Embed Size (px)
Citation preview

El metabolismo es el conjunto de reacciones químicas que ocurren en un organismo. En todo
momento en una célula ocurren miles de reacciones químicas, de manera muy rápida y ordenada,
ajustada a las necesidades que tenga la célula en cada instante. Estas reacciones sólo son
posibles a temperatura ambiente gracias a las enzimas.
Un compuesto que penetra en una célula experimenta una serie de reacciones químicas sucesivas,
cada una controlada por una enzima diferente. Estas secuencias ordenadas de reacciones son las
rutas metabólicas. Cada reacción de una ruta implica un pequeño cambio, así no se provocan
alteraciones graves del medio celular que impedirían el mantenimiento de la estructura de las
macromoléculas.
En algunas rutas metabólicas los sustratos que inician las reacciones siguen una secuencia lineal
hasta llegar a los productos finales. Otras rutas siguen una secuencia cíclica, de manera que los
productos de la última reacción son también sustratos de la reacción inicial. En estos casos los
verdaderos productos finales son compuestos intermediarios.
La energía y las reacciones químicas
Un organismo, una célula o un extracto celular pueden considerarse como un sistema físico que
debe cumplir las leyes de la termodinámica:
La primera ley o de conservación de la energía establece que la energía total de un sistema
y su entorno es constante. En otras palabras, la energía ni se crea ni se destruye; sólo se
transforma.
La segunda ley o de aumento de la entropía (medida del grado de desorden o de azar)
establece que un proceso tiene lugar espontáneamente si aumenta la suma de las entropías
del sistema y de su entorno.
Aunque los seres vivos parecen negar el segundo principio de la termodinámica, ya que son
estructuras ordenadas, en realidad sí que lo cumplen, ya que, al mismo tiempo que ellos se
ordenan, contribuyen en mayor medida al aumento de desorden en el entorno que los rodea,
fundamentalmente, desprendiendo calor.
Como resulta muy difícil medir el grado de desorden del entorno, Gibbs creó la función de
energía libre (G), al combinar las dos leyes de la termodinámica. G es una medida de la
capacidad de un sistema para producir trabajo útil (energía potencial), a presión y temperatura
constantes. La ventaja de G es que permite predecir si una reacción química puede ocurrir
espontáneamente o no:
Sólo puede ser espontánea una reacción exergónica, es decir, aquélla en que la variación de
energía libre es negativa ( G<0).
Una reacción está en equilibrio, es decir, no varía la concentración de sustratos y productos,
si no hay variación de energía libre ( G=0). Si se modifican levemente las condiciones de la
reacción en cualquier sentido, ésta tenderá a regresar al equilibrio, por lo que se comporta
como reversible.
No son espontáneas las reacciones endergónicas, es decir, aquéllas en que la variación de energía
libre es positiva ( G>0). La tendencia espontánea será la producción de la reacción opuesta.

La variación de energía libre es independiente de los pasos intermedios que se hayan
podido producir; sólo depende de los estados inicial y final. Por esta razón, en el
metabolismo pueden ocurrir reacciones endergónicas siempre y cuando vayan acopladas a
reacciones muy exergónicas, de manera que el balance global sea exergónico.
Casi todas las reacciones químicas celulares son reversibles, en el sentido de que pueden
transcurrir en los dos sentidos: desde los sustratos a los productos y al revés. Sólo aquellas
reacciones que están muy alejadas del equilibrio o cuyas condiciones celulares les impiden
aproximarse a él, se comportan como si fueran irreversibles.
El G, para que se produzca una reacción, depende de cuál sea la situación de equilibrio a la que
tiendan reactivos y productos, y de las concentraciones a las que se encuentren inicialmente. Si
en un momento determinado la relación de concentraciones de reactivos a productos es mucho
mayor que en el equilibrio, tenderá a producirse (será exergónica) la transformación de reactivos
en productos. Inversamente, si dicha relación es mucho menor que en el equilibrio, entonces el
proceso espontáneo sería la transformación de productos en reactivos.
La moneda energética: importancia del ATP
Las células contienen diversos compuestos que pueden experimentar reacciones muy exergónicas,
capaces de impulsar procesos que requieran energía. El más importante de estos compuestos es
el ATP (adenosin trifosfato). Una molécula de ATP está formada por una ribosa que lleva unidos,
por un lado, una adenina y, por otro lado, una cadena de tres grupos fosfato.
Para la célula, el ATP es una moneda de energía. Cuando una célula obtiene energía del exterior,
bien a partir de la luz o bien por oxidación de moléculas que toma del exterior, utiliza parte de esa
energía para formar ATP a partir del ADP y fosfato inorgánico. Cuando la célula necesita realizar
algún proceso endergónico, lo acopla a la hidrólisis del ATP para dar ADP y fosfato inorgánico.
