Embed Size (px)
Citation preview

MINERALES TRAZA Y EFICIENCIA REPRODUCTIVA EN OVINOS Maximino Huerta Bravo
Posgrado en Producción Animal, Departamento de Zootecnia, Universidad Autónoma Chapingo, Km. 38.5 Carretera México-Texcoco. Chapingo, México CP 56230
Introducción Los minerales traza reciben su nombre porque las cantidades requeridas en la dieta son pequeñas y su determinación en los cuerpos vivos es difícil. Sin embargo, sus funciones son muy importantes. Por otro lado, los requerimientos de los elementos traza para las funciones reproductivas son muy pequeñas en relación a las necesidades para mantenimiento, crecimiento o producción de leche. La cantidad de premezcla de minerales traza suplementada normalmente varía de 250 a 500 g por tonelada de alimento. En general, los minerales traza son muy densos. Todo lo anterior ocasiona que estos minerales reciban poca importancia por todos los actores involucrados en los procesos de alimentación y que sean los más afectados cuando hay problemas de mezclado. Además, el coeficiente de variación de estos elementos en los alimentos es muy alta, de tal manera que su balanceo en las dietas no sea recomendable, porque la probabilidad de tener dietas deficientes es alta. Por otro lado, algunos ingredientes están muy desbalanceados en minerales o tienen sustancias antinutricionales que interfieren con los minerales traza y con el funcionamiento normal del animal como los granos de destilería, la canola, yuca, algunos forrajes como leucaena. La eficiencia reproductiva se define como el número de corderos destetados por cada 100 ovejas empadradas. Para diversas evaluaciones, incluidas las financieras es conveniente calcular los kilogramos de cordero destetado por oveja al año. Cuando conocemos la cantidad de alimento consumido por la oveja durante el año y su(s) cría(s) hasta el destete, podemos determinar la conversión alimenticia. Este valor puede variar de 16.5 a 32.8 kg de alimento por kilogramo de cordero destetado (Fletcher et al. (1985). Estos valores son demasiados elevados en comparación con el alimento requerido para engordar un cordero destetado hasta el peso de mercado (4 a 6 kg de alimento/kg de peso). Por lo anterior, el objetivo de este documento es destacar la importancia de los minerales traza en la eficiencia reproductiva. Eficiencia reproductiva Las pérdidas potenciales de corderos en la línea de tiempo de la reproducción (Kott, 2006) se pueden clasificar en:
Pérdidas embrionarias (0 a 45 días de gestación)
Pérdidas fetales (45 a 147 días de gestación)
Pérdidas de corderos predestete (parto a 60-120 días de edad)
Pérdidas posdestete
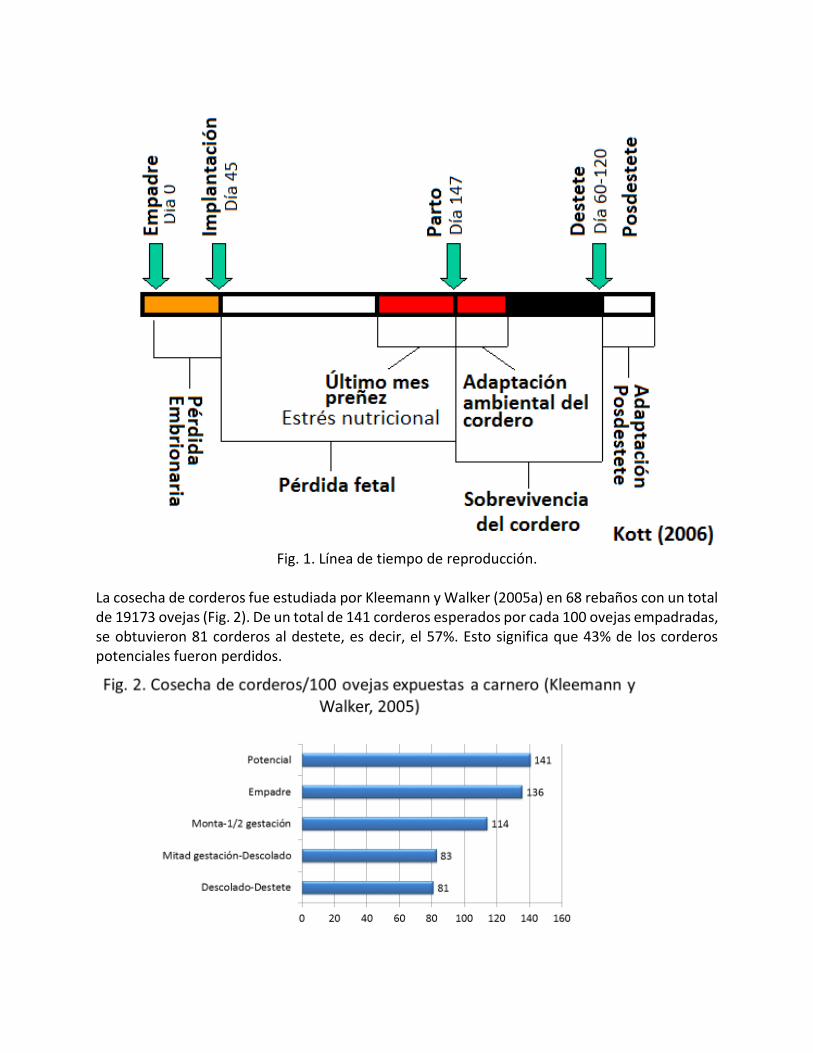
Fig. 1. Línea de tiempo de reproducción.
La cosecha de corderos fue estudiada por Kleemann y Walker (2005a) en 68 rebaños con un total de 19173 ovejas (Fig. 2). De un total de 141 corderos esperados por cada 100 ovejas empadradas, se obtuvieron 81 corderos al destete, es decir, el 57%. Esto significa que 43% de los corderos potenciales fueron perdidos.
