Embed Size (px)
Citation preview

PENGARUH KOMBINASI NAPHTHALENE ACETIC ACID (NAA) DAN KINETIN PADA KULTUR IN VITRO EKSPLAN DAUN TEMBAKAU
(Nicotiana tabacum L. var. Prancak 95)
Daniar robbiani*, Tutik Nurhidayati1, Nurul Jadid1
Program Studi Biologi, Fakultas Matematika Ilmu Pengetahuan Alam Institut Teknologi Sepuluh Nopember Surabaya
ABSTRAK
Penelitian Pengaruh Kombinasi Naphthalene Acetic Acid (NAA) dan Kinetin Terhadap
Kultur In vitro Eksplan Daun Tembakau (Nicotiana tabacum L. var. Prancak 95) bertujuan untuk menentukan kombinasi konsentrasi NAA dan Kinetin yang efektif dan mengetahui pengaruh morfogenesis eksplan dari kombinasi tersebut. Penelitian ini dilaksanakan di Laboratorium Kultur Jaringan Tumbuhan Program Studi Biologi ITS Surabaya. Penelitian ini disusun dalam Rancangan Acak Lengkap (RAL) dengan dua faktor. Faktor pertama konsentrasi NAA, terdiri atas 6 level yaitu 0 ppm; 0,5 ppm; 1 ppm; 1,5 ppm; 2 ppm, dan 2,5 ppm. Faktor kedua konsentrasi Kinetin, terdiri atas 5 level yaitu 0 ppm; 1 ppm; 2 ppm; 3 ppm; and 4 ppm. Hasil menunjukkan bahwa jumlah proliferasi tunas tertinggi diperoleh pada perlakuan dengan 0 ppm NAA dan 4 ppm Kinetin (62,75 tunas/eksplan) dan perlakuan dengan 2,5 ppm NAA dan 0 ppm Kinetin memberikan hasil terbaik untuk proliferasi akar (37,75 akar/eksplan). Kalus yang didapatkan paling dominan berwarna putih kehijauan dan tekstur kompak. Kata kunci: Nicotiana tabacum L. var. Prancak 95, NAA, Kinetin, Kalus, Kultur jaringan tumbuhan
ABSTRACT
The research of the influence of Naphthalene Acetic Acid (NAA) and Kinetin combination on in vitro culture of Tobacco leaf explant (Nicotiana tabacum L. var. Prancak 95) was conducted to determine the effective combination of NAA and Kinetin concentration and to observe the explant morphogenetic effect of its combination. This research was carried out in the plant tissue culture laboratory, department of Biology ITS, Surabaya. This research was designed with completely randomize design with two factors. The first factor was the NAA concentration, consisted of six levels i.e. 0 ppm; 0.5 ppm; 1 ppm; 1,5 ppm; 2 ppm; and 2,5 ppm. The second factor was Kinetin concentration, consisted of five levels i.e. and 0 ppm ; 1 ppm; 2 ppm, 3 ppm, and 4 ppm. The result showed that highest amount of shoot proliferation was obtained from treatment with 0 ppm NAA and 4 ppm Kinetin (62,75 shoot/explant) and the treatment for 2,5 ppm NAA and 0 ppm Kinetin gave the best result for root proliferation (37,75 root/explant). The most dominant callus obtained were white greenish in colour and compact in texture. Key words: Nicotiana tabacum L. var. Prancak 95, NAA, Kinetin, Callus, Plant Tissue Culture *Corresponding author Phone : +6285719149996 e-mail : [email protected] 1 Alamat sekarang : Prodi Biologi, Fak MIPA, Institut Teknologi Sepuluh Nopember, Surabaya
I PENDAHULUAN Penanaman dan penggunaan tembakau di Indonesia Penanaman dan penggunaan tembakau di Indonesia sudah dikenal sejak lama. Komoditi tembakau mempunyai arti yang cukup penting, tidak hanya sebagai
sumber pendapatan bagi para petani, tetapi juga bagi Negara. Tembakau (daunnya) digunakan sebagai bahan pembuatan rokok (Hanum, 2008). Tembakau mengandung alkaloid nikotin yang berdampak buruk bagi kesehatan manusia juga sangat beracun bagi

