Embed Size (px)
Citation preview

Fachhochschule Wiesbaden Standort Geisenheim
Fachbereich Geisenheim
Studiengang Gartenbau-Management
Bachelorarbeit
Musa in Shaded Perennial Crops - Response to
Light Interception
Referent: Prof. Dr. Joachim Heller
Korreferent: Prof. Dr. Jürgen Jaki
Vorgelegt von:
Christian Dold

Geisenheim, den 24.01.2007

Eidesstattliche Erklärung:
Ich erkläre hiermit wahrheitsgemäß, daß ich
- die eingereichte Arbeit selbständig und ohne unerlaubte Hilfsmittel angefertigt habe,
- nur die im Literaturverzeichnis aufgeführten Hilfsmittel benutzt und fremdes
Gedankengut als solches kenntlich gemacht habe,
- alle Personen und Institutionen, die mich bei der Vorbereitung und Anfertigung der
Abhandlung unterstützt haben, genannt habe und
- die Arbeit noch keiner anderen Stelle zur Prüfung vorgelegt habe
Ort, Datum
Unterschrift (Vor- und Zuname)

Acknowledgement
I would like to thank Dr. Charles Staver, Bioversity International, and Dr. Luis Pocasangre,
Centro Agronómico de Investigación y Enseñanza (CATIE) and Bioversity International, for
their supervision, for the opportunity of an internship at CATIE, and for their support.
Furthermore, I would like to thank the BEAF-Group of Deutsche Gesellschaft für Technische
Zusammenarbeit, namely Dr. Marlene Diekmann and Dr. Kasten, for the financial and
administrative support; without it would have not have been possible to do my internship.
Also, I would like to thank Inwent gGmbH, namely Iris Lenzen, for their financial and
administrative support.
Besides, it is a great pleasure to thank Prof. Dr. Joachim Heller for his supervision, not only
during my internship and my bachelor thesis, but also during my studies at the University of
Geisenheim. Furthermore, Prof. Dr. Jürgen Jaki, for his supervision and support during my
bachelor thesis, as well as Prof. Dr. Kai Velten; without him there would be not much of
statistical data.
At last, I would like to thank Pablo Siles – Are you already a PhD?

Seite I
Contents
1 Introduction .................................................................................................. 1
1.1 Formulation of the Problem ............................................................... 1
1.2 Aim of the Work................................................................................. 1
1.3 Structure of the Work......................................................................... 1
2 Light Interception in Tropical Agroforestry Systems..................................... 2
3 Musa in Agroforestry Systems ................................................................... 11
3.1 Importance of Musa on Farm Level in Central America................... 11
3.2 Research Review on Musa in Shade............................................... 13
4 Musa in Shaded Coffee - A Case Study of Costa Rica .............................. 21
4.1 Description ...................................................................................... 21
4.1.1 Botanic garden – Musa ‘Gros Michel’ in Sun and Shade.. 21
4.1.2 Six Varieties in Four Different Light Levels ....................... 23
4.1.3 Light measurements in Agroforestry Systems .................. 24
4.1.4 Interviews ......................................................................... 24
4.2 Material and Methods ...................................................................... 25
4.2.1 Light Measurements ......................................................... 26
4.2.2 Number of Functional Leaves........................................... 28
4.2.3 Length of the Petiole......................................................... 28
4.2.4 Non-destructive Measurements of Leaf Area ................... 29
4.2.5 Destructive Measurements of Leaf Area........................... 29
4.2.6 Leaf Emission Rate (LER) ................................................ 32
4.2.7 Circumference and Height ................................................ 32
4.2.8 Black Sigatoka Leaf Spot.................................................. 33

Seite II
4.2.9 Leaf Tearing ..................................................................... 33
4.2.10 Leaf Angle ........................................................................ 34
4.2.11 Leaf Folding...................................................................... 35
4.3 Results ............................................................................................ 36
4.3.1 Light Measurement in the Botanic Garden ....................... 36
4.3.2 Results Pseudostems III ................................................... 38
4.3.3 Results Pseudostem II...................................................... 44
4.3.4 Results Six Varieties......................................................... 48
4.3.5 Light Measurements in Coffee Agroforestry Systems....... 52
4.3.6 Interviews ......................................................................... 60
5 Discussion.................................................................................................. 60
6 Conclusion ................................................................................................. 67
7 References................................................................................................. 69

Seite III
List of Figures
Fig. 1: Solar radiation at different latitudes with respect to seasonal variation
Fig. 2: Instanenous Photon Flux Density (PFD) at two points of the same gap, Costa Rica
Fig. 3: The exponential attenuation of solar radiation in a plant stand
Fig. 4: Using Beer’s law requires a random structure of the canopy
Fig. 5: Amount of solar energy in four Central American banana production areas
Fig. 6: Comparison of the results of MURRAY (1961), and ISRAELI et al. (1995)
Fig. 7: Summary of different research about cultivar light response curves
Fig. 8: Banana ‘Gros Michel’ mainly asociatied with cacao at CATIE
Fig. 9: The shaded banana suckers in the shade of coffee, Poró and Cedro
Fig. 10: Example for counting the total number of leaves
Fig. 12: Measurement of the angle between leaf sheath and leaf angle
Fig. 13: Measuring the leaf blade angle using a template
Fig. 14: Radiation environment in week 18 in the two treatments above the Pseudostem III
Fig. 15: Radiation environment in week 18 in the two treatments above the Pseudostem II
Fig. 16: Weekly number of leaves of Pseudostems III
Fig. 17: Weekly measurements of Leaf Emission Rate (LER) of Pseudostems III
Fig. 18: Two-week measurements of height (cm) of Pseudostems III averaged data
Fig. 19: Measurements of circumference (cm) of Pseudosems III
Fig. 20: Weekly leaf area of pseudostems II from week 13 to week 27
Fig. 21: Weekly measurements of Leaf Emission Rate (LER) of Pseudostems II
Fig. 22: Measurements of height (cm) from week 13 to week 34 of Pseudostems II
Fig. 23: Total transmitted light (%) in the most shaded plot; above coffee and above banana

Seite IV
Fig. 24: Raditation regime of the four plots above and in height of the banana plants
Fig. 25: Average growth of height of six banana varieties in different light regimes
Fig. 26: Average growth of leaf area of six banana varieties in different light regimes
Fig. 27: Average total transmitted light of the all coffee agroforestry systems
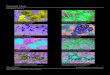
Fig. 28: Light dispersal map of Coffee – Cashá agroforestrial system
Fig. 29: Light dispersal map of Coffee – Roble Coral agroforestry system
Fig. 30: Light dispersal map of Coffee – Poró agroforestry system
Fig. 31: Light dispersal map of Coffee – Roble Coral - Cashá agroforestry system
Fig. 32: Light dispersal map of Coffee – Poró - Cashá agroforestry system
Fig. 33: Light dispersal map of Coffee – Poró – Roble Coral agroforestry system
Fig. 34: Musa AAA Cavendish subgroup; light response curve
Fig. 35: Stations of leaf development
Fig. 36: Gauhl’s modification of Stover’s severity scoring system

Seite V
List of Tables
Tab. 1: Summary of different research about impacts on photosynthesis rate
Tab. 2: Method of growth measurement and frequency
Tab. 3: Impact of Black Sigatoka on Pseudostems III in two different light conditions
Tab. 4: Leaf Tearing of twenty plants comparable in height of Pseudostems III in week 18
Tab. 5: Estimated average leaf area of Pseudostems III from week 18 to week 35
Tab. 6: Impact of Black Sigatoka on Pseudostems II in two different light
Tab. 7: Leaf area of week 26 and estimated total increase of leaf area
Tab. 8: Folding of the leaf blades; measurements at the 2nd leaf of Pseudostems
Tab. 9: Leaf Angle of Pseudostem II; week 20; Leaf 1 to 5
Tab.10: Growth of Height, Circumference, Length of Petiole and Leaf Area of six varieties
Tab. 11: Explanation of the five stages of leaf development
Tab. 12: Soil samples of the botanic garden, ten of each; 0 – 30 cm
Tab. 13: Soil samples of the site with the six varieties; 0 – 10 cm
Tab. 14: Soil samples of the site with the six varieties; 0 – 30 cm
Tab. 15: Amount of dry matter of organic fertilizer in the plots with the six varieties

Seite VI
List of Abbreviations
AN Leaf area of the youngest leaf (equation 10)
A i Leaf area of the oldest green leaf (in equation 10)
A i, N integration of the leaf areas between leaf i and leaf N (in equation 10)
AR Rate of increasing leaf area
CATIE Tropical Agriculture Research and Higher Education Centre
dm Percentage of dry matter in the fruit tissue
DDT Disease development time
GLA Gap Light Analyzer (software)
GPS Global Positioning System
H Radiation on any point on earth at the ground per time unit
H0 Extraterrestical radiation per time unit
Hb Direct solar radiation
Hi Harvest index
i oldest leaf in equation 10
I Incident radiation below the canopy (in PFD)
Io Radiation above the canopy (in PFD)
Ia PFD absorbed by the canopy
Ir PFD reflected by the canopy
Irs PFD reflected by the soil
It PFD transmitted through the canopy
k Extinction coefficient
Kt Cloudiness Index

Seite VII
L Leaf Area Index
LER Leaf Emission Rate
N youngest leaf in equation 10 (its number represents total number of leaves)
P first crop cycle of banana
PAR Photosynthetic Active Radiation
PFD Photosynthetically active photon flux density (µmol photons m-2s-1)
Rp visible waveband between 400nm and 700 nm
Rs total shortwave (broadband) radiation (all wavelengths 0.25 µm to 25 µm)
R1 first ratoon cycle of banana
Tc Temperature coefficient
Y Yield of banana
YLS Youngest leaf spotted
γ Radiation use efficiency

Christian Dold Introduction
Seite 1
1 Introduction
1.1 Formulation of the Problem
Throughout the tropics banana and plantain (Musa spp.) is a very common crop in
agroforestry systems, characteristically cultivated by small scale farmers. The production is
extensive with low financial imputs and labour. Typically, it is associated with perennial
crops like coffee (Coffea sp.) and cacao (Theobroma sp.) and an upper story of timber trees.
Thus, Musa spp. is cultivated in highly shaded conditions. Light interception is one significant
limiting factor (NORGROVE 1998, STOVER and SIMMONDS 1987 204-205, 228-231)
While research is mainly focused on all other production units, the improvement of shaded
banana and plantain yield is widly neglected. The few research which are available are
discriminated; banana yield was increased as well as decreased due to shade. Therefore, the
question stays remain if banana production in agroforestry systems could be improved in
relation to light interception (TURNER 1998).
1.2 Aim of the Work
Aim of this work is to ask and find possible answers on the widely unknown topic how the
production of shaded Musa spp. in agroforestry systems can be improved.
1.3 Structure of the Work
At first, some facts about light interception are necessary to understand light as a main
limiting factor in agroforestry systems. Then, a brief description of the importance of banana
and plantain in agroforestry systems will be shown. Then, the state of art concerning the
research on banana in shade will help to understand the theoretical backround. Besides, a
recent study on banana in shade in the tropical region of Costa Rica can give some results on
the topic. A general discussion will concern with the advantages and disadvantages of banana
production in agroforestry systems, and can give an outlook on further research. In the end,
the conclusion will summarize the main ideas of this work.

