Embed Size (px)
Citation preview

► Meioza, gamety, rozmnožování
► Mendelovská dědičnost
► Chromosomální základy dědičnosti
► Centrální dogma molekulární biologie
Biologie I
Genetika – chromosomální a molekulární základy dědičnosti

► Přenos vlastností/predispozic rodičovského organismu na potomky
► Přenos DNA s geny (chromosomální i extrachromosomální)
► Gen = základní jednotka dědičnosti, na chromosomu umístněn v lokusu
Dědičnost
Rozmnožování
► Nepohlavní (vegetativní, asexuální) = klony (genetické kopie)
► Pohlavní (generativní, sexuální)
Potomek nese [jedinečnou] kombinaci genů získaných od obou rodičů
proces tvorby potomstva (zde ve smyslu celých organismů)
Amoeba dělící se mitoticky
100 µm
Mnohobuněčný nezmar
(Hydra) množící se pučením
(pupen: mitosy, masa
buněk se vyvijí v nového
jedince → oddělení)
např.: rodičovský
jedinec
pupen

v důsledku náhodné kombinace genů (chromosomů rodičů) při oplození se
potomek vzhledem liší od rodičů i sourozenců
Pohlavní rozmnožování a dědičnost
Homo sapiens sapiens
Somatická buňka rodiče: • 46 chromosomů (diploidní; 2n)
• 2 gonosomy (pohlavní chr.)
• 44 autosomů (22 párů)
Potomek:
46 chromosomů (2n) 2 gonosomy (pohlavní chr.)
22 párů autosomů
gameta otec 22 autosomů
+ gonosom
(haploidní; n)
gameta matka 22 autosomů
+ gonosom
(haploidní; n) Karyotyp – soubor chromosomů, rozlišitelné
podle tvaru a proužkování (uspořádaný a graficky znázorněný = karyogram)
zde muž (46, XY)
metafázní chromosomy izolované z leukocytů
po barvení (proužky)
centromera
sesterské
chromatidy
pár homologních
chromosomů
Adapto
váno z
Cam
pbell,
Reece:
Bio
logy 6
th e
ditio
n ©
Pears
on
Educatio
n,
Inc,
publis
hin
g a
s B
enja
min
Cum
min
gs

splynutí gamet (syngamie, oplození)
za vzniku diploidní zygoty
Pohlavní rozmnožování a dědičnost
gameta
Diploidní
mnohobuněčný
potomek
Barvy
MEIOZA OPLOZENÍ
n
n
n
2n 2n zygota
Haploidní stav (gamety = pohlavní buňky )
Diploidní stav
mitozy, dělení
diferenciace
gameta
2. pohlaví
mitoza
Rozmnožovací cyklus živočichů
Ad
ap
tová
no
z C
am
pb
ell,
Ree
ce
: B
iolo
gy 6
th e
ditio
n ©
Pe
ars
on
Ed
uca
tio
n, In
c,
pu
blis
hin
g a
s B
en
jam
in C
um
min
gs

Pohlavní rozmnožování a dědičnost
► Časový nástup meiozy
se v životních cyklech
různých organismů
může lišit:
Většina hub, někteří prvoci a řasy
Rostliny a některé řasy
hlenka (Dictiostelium)
haploidní mnohobuněčný
organismus
mitoza mitoza
gamety
zygota
zygota
mitoza mitoza
gamety spory
haploidní mnohobuněčný
organismus (gametofyt)
diploidní mnohobuněčný
organismus (sporofyt)
mitoza
meioza oplození
meioza oplození
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson Education, Inc, publishing as Benjamin Cummings

MEIOZA – tvorba haploidních buněk rozdělením diplodních
MEIOZA
redukční dělení
(redukce počtu
chromosomů a
separace
sesterských
chromatid)
diploidní
buňka
meiozu předchází
replikace chromosomů
výsledkem jsou
buňky nesoucí
jednu sadu
původních
homologních
autosomů
(+ gonosom)
1 pár homologních chromosomů = 2 možnosti...
pár homologních chromosomů
replikace
pár replikovaných homologních chromosomů
sesterské
chromatidy
2n
2n
n
buňky s 1 replikovaným
chromosomem
separace
chromatid
n
chromosomy
chromatidy
Ad
ap
tová
no
z J
oh
nso
n R
.: B
iolo
gy, 5
th e
ditio
n 1
99
9; ©
Th
e M
cG
raw
-Hill
Com
p.,
In
c.

