Embed Size (px)
Citation preview
Modulo 4
Una proteina in azione

Mioglobina ed emoglobina: un esempio di
funzionamento di una proteina
La funzione della maggior parte della proteine globulari è mediata dall’interazione
reversibile con altre molecole dette ligandi: piccole molecole ma anche altre
macromolecole.
Il sito in cui avviene il contatto con il ligando: sito di legame: è complementare al
ligando stesso per dimensione, forma, carica, e carattere idrofilico o idrofobico.
Rende il legame specifico.
L’ossigeno (O2) ha una bassa solubilità in acqua (sangue = 3ml/L). E’ necessaria
una via per legare l’ossigeno e trasportarlo in sufficiente quantità ai tessuti.
Proteine come la mioglobina e l’emoglobina si sono evolute a questo
fondamentale scopo.
La mioglobina (Mb) e l’emoglobina
(Hb): proteine cruciali nella
conversione dalla vita anaerobica a
quella aerobica.
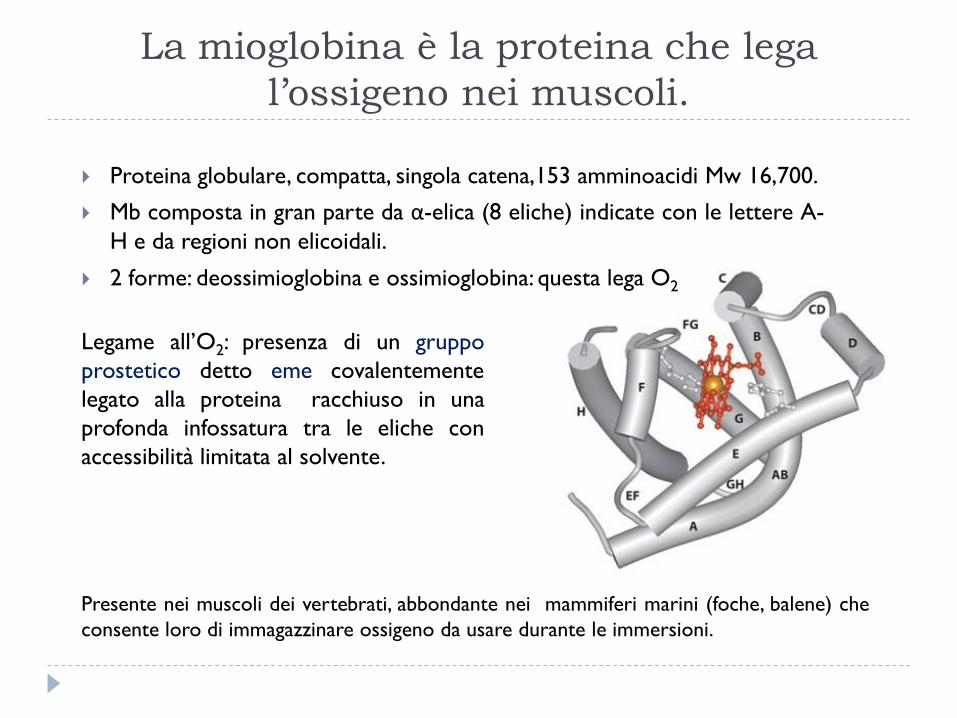
La mioglobina è la proteina che lega
l’ossigeno nei muscoli.
Presente nei muscoli dei vertebrati, abbondante nei mammiferi marini (foche, balene) che
consente loro di immagazzinare ossigeno da usare durante le immersioni.
Proteina globulare, compatta, singola catena,153 amminoacidi Mw 16,700.
Mb composta in gran parte da α-elica (8 eliche) indicate con le lettere A-
H e da regioni non elicoidali.
2 forme: deossimioglobina e ossimioglobina: questa lega O2
Legame all’O2: presenza di un gruppo
prostetico detto eme covalentemente
legato alla proteina racchiuso in una
profonda infossatura tra le eliche con
accessibilità limitata al solvente.
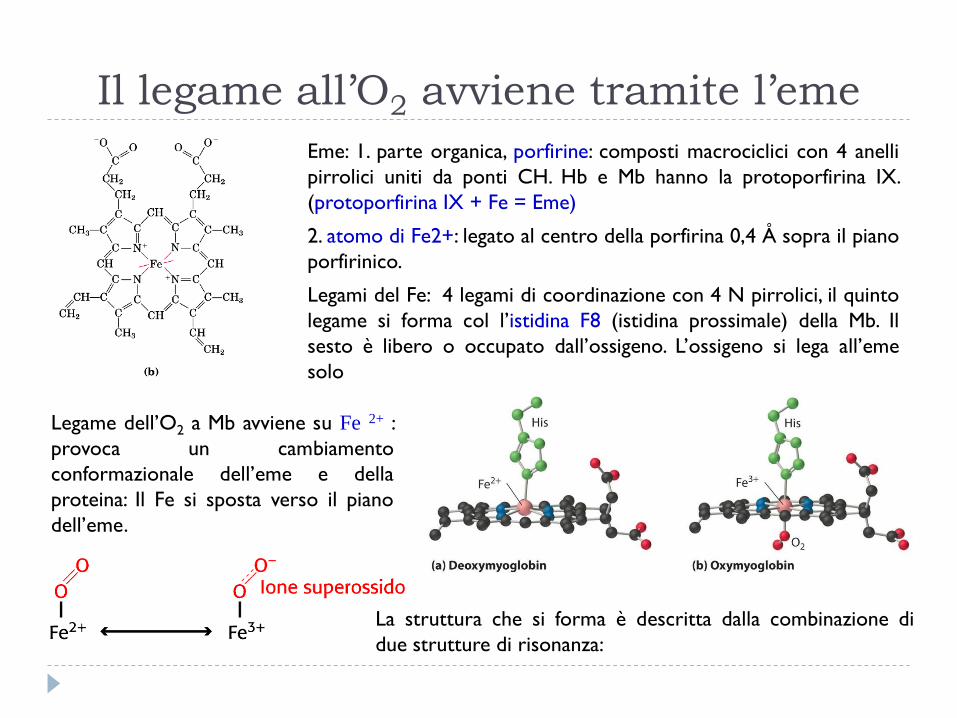
Il legame all’O2 avviene tramite l’eme
Eme: 1. parte organica, porfirine: composti macrociclici con 4 anelli
pirrolici uniti da ponti CH. Hb e Mb hanno la protoporfirina IX.
(protoporfirina IX + Fe = Eme)
2. atomo di Fe2+: legato al centro della porfirina 0,4 Å sopra il piano
porfirinico.
Legami del Fe: 4 legami di coordinazione con 4 N pirrolici, il quinto
legame si forma col l’istidina F8 (istidina prossimale) della Mb. Il
sesto è libero o occupato dall’ossigeno. L’ossigeno si lega all’eme
solo
Legame dell’O2 a Mb avviene su Fe 2+ :
provoca un cambiamento
conformazionale dell’eme e della
proteina: Il Fe si sposta verso il piano
dell’eme.