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 2
2 Light Interception in Tropical Agroforestry Systems
The tropics are defined as the belt around the earth between the Tropic of Cancer (23° 30’
latitude N) and the Tropic of Capricorn (23° 30’ latitude S). As the axis of the earth being
inclined by around 23° 30’, at some time of the year sun rays are perpendincular in the
tropics. Also variation of day length is very little. At the equator day length is always about
12 hours. The difference between the longest and the shortest day is increasing by about 7
minutes per degree in the tropics (NAKASONE and PAULL 1998 3 – 6).
The energy that reaches earth on a perpendicular surface at the mean distance from the sun is
called solar constant. The solar constant is 1.37 kJ m-² s-1.
The amount of solar radiation which reaches earth is depending on latitude and season (see
Fig. 1). In the tropical belt there is a smaller variation of annual solar radiation compared to
higher latitudes. The ratio between the solar energy flux on any latitude on earth to the solar
constant is called geometrical ratio.
Fig. 1: Solar radiation at different latitudes with respect to seasonal variation
Reference: AZAM-ALI and SQUIRE (2001)

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 3
In addition, the amount of solar radiation reaching earth is depending on gases, clouds and
aerosols in the atmosphere which are capable of absorbing and scattering radiation
(NAKASONE and PAULL 1998 3 – 6, AZAM-ALI and SQUIRE 2001 29). Overcast conditions
ocurre especially in the humid tropics where in average only three to five sunshine hours per
day are available (ROBINSON 1996 89).
The cloudiness index gives the severity of scattering and absorbing components in the
atmosphere and can be computed (FRAZER et al. 1999):
Kt = H/H0 (1)
Where Kt is the cloudiness index, H is the incident global radiation on any point on earth at
the ground per time unit, and H0 is the extraterrestical radiation per time unit
The radiation going through the atmosphere can be divided in a direct beam fraction and a
diffuse fraction. “Direct (beam) radiation is the energy that streams from the solar disk and is
neither absorbed nor scattered by the earth’s atmosphere. Diffuse radiation is the portion that
is scattered towards the earth’s surface from all regions of the sky.” (FRAZER et al. 1999).
The more clouds, gases and aerosols in the atmosphere, the higher is the amount of diffuse
raditation. Hence, beam fraction is depending on the cloudiness index:
Hb/H = [1-exp(-3.044Kt2.436)] (2)
Where Hb is the direct radiation, H is the incident global radiation per time unit, Hb/H is the
proportion of direct solar radiation and Kt is the cloudiness Index (FRAZER et al. 1999).
After considering latitude as well as direct beam and diffuse radtiation, the amount of
Photosynthetic Active Radiation (PAR; 0.4 to 0.7 µm waveband) reaching earth is of
importance. It is the radiation which is necessary for photosynthesis and thus, is responsible

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 4
for plant growth. The proportion of PAR to the total incoming shortwave radiation is called
the spectral fraction. In general, in the tropics about 50% of incoming radiation is considered
to be PAR. But with increasing cloud cover (Kt < 0.5) the amount of PAR increases. Thus,
the flux of PAR is also depending on the cloudiness index (R2=0.73) and can be computed
(FRAZER et al. 1999, AZAM-ALI and SQUIRE 2001 29-30).
Rp/Rs=1-exp(-0.499Kt-0.219) (3)
Where Rs is the total shortwave (broadband) radiation contributed by all wavelengths (0.25
µm to 25 µm), Rp is the visible waveband between 400 and 700 nm, and Rp/Rs is the
proportion of PAR on the total global shortwave radiation, and Kt is the cloudiness Index
(Frazer et al. 1999).
On a cloudless day at noon in the tropics the energy input is about 7 – 12 MJ m-2d-1. In the
most tropical regions an average annual solar radiation is 9 MJ m-2d-1 ±20%. Highest annual
solar radiation is in the subtropics (latitude 20° to 30°) (12 MJ m-2d-1), because the subtropics
are less clouded (NAKASONE and PAULL 1998 3 – 6, 21).
For the direct impact of absorbed photons within the PAR waveband and the use of CO2
within photosynthesis, it is common to describe radiation as photosynthetically active photon
flux density (PFD; µmol photons m-2s-1) (KÖRNER 2002 908). Above a tropical forest canopy
on a clear day in the dry season, PFD exceed 2200 µmol m-2 s-1 (CHAZDON et al. 1996 1).
The next question which has to be considered is how PAR is entering the canopy of an
agroforestry stand.
One part of the incoming radiation will be reflected by the canopy depending on its
prosperities. For example, a forest of Picea sp. has a reflection of 20%. The rest of the
radiation will be absorbed by the canopy or transmitted. Below the canopy most of the
incoming radiation is scattered and reflected due to leaves and woody surfaces. Especially
species with needles of fine-fingered leaves are increasing the proportion of diffuse radiation.

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 5
Direct radiation is entering the canopy through gaps, so called sunflecks. Although diffuse
radiation makes up the major part of solar radiation below a canopy, the majority of incoming
photons is of direct radiation. Thereby, sunflecks with longer duration are more likely to reach
full sun conditions than more frequent sunflecks with shorter duration. In addition, few big
gaps contribute more to direct sunbeam than many small gaps. The impact of gap size is not
linear; at the edges as well as at the centre of a gap there are different PFD values. In a
tropical forest it is seldom that daily PFD is above 50% of full sun conditions at the center of
relatively large gaps (see Fig. 2). In Costa Rica within a forest in the centre of a gap of 350m²
just 15% of daily incoming PFD exceeded 500 µmol m-2 s-1 (CHAZDON et al. 1996 9 – 14,
KÖRNER 2002 908).
Fig. 2: Instanenous Photon Flux Density (PFD) at two points of the same gap
with a size of 350 m² within a forest, Costa Rica
Reference: CHAZDON et al. (1996)
How much light the understory crop is receiving is depending on leaf area index (L) and the
angular arrangement of the leaves of both the crop as well as the upper story. “The size of the
intercepting surface depends on the green leaf area index (L) of a crop which can be
expressed as the product of the number of plants per unit of ground area, the number of leaves

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 6
Fig. 3: The exponential attenuation of solar radiation in a plant stand can be
expressed by a Beer’s Law type. Here, Leaf Area Index (L) = 1 to ease the
explanation; downwards L will be accumulated; k in equation 4 is depending
on plant structure (here: k=0.69)
Reference: Körner (2002 910)
per plant and the mean area of leaves per plant.” (AZAM-ALI and SQUIRE 2001 30). In a forest
stand the other upper parts of the plants can also be considered to L as trunks and stems are
reflecting and absorbing light. The impact on L is typically below 1 (KÖRNER 2002 911).
The angle of the leaves can be expressed as the extinction coefficient (k). Plants with narrow
and vertical leaves have a lower value of k than plants with broader and more horizontal
leaves. The attenuation of radiation through a canopy of a plant is exponential and obeys
Beer’s law of exponential decay (see Fig. 3) (AZAM-ALI and SQUIRE 2001 30).

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 7
Fig. 4: Using Beer’s law requires a random structure of the canopy; this could
be a problem where crops are planted in rows with inter-row spaces
REFERENCE: AZAM-ALI and SQUIRE (2001 30)
KÖRNER (2006 910) shows Beer´s law, also used in banana by JIMÉNEZ and LHOMME (1994):
I = I0 *e[-kL] (4)
where I is the incident radiation below the canopy, Io the radiation above the canopy, L the
leaf area index and k the attenuation coefficient.
The use of equation 4 requires a random structure of the canopy so that radiation is at any
level relatively uniform (see Fig. 4). Otherwise it fails (AZAM-ALI and SQUIRE 2001 30).

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 8
Furthermore, JIMÉNEZ and LHOMME (1994) recommend following equation (changed) for
estimating absorbed radiation:
Ia = I0 - Ir - It + Irs (5)
Where Ia is the PFD absorbed by the canopy, I0 is the incident radiation, Ir is the PFD reflected
by the canopy, Irs is the PFD reflected by the soil, and It is the PFD transmitted through the
canopy
The productivity of a plant is not only depending on intercepted radiation but also on dry
matter that is accumulated due to absorbed radiation. This is expressed by the radiation use
efficiency (γ) (NORGROVE 1998, TURNER 1998a)
γ = biomass accumulated / radiation intercepted (6)
Radiation use efficiency is also depending on seasonal variation, stress factors or pests and
diseases, plant development stage as well as some phenological factors (e.g. production of
lipid costs more energy than the production of carbohydrate) (AZAM-ALI and SQUIRE 2001
30).
Following, some practical ideas of agroforestry systems considering light will be discussed.
“Radiation, however, is normally not the factor limiting plant growth in the tropics, except
under circumstances of heavy mist and cloud or shade from vegetation and mountains”
(NAKASONE and PAULL 1998 21). The opinion of the authors might be considering tasks of
conventional horticulture.

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 9
More severe is the statement of CHAZDON et al. (1996 5), considering radiation regime of
tropical forests: “Across the complex matrix of microsites that compose tropical forests, light
availability varies dramatically than any other single plant resource. … Among the
environmental factors that influence plant growth and survival in tropical forests, light
availability is likely to be the resource most frequently limiting growth, survival, and
reproduction.”
Agroforestrial systems might range inbetween these to points of view.
The general idea of an agroforestry system considering radiation is the improvement of
photosynthetic efficiency because the leaves of the crops are also set in a vertical plane.
Therefore, light interception is increased and dispersion of solar radiation is improved.
Besides, shading might have some positive effect on the system, like a decreased weed
pressure. However, the understorey crop has to face an environment in which light
competition is high (NORGROVE 1998).
Fig. 5: Amount of solar energy in four Central American banana production
areas expressed in average Langleys/day (1 Langley = 1g cal/cm²)
Reference: STOVER and SIMMONDS (1987 204)

Christian Dold Light Interception in Tropical Agroforestry Systems
Seite 10
In conclusion, it is not possible to generalize the ambient radiation of the whole tropics, as it
is depending on season, latitude and cloud cover (dry and wet season) (see also Fig. 5)
(STOVER and SIMMONDS 1987 204-205). In addition, radiation interception in agroforestry
systems is much more complicated than in monoculture. Light interception is depending on
the location of each species, the spatial distribution of the leaves, solar angle, and row
orientation. In addition, plants may interact different in reduced light compared to full sun
(AZAM-ALI and SQUIRE 2001 119-121).
Furthermore the amount of diffuse and direct light below the canopy and hence the
distribution and size of gaps in the canopy are of importance. Above all, daily PFD can vary
greatly; the same plant often grows in highly different light regimes (CHAZDON et al. 1996
12). In the special case of shaded banana, the radiation environment can be more distinct, for
banana production takes also place in the subtropics. Therefore it is questionable to adapt
results to different light environments (ISRAELI et al.1995). In addition light environment
differs greatly for banana; the young sucker first growing in the shade of the mother plant will
find a different light environment after harvesting the mother plant (STOVER 1984).

Christian Dold Musa in Agroforestry Systems
Seite 11
3 Musa in Agroforestry Systems
3.1 Importance of Musa on Farm Level in Central America
In Central America and Mexico it is common method producing coffee in agroforestry
systems. Coffee is grown together with different fruit and timber tree species as well as
Musaceae. The farmer´s objective is the establishment of light conditions which benefits
coffee production. Besides shading, farmers have advantage of by-products of the shading
plants. Thereby, Musaceae are of great importance.
The function of Musa spp. in coffee agroforestry systems is wide ranged. Leaves and fruits are
used for fodder; fruits are used for self consumption, and as an important source of whole year
income. It is a fast growing species which is helpful for temporal shading of coffee when
timber trees are still too small, as it could be in newly established fields or after cutting timber
trees (YÉPEZ, 2001, SOTO-PINTO et al., 2000, SCHIBLI 2001).
Especially small-scale farmers prefer the cultivation of Musa; with increasing farm size the
density of Musa decreases. Cultivation methods of banana like desuckering, deleafing and
cutting the pseudostems are seen to be basic labour in a coffee agroforestry system (LÓPEZ et
al. 2003).
Production of banana in agroforestry systems must be distinguished to banana production in
monocoulture. Banana production in agroforestry systems is mostly extensive with small
inputs of labour, fertilizer and pesticides. Thus, compared to high intensive monoculture, there
is different intension of banana growing, for example small scale farmers often produce for
regional or local markets (SCHIBLI 2001). Following some examples will show the importance
of banana in shaded perennial crops.

Christian Dold Musa in Agroforestry Systems
Seite 12
Most of Mexico´s coffee production is based on agroforestry in small or medium farm sizes.
Where coffee is grown in polyculture, Musa is part of the lower strata. Research shows that in
a five layered coffee multistrata system Musa sapientum is the most frequent species in the
second strata (3m to 6m). Only Inga species, which are part of the upper strata, is more
frequent in the whole system (MOGUEL and TOLEDO, 1999, SOTO-PINTO et al. 2000).
More information about Musa in coffee agroforestry systems is available of Nicaragua.
Interviews in northern Nicaragua show that, besides coffee, Musa are of great interest in
agroforestry systems.
Musa is mentioned as the most frequent species of the upper storeys, followed by Inga spp.,
followed by Citrus spp.. The combinations of the upper storey differed slightly, but in 83% of
all cases Musa, guineo blanco (M. acuminata AAA) and plantain (M. acuminata x M.
balbisiana), were included. In addition, Musa made 14% of the gross income of coffee,
together with the advantage to be a whole year crop. 89% of the families used Musa in
subsistence farming (SCHIBLI , 2001).
Thus, the four main uses of Musa in Nicaragua were self consumption, shading of coffee,
trade, and income possibility independent to seasons. Besides, the farmers mention that Musa
has some further benefits; failed leaf abscission, temporal shading, maintaining the humidity,
and fertilizing the soil (SCHIBLI , 2001).
In the region of Talamanca, Costa Rica, organic banana production is managed in agro-
silvyculture systems, either Inga sp. – Theobroma sp.- Musa spp.- systems or Inga sp.- Musa
spp. - systems (SUAREZ and SOMARRIBA 2002). The case of Talamanca is therefore
interesting, as banana production is for export market; the fruits are for the production of
organic baby food. But it is also not representive as the facilities are already given by the
market chain (STAVER 2007; personal communication).

