Embed Size (px)
Citation preview

G. M. Toffolo
Bressanone GNB 2010 1
Gianna Maria Toffolo, Alberto Corradin, Barbara Di Camillo
Dipartimento di Ingegneria dell’Informazione
Università di Padova
Modelli deterministici e modelli stocastici per la Molecular Systems Biology
1
Sommario
• Formulazione di modelli deterministici (ODE) della
trascrizione, traduzione e degradazione
• Modelli per descrivere effetti stocastici
• Esempio: circuito genico del retrovirus HTLV1
2
G. M. Toffolo
Bressanone GNB 2010 2
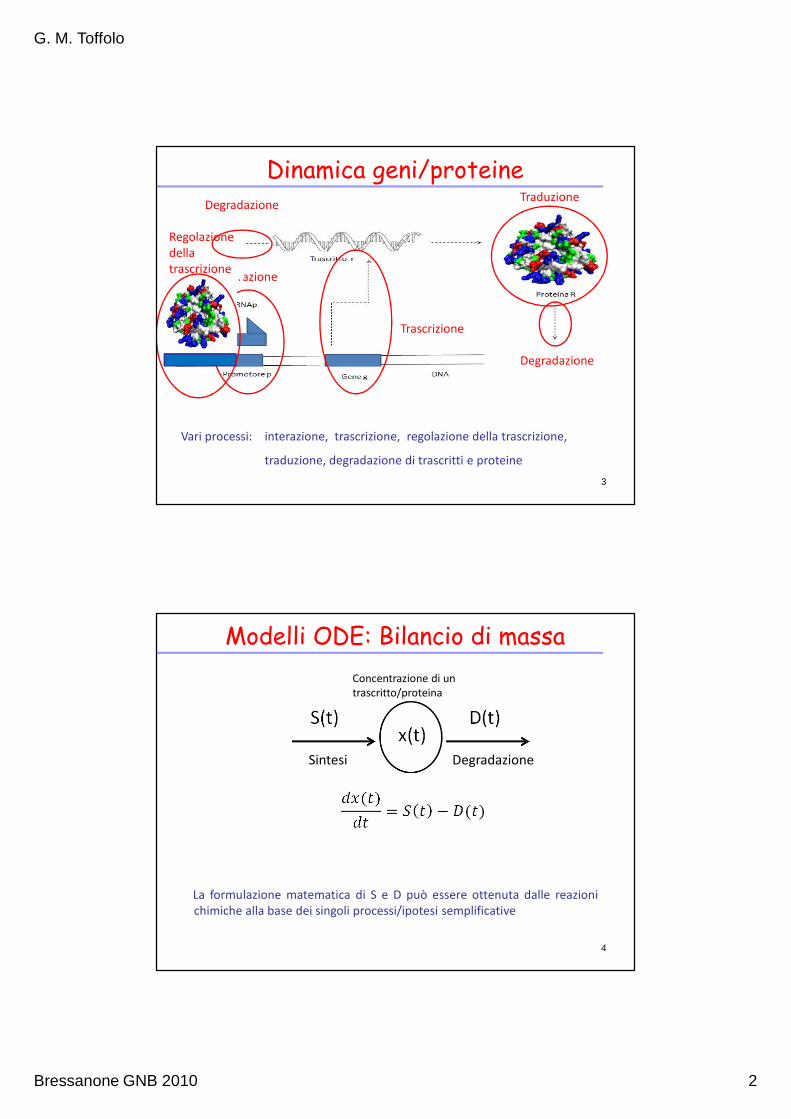
Dinamica geni/proteine
Vari processi: interazione, trascrizione, regolazione della trascrizione,
traduzione, degradazione di trascritti e proteine
Interazione
Trascrizione
Traduzione
Regolazione
della
trascrizione
Degradazione
Degradazione
3
Modelli ODE: Bilancio di massa
La formulazione matematica di S e D può essere ottenuta dalle reazioni
chimiche alla base dei singoli processi/ipotesi semplificative
Sintesi Degradazione
Concentrazione di un
trascritto/proteina
4

G. M. Toffolo
Bressanone GNB 2010 3
Reazioni chimiche: Legge di azione di massa
k
A + B -> C
A, B, C = specie chimiche
k = rate constant [1/tempo]
Legge di azione di massa
