Embed Size (px)
Citation preview

267Laterality is universal among fishesZOOLOGICAL SCIENCE 34: 267–274 (2017) © 2017 Zoological Society of Japan
* Corresponding author. E-mail: [email protected]:10.2108/zs160196
Laterality is Universal Among Fishes but Increasingly Cryptic Among Derived Groups
Michio Hori1, Mifuyu Nakajima2, Hiroki Hata3, Masaki Yasugi4, Satoshi Takahashi5, Masanori Nakae6, Kosaku Yamaoka7, Masanori Kohda8, Jyun-ichi Kitamura9,
Masayoshi Maehata10, Hirokazu Tanaka8,11, Norihiro Okada12,13, and Yuichi Takeuchi14*
1Kyoto University, Yoshida-Honmachi, Sakyo-ku, Kyoto 606-8501, Japan, 2Department of Biology, Stanford University, Stanford, CA 94305, USA, 3Graduate School of Science and Engineering, Ehime University,
Bunkyocho 2-5, Matsuyama 790-8577, Japan, 4National Institute for Basic Biology, Higashiyama 5-1, Myodaiji, Okazaki, Aichi 444-8787, Japan, 5Research Group of Environmental Sciences, Nara Woman’s University,
Kitauoya Nishimachi, Nara 630-8506, Japan, 6Department of Zoology, National Museum of Nature and Science, Amakubo 4-1-1, Tsukuba, Ibaraki 305-0005, Japan, 7Kochi University, Monobe-B, Nankoku, Kochi 783-8502,
Japan, 8Department of Biological Science, Osaka City University, Sugimotocho, Sumiyoshi-ku, Osaka 558-8585, Japan, 9Mie Prefectural Museum, Isshinden-kouzubeta 3060, Tsu, Mie 514-0061, Japan, 10Kobe
Gakuin University, Arise 518, Ikawadanicho, Nishi-ku, Kobe 651-2180, Japan, 11Institute of Ecology and Evolu-tion, University of Bern, Hinterkappelen 3032, Switzerland, 12Department of Life Sciences, National Cheng Kung University, Tainan 701, Taiwan, 13Foundation for Advancement of International Science, Tsukuba 305-
0821, Japan, 14Department of Anatomy, Graduate School of Medicine and Pharmaceutical Sciences, University of Toyama, Sugitani, Toyama 930-0194, Japan
Laterality has been studied in several vertebrates, mainly in terms of brain lateralization and behav-ioral laterality, but morphological asymmetry has not been extensively investigated. Asymmetry in fishes was first described in scale-eating cichlids from Lake Tanganyika, in the form of bilateral dimorphism in which some individuals, when opening their mouths, twist them to the right and others to the left. This asymmetry has a genetic basis, and is correlated with lateralized attack behaviors. This has subsequently been found in fishes from numerous taxa with various feeding habits. The generality of such morphological laterality should thus be investigated in as wide a range of fishes as possible. Using specific indicators of lateral differences in mandibles and head inclination, we find that representative species from all 60 orders of extant gnathostome fishes (both bony and cartilaginous) possess morphological laterality. Furthermore, we identify the same laterality in agnathans (hagfish and lamprey), suggesting that this trait appeared early in fish evolu-tion and has been maintained across fish lineages. However, a comparison of asymmetry among groups of bony fishes reveals, unexpectedly, that phylogenetically more recent-groups possess less asymmetry in body structures. The universality of laterality in fishes indicates a monophyletic origin, and may have been present in the ancestors of vertebrates. Ecological factors, predator–prey interactions in particular, may be key drivers in the evolution and maintenance of dimorphism, and may also be responsible for the cryptic trend of asymmetry in derived groups. Because lung-fish and coelacanths share this trait, it is likely that tetrapods also inherited it. We believe that study of this morphological laterality will provide insights into the behavioral and sensory lateral-ization of vertebrates.
Key words: morphological asymmetry, dimorphism, anti-symmetry, fish evolution, frequency-dependent selection
INTRODUCTION
Behavioral and sensory lateralization is widely recog-nized in various vertebrates (Roger and Andrew, 2002). Studies of lateralization have focused primarily on humans and higher vertebrates, mainly examining cerebral lateral-
ization and related behavioral or sensory differentiation between right and left sides. However, morphological asym-metry, such as differences between the right and left sides of kinetic or sensory systems, has not been explored in depth, except for differences in the brain and nervous sys-tem. Recent studies of laterality in fishes have revealed that behavioral laterality is always associated with morphological asymmetry, which is characteristically represented as dimor-phism in populations, i.e., every population is composed of

268 M. Hori et al.
both lefty and righty individuals at different rates. The goals of this paper are (1) to demonstrate that all fishes have later-ality and, therefore, that the origins of laterality are at least as old as those of fish, and (2) to discuss the significance of the maintenance of dimorphic laterality in most populations. Specifically, we direct our attention to predator–prey interac-tions in biotic communities as the primary factor responsible for the maintenance of dimorphism of laterality in fish.
The first and most notable example of laterally asym-metric bodies in fishes was found in scale-eating cichlids in Lake Tanganyika (Liem and Stewart, 1976; Hori, 1991, 1993; Hata et al., 2013), in which laterality was associated with hunting techniques. Each population consists of righty (the right side of the head faces slightly forward, and the mouth opens to the left) and lefty (vice versa) individuals. Righty (lefty) individuals attack the right (left) sides of their prey fish from behind to tear off scales (Hori, 1993; Takeuchi et al., 2012, 2016). The behavioral laterality associated with bilat-eral asymmetry has also been found in fishes from a wide range of trophic levels and taxa, including cichlids (Hori et al., 2007; Takeuchi and Hori, 2008), gobies (Seki et al., 2000; Yasugi and Hori, 2011), bettas (Takeuchi et al., 2010), large-mouth bass (Nakajima et al., 2007; Yasugi and Hori, 2011), characins (Hata et al., 2011), poeciliids (Matsui et al., 2013), and anglerfish (Yasugi and Hori, 2016). Nearly all of the indi-viduals in these studies could be classified as either righty or lefty individuals.
Laterality appears to have a genetic basis, as indicated by studies on parents and their offspring in wild species (a goby: Seki et al., 2000; two cichlids: Hori et al., 2007) and in breeding experiments (a cichlid: Hata et al., 2012; medaka: Hata et al., 2012; and zebrafish: Hata and Hori, 2012), in which righty and lefty offspring were born in a Mendelian ratio with left dominant over right. Additionally, genetic stud-ies have identified genomic loci corresponding to mandibu-lar asymmetry (Stewart and Albertson, 2010).
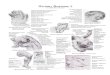
The degree of laterality in individuals has been quanti-fied based primarily on two measures: (i) differences between the right and left mandibles (Hori, 1993; Seki et al., 2000; Takeuchi and Hori, 2008; Yasugi and Hori, 2011; Takeuchi et al., 2012; Hata et al., 2013), more specifically, the height of the posterior ends of the right and left mandi-bles (HMPE) (Hori et al., 2007; Hata et al., 2013) (Fig. 1A), and (ii) the angle θ formed by the head and vertebrae (Hata et al., Yasugi and Hori, 2011; Nakajima et al., 2007) (Fig. 1B), in which the head of each individual inclines laterally, such that one side of the head faces forward (Hori et al., 2007). Laterality identified in every individual using these two meth-ods has been consistent (Yasugi and Hori, 2011; Hata et al., 2013).
In general, lateral asymmetry in species or populations can be divided into three categories based on the frequency distribution of measured laterality: fluctuating asymmetry (FA) with a unimodal and symmetrical distribution, direc-tional asymmetry (DA) with a unimodal distribution shifted from symmetry, and anti-symmetry (AS) with a bimodal distribution (Palmer and Strobeck, 1992). Recent studies examining the asymmetry of fishes have shown bimodal dis-tributions, suggesting AS (Hori et al., 2007; Nakajima et al., 2007; Takeuchi and Hori, 2008; Seki et al., 2000; Takeuchi et al., 2010, 2012, 2016; Hata et al., 2011, 2013; Yasugi and
Hori, 2011, 2012; Matsui et al., 2013). Here, to ascertain the type of asymmetry, we examined frequency distributions of two measures in two species of bony fishes—amberjack and catfish—as arbitrary representatives of dominant taxa in marine and freshwater environments, respectively, as well as in one cartilaginous fish, a stingray.
Next, to assess the prevalence of lateral asymmetry among fishes and to identify trends in the degree of laterality during evolution, we measured the laterality of individuals from at least one arbitrary representative species from all 62 extant orders of fishes (Nelson, 2006). The species exam-ined and the measurements of the 62 orders are listed in the electronic supplementary material (Supplementary Table S1). We measured mandibular asymmetry and head angle
Fig. 1. Bone measurements used to identify fish laterality. (A) The height of the mandible at the posterior end (HMPE) is the distance from A (the deepest point on the suspensoriad articulation facet of the anguloarticular) to RP (the ventral corner of the retroarticular process). (B) Ventral view of a skeleton showing data points for head angle (θ ), defined as the angle between the line extended from VIII–B (dashed red line) and P–B. B: the sagittal posterior end of the basioccipital; P: the center of the posterior portion of the parasphenoid; VIII: the sagittal posterior end of the third vertebra. Both panels show the Japanese amberjack (Seriola quinqueradiata).