Christian Dold Musa in Agroforestry Systems
Seite 13
3.2 Research Review on Musa in Shade
Although Musa is a common crop in agroforestry systems, just a few studies are available
about the effects of shade upon growth and yield of Musa (NORGROVE 1998). Two tasks are
often observed in shaded banana; duration of crop cycle and response on yield.
In the case of crop cycle, most studies have shown that life cycle of banana in shade is more
or less lengthened which leads to delayed yield.
TORQUEBIAU and AKYEAMPONG (1994) found in Kenya that crop cycle of Musa ‘Dwarf
Cavendish’ (AAA) in 70% light, 50% light and 25% light is lengthened up to 5 days, 95 days
and 245 days compared to full sun, respectively.
In Puerto Rico, growth cycle was lengthened in shade; 38% of the bananas in 50% light were
harvested after 16 to 17 months, whereas in sun 65% were harvested in the same period
testing plantains var. ‘Maricongo’, and bananas, ‘Cavendish’ (AAA) as well as the variety
‘Monte-Christo’ (VICENTE-CHANDLER et al. 1966).
In Trinidad, growth cycle of Musa ‘Dwarf Cavendish’ (AAA) was 13.5 months, 11.1 months,
10.7 months and 10.4 months in 20%, 50%, 70% and 100% of sunlight, respectively
(MURRAY, 1961).
ROBINSON and NEL (1988) observed the impact of high densities of Musa (AAA) cv.
‘Williams’ (Cavendish subgroup) in monoculture where radiation is decreased due to mutual
shading. It is concluded that a longer crop cycle is the reason of a slower Leaf Emission Rate
(LER; Method of measuring leaf evolving in banana/time). This is due to the close correlation
of LER and temperature. As temperature is decreased in shade LER is slower.
Growth cycle of banana ends with flowering. Flower initiation of banana has no relation to
photoperiod or temperature (NAKASONE and PAULL 1998 105). ROBINSON and NEL (1988)
assume that flowering needs a minimum efficient leaf area or assimilation potential as a
stimulus for the growing point. The stimulus could interact with assimilate or hormone.
Banana produces in higher densities a higher total number of leaves till flower initiation. The
slower LER together with a higher number of leaves leads to a lengthened crop cycle.

Christian Dold Musa in Agroforestry Systems
Seite 14
The delay seems to be more severe in the following ratoon crop cycles (R) than in the first
crop cycle (P). ISRAELI et al. (1995) tested ‘Grand Nain’ (AAA) bananas in the Jordan Valley
of Israel in different light levels of 80%, 60%, and 30%. Although in P there was no
siginificant delay of yield, the first ratoon (R1) was delayed up to 6 days, 9 days and 15 days
compared to full sun, respectively. It is assumed that the impact on P was less as shade was
established two months before flowering.
The fact that the ratoon crop cycle is lengthened has its reason in the high dependence of the
sucker to the mother plant. A high Leaf area index (L) of the mother plant lengthens crop
cycle and delays sucker growth (ROBINSON and NEL, 1989, found in ECKSTEIN and ROBINSON,
1999). Growth of the sucker is fastened when the canopy of the mother plant is more open
due to higher temperature and solar radiation (ROBINSON and NEL, 1988).
Taking the focus on yield, the studies are discriminated. Although many research have found
that yield is decreased in some amount of shade, there are studies which indicate that yield
might be stable or increase in shaded conditions.
TORQUEBIAU and AKYEAMPONG (1994), observe that highest yield was reached at 50% light,
with 20% higher yield compared to open sun. Bunches have more fruits, but lower weight per
unit, and more rodden fruits.
Also MURRAY (1961) observes highest yields at 50% and 70% with both 2.44 kg compared to
2.18 kg per bunch and month in full sun. But as just two plants per plot were tested, stastitical
data are missing. It is suggested that the use of shade cloth might have a positive effect on the
microclimate and therefore yield was higher in shade.
VICENTE-CHANDLER et al. (1966) mention that banana and plantain yield in 50% light is twice
as high than in sun. Plantains had higher yields with more fruits per bunch and the same
weight per fruit. Also, bananas had higher yields with more hands per bunch.
The reason is that disease impact of Yellow Sigatoka (Mycosphaerella musicola, syn.:
Cercospora musae) in shade is less due to reduced due formation on banana leaves. The
plants in sun were severly damaged, while the leaves in shade were healthier.

Christian Dold Musa in Agroforestry Systems
Seite 15
After seven months, in average the unshaded plantains had 20.7 highly infected leaves,
compared to 0.06 leaves in shade. In addition, 49.4 banana leaves in sun were infected versus
25 leaves in shade.
ISRAELI et al. (2002) compared Grand Nain (AAA) banana under artificial shade (shadehouse
with transparent/white nets with a light interception of 83% and 72%, respectively), in the
Jordan Valley, Israel, to full sun. Yields were similar in all treatments due to a better
microclimate for banana beneath shade cloth. Water pressure was very high and more limiting
than light.
On the other hand, ISRAELI et al. (1995) found that yield was less in all shade levels in P and
R1. In the first cycle at 60% light bunch weight was reduced about 7%; 30% of full sunlight
reduced bunch weight about 32% compared to full sun. In the second cycle, yield was
statistically significant reduced by 12t/ha (-15%) in 80% light, 19t/ha (-24%) in 60% light and
49t/ha (-61%) in 30% light. Shade affects a reduced finger weight; in 30% light in reduced
finger length, and thus banana is not commercially valued. Also ECKSTEIN et al. (1997)
(found in NORGROVE, 1998) observed a reduced bunch weight in shade, in P cycle as well as
R1.
Other factors also indicate that banana production in sun has higher yield. Metabolism of
shaded leaves is less (MURRAY, 1961); stomatal density is reduced in 30% light, especially on
the adaxial side (ISRAELI et al., 1995); and photosynthesis rate of banana leaves (‘Williams’
AAA) decreases greatly on overcast days and even more due to self-shading (ROBINSON,
1996, 89-90).
ROBINSON (1996 83) shows different effects which reduces PAR and thus, photosynthesis
(see Tab. 1).

Christian Dold Musa in Agroforestry Systems
Seite 16
Tab. 1: Summary of different research about impacts on photosynthesis rate; all data
are referring to subtropical climate and AAA Cavendish subgroup
Reference: Robinson (1996 83,87)
Internal Factor Photosynthesis Rate
(µmol CO2 m-2 s-1)
External Factor Photosynthesis Rate
(µmol CO2 m-2 s-1)
Leaf Number Leaf Tearing
1 (youngest) 11.7 Untorn Leaf 20.5
2 18.7 50 mm strip width 18.3
3 21.0 25 mm strip width 16.5
4 20.6 12 mm strip width 13.7
5 19.4 Soil water potential
6 (mid profile) 17.6 Unstressed plants 26.4
Leaf surface (leaf 3, 4 and 5) Stressed for 4 days 24.2
Abaxial/adaxial
(summer)
33.2 / 21.6 Weather Conditions (summer)
Abaxial/adaxial
(winter)
18.8 / 6.1 Sunny day (PAR=1974) 17.2
Cultivar (AAA) (summer) Overcast (PAR = 477) 11.4
Dwarf Cavendish 29.1 Mutual shaded from Canopy (summer)
Grand Nain 28.6 Leaf 3 sunlit (PAR=1652) 20.9
Valery 27.5 Leaf 4 shaded (PAR= 80) 6.8

Christian Dold Musa in Agroforestry Systems
Seite 17
The differences of results in yield are interesting, especially because all reasons are still
unclear. NORGROVE (1998) and TURNER (1998a) try to answer the task with an analytical
approach taking available data on banana in shade into account:
The approach includes the theory of MONTEITH (1977) (found in NORGROVE 1998) which
indicate that accumulated dry matter and intercepted radiation is proportional. “Assuming
everthing remains constant, other than the incident radiation, a 50% reduction in radiation will
cause a 50% reduction in yield“(TURNER 1998a)
TURNER (1998a) shows that the results of MURRAY (1961) as well as ISRAELI et al. (1995) are
not proportional to radiation (see Fig. 6). The reason might be that there is a difference
between incident radiation and intercepted radiation.
Fig. 6: TURNER’s (1998a) approach to compare the results of MURRAY (1961),
P (M) (one crop cycle), and ISRAELI et al. (1995) P (I) and R (I) (first crop
cycle and ratoon crop) with the theory of MONTEITH (1977); no result show 1:1
accuracy
Reference: Turner (1998a)

Christian Dold Musa in Agroforestry Systems
Seite 18
Another approach to understand MONTEITH’s theory (1977) is by following equation
(changed):
Y= {I 0[1-e-kL]γTcHi}/d m (7)
Where Y is the yield, I0 is the incoming radiation, L is the leaf area index, k is the extinction
coefficient, γ is the radiation use efficiency, Tc is the temperature coefficient (between 0 and
1; 0 describes the temperature when growth stops and 1 is the optimum temperature), Hi is the
harvest index (proportion of dry matter of the fruit) and dm is the percentage of dry matter in
the fruit tissue (one possible unit of expressing yield) (TURNER 1998a, TURNER 1998b).
The most probable factors which change in shade considering in equation (7) are k, L and γ
(Tc is assumed not to change; H might not change due to shade) (TURNER 1998a).
NORGROVE (1998) suggested some hypothesises why γ could change. Some of them are
presented as followed, and will be discussed further:
• The PAR saturation point is below full light conditions
• Decreased yield losses due to leaf diseases
• Water and nutrients are less limiting in shade
As already mentioned diseases and pests can decrease γ. In the case of VICENTE-CHANDLER et
al. (1966) the impact of Yellow Sigatoka was less in shaded banana and plantain. Thus, the
losses of Yellow Sigatoka are being more limiting than the reduced radiation. NORGROVE
(1998) also mention that the impact of Black Sigatoka (Mycosphaerella fijiensis (Morelet)),
one major disease in banana production in the tropics, is less in shade than in sun.

Christian Dold Musa in Agroforestry Systems
Seite 19
Also other factors than light might be more limiting.When water (precipitation) and nutrients
are not limited, light is the next most probable limiting factor. Thus, if nutrients or water are
not sufficient in relation to the needs of the plant, the quantity of light will not contribute to
plant growth. In agroforestrial systems the competition of water, nutrients and light is high.
Especially, when high trees are used, the negative impacts for the understory crops are
significant (TORQUEBIAU and AKYEAMPONG, 1994).
Banana is reacting quite fast on water losses; although the plant appears vigour, the stomata
close which has an impact on photosynthesis (Turner 1998a). MURRAY (1961) recommended
banana production in shade, when nutrients are more limiting, as nutrient content of shaded
banana leaves is higher.
This attends also with the study of ISRAELI et al. (2002). The author found that the positive
microclimate (wind protection) and thus, the reduction of water stress, overcome the negative
impact of shade.
Furthermore, NORGROVE (1998) suggests that banana might have a light saturation point
which is below full sunlight; light saturation point is just said to be 20% of light flux density
of a cloudless day in the tropics. Also NAKASONE and PAULL (1998 21) describe Musa as a
C3-Plant; hence, the rate of photosynthesis is limited by ambient carbon dioxide (CO2) rather
than light.
In contrast, ROBINSON (1996, 90) recomnmends a PAR of 1500 – 2000 µmol m-2s-1 to be
adequate for banana production.
TURNER (1998a) sums up different studies describing light response curves of different
cultivars and different subgroups (see Fig. 7). While some studies show light saturating points
below full sunlight, others are not saturated at full sun.
In another study of TURNER (1998b) the author is sure that banana is not saturated at full
sunlight showing some results of Fig. 7. Then, shade will have a high impact on banana.

Christian Dold Musa in Agroforestry Systems
Seite 20
Fig. 7: Summary of different research about cultivar light response curves;
Cavendish, Williams and Gros Michel belong to AAA while Bluggoe belongs
to ABB
Reference: TURNER (1998a)
Apart from γ other factors are likely to change. The extinction coefficient k is changing in
shade due to less (naturally) diurnal folding of banana leaves. Even well irrigated banana
plants fold to avoid overheating. Thus, radiation interception is reduced. This effect is
stronger when plants are water stressed. In addition, k is changing within the depth of the
banana canopy (TURNER 1998a, TURNER 1998b, THOMAS and TURNER 2000).
Besides, leaf area index (L) stays stable or might increase in some amount of shade. ISRAELI
et al. (1995) found no statistical difference in leaf area until 30% light. ROBINSON and NEL
(1988) recognized an almost doubled L of ten plants at flowering in higher densities of 2222
plants per hectare compared to 1000 plants per hectare.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 21
4 Musa in Shaded Coffee - A Case Study of Costa Rica
4.1 Description
Aim of this part of the work is to evaluate the morphological differences of Musa spp. in
shade and light. In addition, the amount of light in different agroforestry system is evaluated.
Therefore, measurements were done from the end of March to the beginning of September in
2007. Furthermore, results of a small number of ten interviews will be presented.
4.1.1 Botanic garden – Musa ‘Gros Michel’ in Sun and Shade
Banana plants in the botanic garden (626 m, N 09.89505°, W 083.65709°) of the Tropical
Agriculture Research and Higher Education Centre (CATIE), Turrialba, Costa Rica are used
for morphological measurements. The variety is Musa ‘Gros Michel’ (AAA). Ten plants in
shade and ten plants in sun are chosen randomized for measurements. The banana plants are
of unknown but different age. The number of suckers and pseudostems at each plant differed
and was very high. In 2007, week 12, in sun and shade the plants had an averaged number of
4 ± 3 and 6 ± 3 sword suckers and maidens up to 1m, 2 ± 1 and 2 ± 1 suckers up to 2m as well
as 4 ± 2 and 3 ± 1 non-fruitbearing and fruit bearing pseudostems higher than 2m,
respectively. Two pseudostems in two different heights at every plant were chosen. Those are
suckers between 1m and 2m (named to Category Pseudostems II), and not flowering
pseudostems with more than 2m (named to Category Pseudostems III). There are no
replications.
The banana plants are mainly associated with cacao plants and trees (Schizolobium
parahybum). The highest pseudostems of banana shade cacao and an upper storey of trees is
shading the banana plants. Therefore, three different storeys can be generally observed.
Where the banana plants are placed in full sun the upper storey of the trees is missing.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 22
Fig. 8: Banana ‘Gros Michel’ mainly asociatied with cacao in the botanical
garden, CATIE; the high pseudostems is shading the cacao plants while
young suckers have to grow in shade of banana and cacao; the shaded part
has an upper storey of Schizolobium parahybum
The banana plants in particular have more complex shade conditions. The shoots of one plant
have different heights. Smaller suckers can be shaded by the upper storeys of cacao or by the
shade trees as well as by the plant itself due to the higher pseudostems. Thus, even in full sun
the smaller suckers are shaded (see Fig. 8).