serangga sehingga nikotin dapat dimanfaatkan oleh manusia sebagai bioinsektisida (Susilowati, 2006). Tembakau juga dapat dimanfaatkan sebagai pewarna kain sutera dengan menggunakan daunnya untuk menjadi larutan celup pada proses pencelupan kain sutera (Santosa, 2007). Keistimewaan dan manfaat yang besar dari tembakau membuat tembakau mempunyai potensi untuk dikembangkan dalam berbagai bidang. Salah satu tanaman yang sering dikembangkan adalah tembakau Madura. Pada saat ini tembakau Madura yang berkembang sebagai bahan baku rokok adalah adalah var. Prancak 95 dan Cangkring 95 (Basuki et al., 1999). Keunggulan tembakau var. Prancak 95 adalah memiliki sifat hasil sedang, mutu tinggi, aromanya khas, kadar nikotin rendah, tahan terhadap penyakit lanas dan sesuai ditanam di lahan tegal dan gunung. Selama ini budidaya tembakau Madura dilakukan secara konvensional. Budidaya tembakau secara konvensional memerlukan proses yang tidak sederhana dan waktu yang relatif lama, selain itu sifat-sifat genetis tidak sama persis seperti induknya. Seiring dengan berkembangnya ilmu pengetahuan, tanaman tembakau juga mulai dibudidayakan melalui program pemuliaan tanaman. Hal tersebut bertujuan untuk meningkatkan jumlah produksi tanaman tembakau. Salah satu program pemuliaan tanaman tembakau tersebut adalah dengan teknik kultur jaringan. Manfaat utama kultur jaringan adalah menghasilkan tanaman baru dalam jumlah yang besar dalam jangka waktu yang relatif singkat dengan sifat dan kualitas yang diharapkan sama dengan induknya (Rahardja, 1995 dalam Yunus, 2007). Salah satu perbanyakan tanaman tembakau secara in vitro yang efisien adalah dengan mengkulturkan organ yaitu eksplan dari daun muda tembakau (Hendaryono, 1994). Penggunaan eksplan dari jaringan muda lebih sering berhasil karena sel-selnya aktif membelah, dinding sel tipis karena belum terjadi penebalan lignin dan selulose yang menyebabkan kekakuan pada sel.
Selain itu faktor lain yang memberikan pengaruh terhadap keberhasilan perbanyakan tanaman secara in vitro adalah zat pengatur tumbuh. Zat pengatur tumbuh yang banyak digunakan dalam kultur jaringan adalah auksin dan sitokinin. NAA (Naphthalene Acetic Acid) merupakan salah satu auksin yang berperan dalam perpanjangan sel. Sedangkan Kinetin (6-furfury amino purine) adalah salah satu sitokinin yang berperan untuk pembelahan sel. Sitokinin bersama-sama dengan auksin akan memberikan pengaruh interaksi terhadap diferensiasi jaringan dalam kultur jaringan tanaman (Hendaryono, 1994).
Penelitian terhadap interaksi antara kinetin dan auksin pada kultur tembakau telah membuktikan adanya peranan dari kedua zat tumbuh ini terhadap pertumbuhan. Kinetin yang berimbang dengan auksin dapat menyebabkan pertumbuhan kalus (Abidin, 1985 dalam Fitrianti, 2006). Jumlah auksin dan sitokinin yang perlu ditambahkan kedalam kultur tergantung kandungan auksin dan sitokinin endogen pada eksplan. Berdasarkan penelitian Maryanto (1987) dalam Suryowinoto (1991) pada kultur tembakau (Nicotiana tabacum) dengan perbandingan auksin : kinetin 5:0 atau 4:1 hanya terjadi pertumbuhan akar saja. Pada jumlah perbandingan sebaliknya yaitu auksin : kinetin 0:5 atau 1:4 hanya terjadi tunas besar, tanpa ada akar sama sekali. Sedangkan menurut Suryowinoto (1991) dalam Hendaryono (1994), pada kultur in vitro daun muda tembakau pada medium MS perlu ditambahkan zat pengatur tumbuh IAA dan Kinetin dengan perbandingan 2:3 ppm.
