Embed Size (px)
Citation preview

Mikrobiologie II - Überblick
•Vorbemerkungen zum Zellmetabolismus•Energiestoffwechsel von Mikroorganismen
Gärungsformen, SubstratkettenphosphorylierungEnergiegewinnung durch die AtmungsketteEnergiegewinnung durch Photosynthese
•Mikroorganismen als InfektionserregerPathogenitätsfaktorenToxine

Mikrobiologie II - Überblick•Energiestoffwechsel von Mikroorganismen Gärungsformen, Substratkettenphosphorylierung
- Glycolyse (Embden-Meyerhoff-Weg)- Pentosephosphat-Weg- Entner-Douderoff-Weg- Phosphoketolase-Weg, Milchsäurebakterien- Reduktion von Pyruvat - Ethanolbildung in Hefen und Bakterien - Propionsäuregärung, Propionibakterien - Ameisensäuregärung, Enterobacteriaceae - Buttersäuregärung, Butanolgärung, Clostridien - Homoacetatfermentation in Clostridien

Mikrobiologie II - Überblick
•Energiestoffwechsel von Mikroorganismen Energiegewinnung durch die Atmungskette
- Zitronensäurezyklus (Krebs-Zyklus)- Elektronentransportkette (Atmungskette)- Anaerober Elektronentransport- Aerobe chemolithotrophe Bakterien- Unvollständige Oxidation- Toxische Wirkung des Sauerstoffs

Mikrobiologie II - Überblick
•Energiestoffwechsel von Mikroorganismen Energiegewinnung durch Photosynthese
- Struktur und Funktion photosynthetischer Reaktionszentren- Calvin-Zyklus- Purpurbakterien- Grüne Bakterien- Cyanobakterien- Halobakterien (Archaeen), Bakterienrhodopsin

Mikrobiologie II - Überblick
•Mikroorganismen als Infektionserreger Pathogenitätsmechanismen: Anheften, Invadieren Bakterielle Toxine
- Diphtherietoxin- Choleratoxin- Pertussis-Toxin- Clostridien-Neurotoxine

Grundprinzipien der Biochemie
Um die Vorgänge des Zellmetabolismus verstehen zukönnen, sind Grundkenntnisse der Thermodynamik(in welche Richtung läuft eine Reaktion, Einfluß vonEnergie und Entropie) und Kinetik (Reaktions-geschwindigkeit) unbedingt erforderlich.

“Energieinhalt” (Freie Standard-Enthalpie) wichtigerEnergiemetaboliten*

“Energieinhalt” (Freie Standard-Enthalpie) wichtigerEnergiemetaboliten
Die Werte geben die Enthalpie des jeweiligen „Gruppen-übertragungspotentials“ an, also die Enthalpie, die in derBindung zum (bei ATP äußersten) Phosphat steckt, alsokeinesfalls eine Art „Gesamtenergie“ des Moleküls (Was wäredas überhaupt? Die Energie, die bei Verbrennung frei würde?Die Energie, die bei Kernreaktionen frei würde?)
In der Tabelle auf der vorigen Seite geht es jeweils um diehydrolytische Abspaltung eines Phosphatrestes (also + 1 H2O,1 H3PO4 geht ab).

Gibb´sche Freie Enthalpie
Zur Erinnerung (Physikalische Chemie, Biochemie)
ΔG = ΔH - TΔS(Freie Enthalpie = Enthalpie - absolute Temperatur * Entropie
Enthalpie H entspricht der Wärme (Wert negativ: exothermeReaktion, Wärme wird frei).Entropie ist ein Maß für die Unordnung, nimmt insgesamtimmer zu (2. Hauptsatz der Thermodynamik)

Gibb´sche Freie Enthalpie
ΔG = ΔH - TΔSEine Reaktion läuft nur ab, wenn die Freie Enthalpie abnimmt.Eine Teilreaktion mit positivem ΔG kann ablaufen, wenn esmit einer anderen Teilreaktion so gekoppelt ist, dass ΔGinsgesamt negativ ist.
Das „Δ“ bedeutet immer den Unterschied von Produkten derchemischen Reaktion zu den Edukten(Ausgangsverbindungen).

Gibb´sche Freie EnthalpieΔG = ΔH - TΔS
Aus der Tabelle der Energiemetaboliten könnte man nunschließen, dass z.B. aus Kreatinphosphat (ΔG°=-43,1 kJ/mol)und ADP ATP und Kreatin entstehen können, nie aberumgekehrt (ATP>ADP ΔG°=-30,5 kJ/mol). Tatsächlich spieltaber die Konzentration der Substanzen noch eineentscheidende Rolle. Diese „°“-Zahlen sind Standardwerte, diebei gleichen Konzentrationen (je 1 molar) gelten. Im Muskelliegt viel ATP und wenig ADP vor, dadurch kommt imGleichgewichtszustand („steady state“) auch Kreatinphosphatin nutzbaren (Energiespeicher) Mengen vor.

Gibb´sche Freie EnthalpieΔG = ΔH - TΔS
Die Definition für „°“-Zahlen mit „alles ist 1 molar“ schließtauch Protonen (H+) ein. Diese Standardbedingungenentsprechen daher einer extrem konzentrierten Säure mitpH=0!Für physiologische (natürliche, zelluläre) Reaktionen nimmtman daher lieber ein °‘ (ΔG °‘) als Standardwert. Dabei istalles 1 molar, nur der pH ist neutral (pH=7, also H+-Konzentration 10-7 molar), und die Konzentration von Wasserist 55 molar (wässriges M). Unterschiede zwischen ΔG ° undΔG °‘ treten nur auf, falls in der Reaktion H+ (oder OH-) oderWasser vorkommt.

