Embed Size (px)
Citation preview

Universidad Rafael Landívar Facultad de Ingeniería Ingeniería Química Industrial Laboratorio de Bioquímica Lic. María Lucía Escobar
Metabolismo de los lípidos Parcial # 3
Zucely Castillo 1022507 Andrea Rímola
Jessica Villagrán 1008508 Julio Zabaleta 1171909
Guatemala, julio 5 de 2012

Metabolismo de los lípidos
Los lípidos son un conjunto de moléculas orgánicas, compuestas principalmente por carbono e hidrógeno y en menor medida oxígeno. Tienen como característica principal el ser hidrófobas y solubles en disolventes orgánicos. Principales Funciones:
• Reserva energética (ej. Trigliceridos) • Estructural (ej. Fosfolípidos) • Reguladora (ej. Hormonas esteroides)
Cuando las reservas energéticas son elevadas, los triacilgliceroles se almacenan mediante un proceso denominado lipogénesis. Cuando las reservas energéticas son bajas, los triacilgliceroles se degradan mediante un proceso denominado lipólisis para formar ácidos grasos y glicerol. Ácidos grasos y Triacilgliceroles
• Los ácidos grasos son fuente de energía. • Las moléculas de triacilgliceroles se digieren dentro del intestino delgado.
Una de las formas como se sintetiza los triacilgliceroles:
Transporte a través membrana plasmática
De células de pared intestinal

Reacciones de los triacilgliceroles:
Los quilomicrones se secretan a la linfa (líquido hístico derivado de la
sangre) pasan de ésta a la sangre en el ducto torácico. Los triacilgliceroles de los quilomicrones circulantes se retiran de la sangre
por células de los tejidos adiposo y muscular (son los principales depósitos de almacenamiento de lípidos del organismo)
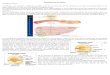
Digestión y absorción de las grasas en el intestino delgado:

• La lipasa de lipoproteína que se sintetiza en la musculatura cardiaca, glándulas mamarias lactantes y tejido adiposo, se transfiere a la superficie del endotelio de los capilares donde convierte los triacilgliceroles de los quilomicrones en ácidos grasos y glicerol.
• Los ácidos grasos son captados por las células y posteriormente se degradan para generar energía, el glicerol viaja en la sangre hasta el hígado donde la enzima cinasa de glicerol lo convierte en glicerol-3-fostato.
• Los remanentes de quilomicrones se retiran de la sangre a través de endocitosis mediada por receptores.
Triacilgliceroles:
• Se sintetizan en el hígado y se almacenan en el tejido adiposo. • Para sintetizarlo se requiere:
– Glicerol-3-fosfato – 2 acil-CoA
La reacción entre lo anterior forma ácido fosfatídico, el cual posteriormente se convierte en diacilglicerol y en una tercera reacción de acilación se forma los triacilgliceroles. Degradación de los ácidos grasos:
• Los ácidos grasos se degradan por la separación secuencial de fragmentos de dos carbonos. Se da la ruptura del enlace entre los átomos de carbono α y β.
• El proceso anterior recibe el nombre de β-oxidación. • En este proceso se forma acetil-CoA. • Se produce dentro de la mitocondrias cuando las cadenas son medianas, si
son muy largas primero ocurre en los peroxisomas para acortarla y luego en la mitocondrias.
Oxidación total del ácido graso:
• La oxidación aerobia de un ácido graso genera un gran número de moléculas de ATP.
• En las plantas la β-oxidación tiene lugar predominantemente en los peroxisomas.
Biosíntesis de los ácidos grasos:
• La síntesis de los ácidos grasos tiene lugar dentro del citoplasma de la mayoría de las células animales.
• El principal lugar donde se llevan a cabo estos procesos es el hígado.

