Embed Size (px)
DESCRIPTION
Las neuronas (del griego νεῦρον, cuerda, nervio) son un tipo de células del sistema nervioso cuya principal característica es la excitabilidad eléctrica de su membrana plasmática ; están especializadas en la recepción de estímulos y conducción del impulso nervioso (en forma de potencial de acción)
Citation preview

Las Neuronas y su funcionalidad
Ms. Ing. Jairo E. Márquez D.
El presente documento tiene como objetivo dar a conocer de forma general, las características de las neuronas y su funcionalidad general, con el fin que el estudiante de ingeniería, conceptualice esta información y relacione con los temas de Inteligencia artificial. Por consiguiente este documento tiene un uso estrictamente académico, en la que se da el crédito a los autores y fuentes que contribuyeron al desarrollo del mismo.
Neurona [1]
Las neuronas (del griego νεῦρον, cuerda, nervio) son un tipo de células del sistema nervioso cuya principal característica es la excitabilidad eléctrica de su membrana plasmática1; están especializadas en la recepción de estímulos y conducción del impulso nervioso (en forma de potencial de acción) entre ellas o con otros tipos celulares, como por ejemplo las fibras musculares de la placa motora.
Las neuronas, se caracterizan por ser altamente diferenciadas. Por lo cual, la mayoría de las neuronas no se dividen una vez alcanzada su madurez; no obstante, una minoría sí lo hace.2 Las neuronas presentan unas características morfológicas típicas que sustentan sus funciones: un cuerpo celular llamado soma o «pericarion», central; una o varias prolongaciones cortas que generalmente transmiten impulsos hacia el soma celular,
1 La membrana plasmática es un bicapa lipídica que delimita todas las células. Es una estructura laminada formada por fosfolípidos, glicolípidos y proteínas, que engloban, delimita, da forma y contribuye a mantener el equilibrio entre el interior (medio intracelular) y el exterior (medio extracelular) de las células. Es similar a las membranas que delimitan los orgánulos de células eucariotas. Está compuesta por dos láminas que sirven de "contenedor" para el citosol y los distintos compartimentos internos de la célula, así como también otorga protección mecánica. Está formada principalmente por fosfolípidos (fosfatidiletanolamina y fosfatidilcolina), colesterol, glúcidos y proteínas (integrales y periféricas).
La principal característica de esta barrera es su permeabilidad selectiva de las moléculas que deben entrar y salir de la célula. De esta forma se mantiene estable el medio intracelular, regulando el paso de agua, iones y metabolitos, a la vez que mantiene el potencial electroquímico (haciendo que el medio interno esté cargado negativamente). La membrana plasmática es capaz de recibir señales que permiten el ingreso de partículas a su interior. Cuando una molécula de gran tamaño atraviesa o es expulsada de la célula y se invagina parte de la membrana plasmática para recubrirlas cuando están en el interior ocurren respectivamente los procesos de endocitosis y exocitosis.

denominadas dendritas; y una prolongación larga, denominada axón o «cilindroeje», que conduce los impulsos desde el soma hacia otra neurona u órgano diana.[2]3
La neurogénesis en seres adultos, fue descubierta apenas en el último tercio del siglo XX. Hasta hace pocas décadas se creía que, a diferencia de la mayoría de las otras células del organismo, las neuronas normales en el individuo maduro no se regeneraban, excepto las células olfatorias. Los nervios mielinados del sistema nervioso periférico también tienen la posibilidad de regenerarse a través de la utilización del neurolema, una capa formada de los núcleos de las células de Schwann.
Una neurona típica consta de: un núcleo voluminoso central, situado en el soma; un pericarion que alberga los orgánulos celulares típicos de cualquier célula eucariota2; y neuritas (esto es, generalmente un axón y varias dendritas) que emergen del pericarion. [2]
Núcleo
Situado en el cuerpo celular, suele ocupar una posición central y ser muy conspicuo (visible), especialmente en las neuronas pequeñas. Contiene uno o dos nucléolos prominentes, así como una cromatina dispersa, lo que da idea de la relativamente alta actividad transcripcional de este tipo celular. La envoltura nuclear, con multitud de poros nucleares, posee una lámina nuclear muy desarrollada. Entre ambos puede aparecer el cuerpo accesorio de Cajal, una estructura esférica de en torno a 1μm de diámetro que corresponde a una acumulación de proteínas ricas en los aminoácidos arginina y tirosina.
Nota 1:[3] Un aminoácido es una molécula orgánica con un grupo amino (-NH2) y un grupo carboxilo (-COOH; ácido). Los aminoácidos más frecuentes y de mayor interés son aquellos que forman parte de las proteínas. Dos aminoácidos se combinan en una reacción de condensación que libera agua formando un enlace peptídico; estos dos "residuos" de aminoácido forman un dipéptido. Si se une un tercer aminoácido se forma un tripéptido y así, sucesivamente, para formar un polipéptido. Esta reacción tiene lugar de manera natural en los ribosomas.
Todos los aminoácidos componentes de las proteínas son L-alfa-aminoácidos. Por lo tanto, están formados por un carbono alfa unido a un grupo carboxilo, a un grupo amino, a un hidrógeno y a una cadena (habitualmente denominada cadena lateral o radical R) de estructura variable, que determina la identidad y las propiedades de cada uno de los diferentes aminoácidos; existen cientos de radicales por lo que se conocen cientos de aminoácidos diferentes, pero sólo 20 (actualmente se consideran 22) forman parte de las proteínas y tienen codones específicos en el código genético.
2 Se denominan células eucariotas a todas las células que tienen su material hereditario, fundamentalmente su información genética, encerrado dentro de una doble membrana, la envoltura nuclear; la cual delimita un núcleo celular.
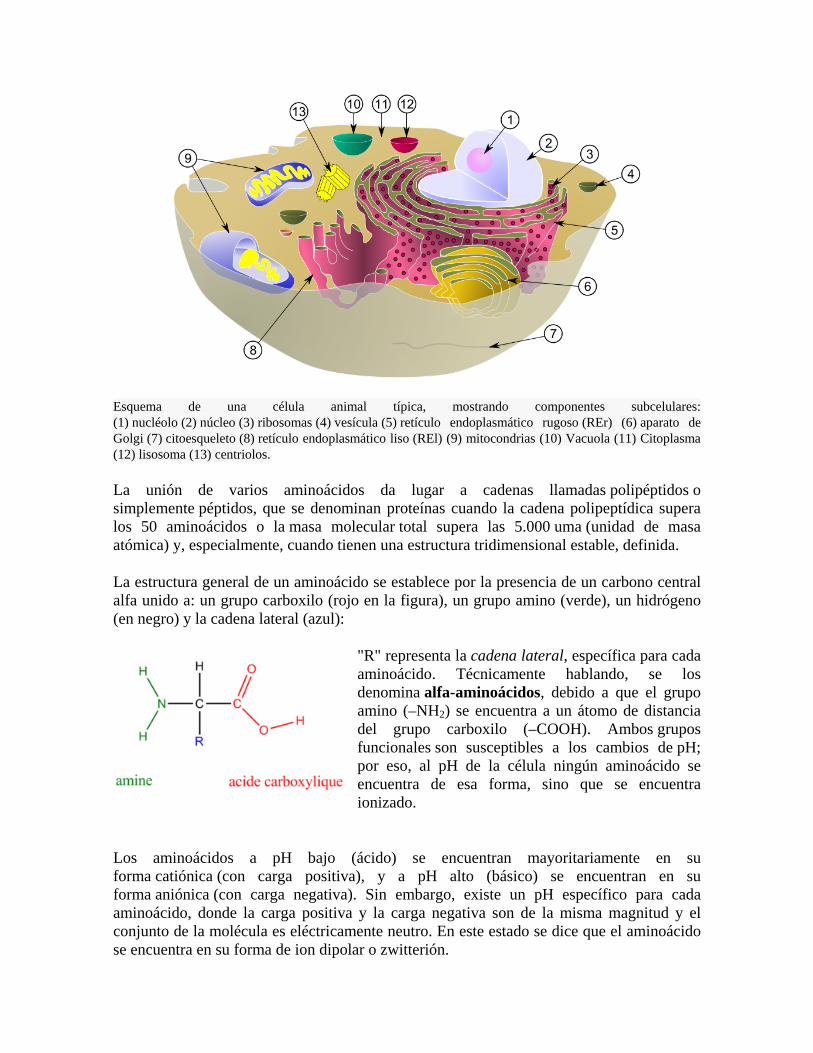
Esquema de una célula animal típica, mostrando componentes subcelulares: (1) nucléolo (2) núcleo (3) ribosomas (4) vesícula (5) retículo endoplasmático rugoso (REr) (6) aparato de Golgi (7) citoesqueleto (8) retículo endoplasmático liso (REl) (9) mitocondrias (10) Vacuola (11) Citoplasma (12) lisosoma (13) centriolos.
La unión de varios aminoácidos da lugar a cadenas llamadas polipéptidos o simplemente péptidos, que se denominan proteínas cuando la cadena polipeptídica supera los 50 aminoácidos o la masa molecular total supera las 5.000 uma (unidad de masa atómica) y, especialmente, cuando tienen una estructura tridimensional estable, definida.
La estructura general de un aminoácido se establece por la presencia de un carbono central alfa unido a: un grupo carboxilo (rojo en la figura), un grupo amino (verde), un hidrógeno (en negro) y la cadena lateral (azul):
"R" representa la cadena lateral, específica para cada aminoácido. Técnicamente hablando, se los denomina alfa-aminoácidos, debido a que el grupo amino (–NH2) se encuentra a un átomo de distancia del grupo carboxilo (–COOH). Ambos grupos funcionales son susceptibles a los cambios de pH; por eso, al pH de la célula ningún aminoácido se encuentra de esa forma, sino que se encuentra ionizado.
Los aminoácidos a pH bajo (ácido) se encuentran mayoritariamente en su forma catiónica (con carga positiva), y a pH alto (básico) se encuentran en su forma aniónica (con carga negativa). Sin embargo, existe un pH específico para cada aminoácido, donde la carga positiva y la carga negativa son de la misma magnitud y el conjunto de la molécula es eléctricamente neutro. En este estado se dice que el aminoácido se encuentra en su forma de ion dipolar o zwitterión.

Notas 2: La arginina (arg o R) es uno de los 20 aminoácidos que se encuentran formando parte de las proteínas. En el tejido hepático, la arginina puede ser sintetizada en el ciclo de la ornitina (o ciclo de la urea). Se clasifica, en población pediátrica, como un aminoácido esencial. Este aminoácido se encuentra involucrado en muchas de las actividades de las glándulas endocrinas. [4]
La palabra tirosina proviene del griego tyros, que significa queso. Se llama así ya que este aminoácido fue descubierto por un químico alemán llamado Justus Von Liebig a partir de la proteína caseína, que se encuentra en el queso. Se conocen tres isómeros distintos del aminoácido tirosina: para-tirosina, meta- tirosina y la orto-tirosina. Aunque la forma más conocida y estudiada es la para-tirosina o también llamada L-tirosina.
La tirosina es uno de los 20 aminoácidos que forman las proteínas. Se clasifica como un aminoácido no esencial en los mamíferos ya que su síntesis se produce a partir de la hidroxilación de otro aminoácido: la fenilalanina. Esto se considera así siempre y cuando la dieta de los mamíferos contenga un aporte adecuado de fenilalanina. Por tanto el aminoácido fenilalanina sí que es esencial.
Pericarion
Diversos orgánulos3 llenan el citoplasma que rodea al núcleo. El orgánulo más notable, por estar el pericarion lleno de ribosomas4 libres y adheridos al retículo rugoso5, es la 3 Se denomina orgánulos (o también organelas, organelos, organoides o elementos celulares) a las diferentes estructuras contenidas en el citoplasma de las células, principalmente las eucariotas, que tienen una forma determinada. La célula procariota carece de la mayor parte de los orgánulos. 4 Los ribosomas son complejos macromoleculares de proteínas y ácido ribonucleico (ARN) que se encuentran en el citoplasma, en las mitocondrias, en retículo endoplasmatico y en los cloroplastos. Son un complejo molecular encargado desintetizar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero (ARNm). Sólo son visibles al microscopio electrónico, debido a su reducido tamaño (29 nm en células procariotas y 32 nm eneucariotas). Bajo el microscopio electrónico se observan como estructuras redondeadas, densas a los electrones. Bajo el microscopio óptico se

llamada sustancia de Nissl, al microscopio óptico, se observan como grumos basófilos6, y, al electrónico, como apilamientos de cisternas del retículo endoplasmático. Tal abundancia de los orgánulos relacionados en la síntesis proteica se debe a la alta tasa biosintética del pericarion.
Estos son particularmente notables en neuronas motoras somáticas, como las del ucerno anterior de la médula espinal o en ciertos núcleos de nervios craneales motores. Los cuerpos de Nissl no solamente se hallan en el pericarion sino también en las dendritas, aunque no en el axón, y es lo que permite diferenciar de dendritas y axones en el neurópilo.
El aparato de Golgi7, que se descubrió originalmente en las neuronas, es un sistema muy desarrollado de vesículas aplanadas y agranulares pequeñas. Es la región donde los productos de la sustancia de Nissl posibilitan una síntesis adicional. Hay lisosomas8 primarios y secundarios (estos últimos, ricos en lipofuscina9, pueden marginar al núcleo en individuos de edad avanzada debido a su gran aumento). [5] Las mitocondrias, pequeñas y redondeadas, poseen habitualmente crestas longitudinales.
observa que son los responsables de la basofilia que presentan algunas células. Están en todas las células (excepto en los espermatozoides). Los ribosomas no se definen como orgánulos, ya que no existen endomembranas en su estructura. Fuente. Ribosomas. http://es.wikipedia.org/wiki/Ribosoma 5 El retículo endoplasmático rugoso (RER), también llamado retículo endoplasmático granular, ergastoplasma o ergatoplasma, es un orgánulo propio de la célula eucariota que participa en la síntesis y el transporte deproteínas en general. En las células nerviosas también se conoce como cuerpos de Nissl. 6 Se denomina basófilo a cualquier célula que se tiñe fácilmente con colorantes básicos (hematoxilina principalmente). Sin embargo, cuando se emplea este término sin ninguna aclaración adicional, suele referirse a uno de los tipos de leucocitos (glóbulos blancos de la sangre) de la familia de los granulocitos. 7 El aparato de Golgi es un orgánulo presente en todas las células eucariotas excepto los glóbulos rojos y las células epidérmicas. 8 Los lisosomas son orgánulos relativamente grandes, formados por el retículo endoplasmático rugoso y luego empaquetadas por el complejo de Golgi, que contienen enzimas hidrolíticas y proteolíticas que sirven para digerir los materiales de origen externo (heterofagia) o interno (autofagia) que llegan a ellos. Es decir, se encargan de la digestión celular. Son estructuras esféricas rodeadas de membrana simple. Son bolsas de enzimas que si se liberasen, destruirían toda la célula. Esto implica que la membrana lisosómica debe estar protegida de estas enzimas. El tamaño de un lisosoma varía entre 0.1–1.2 μm. Kuehnel, W (2003) (en inglés). Color Atlas of Cytology, Histology, & Microscopic Anatomy (4th edición). Thieme. pp. 34. 9 La lipofuscina es un pigmento compuesto por polímero de lípidos y fosfolípidos, derivados de la peroxidación de los lípidos poliinsaturados de las membranas subcelulares. Su importancia radica en que es un signo de vejez celular originado por la acción de los radicales libres. Es normal y no patológico observarla en preparaciones histológicas de tejido cardíaco (en el cono sarcoplásmico de la fibra muscular) y tejido neuronal (en el soma de las neuronas), ya que estas células carecen de capacidad para regenerarse debido a su alto grado de especialización.

Partes internas de una neurona
En cuanto al citoesqueleto10, el pericarion es rico en microtúbulos11 (clásicamente, de hecho, denominados neurotúbulos, si bien son idénticos a los microtúbulos de células no neuronales) [6] y filamentos intermedios (denominados neurofilamentos por la razón antes mencionada).[7] Los neurotúbulos se relacionan con el transporte rápido de las moléculas de proteínas que se sintetizan en el cuerpo celular y que se llevan a través de las dendritas y el axón.
10 El citoesqueleto es un entramado tridimensional de proteínas que provee soporte interno en las células, organiza las estructuras internas de la misma e interviene en los fenómenos de transporte, tráfico y división celular.1 En las células eucariontas, consta de microfilamentos, filamentos intermedios y microtúbulos, mientras que en las procariotas está constituido principalmente por las proteínas estructurales FtsZ y MreB. El citoesqueleto es una estructura dinámica que mantiene la forma de la célula, facilita la movilidad celular (usando estructuras como los cilios y los flagelos), y desempeña un importante papel tanto en el tráfico intracelular (por ejemplo, los movimientos de vesículas y orgánulos) y en la división celular.
11 Los microtúbulos son estructuras tubulares de las células, de 25 nm de diámetro exterior y unos 12 nm de diámetro interior, con longitudes que varían entre unos pocos nanómetros a micrómetros, que se originan en los centros organizadores de microtúbulos y que se extienden a lo largo de todo el citoplasma. Se hallan en las células eucariotas y están formadas por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina.
Los microtúbulos intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis) y que, junto con los microfilamentos y los filamentos intermedios, forman el citoesqueleto. Además, constituyen la estructura interna de los cilios y los flagelos.

Dendritas
Las dendritas son ramificaciones que proceden del soma neuronal que consisten en proyecciones citoplasmáticas envueltas por una membrana plasmática sin envoltura de mielina. En ocasiones, poseen un contorno irregular, desarrollando espinas. Sus orgánulos y componentes característicos son: muchos microtúbulos y pocos neurofilamentos, ambos dispuestos en haces paralelos; muchas mitocondrias; grumos de Nissl, más abundantes en la zona adyacente al soma; retículo endoplasmático liso, especialmente en forma de vesículas relacionadas con la sinapsis.
Axón
El axón es una prolongación del soma neuronal recubierta por una o más células de Schwann en el sistema nervioso periférico de vertebrados, con producción o no de mielina. Puede dividirse, de forma centrífuga al pericarion, en: cono axónico, segmento inicial, resto del axón. [8]
• Cono axónico. Adyacente al pericarion, es muy visible en las neuronas de gran tamaño. En él se observa la progresiva desaparición de los grumos de Nissl y la abundancia de microtúbulos y neurfilamentos que, en esta zona, se organizan en haces paralelos que se proyectarán a lo largo del axón.
• Segmento inicial. En él comienza la mielinización externa. En el citoplasma, a esa altura se detecta una zona rica en material electronodenso en continuidad con la membrana plasmática, constituido por material filamentoso y partículas densas; se asume que interviene en la generación del potencial de acción que transmitirá la señal sináptica. En cuanto al citoesqueleto, posee esta zona la organización propia del resto del axón. Los microtúbulos, ya polarizados, poseen la proteína τ [9] pero no la proteína MAP-2.
• Resto del axón. En esta sección comienzan a aparecer los nódulos de Ranvier y las sinapsis.
Función de las neuronas
Las neuronas tienen la capacidad de comunicarse con precisión, rapidez y a larga distancia con otras células, ya sean nerviosas, musculares o glandulares. A través de las neuronas se transmiten señales eléctricas denominadas impulsos nerviosos.