Gibb´sche Freie Enthalpie
ΔG = ΔH - TΔS
Diese Konzentrationsabhängigkeit macht das „Ziehen“ einerungünstigen Reaktion durch eine günstige erst möglich. Vom„ungünstigen“ Zwischenprodukt liegt im „steady state“immer nur eine geringe Konzentration vor (durch denschnellen Verbrauch durch die „ziehende“ Weiterreaktion).

Ermöglichen “ungünstiger”Reaktionen
Beispiel für ein energetisch ungünstiges Zwischenprodukt, dasdurch günstige Weiterreaktionen ständig abgezogen wird:
Das Gleichgewicht der Aldolasereaktion liegt weit auf Seitendes Fructose-bisphosphats (ΔG°=+22,8 kJ/mol fürAldolspaltung). In der Zelle wird das GAP aber so schnellverbraucht, dass (wegen der extrem niedrigen Konzentrationder Edukte GAP und DHAP) die Spaltung abläuft (ΔG=-5,9kJ/mol für die zellulären Konzentrationsverhältnisse).

Ermöglichen “ungünstiger”Reaktionen
Im Fall der ATP-Bildung: ADP + Pi ATP + H2O,ΔG°=+30,5 kJ/mol, kann der „Trick“ mit der niedrigenKonzentration nicht genutzt werden, da die Zelle eine rechthohe Konzentration des Energielieferanten ATP unbedingtbraucht. Ausweg ist hier die Kopplung an stark„exergonische“ (negatives ΔG) Reaktionen, z.B. dieGAPDH/PGK-Reaktion:GAP + Pi + NAD+ 1,3-Diphosphoglycerat + NADH + H+
1,3-Diphosphoglycerat +ADP 3-Phosphoglycerat + ATP
Die energieliefernde Reaktion ist dabei die Oxidation desGAP: GAP + NAD+ 3-Phosphoglycerat + NADH + H+

In diesem Fall darf kein Enzym die ReaktionADP + Pi ATP + H2O katalysieren, da sonst das aufwendigproduzierte ATP wieder hydrolysieren würde!
Ermöglichen “ungünstiger”Reaktionen

Gleichgewichtsreaktionen
Überhaupt ist es wichtig, sich klar zu machen, dass prinzipiellalle Reaktionen auf einen Gleichgewichtszustand zulaufen,d.h., es sind „am Ende“, im Reaktionsgleichgewicht, auchnoch Ausgangsverbindungen vorhanden. Ganz in eineRichtung verschoben kann das aber dadurch werden, das einProdukt entfernt wird (Weiterreaktion, aber auch, wenn einGas entsteht und entweicht). Dann läuft die Reaktion (imVersuch, dies Produkt nachzuliefern) immer weiter, bis zumVerschwinden der Edukte.

Gleichgewichtsreaktionen
Die Mengenverhältnisse von Edukten und Produkten imGleichgewicht werden durch das Massenwirkungsgesetzbeschrieben:
Für a A + b B x X + y Y
gilt im Reaktionsgleichgewicht:
Kc = [X]x[Y]y/ [A]a[B]b

Gleichgewichtsreaktionen
Konkret für die Kreatinphosphatbildung:
ATP + Kreatin ADP + Kreatinphosphat
Kc = [ADP][Kreatinphosphat]/[ATP][Kreatin]
(die eckigen Klammern bedeuten die Konzentration derjeweiligen Substanz, die Kleinbuchstaben a, b, x, y die Anzahlder Moleküle in der Reaktion, im Beispiel je 1. Dastiefgestellte c zeigt, das die Gleichgewichtskonstante mitKonzentrationen berechnet wurde (es geht auch mitPartialdrucken, das ist dann Kp)).

Gleichgewichtsreaktionen
Zwischen der Gibbschen freien Enthalpie und derGleichgewichtskonstante besteht folgende Beziehung: ΔG° = -RT lnKDabei ist T die absolute Temperatur (0°C = 273,15 K) undR die allgemeine Gaskonstante, also ein fester Wert:R = 8,314 J/(K mol)Für unser Beispiel errechnen wir aus den Enthalpien derTeilreaktionen (+43,1 kJ/mol und -30,5 kJ/mol) dieReaktionsenthalpie mit +12,6 kJ/mol

Gleichgewichtsreaktionen
Zwischen der Gibbschen freien Enthalpie und derGleichgewichtskonstante besteht folgende Beziehung: 12,6 kJ/mol = -0,008314 kJ/(K mol) 310 K* lnKc*bei 37°C
lnKc = -4,889Kc = e-4,889 = 0,00753 (Eulersche Zahl e ca. 2,71)
-------------Kleiner Trick mit natürlichen Logarithmen:ln x = 2,303 log xAlso ist unser Kc (-4,889/2,303) = 10-2,123

Gleichgewichtsreaktionen
Kc = 0,00753Also ist das Produkt der Ausgangskonzentrationen 133 malgrößer als das der Produkte.Klingt schlecht für die Kreatinphosphatbildung. Wenn aber imgut versorgten Muskel das ATP/ADP-Verhältnis 50:1 ist, kannimmerhin (133/50 = 2,66, so wäre das Kreatin/Kreatin-Phosphat-Verhältnis) gut ein Viertel des Kreatins inKreatinphosphat umgewandelt werden. Im Extremfall sollsogar ein ATP/ADP-Verhältnis bis 500 erreicht werdenkönnen, wodurch der Großteil des Kreatins in Kreatinphosphatumgewandelt werden könnte.