• Los ácidos grasos se sintetizan cuando la alimentación tiene poca grasas o muchos carbohidratos o proteínas.
• Los ácidos grasos se sintetizan a partir de la glucosa. • La glucosa se convierte en piruvato en el citoplasma, éste entra al
mitocondrias y se convierte en acetil-CoA y éste se condensa con el oxalacetato para formar citrato.
• Cuando la concentración de citrato es alta, éste pasa al citoplasma donde se fragmenta para formar acetil-CoA y oxalacetato.
• A partir del acetil-CoA se sintetiza el ácido graso.
La síntesis de ácidos grasos se da en el citoplasma de las células animales; pero el hígado es el principal lugar de este proceso; y se sintetizan cuando la alimentación es deficiente de grasas y abundante en carbohidratos o proteínas. La mayoría de los ácidos grasos se sintetizan a partir de la glucosa de los alimentos. La biosíntesis de los ácidos grasos es semejante a su beta-oxidación, pero se diferencia por los siguientes factores:
Localización: La síntesis de los ácidos grasos sucede de forma predominante en el citoplasma, mientras que la beta-oxidación ocurre en las mitocondrias.
Enzimas: Las enzimas que catalizan la síntesis de los ácidos grasos tienen una estructura distinta a las de la beta-oxidación; y forman un complejo multienzimático que se denomina sintasa de ácidos grasos.
Enlace tioéster: Los intermediarios de la síntesis de ácidos grasos están ligados mediante un enlace tioéster a la proteína transportadora del acilo (ACP), la cual es es un componenete de la sintasa de ácidos grasos; mientras que en la beta oxidación se une a la CoASH.
Transportadores electrónicos: Al contrario de la beta-oxidación, la cual produce NADH y FADH2, la síntesis de ácidos grasos consume el NADPH.
Mecanismo Los sustratos para la síntesis de ácidos grasos son la acetil-ACP, que se forma a partir de la acetil-CoA y ACP por acción de una transacetilisa) y la ACP, que se genera a partir de la acetil-CoA, dióxido de carbono y ACP por efecto de ACC y maloniltransacetilasa. Durante cada ciclo de elongación, la molécula de ácido graso naciente se alarga dos carbonos cuando una malonil-ACP se condensa con la cadena acil-ACP creciente en una reacción catalizada por sintetasa de veto-cetoacetil-ACP. A esto sigue una reducción del nuevo grupo carbonilo por una reductasa para formar un alcohol y la eliminación de agua para formar un doble enlace carbono-carbono. Cada ciclo de elogación termina con la reducción del doble enlace carbono-carbono. El proceso concluye hasta la formación de palmiato.

La elogación y la desaturación de los ácidos grasos que se sintetizan en el citoplasma o de los obtenidos en la alimentación se realizan en primera instancia mediante enzimas del ER. La elogación y desaturación de los ácidos grasos son especialmente importantes en la regulación de la fluidez de la membrana y de la síntesis de los precursores de diversos derivados de los ácidos grasos como los eicosanoides. La elogación de los ácidos grasos en el ER, que utiliza unidades de dos carbonos proporcionadas por la malonil-CoA, es un ciclo de reacciones de condensación, reducción, deshidratación semejante al que se observa en la síntesis citoplasmática de los ácidos grasos. Al contrario que en el proceso citoplasmático, los intermediarios del proceso de elogación en el ER son ésteres de CoA. Estas reacciones pueden alargar tanto ácidos grasos saturados e insaturados.