Muchas veces se dice que los enlaces fosfato del ATP son de alta energía: en realidad la gran
energía que se libera de la hidrólisis es consecuencia de las propiedades de sustratos y productos
de la reacción, que la hacen estar muy desplazada en el equilibrio hacia la ruptura de ese enlace.
Figura 1. Esquema de la formación e hidrólisis del ATP y acoplamiento de reacciones endergónicas con exergónicas.
Glucosa
CO2 + H2O
Proteínas
Aminoácidos
Reacciones
exergónicas
ATP
A
A
P P P
Reacciones
endergónicas
P P
Proceso
energéticamente
ADP
+ P
Bajo
Con
tenido
ene
rgético
de las
molécu
las
Alto

Las enzimas que catalizan la hidrólisis del ATP reciben el nombre general de ATPasas y participan en
procesos como movimientos celulares o intercambios de sustancias a través de la membrana.
Para activar un compuesto, para que aumente su energía útil y pueda hacer reacciones que, por sí
solo, serían endergónicas, se le suele transferir un grupo fosfato procedente de la hidrólisis del ATP.
Este proceso se denomina fosforilación. El compuesto, al resultar fosforilado, recibe parte de la
energía almacenada en el ATP y queda más reactivo. Este mecanismo es típico de reacciones de
condensación, que resultarían muy endergónicas si tuvieran que juntarse las moléculas en su
estado normal.
El acoplamiento de reacciones endergónicas con reacciones exergónicas (Figura 1) es uno
de los procesos bioquímicos más importantes, ya que sirve para impulsar muchas reacciones,
transportar sustancias a través de membranas o provocar movimientos.
2. CATABOLISMO Y ANABOLISMO
El metabolismo puede estudiarse como la suma de dos
conjuntos de reacciones (figura 2):
El catabolismo es el conjunto de todas las vías
metabólicas en que los compuestos orgánicos se
degradan. Se pasa de polímeros a monómeros y de
monómeros a compuestos orgánicos más pequeños
o inorgánicos. Son reacciones exergónicas.
En general, estas reacciones implican la rotura de
enlaces covalentes y la oxidación de compuestos.
La energía que se desprende en estos procesos es
utilizada por la célula para fabricar intermediarios
de alta energía, principalmente, en forma de ATP o
de mediadores reducidos (NADH, NADPH, etc.).
El anabolismo es el conjunto de procesos
endergónicos de biosíntesis, opuestos a los del
catabolismo. Utiliza la energía obtenida en las rutas
del catabolismo o en los procesos de captación de
energía luminosa. Estas reacciones incluyen la
formación de enlaces covalentes y la reducción de
moléculas. A partir de moléculas inorgánicas o
moléculas orgánicas sencillas, se sintetizan
monómeros, y a partir de ellos los polímeros.
Figura 2. Esquema global del anabolismo y catabolismo.
3. ENZIMAS
Las enzimas actúan como biocatalizadores que aceleran las reacciones químicas dentro de la
célula sin transformarse ellas mismas en una molécula diferente.
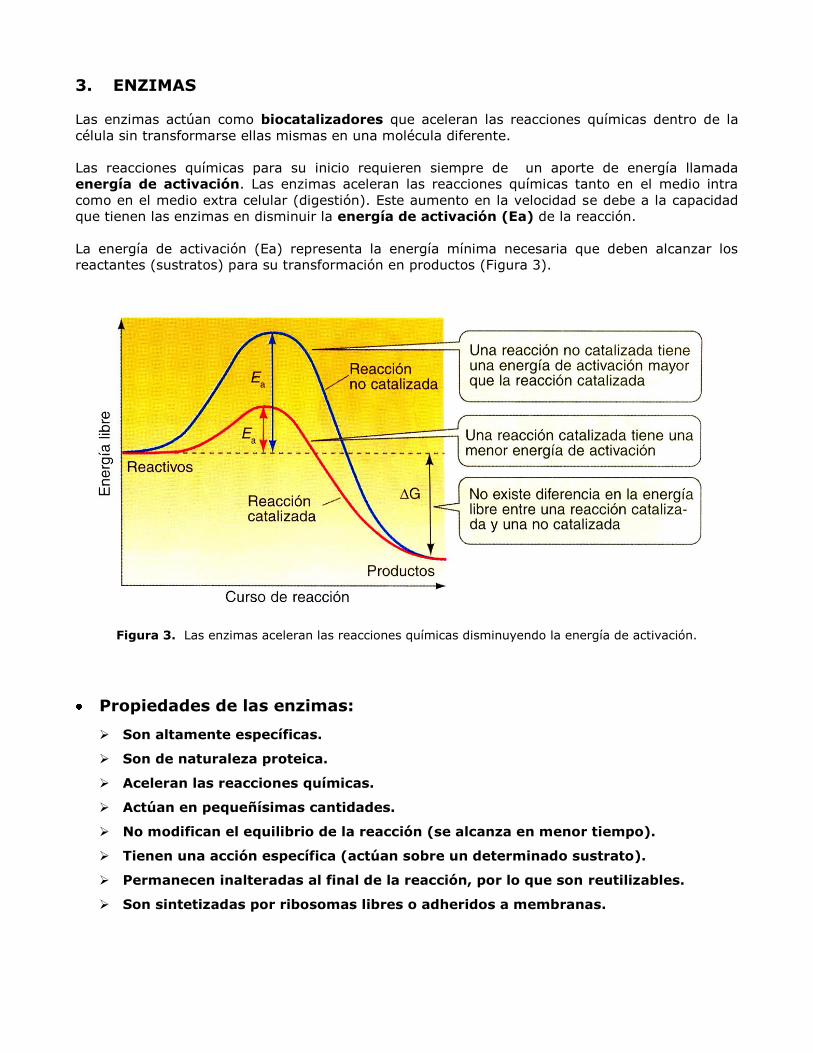
Las reacciones químicas para su inicio requieren siempre de un aporte de energía llamada
energía de activación. Las enzimas aceleran las reacciones químicas tanto en el medio intra
como en el medio extra celular (digestión). Este aumento en la velocidad se debe a la capacidad
que tienen las enzimas en disminuir la energía de activación (Ea) de la reacción.