269Laterality is universal among fishes
(as described above) in 10–12 individuals from one to sev-eral populations of each species, except for five rare species in which only a few individuals (n < 6) were available.
MATERIALS AND METHODS
Collection of fish specimensTo define the frequency distribution of laterality, we collected
approximately 100 individuals each of Japanese amberjack (Seri-ola quinqueradiata, Perciformes), Amur catfish (Silurus asotus, Siluriformes), and Japanese stingray (Dasyatis akajei) as arbitrary representatives of dominant marine and freshwater bony fish taxa, and a marine cartilaginous fish, respectively. Next, to determine the presence and degree of asymmetry in each fish order, we chose at least one arbitrary representative species from all 62 orders of extant fishes based on their availability. We then collected 10–12 individuals from one to several populations of each species, except for five rare species in which only a few individuals (n < 6) were available (see Supplementary Table S1 for details). Animal care and operations were performed in accordance with the Regulation on Animal Experimentation at Kyoto University.
Measurements of morphological laterality in fish specimensLaterality of gnathostome fishes was quantified by two mea-
sures: the difference in the height of the mandible at the posterior end (HMPE) of the right and left mandibles (Fig. 1A for bony fishes and electronic supplementary material, Supplementary Figure S2A for cartilaginous fish) and the angle from the vertebrae to the head (θ in Fig. 1B for bony fishes and Supplementary Figure S2B for cartilaginous fishes) following Hata et al. (2013). In cartilaginous fishes (except in chimaeras), the sustentaculum functions as the mandibular surface of the hyomandibular-mandibular joint, and the nodule on the ventral corner of the mandibular cartilage (VC) func-tions as the attachment point of the depressor mandibularis (Dean and Motta, 2004; Wilga, 2005), such that the HMPE provides a functional measure equivalent to that of bony fishes. Despite its taxonomic position, the sturgeon is grouped with cartilaginous rather than bony fishes in this paper, as its skeleton is cartilaginous and the mechanical structure of its mouthparts resembles that of cartilaginous fishes (Carroll and Wainwright, 2003).
To prepare skeletal material, samples of all specimens, except those of the coelacanth, were gently boiled, and tissues were removed carefully by hand. For agnathan fishes (hagfish and lamprey), the mouthparts (a pair of spinose cartilage and flat carti-laginous bones, respectively, which may be homologous to the mandible of gnathostome fishes; Kuratani, 2012) were measured (Supplementary Figures S5A and S6A). We also measured the angle from the vertebrae to the head (Supplementary Figures S5B and S6B). Specimens of agnathan fishes were moderately boiled to expose mouthparts for measurement and then transparentized to measure θ. On each skeletal specimen of both gnathostome and agnathan fishes, the data points were marked with a fine pen under a binocular microscope, and HMPE and θ were measured using a digital microscope (VHX-100, Keyence Co., Inc., Osaka, Japan) to within 0.005 mm and 0.05° of accuracy, respectively. Data were measured horizontally using a level scope attached to a monocular microscope of our own design. Specifically, specimens were secured with a small clip that had a universal joint connected to a base comprising a magnet, which was then placed on a two-dimensional gonio-stage. Manipulating the gonio-stage and moni-toring via the level scope kept the specimens level. We used the median of three measurements from each specimen. Each mea-surement was obtained by repositioning the specimen on the microscope stage. For the coelacanth, computerized axial tomog-raphy (CAT) scan images of two frozen specimens stored at the Tokyo Institute of Technology were supplied, and the two measures were obtained using a DICOM viewer (INTAGE Realia, K.G.T., Inc.,
Tokyo, Japan). The index of asymmetry (IAS) for the difference between right and left mandible heights was given by [2 × (R − L) / (R + L)] × 100, where R and L are the right and left HMPE, respec-tively. Fish with positive IAS values and θ were defined as righty, and those with negative values were defined as lefty. Note that the definition of laterality used here and in recent studies differs from that used in earlier papers (Hori, 1991, 1993; Seki et al., 2000), which defined individuals with the mouth opening to the right as ‘right-handed’ or ‘dextral’. The terminology used in the present study, ‘lefty’, reflects the fact that the left mandible of such ‘right-handed’ fish is larger than the right mandible (Hori et al., 2007; Takeuchi and Hori, 2008; Yasugi and Hori, 2011, 2012), and that the left eye is dominant (Takeuchi et al., 2010).
Classification and statistical examination of asymmetryThe asymmetries of morphological characteristics were cate-
gorized as FA, DA, or AS for each species using the frequency dis-tributions of the laterality indices. To define the asymmetries of the IAS and θ in each species, we fitted three models to the frequency distributions of our measurements: (1) FA model, which assumes a normal distribution with a mean of 0 and standard deviation (s.d.) equal to the s.d. of the data; (2) DA model, which assumes a normal distribution of both the mean μ (≠ 0) and s.d. of the data; and (3) AS model, which assumes two normal distributions with the same s.d. and means at ±μ (bimodal distribution). The two normal distribu-tions with means +μ and −μ correspond to the frequency distribu-tions of righty and lefty fish, respectively. The means, s.d., and ratios of righty to lefty individuals in each model were estimated by maximum likelihood. For each species, we calculated the Akaike information criterion (AIC) of the three models using the R package IASD 1.0.7. The models with the lowest AIC values were selected as the best explanations of the asymmetries in morphological charac-teristics.
Phylogenetic grouping of fishesThe classification of extant fish orders used here follows
Nelson (2006). For evolutionary analysis, we categorized orders within Osteichthyes (bony fishes) as follows: (1) sarcopterygians (lobe-finned fishes), (2) primitive actinopterygians (ray-finned fishes), and (3) lower, (4) intermediate, and (5) higher teleosts (one of three infraclasses of ray-finned fishes). These categories were based on those proposed by Nelson (2006), but are slightly modi-fied in light of recent phylogenetic hypotheses, as shown in Supple-mentary Figure S1 (Betancur-R et al., 2013). Although Nelson et al. (2016) changed their classification of extant orders from those in Nelson (2006), the fish categories mentioned above are consistent with those of the former (group memberships are shown in Fig. 3A and Supplementary Table S1). Within the teleosts, we also catego-rized fishes into an ecological group ‘deep-sea fishes’ (those living at depths below 200 m) due to their distinctive degrees of asym-metry (see Discussion). The degree of laterality of each category was calculated as the average distance for laterality intensity (DLI), which was measured from the origin to a point (x, y) representing each species, where x is the standardized absolute value of IAS and y is the standardized absolute value of θ.
Degree of lateral asymmetry during growthTo compare the degree of lateral asymmetry among fishes of
various ages, we collected 60 largemouth bass (Micropterus salmoides) individuals of various lengths from a single population in Lake Biwa, Japan, in 2004–2008. We measured the standard length, fish age by counting circuli on the opercular bone (Menon, 1950; Campana, 2001), IAS, and θ of every individual. Then we used analysis of variance (ANOVA) to assess the effects of age on IAS and θ.

270 M. Hori et al.
Fig. 2. Frequency dis-tributions of (A) the index of asymmetry (IAS) of the mandibles and (B) head angle (θ ) of the Japanese amber-jack (Seriola quinquera-diata, n = 100; upper) and Amur catfish (Silurus asotus, n = 110; lower). Lines show the proba-bility densities derived from the three models (see Methods). The model selected by the lowest Akaike informa-tion criterion (AIC) value is indicated by the thick line.
Fig. 3. Degrees of laterality among extant teleost fishes. (A) Laterality quantified by the abso-lute values of the index of asym-metry (IAS) of the mandibles and the head angle (θ ) in each repre-sentative species from all extant orders of teleost fishes. Index numbers correspond to the extant orders of fishes in Nelson (2006). Species are grouped into six cat-egories (deep-sea fishes and five phylogenetic groups) indicated by colors. Each axis was standard-ized by the mean and standard deviation (s.d.) of all original val-ues. (B) Comparison of the dis-tance of laterality intensity (DLI) between the seven species of deep-sea fishes and the other teleost fishes (range, outlier, lower and upper quartiles, and median). P-values are from the Wilcoxon rank sum test. **, P < 0.01. (C) Comparison of DLI among five phylogenetic groups (range, outlier, lower and upper quartiles, and median).