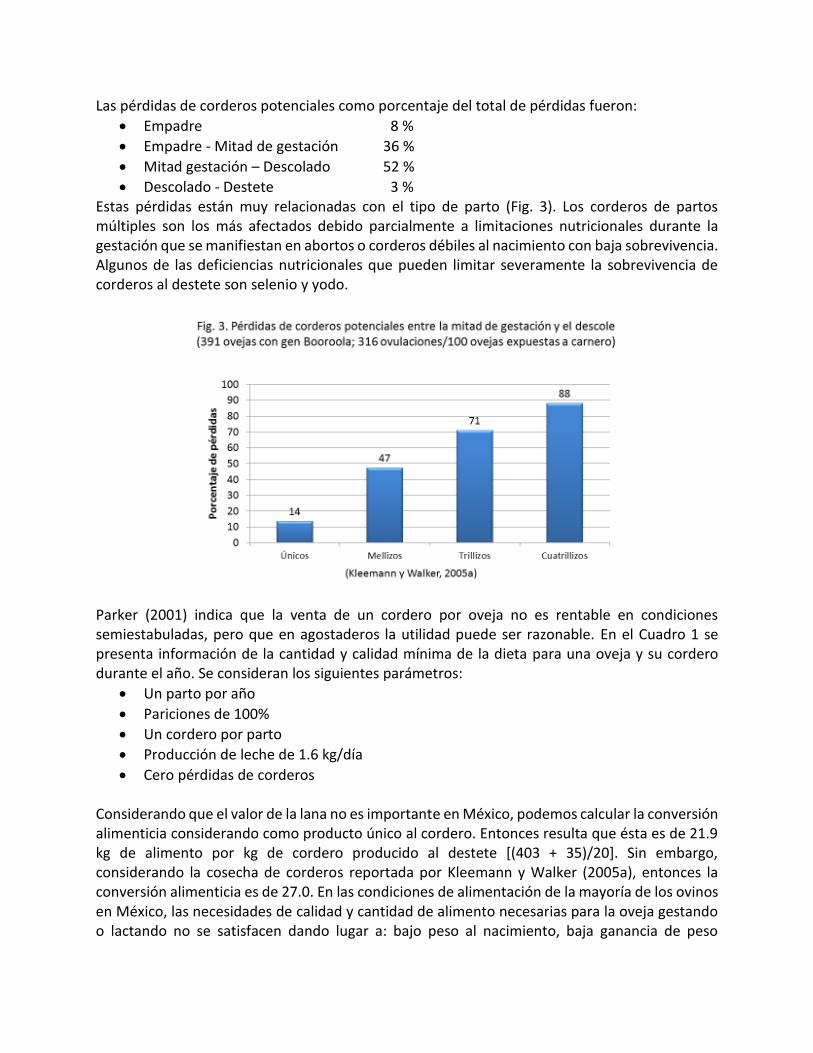
Las pérdidas de corderos potenciales como porcentaje del total de pérdidas fueron:
Empadre 8 %
Empadre - Mitad de gestación 36 %
Mitad gestación – Descolado 52 %
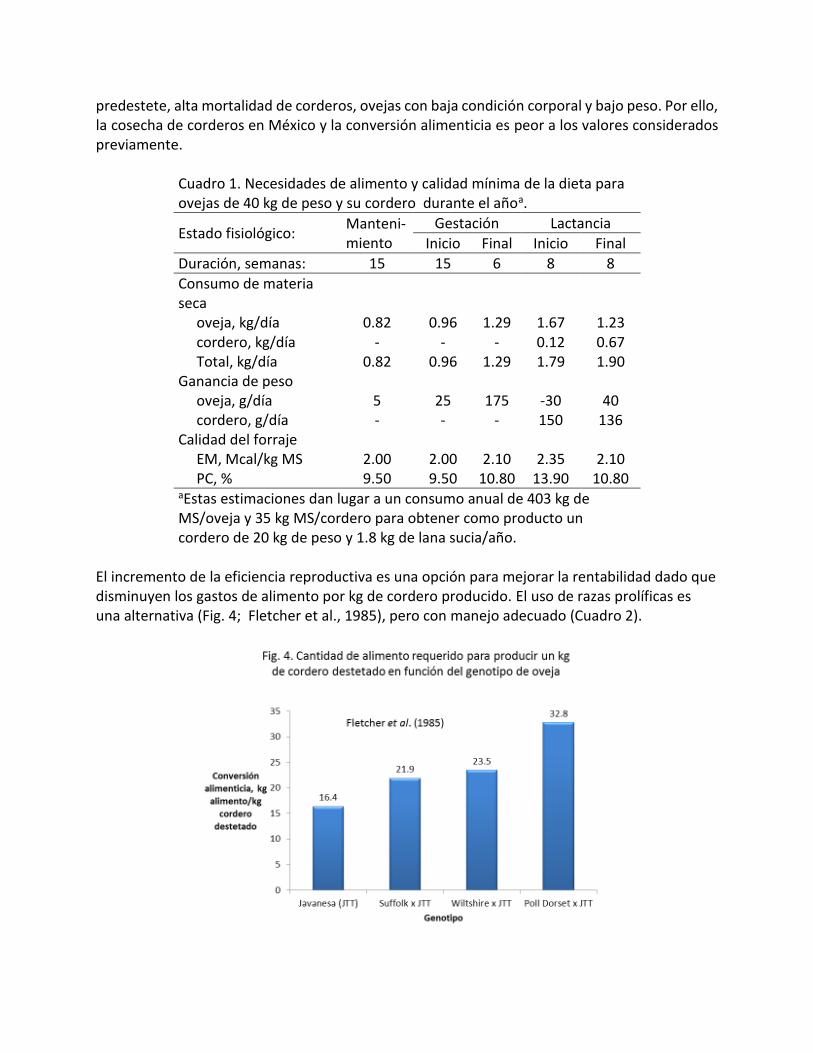
Descolado - Destete 3 % Estas pérdidas están muy relacionadas con el tipo de parto (Fig. 3). Los corderos de partos múltiples son los más afectados debido parcialmente a limitaciones nutricionales durante la gestación que se manifiestan en abortos o corderos débiles al nacimiento con baja sobrevivencia. Algunos de las deficiencias nutricionales que pueden limitar severamente la sobrevivencia de corderos al destete son selenio y yodo.