La energía de activación (Ea) representa la energía mínima necesaria que deben alcanzar los
reactantes (sustratos) para su transformación en productos (Figura 3).
Figura 3. Las enzimas aceleran las reacciones químicas disminuyendo la energía de activación.
Propiedades de las enzimas:
Son altamente específicas.
Son de naturaleza proteica.
Aceleran las reacciones químicas.
Actúan en pequeñísimas cantidades.
No modifican el equilibrio de la reacción (se alcanza en menor tiempo).
Tienen una acción específica (actúan sobre un determinado sustrato).
Permanecen inalteradas al final de la reacción, por lo que son reutilizables.
Son sintetizadas por ribosomas libres o adheridos a membranas.
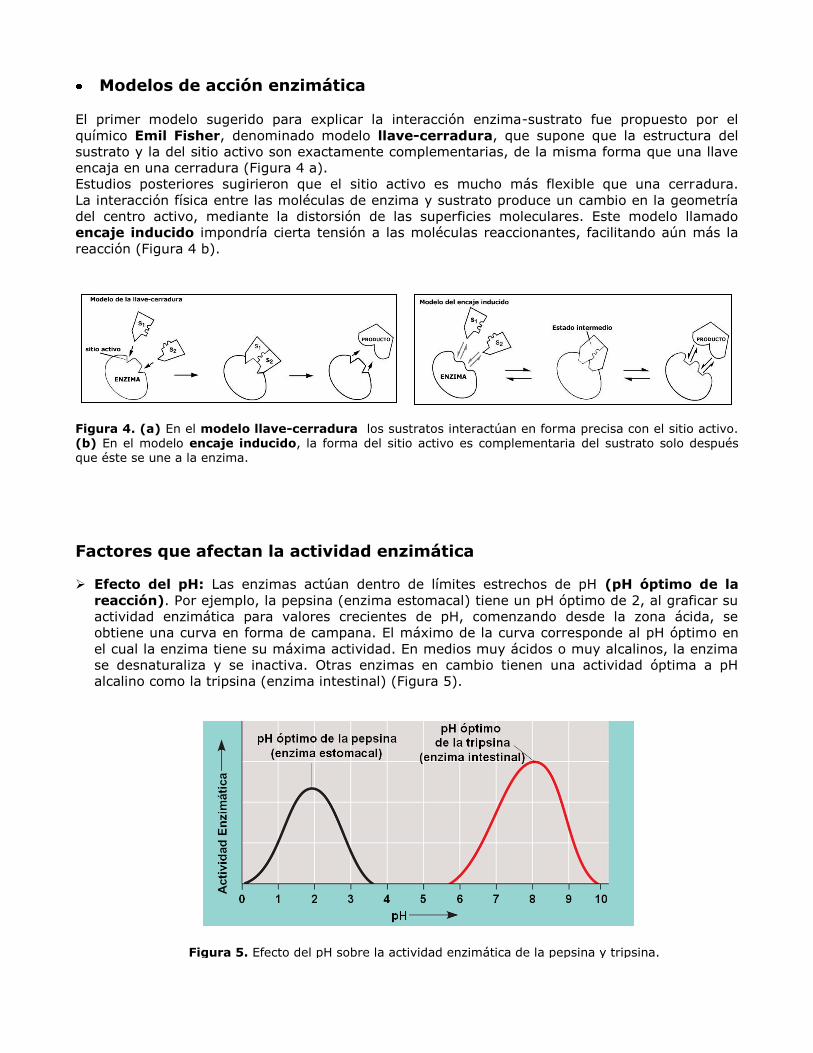
Modelos de acción enzimática
El primer modelo sugerido para explicar la interacción enzima-sustrato fue propuesto por el
químico Emil Fisher, denominado modelo llave-cerradura, que supone que la estructura del
sustrato y la del sitio activo son exactamente complementarias, de la misma forma que una llave
encaja en una cerradura (Figura 4 a).
