Embed Size (px)
Citation preview

BLOQUEIO DA MEIOSE COM BUTIROLACTONA I EM OVÓCITOS BOVINOS: EFEITOS SOBRE A MATURAÇÃO NUCLEAR E CITOPLASMÁTICA
PAULO ROBERTO ADONA
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO – UENF
CAMPOS DOS GOYTACAZES – RJ MARÇO – 2006

BLOQUEIO DA MEIOSE COM BUTIROLACTONA I EM OVÓCITOS BOVINOS: EFEITOS SOBRE A MATURAÇÃO NUCLEAR E CITOPLASMÁTICA
PAULO ROBERTO ADONA
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte das exigências para obtenção do título de Doutor em Produção Animal.
Orientadora: Profª. Drª. Cláudia Lima Verde Leal CAMPOS DOS GOYTACAZES – RJ
MARÇO – 2006

FICHA CATALOGRÁFICA
Preparada pela Biblioteca do CCTA / UENF 028/2006
Adona, Paulo Roberto
Bloqueio da meiose com butirolactona I em ovócitos bovinos : efeitos sobre a maturação nuclear e citoplasmática / Paulo Roberto Adona. – 2006. 78 f. il.
Orientador: Cláudia Lima Verde Leal Tese (Doutorado em Produção Animal) – Universidade Estadual do Norte Fluminense Darcy Ribeiro, Centro de Ciências e Tecnologias Agropecuárias. Campos dos Goytacazes, RJ, 2006. Bibliografia: f. 55-67.
1. Butirolactona I 2. Embrião 3. Maturação 4. Meiose 5. Ovócito bovino I. Universidade Estadual do Norte Fluminense Darcy Ribeiro. Centro de Ciências e Tecnologias Agropecuárias. II. Título.
CDD 636.2082

BLOQUEIO DA MEIOSE COM BUTIROLACTONA I EM OVÓCITOS BOVINOS: EFEITOS SOBRE A MATURAÇÃO NUCLEAR E CITOPLASMÁTICA
PAULO ROBERTO ADONA
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte das exigências para obtenção do título de Doutor em Produção Animal.
Aprovado em 10 de março de 2006. Comissão Examinadora: ___________________________________________________________________
Profª. Drª. Gisele Zoccal Mingoti (Doutorado em Fisiologia) UNESP ___________________________________________________________________
Profª. Drª. Maria Clara Caldas Bussiere (Doutorado em Fisiologia) UENF ___________________________________________________________________
Profº Dr. Reginaldo da Silva Fontes (Doutorado em Fisiologia) UENF ___________________________________________________________________ Profª. Drª. Cláudia Lima Verde Leal (Doutorado Reprodução Animal) USP/FZEA
(Orientador)

ii
AGRADECIMENTOS
Em especial à minha mãe, meu irmão e às minhas irmãs.
Gostaria de expressar minha profunda gratidão e carinho, em especial:
À Rosângela A. Queiroz;
À Cláudia L. V. Leal;
À Margot A. N. Dode;
À Maineide Z. Velasques.
À todos meus amigos pelo carinho e companheirismo. Sem cada um de vocês
nada disso seria possível e a vida não teria sentido.
Alexandre Barreto; Aline S. M. César; Amanda de A. Rocha; Aparecida Mandella; Bethania Lopes; Bruno Fagundes; Camila Cortez; Fabiana Bressan; Felipe C. Braga; Fernando Biaze; Flávio P. Júnior; Giovana K. F. Merighe; Gustavo (Sancho); Hebinho (Baby Sauro) Isabele P. Emanuelli; Kátia L. Schwartz; Kelen S. Viana; Lígia Garcia Mesquita; Marcos R. Chiaratti; Mariene (Bitoca); Moysés S. Miranda; Nilton P. dos Santos; Norma Clea Rodovalho; Patrícia M. Porciúncula; Paula Ripamonte; Pedro Ratto; Roberto Carneiro; Rosemary Bastos; Sílvia G. Matta; Sylvia S. Cortezzi; Tiago H. C. De Bem; Vanessa G. Ueno;

iii
Aos Professores: Maria Clara C. Bussiere, Flávio Vieira Meirelles, Reginaldo
Fontes, Gisele Zoccal Mingoti.
Àqueles que contribuíram direta ou indiretamente para a realização desse
trabalho.
À Universidade Estadual do Norte Fluminense Darcy R. (UENF/CCTA/LMGA);
À Universidade de São Paulo (USP/FZEA/ZAB);
À FAPESP, pelo financiamento do projeto de pesquisa;
À CAPES, pela concessão da bolsa de estudo.

iv
BIOGRAFIA
PAULO ROBERTO ADONA, filho de Ivo Adona e Maria Boniatti Adona,
nasceu em 26 de agosto de 1966, na cidade de Campinas do Sul – RS.
Em março de 1995, ingressou no curso de Biologia da Universidade Católica
Dom Bosco (UCDB), em Campo Grande – MS. Submeteu-se à apresentação de
monografia para conclusão do curso em dezembro de 1998.
Foi selecionado em março de 1997 como estagiário do setor de Reprodução
Animal da Embrapa – CNPGC, sob orientação da Pesquisadora Drª. Margot A. N.
Dode.
Foi aprovado em agosto de 2000 no curso de Pós-graduação em Produção
Animal, a nível de mestrado, da Universidade Estadual do Norte Fluminense
(UENF), Campos dos Goytacazes – RJ, submetendo-se à defesa de tese para a
conclusão do curso em abril de 2002.
Foi aprovado em março de 2002 no curso de Pós-graduação em Produção
Animal, a nível de doutorado, da Universidade Estadual do Norte Fluminense Darcy
Ribeiro (UENF), Campos dos Goytacazes – RJ, submetendo-se à defesa de tese
para a conclusão do curso em março de 2006.

v
CONTEÚDO LISTA DE ABREVIATURAS------------------------------------------------------------------- viii
RESUMO ------------------------------------------------------------------------------------------- x
ABSTRACT ---------------------------------------------------------------------------------------- xii
1. INTRODUÇÃO ----------------------------------------------------------------------------- 01
2. REVISÃO DE LITERATURA ----------------------------------------------------------- 03
2.1. Ovogênese e foliculogênese ----------------------------------------------------- 03
2.2. Maturação ovocitária --------------------------------------------------------------- 04
2.2.1. Maturação nuclear ----------------------------------------------------------- 05
2.2.2. Maturação citoplasmática ------------------------------------------------- 06
2.2.3. Organização estrutural na maturação ---------------------------------- 07
2.2.3.1. Citoesqueleto --------------------------------------------------------- 07
2.2.3.2. Organelas -------------------------------------------------------------- 08
2.2.3.2.1. Mitocôndrias ----------------------------------------------------- 08
2.2.3.2.2. Grânulos corticais --------------------------------------------- 09
2.2.4. Mudanças moleculares e bioquímicas --------------------------------- 10
2.2.4.1. Cinases dependentes de ciclinas (CDKs) --------------------- 11
2.2.4.2. Fator promotor da maturação (MPF) --------------------------- 12
2.2.4.3. Proteína cinase ativada por mitógenos (MAPK) ------------- 13
2.3. Competência ovocitária ------------------------------------------------------------ 15
2.4. Bloqueio meiótico-------------------------------------------------------------------- 17
3. OBJETIVOS -------------------------------------------------------------------------------- 20

vi
4. MATERIAL E MÉTODOS --------------------------------------------------------------- 21
4.1. Coleta de ovários e ovócitos ----------------------------------------------------- 21
4.2. Solução estoque do inibidor ------------------------------------------------------ 22
4.3. Bloqueio da meiose com butirolactona I --------------------------------------- 22
4.4. Determinação do estádio da meiose ------------------------------------------- 22
4.5. Maturação in vitro ------------------------------------------------------------------- 23
4.6. Microtúbulos -------------------------------------------------------------------------- 24
4.7. Microfilamentos ---------------------------------------------------------------------- 24
4.8. Grânulos corticais ------------------------------------------------------------------- 25
4.9. Mitocôndrias -------------------------------------------------------------------------- 25
4.10. Fecundação e cultivo in vitro --------------------------------------------------- 25
4.11. Delineamento experimental ----------------------------------------------------- 27
4.11.1. Experimento 1. Avaliação de concentrações decrescentes de
BL I em meio suplementado com BSA -----------------------------------------------------
27
4.11.2. Experimento 2. Avaliação de concentrações crescentes de BL
I em meio sem suplementação de BSA ----------------------------------------------------
27
4.11.3. Experimento 3. Efeito do bloqueio da meiose na maturação
nuclear ----------------------------------------------------------------------------------------------
28
4.11.4. Experimento 4. Cinética da maturação nuclear pós-bloqueio
da meiose ------------------------------------------------------------------------------------------
29
4.11.5. Experimento 5. Efeito do bloqueio da meiose na organização
dos microtúbulos e dos microfilamentos ---------------------------------------------------
29
4.11.6. Experimento 6. Efeito do bloqueio da meiose na distribuição
dos grânulos corticais e das mitocôndrias -------------------------------------------------
30
4.11.7. Experimento 7. Efeito da BL I no desenvolvimento
embrionário in vitro -------------------------------------------------------------------------------
31
4.12. Análise estatística ----------------------------------------------------------------- 32
5. RESULTADOS ----------------------------------------------------------------------------- 33
5.1. Experimento 1. ----------------------------------------------------------------------- 33
5.2. Experimento 2. ----------------------------------------------------------------------- 34
5.3. Experimento 3. ----------------------------------------------------------------------- 35
5.4. Experimento 4. ----------------------------------------------------------------------- 36
5.5. Experimento 5. ----------------------------------------------------------------------- 38

vii
5.7. Experimento 6. ----------------------------------------------------------------------- 40
5.9. Experimento 7. ----------------------------------------------------------------------- 43
6. DISCUSSÃO ------------------------------------------------------------------------------- 45
7. CONCLUSÕES ---------------------------------------------------------------------------- 54
8. REFERÊNCIAS BIBLIOGRÁFICAS ------------------------------------------------- 55
APÊNDICE ----------------------------------------------------------------------------------------- 68

viii
LISTA DE ABREVIATURAS μg – micrograma;
μL – microlitro;
μM – micromolar;
AI – anáfase I;
AMPc – monofosfato de adenosina cíclica;
ATP – trifosfato de adenosina;
B199 – TCM-199, sais de Earle e 20 mM de bicarbonato de sódio;
BL I – butirolactona I;
BSA – albumina sérica bovina;
CDK – cinase dependente de ciclina;
CIV – cultivo in vitro;
CO2 – dióxido de carbono;
CSF – fator citostático;
DMSO – dimetilsulfóxido;
DNA – ácido desoxirribonucléico;
ERK – cinase regulada por sinal extracelular (MAPK);
FITC – isotiocianato de fluoresceína;
FIV – fecundação in vitro;
FSH – hormônio folículo estimulante;

ix
GDP – guanosina difosfato;
GTP – guanosina trifosfato;
H199 – TCM -199, sais de Earle, 20 mM de bicarbonato de sódio e 25 mM de Hepes;
LH – hormônio luteinizante;
MAPK – proteína cinase ativada por mitógenos;
MAP2K, MAP3K, MAP4K – cinases ativadoras de cinases ativadas por mitógenos
MAT – metáfase I, anáfase I e telófase I;
MBP – proteína básica de mielina;
MEK – cinase regulada por sinal extracelular (MAP2K);
MI – metáfase I;
MII – metáfase II;
Miss – MAPK de interação e estabilização do fuso meiótico;
MIV – maturação in vitro;
mL – mililitro;
mos – oncogene c-mos;
MPF – fator promotor da maturação;
Myt1, Wee1 e CDC25 – proteínas cinases envolvidas na atividade do MPF;
P90RSK – proteína cinase ribossomo S6;
PBS – tampão fosfato salina;
PIV – produção in vitro;
PKA – proteína cinase A;
PKC – proteína cinase C;
PVA – álcool polivinílico;
Raf-1 – MAP3K;
Ras – MAP4K;
RNAm – ácido ribonucléico mensageiro;
SFB – soro fetal bovino;
SOF – fluido de oviduto sintético;
TALP – meio tyrode’s com albumina, lactato e piruvato;
TCM-199 – meio de cultura de tecidos 199;
TI – telófase I;
VG – vesícula germinativa.

x
RESUMO
ADONA, Paulo Roberto, Dr. S., Universidade Estadual do Norte Fluminense Darcy
Ribeiro: março de 2006; Bloqueio da meiose com butirolactona I em ovócitos
bovinos: efeitos sobre a maturação nuclear e citoplasmática; Orientador: Profª. Drª.
Cláudia Lima Verde Leal. Conselheiro: Profª. Drª. Maria Clara Caldas Bussiere.
O presente estudo teve por objetivo avaliar o efeito do uso da butirolactona I
(BL I) na pré-maturação sobre a progressão da meiose, estruturas celulares e
desenvolvimento de ovócitos bovinos cultivados in vitro. Ovários bovinos foram
coletados em frigoríficos e os folículos de 2-6 mm de diâmetro foram aspirados para
a obtenção dos ovócitos. Inicialmente, foram avaliadas diferentes concentrações de
BL I em meio suplementado ou não com BSA. Houve diferença (P < 0,05) nas taxas
de vesícula germinativa (VG) entre os grupos C/24 (0,0%), 25 (65,1%), 50 (84,4%) e
100 μM (97,4%) de BL I suplementados com BSA, mas não houve diferença (P >
0,05) entre os tratados com 10 (94,7%), 15 (97,2%), 20 (98,7%) e 25 μM (98,7%) de
BL I em meio sem BSA. As taxas de maturação (MII) não diferiram (P > 0,05) entre o
controle (91,1%) e os tratados com 10 (B10 = 91,5%) e 100 μM (B100 = 92,4%) de
BL I. Em relação à cinética de MIV pós-bloqueio, observou-se que às 6h de MIV os
grupos B10 (71,4%) e B100 (74,3%) apresentaram taxas de VG inferiores (P < 0,05)
ao controle (97,3%). Com 12h de MIV a maior parte dos ovócitos estava em estádios
intermediários da meiose (MAT) (77,9; 83,1 e 86,4% para controle, B10 e B100,
respectivamente, P > 0,05). Com 18h de MIV as taxas de MII não diferiram (P >

xi
0,05) entre controle (72,0%), B10 (73,7%) e B100 (78,9%). Com relação aos
microtúbulos não foram observadas alterações no fuso meiótico quanto à
organização e formação da placa metafásica nos diferentes grupos de ovócitos
(controle, B10 e B100) e horários (0, 6, 12, 18 e 24h) avaliados. Também não foi
observada nenhuma alteração no arranjo dos microfilamentos no citoplasma dos
ovócitos avaliados nas mesmas condições. Em 100% dos ovócitos não maturados
independente do grupo (C/0h, B10 e B100), os grânulos corticais apresentavam-se
dispersos pelo citoplasma (P > 0,05) dos ovócitos. Após 24h de MIV, 98% dos
ovócitos apresentavam migração dos grânulos corticais para a região periférica do
citoplasma em todos os grupos (P > 0,05). As mitocôndrias nos ovócitos não
maturados dos grupos tratados encontravam-se em sua maioria na periferia dos
ovócitos (81,5 e 86,9%, P > 0,05), mas foi inferior (P < 0,05) ao controle (100%).
Após 24h de MIV, as mitocôndrias migraram por todo o ovócito, sendo o grupo C/24
(81,5%) inferior (P < 0,05) aos grupos B10M (95,2) e B100M (98,2%) que não
diferiram entre si (P > 0,05). As taxas de clivagem (81-87%) não diferiram (P > 0,05)
para os ovócitos submetidos à FIV nos grupos controle, B10 e B100. Com relação à
percentagem de blastocisto no D7, os grupos controle (38,3%) e B10 (41,6%) não
diferiram entre si (P > 0,05), mas ambos foram superiores (P < 0,05) ao grupo B100
(33,0%). A taxa de blastocisto eclodido foi similar (P > 0,05) entre os grupos controle
(19,2%), B10 (17,7%) e B100 (11,0%) no D8. Também não foi observada variação
(P > 0,05) no número médio de células dos blastocistos no D8, entre o controle
(138), B10 (136) e B100 (150). A pré-maturação com BL I não aumentou os índices
de desenvolvimento, porém também não induziu alterações estruturais e não
comprometeu (B10) o desenvolvimento embrionário, sugerindo a possibilidade de
sua aplicação em biotécnicas de PIV de embriões e de clonagem.
Palavras-chave: butirolactona I, embrião, maturação, meiose e ovócito bovino.

xii
ABSTRACT
ADONA, Paulo Roberto, Dr. S., Universidade Estadual do Norte Fluminense Darcy
Ribeiro: março de 2006; Meiosis block using butyrolactone I in bovine oocytes:
Effects on nuclear and cytoplasmic maturation; Supervisor: Prof. Dr. Cláudia Lima
Verde Leal. Counselor: Prof. Dr. Maria Clara Caldas Bussiere.
The present study aimed to assess the effects of butyrolactone I (BL I) in pre-
maturation on meiosis progression, cellular structures and development of bovine
oocytes cultured in vitro. Bovine ovaries were collected in abattoirs and 2-6mm
follicles were aspirated to obtain oocytes. Initially, different BL I concentrations were
evaluated in B199 medium supplemented or not with BSA. Germinal vesicle (GV)
rates were different (P < 0.05) among the groups C/24 (0.0%) 25 (65.1%), 50
(84.4%) and 100 μM (97.4%) BL I in medium supplemented with BSA. However, no
difference (P>0.05) was observed when oocytes were cultured with 10, 15, 20 and 25
μM BL I (94.7, 97.2, 98.7 and 98.7%, respectively) in medium without BSA.
Maturation rates (MII) were similar (P > 0.05) between controls (91.1%) and treated
oocytes (91.5 and 92.4% for B10 and B100, respectively). Regarding maturation
kinetics, VG rates at 6h IVM in controls (97.3%) were superior (P < 0.05) to treated
oocytes (71.4 and 74.3% for B10 and B100, respectively). At 12h most of the oocytes
were at intermediate stages of meiosis (77.9, 83.1 and 86.4%, for control, B10 and
B100, respectively, P>0.05). After 18 h IVM, MII rates were similar (P > 0.05) among
groups (72.0, 73.7 and 78,9% for control, B10 and B100, respectively). Regarding

xiii
microtubules no alterations in meiotic spindle were observed for organization and
formation of metaphase plate in different groups (control, B10 and B100) and IVM
periods (0, 6, 12, 18 and 24h). No alterations in microfilament arrangement were
observed either under the same conditions. All immature oocytes (100%) presented
cortical granules (CG) dispersed throughout the cytoplasm, irrespective of oocyte
group (C/0h, B10 and B100, P > 0.05). After 24 h IVM, also irrespective of oocyte
group, 98% of the oocytes had the CG migrated to the periphery of the cytoplasm (P
> 0.05). Immature oocytes in treated groups presented mitochondria mostly in the
periphery of the oocytes (81.5 and 86.9%, P > 0,05), although at a lower rate than in
control oocytes (100%, P < 0.05). After 24 h IVM, mitochondria migrated throughout
the cytoplasm, but in both treated groups the migration rates were superior (95.2 and
98.2% for B10 and B100, P > 0.05) to controls (81.5%, P < 0.05). Cleavage rates
were not affected (81-87%, P > 0.05) in oocytes fertilized in vitro. However,
blastocyst rates in day 7 were reduced in the B100 group (33.0%, P < 0.05). Controls
and B10 had similar rates (38.3 and 41.6%, respectively, P >0.05). Day 8 hatching
rates (19.2, 17.7 and 11.0% for control, B10 and B100, P > 0.05) and blastocyst cell
numbers (138, 136 and 150 B100 for control, B10 and B100, P > 0.05) were
unaffected. Pre-maturation using BL I did not increase blastocyst rates, but also did
not affect cell structures and development (B10), suggesting the possibility of its use
for in vitro embryo production and cloning.
Palavras-chave: butyrolactone I, embryo, maturation, meiosis and oocyte bovine.

1. INTRODUÇÃO
Os mecanismos pelos quais os ovócitos adquirem a competência para
desenvolverem-se até o estádio de blastocisto ainda não estão totalmente
compreendidos. Há evidências de que a aquisição da competência está
correlacionada com moléculas de RNAs e de proteínas estocadas e processadas
durante as fases de crescimento, “capacitação” (pré-maturação) e maturação do
ovócito. Essas moléculas são usadas para sustentar as fases iniciais da
embriogênese até que a transcrição do DNA do embrião se torne ativa (DE LA
FUENTE e EPPIG, 2001).
Apesar dos muitos esforços para melhorar a produção in vitro (PIV) de
embriões para fins científicos e/ou comerciais, a sua eficiência ainda é relativamente
baixa. Apenas 35-40% dos ovócitos bovinos maturados in vitro (MIV) desenvolvem-
se até o estádio de blastocisto (MAYES e SIRARD, 2001; SIRARD et al., 2006), e
ainda desses, somente 30% chegam a termo após a transferência (WARD et al.,
2002; PARK et al., 2005). Essas baixas taxas são determinadas por vários fatores
decorrentes das etapas que constituem o sistema de PIV de embriões. As condições
de cultura para maturação nuclear e citoplasmática do ovócito certamente têm um
papel fundamental, e uma maturação completa é essencial para o desenvolvimento
embrionário. Portanto, para a obtenção de um sistema que possibilite a produção de
um maior número de embriões é necessária a compreensão dos mecanismos
envolvidos na maturação.
Observou-se que in vivo, os ovócitos, ao final de seu período de crescimento,

2
mas antes do período de maturação propriamente dito, passam por um período
chamado de “capacitação”, o qual parece ser importante para o desenvolvimento
pleno de sua competência. Nesse período ocorrem modificações estruturais e
moleculares que são importantes para o desenvolvimento embrionário (HYTTEL et
al., 1997, DIELEMAN et al., 2002).
Os ovócitos utilizados no sistema de PIV de embriões normalmente já
concluíram sua fase de crescimento e entraram no período de “capacitação”
ovocitária (HYTTEL et al., 1997). Porém, ao serem removidos do ambiente folicular
reiniciam a meiose (KUBELKA et al., 2000) e uma parte dessa população ainda não
concluiu a “capacitação” ovocitária.
Uma das alternativas que vários pesquisadores vêm analisando para estudar
a competência de desenvolvimento dos ovócitos in vitro é o bloqueio da retomada da
meiose antes da maturação. No entanto, pouco se sabe do que ocorre nesse
período. De toda forma, o bloqueio da meiose serviria como ferramenta para estudar
os possíveis fatores envolvidos na indução da “capacitação” após a remoção dos
ovócitos do ambiente folicular, e que poderiam ter efeitos positivos sobre o
desenvolvimento embrionário subseqüente. A obtenção de conhecimento nesse
sentido, por sua vez, poderá contribuir para um melhor embasamento no
desenvolvimento de procedimentos mais eficientes para a PIV de embriões bovinos.