Parker (2001) indica que la venta de un cordero por oveja no es rentable en condiciones semiestabuladas, pero que en agostaderos la utilidad puede ser razonable. En el Cuadro 1 se presenta información de la cantidad y calidad mínima de la dieta para una oveja y su cordero durante el año. Se consideran los siguientes parámetros:
Un parto por año
Pariciones de 100%
Un cordero por parto
Producción de leche de 1.6 kg/día
Cero pérdidas de corderos Considerando que el valor de la lana no es importante en México, podemos calcular la conversión alimenticia considerando como producto único al cordero. Entonces resulta que ésta es de 21.9 kg de alimento por kg de cordero producido al destete [(403 + 35)/20]. Sin embargo, considerando la cosecha de corderos reportada por Kleemann y Walker (2005a), entonces la conversión alimenticia es de 27.0. En las condiciones de alimentación de la mayoría de los ovinos en México, las necesidades de calidad y cantidad de alimento necesarias para la oveja gestando o lactando no se satisfacen dando lugar a: bajo peso al nacimiento, baja ganancia de peso
predestete, alta mortalidad de corderos, ovejas con baja condición corporal y bajo peso. Por ello, la cosecha de corderos en México y la conversión alimenticia es peor a los valores considerados previamente.
Cuadro 1. Necesidades de alimento y calidad mínima de la dieta para ovejas de 40 kg de peso y su cordero durante el añoa.
Estado fisiológico: Manteni- miento
Gestación Lactancia
Inicio Final Inicio Final
Duración, semanas: 15 15 6 8 8
Consumo de materia seca
oveja, kg/día 0.82 0.96 1.29 1.67 1.23 cordero, kg/día - - - 0.12 0.67 Total, kg/día 0.82 0.96 1.29 1.79 1.90 Ganancia de peso oveja, g/día 5 25 175 -30 40 cordero, g/día - - - 150 136 Calidad del forraje EM, Mcal/kg MS 2.00 2.00 2.10 2.35 2.10 PC, % 9.50 9.50 10.80 13.90 10.80 aEstas estimaciones dan lugar a un consumo anual de 403 kg de MS/oveja y 35 kg MS/cordero para obtener como producto un cordero de 20 kg de peso y 1.8 kg de lana sucia/año.
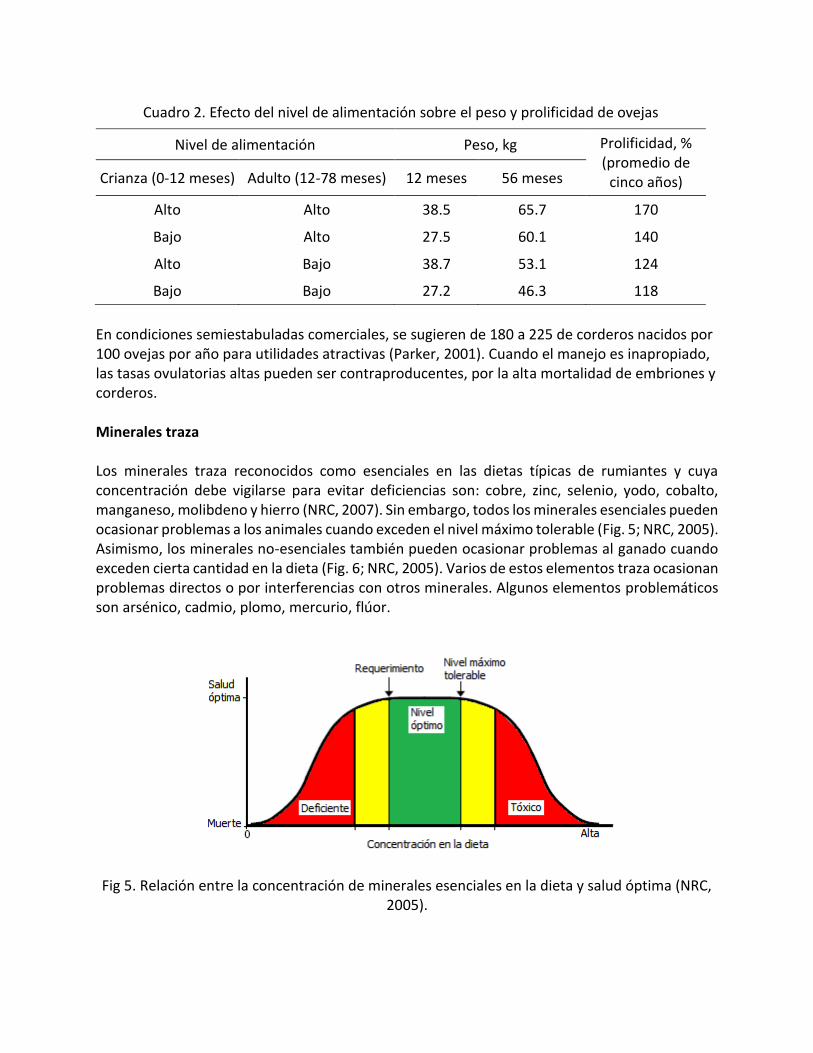
El incremento de la eficiencia reproductiva es una opción para mejorar la rentabilidad dado que disminuyen los gastos de alimento por kg de cordero producido. El uso de razas prolíficas es una alternativa (Fig. 4; Fletcher et al., 1985), pero con manejo adecuado (Cuadro 2).