Meioza I
... 2 páry homologních chromosomů = 4 možnosti...
... 23 párů chromosomů = 8 388 608 možnosti...
... 3 páry homologních chromosomů
homologní páry (není znázorněna replikace)
8 možností meiozy (ne všechny realizovány, 1 meioza = max. 4 buňky/gamety k použití)
► Počet možností při haploidním čísle n chromosomů je 2n.
► 1. Chromosomy/chromatidy se rozchází nezávisle
► 2. Oplození je nahodilé (sejde se vždy 1 kombinace každého rodiče)
a v meioze je variabilita umocněna ještě dalším způsobem
2 zdroje genetické variability potomků
Ad
ap
tová
no
z J
oh
nso
n R
.: B
iolo
gy, 5
th e
ditio
n
19
99
; ©
Th
e M
cG
raw
-Hill
Com
p.,
In
c.

► Meiozu předchází meiotická interfáze
s replikací chromosomů a zdvojením centrosomu
► Každé dělení během meiosy pak probíhá ve 4 fázích (podobně jako mitoza)
1. profáze
2. metafáze
3. anafáze
4. telofáze
► První dělení: meioza I
(tzv. heterotypické dělení – určité odlišnosti od mitozy – složitější v profázi)
► Druhé dělení: meioza II
(tzv. homeotypické dělení – obdoba mitozy)
● interkineze – období mezi meiozou I a meiozou II
MEIOZA – detailnější pohled na dvě po sobě jdoucí dělení

• Kondenzace chromosomů
• Mizí jadérka
• Vznikají kinetochory
• Párování homologních chromosomů
a vznik synapsí (těsný podélný kontakt)
• Později chromosomové páry patrné
jako tetrády (4 prokřížené chromatidy)
• V překříženích výměna homologických
částí nesesterských chromatid
(crossing-over)
• Místo překřížení = chiazma,
v tetrádě obvykle několik chiazmat
• Prodlužování mikrotubulů
• Oddalování centrosomů (=dělícího vřeténko)
• Rozpad jaderného obalu
• Napojování mikrotubulů na kinetochory
1. Profáze I – přehled Meioza I
sesterské chromatidy
dělící vřeténko
páry homologních chromosomů
s vyměněnými úseky
nesesterských chromatid
tetráda
chiazmata
PROFÁZE I
centrosom
Adapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on E
ducatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

1. Profáze I – může představova více než 90% času trvání meiozy
Meioza I
5 stádií:
i) Leptotene – chromosomy přichyceny
k jadernému obalu; homologní poblíž a
silně kondenzují
ii) Zygotene – synapse: tvorba proteino-
vého synaptonemálního komplexu mezi
homologními chromosomy
Dvojice paralelních chromosomů = bivalent
iii) Pachytene – homologické části part-
nerských chromosomů „drženy“ naproti
sobě, rozbalení DNA, crossing-over
iv) Diplotene – synaptonemální komplex
se rozpadá (chiazmata přetrvávají),
chromosomy dekondenzují
a jsou transkričně aktivní
v) Diakineze – chromosomy opět kondenzují chiasmata
(místa
crossing-over)
syn
ap
ton
em
áln
í
ko
mp
lex
synapse
sesterské chromatidy chromosomu
chromatidy
homologního
chromosomu
centromera protilehlé
nesesterské
chromatidy
crossing-over
partnerské homologní chromosomy
Adaptováno z Johnson R.: Biology, 5th edition 1999; © The McGraw-Hill Comp., Inc.

centromera (s kinetochorem)
metafázní destička
napojené kinetochorové
mikrotubuly
PROFÁZE I ANAFÁZE I
• Kinetochorové mikrotubuly jednoho pólu dělícího vřeténka
napojeny k jednomu chromosomu z páru (na jednu stranu centromery)
• chiazmata se posouvají ke koncům chromosomů = terminální chiazmata
(umožňuje oddálení centromer a usnadňuje správné napojení mikrotubulů)
•Tetrády seřazeny v metafázní destičce
2. Metafáze I
METAFÁZE I
astrosféra (živočišná buňka)
nekinetochorové mikrotubuly
chiazmata
Meioza I A
dapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on
Educatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

3. Anafáze I
• Separace homologních chromosomů (zanikají tetrády)
• Každý chromosom (dvě sesterské chromatidy)
transportován k opačnému pólu
sesterské chromatidy (zůstávají spojeny)
PROFÁZE I ANAFÁZE I ANAFÁZE I METAFÁZE I
homologní chromosomy
Meioza I A
dapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on
Educatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