GleichgewichtsreaktionenMit steigenden ΔG-Werten verschiebt sich das Gleichgewichtaber bald extrem zu einer Seite. (Die Tabelle geht von 300 K,also Raumtemperatur aus):

Gleichgewichtsreaktionen
Das schnelle Ausrechnen von Kc aus ΔG° geht mit
1,493ΔG°= Kc für 300 Kund 1,474ΔG°= Kc für 310 K
Wenn der Taschenrechner keine beliebige Basis zumExponenten erlaubt, hilft die Webseite:http://rechneronline.de/logarithmus/

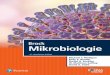
Kinetik
Auch wenn eine Reaktion „thermodynamisch begünstigt“ist, also ein stark negatives ΔG hat, heißt das nicht, das sieauch stattfindet. Zwischen Edukt und Produkt liegen meistenergetisch sehr ungünstige Übergangszustände, die einen„Energieberg“ darstellen, der die Reaktion verhindert.
Kinetik

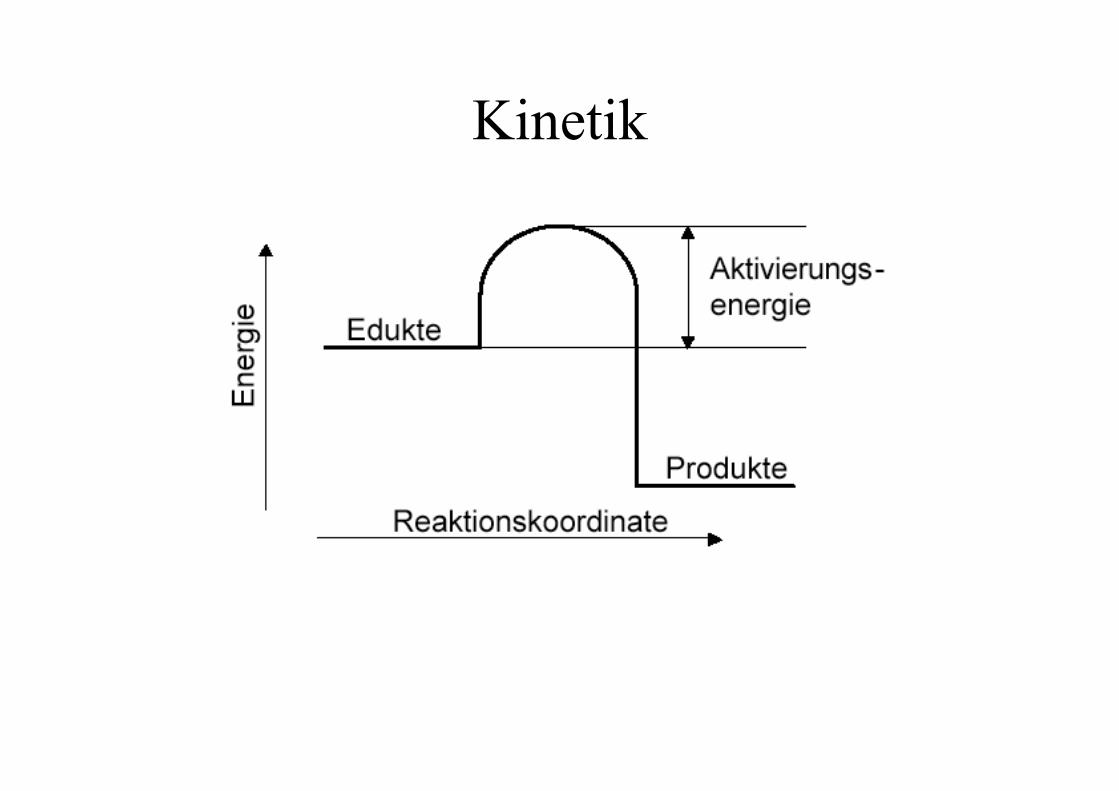
Kinetik - Katalyse
Katalysatoren sind Stoffe, die diesen Energieberg vermindernbzw. Umwege mit weniger ungünstigen Übergangszuständenermöglichen und dadurch chemische Reaktionen enormbeschleunigen oder überhaupt erst möglich machen.
Kinetik - Katalyse

Kinetik - Enzyme
Die effektivsten und in der lebenden Zelle weitaus wichtigstenKatalysatoren sind die Enzyme. Sie biegen die Edukte undbringen sie in Umgebungen (Ladungen, pH, Wasser-abschirmung...), die sie in optimale Reaktionsfähigkeitversetzen.

Enzymkatalysierte Reaktionen
Zwei Punkte sollte man sich dabei immer im Hinterkopfhalten:
1. Zwischen den vielen Komponenten der Zelle ist theoretischeine Unzahl chemischer Reaktionen möglich. Die Enzymelegen fest, welche davon wirklich geschehen. Das müssenkeineswegs die energetisch günstigsten sein.Unterschiedliche Organismen setzen dieselben Ausgangsstoffeganz verschieden um, dazu haben sie verschiedene Sets vonEnzymen.

Enzymkatalysierte Reaktionen
2. Enzyme können nur Reaktionen beschleunigen, aber siekönnen keine Reaktionsgleichgewicht verschieben. Ihr Effektist nur der, dass das Gleichgewicht schneller erreicht werdenkann.Beschleunigt werden immer die Hin- und die Rückreaktion imgleichen Maße!Durch die schon genannte Kopplung mit anderen Reaktionenermöglichen Enzyme aber scheinbar (!) unmögliche,energetisch ungünstige Umsetzungen.

Enzymkatalysierte ReaktionenEnzyme, Proteine mit katalytischen Eigenschaften, sind dieMaschinen der Zelle. Sie haben Fähigkeiten, von denen derChemiker nur träumen kann: Sie können chemischeReaktionen im „wässrigen Medium“ und bei mäßigenTemperaturen (meist 20°-37°C) und neutralem pHdurchführen, die im Labor oft nur bei großer Hitze, inkonzentrierten Säuren oder in organischen Lösungsmittelnablaufen. Außerdem sind Enzymreaktionen hochspezifisch,anders als beim Chemiker entstehen praktisch keine„Nebenprodukte“ (Abfall), die „Ausbeute“ der Reaktion ist100% (davon können Chemiker nur träumen). Außerdem sindEnzyme „stereoselektiv“.