d[C]/dt= k∙[A]∙[B]
5C.M. Guldberg and P. Waage, Videnskabs-Selskabet i Christiana , 1864
Reazione reversibile: Equilibrio chimicokf
A + B ⇌ C
kr
kf = forward rate constant
kr = reverse rate constant
All’equilibrio:
kf ∙[A]∙[B]= kr ∙[C]
[C]= kf /kr∙[A]∙[B]=K∙[A]∙[B]
K= kf /kr = costante d’equilibrio adimensionale
6
G. M. Toffolo
Bressanone GNB 2010 4
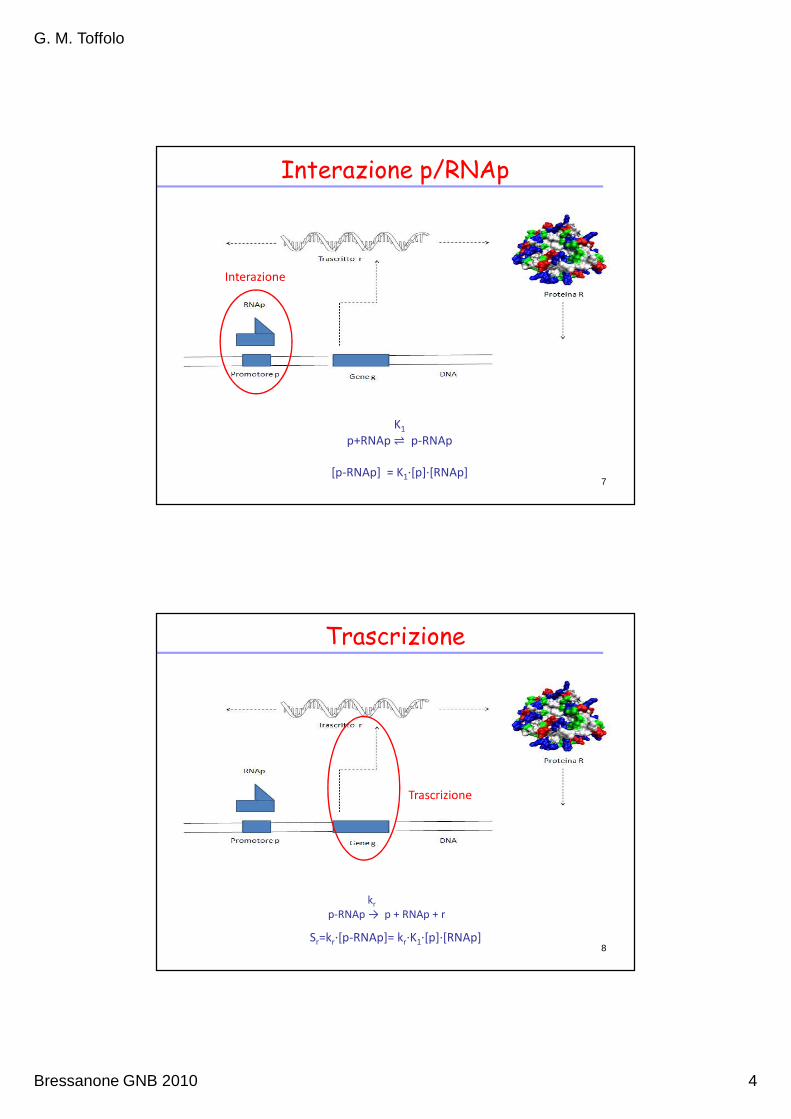
Interazione p/RNAp
K1
p+RNAp ⇌ p-RNAp
[p-RNAp] = K1∙[p]∙[RNAp]
Interazione
7
Trascrizione
Sr=kr∙[p-RNAp]= kr∙K1∙[p]∙[RNAp]
Trascrizione
8
kr
p-RNAp → p + RNAp + r

G. M. Toffolo
Bressanone GNB 2010 5
Regolazione della trascrizione
K2
p+FT⇌ p-FT
K3
p-FT + RNAp ⇌ p-FT-RNAp
ckr
p-FT-RNAp → p + FT + RNAp + r
SrFT= c∙kt ∙K2 ∙K3 ∙[p] ∙[FT] ∙[RNAp]
Regolazione
della
trascrizione
9
Trascrizione totale
Srtot(t)= Sr(t)+Sr
FT(t)
[p]= costante → competizione
Regolazione
della
trascrizione
10
Trascrizione
G. M. Toffolo
Bressanone GNB 2010 6
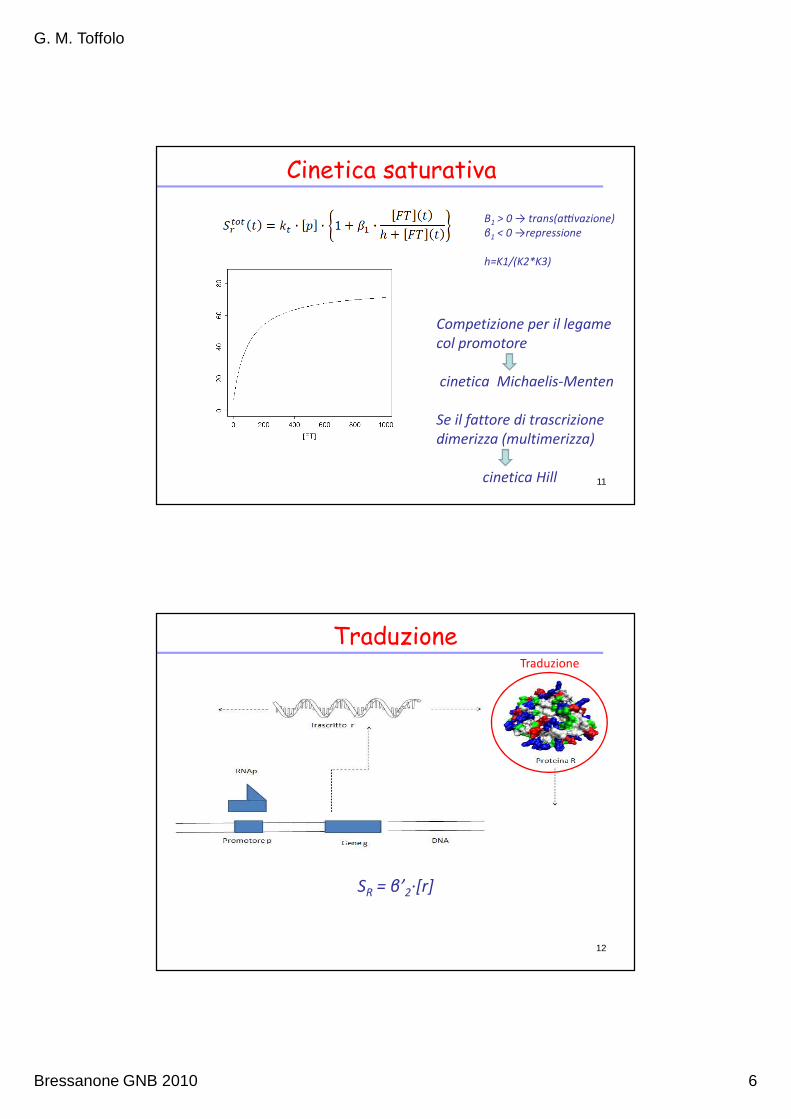
Cinetica saturativa
Competizione per il legame
col promotore
cinetica Michaelis-Menten
Se il fattore di trascrizione
dimerizza (multimerizza)
cinetica Hill 11
Β1 > 0 → trans(a:vazione)
β1 < 0 →repressione
h=K1/(K2*K3)
Traduzione