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 23
4.1.2 Six Varieties in Four Different Light Levels
Besides, first results of growth measurements of six different banana varieties, Gros Michel
(AAA), Plantain (AAB), Bluggoe (ABB), Baby (AA), FHIA-18 (AAAA), and Manzano
(AAB), in different light conditions are presented.
Four plots of 300m² to 400m² are established at CATIE, Turrialba, Costa Rica (9:52:59 N,
83:39:59 W, 624 m). Five young banana suckers (pre-cultivated, 9 to 11 weeks in
greenhouses, except the variety ‘Bluggoe’ where the corms were directly planted) of each
variety are planted (3m to 4m x 3m to 4m) randomized in full sun and three agroforestry
coffee fields (Coffea sp.). The coffee is associated with Poró (Erythrina poepiggiana) and
Cedro (Cedrela odorata). Along the edge of the plots there are six Pinus sp.. Coffee has a
planting distance of 1m x 1.5m. There are no replications. At planting in week 33 2kg of fresh
Poró leaves and 500g of Gallinaza (chicken manure) was applied (see also Appendix IV).
As the bananas were in the sucker stage at planting time, the plants are shaded by two to three
upper storeys; by Poró and Cedro as well as coffee (see Fig. 8). The bananas in plain field are
none or just less shaded by the surrounded area.
Fig. 9: The shaded banana suckers are growing in the shade of coffee as
well as the upper storey of Poró and Cedro

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 24
4.1.3 Light measurements in Agroforestry Systems
At Tropical Agriculture Research and Higher Education Centre (CATIE), Turrialba, Costa
Rica, incoming light in coffee agroforestry stands with a second storey of trees was estimated.
Therefore, six plots were chosen (9° 53’ 44’’ N, 83° 40’ 7’’ W, 600 msnm, 2600 mma). The
plots are of different size, between 1612 m² and 4088 m², but all plots involved a microplot of
288 m², in which light measurements were done. The plots have different tree mixtures,
building the upper canopy. Those are single stands with Erythrina poeppiggiana (Poró),
Terminalia amazonia (Roble Coral), Abarema idiopoda (Cashá), and mixed stands with Poró
together with Roble Coral, Poró together with Cashá as well as Cashá together with Roble
Coral. Poró is a broad-leafed, nitrogen-fixing tree species which is frequently used for shading
in coffee-agroforestry systems in Costa Rica. Cashá is a fine fingered-leafed species with an
umbrella-like habitus, while Roble Coral has small – medium simple leaves with a
columnaris-like habitus.
The plots are fertilized with 2 kg of decomposed coffee residuals. Coffee is planted 1m x 2m,
while the trees have a distance of 6m x 4m. The single Poró treatment has a tree planting
distance of 12m x 8m (transplanted). At time of measurements, Poró was pruned keeping the
trees small with two major branches. In addition, tree density in the other plots is lower due to
losses, and some trees are less established. Coffee has a height of 1.80m to 3m.
4.1.4 Interviews
Ten interviews were done mostly in the region of Turrialba, Costa Rica, but also in the region
of Talamanca, Costa Rica. Mostly small scale farmers were interviewed with an established
simple questionnaire (see Appendix V). The intervieweés are not randomized chosen as it was
not possible to find enough participants for this procedure. Furthermore, a translator was
needed. Together with the small number of interviews, the results are not representative and
can just give an idea of Musa in agroforestry systems in both regions. Besides, light was
measured using a densiometer. The intervieweés are asked for 1 to 2 good and bad sites for
banana production in their fields considering light environment. Then light was measured at
one from the interviewer chosen point.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 25
4.2 Material and Methods
Table 2 gives the frequency of measurements in the botanic garden and the treatments with
the six varieties. Material and methods of the interviews and the light measurements, see 4.2.1
Tab. 2: Method of growth measurement and frequency
Method Botanic Garden Banana
Varieties
Light Measuremensts Once Once
Pseudostem height Two-week
All pseudostems
Twice
Circumference Two-week
Pseudostems III
Twice
Leaf Emission Rate Weekly
All pseudostems
---
Functional leaf number Weekly
All pseudostems
Twice
Length of the petiole --- Twice
Non-destructive measurements of leaf
area
Weekly
Pseudostems II
Twice
Destructive measurements of leaf area Pseudostems II - Once
Pseudostem III - Once
twenty different plants in sun and shade - Once
---
Dead leaves due to Black Sigatoka Weekly
All pseudostems
---

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 26
Disease Development Rate (DDT) Weekly
All pseudostems
---
Youngest Leaf Spotted (YLS) Weekly
All pseudostems
---
Leaf angle Once
Pseudostem II
---
Leaf blade angle Twice
Pseudostem II
---
Leaf tearing Twice
twenty different plants in sun and shade
---
4.2.1 Light Measurements
Light is measured using hemispherical photography (Nikon Coolpix 8700; Nikon FC-E9 fish-
eye Converter, 7 mm f/2.8 approx.183°, equidistant projection).
Hemispherical photography can evaluate light up to 2.5 m and less and is a good possibility to
compare radiation of different sites. It is also an often used method for measuring light in tree
stands. However, it is not possible to measure intercepted light, but incident light (CHAZDON
et al. 1996 10, 13).
In the site with different banana varieties, in total 120 photos were taken above the banana
plants, 30 for each plot. The mean was taken to evaluate average total transmitted light for
each plot. In addition, 12 photos were taken in the most shaded plot above the suckers in
height of the coffee storey. Then, light enivironment was compared in the height of the sucker
to the height above coffee.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 27
In the botanic garden of CATIE, light was measured above the pseudostems. Thus, 10 photos
for each height level are taken and the mean gives average total transmitted light.
In the agroforestry systems light is measured in the height of the coffee every 2m x 2m. Then,
light dispersal maps are made. In total for each plot around 100 photos are taken.
All photos are taken in direction to the geographical north using a compass. Thus, some
abbreviation of 5 -15 degrees NO/NW occurred. The camera is levelled to ensure a photo,
straight to the upper canopy. Little abbreviaition when levelling occurred. Besides, direct
sunbeam led to distortions in light.
The photographies are evaluated with the software Gap Light Analyzer (GLA), Version 2.0
(SFU, Barnaby, Canada; IES, Millbrook, USA).
GLA requires some data for light estimation. Latitude, longitude and elevation were measured
using GPS (Global Positioning System). Magnetic declination of geographic north and
magnetic north was estimated on the homepage of the Geological Survey of Canada,
http://www.geolab.nrcan.gc.ca, clination (FRAZER et al. 1999).
Cloudiness Index, Beam Fraction and Spectral Fraction were estimated as recommended by
FRAZER et al. (1999) (see also equation (1), (2), (3)). The required data was taken from the
weather station of CATIE, Turrialba, Costa Rica. The average daily solar radiaton per month
from 1968 to 2005 was available.
A whole year progression of the sun was desired. The sun position from sunrise to sunset is
measured every 2 minutes. The number of azimuth regions was set by 36 and the number of
zenith regions was set by 10.
Every picture was threshold manually by the same person.
For the interviews, light measurements with densiometer are done field. Canopy closure is
evaluated in the four directions, North, East, South and West, and every count was multiplied
with 1.04 as it is common method. The mean of the four directions gives the overstorey
density.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 28
4.2.2 Number of Functional Leaves
Banana leaves are distinguished between functional and non-functional. Functional leaves of
banana are defined as leaves with a minimum width of 10 cm (MOREAU 1965, found in
STOVER 1979). Only functional leaves are counted (see Fig.10). Fully emerged leaves are
marked as described in ROBINSON (1996 72).
Fig. 10: Example for counting the total number of leaves
Reference: VARGAS et al. (1992)
4.2.3 Length of the Petiole
The length of the petiole is the distance from the pseudostem to the leaf base. Each petiole of
every functional leaf was measured.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 29
4.2.4 Non-destructive Measurements of Leaf Area
Numerous equations for estimating leaf area are published. MURRAY (1960) suggested the
equation:
Leaf Area = Length x Width x 0.8 (8)
STOVER (1982) compared the results of the equation with measuring leaf area using a
planimeter and was accurate within 2%.
Leaf area is measured from every new and fully evolved leaf. Length is measured along the
midrib of the leaf and width at the widest part of the leaf. Only functional leaves are involved
in leaf area measurements. The sum of all leaves gives the total leaf area.
4.2.5 Destructive Measurements of Leaf Area
Leaf area differs relatively to plant development. First, leaf area is exponentially increasing,
followed by a phase of similar leaf area (plateau phase) followed by a short period of
decreased leaf area till flowering (TURNER 2003). STOVER (1979) shows a typical graph of the
increasing leaf area of 15 ‘Grand Nain’ plants (see Fig. 11). Referring on those results,
TURNER (2003) mention an exponential phase of at least 75%.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 30
Fig. 11: Increasing leaf area of 15 'Grand Nain' (AAA) in Honduras, 1976
Reference: STOVER (1979)
TURNER (2003) estimated leaf area including the exponential phase and the number of leaves.
The rate of exponentially increasing leaf area is (changed):
AR = (lnAN - lnAi)/( N-1) (9)
where
AR: rate of increasing leaf area
AN: area of the youngest leaf
A i: area of the oldest green leaf
N: the youngest leaf (and total number of leaves)

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 31
To estimate leaf area between any two leaves without considering the plateau phase
(changed):
A i,N = Ai[(exp(AR*N) - exp(AR*i))/A R] (10)
where Ai,N is the integration of the leaf areas between the oldest leaf (i) at the time of
measurement and the youngest leaf (N), Ai is the area of the oldest leaf on the plant, AR is the
value of equation (9), Leaf i has the value 1 and Leaf N is equivalent to the total number of
leaves.
To use equation (10), measurements of Ai, AN, and N are needed. Then, AR can be computed
(TURNER 2003).
It was just possible to estimate leaf area of the Pseudostems III in the botanic garden with
destructive measurements.
For measuring leaf area of the Pseudostems III, the fifth leaf of twenty other plants in the
botanical garden was cut in week 18. The plants were of the same current height like the
pseudostems, assuming that plant development is related to height and thus similar. Length
and width was measured and leaf area was estimated according to MURRAY (1960) which
represents Ai in equation (10). In week 35 the youngest leaf of the Pseudostems III was cut
and leaf area according to MURRAY (1960) was estimated which represents AN in equation
(10).
Total leaf number (N) is the ratio between the youngest leaf in Week 35 and the 5th leaf in
Week 18 at the Pseudostems III.
Total Leaf Area evolved was estimated from Week 18 to Week 35. After using equation (10)
the mean was taken.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 32
After week 27 pseudostem height and leaf length of Pseudostem II in the botanic garden were
too high for non-destructive measurements. Thus, leaf area need to be evaluated by using
destructive methods.
A i was the youngest leaf in week 27. AN was the youngest leaf in week 35. Total leaf number
(N) was the leaf number between week 27 and week 35. Total leaf area evolved was estimated
from Week 27 to Week 35. After using equation (10) the mean was taken. TURNER (2003)
suggested an overestimation of 10% by using equation (10).
4.2.6 Leaf Emission Rate (LER)
The folded leaves are already produced within the pseudostem. Due to a rapid growth of the
leaf sheath, which can be 4m in 10 days for the variety ’Gros Michel’, the leaves evolve out
of the pseudostem. After evolving the leaf unfolds (CARLIER et al. 2002). BRUN (1963)
defines the unfolding of a banana leaf in five different stages (originally from A to E; now
with a scaling) (see Appendix I). Using BRUN’s scale (1963), the development stage of a leaf
can be defined numerically. The ratio between two measurements is the LER.
4.2.7 Circumference and Height
Height and circumference of the banana pseudostem are correlated to number of hands
(STOVER and SIMMONDS 1987). Height is measured at one side of the corm from the ground
to the upper edge of the pseudostem. The pseudostem ends with the last pair of fully evolved
leaf sheath (ALVAREZ et al. 2001).
Circumference in the botanic garden is measured at 1m height (STOVER 1982, ALVAREZ et al.
2001).
Circumference of the six varieties was measured in 5cm above ground, as the suckers were
too small.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 33
4.2.8 Black Sigatoka Leaf Spot
The effect of Black Sigatoka (Mycosphaerella fijiensis (Morelet)) is estimated using GAUHL´s
(1989) scoring system (see Appendix II). Leaves, where the petiole collapses before Stage 6,
are not taken into account.
In addition, disease development time (DDT) is measured which is the time between a
unfurled leaf in Stage B of BRUN´s scale and the appearance of 10 mature necrotic lesions on
that leave. Moreover, the youngest leaf spotted (YLS) is evaluated, which is the number of
functional leaves beginning at the youngest leaf and ending at the leaf used for DDT
measurements (CARLIER et al. 2002, GAUHL 1989).
4.2.9 Leaf Tearing
Leaf tearing was measured on the second leaf and the fifth leaf of twenty banana plants, ten in
sun and ten in shade in week 18. The measurements were done on twenty different plants as
destructive methods would have an influence on the other measurements. The height of the
plants was similar of those with the Pseudostems III in week 18. Leaves were not damaged
and free of diseases, despite of some lesions due to Sigatoka Negra with disease severity of 1,
using GAUHL´s scoring system.
Torn leaves with strip widths of 12mm, 25mm, and 50mm have a reduced photosynthetic
efficiency per unit of leaf area by 33%, 20% and 11%, respectively (ROBINSON 1996 91).
TURNER (1994) recommends windbreaks when tearing is less than 50 mm (found in
NAKASONE and PAULL 1998). Thus, the stripes were counted and categorized in stripes of 0-
12 mm, 13-25 mm and 26-50 mm. In addition, leaf length and width was measured.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 34
4.2.10 Leaf Angle
As a banana leaf changes its angle from unfurling till collapsing, the leaves are compared due
to their development stage. Therefore, leaf angle was compared in relation to leaf
development starting with the youngest leaf.
The angle between the leaf sheath and the pseudostem is measured using a protractor (Fig.
12). As the pseudostem is not straight but getting narrower to the top, leaf angle was
measured assuming the pseudostem is straight by levelling the protractor.
Fig. 12: Measurement of the angle between leaf sheath and leaf angle,
mention the level at the mark of 90°

