Embed Size (px)
Citation preview

1
1
Short title Biosynthesis of 2-phenylethanol in poplar 1
2
Author for contact details Tobias G Koumlllner 3
4
Separate pathways contribute to the herbivore-induced formation of 5
2-phenylethanol in poplar 6
7
Jan Guumlnthera Nathalie D Lackusa Axel Schmidta Meret Huberab Heike-Jana 8
Stoumldtlera Michael Reichelta Jonathan Gershenzona Tobias G Koumlllnera 9
10
11
aMax Planck Institute for Chemical Ecology Hans-Knoumlll-Strasse 8 D-07745 Jena 12
Germany 13
bPresent address Institute of Plant Biology and Biotechnology University of Muumlnster 14
Schlossplatz 7 48143 Muumlnster 15
16
E-mail addresses 17
JaG jguenthericempgde 18
AS aschmidticempgde 19
MH hubermuni-muensterde 20
NDL nlackusicempgde 21
HJS Heike-JanaSgmxde 22
MR reichelticempgde 23
JoG gershenzonicempgde 24
TGK koellnericempgde 25
26
One-sentence summary 27
Biochemical analysis of the aromatic amino acid decarboxylase family in poplar 28
revealed two enzymes involved in the herbivore-induced formation of 2-29
phenylethanol and 2-phenylethylamine 30
31
32
FOOTNOTES 33
Plant Physiology Preview Published on March 7 2019 as DOI101104pp1900059
Copyright 2019 by the American Society of Plant Biologists
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
2
2
Author contributions 34
TGK JaG and JoG designed the research JaG NDL AS HJS and MR carried out 35
the experimental work JaG NDL MH AS MR and TGK analyzed data TGK and 36
JaG wrote the manuscript All authors read and approved the final manuscript 37
38
Responsibilities of the Author for Contact 39
It is the responsibility of the author for contact to ensure that all scientists who have 40
contributed substantially to the conception design or execution of the work described 41
in the manuscript are included as authors in accordance with the guidelines from the 42
Committee on Publication Ethics (COPE 43
httppublicationethicsorgresourcesguidelines) It is the responsibility of the author 44
for contact also to ensure that all authors agree to the list of authors and the 45
identified contributions of those authors 46
47
Funding information 48
The research was funded by the Max-Planck Society 49
50
51
Corresponding author E-mail koellnericempgde phone +49 (0) 3641 57 1329 52
fax +49 (0) 3641 57 1302 53
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
3
3
ABSTRACT 54
Upon herbivory the tree species western balsam poplar (Populus trichocarpa) 55
produces a variety of phenylalanine-derived metabolites including 2-56
phenylethylamine 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside To 57
investigate the formation of these potential defense compounds we functionally 58
characterized aromatic L-amino acid decarboxylases (AADCs) and aromatic 59
aldehyde synthases (AASs) which play important roles in the biosynthesis of 60
specialized aromatic metabolites in other plants Heterologous expression in 61
Escherichia coli and Nicotiana benthamiana showed that all five AADCAAS genes 62
identified in the P trichocarpa genome encode active enzymes However only two 63
genes PtAADC1 and PtAAS1 were significantly upregulated after leaf herbivory 64
Despite a sequence similarity of about 96 PtAADC1 and PtAAS1 showed different 65
enzymatic functions and converted phenylalanine into 2-phenylethylamine and 2-66
phenylacetaldehyde respectively The activities of both enzymes were 67
interconvertible by switching a single amino acid residue in their active sites A 68
survey of putative AADCAAS gene pairs in the genomes of other plants suggests an 69
independent evolution of this function-determining residue in different plant families 70
RNAi-mediated downregulation of AADC1 in grey poplar (P times canescens) resulted in 71
decreased accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-72
glucopyranoside while the emission of 2-phenylethanol was not influenced To 73
investigate the last step of 2-phenylethanol formation we identified and characterized 74
two P trichocarpa short-chain dehydrogenases PtPAR1 and PtPAR2 which were 75
able to reduce 2-phenylacetaldehyde to 2-phenylethanol in vitro In summary 2-76
phenylethanol and its glucoside may be formed in multiple ways in poplar Our data 77
indicate that PtAADC1 controls the herbivore-induced formation of 2-78
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside in planta while PtAAS1 79
likely contributes to the herbivore-induced emission of 2-phenylethanol 80
81
82
Key words Populus trichocarpa 2-phenylethanol 2-phenylethylamine aromatic 83
amino acid decarboxylase aldehyde synthase aldehyde reductase plant 84
metabolism gene duplication neofunctionalization 85
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
4
4
INTRODUCTION 86
Many plant secondary or specialized metabolites play major roles in plant defense 87
against herbivores Some accumulate as deterrents or toxins as part of direct plant 88
defense while others are released as volatiles and can attract parasitoids or 89
predators of the herbivores a phenomenon that has been termed indirect defense 90
(Unsicker et al 2009) Still others may serve as internal signals to activate the 91
formation of both direct and indirect defenses (Maag et al 2015) Plant specialized 92
metabolites are highly diverse and belong to different compound classes including 93
terpenoids phenylpropanoids benzenoids amino acid derivatives and fatty acid 94
derivatives While some of these metabolites as for example certain terpenes and 95
phenylpropanoids are ubiquitous among plants others are exclusively produced in 96
specific families or even a single genus (Fahey et al 2001 Bieri et al 2006) 97
2-Phenylethanol and its glucoside 2-phenylethyl-β-D-glucopyranoside are amino 98
acid-derived specialized compounds that are widespread among plants The free 99
alcohol is well known for its pleasant lsquorose-likersquo aroma and has been reported to be a 100
flower fruit and vegetative volatile released from a multitude of plant species of 101
more than 50 families (Knudsen et al 2006) As a bioactive compound 2-102
phenylethanol plays diverse roles in plant-insect interactions It acts for example as 103
an attractant for pollinators such as butterflies bees and beetles (Roy and Raguso 104
1997 Honda et al 1998 Imai et al 1998) and mediates direct and indirect plant 105
defenses (Zhu et al 2005 Galen et al 2011) 106
The biosynthesis of 2-phenylethanol from phenylalanine was first elucidated in the 107
yeast Saccharomyces cerevisiae where it proceeds via the Ehrlich pathway 108
(reviewed in Hazelwood et al 2008) This pathway starts with the transamination of 109
an aromatic or branched-chain amino acid followed by a decarboxylation of the 110
resulting α-keto acid The aldehyde formed is finally reduced to yield 2-111
phenylethanol In plants the biosynthesis of 2-phenylethanol has been extensively 112
investigated in rose petunia and tomato (Sakai et al 2007 Kaminaga et al 2006 113
Tieman et al 2006 Tieman et al 2007) The pathways described in these species 114
also start from phenylalanine and involve the formation and subsequent reduction of 115
2-phenylacetaldehyde however the initial reactions are different from those of the 116
Ehrlich pathway (Figure 1) Tomato for example contains several aromatic amino 117
acid decarboxylases (AADCs) that catalyze the formation of 2-phenylethylamine 118 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
5
5
which could be further converted to 2-phenylacetaldehyde by a monoamine oxidase 119
(Tieman et al 2006) In contrast rose and petunia possess a bifunctional aromatic 120
aldehyde synthase (AAS) able to catalyze both phenylalanine decarboxylation and 121
the subsequent oxidative transamination (Kaminaga et al 2006 Sakai et al 2007 122
Farhi et al 2010) The recent identification of an aromatic amino acid transaminase 123
(AAAT) involved in aroma compound formation in melon as well as the 124
characterization of an AAAT and a phenylpyruvate decarboxylase (PPDC) in rose 125
indicate that the Ehrlich pathway might also contribute to the formation of 2-126
phenylethanol and other related alcohols in plants (Gonda et al 2010 Hirata et al 127
2016 Sheng et al 2018) Moreover recent research on poplar suggests another 128
potential route to 2-phenylethanol that involves the P450-catalyzed production of 129
(EZ)-phenylacetaldoxime from phenylalanine (Figure 1 Irmisch et al 2013 Irmisch 130
et al 2014a) 131
AADC and AAS both belong to group II pyridoxal-5rsquo-phosphate (PLP)-dependent 132
enzymes that encompass a large family referred to as the plant aromatic amino acid 133
decarboxylase (AAAD) family (Torrens-Spence et al 2018a Facchini et al 2000) 134
AAADs can be assigned to three major groups depending on their substrate 135
specificity for tryptophan tyrosine or phenylalanine respectively They form mid-136
sized gene families with an average number of about 10 members per plant genome 137
and have been identified in all plant lineages including algae non-vascular land 138
plants gymnosperms and angiosperms (Kumar 2016) Due to their relatively high 139
sequence identities a reliable functional prediction of single AAAD members has 140
been difficult in the past However two recent studies identified amino acid residues 141
that determine the substrate selectivity and catalytic activity of AAAD enzymes The 142
different substrate preferences of tryptophan and tyrosine decarboxylases for 143
example are indicated by a conserved glycine-serine polymorphism in the active site 144
of these enzymes (Torrens-Spence et al 2014a) The difference between AADC and 145
AAS activity is determined by the presence of either a tyrosine or phenylalanine (or 146
other apolar residue) on a catalytic loop proximal to the active site (Torrens-Spence 147
et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) 148
The last step in the formation of 2-phenylethanol common to all of these pathways is 149
the reduction of 2-phenylacetaldehyde This reaction is catalyzed by 2-150
phenylacetaldehyde reductase (PAR) which belongs to the large family of NADPH-151 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
6
6
dependent short-chain dehydrogenasereductases (Tieman et al 2007 Chen et al 152
2011) Tomato for example possesses two PARs (LePAR1 and LePAR2) that 153
accept 2-phenylacetaldehyde as substrate While LePAR1 is highly substrate-154
specific LePAR2 can convert a variety of aromatic and aliphatic aldehydes into the 155
respective alcohols (Tieman et al 2007) Such substrate promiscuity has also been 156
reported for another PAR that is involved in 2-phenylethanol formation in roses (Chen 157
et al 2011) 158
In recent years poplar has been established as a model system for studying plant 159
defenses against herbivores in long-living woody plant species (Peters and 160
Constabel 2002 Arimura et al 2004 Ralph et al 2006 Frost et al 2007 Philippe 161
and Bohlmann 2007 Major and Constabel 2008 Danner et al 2011 Irmisch et al 162
2014b) After leaf herbivory the western balsam poplar (P trichocarpa) accumulates 163
various defense compounds and releases a complex volatile blend that mediates 164
direct and indirect defense reactions (Irmisch et al 2013 McCormick et al 2014) 165
The volatile blend is dominated by terpenes green leaf volatiles and nitrogen-166
containing compounds and contains 2-phenylethanol as one of its main components 167
(Irmisch et al 2013) RNAi and labeling experiments indicated that a significant 168
proportion of herbivore-induced 2-phenylethanol seems to be produced via an oxime-169
dependent pathway (Irmisch et al 2013 Irmisch et al 2014a) however whether 170
poplar AAAD enzymes may also contribute to the formation of 2-phenylethanol and 171
related compounds was unclear 172
In order to elucidate the possible contribution of AAAD-dependent pathways to the 173
formation of 2-phenylethanol in poplar we measured the accumulation of pathway 174
intermediates in herbivore-damaged P trichocarpa leaves using liquid 175
chromatography-tandem mass spectrometry (LC-MSMS) RNA-seq experiments and 176
RT-qPCR were used to identify herbivore-induced AAAD candidate genes 177
Characterization of heterologously produced and purified AAAD proteins 178
overexpression in Nicotiana benthamiana and RNAi-mediated downregulation 179
revealed that two candidate enzymes PtAADC1 and PtAAS1 are involved in 180
herbivore-induced phenylalanine metabolism and 2-phenylethanol formation in 181
poplar 182
183
184 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

2
2
Author contributions 34
TGK JaG and JoG designed the research JaG NDL AS HJS and MR carried out 35
the experimental work JaG NDL MH AS MR and TGK analyzed data TGK and 36
JaG wrote the manuscript All authors read and approved the final manuscript 37
38
Responsibilities of the Author for Contact 39
It is the responsibility of the author for contact to ensure that all scientists who have 40
contributed substantially to the conception design or execution of the work described 41
in the manuscript are included as authors in accordance with the guidelines from the 42
Committee on Publication Ethics (COPE 43
httppublicationethicsorgresourcesguidelines) It is the responsibility of the author 44
for contact also to ensure that all authors agree to the list of authors and the 45
identified contributions of those authors 46
47
Funding information 48
The research was funded by the Max-Planck Society 49
50
51
Corresponding author E-mail koellnericempgde phone +49 (0) 3641 57 1329 52
fax +49 (0) 3641 57 1302 53
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
3
3
ABSTRACT 54
Upon herbivory the tree species western balsam poplar (Populus trichocarpa) 55
produces a variety of phenylalanine-derived metabolites including 2-56
phenylethylamine 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside To 57
investigate the formation of these potential defense compounds we functionally 58
characterized aromatic L-amino acid decarboxylases (AADCs) and aromatic 59
aldehyde synthases (AASs) which play important roles in the biosynthesis of 60
specialized aromatic metabolites in other plants Heterologous expression in 61
Escherichia coli and Nicotiana benthamiana showed that all five AADCAAS genes 62
identified in the P trichocarpa genome encode active enzymes However only two 63
genes PtAADC1 and PtAAS1 were significantly upregulated after leaf herbivory 64
Despite a sequence similarity of about 96 PtAADC1 and PtAAS1 showed different 65
enzymatic functions and converted phenylalanine into 2-phenylethylamine and 2-66
phenylacetaldehyde respectively The activities of both enzymes were 67
interconvertible by switching a single amino acid residue in their active sites A 68
survey of putative AADCAAS gene pairs in the genomes of other plants suggests an 69
independent evolution of this function-determining residue in different plant families 70
RNAi-mediated downregulation of AADC1 in grey poplar (P times canescens) resulted in 71
decreased accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-72
glucopyranoside while the emission of 2-phenylethanol was not influenced To 73
investigate the last step of 2-phenylethanol formation we identified and characterized 74
two P trichocarpa short-chain dehydrogenases PtPAR1 and PtPAR2 which were 75
able to reduce 2-phenylacetaldehyde to 2-phenylethanol in vitro In summary 2-76
phenylethanol and its glucoside may be formed in multiple ways in poplar Our data 77
indicate that PtAADC1 controls the herbivore-induced formation of 2-78
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside in planta while PtAAS1 79
likely contributes to the herbivore-induced emission of 2-phenylethanol 80
81
82
Key words Populus trichocarpa 2-phenylethanol 2-phenylethylamine aromatic 83
amino acid decarboxylase aldehyde synthase aldehyde reductase plant 84
metabolism gene duplication neofunctionalization 85
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
4
4
INTRODUCTION 86
Many plant secondary or specialized metabolites play major roles in plant defense 87
against herbivores Some accumulate as deterrents or toxins as part of direct plant 88
defense while others are released as volatiles and can attract parasitoids or 89
predators of the herbivores a phenomenon that has been termed indirect defense 90
(Unsicker et al 2009) Still others may serve as internal signals to activate the 91
formation of both direct and indirect defenses (Maag et al 2015) Plant specialized 92
metabolites are highly diverse and belong to different compound classes including 93
terpenoids phenylpropanoids benzenoids amino acid derivatives and fatty acid 94
derivatives While some of these metabolites as for example certain terpenes and 95
phenylpropanoids are ubiquitous among plants others are exclusively produced in 96
specific families or even a single genus (Fahey et al 2001 Bieri et al 2006) 97
2-Phenylethanol and its glucoside 2-phenylethyl-β-D-glucopyranoside are amino 98
acid-derived specialized compounds that are widespread among plants The free 99
alcohol is well known for its pleasant lsquorose-likersquo aroma and has been reported to be a 100
flower fruit and vegetative volatile released from a multitude of plant species of 101
more than 50 families (Knudsen et al 2006) As a bioactive compound 2-102
phenylethanol plays diverse roles in plant-insect interactions It acts for example as 103
an attractant for pollinators such as butterflies bees and beetles (Roy and Raguso 104
1997 Honda et al 1998 Imai et al 1998) and mediates direct and indirect plant 105
defenses (Zhu et al 2005 Galen et al 2011) 106
The biosynthesis of 2-phenylethanol from phenylalanine was first elucidated in the 107
yeast Saccharomyces cerevisiae where it proceeds via the Ehrlich pathway 108
(reviewed in Hazelwood et al 2008) This pathway starts with the transamination of 109
an aromatic or branched-chain amino acid followed by a decarboxylation of the 110
resulting α-keto acid The aldehyde formed is finally reduced to yield 2-111
phenylethanol In plants the biosynthesis of 2-phenylethanol has been extensively 112
investigated in rose petunia and tomato (Sakai et al 2007 Kaminaga et al 2006 113
Tieman et al 2006 Tieman et al 2007) The pathways described in these species 114
also start from phenylalanine and involve the formation and subsequent reduction of 115
2-phenylacetaldehyde however the initial reactions are different from those of the 116
Ehrlich pathway (Figure 1) Tomato for example contains several aromatic amino 117
acid decarboxylases (AADCs) that catalyze the formation of 2-phenylethylamine 118 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
5
5
which could be further converted to 2-phenylacetaldehyde by a monoamine oxidase 119
(Tieman et al 2006) In contrast rose and petunia possess a bifunctional aromatic 120
aldehyde synthase (AAS) able to catalyze both phenylalanine decarboxylation and 121
the subsequent oxidative transamination (Kaminaga et al 2006 Sakai et al 2007 122
Farhi et al 2010) The recent identification of an aromatic amino acid transaminase 123
(AAAT) involved in aroma compound formation in melon as well as the 124
characterization of an AAAT and a phenylpyruvate decarboxylase (PPDC) in rose 125
indicate that the Ehrlich pathway might also contribute to the formation of 2-126
phenylethanol and other related alcohols in plants (Gonda et al 2010 Hirata et al 127
2016 Sheng et al 2018) Moreover recent research on poplar suggests another 128
potential route to 2-phenylethanol that involves the P450-catalyzed production of 129
(EZ)-phenylacetaldoxime from phenylalanine (Figure 1 Irmisch et al 2013 Irmisch 130
et al 2014a) 131
AADC and AAS both belong to group II pyridoxal-5rsquo-phosphate (PLP)-dependent 132
enzymes that encompass a large family referred to as the plant aromatic amino acid 133
decarboxylase (AAAD) family (Torrens-Spence et al 2018a Facchini et al 2000) 134
AAADs can be assigned to three major groups depending on their substrate 135
specificity for tryptophan tyrosine or phenylalanine respectively They form mid-136
sized gene families with an average number of about 10 members per plant genome 137
and have been identified in all plant lineages including algae non-vascular land 138
plants gymnosperms and angiosperms (Kumar 2016) Due to their relatively high 139
sequence identities a reliable functional prediction of single AAAD members has 140
been difficult in the past However two recent studies identified amino acid residues 141
that determine the substrate selectivity and catalytic activity of AAAD enzymes The 142
different substrate preferences of tryptophan and tyrosine decarboxylases for 143
example are indicated by a conserved glycine-serine polymorphism in the active site 144
of these enzymes (Torrens-Spence et al 2014a) The difference between AADC and 145
AAS activity is determined by the presence of either a tyrosine or phenylalanine (or 146
other apolar residue) on a catalytic loop proximal to the active site (Torrens-Spence 147
et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) 148
The last step in the formation of 2-phenylethanol common to all of these pathways is 149
the reduction of 2-phenylacetaldehyde This reaction is catalyzed by 2-150
phenylacetaldehyde reductase (PAR) which belongs to the large family of NADPH-151 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
6
6
dependent short-chain dehydrogenasereductases (Tieman et al 2007 Chen et al 152
2011) Tomato for example possesses two PARs (LePAR1 and LePAR2) that 153
accept 2-phenylacetaldehyde as substrate While LePAR1 is highly substrate-154
specific LePAR2 can convert a variety of aromatic and aliphatic aldehydes into the 155
respective alcohols (Tieman et al 2007) Such substrate promiscuity has also been 156
reported for another PAR that is involved in 2-phenylethanol formation in roses (Chen 157
et al 2011) 158
In recent years poplar has been established as a model system for studying plant 159
defenses against herbivores in long-living woody plant species (Peters and 160
Constabel 2002 Arimura et al 2004 Ralph et al 2006 Frost et al 2007 Philippe 161
and Bohlmann 2007 Major and Constabel 2008 Danner et al 2011 Irmisch et al 162
2014b) After leaf herbivory the western balsam poplar (P trichocarpa) accumulates 163
various defense compounds and releases a complex volatile blend that mediates 164
direct and indirect defense reactions (Irmisch et al 2013 McCormick et al 2014) 165
The volatile blend is dominated by terpenes green leaf volatiles and nitrogen-166
containing compounds and contains 2-phenylethanol as one of its main components 167
(Irmisch et al 2013) RNAi and labeling experiments indicated that a significant 168
proportion of herbivore-induced 2-phenylethanol seems to be produced via an oxime-169
dependent pathway (Irmisch et al 2013 Irmisch et al 2014a) however whether 170
poplar AAAD enzymes may also contribute to the formation of 2-phenylethanol and 171
related compounds was unclear 172
In order to elucidate the possible contribution of AAAD-dependent pathways to the 173
formation of 2-phenylethanol in poplar we measured the accumulation of pathway 174
intermediates in herbivore-damaged P trichocarpa leaves using liquid 175
chromatography-tandem mass spectrometry (LC-MSMS) RNA-seq experiments and 176
RT-qPCR were used to identify herbivore-induced AAAD candidate genes 177
Characterization of heterologously produced and purified AAAD proteins 178
overexpression in Nicotiana benthamiana and RNAi-mediated downregulation 179
revealed that two candidate enzymes PtAADC1 and PtAAS1 are involved in 180
herbivore-induced phenylalanine metabolism and 2-phenylethanol formation in 181
poplar 182
183
184 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