La struttura che si forma è descritta dalla combinazione di
due strutture di risonanza:

Ragione funzionale della parte globinica
Funzione della parte proteica: Mb stabilizza il complesso con
l’ossigeno in modo da rilasciare O2 ed evitando l’ossidazione
del Fe.
.His E7 (istidina distale) forma un legame H con la forma di
superossido dell’ossigeno. Svolge un ruolo protettivo
fondamentale dall’ossidazione.
La CO si lega al ferro eminico in
competizione con l’O2. Quando l’eme è
contenuto nella mioglobina, l’affinità per
CO diminuisce di 100 volte
Il legame del CO non è ottimale per l’ingombro sterico
dovuto alla HisE7, , il legame dell’O2 che forma un angolo
di 120° con il piano dell’eme, non viene influenzato
sfavorevolmente all’interno della proteina.
l’Ossigeno rilasciato deve essere allo stato O2 e non come anione superossido (tossico), Fe deve rimanere allo stato ferroso (Fe2+) Mb che contiene Fe3+ (metamioglobina) non è più in grado di legare l’ ossigeno.

Curva di saturazione (legame, ossigenazione)
dell’ossigeno alla mioglobina
Il legame reversibile dell’O2 alla mioglobina, può essere descritto dall’equilibrio:
[ ] Y
[ ] [ ] Mb MbO
MbO
2
2
+ =
[ ] [ ] [ ] MbO
O Mb Kd
2
2 = O2 Mb MbO 2 + La Kd di equilibrio (dissociazione) della ossi Mb:
Y = Siti di legame occupati
Totale dei siti di legame
[O2] trattandosi di gas può
essere espresso come pO2
(pressione parziale dell’O) si
ottiene:
Kd pO
pO
2
2
+ = Y Equazione iperbole rettangolare
[ ] [ ] Kd O
O
2
2
+ = Y
Ricavando [MbO2] dall’equazione 1 si ottiene:
(1)
Con 0<Y<1 Definiamo come Y (saturazione frazionale) il rapporto:
P50 = valore di pO2 al quale Y= 0,5 (50% di saturazione).
Curva di legame dell’O2 alla
mioglobina
1 Torr = 1 mmHg
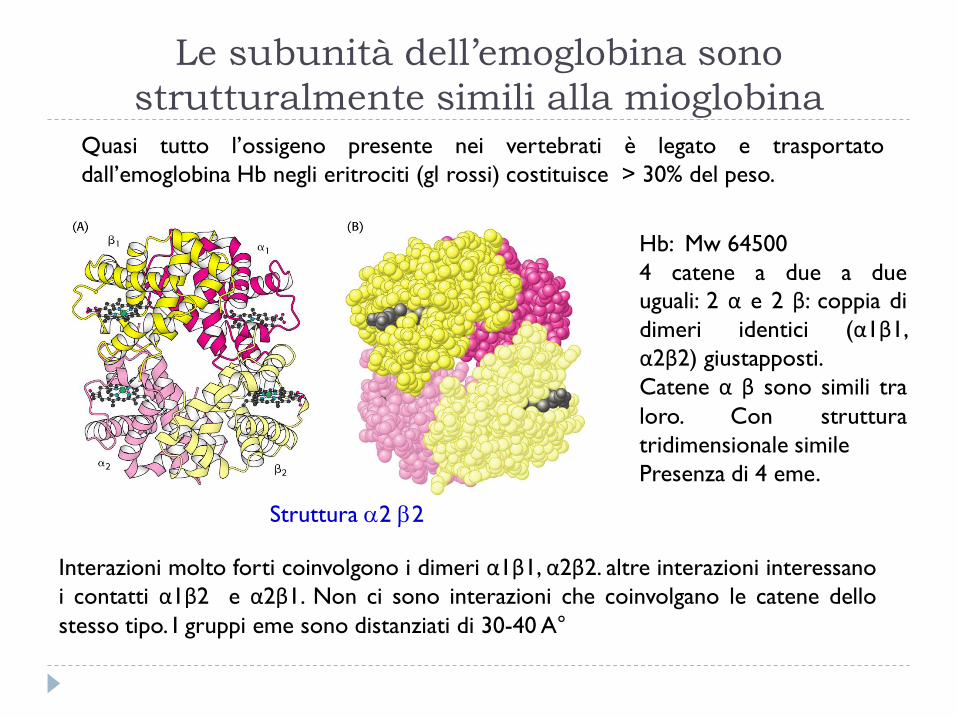
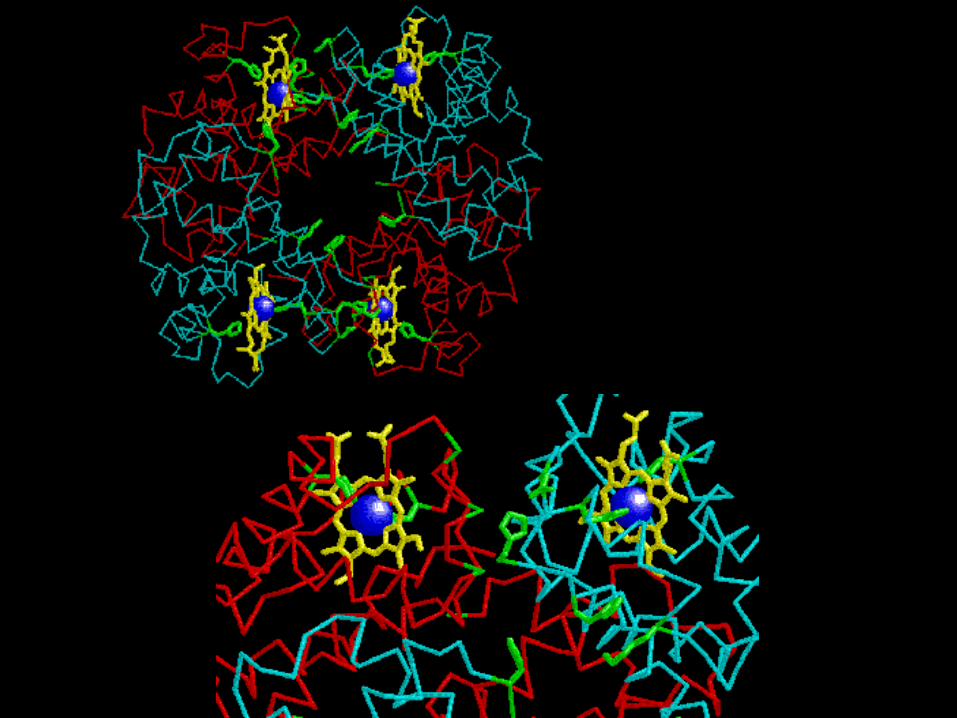
Interazioni molto forti coinvolgono i dimeri α1β1, α2β2. altre interazioni interessano
i contatti α1β2 e α2β1. Non ci sono interazioni che coinvolgano le catene dello
stesso tipo. I gruppi eme sono distanziati di 30-40 A°
Struttura 2 2
Le subunità dell’emoglobina sono
strutturalmente simili alla mioglobina Quasi tutto l’ossigeno presente nei vertebrati è legato e trasportato
dall’emoglobina Hb negli eritrociti (gl rossi) costituisce > 30% del peso.