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 35
4.2.11 Leaf Folding
Banana leaves are easily water stressed, already before any visible signs occur. The further
reaction of water stress is leaf folding and wilting (ROBINSON 1996 90). Leaf blade angle was
compared in sun versus shade. At the beginning of the raining season leaf blade angle of the
second leaf was measured, once in the morning and once in the afternoon. Temperature was
measured at both times. Templates were used with three different angles of 135°, 90° and 45°.
Assuming that a healthy, not water-stressed leaf has a blade angle of 180° degree, four
categories of leaf blade angles were set: 180° - 136°, 135° - 91°, 90 – 46° and lower than 46°.
Leaf blade angle was measured by using the templates at the middle of the leaf at the midrib
(Fig. 12). Abnormal or diseased parts were avoided for measurements as they could also
cause differences in leaf blade angle.
Fig. 13: Measuring the leaf blade angle using a template

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 36
4.3 Results
First the results of the botanic garden will be presented, beginning with light data. The growth
data will be divided in the categories Pseudostems III and Pseudostems II. Then, the results of
the six varieties and the light data of the plots will be presented. Furthermore, the results of
the light measurements in coffee agroforestry systems will be evaluated. At last, data of the
interviews will be shown.
The graphs and statistical measurements (unpaired t-test, standard deviation, standard error,
mean) were done with the Software SigmaPlot 9.0.
4.3.1 Light Measurement in the Botanic Garden
Light measurements of the category Pseudostem III show a significant difference by p =
1.3700* 10-8 in light compared to the shaded plants. In sun the total transmitted light was
about 71% ± 11 standard deviation, compared to 32% ± 4 standard deviation in shade (see
also Fig. 14). To ease the formulations, following the treatments will distuingished between
71% and 32%. Canopy openness was about 50% ± 10 standard deviation in sun compared to
23% ± 2 standard deviation. The low value of canopy openness in sun is due to technical
problems taking the hemispherical photo above the plant canopy, as the first leaf, evolved or
unfurled, is straight. This reduces the value of canopy openness as well as total transmitted
light. In addition, the surrounding area is always contributing to some shade. Therefore, it can
be suggested, that the data might be higher in sun. In addition, direct sunbeam led to
overestimation, especially in the shaded area.
Thus, the absolute numbers might differ, but the relative impact of the upper storey is
predicted.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 37
Fig. 14: Radiation environment in week 18 in the two treatments above the
Pseudostem III out of 10 measurements each; the whiskers show the
standard error
Total transmitted light was significantly different in the two treatments by p = 2.0907 * 10-3.
Mean total transmitted light was about 34% ± 16 standard deviation in sun and 14% ± 4
standard deviation in shade (see also Fig. 15). Canopy openness in sun and shade was 25% ±
8 standard deviation and 11% ± 3 standard deviation, respectively.
Direct sunbeam led to overestimations, especially in the shaded plot. As canopy openness is
low it is questionable, if the abslolut numbers are predictable, as FRAZER (2001) only
recommended hemispherical camera in sites with canopy openness higher than 10%.
The high variety in the data of the sun treatment is due to the highly variable light
environment in which a young sucker grows beneath the mother plant. Furthermore, light
environment changed by time as the young suckers grew further and harvesting changed light
regime by cutting the mother plants.
Nevertheless, the difference between the two treatments could be predicted. Following the
treatments will be distuingished between 34% light and 14% light, to ease the formulations.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 38
Fig. 15: Radiation environment in week 18 in the two treatments above the
Pseudostem II out of 10 measurements each; the whiskers show the standard error
4.3.2 Results Pseudostems III
Three plants in sun flowered before the end of data taking. As growth stops after flowering,
the plants were excluded in these results. Therefore, for the measurements mostly in sun it is
n = 7, while in shade it is n = 10.
Statistically significant more leaves are found in Stage 6 of GAUHL ’s scoring system in 71%
transmitted light than in 32% transmitted light (see Tab. 3). While 63% of evolved leaves died
due to Black Sigatoka in sun only 20% are found in shade in the same period. Although DDT
just differs by 4 days there is a high significant difference between sun and shade, for a high
frequency of infected leaves in sun was found. DDT and YLS data is depending on leaves in

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 39
Stage B of BRUN`s scale of leaf development. 22 leaves in sun were used versus 13 leaves in
shade, which also indicate a reduced infection pressure due to shade.
In both cases the youngest leaf spotted (YLS) was the third to forth leaf, which means that the
impact of Black Sigatoka begins on leaves which are important on plant development.
Tab.3: Impact of Black Sigatoka on Pseudostems III in two different light
conditions from week 23 to week 35; dead leaves due to Sigatoka using
GAUHL ’s scoring table, the total number of leaves evolved in the same
period, Disease Development Time (DDT), Youngest Leaf Spotted
(YLS) and the total number of leaves used for DDT/YLS-
measurements; as three plants in sun flowered, n=7 while in shade
n=10; ns = no statistical difference; the values behind ± is the standard
deviation
Impacts of Sigatoka 71 % Total Transmitted
Light
32% Total Transmitted Light p
Leaves Stage 6 8.4 ± 2.5 3.5 ± 2.3 0.0000027535
Total Leaves Week 23 - 35 13.4 ± 2.5 17.8 ± 2.1 ns
Disease Development Time 50 days ± 11 54 days ± 11 0.0
Youngest Leaf Spotted 3.4 ± 1 3.6 ± 0.77 ns
Total Number YLS/DDT 22 13

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 40
Tab. 4: Leaf Tearing of twenty plants comparable in height of Pseudostems III in
week 18 (3.5 – 3.6 m); the average number of strips in three different
widths on both leaf blades; light conditions and development stage are
assumed to be similar; ns = no statistical difference; the values behind ±
is the standard deviation
Leaf Tearing of the 2nd leaf
Size of strips (cm) 1.2 p 2.5 p 5 p
Sun 0.9 ± 1.1 2.4 ± 2.2 3.9 ± 4.9
Shade 0.7 ± 1.6 ns 1.1 ± 1.1 ns 1.7 ±1.3 ns
Leaf Tearing of the 5th leaf
Size of strips (cm) 1.2 p 2.5 p 5 p
Sun 3.2 ± 4.6 5.5 ± 5.9 10.9 ± 9.3
Shade 0.8 ± 1.1 ns 1 ± 1.2 0.0287 3.5 ± 2.5 0.0254
Tab. 5: Estimated average leaf area of Pseudostems III from week 18 to week 35
using equation (10) (TURNER 2003), weekly leaf number at the plant,
and average total number of leaves evolved in the same period; in sun
three plants started to flower and therefore n=7, while in shade n=10;
the value behind ± is the standard deviation; ns = no statistical
difference TURNER (2003) assumed an overestimation of leaf area by
10%
Estimated Leaf Area (m²) Weekly Leaf Number Total Leaf Number
71 % Light 29.6 ± 7 7.0 ± 0.6 16.3 ± 1.5
32 % Light 24.19 ± 6.08 8.0 ± 0.6 14.5 ± 1.4
p ns 1,6333* 10-4 0,0253

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 41
The second leaf was neither in sun nor in shade severely torn; also there is no significant
difference between the two groups. The fifth leaf in sun was significantly higher torn with
strips of 2.5 cm and 5 cm width than in shade. Leaf tearing is a result of wind; while newly
developed leaves are still untorn; by time the higher impact of wind in the sun plot is tearing
the leaves (see Tab. 4).
Although in 71% total transmitted light the plants produced more leaf area, there is no
statistical significant difference. However, statistically more leaves were produced in 71%
light, while the weekly number of leaves was significant lower by 1 leaf (see Tab. 5).
Thus, the plants in sun had a faster leaf turn over, with less leaves at the plant per week and
more total produced leaves compared to the shaded plants.
Fig. 16 shows the weekly number of leaves in the whole period of week 13 to week 35. In
average, the shaded plants had more leaves per week. But only in week 16, 20, 21, 22 and 24
a statistical difference can be found. This also confirms the longevity of shaded banana
leaves.
Fig. 17 shows the Leaf Emission Rate (LER) from week 18 to week 35. Although LER in sun
was higher, only in week 23 there was statistical significant difference.
Nevertheless, it shows that leaf emission in sun was faster than in shade and therefore total
leaf number was higher.
Over the whole period there was no significant difference in height and circumference.
However, circumference was higher in sun than in shade. Besides, it is interesting that
although banana in sun was smaller in week 13, in average banana pseudostems were higher
in week 35.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 42
Fig. 16: Weekly number of leaves of Pseudostems III; the red dots are
marking the weeks when leaf number was statistically significant different
with p<5%; as 3 plants in sun flowered in 71% Light n=7, and for 32% n=10
Fig. 17: Weekly measurements of Leaf Emission Rate (LER) of Pseudostems
III; averaged data; as three plants in sun flowered, for 71% Light n=7 and
32% Light n=10; the red dots is marking the weeks of statistical significant
differences with p<5%

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 43
Fig. 18: Two-week measurements of height (cm) of Pseudostems III
averaged data; for 71% Light n=7 and for 32% n=10
Fig. 19: Measurements of circumference (cm) of Pseudosems III; averaged
data; as three plants in sun flowered, for 71% Light n=7 and 32% Light n=10

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 44
4.3.3 Results Pseudostem II
Tab. 6: Impact of Black Sigatoka on Pseudostems II in two different light conditions
from week 13 to week 35; dead leaves due to Sigatoka using GAUHL’s
scoring table, the total number of leaves evolved in the same period,
Disease Development Time (DDT), Youngest Leaf Spotted (YLS) and
the total number of leaves used for DDT/YLS-measurements; ns = no
statistical difference; the values behind ± is the standard deviation
Impacts of Sigatoka 34 % Total Transmitted
Light
14% Total Transmitted Light p
Leaves Stage 6 8.4 ± 4.1 3 ± 1.3 3.2480 * 10-3
Total Leaves Week 13 – 35 20.6 ± 2.9 13.5 ± 2.3 9.6530* 10-6
Disease Development Time 41.6 ± 7.8 days 53.4 ± 12.5 days 1.3616 * 10-3
Youngest Leaf Spotted 3.8 ± 0.8 4.1 ± 0.8 ns
Total Number YLS/DDT 30 9
The results of Sigatoka in the category Pseudostem II confirm the results of Sigatoka impact
on Pseudostem III. Again, significantly more leaves were destroyed by Black Sigatoka. 41%
of the leaves in sun died due to Sigatoka compared to 22% in shade in the same period.
Disease Development Time was significantly higher in shade than in sun by around 12 days.
But there was no statistical difference in Youngest Leaf Spotted (Tab. 6).
Much more leaves were used for measurements in sun than in shade because of the fast turn
over of the plants in sun.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 45
Tab. 7: Leaf area of week 26 by non-destructive measurements and estimated total
increase of leaf area using equation (10) (TURNER 2003) (destructive
measurements) from week 28 to 35, weekly leaf number, and total leaf
number from week 13 to 35; average data of n=10; the value behind ±
is the standard deviation; ns = no statistical difference; TURNER (2003)
assumed an overestimation of leaf area by 10%
Leaf Area 34% Light 14% Light p
Leaf Area Week 27 5.81 m² ± 0.97 2.96 m² ± 2.05 8.6916* 10-4
Total Leaf Area Week 28 - 35 7.01 m² ± 2.99 2.93 m² ± 1.54 1.1733 * 10-3
Weekly Leaf Number Week 13 - 35 6.2 ± 0.7 5.5 ± 1.4 ns (5.44%)
Total Leaf Number Week 13 - 35 20.6 ± 2.9 13.5 ± 2.3 9,6530* 10-6
Weekly leaf area was statistically different from the very beginning of data taking and did not
differ until the end of data taking (see Fig. 20). In addition, there is a statistically difference in
estimated leaf area until week 35 (Tab.7). Also the total leaf number differs statistically, and
although weekly leaf number is not statistically different, the p-value is nearby ά = 5% (see
Tab. 6).
Thus, it can be concluded that the young suckers produced more leaves, with a higher weekly
leaf number and a higher leaf area.
Leaf emergence rate (LER) was mostly higher in sun than in shade, and differed statistically
in week 16 to 19, week 21, 22, 24, 25 and 33 (see Fig. 21). Thus, leaf development in sun
was faster than in shade.
Height was similar in sun and shade at the beginning in week 13, but differed statistically at
week 26 until the end of data taking in week 35 (see Fig. 22).
Thus, the whole performance of the young suckers in sun was faster in sun than in shade.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 46
Fig. 20: Weekly leaf area of pseudostems II in m² from week 13 to week 27 of
Pseudostems II out of non-destructive measurements; average
data, in both cases n=10; the red dots are marking the data
which are statistically different with p<5%
Fig. 21: Weekly measurements of Leaf Emission Rate (LER) of Pseudostems
II; averaged data; the red dots is marking the weeks of statistical significant
differences with p<5%

