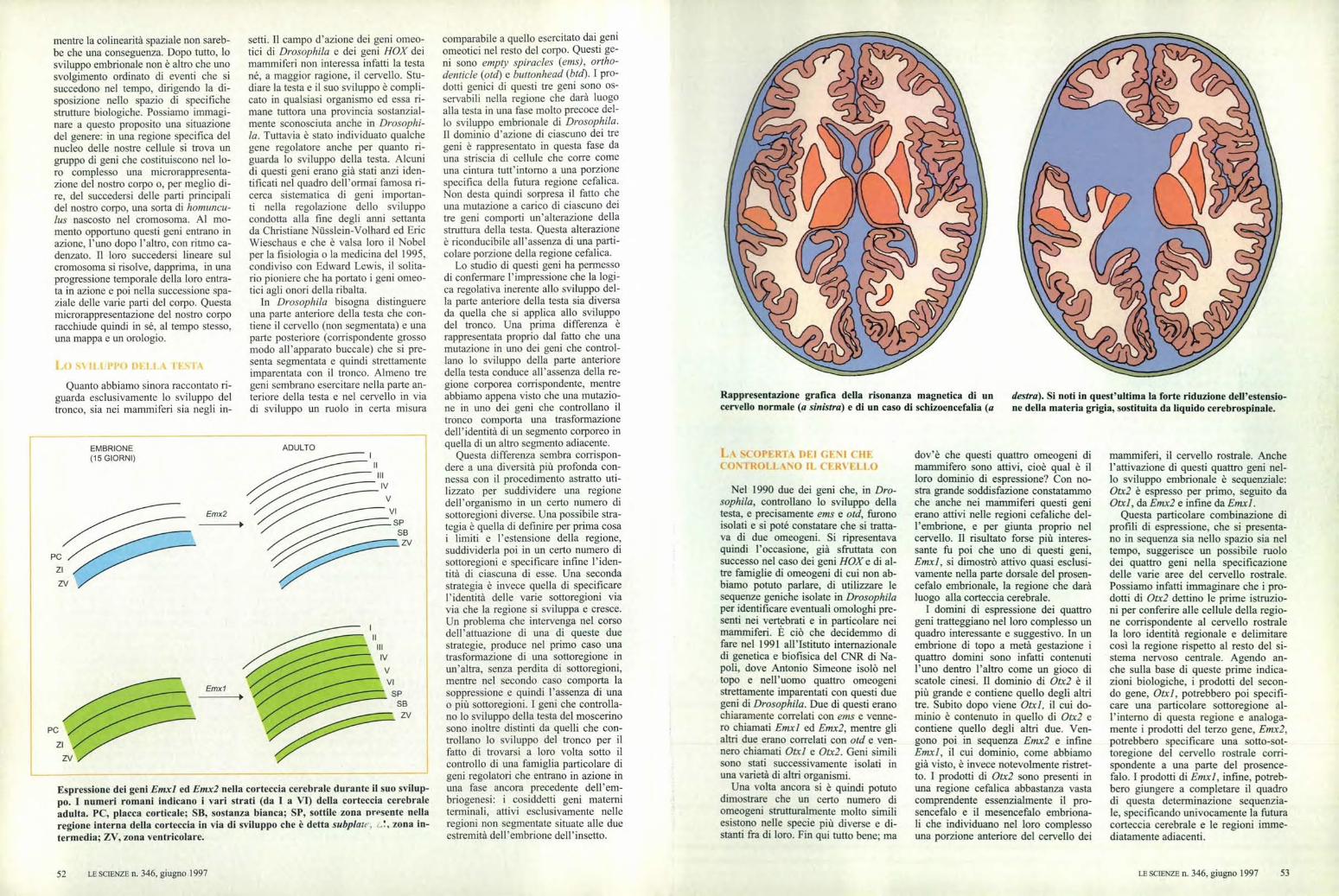
Rappresentazione grafica della risonanza magnetica di un destra). Si noti in quest'ultima la forte riduzione dell'estensio-cervello normale (a sinistra) e di un caso di schizoencefalia (a ne della materia grigia, sostituita da liquido cerebrospinale.