REGULACIÓN DEL METABOLISMO DE LOS ÁCIDOS GRASOS Se emplean mecanismos regulatorios a largo y corto plazo. En la regulación a corto plazo (medida en minutos) las actividades de las moléculas existentes de enzimas reguladoras clave son modificadas por reguladores alostéricos; mediante la modificación covalente a través de hormonas. Cuando los niveles de energía son altos, la beta-oxidación se reprime; igualmente en el hígado cuando la relación insulina/glucagón es alta, la concentración de malonil-CoA es alta y se reprime la beta-oxidación. Los cambios en la regulación a largo plazo del metabolismo de los ácidos grasos, que ocurren en respuesta a las fluctuaciones en la disponibilidad de nutrimentos y en la demanda de energía, son efectuados por modificaciones en la expresión génica. Dos clases de factores de transcripción son componentes notables de un proceso regulatorio intrincado: las SREBP y PPAR. Cada tipo de factor de transcripción cuando se activa, se una a un elemento regulatorio cercano a los genes diana, un proceso que desencadena la unión de moléculas coactivadoras, y posteriormente la transcripción. METABOLISMO DE LOS LÍPIDOS DE MEMBRANA La bicapa lipídica de las membranas celulares está formada en su mayoría por fosfolípido y esfingolípidos. Metabolismo de los Fosfolípidos La mayoría de reacciones de biosíntesis de lípidos se encuentran en el retículo endoplásmatico liso. Cada enzima es una proteína de la membrana que tiene el sitio activo dirigido al citoplasma, por esto la biosíntesis de fosfolípidos se produce en la interfaz de la membrana del retículo endoplásmatico y el citoplasma. La composición de ácidos grasos en los fosfolípidos cambia un poco tras la síntesis. Se cree que este proceso permite a las células ajustar la fluidez de sus membranas. La síntesis de fosfatidiletanolamina comienza cuando la etanolamina entra en la célula y se fosforila de inmediato. Luego reacciona con trifosfato de citidina (CTD) para formar el intermediario activado CDP-etanolamina. Los derivados de CDP tienen una función importante en la transferencia de grupos de cabezas polares en la síntesis de fosfolípidos. La CDP-etanolamina se convierte en fosfatildiletanolamina al reaccionar con diacilglicerol. Esta reacción es catalizada por una enzima del retículo endoplásmatico. La biosíntesis de fosfatidilcolina es similar a la fosfatildiletanolamina, la coolina) necesaria para esta síntesis) se obtiene de la alimentación. La fosfatidilcolina también es sintetizada en el hígado fosfatidiletanolamina, esta se metila tres veces por medio de una enzima lo que da como producto fosfatidilcolina. El donador de metilos es s-adenosilmetionina.

La velocidad a la que se degradan todas las moléculas de una estructura y se sustituyen por moléculas recién sintetizadas, en los fosfolípidos es rápida.

Metabolismo de los Esfingolípidos La síntesis de esfingosina comienza con la condesacion de palmitoil-CoA con la serina para formar 3-cetoesfinganina. A continuacion el NADPH reduce la 3-cetoesfinganina para formar esfinganina, esta se convierte luego en ceramida en dos pasos con la acetil-CoA y FADH2. La esfingomielina se forma cuando la ceramida reacciona con la fosfatidilcolina. Cuando esta reacciona con UDP-glucosa, se produce glucosil ceramida. El galactocerebrosido se sintetiza cuando la ceramida reacciona con UDP-galactosa. Los sulfatidos se sintetizan cuando los galactocerobrosidos reaccionan con 3’-fosfatodenosina-5’-fosfosulfato (PAPS). Los esfingolipidos se degradan dentro de los lisosomas. Cuando las enzimas que se requieren para degradar estas moléculas no se producen o son defectuosas, se produce la esfingolipidosis.

METABOLISMO DE ISOPRENOIDES
Los isoprenoides se encuentran en los eucariotas. Las rutas de biosíntesis de los
isoprenoides producen una enorme variedad de productos en diferentes tipos
celulares y en diferentes especies. A pesar de su diversidad, el comienzo de la
biosíntesis de los isoprenoides parece ser idéntico en la mayoría de las especies

investigadas (p. ej., levaduras, mamíferos y vegetales). (HMG-CoA = p-hidroxi-p-
metilgluta¡ jI-CoA.).
METABOLISMO DEL COLESTEROL
El colesterol forma parte de las membranas plasmáticas de las células eucariotas,
siendo un componente esencial para su estabilidad estructural y funcional. Se
encuentra presente en los tejidos y en el plasma
La mayor parte del colesterol se encuentra en forma esterificada ( ester de
colesterilo). Es transportado en el torrente sanguíneo (plasma) por medio de las
lipoproteínas del plasma. La proporción más elevada de colesterol se encuentra
en las LDL
Funciones:
a. Componente estructural esencial de membranas
b. Componente de la capa externa de las lipoproteínas plasmáticas

c. Precursor de esteroides cumplen importantes funciones fisiológicas, tales
como ácidos biliares, hormonas esteroideas, vitamina D, etc.
(corticoesteroides,hormonas sexuales)
d. Precursor de ácidos biliares( transporte inverso del colesterol)
e. Precursor de vitamina D
f. Constituyente principal de cálculos biliares
g. Procesos patológicos; factor en la génesis de ateroesclerosis