Enzymkatalysierte Reaktionen
Durch die Zusammensetzung des aktiven Zentrums und dieAbschirmung von der Umgebung verlaufen einige Reaktionenim aktiven Zentrum eines Enzyms „in wasserfreiemorganischen Lösungsmittel“ oder bei extremen pH-Werten,während die Umgebung wässrig und pH-neutral ist.

Enzyme der GlycolyseHexokinase oder PhosphotransferasePhosphohexose-IsomerasePhosphofruktokinaseFruktosebisphosphat-Aldolase (Aldolase)TriosephosphatisomeraseGlycerinaldehydphosphat-DehydrogenasePhosphoglyceratkinasePhosphoglyceratmutaseEnolasePyruvatkinase
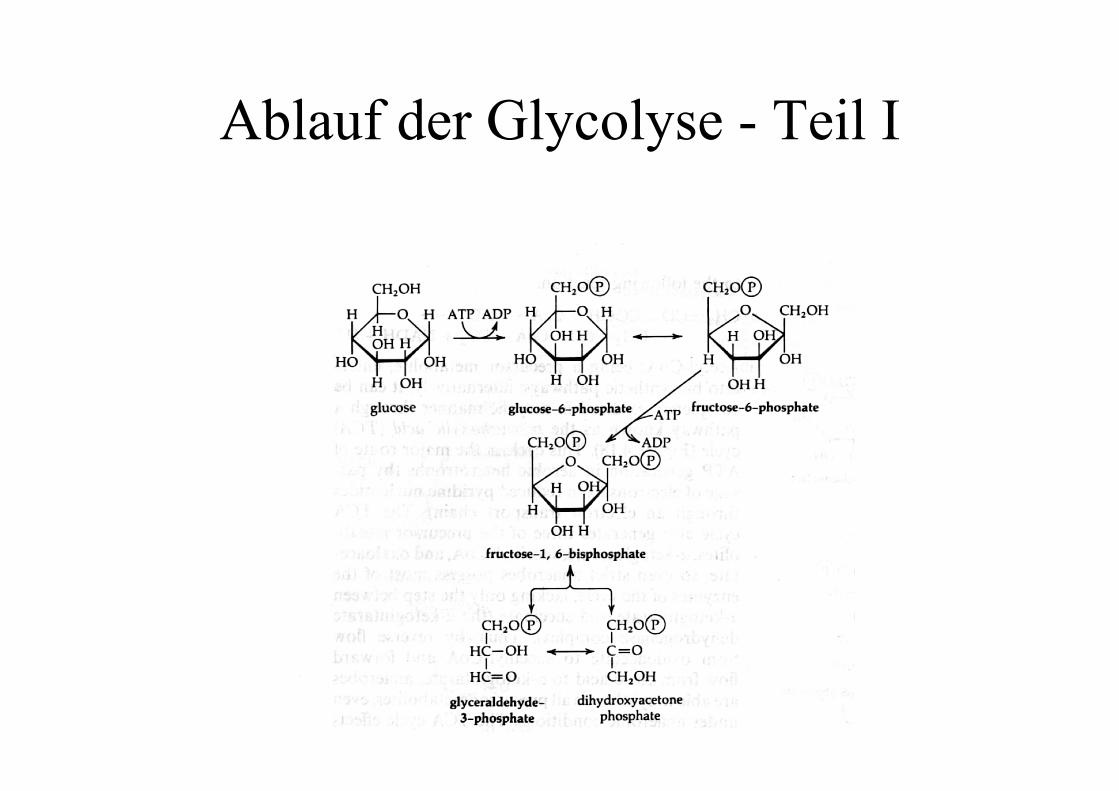
Ablauf der Glycolyse - Teil I

Der Einstieg in die Glycolyse
Bei Eukaryonten wird die Glucose durch Hexokinasephosphoryliert, bei vielen Bakterien durch dasPhosphotransferase-System während der Aufnahme in dieZelle.Durch das Anhängen des Phosphats wird der Zucker für dieZelle viel besser handhabbar: er kann nicht mehr entkommen(Phosphotransferasesystem), und kann durch die Ladung inaktiven Zentren schnell und eindeutig orientiert werden.
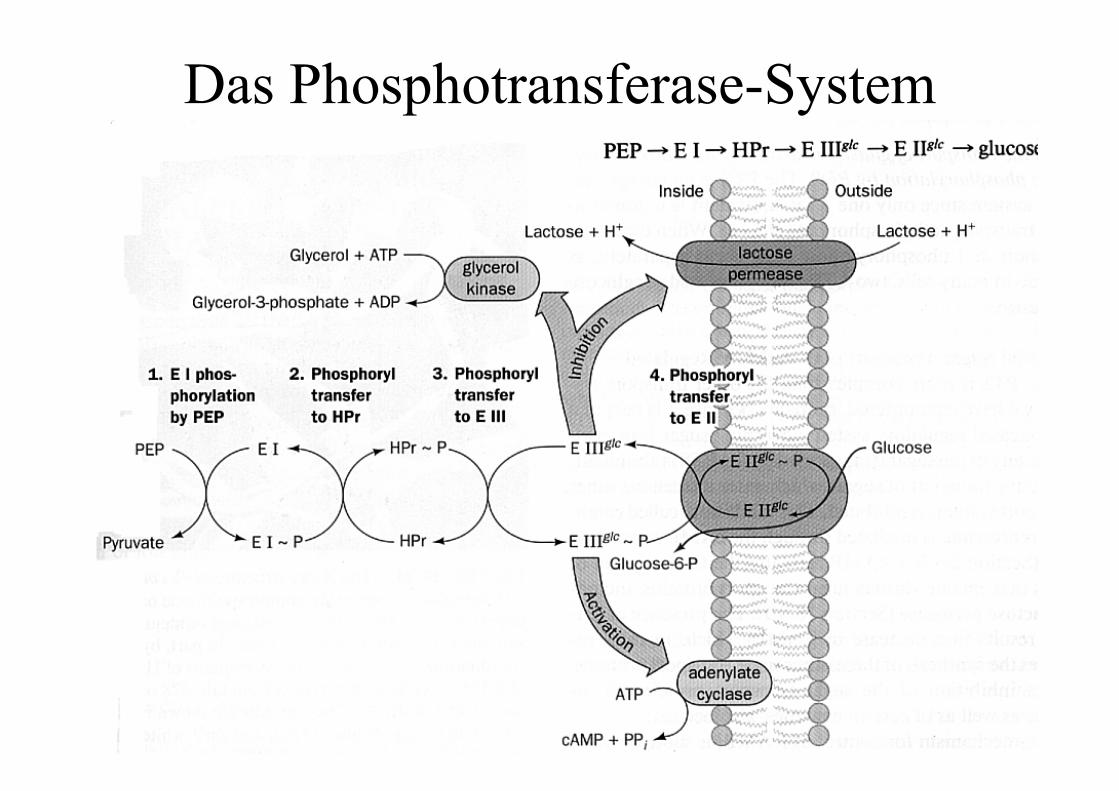
Das Phosphotransferase-System