271Laterality is universal among fishes
RESULTS
Types of asymmetryOur measurements of amberjack and catfish were best
explained by the AS model, which indicates that the focal laterality in both species was AS. Histograms of mandibular asymmetry and head angle showed bimodal distributions in both sexes, with few individuals near 0 (Fig. 2). AS was the best-fitting model for these distributions among the three asymmetry types, per the AIC values. AS has been detected in the laterality of several fish orders (Takeuchi and Hori, 2008; Yasugi and Hori, 2011; Hata et al., 2011, 2013; Matsui et al., 2013) and was also found in a cartilaginous fish, the Japanese stingray, in the present study (Supplementary Fig-ure S2).
Except for the six species with small sample sizes, every species examined was composed of both righty and lefty individuals, and the absolute degree of laterality was always greater than 0 for both measures (Supplementary Table S1). The degree of laterality measured in the present study did not appear to change during growth (Supplementary Figure S3). The distributions of both measures were mostly explained by the AS model, with only three exceptions in mandibular asymmetry (Supplementary Table S1). Among the exceptions, two showed DA, which is likely due to biased proportions of morphs among the samples (one or two indi-viduals of minority morph). The third exception was a flatfish (Pleuronectiformes), which showed FA. However, our mea-surements of head angle in flatfishes demonstrated that each population was composed of both righty and lefty indi-viduals. Flatfishes have DA bodies with both eyes on the same side of the face. The upper and lower jaws on the side with the eyes are usually shorter than those on the opposite side. The IAS of the mandibles in flatfishes may be biased by the restricted form of the mouthparts (Supplementary Figure S4). In the likelihood ratio test between FA and AS, AS was significantly better than FA for both measures among all species with ≥10 samples (Supplementary Table S1). Therefore, all fishes may exhibit laterality.
Although agnathans have no mandibles, their mouthparts and head angle showed AS distributions (Supplementary Figures S5 and S6). Therefore, the phylogenetic occurrence of focal laterality may date back to the origin of fishes.
Trends in fish phylogenyThe absolute values of our two laterality measures (IAS
of the mandibles and θ ) in all orders of extant bony fishes revealed functional and evolutionary trends (Fig. 3A). First, deep-sea fishes had high levels of asymmetry except for one species (jellynose fish). The distances from the origin of the two axes in Fig. 3A to the points representing each spe-cies (DLI) were significantly greater for the seven deep-sea fishes than for the other fishes (Wilcoxon rank sum test, n = 45, P = 0.010; Fig. 3B). Because the exploitation of extreme depths evolved independently among deep-sea fishes (Nelson, 2006), extreme asymmetry seems adaptive for life in the deep sea, as discussed below. Second, the absolute values of IAS and θ in other fishes had a significant negative correlation (Spearman’s rank correlation test, n = 38, ρ = − 0.402, P = 0.012; Fig. 3A), suggesting trade-offs between the two structures in developing laterality.
‘Ancient fishes,’ such as lungfish, coelacanths, and gars, had greater DLIs than ‘modern fishes,’ such as perciforms and scorpaeniforms. We categorized bony fishes (except the deep-sea fishes) into five phylogenetic groups (see Meth-ods). The DLI values had a significant negative correlation with the phylogenetic order of the groups (Spearman’s rank correlation test, n = 38, ρ= − 0.352, P = 0.030; Fig. 3C), implying that the degree of morphological laterality was reduced progressively during the evolution of bony fishes. The phylogenetic trends in laterality among cartilaginous fishes are shown in Supplementary Figure S7; more derived species, such as rays and skates, had lower DLI values.
DISCUSSION
Our results indicate that AS laterality is universally shared among fishes, but that the degree of asymmetry has become increasingly weaker in more derived groups. How-ever, functional (behavioral) laterality seems not to be reduced even in derived groups, since many fishes have prominent behavioral laterality, including cichlids (Hori, 1993; Takeuchi and Hori, 2008), gobies (Seki et al., 2000), largemouth bass (Nakajima et al., 2007; Yasugi and Hori, 2011), and bettas (Takeuchi et al., 2010), all of which belong to derived groups. This cryptic trend in morphological later-ality should be considered in depth. Here, we discuss 1) why morphological and behavioral laterality has developed in fish and why fish have AS rather than DA laterality; 2) the relationship between the two measures we used to evaluate morphological laterality, its implications for body form, and how this relates to the progressive weakening of morpho-logical laterality in fish evolution; 3) the origin and legacy of fish laterality; and 4) the relationship between laterality in fishes and that in aquatic invertebrates.
Behavioral advantage has been proposed as a driving force behind lateral asymmetry among fishes. The general fish body plan facilitates a bilateral motor pattern. The body is propelled by side-to-side undulations. When a fish makes a dashing movement, it first bends its body either to the right or left and subsequently straightens it (Wilga, 2005). The locomotive speed of this initial motion is crucial in predator–prey interactions (Cantalupo et al., 1995; Takeuchi et al., 2012; Yasugi and Hori, 2012). If one side of the body is mechanically dominant, i.e., has more muscle and effective support tissue than the opposite side, the initial use of this side can produce stronger propulsion and higher speed than that generated by non-specialists with completely symmetri-cal bodies (Takeuchi et al., 2012). Therefore, morphological asymmetry and laterally disproportionate usage (behavioral laterality) may be associated and mutually reinforcing.
Although asymmetrical bodies seem advantageous, why are fish dimorphic rather than monomorphic? Behav-ioral laterality inevitably leads to biased directions of attack (Nakajima et al., 2007; Yasugi and Hori, 2012) and evasion (Izvekov et al., 2009; Yasugi and Hori, 2012). These biases may provoke negative, frequency-dependent selection. For instance, of the two lateral morphs among scale-eaters, the numerically dominant morph was more likely to fail at feed-ing than the less common morph, probably because prey fishes are more vigilant on the side more frequently attacked by the major morph (Hori, 1993; Takahashi and Hori, 1994, 1998). Among pursuit-type piscivores, predation success

272 M. Hori et al.
was higher if the laterality of the predator and its prey were opposite (so-called ‘cross predation’) than if they were the same (‘parallel predation’) (Yasugi and Hori, 2011, 2012). Conversely, when luring-type piscivores face prey fish dur-ing a predation event, parallel predation was more success-ful (Yasugi and Hori, 2016). This difference in success also seems to cause an alternating advantage between righties and lefties in both predator and prey fishes and results in the dynamic maintenance of laterality (Nakajima et al., 2004). This advantage is shared by predators that attack the same prey fishes and by prey fishes that share a common predator (Takahashi and Hori, 2005). Natural selection through pred-ator-prey interactions may play a major role in maintaining anti-symmetric dimorphism in fish communities (Stearns and Hoekstra, 2000; Hedrick, 2005; Sinervo and Calsbeek, 2006). We suggest that this is the very reason why morpho-logical laterality has progressively weakened during fish evolution, which we discuss below.
The two measures of fish laterality used here represent the most distinctly asymmetric features. The morphological implications of head angle are intuitive. If the body is asym-metric due to differential development of the right and left sides, the more developed side may be structurally domi-nant over the opposite side. The more developed side should, therefore, be convex, as suggested in Fig. 1. Zebrafish were found to have a trunk muscle volume that was laterally biased in relation to the laterality of the fish (Heuts, 1999), although there was a significant bias towards escaping in one direction at the population level. The mor-phological implication of the IAS of the HMPE can be also understood by applying the principle of leverage; i.e., the HMPE acts as a line between the effort point (i.e., where the ligament is attached) and the fulcrum point of the lever (i.e., the articulation of the mandible) (Westneat, 1990). Thus, the difference between the right and left HMPE may cause the force and speed of opening to differ between the right and left mandibles, causing the mouth to twist as it opens. Because of this morphological asymmetry, the fish can open its mouth toward one side to a greater extent than through head inclination alone. In any lateralized behavior, these fac-tors may work together. For example, in the shrimp-eating cichlid (Neolamprologus fasciatus; Takeuchi and Hori, 2008) and the scale-eating cichlid (Perissodus microlepis; Takeuchi et al., 2016), each morph exhibits an advantage (greater hunting success) when the fish uses the dominant side of its body during foraging. The direction to which the body was bent, the eye used to aim at the target, and the dominant side of the mandible were all consistent.
The negative relationship between the two measures found among extant bony fishes (Fig. 3A) suggests a trade-off between asymmetry in the mandibles and head angle. Although this relationship may reflect the constraints imposed by morphogenesis, it may be ultimately affected by differences in lifestyles and modes of locomotion among fish groups. Fishes with a large head angle, such as silvereye, John Dory, and puffer fish (#42, 54, and 60, respectively, in Fig. 3A), are deep-bodied, whereas those with high IAS val-ues, such as herring, salmon, and pickerel (#27, 35, and 36, respectively, in Fig. 3A), have slender bodies (defined as having a high body length to diameter ratio). Perciforms are highly variable in body shape. Among them, slender fishes,
such as amberjack, are adapted to prolonged high-speed swimming and a pelagic lifestyle, whereas deep-bodied fishes, such as sea bream, are adapted to quick turns and a demersal lifestyle (Azuma, 2006). This relationship between body shape and swimming style may apply to all bony fishes except those with extreme body shapes, such as deep-sea fishes and eels. Thus, we propose that deep-bodied fishes are suited to tilting the head to either side during asymmetric behaviors, whereas slender fishes are suited to having asymmetric mandibles due to their streamlined bodies and smaller head angles adapted to rapid and prolonged cruis-ing. A more symmetrical bodyline is adaptive for high-speed swimming (Gosline, 1971). One of the fastest-moving fishes, the flying fish Prognichthys brevipinnis, has the smallest head angle (Supplementary Table S1).
The universality of morphological laterality in fishes sug-gests a monophyletic origin. We hypothesize that the evolu-tion of fish laterality began with lancelets or agnathans, and that laterality has been maintained over the course of fish evolution. Ecological factors, particularly predator–prey interactions, are likely key drivers in the evolution of lateral dimorphism in every fish population, which should be main-tained by frequency-dependent selection mediated through minority advantage (Hori, 1993; Stearns and Hoekstra, 2000; Hedrick, 2005; Sinervo and Calsbeek, 2006). Although laterality is inevitable for effective locomotion in every fish species, they must also conceal their own laterality from appearance, as such laterality would indicate a preferred attacking direction of predators or escape direction of prey. Laterality may therefore have become increasingly cryptic during fish evolution. In this context, the extreme asymmetry in deep-sea fishes (Fig. 3A) may be explained as follows; since they live in the dark and rely only slightly on vision, they have little need to conceal their morphological laterality.
In addition to the origin of laterality, the remarkable later-ality of sarcopterygians (coelacanths and lungfish; #61 and 62, respectively, in Fig. 3A), which share a common ances-tor with terrestrial tetrapods (amphibians, reptiles, birds, and mammals), provides another implication for the universality of laterality, particularly with respect to the traits inherited by tetrapods from fish. Lateralized behaviors have been evalu-ated in animals ranging from fish to mammals (Roger and Andrew, 2002) with emphasis on brain lateralization, which is thought to facilitate enhanced performance or faster responses (Vallortigara and Rogers, 2005; Dadda et al., 2010). By contrast, the laterality of behaviors in fishes, including hunting techniques (Hori, 1993; Nakajima et al., 2007; Takeuchi and Hori, 2008; Hata et al., 2011; Yasugi and Hori, 2012; Takeuchi et al., 2012, 2016), scouting behaviors (Matsui et al., 2013), and social behaviors (Takahashi and Hori, 2008; Takeuchi et al., 2010), are correlated with the morphological laterality that we have demonstrated here. This documentation of morphological laterality in fish may lead to a deeper understanding of laterality in other verte-brates.
Morphological AS has also been known in aquatic inver-tebrate groups, including Arthropoda and Mollusca (Palmer, 2005). Our recent studies on shrimp (Takeuchi et al., 2008), crayfish (Tobo et al., 2012), and cuttlefish (Lucky et al., 2012) demonstrated that their morphological AS is associated with behavioral laterality, which is involved with escaping from