EMBRIONE
ADULTO(15 GIORNI)
Emxl
PC
ZI
ZV
IV
V
VISPSBZV
Emx2
Espressione dei geni Emxl ed Emx2 nella corteccia cerebrale durante il suo svilup-po. I numeri romani indicano i vari strati (da I a VI) della corteccia cerebraleadulta. PC, placca corticale; SB, sostanza bianca; SP, sottile zona presente nellaregione interna della corteccia in via di sviluppo che è detta subplau „.,!, zona in-termedia; ZV, zona ventricolare.
mentre la colinearità spaziale non sareb-be che una conseguenza. Dopo tutto, losviluppo embrionale non è altro che unosvolgimento ordinato di eventi che sisuccedono nel tempo, dirigendo la di-sposizione nello spazio di specifichestrutture biologiche. Possiamo immagi-nare a questo proposito una situazionedel genere: in una regione specifica delnucleo delle nostre cellule si trova ungruppo di geni che costituiscono nel lo-ro complesso una microrappresenta-zione del nostro corpo o, per meglio di-re, del succedersi delle parti principalidel nostro corpo, una sorta di homuncu-lus nascosto nel cromosoma. Al mo-mento opportuno questi geni entrano inazione, l'uno dopo l'altro, con ritmo ca-denzato. Il loro succedersi lineare sulcromosoma si risolve, dapprima, in unaprogressione temporale della loro entra-ta in azione e poi nella successione spa-ziale delle varie parti del corpo. Questamicrorappresentazione del nostro corporacchiude quindi in sé, al tempo stesso,una mappa e un orologio.
1,0 c vti 1PPfl ni- 1 I A TFST'n
Quanto abbiamo sinora raccontato ri-guarda esclusivamente lo sviluppo deltronco, sia nei mammiferi sia negli in-
setti. Il campo d'azione dei geni omeo-tici di Drosophila e dei geni HOX deimammiferi non interessa infatti la testané, a maggior ragione, il cervello. Stu-diare la testa e il suo sviluppo è compli-cato in qualsiasi organismo ed essa ri-mane tuttora una provincia sostanzial-mente sconosciuta anche in Drosophi-la. Tuttavia è stato individuato qualchegene regolatore anche per quanto ri-guarda lo sviluppo della testa. Alcunidi questi geni erano già stati anzi iden-tificati nel quadro dell'ormai famosa ri-cerca sistematica di geni importan-ti nella regolazione dello sviluppocondotta alla fine degli anni settantada Christiane Ntisslein-Volhard ed EricWieschaus e che è valsa loro il Nobelper la fisiologia o la medicina del 1995,condiviso con Edward Lewis, il solita-rio pioniere che ha portato i geni omeo-tici agli onori della ribalta.
In Drosophila bisogna distinguereuna parte anteriore della testa che con-tiene il cervello (non segmentata) e unaparte posteriore (corrispondente grossomodo all'apparato buccale) che si pre-senta segmentata e quindi strettamenteimparentata con il tronco. Almeno tregeni sembrano esercitare nella parte an-teriore della testa e nel cervello in viadi sviluppo un ruolo in certa misura
comparabile a quello esercitato dai geniomeotici nel resto del corpo. Questi ge-ni sono empty spiracles (ems), ortho-denticle (otd) e buttonhead (6td). I pro-dotti genici di questi tre geni sono os-servabili nella regione che darà luogoalla testa in una fase molto precoce del-lo sviluppo embrionale di Drosophila.Il dominio d'azione di ciascuno dei tregeni è rappresentato in questa fase dauna striscia di cellule che corre comeuna cintura tutt'intorno a una porzionespecifica della futura regione cefalica.Non desta quindi sorpresa il fatto cheuna mutazione a carico di ciascuno deitre geni comporti un'alterazione dellastruttura della testa. Questa alterazioneè riconducibile all'assenza di una parti-colare porzione della regione cefalica.
Lo studio di questi geni ha permessodi confermare l'impressione che la logi-ca regolativa inerente allo sviluppo del-la parte anteriore della testa sia diversada quella che si applica allo sviluppodel tronco. Una prima differenza èrappresentata proprio dal fatto che unamutazione in uno dei geni che control-lano lo sviluppo della parte anterioredella testa conduce all'assenza della re-gione corporea corrispondente, mentreabbiamo appena visto che una mutazio-ne in uno dei geni che controllano iltronco comporta una trasformazionedell'identità di un segmento corporeo inquella di un altro segmento adiacente.