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 47
Fig. 22: Measurements of height (cm) from week 13 to week 34 of
Pseudostems II; average data, in both cases n=10; the red dots are marking
the statistically significant data with p<5%
Tab. 8: Folding of the leaf blades; measurements at the 2nd leaf of Pseudostems II at
the end of the dry season; ns = no statistical difference
Folding of the 2nd Leaf
7 AM; 28.4°C 180° to 136° 135° to 91° 90° to 46° <46°
Sun 10 0 0 0
Shade 10 0 0 0
12.30 PM; 33.8°C 180° to 136° 135 to 91° 90° to 46° <46°
Sun 4 4 2 0
Shade 9 0 1 0

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 48
Tab. 9: Leaf Angle of Pseudostem II; week 20; Leaf 1 to 5; ns = no significant
difference; n was differing as not all plants had five leaves at the time of
measurements
Leaf Angles Leaf 1 Leaf 2 Leaf 3 Leaf 4 Leaf 5
Sun 10° ± 7 15° ± 8 25° ± 11 25° ± 6 24° ± 7
Shade 13° ± 7 30° ± 9 20° ± 6 27° ± 3 25° ± 3
p ns p<5% ns ns ns
A difference in leaf angle and folding of leaf blades has an impact on k. Leaf 1 is a sink, but
leaf 2 to 5 have the highest photosynthesis rate (see also Table 1). But leaves folded more in
sun than in shade. However, there were no statistical significant differences of leaf angle
except of the second leaf (Table 8, Table 9).
4.3.4 Results Six Varieties
In the most shaded plot out of twelve measurements, radiation in height and above the banana
plants is 13% ± 4 standard deviation together with a canopy openness of 8% ± 2 standard
deviation. In contrast, radiation above banana plants in height of coffee is 25% ± 7 standard
deviation and a canopy openness of 18% ± 3 standard deviation (see Fig. 23). Absolut
numbers might differ because of the low canopy openness (FRAZER et al., 2001).
Nevertheless, there is significant difference of both light regimes with p<5%.
In full sun, highly shaded, medium shaded and light shaded plots, mean total transmitted light
is 88% ± 0.5, 37% ± 0.6, 18% ± 1.2, and 13% ± 2.2, respectively (value behnd ± is standard
error) (see also Fig. 24). Canopy openness in full sun, higly shaded, medium shaded and light
shaded plots is 66% ± 0.5, 8% ± 0.3, 11% ± 0.5, and 23% ± 1.2 (standard error), respectively.
The low value of canopy openness in the highly shaded and medium shaded plot might could
contribute that absolute numbers differ (FRAZER et al. 2001).

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 49
Fig. 23: Total transmitted light (%) in the most shaded plot; Coffee: light
above banana plants in height of the coffee plants; Banana: light above and
in height of the same banana plants; the whiskers show the standard error
Fig. 24: Raditation regime of the four plots above and in height of the banana
plants – full sun, light shaded, medium shaded and high shaded out of thirty
measurements each

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 50
Table 10: Growth of Height (cm), Circumference (cm), Length of Petiole (cm) and
Leaf Area (cm²) of six different varieties, Bluggoe (ABB), Baby (AA),
FHIA-18 (AAAA), Plantain (AAB), Gros Michel (AAA) and Manzano
(AAB) in four different light conditions; the data represents ratio of
growth of two measurements at week 34 and week 37; radiation is
defined as total transmitted light above the plant; nd = no data available
Total Trans.
Light Unit Bluggoe Baby FHIA-18 Plantain
Gros
Michel Manzano
88% Height+ 10.8 nd 12.4 6.2 5.4 7
Circumference+ 8.7 nd
2.3 1.6 2.5 2
Leaves+ 0.8 nd
2.2 2 2.4 2
Petiole+ 1.60 nd
0.68 0.77 1.96 0.92
Leaf Area+ 198.48 nd
971.92 478.64 710.4 807.52
37% Height+ 17.25 7.2 20.4 20 23.8 21.4
Circumference+ 5.75 1.8 2 2.7 2.5 2.8
Leaves+ 1.5 2 2.4 2.2 2.4 2.4
Petiole+ 1.28 0.37 1.10 1.84 2.11 2.19
Leaf Area+ 1041.8 541.52 1495.2 1112 1411.2 1033.44
18% Height+ 9.4 13.8 9.2 10.4 22.6 13
Circumference+ 6.7 1.4 1.1 1 1 1.2
Leaves+ 0 1.2 1.6 2 1.6 1
Petiole+ 0 1.42 1.11 1.49 1.64 1.07
Leaf Area+ 0 568.88 1129.28 1301.28 1142.32 600.72
13% Height+ 8.2 6.8 8 15.2 25.4 15
Circumference+ 4 0.7 0.8 0.6 1.14 0.7
Leaves+ 0 0.8 1.6 1.4 1.6 1.2
Petiole+ 0 0.73 0.93 1.20 1.67 1.15
Leaf Area+ 0 370.64 1208 1064.56 1569.44 766.08

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 51
Fig. 25: Average growth of height of six banana varieties (five individuals
each) in three different light environments within three weeks; radiation is
total transmitted light (%)
Fig. 26: Average growth of leaf area of six banana varieties (five individuals
each) in three different light environments within three weeks; radiation is
total transmitted light (%)

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 52
Table 10 shows banana growth data of six varieties in four different light regimes. The variety
‘Baby’ got sunburned in 88% and 35% total transmitted light. The variety ‘Bluggoe’ was
planted as corms. Fig. 24 and Fig. 25 show growth of leaf area and height in 35%, 18% and
13% total transmitted light, respectively. Only the variety ‘Gros Michel’ performed well in
height growth and leaf area growth in all three light environments. Plantain and ‘FHIA-18’
show have similar growth of leaf area. As ‘Baby’ got sunburned the data are not predictable.
‘Bluggoe’ had only in 35% light functional leaves and thus, countable leaf area. However,
three weeks of measurements cannot show reliable data of banana performance.
4.3.5 Light Measurements in Coffee Agroforestry Systems
All six plots have a canopy openness >10%. Direct sunbeam led to overestimation of total
transmitted light. It was visually observed and by working with the hemispherical photos, that
tree density and the surrounded area had an influence on total transmitted light. For light
measurements the treatments are too small. Fig. 27 show the averaged total transmitted light
of all six plots, out of around 100 measurements for each plot. Of all six plots, the single Poró
plot has the best light environment with nearly 60% light. This is not surprising as Poró was
cut before taking hemispherical photographies. In addition, the Poró plot has the lowest tree
density with 12m x 8m.
Fig. 28 to Fig. 33 show light dispersal maps of the six coffee-agroforestry systems. The maps
are not scaled, but the tree distance gives an idea of the original length and width of the plot.
In addition, the direction to north gives the way of the sun. The colours in the dispersal maps
are not standardized. It shows that missing trees in the plots have influence on light
transmitted through the canopy. The most shaded plot is the Cashá – Roble Coral agroforestry
system where tree density is highest.
If and in which severity tree habitus, leaf structure, tree mixing, and planting pattern in
relation to north have influence on light environment can just be assumed. However, in some
places were trees are missing there is no difference in light regime, which could be the
influence of the surrounded area or the way of the sun.

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 53
Comparing light dispersal maps of the Poró-plot and the Cashá – Roble Coral-plot (Fig. 30
and Fig. 31) with both an almost regular planting pattern of 12m x 8m and 6m x 4m,
respectively, that Poró has a less regular dispersed light environment.
Fig. 27: Average total transmitted light of all coffee agroforestry systems;
the whiskers show the standard error

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 54
Fig. 28: Light dispersal map of Coffee – Cashá agroforestry system

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 55
Fig. 29: Light dispersal map of Coffee – Roble Coral agroforestry system

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 56
Fig. 30: Light dispersal map of Coffee – Poró agroforestry system

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 57
Fig. 31: Light dispersal map of Coffee – Roble Coral - Cashá agroforestry
system

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 58
Fig. 32: Light dispersal map of Coffee – Poró - Cashá agroforestry
system

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 59
Fig. 33: Light dispersal map of Coffee – Poró – Roble Coral agroforestry
system

Christian Dold Musa in Shaded Coffee - A Case Study of Costa Rica
Seite 60
4.3.6 Interviews
The typical coffee agroforestry system of the ten visited farms with shaded banana was a
Coffee – Poró – Banana/Plantain – Laurel agroforestry system. In addition, some other timber
and fruit trees like Guava (Inga sp.), Cedro or cacao (Theobromoa cacao) are observed. The
first storey is the coffee, the second and third is either pruned Poró or banana (or fruit trees),
depending on time of pruning. The upper storey is set by Laurel or other timber trees. Only in
Talamanca it is preferred to use Poró unpruned for the upper storey. Field size was in average
3 ha with the smallest field 0.5 ha and the greatest of 9 ha.
The average plant density of the system was 4555 plants/ha (coffee), 162 plants/ha (Poró),
179 plants/ha (banana/plantain) and 37 plants/ha (Laurel). Coffee and Poró is uniform
planted, while Laurel has a more scattered planting pattern. Coffee has a plant density of 1.1m
x 1.25m up to 2m x 2m, and Poró differes between 4m x 5m up to 10m x 10m. The trees of
the upper storey have a minimum plant density of 10m x 10m. Banana and plantain was
planted from 2m x 2.5m up to 6m x 5m. In the case of banana the planting pattern differs;
some farmers grow banana in a uniform, some in a clustered planting pattern.
On each farm, two to four different banana varieties are found. The most used banana
varieties are ‘Gros Michel’ (30%) and ’Congo’ (23%), but also ‘Plantain’ (13%), ‘Gran
Enano’ (10%) and FHIA-varieties (7%) are mentioned.
19 measurements each, for good and for bad sites in relation to light environment for banana
on field are made with the densiometer. Canopy closure not differed with a mean of 13% ± 6
in good sites and 10% ± 5 (value is standard deviation) in bad sites. The reason is that
available light for banana varied greatly in field.

Christian Dold Discussion
Seite 61
5 Discussion
Different factors have to be included for improving banana production in shaded perennial
crops. Those are climate, disease pressure, and interaction of the species within an
agroforestry system, Musa cultivar and amount of solar radiation.
STOVER and SIMMONDS (1987 203) show that within the tropics high variation of solar
radiation is typical. There are differences between dry and wet season, and in duration of
overcast conditions. Some regions do not reach the amount of solar energy which would be
desireable for banana production.
In addition, CHAZDON et al. (1996 1) describes that understory plants in tropical forest are
typically growing in less than 1% of available radiation. Agroforestry systems might have a
better light regime, depending on management and species. But even in relatively large gaps
incoming radiation is lower than of full sun light (see Fig. 2).
Therefore, shaded banana in agroforestry systems would have a great disadvantage. CHAZDON
et al. (1996) say that at the same plant of the lower strata differences of light are high.
ROBINSON (1996 88) found that self-shaded leaves of banana have a lower photosynthetic rate
than exposed leaves due to lower transmitted PAR (see also Tab. 1). An overstorey tree
canopy might have the same impact on banana.
However, there is not enough data about Musa in agroforestry systems although it is often
used by small-scale farmers (NORGROVE 1998). Only VICENTE-CHANDLER et al. (1966) used
an upper story of plants for shading (Inga sp.); the author also describes light regime as highly
variable but in average by 50% of available radiation.
Available data of Musa Cavendish subgroup AAA show that light saturation point is near full
sun conditions (see Fig. 34). TURNER (1998b) describes Musa as light sensitive. Still enough
data of light response of different subgroups and cultivars are missing; light response curve of
Bluggoe (ABB) is similar of those of the Cavendish subgroup; but the cultivar AAA ‘Gros
Michel’ seems to be less susceptible to reduced radiation (see Fig. 7).