Cuadro 2. Efecto del nivel de alimentación sobre el peso y prolificidad de ovejas
Nivel de alimentación Peso, kg Prolificidad, % (promedio de
cinco años) Crianza (0-12 meses) Adulto (12-78 meses) 12 meses 56 meses
Alto Alto 38.5 65.7 170
Bajo Alto 27.5 60.1 140
Alto Bajo 38.7 53.1 124
Bajo Bajo 27.2 46.3 118
En condiciones semiestabuladas comerciales, se sugieren de 180 a 225 de corderos nacidos por 100 ovejas por año para utilidades atractivas (Parker, 2001). Cuando el manejo es inapropiado, las tasas ovulatorias altas pueden ser contraproducentes, por la alta mortalidad de embriones y corderos. Minerales traza Los minerales traza reconocidos como esenciales en las dietas típicas de rumiantes y cuya concentración debe vigilarse para evitar deficiencias son: cobre, zinc, selenio, yodo, cobalto, manganeso, molibdeno y hierro (NRC, 2007). Sin embargo, todos los minerales esenciales pueden ocasionar problemas a los animales cuando exceden el nivel máximo tolerable (Fig. 5; NRC, 2005). Asimismo, los minerales no-esenciales también pueden ocasionar problemas al ganado cuando exceden cierta cantidad en la dieta (Fig. 6; NRC, 2005). Varios de estos elementos traza ocasionan problemas directos o por interferencias con otros minerales. Algunos elementos problemáticos son arsénico, cadmio, plomo, mercurio, flúor.
Fig 5. Relación entre la concentración de minerales esenciales en la dieta y salud óptima (NRC,
2005).
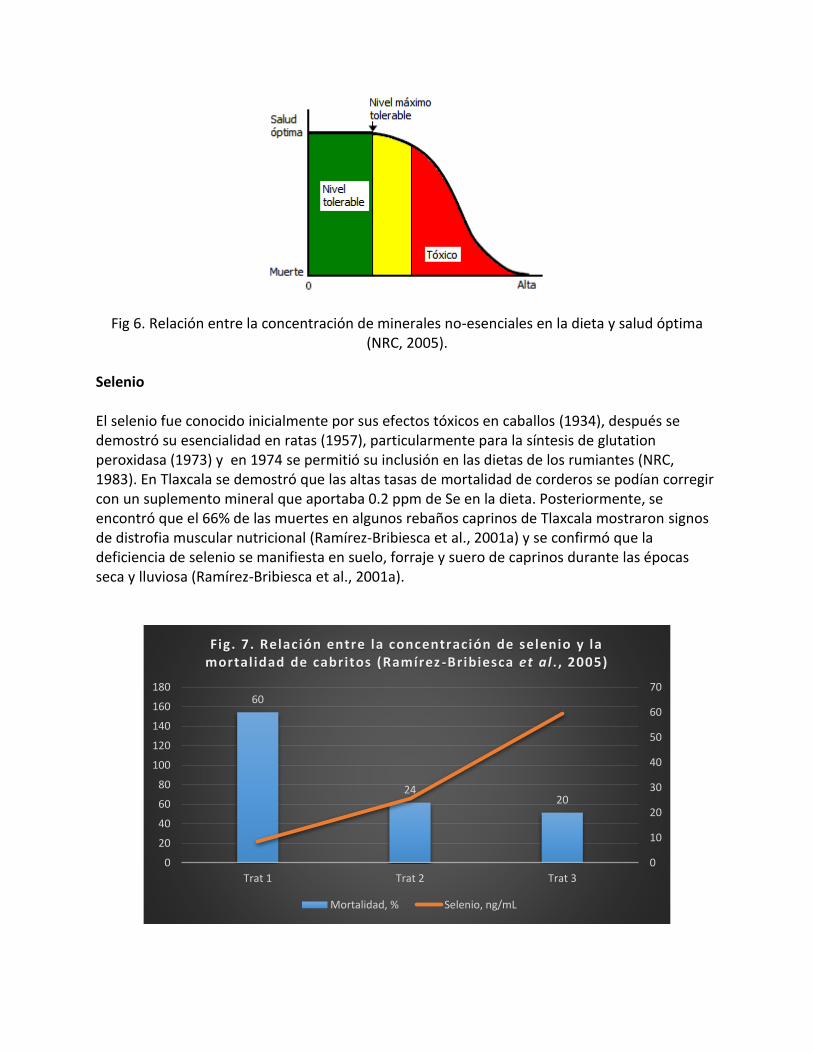
Fig 6. Relación entre la concentración de minerales no-esenciales en la dieta y salud óptima (NRC, 2005).
Selenio El selenio fue conocido inicialmente por sus efectos tóxicos en caballos (1934), después se demostró su esencialidad en ratas (1957), particularmente para la síntesis de glutation peroxidasa (1973) y en 1974 se permitió su inclusión en las dietas de los rumiantes (NRC, 1983). En Tlaxcala se demostró que las altas tasas de mortalidad de corderos se podían corregir con un suplemento mineral que aportaba 0.2 ppm de Se en la dieta. Posteriormente, se encontró que el 66% de las muertes en algunos rebaños caprinos de Tlaxcala mostraron signos de distrofia muscular nutricional (Ramírez-Bribiesca et al., 2001a) y se confirmó que la deficiencia de selenio se manifiesta en suelo, forraje y suero de caprinos durante las épocas seca y lluviosa (Ramírez-Bribiesca et al., 2001a).