Hb: Mw 64500
4 catene a due a due
uguali: 2 α e 2 β: coppia di
dimeri identici (α1β1,
α2β2) giustapposti.
Catene α β sono simili tra
loro. Con struttura
tridimensionale simile
Presenza di 4 eme.
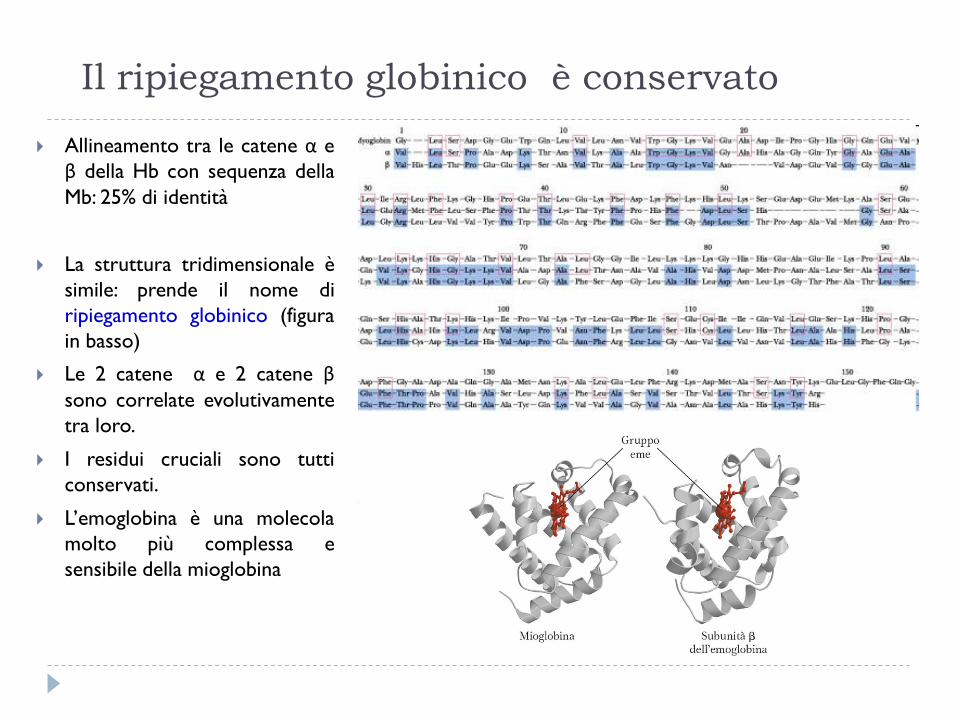
Il ripiegamento globinico è conservato
Allineamento tra le catene α e
β della Hb con sequenza della
Mb: 25% di identità
La struttura tridimensionale è
simile: prende il nome di
ripiegamento globinico (figura
in basso)
Le 2 catene α e 2 catene β
sono correlate evolutivamente
tra loro.
I residui cruciali sono tutti
conservati.
L’emoglobina è una molecola
molto più complessa e
sensibile della mioglobina
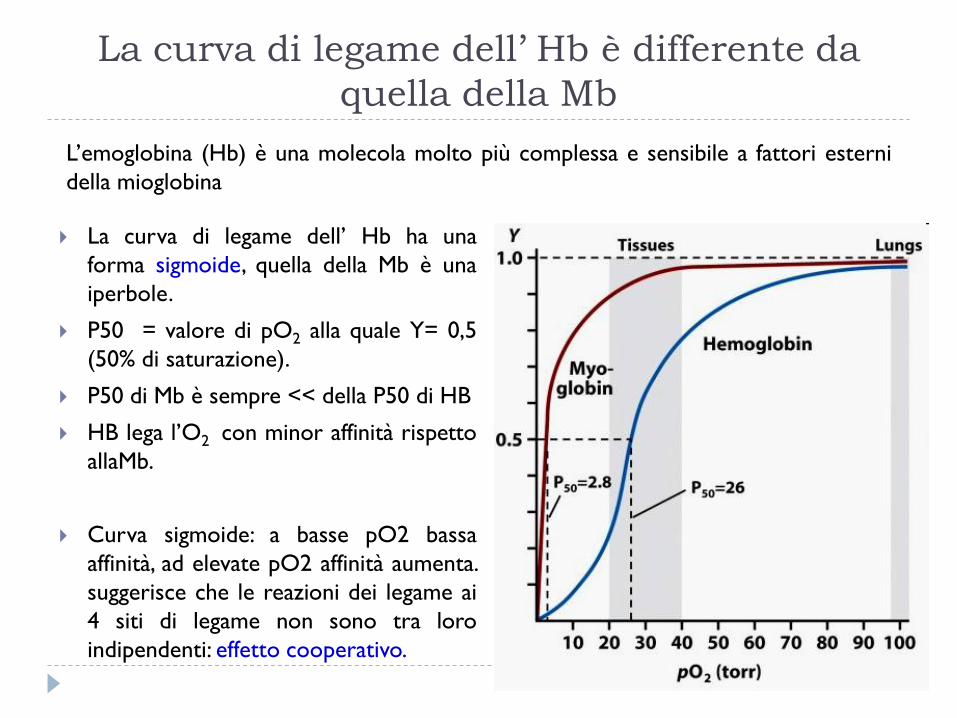
La curva di legame dell’ Hb è differente da
quella della Mb
La curva di legame dell’ Hb ha una
forma sigmoide, quella della Mb è una
iperbole.
P50 = valore di pO2 alla quale Y= 0,5
(50% di saturazione).
P50 di Mb è sempre << della P50 di HB
HB lega l’O2 con minor affinità rispetto
allaMb.
Curva sigmoide: a basse pO2 bassa
affinità, ad elevate pO2 affinità aumenta.
suggerisce che le reazioni dei legame ai
4 siti di legame non sono tra loro
indipendenti: effetto cooperativo.