Questa differenza sembra corrispon-dere a una diversità più profonda con-nessa con il procedimento astratto uti-lizzato per suddividere una regionedell'organismo in un certo numero disottoregioni diverse. Una possibile stra-tegia è quella di definire per prima cosai limiti e l'estensione della regione,suddividerla poi in un certo numero disottoregioni e specificare infine l'iden-tità di ciascuna di esse. Una secondastrategia è invece quella di specificarel'identità delle varie sottoregioni viavia che la regione si sviluppa e cresce.Un problema che intervenga nel corsodell'attuazione di una di queste duestrategie, produce nel primo caso unatrasformazione di una sottoregione inun'altra, senza perdita di sottoregioni,mentre nel secondo caso comporta lasoppressione e quindi l'assenza di unao più sottoregioni. I geni che controlla-no lo sviluppo della testa del moscerinosono inoltre distinti da quelli che con-trollano lo sviluppo del tronco per ilfatto di trovarsi a loro volta sotto ilcontrollo di una famiglia particolare digeni regolatori che entrano in azione inuna fase ancora precedente dell'em-briogenesi: i cosiddetti geni materniterminali, attivi esclusivamente nelleregioni non segmentate situate alle dueestremità dell'embrione dell'insetto.
LA SCOPERTA DEI GENI CHECONTROLLANO IL CERVELLO
Nel 1990 due dei geni che, in Dro-sophila, controllano lo sviluppo dellatesta, e precisamente ems e otd, furonoisolati e si poté constatare che si tratta-va di due omeogeni. Si ripresentavaquindi l'occasione, già sfruttata consuccesso nel caso dei geni HOX e di al-tre famiglie di omeogeni di cui non ab-biamo potuto parlare, di utilizzare lesequenze geniche isolate in Drosophilaper identificare eventuali omologhi pre-senti nei vertebrati e in particolare neimammiferi. È ciò che decidemmo difare nel 1991 all'Istituto internazionaledi genetica e biofisica del CNR di Na-poli, dove Antonio Simeone isolò neltopo e nell'uomo quattro omeogenistrettamente imparentati con questi duegeni di Drosophila. Due di questi eranochiaramente correlati con ems e venne-ro chiamati Emxl ed Emx2, mentre glialtri due erano correlati con otd e ven-nero chiamati Otxl e Otx2. Geni similisono stati successivamente isolati inuna varietà di altri organismi.
Una volta ancora si è quindi potutodimostrare che un certo numero diomeogeni strutturalmente molto similiesistono nelle specie più diverse e di-stanti fra di loro. Fin qui tutto bene; ma
dov'è che questi quattro omeogeni dimammifero sono attivi, cioè qual è illoro dominio di espressione? Con no-stra grande soddisfazione constatammoche anche nei mammiferi questi genierano attivi nelle regioni cefaliche del-l'embrione, e per giunta proprio nelcervello. Il risultato forse più interes-sante fu poi che uno di questi geni,Emxl, si dimostrò attivo quasi esclusi-vamente nella parte dorsale del prosen-cefalo embrionale, la regione che daràluogo alla corteccia cerebrale.
I domini di espressione dei quattrogeni tratteggiano nel loro complesso unquadro interessante e suggestivo. In unembrione di topo a metà gestazione iquattro domini sono infatti contenutil'uno dentro l'altro come un gioco discatole cinesi. Il dominio di Otx2 è ilpiù grande e contiene quello degli altritre. Subito dopo viene Otxl, il cui do-minio è contenuto in quello di 01x2 econtiene quello degli altri due. Ven-gono poi in sequenza Emx2 e infineEmxl, il cui dominio, come abbiamogià visto, è invece notevolmente ristret-to. I prodotti di Otx2 sono presenti inuna regione cefalica abbastanza vastacomprendente essenzialmente il pro-sencefalo e il mesencefalo embriona-li che individuano nel loro complessouna porzione anteriore del cervello dei
mammiferi, il cervello rostrale. Anchel'attivazione di questi quattro geni nel-lo sviluppo embrionale è sequenziale:Obc2 è espresso per primo, seguito daOtxl, da Emx2 e infine da Emxl.