Estudios posteriores sugirieron que el sitio activo es mucho más flexible que una cerradura.
La interacción física entre las moléculas de enzima y sustrato produce un cambio en la geometría
del centro activo, mediante la distorsión de las superficies moleculares. Este modelo llamado
encaje inducido impondría cierta tensión a las moléculas reaccionantes, facilitando aún más la
reacción (Figura 4 b).
Figura 4. (a) En el modelo llave-cerradura los sustratos interactúan en forma precisa con el sitio activo. (b) En el modelo encaje inducido, la forma del sitio activo es complementaria del sustrato solo después que éste se une a la enzima.
Factores que afectan la actividad enzimática
Efecto del pH: Las enzimas actúan dentro de límites estrechos de pH (pH óptimo de la
reacción). Por ejemplo, la pepsina (enzima estomacal) tiene un pH óptimo de 2, al graficar su
actividad enzimática para valores crecientes de pH, comenzando desde la zona ácida, se
obtiene una curva en forma de campana. El máximo de la curva corresponde al pH óptimo en
el cual la enzima tiene su máxima actividad. En medios muy ácidos o muy alcalinos, la enzima
se desnaturaliza y se inactiva. Otras enzimas en cambio tienen una actividad óptima a pH
alcalino como la tripsina (enzima intestinal) (Figura 5).
Figura 5. Efecto del pH sobre la actividad enzimática de la pepsina y tripsina.
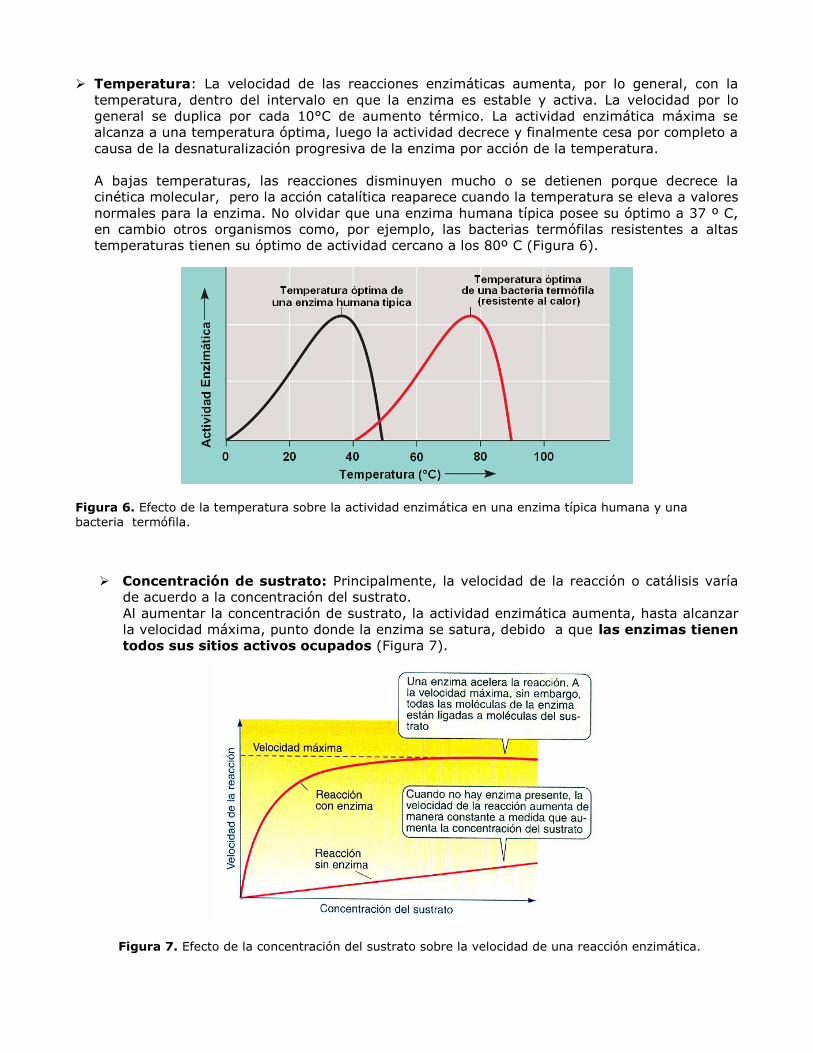
Temperatura: La velocidad de las reacciones enzimáticas aumenta, por lo general, con la
temperatura, dentro del intervalo en que la enzima es estable y activa. La velocidad por lo
general se duplica por cada 10°C de aumento térmico. La actividad enzimática máxima se
alcanza a una temperatura óptima, luego la actividad decrece y finalmente cesa por completo a
causa de la desnaturalización progresiva de la enzima por acción de la temperatura.