Perbandingan konsentrasi yang tepat antara sitokinin dan auksin akan memacu pertumbuhan eksplan kultur in vitro. Oleh karena itu, konsentrasi zat pengatur tumbuh perlu diperhatikan untuk keberhasilan teknik kultur jaringan. Berdasarkan observasi di atas maka dilakukan penelitian kultur jaringan tembakau dengan penambahan dua jenis zat pengatur tumbuh yaitu auksin (NAA) dan Sitokinin (Kinetin) pada berbagai konsentrasi.

II METODOLOGI 1 Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan Februari – Maret 2010 di Laboratorium Kultur Jaringan Program Studi Biologi ITS. Jenis tembakau Madura yang digunakan adalah (Nicotiana tabacum L.var. Prancak 95) yang diperoleh dari PT. Sadhana-Pasuruan. 2 Alat, Bahan, dan Cara Kerja 2.1 Tahap Persiapan a. Sterilisasi Alat dan Ruang Kerja
Semua peralatan baik alat pembuatan media (botol kultur) dan alat inokulasi eksplan (cawan petri, scalpel blade, gunting eksplan, pinset, kertas saring, dll) disterilisasi dengan autoklaf dengan suhu 121oC tekanan 1,5 atm selama 15 menit (Nugroho, 2004). Laminair Air Flow (LAF) disemprot dengan alkohol 70% dan alat-alat yang dimasukkan ke dalam LAF juga harus disemprot dengan alkohol 70%. Ruang tanam disterilisasi dengan sinar UV selama 1 jam sebelum LAF digunakan. Ketika LAF digunakan maka sinar UV harus dimatikan dan blower dihidupkan (Fitrianti, 2006). b. Sterilisasi Eksplan
Sterilisasi permukaan eksplan daun ini ada 2 tahap yaitu sterilisasi tahap I yang dilakukan di ruang persiapan dan sterilisasi tahap II yang dilakukan di LAF. Sterilisasi tahap I meliputi: Daun tembakau muda (daun kedua sampai ketiga dari pucuk) diambil dari green house dibilas dengan air mengalir hingga bersih. Sedangkan sterilisasi tahap II meliputi: Daun tembakau dimasukkan ke dalam 70 % etanol selama 0,5 menit. Kemudian dibilas dengan aquades steril selama 5 menit. Potongan daun tembakau disterilisasi dengan 1% sodium hypochlorite (Bayclin ™) selama ± 10 menit. Kemudian dibilas tiga kali dengan aquades steril selama 5 menit sebanyak 3 kali sambil digojog. Selanjutnya eksplan diambil dengan pinset dan ditiriskan pada kertas saring. (Fowke, L.C. et al, 1983). 2.2 Pembuatan Media a. Pembuatan Stok Zat Pengatur Tumbuh NAA dan Kinetin
Untuk pembuatan larutan stok NAA (MerckTM), dilakukan penimbangan bahan sebanyak 10 mg dan dimasukkan ke dalam erlenmeyer berukuran 100 ml yang diberi aquades sedikit. Sambil diaduk, diteteskan sedikit demi sedikit larutan NaOH 1 N dengan hati-hati sambil dikocok sampai zat pengatur
tumbuh larut benar. Kemudian ditambahkan aquades steril ke dalam erlenmeyer hingga volume mendekati 70 ml sambil dikocok-kocok. Kemudian ditambahkan kembali aquades steril ke dalam erlenmeyer sampai volume menjadi 100 ml. Kemudian larutan dipindahkan ke dalam wadah stok, ditutup rapat dengan aluminium foil dan kapas dan diberi label NAA. Selanjutnya disimpan dalam lemari es. Untuk pembuatan larutan stok Kinetin (TCITM) caranya juga sama seperti pembuatan larutan stok NAA. Apabila zat pengatur tumbuh belum larut, dapat dibantu dengan memanaskannya dengan pemanas atau hot plate (Santoso dan Nursandi, 2003).