TELOFÁZE I a CYTOKINEZE
• Chromosomy pokračují v rozestupu a dosahují pólů
• Každý chromosom je stále tvořen dvěmi sesterským chromatidami
• U některých druhů dekondenzace chromosomů, tvrba jádra i jadérka
• Cytokineze obvykle probíhá souřasně a tvoří se dvě haplodní buňky
Zivočišné buňky: zaškrcení
Rostlinné buňky: fragmosplast
Žádný z chromosomů se již nereplikuje
Meiosis I
Meioza II
Meioza I
Meioza II
► 3. Crossing-over mohl vyměnit části
chromatid = sesterské chromatidy
již nejsou geneticky identické
3. zdroj genetické variability potomků
zaškrcení
Adapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on
Educatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

Meioza II je velmi podobná mitoze
• Tvorba dělího aparátu
• Chromosomy
(dvě sesterské chromatidy)
kondenzované
• Všechny sesterské chromatidy správně napojeny
na kinetochorové mikrotubuly
• Sesterské chromatidy na metafázní destičce
• Sesterské chromatidy
odděleny a transportovány
k pólům
• Chromosomy
(chromatidy) na
opačných pólech
• Rozpad dělícího
aparátu, tvoří se
jádra
• Chromosomy
despiralizují
+ jadérka
Výsledek
4 dceřinné buňky
geneticky odlišné
mezi sebou a i od
mateřské buňky
A CYTOKINEZE
PROFÁZE II METAFÁZE II ANAFÁZE II TELOFÁZE I A CYTOKINEZE TELOFÁZE II
separace sesterských chromatid
zaškrcení
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson Education, Inc, publishing as Benjamin Cummings

Mendelelovská dědičnost
● mnich (od 1843) a později opat
(od 1868) v augistiánském
kláštěře sv. Tomáše v Brně
● od r. 1957 se zbýval křížením
hrachu (Pisum sativum),
dostupný v řadě odrůd
● 1865 práce
Versuche über Pflanzen-Hybride
vcelku bez povšimnutí, znovu objevena 1900
Hugo de Vries (NL)
Erich von Tschermak (A)
Carl Correns (D)
Gregor Johann Mendel (*1822 in Hynčice – †1884 in Brno)
► Křížil čisté (homozygotní) linie (P, F0);
takové křížení označováno hybridizace
► Křížil hybridy F1 (1. filiální generace)
→ F2, v níž byl poměr forem znaku z P = 3:1
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson
Education, Inc, publishing as Benjamin Cummings

důležité termíny:
Genom – kompletní genetický materiál daného organismu
Alely – alternativní formy téhož genu (dvě alely v diploidním organismu)
Homozygot – pár identických alel (čistá linie)
Heterozygot – dvě rozdílné alely jednoho genu
Genotyp – soubor všech alel v organismu
Fenotyp – fyzické a fyziologické rysy (znaky) organismu
alela pro fialové květy
alela pro bílé květy
pár homologních
chromosomů
lokus pro
barvu květu
Adapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on E
ducatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

Mendelův model dědičnosti pro dvě alely jednoho genu
homozygotní linie nesoucí dominantní alelu (P),
která se na rozdíl od recesivní alely (p) projeví („dominantní je funční“)
hybridizace
1. Zákon o uniformitě generace F1
při vzájemném křížení dvou homozygotů jsou
potomci genotypově i fenotypově jednotní
křížení tzv. monohybridů
(jedinců u kterých sledujeme 1 znak)
2. Zákon o jednoduchých štěpných poměrech v F2
při křížení heterozygotů F1 může být potomkovi
předána každá ze dvou alel se stejnou
pravděpodobností nezávislá segregace alel
→ fenotypový štěpný poměr 3:1
Mendelelovská dědičnost
homozygotní P
generace F1
generace F2
vzhled:
genotyp:
gamety:
vzhled:
genotyp:
gamety:
samičí F1 samčí F1
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson Education, Inc,
publishing as Benjamin Cummings

Punnettův čtverec
Rodiče
Ee Ee
vajíčka
sp
em
ie
Pu
nne
ttů
v č
tve
re
c
potomci
E e
E
e
Ee
Ee EE
e e
Alely Fenotypy
nepřirostlý lalůček 3
1
E = nepřirostlý lalůček
e = přirostlý lalůček přirostlý lalůček
● Heterozygotní rodiče
s nepřirostlým lalůčkem (determinováno dominantní alelou E)
● Gamety:
možnost E = ½
možnost e = ½
● Šance zdědit genotypy
EE =½×½=¼
Ee =½×½=¼
eE =½×½=¼
ee =½×½=¼
● Pravděpodobnost u novorozence
75% nepřirostlý a 25% přirostlý lalůček
(3:1)
Mendelelovská dědičnost
Adapto
váno z
Johnson R
.: B
iolo
gy,
5th e
ditio
n 1
999;
© T
he M
cG
raw
-Hill
Com
p.,
Inc.