SR = β’2∙[r]
Traduzione
12
G. M. Toffolo
Bressanone GNB 2010 7
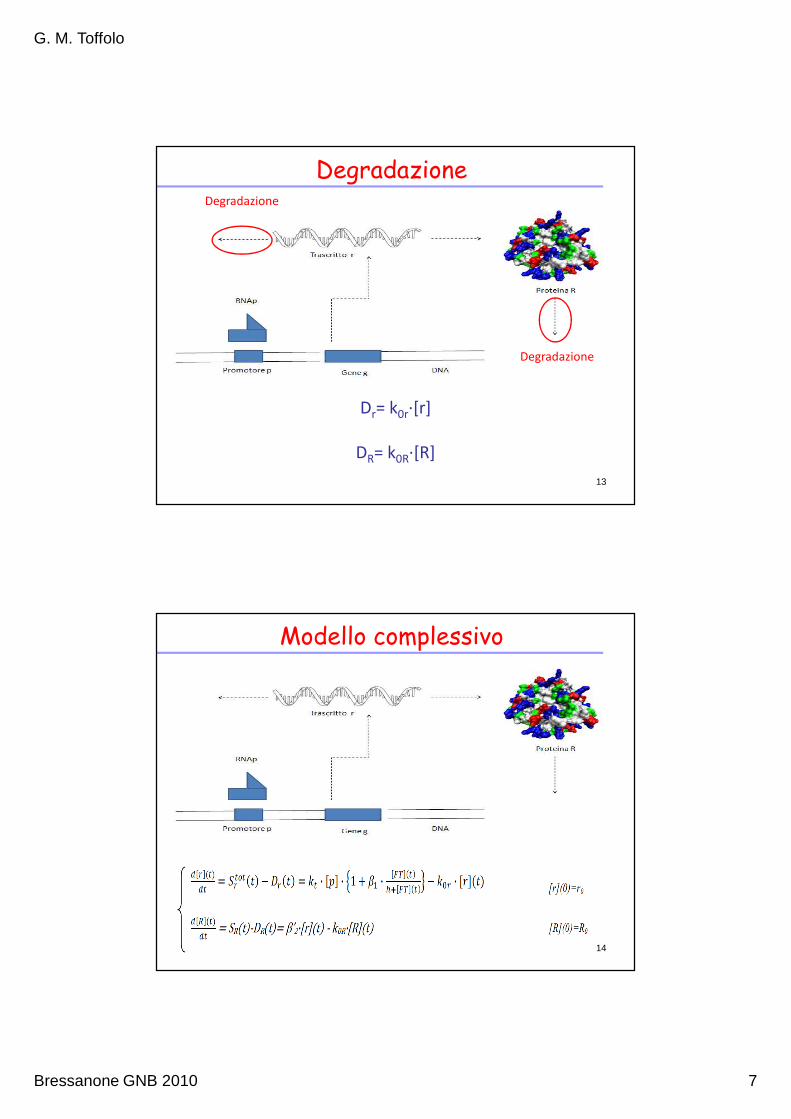
Degradazione
Degradazione
Degradazione
Dr= k0r∙[r]
DR= k0R∙[R]
13
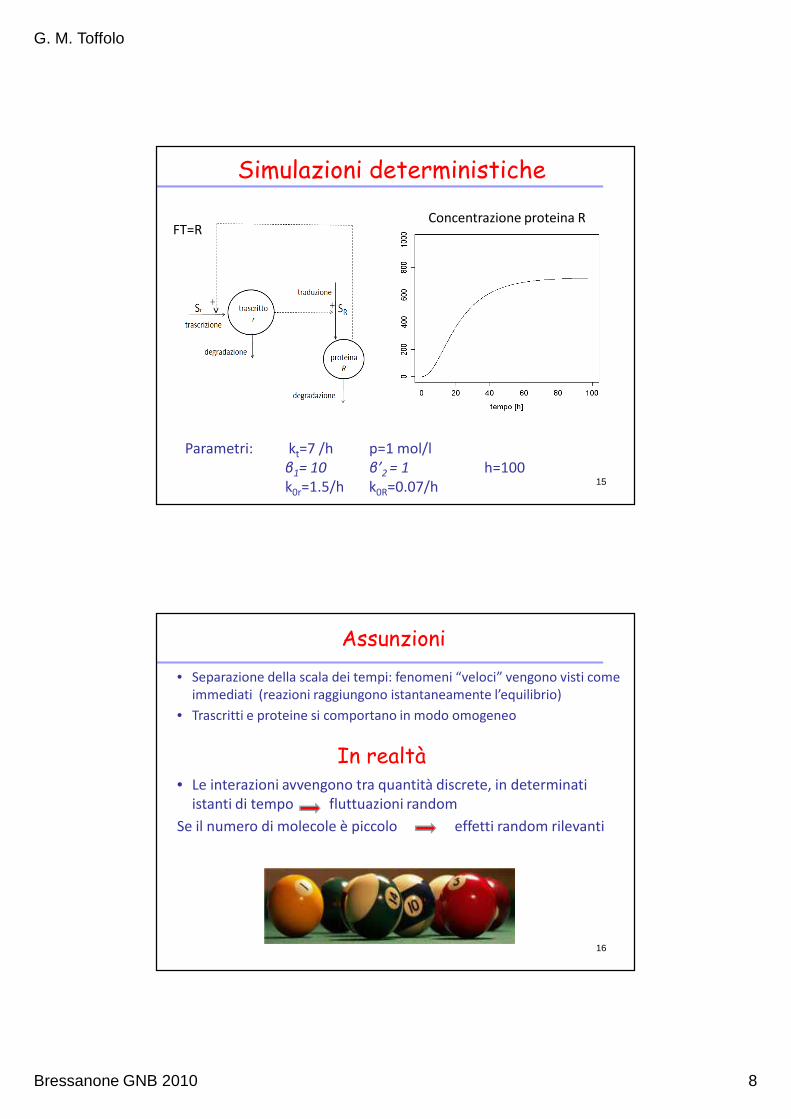
Modello complessivo
14
G. M. Toffolo
Bressanone GNB 2010 8
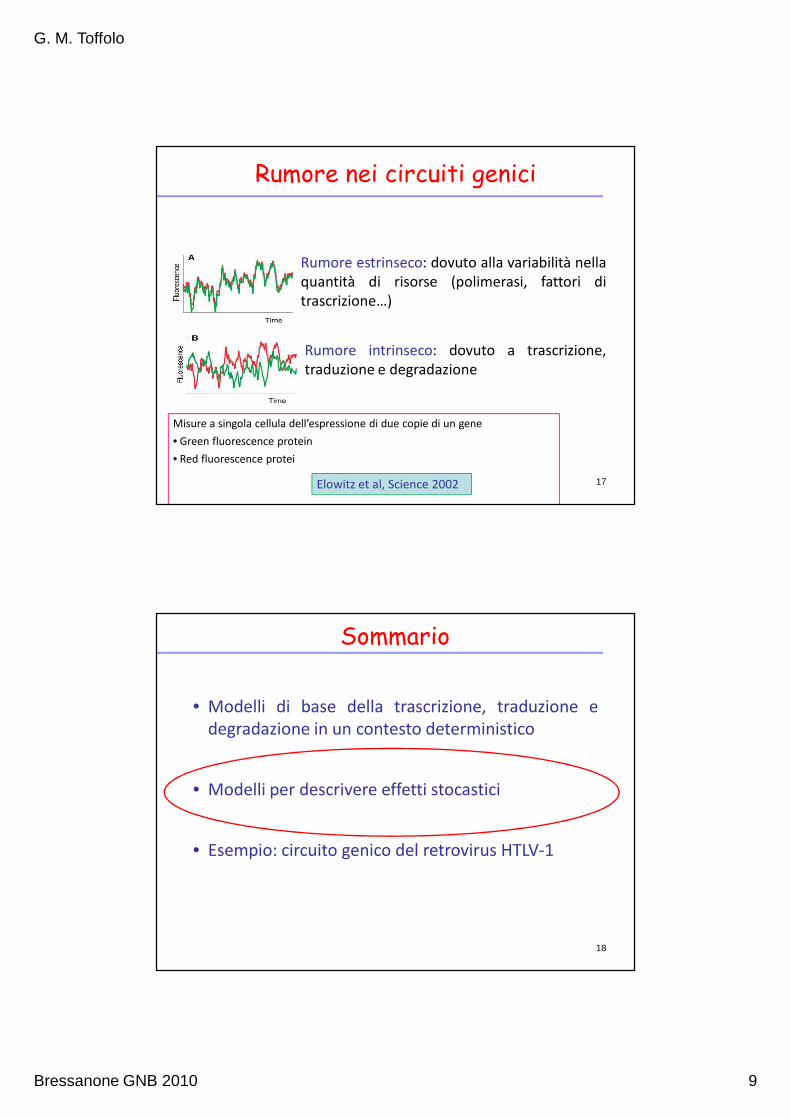
Simulazioni deterministiche
Parametri: kt=7 /h p=1 mol/l
β1= 10 β’2 = 1 h=100
k0r=1.5/h k0R=0.07/h 15
FT=RConcentrazione proteina R
Assunzioni
• Le interazioni avvengono tra quantità discrete, in determinati
istanti di tempo fluttuazioni random
Se il numero di molecole è piccolo effetti random rilevanti
16
• Separazione della scala dei tempi: fenomeni “veloci” vengono visti come
immediati (reazioni raggiungono istantaneamente l’equilibrio)
• Trascritti e proteine si comportano in modo omogeneo
In realtà
G. M. Toffolo
Bressanone GNB 2010 9
Rumore nei circuiti genici
Misure a singola cellula dell’espressione di due copie di un gene
• Green fluorescence protein
• Red fluorescence protei
Rumore estrinseco: dovuto alla variabilità nella
quantità di risorse (polimerasi, fattori di
trascrizione…)
Rumore intrinseco: dovuto a trascrizione,
traduzione e degradazione