Perhitungan perlakuan zat pengatur tumbuh (Lampiran 4) menggunakan rumus pengenceran yaitu: V1.M1 = V2.M2 V1 = volume larutan stok yang dicari M1 = dosis larutan stok yang tersedia V2 = volume medium yang akan dibuat M2 = dosis medium yang akan dibuat
(Hendaryono, 1994).
b. Pembuatan Media Kultur (Media MS) Media kultur yang digunakan dalam
penelitian ini merupakan media MS (Lampiran 1) yang terdiri dari unsur mikro, unsur makro, sukrosa, vitamin, agar, dan zat pengatur tumbuh (NAA dan Kinetin). Untuk pembuatan media MS, erlenmeyer berukuran 1 liter disiapkan lalu sebanyak 500 ml medium MS cair siap pakai yang sudah mengandung unsur mikro, unsur makro, sukrosa, vitamin, dipanaskan sambil diaduk-aduk. Kemudian ditambahkan zat pengatur tumbuh NAA sesuai konsentrasi (dalam hal ini digunakan konsentrasi 0 ppm; 0,5 ppm; 1 ppm; 1,5 ppm; dan 2 ppm) dan Kinetin (dalam hal ini digunakan konsentrasi 0 ppm; 1 ppm; 2 ppm; dan 3ppm) sambil diaduk homogen di atas pemanas. Selanjutnya diukur pH larutan 5,8 menggunakan pH meter. Apabila terlalu asam ditambahkan NaOH dan apabila terlalu basa ditambahkan HCl. Jika pH telah sesuai, ditambahkan agar-agar sebanyak 8 gr dan medium MS cair ditambahkan kembali hingga volume 1 L. Media dididihkan dan diaduk hingga agar-agar larut dan tercampur rata kemudian dibagi media sekitar 25 ml/botol ke dalam botol kultur dalam keadaan masih cair. Botol kultur ditutup rapat dengan penutup plastik dan diautoklaf pada suhu 121oC, tekanan 1,5 atm selama 15 menit. Setelah itu, diberi label sesuai

perlakuan dan disimpan di dalam ruang steril (Hendaryono, 1994).
2.3 Inokulasi Eksplan
Proses inokulasi dilakukan di laminar air flow dengan kondisi aseptik. Alat-alat inokulasi ditata didalam laminar air flow. Setiap alat tersebut dicelupkan ke dalam alkohol 70% dan dipanaskan di atas nyala api bunsen selama 1-2 menit. Bunsen yang akan dipakai hendaknya terisi penuh. Daun Nicotiana tabacum L. dikeluarkan dari botol sterilisasi dan diletakkan pada cawan petri steril yang telah dilapisi kertas tissue/kertas serap steril untuk menyerap aquades. Kemudian daun dipotong-potong persegi di atas petridish dengan ukuran 0,5 - 1 cm2. Eksplan tersebut kemudian diinokulasikan ke dalam botol kultur yang telah berisi media MS modifikasi dengan posisi horizontal (mendatar) dan bagian abaksial menempel pada permukaan medium (Dhaliwal et al., 2004). Media MS modifikasi ini terdiri atas unsur makro, unsur mikro, sukrosa, vitamin, agar, zat pengatur tumbuh NAA dan Kinetin. Setiap botol kultur berisi 2 eksplan. Botol ditutup rapat dan diberi label yaitu tanggal dilakukan inokulasi eksplan dan konsentrasi hormon yang digunakan. Kemudian ditata rapi dalam rak kultur bertingkat. Botol berisi eksplan diinkubasi pada suhu 25-28oC, kelembaban 70% dengan fotoperiode 12 jam terang dan 12 jam gelap selama ± 1 bulan. Setiap kolom rak kultur diberi pencahayaan dengan lampu flourescen 40 watt (Gunawan, 1995).