Testovací (zpětné) křížení
odhalení genotypu křížením
s homozygotem v recesivním stavu
v případě lalůčků raději
Rozbor rodokmenu
pro analýzu rizik u nemocí
děděných jako jednoduché
mendelovské znaky
Mendelelovská dědičnost
přirostlý lalůček nepřirostlý lalůček
rod
ok
me
n
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson Education, Inc, publishing as Benjamin Cummings
dominantní fenotyp;
neznámý genotyp
(možno PP nebo Pp)
recesivní fenotyp;
známý genotyp pp
pro možnost PP
bude fenotyp fialový květ
pro možnost Pp
50% fialový květ
50% bílý květ
hybrid F1 homozygot
P

► neúplná dominance
homozygotní P
generace F1
generace F2
samičí F1 samčí F1
gamety
gamety
červená
CRCR
bílá
CWCW
růžová
CRCW
Adapto
váno z
Cam
pbell,
Reece:
Bio
logy 6
th e
ditio
n ©
Pears
on E
ducatio
n,
Inc, publis
hin
g a
s B
enja
min
Cum
min
gs
CR: alela barva červená
Cw: alela barva bílá
jen 1 alela CR
genotyp F1 se projeví v F2
Štepný poměr 1:2:1
(není třeba zpětné křížení)

3. Zákon o čistotě a plné kombinovatelnosti vloh
vlohy pro znaky a vlastnosti se přenášejí z generace na generaci
v čisté formě a vzájemně se nemísí (nezávislá segregace alel)
Mendelelovská dědičnost
sledujeme 2 znaky současně (oba s možností dominance):
křížení tzv. dihybridů
(sledujeme 2 znaky)
→ fenotypový štěpný poměr 9:3:3:1
(zpětné křížení s yyrr poměr 1:1:1:1)
homozygotní P
generace F1
generace F2
samčí F1
gamety gamety
samičí F1
štěpný poměr 3:1 jako
u jednoho znaku
nebyl pozorován
alely segregují nezávisle
žluté, kulaté
zelené, kulaté
žluté, scvrklé
zelené, scvrklé
Adapto
váno z
Cam
pbell,
Reece:
Bio
logy 6
th e
ditio
n ©
Pears
on E
ducatio
n,
Inc, publis
hin
g a
s B
enja
min
Cum
min
gs

dihybrid
AaBb
A
A A
A A
A
A
AB
ab
Ab
aB
a
a
a
a
a a
a
A
A b A
a a a
B
B
B B
B
B
B
B
B
B a
B
b
b b
A A
b b B B
a a
b
b
b
b
b
A
b
b
► štěpný poměr platí, pokud jsou geny na různých chromosomech
metafáze I dvě možnosti
uspořádání
chromosomů
možné gamety alely
mendelovsky
rozděleny
metafáze II
Mendelelovská dědičnost A
dapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on E
ducatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

Fenotypy po křížení
F1 … všichni červené oči
F2 … červené:bílé = 3:1 (štěpný poměr odpovídá mendelovské dědíčnosti)
ALE: bílé oči pouze u samečků →
→ znak vázaný na chromosom X
Chromosomální základ dědičnosti
Thomas Hunt
Morgan
(1866 – 1945)
Drosophila melanogaster
sameček samička
divoký typ mutant
XY XX
homozygotní
P
generace
F1
vajíčka
generace
F2
spermie
vajíčka spermie
Adaptováno z Campbell, Reece: Biology 6th edition ©
Pearson Education, Inc, publishing as Benjamin Cummings

další Morganův experiment:
Vazba vloh / genů na libovolném chromosomu
Znaky: barva těla a vývoj křídel
Alely:
b+ : normální šedé tělo
b : černé tělo (black)
vg+ : normální křídla
vg : zakrnělá (vestingal) křídla
► Převahu rodičovských fenotypů
vysvětluje vazba genů na jednom
chromosomu.
ale proč pak není poměr 1:1:0:0?
rodiče v
zpětném
křížení
většina
potomků
nebo
homozygotní
P
divoký typ
šedé tělo,
normální křídla
dvojitý mutant
černé tělo,
zakrnělá křídla
dihybrid F1
šedé tělo,
normální křídla
dvojitý mutant
černé tělo,
zakrnělá křídla
ZPĚTNÉ KŘÍŽENÍ
potomstvo
po zpětném
křížení
očekávání:
nezávyslá
segregace
SKUTEČNOST:
rodičovský fen. rekombinovaný fen.
šedé /
normální
černé /
zakrnělá
šedé /
zakrnělá
černé /
normální
fenotyp
tělo / křídla
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson Education, Inc, publishing as Benjamin Cummings