Questa particolare combinazione diprofili di espressione, che si presenta-no in sequenza sia nello spazio sia neltempo, suggerisce un possibile ruolodei quattro geni nella specificazionedelle varie aree del cervello rostrale.Possiamo infatti immaginare che i pro-dotti di Otx2 dettino le prime istruzio-ni per conferire alle cellule della regio-ne corrispondente al cervello rostralela loro identità regionale e delimitarecosì la regione rispetto al resto del si-stema nervoso centrale. Agendo an-che sulla base di queste prime indica-zioni biologiche, i prodotti del secon-do gene, Otxl, potrebbero poi specifi-care una particolare sottoregione al-l'interno di questa regione e analoga-mente i prodotti del terzo gene, Emx2,potrebbero specificare una sotto-sot-toregione del cervello rostrale corri-spondente a una parte del prosence-falo. I prodotti di Emxl, infine, potreb-bero giungere a completare il quadrodi questa determinazione sequenzia-le, specificando univocamente la futuracorteccia cerebrale e le regioni imme-diatamente adiacenti.
52 LE SCIENZE n. 346, giugno 1997
LE SCIENZE n. 346, giugno 1997 53

RACCOGLITORI 1997LE SCIENZE
SCIENTIFICAMERICAN
Sono disponibili i raccoglitori 1997 (volumi LVIII e LIX)per i fascicoli da gennaio (n. 341) a giugno (n. 346)e da luglio (n. 347) a dicembre (n. 352).
Sono ancora disponibili i raccoglitoridal volume L al volume LVII (1993-1996)e raccoglitori non numerati.I raccoglitori si possono richiedere direttamente all'editoreusando l'apposita cartolina allegata alla rivistae versando il relativo importo.Gli ordini vengono evasi solo a pagamento avvenuto.
I raccoglitorisi trovano anche pressoi seguenti punti vendita:
BOLOGNA - Libreria Parolini -Via U. Bassi 14
FIRENZE - Libreria Marzocco -Via de Martelli 221R
GENOVA - Libreria Di Stefano- Via R. Cecc,ardi 40/R
MILANO - Le Scienze S.p.A. -Piazza della Repubblica 8
NAPOLI - Libreria Guida A. -Via PortAlba 20/21
PADOVA- Libreria Cortina -Via F. Marzolo 4
PALERMO - Libreria Dante -Quattro Canti di Città
ROMA - Libreria Feltrinelli -Via V. Emanuele Orlando 78
TORINO - Libreria Zanaboni -Corso Vittorio Emanuele 41
iimisei~~"
9ge tezzoyditine 2. 10.000
rana
LVIIIGAMOW. I Or,mrsime es,r7
In conclusione, i quattro geni di cuistiamo parlando sembrano parteciparein maniera coordinata ai processi checonducono alla regionalizzazione delcervello rostrale e alla specificazionedell'identità cellulare e sovracellularedelle varie aree cerebrali. Abbiamo vi-sto che i geni HOX interpretano un ruo-lo in certa misura corrispondente nel re-gionalizzare il midollo spinale e ilromboencefalo. Non sappiamo ancoraquante altre famiglie geniche esistanoche contribuiscono a regolare lo svilup-po di regioni e sottoregioni del sistemanervoso centrale, ma già possiamo indi-viduare almeno due grandi sistemi diquesto tipo: i geni HOX e il complessodei geni Otx e Emx.
A questa ripartizione, che potremmodefinire anatomo-molecolare del siste-ma nervoso centrale, è molto probabilefaccia riscontro un'analoga ripartizio-ne delle corrispondenti regioni del restodel corpo. Rimane fuori da questo sche-ma una ristretta regione embrionale checorrisponde approssimativamente al fu-turo cervelletto e alle regioni posterio-ri del mesencefalo adulto. Questa regio-ne non appartiene al dominio dei ge-ni HOX e appartiene solo in manieramarginale e transitoria a quello di Obc2.Sembra sempre più probabile che sitratti di una regione particolare che se-gue schemi di sviluppo del tutto pe-culiari e comunque diversi sia da quel-li che valgono per il cervello vero eproprio sia da quelli che valgono per leregioni posteriori del sistema nervosocentrale.