L’emoglobina (Hb) è una molecola molto più complessa e sensibile a fattori esterni
della mioglobina
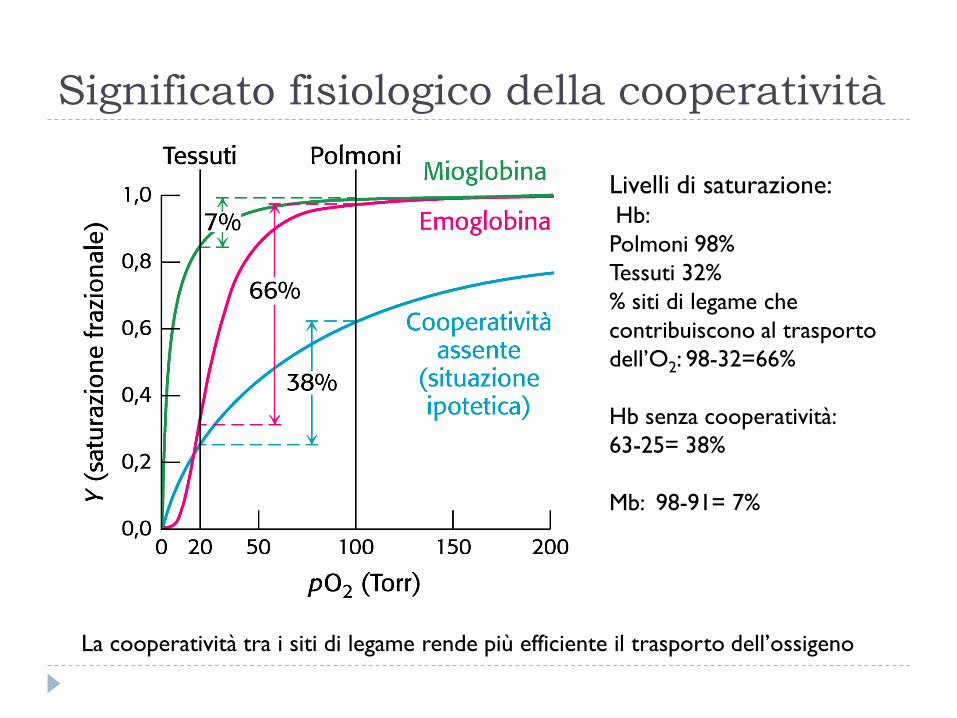
Significato fisiologico della cooperatività
La cooperatività tra i siti di legame rende più efficiente il trasporto dell’ossigeno
Livelli di saturazione: Hb:
Polmoni 98%
Tessuti 32%
% siti di legame che
contribuiscono al trasporto
dell’O2: 98-32=66%
Hb senza cooperatività:
63-25= 38%
Mb: 98-91= 7%
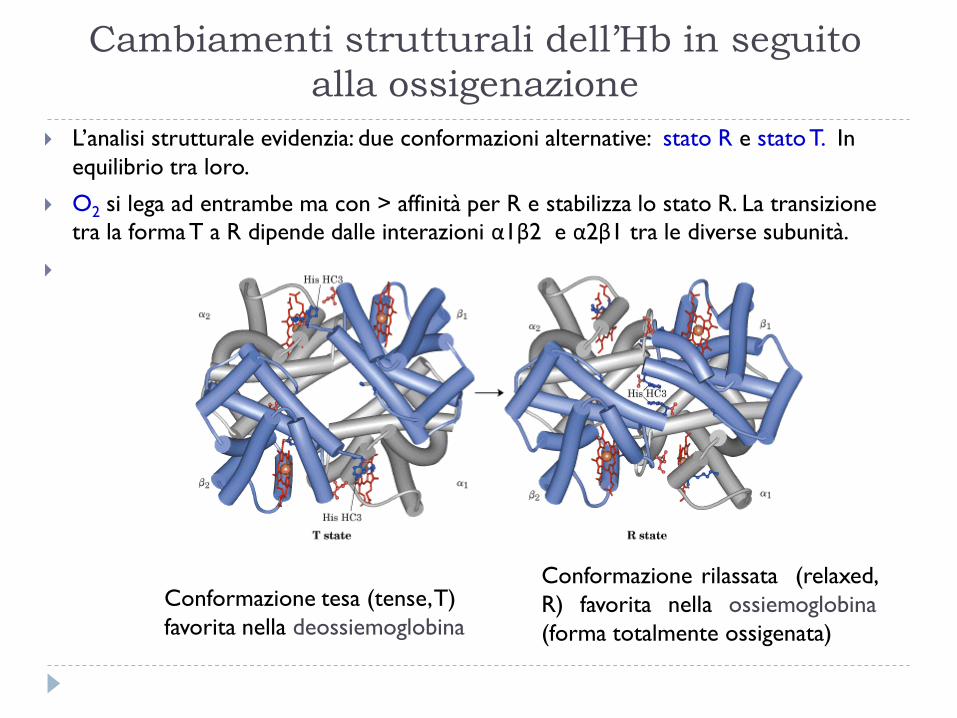
Conformazione tesa (tense, T)
favorita nella deossiemoglobina
Conformazione rilassata (relaxed,
R) favorita nella ossiemoglobina
(forma totalmente ossigenata)
Cambiamenti strutturali dell’Hb in seguito
alla ossigenazione
L’analisi strutturale evidenzia: due conformazioni alternative: stato R e stato T. In
equilibrio tra loro.
O2 si lega ad entrambe ma con > affinità per R e stabilizza lo stato R. La transizione
tra la forma T a R dipende dalle interazioni α1β2 e α2β1 tra le diverse subunità.
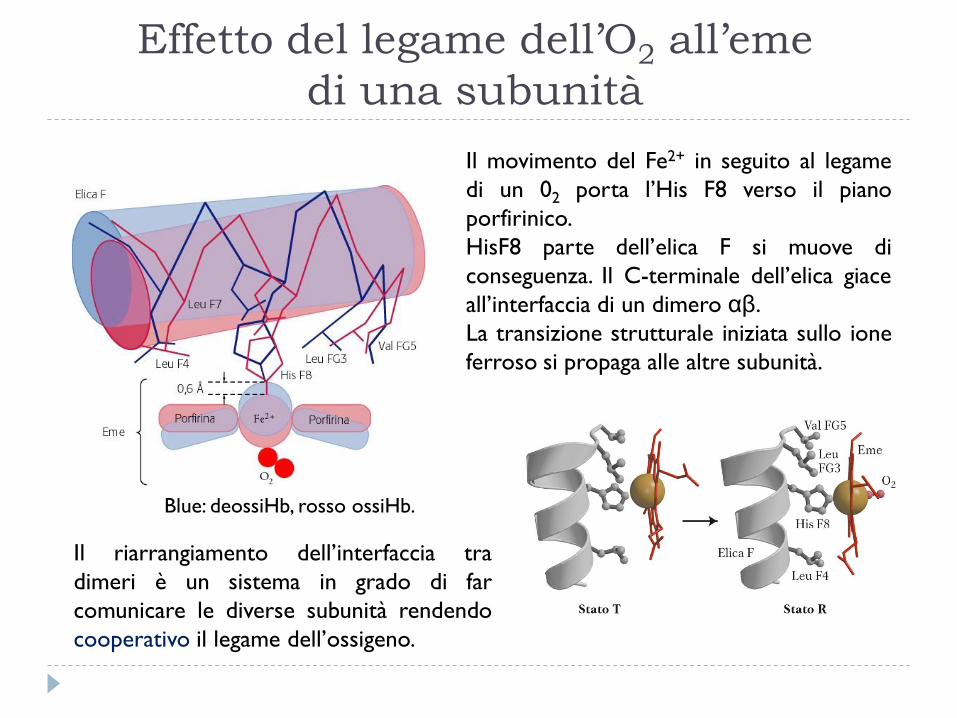
Effetto del legame dell’O2 all’eme
di una subunità
Il riarrangiamento dell’interfaccia tra
dimeri è un sistema in grado di far
comunicare le diverse subunità rendendo
cooperativo il legame dell’ossigeno.
Il movimento del Fe2+ in seguito al legame
di un 02 porta l’His F8 verso il piano
porfirinico.