17Elowitz et al, Science 2002
Sommario
• Modelli di base della trascrizione, traduzione e
degradazione in un contesto deterministico
• Modelli per descrivere effetti stocastici
• Esempio: circuito genico del retrovirus HTLV-1
18

G. M. Toffolo
Bressanone GNB 2010 10
Modello stocastico : una reazionek
Reazione R: X1 + X2 → C
x=[x1, x2] = vettore di stato (n° molecole) processo aleatorio bivariato
h(x) = funzione di rischio prop n° possibili combinazioni
x1∙ x2
Se R avviene: x→ x + + + + νννν
19
Wilkinson DJ , CRC Press, 2006
Modello stocastico : M reazioni
N specie chimiche X1…XN
x(t)= [X1(t),…,XN(t)] : vettore di stato al tempo t. Processo aleatorio
multivariato. La sua evoluzione temporale dipende da:
• qual è la prossima reazione
• quando essa avviene
M reazioni chimiche R1…RM
hj(x(t))= funzione di rischio per la reazione Ri
γi = vettore di cambiamento di stato della reazione Ri
Chemical Master Equation (CME): Facile da scrivere ma difficile
da risolvere
20

G. M. Toffolo
Bressanone GNB 2010 11
Due approcci
Gillespie algorithm: Simulazioni numeriche
Rumore additivo: Funzioni di trasferimento
21
Simulazioni numeriche basate sul campionamento:
• della prossima reazione (sulla base delle funzioni di rischio)