A bajas temperaturas, las reacciones disminuyen mucho o se detienen porque decrece la
cinética molecular, pero la acción catalítica reaparece cuando la temperatura se eleva a valores
normales para la enzima. No olvidar que una enzima humana típica posee su óptimo a 37 º C,
en cambio otros organismos como, por ejemplo, las bacterias termófilas resistentes a altas
temperaturas tienen su óptimo de actividad cercano a los 80º C (Figura 6).
Figura 6. Efecto de la temperatura sobre la actividad enzimática en una enzima típica humana y una
bacteria termófila.
Concentración de sustrato: Principalmente, la velocidad de la reacción o catálisis varía
de acuerdo a la concentración del sustrato.
Al aumentar la concentración de sustrato, la actividad enzimática aumenta, hasta alcanzar
la velocidad máxima, punto donde la enzima se satura, debido a que las enzimas tienen
todos sus sitios activos ocupados (Figura 7).
Figura 7. Efecto de la concentración del sustrato sobre la velocidad de una reacción enzimática.

Sustratos
Enzima Estado de
transición
Producto
A
B
C
Complejo enzima -sustrato
ACTIVIDAD 1
CONTESTE
1. La figura representa una reacción enzimática
a. Si el producto C es sacarosa, qué moléculas serán A y B
..........................................................................................................................
b. ¿Esta reacción enzimática corresponde a una hidrólisis o a una síntesis por
deshidratación?
...........................................................................................................................
c. ¿Por qué la misma enzima no puede catalizar la reacción de izquierda a derecha?
...........................................................................................................................
..........................................................................................................................
..........................................................................................................................
2. El gráfico presenta la actividad relativa de cuatro enzimas a distinto pH
a. ¿Qué ocurre con la conformación espacial de las enzimas;pepsina, tripsina y colinesterasa
entre pH 4 y 6?
..........................................................................................................................
b. A partir de pH 7, ¿la colinesterasa detiene su acción?
..........................................................................................................................
c. ¿Afecta a la papaina la variación de pH entre 4 y 10?
..........................................................................................................................
d. ¿Qué enzima tiene su óptimo a pH ácido?
..........................................................................................................................
e. En el gráfico, ¿cuál es la variable independiente y cuál es la variable independiente?
.........................................................................................................................
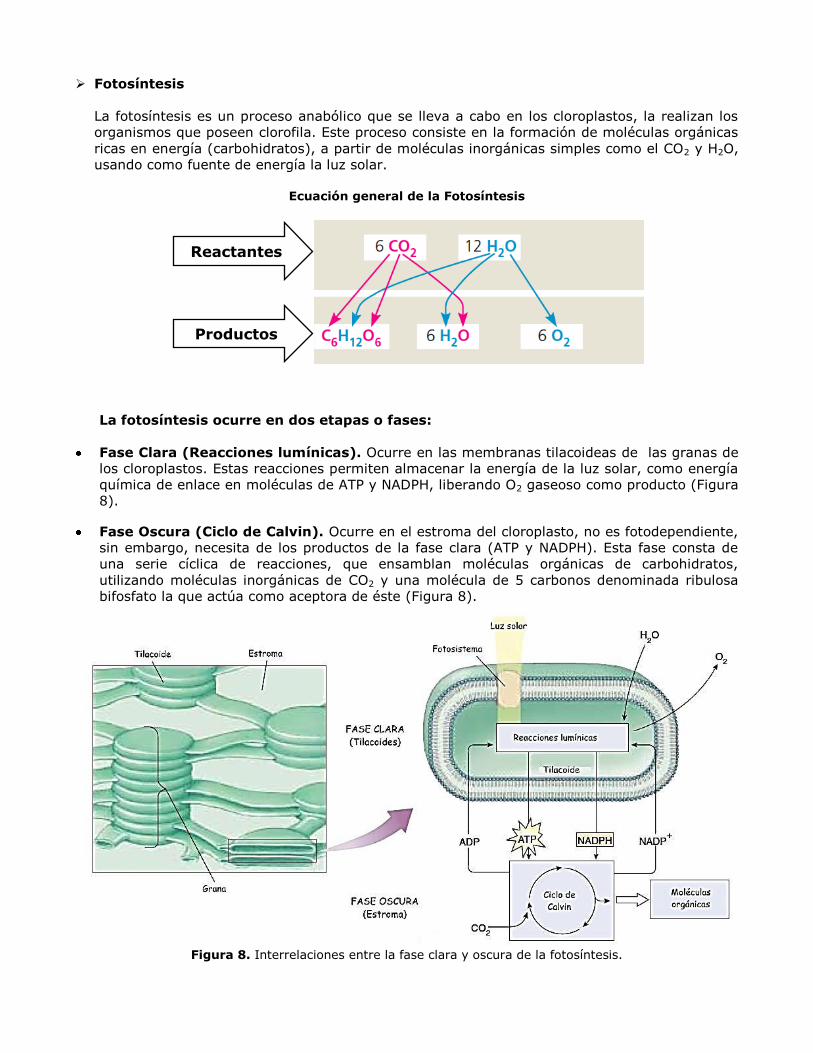
Fotosíntesis
La fotosíntesis es un proceso anabólico que se lleva a cabo en los cloroplastos, la realizan los
organismos que poseen clorofila. Este proceso consiste en la formación de moléculas orgánicas
ricas en energía (carbohidratos), a partir de moléculas inorgánicas simples como el CO2 y H2O,
usando como fuente de energía la luz solar.