HisF8 parte dell’elica F si muove di
conseguenza. Il C-terminale dell’elica giace
all’interfaccia di un dimero αβ.
La transizione strutturale iniziata sullo ione
ferroso si propaga alle altre subunità.
Blue: deossiHb, rosso ossiHb.
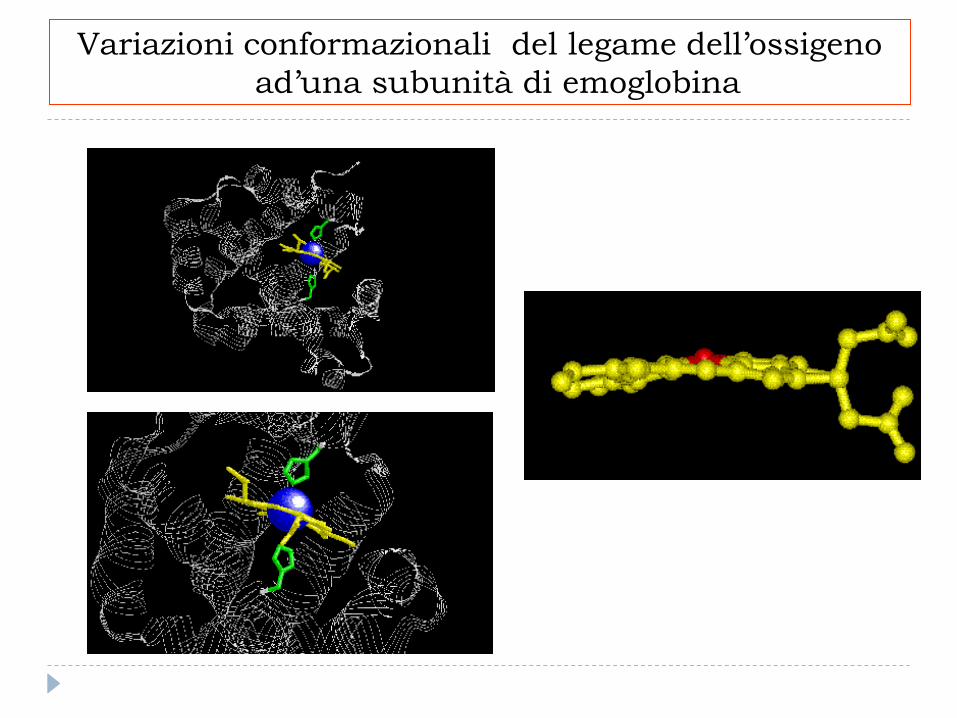
Variazioni conformazionali del legame dell’ossigeno
ad’una subunità di emoglobina
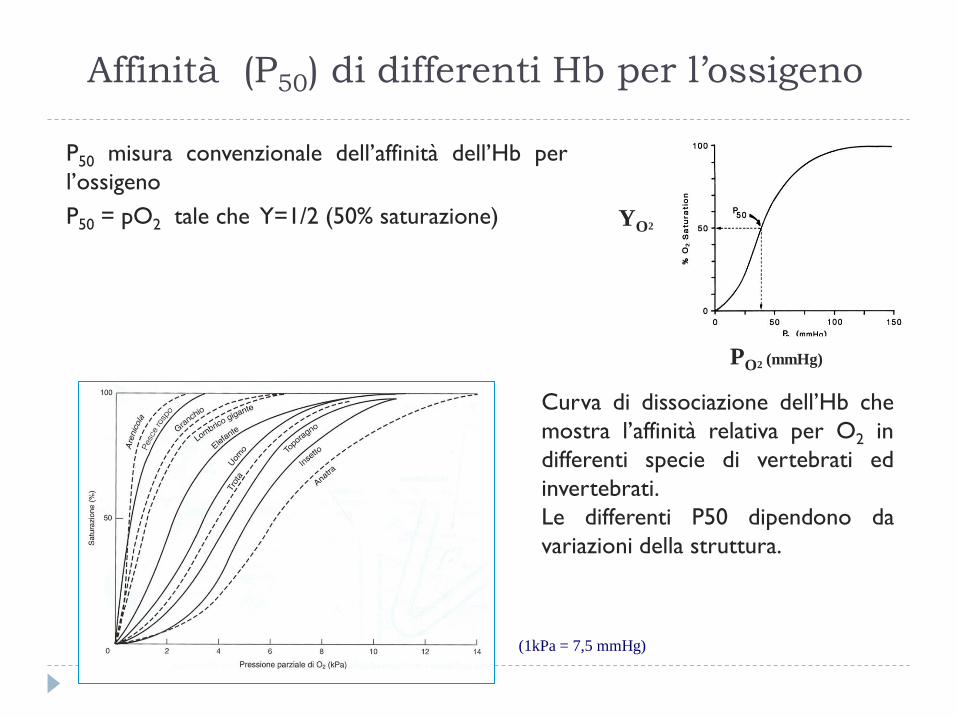
Affinità (P50) di differenti Hb per l’ossigeno
P50 misura convenzionale dell’affinità dell’Hb per
l’ossigeno
P50 = pO2 tale che Y=1/2 (50% saturazione)
YO2
PO2 (mmHg)
Curva di dissociazione dell’Hb che
mostra l’affinità relativa per O2 in
differenti specie di vertebrati ed
invertebrati.
Le differenti P50 dipendono da
variazioni della struttura.