60
2420
0
10
20
30
40
50
60
70
0
20
40
60
80
100
120
140
160
180
Trat 1 Trat 2 Trat 3
Fig. 7. Relación entre la concentración de selenio y la mortal idad de cabritos (Ramírez -Bribiesca et al . , 2005)
Mortalidad, % Selenio, ng/mL
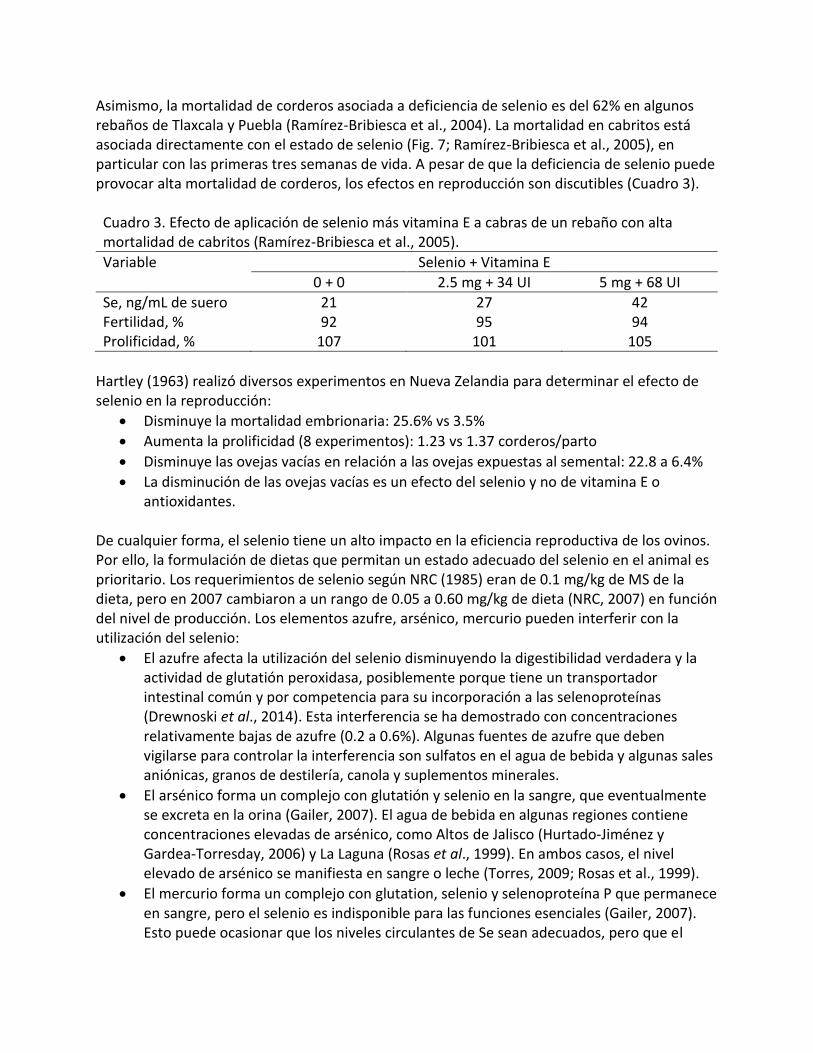
Asimismo, la mortalidad de corderos asociada a deficiencia de selenio es del 62% en algunos rebaños de Tlaxcala y Puebla (Ramírez-Bribiesca et al., 2004). La mortalidad en cabritos está asociada directamente con el estado de selenio (Fig. 7; Ramírez-Bribiesca et al., 2005), en particular con las primeras tres semanas de vida. A pesar de que la deficiencia de selenio puede provocar alta mortalidad de corderos, los efectos en reproducción son discutibles (Cuadro 3). Cuadro 3. Efecto de aplicación de selenio más vitamina E a cabras de un rebaño con alta mortalidad de cabritos (Ramírez-Bribiesca et al., 2005).
Variable Selenio + Vitamina E
0 + 0 2.5 mg + 34 UI 5 mg + 68 UI
Se, ng/mL de suero 21 27 42 Fertilidad, % 92 95 94 Prolificidad, % 107 101 105
Hartley (1963) realizó diversos experimentos en Nueva Zelandia para determinar el efecto de selenio en la reproducción:
Disminuye la mortalidad embrionaria: 25.6% vs 3.5%
Aumenta la prolificidad (8 experimentos): 1.23 vs 1.37 corderos/parto
Disminuye las ovejas vacías en relación a las ovejas expuestas al semental: 22.8 a 6.4%
La disminución de las ovejas vacías es un efecto del selenio y no de vitamina E o antioxidantes.
De cualquier forma, el selenio tiene un alto impacto en la eficiencia reproductiva de los ovinos. Por ello, la formulación de dietas que permitan un estado adecuado del selenio en el animal es prioritario. Los requerimientos de selenio según NRC (1985) eran de 0.1 mg/kg de MS de la dieta, pero en 2007 cambiaron a un rango de 0.05 a 0.60 mg/kg de dieta (NRC, 2007) en función del nivel de producción. Los elementos azufre, arsénico, mercurio pueden interferir con la utilización del selenio:
El azufre afecta la utilización del selenio disminuyendo la digestibilidad verdadera y la actividad de glutatión peroxidasa, posiblemente porque tiene un transportador intestinal común y por competencia para su incorporación a las selenoproteínas (Drewnoski et al., 2014). Esta interferencia se ha demostrado con concentraciones relativamente bajas de azufre (0.2 a 0.6%). Algunas fuentes de azufre que deben vigilarse para controlar la interferencia son sulfatos en el agua de bebida y algunas sales aniónicas, granos de destilería, canola y suplementos minerales.