Rekombinantní
fenotypy
jsou důsledkem
crossing-over
Frekvence
rekombinace
gamety
dihybrid F1
šedé tělo,
normální křídla
dvojitý mutant
černé tělo,
zakrnělá křídla
ZPĚTNÉ
KŘÍŽENÍ
rodičovský fenotyp rekombinovaný fenotyp
replikace
MEIOZA I
a crossing-over
MEIOZA II
separace
chromatid
MEIOZA I a II
crossing-over
neutrální
rekombinantní
chromosomy
vajíčko spermie
vajíčko
spermie
potomci
křížení
(391 / 2300) x 100 = 17%
Adapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on E
ducatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs
Fenotypy (tělo - křídla):
1. gray – normal
šedé – normální
2. Black – vestingal
černé – zakrnělá
3. Gray – vestingal
šedé – zakrnělá
4. Black – normal
černý– normální
{
{
83%
17%

Morganovy zákony
1. Geny jsou lineárně uspořádány v chromosomech.
2. Geny jednoho chromosomu tvoří vazebnou skupinu.
Počet vazebných skupin = počet párů homologních chromosomů
3. Mezi geny homologického páru chromosomů může probíhat
genová výměna prostřednictvím crossing-over.
Frekvence crossing over je přímo úměrná vzdálenosti genů.
případ třech genů na chromosomu II
b, vg a cn (cinnabar, rumělka)
fenotyp cn: světlejší oči než
divoký typ
Frekvence rekombinací:
9% pro cn a b
9,5% pro cn a vg
17% b a vg
Vzdálenější geny – větší šance, že
mezi nimi proběhne crossing-over
chromosom
frekvence
rekombinace
Adaptováno z Campbell, Reece: Biology 6th edition ©
Pearson Education, Inc, publishing as Benjamin Cummings

dihybrid
AaBb
A
A A
A A
A
A
AB
ab
Ab
aB
a
a
a
a
a a
a
A
A b A
a a a
B
B
B B
B
B
B
B
B
B a
B
b
b b
A A
b b B B
a a
b
b
b
b
b
A
b
b
► štěpný poměr platí, pokud jsou geny na různých chromosomech
metafáze I dvě možnosti
uspořádání
chromosomů
možné gamety alely
mendelovsky
rozděleny
metafáze II
► a jedním genem je determinován jeden fenotyp neovlivněný prostředím
Mendelelovská dědičnost A
dapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on E
ducatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

Pleiotropie
Kodominance – vícealelové systémy vs 2n
• Gen ovlivní více než jeden fenotypový znak jedince
Případ dědičnosti krevních skupin:
3 alely pro dědičnost krevních skupin:
Alela IA - antigen A na povrchu erythrocytů
- protilátky proti B v séru (anti-B)
Alela IB - antigen B na povrchu erythrocytů
- protilátky proti A v séru (anti-A)
Alela i - erythrocyt bez antigenu A nebo B
- v séru protilátky proti A i B
- 6 možných genotypů
- fenotypy krevní skupina A, B, AB nebo 0
• IA a IB jsou vůči sobě kodominantní
• IA a IB jsou vůči alele i dominantní
Případ fenylketourie: - gen pro fenylalanin hydroxylasu na autosomu
- mutace = recesivní alela; autosomálně recesivní onemocnění
- porucha přeměny aminokyseliny fenylalaninu na tyrosin
Fenotypové projevy: - mentální retardace, epileptické záchvaty
- bledá pleť, modré oči, světlé vlasy (nedostatek tyrosinu pro tvorbu melaninu)
- časté ekzémy a vyrážky
krevní
skupina
geno-
typy sérum
erythrocyty po přidání séra
krevních skupin ve sloupci
Adaptováno z Campbell, Reece: Biology 6th edition © Pearson Education, Inc, publishing as Benjamin Cummings