(1kPa = 7,5 mmHg)
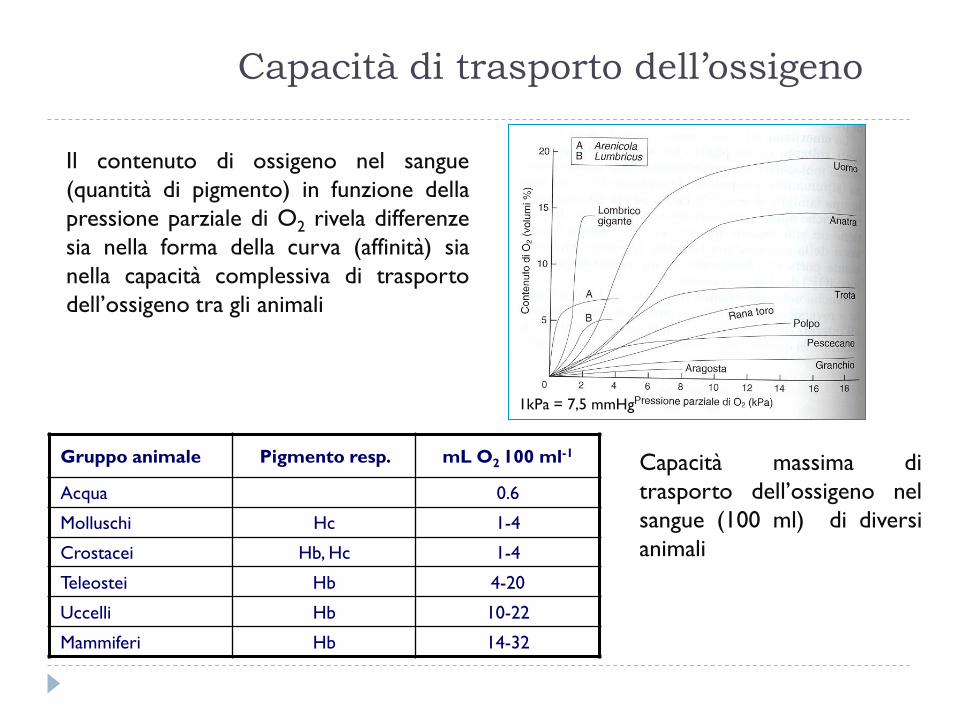
Capacità di trasporto dell’ossigeno
1kPa = 7,5 mmHg
Gruppo animale Pigmento resp. mL O2 100 ml-1
Acqua 0.6
Molluschi Hc 1-4
Crostacei Hb, Hc 1-4
Teleostei Hb 4-20
Uccelli Hb 10-22
Mammiferi Hb 14-32
Il contenuto di ossigeno nel sangue
(quantità di pigmento) in funzione della
pressione parziale di O2 rivela differenze
sia nella forma della curva (affinità) sia
nella capacità complessiva di trasporto
dell’ossigeno tra gli animali
Capacità massima di
trasporto dell’ossigeno nel
sangue (100 ml) di diversi
animali
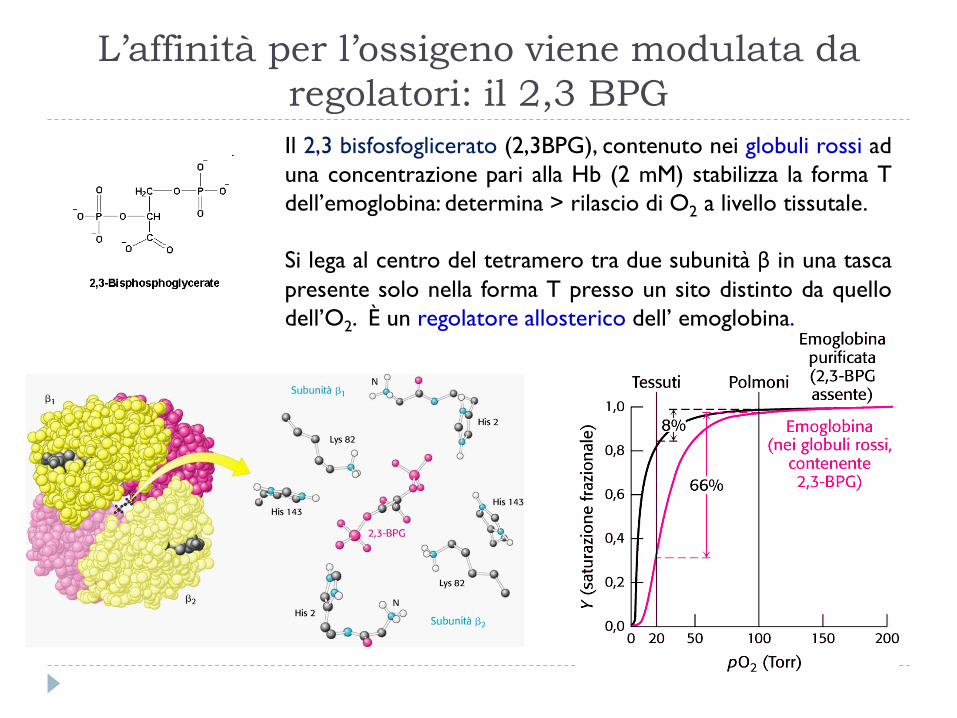
L’affinità per l’ossigeno viene modulata da
regolatori: il 2,3 BPG
Il 2,3 bisfosfoglicerato (2,3BPG), contenuto nei globuli rossi ad
una concentrazione pari alla Hb (2 mM) stabilizza la forma T
dell’emoglobina: determina > rilascio di O2 a livello tissutale.
Si lega al centro del tetramero tra due subunità β in una tasca
presente solo nella forma T presso un sito distinto da quello
dell’O2. È un regolatore allosterico dell’ emoglobina.
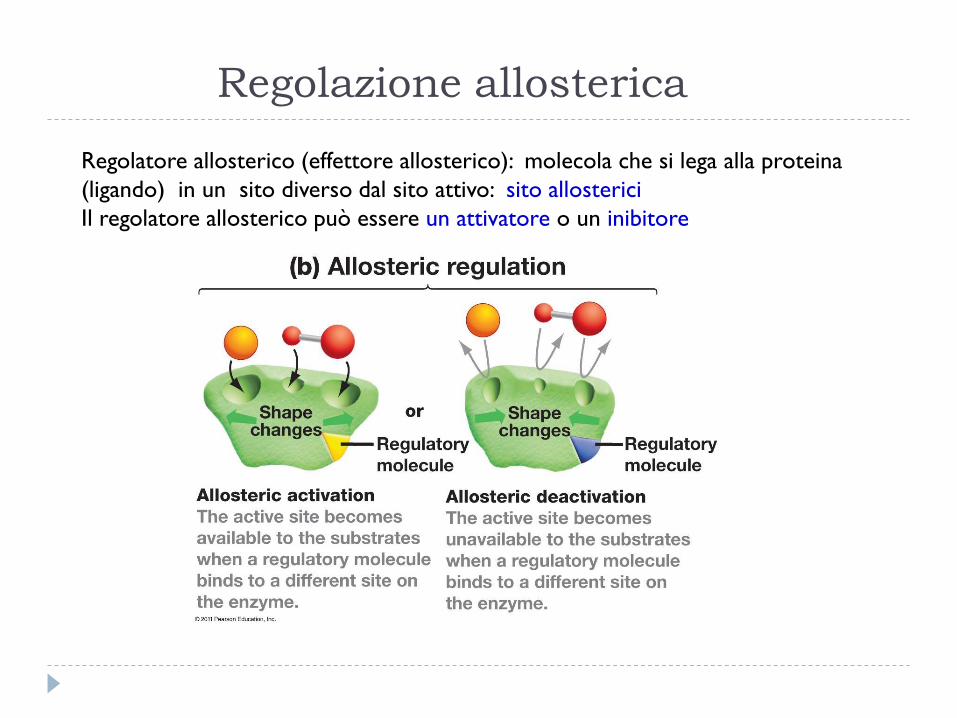
Regolazione allosterica
Regolatore allosterico (effettore allosterico): molecola che si lega alla proteina
(ligando) in un sito diverso dal sito attivo: sito allosterici
Il regolatore allosterico può essere un attivatore o un inibitore
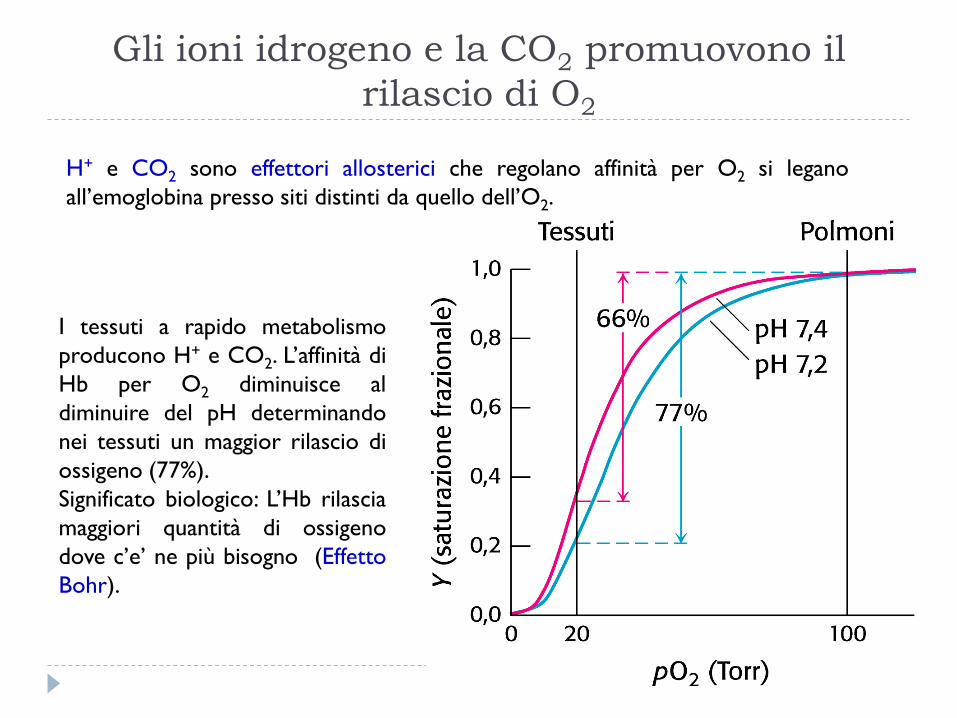
Gli ioni idrogeno e la CO2 promuovono il
rilascio di O2
H+ e CO2 sono effettori allosterici che regolano affinità per O2 si legano
all’emoglobina presso siti distinti da quello dell’O2.
I tessuti a rapido metabolismo
producono H+ e CO2. L’affinità di
Hb per O2 diminuisce al
diminuire del pH determinando
nei tessuti un maggior rilascio di
ossigeno (77%).
Significato biologico: L’Hb rilascia
maggiori quantità di ossigeno
dove c’e’ ne più bisogno (Effetto
Bohr).
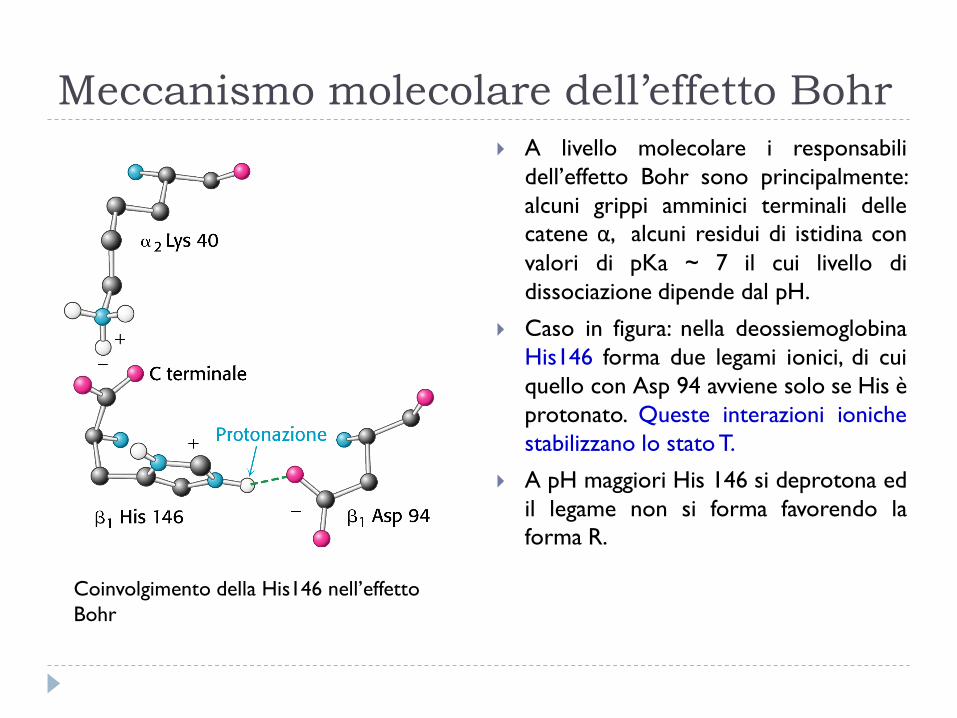
Meccanismo molecolare dell’effetto Bohr
A livello molecolare i responsabili
dell’effetto Bohr sono principalmente:
alcuni grippi amminici terminali delle
catene α, alcuni residui di istidina con
valori di pKa ~ 7 il cui livello di
dissociazione dipende dal pH.
Caso in figura: nella deossiemoglobina
His146 forma due legami ionici, di cui
quello con Asp 94 avviene solo se His è
protonato. Queste interazioni ioniche
stabilizzano lo stato T.
A pH maggiori His 146 si deprotona ed
il legame non si forma favorendo la
forma R.
Coinvolgimento della His146 nell’effetto
Bohr
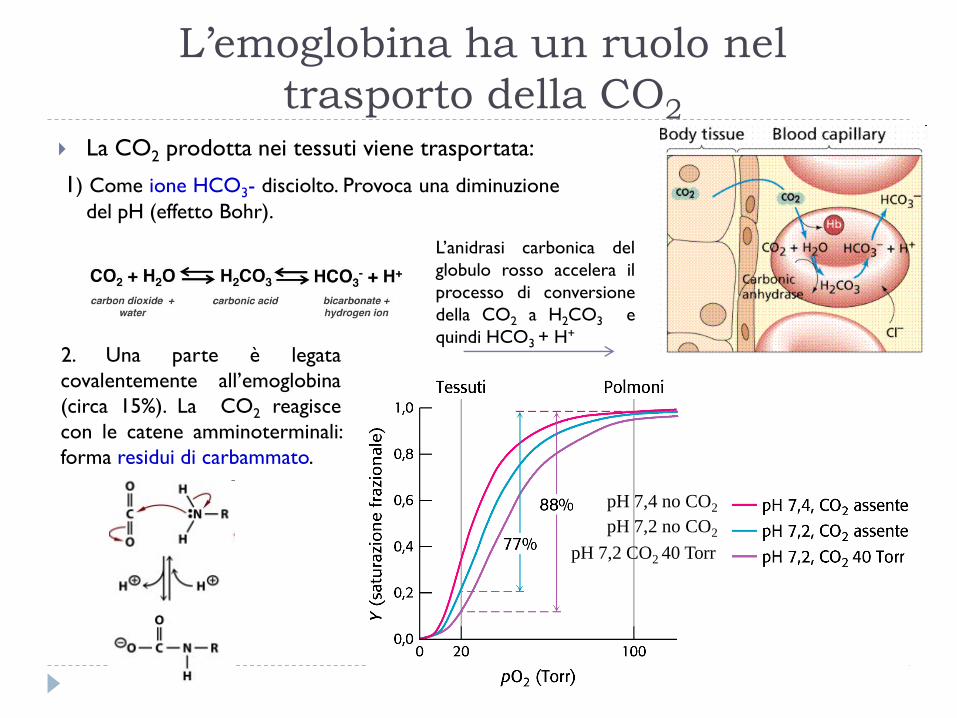
L’emoglobina ha un ruolo nel
trasporto della CO2
La CO2 prodotta nei tessuti viene trasportata:
1) Come ione HCO3- disciolto. Provoca una diminuzione
del pH (effetto Bohr).
2. Una parte è legata
covalentemente all’emoglobina
(circa 15%). La CO2 reagisce
con le catene amminoterminali:
forma residui di carbammato.
pH 7,4 no CO2
pH 7,2 no CO2
pH 7,2 CO2 40 Torr
L’anidrasi carbonica del
globulo rosso accelera il
processo di conversione
della CO2 a H2CO3 e
quindi HCO3 + H+
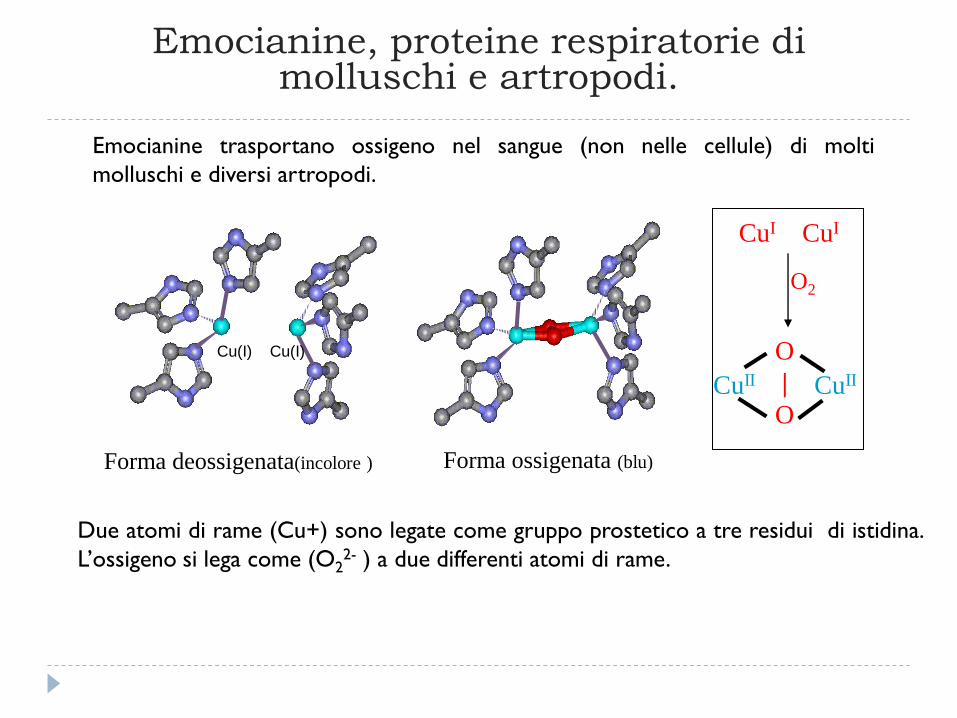
Emocianine, proteine respiratorie di molluschi e artropodi.
Due atomi di rame (Cu+) sono legate come gruppo prostetico a tre residui di istidina.
L’ossigeno si lega come (O22- ) a due differenti atomi di rame.
Forma deossigenata(incolore ) Forma ossigenata (blu)
Cu(I) Cu(I)
CuII CuII
O
|
O
CuI CuI
O2
Emocianine trasportano ossigeno nel sangue (non nelle cellule) di molti
molluschi e diversi artropodi.
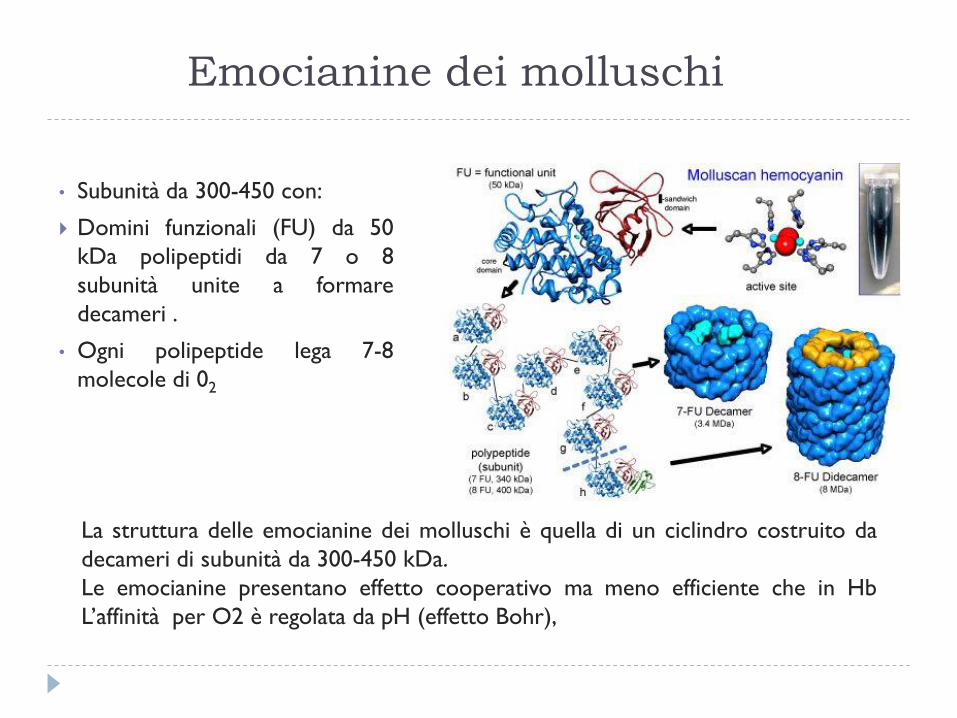
Emocianine dei molluschi
• Subunità da 300-450 con:
Domini funzionali (FU) da 50
kDa polipeptidi da 7 o 8
subunità unite a formare
decameri .
• Ogni polipeptide lega 7-8
molecole di 02
La struttura delle emocianine dei molluschi è quella di un ciclindro costruito da
decameri di subunità da 300-450 kDa.
Le emocianine presentano effetto cooperativo ma meno efficiente che in Hb
L’affinità per O2 è regolata da pH (effetto Bohr),

Concetti principali
Le proteine sono strutture dinamiche soggette a cambiamenti
conformazionali piu o meno accentuati.
La variazioni di pochi residui può avere effetti importanti sulla
funzionalità o sulla modulazione dell’attività della proteina.
Il legame di molecole regolatrici (effettori allosterici) ha un
profondo effetto sull’attività della proteina.
Funzioni simili nei diversi organismi possono essere compiute da
proteine diverse, non evolutivamente correlate (evoluzione
convergente) che possiedono proprietà simili.





