El arsénico forma un complejo con glutatión y selenio en la sangre, que eventualmente se excreta en la orina (Gailer, 2007). El agua de bebida en algunas regiones contiene concentraciones elevadas de arsénico, como Altos de Jalisco (Hurtado-Jiménez y Gardea-Torresday, 2006) y La Laguna (Rosas et al., 1999). En ambos casos, el nivel elevado de arsénico se manifiesta en sangre o leche (Torres, 2009; Rosas et al., 1999).
El mercurio forma un complejo con glutation, selenio y selenoproteína P que permanece en sangre, pero el selenio es indisponible para las funciones esenciales (Gailer, 2007). Esto puede ocasionar que los niveles circulantes de Se sean adecuados, pero que el

animal esté deficiente. Las fuentes de mercurio pueden ser el agua de bebida, la harina de pescado y algunas vacunas.
Por estas interacciones, el requerimiento real de selenio para animales de algún rancho en particular se desconoce. Actualmente se tienen varias opciones para proveer selenio a los animales: selenio en forma orgánica e inorgánica, selenio oral o inyectable, diversas fuentes inorgánicas de selenio como selenito de sodio, selenato de potasio y selenato de bario, bolos de selenio. Algunos aspectos a considerar son:
1. La vida media del selenio en forma inorgánica en el animal es de 10 días (Conrad y Moxon, 1979).
2. El tiempo requerido para restaurar totalmente la actividad de glutation peroxidasa en los glóbulos rojos es ca. 120 días (Zachara et al., 1993).
3. La forma requerida por el animal para incorporar el selenio a las selenoproteínas es la inorgánica. Las formas orgánicas de selenio deben convertirse a inorgánicas.
4. La función del selenio como antioxidante es compartida con zinc, manganeso, cobre, vitamina E, hierro, vitamina A entre otros. Por ello, cuando alguno o varios de estos nutrimentos están deficientes, la necesidad de selenio se incrementa.
5. Los espermatozoides generan una cantidad elevada de oxidantes, así como el embrión y feto. Estos oxidantes ayudan a la función reproductiva, pero también pueden ocasionar daños severos. Por ello, se requiere un balance de todos los elementos.
6. El selenio es importante para transformar la tiroxina (T4) en la forma activa triyodotironina (T3), por lo cual una deficiencia de selenio puede provocar bocio.
Debe considerarse que la concentración de selenio en el agua de bebida puede ser alta en algunas regiones. En 125 pozos de la región de los Altos Jalisco es de 45.1 µg/L de agua (con un rango de 2 a 122; Hurtado y Gardea, 2007). Asimismo, las dietas pueden contener niveles altos de selenio (0.66 a 1.90 mg Se/kg MS), en algunos casos debido a fuentes minerales contaminadas. Zinc El zinc es un elemento cuya esencialidad fue descubierta cuando se inició la inclusión de pasta de soya en las dietas de los cerdos en sustitución de la harina de pescado, dado que las fuentes de proteína de origen animal contienen más zinc que las vegetales. La deficiencia se realza cuando las dietas tienen niveles altos de calcio, por la formación de complejos con fitatos que hacen indisponible al Zn. El zinc es necesario para la expresión o represión de genes, de tal manera que el potencial genético de los animales no se puede expresar completamente. La deficiencia provoca enanismo, hipogonadismo en hembras y machos, fetos deformes y fetos momificados (Apgar y Fitzgerald, 1985). En general, se ha demostrado que todas las etapas de la reproducción pueden ser afectadas por la deficiencia de Zn (Fig. 8; Apgar, 1992). La importancia práctica de la deficiencia de Zn en la reproducción a nivel de campo se desconoce, probablemente porque los requerimientos en condiciones normales son menores

que los contenidos en los forrajes y suplementos y que es un elemento muy controlado en el organismo por sistemas transportadores.
Fig. 8. Efectos de la deficiencia de zinc en la reproducción de animales. Leyenda: a = rata, b = ratón, c = conejillo de indias, d = hámster, e = cerdo, f = ovinos, g = mono Bonet, h = mono Rhesus macho, i = pollo. Cobre El cobre es un elemento con mala fama por los casos de intoxicación de ovinos reportados en México y otros países del mundo como el Reino Unido. La intoxicación se favorece porque los ovinos pueden absorber y retener más cobre que los bovinos > cerdos > aves. El cobre es muy reactivo y puede ayudar a controlar protozoarios, nematodos, hongos y otros microorganismos. Por ello, en aves y cerdos se utiliza como aditivo para mejorar la salud y el comportamiento productivo de los mismos. Las dosis utilizadas son de 30 a 40 veces el requerimiento. Dado que

la absorción del Cu es muy limitada, la mayoría del mismo aparece en las excretas de las aves y cerdos, en concentraciones que pueden representar hasta 100 veces el requerimiento de los ovinos. Cuando las excretas son proporcionadas a ovinos, particularmente corderos que tienen mayor tasa de absorción de este elemento que los animales adultos, la intoxicación y muerte de los animales aparece en corto tiempo porque el animal tiene capacidad limitada para almacenar Cu en el hígado como mecanismo de detoxificación. La deficiencia de cobre es la más común en México y el resto del mundo. Además, dado que las premezclas minerales para ovinos y alimentos comerciales no incluyen suficiente cobre, la deficiencia es cada vez más severa. Los elementos azufre, molibdeno, hierro, zinc, plomo, cadmio y plata interfieren con el metabolismo de selenio, como la absorción intestinal, retención en tejidos, excreción en orina y disponibilidad en tejidos. Esta interacción genera traslape en los requerimientos y máximos tolerables de cobre en la dieta: 1) cuando todos estos elementos y el cobre están balanceados en la dieta, no hay problema; 2) cuando los otros elementos están bajos en la dieta, el cobre se metabolizará mejor y puede haber intoxicación; y 3) cuando alguno o varios de los otros elementos están altos en la dieta, provocan deficiencia de cobre. En ovinos se ha reportado infertilidad, asociada en algunos casos con aborto de fetos pequeños muertos (Howell y Hall, 1970). Los abortos pueden estar asociados a poca actividad de la enzima diamino oxidasa, responsable de la destrucción de histamina (Maintz et al., 2008). Otros problemas son: - Disminuye tasa de concepción - Retraso del estro - Depresión del estro - Dificultades al parto - Retención de placentas - Muerte fetal - Falta de libido y espermatogénesis reducida. Manganeso La deficiencia de manganeso en condiciones prácticas de alimentación es poco común y el nivel máximo tolerable (2000 ppm; NRC, 2005) es muy alto en relación a las cantidades requeridas. Por otro lado, la determinación de manganeso en sangre es complicada. Sin embargo, niveles relativamente bajos de manganeso en la dieta (40 a 200 ppm) pueden provocar interferencia con el metabolismo del hierro cuando hay deficiencia de cobre. Además, niveles altos de manganeso en sangre están asociados con la manifestación de enfermedades neurodegenerativas como scrapie (Bowman et al., 2011). Algunos reportes del efecto de la deficiencia de manganeso en ovinos son:
Retraso o depresión del estro y concepción.