GlucosephosphatisomeraseTriosephosphatisomerase
Bei beiden Enzymen liegt das Gleichgewicht der Isomerebeim „falschen“: GPI 70% Glucose-6-P, TPI sogar 95%DHAP. Die schnellen Reaktionen liefern das benötigteIsomere aber ausreichend nach, so dass beide Schritte nichtbremsend („geschwindigkeitsbestimmend“) wirken.
Die TPI gilt als „ideales Enzym“. Ihre Geschwindigkeit istallein diffusionskontrolliert (das Substrat muß ja mit demEnzym in Kontakt kommen), d.h., die enzymatische Reaktionselbst geht fast unmeßbar schnell.
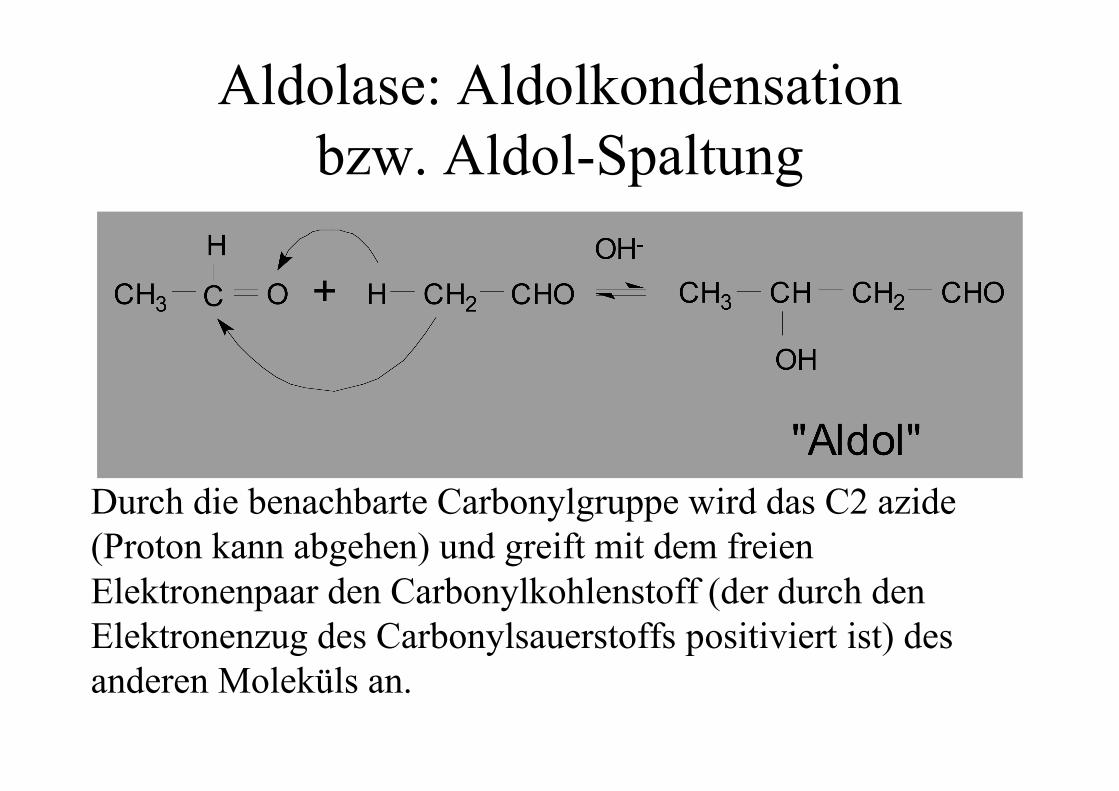
Aldolase: Aldolkondensationbzw. Aldol-Spaltung
Durch die benachbarte Carbonylgruppe wird das C2 azide(Proton kann abgehen) und greift mit dem freienElektronenpaar den Carbonylkohlenstoff (der durch denElektronenzug des Carbonylsauerstoffs positiviert ist) desanderen Moleküls an.