Estos impulsos nerviosos viajan por toda la neurona comenzando por las dendritas, y pasa por toda la neurona hasta llegar a los botones terminales, que pueden conectar con otra neurona, fibras musculares o glándulas. La conexión entre una neurona y otra se denomina sinapsis.
Las neuronas conforman e interconectan los tres componentes del sistema nervioso: sensitivo, motor e integrador o mixto; de esta manera, un estímulo que es captado en alguna región sensorial entrega cierta información que es conducida a través de las neuronas y es analizada por el componente integrador, el cual puede elaborar una respuesta, cuya señal es conducida a través de las neuronas. Dicha respuesta es ejecutada mediante una acción motora, como la contracción muscular o secreción glandular.
El impulso nervioso
Las neuronas transmiten ondas de naturaleza eléctrica originadas como consecuencia de un cambio transitorio de la permeabilidad en la membrana plasmática. Su propagación se debe a la existencia de una diferencia de potencial o potencial de membrana (que surge gracias a las concentraciones distintas de iones a ambos lados de la membrana, según describe el potencial de Nernst [10] ) entre la parte interna y externa de la célula (por lo general de -70mV). La carga de una célula inactiva se mantiene en valores negativos (el interior respecto al exterior) y varía dentro de unos estrechos márgenes. Cuando el potencial de membrana de una célula excitable se despolariza más allá de un cierto umbral (de 65mV a 55mV) la célula genera (o dispara) un potencial de acción. Un potencial de acción es un cambio muy rápido en la polaridad de la membrana de negativo a positivo y vuelta a negativo, en un ciclo que dura unos milisegundos. [11]
Propiedades electrofisiológicas intrínsecas
Hasta finales de los años 80 del siglo XX el dogma de la neurociencia dictaba que sólo las conexiones y los neurotransmisores liberados por las neuronas determinaban la función de una neurona. Las investigaciones realizadas por Rodolfo Llinás con sus colaboradores durante los años 80 sobre vertebrados pusieron de manifiesto que el dogma mantenido hasta entonces era erróneo. En 1988, Rodolfo Llinás presentó el nuevo punto de vista funcional sobre la neurona en su artículo "The Intrinsic Electrophysiological Properties of Mammalian Neurons: Insights into Central Nervous System Function"[12] y que es considerado un manifiesto que marca el cambio de mentalidad en neurociencia respecto al aspecto funcional de las neuronas con más de 1250 citas en la bibliografía científica. El nuevo punto de vista funcional sobre la neurona quedo resumido en lo que hoy es conocido por la Ley de Llinás.

Neurosecreción
Las células neurosecretoras son neuronas especializadas en la secreción de sustancias que, en vez de ser vertidas en la hendidura sináptica, lo hacen en capilares sanguíneos, por lo que sus productos son transportados por la sangre hacia los tejidos diana; esto es, actúan a través de una vía endocrina. [13] Esta actividad está representada a lo largo de la diversidad zoológica: se encuentra en crustáceos, [14] insectos, [15] equinodermos, [16] vertebrados, etc.
Clasificación de las neuronas
Aunque el tamaño del cuerpo celular puede ser desde 5 hasta 135 micrómetros, las prolongaciones o dendritas pueden extenderse a una distancia de más de un metro. El número, la longitud y la forma de ramificación de las dendritas brindan un método morfológico para la clasificación de las neuronas.
Según la forma y el tamaño
Según el tamaño de las prolongaciones, los nervios se clasifican en: [17]
• Poliédricas: como las motoneuronas del asta anterior de la médula. • Fusiformes: las que se encuentran en el doble ramillete de la corteza cerebral. • Estrelladas: como las neuronas aracniforme y estrelladas de la corteza cerebral y las
estrelladas, en cesta y Golgi del cerebelo. • Esféricas: en ganglios espinales, simpáticos y parasimpáticos • Piramidales: presentes en la corteza cerebral.
Según la polaridad
Según el número y anatomía de sus prolongaciones, las neuronas se clasifican en: [18]
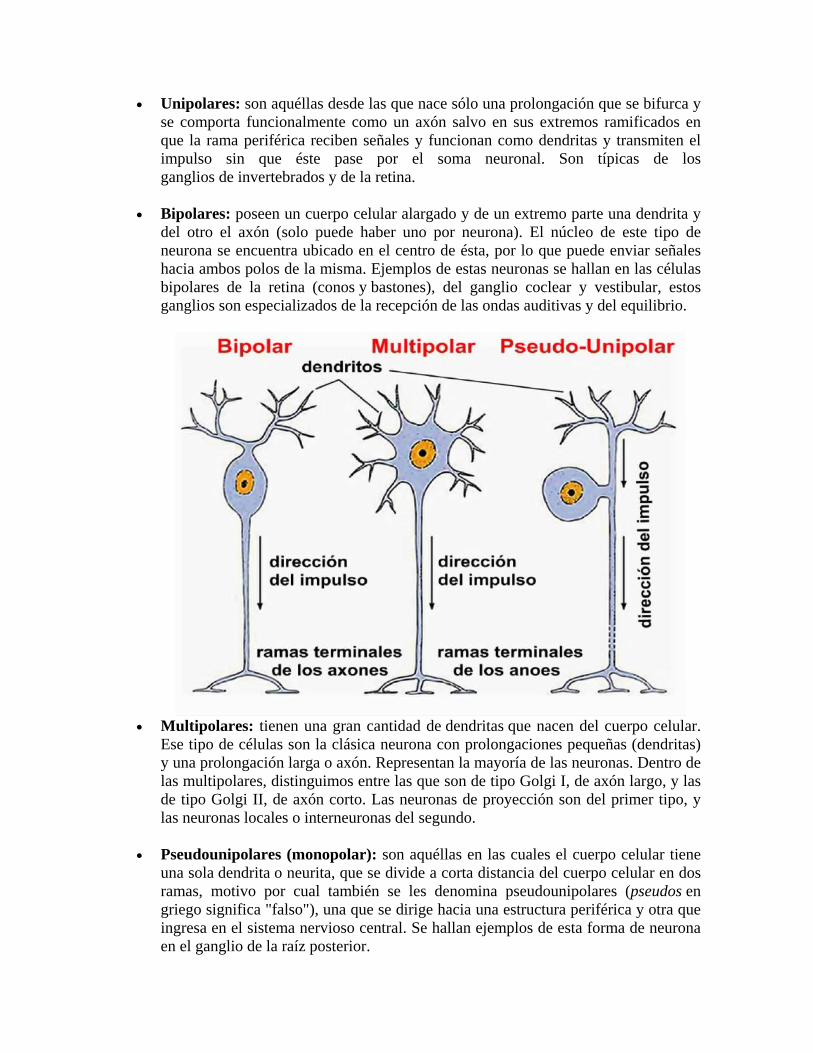
• Unipolares: son aquéllas desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente como un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que éste pase por el soma neuronal. Son típicas de los ganglios de invertebrados y de la retina.
• Bipolares: poseen un cuerpo celular alargado y de un extremo parte una dendrita y del otro el axón (solo puede haber uno por neurona). El núcleo de este tipo de neurona se encuentra ubicado en el centro de ésta, por lo que puede enviar señales hacia ambos polos de la misma. Ejemplos de estas neuronas se hallan en las células bipolares de la retina (conos y bastones), del ganglio coclear y vestibular, estos ganglios son especializados de la recepción de las ondas auditivas y del equilibrio.
• Multipolares: tienen una gran cantidad de dendritas que nacen del cuerpo celular.
Ese tipo de células son la clásica neurona con prolongaciones pequeñas (dendritas) y una prolongación larga o axón. Representan la mayoría de las neuronas. Dentro de las multipolares, distinguimos entre las que son de tipo Golgi I, de axón largo, y las de tipo Golgi II, de axón corto. Las neuronas de proyección son del primer tipo, y las neuronas locales o interneuronas del segundo.
• Pseudounipolares (monopolar): son aquéllas en las cuales el cuerpo celular tiene una sola dendrita o neurita, que se divide a corta distancia del cuerpo celular en dos ramas, motivo por cual también se les denomina pseudounipolares (pseudos en griego significa "falso"), una que se dirige hacia una estructura periférica y otra que ingresa en el sistema nervioso central. Se hallan ejemplos de esta forma de neurona en el ganglio de la raíz posterior.

• Anaxónicas: son pequeñas. No se distinguen las dendritas de los axones. Se
encuentran en el cerebro y órganos especiales de los sentidos.
Según las características de las neuritas
De acuerdo a la naturaleza del axón y de las dendritas, clasificamos a las neuronas en:3
• Axón muy largo o Golgi de tipo I. El axón se ramifica lejos del pericarion. Con axones de hasta 1 m.
• Axón corto o Golgi de tipo II. El axón se ramifica junto al soma celular. • Sin axón definido. Como las células amacrinas de la retina. • Isodendríticas. Con dendritas rectilíneas que se ramifican de modo que las ramas
hijas son más largas que las madres. • Idiodendríticas. Con las dendritas organizadas dependiendo del tipo neuronal; por
ejemplo, como las células de Purkinje del cerebelo. • Alodendríticas. Intermedias entre los dos tipos anteriores.
Según el mediador químico
Las neuronas pueden clasificarse, según el mediador químico, en:18
• Colinérgicas. Liberan acetilcolina. • Noradrenérgicas. Liberan norepinefrina. • Dopaminérgicas. Liberan dopamina. • Serotoninérgicas. Liberan serotonina. • Gabaérgicas. Liberan GABA, es decir, ácido γ-aminobutírico.
Según la función
Las neuronas pueden ser sensoriales, motoras o interneuronas:
• Motoras: Son las encargadas de producir la contracción de la musculatura. • Sensoriales: Reciben información del exterior, ej. Tacto, gusto, visión y las
trasladan al sistema nervioso central. • Interneuronas: Se encargan de conectar entre las dos diferentes neuronas.
Doctrina de la neurona
La doctrina de la neurona, establecida por Santiago Ramón y Cajal a finales del siglo XIX, es el modelo aceptado hoy en neurofisiología. Consiste en aceptar que la base de la función neurológica radica en las neuronas como entidades discretas, cuya interacción, mediada por sinapsis, conduce a la aparición de respuestas complejas. Cajal no solo postuló este principio, sino que lo extendió hacia una «ley de la polarización dinámica», que propugna la transmisión unidireccional de información (esto es, en un sólo sentido, de las dendritas hacia los axones). [19] No obstante, esta ley no siempre se cumple. Por ejemplo, las células

gliales pueden intervenir en el procesamiento de información, [20] e, incluso, las efapsis o sinapsis eléctricas, mucho más abundantes de lo que se creía, [21] presentan una transmisión de información directa de citoplasma a citoplasma. Más aún: las dendritas pueden dirigir una señal sináptica de forma centrífuga al soma neuronal, lo que representa una transmisión en el sentido opuesto al postulado, [22] de modo que sean los axones los que reciban de información (aferencia).
Redes neuronales
Una red neuronal se define como una población de neuronas físicamente interconectadas o un grupo de neuronas aisladas que reciben señales que procesan a la manera de un circuito reconocible. La comunicación entre neuronas, que implica un proceso electroquímico, [23] implica que, una vez que una neurona es excitada a partir de cierto umbral, ésta se despolariza transmitiendo a través de su axón una señal que excita a neuronas aledañas, y así sucesivamente. El sustento de la capacidad del sistema nervioso, por tanto, radica en dichas conexiones. En oposición a la red neuronal, se habla de circuito neuronal cuando se hace mención a neuronas que se controlan dando lugar a una retroalimentación («feedback»), como define la cibernética.
Cerebro y neuronas
El número de neuronas en el cerebro varía drásticamente según la especie estudiada. [24] Se estima que cada cerebro humano posee en torno a 1011 neuronas: es decir, unos cien mil millones. No obstante, Caenorhabditis elegans, un gusano nematodo muy empleado como animal modelo, posee sólo 302; [25] y la mosca de la fruta, Drosophila melanogaster, unas 300.000, que bastan para permitirle exhibir conductas complejas. [26]
La fácil manipulación en el laboratorio de estas especies, cuyo ciclo de vida es muy corto y cuyas condiciones de cultivo poco exigentes, permiten a los investigadores científicos emplearlas para dilucidar el funcionamiento neuronal, puesto que el mecanismo básico de la actividad neuronal es común al de nuestra especie. [27]
Redes neuronales artificiales
El conocimiento de las redes neuronales biológicas ha dado lugar a un diseño empleado en inteligencia artificial. Estas redes funcionan porque cada neurona recibe una serie de entradas a través de interconexiones y emite una salida. Esta salida viene dada por tres funciones: una función de propagación que por lo general consiste en el sumatorio de cada