3
2. REVISÃO DE LITERATURA
2.1. Ovogênese e foliculogênese
A ovogênese refere-se à seqüência de eventos em que as células
germinativas primordiais diferenciam-se em ovogônias, ovócitos primários e
secundários, findando com a fecundação do ovócito maturo (GONSALVES et al.,
2002; VAN DEN HURK e ZHAO, 2005). Já a foliculogênese inicia-se com a
formação dos folículos
primordiais,
progredindo para
folículos primários,
secundários e
terciários, finalizando
com a ovulação de um
ovócito maturo
(GONSALVES et al.,
2002; VAN DEN
HURK e ZHAO, 2005)
(Figura 1). A
diferenciação das
células da linhagem
germinativa feminina ocorre totalmente ou quase totalmente na fase
embrionária/fetal dependendo da espécie (MOORE e PERSAUD, 2000;
GONSALVES et al., 2002) (Tabela 1). Estudos recentes, porém, sugerem que possa
Figura 1. Diagrama simplificado da ovogênese e foliculogênese. Fonte: adaptado de Gonsalves et al., (2002) e Van den Hurk e Zhao (2005).
C. germinativas primordiais
Endoderma
Ovogônia
Ovócito 1°
Folículo primordial Folículo 1º
Folículo 2º
Folículo ovulatório
Meiose IIOvulação
Fecundação
Zigoto
Mitose
FO L I C U L O G Ê N E S E
Blastocisto
O V O G Ê N E S
Dictióteno
Início da meiose I
Prófase I
Prófase I
Folículo 3º Folículos pré-antrais
Folículos antrais
E
Ovócito 2°
Término da meiose I
Término da meiose II

4
haver diferenciação dessas células também em adultos, embora a maior parte das
evidências ainda indique que folículos primários sejam formados apenas na vida
fetal (BUKOVSKY et al., 2005).
Ao nascer, os bovinos dispõem de um número determinado de ovócitos
primários, cada um dos quais com o material genético duplicado (GONSALVES et
al., 2002). O processo de
diferenciação dos ovócitos
primários (imaturo) para
secundários (maturo) é iniciado
na maturidade sexual quando, a
intervalos médios de 21 dias
(bovinos), um ovócito (2n)
conclui a primeira meiose
iniciada na fase fetal. Agora, o ovócito secundário ou maturado (n) encontra-se apto
para a fecundação. Com a fusão do espermatozóide, o ovócito finaliza sua segunda
divisão meiótica (extrusão do segundo corpúsculo polar). Desse modo o material
genético do ovócito é reduzido à metade por meio de duas divisões meióticas
sucessivas. A ploidia retorna à sua conformação original (2n) com a interação
(singamia) do material genético do espermatozóide com o do ovócito, formando
assim o zigoto, que originará o embrião (MOORE e PERSAUD, 2000; GONSALVES
et al., 2002; HAFEZ, 2002).
Tabela 1. Cronologia da ovogênese e foliculogênese em bovinos e ovinos.
Eventos Dias de gestação Bovinos Ovinos
Células germinativas primordiais 35 23 Ovogônias 60 48 Ovócitos primários 75-80 55 Folículo primordial 90-130 90 Folículos primários 140 95 Folículos secundários 210 103 Folículos terciários 230-250 150 Nascimento 280 150
Fonte: McNatty et al., 2000; Gonsalves et al., 2002; Fair, 2003
2.2 . Maturação ovocitária
Durante todo o período do desenvolvimento, desde a formação e crescimento
dos ovócitos e dos folículos, até após o período da dominância folicular, os ovócitos
bovinos permanecem em estádio de prófase da primeira meiose. In vivo, o reinício
da meiose ocorre com o surgimento do pico pré-ovulatório de LH (hormônio
luteinizante) e se dá somente nos ovócitos inteiramente crescidos e meioticamente
competentes dos folículos pré-ovulatórios (FAIR, 2003; RODRIGUEZ e FARIN,
2004). A competência dos ovócitos tem uma estreita relação com as multicamadas
de células do cumulus que os cercam, mantendo uma importante via de

5
comunicação através das junções comunicantes (gap), antes e durante o pico pré-
ovulatório de LH (RODRIGUEZ e FARIN, 2004; GILCHRIST et al., 2004). Logo após
o pico de LH, começa a ocorrer o desaparecimento dessas vias de comunicação
entre o ovócito e as células do cumulus (HYTTEL et al., 1997).
A progressão do ciclo celular do ovócito até o estádio de metáfase da
segunda meiose (metáfase II), tanto in vivo quanto in vitro, é marcada por uma série
de transformações bioquímicas e estruturais no núcleo e no citoplasma do ovócito,
eventos esses que caracterizam a maturação ovocitária (MACHATKOVA et al.,
2004; DE SOUSA et al., 2004). Na transição da prófase I à metáfase II, estabelece-
se uma complexa cascata de fosforilações e desfosforilações de uma série de
proteínas envolvidas no reinício e na regulação da meiose (DEKEL, 2005; DUMONT
et al., 2005). Entre as proteínas que mais se destacam no período da maturação
estão as proteínas do complexo MPF (fator promotor da maturação) e da família
MAPK (proteína cinase ativada por mitógenos). Para efeito de estudo, a maturação
será dividida em maturação nuclear e citoplasmática, apesar desses eventos
ocorrerem concomitantemente.
2.2.1. Maturação nuclear
A diferenciação do ovócito imaturo em maturo envolve uma série de
alterações que são estimuladas principalmente por hormônios gonadotróficos
(MERTON et al., 2003). Sob a influência dos hormônios, o ovócito recomeça seu
ciclo celular progredindo da fase de diplóteno da prófase da primeira meiose
(prófase I), passando pelos estádios de metáfase I, anáfase I, telófase I (término da
primeira divisão meiótica) e progredindo até o estádio de metáfase da segunda
divisão meiótica (MEINECKE et al., 2001). No intervalo que compreende os estádios
de prófase I a metáfase II, os cromossomos condensam e o envelope nuclear é
desfeito marcando o início da maturação nuclear (MEINECKE et al., 2001; JONES,
2004). Dando seqüência, os cromossomos homólogos são divididos em dois grupos,
com a metade do número original de cromossomos. Ao término da primeira divisão
meiótica o citoplasma é dividido assimetricamente (CAN et al., 2003), gerando duas
células de tamanhos diferentes: uma pequena chamada de corpúsculo polar e outra

6
grande, o ovócito secundário. Ao término da maturação nuclear o ovócito permanece
nesse estádio do ciclo celular (metáfase II) até a fecundação (MAYES e SIRARD,
2001) ou ativação partenogenética (YI e PARK, 2005).
Os fatores da ativação (natural ou artificial) dos ovócitos maturos vão
promover o término da segunda divisão meiótica, que se caracteriza pela progressão
da metáfase II até a telófase II com a liberação do segundo corpúsculo polar. Após a
ativação, o futuro embrião prossegue seu ciclo celular por divisão mitótica. In vitro, a
maturação nuclear de ovócitos bovinos de folículos > 2 mm de diâmetro em
condições apropriadas requer de 18 a 24 horas de cultivo (KHATIR et al., 1998;
DODE e ADONA, 2001; ADONA e LEAL, 2004).
2.2.2. Maturação citoplasmática
A competência de desenvolvimento do ovócito em embrião em estádio
adiantado de desenvolvimento é basicamente dependente das modificações
bioquímicas, moleculares e estruturais do citoplasma. Essas alterações na
maturação do ovócito são processos altamente complexos que envolvem vários
eventos simultâneos como síntese de proteínas (KHATIR et al., 1997; SIRARD et al.,
1998), modificações moleculares (KUBELKA et al., 2000) e migração e
reorganização de organelas no citoplasma (STOJKOVIC et al., 2001). Se
compararmos as taxas de
maturação (Tabela 2),
observaremos que a maturação
citoplasmática (taxa de
blastocistos) in vitro está aquém
da maturação nuclear (taxa de
metáfase II) (HASHIMOTO et al., 2002; OYAMADA et al., 2004; ADONA e LEAL
2004). Por outro lado, in vivo, as taxas de blastocistos são de aproximadamente 80%
(RIZOS et al., 2002).
Tabela 2. Comparação entre maturação nuclear e citoplasmática.
Nuclear1 Citoplasmática2
81,0% 33,5% 81,4% 22,2% 90,9% 34,8%
1 Taxa de ovócitos em estádio de metáfase II; 2 Taxa de embriões em estádio de blastocisto, após sete dias de cultivo in vitro. Fonte: Hashimoto et al., 2002; Oyamada et al., 2004; Adona e Leal, 2004

7
2.2.3. Organização estrutural na maturação
2.2.3.1. Citoesqueleto
Microtúbulos, um dos componentes do citoesqueleto, são filamentos
altamente dinâmicos e sua dinâmica se dá pela adição (polimerização) ou remoção
(despolimerização) constante de novas unidades de tubulina α e β. Na transição do
ciclo celular durante a maturação de ovócitos de mamíferos ocorre uma intensa
reorganização dos microtúbulos (KIM et al., 2000).
Durante a maturação meiótica, duas populações de centrossomos regulam
coordenadamente o conjunto de microtúbulos nos eventos nucleares e
citoplasmáticos, tais como formação do eixo meiótico orientado assimetricamente,
segregação dos cromossomos durante a meiose, a extrusão do primeiro corpúsculo
polar e a formação da segunda placa metafásica (ALBERTS et al., 1997; KIM et al.,
2000; CAN et al., 2003; SUN et al., 2004).
Os erros de segregação dos cromossomos durante uma ou outra divisão
meiótica podem resultar em embrião aneuplóide após a fecundação, o que por sua
vez pode ter grave conseqüência para o desenvolvimento (BRUNET et al., 2003;
SUN et al., 2004). Especificamente nos mamíferos, a perda ou ganho de um
cromossomo autossômico resulta em distúrbios fisiológicos e de desenvolvimento
(ABRUZZO e HASSOLD, 1995).
Para o sucesso da fecundação, o fuso meiótico no ovócito deve permanecer
estável e corretamente organizado durante o
bloqueio do ovócito em metáfase II (TERRET et
al., 2003). Há evidências de que as proteínas da
família MAPK (proteína cinase ativada por
mitógenos) e a PKC (proteína cinase C) são
responsáveis pela estabilidade dos microtúbulos
mantendo o fuso meiótico estável em ovócitos de
ratos (HORNE et al., 2003; TONG et al., 2003).
Os substratos da MAPK (Figura 2), tais como a
p90rsk (proteína cinase ribossomo S6) e a MISS
(MAPK de interação e estabilização do fuso)
Figura 2. Proteínas envolvidas na estabilidade do fuso meiótico e na atividade do CSF em camundongos. Fonte: Lefebvre et al., (2002).
mos
MAPKK
MAPK
Miss P90RSK
Estabilidade do fuso MII CSF bloqueio ?

8
parecem participar dessa estabilidade via proteínas mos/.../MAPK (LEFEBVRE et al.,
2002; TERRET et al., 2003).
A rede de microtúbulos é usada por proteínas motoras (famílias das cinesinas
e dineínas) para gerar movimentos periódicos das organelas no interior da célula. As
proteínas motoras dependentes de microtúbulos desempenham uma função
importante no posicionamento das organelas (ALBERTS et al., 1997).
Os microfilamentos são os maiores componentes do citoesqueleto em
ovócitos de mamíferos e fornecem a estrutura para divisão celular (KIM et al., 2000).
Ficam situados principalmente no córtex celular (camada situada logo abaixo da
membrana plasmática rica em actina e uma variedade de proteínas) de ovócitos em
estádio VG (vesícula germinativa) e no eixo meiótico da célula depois do rompimento
da VG (desestruturação da membrana nuclear) (ALBERTS et al., 1997; KIM et al.,
2000). Estão envolvidos na incorporação do espermatozóide, na extrusão do
corpúsculo polar e na migração dos grânulos corticais para o córtex celular durante a
maturação ovocitária em diferentes espécies de mamíferos (CONNORS et al., 1998;
KIM et al., 2000; SUN et al., 2001a).
2.2.3.2. Organelas
2.2.3.2.1. Mitocôndrias
As mitocôndrias são a central bioenergética da célula com função claramente
essencial que define a competência funcional dos ovócitos. São organelas
especializadas que ocupam uma porção substancial do volume citoplasmático das
células de origem materna. São responsáveis pela produção da maior parte da
energia celular em forma de ATP (trifosfato de adenosina), por fosforilação oxidativa
através do metabolismo dos carboidratos e dos ácidos graxos contidos no
citoplasma provenientes do meio interno e externo (WILDING et al., 2001;
CUMMINS, 2004). Nos ovócitos em geral, há uma grande concentração de
mitocôndrias para suportar uma taxa mais elevada de síntese de moléculas dos
processos fisiológicos envolvidos no desenvolvimento. A eficiência da matriz
mitocondrial na conversão do piruvato em ATP é requerida para o processo de
maturação e divisão celular (WILDING et al., 2001). A inabilidade das mitocôndrias

9
de aumentarem e/ou acumularem ATP tem sido ligada ao desenvolvimento anormal
ou ao bloqueio do desenvolvimento embrionário (STEUERWALD et al., 2000).
Durante a mitose, as mitocôndrias são distribuídas aleatoriamente entre as
células filhas e no decorrer da ovogênese há um aumento substancial no número de
mitocôndrias (6000 para 193.000 mitocôndrias no ovócito em humanos) com uma
variação expressiva entre ovócitos do mesmo estádio de desenvolvimento
(REYNIER et al., 2001; CUMMINS, 2004). Porém, se os blastômeros embrionários
não receberem uma população suficiente de mitocôndrias para produção de ATP
podem tornare-se disfuncionais e fragmentados (CUMMINS, 2004). Há uma
correlação similar entre o potencial para o desenvolvimento, índice de ATP e função
mitocondrial tanto nos ovócitos quanto nos embriões bovinos (STOJKOVIC et al.,
2001).
Durante a maturação ovocitária as mitocôndrias modificam sua disposição
dentro do citoplasma celular. Elas são localizadas próximas às gotas de lipídios que
por sua vez aumentam de tamanho com a progressão da maturação (HYTTEL et al.,
1997). Em ovócitos de mamíferos em estádio de VG, as mitocôndrias são
encontradas em maior quantidade na periferia do citoplasma, e com pequenos
grupos dispersos mais ao centro do ovócito (HYTTEL et al., 1997; SUN et al.,
2001b). Já nos ovócitos em estádio metáfase II, as mitocôndrias estão mais
centralizadas no citoplasma (HYTTEL et al., 1997; SUN et al., 2001b).
2.2.3.2.2. Grânulos Corticais
Os grânulos corticais (GC) são organelas produzidas a partir do complexo de
Golgi e estão presentes apenas nos gametas femininos de todos os mamíferos, na
maioria dos vertebrados e em muitos invertebrados (WESSEL et al., 2001; MAYES,
2002). São possuidores de uma população de moléculas que incluem proteases,
glicosidases, enzimas e proteínas estruturais que contribuem para a barreira física e
bioquímica que bloqueia a polispermia (WESSEL et al., 2001).
Os GC são vesículas secretoras não renováveis e com advento da
fecundação o seu conteúdo não é mais sintetizado. Os RNAs que codificam as
diversas moléculas do conteúdo dos GC são degradados seletivamente no período

10
da maturação do ovócito e só voltam a ser transcritos e codificados nos novos
ovócitos do ciclo reprodutivo (WESSEL et al., 2001).
A principal função do conteúdo presente nos GC é de construir
(equinodermos – Ex.: ouriço-do-mar) ou de modificar (mamíferos – Ex.: bovinos) a
matriz extracelular (zona pelúcida) existente nos ovócitos para oferecer uma barreira
bioquímica e mecânica que impede a entrada de mais de um espermatozóide no
ovócito (WESSEL et al., 2001). Os GC são formados nos ovócitos em crescimento,
mas sua redistribuição ocorre no período da maturação (DUCIBELLA et al., 1994; HYTTEL et al., 1997). Primeiramente, os GC podem ser identificados em pequenos
grupos pelo citoplasma dos ovócitos em estádio de VG. Sua migração para a
periferia do ovócito vai ocorrendo com o avanço da maturação. Em estádio de
metáfase II, os GC estão distribuídos no córtex próximo à membrana plasmática
(CONNORS et al., 1998; WESSEL et al., 2001; VELILLA et al., 2004).
2.2.4. Mudanças moleculares e bioquímicas
Durante o crescimento, a “capacitação” e a maturação do ovócito, diversas
moléculas são sintetizadas e armazenadas. Essas moléculas vão dar suporte à
maturação e ao desenvolvimento após a fecundação, até que o genoma do embrião
se torne transcricionalmente ativo e as mensagens derivadas do embrião comecem
a regular a embriogênese (CHA e CHIAN, 1998; MERMILLOD et al., 2000;
MEIRELLES et al., 2004). A maioria dos RNAm (ácido ribonucléico mensageiro) no
ovócito é sintetizada e acumulada durante o período de crescimento do ovócito (DE
SOUSA et al., 1998). Esse metabolismo no ovócito é caracterizado pela transcrição
e pela tradução ativa durante o período pré-ovulatório. Entretanto, a transcrição
gênica bovina cessa antes da ovulação, assim, o ovócito, o zigoto e o embrião com
menos de 16 blastômeros são dependentes do “pool” dos RNAs e das proteínas
acumuladas durante o período de crescimento e de maturação do ovócito
(GANDOLFI e GANDOLFI, 2001; MEMILI e FIRST 2000; LONERGAN et al., 2003).
Ovócitos de muitas espécies dependem da síntese e da ativação de uma
variedade de proteínas para que possam ingressar e prosseguir no processo de
maturação ovocitária. Entre as proteínas mais importantes que regulam o

11
mecanismo da maturação ovocitária estão as proteínas do complexo MPF e as
proteínas da família MAPK (SHENG et al., 2002; TIAN et al., 2002; JONES, 2004).
2.2.4.1. Cinases dependentes de ciclinas (CDKs)
As CDKs são uma família de serina/treonina cinases envolvidas na regulação
do ciclo celular (CDK1, 2, 3, 4, 6 e 7), na transcrição (CDK7, 8 e 9) ou na função
neuronal (CDK5) (SCHANG, 2004; MAPELLI et al., 2005). A atividade da CDK é
dependente da interação com uma ciclina, cujos níveis são regulados
seqüencialmente para assegurar que as fases do ciclo celular prossigam na ordem
correta (ARRIS et al., 2000). Por exemplo, as ciclinas D interagem com as CDKs 4 e
6 durante a fase G1, a ciclina E com a CDK2 no final da fase G1, a ciclina A com a
CDK2 na fase S/G2 e a ciclina B com a CDK1 na fase G2/M (Figuras 3 e 4)
(KNOCKAERT et al., 2002; ARRIS et al., 2000). A falha no controle das CDKs e a
conseqüente perda da função do ponto de verificação do ciclo celular têm sido
relacionadas diretamente à patologia
molecular do câncer (ARRIS et
al.,2000).
Figura 3. Diagrama simplificado da atividade das CDKs/ciclinas na regulação do ciclo celular. Fonte: Knockaert et al., (2002) e Arris et al., (2000)
CDK 2 Ciclina E
CDK 1 Ciclina B
CDK 4/6 Ciclina D
CDK 2 Ciclina A
Mitose
Meiose
ovócito
Figura 4. CDKs envolvidas na regulação do ciclo celular, na transcrição e na função neuronal. Fonte: Knockaert et al., (2002)
CDK1 Ciclina B
Ringo Transição de prófase I para metáfase II Regulação da topoisomerase II
Transição das fases G1/S e S/G2 Duplicação do centrossomo.
Transição das fases G1/S
Fase G1
Apoptose Migração dos neurônios Envolvida em outros eventos celular
Sinal de tradução e transcrição
Fase G1, morte de células neurais.
Ativação da CDK1 e CDK2
Regulação da DK7/ciclina H
Processamento de RNA ou transcrição Apoptose
CDK2
CDK3 Ciclina E?
Ciclina E
Ciclina A
Ciclina D
P35, P25
P39, P29
Ciclina D
Ciclina H
Ciclina C
Ciclina K
Ciclina T
Ciclina L
CDK4
CDK5
CDK6
CDK7
CDK8
CDK9
CDK11
Fase G1
F a s e S
Fase G2
F a s e M

12
2.2.4.2. Fator promotor da maturação (MPF)
O MPF é um heterodímero composto por uma cinase catalítica (p34cdc2 ou
CDK1) e uma subunidade de ciclina B regulatória (ABRIEU et al., 2001; JONES,
2004). Há ao menos três tipos de ciclinas B (B1, B2 e B3), e nos mamíferos parece
que a ciclina B1 é a principal responsável pela atividade do complexo MPF. Embora
a ligação da CDK1 com a ciclina B1 seja necessária, não é o suficiente para
desencadear a atividade do complexo MPF (CASTRO et al., 2001; JONES, 2004),
que é dependente dos mecanismos subseqüentes: fosforilação da CDK1 nos
resíduos de tirosina 15 (Y15),
treonina 14 (T14), treonina 161
(T161) e a posterior
desfosforilação dos resíduos
Y15 e T14 (KIKUCHI et al.,
2000; VAILLANT et al., 2001;
JOSEFSBERG e DEKEL,
2002). Em termos gerais a
CDK1 é fosforilada em Y15 e
T14 pelas proteínas cinases
Myt1 e Wee1, que causam uma
fosforilação inibitória da CDK1.
Nesse estádio o complexo MPF
está formado (pré-MPF), mas
ainda está inativo (Figura 5). A
ativação do MPF depende da
desfosforilação em Y15 e T14
pela proteína fosfatase CDC25
(MPF – ativo) (KIKUCHI et al., 2000; JONES, 2004). O aumento da atividade da
proteína CDC25 e a baixa atividade das proteínas cinases Myt1 e Wee1 são
necessários para a ativação completa do complexo MPF (JONES, 2004).
Figura 5. Diagrama simplificado do mecanismo de ativação do MPF. Fonte: Dekel (2005).
Wee1
Myt1
AMPc
PKC
Síntese
Ovócito em VG
Condensação dos cromossomos Rompimento da VG
Mos -RMAm poliadenilação
Mos síntese, MEK MAPK ativação.
Formação do fuso
Extrusão do 1ª corpúsculo polar
CSF estável Metáfase II bloqueada
Fecundação ou ativação artif. CSF instável
M E I O S E
A T I V A D A
n
Ciclina B
A variação da atividade do MPF pode ser detectada nos ovócitos bovinos
durante a maturação. Sua atividade é baixa em VG passando a ser observada no
início do rompimento da VG. Alcança um pico no estádio de metáfase I e declina sua
MPF
MPF Ativo
MPF Ativo
MPF Ativo
CDK1
M CDK1 E I O S E
B L O Q U E A D A
Ativação
CDC25

13
atividade durante a transição entre os estádios de metáfase I e metáfase II
(KUBELKA et al., 2000), reativando para entrada do ovócito em metáfase II. Sua
inativação nos ovócitos em estádio de metáfase II é induzida pela fecundação ou
pela ativação paternogenética (NEBREDA e FERBY, 2000; KUBELKA et al., 2000;
KIKUCHI et al., 2000; ABRIEU et al., 2001; LEDAN et al., 2001). A inativação
abrupta do MPF é considerada como um disparador necessário para o escape da
meiose bloqueada em metáfase II (TAIEB et al., 1997).
O processo de desorganização do heterodimero do complexo MPF independe
da sua ativação catalítica, que é causada geralmente pela proteólise da ciclina B.
Em ovócitos fecundados de camundongos e de suínos, a degradação da ciclina B foi
claramente relacionada com a inativação do complexo MPF (WINSTON, 1997;
KIKUCHI et al., 1999).
O MPF é um dos principais reguladores das alterações morfológicas durante
a maturação ovocitária, regulando a condensação dos cromossomos, o rompimento
do envelope nuclear, a reorganização dos microtúbulos e outras organelas
citoplasmáticas com a participação de outras proteínas (KIM et al., 2000; KANO et
al., 2000; KRISCHEK e MEINECKE, 2002; KUBELKA et al., 2002; LEFEBVRE et al.,
2002).
2.2.4.3. Proteína cinase ativada por mitógenos (MAPK)
Um segundo grupo importante de proteínas que estão envolvidas na
progressão da meiose pertencem à família das serina/treonina cinases, as proteínas
cinases ativadas por mitógenos (MAPK) (KUBELKA et al., 2000). Essas proteínas
são ativadas dentro de vias específicas de transdução de sinais (Figura 6), por sinais
extracelulares (NEBREDA e FERBY, 2000). Por esta razão, a MAPK também é
chamada de ERK (cinase regulada por sinal extracelular – suas variantes, ERK1/2 –
p44/p42 kDa) (KRISCHEK e MEINECKE, 2002). A via intracelular de transdução de
sinal consiste na proteína mos (do oncogene c-mos) que ativa a MEK (MAPKK) que
fosforila a ERK (MAPK) (CHA e CHIAN, 1998; KRISCHEK e MEINECKE, 2002; SUN
et al., 2002).

14
A ampla faixa de atuação das MAPKs é mediada pela fosforilação de diversos
substratos, incluindo fosfolipases, fatores de transcrição e proteínas do
citoesqueleto. As MAPKs também catalisam fosforilação e a ativação de diversas
proteínas cinases, denominadas de proteínas cinases ativadas pelas MAPKs, que
representam um adicional enzimático de vários espectros em diferentes células
(ROUX e BLENIS, 2004).
As MAPKs são amplamente ativadas por fatores de crescimento e por soro,
com uma ativação menor pelo
estresse, efeito osmótico e pela
desorganização dos microtúbulos.
Uma variedade de estímulos
diferentes pode ativá-las, mas no
geral a ERK1 e ERK2 são ativadas
preferencialmente em resposta aos
fatores de crescimento, enquanto
as cinases JNK (c-jun amino
terminal cinase) e p38 cinase são
mais responsivas aos estímulos de estresse e efeitos osmóticos (CHEN et al., 2001;
ROUX e BLENIS, 2004).
MAP4K GDP GTP
A via MAPK é ativada universalmente durante a maturação meiótica em
ovócitos de vertebrados. Entretanto, o tempo requerido para sua ativação é díspar
nas diferentes espécies (NEBREDA e FERBY, 2000). A ativação da MAPK em
ovócitos bovinos ocorre após 8 horas de cultivo in vitro e apresenta um aumento
gradual até 12–14 horas e se mantém estável até o final da maturação (KUBELKA et
al., 2000).
As duas principais isoformas (ERK1/2) da MAPK são ativadas com a
proximidade do rompimento da VG em ovócito bovinos (KUBELKA et al., 2000). Isso
sugere que a MAPK não é requerida para o reinício da meiose, mas é essencial em
eventos pós-rompimento da VG (KANO et al., 2000; LEFEBVRE et al., 2002).
Porém, a injeção de MAPK ativa em ovócitos de suíno ou de bovino induz o
rompimento da VG, indicando que essa proteína promove o reinício da meiose em
condições especiais (FISSORE et al., 1996; INOUE et al., 1998).
MAP3K MAPKK MAPK Figura 6. Diagrama simplificado das vias de ativação das MAPKs. Fonte: Cha e Chian 1998; Nebreda e Ferby, 2000; Kubelka et al., 2000; Krischek e Meinecke, 2002; Sun et al., 2002.
GTP
Raf-1
MEK
ERK
CSF
GDP
mos
AMPc ⇓
PKA ⇓
Ras

15
2.3. Competência ovocitária
A competência para a progressão da maturação meiótica em ovócitos de
mamíferos é adquirida durante o crescimento folicular (PAVLOK et al., 2000;
MIYANO, 2003). Nesse contexto, para que o ovócito tenha competência para a
maturação tanto nuclear como citoplasmática esse deve completar sua fase de
crescimento. Em bovinos foi demonstrado que folículos de diâmetro acima de 3 mm
de diâmetro contêm ovócitos (110-120 μm de diâmetro) com potencial de
desenvolvimento mais elevado que os de folículos menores (GANDOLFI e
GANDOLFI, 2001). Ovócitos obtidos de folículos maiores de 3 mm de diâmetro
resultam em blastocistos com maior número de células nucleadas, comparados com
os ovócitos obtidos de folículos menores que 3 mm (AVELINO et al., 1998).
Ainda não está totalmente compreendido como os ovócitos adquirem a
competência meiótica durante sua fase de crescimento e se tornam competentes
para reiniciar e terminar a meiose. Há evidências de que a aquisição da competência
meiótica nos ovócitos está correlacionada com as moléculas de RNAs e de proteínas
estocadas durante a fase de crescimento e maturação do ovócito e com o aumento
da funcionalidade das mitocôndrias (DE LA FUENTE e EPPIG, 2001; STOJKOVIC et
al., 2001; TOMEK et al., 2002; CHIAN et al., 2003).
Essas moléculas são usadas para sustentar as fases iniciais do
desenvolvimento embrionário antes que a transcrição do DNA do embrião se torne
ativa (DE LA FUENTE e EPPIG, 2001). Os padrões de síntese protéica aumentam
antes do rompimento da vesícula germinativa em ovócitos durante o cultivo de
maturação, sugerindo que essa síntese de proteínas possa ser importante para o
subseqüente desenvolvimento embrionário (TOMEK et al., 2002; CHIAN et al.,
2003). Isso faz do ovócito uma célula muito especial, completamente diferente das
células somáticas, onde os RNAs e as proteínas se submetem geralmente a uma
rápida rotação de estoque. Para permitir o estoque e o uso oportuno de tais
moléculas armazenadas, vários mecanismos necessitam ser eficaz nesse controle
no ovócito (GANDOLFI e GANDOLFI, 2001; TOMEK et al., 2002).
Em bovinos, os RNAs e as proteínas do ovócito conduzem o desenvolvimento
embrionário inicial após a fecundação até o 4º ciclo celular quando o controle
genômico (estádio de 8 a 16 células) do embrião torna-se evidente. Muitos dos

16
produtos derivados dos transcritos maternos são necessários para preparar o
maquinário biológico do embrião (SIRARD, 2001; DIELEMAN et al., 2002; KAÑKA,
2003).
A competência meiótica em ovócitos de Xenopus sp está correlacionada com
as mudanças no acúmulo e na ativação de moléculas do ciclo celular (JESSUS e
OZON, 2004). Esses mesmos
mecanismos foram observados em
ovócitos em crescimento de suínos
que começam a acumular
moléculas CDK1. Já a ciclina B foi
observada em ovócitos de folículos
antrais pequenos quando cultivados
em meio de maturação (Figura 8),
mas a CDK1 foi negativamente
fosforilada e assim inativada, talvez
pela proteína Myt1 (KANAYAMA et al., 2002; MIYANO, 2003). Nos folículos de 1,0-
1,5 mm de diâmetro, os ovócitos iniciam a meiose, mas param em metáfase I. Eles
são capazes de ativar o MPF, mas não estabelecem a cascata de proteínas da via
MAPK (KANAYAMA et al., 2002; MIYANO, 2003). Os ovócitos em crescimento
desenvolvem primeiramente a habilidade de ativar CDK1 e posteriormente de ativar
a via MAPK. Somente os ovócitos com o crescimento completo possuem
competência para ativar efetivamente as duas vias do ciclo celular.
Folículos 0,5-0,7mm 1,0-1,5 mm 4,0-6,0 mm Ovócitos 104 μm 109 μm 122μm
0 27 42 0 27 42 0 27 42h
histona H1 MBP Cultivo (h)
Figura 8. Mudanças nas atividades da CDK1 e da MAPK durante a maturação de ovócitos de suínos em vários estádios do crescimento folicular. As atividades da CDK1 e da MAPK foram detectadas pela fosforilação de histona H1 e da proteína básica de mielina (MBP), respectivamente. Adaptado a partir de Miyano, 2003.
Na produção de embriões in vitro, a maioria dos ovócitos bovinos são
provenientes de folículos de 2 a 6 mm de diâmetro (ADONA e LEAL, 2004; DALVIT
et al., 2005). A remoção dos ovócitos dos folículos antes de eles finalizarem a
foliculogênese é, provavelmente, um dos fatores envolvidos na baixa produção de
embriões in vitro. Assim sendo, a interrupção da foliculogênese seria um fator
responsável pela redução da competência ovocitária. Para contornar a
foliculogênese interrompida a que os ovócitos são submetidos, uma alternativa seria
mantê-los por um determinado período com a mesma configuração nuclear daquela
encontrada nos folículos. Para tal, os ovócitos seriam submetidos ao bloqueio
meiótico, evitando a retomada da meiose logo após a sua remoção dos folículos,
para que tenham o tempo necessário para sofrerem as mudanças necessárias antes
de serem submetidos à maturação in vitro.

17
2.4. Bloqueio meiótico
A maturação espontânea em ovócitos de bovinos ocorre in vitro após sua
retirada do ambiente folicular, o que se deve, provavelmente, à remoção do sinal
inibidor proveniente do folículo. Estudos sugerem que o fator inibidor que controla a
retomada da meiose seja produzido pelas células da teca e/ou granulosa (células
foliculares) (FOOTE e THIBAULT, 1969; SIRARD et al., 1998). KOTSUJI et al.
(1994) demonstraram que o fator inibidor da meiose em ovócitos bovinos seria
sintetizado pelas células da granulosa, mas que as células da teca contribuiriam na
amplificação desse sinal. O mecanismo que envolve o reinício da meiose também
está associado a redução nas concentrações de AMPc (monofosfato de adenosina
cíclica) (CONTI et al., 1998).
O AMPc é uma molécula sinalizadora intracelular que exerce um importante
controle na meiose em mamíferos, anfíbios e em alguns invertebrados (BILODEAU-
GOESEELS, 2003). A redução do AMPc no interior do ovócito parece estar
envolvida com a ruptura da vesícula germinativa, ao menos in vitro. Assim, os níveis
elevados da AMPc dentro do ovócito mantêm o bloqueio meiótico, visto que a
redução do AMPc é um sinal necessário para a maturação ovocitária (CONTI et al.,
1998; EYERS et al., 2005).
A regulação nas atividades enzimáticas de uma série de proteínas cinases e
fosfatases controla a meiose em ovócitos de mamíferos. Dessa forma, a meiose
pode ser bloqueada por inibidores que mantenham altas concentrações de AMPc no
interior do ovócito (BILODEAU-GOESEELS, 2003), por inibidores não específicos da
síntese protéica (MEINECKE et al., 2001), de proteínas cinases (ANDERIESZ et al.,
2000; DODE e ADONA, 2001) e por inibidores específicos da fosforilação de
proteínas CDKs (ADONA e LEAL, 2004).
In vitro, o reinício da meiose ocorre porque o ovócito é liberado da influência
dos inibidores provenientes do folículo (KOTSUJI et al., 1994). No entanto, a
maturação in vitro resulta em uma redução na produção de embriões, sugerindo que
nem todos os ovócitos conseguem maturar adequadamente (maturação
citoplasmática). Portanto, diferentes protocolos estão sendo usados in vitro para
permitir que todos (ou a maioria) os ovócitos coletados terminem a maturação
nuclear sem afetar a qualidade dos mesmos, a fim de maximizar a produção de

18
embriões (ADONA e LEAL, 2004; SCHOEVERS et al., 2005). O uso de inibidores
fisiológicos ou farmacológicos pode ser uma alternativa in vitro para um cultivo de
pré-maturação antes de submeter os ovócitos à maturação, fecundação e ao
desenvolvimento embrionário.
A ativação das cinases dependentes de ciclinas (CDKs - MPF) é um ponto
chave no reinício da meiose em ovócitos. Na tentativa de sincronizar o
desenvolvimento nuclear nos ovócitos in vitro, alguns estudos demonstraram a
inibição da ativação do MPF pelo aumento de níveis intracelulares de AMPc com
dbcAMP ou hipoxantina (SUN et al., 1999; MA et al., 2003), pela inibição não
específica da síntese protéica com cicloheximida (MEINECKE et al., 2001) ou pela
inibição não específica de proteínas cinases com 6-dimetilaminopurina
(ANDERIESZ et al., 2000; DODE e ADONA, 2001). Embora a inibição da ativação
do MPF seja bem sucedida com o uso dessas drogas, os efeitos não específicos
desses inibidores são prejudiciais para o desenvolvimento subseqüente do ovócito.
As CDKs desempenham um papel central na regulação do ciclo da divisão
celular, o que lhes faz um alvo promissor para o desenvolvimento de agentes
terapêuticos do câncer. Um grande esforço foi feito nos últimos anos na busca de
inibidores específicos de CDKs de baixo peso molecular (BRAÑA et al., 2004). A
butirolactona I é um inibidor natural isolado do fungo Aspergillus terreus, que exibe
atividade antiproliferativa, inibindo seletivamente em mamíferos as cinases CDK2 e
CDK1, que executam um importante papel na progressão do ciclo celular nas fases
G1/S e G2/M, respectivamente (SCHIMMEL et al., 1998; SCHANG, 2004).
Entretanto, tem pouco efeito nas proteínas cinases ativadas por mitógenos, proteína
cinase C, cinases dependentes de AMPc, caseína cinase I e II e no receptor tirosina
cinase do fator de crescimento epidermal (SCHIMMEL et al., 1998; SAX et al., 2002;
BRAÑA et al., 2004). A butirolactona I comporta-se como um inibidor competitivo
pelo local de ligação do ATP na CDK1 (SAX et al., 2002; BRAÑA et al., 2004).
É possível manter os ovócitos bovinos em estádio de vesícula germinativa in
vitro por um determinado período de tempo com o uso de inibidores específicos ou
não de CDKs (butirolactona I, roscovitina, cicloheximida e 6-dimetilaminopurina).
Essas informações são importantes, pois tornam possível fazer um cultivo de pré-
maturação para, posteriormente, induzir a maturação final. Assim, permite ao ovócito
um tempo maior para sofrer as mudanças necessárias à aquisição da competência
meiótica, visto que estes são submetidos a uma foliculogênese interrompida, sendo

19
removidos precocemente dos folículos e ocorrendo a retomada espontânea da
meiose. Entretanto, convém ressaltar que nesse tipo de estudo é importante verificar
não só a efetividade das substâncias em inibir o reinício da meiose, mas também
sua eficiência na reversibilidade e ausência de efeitos negativos sobre o
desenvolvimento embrionário.

20
3. OBJETIVOS
O presente estudo teve por objetivo avaliar o efeito da butirolactona I no
cultivo de pré-maturação sobre a meiose, estruturas celulares e desenvolvimento
dos ovócitos bovinos cultivados in vitro.
Mais especificamente foram avaliados:
1) as condições de redução da concentração de BL I para indução de bloqueio
meiótico e sua reversão (maturação in vitro);
2) a cinética de maturação pós-bloqueio;
3) a organização do citoesqueleto (microtúbulos e microfilamentos) após o
bloqueio meiótico e sua reversão;
4) a distribuição de organelas (grânulos corticais e mitocôndrias) após o bloqueio
meiótico e sua reversão;
5) o efeito do bloqueio meiótico sobre o desenvolvimento embrionário.

21
4. MATERIAL E MÉTODOS
4.1. Coleta de ovários e ovócitos
Ovários de fêmeas bovinas foram coletados em frigoríficos logo após o abate
e transportados em solução fisiológica estéril (NaCl 0,9% – Sigma, S9625) acrescida
de antibióticos (100 UI/mL de penicilina – Sigma, P7794 e 100 ug/mL de
estreptomicina – Sigma, S9137) a uma temperatura de 30º C. No laboratório, os
ovários foram lavados em NaCl 0,9% e os folículos com diâmetro de 2 – 6 mm
foram aspirados com auxílio de uma agulha de 18 “G” (1,20 X 40 mm) conectada a
uma seringa descartável de 10 mL. O líquido folicular contendo os ovócitos foi
depositado em tubos cônicos de 50 mL (Cellstar, 227261) e mantido em repouso
para decantação por 5 minutos. A porção superior do líquido foi retirada e na porção
restante foram adicionados 3-5 mL de meio H199 (TCM199 com 25 mM de Hepes –
Gibco, 12340030) acrescido de 100 UI/mL de penicilina e 100 ug/mL de
estreptomicina; suplementado com 1% de soro fetal bovino – Gibco, 26140-079).
Posteriormente, o material do tubo foi transferido para uma placa de Petri (60 x 15
mm – Falcon, 353002), onde foi feita a busca dos ovócitos sob estereomicroscópio
para avaliação e classificação dos mesmos. Somente ovócitos classificados como
graus I e II (DE LOOS et al., 1991), contendo três ou mais camadas de células do
cumulus oophorus e citoplasma uniforme, foram utilizados para os experimentos.

22
4.2. Solução estoque do inibidor
O inibidor de cinases dependentes de ciclinas (CDKs) butirolactona I (Biomol,
87414-49-1) foi preparado em solução estoque de 50 mM em dimetilsulfóxido
(DMSO – Sigma, D5879) e rediluído para 5 mM com B199 [TCM199 com sais de
Earle e com 20 mM de bicarbonato de sódio (Gibco-11150-067) suplementado com
10 μg/mL de gentamicina (Sigma, G1272)], aliquotado em tubos para
microcentrífuga e conservado no freezer a -20º C.
4.3. Bloqueio da meiose com butirolactona I
Para o bloqueio da meiose os ovócitos bovinos foram cultivados in vitro com
butirolactona I (BL I) diluída em meio B199 (suplementado com 0,2 mM de piruvato –
Sigma, P5280) na concentração apropriada do experimento (descrita
posteriormente). O cultivo foi feito em gotas de 100 µl de meio de bloqueio (±20
ovócitos por gota), sob óleo mineral (Sigma, M8410) a 38,5º C em atmosfera de 5%
de CO2 em ar.
4.4. Determinação do estádio da meiose
Para determinação do estádio da meiose, os ovócitos foram desnudados
(remoção das células do cumulus oophorus) em tubo de 5 mL com 0,3 mL de PBS
acrescida de 1% de SFB (soro fetal bovino) e agitados no “vortex” por 4 minutos
(Figura 9). Os ovócitos desnudos foram fixados entre lâmina e lamínula por 24 horas
em etanol (Synth, A1084.01.BJ) e ácido acético (Synth, A1019.01.BJ) 3:1. Após a
fixação, foram corados com lacmóide 0,004% (Aldrich, 274720) e observados em
microscópio de contraste de fase para determinação dos estádios da meiose. Os
ovócitos foram classificados de acordo com a configuração nuclear em vesícula
germinativa (VG), metáfase I (M I), anáfase I (A I), telófase I (T I) e metáfase II (M II)

23
(Figura 10). A taxa de bloqueio foi avaliada como a percentagem de ovócitos em VG
e a de reversão como a percentagem de ovócitos em M II.
Figura 9. Procedimentos para fixação e coloração de ovócitos.
3) Pressionar a lamínula sobre a lâmina suavemente até fazer uma pressão nos ovócitos – pouca pressão os ovócitos ficam soltos entre a lâmina/lamínula; o inverso os ovócitos rompem. 4) Colocar a lâmina no fixador 3:1 (álcool etílico e ácido acético). 5) Deixar fixando por no mínimo 24 h.
6) coloração com lacmóide ou orceina acética
a) Colocar a lâmina em um ângulo de 45°, adicionar o corante na parte superior da lâmina; b) Corar por ±5 minutos; c) se for necessário remover o excesso de corante com o próprio fixador; d) Não deixar secar a lâmina – vedar com esmalte. (corante: 40 mL ácido acético, 60 mL água + 0,004% do corante – aquecer para dissolver melhor e filtrar)
2) Preparação das lâminas para fixação dos ovócitos
Lâmina: Fazer uma gota de ±4μL
com ± 25 ovócitos.
Lamínula: Colocar pequenas gotas
de cola (silicone) nas bordas.
1) Procedimento para desnudar os ovócitos
Em um tubo de 5 mL adicionar 0,3 mL de PBS e os ovócitos. Agitar no vortex (velocidade 6-7) por 4 minutos; lavar a parede do tubo com 2 mL de meio; transferir o conteúdo para uma placa de Petri para seleção dos ovócitos.
P B S
A B C D
400X 400X 200X 200X
Figura 10. Classificação da meiose de ovócitos bovinos. A) vesícula germinativa; B) metáfase I; C) anáfase I/telófase I; D) metáfase II. Seta vermelha: placa metafásica e seta azul: 1° corpúsculo polar.
4.5. Maturação in vitro
Após o período de bloqueio, os ovócitos foram lavados três vezes em meio
livre de inibidor e transferidos para o meio de maturação [B199 suplementado com
10% de SFB (soro fetal bovino), 5,0 µg/mL de LH (hormônio luteinizante, Bioniche –
Lutropin-v), 0,5 µg/mL de FSH (hormônio folículo estimulante, Bioniche – Folltropin-
v), 0,2 mM de piruvato e 10 μg/mL de gentamicina]. Como controle, um grupo de
ovócitos foi submetido à maturação sem sofrer cultivo prévio de bloqueio. O cultivo

24
de maturação in vitro (MIV) foi feito em gotas de 100 µl de meio de maturação (±20
ovócitos por gota), sob óleo mineral a 38,5 º C e atmosfera de 5% de CO2 em ar.
4.6. Microtúbulos
Para avaliação dos microtúbulos os ovócitos foram desnudados e fixados em
3,7% de paraformaldeído (Sigma, P6148) acrescido de 0,6% Triton X-100 (USB,
9002-93-1) em PBS (livre de cálcio e magnésio) com 0,1% de PVA (álcool polivinílico
- Sigma, P8136) por 30 minutos; lavados três vezes em PBS com 0,1% de PVA
(PP); bloqueados com 3% de soro de cabra (Invitrogen, 16210-064) em PP por 45
minutos; corados com anticorpo anti-alfa tubulina conjugada com FITC (Sigma,
F23168) (1:100) em PP por uma hora; lavados por três vezes em PP; corados com
10 μg/mL de iodeto de propídio (Sigma, P4170) em PP por 15 minutos; lavados três
vezes em PP e montados entre lâmina e lamínula com glicerol (USB, 56-81-5) para a
avaliação sob microscópio de epifluorescência.
4.7. Microfilamentos Para avaliação dos microfilamentos os ovócitos foram desnudados e fixados
em 3,7% de paraformaldeído mais 0,6% Triton X-100 em PP por 30 minutos; lavados
três vezes em PP; colorados com 1 μg/mL de faloidina conjugada com FITC (P5282)
mais 10 μg/mL de iodeto de propídio em PP por 30 minutos; lavados três vezes em
PP e montados entre lâmina e lamínula com glicerol para a avaliação sob
microscópio de epifluorescência.

25
4.8. Grânulos corticais
Para avaliação dos grânulos corticais os ovócitos foram desnudados e a zona
pelúcida foi removida com 0,5% protease (Sigma, P8811) em PBS (livre de cálcio e
magnésio) por ±6 minutos a 38,5 °C; fixados em 3,7% de paraformaldeído em PP
por 30 minutos; incubados em solução de bloqueio [SB: PBS suplementado com
0,1% de BSA, 0,751% de glicina (Sigma, G8790) e 0,2% de azida sódica (Research
Organics, 0939S)] por duas horas; permeabilizados com 0,1% Triton X-100 em SB
por 5 minutos; lavados três vezes em SB; corados com 1 μg/mL de lectina – Lens
culinaris (Sigma, L9262) e 10 μg/mL de iodeto de propídio em SB por 15 minutos;
lavados três vezes em SB e montados entre lâmina e lamínula com glicerol para a
avaliação sob microscópio de epifluorescência.
4.9. Mitocôndrias
Para avaliação das mitocôndrias os ovócitos foram desnudados; corados com
0,5 µM de Mitotracker red (Molecular probes) e 10 µg/mL Hoechst 33342 (Sigma,
B2261) em PP por 20 minutos; lavados três vezes em PP e montados entre lâmina e
lamínula para a avaliação sob microscópio de epifluorescência.
4.10. Fecundação e cultivo in vitro
Para a fecundação in vitro (FIV) foi utilizado sêmen congelado de um mesmo
touro e da mesma partida, o qual foi preparado segundo a técnica de gradiente
Percoll (Amersham Biosciences, 17-0891-01). Em um tubo cônico de 15 mL (Tedia
Brazil, 1475-1611.50), foi adicionado 1 mL de Percoll 90% e, em seguida, 1 mL de
Percoll 45% (Apêndice). O gradiente foi mantido na estufa de CO2 por uma hora
antes de sua utilização. O sêmen foi descongelado a uma temperatura de 37° C por
30 segundos e, em seguida, adicionado na porção superior do gradiente de Percoll e

26
centrifugado por 10-12 minutos a 650 “g”. Após a retirada do sobrenadante foram
adicionados 4 mL de meio TALP-sp (Apêndice), e uma nova centrifugação de 2
minutos a 200 “g” foi realizada para remover o excedente do Percoll.
Os espermatozóides foram adicionados na gota de fecundação a uma
concentração final de 2 x 106 espermatozóides/mL. A fecundação foi feita em TALP–
FIV (Apêndice) suplementado com 2 µM penicilamina (Sigma, P4875), 1 µM
hipotaurina (Sigma, H1384), 250 µM epinefrina (Sigma, E1635) e 20 µg/mL heparina
(181 UI/mg, USB, 9041-08-1). Os espermatozóides e os ovócitos foram co-
incubados em gotas de 100 µL de meio sob óleo mineral em placa de Petri (35 x 10
mm. Corning, 430165) por um período de 18 horas.
Após as 18 horas de FIV, os ovócitos foram parcialmente desnudados com
auxílio de um micro-pipetador de 50 µL. Após o desnudamento parcial, os ovócitos
foram transferidos para o meio de cultivo in vitro SOF (fluido de oviduto sintético -
Apêndice) em gotas de 100 µL e com 48 horas de cultivo in vitro (D2), foi avaliada a
taxa de clivagem. Os ovócitos clivados permaneceram no meio de cultivo in vitro e
os demais foram removidos e descartados. No quinto dia (D5) de cultivo in vitro foi
feita a troca (feeding) parcial (40%) do meio de cultivo e a partir do sétimo (D7) e do
oitavo dia (D8) foi feita a avaliação das taxas de formação de blastocisto (D7), de
eclosão (D8) e a contagem do número de células dos embriões eclodidos (D8). O
cultivo in vitro dos embriões foi feito em estufa de CO2 a 38,5ºC em atmosfera de 5%
de CO2 em ar.
Para a contagem do número de células dos embriões em estádio de
blastocisto eclodido, eles foram corados com 10 µg/mL de Hoechst 33342 em PP
por 15 minutos; lavados três vezes em PP e montados entre lâmina e lamínula para
a avaliação sob microscópio de epifluorescência.

27
4.11. Delineamento experimental
4.11.1. Experimento 1: Avaliação de concentrações decrescentes de BL I em meio suplementado com BSA.
Nesse experimento foram avaliadas concentrações decrescentes de BL I
(100, 50 e 25 μM) em meio de cultivo (B-199) suplementado com albumina sérica
bovina (BSA, Sigma, A9647) (Figura 11), para avaliar a capacidade da BL I em
promover o bloqueio meiótico em diferentes concentrações. Os ovócitos foram
cultivados na presença ou ausência (controle) de BL I por um período de 24 h. Após
o período de cultivo in vitro, os ovócitos foram desnudados, fixados, corados e
avaliados sob microscópio de contraste de fase para determinação das taxas de
bloqueio da meiose (VG). Um grupo de ovócitos foi fixado logo após a aspiração
(controle zero hora).
Figura 11. Esquema representativo do delineamento do experimento 1.
25 μM de BL I
50 μM de BL I
100 μM de BL I
fixados
Controle C/0
24 h de cultivo com BSA
desnudados
corados
avaliados
Controle C/24
Avaliado logo após a aspiração
4.11.2. Experimento 2: Avaliação de concentrações crescentes de BL I em meio sem suplementação de BSA
Nessa avaliação, não foi usada suplementação protéica (BSA) no meio de
bloqueio (Figura 12). O bloqueio da meiose foi feito em meio B199 com diferentes
concentrações de BL I (10, 15, 20 e 25 μM) com o intuito de investigar a capacidade
do inibidor em bloquear a meiose em concentrações ainda mais baixas, porque
segundo KUBELKA et al., (2000) o inibidor (BL I) poderia interagir com o BSA,
reduzindo sua disponibilidade no meio. Com base nesse estudo os ovócitos bovinos
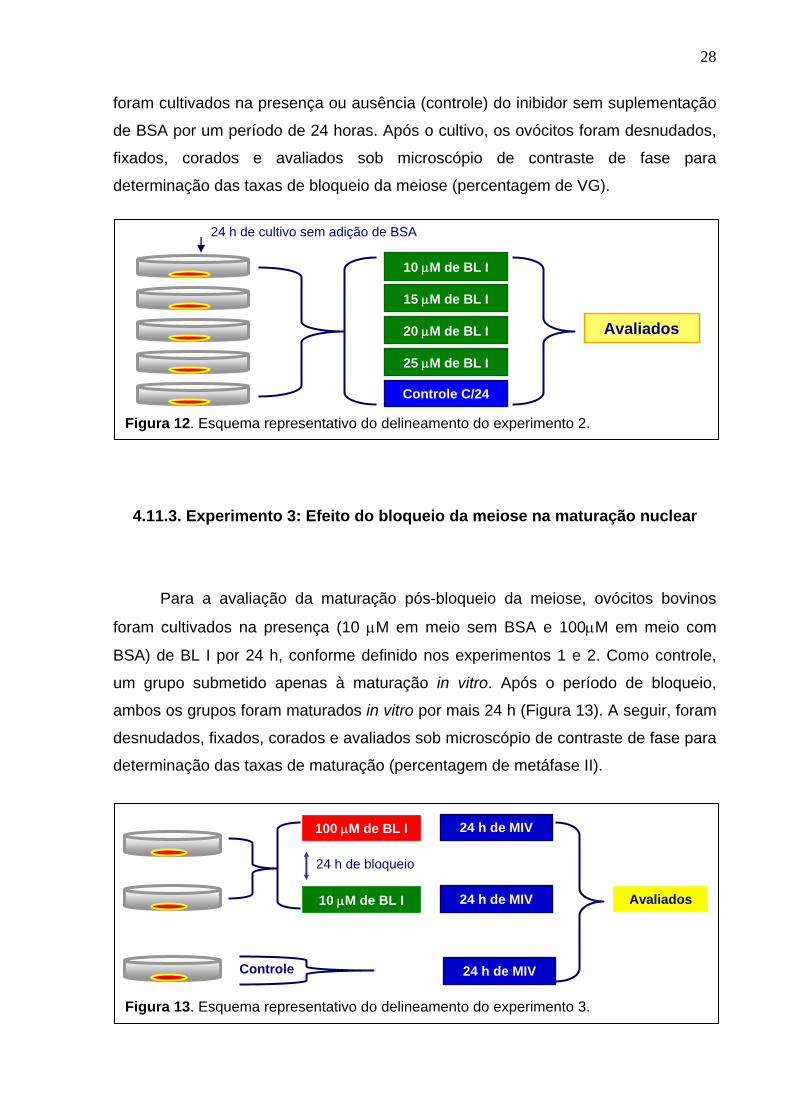
28
foram cultivados na presença ou ausência (controle) do inibidor sem suplementação
de BSA por um período de 24 horas. Após o cultivo, os ovócitos foram desnudados,
fixados, corados e avaliados sob microscópio de contraste de fase para
determinação das taxas de bloqueio da meiose (percentagem de VG).
Figura 12. Esquema representativo do delineamento do experimento 2.
10 μM de BL I
15 μM de BL I
20 μM de BL I
25 μM de BL I
Avaliados
Controle C/24
24 h de cultivo sem adição de BSA
4.11.3. Experimento 3: Efeito do bloqueio da meiose na maturação nuclear
Para a avaliação da maturação pós-bloqueio da meiose, ovócitos bovinos
foram cultivados na presença (10 μM em meio sem BSA e 100μM em meio com
BSA) de BL I por 24 h, conforme definido nos experimentos 1 e 2. Como controle,
um grupo submetido apenas à maturação in vitro. Após o período de bloqueio,
ambos os grupos foram maturados in vitro por mais 24 h (Figura 13). A seguir, foram
desnudados, fixados, corados e avaliados sob microscópio de contraste de fase para
determinação das taxas de maturação (percentagem de metáfase II).
Figura 13. Esquema representativo do delineamento do experimento 3.
10 μM de BL I
100 μM de BL I
Avaliados
24 h de bloqueio
24 h de MIV
24 h de MIV
24 h de MIV
Controle

29
4.11.4. Experimento 4: Cinética da maturação nuclear pós-bloqueio da meiose
Nesse experimento, os ovócitos foram bloqueados por um período de 12
horas na presença (100 μM de BL I) ou na ausência (10 μM de BL I) de BSA e
avaliados ao longo da MIV, nos horários de 6, 12 e 18 horas de maturação (Figura
14). Os períodos de bloqueio da meiose e da maturação (reversão) foram baseados
no estudo feito por ADONA e LEAL (2006). Após cada período de maturação (6, 12
e 18 horas), os ovócitos foram desnudados, fixados, corados e avaliados sob
microscópio de contraste de fase para determinação do estádio de maturação
nuclear. Um grupo de ovócitos submetido somente à MIV também foi avaliado nos
mesmos períodos dos tratados com BL I.
Horários das avaliações Figura 14. Esquema representativo do delineamento do experimento 4.
12 horas de bloqueio
10 μM de BL I
100 μM de BL I
12 h de MIV
18 h de MIV
MIV
Controle
Avaliações
CCiinnééttiiccaa ddaa mmaattuurraaççããoo ppóóss--bbllooqquueeiioo ddaa mmeeiioossee
6 h de MIV
4.11.5. Experimento 5: Efeito do bloqueio da meiose na organização dos microtúbulos e dos microfilamentos
Nesse experimento foram avaliados os efeitos do bloqueio meiótico em
condições de baixa e alta concentração de BL I (definidos nos experimentos 1 e 2)
na organização dos microtúbulos e dos microfilamentos. Os ovócitos foram
bloqueados por 24 horas e posteriormente cultivados em meio de maturação.
Ovócitos foram coletados às 0, 6, 12, 18 e 24 horas de pós-bloqueio para a
avaliação da organização dos microtúbulos, microfilamentos. A progressão meiótica
foi avaliada concomitantemente (Figura 15). O controle foi avaliado nos mesmos

30
horários ao longo da MIV sem ter sofrido inibição prévia. Um grupo de ovócitos foi
avaliado logo após a aspiração (controle 0h) e um outro no final das 24 horas de
bloqueio (antes do início da MIV – controle 24h).
Figura 15. Esquema representativo do delineamento do experimento 5.
24 horas de bloqueio
10 μM de BL I
100 μM de BL I
6 h de MIV
12 h de MIV
18 h de MIV
MIV
Controle
Avaliações
Horários das avaliações
0 h de MIV
24 h de MIV
Avaliação dos microtúbulos e dos microfilamentos
4.11.6. Experimentos 6: Efeito do bloqueio da meiose na distribuição dos grânulos corticais e das mitocôndrias.
Nesse experimento foi avaliado o efeito da BL I na distribuição dos grânulos
corticais e das mitocôndrias nos ovócitos tratados com duas concentrações de BL I,
uma na ausência (10 μM) e outra na presença (100 μM) de BSA. Os ovócitos foram
bloqueados por 24 horas e após esse período de bloqueio foram divididos em dois
grupos, uns para avaliação imediata (24 horas de bloqueio) e os demais cultivados
em meio de maturação e coletados somente depois de 24 horas de MIV (Figura 16),
para a avaliação do efeito na distribuição dos grânulos corticais e das mitocôndrias
nos ovócitos tratados com o inibidor do ciclo celular. No grupo controle, a avaliação
foi feita em dois momentos, um logo após a aspiração (zero hora) e uma outra no
final da maturação (24 horas de MIV).
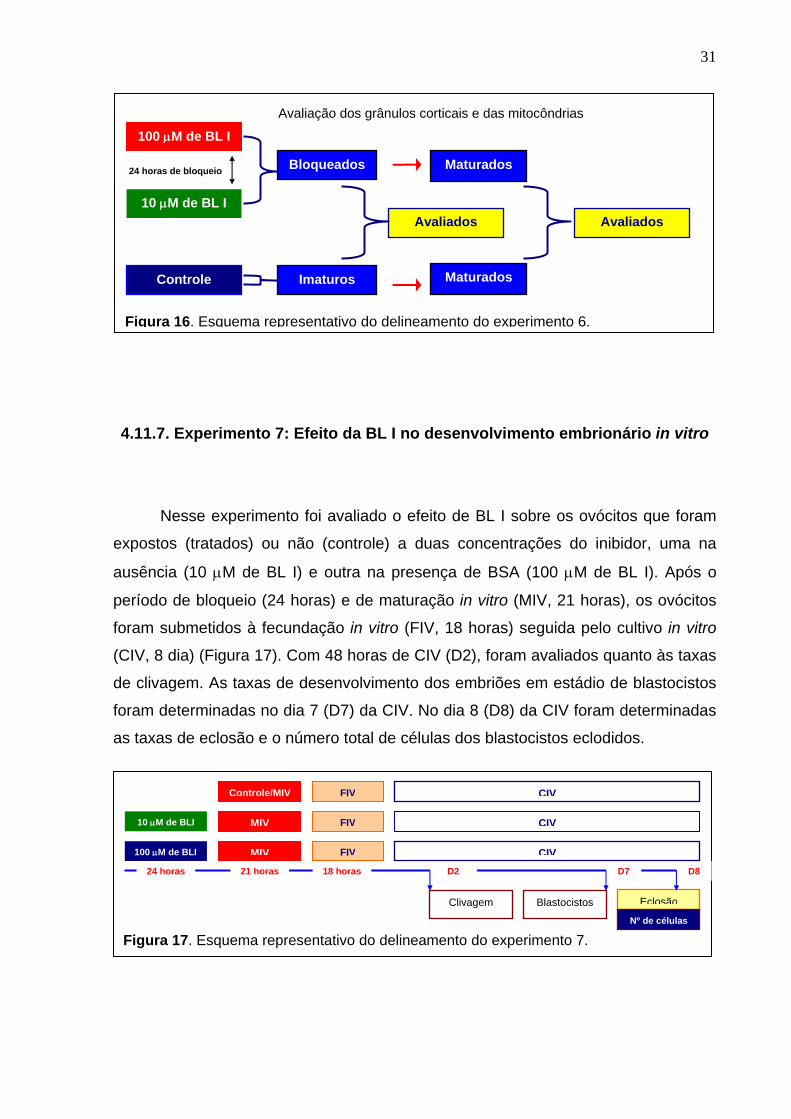
31
Figura 16. Esquema representativo do delineamento do experimento 6.
24 horas de bloqueio
Controle
10 μM de BL I
100 μM de BL I
Avaliados
Bloqueados
Imaturos
Maturados
Avaliados
Maturados
Avaliação dos grânulos corticais e das mitocôndrias
4.11.7. Experimento 7: Efeito da BL I no desenvolvimento embrionário in vitro
Nesse experimento foi avaliado o efeito de BL I sobre os ovócitos que foram
expostos (tratados) ou não (controle) a duas concentrações do inibidor, uma na
ausência (10 μM de BL I) e outra na presença de BSA (100 μM de BL I). Após o
período de bloqueio (24 horas) e de maturação in vitro (MIV, 21 horas), os ovócitos
foram submetidos à fecundação in vitro (FIV, 18 horas) seguida pelo cultivo in vitro
(CIV, 8 dia) (Figura 17). Com 48 horas de CIV (D2), foram avaliados quanto às taxas
de clivagem. As taxas de desenvolvimento dos embriões em estádio de blastocistos
foram determinadas no dia 7 (D7) da CIV. No dia 8 (D8) da CIV foram determinadas
as taxas de eclosão e o número total de células dos blastocistos eclodidos.
Figura 17. Esquema representativo do delineamento do experimento 7.
Eclosão
Nº de células
Clivagem Blastocistos
Controle/MIV FIV CIV
24 horas 21 horas 18 horas D2 D7 D8
MIV CIVFIV10 μM de BLI
MIV FIV CIV100 μM de BLI

32
4.12. Análise estatística
Os dados foram avaliados utilizando metodologia de quadrados mínimos, por
meio do procedimento PROC GLM do programa Statistical Analysis System, versão
9.1.3 (SAS, 1995). As variáveis foram transformadas, de acordo a função arco seno
(raiz - percentagens), de acordo com as recomendações de Banzatto e Kronca
(1989). Os dados dos números de células dos blastocistos não foram transformados,
sendo analisados em escala original. Para os resultados significativos nas análises
de variância foi utilizado o teste t de Student como procedimento de comparações
múltiplas entre os grupos. O nível de significância foi de 5% para evidenciar
diferenças entre os tratamentos.

33
5. RESULTADOS
5.1. Experimento 1
Nesse experimento foram avaliadas diferentes concentrações de butirolactona
I (BL I) em meio de cultivo B199 suplementado com albumina sérica bovina (BSA),
visando determinar a concentração mais adequada em estudos de bloqueio meiótico
em ovócitos bovinos.
A maioria dos ovócitos tratados nas concentrações de 100 (B100), 50 (B50) e
25 µM (B25) de BL I permaneceram bloqueados na meiose em estádio de vesícula
germinativa (VG), nas proporções de 97,4; 84,4 e 65,1%; respectivamente. No grupo
controle após 24 h de cultivo (C/24) não foram observados ovócitos em estádio de
VG (0%). Dos ovócitos avaliados logo após a aspiração folicular (controle zero hora
– C/0), 100% dos mesmos encontravam-se em estádio VG (Tabela 3). Os grupos
C/24 (0%), B100 (97,4%), B50 (84,4%) e B25 (65,1%) diferiram entre si (P < 0,05)
para o percentual de ovócitos em VG, com as taxas de bloqueio meiótico diminuindo
com a redução da concentração do inibidor. O grupo B100 (97,4%) e o C/0 (100%)
por outro lado, foram similares entre si (P > 0,05).
Foram observados ovócitos nas fases intermediárias da meiose (MAT,
metáfase I, anáfase I e telófase I), somente no grupo C/24, no demais tratamentos
os ovócitos estavam em VG ou MII (metáfase II). A taxa de ovócitos em MAT do
grupo C/24 (10,4%) diferiu (P < 0,05) dos demais tratamentos (0%).
Com relação aos percentuais de ovócitos que atingiram o estádio de MII, ou
seja, que concluíram a maturação in vitro (MIV), houve diferença (P < 0,05) nas
taxas de MII entre os ovócitos dos grupos C/24 (89,6%), B100 (2,6%), B50 (15,6%) e

34
B25 (34,9%) após 24 horas de cultivo, com a redução da taxa de MII com o aumento
da concentração de BL I. Não houve diferença (P > 0,05) entre os grupos C/0 (0%) e
B100 (2,6%) para o percentual de ovócitos em estádio de MII.
Tabela 3. Estádios da meiose de ovócitos bovinos submetidos ao bloqueio meiótico com concentrações decrescentes de BL I (100, 50 e 25 µM) em meio de cultivo B199 suplementado com BSA.
Estádios da meiose
Tratamentos* Ovócitos VG1
Nº (%) MAT2
Nº (%) MII3
Nº (%) C/0 75 75 (100)a 0 (0,0)b 0 (0,0)d
C/24 77 0 (0,0)d 8 (10,4 ± 1,4)a 69 (89,6 ± 1,4)a
B100 78 76 (97,4 ± 3,9)a 0 (0,0)b 2 (2,6 ± 3,9)d
B50 77 65 (84,4 ± 1,8)b 0 (0,0)b 12 (15,6 ± 1,8)c
B25 83 54 (65,1 ± 4,0)c 0 (0,0)b 29 (34,9 ± 4,0)b
1Vesícula germinativa (VG); 2metáfase I, anáfase I e telófase I (MAT); 3metáfase II (MII); *controle 0 hora (C/0), ovócitos avaliados logo após a aspiração folicular; controle 24 horas (C/24), ovócitos cultivados por 24 horas em meio B199 suplementado com BSA; tratados (B25, B50 e B100), ovócitos cultivados por 24 horas com 25, 50 e 100 μM de BL I em meio B199 suplementado com BSA. Resultado de três réplicas. a-d Letras diferentes na mesma coluna indicam diferença entre tratamentos (P < 0,05) e o símbolo ± refere-se ao desvio padrão da média.
5.2. Experimento 2
No experimento anterior não foi possível reduzir a concentração de BL I sem
perder a eficiência do bloqueio. Então, procurou-se reduzir a concentração de BL I
em meio de cultivo sem suplementação de BSA, verificando se o bloqueio nessa
condição de cultivo seria eficiente na manutenção da meiose bloqueada. Após 24
horas de bloqueio (sem adição de BSA) a maioria dos ovócitos tratados nas
concentrações de 10 (B10), 15 (B15), 20 (B20) e 25 µM (B25) de BL I
permaneceram em estádio de VG. Houve diferença (P < 0,05) do grupo C/24
(13,7%) em relação aos tratados com B10 (94,9%), B15 (97,2%), B20 (98,7%) e B25
(98,7%) para o percentual de ovócitos em estádio de VG (Tabela 4), os quais não
diferiram entre si (P > 0,05). Após 24 horas de cultivo foram observados ovócitos nos
estádios de MAT, somente no grupo C/24 (11,0%, P 0 < 0,05).

35
Os percentuais de ovócitos que atingiram o estádio de MII após 24 horas de
cultivo diferiram (P < 0,05) entre o grupo C/24 (75,3%) e os grupos B10 (5,1%), B15
(2,8%), B20 (1,3%) e B25 (1,3%), que não diferiram entre si (P > 0,05).
Pode-se observar que na ausência de BSA, concentrações mais baixas de BL
I (entre 10 e 25 µM) são igualmente eficientes no bloqueio da meiose de ovócitos
bovinos cultivados in vitro.
Tabela 4. Estádios da meiose de ovócitos bovinos submetidos ao bloqueio meiótico com diferentes concentrações de BL I (10, 15, 20 e 25 µM) em meio de cultivo B199 sem suplementação de BSA.
Estádios da meiose
Tratamentos* Ovócitos VG1
Nº (%) MAT2
Nº (%) MII3
Nº (%) C/24 73 10 (13,7 ± 2,4)b 8 (11,0 ± 3,5)a 55 (75,3 ± 1,1)a
B10 78 74 (94,9 ± 2,6)a 0 (0,0)b 4 (5,1 ± 2,6)b
B15 72 70 (97,2 ± 2,3)a 0 (0,0)b 2 (2,8 ± 2,3)b
B20 76 75 (98,7 ± 2,2)a 0 (0,0)b 1 (1,3 ± 2,2)b
B25 76 75 (98,7 ± 2,4)a 0 (0,0)b 1 (1,3 ± 2,4)b
1Vesícula germinativa (VG); 2metáfase I, anáfase I e telófase I (MAT); 3metáfase II (MII); *controle 24 h (C/24), ovócitos cultivados por 24 h em meio B199 sem suplementação de BSA; tratados (B10, B15, B20 e B25), ovócitos cultivados por 24 h com 10, 15, 20 e 25 μM de BL I em meio B199 sem suplementação de BSA. Resultado de três réplicas. a-b Letras diferentes na mesma coluna indicam diferença entre tratamentos (P < 0,05) e o símbolo ± refere-se ao desvio padrão da média.
5.3. Experimento 3
Como foi possível bloquear a meiose dos ovócitos bovinos com eficiência, em
meio de bloqueio na presença e na ausência de BSA, fez-se necessário averiguar se
a reversibilidade da meiose nos ovócitos (maturação) seria afetada pelo bloqueio
meiótico nas duas condições de cultivo.
Os ovócitos foram bloqueados por 24 h nas concentrações de 10 µM (B10) e
100 µM BL I (B100) em meio de cultivo suplementado (B100) ou não (B10) com BSA
e maturados em meio de MIV por igual período. Um pequeno percentual dos
ovócitos permaneceu nos estádios de VG (3,3; 0,0 e 1,1%) e de MAT (5,6; 8,5 e
6,5%) após 24 h de MIV (P > 0,05) nos grupos C/24, B10 e B100, respectivamente.
O percentual de maturação nuclear (taxa de metáfase II) foi de 91,0; 91,4 e 92,3%

36
para os grupos C/24, B10 e B100, respectivamente. As taxas de VG, MAT e MII não
diferiram (P > 0,05) entre os tratamentos após 24 h de MIV (Tabela 5).
O uso de BL I em baixa concentração (10 µM) na ausência de BSA não reduz
a taxa de MIV (MII) pós-bloqueio da meiose, indicando ser tão eficaz quanto a
concentração usual de 100 µM em meio suplementado com BSA, ao menos no que
se refere a esse parâmetro.
Tabela 5. Estádios da meiose de ovócitos bovinos submetidos ao bloqueio meiótico nas concentrações de 10 e 100 µM de BL I em meio de cultivo B199 suplementado ou não com BSA.
Estádios da meiose
Tratamentos* Ovócitos VG1
Nº (%) MAT2
Nº (%) MII3
Nº (%) C/24 90 3 (3,3 ± 0,4) 5 (5,6 ± 2,3) 82 (91,1 ± 2,6)
B10 94 0 (0,0) 8 (8,5 ± 2,1) 86 (91,5 ± 2,1)
B100 92 1 (1,1 ± 1,9) 6 (6,5 ± 3,1) 85 (92,4 ± 1,8)
1Vesícula germinativa (VG); 2metáfase I, anáfase I e telófase I (MAT); 3metáfase II (MII); *controle 24 horas (C/24), ovócitos cultivados por 24 horas em meio de maturação in vitro; tratados (B10 e B100), ovócitos cultivados por 24 h com 10 e 100 μM de BL I em meio B199 suplementado (B100) ou não (B10) com BSA seguidos de MIV por 24 h. Resultado de três réplicas. Não houve diferença entre tratamentos (P > 0,05) e o símbolo ± refere-se ao desvio padrão da média.
5.4. Experimento 4
Uma vez determinados nos experimentos anteriores que o bloqueio meiótico
com BL I na presença e na ausência de BSA e a reversibilidade dos mesmos foi
eficiente, fez-se necessário fazer um acompanhamento mais detalhado da
maturação nuclear para averiguar um possível efeito do bloqueio sobre a progressão
meiótica (cinética) durante a maturação in vitro.
Os ovócitos foram bloqueados por 12 h com B10 e B100 e transferidos para o
meio de MIV e avaliados em diferentes horários (6, 12 e 18 h) de MIV. O grupo
controle foi avaliado nos mesmos horários sem ser submetido ao bloqueio com BL I.
As taxas de VG observadas com 6 h de MIV pós-bloqueio (Tabela 6) nos
grupos controle (97,3%) e tratados com B10 (71,4%) e B100 (74,3%) apresentaram
diferença (P < 0,05) entre o controle e os tratados. O percentual de MAT às 6 h de
MIV também apresentou diferença (P < 0,05) somente entre o controle (2,7%) e os
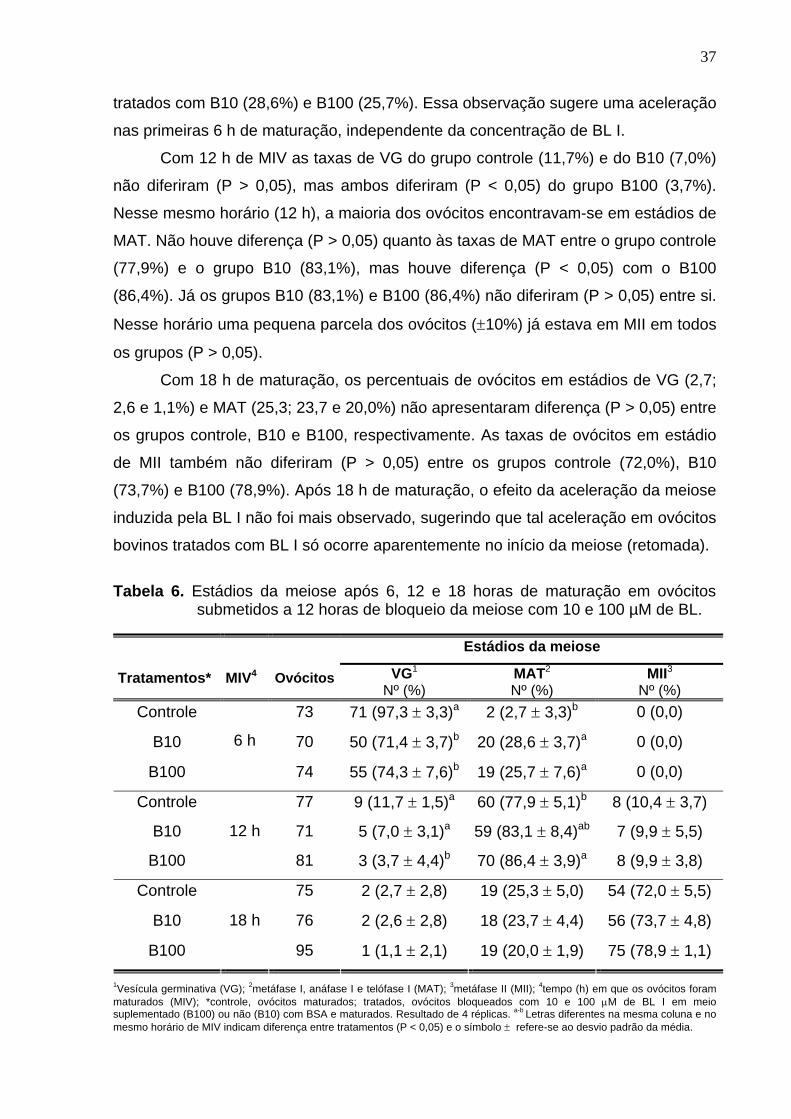
37
tratados com B10 (28,6%) e B100 (25,7%). Essa observação sugere uma aceleração
nas primeiras 6 h de maturação, independente da concentração de BL I.
Com 12 h de MIV as taxas de VG do grupo controle (11,7%) e do B10 (7,0%)
não diferiram (P > 0,05), mas ambos diferiram (P < 0,05) do grupo B100 (3,7%).
Nesse mesmo horário (12 h), a maioria dos ovócitos encontravam-se em estádios de
MAT. Não houve diferença (P > 0,05) quanto às taxas de MAT entre o grupo controle
(77,9%) e o grupo B10 (83,1%), mas houve diferença (P < 0,05) com o B100
(86,4%). Já os grupos B10 (83,1%) e B100 (86,4%) não diferiram (P > 0,05) entre si.
Nesse horário uma pequena parcela dos ovócitos (±10%) já estava em MII em todos
os grupos (P > 0,05).
Com 18 h de maturação, os percentuais de ovócitos em estádios de VG (2,7;
2,6 e 1,1%) e MAT (25,3; 23,7 e 20,0%) não apresentaram diferença (P > 0,05) entre
os grupos controle, B10 e B100, respectivamente. As taxas de ovócitos em estádio
de MII também não diferiram (P > 0,05) entre os grupos controle (72,0%), B10
(73,7%) e B100 (78,9%). Após 18 h de maturação, o efeito da aceleração da meiose
induzida pela BL I não foi mais observado, sugerindo que tal aceleração em ovócitos
bovinos tratados com BL I só ocorre aparentemente no início da meiose (retomada).
Tabela 6. Estádios da meiose após 6, 12 e 18 horas de maturação em ovócitos
submetidos a 12 horas de bloqueio da meiose com 10 e 100 µM de BL.
Estádios da meiose
Tratamentos* MIV4 Ovócitos VG1
Nº (%) MAT2
Nº (%) MII3
Nº (%) Controle 73 71 (97,3 ± 3,3)a 2 (2,7 ± 3,3)b 0 (0,0)
B10 70 50 (71,4 ± 3,7)b 20 (28,6 ± 3,7)a 0 (0,0)
B100
6 h
74 55 (74,3 ± 7,6)b 19 (25,7 ± 7,6)a 0 (0,0)
Controle 77 9 (11,7 ± 1,5)a 60 (77,9 ± 5,1)b 8 (10,4 ± 3,7)
B10 71 5 (7,0 ± 3,1)a 59 (83,1 ± 8,4)ab 7 (9,9 ± 5,5)
B100
12 h
81 3 (3,7 ± 4,4)b 70 (86,4 ± 3,9)a 8 (9,9 ± 3,8)
Controle 75 2 (2,7 ± 2,8) 19 (25,3 ± 5,0) 54 (72,0 ± 5,5)
B10 76 2 (2,6 ± 2,8) 18 (23,7 ± 4,4) 56 (73,7 ± 4,8)
B100
18 h
95 1 (1,1 ± 2,1) 19 (20,0 ± 1,9) 75 (78,9 ± 1,1)
1Vesícula germinativa (VG); 2metáfase I, anáfase I e telófase I (MAT); 3metáfase II (MII); 4tempo (h) em que os ovócitos foram maturados (MIV); *controle, ovócitos maturados; tratados, ovócitos bloqueados com 10 e 100 μM de BL I em meio suplementado (B100) ou não (B10) com BSA e maturados. Resultado de 4 réplicas. a-b Letras diferentes na mesma coluna e no mesmo horário de MIV indicam diferença entre tratamentos (P < 0,05) e o símbolo ± refere-se ao desvio padrão da média.

38
5.5. Experimento 5
Nesse experimento foram avaliadas a configuração da maturação nuclear,
organização dos microtúbulos e a disposição dos microfilamentos nos ovócitos
submetidos ao bloqueio meiótico com B10 e B100. Os ovócitos foram bloqueados
por 24 horas, transferidos para o meio de maturação (MIV) e avaliados em diferentes
horários de MIV (0, 6, 12, 18 e 24 horas). O grupo controle foi avaliado nos mesmos
horários sem ser submetido a uma pré-maturação com BL I.
Os resultados da configuração da maturação nuclear não serão descritos aqui
por que seguiram o mesmo padrão observado na cinética da maturação nuclear pós-
bloqueio. As observações feitas sugerem uma aceleração no início da retomada da
meiose nos ovócitos tratados com BL I, mas no decorrer da MIV essa aceleração
não foi mais evidenciada, confirmando os resultados do experimento 4.
A marcação positiva dos microtúbulos foi observada por todo o citoplasma
dos ovócitos em todos os grupos e horários. Uma marcação mais intensa dos
microtúbulos foi observada em MAT e MII junto aos cromossomos (fuso meiótico)
nos ovócitos que haviam iniciado o rompimento da VG (Figura 18). Esse mesmo
padrão foi observado em todos os tratamentos (controle, B10 e B100) e horários (6,
12, 18 e 24 h). Não foram observadas alterações no fuso meiótico quanto à
organização e formação da placa metafásica nos diferentes grupos de ovócitos e
horários dos tratamentos. Dos 653 ovócitos avaliados nesse experimento apenas um
ovócito tratado com B100 após 24 horas de maturação pós-bloqueio apresentou
alteração na placa metafásica, o que é importante para demonstrar a segurança e a
efetividade da droga.
Com relação aos microfilamentos, observou-se uma marcação por todo o
citoplasma dos ovócitos em todos os grupos e horários (Figura 19). Uma marcação
mais intensa dos microfilamentos foi observada na região onde o corpúsculo polar foi
liberado e no córtex celular, nos horários de 18 e 24 horas maturação, para todos os
grupos (controle, B10 e B100). Não foi observada nenhuma diferença ou alteração
no arranjo dos microfilamentos no citoplasma dos ovócitos avaliados nos diferentes
grupos e horários de maturação, indicando a segurança na utilização dessa droga.
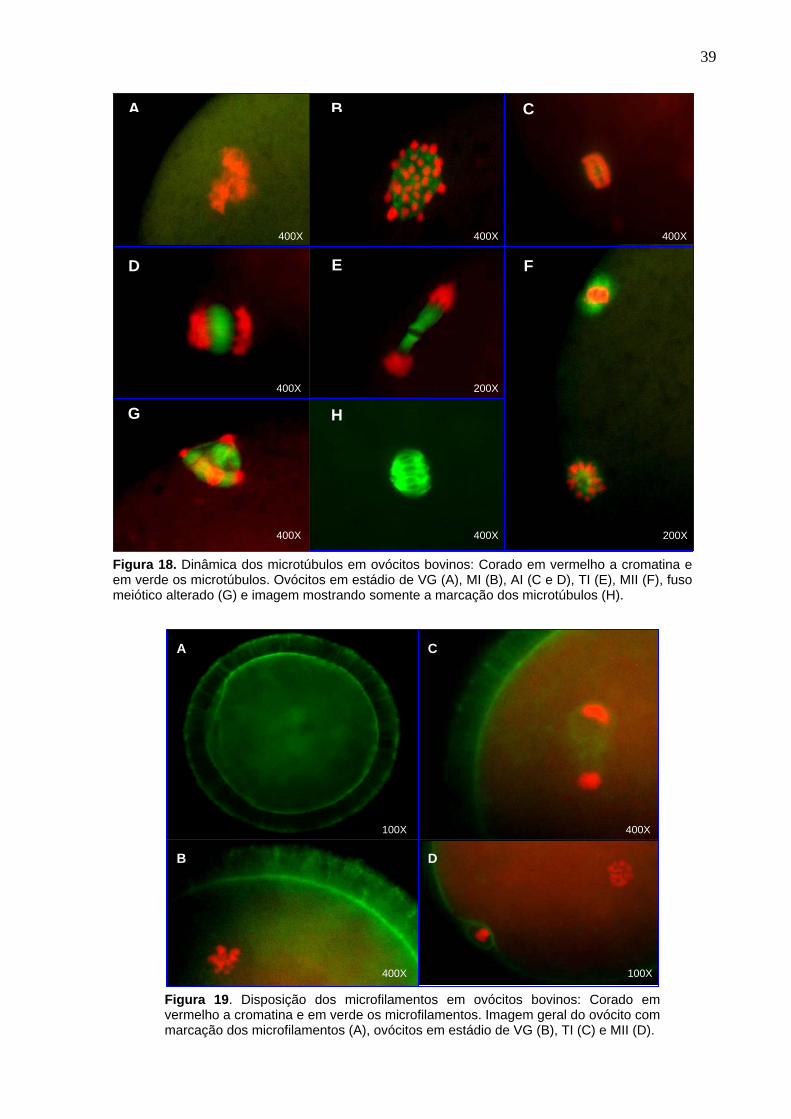
39
A B
400X
400X 400X
200X
400X
400X
200X
C
D F
G
400X
E
H
Figura 18. Dinâmica dos microtúbulos em ovócitos bovinos: Corado em vermelho a cromatina e em verde os microtúbulos. Ovócitos em estádio de VG (A), MI (B), AI (C e D), TI (E), MII (F), fuso meiótico alterado (G) e imagem mostrando somente a marcação dos microtúbulos (H).
A
100X
400X
C
B D
100X
400X
Figura 19. Disposição dos microfilamentos em ovócitos bovinos: Corado em vermelho a cromatina e em verde os microfilamentos. Imagem geral do ovócito com marcação dos microfilamentos (A), ovócitos em estádio de VG (B), TI (C) e MII (D).

40
5.6. Experimento 6
A configuração da maturação nuclear, migração dos grânulos corticais e
mitocôndrias foram avaliadas nos ovócitos submetidos ao bloqueio da meiose com
BL I e nos ovócitos maturados pós-bloqueio da meiose. Os ovócitos do grupo
controle (não bloqueados) foram avaliados logo após a aspiração folicular e após a
MIV.
As taxas de ovócitos em estádio de VG para os grupos C/0, B10 e B100
foram de ~98,4%. Os percentuais de ovócitos em estádio de VG não diferiram (P >
0,05) entre os grupos de ovócitos avaliados. Após 24 h de MIV ~96,6% (P > 0,05)
ovócitos encontravam-se no estádio de MII para os grupos C/24, B10 e B100.
Em 100% dos ovócitos imaturos (não maturados) independente dos grupos
(C/0, B10 e B100, P > 0,05), os clusters de grânulos corticais apresentavam-se
dispersos pelo citoplasma dos ovócitos (distribuição geral). Mas, após 24 horas de
maturação, também independente dos grupos, 98% (P > 0,05) dos ovócitos
apresentavam migração dos grânulos corticais para a região periférica do citoplasma
(distribuição periférica) (Tabela 7, Figura 20).
As mitocôndrias encontravam-se em sua maioria na região periférica do
citoplasma nos ovócitos imaturos. No entanto, nos grupos tratados a proporção (81,5
e 86,9%, P > 0,05) de ovócitos com mitocôndrias localizadas na região mais
periférica do citoplasma foi inferior (P < 0,05) ao controle (100%). Após 24 horas de
maturação, as mitocôndrias migraram por todo o citoplasma do ovócito. No grupo
C/24 (81,5%) a migração das mitocôndrias foi inferior (P < 0,05) aos grupos B10M
(95,2) e B100M (98,2%) que não diferiram (P > 0,05) entre si (Tabela 8, Figura 21).
De acordo com os resultados, sugere-se que embora o ciclo celular meiótico
estivesse bloqueado com BL I, o que também bloqueou a migração dos grânulos
corticais, a migração das mitocôndrias não foi totalmente bloqueada. No entanto, o
bloqueio não interfere na migração posterior dos grânulos corticais e das
mitocôndrias quando os ovócitos são submetidos à maturação in vitro.
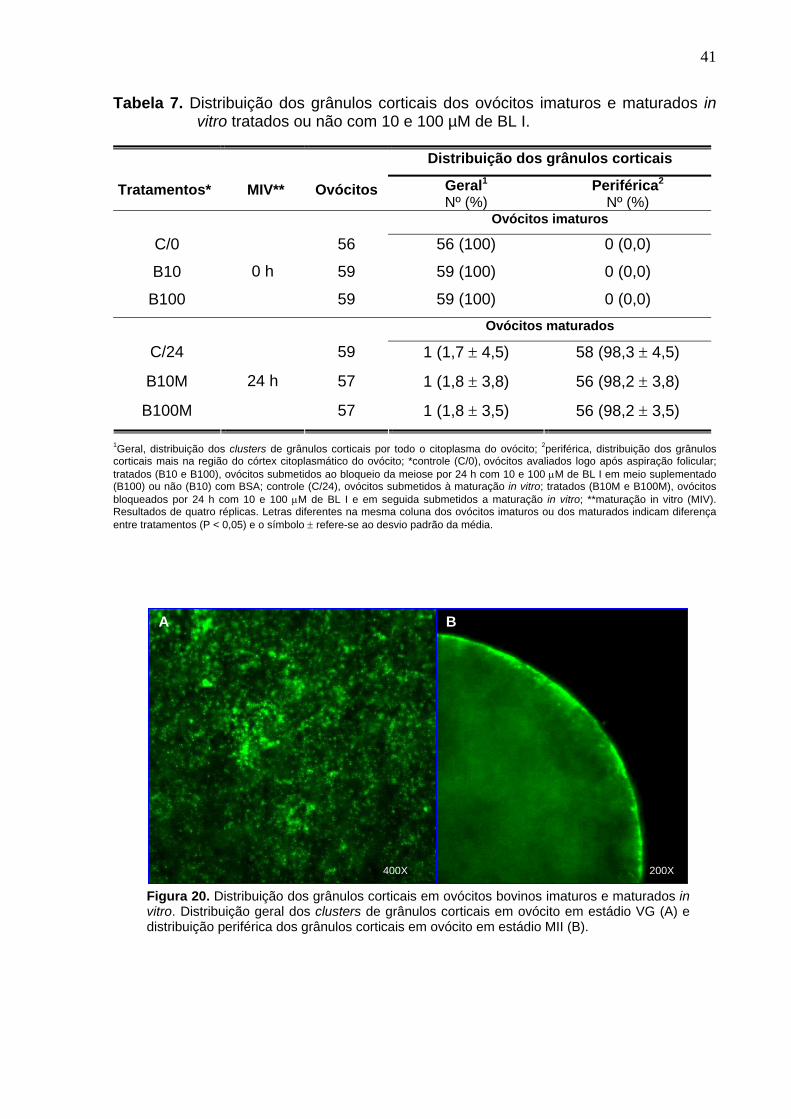
41
Tabela 7. Distribuição dos grânulos corticais dos ovócitos imaturos e maturados in vitro tratados ou não com 10 e 100 µM de BL I.
Distribuição dos grânulos corticais
Tratamentos* MIV** Ovócitos Geral1
Nº (%) Periférica2
Nº (%) Ovócitos imaturos
C/0 56 56 (100) 0 (0,0)
B10 59 59 (100) 0 (0,0)
B100
0 h
59 59 (100) 0 (0,0)
Ovócitos maturados
C/24 59 1 (1,7 ± 4,5) 58 (98,3 ± 4,5)
B10M 57 1 (1,8 ± 3,8) 56 (98,2 ± 3,8)
B100M
24 h
57 1 (1,8 ± 3,5) 56 (98,2 ± 3,5)
1Geral, distribuição dos clusters de grânulos corticais por todo o citoplasma do ovócito; 2periférica, distribuição dos grânulos corticais mais na região do córtex citoplasmático do ovócito; *controle (C/0), ovócitos avaliados logo após aspiração folicular; tratados (B10 e B100), ovócitos submetidos ao bloqueio da meiose por 24 h com 10 e 100 μM de BL I em meio suplementado (B100) ou não (B10) com BSA; controle (C/24), ovócitos submetidos à maturação in vitro; tratados (B10M e B100M), ovócitos bloqueados por 24 h com 10 e 100 μM de BL I e em seguida submetidos a maturação in vitro; **maturação in vitro (MIV). Resultados de quatro réplicas. Letras diferentes na mesma coluna dos ovócitos imaturos ou dos maturados indicam diferença entre tratamentos (P < 0,05) e o símbolo ± refere-se ao desvio padrão da média.
200X400X
A B Figura 20. Distribuição dos grânulos corticais em ovócitos bovinos imaturos e maturados in
vitro. Distribuição geral dos clusters de grânulos corticais em ovócito em estádio VG (A) e distribuição periférica dos grânulos corticais em ovócito em estádio MII (B).
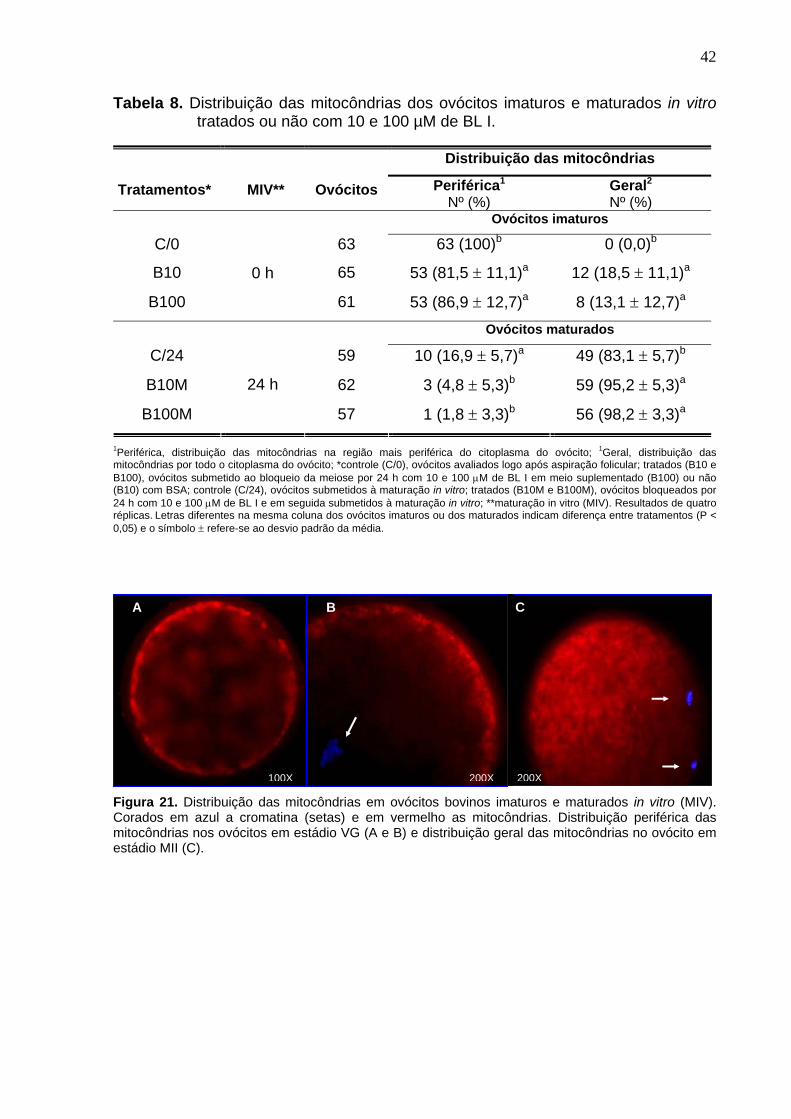
42
Tabela 8. Distribuição das mitocôndrias dos ovócitos imaturos e maturados in vitro tratados ou não com 10 e 100 µM de BL I.
Distribuição das mitocôndrias
Tratamentos* MIV** Ovócitos Periférica1
Nº (%) Geral2
Nº (%) Ovócitos imaturos
C/0 63 63 (100)b 0 (0,0)b
B10 65 53 (81,5 ± 11,1)a 12 (18,5 ± 11,1)a
B100
0 h
61 53 (86,9 ± 12,7)a 8 (13,1 ± 12,7)a
Ovócitos maturados
C/24 59 10 (16,9 ± 5,7)a 49 (83,1 ± 5,7)b
B10M 62 3 (4,8 ± 5,3)b 59 (95,2 ± 5,3)a
B100M
24 h
57 1 (1,8 ± 3,3)b 56 (98,2 ± 3,3)a
1Periférica, distribuição das mitocôndrias na região mais periférica do citoplasma do ovócito; 1Geral, distribuição das mitocôndrias por todo o citoplasma do ovócito; *controle (C/0), ovócitos avaliados logo após aspiração folicular; tratados (B10 e B100), ovócitos submetido ao bloqueio da meiose por 24 h com 10 e 100 μM de BL I em meio suplementado (B100) ou não (B10) com BSA; controle (C/24), ovócitos submetidos à maturação in vitro; tratados (B10M e B100M), ovócitos bloqueados por 24 h com 10 e 100 μM de BL I e em seguida submetidos à maturação in vitro; **maturação in vitro (MIV). Resultados de quatro réplicas. Letras diferentes na mesma coluna dos ovócitos imaturos ou dos maturados indicam diferença entre tratamentos (P < 0,05) e o símbolo ± refere-se ao desvio padrão da média.
200X
C
100X
A
200X
B
Figura 21. Distribuição das mitocôndrias em ovócitos bovinos imaturos e maturados in vitro (MIV). Corados em azul a cromatina (setas) e em vermelho as mitocôndrias. Distribuição periférica das mitocôndrias nos ovócitos em estádio VG (A e B) e distribuição geral das mitocôndrias no ovócito em estádio MII (C).
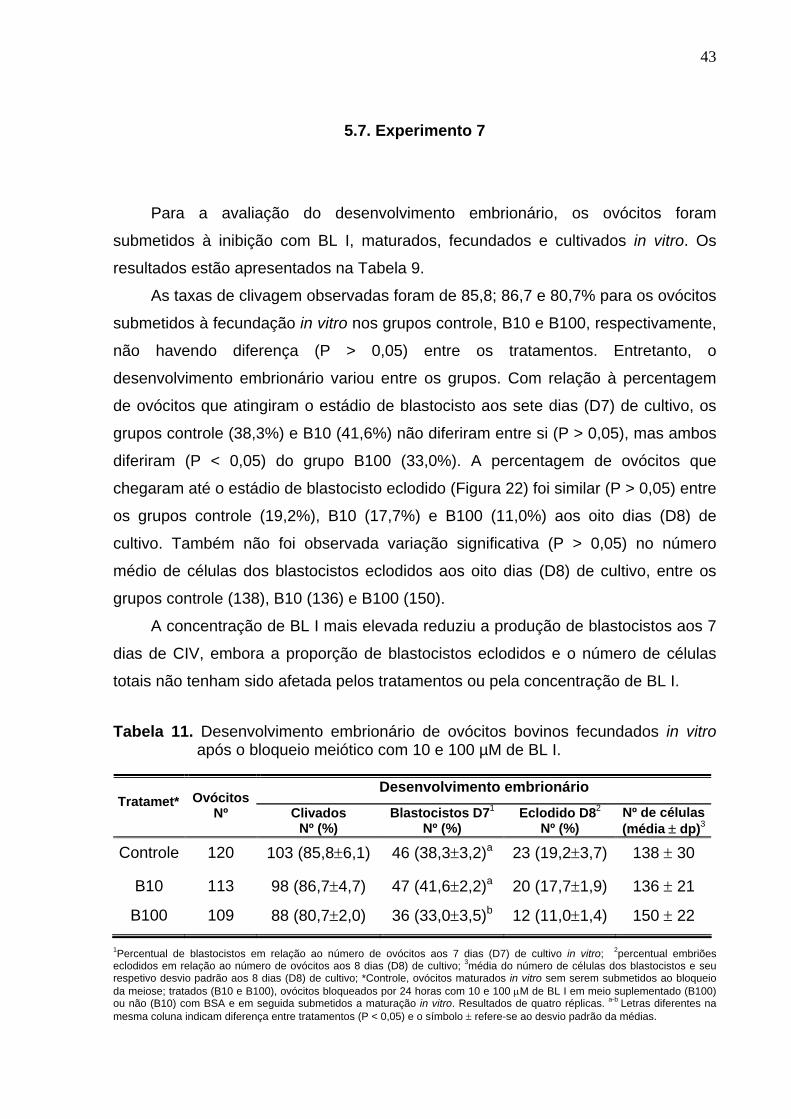
43
5.7. Experimento 7
Para a avaliação do desenvolvimento embrionário, os ovócitos foram
submetidos à inibição com BL I, maturados, fecundados e cultivados in vitro. Os
resultados estão apresentados na Tabela 9.
As taxas de clivagem observadas foram de 85,8; 86,7 e 80,7% para os ovócitos
submetidos à fecundação in vitro nos grupos controle, B10 e B100, respectivamente,
não havendo diferença (P > 0,05) entre os tratamentos. Entretanto, o
desenvolvimento embrionário variou entre os grupos. Com relação à percentagem
de ovócitos que atingiram o estádio de blastocisto aos sete dias (D7) de cultivo, os
grupos controle (38,3%) e B10 (41,6%) não diferiram entre si (P > 0,05), mas ambos
diferiram (P < 0,05) do grupo B100 (33,0%). A percentagem de ovócitos que
chegaram até o estádio de blastocisto eclodido (Figura 22) foi similar (P > 0,05) entre
os grupos controle (19,2%), B10 (17,7%) e B100 (11,0%) aos oito dias (D8) de
cultivo. Também não foi observada variação significativa (P > 0,05) no número
médio de células dos blastocistos eclodidos aos oito dias (D8) de cultivo, entre os
grupos controle (138), B10 (136) e B100 (150).
A concentração de BL I mais elevada reduziu a produção de blastocistos aos 7
dias de CIV, embora a proporção de blastocistos eclodidos e o número de células
totais não tenham sido afetada pelos tratamentos ou pela concentração de BL I.
Tabela 11. Desenvolvimento embrionário de ovócitos bovinos fecundados in vitro após o bloqueio meiótico com 10 e 100 µM de BL I.
Desenvolvimento embrionário
Tratamet* Ovócitos Nº Clivados
Nº (%) Blastocistos D71
Nº (%) Eclodido D82
Nº (%) Nº de células (média ± dp)3
Controle 120 103 (85,8±6,1) 46 (38,3±3,2)a 23 (19,2±3,7) 138 ± 30
B10 113 98 (86,7±4,7) 47 (41,6±2,2)a 20 (17,7±1,9) 136 ± 21
B100 109 88 (80,7±2,0) 36 (33,0±3,5)b 12 (11,0±1,4) 150 ± 22
1Percentual de blastocistos em relação ao número de ovócitos aos 7 dias (D7) de cultivo in vitro; 2percentual embriões eclodidos em relação ao número de ovócitos aos 8 dias (D8) de cultivo; 3média do número de células dos blastocistos e seu respetivo desvio padrão aos 8 dias (D8) de cultivo; *Controle, ovócitos maturados in vitro sem serem submetidos ao bloqueio da meiose; tratados (B10 e B100), ovócitos bloqueados por 24 horas com 10 e 100 μM de BL I em meio suplementado (B100) ou não (B10) com BSA e em seguida submetidos a maturação in vitro. Resultados de quatro réplicas. a-b Letras diferentes na mesma coluna indicam diferença entre tratamentos (P < 0,05) e o símbolo ± refere-se ao desvio padrão da médias.
44
Figura 22. Blastocisto eclodido em D8 corado com Hoechst para visualização dos núcleos para contagem de células.
100X

45
6. DISCUSSÃO
O bloqueio meiótico visa manter os ovócitos em estádio de vesícula
germinativa (VG) para que os mesmos tenham um tempo adicional para sofrerem
as modificações que ocorrem antes da maturação propriamente dita (HYTTEL et al.,
1997) e adquirir competência para o desenvolvimento (KUBELKA et al., 2000;
MERMILLOD et al., 2000).
Um dos tratamentos usualmente utilizados é o cultivo pré-maturação na
presença de inibidores de cinases dependentes de ciclinas (MERMILLOD et al.,
2000; PONDERATO et al., 2001; KUBELKA et al., 2002), o que, no entanto pode
causar danos ultra-estruturais (FAERGE et al., 2001; FAIR et al., 2002; LONERGAN
et al., 2003). Tal tratamento, embora não reduza (IMAI et al., 2002; BEKER-VAN
WOUDENBERG et al., 2005) e possa até aumentar o desenvolvimento a blastocisto
(HASHIMOTO et al., 2002; COY et al., 2005a), pode ter efeitos posteriores
desconhecidos.
O presente trabalho propôs reduzir a concentração de BL I usualmente
utilizada para indução de bloqueio meiótico (100 μM), no intuito de reduzir riscos de
alterações nos ovócitos e possivelmente aumentar o desenvolvimento embrionário.
No primeiro experimento foram avaliadas concentrações decrescentes de BL
I (100, 50 e 25 μM) e seu efeito na indução de bloqueio meiótico. Observou-se que
à medida que se reduzia a concentração do inibidor, reduzia-se também a eficiência
do bloqueio meiótico. KUBELKA et al. (2000) também observaram que a indução de
bloqueio meiótico com BL I é dependente de concentração do inibidor e que a

46
presença de BSA no meio faz com que a concentração necessária para induzir o
bloqueio seja maior do que aquela necessária com PVA. Os autores sugerem que a
BL I poderia se ligar ao BSA, reduzindo sua disponibilidade no meio.
Para poder reduzir a concentração de BL I, testou-se a utilização de meio
sem adição de BSA e concentrações ainda mais baixas de inibidor. Nesse
experimento, todas as concentrações foram igualmente eficientes em manter os
ovócitos em VG, demonstrando que de fato o BSA interfere na ação da droga. Na
sua ausência, uma concentração 10 vezes mais baixa (10 μM) pode ser usada para
manter um bloqueio meiótico eficaz, visto que 95% dos ovócitos permaneceram em
VG após 24h de cultivo. Esse valor é bastante próximo ao obtido com alta
concentração de BL I (100 μM) na presença de BSA (97%). WU et al. (2002)
utilizando ovócitos de suínos e uma concentração de 12,5 μM BL I, próxima da que
utilizamos, também obtiveram sucesso no bloqueio meiótico. PONDERATO et al.,
(2001) também conseguiram taxas similares de bloqueio usando uma mistura de BL
I e roscovitina (ROS) (12,5 e 6,25 μM, respectivamente) numa concentração total de
18,75 μM de inibidores no meio. Essa concentração, porém, é quase o dobro da
menor concentração utilizada no presente estudo.
Visto que as taxas de bloqueio de 100 μM BL em meio com BSA e 10 μM BL
I em meio sem BSA foram bastante próximas, fez-se necessário verificar se a MIV
subseqüente ao bloqueio também seria similar, uma vez que utilizando baixa
concentração de BL I, o cultivo era feito por 24 h sem adição de macromoléculas, o
que poderia prejudicar a maturação dos ovócitos. Nesse experimento, porém,
observou-se que as taxas de maturação nuclear (progressão à MII) foram similares
para os dois sistemas de bloqueio e também para os controles não bloqueados
antes da MIV (~90%). O uso da mistura de inibidores (BL I + ROS) em baixas
concentrações também não prejudicou a progressão meiótica (>90%) de ovócitos
bovinos (PONDERATO et al., 2001). Esses autores também excluíram o BSA do
meio de bloqueio. Em ovócitos de suínos, o uso de concentração similar (12,5 μM
em meio contendo PVA e EGF) também não afetou a taxa de MII (83,4%) (WU et
al., 2002). Embora não haja descrição do uso de concentração tão baixa de BL I em
ovócitos bovinos, os resultados mostram que esse tratamento pode ser utilizado
para bloquear eficientemente a meiose de forma totalmente reversível.
Alguns pesquisadores relatam que o uso de inibidores como a BL I e ROS
causam uma aceleração da progressão meiótica durante a MIV (LAGUTINA et al.,

47
2002; HASHIMOTO et al., 2002; ADONA e LEAL, 2004). No experimento seguinte
foi avaliada a cinética da maturação pós-bloqueio de ambos os sistemas de cultivo,
para averiguar se uma concentração mais baixa de BL I reduziria esse efeito.
Observou-se uma aceleração na retomada da meiose em ambos os tratamentos em
comparação com os controles não bloqueados, uma vez que às 6 h de MIV, a
proporção de ovócitos em VG (~70%) foi inferior ao controle (97%). Essa diferença,
dos ovócitos tratados estarem em estádios mais avançados da meiose, se manteve
às 12h, porém às 18h de MIV todos os ovócitos apresentaram taxas similares de
MII. Isso indica que a aceleração que ocorreu no reinício da meiose, já não foi mais
observada na segunda metade do período de maturação. PONDERATO et al.
(2001) observaram um padrão similar ao usar a mistura de inibidores em baixas
concentrações, onde houve uma aceleração inicial de ~ 4 h na quebra da VG, mas
às 18h, os ovócitos estavam em estádios similares da meiose. Em suínos, tal
fenômeno também foi observado (MARCHAL et al., 2001; WU et al., 2002).
Essa aceleração pode ser um padrão do tipo de droga e não um efeito da
concentração, pois é observado em diferentes espécies, diferentes inibidores de
CDK (BLI, ROS e a mistura de ambos) e concentrações diferentes como 10 e 100
μM.
Alguns estudos mostram que o bloqueio da meiose de ovócitos não inibe
completamente a síntese (MARCHAL et al., 2001) e fosforilação protéicas
(VIGNERON et al., 2004a) ou a transcrição (LEQUARRE et al., 2004). WU et al.
(2002) ainda observaram que a MAPK, embora inibida durante o bloqueio, é
rapidamente ativada após a remoção do inibidor. Isso indica que durante o bloqueio
pode haver acúmulo de fatores necessários à progressão meiótica.
O efeito dessa aceleração, porém, não parece ser deletério, pois as taxas de
desenvolvimento à blastocisto usando tais tratamentos não foram prejudicadas
(BEKER-VAN WOUDENBERG et al., 2005) e podem até melhorar o
desenvolvimento à blastocisto (COY et al., 2005a). Além disso, recentemente foi
relatado o nascimento de leitões produzidos in vitro a partir de ovócitos bloqueados
antes da MIV (COY et al., 2005b). O significado da aceleração da meiose ainda não
está bem esclarecido.
Nos experimentos seguintes buscou-se averiguar o efeito de tais tratamentos
sobre estruturas celulares que sofrem modificações durante o processo normal de
maturação e que são alguns dos indicativos de maturação citoplasmática. Sabe-se

48
que o citoesqueleto (microtúbulos e microfilamentos) (CONNORS et al., 1998; KIM
et al., 2000; SUN et al., 2001ac) e a disposição de organelas como grânulos
corticais (CONNORS et al., 1998; WESSEL et al., 2001; VELLILA et al., 2004) e
mitocôndrias estão relacionados à competência dos ovócitos (HYTTEL et al., 1997;
SUN et al., 2001b; WILDING et al., 2001).
Nesse estudo foi observada uma marcação positiva dos microtúbulos por
todo o citoplasma dos ovócitos controles e inibidos (10 ou 100 μM) ao longo de toda
a maturação in vitro pós-bloqueio. Uma marcação mais intensa dos microtúbulos foi
observada no período em que normalmente se inicia o rompimento do envelope
nuclear, permanecendo junto aos cromossomos no estádio de MII, organizando o
eixo meiótico independente dos tratamentos. Não foram observadas alterações no
fuso meiótico quanto à organização e formação da placa metafásica nos diferentes
grupos de ovócitos e horários dos tratamentos. Esses resultados são similares aos
observados por KIM et al. (2000) na avaliação dos microtúbulos em ovócitos
bovinos maturados in vitro, sugerindo que o inibidor não parece afetar a
organização dos microtúbulos.
WU et al. (2002), também não observaram alterações em ovócitos de suínos
expostos à baixa concentação de BL I. Por outro lado, JU et al. (2003), usando ROS
também em ovócitos suínos, observaram alterações no citoesqueleto, mas só em
concentrações mais altas, assim como ALBARRACÍN et al. (2005) em ovócitos de
bezerras. JU et al. (2003), porém, observaram alterações mais intensas na
cromatina. Nesse estudo houve apenas um ovócito com uma alteração do fuso,
envolvendo os microtúbulos e a cromatina no grupo tratado com 100 μM BL I, não
sendo, porém, significativo.
Essa diferença entre relatos pode se dever a diferentes condições de cultivo,
inibidores, animais, espécies ou ainda à forma de avaliação. Esses estudos foram
feitos por microscopia confocal, enquanto que o nosso foi feito por microscopia de
epifluorescência, que gera imagens menos nítidas que o confocal.
A marcação dos microfilamentos também foi observada por todo o citoplasma
dos ovócitos em todos os tratamentos e horários. Uma marcação mais intensa dos
microfilamentos foi observada no córtex e na região onde o corpúsculo polar foi
liberado, nos horários de 18 e 24 horas de maturação. Esses resultados são
similares aos encontrados por KIM et al. (2000) e WU et al. (2002) que observaram
os microfilamentos no córtex celular, em torno da VG e por todo o citoplasma do

49
ovócito. Não foi observada nenhuma diferença ou alteração no arranjo dos
microfilamentos no citoplasma dos ovócitos avaliados nos diferentes grupos e
horários de maturação, sendo esses resultados similares aos encontrados em
ovócitos de suínos tratados com BL I (WU et al., 2002).
Os dados dos microtúbulos e microfilamentos sugerem que o bloqueio
meiótico nas condições usadas nesse estudo não afetaram sua organização nos
ovócitos maturados in vitro pós-bloqueio da meiose, sendo um fato importante para
sua utilização, pois o arranjo dos microtúbulos e microfilamentos estão associados e
interagem com as mudanças cromossômicas durante a maturação do ovócito (KIM
et al., 2000; BRUNET e MARO, 2005). Com o ovócito bloqueado em estádio de VG
os cromossomos estão descondensados e o MPF está inativo (KUBELKA et al.,
2000). Nesse estádio celular, a dinâmica do citoesqueleto também permanece
inativa no que se refere aos eventos da divisão celular. Com a supressão do
bloqueio meiótico, porém, há o rompimento da VG, a condensação dos
cromossomos e a ativação do MPF (KUBELKA et al., 2000), eventos que promovem
a ativação do citoesqueleto (BRUNET e MARO, 2005). Os microtúbulos e os
microfilamentos não são requeridos para o rompimento da VG, mas, são importantes
na formação da placa metafásica e para a extrusão do corpúsculo polar (KIM et al.,
2000).
Os grânulos corticais nos ovócitos imaturos estão distribuídos por todo o
citoplasma celular e sua migração para a região periférica do ovócito (córtex) vai
ocorrendo com o avanço da maturação (CONNORS et al., 1998; WESSEL et al.,
2001; VELILLA et al., 2004). Nos ovócitos bloqueados com BL I por 24 horas,
independente dos tratamentos, não foi constatada migração dos grânulos corticais
para o córtex celular, mantendo a mesma distribuição de ovócitos imaturos. Esse
bloqueio da migração sugere que o MPF estaria envolvido, uma vez que, estando
inibida pela BL I, a migração dos grânulos corticais não ocorreu. WESSEL et al.
(2002) sugerem essa relação, uma vez, que utilizando ovócitos de ouriço-do-mar
bloqueados com ROS, também não houve migração de grânulos corticais.
A ausência de migração também pode estar relacionada a “inatividade” dos
microfilamentos que são responsáveis pela migração dos mesmos para região do
córtex celular (CONNORS et al., 1998; SUN et al., 2001a). Essa “inatividade” pode
estar associada ao bloqueio meiótico causada pelo MPF inativo, o qual não induz a
dinâmica do citoesqueleto (BRUNET e MARO, 2005), responsável pelo movimento

50
de organelas no interior do citoplasma celular (CONNORS et al., 1998; SUN et al.,
2001a; WESSEL et al., 2002).
Um estudo, porém, observou que a utilização de um enantiômero da ROS
bloqueia a meiose, mas não totalmente a migração de grânulos corticais, pois uma
parcela dos ovócitos tratados apresentou migração (COY et al., 2005a). Essa
contradição pode ser resultado de diferenças nas condições de cultivo, tipo de droga
ou de avaliação ou bloqueio menos eficiente.
Em ovócitos maturados in vivo ou in vitro os grânulos corticais são
encontrados em sua maioria na região do córtex celular (CONNORS et al., 1998;
WESSEL et al., 2001; VELILLA et al., 2004), sendo um dos indicativos de maturação
citoplasmática. A migração dos grânulos corticais para a região do córtex celular,
nos ovócitos maturados in vitro após o bloqueio da meiose com BL I foi similar aos
ovócitos submetidos somente à maturação in vitro. Esses resultados condizem com
os descritos para ovócitos maturados in vitro (VELILLA et al., 2004), sugerindo que o
bloqueio do ciclo meiótico pela BL I nos ovócitos bovinos, nas concentrações
apresentadas nesse estudo, não afeta a migração dos grânulos corticais no pós-
bloqueio meiótico. COY et al. (2005a) também observaram migração normal após a
MIV de ovócitos bloqueados com ROS. Essa migração dos grânulos corticais para o
córtex celular também é um indicativo de que os microfilamentos não foram afetados
pelo bloqueio meiótico, uma vez que são eles os responsáveis pela migração dos
mesmos (CONNORS et al., 1998; SUN et al., 2001a).
A distribuição das mitocôndrias para as células-filhas durante a divisão celular
é uma característica essencial para a proliferação das células. Elas têm um papel
central no fornecimento de ATP para a maturação, fecundação e para o
desenvolvimento embrionário (SUN et al., 2001b; WILDING et al., 2001). Durante a
maturação ovocitária as mitocôndrias alteram sua disposição no citoplasma celular.
Em ovócitos bovinos (HYTTEL et al., 1997) e suínos (SUN et al., 2001b) em estádio
de VG as mitocôndrias estão distribuídas em sua maioria na periferia do citoplasma
com grupos dispersos também mais ao centro do ovócito. Já nos ovócitos maturados
em estádio metáfase II as mitocôndrias estão mais centralizadas no citoplasma
(HYTTEL et al., 1997; SUN et al., 2001b). Esse mesmo padrão de distribuição das
mitocôndrias nos ovócitos imaturos e maturados foi observado nesse estudo, com
uma possível diferença na distribuição das mitocôndrias nos ovócitos maturados,
que segundo HYTTEL et al. (1997) estão mais centralizadas no citoplasma do

51
ovócito, mas nesse estudo estavam distribuídas por todo citoplasma. Essa variação
pode ser devido à diferença técnica de avaliação aplicada nos dois estudos
(epifluorescência nesse e microscopia eletrônica de transmissão no outro).
Houve diferença na distribuição das mitocôndrias entre os ovócitos imaturos
do grupo controle C/0 h (100% dos ovócitos recém-aspirados com mitocôndrias
distribuídas em sua maioria na região periférica do citoplasma) em relação aos
tratamentos com B10 e B100 (81,5% e 86,9%, respectivamente, com esse tipo de
distribuição). Ou seja, em 13-19% dos ovócitos mantidos em VG com BL I houve
migração das mitocôndrias para um padrão similar ao de ovócitos maturados. Isso
sugere que essa migração poderia não ser completamente controlada pelo MPF (ou
MAPK que é indiretamente inibido pela BL I – KUBELKA et al., 2000), isto é, outra(s)
via(s) poderia(m) participar desse controle. WU et al. (2002), também observaram
migração de mitocôndrias em ovócitos de suínos bloqueados. VIGNERON et al.
(2004b) observaram que ovócitos bovinos bloqueados com ROS acumulam algumas
moléculas relacionadas com o ciclo celular como a Aurora A cinase, JNK e PI3K. Se
essas moléculas têm de fato relação com a migração das mitocôndrias deve ser
testado.
A distribuição das mitocôndrias no interior do ovócito é mediada pelos
microtúbulos (SUN et al., 2001b) que são usados por proteínas motoras que geram
movimento das organelas no interior da célula (ALBERTS et al., 1997). Embora, o
ciclo celular meiótico estivesse bloqueado foi observada migração das mitocôndrias,
indicando que mesmo com ciclo celular bloqueado o mecanismo que gera o
movimento das mitocôndrias estava ativo em parte dos ovócitos submetidos ao
bloqueio meiótico. Em ovócitos de suínos tratados com BL I também foi observada a
translocação das mitocôndrias no interior do citoplasma celular (WU et al., 2002).
Esses resultados indicam que a translocação das mitocôndrias pode ser, ao menos
em parte, independente da maturação nuclear e da ativação do MPF (WU et al.,
2002).
Nos ovócitos maturados também foi observada diferença no padrão da
distribuição das mitocôndrias no citoplasma entre o controle C/24 (81,5% dos
ovócitos com mitocôndrias distribuídas por todo citoplasma) e os tratamentos com
B10M e B100M (95,2% e 98,2, respectivamente). Isso indica que houve uma maior
migração das mitocôndrias após a remoção do inibidor, provavelmente ocasionada
pela parcela de ovócitos que já havia iniciado a migração mesmo durante o bloqueio.

52
No que se refere ao desenvolvimento embrionário, no presente estudo as
diferenças se revelaram somente mais tardiamente na formação de blastocistos,
visto que as taxas de clivagem foram elevadas (80-86%) e semelhantes em todos os
tratamentos. Pela observação dos resultados do percentual de ovócitos que
alcançaram o estádio de blastocisto, verificou-se um número maior de embriões nos
grupos controle (38,3%) e B10 (41,6%) comparado ao B100 (33,0%). No entanto, os
percentuais de embriões eclodidos e o número médio de células dos blastocistos
não diferiram entre os tratamentos. A maturação citoplasmática avaliada pela
formação de blastocistos aos 7 dias de cultivo, porém, foi afetada pelo inibidor BL I
na concentração de 100 μM de BL I (B100), o que foi demonstrado pelo
comprometimento no desenvolvimento embrionário. As prováveis causas da baixa
eficiência na produção de embriões podem estar correlacionadas com as
concentrações do inibidor. LONERGAN et al. (2000) demonstraram que o bloqueio
meiótico com concentração elevada de BL I (150 μM) tem efeitos adversos no
desenvolvimento embrionário de ovócitos bovinos. Entre as alterações observadas
por microscopia eletrônica no estudo feito por LONERGAN et al. (2003) em ovócitos
bovinos tratados com BL I (100 μM) ou com roscovitina (25 μM), as células do
cumulus apresentavam-se mais alongadas, o núcleo do ovócito ondulado e
invaginado, as mitocôndrias com o formato bastante modificado (pleomorficas), o
reticulo endoplasmático aumentado e alguns dos grânulos corticais com aparência
de degenerados. Essas alterações foram bem mais acentuadas nos ovócitos
tratados com roscovitina (ROS). Em concordância com essa observação, pode-se
presumir que estaria ocorrendo uma correlação entre a maior dose do inibidor e uma
maior toxicidade, visto que a menor concentração de BL I (B10) não prejudicou a
produção de embriões (41,6%) quando comparado ao controle (38,3%).
Por outro lado, a maior parte dos estudos usando 100 μM de BL I ou não
reduz (PONDERATO et al., 2001; IMAI et al., 2002, LONERGAN et al., 2000;
BEKER-VAN WOUNDERBERG et al., 2005) ou até pode aumentar o
desenvolvimento até o estádio de blastocisto (HASHIMOTO et al., 2002; COY et al.,
2005b). Além disso, há relato de desenvolvimento fetal (PONDERATO et al., 2002) e
nascimento (COY et al., 2005b) com tratamentos similares. Apenas um estudo de
MARCHAL et al. (2001) com ovócitos suínos com ROS também obteve redução da
produção de blastocistos. Mesmo resultados anteriores de nosso laboratório não
mostraram essa redução (ADONA e LEAL, 2004). As razões para essa discrepância

53
não são claras, mas poderia ser causada por variação sazonal na qualidade dos
ovócitos que poderiam estar mais sensíveis ao uso da concentração mais alta do
inibidor.
Embora tenha havido redução da formação de blastocistos apenas no grupo
bloqueado com 100 μM BL I, a taxa de eclosão e o número de células não foi
afetado negativamente por nenhum tratamento, indicando que apesar de poder
afetar a proporção de blastocistos formados, a qualidade dos mesmos não foi. Isso
pode indicar um efeito do tratamento sobre a qualidade dos ovócitos, mas não dos
embriões gerados. RIZOS et al. (2002) observaram que a proporção de embriões
produzidos por FIV é afetada pela qualidade intrínseca do ovócito, mas que a
qualidade dos embriões é afetada pelo cultivo in vitro. A falta de efeito negativo
sobre o número de células de embriões obtidos de ovócitos bloqueados também foi
observada por MERMILLOD et al. (2000). HASHIMOTO et al. (2002), porém,
obtiveram aumento do número de células. PONDERATO et al. (2001), avaliando a
qualidade de embriões por taxa de eclosão após vitrificação de blastocistos também
não observaram diferenças entre embriões de ovócitos bloqueados ou não.
Os resultados do desenvolvimento embrionário aqui apresentados estão de
acordo com a literatura (MERMILLOD et al., 2000; PONDERATO et al., 2001;
PONDERATO et al., 2002; ADONA e LEAL, 2004) apesar de não ter sido
encontrado nenhum estudo em ovócitos de bovinos com a concentração próxima de
10 μM BL I (B10). PONDERATO et al. (2001) em seu estudo misturou os inibidores
ROS (12,5 μM) e BL I (6,25 μM), reduzindo a concentração dos mesmos, tendo bons
resultados de desenvolvimento embrionário, mas similar ao de ovócitos não trados.
De toda forma, o uso de inibidor em baixa concentração na ausência de
macromoléculas não afetou negativamente o desenvolvimento embrionário.
O cultivo de pré-maturação (B10) não melhorou e nem piorou os percentuais
de embriões que chegaram ao estádio de blastocistos em relação aos ovócitos não
tratados com BL I. Mais estudos são necessários para a melhor utilização do inibidor
(BL I), buscando melhorar a eficiência da maturação dos ovócitos bovinos in vitro,
adicionando aditivos durante o período de bloqueio (antioxidantes, hormônios,
fatores de crescimento ou fatores que estão envolvidos com a competência do
ovócito) para estudar os mecanismos envolvidos na maturação citoplasmática.

54
7. CONCLUSÕES
De acordo com os resultados obtidos nesse estudo, pode-se concluir que:
1) ovócitos bovinos podem ter a meiose bloqueada de forma eficaz e
reversível com baixa concentração de BL I, desde que na ausência de BSA;
2) a cinética da maturação in vitro pós-bloqueio da meiose é acelerada
mesmo com baixa concentração de BL I;
3) a organização dos microtúbulos e microfilamentos não foi afetada pelo
bloqueio meiótico com BL I sendo, sob esse aspecto, segura para ser utilizada na
pré-maturação;
4) a migração dos grânulos corticais foi inibida durante o bloqueio, mas não
foi alterada no pós-bloqueio; enquanto a translocação das mitocôndrias não foi
completamente inibida pelo bloqueio da meiose;
5) a BL I pode prejudicar ou não o desenvolvimento embrionário, dependendo
da concentração empregada;
6) a BL I pode ser utilizada em baixa concentração para bloquear de forma
segura e eficaz a meiose em ovócitos bovinos, prestando-se como ferramenta para
estudar as condições de cultivo para a produção de embriões in vitro.

55
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABRIEU A, DORÉE M, FISHER D. (2001). The interplay between cyclin-B–Cdc2
kinase (MPF) and MAP kinase during maturation of oocytes. Journal of Cell
Science. 114:257-267.
ABRUZZO MA, HASSOLD TJ. (1995). Etiology of nondisjunction in humans. Environmental and Molecular Mutagenesis. 25:38-47. Suppl 26.
ADONA PR, LEAL CLV. (2004). Meiotic inhibition with different cyclin-dependent
kinase inhibitors in bovine oocytes and its effects on maturation and embryo
development. Zygote. 12:197–204.
ADONA PR, LEAL CLV. (2006). Effect of concentration and exposure period to
butirolactone I on meiosis progression in bovine oocytes. Arquivo Brasileiro de
Medicina Veterinária e Zootecnia. Aceito para publicação.
ALBERTS B, BRAY D, LEWIS J, RAFF M, ROBERTS K, WATSON JD. (1997).
Biologia Molecular da Célula. Tradução de Amauri Braga Simonetti. Artes
Medicas, 3ª ed. Porto Alegre, p.1294.
ALBRARRACÍN JL, MOARATÓ R, IZQUIERDO D, MOGAS T. (2005). Effects of
roscovitine on the nuclear and cytoskeletal components of calf oocytes and their
subsequent development. Theriogenology, 64:1740-1755.
ANDERIESZ C, FONG C-Y, BONGSO A, TROUNSON AO. (2000). Regulation of
human and mouse oocyte maturation in vitro with 6-dimethylaminopurine. Human
Reproduction. 15:379-388.
ARRIS CE, BOYLE FT, CALVERT AH, CURTIN NJ, ENDICOTT JA, GARMAN EF,
GIBSON AE, GOLDING BT, GRANT S, GRIFFIN RJ, JEWSBURY P, JOHNSON
LN, LAWRIE AM, NEWELL DR, NOBLE MEM, EDWARD A. SAUSVILLE EA,
SCHULTZ R, YU W. (2000). Identification of novel purine and pyrimidine cyclin-
dependent kinase inhibitors with distinct molecular interactions and tumor cell
growth inhibition profiles. Journal of Medicinal Chemistry. 43:2797-2804.
AVELINO KB, BORDIGNOW V, SMITH LC. (1998). Corpum luteum and follicular
size on the bovine oocytes competence to result in blastocyst. Arquivo da
Faculdade de Veterinária – UFRGS, Porto Alegre, 26:223 (supl.).
BANZATTO DA, KRONCA SN. (1989). Experimentação Agrícola. Edição. Funep,
Jaboticabal, FCAV-UNESP, 247 p.

56
BEKER-VAN WOUDENBERG AR, ZEINSTRA EC, ROELEN BAJ,
COLENBRANDER B, BEVERS MM. (2005). Developmental competence of
bovine oocytes after specific inhibition of MPH kinase activity: effect of estradiol
supplementation and follicle size. Animal Reproduction Science, disponível on
line em 12/07/2005 no site. www.sicencedirect.com/science/journal/03784320.
BILODEAU-GOESEELS S. (2003). Effects of phosphodiesterase inhibitors on
spontaneous nuclear maturation and cAMP concentrations in bovine oocytes.
Theriogenology. 60:1679-1690.
BRAÑA MF, GARCÍA ML, LÓPEZ B, DE PASCUAL-TERESA B, RAMOS A,
POZUELO JM, DOMÍNGUEZ MT. (2004). Synthesis and biological evaluation of
analogues of butyrolactone I and molecular model of its interaction with CDK2.
Organic & Biomolecular Chemistry. 2: 1864-1871.
BRUNET S, G. PAHLAVAN G, TAYLOR S, MARO B. (2003). Functionality of the
spindle checkpoint during the first meiotic division of mammalian oocytes. Reproduction. 126:443–450.
BRUNET S, MARO B. (2005). Cytoskeleton and cell cycle control during meiotic
maturation of the mouse oocyte: integrating time and space. Reproduction.
130:801-811.
BUKOVSKY A, CAUDLE MR, SVETLIKOVA M, WIMALASENA J, AYALA ME,
DOMINGUEZ R. (2005). Oogenesis in adult mammals, including humans.
Endocrine, 26:1-15.
CAN A, SEMIZ O, ÇINAR O. (2003). Centrosome and microtubule dynamics during
early stages of meiosis in mouse oocytes. Molecular Human Reproduction. 9:749-
756.
CASTRO A, PETER M, MAGNAGHI-JAULIN L, VIGNERON S, GALAS S, LORCA T,
LABBE J-C. (2001). Cyclin B/cdc2 induces c-mos stability by direct
phosphorylation in Xenopus oocytes. Molecular Biology of the Cell. 12:2660–
2671.
CHA K-Y, CHIAN R-C. (1998). Maturation in vitro of immature human oocytes for
clinical use. Human Reproduction Update. 4:103-120.
CHEN Z, GIBSON TB, ROBINSON F, SILVESTRO L, PEARSON G, XU B-E,
WRIGHT A, VANDERBILT C, COBB MH. (2001). MAP Kinases. Chemical
Reviews. 101:2449-2476.

57
CHIAN RC, BUCKETT WB, TAN S-L. (2003). In-vitro maturation of human oocytes.
Reproductive BioMedicine Online. 8:148-166.
CONNORS SA, KANATSU-SHINOHARA M, SCHULTZ RM, KOPF GS. (1998).
Involvement of the cytoskeleton in the movement of cortical granules during
oocyte maturation, and cortical granule anchoring in mouse eggs. Developmental
Biology. 200:103–115.
CONTI M, ANDERSEN CB, RICHARD FJ, SHITSUKAWA K, TSAFRIRI A. (1998).
Role of cyclic nucleotide phosphodiesterases in resumption of meiosis. Molecular
and Cellular Endocrinology. 145:9–14.
COY P, ROMAR R, PAYTON RR, McCANN L, SAXTON AM, EDWARDS JL.
(2005a). Maintenance of meiotic arrest in bovine oocytes using the S-enantiomer
of roscovitine: effects on maturation, fertilization and subsequent embryo
development in vitro. Reproduction. 129:19-26.
COY P, ROMAR R, RUIZ S, CÁNOVAS S, GADEA J, VÁZQUEZ FG, MATÁS C.
(2005b). Birth of piglets after transferring of in vitro-produced embryos pre-
matured with R-roscovitine. Reproduction. 129:747-755.
CUMMINS JM. (2004). The role of mitochondria in the establishment of oocyte
functional competence. European Journal of Obstetrics & Gynecology and
Reproductive Biology. Suppl. 115:S23-29.
DALVIT G, LLANES SP, DESCALZO A, INSANI M, BECONI M, CETICA P. (2005).
Effect of alpha-tocopherol and ascorbic acid on bovine oocyte in vitro maturation.
Reproduction in Domestic Animals. 40:93-97.
DE LA FUENTE R, EPPIG JJ. (2001).Transcriptional Activity of the mouse oocyte
genome: companion granulosa cells modulate transcription and chromatin
remodeling. Developmental Biology. 229:224–236. DE LOOS F, KASTROP P, VAN BENEDEN TH, KRUIP TA. (1991). Heterologous
cell contacts and metabolic coupling in bovine cumulus oocyte complexes.
Molecular Reproduction and Development. 28: 255-259.
DE SOUSA PA, CAVENEY A, WESTHUSIN ME, WATSON AJ. (1998).Temporal
patterns of embryonic gene expression and their dependence on oogenetic
factors. Theriogenology. 49:115-128.
DE SOUSA PA, MARTINS DA SILVA SJ, ANDERSON RA. (2004). Neurotrophin
signaling in oocyte survival and developmental competence: A paradigm for
cellular toti-potency. Cloning and Stem Cells. 6:375-385.

58
DEKEL N. (2005). Cellular, biochemical and molecular mechanisms regulating
oocyte maturation. Molecular and Cellular Endocrinology. 234:19–25
DIELEMAN SJ, HENDRIKSEN PJM, VIUFF D, THOMSEN PD, HYTTEL P, et al.,
(2002). Effects of in vivo prematuration and in vivo final maturation on
developmental capacity and quality of pre-implantation embryos. Theriogenology.
57:5-20.
DODE MAN, ADONA PR. (2001). Developmental capacity of Bos indicus oocytes
after inhibition of meiotic resuption by 6-dimethyminopurine. Animal Reproduction
Science. 65:171-180.
DUCIBELLA T, DUFFY P, BUETOW J. (1994). Quantification and localization of
cortical granules during oogenesis in the mouse. Biology of Reproduction. 50:467-
473.
DUMONT J, UMBHAUER M, RASSINIER P, HANAUER A, VERLHAC, M-H. (2005).
p90Rsk is not involved in cytostatic factor arrest in mouse oocytes. The Journal of
Cell Biology. 1-5.
EYERS PA, LIU J, HAYASHI NR, LEWELLYN AL, GAUTIER J, MALLER JL. (2005).
Regulation of the G(2)/M transition in Xenopus oocytes by the cAMP-dependent
protein kinase. The Journal Of Biological Chemistry. 280:24339-24346.
FAERGE I, MAYES M, HYTTEL P, SISRARD MA. (2001). Nuclear ultrastructure in
bovine oocytes after inhibition of meiosis by chemical and biological inhibitors.
Molecular Reproduction and Development, 59:459-467.
FAIR T, HYTTEL P, MOTLIK J, BOLAND M, LONERGAN P. (2002). Maintenance of
meiotic arrest in bovine oocytes in vitro using butyrolactone I: Effects on oocyte
ultrastructure and nucleolus. Molecular Reproduction and Development, 62:375-
386.
FAIR T. (2003). Follicular oocyte growth and acquisition of developmental
competence. Animal Reproduction Science. 78:203–216.
FISSORE RA, HE CL, VANDE WOUDE GF. (1996). Potential role of mitogen-
activated protein kinase during meiosis resumption in bovine oocytes. Biology of
Reproduction. 55:1261-1270.
FOOTE WD, THIBAULT C. (1969). Recherches experimentales sur la maturation in
vitro des ovocytes de truie et de veau. Ann. Biol. Anim. Biochim. Biophys. 3:329-
349.
GANDOLFI, BTAL, GANDOLFI F. (2001). The maternal legacy to the embryo:

59
cytoplasmic components and their effects on early development. Theriogenology,
55:1255-1276.
GILCHRIST RB, RITTER LJ, ARMSTRONG DT. (2004). Oocyte–somatic cell
interactions during follicle development in mammals. Animal Reproduction
Science. 82:431–446.
GONSALVES PBD, VISINTIN JA, OLIVEIRA MAL, MONTAGNER MM, DA COSTA
LFS. (2002). Produção in vitro de embriões. In: Gonsalves PBD, Figueiredo JR,
Freitas VJ. Biotécnicas aplicadas à reprodução animal. 1º edição, Livraria Varela.
p.195-226.
HAFEZ ESE. (2002). Reprodução Animal, 7ª Edição, Editora Manole LTDA. 380p.
HASHIMOTO S, MINAMI N, TAKAKURA R, IMAI H. (2002). Bovine immature
oocytes acquire developmental competence during meiotic arrest in vitro. Biology
of Reproduction. 66:1696–1701.
HORNE MM, GUADAGNO TM. (2003). A requirement for MAP kinase in the
assembly and maintenance of the mitotic spindle. The Journal of Cell Biology.
161:1021–1028
HYTTEL P, FAIR T, CALLESEN H, GREVE T. (1997). Oocyte growth capacitation
and final maturation in cattle.Theriogenology. 47:23-32.
IMAI K, KOBAYASHI S, KANEYAMA K, KOJIMA T, NAGAI T. (2002). Effects of
butyrolactone I on GVBD in bovine oocytes and subsequent maturation,
fertilization and development in vitro. Journal Reproduction and Development,
48:249-255.
INOUE M, NAITO K, NAKAYAMA T, SATO E. (1998). Mitogen-activated protein
kinase translocates into the germinal vesicle and induces germinal vesicle
breakdown in porcine oocytes. Biology of Reproduction. 58:130-136.
JESSUS C, OZON R. (2004). How does Xenopus oocyte acquire its competence to
undergo meiotic maturation. Biology of the cell. 96:187-192.
JONES KT. (2004). Turning it on and off: M-phase promoting factor during meiotic
maturation and fertilization. Molecular Human Reproduction. 10:1-5.
JOSEFSBERG LB-Y, DEKEL N. (2002). Translational and post-translational
modifications in meiosis of the mammalian oocyte. Molecular and Cellular
Endocrinology. 187:161–171.

60
JU JC, TSAY C, RUAN CW. (2003). Alterations and reversibility in the chromatin,
cytoskeleton and development of pig oocytes treated with roscovitine. Molecular
Reproduction and Development. 64:482-491.
KANAYAMA N, MIYANO T, LEE J. (2002). Acquisition of meiotic competence in
growing pig oocytes correlates with their ability to activate Cdc2 kinase and MAP
kinase. Zygote. 10:261–270.
KAÑKA J. (2003). Gene expression and chromatin structure in the pre-implantation
embryo. Theriogenology. 59:3-19.
KANO F, TAKENAKA K, YAMAMOTO A, NAGAYAMA K, NISHIDA E, MURATA M.
(2000). MEK and Cdc2 kinase are sequentially required for golgi disassembly in
MDCK cells by the mitotic xenopus extracts. The Journal of Cell Biology.
149:357–368
KHATIR H, CAROLAN C, LONERGAN P, MERMILLOD P. (1997). Characterization
of calf follicular fluid and its ability to support cytoplasmic maturation of cow and
calf oocytes. Journal of Reproduction fertility. 111:267-275.
KHATIR H, LONERGAN P, MERMILLOD P. (1998). Kinetics of nuclear maturation
and profiles of oocytes from prepubertal and adult cattle during in vitro maturation.
Theriogenology. 50:917-929.
KIKUCHI K, NAITO K, NOGUCHI J, SHIMADA A, KANEKO K, YAMASHITA M,
TOJO H, TOYODA Y. (1999). Inactivation of p34cdc2 kinase by the accumulation
of its phosphorylated forms in porcine oocytes matured and aged in vitro. Zygote.
7:173–179.
KIKUCHI K, NAITO K, NOGUCHI J, SHIMADA A, KANEKO H, YAMASHITA M, AOKI
F, TOJO H, TOYODA Y. (2000). Maturation/M-phase promoting factor: A
regulator of aging in porcine oocytes. Biology of Reproduction. 63:715-722.
KIM N-H, CHO SK, CHOI SH, KIM EY, PARK S, PILL LIM JH. (2000). The
distribution and requirements of microtubules an microfilaments in bovine oocytes
in vitro maturation. Zygote. 8:25-32.
KNOCKAERT M, GREENGARD P, MEIJER L. (2002). Pharmacological inhibitors of
cyclin-dependent kinases. TRENDS in Pharmacological Sciences. 23:417-425.
KOTSUJI, F.; KUBO, M.; TOMINAGA, T. (1994) Effect of interactions between
granulose and thecal cells on meiotic arrest in bovine oocytes. Journal of
Reproduction and Fertility. 100:151-156.

61
KRISCHEK C, MEINECKE B. (2002). In vitro maturation of bovine oocytes requires
polyadenylation of mRNAs coding proteins for chromatin condensation, spindle
assembly, MPF and MAP kinase activation. Animal Reproduction Science.
73:129–140.
KUBELKA M, ANGER M, KALOUS J, SCHULTZ RM, MOTLIK J. (2002).
Chromosome condensation in pig oocytes: lack of a requirement for either cdc2
kinase or MAP kinase activity. Molecular Reproduction and Development. 63:110-
118.
KUBELKA M, MOTLIK J, SCHULTZ RM, PAVLOK A. (2000). Butyrolactone I
reversibly inhibits meiotic maturation of bovine oocytes, without influencing
chromosome condensation activity. Biology of Reproduction. 62:292-302.
LAGUTINA I, PONDERATO N, LAZZARI G, GALLI C. (2002). Kinetics of oocyte
maturation and subsequent development of IVF, parthenogenetic, and NT bovine
embryos after meiotic inhibition with roscovitinre. Cloning and Stem Cells. 4:113-
119.
LEDAN E, POLANSKI Z, TERRET M-E, MARO B. (2001). Meiotic Maturation of the
Mouse Oocyte Requires an Equilibrium between Cyclin B Synthesis and
Degradation. Developmental Biology. 232:400–413.
LEFEBVRE C, TERRET ME, DJIANE A, RASSINIER P, MARO B, VERLHAC M-H.
(2002). Meiotic spindle stability depends on MAPK-interacting and spindle-
stabilizing protein (MISS), a new MAPK substrate. The Journal of Cell Biology.
157:603–613.
LEQUARRE AS, TRAVERSO JM, MARCHANDISE J, DONAAY I. (2004). Poly(A)
RNA is reduced by half during bovine oocyte maturation but increases when
meiotic arrest is maintained with CDK inhibitors. Biology of Reproduction. 71:425-
431.
LONERGAN P, DINNYÉS A, FAIR T, YANG X, BOLAND M. (2000). Bovine oocyte
and embryo development following meiotic inhibition with butyrolactone I.
Molecular Reproduction and Development, 57:204-209.
LONERGAN P, FAERGE I, HYTTEL PM, BOLAND M, FAIR T. (2003). Ultrastructural
modifications in bovine oocytes maintained in meiotic arrest in vitro using
roscovitine or butyrolactone. Molecular Reproduction and Development. 64:369–
378.

62
MA S, LAN G, MIAO Y, WANG Z, CHANG Z, LUO M, TAN J. (2003). Hypoxanthine
(HX) inhibition of in vitro meiotic resumption in goat oocytes. Molecular
Reproduction and Development. 66:306-313.
MACHATKOVA M, KRAUSOVA K, JOKESOVA E, TOMANEK M. (2004).
Developmental competence of bovine oocytes: effects of follicle size and the
phase of follicular wave on in vitro embryo production. Theriogenology. 61:329–
335.
MAPELLI M, MASSIMILIANO L, CROVACE C, SEELIGER MA, TSAI L-H, MEIJER L,
MUSACCHIO A. (2005). Mechanism of CDK5/p25 Binding by CDK Inhibitors.
Journal of Medicinal Chemistry. 48:671-679.
MARCHAL R, TOMANEK M, TERQUI M, MERMILLOD P. (2001). Effect of cell cycle
depenent kinases inhibitor on nuclear and cytoplasmic maturation of porcine
oocytes. Molecular Reproduction and Development. 60:65-73.
MAYES M, SIRARD MA. (2001). The influence of cumulus-oocyte complex
morphology and meiotic inhibitors on the kinetics of nuclear maturation in cattle.
Theriogenology. 55:911-922.
MAYES M. (2002). The meiotic arrest of bovine oocytes. These.
www.theses.ulaval.ca/2002/20201/20201.pdf
McNATTY KP, FIDLER AE, JUENGEL JL, QUIRKE LD, SMITH PR, HEATH DA,
LUNDY T, O’CONNELL A, TISDALL DJ. (2000). Growth and paracrine factors
regulating follicular formation and cellular function. Molecular and Cellular
Endocrinology. 163:11–20.
MEINECKE B, JANAS U, PODHAJSKY E, MEINECKE-TILLMANN S. (2001).
Histone H1 and MAP kinase activities in bovine oocytes following protein
synthesis inhibition. Reproduction in Domestic Animals. 36:183-188.
MEIRELLES FV, CAETANO AR, WATANABE YF, RIPAMONTE P, CARAMBULA
SF, MERIGHE GK, GARCIA SM. (2004). Genome activation and developmental
block in bovine embryos. Animal Reproduction Science. 82:13-20.
MEMILI E, FIRST NL. (2000). Zygotic and embryonic gene expression in cow: a
review of timing and mechanisms of early gene expression as compared with
other species. Zygote. 8:87-96.
MERMILLOD P, TOMANEK M, MARCHAL R, MEIJER L. (2000). High
developmental competence of cattle oocytes maintained at the germinal vesicle
stage for 24 hours in culture by specific inhibition of MPF kinase activity.

63
Molecular Reproduction and Development. 55:89-95.
MERTON JS, DE ROOS AP, MULLAART E, DE RUIGH L, KAAL L, VOS PL,
DIELEMAN SJ. (2003). Factors affecting oocyte quality and quantity in
commercial application of embryo technologies in the cattle breeding industry.
Theriogenology. 59:651-74.
MIYANO T. (2003). Bringing up small oocytes to eggs in pigs and
cows.Theriogenology. 59: 61-72. MOORE e PERSAUD (2000). Embriologia básica. 5ª edição Guanabara Koogan.
453p.
NEBREDA AR, FERBY I. (2000). Regulation of the meiotic cell cycle in oocytes.
Current Opinion in Cell Biology. 12:666–675.
OYAMADA T, IWAYAMA H, FUKUI Y. (2004). Additional effect of epidermal growth
factor during in vitro maturation for individual bovine oocytes using a chemically
defined medium. Zygote. 12:143-150.
PARK Y-S, KIM S-S, KIM J-M, PARK H-D, BYUN M-D. (2005). The effects of
duration of in vitro maturation of bovine oocytes on subsequent development,
quality and transfer of embryos. Theriogenology. 64:123-134.
PAVLOK A. (2000). D-penicillamine and granulosa cells can effectively extend the
fertile life span of bovine frozen-thawed spermatozoa in vitro: effect on fertilization
and polyspermy. Theriogenology. 55:1135-1146.
PONDERATO N, CROTTI G, TURINI P, DUCHI R, GALLI C, LAZZARI G. (2002).
Embryonic and foetal development of bovine oocytes treated with a combination
of butyrolactone I and roscovitine inan enriched medium prir to IVM and IVF.
Molecular Reproduction and Development. 62:513-518.
PONDERATO N, LAGUTINA I, CROTTI G, TURINI P, GALLI C, LAZZARI G. (2001).
Bovine oocytes treated prior to in vitro maturation with combination of
butyrolactone I and roscovitine at low doses maintain a normal developmental
capacity. Molecular Reproduction and Development. 60:579-585.
REYNIER P, MAY-PANLOUP P, CHRÉTIEN M-F, MORGAN CJ, JEAN M,
SAVAGNER F, BARRIÈRE P, MALTHIÈRY Y. (2001). Mitochondrial DNA content
affects the fertilizability of human oocytes. Molecular Human Reproduction. 7:425-
429.
RIZOS D, WARD F, DUFFY P, BOLAND M, LONERGAN, P. (2002). Consequences
of bovine oocyte maturation, fertilization or early embryo development in vitro

64
versus in vivo: Implications for blastocyst yield and blstocyst quality. Molecular
Reproduction and Development, 61:234-248.
RODRIGUEZ KF, FARIN CE. (2004). Gene transcription and regulation of oocyte
maturation. Reproduction fertility and Development. 16:55-67. ROUX PP, BLENIS J. (2004). ERK and p38 MAPK-activated protein kinases: a
family of protein kinases with diverse biological functions. Microbiology and
Molecular Biology. 68:320–344.
SAS (1995). USER’S GUIDE: basic and statistic. Cary,NC: SAS,. 1.686 p.
SAX JK, DASH BC, HONG R, DICKER DT, EL-DEIRY WS. (2002). The Cyclin-
Dependent kinase inhibitor butyrolactone is a potent inhibitor of p21WAF1/CIP1
expression. Cell Cycle 1:90-96.
SCHANG LM. (2004). Effects of pharmacological cyclin-dependent kinase inhibitors
on viral transcription and replication. Biochimica et Biophysica Acta (BBA).
1697:197-209.
SCHIMMEL TG, COFFMAN AD, PARSONS SJ. (1998). Effect of Butyrolactone I on
the Producing Fungus, Aspergillus terreus. Applied and Environmental
Microbiology. 64:3707-3712.
SCHOEVERS EJ, BEVERS MM, ROELEN BAJ, COLENBRANDER B. (2005).
Nuclear and cytoplasmic maturation of sow oocytes are not synchronized by
specific meiotic inhibition with roscovitine during in vitro maturation.
Theriogenology. 63:1111–1130.
SHENG J, KUMAGAI A, DUNPHY WG, VARSHAVSKY A. (2002). Dissection of c-
mos degron. The EMBO Journal. 21:6061-6071.
SIRARD MA, RICHARD F, BLONDIN P, ROBERT C. (2006). Contribution of the
oocyte to embryo quality. Theriogenology. 65 :126–136.
SIRARD MA, RICHARD F, MAYES M. (1998). Controlling meiotic resumption in
bovine oocytes: a review. Theriogenology. 49:483-497.
SIRARD MA. (2001). Resumption of meiosis: mechanism involved in meiotic
progression and its relation with developmental competence. Theriogenology.
55:1241-1254.
STEUERWALD N, BARRITT JA, ADLER R, MALTER H, SCHIMMEL T, COHEN J,
BRENNER CA. (2000). Quantification of mtDNA in single oocytes, polar bodies
and subcellular components by real-time rapid cycle fluorescence monitored
PCR. Zygote. 9:209–215.

65
STOJKOVIC M, MACHADO SA, SOTOJKOVIC P, ZAKHARTCHENKO V, HUTZLER
P, GONÇALVES PB, WOLF E. (2001). Mitochondrial distribution and adenosine
triphosphate content of bovine oocytes before and after in vitro maturation:
correlation with morphological criteria and developmental capacity after in vitro
fertilization and culture. Biology of Reproduction. 64:904-909.
SUN QY, LAI L, PARK KW, KÜHHOLZER B, PRATHER RS, SCHATTEN H. (2001a).
Dynamic events are differently mediated by microfilaments, microtubules, and
mitogen-activated kinase during porcine oocyte maturation and fertilization in
vitro. Biology of Reproduction. 64:879-889.
SUN QY, LAI L, WU GM, PARK KW, DAY BN, PRATHER RS, SCHATTEN H.
(2001c). Microtubule assembly after treatment of pig oocytes with taxol:
correlation with chromosomes, gamma-tubulin, and MAP kinase. Molecular
Reproduction and Development. 60:481-490.
SUN QY, LU Q, BREITBART H, CHEN DY. (1999). CAMP inhibits mitogen-activated
protein (MAP) kinase activation and resumption of meiosis, but exerts no effects
after spontaneous germinal vesicle breakdown (GVBD) in mouse oocytes.
Reproduction, fertility, and development.11:81-86.
SUN QY, WU GM, LAI L, BONK A, CABOT R, PARK KW, DAY BN, PRATHER RS,
SCHATTEN H. (2002). Regulation of mitogen-activated protein kinase
phosphorylation, microtubule organization, chromatin behavior, and cell cycle
progression by protein phosphatases during pig oocyte maturation and fertilization
in vitro. Biology of Reproduction. 66:580-588.
SUN QY, WU GM, LAI L, PARK KW, CABOT R, CHEONG HT, DAY BN, PRATHER
RS, SCHATTEN H. (2001b). Translocation of active mitochondria during pig
oocyte maturation, fertilization and early embryo development in vitro.
Reproduction. 122:155-163.
SUN X-F, WANG W-H, KEEFE DL. (2004). Overheating is detrimental to meiotic
spindles within in vitro matured human oocytes. Zygote. 12:65–70.
TAIEB R, THIBIER C, JESSUS C. (1997). On cyclins, oocytes, and eggs. Molecular
Reproduction and Development. 48:397–411.
TERRET ME, LEFEBVRE C, DJIANE A, RASSINIER P, MOREAU J, MARO B,
VERLHAC M-H. (2003). DOC1R: a MAP kinase substrate that control microtubule
organization of metaphase II mouse oocytes. Development. 130:5169-5177.

66
TIAN XC, LONERGAN P, JEONG BS, EVANS AC, YANG X. (2002). Association of
MPF, MAPK, and nuclear progression dynamics during activation of young and
aged bovine oocytes. Molecular Reproduction and Development. 62:132-138.
TOMEK W, TORNER H, KANITZ W. (2002). Comparative analysis of protein
synthesis, transcription and cytoplasmic polyadenylation of mRNA during
maturation of bovine oocytes in vitro. Reproduction in Domestic Animals. 37:86-
91.
TONG C, FAN HY, CHEN DY, SONG XF, SCHATTEN H, SUN QY. (2003). Effects of
MEK inhibitor U0126 on meiotic progression in mouse oocytes: microtuble
organization, asymmetric division and metaphase II arrest. Cell research. 13:375-
383. VAILLANT MF, HACCARD O, OZON R, JESSUS C. (2001). Interplay between Cdc2
Kinase and the c-Mos/MAPK Pathway between Metaphase I and Metaphase II in
Xenopus Oocytes. Developmental Biology. 231:279–288.
VAN DEN HURK R, ZHAO J. (2005). Formation of mammalian oocytes and their
growth, differentiation and maturation within ovarian follicles. Theriogenology.
63:1717-1751.
VELILLA E, IZQUIERDO D, RODRIGUEZ-GONZALEZ E, LOPEZ-BEJAR M, VIDAL
F, PARAMIO MT. (2004). Distribution of prepubertal and adult goat oocyte cortical
granules during meiotic maturation and fertilisation: ultrastructural and
cytochemical study. Molecular Reproduction and Development. 68:507-514.
VINGNERON C, PERREAU C, DALBIÉS-TRAN, JOLY C, HUMBLOT P, UZBEKOVA
S, MERMILLOD P. (2004a). Protein synthesis and mRNA sotarage in cattle
oocytes maintained under meiotic block by roscovitine inhibition of MPF activity.
Molecular Reproduction and Development. 69:457-465.
VINGNERON C, PERREAU C, DUPONT J, UZBEKOVA S, PRINGENT C,
MERMILLOD P. (2004b). Several signaling pathways are involved in the control
of cattle oocyte maturation. Molecular Reproduction and Development. 69:466-
474.
WARD F, ENRIGHT B, RIZOS D, BOLAND M, LONERGAN P. (2002). Optimization
of in vitro bovine embryo production: effect of duration of maturation, length of
gamete co-incubation, sperm concentration and sire. Theriogenology. 57:2105-
2117.

67
WESSEL GM, BROOKS JM, GREEN E, HALEY S, VORONINA E, WONG J,
ZAYDFUDIM V, CONNER S. (2001). The Biology of Cortical Granules.
Department of Molecular Biology, Cell Biology & Biochemistry. Brown University.
Providence, Rhode Island, 02912. E-mail: [email protected] 90p.
WESSEL GM, CONNER SD, BERG L. (2002). Cortical granule translocation is
microfilament mediated and linked to meiotic maturation in the sea urchin oocyte.
Development, 126:4315-4325.
WILDING M, DALE B, MARINO M, DI MATTEO L, ALVIGGI C, PISATURO ML,
LOMBARDI L, DE PLACIDO G. (2001). Mitochondrial aggregation patterns and
activity in human oocytes and preimplantation embryos. Human Reproduction.
16:909-917.
WINSTON N J. (1997). Stability of cyclin B protein during meiotic maturation and first
mitotic cell division in mouse oocytes. Biology of the Cell. 89:211–219.
WU GM, SUN QY, MAO J, LAI L, PARK KW, PRATHER RS, DIDION BA, DAY BN.
(2002). High developmental competence of pig oocytes after meiotic inhibition
with a specific M-phase promoting factor kinase inhibitor, butyrolactone I. Biology
of Reproduction. 67:170-177.
YI YJ, PARK CS. (2005). Parthenogenetic development of porcine oocytes treated by
ethanol, cycloheximide, cytochalasin B and 6-dimethylaminopurine. Animal
Reproduction Science. 86:297-304.