SÍNTESIS DEL COLESTEROL
El colesterol se sintetiza en el hígado. La síntesis de colesterol se divide en tres
fases:
1. Fase I: Formación de HMG- CoA (β-hidroxi-β-metilglutari-CoA) a partir de
acetil-CoA.
2. Fase II: Conversión de HMG- CoA en escualeno
3. Fase III: Conversión de escualeno en colesterol
FASE 1: Formación de HMG- CoA (β-hidroxi-β-metilglutari-CoA) a partir de
acetil-CoA.
La primera fase es un proceso citoplásmico. El cual inicia con la condensación de
dos moléculas de acetil- CoA para formar β- cetobutiril CoA también conocida
como acetoacetil CoA, esta reacción se encuentra catalizada por la tiolasa.
Seguidamente la β- cetobutiril CoA se condensa con otra molécula de acetil- CoA
formando la β-hidroxi- β- cetobutiril CoA (HMG-CoA) y catalizada por β-hidroxi- β-
cetobutiril CoA sintasa.
FASE II: Conversión de HMG- CoA en escualeno
La segunda fase comienza con la reducción de la HMG-CoA formada en la fase I
para formar mevalonato, el NADPH cumple su función como agente reductor.
A partir de MEVALONATO se obtienen los ISOPRENOS ACTIVADOS, estos
pasos implican gasto de ATP. Los ISOPRENOS se obtienen por reacciones
CABEZA-COLA. Los Isoprenos activados reaccionan entre sí para formar
ISOPRENOIDES de:
10 C: GERANIL-PP
15 C: FARNESIL-PP

Dos moléculas de FARNESIL-PP se condensan para formar una de 30 C:
ESCUALENO que se asimila a la estructura abierta del COLESTEROL.

FASE III: Conversión de escualeno en colesterol
En la tercera fase empieza con la unión del escualeno a una proteína
transportadora citoplásmica específica llamada proteína transportadora de
esteroles. La conversión del escualeno en lanosterol tiene lugar con el
intermediario unido a esta proteína. Las actividades enzimáticas que se requieren
para la formación del epóxido dependiente de oxígeno (escualeno
monooxigenasa) y la posterior ciclación (2,3-oxidoescualeno lanosterol ciclasa)
que dan lugar a la síntesis de lanosterol se encuentran en los microsomas. La
escualeno monooxigenasa requiere para su actividad NADPH y FAD. Tras su
síntesis, el lanosterol se une a una segunda proteína transportadora, a la que
permanece unido durante las reacciones restantes. Todas las actividades
enzimáticas que catalizan las 20 reacciones restantes necesarias para convertir el
lanosterol en colesterol están embebidas en las membranas microsómicas. En un
conjunto de transformaciones que utilizan el NADPH y el oxígeno, el lanosterol se
convierte en 7-deshidrocolesterol. Este producto posteriomente se reduce por el
NADPH para formar colesterol

REGULACIÓN DEL COLESTEROL
La regulación de la biosíntesis del colesterol se ejerce fundamentalmente en el
hígado. El aumento de colesterol en la diete produce una inhibición de la síntesis
hepática, que se lleva a cabo en la primera etapa de formación de mevolanato, por
inhibición de la HMG-CoA reductasa, que actúa como enzima limitante.
Por otra parte, la síntesis de colesterol también se ve inhibida como consecuencia
del efecto que ejercen sus niveles intracelulares sobre la síntesis y expresión del
receptor celular LDL, que bloquea la incorporación de colesterol desde dichas
lipoproteínas plasmáticas.
DEGRADACIÓN DEL COLESTEROL
El mecanismo más importante para degradar y eliminar el colesterol es la síntesis
de ácidos biliares. Ésta, que tiene lugar en el hígado, se esquematiza en la Figura
12-28. La conversión de colesterol en 7-ct-hidrocolesterol, que cataliza la
colesterol-7-hidroxilasa (una enzima microsómica), es la reacción limitante de la
velocidad de la síntesis de ácidos biliares. En reacciones posteriores, el doble
enlace de C-S se reagrupa y reduce y se introduce otro grupo hidroxilo. Los
productos de este proceso, el ácido cólico y el ácido desoxicólico, se convierten en
sales biliares por enzimas microsómicas que catalizan reacciones de conjugación.
REFERENCIAS
Mckee (2009) Bioquímica, Las Bases Moleculares para la Vida (4ª. Ed.).
México, Mc Graw Hill