Ablauf der Glycolyse - Teil II

Reaktion der GAPDH
Die Glycerinaldehydphosphat-Dehydrogenase sorgt für dasEntstehen einer energiereichen Phosphatbindung, die imfolgenden Schritt zur ATP-Bildung genutzt wird.Dabei bildet das GAP mit einer Cys-SH-Gruppe des Enzymszunächst ein energiearmes Thiohalbacetal. Durch Oxidation(mit NAD+) wird daraus ein energiereicher Thioester, derdurch Phosphorolyse zu einem Säureanhydrid gespalten wird.
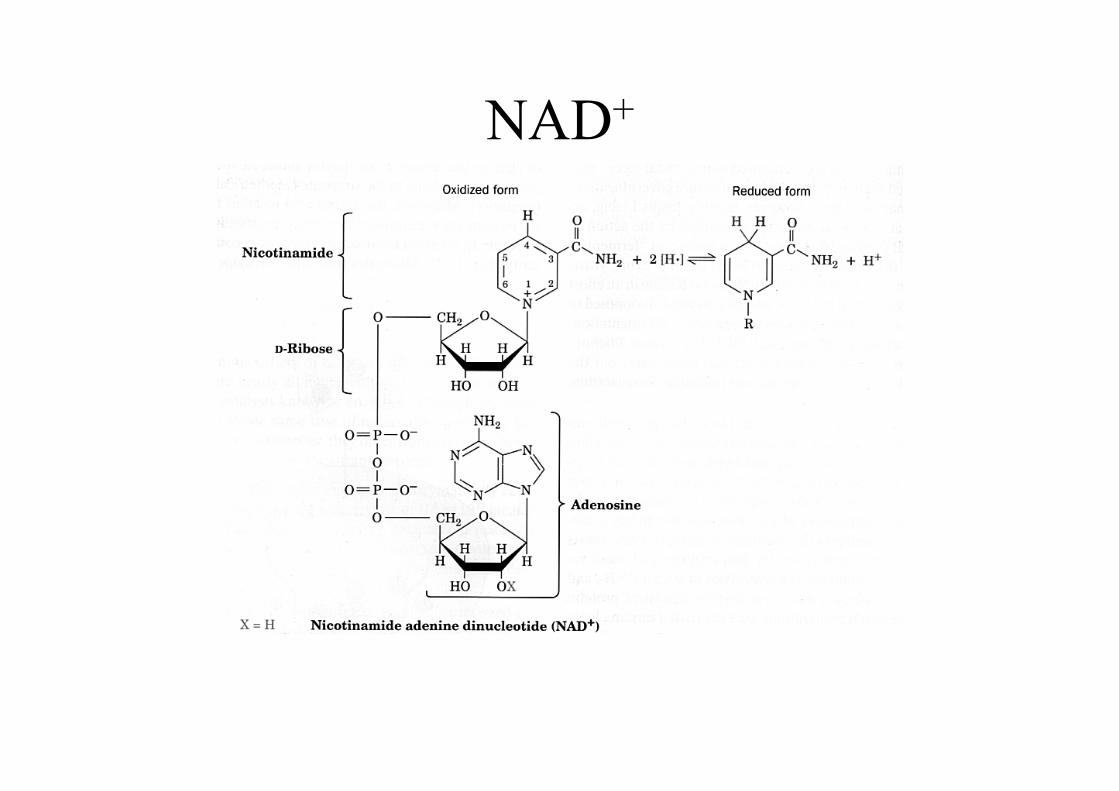
NAD+

GAPDH
Statt Phosphat kann die GAPDH auch „versehentlich“ Arsenateinbauen. Das Produkt, 1-Arseno-3-Phosphoglycerat, zerfälltin Wasser zu 3-PG, es wird kein ATP gebildet, damit fällt derEnergiegewinn durch die Glycolyse komplett aus. Arsenat istalso ein Energiegift.

PhosphoglyceratmutaseBeim „Wandern“ des Phosphats von -3 nach -2 wird erst einenzymgebundenes Phosphat auf 2 übertragen (es entsteht 2,3-Bisphosphoglycerat), dann das 3-P aufs Enzym rück-übertragen (und landet auf dem nächsten Phosphoglycerat).Zur Erstphosphorylierung des Enzyms und um es bei Verlustdes Phosphats (wenn versehentlich 2,3-BPG entkommt) zuregenerieren, werden kleine Mengen 2,3-BPG benötigt (diedie PGM mit einer BPG-Mutase-Aktivität aus 1,3-BGP bildet,dasselbe passiert in den Erythrocyten mit einer spezifischenBPG-Mutase in großem Umfang (2,3-BPG erleichtert O2-Abgabe vom Hämoglobin)).

Phosphoenolpyruvat
Bei der zweiten Energiegewinnung in der Glycolyse entstehtdie energiereiche Verbindung PEP durch Wasserabspaltungaus dem energiearmen 2-Phosphoglycerat (Enzym Enolase).Die hohe Energie (ΔG=-61,9 kJ/Mol, ATP-Bildung ΔG=-30,5kJ/Mol) steckt nur zu einem kleinen Teil (ΔG=-16 kJ/Mol) inder Phosphatbindung, sondern überwiegend in derTautomerisierung des Enols ins Keton (ΔG=-46 kJ/Mol) .