Ecuación general de la Fotosíntesis
La fotosíntesis ocurre en dos etapas o fases:
Fase Clara (Reacciones lumínicas). Ocurre en las membranas tilacoideas de las granas de
los cloroplastos. Estas reacciones permiten almacenar la energía de la luz solar, como energía
química de enlace en moléculas de ATP y NADPH, liberando O2 gaseoso como producto (Figura
8).
Fase Oscura (Ciclo de Calvin). Ocurre en el estroma del cloroplasto, no es fotodependiente,
sin embargo, necesita de los productos de la fase clara (ATP y NADPH). Esta fase consta de
una serie cíclica de reacciones, que ensamblan moléculas orgánicas de carbohidratos,
utilizando moléculas inorgánicas de CO2 y una molécula de 5 carbonos denominada ribulosa
bifosfato la que actúa como aceptora de éste (Figura 8).
Figura 8. Interrelaciones entre la fase clara y oscura de la fotosíntesis.
Reactantes
Productos
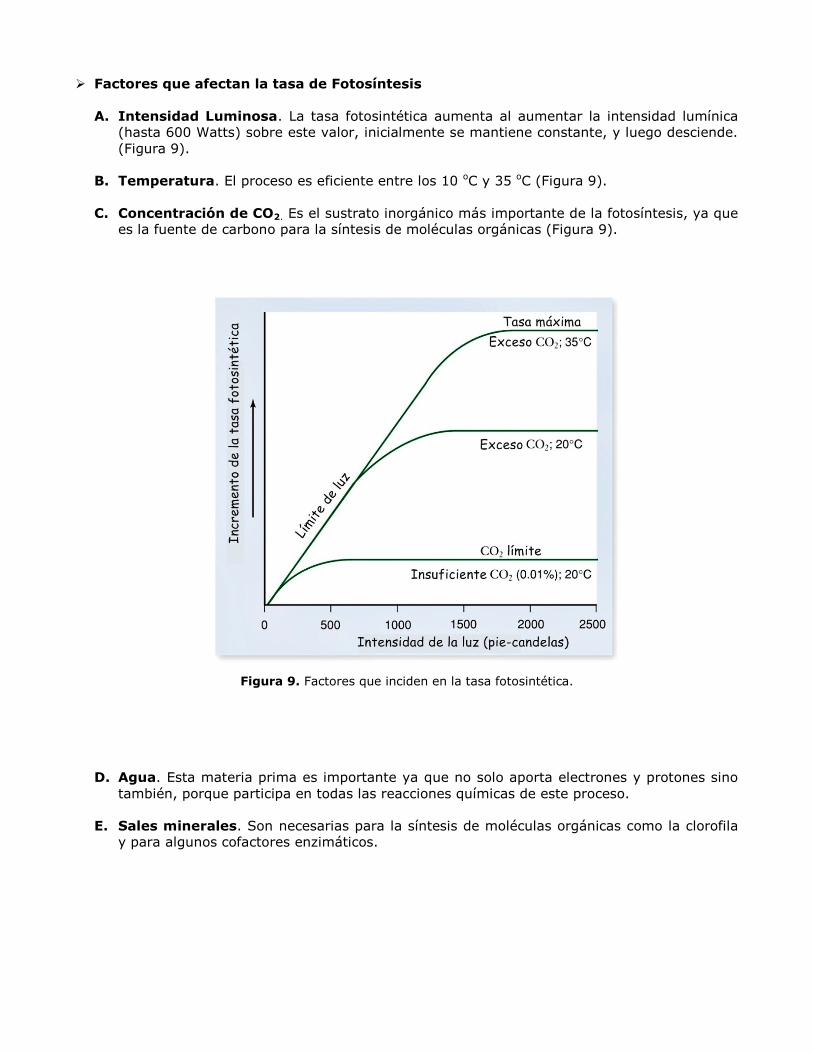
Factores que afectan la tasa de Fotosíntesis
A. Intensidad Luminosa. La tasa fotosintética aumenta al aumentar la intensidad lumínica
(hasta 600 Watts) sobre este valor, inicialmente se mantiene constante, y luego desciende.