Polygenní dědičnost
Pozn. pro znaky, měnící se v populaci plynule = znaky kvantitativní – vělikost těla, barva kůže
(rozdíl proti kvalitativním znakům = diskrétní znaky – např. krevní skupina)
Vliv prostředí na fenotyp
Genové interakce
• Účinky více genů na jednu fenotypovou vlastnost
• Fenotyp je dán součtem účinků genů (aditivní, kumulativní efekt)
norma reakce = různost fenotypů, které mohou vzniknout
z jediného genotypu vlivem vnějšího prostředí
norma reakce nulová – např. krevní skupiny, barva květu hrachu
norma reakce nenulová – např. barva květu hortenzie
zásaditá kyselá
půda
• Na sledovaném kvalitativním znaku se podílí více než jeden gen
• Alely dominantní a recesivní

Mendelelovská dědičnost
→ fenotypový štěpný poměr 9:3:3:1
(zpětné křížení s yyrr poměr 1:1:1:1)
homozygotní P
generace F1
generace F2
samčí F1
gamety gamety
samičí F1
štěpný poměr 3:1 jako
u jednoho znaku
nebyl pozorován
alely segregují nezávisle
žluté, kulaté
zelené, kulaté
žluté, scvrklé
zelené, scvrklé
Adapto
váno z
Cam
pbell,
Reece:
Bio
logy 6
th e
ditio
n ©
Pears
on E
ducatio
n,
Inc, publis
hin
g a
s B
enja
min
Cum
min
gs

► interakce bez změny štěpných poměrů 9:3:3:1 – reciproká interakce
oba geny se projevují samostatně, interakce až na úrovni fenotypu
Barva plodu papriky:
Alely: R – červený pigment ANO
r – červený pigment NE
Cl – rozklad chlorofylu ANO
cl – rozklad chlorofylu NE
(pomlčky v obr. = jakákoliv alela)
objevují se fenotypy, které nebyly
u rodičovských rostlin (P1 nebo P2)
(pomlčka = jakákoliv alelu)
výsledek zpětného křížení s rrclcl je 1:1:1:1
x x
Genové interakce
Převzato z Nečásek a kol. Obecná genetika. SPN Praha 1979

► dominantní epistaze – dominantní alela epistatického genu potlačí projev
hypostatického genu (obdoba dominance)
Barva květů jiřiny:
Alely: Y – žlutý pigment ANO
I – pigment ‘slonová kost’ ANO
y a i – pigment NE
štěpný poměr v F
2 12:3:1
přítomnost žlutého pigmentu překryje
barvu slonové kosti
výsledek zpětného křížení s yyii je 2:1:1
Genové interakce
Převzato z Nečásek a kol. Obecná genetika. SPN Praha 1979

► recesivní epistaze – gen v homozygotně recesivní stavu potlačí/neumožní
projev alely druhého genu
Barva květů šalvěje:
Alely: P / p – červený pigment ANO / NE
A / a – methylace červeného
pigmentu = fialová ANO / NE
štěpný poměr v F
2 9:3:4
výsledek zpětného křížení s ppaa je 1:1:2
Genové interakce
Převzato z Nečásek a kol. Obecná genetika. SPN Praha 1979

► komplementarita – alespoň jedna dominantní alela obou genů
Barva květů hrachoru:
Alely: C a R – červený pigment ANO
produkt C je stále bezbarvý
štěpný poměr v F
2 9:7
výsledek zpětného křížení s ccrr je 1:3
Genové interakce
Převzato z Nečásek a kol. Obecná genetika. SPN Praha 1979

► inhibice – dominantní alela “inhibitoru” zamezí projevu druhého genu
(inhibitor sám jiný projev nemá)
Barva peří kura domácího:
Alely: C / c – červený melanin ANO / NE
I / i – inhibice depozice melaninu
v keratinu peří ANO / NE
štěpný poměr v F
2 13:3
výsledek zpětného křížení s ccii je 3:1
Genové interakce
Převzato z Nečásek a kol. Obecná genetika. SPN Praha 1979

► duplicita (nekumulativní) – dva geny téhož účinku
a postačuje jedna dominantní alela
Tvar šešule kokošky patuší tobolky:
Alely: T1 nebo T2 – zajistí normální tvar
t1 a t2 v homozygotně recesivním
stavu – špičatý tvar
štěpný poměr v F
2 15:1
výsledek zpětného křížení s ccii je 3:1
Genové interakce
Převzato z Nečásek a kol. Obecná genetika. SPN Praha 1979

Polygenní dědičnost
Zjednodušený model polygenní dědičnosti barvy kůže:
3 geny:
- alely A, B, C – tmavá kůže (v obr. černé tečky bez rozlišení alel)
- alely a, b, c – světlá kůže
(v obr. bílé tečky bez rozlišení alel)
- vztah neúplné dominance
například zbarvení
AaBbCc = AABbcc
barvu kůže ovlivní
i vlivy prostředí
(→spojitá Gausova křivka)
Adapto
váno z
Cam
pbell, R
eece: B
iolo
gy 6
th editio
n ©
Pears
on E
ducatio
n, In
c, p
ublis
hin
g a
s B
enja
min
Cum
min
gs