Enolase
Enolase benötigt zur Substratbindung Mg2+. Fluorid (F-) bildetmit dem Magnesium einen Komplex im aktiven Zentrum desEnzyms und blockiert es so. Darauf beruht die Giftigkeit vonFluorid (wieder ein Energiegift).
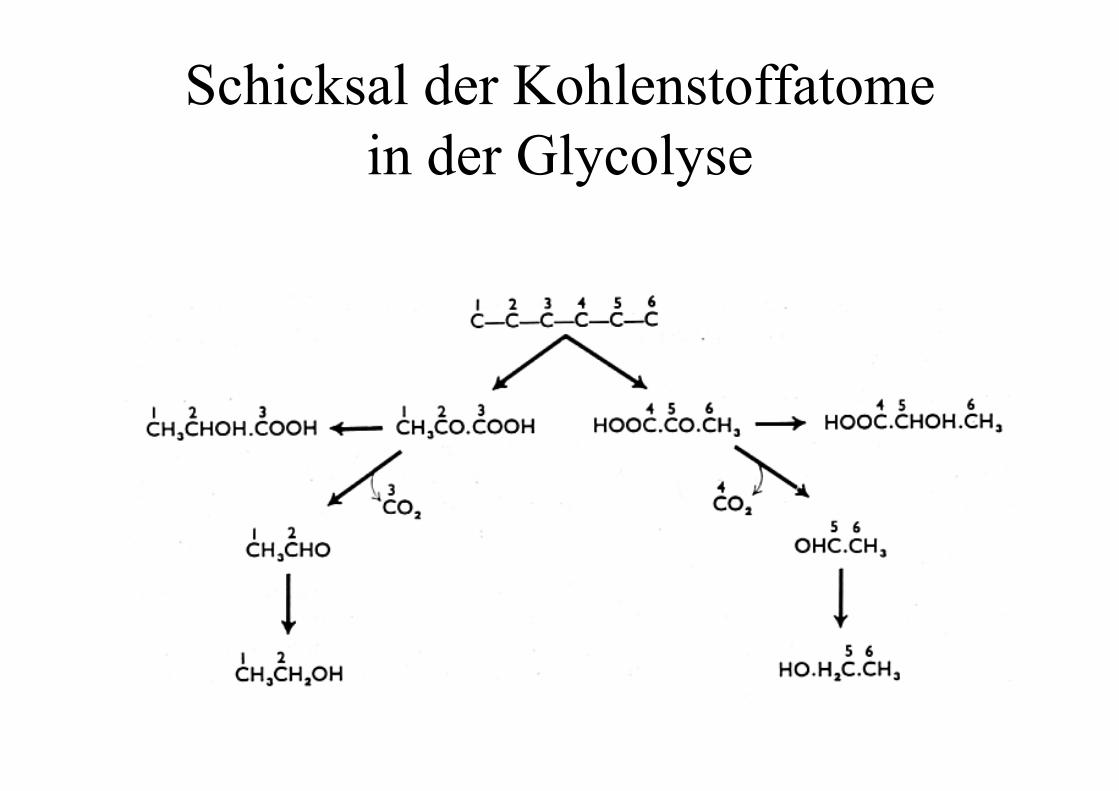
Schicksal der Kohlenstoffatomein der Glycolyse

GleichgewichtsreaktionenUm zu überprüfen, ob eine Reaktion mit den gemessenenKonzentrationen von Substraten und Produkten in diegewünschte Richtung läuft (ob diese Reaktionsrichtung einnegatives ΔG hat), modifiziert man die ΔG°-Gleichung zu:
ΔG = ΔG° RT lnQ
wobei Q das wirkliche Konzentrationsverhältnis vonProdukten/Substraten ist. Ergibt ΔG einen negativen Wert,läuft die Reaktion in die „richtige“ Richtung. Die Beziehungvon K und ΔG° gilt auch für Q und ΔG, eine Verschiebungdes Konzentrationsverhältnisses um 10x ändert die freieEnthalpie um 5,7 kJ/mol.
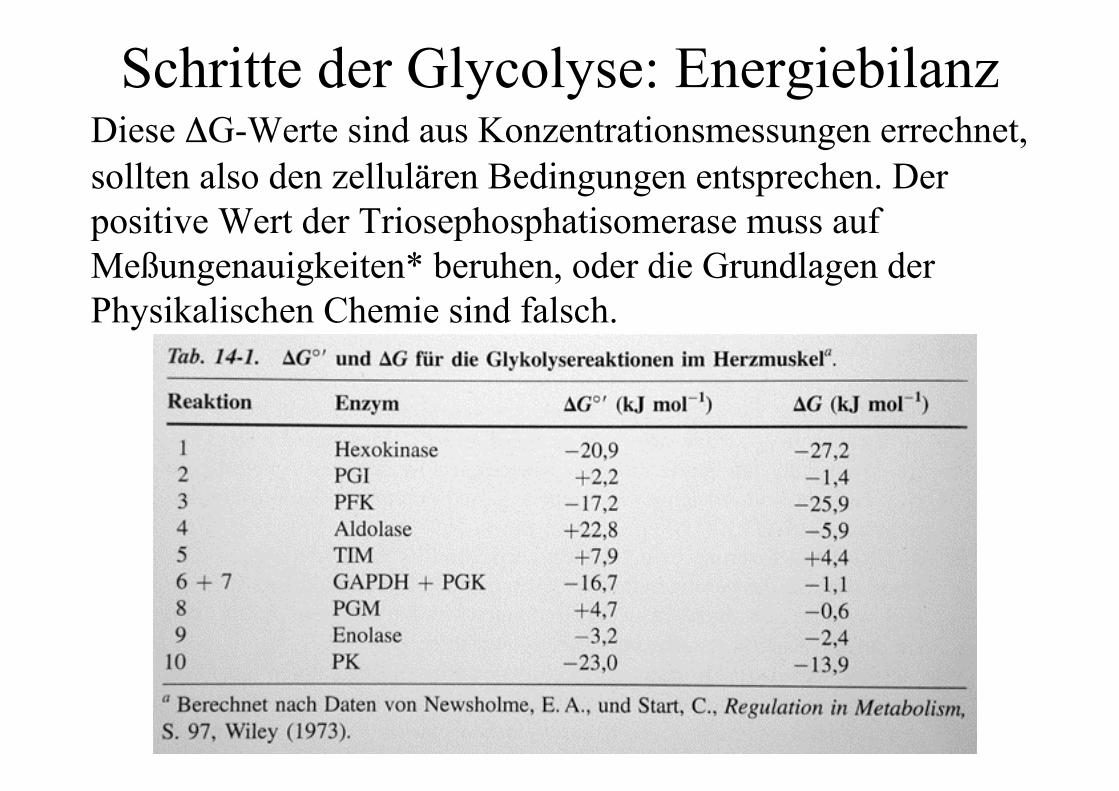
Schritte der Glycolyse: EnergiebilanzDiese ΔG-Werte sind aus Konzentrationsmessungen errechnet,sollten also den zellulären Bedingungen entsprechen. Derpositive Wert der Triosephosphatisomerase muss aufMeßungenauigkeiten* beruhen, oder die Grundlagen derPhysikalischen Chemie sind falsch.

Schritte der Glycolyse: Energiebilanz* Da hier die GAP-Menge ja extrem niedrig ist (sowohl beiAldolase wie bei TIM liegt das Gleichgewicht ja weit auf deranderen Seite), bewirkt hier ein kleiner Bestimmungsfehler(ist hier zu viel oder zu wenig GAP bestimmt?) eine starkeVerschiebung des Enthalpiewertes.
Bei einem ATP:ADP-Verhältnis von 50:1 werden alle ATP-verbrauchenden Reaktionen um -10,08 kJ/mol (bei 310 K, -9,775 kJ/mol bei 300 K) begünstigt, und alle ATP-bildendenum den gleichen Betrag ungünstiger.Ein NADH:NAD+-Verhältnis von 1:1000 erleichtertOxidationen (GAPDH) und erschwert Reduktionen um17,8 kJ/mol (bei 310 K).
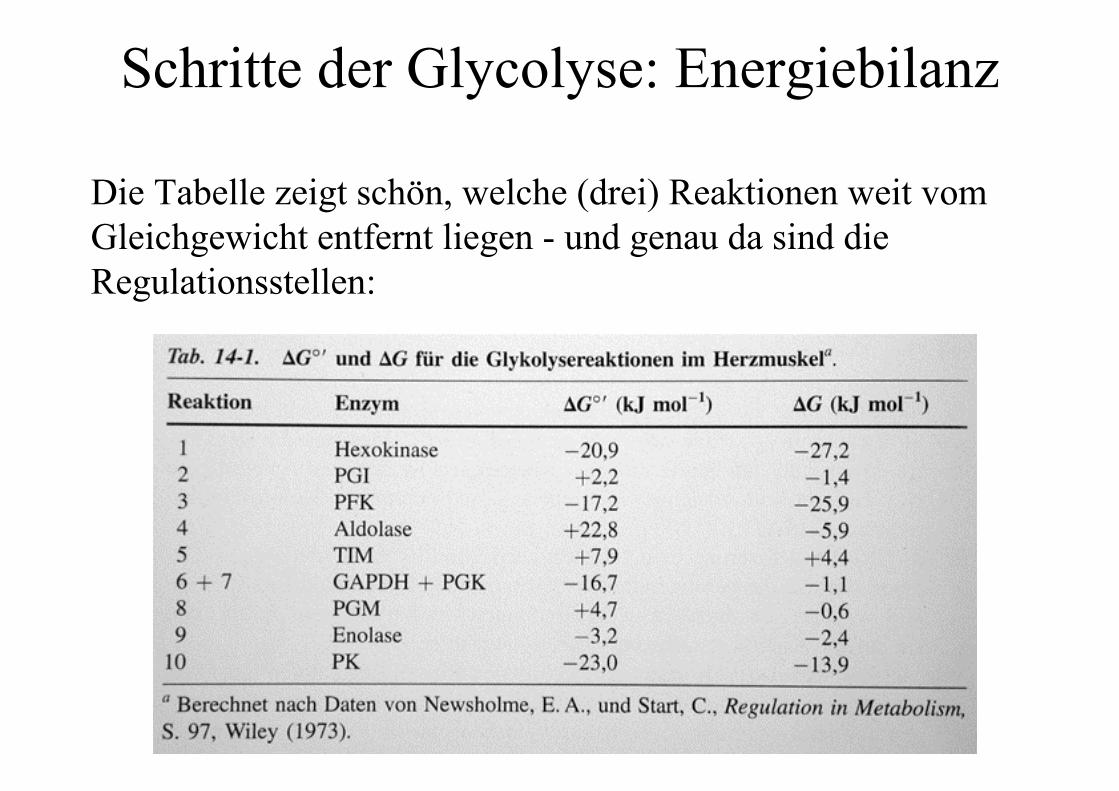
Schritte der Glycolyse: Energiebilanz
Die Tabelle zeigt schön, welche (drei) Reaktionen weit vomGleichgewicht entfernt liegen - und genau da sind dieRegulationsstellen:

Regulationsschritte in derGlycolyse
Zur Regulation eines Ablaufes bieten sich die geschwindig-keitsbestimmenden Schritte an. An zwei Stellen unterscheidensich Glycolyse und der inverse Weg, die Gluconeogenese: beider Umsetzung Fructose-6-P und Fructose-1,6-bisphosphatund den Schritten zwischen Phosphoenolpyruvat und Pyruvat.In beiden Fällen erfordert der „Rückweg“ mehr Energie alsbeim „Hinweg“ gewonnen wird. Liefen beide Prozessegleichzeitig, würde als Nettobilanz Energie verschwendet(„futile cycle“).

Regulationsschritte in derGlycolyse
Daher sind die Phosphofruktokinase und die Pyruvatkinasedie Haupt-Regulationspunkte der Glycolyse, dazu kommt derEintritt in die Glycolyse, die Phosphorylierung der Glucose(deren Rückweg durch Hydrolyse des Phosphats ebenfalls zuEnergieverlust führt).

Bilanz der GlycolyseBeim Umsatz von einer Hexose (Glucose, Fructose...) zu zweiPyruvat werden zwei ATP reingesteckt, aber vier gewonnen:Gesamtgewinn 2 ATP im Verlauf der Substratmetaboli-sierung: „Substratkettenphosphorylierung“.Außerdem sind zwei NAD+ zu NADH reduziert worden. Umdie Redox-Bilanz neutral zu halten (die NAD+ wiederzu-gewinnen), werden die Reduktionsäquivalente entweder aufdie Produkte rückübertragen (Gärung) oder in derAtmungskette auf Elektronenakzeptoren (meist O2)übertragen. Bei diesem Prozeß kommt es zu einer weiterenEnergieausnützung (ATP-Bildung).