entrada multiplicada por el peso de su interconexión; una función de activación,12 que modifica a la anterior y que puede no existir, siendo en este caso la salida la misma función de propagación; y una función de transferencia13, que se aplica al valor devuelto por la función de activación. Se utiliza para acotar la salida de la neurona y generalmente viene dada por la interpretación que queramos darle a dichas salidas. [28]
12 La Función de Activación de un nodo define la salida de un nodo dada una entrada o un conjunto de entradas. Se podría decir que un circuito estándar de computador se comporta como una red digital de funciones de activación al activarse como "ON" (1) u "OFF" (0), dependiendo de la entrada. Esto es similar al funcionamiento de un Perceptrón en una Red neuronal artificial.
En las redes neurales inspiradas sobre la biología, la función de activación es usualmente una abstracción representando una tasa de potencial de activación gatillándose en la celda. En su forma simplificada, esta función es binaria, esto es, se activa la neurona o no. La función se ve como Φ(vi) = U(vi), donde U es la función escalón. En este caso, un gran número de neuronas deben ser usadas en computación más allá de la separación lineal de las categorías.
Una función rampa también puede ser usada para reflejar el incremento del potencial de activación que ocurre cuando la entrada se incrementa. La función podría ser de la forma Φ(vi) = µvi, donde μ es la pendiente. Esta función de activación es lineal, y por consiguiente tiene los mismos problemas que la función binaria. En adición, las redes neurales construidas usando este modelo tienen convergencia inestable porque a la larga, las entradas a la neurona tienden a incrementarse sin límite, esta función no es normalizable.
Los problemas mencionados anteriormente, pueden ser manejados usando una función de activación sigmoidal. Un modelo realístico permanece en cero hasta que una entrada es recibida, en este punto la frecuencia de activación se incrementa rápidamente, pero gradualmente llega a serasíntota cuando la frecuencia es 100%. Matemáticamente, esto se ve como Φ(vi) = U(vi)tanh(vi), donde la función de tangente hiperbólica puede también ser cualquier función sigmoidal. Esta conducta es realísticamente reflejada en la neurona, ya que las neuronas no pueden físicamente activarse más rápido que una cierta tasa.
El modelo final que es usado en perceptrones multicapa es el modelo de activación sigmoidal en la forma de tangente hiperbólica. Dos formas de esta función son comúnmente usados: Φ(vi) = tanh(vi), cuyos rangos son normalizados desde -1 hasta 1, y Φ(vi) = (1+exc(-vi))-1 es verticalmente normalizado desde 0 a 1. El último modelo es frecuentemente considerado más biológicamente realístico, pero tiene dificultades teóricas y experimentales en ciertos tipos de problemas computacionales.
Fuente. Haykin, Simon (1998). Neural Networks: A Comprehensive Foundation (2 edición). Prentice Hall.
13 Una función de transferencia es un modelo matemático que a través de un cociente relaciona la respuesta de un sistema (modelada) a una señal de entrada o excitación (también modelada). En la teoría de control, a menudo se usan las funciones de transferencia para caracterizar las relaciones de entrada y salida de componentes o de sistemas que se describen mediante ecuaciones diferenciales lineales e invariantes en el tiempo. Es por eso que se puede definir matemáticamente como:
La función de trasferencia de un sistema lineal e invariante en el tiempo (LTI), se define como el cociente entre la transformada de Laplace de la salida y la transformada de Laplace de la entrada, bajo la suposición de que las condiciones iniciales son nulas.
Potencial de acción
Un potencial de acción o también llamado impulso eléctrico, es una onda de descarga eléctrica que viaja a lo largo de la membrana celular modificando su distribución de carga eléctrica. Los potenciales de acción se utilizan en el cuerpo para llevar información entre unos tejidos y otros, lo que hace que sean una característica microscópica esencial para la vida de los animales. Pueden generarse por diversos tipos de células corporales, pero las más activas en su uso son las células del sistema nervioso para enviar mensajes entre células nerviosas (sinapsis) o desde células nerviosas a otros tejidos corporales, como el músculo o las glándulas.
Muchas plantas también generan potenciales de acción que viajan a través del floema para coordinar su actividad. La principal diferencia entre los potenciales de acción de animales y plantas es que las plantas utilizan flujos de potasio y calcio mientras que los animales utilizan potasio y sodio.
Los potenciales de acción son la vía fundamental de transmisión de códigos neurales. Sus propiedades pueden frenar el tamaño de cuerpos en desarrollo y permitir el control y coordinación centralizados de órganos y tejidos
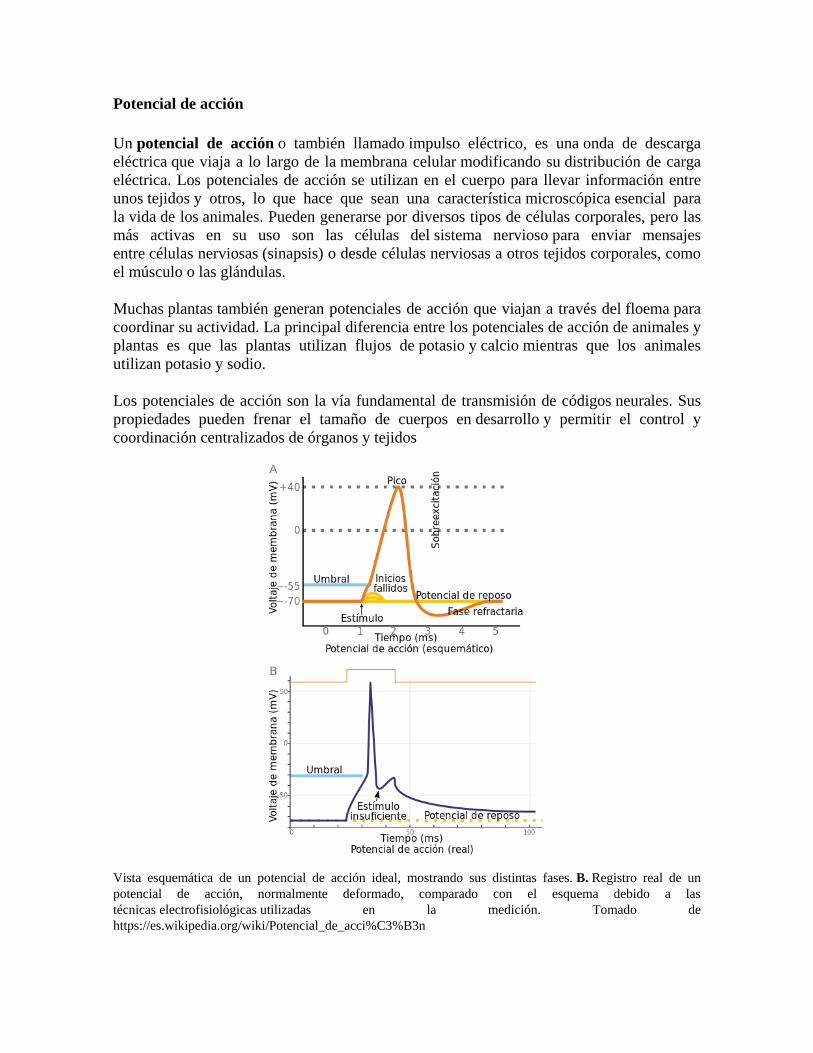
Vista esquemática de un potencial de acción ideal, mostrando sus distintas fases. B. Registro real de un potencial de acción, normalmente deformado, comparado con el esquema debido a las técnicas electrofisiológicas utilizadas en la medición. Tomado de https://es.wikipedia.org/wiki/Potencial_de_acci%C3%B3n
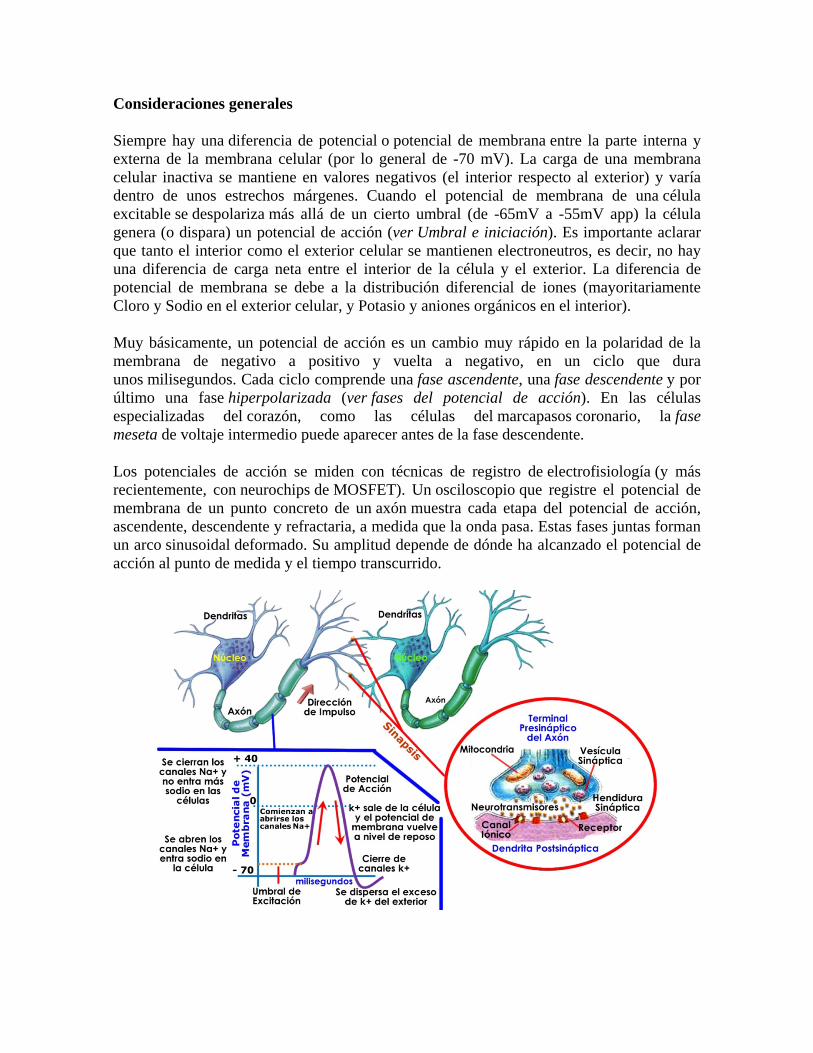
Consideraciones generales
Siempre hay una diferencia de potencial o potencial de membrana entre la parte interna y externa de la membrana celular (por lo general de -70 mV). La carga de una membrana celular inactiva se mantiene en valores negativos (el interior respecto al exterior) y varía dentro de unos estrechos márgenes. Cuando el potencial de membrana de una célula excitable se despolariza más allá de un cierto umbral (de -65mV a -55mV app) la célula genera (o dispara) un potencial de acción (ver Umbral e iniciación). Es importante aclarar que tanto el interior como el exterior celular se mantienen electroneutros, es decir, no hay una diferencia de carga neta entre el interior de la célula y el exterior. La diferencia de potencial de membrana se debe a la distribución diferencial de iones (mayoritariamente Cloro y Sodio en el exterior celular, y Potasio y aniones orgánicos en el interior).
Muy básicamente, un potencial de acción es un cambio muy rápido en la polaridad de la membrana de negativo a positivo y vuelta a negativo, en un ciclo que dura unos milisegundos. Cada ciclo comprende una fase ascendente, una fase descendente y por último una fase hiperpolarizada (ver fases del potencial de acción). En las células especializadas del corazón, como las células del marcapasos coronario, la fase meseta de voltaje intermedio puede aparecer antes de la fase descendente.
Los potenciales de acción se miden con técnicas de registro de electrofisiología (y más recientemente, con neurochips de MOSFET). Un osciloscopio que registre el potencial de membrana de un punto concreto de un axón muestra cada etapa del potencial de acción, ascendente, descendente y refractaria, a medida que la onda pasa. Estas fases juntas forman un arco sinusoidal deformado. Su amplitud depende de dónde ha alcanzado el potencial de acción al punto de medida y el tiempo transcurrido.

El potencial de acción no se mantiene en un punto de la membrana plasmática, sino que viaja a lo largo de la membrana (ver propagación). Puede desplazarse a lo largo de un axón a mucha distancia, por ejemplo transportando señales desde el cerebro hasta el extremo de la médula espinal. En animales grandes como las jirafas o las ballenas la distancia puede ser de varios metros.
La velocidad y simplicidad de los potenciales de acción varía según el tipo celular e incluso entre células del mismo tipo. Aun así, los cambios de voltaje tienden a tener la misma amplitud entre ellas. En una misma célula, varios potenciales de acción consecutivos son prácticamente indistinguibles. La "causa" del potencial de acción es el intercambio de iones a través de la membrana celular. Primero, un estímulo abre los canales de sodio. Dado que hay algunos iones de sodio en el exterior, y el interior de la neurona es negativo con relación al exterior, los iones de sodio entran rápidamente a la neurona. Recuerda que el sodio tiene una carga positiva, así que la neurona se vuelve más positiva y empieza a despolarizarse. Los canales de potasio de demoran un poco más en abrirse; una vez abiertos el potasio sale rápidamente de la célula, revirtiendo la despolarización. Más o menos en este momento, los canales de sodio empiezan a cerrarse, logrando que el potencial de acción vuelva a -70 mV (repolarización). En realidad el potencial de acción va más allá de -70 mV (hiperpolarización), debido a que los canales de potasio se quedan abiertos un poco más. Gradualmente las concentraciones de iones regresan a los niveles de reposo y la célula vuelve a -70 mV
Mecanismo subyacente
Potencial de membrana en reposo
Cuando la célula no está estimulada por corrientes despolarizantes supraumbrales, se dice que se encuentra en un potencial de membrana en reposo.
La membrana celular está compuesta mayoritariamente por una bicapa de fosfolípidos altamente hidrofóbica, que impide el paso libre de partículas cargadas como los iones. Por lo cual esta bicapa de fosfolipidos se comporta como un condensador, separando cargas (dadas por los iones en disolución) a una distancia de aproximadamente 4nm. Esto permite la mantención del potencial de membrana a lo largo del tiempo. El potencial de membrana se debe a la distribución diferencial de iones entre el interior y el exterior celular. Este potencial de membrana es mantenido a lo largo del tiempo por el transporte activo de iones por parte de bombas, tales como la bomba sodio-potasio y la bomba de calcio. Estas proteínas usan la energía de hidrólisis de ATP para transportar iones en contra de su gradiente electroquímico, manteniendo así los gradientes de concentraciones iónicas que definen el potencial de membrana.

La membrana celular hidrofóbica impide que las moléculas cargadas difundan a través de ella, manteniendo una diferencia de potencial. Tomado de https://es.wikipedia.org/wiki/Potencial_de_acci%C3%B3n
Fases del potencial de acción
Las variaciones potencial de membrana durante el potencial de acción son resultado de cambios en la permeabilidad de la membrana celular a iones específicos (en concreto, sodio y potasio) y por consiguiente cambios en las concentraciones iónicas en los compartimientos intracelular y extracelular. Estas relaciones están matemáticamente definidas por la ecuación de Goldman, Hodgkin y Katz (GHK).
Los cambios en la permeabilidad de la membrana y el establecimiento y cese de corrientes iónicas durante el potencial de acción refleja la apertura y cierre de los canales iónicos que forman zonas de paso a través de membrana para los iones. Las proteínas que regulan el paso de iones a través de la membrana responden a los cambios de potencial de membrana.
En un modelo simplificado del potencial de acción, el potencial de reposo de una parte de la membrana se mantiene con el canal de potasio. La fase ascendente o de despolarización del potencial de acción se inicia cuando el canal de sodio dependiente de potencial se abre, haciendo que la permeabilidad del sodio supere ampliamente a la del potasio. El potencial de membrana va hacia ENa. En algunas células, como las células del

marcapasos coronario, la fase ascendente se genera por concentración de calcio más que de sodio.
Tras un corto intervalo, el canal de potasio dependiente de voltaje (retardado) se abre, y el canal de sodio se inactiva. Como consecuencia, el potencial de membrana vuelve al estado de reposo, mostrado en el potencial de acción como una fase descendente.
Debido a que hay más canales de potasio abiertos que canal de sodio (los canales de potasio de membrana y canales de potasio dependientes de voltaje están abiertos, y el canal de sodio está cerrado), la permeabilidad al potasio es ahora mucho mayor que antes del inicio de la fase ascendente, cuando sólo los canales de potasio de membrana estaban abiertos. El potencial de membrana se acerca a EK más de lo que estaba en reposo, haciendo que el potencial esté en fase refractaria. El canal de potasio retardado dependiente de voltaje se cierra debido a la hiperpolarización, y la célula regresa a su potencial de reposo.
Las fases ascendente y descendente del potencial de acción se denominan a veces despolarización e hiperpolarización respectivamente. Técnicamente, la despolarización es cualquier cambio en el potencial de membrana que lleve la diferencia de potencial a cero. Igualmente, la hiperpolarización es cualquier cambio de potencial que se aleje de cero. Durante la fase ascendente, el potencial de membrana primero se aproxima a cero, y luego se hace más positivo; así, la fase ascendente incluye tanto despolarización como hiperpolarización. Aunque es técnicamente incorrecto denominar las fases ascendente y descendente como despolarización e hiperpolarización, es común verlo entre profesores, físicos y libros de neurociencia.
Potencial de membrana [29] El potencial de membrana es la diferencia de potencial14 a ambos lados de una membrana que separa dos soluciones de diferente concentración de iones, como 14 La tensión eléctrica, o voltaje, diferencia de potencial eléctrico o tensión eléctrica (denotado dV y medido en voltios o julios por coulomb) es la diferencia de potencial entre dos puntos o la diferencia de potencial de energía eléctrica por la unidad de carga eléctrica entre dos puntos. También se puede definir como el trabajo por unidad de carga ejercido por el campo eléctrico sobre una partícula cargada para moverla entre dos posiciones determinadas.
La tensión es independiente del camino recorrido por la carga y depende exclusivamente del potencial eléctrico de los puntos A y B en el campo eléctrico, que es un campo conservativo.
Si dos puntos que tienen una diferencia de potencial se unen mediante un conductor, se producirá un flujo de electrones. Parte de la carga que crea el punto de mayor potencial se trasladará a través del conductor al punto de menor potencial y, en ausencia de una fuente externa (generador), esta corriente cesará cuando ambos puntos igualen su potencial eléctrico (ley de Henry). Este traslado de cargas es lo que se conoce

la membrana celular que separa el interior y el exterior de una célula. Cuando se habla de potenciales de membrana, se debería de hablar del "potencial de difusión" o "potencial de unión líquida"15. Al estudiar los potenciales de membrana desde un punto de vista teórico, debemos conocer:
Potencial de Nernst
El potencial de reposo de una célula es producido por diferencias en la concentración de iones dentro y fuera de la célula y por diferencias en la permeabilidad de la membrana celular a los diferentes iones. El potencial de equilibrio de Nernst relaciona la diferencia de potencial a ambos lados de una membrana biológica en el equilibrio con las características relacionadas con los iones del medio externo e interno y de la propia membrana.
como corriente eléctrica. Cuando se habla sobre una diferencia de potencial en un sólo punto, o potencial, se refiere a la diferencia de potencial entre este punto y algún otro donde el potencial se defina como cero.
15 El potencial de unión líquida se produce cuando dos disoluciones de diferentes concentraciones están en contacto entre sí a través de una membrana semipermeable. Se origina en la interfase una diferencia de potencial llamada potencial de unión. Es muy pequeña, del orden de unos pocos milivoltios. La disolución más concentrada tendrá una tendencia a migrar hacia la relativamente menos concentrada y si los iones no difunden a igual velocidad se producirá una diferencia de potencial.

Generación de un potencial de membrana. Para una explicación detallada, véase el texto.
El potencial de Nernst se establece entre disoluciones separadas por una membrana semipermeable. Por ejemplo, KCl (cloruro de potasio), una sal, en medio acuoso se disocia en K+y Cl- en relación 1:1, compensando las cargas positivas de los cationes potasio con las negativas de los aniones cloruro, por lo que la disolución será eléctricamente neutra. De existir una membrana biológica selectivamente permeable al K+ en el interior de la solución, los K+ difundirán libremente a un lado y a otro de la membrana. Sin embargo, como hay más iones en el compartimento 1, inicialmente fluirán más iones K+ del 1 al 2 que del 2 al 1. Como el Cl- no puede difundir a través de la membrana, pronto hay un exceso de carga positiva en el compartimento 2 y un exceso de carga negativa en el 1. El fluido en cada compartimento permanece con una carga neutra, si bien las cargas en exceso se concentran a lo largo de la membrana. Las capas de carga positiva y negativa a cada lado de la membrana producen una diferencia de potencial V=V1 –V2 a través de la membrana y un campo eléctrico E, que retarda el flujo de iones positivos del compartimento 1 al 2 y que acelera su flujo del compartimento 2 al 1.
En este sistema, tras un tiempo se alcanzará el equilibrio dinámico en el que exista un flujo de K+ idéntico del 2 al 1 como del 1 al 2. Este equilibrio depende de la diferencia de concentración que favorece el movimiento del 1 al 2 y de la diferencia de potencial que favorece la difusión del 2 al 1. La diferencia de potencial V en el equilibrio viene dada, en función de las concentraciones y de los iones de K+ en los dos compartimentos, mediante:
Éste es el potencial de equilibrio de Nernst. Posee un valor negativo cuando la membrana es permeable a los iones positivos, y positivo cuando lo es a los aniones. Aquí k es la constante de Boltzmann [1.3806504x10-23 J/K] y la T la temperatura absoluta. La magnitud kT es proporcional a la energía cinética media de los iones en solución
y es proporcional al flujo neto de iones debido a la diferencia de concentración. La magnitud eV es proporcional al flujo neto de iones debido a la diferencia de potencial. Así, la ecuación antes citada es la condición para que estos dos flujos sean iguales y opuestos.
Modificaciones: Temperatura de 37 ºC
A la temperatura corporal de 37 ºC el valor de KT/e es:
De modo que el potencial de Nernst es
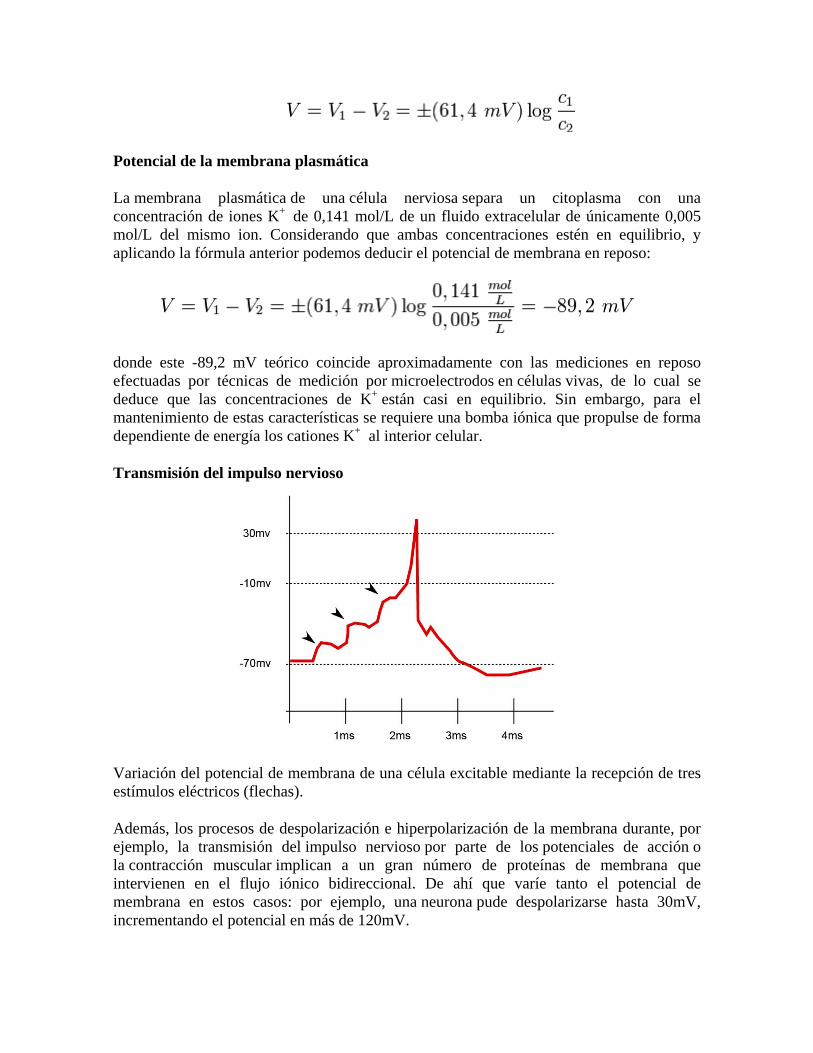
Potencial de la membrana plasmática
La membrana plasmática de una célula nerviosa separa un citoplasma con una concentración de iones K+ de 0,141 mol/L de un fluido extracelular de únicamente 0,005 mol/L del mismo ion. Considerando que ambas concentraciones estén en equilibrio, y aplicando la fórmula anterior podemos deducir el potencial de membrana en reposo:
donde este -89,2 mV teórico coincide aproximadamente con las mediciones en reposo efectuadas por técnicas de medición por microelectrodos en células vivas, de lo cual se deduce que las concentraciones de K+ están casi en equilibrio. Sin embargo, para el mantenimiento de estas características se requiere una bomba iónica que propulse de forma dependiente de energía los cationes K+ al interior celular.
Transmisión del impulso nervioso
Variación del potencial de membrana de una célula excitable mediante la recepción de tres estímulos eléctricos (flechas).
Además, los procesos de despolarización e hiperpolarización de la membrana durante, por ejemplo, la transmisión del impulso nervioso por parte de los potenciales de acción o la contracción muscular implican a un gran número de proteínas de membrana que intervienen en el flujo iónico bidireccional. De ahí que varíe tanto el potencial de membrana en estos casos: por ejemplo, una neurona pude despolarizarse hasta 30mV, incrementando el potencial en más de 120mV.

Ecuación de Goldman-Hodgkin-Katz (aplicable a membranas permeables a múltiples iones)
Tal como se vio anteriormente, la ecuación de Nerst solo calcula el potencial de difusión para un ion en particular, es decir, se asume que en el medio externo sólo existe un tipo de iones (por ejemplo: Na+).
Se sabe que tanto en los medios intra y extra celular existen múltiples iones tales como: Na+, K+, Cl-, Mg2+, entre otros, por lo tanto es necesario disponer de una fórmula que calcule dicho potencial para todos los iones presentes en el líquido extracelular.
Se sabe que la membrana celular es permeable a múltiples iones diferentes, por lo tanto al momento en que dichos iones difunden se genera un potencial de membrana que depende de tres factores:
1. La polaridad de la carga de cada uno de los iones a difundir. 2. La permeabilidad de la membrana a cada uno de los iones. 3. Las concentraciones de los mismos tanto en el exterior como en el interior de la
membrana.
La Ecuación de Goldman (también llamada de Goldman - Hodgkin - Katz) calcula el Potencial de la membrana en el interior de la célula cuando participan dos iones positivos univalentes (K+ y Na+) y un ion negativo también univalente (Cl-).
*Ecuación de Goldman
** donde:
- C = Concentración del ion
- P = Permeabilidad de la membrana al ion
Aclaraciones:
1. Los iones sodio, potasio y cloruro son los iones más importantes que participan en la generación del potencial de membrana en las fibras nerviosas y musculares. El gradiente de concentración de cada uno de los iones a través de la membrana ayuda a determinar el voltaje del potencial de membrana.
2. La permeabilidad de la membrana a cada uno de los iones determina el grado de importancia de cada uno de ellos, es decir, si la membrana por algún motivo solo es

permeable por ejemplo al sodio el potencial de membrana será igual al Potencial de Nerst para el sodio.
3. Un gradiente de concentración positivo en el interior de la membrana causa electronegatividad en el interior de la misma; esto explica que si hay por ejemplo una mayor concentración de iones sodio en el interior de la membrana, habrá por lo tanto mayor difusión del mismo, desde el interior hasta el exterior de la membrana, generando un déficit de cargas positivas en el interior de la membrana, lo cual dotará a dicho medio, de carga negativa.
4. Los cambios rápidos de concentración de los iones sodio y potasio son los principales responsables de la transmisión nerviosa.
El potencial de membrana no es el mismo en todas las células, dependiendo del origen de las mismas. Existen células que tienen -50mV y otras, como por ejemplo las musculares, que oscilan entre -50 y 60mV.
En el organismo existen dos espacios: el intercelular o extracelular y el intracelular. En el extracelular o líquido intersticial, el anión más abundante es el ion cloruro. En el medio intracelular o citoplasma, los aniones más abundantes son las proteínas, que en las condiciones del pH celular interno, están ionizadas negativamente por liberación de iones hidrógeno, H+. El catión más abundante en el líquido intersticial es el ion sodio, y en el citoplasma es el ion potasio.
El desequilibrio iónico que produce la polarización de la membrana es debido a la distinta permeabilidad que presenta frente a cada uno de estos iones. El ion de potasio atraviesa la membrana libremente; la permeabilidad para el sodio es menor, y además es expulsado por medio de un transporte activo llamado bomba de sodio-potasio. Las proteínas, debido a su tamaño, no pueden atravesar libremente la membrana. Toda esta dinámica establece una diferencia de potencial en condiciones de reposo, de unos -90mV.
Umbral e iniciación
Los potenciales de acción se desencadenan cuando una despolarización inicial alcanza un umbral. Este potencial umbral varía, pero normalmente está en torno a -55 a -50 milivoltios sobre el potencial de reposo de la célula, lo que implica que la corriente de entrada de iones sodio supera la corriente de salida de iones potasio. El flujo neto de carga positiva que acompaña los iones sodio despolariza el potencial de membrana, desembocando en una apertura de los canales de sodio dependientes de voltaje. Estos canales aportan un flujo mayor de corrientes iónicas hacia el interior, aumentando la despolarización en una retroalimentación positiva que hace que la membrana llegue a niveles de despolarización elevados.
El umbral del potencial de acción puede variar cambiando el equilibrio entre las corrientes de sodio y potasio. Por ejemplo, si algunos de los canales de sodio están inactivos, determinado nivel de despolarización abrirá menos canales de sodio, y aumenta así el

umbral de despolarización necesario para iniciar el potencial de acción. Esta es el principio del funcionamiento del periodo refractario (ver periodo refractario).
Los potenciales de acción son muy dependientes de los equilibrios entre iones sodio y potasio (aunque hay otros iones que contribuyen minoritariamente a los potenciales, como calcio y cloro), y por ello los modelos se hacen utilizando sólo dos canales iónicos transmembrana: un canal de sodio dependiente de voltaje y un canal de potasio pasivo. El origen del umbral del potencial de acción puede visualizarse en la curva I/V (imagen) que representa las corrientes iónicas a través de los canales frente al potencial de membrana. (La curva I/V representada en la imagen es una relación instantánea entre corrientes. Se muestra el pico de corrientes a determinado voltaje, registrado antes de que ocurra ninguna inactivación (1 ms tras alcanzar ese voltaje para el sodio). También es importante apuntar que la mayoría de voltajes positivos del gráfico sólo pueden conseguirse por medios artificiales, mediante la aplicación de electrodos a las membranas).
Gráfica de corrientes (flujo iónico) frente a voltaje (potencial de membrana), ilustrando el umbral (flecha roja) del potencial de acción de una célula ideal. Tomado de http://es.wikipedia.org/wiki/Potencial_de_Nernst
Destacan cuatro puntos en la curva I/V indicados por las flechas de la figura:
1. La flecha verde indica el potencial de reposo de la célula y el valor del potencial de equilibrio para el potasio (Ek). Debido a que el canal K+ es el único abierto con esos valores de voltaje negativos, la célula se mantendrá en Ek. Aparecerá un potencial de reposo estable con cualquier voltaje en que el sumatorio I/V (línea verde) cruce el punto de corriente nula (eje x) con una pendiente positiva, como hace en la flecha verde. Esto es debido a que cualquier perturbación del potencial de membrana hacia valores negativos significará corrientes netas de entrada que despolarizarán la célula más allá del punto de cruce, mientras que cualquier perturbación hacia valores positivos significará corrientes netas de salida que hiperpolarizarán la célula. Así, cualquier cambio del potencial de membrana de pendiente positiva tiende a devolver a la célula al valor de cruce con el eje.
2. La flecha amarilla indica el equilibrio del potencial Na+ (ENa). En este sistema de
dos iones, el ENa es el límite natural del potencial de membrana del que la célula no puede pasar. Los valores de corrientes en el gráfico que exceden este límite se han medido de forma artificial obligando a la célula a sobrepasarlo. Aun así, el ENa sólo podría alcanzarse si la corriente de potasio no existiese.
3. La flecha azul indica el voltaje máximo que puede alcanzar el pico del potencial de acción. Es el potencial de membrana máximo que la célula en estado natural puede alcanzar, y no puede llegar al ENa debido a la acción contraria de los flujos de potasio.
4. La flecha roja indica el umbral del potencial de acción. Es el punto donde el Isum se cambia a un flujo neto hacia el interior. Destaca que en este punto se atraviesa el punto de flujo neto cero, pero con pendiente negativa. Cualquier "punto de corte con pendiente negativa" del nivel de flujo cero en el gráfico I/V es un punto inestable. Si el voltaje en este punto es negativo, el flujo va hacia el exterior y la célula tiende a volver al potencial de reposo. Si el voltaje es positivo, el flujo va hacia el interior y tiende a despolarizar la célula. Esta despolarización implica mayor flujo hacia el interior, haciendo que los flujos de sodio se realimenten. El punto en el que la línea verde alcanza el valor más negativo es cuando todos los canales de sodio están abiertos. La despolarización más allá de este punto baja las corrientes de sodio ya que la fuerza eléctrica disminuye a medida que el potencial de membrana se acerca a ENa.
Microfotografía de una red neuronal biológica, cuyas células han sido teñidas de material fosforescente para su mejor identificación.

El umbral del potencial de acción se confunde a veces con el umbral de la apertura de canales de sodio. Es una incorrección, ya que los canales de sodio carecen de umbral. Por el contrario, se abren en respuesta a la despolarización aleatoriamente. La despolarización no implica tanto la apertura de los canales como el incremento de la probabilidad de que se abran. Incluso en potenciales de hiperpolarización, un canal de sodio puede abrirse esporádicamente. Además, el umbral del potencial de acción no es el voltaje a la que el flujo de iones sodio se hace importante; es el punto en que excede el flujo de potasio.
Biológicamente, en las neuronas la despolarización se origina en las sinapsis dendríticas. En principio, los potenciales de acción podrían generarse en cualquier punto a lo largo de la fibra nerviosa. Cuando Luigi Galvani descubrió la electricidad animal haciendo que la pierna de una rana muerta volviese a la vida tocando el nervio ciático con un escalpelo, aplicándole sin darse cuenta una carga electrostática negativa e iniciando un potencial de acción.
Modelo del circuito
A. Un circuito básico RC (resistencia/condensador) superpuesto sobre una membrana bicapa, muestra la relación entre ambos. B. Se pueden utilizar circuitos más elaborados para representar modelos de membranas con canales iónicos, como este ejemplo con canales de sodio (azul) y potasio (verde). Tomado de https://es.wikipedia.org/wiki/Potencial_de_acci%C3%B3n
Las membranas celulares con canales iónicos pueden representarse con un modelo de circuito RC para entender mejor la propagación de potenciales de acción en membranas biológicas. En estos circuitos, la resistencia representa los canales iónicos de membrana, mientras que el condensador representa el aislamiento de la membrana lipídica. Los potenciómetros indican los canales iónicos regulados por voltaje, ya que su valor cambia con el voltaje. Una resistencia de valor fijo representa los canales de potasio que mantienen el potencial de reposo. Los gradientes de sodio y potasio se indican en el modelo como fuentes de voltaje (pila).
Propagación
En los axones amielínicos, los potenciales de acción se propagan como una interacción pasiva entre la despolarización que se desplaza por la membrana y los canales de sodio regulados por voltaje.

Cuando una parte de la membrana celular se despolariza lo suficiente como para que se abran los canales de sodio dependientes de voltaje, los iones de sodio entran en la célula por difusión facilitada. Una vez dentro, los iones positivos de sodio impulsan los iones próximos a lo largo del axón por repulsión electrostática, y atraen los iones negativos desde la membrana adyacente.
Como resultado, una corriente positiva se desplaza a lo largo del axón, sin que ningún ion se esté desplazando muy rápido. Una vez que la membrana adyacente está suficientemente despolarizada, sus canales de sodio dependientes de voltaje se abren, realimentando el ciclo. El proceso se repite a lo largo del axón, generándose un nuevo potencial de acción en cada segmento de la membrana.
Los potenciales de acción de membrana pueden representarse uniendo varios circuitos RC, cada uno representando un trozo de membrana. Tomado de https://es.wikipedia.org/wiki/Potencial_de_acci%C3%B3n
Velocidad de propagación
Los potenciales de acción se propagan más rápido en axones de mayor diámetro, si los demás parámetros se mantienen. La principal razón para que ocurra es que la resistencia axial de la luz del axón es menor cuanto mayor sea el diámetro, debido a la mayor relación entre superficie total y superficie de membrana en un corte transversal. Como la superficie de la membrana es el obstáculo principal para la propagación del potencial en axones

amielínicos, el incremento de esta tasa es una forma especialmente efectiva de incrementar la velocidad de la transmisión.
Un ejemplo extremo de un animal que utiliza el aumento de diámetro de axón como regulador de la velocidad de propagación del potencial de membrana es el calamar gigante. El axón del calamar gigante controla la contracción muscular asociada con la respuesta de evasión de depredadores del animal. Este axón puede sobrepasar 1 mm de diámetro, y posiblemente sea una adaptación para permitir una activación muy rápida del mecanismo de escape. La velocidad de los impulsos nerviosos en estas fibras es una de las más rápidas de la naturaleza, para los que poseen neuronas amielínicas.
Conducción saltatoria
En axones mielínicos, la conducción saltatoria es el proceso por el que los potenciales de acción parecen saltar a lo largo del axón, siendo regenerados sólo en unos anillos no aislados (los nodos de Ranvier16). La conducción saltatoria incrementa la velocidad de conducción nerviosa sin tener que incrementar significativamente el diámetro del axón.
Ha desempeñado un papel importante en la evolución de organismos más complejos cuyos sistemas nerviosos necesitan transmitir rápidamente potenciales de acción a largas distancias. Sin conducción saltatoria, la velocidad de conducción requeriría incrementos
16 En la fisiología de una neurona se llaman nodos de Ranvier a las interrupciones que ocurren a intervalos regulares a lo largo de la longitud del axón en la vaina de mielina que lo envuelve. Son pequeñísimos espacios, de un micrómetro de longitud, que exponen a la membrana del axón al líquido extracelular. Fueron descubiertos por el patólogo francés y anatomista Louis-Antoine Ranvier (1835-1922).
Sirven para que el impulso nervioso se traslade con mayor velocidad, de manera saltatoria y con menor posibilidad de error. La aislación que provee la célula de Schwann (en el sistema nervioso periférico) o eloligodendrocito (en el sistema nervioso central) hace posible que el potencial de acción salte de un nodo al siguiente, a diferencia de tener que hacerlo a pequeños incrementos a lo largo del axón si estuviere desprovisto de la vaina de mielina. [Instituto Latinoamericano de la Comunicación Educativa]
El potencial de acción de la neurona logra saltar de un nodo al otro debido a que éstos, a diferencia del resto de la neurona, están dotados de agrupaciones de canales de sodio y potasio.

drásticos en el diámetro del axón, a tal punto que podrían resultar en la formación de sistemas nerviosos excesivamente grandes para los cuerpos que deben alojarlos.
Mecanismo detallado
El principal obstáculo para la velocidad de transmisión en axones amielínicos es la capacitancia de la membrana. La capacidad de un condensador puede disminuirse bajando el área de un corte transversal de sus placas, o incrementando la distancia entre las placas. El sistema nervioso utiliza la mielinización para reducir la capacitancia de la membrana. La mielina es una vaina protectora creada alrededor de los axones por las células de Schwann17 y los oligodendrocitos18, células de la neuroglía que aplastan sus citoplasmas formando láminas de membrana y plasma. Estas láminas se arrollan en el axón, alejando las placas conductoras (el plasma intra y extracelular) entre sí, disminuyendo la capacitancia de la membrana.
El aislamiento resultante redunda en una conducción rápida (prácticamente instantánea) de los iones a través de las secciones mielinizadas del axón, pero impide la generación de potenciales de acción en estos segmentos. Los potenciales de acción sólo se vuelven a producir en los nodos de Ranvier desmielinizados, que se sitúan entre los segmentos mielinizados. En estos anillos hay un gran número de canales de sodio dependientes de
17 Las Células de Schwann son células gliales periféricas que se forman en la cresta neural embrionaria y acompañan a la neurona durante su crecimiento y desarrollo. Recubren a las prolongaciones (axones) de las neuronas formándoles una vaina aislante de mielina.
Acompañando a los axones nerviosos de pequeño diámetro, las células de Schwann son amielínicas (sin mielina) y pueden alojar varios axones. [Escuela de Medicina, Pontificia Universidad Católica de Chile]
Por el contrario, los axones de mayor diámetro, son envueltos por células de Schwann mielínicas, es decir que sí poseen mielina. A lo largo de los axones, en su envoltura mielínica se producen bandas circulares sin mielina, lo cual parece coincidir con el límite entre las células de Schwann. Estas bandas sin mielina se llaman nodos de Ranvier. La mielina se compone de capas concéntricas de la membrana de las células de Schwann que rodean de manera espiral al axón de la neurona. (ver ilustración) Fuente. Células de Schwann http://es.wikipedia.org/wiki/C%C3%A9lula_de_Schwann [On Line] consultado el 20 de julio de 2014.
18 Los oligodendrocitos con células gliares del sistema nervioso central que recubren los axones mielinicos y amielinicos de las neuronas.

voltaje (hasta cuatro órdenes de magnitud superior a la densidad de axones amielínicos), que permiten que los potenciales de acción se regeneren de forma eficaz en ellos.
Debido a la mielinización, los segmentos aislados del axón actúan como un cable pasivo: conducen los potenciales de acción rápidamente porque la capacitancia de la membrana es muy baja, y minimizan la degradación de los potenciales de acción porque la resistencia de la membrana es alta. Cuando esta señal que se propaga de forma pasiva alcanza un nodo de Ranvier, inicia un potencial de acción que viaja de nuevo de forma pasiva hasta que alcanza el siguiente nodo, repitiendo el ciclo.
Minimización de daños
La longitud de los segmentos mielinizados de un axón es importante para la conducción saltatoria. Deben ser tan largos como sea posible para optimizar la distancia de la conducción pasiva, pero no lo suficiente como para que la disminución en la intensidad de la señal sea tanta que no alcance el umbral de sensibilidad en el siguiente nodo de Ranvier. En realidad, los segmentos mielinizados son lo suficientemente largos para que la señal que se propaga pasivamente recorra al menos dos segmentos manteniendo una amplitud de señal suficiente como para iniciar un potencial de acción en el segundo o tercer nodo. Así se eleva el factor de seguridad de la conducción saltatoria, permitiendo que la transmisión traspase nodos en caso de que estén dañados.
Enfermedad
Algunas enfermedades afectan la conducción saltatoria y disminuyen la velocidad de desplazamiento de un potencial de acción. La más conocida de todas estas enfermedades es la esclerosis múltiple, en la que los daños en la mielina imposibilitan el movimiento coordinado.
Periodo Refractario
Se define como el momento en el que la célula excitable no responde ante un estímulo y por lo tanto no genera un nuevo Potencial de Acción. Se divide en dos: Periodo Refractario Absoluto (o Efectivo) y Periodo Refractario Relativo.
El Periodo Refractario Absoluto es aquel en el que los Canales de Na+ sensibles a voltaje se encuentran "inactivados", por lo que se inhibe el transporte de iones sodio.
En cambio el Periodo Refractario Relativo se da en alguna parte de la Fase de Repolarización, en donde los Canales de Na+ paulatinamente comienzan a cerrarse para así comenzar a abrirse y transportar nuevamente sodio, por lo que al agregar un estímulo

excitatorio muy intenso se puede provocar que los canales que se encuentran cerrados en ese momento se abran y generen un nuevo Potencial de Acción. El Periodo Refractario Relativo termina después de la fase de Hiperpolarización (o Postpotencial) en donde todos los Canales de Na+ sensibles a Voltaje están cerrados y disponibles para un nuevo estímulo.
También existe un Período Refractario Efectivo, que sólo se observa en las células musculares cardíacas (esto se debe a que las células se encuentran formando un sincitio celular). En este caso, la célula se despolariza normalmente, pero no puede conducir dicho estímulo a las células vecinas a ella. Este período refractario, es un parámetro muy útil en la evaluación de drogas antiarrítmicas.
El Periodo Refractario varía de célula a célula, y es una de las características que permiten decir si una célula es más o menos excitable que otra. En otros casos como el músculo cardiaco, su amplio Periodo Refractario le permite la increíble capacidad de no tetanizarse.
Ventaja evolutiva
Con respecto a las causas por las que la naturaleza ha desarrollado esta forma de comunicación, esto se responde al considerar la transmisión de información a gran distancia a través de un axón nervioso. Para llevar la información de un extremo a otro del axón, han de superarse leyes físicas como las que condicionan el movimiento de señales eléctricas en un cable. Debido a la resistencia eléctrica y a la capacitancia del cable, las señales tienden a degradarse a lo largo del cable. Estas propiedades denominadas propiedades del cable determinan los límites físicos a los que pueden llegar las señales.
El correcto funcionamiento del cuerpo necesita que las señales lleguen de un extremo a otro del axón sin pérdidas en el camino. Un potencial de acción no sólo se propaga a lo largo del axón, sino que se regenera por el potencial de membrana y corrientes iónicas en cada estrechamiento de la membrana en su camino. En otras palabras, la membrana nerviosa regenera el potencial de membrana en toda su amplitud a medida que la señal recorre el axón, superando los límites que impone la teoría de líneas de transmisión.
Sinapsis [30]
La sinapsis (del gr. σύναψις, "enlace") es una unión intercelular especializada entre neuronas1 o entre una neurona y una célula efectora (casi siempre glandular o muscular). En estos contactos se lleva a cabo la transmisión del impulso nervioso. Éste se inicia con una descarga química que origina una corriente eléctrica en la membrana de la célula presináptica (célula emisora); una vez que este impulso nervioso alcanza el extremo del axón (la conexión con la otra célula), la propia neurona segrega un tipo de compuestos químicos (neurotransmisores) que se depositan en el espacio sináptico (espacio intermedio entre esta neurona transmisora y la neurona postsináptica o receptora). Estas sustancias

segregadas o neurotransmisores (noradrenalina y acetilcolina19 entre otros) son los encargados de excitar o inhibir la acción de la otra célula llamada célula post sináptica.
Estos enlaces químico-eléctricos están especializados en el envío de cierto tipo de señales de pervivencia, las cuales afectan a otras neuronas, a células no neuronales como las musculares o glandulares.
Existen dos tipos de actividad base distinta, la actividad de pervivencia y la actividad de supervivencia.
La actividad sináptica de pervivencia se desarrolla en estos contextos:
• Entre dos neuronas: al estímulo lo portan los neurotransmisores de tipo aminoácido. • Entre una neurona y una célula muscular: al estímulo lo portan los
neurotransmisores de tipo éster. • Entre una neurona y una célula secretora: al estímulo lo portan los
neurotransmisores de tipo neuropéptido.
La actividad sináptica de supervivencia se desarrolla en estos contextos:
• En la actividad procreadora. • En la actividad de consumo alimenticio. • En la actividad de conservación homeostática extrema.
La sinapsis se produce en el momento en que se registra actividad químico-eléctrica presináptica y otra postsináptica. Si esta condición no se da, no se puede hablar de sinapsis." En dicha acción se liberan neurotransmisores" ionizados con base química, cuya cancelación de carga provoca la activación de receptores específicos que, a su vez, generan otro tipo de respuestas químico-eléctricas.
Cada neurona se comunica, al menos, con otras mil neuronas y puede recibir,
19 La acetilcolina (ACh o ACo) es un neurotransmisor que fue aislado y caracterizada farmacológicamente por Henry Hallett Dale en 1914, y confirmada como un neurotransmisor (el primero en ser identificado) por Otto Loewi; por su trabajo recibieron en 1936 el premio Nobel en fisiología y medicina.
La acetilcolina está ampliamente distribuida en el sistema nervioso central y en el sistema nervioso periférico. Su función, al igual que otros neurotransmisores, es mediar en la actividad sináptica del sistema nervioso.

simultáneamente, hasta diez veces más conexiones de otras. Se estima que en el cerebro humano adulto hay por lo menos 1014 conexiones sinápticas (aproximadamente, entre 100 y 500 billones). En niños alcanza los 1000 billones. Este número disminuye con el paso de los años, estabilizándose en la edad adulta.
Las sinapsis permiten a las neuronas del sistema nervioso central formar una red de circuitos neuronales. Son cruciales para los procesos biológicos que subyacen bajo la percepción y el pensamiento. También son el sistema mediante el cual el sistema nervioso conecta y controla todos los sistemas del cuerpo.
Sinapsis tripartita
De acuerdo con las últimas investigaciones relacionadas con los astrocitos, esta sinapsis constaría de tres elementos: los pre y postsinápticos neuronales y los astrocitos cercanos, que funcionarían como reguladores en la transferencia de información en el interior del sistema nervioso.
Histología
Desde el punto de vista histológico y funcional, una neurona tiene tres zonas principales: el cuerpo o soma, las dendritas y el axón. Estos dos últimos elementos son los encargados de establecer las relaciones sinápticas: las dendritas son como antenas o tentáculos que reciben la mayoría de la información que proviene de otras células; el axón, por su parte, es el cable con el que una neurona se conecta a otras.
Las conexiones pueden establecerse a muy corto alcance, a unos cientos de micrómetros a la redonda, o a distancias mayores. La moto neuronas de la espina dorsal, por ejemplo, se comunican directamente con órganos como los músculos para dar lugar al movimiento (sinapsis neuromuscular).
Una sinapsis prototípica, como las que aparecen en los botones dendríticos, consiste en unas proyecciones citoplasmáticas con forma de hongo desde cada célula que, al juntarse, los extremos de ambas se aplastan uno contra otro. En esta zona, las membranas celulares de ambas células se juntan en una unión estrecha que permite a las moléculas de señal llamadas neurotransmisores y pasar rápidamente de una a otra célula por difusión. El canal de unión de la neurona postsináptica es de aproximadamente 20 nm de ancho, y se conoce como hendidura sináptica.
Estas sinapsis son asimétricas tanto en su estructura como en su funcionamiento. Sólo la neurona presináptica segrega los neurotransmisores, que se unen a los receptores transmembrana que la célulapostsináptica tiene en la hendidura. El terminal nervioso presináptico (también llamado botón sináptico o botón) normalmente emerge del extremo de un axón, mientras que la zona postsináptica normalmente corresponde a una dendrita, al cuerpo celular o a otras zonas celulares.
La zona de la sinapsis donde se libera el neurotransmisor se denomina zona activa. En las zonas activas, las membranas de las dos células adyacentes están unidas estrechamente mediante proteínas de adhesión celular. Justo tras la membrana de la célula postsináptica aparece un complejo de proteínas entrelazadas denominado densidad postsináptica.
Las proteínas de la densidad postsináptica cumplen numerosas funciones, que van desde el anclaje y movimiento de receptores de neurotransmisores de la membrana plasmática, hasta el anclaje de varias proteínas reguladoras de la actividad de estos receptores.
Tipos de sinapsis
Sinapsis eléctrica
Una sinapsis eléctrica es aquella en la que la transmisión entre la primera neurona y la segunda no se produce por la secreción de un neurotransmisor, como en las sinapsis químicas (véase más abajo), sino por el paso de iones de una célula a otra a través de uniones gap, pequeños canales formados por el acoplamiento de complejos proteicos, basados en conexinas, en células estrechamente adheridas.
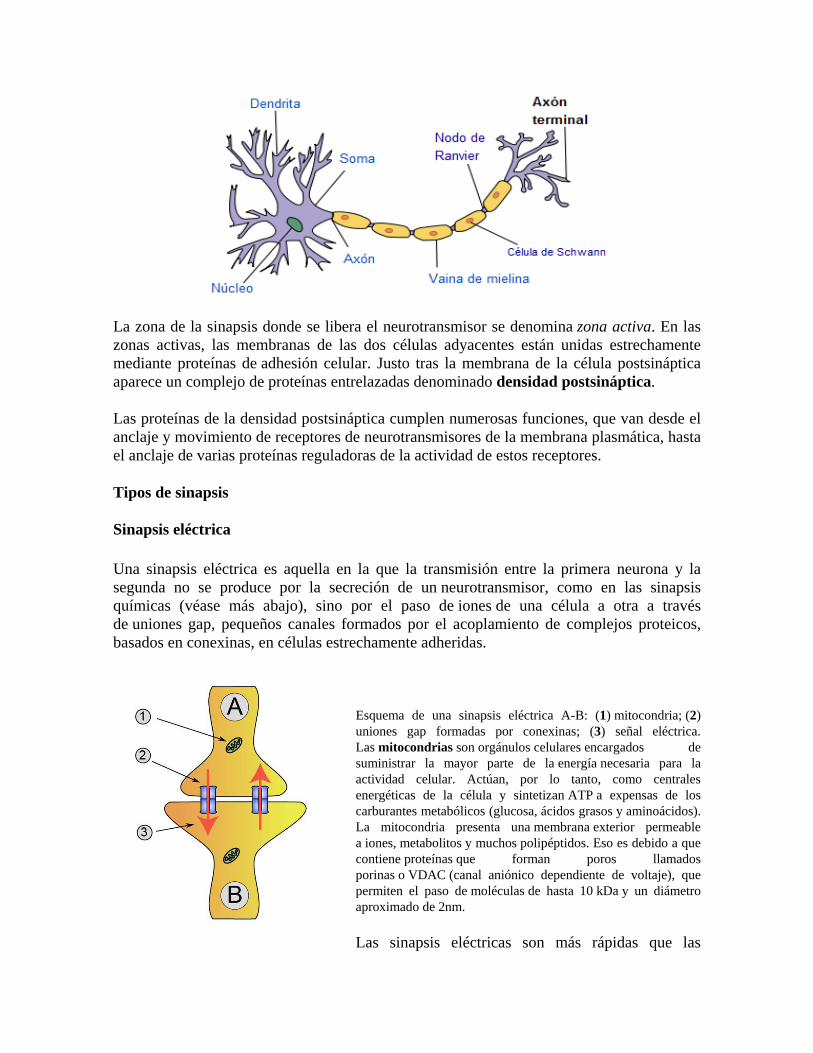
Esquema de una sinapsis eléctrica A-B: (1) mitocondria; (2) uniones gap formadas por conexinas; (3) señal eléctrica. Las mitocondrias son orgánulos celulares encargados de suministrar la mayor parte de la energía necesaria para la actividad celular. Actúan, por lo tanto, como centrales energéticas de la célula y sintetizan ATP a expensas de los carburantes metabólicos (glucosa, ácidos grasos y aminoácidos). La mitocondria presenta una membrana exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros llamados porinas o VDAC (canal aniónico dependiente de voltaje), que permiten el paso de moléculas de hasta 10 kDa y un diámetro aproximado de 2nm.
Las sinapsis eléctricas son más rápidas que las

sinapsis químicas pero menos plásticas; por lo demás, son menos propensas a alteraciones o modulación porque facilitan el intercambio entre los citoplasmas de iones y otras sustancias químicas. En los vertebrados son comunes en el corazón y el hígado.
Las sinapsis eléctricas tienen tres ventajas muy importantes:
1. Las sinapsis eléctricas poseen una transmisión bidireccional de los potenciales de acción, en cambio la sinapsis química solo posee la comunicación unidireccional.
2. En la sinapsis eléctrica hay una sincronización en la actividad neuronal lo cual hace posible una coordinada acción entre ellas.
3. La comunicación es más rápida en la sinapsis eléctricas que en las químicas, debido a que los potenciales de acción pasan a través del canal proteico directamente sin necesidad de la liberación de los neurotransmisores.
Sinapsis química
La sinapsis química se establece entre células que están separadas entre sí por un espacio de unos 20-30 nanómetros (nm), la llamada hendidura sináptica.
La liberación de neurotransmisores es iniciada por la llegada de un impulso nervioso (o potencial de acción), y se produce mediante un proceso muy rápido de secreción celular: en el terminal nervioso presináptico, las vesículas que contienen los neurotransmisores permanecen ancladas y preparadas junto a la membrana sináptica. Cuando llega un potencial de acción se produce una entrada de iones calcio a través de los canales de calcio dependientes de voltaje.
La suma de los impulsos excitatorios e inhibitorios que llegan por todas las sinapsis que se relacionan con cada neurona (1000 a 200.000) determina si se produce o no la descarga del potencial de acción por el axón de esa neurona.

Los iones de calcio inician una cascada de reacciones que terminan haciendo que las membranas vesiculares se fusionen con la membrana presináptica y liberando su contenido a la hendidura sináptica. Los receptores del lado opuesto de la hendidura se unen a los neurotransmisores y fuerzan la apertura de los canales iónicos cercanos de la membrana postsináptica, haciendo que los iones fluyan hacia o desde el interior, cambiando el potencial de membrana local. El resultado es excitatorio en caso de flujos de despolarización, o inhibitorio en caso de flujos dehiperpolarización. El que una sinapsis sea excitatoria o inhibitoria depende del tipo o tipos de iones que se canalizan en los flujos postsinápticos, que a su vez es función del tipo de receptores y neurotransmisores que intervienen en la sinapsis.
Clases de transmisión sináptica
Se distinguen tres tipos principales de transmisión sináptica; los dos primeros mecanismos constituyen las fuerzas principales que rigen en los circuitos neuronales:
• transmisión excitadora: aquella que incrementa la posibilidad de producir un potencial de acción;
• transmisión inhibidora: aquella que reduce la posibilidad de producir un potencial de acción;
• transmisión moduladora: aquella que cambia el patrón y/o la frecuencia de la actividad producida por las células involucradas.
Fuerza sináptica
La fuerza de una sinapsis viene dada por el cambio del potencial de membrana que ocurre cuando se activan los receptores de neurotransmisores postsinápticos. Este cambio de voltaje se denomina potencial postsináptico, y es resultado directo de los flujos iónicos a través de los canales receptores postsinápticos. Los cambios en la fuerza sináptica pueden ser a corto plazo y sin cambios permanentes en las estructuras neuronales, con una duración de segundos o minutos, o de larga duración (potenciación a largo plazo o LTP), en que la activación continuada o repetida de la sinapsis implica que los segundos mensajeros inducen la síntesis proteica en el núcleo de la neurona, alterando la estructura de la propia neurona. El aprendizaje y la memoria podrían ser resultado de cambios a largo plazo en la fuerza sináptica, mediante un mecanismo de plasticidad sináptica.
Integración de señales sinápticas Generalmente, si una sinapsis excitatoria es fuerte, un potencial de acción en la neurona presináptica iniciará otro potencial en la célula postsináptica. En una sinapsis débil, el potencial excitatorio postsináptico ("PEPS")20 no alcanzará el umbral para la iniciación
20 Un potencial excitatorio postsináptico (PEPS) es un incremento temporal en el potencial de membrana postsináptico causado por el flujo de iones cargados positivamente hacia dentro de la célula postsináptica. Existen los potenciales inhibitorios postsinápticos (PIPS), que normalmente se originan con el flujo de iones negativos hacia el lúmen celular. Los PEPS pueden también originarse por un descenso en el
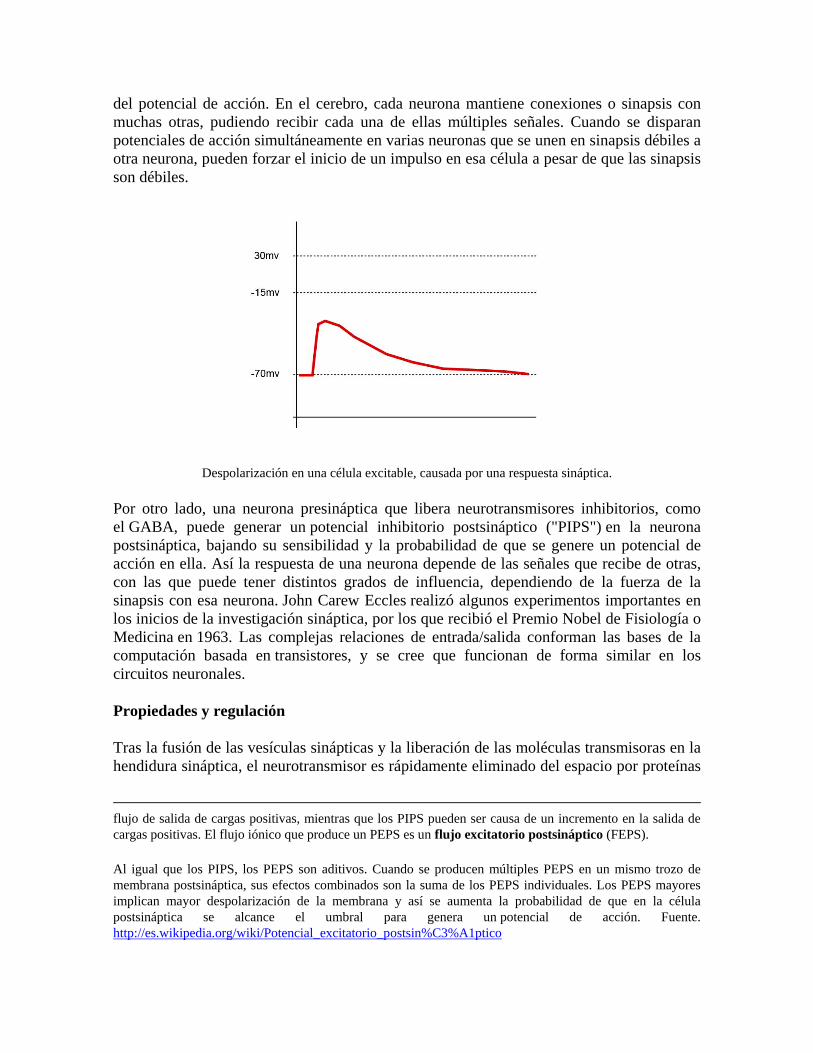
del potencial de acción. En el cerebro, cada neurona mantiene conexiones o sinapsis con muchas otras, pudiendo recibir cada una de ellas múltiples señales. Cuando se disparan potenciales de acción simultáneamente en varias neuronas que se unen en sinapsis débiles a otra neurona, pueden forzar el inicio de un impulso en esa célula a pesar de que las sinapsis son débiles.
Despolarización en una célula excitable, causada por una respuesta sináptica.
Por otro lado, una neurona presináptica que libera neurotransmisores inhibitorios, como el GABA, puede generar un potencial inhibitorio postsináptico ("PIPS") en la neurona postsináptica, bajando su sensibilidad y la probabilidad de que se genere un potencial de acción en ella. Así la respuesta de una neurona depende de las señales que recibe de otras, con las que puede tener distintos grados de influencia, dependiendo de la fuerza de la sinapsis con esa neurona. John Carew Eccles realizó algunos experimentos importantes en los inicios de la investigación sináptica, por los que recibió el Premio Nobel de Fisiología o Medicina en 1963. Las complejas relaciones de entrada/salida conforman las bases de la computación basada en transistores, y se cree que funcionan de forma similar en los circuitos neuronales.
Propiedades y regulación
Tras la fusión de las vesículas sinápticas y la liberación de las moléculas transmisoras en la hendidura sináptica, el neurotransmisor es rápidamente eliminado del espacio por proteínas
flujo de salida de cargas positivas, mientras que los PIPS pueden ser causa de un incremento en la salida de cargas positivas. El flujo iónico que produce un PEPS es un flujo excitatorio postsináptico (FEPS).
Al igual que los PIPS, los PEPS son aditivos. Cuando se producen múltiples PEPS en un mismo trozo de membrana postsináptica, sus efectos combinados son la suma de los PEPS individuales. Los PEPS mayores implican mayor despolarización de la membrana y así se aumenta la probabilidad de que en la célula postsináptica se alcance el umbral para genera un potencial de acción. Fuente. http://es.wikipedia.org/wiki/Potencial_excitatorio_postsin%C3%A1ptico

especializadas en su reciclaje, situadas en las membranas tanto presináptica como postsináptica. Esta recaptación evita la desensibilización de los receptores postsinápticos y asegura que los potenciales de acción subsiguientes generen un PEP de la misma intensidad. La necesidad de una recaptación y el fenómeno de la desensibilización en los receptores y canales iónicos significa que la fuerza de la sinapsis puede disminuir si un tren de potenciales de acción llega en una sucesión rápida, un fenómeno que hace que exista una dependencia de la frecuencia en las sinapsis. El sistema nervioso se aprovecha de esta propiedad para computaciones, y puede ajustar las sinapsis mediante la fosforilación de las proteínas implicadas. El tamaño, número y tasa de reposición de las vesículas también está sujeto a regulación, así como otros muchos aspectos de la transmisión sináptica. Por ejemplo, un tipo de fármaco conocido como inhibidores selectivos de la recaptación de serotonina o SSRI afectan a ciertas sinapsis inhibiendo la recaptación del neurotransmisor serotonina. Por el contrario, un neurotransmisor excitatorio muy importante, la acetilcolina, no es recaptada, pero es eliminada por acción de la enzima acetilcolinesterasa.
El papel de las sinapsis en los fenómenos plásticos
La modificación de los parámetros sinápticos puede modificar el comportamiento de los circuitos neurales y la interacción entre los diferentes módulos que componen el sistema nervioso (modal). Dichos cambios están englobados en un fenómeno conocido como neuroplasticidad.
La plasticidad neuronal, también denominada neuroplasticidad, plasticidad neural o plasticidad sináptica, es la propiedad que emerge de la naturaleza y funcionamiento de las neuronas cuando éstas establecen comunicación, y que modula la percepción de los estímulos con el medio, tanto los que entran como los que salen. [31] Esta dinámica deja una huella al tiempo que modifica la eficacia de la transferencia de la información a nivel de los elementos más finos del sistema. Dichas huellas son los elementos de construcción de la cosmovisión, [32] en donde lo anterior modifica la percepción de lo siguiente [33] o también una neurona plástica.
El cerebro es considerado, en el caso de la plasticidad neuronal, como un órgano extremadamente dinámico en permanente relación con el ambiente, por un lado, y con los hechos psíquicos o los actos del sujeto, por otro. [34] Esto significa que la red neuronal es extremadamente sensible a los cambios y a la contingencia. La interacción de los diferentes acontecimientos acaecidos en las distintas zonas de la psiquis, modula el acontecimiento y las potencialidades de la experiencia, que siempre pueden modificar el estado anterior. [35]
Plasticidad sináptica a corto plazo en vertebrados
Lo más probable es que todas las sinapsis químicas son capaces de sufrir cambios plásticos. Los mecanismos de la plasticidad sináptica en la sinapsis de los mamíferos se desarrollan en escalas temporales que varían desde los milisegundos hasta días, semanas o más. Las formas de plasticidad a corto plazo (duran minutos o menos) se han estudiado con más detalle en las sinapsis musculares periféricas. La activación repetida de la unión neuromuscular desencadena varios cambios que varían en dirección y duración.
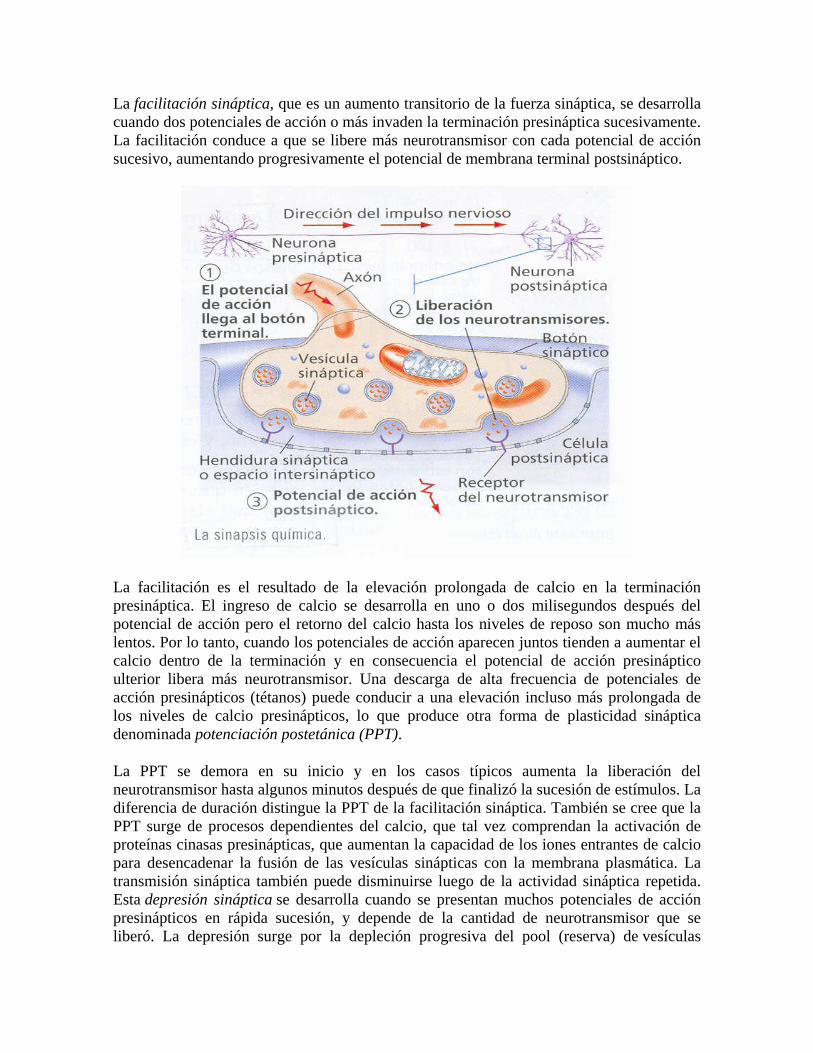
La facilitación sináptica, que es un aumento transitorio de la fuerza sináptica, se desarrolla cuando dos potenciales de acción o más invaden la terminación presináptica sucesivamente. La facilitación conduce a que se libere más neurotransmisor con cada potencial de acción sucesivo, aumentando progresivamente el potencial de membrana terminal postsináptico.
La facilitación es el resultado de la elevación prolongada de calcio en la terminación presináptica. El ingreso de calcio se desarrolla en uno o dos milisegundos después del potencial de acción pero el retorno del calcio hasta los niveles de reposo son mucho más lentos. Por lo tanto, cuando los potenciales de acción aparecen juntos tienden a aumentar el calcio dentro de la terminación y en consecuencia el potencial de acción presináptico ulterior libera más neurotransmisor. Una descarga de alta frecuencia de potenciales de acción presinápticos (tétanos) puede conducir a una elevación incluso más prolongada de los niveles de calcio presinápticos, lo que produce otra forma de plasticidad sináptica denominada potenciación postetánica (PPT).
La PPT se demora en su inicio y en los casos típicos aumenta la liberación del neurotransmisor hasta algunos minutos después de que finalizó la sucesión de estímulos. La diferencia de duración distingue la PPT de la facilitación sináptica. También se cree que la PPT surge de procesos dependientes del calcio, que tal vez comprendan la activación de proteínas cinasas presinápticas, que aumentan la capacidad de los iones entrantes de calcio para desencadenar la fusión de las vesículas sinápticas con la membrana plasmática. La transmisión sináptica también puede disminuirse luego de la actividad sináptica repetida. Esta depresión sináptica se desarrolla cuando se presentan muchos potenciales de acción presinápticos en rápida sucesión, y depende de la cantidad de neurotransmisor que se liberó. La depresión surge por la depleción progresiva del pool (reserva) de vesículas

sinápticas disponibles para la fusión en esta circunstancia. Durante la depresión sináptica, la fuerza de la sinapsis declina hasta que este pool puede recuperarse mediante los mecanismos involucrados el reciclado de las vesículas sinápticas.
Plasticidad sináptica a largo plazo en vertebrados
La facilitación, la depresión y la potenciación postetánica pueden modificar brevemente la transmisión sináptica pero no pueden proporcionar las bases para las memorias u otras manifestaciones de plasticidad conductual que persisten durante meses, semanas o años. Algunos patrones de actividad sináptica en el sistema nervioso central producen un aumento prolongado en la fuerza sináptica conocido como potenciación a largo plazo (PLP), mientras que otros patrones de actividad generan una disminución prolongada de la fuerza sináptica, conocida como depresión a largo plazo (DLP). [36]
Sinapsis inmunitarias
Por analogía con las sinapsis descritas, el encuentro entre una célula antigénica y un linfocito se denomina a veces sinapsis inmunitaria.
Las células presentadoras de antígeno (CPA, o por sus siglas en Inglés: APC) son un grupo diverso de células del sistema inmunitario cuya función es la de captar, procesar y, como su nombre los indica, presentar moléculas antigénicas sobre sus membranas para que sean reconocidos, en especial por linfocitos T. El resultado de la interacción entre una CPA y un linfocito T correspondiente inicia las respuestas inmunitarias antigénicas.
Célula glial [37]
Las células gliales (conocidas también genéricamente como glía o neuroglía) son células nodriza del sistema nervioso que desempeñan, de forma principal, la función de soporte de las neuronas; intervienen activamente, además, en el procesamiento cerebral de la información en el organismo.
Las células gliales controlan, fundamentalmente, el microambiente celular en lo que respecta a la composición iónica, los niveles de neurotransmisores y el suministro de citoquinas y otros factores de crecimiento.
La proporción de neuronas y de células gliales en el cerebro varía entre las diferentes especies (aprox. 10:1 en la mosca doméstica, 1:1 en el cocodrilo y 1:10 en el hombre).
La palabra glía deriva del griego bizantino γλία, cuyo significado era "liga", "unión" o, también, «pegamento». [38]
Canal central de la médula espinal, se observan células ependimarias y neurogliales.
La glía cumple funciones de sostén y nutrición (en el sistema nervioso no existe tejido conjuntivo). Estas células han seguido un desarrollo filogénico y ontogénico diferente al de las neuronas. Debido a que son menos diferenciadas que las neuronas, conservan la capacidad mitótica y son las que se encargan de la reparación y regeneración de las lesiones del sistema nervioso.
Son, igualmente, fundamentales en el desarrollo de las redes neuronales desde las fases embrionales, pues desempeñan el papel de guía y control de las migraciones neuronales en las primeras fases de desarrollo; asimismo, establecen la regulación bioquímica del crecimiento y desarrollo de los axones y dendritas.
También, son las encargadas de servir de aislante en los tejidos nerviosos, al conformar las vainas de mielina que protegen y aíslan los axones de las neuronas.
Mantienen las condiciones homeostáticas (oxígeno y nutrientes) y regulan las funciones metabólicas del tejido nervioso, además de proteger físicamente las neuronas del resto de tejidos y de posibles elementos patógenos, al conformar la barrera hematoencefálica.
Aunque por mucho tiempo se consideró a las células gliales como elementos pasivos en la actividad nerviosa, trabajos recientes demuestran que son participantes activas de la transmisión sináptica, actuando como reguladoras de los neurotransmisores (liberando factores como ATP y sus propios neurotransmisores. Además, las células gliales parecen conformar redes “paralelas” con conexiones sinápticas propias (no neuronales). [39]
La glía reactiva
En cuanto se produce un daño en el sistema nervioso, la glía reacciona cambiando su estado normal al de glía reactiva, precediendo por lo general la activación microglia a la astroglial. La reactividad glial tiene inicialmente como objeto el reparar daños y normalizar niveles de nutrimentos y neurotransmisores; sin embargo, termina por generar lesiones secundarias que pueden llegar a cronificar la patología: provoca muerte neuronal secundaria, ampliando

la zona lesionada hasta el punto de verse afectados grupos neuronales que habían quedado intactos hasta el momento.
La glía reactiva presenta, externamente, células de mayor tamaño que cuando están en reposo y que expresan más filamentos intermedios.
El proceso de reactividad glial implica el reclutamiento y coactivación de células inmunitarias procedentes de la sangre.
Clasificación
Clasificación topográfica
Según su ubicación dentro del sistema nervioso, podemos clasificar a las células gliales en dos grandes grupos:
Glía central
Se encuentra en el Sistema Nervioso Central - SNC (cerebro, cerebelo, tronco cerebral y médula espinal):
• Astrocitos • Oligodendrocitos • Microglía • Células Ependimarias
Glía periférica
Se encuentra en el Sistema Nervioso Periférico - SNP, (ganglios nerviosos, nervios y terminaciones nerviosas):
• Células de Schwann • Células capsulares • Células de Müller
Clasificación morfo-funcional
Por su morfología o función, entre las células gliales se distinguen las células macrogliales (astrocitos, oligodendrocitos y células ependimarias) y las células microgliales (entre el 10 y el 15% de la glía).
Microglía
Son menos que las células de macroglia. Se clasifican según la forma:
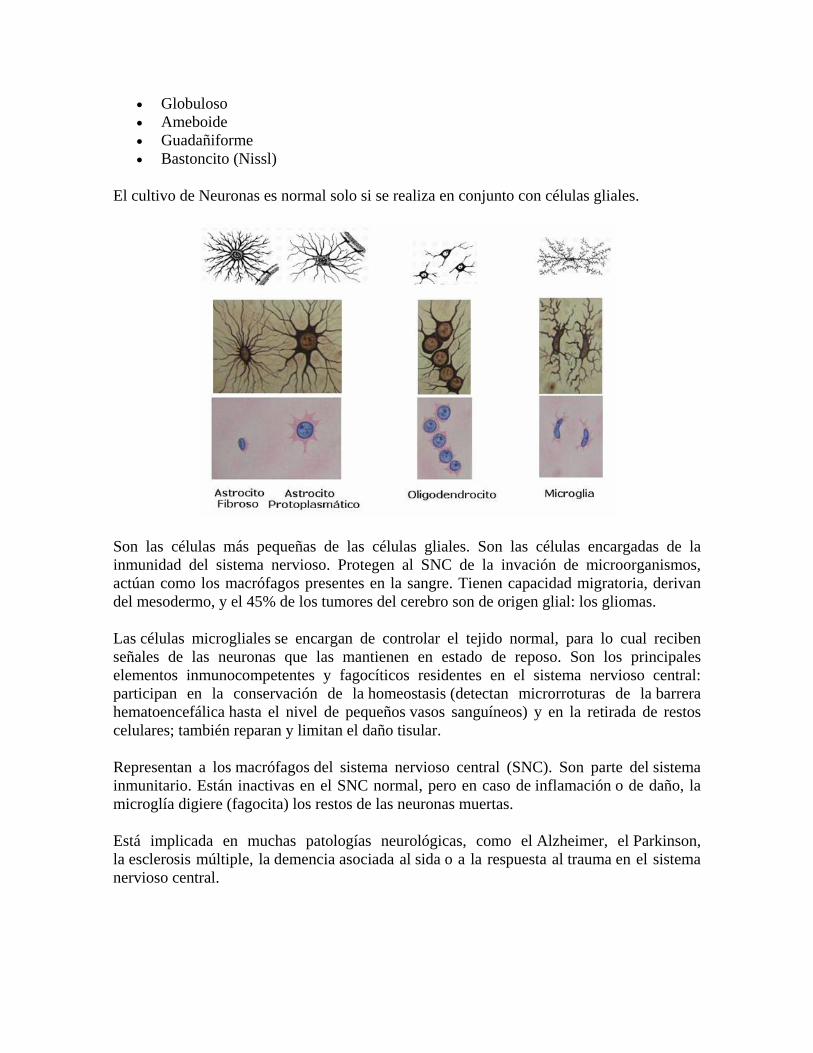
• Globuloso • Ameboide • Guadañiforme • Bastoncito (Nissl)
El cultivo de Neuronas es normal solo si se realiza en conjunto con células gliales.
Son las células más pequeñas de las células gliales. Son las células encargadas de la inmunidad del sistema nervioso. Protegen al SNC de la invación de microorganismos, actúan como los macrófagos presentes en la sangre. Tienen capacidad migratoria, derivan del mesodermo, y el 45% de los tumores del cerebro son de origen glial: los gliomas.
Las células microgliales se encargan de controlar el tejido normal, para lo cual reciben señales de las neuronas que las mantienen en estado de reposo. Son los principales elementos inmunocompetentes y fagocíticos residentes en el sistema nervioso central: participan en la conservación de la homeostasis (detectan microrroturas de la barrera hematoencefálica hasta el nivel de pequeños vasos sanguíneos) y en la retirada de restos celulares; también reparan y limitan el daño tisular.
Representan a los macrófagos del sistema nervioso central (SNC). Son parte del sistema inmunitario. Están inactivas en el SNC normal, pero en caso de inflamación o de daño, la microglía digiere (fagocita) los restos de las neuronas muertas.
Está implicada en muchas patologías neurológicas, como el Alzheimer, el Parkinson, la esclerosis múltiple, la demencia asociada al sida o a la respuesta al trauma en el sistema nervioso central.

Fueron descritas por vez primera por F. Robertson y Franz Nissl como staebchenzellen, esto es, células alargadas. Pío del Río Hortega las diferenció de las otras células gliales y les dio su nombre actual.
Macroglía
Representa en número la mayoría de las células gliales.
Se dividen en:
• Astrocitos: 1. Protoplasmáticos
1. Satélites Neuronales 2. Satélites Vasculares
2. Intermedios 3. Fibrosos
1. Sustancia Blanca 2. Hipofisis (pituicitos)
• Oligodendrocito 1. I de Robertson 2. II de Cajal 3. III de Paladino 4. IV de Del Rio Hortega
• Epitelio Ependimario
Componentes del SNC [Sistema Nervioso Central]
• Astrocitos:
Los astrocitos son las principales y más numerosas células gliales, sobre todo en los organismos más evolucionados. Se trata de células de linaje neuroectodérmico21 que asumen un elevado número de funciones clave para la realización de la actividad nerviosa. Derivan de las células encargadas de dirigir la migración de precursores durante el desarrollo (glía radial) y se originan en las primeras etapas del desarrollo del sistema nervioso central.
Se encargan de aspectos básicos para el mantenimiento de la función neuronal, entrelazándose alrededor de la neurona para formar una red de sostén, y actuando así como una barrera filtradora entre la sangre y la neurona. Cuando existe destrucción neuronal (por ejemplo, tras sufrir un accidente cerebro-vascular), también actúan como liberadores del factor de crecimiento nervioso que, a modo de abono biológico, facilita la regeneración de las conexiones neuronales.
21 De una de las tres regiones diferenciadas del embrión: el ectodermo (las otras dos son el endodermo y el mesodermo).

Los astrocitos también son células de cuerpos pequeños y extensos procesos dendriformes. Los protoplasmáticos se encuentran en la sustancia gris y rodean los cuerpos neuronales, las dendritas y a las sinapsis.
Este tipo de célula posee prolongaciones que se adosan a los vasos sanguíneos que se llaman pies chupadores. Al principio, Camilo Golgi, postuló que la función de estas extensiones, teniendo en cuenta su relación con los vasos, servía para extraer nutrientes de la sangre.
Paul Erlich describió una barrera existente entre la circulación sanguínea y el tejido nervioso llamada Barrera Hemato Encefálica (BHE). Comprobó dicha postulación con un sencillo experimento que consistió en inyectar tinta china en el sistema circulatorio, dicha tintura teñía todos los tejidos del cuerpo menos el tejido nervioso.
Si tenemos en cuenta que el consumo neuronal de glucosa es de 5mg de glucosa por 100 gr de tejido en un minuto, podemos inferir en que los nutrientes llegan a las neuronas por otros caminos, ya que los pies chupadores no serían suficientes para semejante demanda. La microscopía electrónica (ME) ha demostrado que solamente el 85% de los vasos estan rodeados por astrocitos.
Las funciones actualmente aceptadas de los Astrocitos son múltiples:
1. Producen el Factor Trófico Neuronal (NGF), factor indispensable para que las neuronas crezcan en cultivo, descubierto por Rosa Moltacini, laboratorios Carlos Chagas en Brazil, premio Nobel 1986.
2. Mantenimiento del fluido extracelular, consumiendo o captando K+ a través de su membrana que es sumamente permeable.
3. Compartimentalizan las Sinapsis, aislándolas de las sinapsis contiguas. 4. Probable participación en la selección de nutrientes.

5. Mantenimiento del equilibrio iónico del tejido extracelular.
Los fibrosos son denominados también células araña �, ya que se as prolongaciones escasas y largas, su citoplasma está repleto de filamentos y se distribuyen mayormente en la sustancia blanca.
• Oligodendrocitos (oligodendroglía)
Los oligodendrocitos o en conjunto oligodendroglía son más pequeños que los astrocitos y tienen pocas prolongaciones. Además de la función de sostén y unión, se encargan de formar la vaina de mielina que envuelve los axones neuronales en el sistema nervioso central. Producen la mielina a nivel central, por lo tanto se encuentran en la sustancia blanca y favorecen la conducción saltatoria. Son las células homólogas a las célulasde Schwann (teloglia) que forman la mielina a nivel periférico y se diferencian de ellas en que los oligodendrocitos mielinizan varios axones a diferencia de las de Schwann que lo hacen solo en uno.
Son células que están siendo ampliamentes estudiadas actualmente con el objeto de buscar tratamiento para enfermedades desmielinizantes del humano.
• Células ependimarias (ependimocitos)
Las células del epitelio ependimario (epéndimocitos, tanicitos) revisten los ventrículos del encéfalo y del conducto ependimario de la médula espinal que contienen al líquido cefaloraquídeo (LCR).
Los tanicitos son células de contacto entre el tercer ventrículo del cerebro y la eminencia media hipotalámica. Su función no es bien conocida, y se les ha atribuido un papel de transporte de sustancias entre el LCR del tercer ventrículo y el sistema porta hipofisiario. Pueden considerarse una variedad especializada de células ependimarias.
Las células del epitelio coroídeo producen líquido cefalorraquídeo (LCR), a nivel de los plexos coroídeos, en los ventrículos cerebrales.
• Células de Müller
Representan el principal componente glial de la retina en los vertebrados. Se relacionan con el desarrollo, organización y función de la retina. Puede que tengan algo que ver con el crecimiento del ojo y que intervengan en la modulación del procesamiento de la información en las neuronas circundantes. Sin embargo, estudios recientes realizados en la Universidad de Leipzig (Alemania) han revelado que las células de Müller cumplen importantes funciones en la retina relacionados con la luz. Éstas actuarían a modo de "filtro" de la luz que incide sobre el ojo, de modo que a la retina llegaría una imagen más nítida de la que entraría si ésta tuviera que atravesar las distintas capas retinales. Pese a que

este descubrimiento no tiene más aplicación que romper el antiguo dogma de la visión en los seres vivos, puede que tenga utilidad al momento de tratar la ceguera.
Componentes del SNP
• Células satélite
Las células satélite, proporcionan soporte físico, protección y nutrición para las neuronas ganglionares de los ganglios nerviosos craneales, espinales y autonómicos en el sistema nervioso periférico - (SNP).
• Células de Schwann
Las células de Schwann se encargan de proporcionar aislamiento (mielina) a las neuronas del sistema nervioso periférico (SNP). Son el equivalente periférico de los oligodendrocitos del SNC. Hay que tener en cuenta que el sistema nervioso central está compuesto por el encéfalo y la médula espinal, y el periférico por los nervios que salen de la médula espinal.
Capacidad de división
Un malentendido común sobre las células gliales es que en su conjunto, estas conservan su capacidad mitótica en el sistema nervioso maduro (a diferencia de las neuronas). Esto debido a la constatación de la aparición y proliferación de tejido glial remplazando las neuronas luego de lesiones o traumas que implican daños neuronales. Los estudios muestran que células gliales maduras y bien diferenciadas, como los astrocitos o los oligodendrocitos, no retienen esta capacidad. Solo las células precursoras de los oligodendrocitos residentes en los tejidos del sistema nervioso maduro conservan esta peculiaridad.
La Neuroglia [40]
Son células no excitables, donde la relación de neuronas y neuroglia es de 1:10. La neuroglia ocupa la mitad del volumen del SN.
Estas células fueron identificadas y bautizadas por Virchow a mediados del siglo XIX. Dijo que tenían un rol pasivo en el Sistema Nervioso.
No tiene axones ni establece sinapsis. No participan en la transmisión de señales rápidas en el SN. Con otra glía tiene contactos de membrana que facilita la transformación de iones, y conserva la capacidad de dividirse durante toda la vida siendo el origen frecuente de tumores benignos y malignos.
Se acumula en el SN donde se produce destrucción de neuronas por causas patógenas.

Una envoltura glial rodearía las neuronas y sus prolongaciones. El numero de neuroglias que rodea una motoneurona varía en función a la prolongación del axón y aumenta con la actividad de la motoneurona.
Neuroglia proviene del griego, neuro: neuronas y glia: unión, cola. Se divide en Macroglia y Microglia.
Funciones de la Neuroglia en general (según Loyber) :
1. Es un tejido de sostén del tejido nervioso. 2. Interviene en la mielinización de los axones. 3. Interviene en la formación de la BHE por intermedio de los pies chupadores o
vasculares de los astrocitos. 4. Cumpliría un papel orientador en la migración de las neuronas durante el desarrollo
del SN. 5. Parecería que controla la concentración de potasio extracelular. 6. Puede absorber o secretar ciertos intermediarios químicos ,como el Gaba y el ácido
glutámico. 7. Posee una gran actividad ATP de membrana aunque no se conoce bien que relación
puede tener ese hecho con la función neuronal. 8. Interviene en la desintoxicación del amoniaco cuyo exceso es perjudicial pata el SN
pudiendo ese exceso originar alteraciones neurológicas. 9. Almacena glucógeno indispensable para el funcionamiento de las neuronas. 10. Tiene capacidad para sintetizar proteínas necesarias para la conservación de los
axones de las neuronas. 11. Los ependimocitos intervienen en la depuración de los desechos neuronales .por sus
cílias contribuyen a la circulación del liquido cefaloraquídeo y también desempeñan un papel activo en su formación.

12. Además de las funciones descriptas, las células neurológicas podrían tener funciones mas elevadas, como la de ser responsable en ciertos tipos de inhibición presináptica y también la de intervenir en los cambios a largo plazo relacionados con la memoria.
Neurotransmisor [41]
Un neurotransmisor (o neuromediador) es una biomolécula que transmite información de una neurona (un tipo de célula del sistema nervioso) a otra neurona consecutiva, unidas mediante una sinapsis. El neurotransmisor se libera por las vesículas en la extremidad de la neurona presináptica durante la propagación del impulso nervioso, atraviesa el espacio sináptico y actúa cambiando el potencial de acción en la neurona siguiente (denominada postsináptica) fijándose en puntos precisos de su membrana plasmática.
La sinapsis permite a las neuronas comunicarse entre sí, transformando una señal eléctrica en otra química.
En sentido estricto según la definición de hormona22 del Nobel de Medicina Roger Guillemin, un neurotransmisor sería una hormona de secreción paracrina23 liberada por las
22 Las hormonas son sustancias secretadas por células especializadas, localizadas en glándulas de secreción interna o glándulas endocrinas (carentes de conductos), o también por células epiteliales e intersticiales cuyo fin es la de afectar la función de otras células. También hay hormonas que actúan sobre la misma célula que las sintetiza (autocrinas). Hay algunas hormonas animales y sexuales que ayudan al funcionamiento de la tiroxitonina hormona glandial que no secreta mayoría de glándulas al cuerpo en la anzima a y su ciclo de reproducción hormonas vegetales como las auxinas, ácido abscísico, citoquinina, giberelina y el etileno.
Son transportadas por vía sanguínea o por el espacio intersticial, solas (biodisponibles) o asociadas a ciertas proteínas (que extienden su vida media al protegerlas de la degradación) y hacen su efecto en determinados partes del cuerpo no hay que olvidar esto en el taxismo y tropismo y sus ciclos cliontologiticos órganos o tejidos diana (o blanco) a distancia de donde se sintetizaron, sobre la misma célula

neuronas. Aunque debido a sus características específicas, el neurotransmisor a menudo es considerado una forma de comunicación celular distinto de las hormonas, aunque la distinción entre uno y otro es difusa.
Una hormona es cualquier sustancia que liberada por una célula actuase sobre otra célula, tanto cercana como lejana, e independientemente de la singularidad o ubicuidad de su origen y sin tener en cuenta la vía utilizada para su transporte, sea circulación sanguínea, flujo axoplasmático o espacio intersticial.
Procesos bioquímicos asociados a la neurotransmisión
• Síntesis del neurotransmisor por las neuronas presinápticas. Participan las células gliales. Según la naturaleza del neurotransmisor, éste se puede sintetizar en el soma neuronal o en las terminaciones nerviosas. Algunos neurotransmisores se sintetizan directamente en las terminaciones nerviosas gracias a enzimas que se han sintetizado en el soma y se han transportado a estas terminaciones. A través del interior del axón fluye una corriente de sustancias libres o encerradas en vesículas, que pueden ser precursores tanto de los neurotransmisores o sus enzimas, llamada flujo axónico.
• Almacenamiento del neurotransmisor en vesículas de la terminación sináptica.
• Liberación del neurotransmisor por exocitosis, que es calciodependiente. Cuando llega un impulso nervioso a la neurona presináptica, ésta abre los canales de calcio, entrando el ion en la neurona y liberándose el neurotransmisor en el espacio sináptico. El calcio además de iniciar la exocitosis, activa el traslado de las vesículas a los lugares de su liberación con la ayuda de proteínas de membrana plasmática y de la membrana vesicular. Cuando entra el calcio en la neurona, se activa una enzima llamada calmodulina que es una proteinquinasa, encargada de fosforilar a la sinapsina I, situada en la membrana de las vesículas y que las une a los filamentos de actina. Cuando la sinapsina I es fosforilada, las vesículas sinápticas se despegan de la actina y se movilizan hacia los sitios donde deban vaciarse. La fusión de la membrana vesicular con la membrana plasmática es un proceso complejo en el que intervienen varias proteínas como la sinaptobrevina, sinaptotagmina, rab-3 (de la membrana vesicular) sintaxina, SNAP-25, n-sec 1 (de la membrana plasmática) y factor sensible a n-etilmaleimida (NSF) con actividad ATP-asa. Este conjunto de proteínas, forman el complejo SNARE que forma un poro en la membrana
que la sintetiza (acción autócrina) o sobre células contiguas (acción paracrina) interviniendo en la comunicación celular.
23 La liberación paracrina se refiere a un tipo de comunicación celular por secreción química que afecta a una célula vecina a la célula emisora, como es el caso de muchas hormonas, por ejemplo. [Sociedad Argentina de Terapia Intensiva] La sustancia secretada difunde en dirección de los receptores específicos sobre las células adyacentes a la célula que la sintetizó. La liberación paracrina es un tipo de comunicación celular que emplea mensajeros químicos.