3
3
ABSTRACT 54
Upon herbivory the tree species western balsam poplar (Populus trichocarpa) 55
produces a variety of phenylalanine-derived metabolites including 2-56
phenylethylamine 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside To 57
investigate the formation of these potential defense compounds we functionally 58
characterized aromatic L-amino acid decarboxylases (AADCs) and aromatic 59
aldehyde synthases (AASs) which play important roles in the biosynthesis of 60
specialized aromatic metabolites in other plants Heterologous expression in 61
Escherichia coli and Nicotiana benthamiana showed that all five AADCAAS genes 62
identified in the P trichocarpa genome encode active enzymes However only two 63
genes PtAADC1 and PtAAS1 were significantly upregulated after leaf herbivory 64
Despite a sequence similarity of about 96 PtAADC1 and PtAAS1 showed different 65
enzymatic functions and converted phenylalanine into 2-phenylethylamine and 2-66
phenylacetaldehyde respectively The activities of both enzymes were 67
interconvertible by switching a single amino acid residue in their active sites A 68
survey of putative AADCAAS gene pairs in the genomes of other plants suggests an 69
independent evolution of this function-determining residue in different plant families 70
RNAi-mediated downregulation of AADC1 in grey poplar (P times canescens) resulted in 71
decreased accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-72
glucopyranoside while the emission of 2-phenylethanol was not influenced To 73
investigate the last step of 2-phenylethanol formation we identified and characterized 74
two P trichocarpa short-chain dehydrogenases PtPAR1 and PtPAR2 which were 75
able to reduce 2-phenylacetaldehyde to 2-phenylethanol in vitro In summary 2-76
phenylethanol and its glucoside may be formed in multiple ways in poplar Our data 77
indicate that PtAADC1 controls the herbivore-induced formation of 2-78
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside in planta while PtAAS1 79
likely contributes to the herbivore-induced emission of 2-phenylethanol 80
81
82
Key words Populus trichocarpa 2-phenylethanol 2-phenylethylamine aromatic 83
amino acid decarboxylase aldehyde synthase aldehyde reductase plant 84
metabolism gene duplication neofunctionalization 85
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
4
4
INTRODUCTION 86
Many plant secondary or specialized metabolites play major roles in plant defense 87
against herbivores Some accumulate as deterrents or toxins as part of direct plant 88
defense while others are released as volatiles and can attract parasitoids or 89
predators of the herbivores a phenomenon that has been termed indirect defense 90
(Unsicker et al 2009) Still others may serve as internal signals to activate the 91
formation of both direct and indirect defenses (Maag et al 2015) Plant specialized 92
metabolites are highly diverse and belong to different compound classes including 93
terpenoids phenylpropanoids benzenoids amino acid derivatives and fatty acid 94
derivatives While some of these metabolites as for example certain terpenes and 95
phenylpropanoids are ubiquitous among plants others are exclusively produced in 96
specific families or even a single genus (Fahey et al 2001 Bieri et al 2006) 97
2-Phenylethanol and its glucoside 2-phenylethyl-β-D-glucopyranoside are amino 98
acid-derived specialized compounds that are widespread among plants The free 99
alcohol is well known for its pleasant lsquorose-likersquo aroma and has been reported to be a 100
flower fruit and vegetative volatile released from a multitude of plant species of 101
more than 50 families (Knudsen et al 2006) As a bioactive compound 2-102
phenylethanol plays diverse roles in plant-insect interactions It acts for example as 103
an attractant for pollinators such as butterflies bees and beetles (Roy and Raguso 104
1997 Honda et al 1998 Imai et al 1998) and mediates direct and indirect plant 105
defenses (Zhu et al 2005 Galen et al 2011) 106
The biosynthesis of 2-phenylethanol from phenylalanine was first elucidated in the 107
yeast Saccharomyces cerevisiae where it proceeds via the Ehrlich pathway 108
(reviewed in Hazelwood et al 2008) This pathway starts with the transamination of 109
an aromatic or branched-chain amino acid followed by a decarboxylation of the 110
resulting α-keto acid The aldehyde formed is finally reduced to yield 2-111
phenylethanol In plants the biosynthesis of 2-phenylethanol has been extensively 112
investigated in rose petunia and tomato (Sakai et al 2007 Kaminaga et al 2006 113
Tieman et al 2006 Tieman et al 2007) The pathways described in these species 114
also start from phenylalanine and involve the formation and subsequent reduction of 115
2-phenylacetaldehyde however the initial reactions are different from those of the 116
Ehrlich pathway (Figure 1) Tomato for example contains several aromatic amino 117
acid decarboxylases (AADCs) that catalyze the formation of 2-phenylethylamine 118 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
5
5
which could be further converted to 2-phenylacetaldehyde by a monoamine oxidase 119
(Tieman et al 2006) In contrast rose and petunia possess a bifunctional aromatic 120
aldehyde synthase (AAS) able to catalyze both phenylalanine decarboxylation and 121
the subsequent oxidative transamination (Kaminaga et al 2006 Sakai et al 2007 122
Farhi et al 2010) The recent identification of an aromatic amino acid transaminase 123
(AAAT) involved in aroma compound formation in melon as well as the 124
characterization of an AAAT and a phenylpyruvate decarboxylase (PPDC) in rose 125
indicate that the Ehrlich pathway might also contribute to the formation of 2-126
phenylethanol and other related alcohols in plants (Gonda et al 2010 Hirata et al 127
2016 Sheng et al 2018) Moreover recent research on poplar suggests another 128
potential route to 2-phenylethanol that involves the P450-catalyzed production of 129
(EZ)-phenylacetaldoxime from phenylalanine (Figure 1 Irmisch et al 2013 Irmisch 130
et al 2014a) 131
AADC and AAS both belong to group II pyridoxal-5rsquo-phosphate (PLP)-dependent 132
enzymes that encompass a large family referred to as the plant aromatic amino acid 133
decarboxylase (AAAD) family (Torrens-Spence et al 2018a Facchini et al 2000) 134
AAADs can be assigned to three major groups depending on their substrate 135
specificity for tryptophan tyrosine or phenylalanine respectively They form mid-136
sized gene families with an average number of about 10 members per plant genome 137
and have been identified in all plant lineages including algae non-vascular land 138
plants gymnosperms and angiosperms (Kumar 2016) Due to their relatively high 139
sequence identities a reliable functional prediction of single AAAD members has 140
been difficult in the past However two recent studies identified amino acid residues 141
that determine the substrate selectivity and catalytic activity of AAAD enzymes The 142
different substrate preferences of tryptophan and tyrosine decarboxylases for 143
example are indicated by a conserved glycine-serine polymorphism in the active site 144
of these enzymes (Torrens-Spence et al 2014a) The difference between AADC and 145
AAS activity is determined by the presence of either a tyrosine or phenylalanine (or 146
other apolar residue) on a catalytic loop proximal to the active site (Torrens-Spence 147
et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) 148
The last step in the formation of 2-phenylethanol common to all of these pathways is 149
the reduction of 2-phenylacetaldehyde This reaction is catalyzed by 2-150
phenylacetaldehyde reductase (PAR) which belongs to the large family of NADPH-151 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
6
6
dependent short-chain dehydrogenasereductases (Tieman et al 2007 Chen et al 152
2011) Tomato for example possesses two PARs (LePAR1 and LePAR2) that 153
accept 2-phenylacetaldehyde as substrate While LePAR1 is highly substrate-154
specific LePAR2 can convert a variety of aromatic and aliphatic aldehydes into the 155
respective alcohols (Tieman et al 2007) Such substrate promiscuity has also been 156
reported for another PAR that is involved in 2-phenylethanol formation in roses (Chen 157
et al 2011) 158
In recent years poplar has been established as a model system for studying plant 159
defenses against herbivores in long-living woody plant species (Peters and 160
Constabel 2002 Arimura et al 2004 Ralph et al 2006 Frost et al 2007 Philippe 161
and Bohlmann 2007 Major and Constabel 2008 Danner et al 2011 Irmisch et al 162
2014b) After leaf herbivory the western balsam poplar (P trichocarpa) accumulates 163
various defense compounds and releases a complex volatile blend that mediates 164
direct and indirect defense reactions (Irmisch et al 2013 McCormick et al 2014) 165
The volatile blend is dominated by terpenes green leaf volatiles and nitrogen-166
containing compounds and contains 2-phenylethanol as one of its main components 167
(Irmisch et al 2013) RNAi and labeling experiments indicated that a significant 168
proportion of herbivore-induced 2-phenylethanol seems to be produced via an oxime-169
dependent pathway (Irmisch et al 2013 Irmisch et al 2014a) however whether 170
poplar AAAD enzymes may also contribute to the formation of 2-phenylethanol and 171
related compounds was unclear 172
In order to elucidate the possible contribution of AAAD-dependent pathways to the 173
formation of 2-phenylethanol in poplar we measured the accumulation of pathway 174
intermediates in herbivore-damaged P trichocarpa leaves using liquid 175
chromatography-tandem mass spectrometry (LC-MSMS) RNA-seq experiments and 176
RT-qPCR were used to identify herbivore-induced AAAD candidate genes 177
Characterization of heterologously produced and purified AAAD proteins 178
overexpression in Nicotiana benthamiana and RNAi-mediated downregulation 179
revealed that two candidate enzymes PtAADC1 and PtAAS1 are involved in 180
herbivore-induced phenylalanine metabolism and 2-phenylethanol formation in 181
poplar 182
183
184 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

4
4
INTRODUCTION 86
Many plant secondary or specialized metabolites play major roles in plant defense 87
against herbivores Some accumulate as deterrents or toxins as part of direct plant 88
defense while others are released as volatiles and can attract parasitoids or 89
predators of the herbivores a phenomenon that has been termed indirect defense 90
(Unsicker et al 2009) Still others may serve as internal signals to activate the 91
formation of both direct and indirect defenses (Maag et al 2015) Plant specialized 92
metabolites are highly diverse and belong to different compound classes including 93
terpenoids phenylpropanoids benzenoids amino acid derivatives and fatty acid 94
derivatives While some of these metabolites as for example certain terpenes and 95
phenylpropanoids are ubiquitous among plants others are exclusively produced in 96
specific families or even a single genus (Fahey et al 2001 Bieri et al 2006) 97
2-Phenylethanol and its glucoside 2-phenylethyl-β-D-glucopyranoside are amino 98
acid-derived specialized compounds that are widespread among plants The free 99
alcohol is well known for its pleasant lsquorose-likersquo aroma and has been reported to be a 100
flower fruit and vegetative volatile released from a multitude of plant species of 101
more than 50 families (Knudsen et al 2006) As a bioactive compound 2-102
phenylethanol plays diverse roles in plant-insect interactions It acts for example as 103
an attractant for pollinators such as butterflies bees and beetles (Roy and Raguso 104
1997 Honda et al 1998 Imai et al 1998) and mediates direct and indirect plant 105
defenses (Zhu et al 2005 Galen et al 2011) 106
The biosynthesis of 2-phenylethanol from phenylalanine was first elucidated in the 107
yeast Saccharomyces cerevisiae where it proceeds via the Ehrlich pathway 108
(reviewed in Hazelwood et al 2008) This pathway starts with the transamination of 109
an aromatic or branched-chain amino acid followed by a decarboxylation of the 110
resulting α-keto acid The aldehyde formed is finally reduced to yield 2-111
phenylethanol In plants the biosynthesis of 2-phenylethanol has been extensively 112
investigated in rose petunia and tomato (Sakai et al 2007 Kaminaga et al 2006 113
Tieman et al 2006 Tieman et al 2007) The pathways described in these species 114
also start from phenylalanine and involve the formation and subsequent reduction of 115
2-phenylacetaldehyde however the initial reactions are different from those of the 116
Ehrlich pathway (Figure 1) Tomato for example contains several aromatic amino 117
acid decarboxylases (AADCs) that catalyze the formation of 2-phenylethylamine 118 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
5
5
which could be further converted to 2-phenylacetaldehyde by a monoamine oxidase 119
(Tieman et al 2006) In contrast rose and petunia possess a bifunctional aromatic 120
aldehyde synthase (AAS) able to catalyze both phenylalanine decarboxylation and 121
the subsequent oxidative transamination (Kaminaga et al 2006 Sakai et al 2007 122
Farhi et al 2010) The recent identification of an aromatic amino acid transaminase 123
(AAAT) involved in aroma compound formation in melon as well as the 124
characterization of an AAAT and a phenylpyruvate decarboxylase (PPDC) in rose 125
indicate that the Ehrlich pathway might also contribute to the formation of 2-126
phenylethanol and other related alcohols in plants (Gonda et al 2010 Hirata et al 127
2016 Sheng et al 2018) Moreover recent research on poplar suggests another 128
potential route to 2-phenylethanol that involves the P450-catalyzed production of 129
(EZ)-phenylacetaldoxime from phenylalanine (Figure 1 Irmisch et al 2013 Irmisch 130
et al 2014a) 131
AADC and AAS both belong to group II pyridoxal-5rsquo-phosphate (PLP)-dependent 132
enzymes that encompass a large family referred to as the plant aromatic amino acid 133
decarboxylase (AAAD) family (Torrens-Spence et al 2018a Facchini et al 2000) 134
AAADs can be assigned to three major groups depending on their substrate 135
specificity for tryptophan tyrosine or phenylalanine respectively They form mid-136
sized gene families with an average number of about 10 members per plant genome 137
and have been identified in all plant lineages including algae non-vascular land 138
plants gymnosperms and angiosperms (Kumar 2016) Due to their relatively high 139
sequence identities a reliable functional prediction of single AAAD members has 140
been difficult in the past However two recent studies identified amino acid residues 141
that determine the substrate selectivity and catalytic activity of AAAD enzymes The 142
different substrate preferences of tryptophan and tyrosine decarboxylases for 143
example are indicated by a conserved glycine-serine polymorphism in the active site 144
of these enzymes (Torrens-Spence et al 2014a) The difference between AADC and 145
AAS activity is determined by the presence of either a tyrosine or phenylalanine (or 146
other apolar residue) on a catalytic loop proximal to the active site (Torrens-Spence 147
et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) 148
The last step in the formation of 2-phenylethanol common to all of these pathways is 149
the reduction of 2-phenylacetaldehyde This reaction is catalyzed by 2-150
phenylacetaldehyde reductase (PAR) which belongs to the large family of NADPH-151 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
6
6
dependent short-chain dehydrogenasereductases (Tieman et al 2007 Chen et al 152
2011) Tomato for example possesses two PARs (LePAR1 and LePAR2) that 153
accept 2-phenylacetaldehyde as substrate While LePAR1 is highly substrate-154
specific LePAR2 can convert a variety of aromatic and aliphatic aldehydes into the 155
respective alcohols (Tieman et al 2007) Such substrate promiscuity has also been 156
reported for another PAR that is involved in 2-phenylethanol formation in roses (Chen 157
et al 2011) 158
In recent years poplar has been established as a model system for studying plant 159
defenses against herbivores in long-living woody plant species (Peters and 160
Constabel 2002 Arimura et al 2004 Ralph et al 2006 Frost et al 2007 Philippe 161
and Bohlmann 2007 Major and Constabel 2008 Danner et al 2011 Irmisch et al 162
2014b) After leaf herbivory the western balsam poplar (P trichocarpa) accumulates 163
various defense compounds and releases a complex volatile blend that mediates 164
direct and indirect defense reactions (Irmisch et al 2013 McCormick et al 2014) 165
The volatile blend is dominated by terpenes green leaf volatiles and nitrogen-166
containing compounds and contains 2-phenylethanol as one of its main components 167
(Irmisch et al 2013) RNAi and labeling experiments indicated that a significant 168
proportion of herbivore-induced 2-phenylethanol seems to be produced via an oxime-169
dependent pathway (Irmisch et al 2013 Irmisch et al 2014a) however whether 170
poplar AAAD enzymes may also contribute to the formation of 2-phenylethanol and 171
related compounds was unclear 172
In order to elucidate the possible contribution of AAAD-dependent pathways to the 173
formation of 2-phenylethanol in poplar we measured the accumulation of pathway 174
intermediates in herbivore-damaged P trichocarpa leaves using liquid 175
chromatography-tandem mass spectrometry (LC-MSMS) RNA-seq experiments and 176
RT-qPCR were used to identify herbivore-induced AAAD candidate genes 177
Characterization of heterologously produced and purified AAAD proteins 178
overexpression in Nicotiana benthamiana and RNAi-mediated downregulation 179
revealed that two candidate enzymes PtAADC1 and PtAAS1 are involved in 180
herbivore-induced phenylalanine metabolism and 2-phenylethanol formation in 181
poplar 182
183
184 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

5
5
which could be further converted to 2-phenylacetaldehyde by a monoamine oxidase 119
(Tieman et al 2006) In contrast rose and petunia possess a bifunctional aromatic 120
aldehyde synthase (AAS) able to catalyze both phenylalanine decarboxylation and 121
the subsequent oxidative transamination (Kaminaga et al 2006 Sakai et al 2007 122
Farhi et al 2010) The recent identification of an aromatic amino acid transaminase 123
(AAAT) involved in aroma compound formation in melon as well as the 124
characterization of an AAAT and a phenylpyruvate decarboxylase (PPDC) in rose 125
indicate that the Ehrlich pathway might also contribute to the formation of 2-126
phenylethanol and other related alcohols in plants (Gonda et al 2010 Hirata et al 127
2016 Sheng et al 2018) Moreover recent research on poplar suggests another 128
potential route to 2-phenylethanol that involves the P450-catalyzed production of 129
(EZ)-phenylacetaldoxime from phenylalanine (Figure 1 Irmisch et al 2013 Irmisch 130
et al 2014a) 131
AADC and AAS both belong to group II pyridoxal-5rsquo-phosphate (PLP)-dependent 132
enzymes that encompass a large family referred to as the plant aromatic amino acid 133
decarboxylase (AAAD) family (Torrens-Spence et al 2018a Facchini et al 2000) 134
AAADs can be assigned to three major groups depending on their substrate 135
specificity for tryptophan tyrosine or phenylalanine respectively They form mid-136
sized gene families with an average number of about 10 members per plant genome 137
and have been identified in all plant lineages including algae non-vascular land 138
plants gymnosperms and angiosperms (Kumar 2016) Due to their relatively high 139
sequence identities a reliable functional prediction of single AAAD members has 140
been difficult in the past However two recent studies identified amino acid residues 141
that determine the substrate selectivity and catalytic activity of AAAD enzymes The 142
different substrate preferences of tryptophan and tyrosine decarboxylases for 143
example are indicated by a conserved glycine-serine polymorphism in the active site 144
of these enzymes (Torrens-Spence et al 2014a) The difference between AADC and 145
AAS activity is determined by the presence of either a tyrosine or phenylalanine (or 146
other apolar residue) on a catalytic loop proximal to the active site (Torrens-Spence 147
et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) 148
The last step in the formation of 2-phenylethanol common to all of these pathways is 149
the reduction of 2-phenylacetaldehyde This reaction is catalyzed by 2-150
phenylacetaldehyde reductase (PAR) which belongs to the large family of NADPH-151 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
6
6
dependent short-chain dehydrogenasereductases (Tieman et al 2007 Chen et al 152
2011) Tomato for example possesses two PARs (LePAR1 and LePAR2) that 153
accept 2-phenylacetaldehyde as substrate While LePAR1 is highly substrate-154
specific LePAR2 can convert a variety of aromatic and aliphatic aldehydes into the 155
respective alcohols (Tieman et al 2007) Such substrate promiscuity has also been 156
reported for another PAR that is involved in 2-phenylethanol formation in roses (Chen 157
et al 2011) 158
In recent years poplar has been established as a model system for studying plant 159
defenses against herbivores in long-living woody plant species (Peters and 160
Constabel 2002 Arimura et al 2004 Ralph et al 2006 Frost et al 2007 Philippe 161
and Bohlmann 2007 Major and Constabel 2008 Danner et al 2011 Irmisch et al 162
2014b) After leaf herbivory the western balsam poplar (P trichocarpa) accumulates 163
various defense compounds and releases a complex volatile blend that mediates 164
direct and indirect defense reactions (Irmisch et al 2013 McCormick et al 2014) 165
The volatile blend is dominated by terpenes green leaf volatiles and nitrogen-166
containing compounds and contains 2-phenylethanol as one of its main components 167
(Irmisch et al 2013) RNAi and labeling experiments indicated that a significant 168
proportion of herbivore-induced 2-phenylethanol seems to be produced via an oxime-169
dependent pathway (Irmisch et al 2013 Irmisch et al 2014a) however whether 170
poplar AAAD enzymes may also contribute to the formation of 2-phenylethanol and 171
related compounds was unclear 172
In order to elucidate the possible contribution of AAAD-dependent pathways to the 173
formation of 2-phenylethanol in poplar we measured the accumulation of pathway 174
intermediates in herbivore-damaged P trichocarpa leaves using liquid 175
chromatography-tandem mass spectrometry (LC-MSMS) RNA-seq experiments and 176
RT-qPCR were used to identify herbivore-induced AAAD candidate genes 177
Characterization of heterologously produced and purified AAAD proteins 178
overexpression in Nicotiana benthamiana and RNAi-mediated downregulation 179
revealed that two candidate enzymes PtAADC1 and PtAAS1 are involved in 180
herbivore-induced phenylalanine metabolism and 2-phenylethanol formation in 181
poplar 182
183
184 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

6
6
dependent short-chain dehydrogenasereductases (Tieman et al 2007 Chen et al 152
2011) Tomato for example possesses two PARs (LePAR1 and LePAR2) that 153
accept 2-phenylacetaldehyde as substrate While LePAR1 is highly substrate-154
specific LePAR2 can convert a variety of aromatic and aliphatic aldehydes into the 155
respective alcohols (Tieman et al 2007) Such substrate promiscuity has also been 156
reported for another PAR that is involved in 2-phenylethanol formation in roses (Chen 157
et al 2011) 158
In recent years poplar has been established as a model system for studying plant 159
defenses against herbivores in long-living woody plant species (Peters and 160
Constabel 2002 Arimura et al 2004 Ralph et al 2006 Frost et al 2007 Philippe 161
and Bohlmann 2007 Major and Constabel 2008 Danner et al 2011 Irmisch et al 162
2014b) After leaf herbivory the western balsam poplar (P trichocarpa) accumulates 163
various defense compounds and releases a complex volatile blend that mediates 164
direct and indirect defense reactions (Irmisch et al 2013 McCormick et al 2014) 165
The volatile blend is dominated by terpenes green leaf volatiles and nitrogen-166
containing compounds and contains 2-phenylethanol as one of its main components 167
(Irmisch et al 2013) RNAi and labeling experiments indicated that a significant 168
proportion of herbivore-induced 2-phenylethanol seems to be produced via an oxime-169
dependent pathway (Irmisch et al 2013 Irmisch et al 2014a) however whether 170
poplar AAAD enzymes may also contribute to the formation of 2-phenylethanol and 171
related compounds was unclear 172
In order to elucidate the possible contribution of AAAD-dependent pathways to the 173
formation of 2-phenylethanol in poplar we measured the accumulation of pathway 174
intermediates in herbivore-damaged P trichocarpa leaves using liquid 175
chromatography-tandem mass spectrometry (LC-MSMS) RNA-seq experiments and 176
RT-qPCR were used to identify herbivore-induced AAAD candidate genes 177
Characterization of heterologously produced and purified AAAD proteins 178
overexpression in Nicotiana benthamiana and RNAi-mediated downregulation 179
revealed that two candidate enzymes PtAADC1 and PtAAS1 are involved in 180
herbivore-induced phenylalanine metabolism and 2-phenylethanol formation in 181
poplar 182
183
184 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

7
7
185
RESULTS 186
Potential precursors and metabolites of 2-phenylethanol accumulate in 187
herbivore-damaged P trichocarpa leaves 188
In order to elucidate the metabolic processes leading to 2-phenylethanol formation in 189
poplar we measured potential precursors and pathway intermediates in undamaged 190
and gypsy moth (Lymantria dispar) caterpillar-damaged P trichocarpa leaves (Figure 191
2) Phenylalanine and phenylpyruvic acid were present in both undamaged and 192
herbivore-damaged leaves and showed no changes in response to herbivory (Figure 193
2AB) The accumulation of 2-phenylethylamine however was significantly increased 194
upon herbivory and mirrored the herbivore-induced emission of 2-phenylethanol 195
(Figure 2CE) 2-Phenylacetaldehyde could not be detected suggesting a rapid 196
reduction to 2-phenylethanol in poplar 197
It has been shown that 2-phenylethanol can be converted to 2-phenylethyl-β-D-198
glucopyranoside in roses (Hayashi et al 2004) Moreover transgenic poplars 199
overexpressing rose RhPAAS and PAR accumulated large amounts of this glucoside 200
(Costa et al 2013) However whether 2-phenylethyl-β-D-glucopyranoside also 201
occurs in wild type poplars was unclear We could detect 2-phenylethyl-β-D-202
glucopyranoside both in undamaged and herbivore-damaged leaves and the 203
accumulation of this compound was slightly but significantly induced upon herbivory 204
(Figure 2D) 205
206
Identification of putative aromatic amino acid decarboxylase (AAAD) family 207
genes in P trichocarpa 208
A BLAST analysis with RhPAAS (Farhi et al 2010) as query revealed five putative 209
AAAD genes in the genome of P trichocarpa (Tuskan et al 2006) (Figure 3) 210
Considering the function-determining tyrosine-phenylalanine polymorphism in the 211
active sites of their encoded proteins the genes were named PtAADC1 212
(Potri013G052800) PtAADC2 (Potri016G114300) PtAADC3 (Potri004G036200) 213
PtAAS1 (Potri013G052900) and PtAAS2 (Potri002G255600) (Supplemental Figure 214
1) Two genes PtSDC1 (Potri005G190500) and PtSDC2 (Potri002G069800) that 215
also appeared in the BLAST search were similar to histidine (HDC) and serine (SDC) 216
decarboxylase genes (Figure 3) However since SDC-like proteins from barrelclover 217 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

8
8
(Medicago truncatula) and chickpea (Cicer arietinum) have recently been 218
characterized as AAS enzymes (Torrens-Spence et al 2014a) poplar PtSDC1 and 219
PtSDC2 were also chosen as potential AAS gene candidates for further 220
characterization 221
222
Biochemical characterization of poplar AAAD and serine decarboxylase (SDC) 223
enzymes 224
The complete open reading frames of the candidate genes were amplified from 225
cDNA made from herbivore-damaged leaves of P trichocarpa and cloned into the 226
vector pET100D-TOPO After heterologous expression in Escherichia coli 227
recombinant proteins carrying an N-terminal His6-Tag were affinity purified and 228
incubated with phenylalanine tyrosine or tryptophan as potential substrates and PLP 229
as cofactor Both PtAAS1 and PtAAS2 had aldehyde synthase activity and catalyzed 230
the conversion of phenylalanine to 2-phenylacetaldehyde (Table 1) Formation of 2-231
phenylethylamine however could not be detected (Supplemental Figure 2) While 232
PtAAS1 was highly substrate specific and exclusively accepted phenylalanine 233
PtAAS2 showed activity with all tested aromatic amino acids producing the 234
respective aldehydes (Supplemental Figure 3) Notably the amino acid residue that 235
has been reported to determine the substrate specificity of tyrosine and tryptophan 236
decarboxylases (Torrens-Spence et al 2014b) differed between PtAAS1 (serine 237
360) and PtAAS2 (glycine 358) (Supplemental Figure 1) This amino acid change 238
might explain the observed differences in substrate promiscuity In contrast to 239
PtAAS1 and PtAAS2 PtAADC1 PtAADC2 and PtAADC3 showed no aldehyde 240
synthase activity but were able to catalyze the decarboxylation of different aromatic 241
amino acids The Km and kcat values revealed that PtAADC1 and PtAADC2 most 242
likely act as phenylalanine decarboxylases in planta while PtAADC3 is likely a 243
tyrosine decarboxylase (Table 1) To test for potential aromatic amino acid 244
transaminase activity of poplar AAAD candidates we incubated the enzymes with 245
phenylalanine in the presence of the ammonia acceptor α-ketoglutarate However no 246
formation of phenylpyruvic acid could be observed 247
To determine if the putative serine decarboxylases PtSDC1 and PtSDC2 exhibit 248
decarboxylase activity towards aromatic amino acids as previously described for 249
SDC-like enzymes in M truncatula and C arietinum (Torrens-Spence et al 2014a) 250 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

9
9
the enzymes were heterologously expressed and tested with phenylalanine tyrosine 251
tryptophan histidine and serine as potential substrates Both enzymes converted 252
serine into ethanolamine but showed no activity with aromatic amino acids and 253
histidine (Supplemental Figure 4) indicating that PtSDC1 and PtSDC2 likely function 254
as SDCs in planta Thus they were not further considered in this study 255
256
PtAAS1 and PtAADC1 are interconvertible by the change of one amino acid 257
PtAAS1 and PtAADC1 differ in a characteristic amino acid residue that has been 258
described as function-dictating residue in AADC and AAS enzymes from other plants 259
(Torrens-Spence et al 2013 Bertoldi et al 2002 Torrens-Spence et al 2018b) To 260
demonstrate that this tyrosine-phenylalanine polymorphism also determines the 261
decarboxylase and aldehyde synthase activity of PtAADC1 and PtAAS1 262
respectively we constructed the mutant enzymes PtAADC1 (Y338F) and PtAAS1 263
(F338Y) by in vitro mutagenesis and incubated them with phenylalanine as substrate 264
As expected both mutations led to a complete interconversion of the wild type 265
activities While PtAADC1 (Y338F) produced exclusively 2-phenylacetaldehyde 266
PtAAS1 (F338Y) had only decarboxylase activity and produced 2-phenylethylamine 267
(Figure 4A) 268
269
The tyrosine-phenylalanine-determined shift in AADCAAS enzyme activity 270
evolved independently in different plant families 271
Within the poplar AADCAAS clade PtAAS1 PtAADC1 and PtAADC2 form a small 272
subfamily with high nucleotide sequence identity of about 96-98 (Figure 3) While 273
PtAADC2 was found to be located on chromosome 16 PtAADC1 and PtAAS1 were 274
located close to each other on chromosome 13 (Figure 4B) This and the fact that 275
PtAADC1 and PtAAS1 have no other open reading frames between them in the 276
genomic sequence suggest a recent gene duplication that was subsequently followed 277
by neofunctionalization through the mutation of the function-determining amino acid 278
A survey of putative AADCAAS sequences in other available plant genomes 279
revealed the presence of comparable AADCAAS pairs in at least ten species from 280
diverse plant families including monocotyledonous and dicotyledonous plants 281
(Supplemental Figures 5 and 6) The putative AADC and AAS genes in each of these 282
pairs were highly similar to each other and grouped in a species-specific manner 283 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

10
10
indicating an independent evolution of the tyrosine-phenylalanine shift in different 284
plant families or even genera 285
286
Functional analysis of PtAADC1 and PtAAS1 in planta 287
To verify the different enzymatic functions of the characterized poplar phenylalanine-288
using AAAD enzymes in a heterologous plant system PtAADC1 and PtAAS1 were 289
chosen for transient expression in N benthamiana Plants overexpressing PtAAS1 290
showed a significant emission of 2-phenylethanol while wild type plants and plants 291
expressing either eGFP or PtAADC1 produced only trace amounts of this compound 292
if any (Figure 5A) In addition PtAAS1 plants accumulated 2-phenylethyl-β-D-293
glucopyranoside (Figure 5E) The overexpression of PtAADC1 however resulted in 294
the accumulation of 2-phenylethylamine tyramine tryptamine and 2-phenylethyl-β-295
D-glucopyranoside (Figure 5B-E) 2-Phenylacetaldehyde 4-hydroxy-296
phenylacetaldehyde and indole-3-acetaldehyde could not be detected in the tested 297
lines Altogether the exclusive emission of 2-phenylethanol from PtAAS1 plants and 298
the different accumulation levels of the aromatic amines in PtAADC1-overexpressing 299
plants corresponded well to the enzymatic activities and kinetic parameters that were 300
determined in vitro (Table 1 Figure 5) Moreover the results suggest that the 301
reactive aldehydes can be rapidly converted to less reactive alcohols or other 302
compounds in vivo even in a heterologous plant system 303
304
Expression of PtAADC1 and PtAAS1 is induced in response to herbivory in P 305
trichocarpa leaves 306
In order to study the expression of poplar AAAD genes we sequenced the 307
transcriptomes of herbivore-damaged and undamaged P trichocarpa leaves and 308
mapped the quality-trimmed sequence reads to the P trichocarpa genome version 309
v30 (httpsphytozomejgidoegov Tuskan et al 2006) All AAAD genes were 310
found to be expressed in poplar leaves (Figure 6) However the transcript 311
accumulation of PtAADC1 and PtAAS1 was highly upregulated in herbivore-312
damaged leaves in comparison to undamaged control leaves with fold changes of 313
294 and 124 respectively (Figure 6) Expression of PtAADC2 and PtSDC2 was also 314
increased in response to the herbivore treatment but showed lower fold increases 315
(64 and 16 respectively) PtAAS2 showed a significant reduction in gene 316 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

11
11
expression upon herbivory (fold change -15) while expression of PtAADC3 and 317
PtSDC1 were not influenced by the treatment (Figure 6) The RNA-Seq results for 318
poplar AAAD genes were confirmed by RT-qPCR (Supplemental Figure 7) The 319
highly similar genes PtAAS1 PtAADC1 and PtAADC2 could not be targeted by 320
specific primers and were thus amplified using a universal primer pair Extensive 321
sequencing of cloned amplicons that encompassed several polymorphisms including 322
the function-determining codon allowed a reliable assignment of the amplicons to 323
specific genes The RT-qPCR results confirmed the upregulation of PtAAS1 and 324
PtAADC1 gene expression in response to herbivory (Supplemental Table 2) 325
Transcripts of PtAADC2 however could not be detected in control and herbivore-326
damaged leaves Notably total expression of PtAAS1 in herbivore-damaged leaves 327
(925 of sequenced amplicons) was over ten times higher than PtAADC1 (75 of 328
sequenced amplicons) 329
330
RNAi-mediated downregulation of PtAADC1 in P times canescens affects the 331
formation of 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 332
To investigate the function of herbivore-induced AAAD genes in poplar we aimed to 333
generate RNAi lines with reduced AADC1AAS1 gene expression by transformation 334
of a DNA fragment complementary to AADC1 AADC2 and AAS1 Since there is no 335
established transformation method for P trichocarpa we chose grey poplar (P times 336
canescens) for transformation a species commonly used for the generation of 337
transgenic poplar trees (Lepleacute et al 1992) Grey poplar is a hybrid species derived 338
from a cross between silver poplar (P alba) and quaking aspen (P tremula) and 339
BLAST searches with the different P trichocarpa AAAD genes as query and the 340
genomes of P times canescens P alba and P tremula 341
(httpaspendbugaeduindexphpdatabasesspta-717-genome Zhou et al 2015 342
Xue et al 2015) as templates revealed that all three species possess putative 343
AADC1 AADC2 and AAS1 orthologues (Supplemental Figure 8) Interestingly the 344
function-dictating tyrosine of P trichocarpa PtAADC2 was changed into 345
phenylalanine in P times canescens PcanAADC2 and in P tremula PtremAADC2 346
suggesting that both orthologues likely function as aldehyde synthases However 347
sequencing of cloned qPCR fragments that were amplified with a primer pair specific 348
for AADC1 AADC2 and AAS1 from cDNA made from herbivore-damaged P times 349 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

12
12
canescens leaves revealed exclusively PcanAADC1 amplicons (Supplemental Figure 350
9) This indicates that the two aldehyde synthase genes PcanAAS1 and PcanAADC2 351
are likely not expressed in herbivore-damaged leaves of P times canescens 352
Populus times canescens PcanAADC1 RNAi lines as well as wild type trees and trees 353
carrying an empty vector were subjected to L dispar herbivory and the emission of 2-354
phenylethanol and the accumulation of 2-phenylethylamine and 2-phenylethyl-β-D-355
glucopyranoside in herbivory-induced leaves were measured Downregulation of 356
PcanAADC1 in the transgenic lines led to a significantly reduced accumulation of 357
herbivore-induced 2-phenylethylamine and 2-phenylethyl-β-D-glucopyranoside 358
(Figure 7A Supplemental Figure 10) The emission of 2-phenylethanol however 359
remained constant in comparison to wild type and empty vector trees Expression 360
levels of PcanAAS2 were not affected by the RNAi approach (Supplemental Figure 361
10) Thus the observed metabolic changes in the transgenic lines are most likely 362
caused by the downregulation of PcanAADC1 gene expression 363
One of the most characteristic volatiles released from herbivore-damaged poplars is 364
the nitrile benzyl cyanide (McCormick et al 2014) Previous studies showed that this 365
compound is mainly formed from phenylalanine via (EZ)-phenylacetaldoxime 366
through the action of two P450s from the CYP79 and CYP71 families (Irmisch et al 367
2013 Irmisch et al 2014a) Since 2-phenylethylamine has been proposed as 368
alternative substrate for benzyl cyanide (Tieman et al 2006) we measured the 369
emission of the volatile nitrile in herbivore-damaged PcanAADC1 RNAi lines 370
Downregulation of PcanAADC1 expression and the resulting decrease in 2-371
phenylethylamine accumulation did not influence benzyl cyanide emission 372
(Supplemental Figure 11) suggesting that this nitrile is exclusively produced via the 373
oxime-dependent pathway in poplar 374
375
Poplar aldehyde reductases PtPAR1 and PtPAR2 convert 2-376
phenylacetaldehyde into 2-phenylethanol in vitro 377
The findings that 2-phenylacetaldehyde could neither be detected in herbivore-378
damaged poplar leaves nor in PtAAS1-overexpressing N benthamiana plants 379
suggest a rapid and highly efficient conversion of the aldehyde into the 380
corresponding alcohol in planta likely catalyzed by a NADPH-dependent reductase 381
(PAR) as already reported in rose and tomato (Tieman et al 2007 Chen et al 382 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

13
13
2011) To identify putative poplar PARs we performed a TBLASTN analysis with the 383
tomato gene LePAR1 (Tieman et al 2007) as query and the P trichocarpa genome 384
as template A dendrogram of putative poplar reductases and characterized 385
reductases from other plants was generated and the five poplar genes that grouped 386
together with either LePAR1 (PtPAR1 PtPAR2) or aliphatic aldehyde reductases 387
(PtAAR1 PtAAR2 PtAAR3) were considered for further biochemical characterization 388
(Supplemental Figure 12) Heterologously expressed and purified enzymes were 389
tested with a variety of aromatic and aliphatic aldehydes as potential substrates in 390
the presence of the cosubstrate NADPH PtPAR1 and PtPAR2 exhibited enzyme 391
activity with 2-phenylacetaldehyde and produced 2-phenylethanol (Table 2 392
Supplemental Figure 13) However they were also able to reduce other aromatic and 393
aliphatic aldehyde substrates In contrast PtAAR1 PtAAR2 and PtAAR3 were only 394
active with a few aliphatic aldehydes and produced no 2-phenylethanol (Table 2 395
Supplemental Figure 13) Gene expression analysis revealed that all five aldehyde 396
reductase genes were constitutively expressed in P trichocarpa leaves 397
(Supplemental Figure 14) Their transcript accumulation was not influenced by the 398
herbivore treatment 399
400
401
DISCUSSION 402
In response to herbivore feeding poplar produces several aromatic metabolites 403
including 2-phenylethanol 2-phenylethyl-β-D-glucopyranoside and 2-404
phenylethylamine that may function in direct and indirect plant defense and defense 405
signaling Aromatic amino acid decarboxylases (AADC) and aromatic aldehyde 406
synthases (AAS) have been shown to be involved in the biosynthesis of 407
phenylalanine-derived specialized compounds in other plants (Kaminaga et al 2006 408
Tieman et al 2007 Hirata et al 2012) We aimed to study AADCAAS enzymes in 409
P trichocarpa to elucidate their roles in the herbivore-induced metabolism of 410
phenylalanine 411
412
PtAAS1 likely contributes to the formation of herbivore-induced 2-413
phenylethanol in P trichocarpa 414
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

14
14
Aromatic aldehyde synthases are bifunctional enzymes that catalyze the 415
decarboxylation and subsequent oxidative deamination of aromatic amino acids In 416
roses and petunia AAS enzymes have been shown to produce 2-417
phenylacetaldehyde as substrate for the formation of the corresponding alcohol 2-418
phenylethanol (Kaminaga et al 2006 Sakai et al 2007 Farhi et al 2010) P 419
trichocarpa contains two putative AAS (Figure 3) and biochemical characterization of 420
the recombinant enzymes confirmed their aldehyde synthase activity (Table 1) While 421
PtAAS1 accepted exclusively phenylalanine as a substrate PtAAS2 showed activity 422
with phenylalanine tyrosine and tryptophan However the similar Km values of 423
PtAAS1 and PtAAS2 for phenylalanine suggest that both enzymes catalyse the 424
formation of 2-phenylacetaldehyde in vivo (Table 1) In contrast to PtAAS2 which 425
was constitutively expressed transcript accumulation of PtAAS1 was highly 426
upregulated in herbivore-damaged leaves of P trichocarpa and mirrored the 427
herbivore-induced emission of 2-phenylethanol (Figures 2 and 6) This indicates that 428
PtAAS1 likely contributes to the formation of 2-phenylethanol upon herbivory by 429
providing the precursor 2-phenylacetaldehyde 430
Recent studies demonstrated that the herbivore-induced production of 2-431
phenylethanol in poplar is also mediated by an alternative pathway that involves the 432
formation of (EZ)-phenylacetaldoxime (Irmisch et al 2013 Irmisch et al 2014 433
Figure 1) This oxime is biosynthesized from phenylalanine only upon herbivory by 434
the P450 enzyme CYP79D6v3 Downregulation of CYP79D6v3 in P times canescens led 435
to decreased emission of herbivore-induced 2-phenylethanol while N benthamiana 436
plants overexpressing CYP79D6v3 produced the alcohol suggesting a role of (EZ)-437
phenylacetaldoxime as an alternative precursor for 2-phenylethanol in planta (Irmisch 438
et al 2013) Indeed it has been proposed that (EZ)-phenylacetaldoxime can be 439
converted to 2-phenylacetaldehyde by a transoximase activity (Soumlrenssen et al 440
2018) or even directly to 2-phenylethanol by an alcohol dehydrogenase (Ferreira-441
Silva et al 2010) The enzymatic machinery behind these reactions however is still 442
not known Since the Km value of CYP79D6v3 for phenylalanine (Km = 744 microM) was 443
similar to that of PtAAS1 (Km = 460 microM) we hypothesize that both enzymes and 444
pathways contribute to the herbivore-induced formation of 2-phenylethanol in P 445
trichocarpa Notably the related poplar species P times canescens lacked expression of 446
AAS1 It is thus likely that P times canescens produces the alcohol exclusively via the 447 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

15
15
oxime-dependent pathway upon herbivory This assumption is strengthened by the 448
fact that the knockdown of AADC1 in P times canescens had no effect on the herbivore-449
induced emission of 2-phenylethanol (Figure 7) ruling out a potential contribution of 450
this enzyme to the formation of volatile 2-phenylethanol 451
452
PtAADC1 generates 2-phenylethylamine which can be metabolized into 2-453
phenylethyl-β-D-glucopyranoside in vivo 454
Plant aromatic amines and derived metabolites play important roles as signalling and 455
defence compounds and are usually produced by AADC enzymes (reviewed in 456
Facchini et al 2000) We identified and characterized three AADCs in P trichocarpa 457
that accepted various aromatic amino acids as substrates and converted them into 458
the corresponding aromatic amines in vitro Kinetic characterization revealed that 459
PtAADC1 and PtAADC2 preferred phenylalanine as substrate while PtAADC3 460
preferred tyrosine (Table 1) N benthamiana plants overexpressing PtAADC1 461
produced 2-phenylethylamine and minor amounts of tyramine and tryptamine 462
confirming the kinetic parameters determined in vitro RNAi-mediated knockdown of 463
AADC1 in P times canescens led to decreased levels of herbivore-induced 2-464
phenylethylamine and 2-phenylethyl-β-D-glucopyranoside while the emission of 2-465
phenylethanol was not influenced (Figure 7) These results indicate I) that PtAADC1 466
produces 2-phenylethylamine upon herbivory in planta and II) that this compound is 467
further metabolized most likely via 2-phenylacetaldehyde and 2-phenylethanol into 2-468
phenylethyl-β-D-glucopyranoside The presence of separate pathways to herbivore-469
induced 2-phenylethanol and 2-phenylethyl-β-D-glucopyranoside (both constitutive 470
and induced) suggests that these two metabolites have different functions in the 471
plant even if the metabolic fate of 2-phenylethyl-β-D-glucopyranoside is eventually to 472
be hydrolyzed to 2-phenylethanol Since emission of the free alcohol was not 473
influenced by the AADC1 knockdown we propose that the AADC-dependent 2-474
phenylethanol pathway in contrast to the AAS-mediated route is tightly associated 475
with a glucosyltransferase as part of a protein complex The formation of such 476
complexes or metabolons which can prevent the release of unstable or toxic 477
pathway intermediates has been described for the cyanogenic glycoside pathway in 478
sorghum (Sorghum bicolor) where a glucosyltransferase is also included (Nielsen et 479
al 2008 Laursen et al 2016) 480 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

16
16
481
Constitutive 2-phenylethyl-β-D-glucopyranoside might be produced via PtAAS2 482
or the Ehrlich pathway 483
Undamaged leaves of P trichocarpa were found to accumulate significant amounts 484
of 2-phenylethyl-β-D-glucopyranoside (Figure 2) Since PtAAS2 was the only AAAD 485
gene with a considerable constitutive expression we hypothesize that PtAAS2 486
provides 2-phenylacetaldehyde as substrate for the formation of 2-phenylethyl-β-D-487
glucopyranoside in undamaged poplar leaves (Figure 2) However the accumulation 488
of phenylpyruvic acid in these leaves indicates that the Ehrlich pathway might also 489
contribute to the formation of 2-phenylethyl-β-D-glucopyranoside An aromatic amino 490
acid transaminase has recently been reported in melon Its characterization and gene 491
expression pattern suggest that the catabolism of aromatic amino acids into aroma 492
volatiles likely proceeds via an initial transamination rather than decarboxylation or 493
decarboxylationdeamination in this species (Gonda et al 2010) Moreover roses 494
were shown to possess both the AADC-dependent pathway and the phenylpyruvic 495
acid-dependent pathway for the formation of floral 2-phenylethanol While the former 496
pathway is active throughout the whole year the latter is induced exclusively during 497
hot seasons and has been discussed as an adaptation to summer environmental 498
conditions (Hirata et al 2016) The presence of multiple 2-phenylethanol pathways 499
in poplar may also enable the tree to react to different environmental stresses by 500
producing either 2-phenylethanol or 2-phenylethyl-β-D-glucopyranoside 501
independently 502
503
PtAAS1 and PtAADC1 have evolved from a common ancestor by gene 504
duplication and neofunctionalization 505
The emergence of large gene families such as terpene synthases cytochrome P450 506
monooxygenases UDP-glucosyltransferases methyltransferases and acyl 507
transferases that encode enzymes with diverse substrate specificity regiospecificity 508
or catalytic activity created the basis for the chemical diversity of specialized 509
metabolism in plants (Leong and Last 2017) These gene families evolved through 510
repeated duplications of single genes chromosomes or even whole genomes 511
followed by neofunctionalization of resulting gene copies (Moghe and Kruse 2018) It 512
has been shown that single amino acid changes can dramatically alter the specificity 513 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

17
17
or activity of enzymes involved in specialized metabolism (eg Koumlllner et al 2004 514
Irmisch et al 2015 Johnson et al 2001 Junker et al 2013 Fan et al 2017) 515
suggesting that novel enzyme functions may arise in relatively short time periods by 516
the accumulation of only a few nucleotide mutations PtAADC1 and PtAAS1 provide 517
an example of rapid evolutionary changes They form a gene cluster on chromosome 518
13 of the P trichocarpa genome (Tuskan et al 2006) and their high similarity to each 519
other indicates a recent tandem gene duplication event However PtAADC1 and 520
PtAAS1 have already come to encode different enzymatic activities catalysing the 521
decarboxylation and decarboxylationoxidation of phenylalanine respectively Recent 522
studies reported a tyrosinephenylalanine switch that determines the catalytic activity 523
of AADC and AAS enzymes respectively in different species (Bertoldi et al 2002 524
Torrens-Spence et al 2013) In vitro mutagenesis experiments performed in our 525
study confirmed the role of this residue as a function-dictating element in PtAADC1 526
and PtAAS1 (Figure 4) Interestingly a survey of putative AADCAAS genes in a set 527
of sequenced vascular plants (httpwwwphytozomenet) revealed similar 528
AADCAAS gene pairs that differed in the function-dictating residue in at least ten 529
species from monocotyledonous and dicotyledonous plants (Supplemental Figure 5) 530
Although none of these enzymes has been characterized so far their sequence 531
similarities with already published AADCAAS from other plants suggest similar 532
catalytic functions (Supplemental Figure 6) The presence of the putative AADCAAS 533
gene pairs indicates an independent occurrence of the tyrosinephenylalanine-534
mediated activity switch of AADCAAS enzymes in single genera or even single 535
species of the angiosperms The multiple evolution of this trait as well as its 536
conservation in numerous plant lineages suggest different but important functions of 537
AADCAAS enzyme products in plant metabolism 538
539
PtPAR1 and PtPAR2 may participate in the biosynthesis of 2-phenylethanol 540
The last reaction of the various 2-phenylethanol pathways is the reduction of 2-541
phenylacetaldehyde catalyzed by 2-phenylacetaldehyde reductase (Watanabe et al 542
2002 Hirata et al 2012 Figure 1) We identified a group of five genes in the 543
genome of P trichocarpa that showed significant sequence similarity to the recently 544
reported PAR genes from tomato and damask rose (Tieman et al 2007 Chen et al 545
2011) Heterologous expression and enzyme characterization revealed PAR activity 546 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

18
18
for two candidates PtPAR1 and PtPAR2 However both enzymes also accepted a 547
variety of other aromatic and aliphatic aldehydes as substrates and converted them 548
into the corresponding alcohols (Table 2 Supplemental Figure 12) Substrate 549
promiscuity has been described for tomato LePAR2 and rose PAR1 (Tieman et al 550
2007 Chen et al 2011) as well as for short-chain dehydrogenasesreductases 551
involved in other metabolic processes (Oberschall et al 2000 Yamaguchi et al 552
2011 Sengupta et al 2015 Jain et al 2016) Since PtPAR1 and PtPAR2 showed 553
high constitutive expression that remained constant upon herbivory (Supplemental 554
Figure 12) they might contribute to the constitutive as well as herbivore-induced 555
formation of 2-phenylethanol and its glucoside in planta However we cannot rule out 556
that other members of the large short-chain dehydrogenasereductase family of 557
poplar (Supplemental Figure 10) are also involved in the biosynthesis of these 558
compounds 559
560
Herbivory-induced emission of 2-phenylethanol and accumulation of 2-561
phenylethylamine might serve as defensive weapons or signals in P 562
trichocarpa 563
Herbivore-induced volatiles play important roles in plant defence They can 564
negatively impact the attacking herbivore directly by toxicity or deterrence Or they 565
can act as indirect defences by functioning as cues for natural enemies of the 566
herbivore 2-Phenylethanol has been shown to be involved in both processes 567
Florivorous ants for example that feed on flowers of the alpine skypilot (Polemonium 568
viscosum) are strongly repelled by higher concentrations of 2-phenylethanol (Galen 569
et al 2011) On the other hand the aphid-feeding lacewing Chrysoperla carnea is 570
attracted by 2-phenylethanol released from host plants of the aphid (Zhu et al 571
2005) suggesting a function of this compound in indirect defense We propose that 572
2-phenylethanol might also be involved in direct or indirect defense in poplar The 573
volatile blend emitted from L dispar-infested poplar leaves has been shown to attract 574
Glyptapanteles liparidis parasitoids that oviposit on L dispar caterpillars (McCormick 575
et al 2014) and 2-phenylethanol one of the prominent blend components could 576
influence the behavior of the parasitic wasp Additionally 2-phenylethanol has been 577
shown to act as antimicrobial agent (Zhu et al 2011 Liu et al 2014) and thus might 578
protect poplar plants against pathogens associated with herbivores (Chung et al 579 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

19
19
2013 Zhu et al 2014) Alternatively 2-phenylethanol could also act as an internal 580
plant signal to indicate the presence of herbivore damage to other organs Other 581
herbivore-induced natural products especially volatiles have been shown to play 582
roles as defence signals (Maag et al 2015) In woody plants such as poplar where 583
transport through the vascular system involves long distances between branches 584
volatile signals may be important in providing more rapid information about the 585
location of herbivory (Frost et al 2007) 586
Aromatic amines and related condensation products such as hydroxycinnamic acid 587
amides are known to be of central importance in defence responses upon wounding 588
and pathogen attack (Facchini et al 2002 Macoy et al 2015) They have been 589
shown to accumulate after tissue damage in a variety of plants including for example 590
Arabidopsis tomato pepper and potato (Muroi et al 2009 Campos et al 2014 591
Newman et al 2001 Schmidt et al 1998) In potato hydroxycinnamic acid amides 592
are induced by Phytophthora infestans infection and elicitor-treated potato cell 593
cultures also accumulate significant amounts of these compounds (Schmidt et al 594
1998 Schmidt et al 1999) Similar effects have been observed in chitosan-treated 595
potato suspension cultures suggesting that insect feeding might also trigger related 596
plant responses (Villegas and Brodelius 1990) Whether aromatic amines 597
biosynthesized by the herbivore-induced enzyme PtAADC1 might also act as 598
substrates for other defence compounds such as hydroxycinnamic acid amides in 599
poplar should be investigated in future research 600
601
602
MATERIALS AND METHODS 603
Plant treatment 604
Western balsam poplar (Populus trichocarpa) (genotype Muhle Larsen) trees were 605
grown from monoclonal stem cuttings in a greenhouse (24 degC 60 relative humidity 606
16 h8 h lightdark cycle) in a 11 mixture of sand and soil (Klasmann potting 607
substrate Klasmann-Deilmann Geeste Germany) until they reached 1 m in height 608
Wildtype and transgenic grey poplar (P times canescens clone INRA 717-1B4) plants 609
were amplified by micropropagation as described by Behnke et al (2007) Saplings 610
of around 10 cm high were repotted to soil (Klasmann potting substrate) and 611
propagated in a controlled environment chamber for one month (day 22 degC night 18 612 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

20
20
degC 65 rel humidity 16 h8 h lightdark cycle) before they were transferred to the 613
green house 614
For herbivore treatment of poplar plants were enclosed in a PET bag 615
(ldquoBratschlauchrdquo Toppits Germany) with (herbivory) or without (control) 10 gypsy 616
moth (Lymantria dispar) caterpillars (larval stage L4) for 24 hours (start of the 617
treatment ~ 4 pm of day 1 end of the treatment ~ 4 pm of day 2) 618
Lymantria dispar egg batches were provided by Hannah Nadel United States 619
Department of Agriculture- Animal and Plant Health Inspection Service (Buzzardrsquos 620
Bay MA USA) After hatching the caterpillars were reared on an artificial diet (gypsy 621
moth diet MP Biomedicals Illkirch France) until L4 stage Caterpillars were starved 622
in single cups for 1 day prior to the experiment 623
624
RNA extraction and cDNA synthesis 625
Poplar leaf material was harvested immediately after the herbivore treatment flash-626
frozen in liquid nitrogen and stored at -80 degC until further processing After grinding 627
the frozen leaf material in liquid nitrogen to a fine powder with a Vibratory Micro Mill 628
(Pulverisette-0 Fritsch Germany) total RNA was isolated using an InviTrap Spin 629
Plant RNA kit (Stratec Berlin Germany) according to manufacturerrsquos instructions 630
RNA concentration was assessed using a spectrophotometer (NanoDrop 2000c 631
Thermo Scientific Wilmington DE USA) Prior to cDNA synthesis 1 microg of RNA was 632
DNase treated using 1 U of DNase (Thermo Fisher Scientific 633
httpswwwthermofishercom) Single-stranded cDNA was prepared using 634
SuperScriptTM III reverse transcriptase and oligo (dT20) primers (Thermo Fisher 635
Scientific) 636
637
Identification and cloning of putative AAAD genes 638
Putative AAAD genes were identified using a BLAST search against the P 639
trichocarpa genome (Tuskan et al 2006) version 30 640
(httpwwwphytozomenetpoplar) with rose RhPAAS (GenBank Accession 641
ABB04522) as template The complete ORF of the resulting candidates PtAAS1 642
PtAAS2 PtAADC1 PtAADC2 and PtAADC3 were amplified from cDNA made from 643
L dispar-damaged P trichocarpa leaves using a Phusionreg High Fidelity DNA 644
Polymerase (New England Biolabs) and inserted into the pCRtrade-Blunt II-TOPOreg 645 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

21
21
vector (Thermo Fisher Scientific) Primer sequence information is available in 646
Supplemental Table 3 647
648
Identification and cloning of putative PAR genes 649
For the identification of putative PAR and AAR genes a BLAST search was carried 650
out as described above with LePAR1 (Gene Bank Accession EF6134921) as query 651
The complete ORFs of the resulting candidates PtAAR1 PtAAR2 PtAAR3 PtPAR1 652
and PtPAR2 were amplified from cDNA made from L dispar-damaged P trichocarpa 653
leaves using a Phusionreg High Fidelity DNA Polymerase (New England Biolabs) and 654
inserted into the pET100D-TOPOreg vector (Thermo Fisher Scientific) Primer 655
sequence information is available in Supplemental Table 3 656
657
Phylogenetic analysis and amino acid alignment 658
Nucleotide sequence alignments were constructed with the Muscle (codon) algorithm 659
(gap open -29 gap extend 0 hydrophobicity multiplier 15 clustering method 660
upgmb) implemented in MEGA6 (Tamura et al 2013) Phylogenetic trees were 661
generated with MEGA6 using the Maximum Likelihood method (modelmethod 662
Kimura-2-parameter model substitutions type nucleotide rates among sites uniform 663
rates gapsmissing data treatment partial deletion site coverage cutoff 80) An 664
amino acid alignment of poplar AAAD enzymes characterized in this study together 665
with Arabidopsis AtAAS and AtTyrDC (Gutensohn et al 2011 Lehmann and 666
Pollmann 2009) was constructed and visualized with BioEdit 667
(httpwwwmbioncsuedubioeditbioedithtml) and the ClustalW algorithm 668
669
RNA-Sequencing and RT-qPCR analysis 670
Total RNA was extracted from leaf material as described above TruSeq RNA-671
compatible libraries were prepared and PolyA enrichment was performed before 672
sequencing the eight transcriptomes of P trichocarpa (four trees (biological 673
replicates) for each the control and herbivory treatment) on an IlluminaHiSeq 2500 674
sequencer (Max Planck Genome Centre Cologne Germany) with 18 Mio reads per 675
library 100 base pair single end Trimming of the obtained Illumina reads and 676
mapping to the poplar gene model version 30 677
(httpsphytozomejgidoegovpzportalhtml) were performed with the program CLC 678 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

22
22
Genomics Workbench (Qiagen Bioinformatics) (mapping parameter length fraction 679
07 similarity fraction 09 max number of hits 25) Empirical analysis of digital gene 680
expression (EDGE) implemented in the program CLC Genomics Workbench was 681
used for gene expression analysis 682
For validation of the RNA-sequencing results quantitative real-time PCR was carried 683
out with specific primers for each gene Therefore cDNA was prepared as described 684
above and diluted 110 with sterile water For the amplification of gene fragments 685
with a length of about 150-300 bp primer pairs were designed having a Tm ge 60 degC 686
a GC content between 40 - 55 and a primer length in the range of 20 - 25 nt (see 687
Supplemental Table 3 for primer information) Primer pair efficiency was tested by 688
measuring Ct values in stepwise diluted cDNA Amplicon identity was validated via 689
sequencing of cloned fragments Samples were run in triplicate using Brilliant III 690
SYBR Green qPCR Master Mix (Agilent) The following PCR conditions were applied 691
for all reactions initial incubation at 95 degC for 3 min followed by 40 cycles of 692
amplification (95 degC for 20 s and 60 degC for 20 s) SYBR Green fluorescence was 693
measured at the end of each amplification cycle Melting curves were recorded within 694
a range of 55 to 95 degC at the end of a total of 40 cycles All samples were run on a 695
Bio-Rad CFX Connecttrade Real-Time PCR Detection System (Bio-Rad Laboratory 696
Hercules CA USA) in 96-well Hard-Shellreg PCR plates with 6 biological replicates 697
The relative normalized expression compared to the housekeeping gene ubiquitin 698
(Irmisch et al 2013) was analyzed using the Bio-Rad CFX Manager 31 699
700
Biochemical characterization of poplar AAAD enzymes 701
For heterologous expression in Escherichia coli the complete ORFs of the candidate 702
AAAD genes were subcloned into the pET100D-TOPOreg vector and expressed in E 703
coli BL21(DE3) cells (Thermo Fisher Scientific) as N-terminal His6-Tag fusion 704
proteins Cells were grown in TB medium (12 wv tryptone 24 wv yeast 705
extract 05 (vv) glycerol adjusted to pH 72 via potassium buffer 50 microgmL 706
carbenicillin) at 32 degC and 220 rpm until an OD600 of 04 was reached Expression 707
was induced by adding 04 mM IPTG and the cultures were grown overnight at 18degC 708
under continuous shaking at 220 rpm Cells were harvested by centrifugation (5000 times 709
g 10 min 4 degC) the supernatant was decanted and the pellet was resuspended in 710
chilled lysis buffer (20 mM imidazole pH 72 100 mM NaCl 50 mM KH2PO4 2 711 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

23
23
(vv) glycerol 10 mM MgCl2 03 mgmL lysozyme 100 uL Benzonase (Thermo 712
Fisher Scientific) 1 gL proteinase inhibitor HP (Serva wwwservade)) The cells 713
were frozen in liquid nitrogen and thawed in a water bath at 25 degC a total of 5 times 714
and additionally disrupted by a 3 times 30 s treatment with a sonicator (Bandelin 715
UW2070 Berlin Germany 70 intensity) The resulting slurry was pelleted via 716
centrifugation (12000 times g 20 min 4 degC) and the supernatant was transferred to a 717
new reaction tube Soluble protein was purified from the supernatant using a Poly-718
Prepreg Chromatography gravity flow column (Bio-Rad www-bio-radcom) with 500 microL 719
HisPurtrade Cobalt Resin (Thermo Fischer Scientific) according to the manufacturerrsquos 720
instructions (equilibration buffer 100 mM NaCl 50 mM Tris-HCl 2 vv glycerol 20 721
mM imidazole pH 72 washing buffer 100 mM NaCl 50 mM Tris-HCl 2 vv 722
glycerol 40 mM imidazole pH 72 elution buffer 100 mM NaCl 50 mM Tris-HCl 2 723
vv glycerol 150 mM imidazole pH 72) The enzyme-containing eluate was desalted 724
via an Illustra NAP-10 gravity flow column (GE Healthcare) into a storage buffer (50 725
mM Tris-HCl 01 mM EDTA pH 80) and immediately mixed 11 with a PLP-726
containing buffer (50 mM Tris-HCl 02 mM PLP 01 mM EDTA pH 80) Protein 727
concentration was determined via a PCA-enhanced Biuret assay as stated by the 728
manufacturer (Rotireg-Quant universal Karl Roth GmbH Germany) Protein purity and 729
identity was confirmed via SDS-PAGE and colloidal Coomassie staining (Rotireg-Blue 730
Karl Roth GmbH Germany) The relative abundance of the enzymes defined by 731
densitometry (GS-900 Calibrated Densitometer Bio-Rad Germany) was calculated 732
for the determination of kcat values 733
Enzyme assays were performed in closed glass tubes containing approximately 1 microg 734
of purified enzyme and 1 mM of substrate in a total volume of 200 microL of PLP buffer 735
The reaction was carried out for 30 minutes under shaking at 300 rpm and 32 degC and 736
was stopped by adding 800 microl of 100 methanol After placing on ice for 30 minutes 737
the denatured enzymes were removed by centrifugation (4000 times g 4 degC 10 min) 738
and the supernatant was transferred into a glass vial or a 96 Deep-Well Agilent 41 739
mm plate for LC-MSMS analysis 740
For the determination of the kinetic parameters assays were carried out in triplicate 741
and with varying substrate concentrations The enzyme concentration and the 742
incubation times were chosen so that the reaction velocity was linear during the 743
incubation time 744 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

24
24
745
Biochemical characterization of poplar PAR enzymes 746
The complete ORFs of the candidate AAR and PAR genes were cloned into the 747
pET100D-TOPOreg vector and expressed in E coli BL21(DE3) cells (Thermo Fisher 748
Scientific) as N-terminal His6-Tag fusion proteins as described above for AADC and 749
AAS enzymes Soluble protein was purified from the supernatant as described by 750
Irmisch et al (2015) Protein purity and identity was confirmed via SDS-PAGE and 751
colloidal Coomassie staining (Rotireg-Blue Karl Roth GmbH Germany) Enzyme 752
assays were performed in closed glass tubes containing approximately 1 microg of 753
purified enzyme 1 mM of substrate and 1 mM of NADPH in a total volume of 100 microL 754
of reaction buffer (10 glycerol (vv) 10 mM TrisHCl pH = 75 1 mM dithiothreitol) 755
and were carefully overlaid with 150 microL hexane and incubated for 15 hours After 756
incubation enzyme assays were thoroughly mixed the hexane phase was 757
transferred to a new glass vessel and 1 microL was analyzed as described in Irmisch et 758
al (2015) 759
760
Site-directed mutagenesis of PtAAS1 and PtAADC1 761
For site-directed mutagenesis of PtAAS1 and PtAADC1 specific primers possessing 762
the desired mutated codon were designed after the method described by Ho et al 763
1989 (Supplemental Table 3) The vectors pET100D-TOPOregPtAADC1 and 764
pET100D-TOPOregPtAAS1 were used as templates for a Phusionreg High Fidelity DNA 765
PCR The elongation time was adapted to the total length of the template vector 766
according to the instructions of the supplier The double-nicked vector was directly 767
transformed into chemically competent E coli cells (TOP10 One Shot Thermo Fisher 768
Scientific) and screened for successful mutagenesis by sequencing of plasmids 769
purified from single colonies Plasmids containing the desired mutations were 770
transformed into E coli BL21(DE3) cells (Thermo Fisher Scientific) and heterologous 771
expression protein purification and activity tests were performed as described 772
above 773
774
Transient Expression of PtAADC1 and PtAAS1 in Nicotiana benthamiana 775
For expression in N benthamiana the coding regions of PtAADC1 and PtAAS1 were 776
amplified from the respective pET100D-TOPOreg vectors and inserted into the 777 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

25
25
pCAMBiA2300U vector as described in Irmisch et al (2013) The pCAMBiA 2300U 778
vector and vectors carrying eGFP and p19 were kindly provided by the group of D 779
Werck-Reichhart Strasbourg France After verification of the sequence integrity 780
pCAMBiA vectors were separately transferred into Agrobacterium tumefaciens strain 781
C58p90 Protein expression was confirmed by fluorescence analysis (microscope) of 782
N benthamiana plants transformed with eGFP Five milliliters of an overnight culture 783
(220 rpm 28 degC) were used to inoculate 200 mL of Luria-Bertani medium (50 microgmL 784
kanamycin 25 microgmL rifampicin 25 microgmL gentamicin) for overnight growth The 785
following day the cultures were centrifuged (4000 times g 5 min) and the cells were 786
resuspended in infiltration buffer (10 mM MES 10 mM MgCl2 and 100 microM 787
acetosyringone pH 56) to reach a final OD600 of 05 After shaking for at least 2 h at 788
room temperature the cultures carrying PtAADC1 PtAAS1 or eGFP were mixed 789
with an equal volume of cultures carrying pBINp19 For transformation 3- to 4-week-790
old N benthamiana plants were dipped upside down in an Agrobacterium solution 791
and vacuum was applied to infiltrate the leaves Infiltrated plants were placed in a 792
room without direct sunlight Three days after transformation plants were placed 793
under low direct light (LED 40) for three more days Volatiles were measured on 794
the sixth day after transformation For volatile collection plants were separately 795
placed in gas-tight 3-liter glass desiccators Charcoal-filtered air was pumped into the 796
desiccators at a flow rate of 1 Lmin and the air left the desiccators through a filter 797
packed with 30 mg of PorapaqQ (ARS Gainesville FL USA) Volatiles were 798
collected for 6 h (10 AM to 4 PM) The volatile compounds were desorbed from the 799
filters and analyzed by gas chromatography-mass spectrometry (GC-MS) and gas 800
chromatographyndashflame ionization detection (GC-FID) as described below Plants 801
were harvested after the volatile collection ground in liquid nitrogen and stored at -802
80 degC until further analysis 803
804
Vector construction and transformation of poplar 805
The construction of the binary vector was described by Leveacutee et al (2009) The 806
transformation of the P times canescens clone INRA 7171-B4 followed a protocol 807
published by Meilan and Ma (2007) To target only AADC1AAS1 mRNA a fragment 808
between position 455 and 765 of the coding sequence of PtAADC1 was selected 809
Transgenic RNAi poplar plants were amplified by micropropagation as described by 810 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

26
26
Behnke et al (2007) To test the level of transgenicity RT-qPCR analysis was done 811
on wild type vector control (pCAMBIA) and RNAi plants 812
813
Chemical conversion of PtAAS enzyme products into aldoximes 814
For the identification of reactive aromatic aldehydes PtAAS enzymes were incubated 815
with single amino acids as substrates overlaid with 400 microl of pure toluol and 816
incubated for 1 hour at 32 degC The aromatic aldehydes produced by the enzymes 817
were extracted into the toluol phase and transferred to a new glass vial The toluol 818
phase was incubated with an aqueous solution of 100 mM hydroxylamine (pH = 70) 819
at 25 degC for one hour as described earlier (Irmisch et al 2014 Hofmann et al 2018) 820
to form the corresponding aldoxime The toluol phase was then transferred to a fresh 821
glass vial and dried down under a continuous flow of nitrogen The pellet was 822
dissolved in 200 microl methanol and analyzed via LC-MSMS as described in Irmisch et 823
al (2013) 824
825
LC-MSMS analysis of plant methanol extracts 826
Metabolites were extracted from ground plant material (poplar or N benthamiana) 827
with methanol (101 vw) Analytes were separated using an Agilent 1200 HPLC 828
system on a Zorbax Eclipse XDB-C18 column (50 times 46 mm 18 microm (Agilent 829
Technologies)) Mobile phase consisted of formic acid (005 vv) in water (A) and 830
acetonitrile (B) The column temperature was maintained at 25 degC HPLC parameters 831
are given in Supplemental Table 4 832
The liquid chromatography was coupled to an API-5000 tandem mass spectrometer 833
(Applied Biosystems Foster City CA USA) equipped with a Turbospray ion source 834
(ion spray voltage 5500 eV turbo gas temperature 700 degC nebulizing gas 70 psi 835
curtain gas 35 psi heating gas 70 psi collision gas 2 psi) Multiple reaction 836
monitoring (MRM) was used to monitor a parent ion rarr product ion reaction for each 837
analyte Detailed parameters are given in Supplemental Table 5 Identification and 838
quantification of compounds were performed using standard curves made from 839
authentic and commercially available standards of 2-phenylethylamine tyramine 840
tryptamine 2-phenylethyl-β-D-glucopyranoside phenylpyruvic acid and 841
phenylalanine (Supplemental Table 6) Relative quantification of ethanolamine 842
(Supplemental Figure 4) was carried out under the conditions mentioned in 843 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

27
27
Supplemental Tables 4 and 5 Ethanolamine was identified via an authentic standard 844
(Supplemental Table 6) 845
846
GC-MSFID analysis of plant volatiles PAR and AAR products 847
A Hewlett-Packard model 6890 gas chromatograph (GC) was employed with He 848
(MS) or H2 (FID) as carrier gas at 2 mLmin splitless injection (injector temperature 849
230 degC injection volume 1 microL) and a DB-5MS column (Agilent Santa Clara USA 30 850
m times 025 mm times 025 microm) The GC oven temperature was held for 2 min and then 851
increased to 225 degC with a gradient of 5 degCmin held for another 2 min and then 852
further increased to 250 degC with 100 degCmin and a hold for 1 min The coupled mass 853
spectrometer (MS) was a Hewlett-Packard model 5973 with a quadrupole mass 854
selective detector (transfer line temperature 230 degC source temperature 230 degC 855
quadrupole temperature 150 degC ionization potential 70 eV scan range of 40-400 856
atomic mass units) Quantification was performed with the trace of a flame ionization 857
detector (FID) operated at 300 degC For analysis of PAR and AAR reaction products 1 858
microl of the hexane phase covering the enzyme assays was injected into GC-MS and 859
analyzed as described above 2-Phenylethanol and 2-phenylacetaldehyde were 860
identified via authentic standards (Supplemental Table 6) 861
862
TDU-GC-MS analysis of PtAAS enzyme products 863
PtAAS enzyme products were collected from the headspace of the assays with 864
PDMS silicone tubes as described by Kallenbach et al (2014) For the determination 865
of Km values enzyme assays as described above were incubated with 866
preconditioned PDMS silicone tubes in an air-tight vial for 30 minutes After the 867
incubation PDMS tubes were immediately analyzed using thermal desorption GC-868
MS For preparing a standard curve 2-phenylacetaldehyde was dissolved in 100 869
ethanol and added in specific amounts to the buffer system used for the enzyme 870
assays As an enzyme control 1 microg of BSA was added per assay and total volume of 871
the assay was set to 200 microl 872
A Shimadzu thermal desorption unit TD-20 combined with a GC-MS-QP2010 Ultra 873
gas chromatograph was employed with He as carrier gas Thermal desorption of 874
PDMS tubes was carried out with a sample flow of 60 mLmin a desorption 875
temperature of 200 degC a valve temperature of 250 degC and a transfer line 876 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

28
28
temperature of 230 degC The trap tube was cooled (-20 degC) and upon completed 877
desorption heated to 230 degC to release the sample onto the GC column The gas 878
chromatograph was operated at a column flow rate of 15 mLmin (helium) split 879
injection (split ratio 20) and a ZB-5MS column (Zebron Phenomenex California 880
USA 30 m times 025 mm times 025 microm) The GC oven temperature was set to 45 degC held 881
for 3 min then increased to 180 degC with a gradient of 6 degCmin finally increased to 882
300 degC with a 100 degCmin increment and held for another 23 min The coupled mass 883
spectrometer with a quadrupole mass selective detector was operated in Scan mode 884
selective for 33 ndash 350 atomic mass units (interface temperature 250 degC source 885
temperature 230 degC ionization potential 70 eV) 886
887
Statistical analysis 888
Statistical analysis was carried out as described in the figure legends for the 889
respective experiments Studentacutes t-tests Mann-Whitney Rank Sum tests Kruskal-890
Wallis One Way ANOVA and Tukey tests were performed with SigmaPlot 120 891
EDGE tests were performed with CLC Genomics Workbench Data visualization was 892
done with SigmaPlot 120 and R Studio Version 099903 (R Development Core 893
Team httpwwwR-projectorg) 894
895
896
ACCESSION NUMBERS 897
Sequence data for PtAAS1 (MK440562) PtAAS2 (MK440561) PtAADC1 898
(MK440568) PtAADC2 (MK440567) PtAADC3 (MK440566) PtSDC1 (MK440558) 899
PtSDC2 (MK440569) PtPAR1 (MK440560) PtPAR2 (MK440559) PtAAR1 900
(MK440565) PtAAR2 (MK440564) and PtAAR3 (MK440563) can be found in the 901
GenBank under the corresponding identifiers Raw reads of the RNAseq experiment 902
were deposited in the NCBI Sequence Read Archive (SRA) under the accession 903
PRJNA516861 904
905
Supplemental Data 906
Supplemental Figure S1 Amino acid alignment of PtAAS and PtAADC enzymes 907
from Populus trichocarpa and characterized AtAAS and AtTyrDC from Arabidopsis 908
thaliana 909 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

29
29
Supplemental Figure S2 PtAAS1 and PtAAS2 do not release 2-phenylethylamine 910
Supplemental Figure S3 Detection of poplar AAS products via the conversion to 911
corresponding aldoximes 912
Supplemental Figure S4 PtSDC1 and PtSDC2 convert L-serine to ethanolamine 913
Supplemental Figure S5 Phylogeny and comparison of plant AAAD gene clusters 914
Supplemental Figure S6 Dendrogram analysis of putative AADCAAS gene pairs 915
and characterized AADC AAS and GluSerHis-decarboxylase genes from other 916
plants 917
Supplemental Figure S7 Transcript accumulation of PtAAS1 PtAADC1 PtAADC2 918
PtAAS2 and PtAADC3 in caterpillar-damaged (herb) and undamaged (ctr) leaves of 919
Populus trichocarpa 920
Supplemental Figure S8 Dendrogram analysis of AAAD genes from Populus 921
trichocapra P times canescens P tremula and P alba 922
Supplemental Figure S9 Transcript accumulation of AAS1 AADC1 and AADC2 in 923
gypsy moth (Lymantria dispar) caterpillar-damaged Populus times canescens leaves 924
Supplemental Figure S10 Transcript accumulation of PcanAADC1 (A) and 925
PcanAAS2 (B) in caterpillar-damaged leaves of control and AADC1AADC2AAS1 926
RNAi lines of Populus times canescens 927
Supplemental Figure S11 The RNAi-mediated knockdown of PcanAADC1 in 928
Populus times canescens does not influence the herbivore-induced emission of benzyl 929
cyanide 930
Supplemental Figure S12 Dendrogram analysis of putative reductases from poplar 931
and characterized reductases from other plants 932
Supplemental Figure S13 PtPAR1 and PtPAR2 convert 2-phenylacetaldehyde into 933
2-phenylethanol in vitro 934
Supplemental Figure S14 Transcript accumulation of putative aldehyde reductase 935
genes in Lymantria dispar-damaged and undamaged Populus trichocarpa leaves 936
Supplemental Table S1 Accession numbers of genes used for phylogenetic 937
analysis 938
Supplemental Table S2 Expression of PtAAS1 PtAADC1 and PtAADC2 in 939
undamaged and herbivore-damaged Populus trichocarpa leaves 940
Supplemental Table S3 Oligonucleotides used for isolation and RT-qPCR analysis 941
of poplar AAS AADC SDC PAR and AAR genes 942 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

30
30
Supplemental Table S4 HPLC gradients used for separation and analysis of 943
metabolites 944
Supplemental Table S5 MSMS parameters used for LC-MSMS analysis 945
Supplemental Table S6 Compounds used as substrates for enzyme assays and as 946
standards for LC-MSMS and GC-MS quantification 947
948
949
ACKNOWLEDGEMENTS 950
We thank Tamara Kruumlgel Danny Kessler and all the MPI-CE gardeners for their help 951
with rearing the poplar and Nicotiana benthamiana plants D Werck-Reichhart 952
Strasbourg France is thanked for kindly providing the pCAMBIA vectors and for the 953
nice introduction to the USER cloning system We thank Tabea Kroumlber Daniel Veit 954
Natascha Rauch and Marion Staumlger for excellent technical assistance Additionally 955
we thank Sybille B Unsicker (MPI-CE) and her group members for rearing L dispar 956
caterpillars Alexander Haverkamp is thanked for help with the data visualization 957
This research was funded by the Max Planck Society 958
959
960
TABLES 961
Table 1 Kinetic parameters of Populus trichocarpa PtAAS1 PtAAS2 PtAADC1 962
PtAADC2 and PtAADC3 Enzymes were heterologously expressed in Escherichia 963
coli purified and incubated with phenylalanine tyrosine or tryptophan Kinetic 964
parameters are shown as means plusmn SE (n = 3) Volatile phenylacetaldehyde was 965
analyzed via TDU-GC-MS while all other compounds were measured using LC-966
MSMS Compounds marked with asterisks were chemically converted into the 967
corresponding aldoximes before LC-MSMS analysis (see method section) 968
Enzyme Substrate Product Km
(mM) V
max
(nmolmicrog-1
min-1
) k
cat
(s-1
) k
cat K
m
(s-1
mM-1
)
PtAAS1
Phe PHA 046 plusmn 002 - - - Phe PEA - - - - Tyr 4-OH-PHA - - - - Trp IAAld - - - -
PtAAS2 Phe PHA 069 plusmn 039 - - - Phe PEA - - - -
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

31
31
Tyr 4-OH-PHA detected - - - Trp IAAld detected - - -
PtAADC1
Phe PHA - - - - Phe PEA 017 plusmn 003 430 plusmn 12 039 23 Tyr TyrA 20 plusmn 02 854 plusmn 39 008 004 Trp TrpA 30 plusmn 04 77 plusmn 05 0007 0002
PtAADC2
Phe PHA - - - - Phe PEA 019 plusmn 001 089 plusmn 001 081 42 Tyr TyrA 12 plusmn 02 148 plusmn 7 014 011 Trp TrpA - - - -
PtAADC3
Phe PHA - - - - Phe PEA 23 plusmn 02 67 plusmn 02 014 0003
Tyr TyrA 0043 plusmn 0004 361 plusmn 006 0003 008
Trp TrpA - - - - 969
970
Table 2 Enzymatic activity of poplar aldehyde reductases Enzymes were 971
heterologously expressed in Escherichia coli purified and tested with different 972
aldehydes as potential substrates in the presence of NADPH Enzyme products were 973
extracted with hexane and analyzed using GC-MS - no activity detected + activity 974
detected 975
Substrate Product PtPAR1 PtPAR2 PtAAR1 PtAAR2 PtAAR3 citronellal citronellol + + + + +
geranial geraniol + + - - -
nonanal nonanol + + + - -
decanal decanol + + + - +
hexanal hexanol + + - - -
(E)-2-hexenal (E)-2-hexenol + + - - -
benzaldehyde benzyl alcohol + + - - -
2-phenylacetaldehyde 2-phenylethanol + + - - - 976
977
FIGURE LEGENDS 978
PHA phenylacetaldehyde PEA 2-phenylethylamine 4-OH-PHA 4-hydroxy phenylacetaldehyde IAAld indole-3-acetaldehyde TyrA tyramine TrpA tryptamine
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

32
32
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar 979
AAAT aromatic amino acid transaminase AADC aromatic amino acid 980
decarboxylase CYP79 cytochrome P450 family 79 enzyme AAS aromatic 981
aldehyde synthase PPDC phenylpyruvic acid decarboxylase MAO monoamine 982
oxidase TOX transoximase PAR 2-phenylacetaldehyde reductase UGT UDP-983
glucuronosyltransferase β-Glu β-glucosidase Solid lines indicate well-established 984
reactionsenzymes while dashed lines indicate reactions hypothesized in the 985
literature 986
987
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and 988
release phenylalanine-derived metabolites The accumulation of L-phenylalanine 989
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-990
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in 991
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material 992
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were 993
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n 994
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-995
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064) 996
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-997
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T = 998
585) DW dry weight ns not significant nd not detected 999
1000
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family 1001
genes from Populus trichocarpa and characterized AADC AAS and 1002
GluSerHis-decarboxylase genes from other plants Substrates of each clade are 1003
indicated in parentheses The tree was inferred by using the maximum likelihood 1004
method and n = 1000 replicates for bootstrapping Bootstrap values are shown next 1005
to each node The tree is drawn to scale with branch lengths measured in the 1006
number of substitutions per site Genes described in this study are shown in red and 1007
bold with corresponding gene identifiers (italics) Accession numbers are given in 1008
Supplemental Table 1 1009
1010
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

33
33
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of a single 1011
amino acid and are likely derived by tandem gene duplication (A)The 1012
recombinant proteins PtAAS1 PtAADC1 and the corresponding mutant enzymes 1013
PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated with L-phenylalanine in the 1014
presence of PLP Enzyme products were analyzed using GC-MS (2-1015
phenylacetaldehyde) and LC-MSMS (2-phenylethylamine) (B) PtAADC1 and 1016
PtAAS1 form a gene cluster on chromosome 13 Both genes are oriented in the 1017
same direction and there are no coding regions between them Sequence data were 1018
retrieved from the P trichocarpa genome assembly version 30 (wwwphytozomenet 1019
Tuskan et al 2006) 1020
1021
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana 1022
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation 1023
of 2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-1024
D- glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium 1025
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the 1026
headspaces of plants were collected and analyzed via GC-MS and GC-FID 1027
Additionally plants were harvested extracted with methanol and metabolite 1028
accumulation was analyzed via LC-MSMS Different letters above each bar indicate 1029
statistically significant differences as assessed by Kruskal-Wallis One Way ANOVA 1030
and Tukey tests 2-phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1031
1694 P le 0001) tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 1032
0001) 2-phenylethyl-β-D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are 1033
shown (n = 6) FW fresh weight 1034
1035
Figure 6 Transcript accumulation of AAAD and SDC genes in Lymantria 1036
dispar-damaged and undamaged Populus trichocarpa leaves Gene expression 1037
in herbivore-damaged (herb) and undamaged (ctr) leaves was analyzed by Illumina 1038
sequencing and mapping the reads to the transcripts of the P trichocarpa genome 1039
version v30 Expression was normalized to RPKM Significant differences in EDGE 1040
tests are visualized by asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 1041
823213E-12 weighted difference (WD) = 486618E-05) PtAAS2 (P = 0039452261 1042
WD = -111092E-05) PtAADC1 (P = 899025E-09 WD = 156932E-05) PtAADC2 (P 1043 httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

34
34
= 0031269756 WD = 202404E-06) PtAADC3 (P = 1 WD = -435895E-08) 1044
PtSDC1 (P = 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012 WD = 1045
108813E-05) 1046
1047
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile 1048
Populus times canescens calli were transformed via Agrobacterium tumefaciens 1049
Nontransgenic trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) 1050
were subjected to Lymantria dispar feeding 2-Phenylethanol was collected from the 1051
headspace and analyzed using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-1052
β-D-glucopyranoside were extracted with methanol from ground plant material and 1053
analyzed via LC-MSMS Biological replicates (nb) and technical replicates (nt) of EV 1054
lines and RNAi lines were used to test for statistical differences WT nb = 4 EV nb = 1055
3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate statistical significance as assessed 1056
by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-Phenylethylamine (P lt 1057
0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T = 547) 2-1058
phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown 1059
Each data point is represented by a circle FW fresh weight ns ndash not significant 1060
1061
1062
1063
COMPETING INTERESTS 1064
The authors declare that they have no competing interests 1065
1066
LITERATURE CITED 1067
Arimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars 1068 (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles 1069 in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional 1070 characterization and patterns of gene expression of (-)-germacr Plant J 37 603ndash616 1071
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A 1072 Bohlmann J and Schnitzler JP (2007) Transgenic non-isoprene emitting poplars 1073 donrsquot like it hot Plant J 51 485ndash499 1074
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of 1075 tyrosine 332 to phenylalanine converts dopa decarboxylase into a decarboxylation-1076 dependent oxidative deaminase J Biol Chem 277 36357ndash36362 1077
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild 1078 Erythroxylum species J Ethnopharmacol 103 439ndash447 1079
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and 1080 Belleacutes JM (2014) Transgenic Tomato Plants Overexpressing Tyramine N -1081 Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and 1082
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

35
35
Enhanced Resistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1083 1159ndash1169 1084
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S 1085 and Watanabe N (2011) Functional characterization of rose phenylacetaldehyde 1086 reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 1087 2-phenylethanol J Plant Physiol 168 88ndash95 1088
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS 1089 and Felton GW (2013) Herbivore exploits orally secreted bacteria to suppress plant 1090 defenses Proc Natl Acad Sci 110 15728ndash15733 1091
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and 1092 Lewis NG (2013) Transgenic hybrid poplar for sustainable and scalable production of 1093 the commodityspecialty chemical 2-phenylethanol PLoS One 8 1094
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker 1095 SB and Koumlllner TG (2011) Four terpene synthases produce major compounds of 1096 the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 1097 72 897ndash908 1098
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism 1099 physiology and biochemistry Can J Bot 80 577ndash589 1100
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid 1101 decarboxylases Evolution biochemistry regulation and metabolic engineering 1102 applications Phytochemistry 54 121ndash138 1103
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution 1104 of glucosinolates and isothiocyanates amoung plants Phytochemistry 56 5ndash51 1105
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped 1106 pathway creates metabolic innovation in tomato trichomes through BAHD enzyme 1107 promiscuity Nat Commun 8 1ndash13 1108
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and 1109 Vainstein A (2010) Identification of rose phenylacetaldehyde synthase by functional 1110 complementation in yeast Plant Mol Biol 72 235ndash245 1111
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-1112 promiscuity of alcohol dehydrogenases reduction of phenylacetaldoxime to the alcohol 1113 Tetrahedron 66 3410ndash3414 1114
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC 1115 (2007) Within-plant signalling via volatiles overcomes vascular constraints on systemic 1116 signalling and primes responses against herbivores Ecol Lett 10 490ndash8 1117
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-1118 Dependent Impacts of a Floral Volatile Compound on Pollinators Larcenists and the 1119 Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 1120 258ndash272 1121
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein 1122 S Giovannoni JJ Katzir N and Lewinsohn E (2010) Branched-chain and 1123 aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1124 1123 1125
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C 1126 Hiratake J Sakata K Takada M Ogawa K and Watanabe N (2004) Emission 1127 of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these 1128 compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013 1129
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) 1130 The Ehrlich pathway for fusel alcohol production A century of research on 1131 Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 1132 74 8 (2259-2266)) Appl Environ Microbiol 74 3920 1133
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N 1134 (2012) Functional characterization of aromatic amino acid aminotransferase involved in 1135 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 1136
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

36
36
444ndash451 1137 Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J 1138
Suzuki H Ishikawa T Dohra H and Watanabe N (2016) Seasonal induction of 1139 alternative principal pathway for rose flower scent Sci Rep 6 1140
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed 1141 mutagenesis by overlap extension using the polymerase chain reaction Gene 77 51ndash1142 59 1143
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled 1144 indole-3-acetic acid precursors J Label Compd Radiopharm 18 1491ndash1495 1145
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From 1146 Ligustrum japonicum that Stimulate Flower-Visiting by Cabbage Butterfly Pieris rapae 1147 J Chem Ecol 24 2167ndash2180 1148
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia 1149 communis to 2-phenylethanol a major volatile component from host flowers Rosa spp 1150 J Chem Ecol 24 1491ndash1497 1151
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M 1152 Schneider B Block K Schnitzler J-P Gershenzon J Unsicker SB and 1153 Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 1154 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar 1155 Defense Plant Cell 25 4737ndash4754 1156
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA 1157 Gershenzon J Unsicker SB and Koumlllner TG (2014a) Herbivore-induced poplar 1158 cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which 1159 repel a generalist caterpillar Plant J 80 1095ndash1107 1160
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene 1161 synthases and their contribution to herbivore-induced volatile emission in western 1162 balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16 1163
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG 1164 (2015) One amino acid makes the difference The formation of ent-kaurene and 16α-1165 hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262 1166
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein 1167 CaARP functions as aldo keto reductase to scavenge cytotoxic aldehydes Plant Mol 1168 Biol 90 171ndash187 1169
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a 1170 single amino acid changes the substrate specificity of dihydroflavonol 4-reductase Plant 1171 J 25 325ndash333 1172
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the 1173 key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front 1174 Plant Sci 4 1ndash11 1175
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A 1176 robust simple high-throughput technique for time-resolved plant volatile analysis in field 1177 experiments Plant J 78 1060ndash1072 1178
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional 1179 homotetrameric enzyme that catalyzes phenylalanine decarboxylation and oxidation J 1180 Biol Chem 281 23357ndash23366 1181
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of 1182 Floral Scent The Botanical Review 72 1ndash120 1183
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of 1184 Sesquiterpenes Emitted from Two Zea mays Cultivars Is Controlled by Allelic Variation 1185 of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes 1186 Plant Cell 16 1115 LP-1131 1187
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent 1188 Decarboxylase Genes Front Plant Sci 07 1ndash8 1189
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense 1190 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

37
37
compound dhurrin in sorghum Science 354 890ndash893 1191 Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-1192
induced tyrosine decarboxylase from Arabidopsis thaliana FEBS Lett 583 1895ndash1900 1193 Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation 1194
evolution of plant specialized metabolism Curr Opin Struct Biol 47 105ndash112 1195 Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A 1196
Cornu D and Jouanin L (1995) Toxicity to Chrysomela tremulae (Coleoptera 1197 Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol 1198 Breed 1 319ndash328 1199
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) 1200 Expression profiling and functional analysis of Populus WRKY23 reveals a regulatory 1201 role in defense New Phytol 184 48ndash70 1202
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) 1203 Mechanisms of action for 2-phenylethanol isolated from Kloeckera apiculata in control of 1204 Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14 1205
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor 1206 Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against 1207 Herbivores Plant Physiol 146 888ndash903 1208
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and 1209 defensive signals in plants Some metabolites serve both roles BioEssays 37 167-174 1210
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of 1211 hydroxycinnamic acid amide in plants J Plant Biol 58 156ndash163 1212
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB 1213 (2014) The timing of herbivore-induced volatile emission in black poplar (Populus nigra) 1214 and the influence of herbivore age and identity affect the value of individual volatiles as 1215 cues for herbivore enemies BMC Plant Biol 14 1216
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 1217 2 In K Wang ed (Humana Press Totowa NJ) pp 143ndash151 1218
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism 1219 Challenges and prospects in the genomics era Am J Bot 105 959ndash962 1220
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and 1221 Nishioka T (2009) Accumulation of hydroxycinnamic acid amides induced by 1222 pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis 1223 thaliana Planta 230 517ndash527 1224
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM 1225 (2001) Induction of hydroxycinnamoyl-tyramine conjugates in pepper by Xanthomonas 1226 campestris a plant defense response activated by hrp gene-dependent and hrp gene-1227 independent mechanisms Mol Plant Microbe Interact 14 785ndash792 1228
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation 1229 in dhurrin biosynthesis Phytochemistry 69 88ndash98 1230
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D 1231 and Horvaacuteth G V (2000) A novel aldosealdehyde reductase protects transgenic 1232 plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash1233 446 1234
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced 1235 condensed tannin synthesis Cloning and expression of dihydroflavonol reductase from 1236 trembling aspen (Populus tremuloides) Plant J 32 701ndash712 1237
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J 1238 Bot 85 1111ndash1126 1239
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) 1240 interacting with forest tent caterpillars (Malacosoma disstria) Normalized and full-length 1241 cDNA libraries expressed sequence tags and a cDNA microarray for the study of 1242 insect-induced defences Mol Ecol 15 1275ndash1297 1243
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system 1244 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

38
38
Oecologia 109 414ndash426 1245 Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M 1246
and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral 1247 scent compound from L-phenylalanine by two key enzymes a PLP-dependent 1248 decarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 1249 2408 1250
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) 1251 Cloning and expression of a potato cDNA encoding hydroxycinnamoyl-CoA Tyramine 1252 N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280 1253
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of 1254 hydroxycinnamoyltyramines in cell suspension cultures of Solanum tuberosum Planta 1255 205 51ndash55 1256
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as 1257 multi-tasking soldiers involved in diverse plant metabolic processes and stress defense 1258 A structure-function update J Plant Physiol 179 40ndash55 1259
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) 1260 Cloning and Functional Verification of Genes Related to 2-Phenylethanol Biosynthesis 1261 in Rosa rugosa Genes 9 576 1262
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized 1263 Chameleons in General and Specialized Plant Metabolism Mol Plant 11 95ndash117 1264
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 1265 Molecular evolutionary genetics analysis version 60 Mol Biol Evol 30 2725ndash2729 1266
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) 1267 Tomato aromatic amino acid decarboxylases participate in synthesis of the flavor 1268 volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash1269 8292 1270
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato 1271 phenylacetaldehyde reductases catalyze the last step in the synthesis of the aroma 1272 volatile 2-phenylethanol Phytochemistry 68 2660ndash2669 1273
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) 1274 Biochemical Evaluation of the Decarboxylation and Decarboxylation-Deamination 1275 Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash1276 2387 1277
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) 1278 Investigation of a substrate-specifying residue within Papaver somniferum and 1279 Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43 1280
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) 1281 Diverse functional evolution of serine decarboxylases identification of two novel 1282 acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant 1283 Biol 14 247 1284
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) 1285 Structural basis for independent origins of new catalytic machineries in plant AAAD 1286 proteins bioRxiv 404970 1287
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway 1288 Elucidation and Heterologous Reconstitution of Rhodiola Salidroside Biosynthesis Mol 1289 Plant 11 205ndash217 1290
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp 1291 Gray) Science 313 1596ndash1605 1292
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of 1293 vegetative volatiles in plant defense against herbivores Curr Opin Plant Biol 12 479ndash1294 485 1295
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell 1296 Suspension Cultures Enzyme 414ndash420 1297
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation 1298 httpsplantphysiolorgDownloaded on March 22 2021 - Published by
Copyright (c) 2020 American Society of Plant Biologists All rights reserved
39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

39
39
to improve next-generation sequencing data analysis and genome editing efficiency in 1299 Populus tremula times alba 717-1B4 Tree Genet Genomes 11 1300
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and 1301 Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of 1302 [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during flower 1303 opening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 1304 943ndash947 1305
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) 1306 NADPH-dependent reductases involved in the detoxification of reactive carbonyls in 1307 plants J Biol Chem 286 6999ndash7009 1308
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs 1309 for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-1310 coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301 1311
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect 1312 plant responses to herbivory New Phytol 204 315ndash321 1313
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) 1314 Attraction of two lacewing species to volatiles produced by host plants and aphid prey 1315 Naturwissenschaften 92 277ndash281 1316
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B 1317 (2011) Antityrosinase and antimicrobial activities of 2-phenylethanol 2-1318 phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302 1319
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Figure 1 Proposed pathways for the biosynthesis of 2-phenylethanol in poplar
AAAT aromatic amino acid transaminase AADC aromatic amino acid decarboxylase
CYP79 cytochrome P450 family 79 enzyme AAS aromatic aldehyde synthase PPDC
phenylpyruvic acid decarboxylase MAO monoamine oxidase TOX transoximase PAR
2-phenylacetaldehyde reductase UGT UDP-glucuronosyltransferase β-Glu β-
glucosidase Solid lines indicate well-established reactionsenzymes while dashed lines
indicate reactions hypothesized in the literature
phenylpyruvic acid
2-phenylethylamine
2-phenylacetaldehyde
AAS PAR
AAAT PPDC
AADC MAO
TOX CYP79
(EZ)-phenylacetaldoxime
2-phenylethyl-β-D-glucopyranoside
2-phenylethanol
L-phenylalanine
UGT β-Glu
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Figure 2 Herbivore-damaged leaves of Populus trichocarpa accumulate and
release phenylalanine-derived metabolites The accumulation of L-phenylalanine
(A) phenylpyruvic acid (B) 2-phenylethylamine (C) and 2-phenylethyl-β-D-
glucopyranoside (D) and the emission of 2-phenylethanol (E) were analyzed in
Lymantria dispar damaged (herb) and undamaged control (ctr) leaves Plant material
was extracted with methanol and analyzed via LC-MSMS (A - D) Volatiles were
collected for 8 hours and analyzed via GC-FID (E) Means plusmn SE are shown (A - D n
= 10 E n = 9) Asterisks indicate statistical significance as assessed by Studentacutes t-
test or Mann-Whitney Rank Sum Tests L-phenylalanine (P = 0530 t = -064)
phenylpyruvic acid (P = 0772 t = -0294) 2-phenylethylamine (P lt 0001 T = 55) 2-
phenylethyl-β-D-glucopyranoside (P = 0011 T = 71) 2-phenylethanol (P = 0005 T =
585) DW dry weight ns not significant nd not detected
0
1
2
3
4
Em
issio
n
(ngg
FW
h)
ctr herb
nd
2-phenylethylamine C
ctr herb
0
1
2
3
4
5
6
Co
nc
en
trati
on
(microgg
DW
)
2-phenylethyl-β-D-
glucopyranoside D
ctr herb
0
100
200
300
400
500
Co
ncen
trati
on
(microgg
DW
)
E 2-phenylethanol
ns
ctr herb 0
10
20
30
40
50
L-phenylalanine
Co
nc
en
trati
on
(microg
g D
W)
A
ns
ctr herb
0
1
2
3
4
5
6
phenylpyruvic acid
Co
nc
en
trati
on
(microgg
DW
)
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Figure 3 Dendrogram analysis of aromatic amino acid decarboxylase family
genes from Populus trichocarpa and characterized AADC AAS and GluSerHis-
decarboxylase genes from other plants Substrates of each clade are indicated in
parentheses The tree was inferred by using the maximum likelihood method and n =
1000 replicates for bootstrapping Bootstrap values are shown next to each node The
tree is drawn to scale with branch lengths measured in the number of substitutions per
site Genes described in this study are shown in red and bold with corresponding gene
identifiers (italics) Accession numbers are given in Supplemental Table 1
CaTDC1
CaTDC2
OpTDC
CrTDC
PtAAS1 (Potri013G052900)
PtAADC1 (Potri013G052800)
PtAADC2 (Potri016G114300)
PtAAS2 (Potri002G255600)
AtAAS
AtTyrDC
RhAADC
RhPAAS
PtAADC3 (Potri004G036200)
PcTyrDC2
PcTyrDC4
PsTyrDC9
TfTyrDC13
PhPAAS
LeAADC1A
LeAADC1B
LeAADC2
SlHDC
CaAAS
MtAAS
AtSDC
BnSDC
PtSDC2 (Potri002G069800)
PtSDC1 (Potri005G190500)
PhGAD
AtGAD1
AtGAD2
PhPPY-AT
84
100
100
100
99
99
99
98
85
46
100
51
47
100
100
100
100
91
98
77
92
88
100
97
49
51
95
64
93
02
AADCaromatic
aldehyde synthase
(AAS)
(PheTyr)
amino acid
decarboxylase
(SerHis)
+
AAS
amino acid
decarboxylase
(Glu)
aromatic amino acid
decarboxylase (AADC)
(Trp)
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Rela
tive a
bu
nd
an
ce
(TIC
x 1
00
00)
Retention time (min)
A 2-phenylacetaldehyde
0
2
4
6
106 108 110 112
8
10
12
14 PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F)
PtAAS1
PtAAS1
(F338Y)
PtAADC1
PtAADC1
(Y338F) R
ela
tive
in
ten
sit
y
(cps x
100
00
0)
2-phenylethylamine
Retention time (min)
1
10
20
30
10 30 50 70
40
50
60
80
Figure 4 PtAAS1 and PtAADC1 are interconvertible by the mutation of
a single amino acid and are likely derived by tandem gene duplication
(A)The recombinant proteins PtAAS1 PtAADC1 and the corresponding
mutant enzymes PtAAS1 (F338Y) and PtAADC1 (Y338F) were incubated
with L-phenylalanine in the presence of PLP Enzyme products were
analyzed using GC-MS (2-phenylacetaldehyde) and LC-MSMS (2-
phenylethylamine) (B) PtAADC1 and PtAAS1 form a gene cluster on
chromosome 13 Both genes are oriented in the same direction and there are
no coding regions between them Sequence data were retrieved from the P
trichocarpa genome assembly version 30 (wwwphytozomenet Tuskan et
al 2006)
chromosome 13
PtAADC1 PtAAS1
coding region
untranslated region
B
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Figure 5 Overexpression of PtAAS1 PtAADC1 and PtAADC3 in Nicotiana
benthamiana alters the emission of 2-phenylethanol (A) and the accumulation of
2-phenylethylamine (B) tyramine (C) tryptamine (D) and 2-phenylethanol-β-D-
glucopyranoside (E) N benthamiana plants were infiltrated with Agrobacterium
tumefaciens carrying either PtAAS1 or PtAADC1 Five days after infiltration the
headspaces of plants were collected and analyzed via GC-MS and GC-FID Additionally
plants were harvested extracted with methanol and metabolite accumulation was
analyzed via LC-MSMS Different letters above each bar indicate statistically significant
differences as assessed by Kruskal-Wallis One Way ANOVA and Tukey tests 2-
phenylethanol (H = 19867 P le 0001) 2-phenylethylamine (H = 1694 P le 0001)
tyramine (H = 18267 P le 0001) tryptamine (H = 18645 P le 0001) 2-phenylethyl-β-
D-glucopyranoside (H = 216 P le 0001) Means plusmn SE are shown (n = 6) FW fresh
weight
Co
nc
en
trati
on
(microgg
FW
)
2-phenylethylamine B
Co
nc
en
trati
on
(ngg
FW
)
Co
ncen
trati
on
(microgg
FW
)
2-phenylethyl-β-
D-glucopyranoside
E
Co
ncen
trati
on
(ngg
FW
)
tryptamine D
tyramine C 2-phenylethanol A
Em
iss
ion
(ngh
pla
nt)
0
200
400
600
800
a a a
b
0
02
04
06
08
10
12
a a a
b
0
50
100
150
200
250
300
350
a a a
b
0
20
40
60
80
100
b b
a
b
0
2
4
6
8
10
a a
c
b
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Figure 6 Transcript accumulation of AAAD and SDC genes in
Lymantria dispar-damaged and undamaged Populus
trichocarpa leaves Gene expression in herbivore-damaged
(herb) and undamaged (ctr) leaves was analyzed by Illumina
sequencing and mapping the reads to the transcripts of the P
trichocarpa genome version v30 Expression was normalized to
RPKM Significant differences in EDGE tests are visualized by
asterisks Means plusmn SE are shown (n = 4) PtAAS1 (P = 823213E-
12 weighted difference (WD) = 486618E-05) PtAAS2 (P =
0039452261 WD = -111092E-05) PtAADC1 (P = 899025E-09
WD = 156932E-05) PtAADC2 (P = 0031269756 WD =
202404E-06) PtAADC3 (P = 1 WD = -435895E-08) PtSDC1 (P
= 0576261262 WD = 161294E-06) PtSDC2 (P = 0015936012
WD = 108813E-05)
ctr
herb
Exp
ress
ion
(R
PK
M)
0
10
20
30
40
50
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Figure 7 RNAi-mediated knockdown of AADC1 in Populus times canescens Sterile
Populus times canescens calli were transformed via Agrobacterium tumefaciens Nontransgenic
trees (WT) empty vector lines (EV-2-4) and RNAi lines (RNAi-1-4) were subjected to
Lymantria dispar feeding 2-Phenylethanol was collected from the headspace and analyzed
using GC-MSFID 2-Phenylethylamine and 2-phenylethyl-β-D-glucopyranoside were
extracted with methanol from ground plant material and analyzed via LC-MSMS Biological
replicates (nb) and technical replicates (nt) of EV lines and RNAi lines were used to test for
statistical differences WT nb = 4 EV nb = 3 nt = 5 RNAi nb = 4 nt = 5 Asterisks indicate
statistical significance as assessed by Studentacutes t-test or Mann-Whitney Rank Sum Tests 2-
Phenylethylamine (P lt 0001 t = 7940) 2-phenylethyl-β-D-glucopyranoside (P = 0011 T =
547) 2-phenylethanol (P = 0509 T = 404) Medians plusmn quartiles and outliers are shown Each
data point is represented by a circle FW fresh weight ns ndash not significant
2-phenylethylamine 2-phenylethyl-β-D-glucopyranoside
Co
ncen
trati
on
(micro
gg
FW
)
0
2
4
6
8
Em
issio
n (
ngh
pla
nt)
2-phenylethanol
ns
0
1
2
3
4
Co
ncen
trati
on
(micro
gg
FW
)
0
5
10
15
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Parsed CitationsArimura GI Huber DPW and Bohlmann J (2004) Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa x deltoides) cDNA cloning functional characterization andpatterns of gene expression of (-)-germacr Plant J 37 603ndash616
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Behnke K Ehlting B Teuber M Bauerfeind M Louis S Haumlnsch R Polle A Bohlmann J and Schnitzler JP (2007)Transgenic non-isoprene emitting poplars dont like it hot Plant J 51 485ndash499
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bertoldi M Gonsalvi M Contestabile R and Voltattorni CB (2002) Mutation of tyrosine 332 to phenylalanine converts dopadecarboxylase into a decarboxylation-dependent oxidative deaminase J Biol Chem 277 36357ndash36362
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Bieri S Brachet A Veuthey JL and Christen P (2006) Cocaine distribution in wild Erythroxylum species J Ethnopharmacol 103439ndash447
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Campos L Lisoacuten P Loacutepez-Gresa MP Rodrigo I Zacareacutes L Conejero V and Belleacutes JM (2014) Transgenic Tomato PlantsOverexpressing Tyramine N -Hydroxycinnamoyltransferase Exhibit Elevated Hydroxycinnamic Acid Amide Levels and EnhancedResistance to Pseudomonas syringae Mol Plant-Microbe Interact 27 1159ndash1169
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chen XM Kobayashi H Sakai M Hirata H Asai T Ohnishi T Baldermann S and Watanabe N (2011) Functionalcharacterization of rose phenylacetaldehyde reductase (PAR) an enzyme involved in the biosynthesis of the scent compound 2-phenylethanol J Plant Physiol 168 88ndash95
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Chung SH Rosa C Scully ED Peiffer M Tooker JF Hoover K Luthe DS and Felton GW (2013) Herbivore exploits orallysecreted bacteria to suppress plant defenses Proc Natl Acad Sci 110 15728ndash15733
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Costa MA Marques J V Dalisay DS Herman B Bedgar DL Davin LB and Lewis NG (2013) Transgenic hybrid poplar forsustainable and scalable production of the commodityspecialty chemical 2-phenylethanol PLoS One 8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Danner H Boeckler GA Irmisch S Yuan JS Chen F Gershenzon J Unsicker SB and Koumlllner TG (2011) Four terpenesynthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa Phytochemistry 72897ndash908
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Hagel J and Zulak KG (2002) Hydroxycinnamic acid amide metabolism physiology and biochemistry Can J Bot 80577ndash589
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Facchini PJ Huber-Allanach KL and Tari LW (2000) Plant aromatic L-amino acid decarboxylases Evolution biochemistryregulation and metabolic engineering applications Phytochemistry 54 121ndash138
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fahey JW Zalcmann a T and Talalay P (2001) The chemical diversity and distribution of glucosinolates and isothiocyanatesamoung plants Phytochemistry 56 5ndash51
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Fan P Miller AM Liu X Jones AD and Last RL (2017) Evolution of a flipped pathway creates metabolic innovation in tomatotrichomes through BAHD enzyme promiscuity Nat Commun 8 1ndash13
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Farhi M Lavie O Masci T Hendel-Rahmanim K Weiss D Abeliovich H and Vainstein A (2010) Identification of rosephenylacetaldehyde synthase by functional complementation in yeast Plant Mol Biol 72 235ndash245
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ferreira-Silva B Lavandera I Kern A Faber K and Kroutil W (2010) Chemo-promiscuity of alcohol dehydrogenases reduction ofphenylacetaldoxime to the alcohol Tetrahedron 66 3410ndash3414
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Frost CJ Appel M Carlson JE De Moraes CM Mescher MC and Schultz JC (2007) Within-plant signalling via volatilesovercomes vascular constraints on systemic signalling and primes responses against herbivores Ecol Lett 10 490ndash8
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Galen C Kaczorowski R Todd SL Geib J and Raguso RA (2011) Dosage-Dependent Impacts of a Floral Volatile Compound onPollinators Larcenists and the Potential for Floral Evolution in the Alpine Skypilot Polemonium viscosum Am Nat 177 258ndash272
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Gonda I Bar E Portnoy V Lev S Burger J Schaffer AA Tadmor Y Gepstein S Giovannoni JJ Katzir N and LewinsohnE (2010) Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L fruit 61 1111ndash1123
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hayashi S Yagi K Ishikawa T Kawasaki M Asai T Picone J Turnbull C Hiratake J Sakata K Takada M Ogawa K andWatanabe N (2004) Emission of 2-phenylethanol from its β-D-glucopyranoside and the biogenesis of these compounds from [2H8] L-phenylalanine in rose flowers Tetrahedron 60 7005ndash7013
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hazelwood LA Daran JM Van Maris AJA Pronk JT and Dickinson JR (2008) The Ehrlich pathway for fusel alcoholproduction A century of research on Saccharomyces cerevisiae metabolism (Applied and Environmental Microbiology (2008) 74 8(2259-2266)) Appl Environ Microbiol 74 3920
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Ishida H Tomida K Sakai M Hara M and Watanabe N (2012) Functional characterization of aromatic aminoacid aminotransferase involved in 2-phenylethanol biosynthesis in isolated rose petal protoplasts J Plant Physiol 169 444ndash451
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hirata H Ohnishi T Tomida K Ishida H Kanda M Sakai M Yoshimura J Suzuki H Ishikawa T Dohra H and Watanabe N(2016) Seasonal induction of alternative principal pathway for rose flower scent Sci Rep 6
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ho SN Hunt HD Horton RM Pullen JK and Pease LR (1989) Site-directed mutagenesis by overlap extension using thepolymerase chain reaction Gene 77 51ndash59
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Hofmann F Rausch T and Hilgenberg W (1981) Preparation of radioactively labelled indole-3-acetic acid precursors J LabelCompd Radiopharm 18 1491ndash1495
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Honda K Ocircmura H and Hayashi N (1998) Identification of Floral Volatiles From Ligustrum japonicum that Stimulate Flower-Visitingby Cabbage Butterfly Pieris rapae J Chem Ecol 24 2167ndash2180
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Imai T Maekawa M Tsuchiya S and Fujimori T (1998) Field attraction of Hoplia communis to 2-phenylethanol a major volatilecomponent from host flowers Rosa spp J Chem Ecol 24 1491ndash1497
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Boeckler GA Schmidt A Reichelt M Schneider B Block K Schnitzler J-P Gershenzon JUnsicker SB and Kollner TG (2013) Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze theFormation of Volatile Aldoximes Involved in Poplar Defense Plant Cell 25 4737ndash4754
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Clavijo McCormick A Guumlnther J Schmidt A Boeckler GA Gershenzon J Unsicker SB and Koumlllner TG (2014a)Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalistcaterpillar Plant J 80 1095ndash1107
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Jiang Y Chen F Gershenzon J and Koumlllner TG (2014b) Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa) BMC Plant Biol 14 1ndash16
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Irmisch S Muumlller AT Schmidt L Guumlnther J Gershenzon J and Koumlllner TG (2015) One amino acid makes the difference Theformation of ent-kaurene and 16α-hydroxy-ent-kaurane by diterpene synthases in poplar BMC Plant Biol 15 262
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Jain D Khandal H Paul J and Debasis K (2016) A pathogenesis related-10 protein CaARP functions as aldo keto reductase toscavenge cytotoxic aldehydes Plant Mol Biol 90 171ndash187
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Johnson ET Ryu S Yi H Shin B Cheong H and Choi G (2001) Alteration of a single amino acid changes the substratespecificity of dihydroflavonol 4-reductase Plant J 25 325ndash333
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Junker A Fischer J Sichhart Y Brandt W and Draumlger B (2013) Evolution of the key alkaloid enzyme putrescine N-methyltransferase from spermidine synthase Front Plant Sci 4 1ndash11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kallenbach M Oh Y Eilers EJ Veit D Baldwin IT and Schuman MC (2014) A robust simple high-throughput technique fortime-resolved plant volatile analysis in field experiments Plant J 78 1060ndash1072
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kaminaga Y et al (2006) Plant phenylacetaldehyde synthase is a bifunctional homotetrameric enzyme that catalyzes phenylalaninedecarboxylation and oxidation J Biol Chem 281 23357ndash23366
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Knudsen JT Eriksson R and Gershenzon J (2006) Diversity and Distribution of Floral Scent The Botanical Review 72 1ndash120Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Koumlllner TG Schnee C Gershenzon J and Degenhardt J (2004) The Variability of Sesquiterpenes Emitted from Two Zea maysCultivars Is Controlled by Allelic Variation of Two Terpene Synthase Genes Encoding Stereoselective Multiple Product Enzymes PlantCell 16 1115 LP-1131
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Kumar R (2016) Evolutionary Trails of Plant Group II Pyridoxal Phosphate-Dependent Decarboxylase Genes Front Plant Sci 07 1ndash8Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Laursen T et al (2016) Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum Science 354890ndash893
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lehmann T and Pollmann S (2009) Gene expression and characterization of a stress-induced tyrosine decarboxylase fromArabidopsis thaliana FEBS Lett 583 1895ndash1900
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leong BJ and Last RL (2017) Promiscuity impersonation and accommodation evolution of plant specialized metabolism CurrOpin Struct Biol 47 105ndash112
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Lepleacute JC Bonadeacute-Bottino M Augustin S Pilate G Lecirc Tacircn VD Delplanque A Cornu D and Jouanin L (1995) Toxicity toChrysomela tremulae (Coleoptera Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor Mol Breed 1319ndash328
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Leveacutee V Major I Levasseur C Tremblay L MacKay J and Seacuteguin A (2009) Expression profiling and functional analysis ofPopulus WRKY23 reveals a regulatory role in defense New Phytol 184 48ndash70
Pubmed Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Google Scholar Author Only Title Only Author and Title
Liu P Cheng Y Yang M Liu Y Chen K Long CA and Deng X (2014) Mechanisms of action for 2-phenylethanol isolated fromKloeckera apiculata in control of Penicillium molds of citrus fruits BMC Microbiol 14 1ndash14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Major IT and Constabel CP (2008) Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals BiochemicalDiversity and Multiplicity in Defense against Herbivores Plant Physiol 146 888ndash903
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Maag D Erb M Koumlllner TG and Gershenzon J (2015) Defensive weapons and defensive signals in plants Some metabolitesserve both roles BioEssays 37 167-174
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Macoy DM Kim WY Lee SY and Kim MG (2015) Biotic stress related functions of hydroxycinnamic acid amide in plants J PlantBiol 58 156ndash163
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
McCormick AC Boeckler GA Koumlllner TG Gershenzon J and Unsicker SB (2014) The timing of herbivore-induced volatileemission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cuesfor herbivore enemies BMC Plant Biol 14
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Meilan R and Ma C (2007) Poplar (Populus spp) BT - Agrobacterium Protocols Volume 2 In K Wang ed (Humana Press TotowaNJ) pp 143ndash151
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Moghe GD and Kruse LH (2018) The study of plant specialized metabolism Challenges and prospects in the genomics era Am JBot 105 959ndash962
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Muroi A Ishihara A Tanaka C Ishizuka A Takabayashi J Miyoshi H and Nishioka T (2009) Accumulation of hydroxycinnamicacid amides induced by pathogen infection and identification of agmatine coumaroyltransferase in Arabidopsis thaliana Planta 230517ndash527
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Newman M a von Roepenack-Lahaye E Parr A Daniels MJ and Dow JM (2001) Induction of hydroxycinnamoyl-tyramineconjugates in pepper by Xanthomonas campestris a plant defense response activated by hrp gene-dependent and hrp gene-independent mechanisms Mol Plant Microbe Interact 14 785ndash792
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Nielsen KA Tattersall DB Jones PR and Moslashller BL (2008) Metabolon formation in dhurrin biosynthesis Phytochemistry 69 88ndash98
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Oberschall A Deaacutek M Toumlroumlk K Sass L Vass I Kovaacutecs I Feheacuter A Dudits D and Horvaacuteth G V (2000) A novel aldosealdehydereductase protects transgenic plants against lipid peroxidation under chemical and drought stresses Plant J 24 437ndash446
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Peters DJ and Constabel CP (2002) Molecular analysis of herbivore-induced condensed tannin synthesis Cloning and expressionof dihydroflavonol reductase from trembling aspen (Populus tremuloides) Plant J 32 701ndash712
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Philippe RN and Bohlmann J (2007) Poplar defense against insect herbivores Can J Bot 85 1111ndash1126Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Ralph S et al (2006) Genomics of hybrid poplar (Populus trichocarpa times deltoides) interacting with forest tent caterpillars(Malacosoma disstria) Normalized and full-length cDNA libraries expressed sequence tags and a cDNA microarray for the study ofinsect-induced defences Mol Ecol 15 1275ndash1297
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Roy BA and Raguso RA (1997) Olfactory versus visual cues in a floral mimicry system Oecologia 109 414ndash426Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sakai M Hirata H Sayama H Sekiguchi K Itano H Asai T Dohra H Hara M and Watanabe N (2007) Production of 2-phenylethanol in roses as the dominant floral scent compound from L-phenylalanine by two key enzymes a PLP-dependentdecarboxylase and a phenylacetaldehyde reductase Biosci Biotechnol Biochem 71 2408
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Grimm R Schmidt J Scheel D Strack D and Rosahl S (1999) Cloning and expression of a potato cDNA encodinghydroxycinnamoyl-CoAthinsp Tyramine N-(hydroxycinnamoyl)transferase J Biol Chem 274 4273ndash4280
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Schmidt A Scheel D and Strack D (1998) Elicitor-stimulated biosynthesis of hydroxycinnamoyltyramines in cell suspension culturesof Solanum tuberosum Planta 205 51ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sengupta D Naik D and Reddy AR (2015) Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plantmetabolic processes and stress defense A structure-function update J Plant Physiol 179 40ndash55
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Sheng L Zeng Y Wei T Zhu M Fang X Yuan X Luo Y and Feng L (2018) Cloning and Functional Verification of GenesRelated to 2-Phenylethanol Biosynthesis in Rosa rugosa Genes 9 576
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Soslashrensen M Neilson EHJ and Moslashller BL (2017) Oximes Unrecognized Chameleons in General and Specialized PlantMetabolism Mol Plant 11 95ndash117
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tamura K Stecher G Peterson D Filipski A and Kumar S (2013) MEGA6 Molecular evolutionary genetics analysis version 60Mol Biol Evol 30 2725ndash2729
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman D Taylor M Schauer N Fernie AR Hanson AD and Klee HJ (2006) Tomato aromatic amino acid decarboxylasesparticipate in synthesis of the flavor volatiles 2-phenylethanol and 2-phenylacetaldehyde Proc Natl Acad Sci 103 8287ndash8292
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Tieman DM Loucas HM Kim JY Clark DG and Klee HJ (2007) Tomato phenylacetaldehyde reductases catalyze the last stepin the synthesis of the aroma volatile 2-phenylethanol Phytochemistry 68 2660ndash2669
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Liu P Ding H Harich K Gillaspy G and Li J (2013) Biochemical Evaluation of the Decarboxylation andDecarboxylation-Deamination Activities of Plant Aromatic Amino Acid Decarboxylases J Biol Chem 288 2376ndash2387
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP Lazear M Von Guggenberg R Ding H and Li J (2014a) Investigation of a substrate-specifying residuewithin Papaver somniferum and Catharanthus roseus aromatic amino acid decarboxylases Phytochemistry 106 37ndash43
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-Spence MP von Guggenberg R Lazear M Ding H and Li J (2014b) Diverse functional evolution of serinedecarboxylases identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates BMC Plant Biol14 247
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Chiang Y Smith T Vicent MA and Wang Y (2018a) Structural basis for independent origins of new catalyticmachineries in plant AAAD proteins bioRxiv 404970
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Torrens-spence MP Li F Carballo V and Weng J (2018b) Complete Pathway Elucidation and Heterologous Reconstitution ofRhodiola Salidroside Biosynthesis Mol Plant 11 205ndash217
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved

Google Scholar Author Only Title Only Author and Title
Tuskan GA et al (2006) The Genome of Black Cottonwood Populus trichocarpa (Torr amp Gray) Science 313 1596ndash1605Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Unsicker SB Kunert G and Gershenzon J (2009) Protective perfumes the role of vegetative volatiles in plant defense againstherbivores Curr Opin Plant Biol 12 479ndash485
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Villegas M and Brodelius PE (1990) Hydroxycinnamoyltransferase in Plant Cell Suspension Cultures Enzyme 414ndash420Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Xue LJ Alabady MS Mohebbi M and Tsai CJ (2015) Exploiting genome variation to improve next-generation sequencing dataanalysis and genome editing efficiency in Populus tremula times alba 717-1B4 Tree Genet Genomes 11
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Watanabe S Hayashi K Yagi K Asai T Mactavish H Picone J Turnbull C and Watanabe N (2002) Biogenesis of 2-phenylethanol in rose flowers Incorporation of [2H8] L -phenylalanine into 2-phenylethanol and its β-D-glucopyranoside during floweropening of Rosa acuteHoh-Junacute and Rosa damascena Mill Biosci Biotechnol Biochem 66 943ndash947
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Yamauchi Y Hasegawa A Taninaka A Mizutani M and Sugimoto Y (2011) NADPH-dependent reductases involved in thedetoxification of reactive carbonyls in plants J Biol Chem 286 6999ndash7009
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhou X Jacobs TB Xue LJ Harding SA and Tsai CJ (2015) Exploiting SNPs for biallelic CRISPR mutations in the outcrossingwoody perennial Populus reveals 4-coumarate CoA ligase specificity and redundancy New Phytol 208 298ndash301
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu F Poelman EH and Dicke M (2014) Insect herbivore-associated organisms affect plant responses to herbivory New Phytol204 315ndash321
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu J Obrycki JJ Ochieng SA Baker TC Pickett JA and Smiley D (2005) Attraction of two lacewing species to volatilesproduced by host plants and aphid prey Naturwissenschaften 92 277ndash281
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
Zhu YJ Zhou HT Hu YH Tang JY Su MX Guo YJ Chen QX and Liu B (2011) Antityrosinase and antimicrobial activitiesof 2-phenylethanol 2-phenylacetaldehyde and 2-phenylacetic acid Food Chem 124 298ndash302
Pubmed Author and TitleGoogle Scholar Author Only Title Only Author and Title
httpsplantphysiolorgDownloaded on March 22 2021 - Published by Copyright (c) 2020 American Society of Plant Biologists All rights reserved
![Ppt0000006 [Read-Only] - GTP Kerry Corish... · 2010. 8. 17. · Title: Microsoft PowerPoint - Ppt0000006 [Read-Only] Author: Grant Created Date: 8/17/2010 7:38:37 AM](https://img.pdfslide.tips/doc/110x75/5fddfe0c53c4152caf0ac5f1/ppt0000006-read-only-gtp-kerry-corish-2010-8-17-title-microsoft-powerpoint.jpg)
![ПРЕЗЕНТАЦИЯ 2020 год.pptx [Read-Only]therapy.irkutsk.ru/doc/city/bratsk2.pdfMicrosoft PowerPoint - ПРЕЗЕНТАЦИЯ 2020 год.pptx [Read-Only] Author Farid Created](https://img.pdfslide.tips/doc/110x75/5f83d9dfc5429b226f490dbc/-2020-pptx-read-only-microsoft-powerpoint-.jpg)

![Kendall Krieg Strategic Approach to Fire Safety.ppt … Safety.pdf · Title: Microsoft PowerPoint - Kendall Krieg Strategic Approach to Fire Safety.ppt [Read-Only] Author: NMcMartin](https://img.pdfslide.tips/doc/110x75/5a727e8c7f8b9ac0538da4d7/kendall-krieg-strategic-approach-to-fire-safetyppt-wwwasasiorgpapers2005holey-fire.jpg)
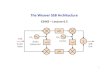
![S S Gavhane.. LIC DO Satara Mob - 9423867070 Agency... · Title: Microsoft PowerPoint - LIC ????? ?? ????? ?????.ppt [Read-Only] [Compatibility Mode] Author: SAGAR Created Date: 7/18/2020](https://img.pdfslide.tips/doc/110x75/60dcaf7fc75df105e1136f15/s-s-gavhane-lic-do-satara-mob-agency-title-microsoft-powerpoint-lic.jpg)
![001 - Energy Media Event Santovito [Read-Only]€¦ · Title: Microsoft PowerPoint - 001 - Energy Media Event_Santovito [Read-Only] Author: elicamp Created Date: 10/29/2014 3:28:49](https://img.pdfslide.tips/doc/110x75/601f66f51c91cf399301831c/001-energy-media-event-santovito-read-only-title-microsoft-powerpoint-001.jpg)

![Positive Impact Finance Principles · Maria Eugenia Sosa Taborda Sustentabilidade & Negócios Inclusivos maria.taborda@itau-unibanco.com.br. Title: Ppt0000042.ppt [Read-Only] Author:](https://img.pdfslide.tips/doc/110x75/5ec0595f11f34c3b1b71f558/positive-impact-finance-maria-eugenia-sosa-taborda-sustentabilidade-negcios.jpg)
![ProMosaik înseamnă pentru noi o lume colorată, plină de ... · Title: Microsoft PowerPoint - Ppt0000081.ppt [Read-Only] Author: popi Created Date: 3/6/2014 3:16:51 PM](https://img.pdfslide.tips/doc/110x75/5b15208d7f8b9a54488df4dc/promosaik-inseamna-pentru-noi-o-lume-colorata-plina-de-title-microsoft.jpg)

![Ppt0000001 [Read-Only]€¦ · Title: Microsoft PowerPoint - Ppt0000001 [Read-Only] Author: City Created Date: 9/12/2010 10:04:34 PM](https://img.pdfslide.tips/doc/110x75/5fbdb9ac0fd3b646a9340486/ppt0000001-read-only-title-microsoft-powerpoint-ppt0000001-read-only-author.jpg)
![02 SDK Présentation déconstruction Eric Corrignan [Read-Only] · Microsoft PowerPoint - 02_SDK_Présentation déconstruction_Eric Corrignan [Read-Only] Author: petiteville Created](https://img.pdfslide.tips/doc/110x75/5fd84f806de47573fa55bea1/02-sdk-prsentation-dconstruction-eric-corrignan-read-only-microsoft-powerpoint.jpg)
![[DRAFT ONLY]httpAssets... · Title [DRAFT ONLY] Author: Administrator Created Date: 4/18/2013 2:23:48 PM](https://img.pdfslide.tips/doc/110x75/5fe8a4b718eeda7f884d2d15/draft-only-httpassets-title-draft-only-author-administrator-created-date.jpg)



![powerpoint to print.ppt [Read-Only]lakshminarayanlenasia.com/articles/Science-Spirituality... · 2015-07-12 · Title: Microsoft PowerPoint - powerpoint_to_print.ppt [Read-Only] [Compatibility](https://img.pdfslide.tips/doc/110x75/5f2b759854643f69da3ee293/powerpoint-to-printppt-read-onlyl-2015-07-12-title-microsoft-powerpoint-.jpg)
![Desmond ESET (002) [Read-Only]desmond.ro/wp-content/uploads/2017/10/06-GDPR-28... · Title: Microsoft PowerPoint - Desmond ESET (002) [Read-Only] Author: iulia Created Date: 10/2/2017](https://img.pdfslide.tips/doc/110x75/5e512f4512e8b819c93da71a/desmond-eset-002-read-only-title-microsoft-powerpoint-desmond-eset-002.jpg)

![Ppt0000007.ppt [Read-Only] · 2019. 6. 19. · Title: Microsoft PowerPoint - Ppt0000007.ppt [Read-Only] Author: Admin Created Date: 10/26/2012 9:52:00 AM](https://img.pdfslide.tips/doc/110x75/5fd28ba38f69d706537f39e1/read-only-2019-6-19-title-microsoft-powerpoint-read-only-author-admin.jpg)
![powerpoint to print.ppt [Read-Only]libroesoterico.com/biblioteca/islam/Epoca Medieval... · Title: Microsoft PowerPoint - powerpoint_to_print.ppt [Read-Only] [Compatibility Mode]](https://img.pdfslide.tips/doc/110x75/601dc0ed4012007aa4288685/powerpoint-to-printppt-read-only-medieval-title-microsoft-powerpoint-.jpg)
![Christmas in Poland.pptx [Read-Only] - StCharles in Poland_12-01... · 2017. 1. 12. · in Poland . Title: Microsoft PowerPoint - Christmas in Poland.pptx [Read-Only] Author: paul](https://img.pdfslide.tips/doc/110x75/5fccfc4249152e16031fb840/christmas-in-read-only-stcharles-in-poland12-01-2017-1-12-in-poland.jpg)