3. Rancangan Penelitian dan Hipotesis
3.1 Rancangan Penelitian Rancangan penelitian ini menggunakan
rancangan acak lengkap (RAL) pola faktorial yang terdiri dari 2 faktor (Faktor 1= konsentrasi NAA dan Faktor 2=konsentrasi Kinetin) dan masing-masing 4 kali ulangan. Rancangan penelitian disajikan pada tabel 2. 3.2 Uji Kuantitatif
Jika eksplan yang ditumbuhkan menghasilkan tunas atau akar, maka akan dihitung jumlah tunas dan jumlah akar, selanjutnya seluruh data yang diperoleh dianalisis dengan menggunakan ANOVA dan jika ada pengaruh maka dilanjutkan dengan uji Tukey dengan tingkat kesalahan 5% menggunakan Minitab.
Adapun hipotesis dalam penelitian ini adalah sebagai berikut :
H0 = Tidak ada pengaruh kombinasi zat pengatur tumbuh NAA dan Kinetin terhadap morfogenesis eksplan tanaman tembakau.
H1 = Ada pengaruh kombinasi zat pengatur tumbuh NAA dan Kinetin terhadap morfogenesis eksplan tanaman tembakau. Sedangkan variabel yang digunakan
adalah : Variabel bebas : perbandingan konsentrasi
zat pengatur tumbuh NAA dan Kinetin
Variabel terikat : jumlah akar dan jumlah tunas
Variabel terkendali : pH, suhu, kelembaban, dan pencahayaan.
3.3 Uji Kualitatif a. Kalus
Jika eksplan yang ditumbuhkan menghasilkan kalus maka dilakukan pengamatan secara deskriptif yaitu morfologi kalus. Menurut Ali et al. (2007) warna kalus yang terbentuk antara lain kuning, kehijauan, dan hijau terang sedangkan tekstur kalus yang terbentuk adalah lunak, keras, dan kompak (Lampiran 3). b. Respon Organogenesis dan Callogenesis
Respon organogenesis dan callogenesis dilakukan pengukuran persentase pertumbuhan eksplan yang meliputi respon callogenesis (eksplan membentuk kalus), respon organogenesis tidak langsung (eksplan berkalus bertunas, eksplan berkalus berakar, eksplan berkalus bertunas berakar), dan respon organogenesis langsung (eksplan bertunas, eksplan berakar, eksplan bertunas dan berakar). IV HASIL DAN PEMBAHASAN 4.1 Respon Callogenesis Kultur In Vitro
Nicotiana tabacum L. var Prancak 95 Penambahan zat pengatur tumbuh NAA
dan Kinetin pada kultur in vitro eksplan daun tembakau Madura (Nicotiana tabacum L. var. Prancak 95) dengan konsentrasi yang berbeda yaitu sebanyak 30 kombinasi memberikan respon organogenesis dan callogenesis yang bervariasi. Penelitian dilakukan selama 30 hari untuk mengetahui respon morfogenesis dari Nicotiana tabacum var. Prancak 95 yang ditandai dengan terbentuknya kalus, tunas, dan akar dalam bentuk persentase (Tabel 4.1).
Selain itu pengamatan secara deskriptif juga dilakukan untuk mengetahui adanya respon callogenesis, organogenesis langsung dan tidak langsung. Callogenesis merupakan respon awal

yang ditandai dengan terbentuknya kalus yang mulai terbentuk pada bagian tepi eksplan (bagian perlukaan) bagian atas maupun bagian bawah yang bersentuhan dengan media, tetapi kalus lebih cepat terbentuk pada bagian yang bersentuhan dengan media, yaitu bagian abaksial daun. Hal ini kemungkinan berkaitan dengan proses pengambilan nutrisi medium oleh eksplan. Penyerapan unsur hara akan lebih baik karena terjadi kontak langsung antara media dengan bagian abaksial daun. Munculnya kalus pada bagian yang terluka diduga karena adanya rangsangan dari jaringan pada eksplan untuk menutupi lukanya. Hal ini sesuai pendapat dari Thomas dan Davey (1975) dalam George and Sherington (1993), mengemukakan bahwa pembelahan sel yang mengarah pada terbentuknya kalus terjadi dari adanya respon terhadap luka dan suplai hormon alamiah atau buatan dari luar ke dalam eksplan. Pada perlakuan 0 ppm NAA dan 0 ppm Kinetin eksplan memberikan respon dengan penambahan volume yaitu eksplan agak membesar dan melengkung dan terbentuk tonjolan kecil yaitu kalus tanpa disertai adanya respon organogenesis yang nantinya akan tumbuh menjadi tunas, atau akar (Gambar 4.1).