Christian Dold Discussion
Seite 62
In this study, young suckers of ‘Gros Michel’ performed best out of six cultivars in three
different shade level. However, just two measurements in-between three weeks cannot predict
shade responsibility of a cultivar. In a shaded agroforestry system acosiated with cacao,
banana shows better growth in high radiation (Category Pseudostems II). Although the young
suckers were in the same height leaf area was significantly higher in increased radiation with
more evolved leaves, same weekly leaf number and LER. Hence, growth was faster.
This gives another view of the results of Category Pseudostem III; although leaf area was
higher in 71% of light there was no significant difference. Weekly leaf number in shade was
higher but total amount of evolved leaves was higher in sun. This shows that in sun leaf turn
over is faster and longevity of leaves in shade is higher. The longevity of the leaf seems to be
the major significant effect (TURNER 1998a).
There was also no significant difference of growth, circumference and LER although all data
were better in sun.
This could assume that banana in shade might overcome the negative impacts by time of plant
development; but I suggest that the assumption that pseudostems in sun and shade, which
have a similar plant height are also of a similar development stage, is wrong. Results of
Category II show this dramatically. Also all available data show that banana perform slower
in shade. In addition, three plants in sun started to flower which also indicates that plant
development is faster.
Nutrients and precipitation were similar; nitrogen measurements are missing but organic
matter was higher in shade than in sun. Assuming that intercompetition between the species
were similar too, light was the most limiting factor. Also pseudostem number not differed
significantly, but age of the plants was unknown.
Referring again to the light response curve of ‘Gros Michel’ in Fig. 7, I suggest that light
regime was to still too low in the shaded treatment; in fact any other assumption cannot be
predicted.

Christian Dold Discussion
Seite 63
Fig. 34: PAR (summer; subtropical climate) and rate of photosynthesis in
Musa AAA Cavendish subgroup; light response curve
Reference: Data from Robinson (1996 87)
Looking at all available results describing a better banana production in shade, we see that in
every case there was a special situation. In the work of ISRAELI et al. (2002) water was more
limiting than light, VICENTE-CHANDLER et al. (1966) describes the lower impact of Yellow
Sigatoka as the reason for good yield; the results of MURRAY (1961) lacks vertification, and
no reason for better banana performance in the work of TORQUEBIAU and AKYEAMPONG
(1994) was found. Furthermore none work exceeded ratoon crop phase; this is especially
important as other papers describing the negative impact of shade on banana is increasing in
the ratoon crops (ISRAELI et al. 1995, STOVER and SIMMONDS 1987 205). In addition, it would
have been desireable to describe yield in units per time, as harvest time is delayed in shade.
Therefore, banana production in some amount of shade could only be recommended in special
cases. When water or nutrients are more limiting than radiation it seems that reduced light of
50% to 80% of full sun might have positive effects (ISRAELI et al. 2002, MURRAY 1961,
TORQUEBIAU and AKYEAMPONG, 1994). In addition, fertilization of nitrogen has a direct
impact on photosynthesis rate (KÖRNER 2002 911).

Christian Dold Discussion
Seite 64
In addition, disease pressure of Yellow Sigatoka (VICENTE-CHANDLER et al. 1966) and Black
Sigatoka is decreased in shade. Results of Category Pseudostem II and Category Pseudostem
III show much evident data: there is a significant difference in disease severity between
banana in sun and shade. Disease development time is faster in sun and significant more
leaves died due to Black Sigatoka. In addition, it was observed that fruit-bearing banana in
sun lost all leaves due to Black Sigatoka. Although no statistical data are available this
assesses to the observations of VICENTE-CHANDLER et al. (1966). In the case of this study it
seems that banana in sun has overcome the negative disease effect due to a fast leaf turnover.
But in the case of VICENTE-CHANDLER et al. (1966) yield was higher, as the impact of Yellow
Sigatoka disease was higher than the impact of reduced radiation.
This is especially interesting for small scale farmers, who are more likely to produce banana
in agroforestry systems, fertilizer or pesticides are expensive or not available. Besides, in
agroforestry systems the competition of water, nutrients and light is high. Hence, an
agroforestry system with sufficient light (reduced light competition) for banana could be a
possible cultivation method, when other factors are more limiting, and a longer development
period is less important.
Furthermore, the impact of leaf tearing was significantly higher at plants in sun than those in
shade. However, the severity of leaf tearing could not be evaluated and can just be
assumed.Torn leaves have a reduced photosynthesis (see Table 1). Windbreaks are
recommended when leaves are torn in strips narrower than 50 mm (ROBINSON 1996 92). But
ROBINSON (1996 92) is also aware of the negative impact of shade referring to ISRAELI et al.
(1995). Therefore, it needs to be evaluated if reduced leaf tearing on windy sites due to
agroforestry systems might have a positive effect. In this study it seems that the plants in sun
have overcome the negative impact of torn leaves.
Also, the analytical approach using equation (7) is of importance. TURNER (1998a) computed
that half of yield could contribute on intercepted radiation. Besides, TURNER (1998b) found
that k is changing within a banana canopy. TURNER and THOMAS (1995) found that leaves

Christian Dold Discussion
Seite 65
fold less in shade and in irrigated plants. Water stress changes k due to folding which has an
impact on intercepted radiation. In this study folding of banana leaves was higher in sun than
in shaded plants.
k is also depending on the amount of diffuse radiation (KÖRNER 2002 910). TURNER (1998a)
show a higher k of banana in shade where diffuse radiation is higher.
In addition, leaf angle has an impact on k as a more horizontal leaf can intercepted more light.
Here, no significant difference of leaf angle despite of leaf 2 was found. However, all
measurements were just made once; a broader analysis would have shown more reliable data.
In addition, phyllotaxy and leaf slope have an impact on k, which needs to be considered.
Also L is one important factor in equation (7). In this study no measurements of L were done,
but leaf area of plants in sun in both categories where higher. TURNER (1998a) shows a higher
L in full sun than in 30% light. ISRAELI et al. (1995) found no difference in leaf area until 30%
of light. In this study leaf area was lower in 32% light, but light environment might differ. A
value of L higher than 4.5 to 5 not contributes to higher yield (TURNER 1998a).
The amount of incident light in agroforestry systems is depending on different factors (see
Chapter 2) which makes it difficult to estimate light. In banana monoculture STOVER (1984)
defined plant density with L and PAR transmitted to the ground rather than plant density. This
could be also a good possibility for banana production in agroforestry systems. With equation
4 of Beer’s Law it is possible to estimate at which total L photo flux density is below light
compensation point (by using light saturation curves; see also Fig. 3) (KÖRNER 2002 910).
STOVER (1984) found in Honduras that in at least five year old plantations a leaf area index
between four and five was best for banana production together with a transmitted PAR to the
ground of 14% to 18%.
In an agroforestry system L and PAR measurements could give reliable data at what amount
of shade banana production is rapidly decreasing. Measurements with hemispherical camera
above banana canopy at harvest could give L of the upper story. After harvest it is easy to
compute L of the banana. As canopy openness might be too low below the mother plant, light
sensors could give PAR at ground and sucker level. Then also k can be evaluated using
equation (4) of JIMENEZ and LHOMME (1994).

Christian Dold Discussion
Seite 66
In addition, management of the agroforestry system like pruning the upper story and plant
pattern might help increasing light regime as direct sunbeam contribute most to total daily
PFD.
In this study, a coffee-Poró agroforestry system with tree density of 12m x 8m in transplanted
planting pattern and with rows in North-South direction shows mean total transmitted light of
60% after pruning and treated with two major branches (see Fig. 30). This is the amount of
around 100 trees/ha. All other plots have a light environment less than 60% with a tree
density of 6m x 4m, or have an unequal light dispersion (see Fig. 28, 29, 31 to 33).
Comparing the light data with previous studies of shaded banana, it seems that only the Poró
plot could have just enough radiation for successful banana growing without some
disadvantages of reduced radiation.
However, because of direct sunbeam, some overestimation ocurre; hence total light is lower
than estimated. In addition, light measurements were taken after pruning; therefore, radiation
was hightest at data taking and will be reduced when Poró starts to grow again. Thus, the
author suggests that a tree density lower than 100 Poró/ha could give sufficient light for
banana production in agroforestry systems.
Furthermore, light regime changed, when trees are missing in the regular pattern. Banana
could be planted in places where tree losses ocurre, for example when trees are cut for timber.
This assesses with experiences from Nicaragua (SCHIBLI 2001). However, the light dispersal
maps show that missing trees not always bring more light inside the plot. If this is due to
planting pattern to North direction, canopy and leaf structure of the trees or the surrounded
area, cannot be predicted. Thus, in general the author suggests that light measurements need
to be done more accurate in bigger plots or with evaluating the impact of the surrounding area
together with the planting pattern and additional measurements of canopy diameters.

Christian Dold Discussion
Seite 67
Considering the interviews, the results are not representative. In literature, a coffee (or cacao)-
Poró-banana – Cedro agroforestry system is described for Centralamerican and Carribean
farms. Poró is pruned twice a year in a height of 4m to 6m. Tree density of Poró is 250
trees/ha (SCHÜTT and LANG 1994 307). Another study of Costa Rica shows that farm size
differs between 4.8 ha to 17.5 ha. Coffee density varies from 3355 plants/ha to 6172
plants/ha. Overall tree density is 130 to 208 trees/ha and banana density is 149 up to 343
plants/ha (ESPINOZA 1985 found in KAPP 1998 127). This would partly assess to the
information of the interviews, and is a much higher tree density than the results of light
measurements in coffee – agroforestry system let suggest. Also ESPINOZA (1985) recommends
lower densities of all upper storeys of maximum 300 – 340 plants/ha, where bananas should
not exceed the amount of 120 plants/ha (because of negative impact on coffee production).
However, the author suggests that more interviews have to be made about banana production
in shade to confirm the presented interviews. A randomized interviewing has to be established
and a greater number of participants are needed. The idea of making densiometer light
measurements on subjective observations of light seems not to give reliable data on banana in
shade in field. Thus, a more structured measurement of light in a representative part of the
field should be preferred.

Christian Dold Conclusion
Seite 68
6 Conclusion
The improvement of banana and plantain in shade is one major task which needs to be more
understood, for in the tropical regions it is common to cultivate banana and plantain in
agroforestrial systems as a stable crop.
In agroforestry systems, typically banana has to face a light environment with high
competition. Banana and plantain is considered to be a light sensitive plant. The amount of
direct radiation, which is dramatically reduced below a tree canopy, is mostly contributing to
total photon flux density (PFD). However, light response of some Musa genetical subgroups
and cultivars is still unknown.
It is common sense that yields are delayed due to reduced radiation. In Costa Rica Musa ’Gros
Michel’ in sun was significantly faster in plant development than in shade. However, some
studies show that yield is increased in some amount of shade. But in every study some special
circumstances have to be considered. Besides, only one crop cycle was observed.
Therefore, banana in shade seems only to perform good when other factors like water,
nutrients or disease pressure of Sigatoka are more limiting than light. In Costa Rica, disease
development of Black Sigatoka was significantly delayed with less infected leaves.
If a reduced photosynthesis rate due to decreased leaf tearing or less leaf folding in shade can
overcome reduced light is questionable. It was observed that there are more untorn leaves and
less leaf folding due to the wind protection and shade of an upper story, although plant
development in sun was faster.
The analytical approach why in some cases banana performs better in shade show that three
factors, leaf area index (L), extinction coefficient (k) and radiation use efficiency (γ) are
mostly contributing to yield considering radiation. γ is also depending on water, nutrients and
disease pressure, L does not differ until highly reduced radiation, and k has a higher value in
reduced radiation.
Considering the practical improvement of banana in shade, the idea of evaluating plant
density of banana monoculture with data on leaf area index and photosynthetic active
radiation (PAR) could also be used in agroforestry systems.

Christian Dold References
Seite 69
7 References
AZAM -ALI , S.N. and SQUIRE , G.R. (2002): Principles of Tropical Agronomy; ISBN
085199136X; p. 236; CABI Publishing Wallingford UK
ALVAREZ , C. E., ORTEGA , A., FERNÁNDEZ, M., ANTONIO , A., BORGES. A. (2001): Growth,
yield and leaf nutrient content of organically grown banana plants in the Canary islands,
Fruits, vol. 56, p. 17–26
BRUN, J. (1963): La cercosporiose du bananier en Guineé, Étude de la phase ascosporée du
Mycosphaerella Leach, Thése doctoral ; Fac. Scl. Univ. Paris Sud/Orsay
CARLIER J., D. DE WAELE AND J.V. ESCALANT (2002): Global evaluation of Musa
germplasm for resistance to Fusarium wilt, Mycosphaerella leaf spot diseases and nematodes,
INIBAP Technical Guidelines 6, The International Network for the Improvement of Banana
and Plantain, Montpellier, France, INIBAP ISBN: 2-910810-52-6
CHAZDON , R.L., PEARCY , R.W., LEE, D.W., FETCHER , N. (1996): Photosynthetic Responses
of Tropical Forest Plants to Contrasting Light Environments; in: Chazdon, R.L., Mulkey, S.S.,
Smith, A.P. (Ed.) (1996): Tropical Forest Plant Ecophysiology; Chapter 1; ISBN 0-412-
03571-5675 p. Chapman & Hall, London, England
ECKSTEIN , K. and ROBINSON, J.C. (1994): Leaf tearing influences physiological efficiency
and yield of bananas Instituut vir Tropiese en Subtropiese Gewasse, Privaatsak X11208,
Nelspruit 1200, South Africa, Inligtingsbulletin - Instituut vir Tropiese en Subtropiese
Gewasse, 1994, No. 266, pp. 8-9, 1 pl.