• dell’istante in cui avverrà (il numero di eventi nell’intervallo
[0,t] è un processo di Poisson)
Algoritmo di Gillespie
22
Poisson distribution
Gillespie DT J Comput Physics, 1976
G. M. Toffolo
Bressanone GNB 2010 12
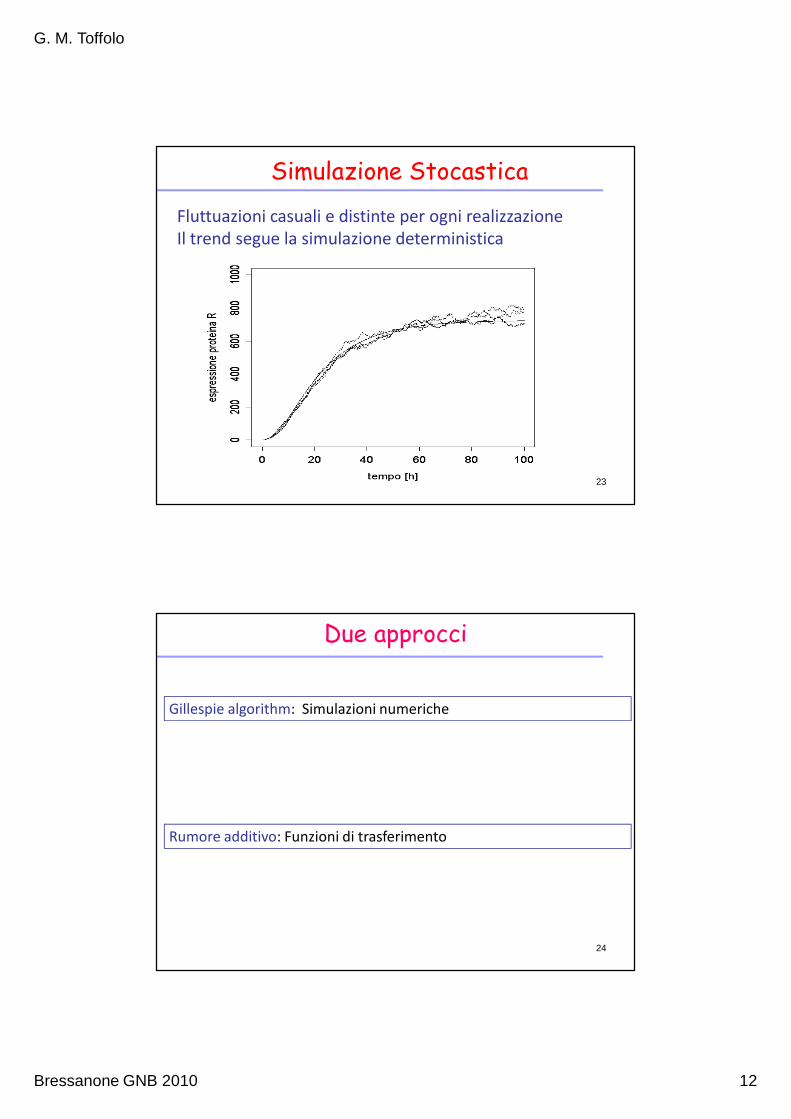
Simulazione Stocastica
Fluttuazioni casuali e distinte per ogni realizzazione
Il trend segue la simulazione deterministica
23
Due approcci
Gillespie algorithm: Simulazioni numeriche
Rumore additivo: Funzioni di trasferimento
24

G. M. Toffolo
Bressanone GNB 2010 13
Rumore additivo “in ingresso”
Sorgenti di rumore “shot” in trascrizione, traduzione e
degradazione.
Ipotesi:
• sistema lineare (o linearizzato)
• rumore in degradazione scorrelato da rumore nella
sintesi
Shot noise:
• processo di Poisson a campioni scorrelati
• PSD(f) =costante= 2*turn-over
25Simpson et al, Proc Natl Acad Sci, 2003
Funzioni di trasferimento
Sorgenti
introdotte
Rumore da
stimare
26
sistema linearizzato nell’intorno
dello stato stazionario
G. M. Toffolo
Bressanone GNB 2010 14
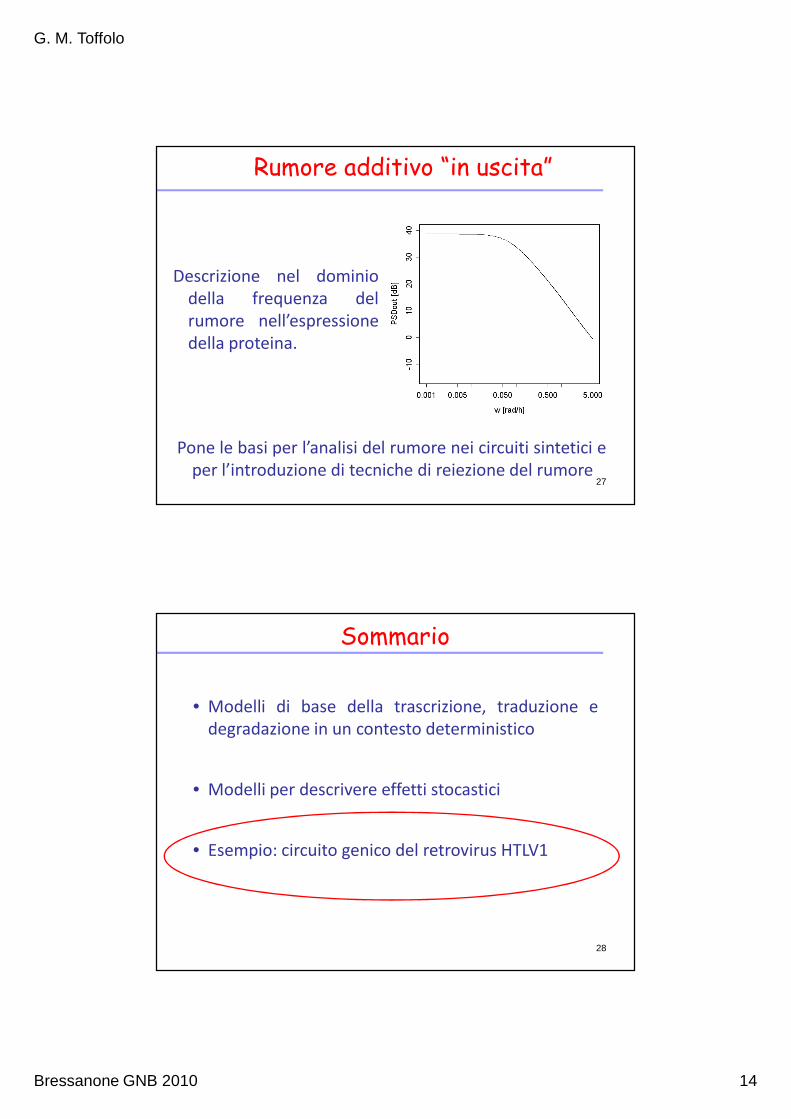
Rumore additivo “in uscita”
Descrizione nel dominio
della frequenza del
rumore nell’espressione
della proteina.
27
Pone le basi per l’analisi del rumore nei circuiti sintetici e
per l’introduzione di tecniche di reiezione del rumore
Sommario
• Modelli di base della trascrizione, traduzione e
degradazione in un contesto deterministico
• Modelli per descrivere effetti stocastici
• Esempio: circuito genico del retrovirus HTLV1
28
G. M. Toffolo
Bressanone GNB 2010 15
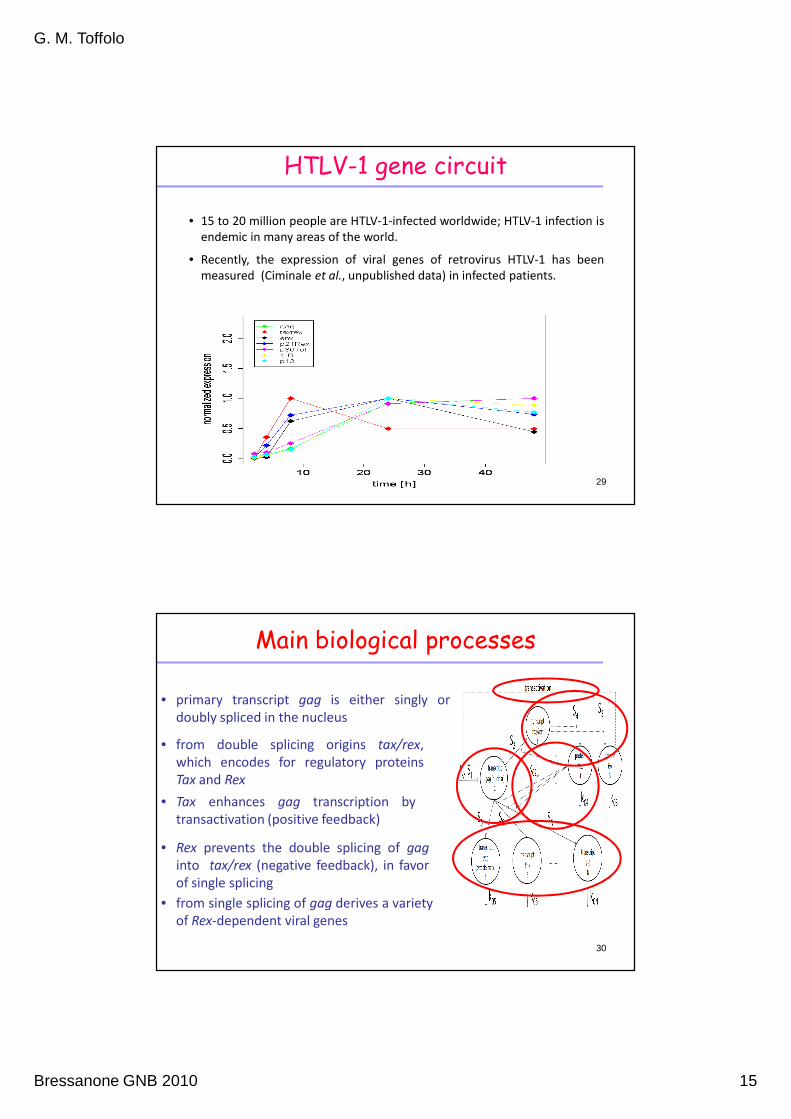
HTLV-1 gene circuit
• 15 to 20 million people are HTLV-1-infected worldwide; HTLV-1 infection is
endemic in many areas of the world.
• Recently, the expression of viral genes of retrovirus HTLV-1 has been
measured (Ciminale et al., unpublished data) in infected patients.
29
Main biological processes
• primary transcript gag is either singly or
doubly spliced in the nucleus
• from double splicing origins tax/rex,
which encodes for regulatory proteins
Tax and Rex
• Tax enhances gag transcription by
transactivation (positive feedback)
• Rex prevents the double splicing of gag
into tax/rex (negative feedback), in favor
of single splicing
• from single splicing of gag derives a variety
of Rex-dependent viral genes
30

G. M. Toffolo
Bressanone GNB 2010 16
Chemical kinetics: From reactions Chemical kinetics: From reactions to equationsto equations
• Syntheses are due to several interactions
• Each interaction is interpreted by a chemical reaction
• From reactions to mathematical formulations by mass action law
Promotor Gene
FT and e co-activator
31
Minimal rate of transcription SMinimal rate of transcription S1111(t)(t)• Several chemical equilibria: interactions with HTLV-1 promoter of TFs
NTPrS= Non-transactivated Promoter Site
• Irreversible reaction of transcription
• Mathematical formulation Macroparameter S
32

G. M. Toffolo
Bressanone GNB 2010 17
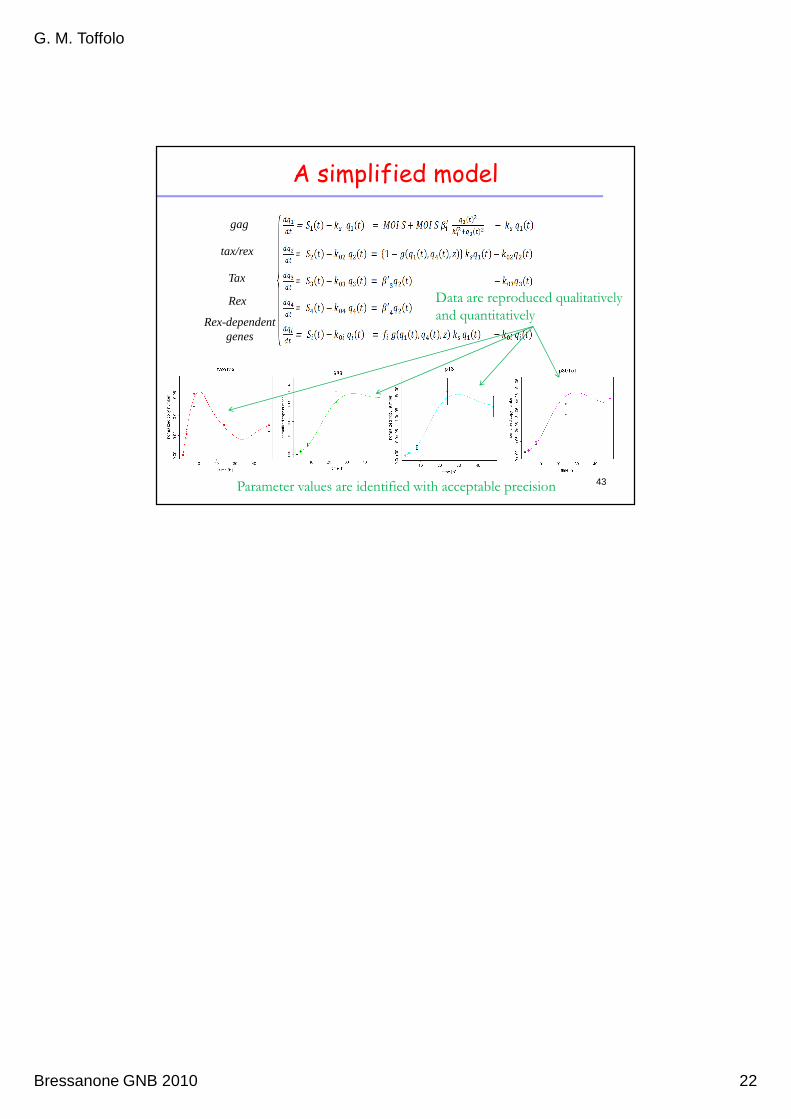
The model
gag
tax/rex
Tax
Rex
Rex-dependent genes
33
Parameter values
Data are reproduced qualitatively and quantitatively
34
• The model is not a priori uniquely identifiable from available data
• Some parameters were fixed to nominal values
• The model becomes uniquely dentifiable
• The remaining parameters were estimated from data with acceptable
precision

G. M. Toffolo
Bressanone GNB 2010 18
Is HTLV-1 a potential synthetic oscillator for eukaryotes ?
• The kinetics of HTLV-1 viral gene tax/rex are
characterized by positive and negative
feedback phenomena as synthetic oscillators
delivered into prokariotes
• Retroviruses, like HTLV-1, can penetrate into
the nucleus, where synthetic gene circuits
have to be delivered in eukaryotes
35
Tunable system parameters
• Protein Tax degradation, by
modulating ubiquitination
• MOI=n° of viral genomes
integrated in the host
genome, regulated by virus
delivery
36
G. M. Toffolo
Bressanone GNB 2010 19
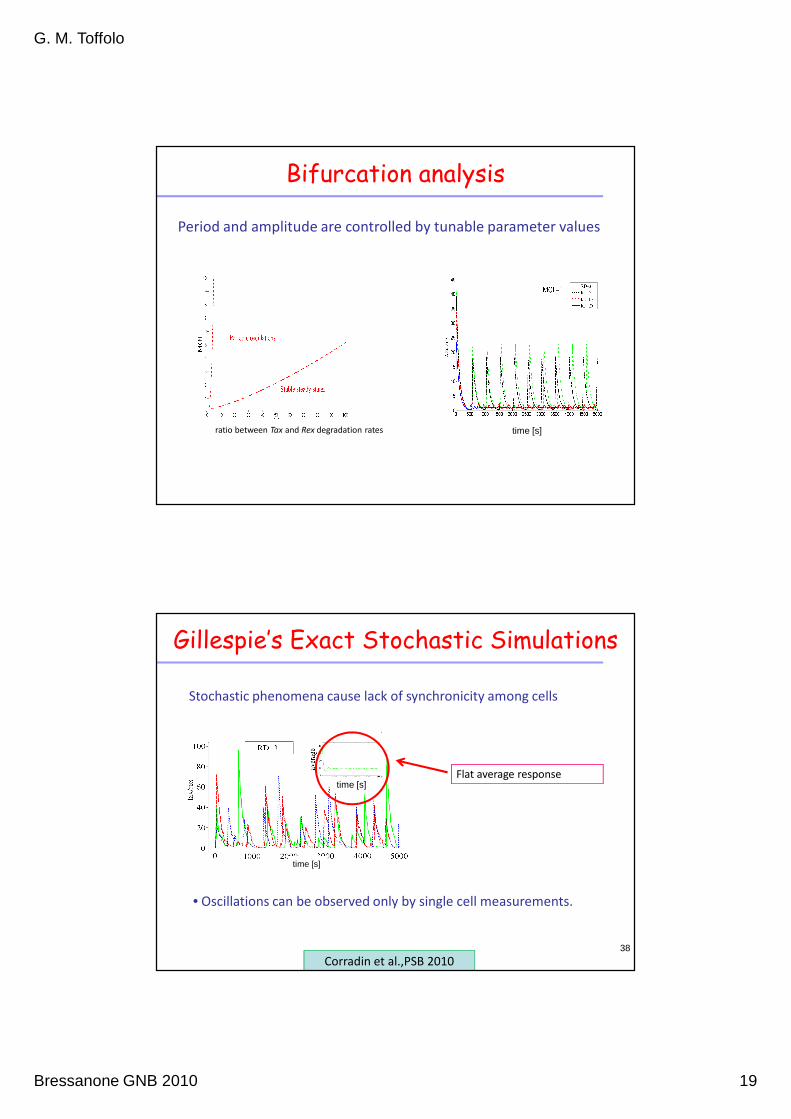
Bifurcation analysis
ratio between Tax and Rex degradation rates
Period and amplitude are controlled by tunable parameter values
time [s]
Gillespie’s Exact Stochastic Simulations
Stochastic phenomena cause lack of synchronicity among cells
time [s]
Corradin et al.,PSB 2010
• Oscillations can be observed only by single cell measurements.