Gambar 4.1. Eksplan dengan perlakuan 0 ppm NAA dan 0 ppm Kinetin
Berdasarkan Tabel 4.1, respon callogenesis pada eksplan Nicotiana tabacum L. terjadi pada semua perlakuan dengan persentase 100% dan 25 %, kecuali pada perlakuan 0 ppm NAA dengan penambahan Kinetin dengan persentase 0% (Tabel 4.1).
Tumbuhnya kalus pada perlakuan 0 ppm
NAA dan 0 ppm Kinetin disebabkan adanya pengaruh hormon endogen yang terkandung dalam eksplan itu sendiri. Hormon endogen tersebut juga mampu memacu sel untuk berkembang dan memperbanyak diri tetapi waktu yang dibutuhkan cenderung lama karena jumlah hormon yang tidak tersedia secara pasti. Hal ini membuktikan bahwa terbentuknya kalus sangat dipengaruhi oleh peran jenis zat pengatur tumbuh. Menurut Zulfiqar et al., (2009) kondisi tersebut membuktikan bahwa pertumbuhan dan morfogenesis tanaman secara in vitro dikendalikan oleh keseimbangan dan interaksi dari ZPT yang ada dalam eksplan baik endogen maupun eksogen yang diserap dari media.
Pada perlakuan Kinetin tanpa penambahan NAA, tidak terbentuk kalus. Sedangkan pada perlakuan NAA tanpa penambahan Kinetin terbentuk kalus, begitupula pada perlakuan dengan interaksi NAA dan Kinetin. Berarti dalam kasus ini NAA lebih berperan dalam pembentukan kalus daripada Kinetin. NAA (auksin) akan merangsang pertumbuhan sel-sel eksplan, sehingga auksin akan cenderung membentuk kalus karena terbentuknya kalus berawal dari pembelahan sel pada daerah meristematik yang tidak terspesialisasi. Pada awal respon pertumbuhan, auksin akan memicu pemanjangan sel melalui pelonggaran selulosa dinding sel. Pemanjangan sel ini sebagai respon terhadap NAA, namun sel tersebut tidak dapat membelah karena tidak ada penambahan Kinetin. Jika hanya Kinetin saja yang ditambahkan ke
Kalus

dalam medium kultur, maka tidak akan ada pengaruh apapun tehadap tumbuhnya kalus karena Kinetin lebih berperan terhadap pembelahan sel serta diferensiasi terbentuknya tunas. Namun, jika Kinetin ditambahkan bersama-sama dengan auksin maka sel-sel akan mengalami pembelahan dan perkembangan secara terus menerus. Rasio sitokinin terhadap auksin mengontrol diferensiasi sel. Ketika konsentrasi kedua hormon tersebut hampir sama, massa sel akan terus bertambah (terbentuk kalus). Penggunaan medium MS dengan penambahan NAA dengan konsentrasi lebih tinggi daripada konsentrasi Kinetin dapat menginduksi proliferasi kalus. Zia et al., (2007) menyebutkan bahwa bahwa kalus dapat tumbuh pada beragam konsentrasi dan jenis zat pengatur tumbuh, namun untuk zat pengatur tumbuh Kinetin pada eksplan daun, kalus hanya tumbuh pada Kinetin dengan konsentrasi yang rendah yaitu dibawah 1 ppm, dan diatas 1 ppm kalus tidak dapat tumbuh. Sedangkan pada zat pengatur tumbuh NAA, kalus dapat tumbuh di semua konsentrasi dan sebagian besar menunjukkan persentase 100%. Selain itu, interaksi zat pengatur tumbuh juga berpengaruh tehadap warna dan tekstur kalus. Adapun tekstur dan warna kalus disajikan pada Tabel 4.2
Morfologi kalus yang terbentuk pada pengamatan selama 30 hari meliputi warna putih, putih kehijauan, hijau, dan coklat. Sedangkan tekstur yang terbentuk yaitu kompak dan remah (Gambar 4.2).