Christian Dold References
Seite 70
ECKSTEIN , K. and ROBINSON, J.C. (1999): The influence of the mother plant on sucker
growth, development and photosynthesis in banana (Musa AAA; Dwarf Cavendish); Journal
of Horticultural Science & Biotechnology (1999) 74 (3), pp. 347 - 350
ECKSTEIN , K., ROBINSON, K., FRASER, C. (1997): Physiological responses of banana (Musa
AAA; Cavendish sub-group) in the subtropics. VII Effects of windbreak shading on
phenology, physiology and yield, Journal of Horticultural Science 72, pp. 389-396
ESPINOZA, L. (1985): Untersuchungen über die Bedeutung der Baumkomponente bei
agroforstlichen Kaffeeanbau an Beispielen aus Costa Rica. Göttinger Beiträge zur Land- und
Forstwirtschaft in den Tropen und Subtropen, Heft 10. Institut für Waldbau der Universität
Göttingen, Göttingen, p. 164
FRAZER , G.W., CANHAM , C.D., LERTZMAN , K.P. (1999): Gap Light Analyzer (GLA),
Version 2.0: Imaging software to extract canopy structure and gap light transmission indices
from true-colour fisheye photographs, users manual and program documentation. Copyright
© 1999: Simon Fraser University, Burnaby, British Columbia, and the Institute of Ecosystem
Studies, Millbrook, New York.
FRAZER , G.W.; FOURNIER , R. A.; TROFYMOW , J.A.; HALL , R.J (2001): A comparison of
digital and film fisheye photography for analysis of forest canopy structure and gap light
transmission, Agricultural and Forest Meteorology 109 (2001), pp. 249 – 263, Elsevier
Science B.V.
GAUHL , F. (1989): Untersuchungen zur Epidemiologie und Ökologie der Schwarzen Sigatoka
Krankheit (Mycosphaerella fijiensis Morelet) an Kochbananen (Musa sp.) in Costa Rica.
Thesis. Univ. Gottingen (West Germany), 128p

Christian Dold References
Seite 71
ISRAELI , Y., PLAUT , Z., SCHWARTZ , A. (1995): Effect of shade on banana morphology,
growth and production, Scientia Horticulturae 62 (1995) 45-46, pp.45 - 56
ISRAELI , Y., ZOHAR , C., ARZI , A., NAMERI , N., SHAPIRA , O., LEVI , Y. (2002): Growing
banana Ander shade screens as a mean of saving irrigation water: preliminary results, Acorbat
Memorias XV. Realizada en Cartagena de Indias, Colombia, 27 de octubre al 02 noviembre
2002. Medellín (COL): Asociación de Bananeros de Colombia AUGURA, 2002, pp. 384 -
389
JIMENEZ O.F. AND LHOMME , J.P. (1994): Rainfall interception and Radiation Regime in a
Plantain Canopy; Fruits Vol. 49 (2), pp. 133 - 138
KAPP, G. (1998): Bäuerliche Forst- und Agroforstwirtschaft in Zentralamerika:
Untersuchungen über forstliche und agroforstiche Produktionssysteme unter besonderer
Berücksichtigung des feuchten Tieflands von Costa Rica und Panama, Freiburg, Habil. Schr.,
1998, ISBN 3-8236-1300-6, p. 303
KÖRNER (2002): Pflanzen im Lebensraum; in: Straßburger, Lehrbuch der Botanik für
Hochschulen; Chapter 13.; 1123 p.; Spektrum Akademischer Verlag Heidelberg, Berlin,
ISBN 3-8274-1010-X
LÓPEZ, A., OROZCO, L., SOMMARIBA , E., BONILLA , G. (2003): Tipologías y manejo de
fincas cafetaleras en los municipios de San Ramón y Matagalpa, Nicaragua, Agroforesteria en
las Americas, Vol. 10, No. 37-38, 2003
MARÍN VARGAS, D.; ROMERO CALDERÓN , R (1992): El combate de la Sigatoka negra,
Corbana, Departamento de Investigaciones, Boletin No. 4

Christian Dold References
Seite 72
MOGUEL , P., TOLEDO, V.M. (1999): Biodiversity Conservation in Traditional Coffee
Systems of Mexio, Conservation Biology, Volume 13, No. 1, Feb. 1999, pp. 11 -21
MOREAU, B. (1965): La Croissance et le Développement du Bananier 'Gros Michel' en
Èquateur, Fruits, Vol. 20, No. 5
MONTEITH , J.L. (1977) : Climate and the efficiency of crop production in Britain,
Philosophical Transactions of the Royal Society of London B 281: 277 – 294
MURRAY , D.B. (1960): The Effect of Deficiencies of the Major Nutrients on Growth and Leaf
Analysis of the Banana, Tropical Agriculture (Trinidad), Vol. 37, No. 2, pp. 98 – 107
NAKASONE , H.Y., and PAULL , R.E. (1998): Crop Production Science in Horticulture 7;
Tropical Fruits, CAB International, Biddles Ltd., Guildford and King´s Lynns, ISBN 0 85199
254 4, 445 p.
NORGROVE, L. (1998): Musa in multistrata systems: Focus on shade, Infomusa, The
International Magazine on Banana and Plantain, Vol. 7, No. 1, June 1998, Technical Center
for Agricultural and Rural Cooperation (CTA)
ROBINSON, J.C. (1996): Crop Production Science in Horticulture 5; Banana and Plantains,
248 p, Wallingford, U.K.: CAB International, ISBN13: 978-0-85198-985-3
ROBINSON, J.C. AND NEL , D.J. (1988): Plant density studies with banana (cv. Williams) in a
subtropical climate. I. Vegetative morphology, phenology and plantation microclimate,
Journal of Horticultural Science (1988) 63 (2), pp. 303-313

Christian Dold References
Seite 73
ROBINSON, J.C. AND NEL , D.J. (1989): Plant density studies with banana (cv. Williams) in a
subtropical climate. II. Compnents of yield and seasonal distribution of yield, Journal of
Horticultural Science 67, pp. 403-410
SCHIBLI , C. (2001): Percepciones de familias productoras de sus sistemas agroforestales con
café, en el norte de Nicaragua, Master-Thesis, CATIE-MIP AF (NORAD), FUMDEC, I.M.C
SCHÜTT and LANG (1994): IN SCHÜTT, WEISBERGER, SCHUCK, LANG, STIM , ROLOFF (1994):
Bäume der Tropen, ISBN 3-933203-79-1, p. 687
SOTO-PINTO , L., PERFECTO, I., CASTILLO -HERNANDEZ, J., CABALLERO -NIETO , J. (2000):
Shade effect on coffee production at the northern Tzeltal zone of the state of Chiapas, Mexico,
Agriculture, Ecosystems and Evironment 80 (2000) 61 - 69
SÚAREZ, A., SOMARRIBA , E. (2002): Aprovechamiento sostenible de madera de Cordia
alliodora de regeneración natural en cacaotales y bananales de indígenas de Talamanca, Costa
Rica, Agroforesteria en las Americas 2002, Vol. 9, No. 35-36
STAVER , C. (2007): personal communication about the region of Talamanca, Costa Rica in
May 2007
STOVER , R.H. (1979): Pseudostem Growth, Leaf Production and Flower Initiation in the
Grand Nain Banana, Tropical Agriculture Research Servicees (SIATSA) Bulletin No. 8, La
Lima, Cortés, Honduras, C.A., 37 pp.
STOVER , R.H. (1982): ‘Valery’ and ‘Grand Nain’: plant and foliage characteristics and a
proposed banana ideotype, Tropical Agriculture (Trinidad), Vol. 59, No.4, pp. 303 - 305

Christian Dold References
Seite 74
STOVER , R.H. (1984): Canopy Management in Valery and Grand Nain using leaf area index
and photosynthetically active radiation measurements; Fruits Vol. 39, No. 2, 1984, pp. 89 - 93
STOVER , R. H. AND SIMMONDS , N. W. (1987): Bananas, 3rd edition, Longman Scientific &
Technical, Harlow U.K. ISBN 0 582 46357 2
TURNER, D.W. (1998a): The impact of evironmental factors on the development and
productivity of bananas and plantains, Memorias XIII Reunion Acorbat (Association fpr
Cooperation in Banana Research in the Caribbean and Tropical America), Guayaquil –
Ecuador, Noviembre 23 – 27 de 1998, Ed.: Ing. Luis Hidalgo Arizaga, CONABAN
Corporación Nacional de Bananeros
TURNER, D.W. (1998b): Ecophysiolgy of Bananas: The Generation and Functioning of the
Leaf Canopy; Proc. Int. Symp. Banana in Subtropics, Ed.: V. Galán Sauco; Acta Horticulturae
490, ISHS 1998; pp. 211 – 221
TURNER, D.W. (1994): Daily variation in Banana Leaf Growth; Australian Journal of
Experimental Agriculture and Animal Husbandry 10, pp. 231 - 234
TURNER, D.W. (2003): An integral method for estimating total leaf area in bananas,
InfoMusa, The International Journal on Banana and Plantain, Vol. 12, No. 2
THOMAS , D.S. and TURNER, D.W. (2001): Banana (Musa sp.) leaf gas exchange and
chlorophyll fluorescence in response to soil drought, shading and lamina folding; Scientia
Horticulturae 90 (2001), Elsevier Science 2001, pp. 93 - 108
YÉPEZ PACHECO, C. (2001): Selección de Árboles para Sombra en Cafetales Diversificados
de Chiapas, Thesis Magister Scientiae, CATIE

Appendices

Christian Dold Appendices
Seite A 1
Appendix I
Tab. 11: Explanation of the five stages of leaf development
Reference: according to Brun (1963), quoted by Carlier et al., 2002
0 (A) 0.2 (B) 0.4 (C) 0.6 (D) 0.8 (E)
Fig. 35: Stations of leaf development
References: according to Brun (1963), found in Vargas et al. (1992), changed
Stage
A
The ‘cigar’, about 10 cm in length, is still joined to the preceding leaf
Stage
B
The ‘cigar’ is bigger, but has not yet reached its full length
Stage
C
The ‘cigar’ is completely free. It reaches its full length and the diameter of its
apex has considerably increased following the loosening of the spiral
Stage
D
The left-hand side has already unfolded, and spreading takes place at the
extreme apex
Stage
E
The upper part of the leaf has unfolded and the base is in an open cornet shape

Christian Dold Appendices
Seite A 2
Appendix II
Fig. 36: Gauhl’s modification of Stover’s severity scoring system; leaves with
infected parts over 50% of the leaf area (only necrosis) are seen as being
destroyed by Sigatoka Negra
References: Carlier et al., (2002)

Christian Dold Appendices
Seite A 3
Appendix III
Tab. 12: Soil samples of the botanic garden, ten of each; 0 – 30 cm
Soil Samples 0 – 30 cm Sun Shade
P cmol(+)/l 13 41
K cmol(+)/l 0.28 0.27
Mg cmol(+)/l 1.3 1.99
Fe mg/l 174 222
Organic matter (%) 5.25 6.56
pH 4.9 5.1

Christian Dold Appendices
Seite A 4
Appendix IV
Tab. 13: Soil samples of the site with the six varieties; 0 – 10 cm
Soil Samples 0 –10cm 13% Light 18% Light 35% Light 88% Light
pH 5.0 5.1 4.8 5.2
N (%) 0.33 0.38 0.35 0.22
P cmol(+)/l 60.5 42.4 33.7 4.5
K cmol(+)/l 0.65 0.66 0.63 0.12
Mg cmol(+)/l 1.24 1.2 1.21 1.53
Fe mg/l 257 241 252 176
Tab. 14: Soil samples of the site with the six varieties; 0 – 30 cm
Soil Samples 0-30cm 13% Light 18% Light 35% Light 88% Light
pH 4.5 4.7 4.6 5.1
N (%) 0.26 0.23 0.26 0.28
P cmol(+)/l 17.8 15.1 11.4 7.1
K cmol(+)/l 0.41 0.47 0.4 0.23
Mg cmol(+)/l 0.63 0.74 0.61 1.8
Fe mg/l 216 175 182 216
Tab. 15: Amount of dry matter of organic fertilizer used for planting in the plots with
the six varieties
Gallinaza dry matter (%) Poró dry matter (%)
82.23 ± 2.05 33.2 ± 3.67

Christian Dold Appendices
Seite A 5
Appendix V
Name
Field Size
Soil prosperities
Coffee:
Planting Distance Planting Pattern (scattered/uniform/clustered)
Storey No. Age
Cacao:
Planting Distance Planting Pattern (scattered/uniform/clustered)
Storey No. Age
Species_1:
Planting Distance Planting Pattern (scattered/uniform/clustered)
Storey No. Age
Species_2:
Planting Distance Planting Pattern (scattered/uniform/clustered)
Storey No. Age
Species_3:
Planting Distance Planting Pattern (scattered/uniform/clustered)
Storey No. Age
Musa:
Planting Distance Planting Pattern (scattered/uniform/clustered)
Storey No. Age:
Variety_1: Variety_2: Variety_3: