Embed Size (px)
DESCRIPTION
ffcghf
Citation preview
9/12/2015
1
LĄSTELIŲ GYVYBINIO CIKLOKONTROLĖ
Dauguma daugialąsčio organizmo ląstelių yra diferencijuotos specialių funkcijų atlikimui ir nesidalija. Tokios ląstelės sustabdo savo ciklą ir patenka į specializuotą G1 periodą vadinamą ramybės periodu – G0
Interfazė {G1, S, G2, G0} + Mitozė {kariokinezė, citokinezė}
Interfazėje skiriami 3 periodai: I periodas vadinamas G1 arba presintetiniu. Jis prasideda tuoj po pasidalijimo. Ląstelėje kaupiama RNR, baltymai, lipidai, angliavandeniai. Auga membranos, didėja branduolys, ląstelė pasiekia suaugusios dydį, funkciškai bei struktūriškai diferencijuojasi. Ląstelėje išlieka mitozėje susiformavęs diploidinis chromosomų rinkinys, kuriame vieną chromosomą sudaro viena chromatidė (2n, 2c).
II periodas vadinamas S arba sintezės. Šiam periodui būdinga DNR replikacija ir chromosominių struktūrų sudvigubėjimas. Taip pat sintezuojami chromosominiai baltymai.
III periodas vadinamas G2 arbapriešmitotiniu. Jame išlieka diploidinis dvigubų chromosomų rinkinys (2n, 4c). Ląstelėje kaupiama energija, sintezuojama ATP, RNR, baltymai. Dvigubėja mitochondrijos, kiti organoidai. Ląstelė ruošiasi dalijimuisi.
Ląstelės gyvybinis ciklas
9/12/2015
2
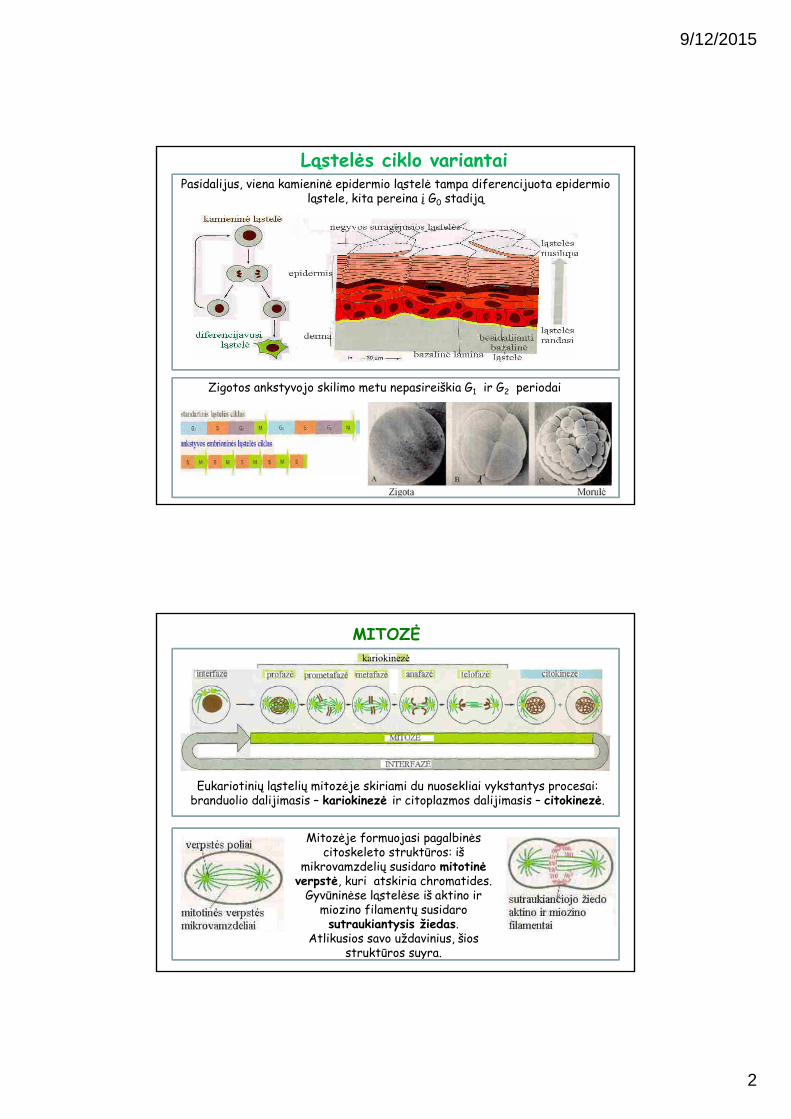
Pasidalijus, viena kamieninė epidermio ląstelė tampa diferencijuota epidermio ląstele, kita pereina į G0 stadiją
Zigotos ankstyvojo skilimo metu nepasireiškia G1 ir G2 periodai
Ląstelės ciklo variantai
MITOZĖ
Mitozėje formuojasi pagalbinės citoskeleto struktūros: iš
mikrovamzdelių susidaro mitotinė verpstė, kuri atskiria chromatides.
Gyvūninėse ląstelėse iš aktino ir miozino filamentų susidaro sutraukiantysis žiedas.
Atlikusios savo uždavinius, šios struktūros suyra.
Eukariotinių ląstelių mitozėje skiriami du nuosekliai vykstantys procesai: branduolio dalijimasis – kariokinezė ir citoplazmos dalijimasis – citokinezė.

9/12/2015
3
Dukterinės centrosomos atsiskiria ir juda link priešingų ląstelės polių. Nuo centrosomų visomis kryptimis tįsta mikrovamzdeliai.
Kiekviena centrosoma sukuria savo mikrovamzdelių komplektą, kurie sąveikaudami sudaro dipolio pavidalo mitotinę verpstę.
Interfazės S periodo gale ląstelė dvigubina centrosomas.
Kol dar nesuiręs branduolio apvalkalas, dalis mikrovamzdelių stabilizuojami: kai kuriuos mikrovamzdelius, vadinamus poliniais, suriša specialūs baltymai.
PROFAZĖ
Įvyksta staigus branduolio apvalkalo (membranos, branduolio porų bei laminų) suirimas į smulkius fragmentus.
Branduolio išorėje buvę mikrovamzdeliai dabar gali susijungti su chromosomomis.
Kinetochoriniai mikrovamzdeliai susijungia su chromosomis per specialų baltymų kompleksą, vadinamą kinetochoru.
Kiekviena sudvigubėjusi chromosoma turi du kinetochorus, atgręžtus į priešingas puses; todėl kiekviena replikuota chromosoma tampa susieta su abiem verpstės poliais.
Mikrovamzdelių, susietų su kiekvienu kinetochorų, skaičius yra įvairus, pvz., žmogui – 20-40, mielėms – 1.
PROMETAFAZĖ
9/12/2015
4
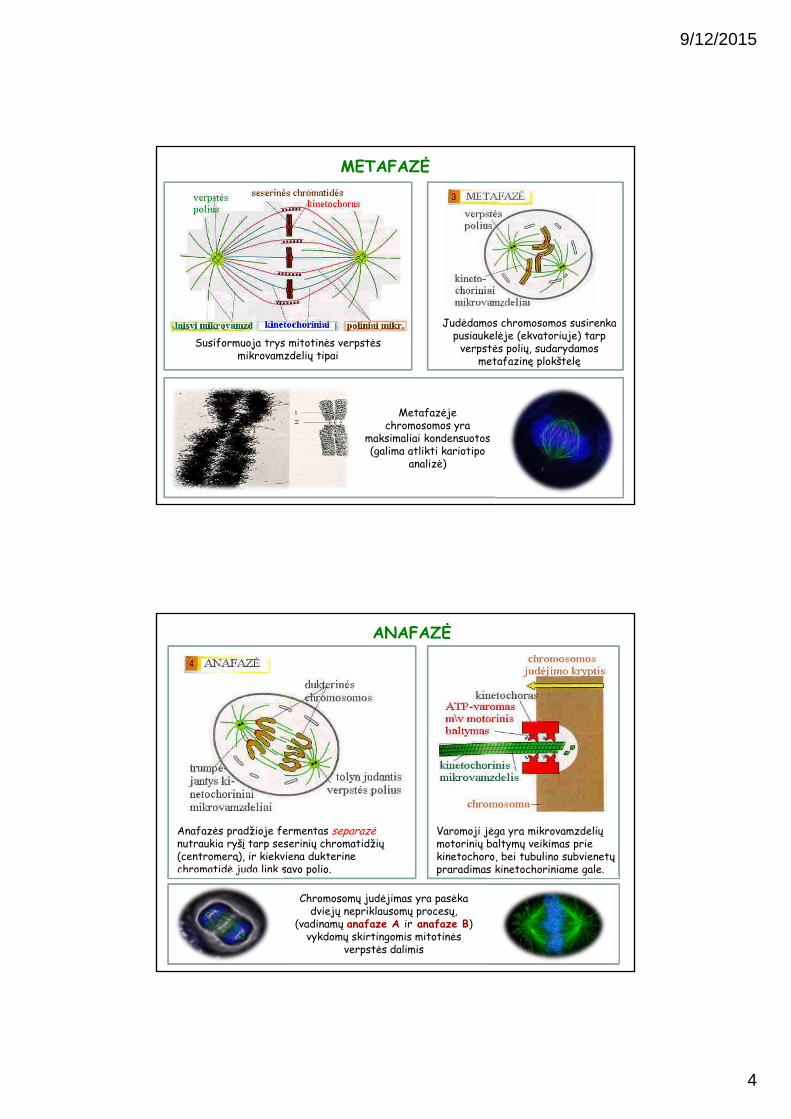
Susiformuoja trys mitotinės verpstės mikrovamzdelių tipai
Metafazėje chromosomos yra
maksimaliai kondensuotos (galima atlikti kariotipo
analizė)
METAFAZĖ
Judėdamos chromosomos susirenkapusiaukelėje (ekvatoriuje) tarp
verpstės polių, sudarydamos metafazinę plokštelę
Chromosomų judėjimas yra pasėka dviejų nepriklausomų procesų,
(vadinamų anafaze A ir anafaze B)vykdomų skirtingomis mitotinės
verpstės dalimis
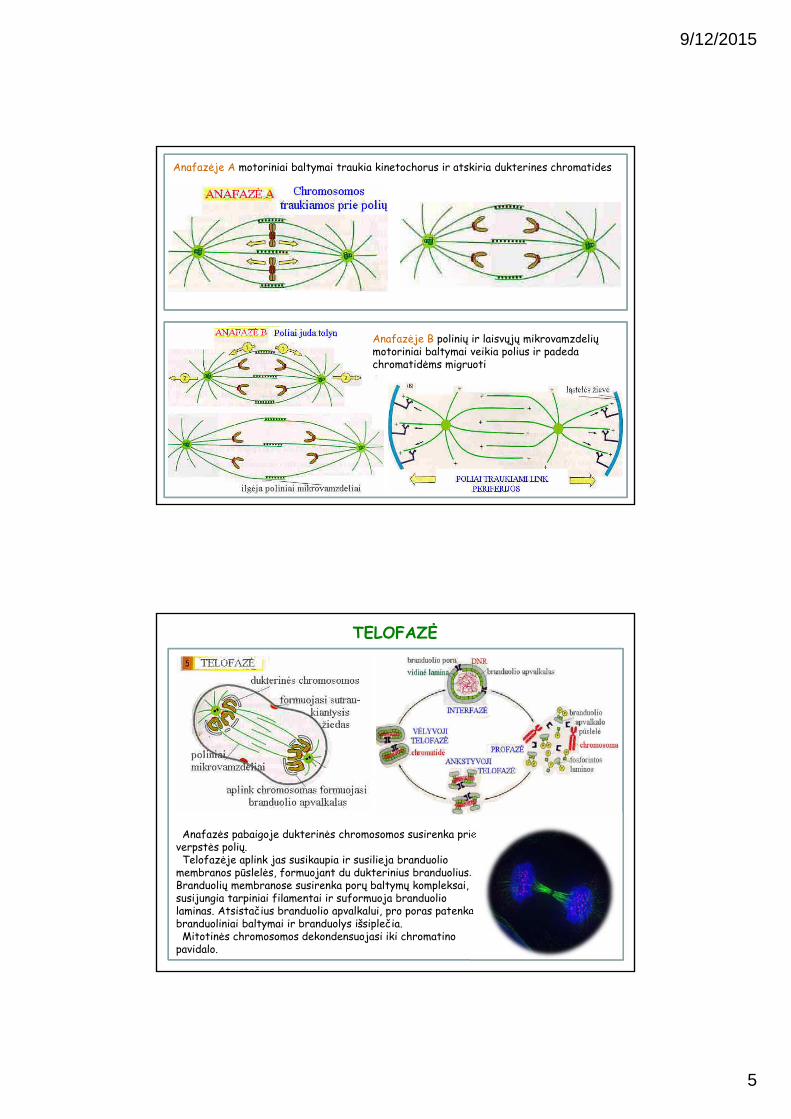
ANAFAZĖ
Anafazės pradžioje fermentas separazėnutraukia ryšį tarp seserinių chromatidžių (centromerą), ir kiekviena dukterine chromatidė juda link savo polio.
Varomoji jėga yra mikrovamzdelių motorinių baltymų veikimas prie kinetochoro, bei tubulino subvienetų praradimas kinetochoriniame gale.
9/12/2015
5
Anafazėje B polinių ir laisvųjų mikrovamzdelių motoriniai baltymai veikia polius ir padeda chromatidėms migruoti
Anafazėje A motoriniai baltymai traukia kinetochorus ir atskiria dukterines chromatides
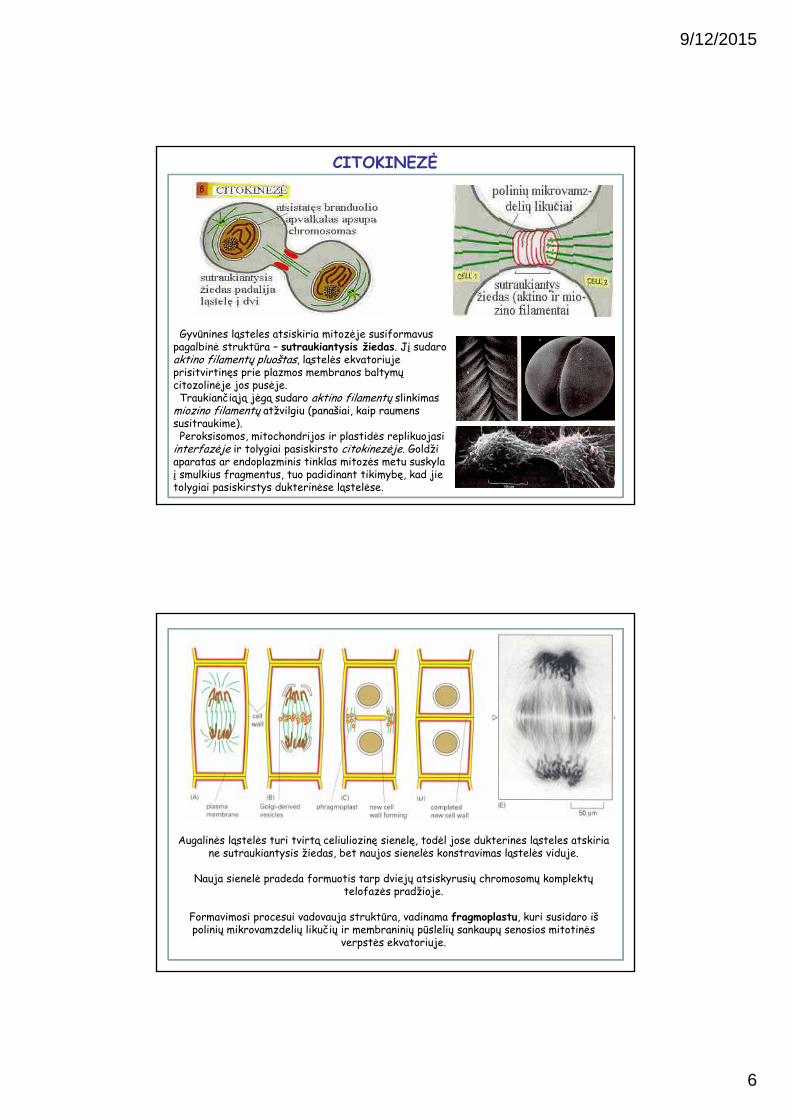
Anafazės pabaigoje dukterinės chromosomos susirenka prie verpstės polių. Telofazėje aplink jas susikaupia ir susilieja branduolio
membranos pūslelės, formuojant du dukterinius branduolius. Branduolių membranose susirenka porų baltymų kompleksai, susijungia tarpiniai filamentai ir suformuoja branduolio laminas. Atsistačius branduolio apvalkalui, pro poras patenka branduoliniai baltymai ir branduolys išsiplečia. Mitotinės chromosomos dekondensuojasi iki chromatino
pavidalo.
TELOFAZĖ
9/12/2015
6
CITOKINEZĖ
Gyvūnines ląsteles atsiskiria mitozėje susiformavus pagalbinė struktūra – sutraukiantysis žiedas. Jį sudaro aktino filamentų pluoštas, ląstelės ekvatoriuje prisitvirtinęs prie plazmos membranos baltymų citozolinėje jos pusėje. Traukiančiąją jėgą sudaro aktino filamentų slinkimas
miozino filamentų atžvilgiu (panašiai, kaip raumens susitraukime).Peroksisomos, mitochondrijos ir plastidės replikuojasi
interfazėje ir tolygiai pasiskirsto citokinezėje. Goldži aparatas ar endoplazminis tinklas mitozės metu suskyla į smulkius fragmentus, tuo padidinant tikimybę, kad jie tolygiai pasiskirstys dukterinėse ląstelėse.
Augalinės ląstelės turi tvirtą celiuliozinę sienelę, todėl jose dukterines ląsteles atskiria ne sutraukiantysis žiedas, bet naujos sienelės konstravimas ląstelės viduje.
Nauja sienelė pradeda formuotis tarp dviejų atsiskyrusių chromosomų komplektų telofazės pradžioje.
Formavimosi procesui vadovauja struktūra, vadinama fragmoplastu, kuri susidaro iš polinių mikrovamzdelių likučių ir membraninių pūslelių sankaupų senosios mitotinės
verpstės ekvatoriuje.
b

9/12/2015
7
Besiformuojant lytinėmsląstelėms (gametoms), stebimaypatinga mitotinio dalijimosimodifikacija, vadinama mejoze.
Mejozės metu į dukterinesląsteles patenka tik po vienąchromosomą iš kiekvienoshomologinių chromosomų poros.
Toks pusiau sumažėjęschromosomų skaičius vadinamashaploidiniu rinkiniu.
Mejozėje, dėl paveldimosiosmedžiagos persiskirstymo(rekombinacijos) homologinėsechromosomose, susidaro įvairustėvinių ir motininių chromosomųsantykis haploidiniuose gametųrinkiniuose.
MEJOZĖ
MEJOZĖ
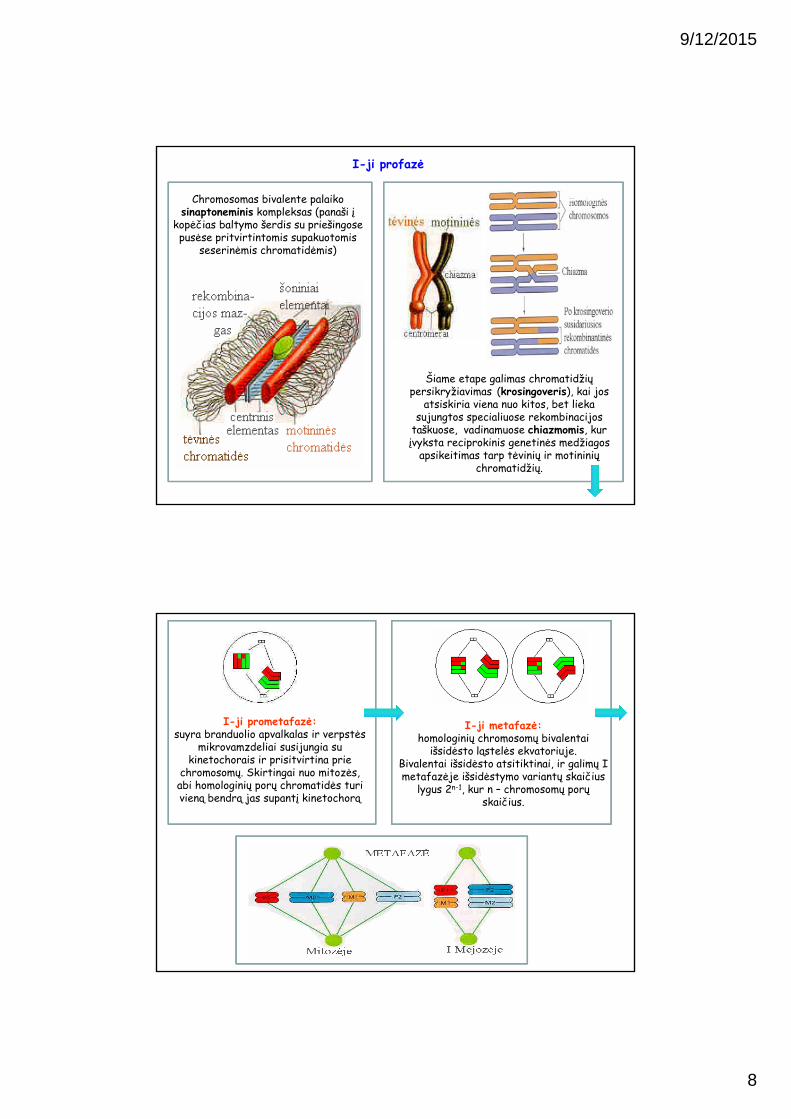
I-ji profazė:kaip ir įprastoje mitozėje, replikuojasi chromosomos, sudvigubėja centrosomos ir formuojasi verpstė.
Tačiau prieš suyrant branduolio apvalkalui, kiekviena replikavusi
chromosoma “susiranda” savo homologą ir įvyksta jų konjugacija.
Chromosomos konjuguoja savo homologinėmis sritimis: vienos
chromosomų poros chromatidės tiksliai prisijungia prie atitinkamų homologinių
chromatidžių formuojant bivalentus(tetrados).
9/12/2015
8
Chromosomas bivalente palaiko sinaptoneminis kompleksas (panaši į
kopėčias baltymo šerdis su priešingose pusėse pritvirtintomis supakuotomis
seserinėmis chromatidėmis)
Šiame etape galimas chromatidžiųpersikryžiavimas (krosingoveris), kai jos
atsiskiria viena nuo kitos, bet lieka sujungtos specialiuose rekombinacijos
taškuose, vadinamuose chiazmomis, kur įvyksta reciprokinis genetinės medžiagos
apsikeitimas tarp tėvinių ir motininių chromatidžių.
I-ji profazė
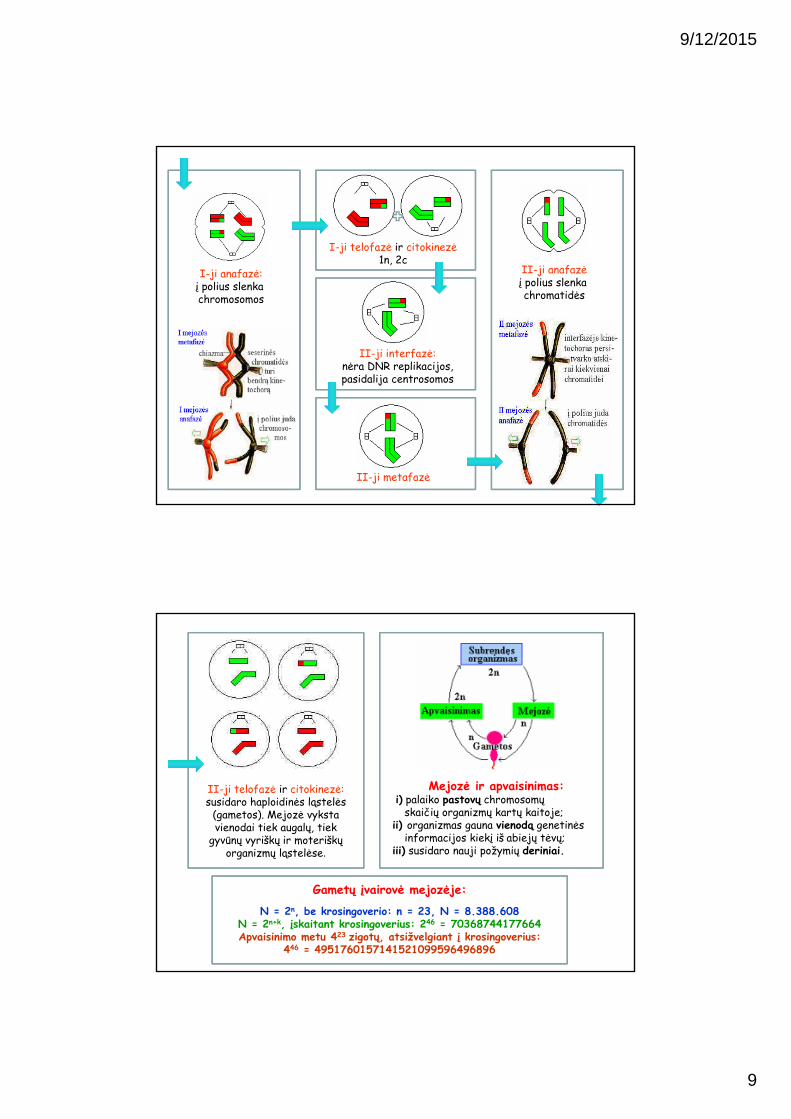
I-ji prometafazė:suyra branduolio apvalkalas ir verpstės
mikrovamzdeliai susijungia su kinetochorais ir prisitvirtina prie
chromosomų. Skirtingai nuo mitozės, abi homologinių porų chromatidės turi vieną bendrą jas supantį kinetochorą
I-ji metafazė:homologinių chromosomų bivalentai
išsidėsto ląstelės ekvatoriuje. Bivalentai išsidėsto atsitiktinai, ir galimų I metafazėje išsidėstymo variantų skaičius
lygus 2n-1, kur n – chromosomų porų skaičius.
9/12/2015
9
I-ji anafazė:į polius slenka chromosomos
I-ji telofazė ir citokinezė1n, 2c
II-ji interfazė:nėra DNR replikacijos,pasidalija centrosomos
II-ji metafazė
II-ji anafazėį polius slenka chromatidės
Mejozė ir apvaisinimas:i) palaiko pastovų chromosomų
skaičių organizmų kartų kaitoje;ii) organizmas gauna vienodą genetinės
informacijos kiekį iš abiejų tėvų;iii) susidaro nauji požymių deriniai.
Gametų įvairovė mejozėje:
N = 2n, be krosingoverio: n = 23, N = 8.388.608N = 2n+k, įskaitant krosingoverius: 246 = 70368744177664Apvaisinimo metu 423 zigotų, atsižvelgiant į krosingoverius:
446 = 4951760157141521099596496896
II-ji telofazė ir citokinezė:susidaro haploidinės ląstelės
(gametos). Mejozė vyksta vienodai tiek augalų, tiek
gyvūnų vyriškų ir moteriškų organizmų ląstelėse.

9/12/2015
10
LĄSTELĖS CIKLO KONTROLĖS SISTEMALąstelės ciklo kontrolę atlieka baltymai,
kurie nepriklauso nuo baltymų, vykdančių dalijimosi procesus.Baltymų aktyvumas keičiamas juos
fosforinant proteino kinazėmis ir defosforinant fosfatazėmis.Tačiau skirtingos kinazės pačios yra
„įjungiamos“/„išjungiamos“ tam tikruose ciklo perioduose ir šį fermentinio aktyvumo kitimą valdo spec. baltymų rūšis – ciklinai.Todėl ląstelės ciklo kontrolės sistemos kinazės vadinamos ciklino priklausomomis(Cpk).
Ląstelės ciklo kontrolės sistema (LCKS), aktyvuoja fermentus bei kitus baltymus, reikalingus kiekvieno ciklo eigai, ir deaktyvuoja juos, procesui pasibaigus.
LCKS taip pat užtikrina, kad kiekviena stadija pasibaigs, prieš prasidedant sekančiai.
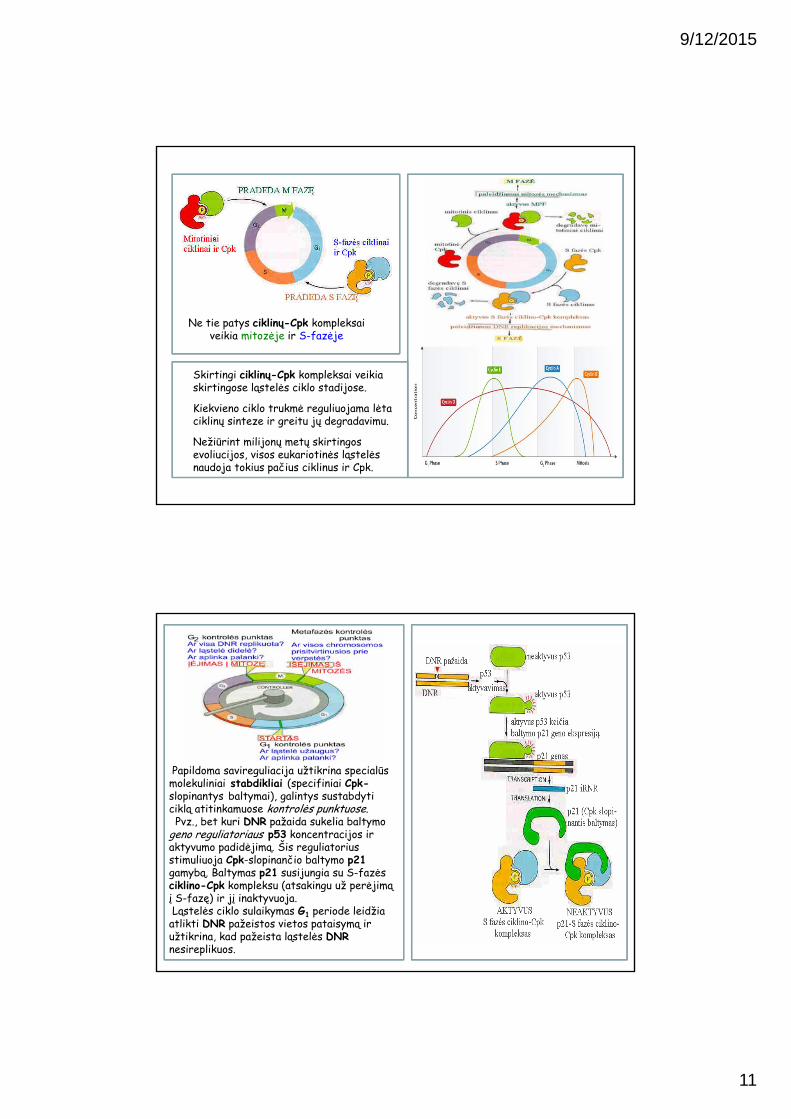
Baltyminis kompleksas, sudarytas iš dviejų subvienetų – reguliacinio (M-ciklino) ir fermentinio (mitotinės Cpk) vadinamas M-fazę pagreitinančiu faktorium (MPF).
MPF aktyvumo ir ciklino koncentracijos periodiškumas
Didžiausias MPF aktyvumas sutampa su ciklino koncentracijos maksimumu
Būtent ciklinų koncentracijos kitimai nusako ląstelės ciklo sinchronizacija, nes kinazių yra visada, o ciklinų kiekiai kinta.
Pvz., mitotinio ciklino komponento sintezė prasideda pasidalijus ląstelei ir tęsiasi visą interfazę. Palaipsniui kylant ciklino koncentracijai, jis suaktyvina MPFir visas mitozei reikalingas kinazės (chromosomų kondensacijai, branduolio apvalkalo suirimui, mikrovamzdelių verpstės formavimui ir t.t.).
Staigią ciklino destrukciją mitozėje užtikrina pačių kinazių aktyvumas, nes jos aktyvuoja ir proteolitinius fermentus, kurie ardo cikliną.
Sumažėjus ciklino koncentracijai, mažėja kinazių aktyvumas, neveikia cikliną ardantys fermentai, ciklinas vėl pradeda kauptis, aktyvuojamos kinazės, ir procesas kartojasi.
9/12/2015
11
Ne tie patys ciklinų-Cpk kompleksai veikia mitozėje ir S-fazėje
Skirtingi ciklinų-Cpk kompleksai veikia skirtingose ląstelės ciklo stadijose.
Kiekvieno ciklo trukmė reguliuojama lėta ciklinų sinteze ir greitu jų degradavimu.
Nežiūrint milijonų metų skirtingos evoliucijos, visos eukariotinės ląstelės naudoja tokius pačius ciklinus ir Cpk.
Papildoma savireguliacija užtikrina specialūs molekuliniai stabdikliai (specifiniai Cpk-slopinantys baltymai), galintys sustabdyti ciklą atitinkamuose kontrolės punktuose.Pvz., bet kuri DNR pažaida sukelia baltymo
geno reguliatoriaus p53 koncentracijos ir aktyvumo padidėjimą. Šis reguliatorius stimuliuoja Cpk-slopinančio baltymo p21gamybą. Baltymas p21 susijungia su S-fazės ciklino-Cpk kompleksu (atsakingu už perėjimą į S-fazę) ir jį inaktyvuoja. Ląstelės ciklo sulaikymas G1 periode leidžia
atlikti DNR pažeistos vietos pataisymą ir užtikrina, kad pažeista ląstelės DNRnesireplikuos.

9/12/2015
12
DAUGIALĄSČIO ORGANIZMO LĄSTELIŲ KIEKIO KONTROLĖSignalai iš kitų ląstelių (augimo faktoriai) per membranos receptorius stimuliuoja
ląstelių dalijimąsi, kad jis vyktu tik tada, jei tai pageidaujama organizmui.
Retinoblastomos (Rb) baltymas yra susijungęs su tam tikru baltymu geno reguliatorium ir blokuoja ląstelės dalijimuisi reikalingų baltymų gamybą. Ekstraląsteliniai augimo faktoriai aktyvuoja G1 periodo (S-fazės) ciklino-Cpkkompleksus. Pastarieji fosforina Rb-baltymą, jis atpalaiduoja reguliacinį baltymą, ir aktyvuojama dalijimuisi reikalingų genų transkripciją.