Incremento de abortos.
Menor peso al nacer.
Yodo El yodo es requerido para la síntesis de las hormonas Tiroxina (T4) y Triyodotironina (T3), hormonas muy importantes en el metabolismo de los animales. Este elemento tiene un impacto tremendo en la eficiencia reproductiva (Cuadro 4): concepción, abortos y sobrevivencia de los corderos.
Cuadro 4. Comportamiento reproductivo de ovinos y caprinos
suplementados con yodo (Tomado de Minson, 1990).
Variable Caprinos Ovinos
Testigo Yodo Testigo Yodo
Éxito de la 1ª inseminación 27 73 - -
Tasa final de concepción, % 79 83 87 100
Abortos, % 47 0 15 0
Corderos con bocio, % - - 100 0
Corderos sobrevivientes 1ª sem., % - - 50 72
Este elemento ha sido estudiado ampliamente en humanos, porque la deficiencia de yodo en los alimentos es normal y provoca bocio. Para disminuir el problema, se decretó que la sal de mesa debe estar yodatada. En animales, el problema se agrava por el consumo de forrajes con sustancias inhibidoras del metabolismo de yodo (Cuadro 5). Cuadro 5. Efecto de la deficiencia de yodo en ovejas sobre el peso al nacer, peso de tiroides y
sobrevivencia de sus corderos (Tomado de Minson, 1990).
Peso al nacer, kg Tiroides, g Sobrevivencia, %
Especie Testigo Yodo Testigo Yodo Testigo Yodo
Brassica oleracea 4.0 4.8 a - 83 87
3.5 4.2 17 3 22 82
- - 10 2 41 90
- - 23 6 - -
Cynodon aethiopicus 2.7 3.6 36 2 30 75
Medicago sativa 3.5 3.9 - - 64 84
Promedio 3.4 4.1 21 3 48 84
Sargison y West (1998) consideran la deficiencia de yodo como un problema emergente porque algunas variedades de forraje liberadas “recientemente” tienen alto contenido de sustancias bociogénicas (Cuadro 6).

Cuadro 6. Magnitud del problema de la deficiencia de yodo (Sargison y West, 1998)
Rebaño Mortalidad perinatal, % Peso tiroides/Peso vivo, g/kg
A 25 0.42
B 50 1.14
C 66 8.50
Los signos clínicos de la deficiencia de yodo en fetos son:
Menor peso
Ausencia de crecimiento de lana
Bocio
Deformaciones del esqueleto
Grados variables de subluxaciones en uniones de las patas
Menor maduración del hueso
Retraso de desarrollo del cerebro En animales adultos se presentan los siguientes problemas:
El desarrollo fetal se puede detener en cualquier etapa dando lugar a muerte fetal temprana y resorción, abortos y natimortos, crías débiles, sin pelo o lana.
Gestaciones y partos prolongados
Retención de membranas fetales
Supresión de estros
Estros irregulares
Mayores servicios por concepción
Menor libido en sementales
Menor calidad del semen Cobalto El cobalto es utilizado por los microorganismos del rumen para sintetizar vitamina B12. Por ello, las funciones del cobalto corresponden a las de la vitamina B12. Este elemento ha sido poco estudiado en México, a pesar de que la deficiencia fue descubierta en ovinos desde 1935 en Australia (Suttle, 2010). La deficiencia de cobalto impide el comportamiento reproductivo. Esto se debe a que los animales pierden demasiado peso y condición corporal, factores que explican 39.3 y 12.7% de la variación en tasa de ovulación. Las ovulaciones por 100 ovejas se incrementaron en 1.8 por kilogramo de incremento de peso vivo al empadre en 68 rebaños (Kleeman y Walker, 2005b). La utilización del ácido propiónico, producto de la digestión de carbohidratos de la dieta en el rumen, depende de la vitamina B12. Dado que el propionato es el principal precursor de la glucosa en el cuerpo, todos los procesos en que se involucra la glucosa se ven afectados, como síntesis de ADN y ARN, síntesis de grasa y síntesis de lactosa. El ácido propiónico que no se

metaboliza se acumula y se excreta en orina como ácido metilmalónico. El ácido propiónico acumulado inhibe el consumo de alimento, que a su vez provoca mobilización de reservas corporales. Literatura citada Apgar, J. 1992. Zinc and reproduction: an update. J. Nutritional Biochemistry 2: 266. Apgar, J. and J. A. Fitzgerald. 1985. Effect on the ewe and lamb of low zinc intake throughout pregnancy.