68
APÊNDICE
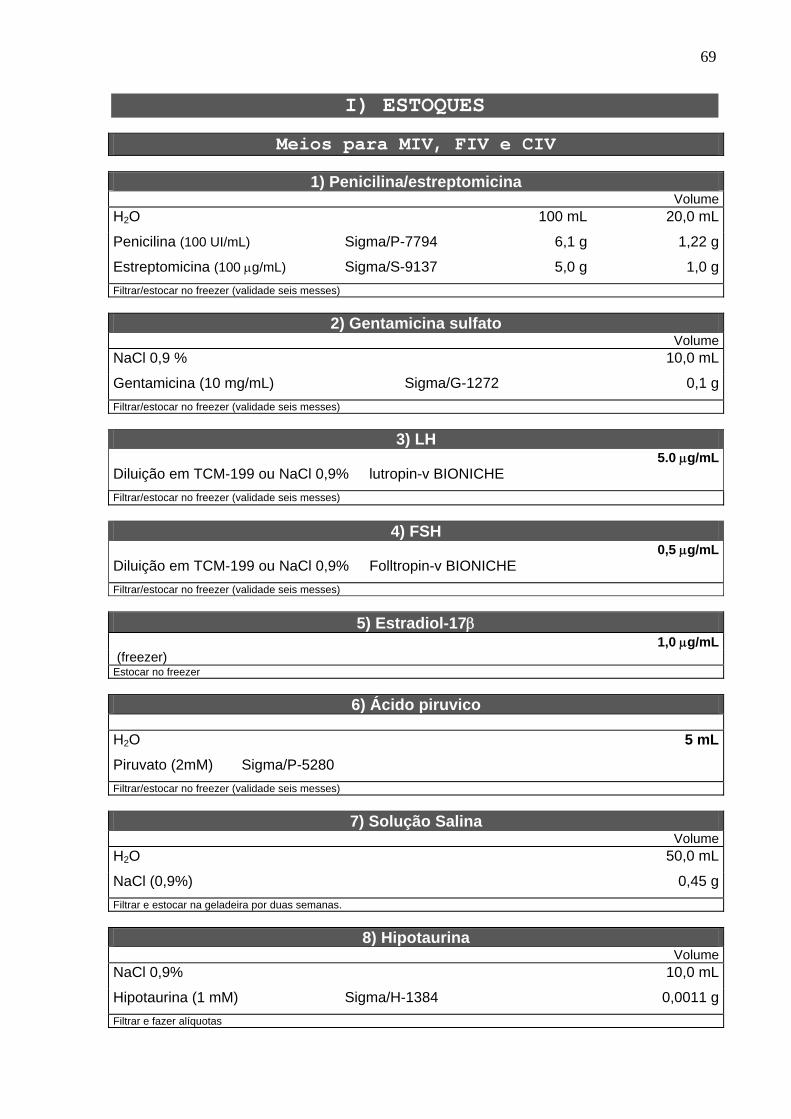
69
I) ESTOQUES
Meios para MIV, FIV e CIV
1) Penicilina/estreptomicina Volume
H2O 100 mL 20,0 mL
Penicilina (100 UI/mL) Sigma/P-7794 6,1 g 1,22 g
Estreptomicina (100 μg/mL) Sigma/S-9137 5,0 g 1,0 gFiltrar/estocar no freezer (validade seis messes)
2) Gentamicina sulfato Volume
NaCl 0,9 % 10,0 mL
Gentamicina (10 mg/mL) Sigma/G-1272 0,1 gFiltrar/estocar no freezer (validade seis messes)
3) LH 5.0 μg/mL
Diluição em TCM-199 ou NaCl 0,9% lutropin-v BIONICHE Filtrar/estocar no freezer (validade seis messes)
4) FSH 0,5 μg/mL
Diluição em TCM-199 ou NaCl 0,9% Folltropin-v BIONICHE Filtrar/estocar no freezer (validade seis messes)
5) Estradiol-17β 1,0 μg/mL
(freezer) Estocar no freezer
6) Ácido piruvico
H2O 5 mLPiruvato (2mM) Sigma/P-5280 Filtrar/estocar no freezer (validade seis messes)
7) Solução Salina Volume
H2O 50,0 mL
NaCl (0,9%) 0,45 gFiltrar e estocar na geladeira por duas semanas.
8) Hipotaurina Volume
NaCl 0,9% 10,0 mL
Hipotaurina (1 mM) Sigma/H-1384 0,0011 gFiltrar e fazer alíquotas

70
9) Epinefrina (Proteger da luz) Volume
H2O 40,0 mL
Acido lático (60%) 104 μL
Metabissulfito (Na2S2O5) Sigma/S-9000 0,04 g
Corrigir o pH para 4,0 com HCl (1M) Acrescentar a Epinefrina (250 mM) Sigma/E-1635 0,00183 gFiltrar e fazer alíquotas
10) Penicilamina
VolumeNaCl 0,9 % 10,0 mL
Penicilamina (2 mM) Sigma/P-4875 0,003 gFiltrar e fazer alíquotas
11) PHE (Proteger da luz) Volume
NaCl (0,9%) 8,0 mL
Penicilamina 5,0 mL
Hipotaurina 5,0 mL
Epinefrina 2,0 mLEstocar em alíquotas de 250 μL no freezer por seis meses (proteger da luz).
12) Heparina Volume
NaCl 0,9 % 10,0 mL
Heparina (3 mg/mL) USB/CAS:9041-08-1 0,03 gFiltrar/estocar no freezer alíquotas de ± 30 μL (validade seis messes)
13) CaCl2.2H2O
VolumeH2O 5,0 mL
CaCl2.2H2O 0,155 gFiltrar/estocar na geladeira (validade seis messes)
14) Sodium tri citrate
VolumeH2O 5,0 mL
Sodium tri citrate (1000X) Merck/1.06448.1000 0,5 gFiltrar/estocar no freezer (validade seis messes)
15) Hemi cálcio–lactato
VolumeH2O 3,0 mL
Hemi cálcio–lactato (100X) 0,164 gFiltrar/estocar no freezer (validade seis messes)
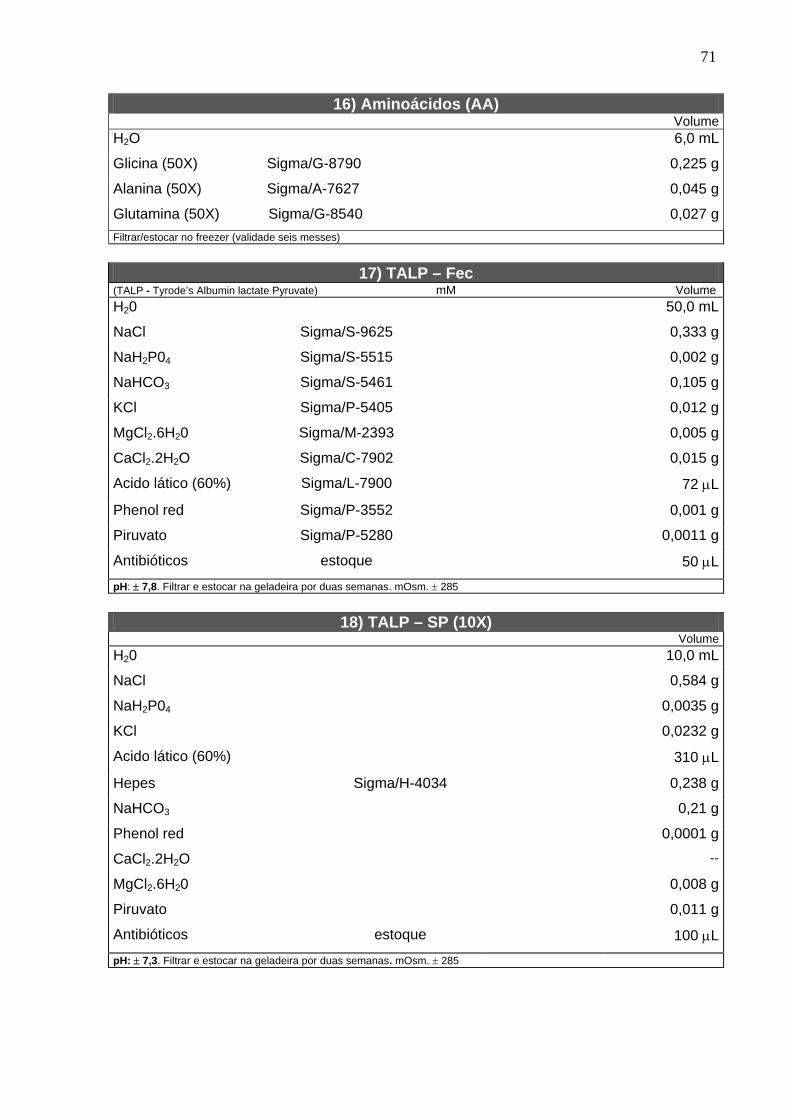
71
16) Aminoácidos (AA) VolumeH2O 6,0 mL
Glicina (50X) Sigma/G-8790 0,225 g
Alanina (50X) Sigma/A-7627 0,045 g
Glutamina (50X) Sigma/G-8540 0,027 gFiltrar/estocar no freezer (validade seis messes)
17) TALP – Fec
(TALP - Tyrode’s Albumin lactate Pyruvate) mM Volume H20 50,0 mL
NaCl Sigma/S-9625 0,333 g
NaH2P04 Sigma/S-5515 0,002 g
NaHCO3 Sigma/S-5461 0,105 g
KCl Sigma/P-5405 0,012 g
MgCl2.6H20 Sigma/M-2393 0,005 g
CaCl2.2H2O Sigma/C-7902 0,015 g
Acido lático (60%) Sigma/L-7900 72 μL
Phenol red Sigma/P-3552 0,001 g
Piruvato Sigma/P-5280 0,0011 g
Antibióticos estoque 50 μLpH: ± 7,8. Filtrar e estocar na geladeira por duas semanas. mOsm. ± 285
18) TALP – SP (10X)
VolumeH20 10,0 mL
NaCl 0,584 g
NaH2P04 0,0035 g
KCl 0,0232 g
Acido lático (60%) 310 μL
Hepes Sigma/H-4034 0,238 g
NaHCO3 0,21 g
Phenol red 0,0001 g
CaCl2.2H2O --
MgCl2.6H20 0,008 g
Piruvato 0,011 g
Antibióticos estoque 100 μL pH: ± 7,3. Filtrar e estocar na geladeira por duas semanas. mOsm. ± 285
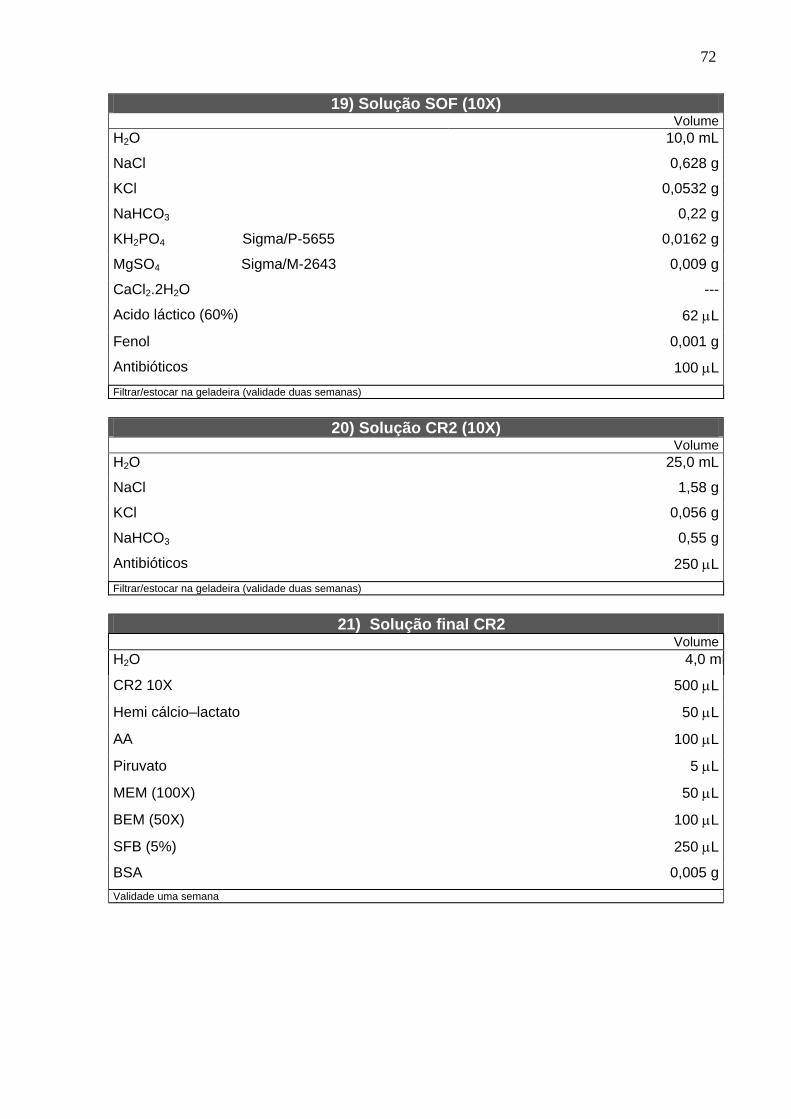
72
19) Solução SOF (10X) VolumeH2O 10,0 mL
NaCl 0,628 g
KCl 0,0532 g
NaHCO3 0,22 g
KH2PO4 Sigma/P-5655 0,0162 g
MgSO4 Sigma/M-2643 0,009 g
CaCl2.2H2O ---
Acido láctico (60%) 62 μL
Fenol 0,001 g
Antibióticos 100 μLFiltrar/estocar na geladeira (validade duas semanas)
20) Solução CR2 (10X)
VolumeH2O 25,0 mL
NaCl 1,58 g
KCl 0,056 g
NaHCO3 0,55 g
Antibióticos 250 μLFiltrar/estocar na geladeira (validade duas semanas)
21) Solução final CR2
VolumeH2O 4,0 mL
CR2 10X 500 μL
Hemi cálcio–lactato 50 μL
AA 100 μL
Piruvato 5 μL
MEM (100X) 50 μL
BEM (50X) 100 μL
SFB (5%) 250 μL
BSA 0,005 gValidade uma semana
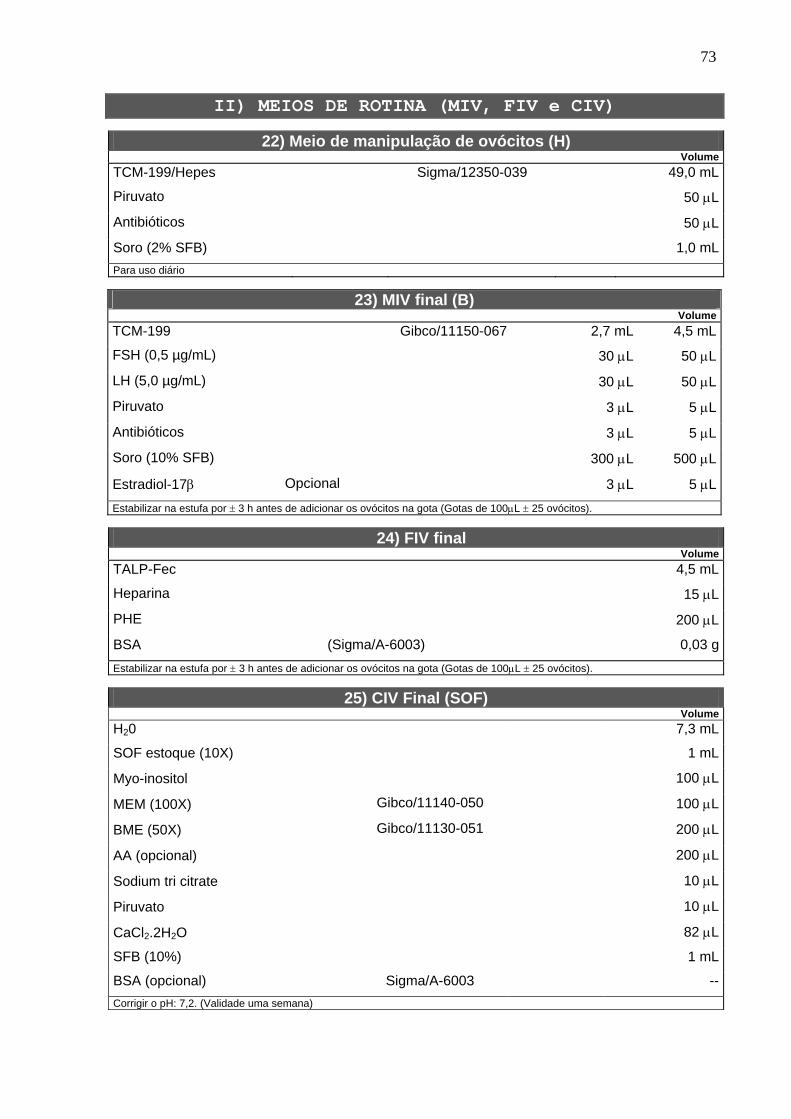
73
II) MEIOS DE ROTINA (MIV, FIV e CIV)
22) Meio de manipulação de ovócitos (H) Volume
TCM-199/Hepes Sigma/12350-039 49,0 mL
Piruvato 50 μL
Antibióticos 50 μL
Soro (2% SFB) 1,0 mLPara uso diário
23) MIV final (B) Volume
TCM-199 Gibco/11150-067 2,7 mL 4,5 mL
FSH (0,5 µg/mL) 30 μL 50 μL
LH (5,0 µg/mL) 30 μL 50 μL
Piruvato 3 μL 5 μL
Antibióticos 3 μL 5 μL
Soro (10% SFB) 300 μL 500 μL
Estradiol-17β Opcional 3 μL 5 μLEstabilizar na estufa por ± 3 h antes de adicionar os ovócitos na gota (Gotas de 100μL ± 25 ovócitos).
24) FIV final Volume
TALP-Fec 4,5 mL
Heparina 15 μL
PHE 200 μL
BSA (Sigma/A-6003) 0,03 g Estabilizar na estufa por ± 3 h antes de adicionar os ovócitos na gota (Gotas de 100μL ± 25 ovócitos).
25) CIV Final (SOF) Volume
H20 7,3 mL
SOF estoque (10X) 1 mL
Myo-inositol 100 μL
MEM (100X) Gibco/11140-050 100 μL
BME (50X) Gibco/11130-051 200 μL
AA (opcional) 200 μL
Sodium tri citrate 10 μL
Piruvato 10 μL
CaCl2.2H2O 82 μL
SFB (10%) 1 mL
BSA (opcional) Sigma/A-6003 --Corrigir o pH: 7,2. (Validade uma semana)
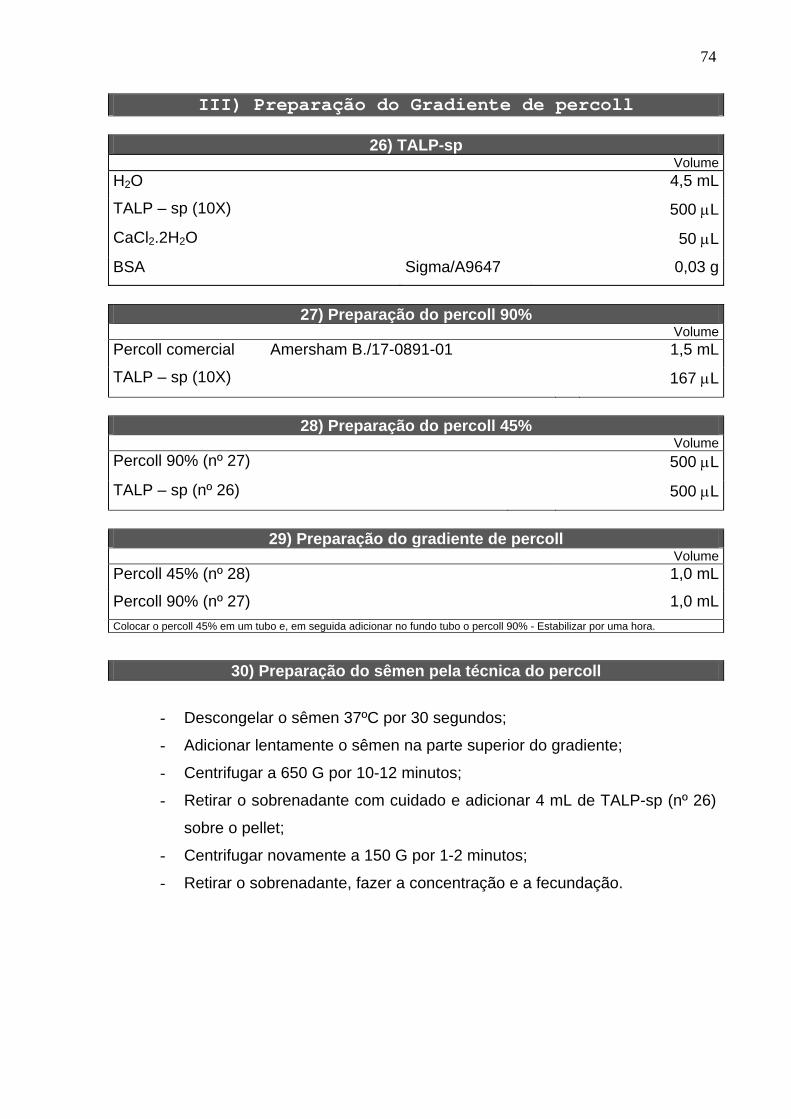
74
III) Preparação do Gradiente de percoll
26) TALP-sp VolumeH2O 4,5 mL
TALP – sp (10X) 500 μL
CaCl2.2H2O 50 μL
BSA Sigma/A9647 0,03 g
27) Preparação do percoll 90%
VolumePercoll comercial Amersham B./17-0891-01 1,5 mL
TALP – sp (10X) 167 μL
28) Preparação do percoll 45%
VolumePercoll 90% (nº 27) 500 μL
TALP – sp (nº 26) 500 μL
29) Preparação do gradiente de percoll
VolumePercoll 45% (nº 28) 1,0 mL
Percoll 90% (nº 27) 1,0 mLColocar o percoll 45% em um tubo e, em seguida adicionar no fundo tubo o percoll 90% - Estabilizar por uma hora.
30) Preparação do sêmen pela técnica do percoll
- Descongelar o sêmen 37ºC por 30 segundos;
- Adicionar lentamente o sêmen na parte superior do gradiente;
- Centrifugar a 650 G por 10-12 minutos;
- Retirar o sobrenadante com cuidado e adicionar 4 mL de TALP-sp (nº 26)
sobre o pellet;
- Centrifugar novamente a 150 G por 1-2 minutos;
- Retirar o sobrenadante, fazer a concentração e a fecundação.

75
31) Cálculo para concentração de espermatozóides (spz) - Prepara um micro tubo com 190 μL de água;
- Retira-se uma amostra de 10 μL Spz para concentração;
- Homogeneizar bem;
- Fazer a contagem de Spz na câmara de Neubauer;
- Contar 5 quadrante de cada lado da câmara;
- Somar o nº de Spz contados nos dois lados da câmara e fazer a média;
- Fazer os cálculos - CI . VI = CF . VF;
- CI = concentração inicial – nº de Spz contado na câmara (média ex. ~46);
- VI = volume inicial –?;
- CF = concentração/final Spz/mL utilizado na fecundação (2,0 .106 Spz/mL);
- VF = volume final – volume da gota de FIV (Ex. Gota de 100 μL);
- Fazer os cálculos para saber o volume há ser utilizado na FIV;
- Exemplo. VI = CF . VF ⇒ VI = 2,0 .106 . 100
CI ~46.106
- VI = ~4,3μL → Volume de Spz utilizado para inseminação dos ovócitos.
Preparação do sêmen pela técnica Swim – up
32) TALP-sp volumeH2O 9,0 mL
TALP – sp (10X) 1,0 mL
CaCl2.2H2O 50 μL
BSA 0,06 gDeixar na estufa até dissolver totalmente o BSA
33) Procedimentos de preparação do sêmen pelo Swim – up
- Descongelar o sêmen 37ºC por 30 segundos;
- Colocar 1mL de TALP-sp (nº 32) em um tubo e, em seguida adicionar o
sêmen no fundo do tubo;
- Deixar o tubo na estufa inclinado a 45º por uma hora;
- Transferir 850 μL da fração superior para outro tubo com 4mL de TALP-sp;
- Centrifugar a 120 G por 5 minutos;
- Retirar o sobrenadante e ressuspender o pellet com 4 mL de TALP-sp;
- Centrifugar novamente a 120 G por 5 minutos;
- Retirar o sobrenadante, fazer a concentração e a fecundação.

76
IV) Meios para coloração
34) Solução de PBS pH - 7.4 – filtrar/estocar na geladeira (validade dois messes) volumeH2O (q.s.p.) 500 mL
NaCl 4.0 g
Na2HPO4 7H2O Nuclear/3168 (Obs: Na2HPO4 = 0,575g) 1,08 g
D-glicose Sigma/G-7021 0.5 g
KCl 0.1 g
KH2PO4 0.1 g
MgCl2 6H2O 0.05 g
Piruvato de sódio 0.018 g
Estreptomicina 0.025 g
Penicilina 0.029 g
Vermelho fenol 0.002 g
CaCl2.2H2O 0.066 g
35) Solução Bloqueio (SB)
VolumePBS 100 mL
BSA 0,1 g
Glicina 0,751 g
Azida sódica Sigma/S-8032 0,2 g
36) Coloração dos Grânulos corticais
1- Desnudar os ovócitos em PBS + 1% de SFB (vortex 4 min);
2- Remoção da zona pelúcida: PBS + 0,5% de protease (±8 min);
3- Fixação: PBS + 3% de paraformaldeído (PFD) (30 min)(Sigma/P6148);
4- Incubação: SB (2h);
5- Permeabilização: SB + 0,1% de Triton X-100 (5 min)(USB/9002-93-1);
6- Coloração: SB com 1 μg/mL de lens culinaris (Sigma/L9262) + 10 μg/mL de Iodeto
de propídio (15 min) (Sigma/P4170);
7- Lavar os ovócitos 3 vezes em SB;
8- Montagem das lâminas com glicerol (USB/CAS.56-81-5);
9- Visualização em microscópio de epifluorescência.
“lens culinaris” aglutinina conjugada a FITC (esta lecitina se liga especificamente à α-D-manose presente nos grânulos
corticais).

77
37) Coloração do Citoesqueleto (microtúbulos)
1- Solução PP: PBS + 0,1% de PVA;
2- Desnudar os ovócitos em PBS + 1% de SFB (vortex 4 min);
3- Fixação: PBS com 3% de PFD + 0.6% de Triton X-100 (30 min);
4- Bloqueio: PP com 3% de Soro de Cabra (45 min);
5- Coloração: PP com α-Tubulina conjugada com FITC (1:100)(1h);
6- Coloração: PP com 10 μg/mL de Iodeto de propídio (15min);
7- Lavar os ovócitos 3 vezes em PP;
8- Montagem das lâminas em glicerol;
9- Visualização em microscópio de epifluorescência.
38) Coloração do Citoesqueleto (microfilamentos)
1- Solução PP: PBS + 0,1% de PVA;
2- Desnudar os ovócitos em PBS + 1% de SFB (vortex 4 min);
3- Fixação: PBS com 3% de PFD + 0.6% de Triton X-100 (30 min);
4- Coloração: PP + 1 μg/mL de Faloidina conjugada com FITC + 10 μg/mL de Iodeto de
propídio (30 min);
5- Lavar os ovócitos 3 vezes em PP;
6- Montagem das lâminas em glicerol;
7- Visualização em microscópio de epifluorescência.
39) Coloração das Mitocôndrias
1- Desnudar os ovócitos em PBS + 1% de SFB (vortex 4 min);
2- Coloração: TCM-199/hepes + 10% de SFB com 0,5 μM de Mitotracker red + 10 μg/mL
de hoechst (20 min);
3- Lavar os ovócitos 3 vezes em TCM-199/hepes + 2% de SFB;
4- Montagem das lâminas;
5- Visualização em microscópio de epifluorescência.
40) Recomendações gerais
- Usar produtos de marcas de confiança (Sigma e Gibco etc.);
- Usar Substâncias “embryo tested” preferencialmente;
- Usar água testada para PIV de embriões;
- Filtrar sempre;
- Deixar os meios de MIV, FIV e CIV estabilizando em overnight;
- Reduzir um pouco incidência de luz na sala de trabalho;

78
- As manipulações devem ser as mais rápidas possíveis;
- Se possível não usar material de vidro ou plástico reciclado;
- Cuidado com oscilações de temperatura durante os procedimentos, se necessário
usar placa aquecedora;
- Atenção com o pH dos meios – pH entre 7,2 e 7,4;
- Ajustar a osmolaridade – mOsm entre 280 – 290.
Para corrigir o pH usar 1M HCl ou 1M NaOH
Para corrigir a osmolaridade usar NaCl (↑ mOsm) ou H2O (↓ mOsm)





