(Figura 9).
B. Temperatura. El proceso es eficiente entre los 10 oC y 35 oC (Figura 9).
C. Concentración de CO2. Es el sustrato inorgánico más importante de la fotosíntesis, ya que
es la fuente de carbono para la síntesis de moléculas orgánicas (Figura 9).
Figura 9. Factores que inciden en la tasa fotosintética.
D. Agua. Esta materia prima es importante ya que no solo aporta electrones y protones sino
también, porque participa en todas las reacciones químicas de este proceso.
E. Sales minerales. Son necesarias para la síntesis de moléculas orgánicas como la clorofila
y para algunos cofactores enzimáticos.

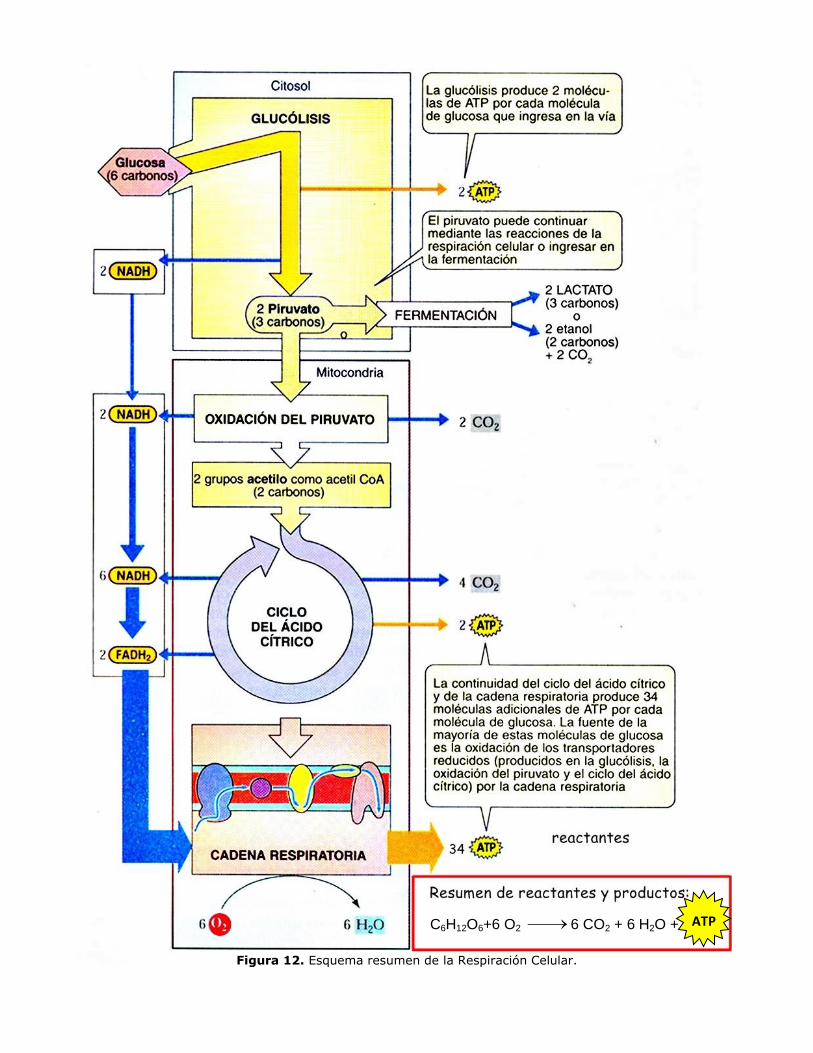
RESPIRACIÓN CELULAR: CATABOLISMO DE LA GLUCOSA
Las células almacenan glúcidos y lípidos de manera especial para poder utilizarlos, en caso
necesario, como reservas energéticas. Los glúcidos se almacenan en forma de grandes inclusiones
de polisacáridos y los lípidos en forma de gotas de triacilglicéridos. A partir de ellos se producen
las rutas de oxidación en las que los primeros tramos son específicos y los últimos son comunes.
La oxidación de los polisacáridos comienza con la despolimerización para obtener unidades
sencillas de monosacáridos. Estos últimos se degradan hasta constituir una molécula más sencilla
y más oxidada denominada ácido pirúvico. Esta ruta, denominada glucólisis, es común a la
mayor parte de los organismos, y en algunos que viven en ausencia de oxígeno (anaerobios). La
oxidación no continúa; en su lugar, el ácido pirúvico se modifica ligeramente. Estos procesos se
denominan fermentaciones.