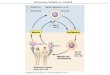
Centrální dogma molekulární biologie
3' 5'
A G G G A C C C C
T C G C T G G G G
5' 3'
templátový řetězev
(pro transkriptci) transkripce
3' 5'
mRNA
DNA
A G G G A C C C C
codon 1 codon 2 codon 3
polypeptid
translace
N N N C C C C C
R1 R2 R3
Serin Asparagin Prolin
O O O
replikace
+ r
RN
A &
tR
NA

Replikace je semikonzervativní
Každý řetězec původní dvojšroubovice
slouží jako templát (matrice) pro syntézu
nového komplementárního vlákna
Nové řetězce jsou syntetizovány jako
Vedoucí řetězec (leading strand):
kontinuální molekula
Opožďující se řetězec (lagging s.):
diskontinuální syntéza krátkých DNA
(Okazakiho fragmenty); pro dokončení
celého vlákna - degradace primerů RNA,
dosyntetizování DNA a spojení (ligace)
G C
A T
G C
G C
P
P
P
P
P
P
P
P
P
P
P
C
C C
C
H H
H
H H
OH
OH O
3
konec
5 konec
3 konec
templátový řetězec nový řetězec
primer RNA
3
5
3
5
5
helikasa rozbaluje dsDNA
= replikační vidlice
leading
strand
lagging strand
DNA polymerasa
DNA ligasa Okazakiho
fragment
5
7
6
4
3
2
1 4
3 2
1
5
konec
3
DNA polymerasa připojí
další nukleotid na 3’
uhlík (-OH) ribosy
nukleotidu předchozího.
DNA polymerasa
3
původní řetězec
(templát)
původní
DNA
● Nový řetězec syntetizován
na základě komplementarity bazí
● Replikaci katalyzuje
DNA polymerasa
(v počátku replikace vyžaduje primer RNA
poskytující 3’OH ribosy)
● DNA se rozbalí a dojde
k oddělení vláken
Nerozbalené
dvojšroubovice
DNA
G
G
G
T
A
A
C
C
3' 5'
A T
C G
A T A
G
G C C G
A
oblast
dokončené
replikace původní
řetězec
nový
řetězec
C C A A
T T
G G
T A
T A
C G
A T
A T
A
C G A T A
T A
T A
G C
G A
G
T
A
C
G
C
G
A
oblast
probíhající
replikace
C

počátek replikace
replikace
probíhá oběma
směry
replikace
ukončena
replikační vidlice replikační bublina
parentální řetězec
nový řetězec
2 nové
duplexy
DNA
Replikace:
Prokaryota
Eukaryota
● jeden počátek replikace
na cirkulárním chromosomu
● jedna „replikační bublina“
rostoucí oběma směry
● mnoho počátků replikace
na lineárním chromosomu
● replikační vidlice rostou
oběma směry a tvoří replikační
bublinu
● po setkání vidlic/bublin jsou
nové řetězce kovalentně spojeny
versus

Transkripce
netemplátový
řetezec DNA templátový
řetězec DNA
5'
C
C G
T A
A T
G G
G
A
C
C
A
G RNA
polymerasa
RNA
(transcript)
C
A T
C G
T A
RNA může být
dále procesována
(upravena)
3
'
3'
5'
● Genetická informace je přepsána
(transkribována) do sekvence RNA
● Přepis začíná sestavením komplexu RNA
polymerasy na promotoru genu a oddělením
vláken DNA (promotor: specifická sekvence
před přepisovanou oblastí)
● Přepis končí terminací transkripce
(specifické sekvence nebo proteinové faktory)
● Transkripce genu není nahodilá
- konstitutivní – stálá (housekeeping geny)
nebo může
- podléhat indukci
- podléhat represi