time [s]Flat average response
38
G. M. Toffolo
Bressanone GNB 2010 20
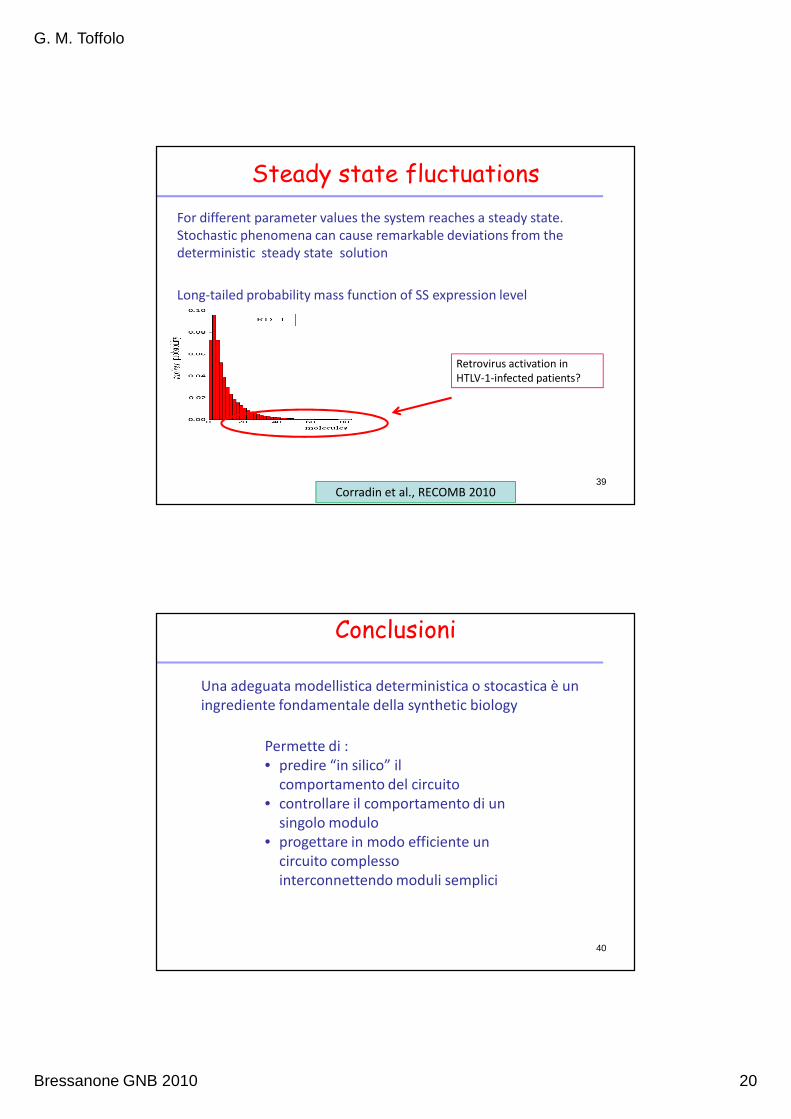
Steady state fluctuations
Long-tailed probability mass function of SS expression level
Retrovirus activation in
HTLV-1-infected patients?
Corradin et al., RECOMB 2010
For different parameter values the system reaches a steady state.
Stochastic phenomena can cause remarkable deviations from the
deterministic steady state solution
39
40
Conclusioni
Permette di :
• predire “in silico” il
comportamento del circuito
• controllare il comportamento di un
singolo modulo
• progettare in modo efficiente un
circuito complesso
interconnettendo moduli semplici
Una adeguata modellistica deterministica o stocastica è un
ingrediente fondamentale della synthetic biology

G. M. Toffolo
Bressanone GNB 2010 21
Conclusioni
41
L’impiego pratico di un modello richiede di :
• formulare le equazioni
• individuare valori numerici per i parametri
Spesso si dispone di valori solo approssimati per i parametri
L’esigenza di una caratterizzazione affidabile di un circuito richiede
• progetto di esperimenti adeguati rispetto alla complessità del
modello
• analisi di identificabilità
• stima dei valori dei parametri/precisione
• validazione del modello
Metodi di formulazione, identificazione e validazione di modelli
trovano nella synthetic biology un campo applicativo di grande
interesse.
Modelli deterministici Vs stocastici
• Reti geniche e biochimiche coinvolgono l’interazione di numeri
interi di molecole che reagiscono in istanti casuali, causando
fluttuazioni casuali nell’espressione delle singole specie chimiche
• Se il numero di molecole è piccolo,le fluttuazioni possono
comportare differenze facilmente identificabili tra cloni
• Inoltre si possono verificare discrepanze significative tra le
predizioni del modello deterministico e il reale comportamento del
sistema.
• Effetti stocastici nell’espressione genica possono avere importanti
implicazioni biologiche42
G. M. Toffolo
Bressanone GNB 2010 22
A simplified model
Data are reproduced qualitatively
and quantitatively
gag
tax/rex
Tax
Rex
Rex-dependent genes
43Parameter values are identified with acceptable precision