273Laterality is universal among fishes
predators or attacking prey. Like fish, these animals move actively in aquatic environments. They suffer minimal effects from gravity due to buoyancy, which may allow them to develop bilateral body asymmetry. By contrast, in terrestrial environments, the effect of gravity may force animals to have a laterally symmetric body supported by limbs in a bal-anced arrangement. Morphological laterality in terrestrial animals may thus have been weakened, although functional (behavioral) laterality seems to remain, as many studies indicate (e.g., Rogers and Andrew, 2002). Although it is dif-ficult to speculate on the phylogenetic relationship of lateral-ity between aquatic invertebrates and fishes, it is possible that all members of Bilateria share laterality as a homology, given the commonness of morphological laterality among invertebrate phyla (Palmer, 2005). In addition, Arthropoda and Mollusca are phylogenetically older than fishes (Moody and Zhuravlev, 2001), and behavioral laterality seems to have already existed among Cambrian trilobites and/or their predators (Babcock, 1993); fish did not appear until the end of this period. Therefore, it is possible that laterality devel-oped first in Arthropoda, and then probably in Cephalopoda (e.g., Nautiloidea in the Ordovician: Babcock, 1993). The Cambrian also saw the explosive development of predator–prey interactions, which were driven by newly acquired eye-sight in most animal phyla (Gould, 1989). We speculate that the development of the anti-symmetric laterality in Bilateria may be closely linked to such interspecific interactions where appearance is crucial to detect prey and/or escape from predators. Although it is unclear whether the laterality of fish is homologous with that of aquatic invertebrates, the anti-symmetric laterality of fish may have been developed through predator-prey interactions in ancient aquatic com-munities. Understanding the laterality of fish may also shed new light on the community structures of aquatic ecosys-tems.
ACKNOWLEDGMENTS
We thank all the institutes, museums, aquariums, researchers, and fishermen who provided the fish materials listed in Supplemen-tary Table S1, and Drs. M. Aibara, R. Takahashi, and N. Ogihara for the CAT scan image data analysis of the coelacanth. This work was funded by grants from the Ministry of Education, Culture, Sports, Science and Technology, Japan (21st Century COE Program (A14), Global COE Program (A06), Priority Areas (14087203), and KAKENHI (21370010, 15H05230, 16H05773, 17K14934)).
COMPETING INTERESTS
The authors have no competing interests to declare.
AUTHOR CONTRIBUTIONS
MH planned and headed the project, and wrote the manuscript. MNakajima collected the materials and refined the manuscript. MH, HH, MY, MNakae, KY, MK, JK, MM, HT, and YT collected materials and carried out the measurements, ST and HT cover the statistical examination, and NO obtained coelacanth specimens and their CT image data. All authors discussed the results and commented on the manuscript.
SUPPLEMENTARY MATERIALS
Supplementary materials for this article are available online (URL: http://www.bioone.org/doi/suppl/10.2108/zs160196).
Supplementary Table S1. List of species analyzed and
model-fitting results.Supplementary Figure S1. Classification and phylogenetic
tree of the extant orders of teleost fishes used in this study.Supplementary Figure S2. Laterality of the Japanese sting-
ray (Dasyatis akajei).Supplementary Figure S3. Relationship between growth and
laterality in largemouth bass (Micropterus salmoides).Supplementary Figure S4. Laterality of the flatfishes.Supplementary Figure S5. Laterality of the hagfish.Supplementary Figure S6. Laterality of the lamprey.Supplementary Figure S7. Degrees of laterality in extant car-
tilaginous fish orders and sturgeons.
REFERENCES
Azuma A (2006) The biokinetics of flying and swimming 2nd ed, American Institute of Aeronautics and Astronautics, Virginia
Babcock LE (1993) Trilobite malformations and the fossil record of behavioral asymmetry. J Paleontol 67: 217–229
Betancur-RR, Broughton RE, Wiley EO, Carpenter K, López JA, Li C, et al. (2013) The Tree of Life and a New Classification of Bony Fishes. PLOS Currents Tree of Life
Campana S (2001) Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59: 197–242
Cantalupo C, Bisazza A, Vallortigara G (1995) Lateralization of predator-evasion response in a teleost fish (Girardinus falcatus). Neuropsychologia 33: 1637–1646
Carroll AM, Wainwright PC (2003) Functional morphology of prey capture in the sturgeon, Scaphirhynchus albus. J Morphol 256: 270–284
Dadda M, Koolhaas WH, Domenici P (2010) Behavioural asymme-try affects escape performance in a teleost fish. Biol Lett 6: 414–417
Dean N, Motta PJ (2004) Anatomy and functional morphology of the feeding apparatus of the lesser electric ray, Narcine brasiliensis(Elasmobranchii: Batoidea). J Morphol 262: 462–483
Gosline WA (1971) Functional Morphology and Classification of Teleostean Fishes. The university press of Hawaii, Hawaii
Gould SJ (1989) Wonderful life; the Burgess shale and the nature of history. WW Norton, New York
Hata H, Hori M (2012) Inheritance patterns of morphological lateral-ity in mouth opening of zebrafish, Danio rerio. Laterality 17: 741–754
Hata H, Yasugi M, Hori M (2011) Jaw laterality and related handed-ness in the hunting behavior of a scale-eating characin, Exodon paradoxus. PLoS ONE 6: e29349
Hata H. Takahashi R, Ashiwa H, Awata S, Takeyama T, Kohda M, Hori M (2012) Inheritance patterns of lateral dimorphism exam-ined by breeding experiments with the Tanganyikan cichlid (Julidochromis transcriptus) and the Japanese medaka (Oryzias latipes). Zool Sci 29: 49–53
Hata H, Yasugi M, Takeuchi Y, Hori M (2013) Distinct lateral dimor-phism in the jaw morphology of the scale-eating cichlids, Perrisodus microlepis and P. straeleni. Ecol Evol 3: 4641–4647
Hedrick PW (2005) Genetics of populations 3rd ed, Jones and Bartlett Publishers, Boston
Heuts B (1999) Lateralization of trunk muscle volume, and lateral-ization of swimming turns of fish responding to external stimuli. Behav Process 47: 113–124
Hori M (1991) Feeding relationships among cichlid fishes in Lake Tanganyika: effects of intra- and interspecific variation of feed-ing behavior on their coexistence. Ecol Int Bull 19: 89–101
Hori M (1993) Frequency-dependent natural selection in the hand-edness of scale-eating cichlid fish. Science 260: 216–219
Hori M, Ochi H, Kohda M (2007) Inheritance pattern of lateral dimor-phism in two cichlids (a scale eater, Perissodus microlepis, and

274 M. Hori et al.
an herbivore, Neolamprologus moorii) in Lake Tanganyika. Zool Sci 24: 489–492
Inoue JG, Miya M (2001) Phylogeny of the basal teleosts, with spe-cial reference to the Elopomorpha. Japan J Ichthyol 48: 75–91
Izvekov EI, Chebotareva Yu V, Izyumov Yu G, Nepomnyashechikh VA, Medyantseva EN (2009) Behavioral and morphological asymmetries in roach Rutilus rutilus (Cyprinidae: Cypriniformes) underyearlings. J Ichthyol 49: 88–95
Kuratani S (2012) Evolution of the vertebrate jaw: comparative embryology and molecular developmental biology reveal the factors behind evolutionary novelty. J Anat 205: 335–347
Liem KF, Stewart DJ (1976) Evolution of the scale-eating cichlid fishes of Lake Tanganyika: a generic revision with a description of a new species. Bull Mus Compar Zool 147: 319–350
Lucky NS, Ihara R, Yamaoka K, Hori M (2012) Behavioral laterality and morphological asymmetry in the cuttlehish, Sepia lycidas. Zool Sci 29: 286–292
Matsui S, Takeuchi Y, Hori M (2013) Relation between morphologi-cal antisymmetry and behavioral laterality in a Poeciliid fish. Zool Sci 30: 613–618
Menon MO (1950) The use of bones other than otoliths in determin-ing the age and growth rate of fishes. J Cons Int Explor Mer 16: 311–335
Moody RTJ, Zhuravlev AY (2001) Atlas of the evolving earth. Andromeda Oxford Ltd, Oxfordshire
Nakajima M, Matsuda H, Hori M (2004) Persistence and fluctuation of lateral dimorphism in fishes. Am Nat 163: 692–698
Nakajima M, Yodo T, Katano O (2007) Righty fish are hooked on the right side of their mouths – observations from an angling exper-iment with largemouth bass, Michroperus salmoides. Zool Sci 24: 855–859
Near TJ, Dornburg A, Eytan RI, Keck BP, Smith WL, Kuhn KL, et al. (2013) Phylogeny and tempo of diversification in the superra-diation of spiny-rayed fishes. PNAS Early Edition: 1–6
Nelson JS (2006) Fishes of the World 4th ed, Wiley, New JerseyNelson JS, Grande TC, Wilson MVH (2016) Fishes of the World 5th
ed, Wiley, New JerseyPalmer AR, Strobeck C (1992) Fluctuating asymmetry as a mea-
sure of development stability: implications of non-normal distri-butions and power of statistical tests. Acta Zool Fennica 191: 57–72
Roger LJ, Andrew RJ (2002) Comparative Vertebrate Lateralization. Cambridge University Press, London
Seki S, Kohda M, Hori M (2000) Asymmetry of mouth morph of a freshwater goby, Rhinogobius flumineus. Zool Sci 17: 1321–1325
Sinervo, B, Calsbeek R (2006) The developmental, physiological, neural, and genetical causes and consequences of frequency-dependent selection in the wild. Annu Rev Ecol Evol Syst 37: 581–610
Stearns SC, Hoekstra RF (2000) Evolution: An Introduction. Oxford University Press, Oxford
Stewart TA, Albertson RC (2010) Evolution of a unique predatory
feeding apparatus: functional anatomy, development and a genetic locus for jaw laterality in Lake Tanganyika scale-eating cichlids. BMC Biology 8: 8
Takahashi S, Hori M (1994) Unstable evolutionarily stable strategy and oscillation: a model on lateral asymmetry in scale-eating cichlids. Am Nat 144: 1001–1020
Takahashi S, Hori M (1998) Oscillation maintains polymorphisms - a model of lateral asymmetry in two competing scale-eating cichlids. J Theor Biol 195: 1–12
Takahashi S, Hori M (2005) Coexistence of competing species by the oscillation polymorphism. J Theor Biol 235: 591–596
Takahashi T, Hori M (2008) Evidence of disassortative mating in a Tanganyikan cichlid fish and its role in the maintenance of intra-population dimorphism. Biol Lett 4: 497–499
Takeuchi Y, Hori M (2008) Behavioural laterality in the shrimp-eating cichlid fish, Neolamprologus fasciatus, in Lake Tanganyika. Anim Behav 75: 1359–1366
Takeuchi Y, Tobo S, Hori M (2008) Morphological asymmetry of the abdomen and behavioral laterality in atyid shrimps. Zool Sci 25: 355–363
Takeuchi Y, Hori M, Myint O, Kohda M (2010) Lateral bias of agonis-tic responses to mirror images and morphological asymmetry in the Siamese fighting fish (Betta splendens). Behav Brain Res 208: 106–111
Takeuchi Y, Hori M, Oda Y (2012) Lateralized kinematics of preda-tion behavior in a Lake Tanganyika scale-eating cichlid fish. PLoS ONE 7: e29272
Takeuchi Y, Hori M, Tada S, Oda Y (2016) Acquisition of lateralized predation behavior associated with development of mouth asymmetry in a Lake Tanganyika scale-eating cichlid fish. PLoS ONE 11: e0147476
Tobo S, Takeuchi Y, Hori M (2012) Morphological asymmetry and behavioral laterality in the crayfish, Procambarus clarkii. Ecol Res 27: 53–59
Vallortigara G, Rogers LJ (2005) Survival with an asymmetrical brain: advantages and disadvantages of cerebral lateralization. Behav Brain Sci 28: 575–633
Westneat M (1990) Feeding mechanics of teleost fishes (Labridae, Perciformes) - a test of 4-bar linkage models. J Morphol 205: 269–295
Wilga CD (2005) Morphology and evolution of the jaw suspension in Lamniform sharks. J Morphol 265: 102–119
Yasugi M, Hori M (2011) Predominance of cross-predation between lateral morphs in a largemouth bass and a freshwater goby. Zool Sci 28: 869–874
Yasugi M, Hori M (2012) Lateralized behavior in the attacks of large-mouth bass on Rhinogobius gobies corresponding to their morphological antisymmetry. J Exper Biol 215: 2390–2398
Yasugi M, Hori M (2016) Predominance of parallel- and cross-pre-dation in anglerfish. Mar Ecol 37: 576–587
(Received November 16, 2016 / Accepted April 6, 2017)

Corrigendum: Laterality is universal among fishes but increasingly cryptic among derived groups
Michio Hori, Mifuyu Nakajima, Hiroki Hata, Masaki Yasugi, Satoshi Takahashi, Masanori Nakae, Kosaku Yamaoka, Masanori Kohda, Jyun-ichi Kitamura, Masayoshi Maehata, Hirokazu Tanaka, Norihiro Okada, and Yuichi Takeuchi
There was an error in Figure 2 in the published article. The model selected by the lowest Akaike information criterion value is indicated by the thick line (AS model).
Fig. 2. Frequency distributions of (A) the index of asymmetry (IAS) of the mandibles and (B) head angle (θ) of the Japanese amberjack (Seriola quinqueradiata, n = 100; upper) and Amur catfish (Silurus asotus, n = 110; lower). Lines show the probability densities derived from the three models (see Methods). The model selected by the lowest Akaike information criterion (AIC) value is indicated by the thick line.

Supplementary information
Laterality is universal among fishes but increasingly cryptic among derived groups
Supplementary Table 1. List of the species analyzed and the model-fitting results. Fish species analyzed as represen-tatives of each extant fish orders are listed with both Latin and common names, sample sizes, Akaike information crite-rion (AIC) values of the model fitted to the two measure-ments (bold: AIC of the selected model), sampling locations, and sources. P-values in LRT are from likelihood ratio test between FA model and AS model. Common names follow those used by FishBase (http://www.fishbase.org/). The index numbers of fish orders follow the classification used in Nelson (2006). Phylogenetic groupings for teleosts in this study, however, are slightly modified from Nelson’s system based on Betancur-R et al. (2013).Supplementary Figure 1. Classification and phylogenetic tree of the extant orders of teleost fishes used in this study. Phylogenetic trees are modified following Betancur-R et al. (2013). The classification is based on Nelson (2006), with a slight modification following Betancur-R et al. (2013) as shown in Supplementary Table 1 (colors are the same as those in Fig. 3A). The blue plot shows deep-sea fishes.Supplementary Figure 2. Laterality of the Japanese stingray (Dasyatis akajei). (A, B) Cartilaginous bones showing the data points used for measurements. (A) Ventral view of the jaw arch and a close-up of the left mandibular cartilage showing the data points for the index of asymmetry (IAS) of the mandibles. The arrow indicates the height of the mandible at the posterior end (HMPE), which is the distance from the postero-dorsal edge of the sustentaculum in the mandibular cartilage (PE) to the ventral corner of the man-dibular cartilage (VC). In cartilaginous fishes (except in chi-maeras), the sustentaculum functions as the mandibular surface of the hyomandibular-mandibular joint, and the nod-ule on the VC functions as the attachment point of the depressor mandibularis (Dean and Motta, 2004; Wilga 2005), such that the HMPE provides a functional measure equivalent to that of teleost fishes. (B) Ventral view of head and vertebrae showing data points for the head angle (θ ), which is the angle between the head and vertebrae on the ventral side of the anterior part of the skeleton. CC: the small foramen near the center of the cranial cartilage; PC: the mid-line of the postero-ventral margin of the cranium; PV: the first large process on the ventral sagittal line of the vertebra. (C, D) Frequency distributions of the (C) IAS of the mandibles and (D) θ in the Japanese stingray. Lines show the probabil-ity densities derived from the three models (see Methods). The model selected by the lowest Akaike information crite-rion (AIC) value is indicated by the thick line.Supplementary Figure 3. Relationship between growth and laterality in largemouth bass (Micropterus salmoi-des). The absolute values of the index of asymmetry (IAS) of mandibles and the head angles (θ ) in three size classes did not differ significantly (ANOVA, P = 0.2379 for IAS and P = 0.7791 for θ ). Individuals were sampled from the same pop-ulation in Lake Biwa, Japan, in 2004–2008. Vertical bars indicate standard deviations (s.d.). Small fish were primarily 0 + (n = 20, standard length [SL; mean ± s.d.] = 76.1 ± 4.1
mm), medium fish were primarily 1+ (n = 20, SL = 125.1 ± 13.6 mm), and large fish were primarily ≥ 2 + (n = 20, SL = 173.4 ± 16.8 mm).Supplementary Figure 4. Laterality of the flatfishes. Fre-quency distributions of the (A) index of symmetry (IAS) of the mandibles and (B) head angles in flathead flounder (Hip-poglossoides dubius; upper) and olive flounder (Paralich-thys olivaceus; lower). The lines are the same as those used in Figure 2. In both species, the model with the best fit to the distributions of IAS was fluctuating asymmetry, whereas the best model for head angle was anti-symmetry (see Supple-mentary Table 1).Supplementary Figure 5. Laterality of the hagfish. (A, B) Data for measurements from inshore hagfish (Eptatretus burgeri). (A) Ventral view of the mouthparts of a moderately boiled fish sample. A pair of flat cartilaginous structures behind the tooth plates is exposed. FC: the length of the flat cartilaginous structure used for the index of asymmetry (IAS). (B) Dorsal view of the anterior part of the body in a transparent preparation showing data for the head angle. CN: the center of the cartilaginous rings of the nasal tube; MC: the posterior end of the membranous cranium; DP: the dorsal datum point equidistant from MC to CN–MC on the sagittal line. Head angle (θ ) is defined as the angle of the line extending anteriorly from MC–DP to CN–MC. A fish with a clockwise positive angle is defined as left type, whereas a fish with a negative angle is defined as right type. The defini-tions of right and left types are consistent with those shown in the ventral view of Figure 1. (C, D) Frequency distributions of (C) IAS of the flat cartilaginous structure and (D) θ in inshore hagfish. The lines are the same as those used in Figure 2. Both distributions showed anti-symmetry (see Supplementary Table 1).Supplementary Figure 6. Laterality of the lamprey. (A, B) Data points used for measurements in arctic lamprey (Lampetra japonica). (A) Ventral view of a mouthpart in which a pair of spinose cartilaginous structures behind the suction cup are exposed in a moderately boiled fish sample. SC: the length of the spinose cartilage used for the index of asymmetry (IAS). (B) Dorsal view of the anterior part of the body in a transparent specimen showing the data points used for head angle (θ ) on the ventral sagittal line. PD: the anterior end of the posterior dorsal cartilage; PO: the poste-rior end of the postero-occipital cartilaginous ring; DP: the dorsal datum point equidistant to PD–PO. θ is defined as the angle from the line extending anteriorly from PO–DP to PD–PO. A fish with a clockwise positive angle is defined as left type, whereas a fish with a negative angle is defined as right type. (C, D) Frequency distributions of the (C) IAS of spinose cartilaginous structures and (D) θ in arctic lamprey. The lines are the same as those in Figure 2. Both distributions showed anti-symmetry (see Supplementary Table 1).Supplementary Figure 7. The degree of laterality in extant cartilaginous fish orders and sturgeons. Two measures of the laterality, the index of asymmetry (IAS) of the mandibles and the head angle, are displayed as in Fig-ure 3A. Fish species are grouped into four color-coded cat-egories.
Supplementary References
1. Betancur-RR, Broughton RE, Wiley EO, Carpenter K, Lopez JA,

Li C, Holcroft NI, Arcila D, Sanciangco M, Cureton J, Zhang F, Buser T, Campbell M, Rowley T, Ballesteros JA, Lu G, Grande T, Arratia G, Ortí G (2013) The tree of life and a new classifica-tion of bony fishes. PLoS Currents Tree of Life.
2. Nelson JS (2006) Fishes of the World 4th ed. Wiley, New Jersey3. Dean MN, Motta PJ (2004) Anatomy and functional morphology
of the feeding apparatus of the lesser electric ray, Narcine brasiliensis (Elasmobranchii: Batoidea). J Morphol 262: 462–483
4. Wilga C (2005) Morphology and evolution of the jaw suspension in Lamniform sharks. J Morphol 265: 102–119

Su
pp
lem
enta
ry T
able
S1.
Li
st o
f spe
cies
ana
lyze
d an
d m
odel
-fitt
ing
resu
lts. F
ish
spec
ies
anal
yzed
as
repr
esen
tativ
es o
f eac
h ex
tant
fish
ord
ers
are
liste
d w
ith b
oth
Lat
in a
nd c
omm
on
nam
es, s
ampl
e si
zes,
Aka
ike
info
rmat
ion
crite
rion
(A
IC)
valu
es o
f the
mod
el fi
tted
to th
e tw
o m
easu
rem
ents
(bo
ld: A
IC o
f the
sel
ecte
d m
odel
), s
ampl
ing
loca
tions
, and
sou
rces
. P-v
alue
s in
LR
T a
re fr
om li
kelih
ood
ratio
test
bet
wee
n FA
mod
el a
nd A
S m
odel
. Com
mon
nam
es fo
llow
thos
e us
ed b
y F
ishB
ase
(htt
p://w
ww
.fish
base
.org
/).T
he in
dex
num
bers
of fi
sh o
rder
s fo
llow
th
e cl
assi
ficat
ion
used
in N
elso
n (2
00
6). P
hylo
gene
tic g
roup
ings
for
tele
osts
in th
is s
tudy
, how
ever
, are
slig
htly
mod
ified
from
Nel
son’
s sy
stem
bas
ed o
n B
etan
cur-
R e
t al.
(201
3).
No.
of
Gro
up in
Sam
ple
|IAS|
|an
gle|
DLI
AIC
in IA
S of
man
dibl
esA
IC in
ang
le b
etw
een
the
head
and
ver
tebr
aeLo
calit
y of
mat
eria
ls
So
urce
of m
ater
ials
orde
r #Fi
g. 3
Ord
er N
ame
Spec
ies N
ame
‡En
glis
h N
ame
size
*m
ean
S.D
.m
ean
S.D
.FA
mod
elD
Am
odel
ASm
odel
LRT
pV o
fA
S to
FA
FAm
odel
DA
mod
elA
Smod
elLR
T pV
of
AS
to F
A
1-
Myx
inifo
rmes
Epta
tret
us b
urge
riIn
shor
e ha
gfis
h11
(5)
3.65
1.09
1.82
0.76
-62
.63
64.2
152.36
< 0.
001
48.1
850
.17
44.42
0.01
6Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
2-
Petro
myz
ontif
orm
esLa
mpe
tra
japo
nica
Arc
tic la
mpr
ey11
(5)
4.11
1.94
4.06
1.28
-66
.52
67.7
664.97
0.06
065
.08
67.0
755.91
< 0.
001
Nor
ther
n Ja
pan
Purc
hase
d fr
om fi
sher
man
3-
Chi
mae
rifor
mes
Chi
mae
ra p
hant
asm
aSi
lver
chi
mae
ra10
(7)
4.82
1.33
2.63
0.85
-62
.58
63.8
551.93
< 0.
001
50.6
852
.21
41.94
0.00
1Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
4-
Het
erod
ontif
orm
esH
eter
odon
tus j
apon
icus
bullh
ead
shar
k10
(4)
3.56
1.51
2.10
0.98
-57
.43
59.3
654.55
0.02
747
.20
48.2
745.79
0.06
5Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
5-
Ore
ctol
obifo
rmes
Chi
losc
ylliu
m p
unct
atum
Bro
wnb
ande
d ba
mbo
osha
rk10
(6)
2.42
0.74
1.77
0.83
-53
.66
55.5
043.77
< 0.
001
47.9
549
.75
46.22
0.05
3Ea
st C
hina
Sea
Cou
rtesy
of D
r. K
. Sat
o (O
kina
wa
Chu
raum
i Aqu
ariu
m)
6-
Lam
nifo
rmes
Lam
na d
itrop
isSa
lmon
shar
k11
(1)
4.08
2.03
2.74
0.86
-66
.58
62.02
65.8
90.
101
56.4
250
.43
47.04
< 0.
001
The
Paci
fic o
f eas
tern
Purc
hase
d fr
om fi
sher
man
7-
Car
char
hini
form
esM
uste
lus g
rise
usSp
otle
ss sm
ooth
-hou
nd11
(3)
2.86
0.86
1.69
0.57
-57
.30
55.3
947.08
< 0.
001
45.9
845
.39
38.13
0.00
2Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
8-
Hex
anch
iform
esH
eptr
anch
ias p
erlo
Shar
pnos
e se
veng
ill sh
ark#
5 (5
)5.
241.
312.
420.
35-
--
--
--
--
East
Chi
na S
eaPr
eser
ved
in H
okka
ido
Uni
vers
ity M
useu
m9
-Ec
hino
rhin
iform
esEc
hino
rhin
us b
rucu
sB
ram
ble
shar
k#1
(1)
3.46
-1.
80-
--
--
--
--
-So
uth
Afr
ica
Pres
erve
d in
Hok
kaid
o U
nive
rsity
Mus
eum
10-
Squa
lifor
mes
Squa
lus a
cant
hias
Pike
d do
gfis
h10
(3)
3.61
1.30
1.21
0.53
-57
.25
57.3
651.54
0.00
535
.94
37.2
933.67
0.03
9Th
e Pa
cific
of e
aste
rnPu
rcha
sed
from
fish
erm
an11
-Sq
uatin
iform
esSq
uatin
a ja
poni
caJa
pane
se a
ngel
shar
k10
(5)
3.43
1.44
2.63
1.10
-56
.64
58.2
753.52
0.02
451
.34
53.3
448.03
0.02
1Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
12-
Pris
tioph
orifo
rmes
Pris
tioph
orus
japo
nicu
sJa
pane
se sa
wsh
ark#
5 (1
)4.
621.
433.
190.
85-
--
--
--
--
East
Chi
na S
eaPr
eser
ved
in H
okka
ido
Uni
vers
ity M
useu
m13
-To
rped
inifo
rmes
Nar
ke ja
poni
caJa
pane
se sl
eepe
r ray
11 (4
)3.
491.
371.
380.
66-
62.2
762
.91
57.33
0.00
842
.54
42.4
941.30
0.07
2Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
14-
Pris
tifor
mes
Pris
tis m
icro
don
Larg
etoo
th sa
wfis
h#1
(0)
3.69
-1.
76-
--
--
--
--
-Th
e co
ast o
f Aus
tralia
Pres
erve
d in
KA
IYU
KA
N A
quar
ium
15-
Raj
iform
esO
kam
ejei
ken
ojei
Oce
llate
spot
skat
e10
(4)
3.36
1.33
1.13
0.47
-56
.08
58.0
851.90
0.01
334
.43
35.0
831.31
0.02
4Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
16-
Myl
ioba
tifor
mes
Das
yatis
aka
jei
Japa
nese
stin
gray
80 (3
9)3.
862.
000.
950.
51-
464.
0546
4.45
449.51
< 0.
001
241.
2724
2.07
232.96
0.00
1Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
172
Poly
pter
iform
esPo
lypt
erus
end
liche
riSa
ddle
d bi
chir
10 (7
)4.
741.
112.
030.
763.
735
62.0
461
.84
48.24
< 0.
001
45.8
747
.15
40.82
0.00
8La
ke T
anga
nyik
a &
oth
erC
olle
cted
by
ours
elve
s & P
urch
ased
from
pet
shop
18-
Aci
pens
erifo
rmes
Acip
ense
r sp
.St
urge
on12
(7)
4.42
1.74
2.23
0.95
-73
.36
74.1
867.02
0.00
457
.26
59.0
452.65
0.01
0W
este
rn Ja
pan
Purc
hase
d fr
om fa
rmer
192
Lepi
sost
eifo
rmes
Atra
ctos
teus
spat
ula
Alli
gato
r gar
10 (6
)4.
751.
833.
061.
074.
691
62.9
064
.65
58.30
0.01
053
.92
55.0
847.66
0.00
4So
uth
Am
eric
aPu
rcha
sed
from
pet
shop
202
Am
iifor
mes
Amia
cal
vaB
owfin
10 (5
)4.
951.
123.
040.
914.
741
62.8
764
.85
48.52
< 0.
001
53.4
455
.23
44.29
< 0.
001
Geo
rgia
, USA
Cou
rtesy
of D
r. M
. Kib
bey
& P
urch
ased
from
pet
shop
213
Hio
dont
iform
esH
iodo
n te
rgis
usM
oone
ye10
(4)
4.86
1.36
2.76
0.58
4.43
762
.77
64.4
352.41
< 0.
001
51.1
252
.91
35.37
< 0.
001
Ohi
o, U
SAC
ourte
sy o
f Dr.
M. K
ibbe
y (O
hio
Stat
e U
nive
rsity
)22
3O
steo
glos
sifo
rmes
Ost
eogl
ossu
mA
row
ana
10 (3
)5.
751.
251.
750.
743.
992
65.8
264
.75
50.65
< 0.
001
43.1
642
.69
40.09
0.02
4B
rasi
lPu
rcha
sed
from
pet
shop
233
Elop
iform
esM
egal
ops c
ypri
noid
esIn
do-P
acifi
c ta
rpon
10 (3
)4.
801.
972.
150.
703.
862
63.3
162
.63
59.75
0.01
846
.72
46.3
139.13
0.00
2C
ultu
red
in S
outh
east
Asi
aPu
rcha
sed
from
pet
shop
243
Alb
ulifo
rmes
Albu
la v
irga
taB
onef
ish
10 (6
)2.
611.
082.
110.
563.
021
51.1
453
.10
47.72
0.02
046
.04
47.4
834.80
< 0.
001
Haw
aii,
USA
Cou
rtesy
of D
r. A
. Ros
site
r (W
aiki
ki A
quar
ium
)25
3A
ngui
llifo
rmes
Angu
illa
japo
nica
Japa
nese
eel
10 (7
)4.
781.
181.
810.
723.
576
62.2
662
.81
49.54
< 0.
001
43.6
744
.29
39.73
0.01
5W
este
rn o
f Jap
anC
olle
cted
by
ours
elve
s & P
urch
ase
from
fish
ery
26D
eep
Sacc
opha
ryng
iform
eEu
ryph
aryn
xPe
lican
eel
#1
(0)
11.0
7-
3.24
-7.
601
--
--
--
--
The
Paci
fic o
f sou
ther
nC
ourte
sy o
f JA
MST
EC27
3C
lupe
iform
esC
lupe
a ha
reng
usA
tlant
ic h
errin
g10
(3)
6.29
2.81
1.62
0.63
4.18
168
.97
69.7
466.65
0.03
841
.47
41.9
137.12
0.01
2N
orw
ayPu
rcha
sed
from
fish
erm
an28
3G
onor
ynch
iform
esG
onor
ynch
us a
bbre
viat
usB
ighe
ad b
eake
d sa
ndfis
h11
(7)
5.21
1.47
2.18
1.01
4.06
370
.37
71.9
259.01
< 0.
001
52.5
253
.97
50.57
0.04
7Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves &
Pur
chas
e fr
om fi
sher
y29
3C
yprin
iform
esC
ypri
nus c
arpi
oC
omm
on c
arp
10 (5
)6.
062.
761.
990.
844.
308
68.2
970
.28
66.42
0.04
945
.81
47.3
342.74
0.02
4W
este
rn Ja
pan
Purc
hase
d fr
om fi
sher
man
303
Cha
raci
form
esPy
goce
ntru
s nat
tere
riR
ed p
iranh
a10
(6)
3.54
1.30
1.83
0.57
3.06
456
.90
57.5
451.44
0.00
643
.36
44.4
135.13
0.00
1So
uth
Am
eric
aPu
rcha
sed
from
pet
shop
313
Silu
rifor
mes
Silu
rus a
sotu
sFa
r Eas
tern
cat
fish
110
(53)
6.00
3.65
1.61
0.91
4.02
774
2.15
743.
06736.88
0.00
744
9.28
450.
12439.73
< 0.
001
Wes
tern
of J
apan
Col
lect
ed b
y ou
rsel
ves
323
Gym
notif
orm
esAp
tero
notu
s alb
ifron
sB
lack
gho
st10
(4)
5.72
1.26
1.88
0.64
4.06
765
.74
67.7
350.88
< 0.
001
43.5
545
.54
30.89
< 0.
001
Cul
ture
d in
Sou
thea
st A
sia
Purc
hase
d fr
om p
et sh
op33
3A
rgen
tinifo
rmes
Glo
ssan
odon
Dee
psea
smel
t10
(5)
4.42
1.76
1.55
0.65
3.21
361
.58
63.3
557.48
0.01
440
.78
42.5
337.71
0.02
4Se
a of
Japa
nPu
rcha
sed
from
fish
erm
an34
3O
smer
iform
esPl
ecog
loss
us a
ltive
lisA
yu12
(5)
4.43
1.21
2.28
0.67
3.82
572
.66
74.5
759.29
< 0.
001
56.7
758
.45
44.98
< 0.
001
Wes
tern
Japa
nC
olle
cted
by
ours
elve
s35
3Sa
lmon
iform
esO
ncor
hync
hus k
eta
Chu
m sa
lmon
10 (2
)5.
281.
691.
970.
603.
928
64.6
263
.83
56.78
0.00
244
.82
44.4
536.11
0.00
1N
orth
ern
Japa
nPu
rcha
sed
from
fish
erm
an36
3Es
ocifo
rmes
Esox
am
eric
anus
Red
fin p
icke
rel
10 (4
)6.
351.
392.
040.
684.
482
67.8
068
.84
52.84
< 0.
001
45.7
147
.21
38.71
0.00
3O
hio,
USA
Cou
rtesy
of D
r. M
. Kib
bey
(Ohi
o St
ate
Uni
vers
ity)
37D
eep
Stom
iifor
mes
Sigm
ops g
raci
leSl
ende
r fan
gjaw
11 (5
)6.
761.
262.
551.
215.
0575
.64
77.3
955.46
< 0.
001
56.0
656
.17
54.53
0.06
0Th
e Pa
cific
of s
outh
ern
Cou
rtesy
of J
AM
STEC
38D
eep
Ate
leop
odifo
rmes
Atel
eopu
s jap
onic
usPa
cific
jelly
nose
fish
12 (1
0)3.
671.
251.
920.
613.
197
68.5
662
.36
59.99
0.00
152
.90
50.7
742.94
< 0.
001
The
Paci
fic o
f sou
ther
nC
olle
cted
by
ours
elve
s39
4A
ulop
iform
esAu
lopu
s jap
onic
usJa
pane
se th
read
-sai
l fis
h12
(6)
3.38
1.57
1.73
0.80
2.91
67.6
269
.11
65.38
0.03
951
.58
53.4
649.18
0.03
6Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
40D
eep
Myc
toph
iform
esD
iaph
us w
atas
eiW
atas
es la
nter
nfis
h10
(6)
5.22
1.62
4.01
1.68
5.82
64.3
466
.05
55.90
0.00
159
.76
61.3
156.50
0.02
2Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
414
Lam
prid
iform
esLa
mpr
is g
utta
tus
Opa
h10
(3)
2.91
1.72
2.11
0.80
3.11
354
.75
51.71
56.0
80.
413
46.6
746
.45
41.74
0.00
8Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
424
Poly
mix
iifor
mes
Poly
mix
ia ja
poni
caSi
lver
eye
10 (5
)2.
881.
122.
661.
243.
694
52.9
554
.35
48.49
0.01
151
.91
53.7
550.47
0.06
4Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves
434
Perc
opsi
form
esPe
rcop
sis o
mis
com
aycu
sTr
out-p
erch
10 (6
)4.
471.
082.
130.
823.
707
60.9
062
.44
47.86
< 0.
001
46.9
348
.12
42.37
0.01
0O
hio,
USA
Cou
rtesy
of D
r. M
. Kib
bey
(Ohi
o St
ate
Uni
vers
ity)
444
Gad
iform
esG
adus
mac
roce
phal
usPa
cific
cod
10 (7
)2.
880.
962.
250.
923.
251
52.5
853
.57
45.39
0.00
248
.11
48.0
744.65
0.01
9Th
e Pa
cific
of n
orth
ern
Purc
hase
d fr
om fi
sher
man
454
Oph
idiif
orm
esSi
rem
bo im
berb
isG
olde
n cu
sk10
(6)
3.34
1.00
2.41
1.02
3.56
155
.35
55.3
346.24
< 0.
001
49.5
951
.56
46.62
0.02
6Th
e Pa
cific
of s
outh
ern
Col
lect
ed b
y ou
rsel
ves &
Pur
chas
ed fr
om fi
sher
y46
4B
atra
choi
difo
rmes
Batr
acho
moe
usTh
ree-
spin
ed to
adfis
h10
(3)
6.52
1.46
1.84
0.60
4.43
368
.35
67.5
253.81
< 0.
001
43.5
844
.47
36.08
0.00
2So
uthe
ast A
sia
Purc
hase
d fr
om p
et sh
op47
Dee
pLo
phiif
orm
esLo
phiu
s litu
lon
Ang
lerf
ish
12 (8
)7.
842.
533.
411.
156.
222
86.5
885
.94
75.97
< 0.
001
66.7
264
.99
57.13
< 0.
001
The
Paci
fic o
f sou
ther
nC
olle
cted
by
ours
elve
s & P
urch
ased
from
fish
ery
485
Mug
ilifo
rmes
Mug
il ce
phal
usFl
athe
ad m
ulle
t11
(4)
4.40
1.51
2.35
0.87
3.87
867
.02
68.7
059.45
0.00
253
.44
53.2
847.41
0.00
5W
este
rn Ja
pan
Cou
rtesy
of D
r.M. N
akam
ura
(Cor
bicu
la R
esea
rch
Inst
itute
)49
5A
ther
inifo
rmes
Athe
rion
ely
mus
Bea
rded
silv
ersi
de10
(6)
4.17
0.88
3.08
0.86
4.52
59.3
561
.09
43.77
< 0.
001
53.5
955
.58
43.16
< 0.
001
The
Paci
fic o
f sou
ther
nC
olle
cted
by
ours
elve
s50
5B
elon
iform
esSt
rong
ylur
a an
asto
mel
laN
eedl
efis
h10
(8)
3.85
1.14
1.96
0.66
3.30
558
.15
56.4
048.82
< 0.
001
44.8
840
.44
38.01
0.00
3W
este
rn Ja
pan
Cou
rtesy
of D
r.M. N
akam
ura
(Cor
bicu
la R
esea
rch
Inst
itute
)50
-2-
Bel
onifo
rmes
Prog
nich
thys
bre
vipi
nnis
Shor
tfin
flyin
gfis
h10
(6)
4.28
0.92
1.30
0.27
-59
.93
61.6
344.66
< 0.
001
36.0
537
.67
20.01
< 0.
001
The
Paci
fic o
f sou
ther
nPu
rcha
sed
from
fish
erm
an51
5C
yprin
odon
tifor
mes
Gam
busi
a af
finis
Mos
quito
fish
10 (5
)5.
130.
731.
710.
233.
665
63.2
964
.66
39.93
< 0.
001
41.2
443
.23
16.62
< 0.
001
Wes
tern
Japa
nC
olle
cted
by
ours
elve
s52
Dee
pSt
epha
nobe
ryci
form
Poro
mitr
a cr
assi
ceps
Cre
sted
big
scal
e10
(6)
4.96
1.13
4.81
2.25
6.61
562
.91
64.7
748.69
< 0.
001
63.7
565
.56
62.23
0.06
1Th
e Pa
cific
of n
orth
ern
Pres
erve
d in
Hok
kaid
o U
nive
rsity
Mus
eum
53D
eep
Ber
ycifo
rmes
Bery
x sp
lend
ens
Sple
ndid
alfo
nsin
o11
(6)
2.97
1.25
2.92
1.42
4.00
553
.76
53.4
350.56
0.02
353
.90
55.1
652.69
0.07
3Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
544
Zeifo
rmes
Zeus
fabe
rJo
hn d
ory
12 (6
)2.
200.
632.
711.
233.
589
55.8
957
.89
43.77
< 0.
001
62.1
964
.09
59.54
0.03
1Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
555
Gas
tero
stei
form
esG
aste
rost
eus
sp.
Thre
e-sp
ined
stic
kleb
ack
10 (5
)6.
902.
021.
520.
384.
443
69.8
371
.81
60.34
< 0.
001
39.3
841
.35
26.91
< 0.
001
Nor
ther
n Ja
pan
(the
Sea
ofC
ourte
sy o
f Dr.
S. M
ori (
Gifu
Eco
nom
y U
nive
rsity
)56
5Sy
nbra
nchi
form
esM
asta
cem
belu
s moo
rii
Spin
y ee
l12
(3)
3.82
1.36
1.95
0.41
3.28
469
.66
70.4
262.13
0.00
252
.63
51.9
133.30
< 0.
001
Lake
Tan
gany
ika
Col
lect
ed b
y ou
rsel
ves
575
Scor
paen
iform
esSe
bast
iscu
s mar
mor
atus
Fals
e ke
lpfis
h10
(5)
3.51
0.81
2.17
0.85
3.37
656
.06
58.0
242.05
< 0.
001
47.3
249
.32
43.13
0.01
3Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
585
Perc
iform
esLa
teol
abra
x ja
poni
cus
Japa
nese
seap
erch
11 (3
)4.
461.
401.
600.
523.
269
67.1
465
.32
57.92
< 0.
001
44.5
843
.90
35.89
0.00
1Th
e Pa
cific
of s
outh
ern
Purc
hase
d fr
om fi
sher
man
58-2
-Se
riol
a qu
inqu
erad
iata
Japa
nese
am
berja
ck10
0 (4
4)2.
831.
641.
510.
72-
523.
0851
9.15
516.29
0.00
338
8.44
388.
68359.58
< 0.
001
Sea
of Ja
pan
Purc
hase
d fr
om fi
sher
man
58-3
-Pe
riss
odus
mic
role
pis
L. T
anga
nyik
a sc
ale-
eate
r10
(3)
9.63
1.56
1.99
0.41
-75
.86
76.5
950.74
< 0.
001
45.1
544
.84
37.51
0.00
2La
ke T
anga
nyik
aC
olle
cted
by
ours
elve
s59
5Pl
euro
nect
iform
esH
ippo
glos
soid
es d
ubiu
sFl
athe
ad fl
ound
er50
(28)
4.11
3.32
2.23
1.05
3.65
5310.37
312.
2631
2.37
1.00
023
4.13
234.
55219.42
< 0.
001
Wes
tern
Japa
nPu
rcha
sed
from
fish
erm
an59
-2-
Para
licht
hys o
livac
eus
Oliv
e flo
unde
r51
(21)
4.55
4.00
2.78
1.62
-330.43
330.
8533
2.43
1.00
026
6.02
265.
99263.35
0.03
1W
este
rn Ja
pan
Purc
hase
d fr
om fi
sher
man
605
Tetra
odon
tifor
mes
Taki
fugu
rubr
ipes
Japa
nese
puf
ferf
ish
10 (4
)3.
910.
932.
890.
784.
2458
.18
60.0
044.77
< 0.
001
52.2
853
.94
41.38
< 0.
001
Wes
tern
Japa
nPu
rcha
sed
from
fish
erm
an61
1C
oela
cant
hifo
rmes
Latim
eria
cha
lum
nae
Coe
laca
nth#
2 (2
)4.
200.
134.
330.
815.
889
--
--
--
--
Coa
st o
f Tan
zani
aPr
eser
ved
in T
okyo
Inst
itute
of T
echn
olog
y62
1C
erat
odon
tifor
mes
Prot
opte
rus a
ethi
opic
usM
arbl
ed lu
ngfis
h11
(5)
3.68
1.43
2.88
1.21
4.15
963
.42
64.6
658.40
0.00
858
.24
59.8
254.54
0.01
7Ea
st A
fric
aC
olle
cted
by
ours
elve
s & P
urch
ased
from
pet
shop
# N
umbe
rs o
f ord
er fo
llow
Nel
son
(200
2).
‡ Sp
ecie
s who
se sa
mpl
es a
re le
ss th
an 1
0.
* Fi
gure
s in
pare
nthe
ses i
ndic
ate
the
num
ber o
f rig
hty
indi
vidu
als i
n th
e sa
mpl
es.

Supplementary Figure S1. Classification and phylogenetic tree of the extant orders of teleost fishes used in this study. Phylogenetic trees are modified following Betancur-R et al. (2013). The classification is based on Nelson (2006), with a slight modification following Betancur-R et al. (2013) as shown in Supplementary Table 1 (colors are the same as those in Fig. 3A). The blue plot shows deep-sea fishes.

Supplementary Figure S2. Laterality of the Japanese stingray (Dasyatis akajei). (A, B) Cartilaginous bones showing the data points used for measurements. (A) Ventral view of the jaw arch and a close-up of the left mandibular cartilage showing the data points for the index of asymmetry (IAS) of the mandibles. The arrow indicates the height of the mandible at the posterior end (HMPE), which is the distance from the postero-dorsal edge of the sustentaculum in the mandibular cartilage (PE) to the ventral corner of the mandibular cartilage (VC). In car-tilaginous fishes (except in chimaeras), the sustentaculum functions as the mandibular surface of the hyomandibular-mandibular joint, and the nodule on the VC functions as the attachment point of the depressor mandibularis (Dean and Motta, 2004; Wilga 2005), such that the HMPE provides a functional measure equivalent to that of teleost fishes. (B) Ventral view of head and vertebrae showing data points for the head angle (θ ), which is the angle between the head and vertebrae on the ventral side of the anterior part of the skeleton. CC: the small fora-men near the center of the cranial cartilage; PC: the midline of the postero-ventral margin of the cranium; PV: the first large process on the ventral sagittal line of the vertebra. (C, D) Frequency distributions of the (C) IAS of the mandibles and (D) θ in the Japanese stingray. Lines show the probability densities derived from the three models (see Methods). The model selected by the lowest Akaike information criterion (AIC) value is indicated by the thick line.

Supplementary Figure S3. Relationship between growth and laterality in large-mouth bass (Micropterus salmoides). The absolute values of the index of asymmetry (IAS) of mandibles and the head angles (θ ) in three size classes did not differ sig-nificantly (ANOVA, P = 0.2379 for IAS and P = 0.7791 for θ ). Individuals were sam-pled from the same population in Lake Biwa, Japan, in 2004–2008. Vertical bars indicate standard deviations (s.d.). Small fish were primarily 0 + (n = 20, standard length [SL; mean ± s.d.] = 76.1 ± 4.1 mm), medium fish were primarily 1+ (n = 20, SL = 125.1 ± 13.6 mm), and large fish were primarily ≥ 2 + (n = 20, SL = 173.4 ± 16.8 mm).
Supplementary Figure S4. Laterality of the flatfishes. Frequency distributions of the (A) index of symmetry (IAS) of the mandibles and (B) head angles in flathead flounder (Hippoglossoides dubius; upper) and olive flounder (Paralichthys olivaceus; lower). The lines are the same as those used in Fig. 2. In both species, the model with the best fit to the distributions of IAS was fluctuating asymmetry, whereas the best model for head angle was anti-symmetry (see Supplementary Table 1).

Supplementary Figure S5. Laterality of the hagfish. (A, B) Data for measurements from inshore hagfish (Eptatretus burgeri). (A) Ventral view of the mouthparts of a moderately boiled fish sample. A pair of flat cartilaginous structures behind the tooth plates is exposed. FC: the length of the flat cartilaginous structure used for the index of asymmetry (IAS). (B) Dorsal view of the anterior part of the body in a transparent preparation showing data for the head angle. CN: the center of the cartilaginous rings of the nasal tube; MC: the posterior end of the mem-branous cranium; DP: the dorsal datum point equidistant from MC to CN–MC on the sagittal line. Head angle (θ ) is defined as the angle of the line extending anteriorly from MC–DP to CN–MC. A fish with a clockwise positive angle is defined as left type, whereas a fish with a negative angle is defined as right type. The definitions of right and left types are consistent with those shown in the ventral view of Fig. 1. (C, D) Frequency distributions of (C) IAS of the flat cartilaginous structure and (D) θ in inshore hagfish. The lines are the same as those used in Fig. 2. Both distributions showed anti-symmetry (see Supplementary Table 1).

Supplementary Figure S6. Laterality of the lamprey. (A, B) Data points used for measurements in arctic lamprey (Lampetra japonica). (A) Ventral view of a mouthpart in which a pair of spinose cartilaginous structures behind the suction cup are exposed in a moderately boiled fish sample. SC: the length of the spinose cartilage used for the index of asymmetry (IAS). (B) Dorsal view of the anterior part of the body in a transparent specimen showing the data points used for head angle (θ ) on the ventral sagittal line. PD: the anterior end of the posterior dorsal cartilage; PO: the posterior end of the postero-occipital cartilaginous ring; DP: the dorsal datum point equidistant to PD–PO. θ is defined as the angle from the line extending anteriorly from PO–DP to PD–PO. A fish with a clockwise positive angle is defined as left type, whereas a fish with a negative angle is defined as right type. (C, D) Frequency distributions of the (C) IAS of spinose cartilaginous structures and (D) θ in arctic lamprey. The lines are the same as those in Fig. 2. Both distributions showed anti-symmetry (see Supplementary Table 1).

Supplementary Figure S7. Degrees of laterality in extant cartilaginous fish orders and sturgeons. Two measures of the laterality, the index of asymmetry (IAS) of the mandibles and the head angle, are displayed as in Fig. 3A. Fish species are grouped into four color-coded categories.