plasmática y permite la fusión de ambas membranas y la salida de la sustancia como el contenido vesicular al espacio sináptico.
• Activación del receptor del neurotransmisor situado en la membrana seminsefalica de la neurona postsináptica. El receptor postsináptico es una estructura proteica que desencadena una respuesta. Los neurorreceptores pueden ser:
Receptores ionotrópicos: Producen una respuesta rápida al abrir o cerrar canales iónicos, que producen despolarizaciones, generando potenciales de acción, respuestas excitatorias, producen hiperpolarizaciones o respuestas inhibitorias. En el primer caso, actúan canales de cationes monoiónicos como los de Sodio y Potasio, mientras que en el segundo caso, son los canales de Cloruro los que se activan. Receptores metabotrópicos: Liberan mensajeros intracelulares, como AMP cíclico, Calcio, y fosfolípidos por el mecanismo de transducción de señales. Estos segundos mensajeros activan proteínas quinasas, las cuales, fosforilan activando o desactivando canales al interior de la célula. En el caso de una despolarización, son los canales de Potasio que se cierran, en caso de hiperpolarización, los mismos canales son abiertos produciendo el aumento de cationes intracelulares.
• Iniciación de las acciones del segundo mensajero.
• Inactivación del neurotransmisor, ya sea por degradación química o por reabsorción en las membranas. En el espacio sináptico, existen enzimas específicas que inactivan al neurotransmisor. Además, las neuronas presinápticas tienen receptores para el neurotransmisor que lo recaptan introduciéndolo y almacenándolo de nuevo en vesículas para su posterior vertido.
Existen súper familias de receptores para cada uno de los diferentes tipos de neurotransmisores. Las drogas de acción cerebral actúan en alguna o algunas de estas etapas.
Clasificación
Los neurotransmisores se pueden agrupar en neurotrasmisores propiamente dichos, y en neuromoduladores. Estos últimos son sustancias que actúan de forma similar a los neurotransmisores; la diferencia radica en que no están limitados al espacio sináptico, sino que se difunden por el fluido extraneuronal e intervienen directamente en las consecuencias postsinápticas de la neurotransmisión. Teniendo en cuenta su composición química se pueden clasificar en: [42]
• Colinérgicos: acetilcolina • Adrenérgicos: que se dividen a su vez en catecolaminas, ejemplo adrenalina o
epinefrina, noradrenalina o norepinefrina y dopamina; indolaminas serotonina, melatonina e histamina

• Aminoacidérgicos: GABA, taurina, ergotioneina, glicina, beta alanina, glutamato y aspartato.
• Peptidérgicos: endorfina, encefalina, vasopresina, oxitocina, orexina, neuropéptido Y, sustancia P, dinorfina A, somatostatina, colecistoquinina, neurotensina, hormona luteinizante, gastrina y enteroglucagón.
• Radicales libres: oxido nítrico (NO), monóxido de carbono (CO), adenosin trifosfato (ATP) y ácido araquidónico.
Principales neurotransmisores
• Acetilcolina (ACh). Se localizan en: o Neuronas motoras en médula espinal → unión neuromuscular o Proscencéfalo basal → numerosas áreas de la corteza o Interneuronas en el cuerpo estriado o Sistema nervioso autónomo → neuronas preganglionares del SNA simpático
y parasimpático, y postganglionares del parasimpático.
• Dopamina. Se localizan en: o Sustancia negra → vía nigroestriada del cuerpo estriado, sistema límbico y
numerosas áreas de la corteza) o Núcleo arcuato del hipotálamo → hipófisis anterior a través de las venas
portales
• Noradrenalina (NE). Se localizan en: o Lucus Ceruleus de la protuberancia → sistema límbico, hipotálamo, corteza o Bulbo raquídeo → locus coeruleus, médula espinal o Neuronas posganglionares del sistema nervioso simpático
• Serotonina. Se localizan en: o Núcleos del rafe protuberancial → múltiples proyecciones o Bulbo raquídeo/Protuberancia → asta posterior de la médula espinal
• Ácido γ-aminobutírico (GABA). Se localizan en: o Principal neurotransmisor inhibidor del cerebro; interneuronas corticales
muy extendidas y vías de proyecciones largas.
• Glicina. Se localizan en: o Principal neurotransmisor inhibidor de la médula espinal
• Glutamato. Se localizan en: o Principal neurotransmisor excitador; localizado por todo el SNC, incluso en
células piramidales corticales.

Sistema nervioso central [43]
El sistema nervioso central (SNC) está constituido por el encéfalo y la médula espinal. Están protegidos por tres membranas: duramadre (membrana externa), aracnoides (intermedia), piamadre (membrana interna) denominadas genéricamente meninges. Además, el encéfalo y la médula espinal están protegidos por envolturas óseas, que son el cráneo y la columna vertebral respectivamente. Se trata de un sistema muy complejo, ya que se encarga de percibir estímulos procedentes del mundo exterior así como transmitir impulsos a nervios y a músculos instintivamente.
Las cavidades de estos órganos (ventrículos en el caso del encéfalo y conducto ependimario en el caso de la médula espinal) están llenas de un líquido incoloro y transparente, que recibe el nombre de líquido cefalorraquídeo. Sus funciones son muy variadas: sirve como medio de intercambio de determinadas sustancias, como sistema de eliminación de productos residuales y para mantener el equilibrio iónico adecuado, transporta el oxígeno y la glucosa desde la sangre hasta las neuronas y también es muy importante como sistema amortiguador mecánico.
Las células que forman el sistema nervioso central se disponen de tal manera que dan lugar a dos formaciones muy características: la sustancia gris, constituida por el soma de las neuronas y sus dendritas, además de por fibras amielinicas., y la sustancia blanca, formada principalmente por las prolongaciones nerviosas (dendritas y axones), cuya función es conducir la información, además de por fibras mielínicas que son las que le confieren ese color que presentan.
En resumen, todos los animales cuyo cuerpo posee un sistema nervioso central están dotados de mecanismos nerviosos encargados de recibir y procesar las sensaciones recogidas por los diferentes sentidos y de transmitir las órdenes de respuesta de forma precisa a los distintos efectores. Se puede afirmar que el sistema nervioso central es uno de los más importantes de todos los sistemas que se encuentran en dichos cuerpos.
Debido a la gran especialización de sus células, el sistema nervioso central no se va a regenerar o tiene muy limitada esa capacidad, en comparación con el sistema nervioso periférico.
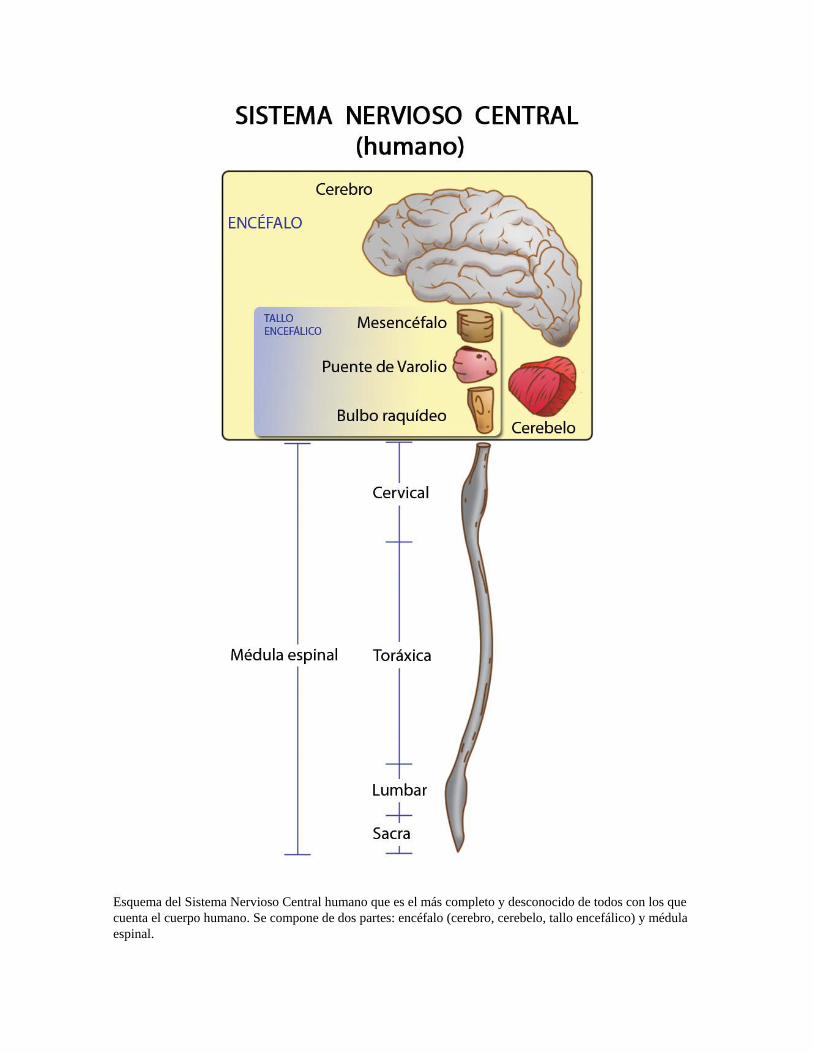
Esquema del Sistema Nervioso Central humano que es el más completo y desconocido de todos con los que cuenta el cuerpo humano. Se compone de dos partes: encéfalo (cerebro, cerebelo, tallo encefálico) y médula espinal.

Enfermedades neurodegenerativas [44]
• Esclerosis múltiple: trastorno caracterizado por episodios discretos de déficit neurológico recurrentes con desmielinización por mecanismos autoinmunes o inmunitarios. Aparece a cualquier edad, aunque es raro en la infancia o después de los 50 años, afectando a mujeres en una proporción 2.1 en relación a los hombres. La lesión se observa en el encéfalo y la médula espinal con un mayor riesgo—hasta 15 veces más elevado—si hay antecedentes familiares. Esta posible susceptibilidad genética parece favorecer una respuesta inmunitaria inadecuada frente a infecciones virales. Las lesiones se caracterizan por infiltrados de células B, células T y macrófagos. Las regiones de desmielinización por lo general se encuentran en los ángulos de los ventrículos laterales, en los pedúnculos y el tronco del encéfalo, causando debilidad de los miembros, visión borrosa o pérdida de la visión unilateral, incoordinación y sensibilidad anormal.
• Enfermedad de Alzheimer: es la más frecuente de las enfermedades neurodegenerativas y la primera causa de demencia, de aparición esporádica, aunque entre un 5-10% son de carácter familiar y la incidencia aumenta con la edad, haciéndose mayor en personas de más de 85 años de edad. Se caracteriza por una falta de memoria progresiva por degeneración de la corteza, de asociación temporal y parietal causando también trastornos afectivos. El cerebro luce más pequeño y de menor peso con atrofia de las circunvoluciones y agrandamiento de los surcos en loshemisferios cerebrales y de los ventrículos compensando la pérdida encefálica. Bajo el microscopio se aprecia angiopatía amiloide cerebral, es decir depósitos en las arterias cerebrales de material amiloide, así como placas seniles que son colecciones focales esféricas de extensiones neuríticas dilatadas y tortuosas y nódulos u ovillos neurofibrilares con forma de llama que son haces de filamentos situados en el citoplasma de las neuronas que desplazan o rodean al núcleo celular.
• Parkinsonismo: pertenecen al grupo de enfermedades neurovegetativas que afectan a los ganglios basales produciendo un trastorno del movimiento, apreciándose rigidez y lentitud en los movimientos voluntarios (bradicinesia) y temblor de reposo. La gravedad de la enfermedad depende de la pérdida de las células nerviosas de la sustancia negra mesencefálica que contienen neuromelanina. Se aprecia una hipopigmentación de la sustancia negra y del locus coeruleus con células dopaminérgicas que contienen melanina. Las células supervivientes de la sustancia negra contienen inclusiones intracitoplasmáticas llamadas cuerpos de Lewy.
• Enfermedad de Huntington: un trastorno de movimientos de tipo coreiforme y demencia en pacientes entre 20-50 años con un factor genético de herencia autosómico dominante por un gen causante localizado en el brazo corto del cromosoma 4. Se atrofia el núcleo caudado y el putamen secundario a la pérdida celular y gliosis.

REFERENCIAS [1] Neurona. (2012, julio 17). [On line] Disponible en http://es.wikipedia.org/wiki/Neurona [2] Paniagua, R.; Nistal, M.; Sesma, P.; Álvarez-Uría, M.; Fraile, B.; Anadón, R. y José Sáez, F. (2002). Citología e histología vegetal y animal. McGraw-Hill Interamericana de España, S.A.U. [3] Aminoácido.(2012, julio 23). [On line]. Disponible en http://es.wikipedia.org/wiki/Amino%C3%A1cido [4] Arginine: biochemistry, physiology, and therapeutic implications. Barbul A.; JPEN J Parenter Enteral Nutr. 1986 Mar-Apr;10(2):227-38. [5] Jeffrey N. KellerEdgardo Dimayugab, Qinghua Chena, Jeffrey Thorpea, Jillian Geeb and Qunxing Ding. Autophagy, proteasomes, lipofuscin, and oxidative stress in the aging brain The International Journal of Biochemistry & Cell Biology. Volume 36, Issue 12, December 2004, Pages 2376-2391. [6] Weisenberg RC. 1972. Microtubule formation in vitro in solutions containing low calcium concentrations. Science 177:1104–5. [7] Ibid. [8] Paniagua, R.; Nistal, M.; Sesma, P.; Álvarez-Uría, M.; Fraile, B.; Anadón, R. y José Sáez, F. (2002). Citología e histología vegetal y animal. McGraw-Hill Interamericana de España, S.A.U. [9] M Goedert, M G Spillantini, and R A Crowther. Cloning of a big tau microtubule-associated protein characteristic of the peripheral nervous system. Proc Natl Acad Sci U S A. 1992 March 1; 89(5): 1983–1987. [10] Cromer, A.H. (1996). Física para ciencias de la vida. Reverté ediciones. ISBN para España 84-291-1808-X. [11] Bear MF, Connors BW, Paradiso M.A (2002). Neurociencia: explorando el cerebro. Barcelona: Masson. [12] Llinás, Rodolfo (1988). «The Intrinsic Electrophysiological Properties of Mammalian Neurons: Insights into Central Nervous System Function».Science 242: p. 1654-1664. [13] Kardong, K. V (1999). Vertebrados. Anatomía comparada, función, evolución. McGraw-Hill Interamericana de España, S.A.U. [14] Dorothy E. Bliss, James B. Durand and John H. Welsh. Neurosecretory systems in decapod Crustacea Cell and Tissue Research. Volume 39, Number 5 / septiembre de 1954.

[15] T A Miller. [Neurosecretion and the Control of Visceral Organs in Insects http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.en.20.010175.001025] Annual Review of Entomology Vol. 20: 133-149 (Volume publication date January 1975) [16] COBB J. L. S.. Neurohumors and neurosecretion in echinoderms: a review. Comparative biochemistry and physiology. C. Comparative pharmacology. [17] Paniagua et al. [18] Ibid. [19] Sabbatini R.M.E. April-July 2003. Neurons and Synapses: The History of Its Discovery. Brain & Mind Magazine, 17. Consultado el 19 de marzo de 2007. [20] Witcher M, Kirov S, Harris K (2007). «Plasticity of perisynaptic astroglia during synaptogenesis in the mature rat hippocampus.». Glia 55 (1): pp. 13–23. [21] Connors B, Long M (2004). «Electrical synapses in the mammalian brain.». Annu Rev Neurosci 27: pp. 393–418. [22] Djurisic M, Antic S, Chen W, Zecevic D (2004). «Voltage imaging from dendrites of mitral cells: EPSP attenuation and spike trigger zones.». J Neurosci 24 (30): pp. 6703–14. [23] Cromer, A.H. (1996). Física para ciencias de la vida. Reverté ediciones. ISBN para España 84-291-1808-X. [24] Williams, R and Herrup, K (2001). "The Control of Neuron Number." Publicado en The Annual Review of Neuroscience 11:423–453 (1988). Last revised Sept 28, 2001. Consultado de http://www.nervenet.org/papers/NUMBER_REV_1988.html on May 12, 2007. [25] Chris Li, Kyuhyung Kim and Laura S. Nelson. (1999). FMRFamide-related neuropeptide gene family in Caenorhabditis elegans Brain Research, Volume 848, Issues 1-2, 27 November, Pages 26-34. [26] Paul Patrick Gordon Bateson, Peter H. Klopfer, Nicholas S. Thompson (1993): Perspectives in Ethology. Springer. [27] Bear MF, Connors BW, Paradiso M.A (2002). Neurociencia: explorando el cerebro. Barcelona: Masson. [28] Gurney, K. (1997) An Introduction to Neural Networks London: Routledge. ISBN 1-85728-673-1 (hardback) or ISBN 1-85728-503-4 (paperback). [29] Potencial de membrana. (2015, Septiembre 20), [On Line]. Disponible en http://es.wikipedia.org/wiki/Potencial_de_Nernst

[30] Sinapsis. (2015, agosto 28), [On Line]. Disponible en http://es.wikipedia.org/wiki/Sinapsis [31] Morris, R.G.M. et al., "Elements of a neurobiological theory of the hippocampus: the role of activity dependents synaptic plasticity in memory", Phil. Trans. R. Soc. Lond. B, Nº 358, 2003, pp. 773-786. [32] Kandel, E.R., Psychotherapy and the single synapse: the impact of psychiatric thought on neurobiological research, J.Neuropsychiatry Clin. Neurosci, 13: 2, 2001, pp. 290-300. [33] François Ansermet & Pierre Magistretti: A cada cual su cerebro. Plasticidad neuronal e inconsciente. Discusiones. pp. 47. [34] Blake, D.T., Byl, N.N., Mercenich, M., Representation of the hand in the cerebral cortex, Behavioral Brain Research, Nº135, 2002, pp. 179-184. [35] François Ansermet & Pierre Magistretti: A cada cual su cerebro. Plasticidad neuronal e inconsciente. Discusiones. pp. 22. [36] Purves, Augustine, Fitzpatrick, Hall, LaMantia, Williams: Neurociencia.pp-647. [37] Célula Glial. (2015, Septiembre 3), [On Line]. Disponible en http://es.wikipedia.org/wiki/Neurogl%C3%ADa [38] Cf. Dale Purves et alii (2008). Neurociencia, Panamericana, Madrid, pág. 9. [39] Ibid. [40] Sonzini Astudillo Bernardo. La Neuroglia. (2014). [On Line]. Disponible en http://www.guiasdeneuro.com.ar/la-neuroglia/ [41] Neurotransmisor. (2015, Septiembre 15), [On Line]. Disponible en http://es.wikipedia.org/wiki/Neurotransmisor [42] Luis Samper, Neuroquímica cerebral: “Las moléculas y la conducta”. Biosalud, Revista de Ciencias básicas. [43] Sistema nervioso central. (2014, Diciembre 14), [On Line]. Disponible en http://es.wikipedia.org/wiki/Sistema_nervioso_central [44] Ibid.