MECCANISMI DI REGOLAZIONIDELLA CORTECCIA CEREBRALE
Fin dal momento della loro pri-ma identificazione i geni Emx sonosembrati particolarmente interessanti.Emxl è espresso quasi esclusivamentenella regione della corteccia cerebralementre Emx2 è espresso in una regio-ne prosencefalica appena un po' piùampia. È molto difficile che un generegolatore sia attivo soltanto in unaspecifica regione del corpo perché igeni regolatori sono necessari un po'dappertutto, per compiere magari fun-zioni un po' diverse nei diversi tessuti.I due geni Emx sono tra i pochissimiche sono attivi soltanto in una regioneristretta del corpo e nel caso particola-re del cervello. Per questo motivo èsembrato fondamentale analizzare indettaglio l'espressione di questi duegeni nella corteccia cerebrale in via disviluppo.
Quello che si è trovato è che durantelo sviluppo embrionale Emx2 è attivosolo nei precursori dei neuroni corticaliche si stanno replicando all'interno del-
la zona germinativa ventricolare, men-tre è completamente muto nei neuroniche hanno smesso di dividersi e si van-no differenziando. Viene naturale sup-porre che l'azione di questo gene sia ri-chiesta per regolare la moltiplicazionedi queste cellule e che non sia invecepiù richiesta quando tale processo dimoltiplicazione è concluso. La moltipli-cazione delle cellule della corteccia de-ve essere accuratamente regolata. Il nu-mero finale di neuroni corticali non de-ve essere troppo basso, altrimenti addiocomplessità architettonica e ricchezzadi connessioni, ma neppure troppo alto,perché ogni singolo neurone deve di-sporre di un suo proprio spazio per dif-ferenziarsi, maturare e stabilire i suoicontatti. Inoltre, poiché abbiamo vistoche la fase di replicazione di queste cel-lule ne regola indirettamente anche lasuccessiva migrazione, un'alterazionenel controllo di quella potrebbe risultarein un difetto di questa e quindi in unaperturbazione nell'architettura comples-siva della corteccia.
Per avere un quadro più preciso dellefunzioni di Emx2 è necessario andare avedere che cosa succede in un organi-smo in cui questo gene non operi nelladovuta maniera, cioè in un organismoche porti una mutazione in questo gene.Una strategia sperimentale molto po-tente è quella di produrre artificialmen-te dei ceppi di topi mancanti dei pro-dotti di questo gene. I primi dati con-fermano che l'azione di Emx2 è effetti-vamente richiesta per il corretto svilup-po della corteccia cerebrale ed è addi-rittura critica per lo sviluppo della re-gione corticale corrispondente all'ippo-campo. I topi mutanti in questione han-no infatti una corteccia di dimensioniridotte, sia per quanto riguarda l'esten-sione sia per lo spessore, una regioneippocampale mal definita e mancanocompletamente del giro dentato, unastruttura facilmente riconoscibile loca-lizzata in questa regione.
In considerazione della complessi-tà delle strutture interessate, l'analisidettagliata delle caratteristiche anato-mo-funzionali di questi topi mutanti ri-chiederà un certo tempo. Nel frattempoperò il caso ha voluto che si riuscisse aidentificare una malformazione conge-nita umana nella quale è mutato Emx2.Questo è stato possibile grazie a unastretta collaborazione del mio gruppocon l'équipe di Armando Cama, del-l'Ospedale Gaslini di Genova. Analiz-zando il patrimonio genetico di un cer-to numero di individui portatori di undifetto congenito della corteccia, chia-mato schizoencefalia, abbiamo scoper-to che la maggioranza di questi portavauna mutazione nel gene Emx2. La schi-zoencefalia è una malformazione mol-
to rara, caratterizzata dalla presenza diveri e propri buchi nella corteccia cere-brale. A seconda dei casi questi buchipossono essere di dimensioni ridotteoppure interessare una grossa porzionedella superficie cerebrale; conseguen-temente il quadro clinico può andaredalla saltuaria comparsa di modestecrisi epilettiche a una grave compro-missione delle capacità intellettive emotorie.
UNA IPOTESI EVOLUTIVA
Queste osservazioni sembrano quindiconfermare l'ipotesi che, nella specieumana, Emx2 controlli la moltiplicazio-ne delle cellule nervose della cortecciae fanno sperare di poter arrivare a dimo-strare il coinvolgimento di questo geneo di altri strettamente correlati in feno-meni patologici più diffusi e più social-mente rilevanti, come l'idrocefalia, i tu-mori cerebrali o l'epilessia.
È interessante considerare infine unaspetto dell'azione di Emx2 o di altrigeni di questo tipo connesso con l'evo-luzione del cervello. Si ritiene oggi chemolti cambiamenti evolutivi siano ri-conducibili a un'alterata regolazio-ne dell'espressione di alcuni geni giàesistenti, piuttosto che alla compar-sa sulla scena biologica di geni com-pletamente nuovi. Questo sembra esse-re vero soprattutto per un organo comeil cervello che si è evoluto con un rit-mo particolarmente accelerato. Vale lapena quindi di domandarsi che cosasuccederebbe se un gene che controllala proliferazione dei neuroni corticalicome Emx2 si disattivasse con un certoritardo in una nuova specie rispetto auna già esistente. Si può assumere chequesta nuova specie avrebbe più neu-roni corticali della precedente e con-seguentemente una corteccia più riccadi cellule. Questo fatto di per sé nonconferirebbe all'animale una capacitàmentale superiore, ma certamente po-trebbe rappresentare un primo passo inquesta direzione. Se nello stesso tempoun certo numero di altri geni - è diffici-le dire quanti - cambiasse le sue abitu-dini, si potrebbe cominciare a pensarea un fenomeno evolutivo di una certarilevanza.
Questa per ora è solo un'ipotesi, masi ha l'impressione che i tempi possanoessere maturi per cominciare a gettareuno sguardo su questi affascinanti pro-cessi. Emx2 è un gene molto antico,presente in tutti i vertebrati che sonostati analizzati; è concepibile che unapiccola alterazione nella sua regolazio-ne possa avere avuto conseguenze rile-vanti sulla formazione della corteccia equindi sulla sua struttura.
Ed Emxl? Della funzione di questo
gene, di origine più recente e quasicertamente derivato da Emx2 per unevento di duplicazione genica e succes-siva differenziazione, si sa ancora po-co, ma si intuisce che il suo ruolo possaessere ancora più sottile e specifico diquello di Emx2. A differenza di Emx2,Emxl è attivo infatti nei neuroni dellacorteccia cerebrale, e in quelli soli, inogni momento della loro vita: prima inquelli che si stanno replicando, poi inquelli che stanno migrando e differen-ziandosi e infine in quelli che si sonogià differenziati e che sono quindi ma-turi e funzionanti all'interno di un cer-vello adulto. Riesce difficile immagina-re quale ruolo possa esercitare lo stessogene in cellule impegnate in fasi cosidiverse della loro esistenza. Una possi-bilità è che esso contribuisca a specifi-care l'identità delle cellule della cortec-cia, distinte per esempio da quelle deigangli basali che si trovano nelle regio-ni ventrali dei due emisferi e che svol-gono funzioni ben diverse. È comun-que certo che Emx I appare al momentocome il gene della corteccia per eccel-lenza. Il futuro ci dirà se esso concorrain modo essenziale a determinare lestraordinarie proprietà della parte piùevoluta del nostro cervello.
EDOARDO BONCINELLI, direttoredi ricerca del CNR presso il Centro perlo studio della farmacologia cellulare emolecolare, è capo del Laboratorio dibiologia molecolare dello sviluppo al-l'Istituto scientifico Ospedale San Raf-faele di Milano. Fino al 1991 ha lavora-to all'Istituto internazionale di geneticae biofisica di Napoli, dove ha percorsole tappe fondamentali della sua carrieradi genetista e biologo molecolare.
KANDEL E. e SCHWARTZ i., Principidi neuroscienze, Casa Editrice Ambro-siana, 1988.
SIMEONE A., ACAMPORA D., GULISANOM., STORNAIUOLO A. e BONCINELLI E.,Nested Expression Domains of FourHomoeobox Genes in the DevelopingRostral Brain in «Nature», 358, pp.687-690, 1992.
CHANGEUX 1.-P., L'uomo neuronale,Feltrinelli, 1993.
BONCINELLI E., Biologia dello svilup-po, La Nuova Italia Scientifica, 1994.
BRUNELLI S., FAIELLA A., CAPRA V., NI-GRO V., SIMEONE A., CAMA A. e BONCI-NELLI E., Germline Mutations in theHomeobox Gene Emx2 in Patients withSevere Schizencephaly in «Nature Ge-netics», 12, pp. 94-96, 1996.
54 LE SCIENZE n. 346, giugno 1997LE SCIENZE n. 346, giugno 1997 55