Transkripce do sekvence mRNA:
Prokaryota
Eukaryota
● mRNA je v podstatě připravena na translaci bez prav
● možnost kontinuální polycistronní mRNA:
transkripce operonu (tandemně uspořádané
kódující sekvence několika proteinů pod
kontrolou jednoho promotoru) versus
● Pre-mRNA obsahuje kódující sekvence (exony) a
intervenující nekódující sekvence (introny)
● Transkript je dále vybaven nekódovanými nukleotidy
(na 3‘ konci nukletidem tvořícím „čepičku“ a
na 5’ konci je modifikován polyadenylací
● Introny jsou vystřiženy (sestřih RNA, splicing)
a exony spojeny v kontinuální kódující řetězec
introns occuring in between exons are to be
excised through RNA splicing
● Se střih probíhá v jádře, katalyzován
nukleoproteinovým komplexem (spliceosomem)
exon
intron intron
exon exon DNA
transkripce
exon
intron intron
exon exon
5' 3' pre-mRNA
exon exon exon
intron intron čepička poly-A 5' 3'
exon exon exon
spliceosome
čepička poly-A
sestřih
pre-mRNA
intron RNA
5' 3'
čepička poly-A
mRNA
jaderný pór
jádro
cytoplasma
5' 3'

Translace na ribosomu
amino
kyselina
3
5
vodíkové
můstky
anti-
kodon
mRNA
5' kodon
3'
2. báze 3.
báze
1.
báze U C G A
U
C
A
G
UUU
fenylalanin
UCU
serin
UAU
tyrosin
UGU
cystein
UUC
fenylalanin
UCC
serin
UAC
tyrosin
UGU
cystein
UCA
serin UUA
leucin
UCG
serin
UUG
leucin UGG
tryptofan
UGA
stop
UAA
stop
UAG
stop
U
C
A
G
CUU
leucin
CUC
leucin
CUA
leucin
CUG
leucin
CCU
prolin
CCC
prolin
CCA
prolin
CCG
prolin
CAC
histidin
CAA
glutamin
CAG
glutamin
CAU
histidin
CGA
arginin
CGG
arginin
CGU
arginin
CGC
arginine
U
C
A
G
AUG (start)
methionin
AUU
isoleucin
AUC
isoleucin
AUA
isoleucin
ACU
threonin
ACC
threonin
ACA
threonin
ACG
threonin
AAU
asparagin
AAC
asparagin
AAA
lysin
AAG
lysin
AGU
serin
AGC
serin
AGA
arginin
AGG
arginin
U
C
A
G
GUU
valin
GUC
valin
GUA
valin
GUG
valin
GCU
alanin
GCC
alanin
GCA
alanin
GCG
alanin
GAU
aspartát
GAC
aspartát
GAA
glutamát
GAG
glutamát
GGU
glycin
GGC
glycin
GGA
glycin
GGG
glycin
U
C
A
G
molekula tRNA má 2 vazebná místa:
● antikodon (H-vazby, komplementarita)
● místo pro vazbu aminokyseliny (kovaletní vazba)
● kodon tvořen
3 bázemi
● 64 možných
kombinací:
● 3 kodony jako
info pro STOP
translaci
● 61 jako info o
identitě
připojované
aminokyseliny
● prerekvizity: ribosomy, mRNA, tRNA, energie, aminokyseliny a info o jejich pozici

Malá podjednotka ribosomu
váže mRNA; iniciační tRNA
(pro Met) se váže na iniciační
Kodon AUG.
Připojuje se velká ribosomální
podjednotka; iniciační tRNA
vázána v místě P; místo A
připraveno pro vazbu
další tRNA.
1. Iniciace (první kodon obvykle AUG)
Met
methionin
tRNA
mRNA
malá
velká
podjednotka ribosomu
3'
5'
místo P
místo A místo E
Met
U A C A U G
start kodon 5' 3'
3. Terminace
Uvolňovací faktor štěpí vazbu
mezi polypeptidem a poslední
tRNA – protein uvolněn.
3
5
Ribosom přeložil mRNA
až po STOP – není tRNA
váží se terminační faktory
U A
U A U G A
stop codon 5' 3'
Asp
Ala
Trp
Val Glu
terminační
(uvolňovací)
faktory
Do místa A přichází
tRNA–aminokyselina
Dvě tRNAs jsou na
ribosomu současně
(místo P a A)
Tvoří se
peptidová vazba
Posun mRNA a “prázdná”
tRNA opouští ribosom z
místa E (uvolněno místo A)
1 2 3 4
2. Elongace
peptidová
vazba Met
Ala
Trp
Ser
Val
U A
C A U G G A C
3 3
C G
antikodon
tRNA
asp
U
Met
Ala Trp
Ser
Val
U A
C A U G G A C
C U G
Asp
6
U A
C A U G G A C
C U G
Met
Val
Asp
Ala
Trp
Ser
další
peptidová
vazba
6 3
G A C C U G
A U G A C C
Met
Val
Asp
Ala
Trp
Ser Thr
6 3
3 stádia translace

Genová exprese u eukaryot