Baltymai – augimo faktoriai ir jų veikimasAugimo faktorius Veikiami audiniai
Trombocitų kilmės (PDGF)Epidermalinis (EGF)Fibroblastų (FGF)Hepatocitų (HGF)EritropoetinasNervų (NDF)
Jungiamojo audinio ląstelių dalijimasisEpidermio ląstelių dalijimasisFibroblastų dalijimasisKepenų ląstelių dalijimasisBesivystančių eritrocitų dalijimasis ir diferenciacija.Palaiko nervinių ląstelių išlikimą
Ląstelės turi ribotą dalijimųsi skaičių (ląstelių senėjimas, Hayflick limit). Pvz., žmogaus gemalo fibroblastai nustoja dalintis po 80 ciklų, net ir veikiami augimo faktoriais. Ląstelėje kaupiasi proteino kinazes slopinantys baltymai, mažėja Cpk kiekis, ir ląstelė pereina į G0periodą. Senėjimas priklauso nuo amžiaus: 40-mečio žmogaus fibroblastas nustoja dalintis po 40 ciklų, o 80-mečio – po 30.
Augimo faktoriai įvairiomis kombinacijomis selektyviai reguliuoja kiekvieno ląstelių tipodalijimąsi ir išlikimą. Jei ląstelė negauna išorinių signalų, joje aktyvuojama intraląstelinė suicidinė programa ir ląstelė miršta.
Senėjimas siejamas su telomeromis –pasikartojančiomis DNR sekomis (pvz., TTAGGG) chromosomų galuose. Šios sekos nesireplikuoja, ir sintezuojamos fermentu telomeraze. Ląstelių senėjimas koreliuoja su šių
sekų skaičiaus sumažėjimu (t.y., ląstelė nesugeba išlaikyti telomerų ilgio).
9/12/2015
13
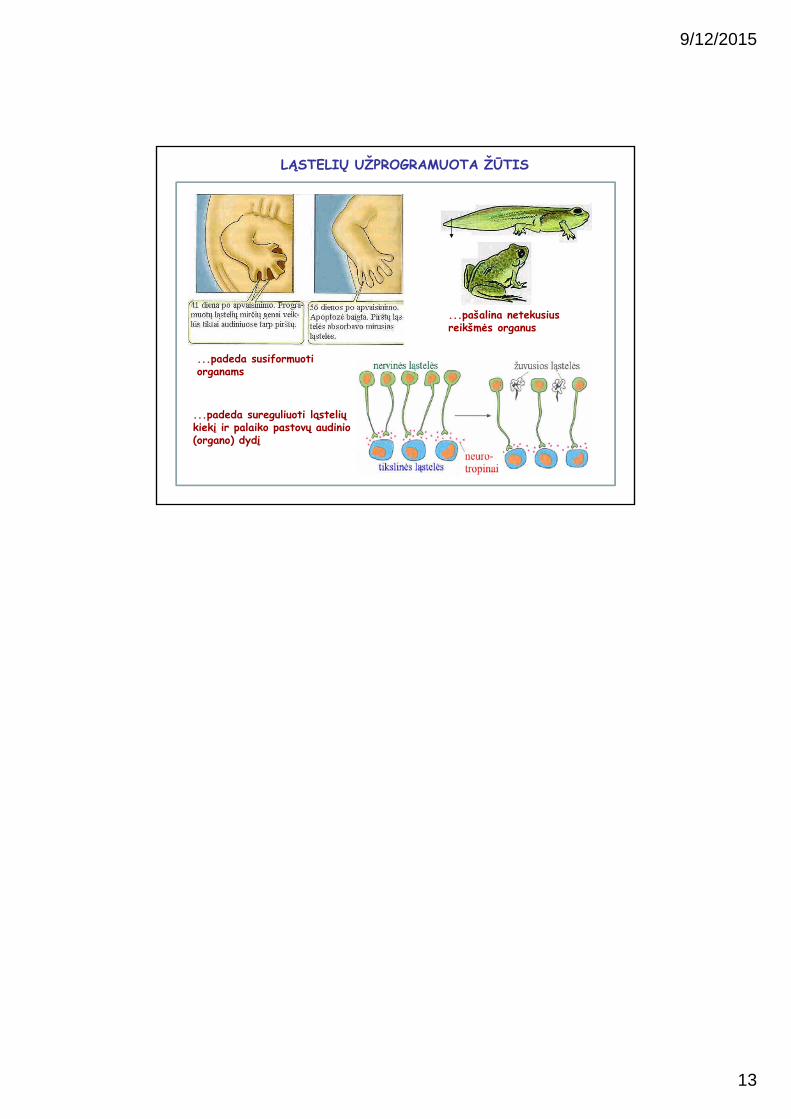
LĄSTELIŲ UŽPROGRAMUOTA ŽŪTIS
...padeda susiformuotiorganams
...pašalina netekusius reikšmės organus
...padeda sureguliuoti ląstelių kiekį ir palaiko pastovų audinio (organo) dydį