Journal of Animal Science 60:1530 – 1538. Bedwal, R. S. and A. Bahuguna. 1994. Zinc, copper and selenium in reproduction. Experientia 50(7): 62-
64. Bowman, A. B., G. F. Kwakyea, E. Herrero Hernández, M. Aschner. 2011. Role of manganese in
neurodegenerative diseases. Journal of Trace Elements in Medicine and Biology 25: 191– 203. Conrad, H. R. and A. L. Moxon. 1979. Transfer of dietary selenium to milk. J. Dairy Sci. 62:404. Cruz, H. E. 2009. Suplementación de cobre a ovejas criollas. Tesis Maestría en Ciencias, Universidad
Autónoma Chapingo, Chapingo, México. 81 p. Drewnoski, M. E., D. J. Pogge and S. L. Hansen. 2014. High-sulfur in beef cattle diets: A review. J. Anim.
Sci. 2014.92:3763–3780 Gailer, J. 2007. Arsenic-selenium and mercury-selenium bonds in biology. A Review. Coordination
Chemistry Reviews 251: 234-254. Hartley, W. J. 1963. Selenium and ewe fertility. Proceedings of the New Zealand Society of Animal
Production 23: 20 - 27. Hurtado-Jiménez, R. y J. Gardea Torresdey. 2007. Evaluación de la exposición a selenio en los Altos de
Jalisco, México. Salud Pública de México. 49(4): 312-315. Hurtado-Jiménez, R., and J. L. Gardea-Torresdey. 2006. Arsenic in drinking water in the Los Altos de
Jalisco region of Mexico. Revista Panamericana de Salud Pública 20(4): 236-247. Kleemann, D. O, and S. K. Walker. 2005a. Fertility in South Australian commercial Merino flocks: sources
of reproductive wastage. Theriogenology 63: 2075-2088. Kott, R. 2006. Montana Farm Flock Sheep Production Handbook. Montana State University.
http://animalrangeextension.montana.edu/articles/sheep/Flock%20Handbook/Handbook-TOC.htm. Consultada el 12 de septiembre de 2014.
Madera López, M. 2009. Suplementación de selenio en corderos destetados. Tesis Maestría en Ciencias, Universidad Nacional Autónoma de México, Cuautitlán Izcalli, México.
Maintz, L., V. Schwarzer, T. Bicher, K. van der Ven and N. Novak. 2008. Effect of histamine and diamine oxidase activities on pregnancy: a critical review. Human Reproduction Update 14(5): 485-495
NRC. 2005. Mineral Tolerance of Animals. 2nd Rev. Ed. The National Academies Press, Washington, D.C. 510 p.
NRC. 2007. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids. The National Academies Press, Washington, D.C. 362 p.
Parker, R. 2001. The Sheep Book. A Handbook for the Modern Shepherd. Ohio University Press. Athens, OH. 321 p.
Qureshi, I. Z. and Q. Abbas. 2013. Modulation of testicular and whole blood trace element concentrations in conjunction with testosterone release following kisspeptin administration in male rabbits (Oryctolagus cuniculus). Biological Trace Element Research 154(2): 210-216.
Ramírez-Bribiesca, E., E. Hernández Camacho, L. M. Hernández Calva y J. L. Tórtora Pérez. 2004. Efecto de un suplemento parenteral con selenito de sodio en la mortalidad de corderos y los valores hemáticos de selenio. Agrociencia 38(1): 43-51.

Ramírez-Bribiesca, J. E. J. L. Tórtora, M. Huerta, L. M. Hernández, R. López, M. M. Crosby. 2005. Effect of selenium-vitamin E injection in selenium-deficient dairy goats and kids on the Mexican plateau. Arq. Bras. Med. Vet. Zootec. 57(1):77-84.
Ramírez-Bribiesca, J. E., J. L. Tórtora, M. Huerta, A. Aguirre, L. M. Hernández. 2001b. Diagnosis of selenium status in grazing dairy goats in the Mexican plateau. Small Ruminant Research 41:81-85.
Ramírez-Bribiesca, J. E., J. L. Tórtora, L. M. Hernández, and M. Huerta. 2001a. Main causes of mortalities in dairy goat kids from the Mexican plateau. Small Ruminant Research 41:77-80.
Rosas, I., R. Belmont, A. Armienta and A. Baez. 1999. Arsenic concentrations in water, soil, milk and forage in Comarca Lagunera, México. Water, Air, and Soil Pollution 112: 133–149.
Sargison, N. D. and D. M. West. 1998. Iodine deficiency: an emerging problem in New Zealand sheep flocks? Proceedings of the New Zealand Society of Animal Production 58:202-204.
Suttle, N. F. 2010. Mineral Nutrition of Livestock. 4th Ed. CABI. Oxfordshire, UK. 587 p. Torres Lechuga, M. E. 2013. Diagnóstico mineral de unidades de producción ovina en Tepatitlán, Jalisco.
Tesis Maestría en Ciencias, Universidad Autónoma Chapingo, Chapingo, México. 49 p. Vázquez-Armijo, J. F. R. Rojo, A. Z. M. Salem, D. López, J. L. Tinoco, A. González, N. Pescador and I. A.
Domínguez-Vara. 2011. Trace elements in sheep and goats reproduction: a review. Tropical and Subtropical Agroecosystems, 14: 1 – 13.
Zachara, B. A. U. Trafikowska, H. Labedzka, and J. Mikolajczak. 1993. Effect of dietary Se intake on blood Se levels and glutathione peroxidase activities in lambs. Small Ruminant Research 9(4): 331-340.





