En los organismos que viven en medios con oxígeno (aerobios) el ácido pirúvico se transforma en
acetil-coenzima A. El acetil-coenzima A procedente de la glucólisis pasa a un proceso cíclico
denominado ciclo de Krebs, durante el cual los dos carbonos de los grupos acetilo son
totalmente oxidados, y en esta transformación se origina dióxido de carbono.
Durante el ciclo de Krebs y en las rutas anteriores se desprenden protones y electrones, los cuales
son capturados por diversos coenzimas que se reducen y posteriormente se vuelven a oxidar
mediante una serie de reacciones redox acopladas que concluyen con la transferencia de los
electrones al oxígeno, que al tomar los dos protones (H+) forma agua. La energía desprendida
durante esta reacción se utiliza para sintetizar ATP a partir de ADP y Pi.
Etapas de la Respiración Celular Aeróbica: C6 H12 O6 + 6O2 6CO2 + 6H2O + E
Las múltiples reacciones que ocurren en la célula se organizan en los siguientes pasos;
Glucólisis, oxidación del piruvato, (formación del acetil CoA), ciclo de Krebs, transporte
de electrones y fosforilación oxidativa y se detallan en el resumen (Ver página 13). El
transporte de electrones y fosforilación oxidativa se revisan en el ANEXO página 14).
Figura 10. Respiración aerobia
ATP
calor

FERMENTACIÓN: La alternativa anaeróbica a la respiración aeróbica.
Las células musculares normalmente utilizan, la respiración aeróbica, obteniendo 38 ATP por
molécula de glucosa, pero también son capaces de sobrevivir sin O2, con las dos moléculas de ATP
de la glucólisis. El inconveniente de utilizar esta vía, está en el suministro de NAD (oxidado), que
debe ser capaz de reponer el NADH (reducido).
Las células musculares mantienen el suministro de NAD (oxidado), a costa de la reducción del
ácido pirúvico obtenido en la glucólisis. De esta manera, el ácido pirúvico queda como ácido
láctico (lactato). La producción de ácido láctico a partir de la glucosa se denomina
fermentación láctica.
Las levaduras también utilizan normalmente la vía aeróbica, pero son capaces de vivir en
ambientes sin oxígeno, realizando la fermentación alcohólica, en la cual produce etanol y
libera CO2.
Figura 11. Reacciones químicas de la Fermentación. A. Fermentación Acido Láctica. B. Fermentación Alcohólica.
Tabla 1. Rendimiento energético de la oxidación completa de la glucosa a CO2 y H2O. Se sintetizan 3 moléculas de ATP, al oxidar una molécula de NADH y 2 moléculas de ATP al oxidar una molécula de FADH2.
ETAPAS TRANSPORTADORES REDUCIDOS NÚMERO DE ATP
Glucólisis 2 NADH 2 ATP
Formación de Acetil CoA 2 NADH Ciclo de Krebs 6 NADH
2 FADH2 2 ATP
Fosforilación Oxidativa 34 ATP
Total : 38 ATP
A B
Glucólisis Glucólisis
Reacciones de
fermentación
Reacciones de
fermentación
C6H12O6+6 O2 6 CO2 + 6 H2O + ATP
Figura 12. Esquema resumen de la Respiración Celular.

ACTIVIDAD 2
Respiración celular aeróbica.
a) Sobre la glucólisis:
¿Dónde se realiza?:……………………………………………………………………………………………………………………
¿Necesita O2? …………………………………………………………………………………………………………………………….
¿Cuáles son sus productos?………………………………………………………………………………………………………
b) Oxidación del piruvato:
¿Dónde se realiza?:……………………………………………………………………………………………………………………
¿Cuáles son sus productos?………………………………………………………………………………………………………
c) Ciclo de Krebs:
¿Dónde se realiza?:……………………………………………………………………………………………………………………
¿Cuáles son sus productos?………………………………………………………………………………………………………
d) Fosforilación oxidativa:
¿Dónde se realiza?:……………………………………………………………………………………………………………………
¿Cuántos NADH y FADH2 se oxidan?…………………………………………………………………………………………
¿Cuántos ATP se producen?………………………………………………………………………………………………………
Glucólisis
Glucosa 2
piruvato
+ 2 ATP por fosforilación
al nivel de sustrato
+ 2 ATP 34 ATP
por fosforilación
al nivel de sustrato
por fosforilación oxidativa
máximo por glucosa Aproximadamente
38 ATP
2
Acetil
COA
Ciclo
del ácido
cítrico
Fosforilación
oxidativa: transporte
de electrones y
quimiósmosis
2 NADH
2 NADH
o
2 FADH2
2 NADH 6 NADH 2 FADH2
MITOCONDRIA CITOSOL Las lanzaderas de electrones
se extienden en la membrana.





