Morfologi kalus yang terbentuk dominan berwarna putih kehijauan dengan tekstur kompak. Respon eksplan yang berasal dari media dengan penambahan Kinetin memiliki tekstur yang lebih kompak dan dominan berwarna putih kehijauan. Dalam penelitian ini dominan kalus yang muncul dari daerah bekas pelukaan berwarna putih, lama-kelamaan akan berubah berwarna putih kehijauan dan kemudian berdiferensiasi menjadi organ-organ vegetatif yakni tunas dan akar. Hal tersebut terjadi kemungkinan karena adanya kesesuaian antara medium, zat pengatur tumbuh yang digunakan dengan faktor lingkungan. Warna hijau ini disebabkan kalus mengandung klorofil, akibat interaksi NAA dan Kinetin terutama Kinetin (sitokinin) yang berperan dalam pembentukan klorofil pada kalus serta faktor lingkungan yaitu paparan cahaya.. Hal ini sesuai dengan pendapat Leupin (2000) bahwa perubahan warna kalus menjadi putih kehijauan, kemungkinan pada sel kalus sudah mulai terbentuk klorofil. Kalus yang berwarna putih tidak mengandung kloroplas, tetapi mengandung plastid yang berisi butir pati yang sedikit-demi sedikit tumbuh menjadi sistem membran yang jelas yang akhirnya terbentuklah butir-butir klorofil dengan paparan cahaya, sehingga kalus menjadi berwarna hijau. Hal tersebut sesuai dengan pernyataan George & Sherrington (1993), bahwa cahaya putih dapat merangsang pembentukan kalus dan

organogenesis dalam kultur jaringan tumbuhan.
Kalus kompak memiliki struktur yang terorganisasi dan ditandai dengan nodul berwarna hijau dan sangat baik untuk regenerasi planlet. Tekstur kalus kompak merupakan efek dari sitokinin yang berperan dalam transport zat hara. Sistem transport sitokinin dari bagian basal ke apeks akan membawa air dan zat hara melalui pembuluh pengangkut dan mempengaruhi potensial osmotik dalam sel. Penambahan sukrosa dalam medium akan mengalir melalui pembuluh floem dan menimbulkan tekanan turgor. Tekanan tersebut muncul akibat adanya perbedaan konsentrasi larutan, sehingga air dan zat hara (sukrosa) dari medium akan masuk kedalam sel melalui cara osmosis. Hal ini akan membuat dinding-dinding sel semakin kaku, sehingga sel kalus akan menjadi kompak. Sukrosa yang merupakan karbohidrat sebagai cadangan makanan ini akan diubah menjadi pati yang digunakan sebagai energi pada proses morfogenesis eksplan, sehingga dapat membantu sel untuk terus membelah. Menurut Purwianingsih (2007), struktur kalus yang kompak dan terjadi perubahan warna kekuningan atau kehijauan, mengindikasikan terjadinya diferensiasi sel. Kalus remah didapatkan pada perlakuan dengan NAA lebih tinggi dari Kinetin dan berwarna coklat karena terdapat pengaruh komposisi medium dan zat pengatur tumbuh. Kalus remah ini terjadi melalui proses pertumbuhan yang mengarah pada pembentukan sel-sel yang berukuran kecil dan berikatan longgar. Dalam hal ini, auksin memiliki peran terhadap pembentukan kalus remah. NAA menstimulasi pemanjangan sel dengan cara penambahan plastisitas dinding sel menjadi longgar, sehingga air dapat masuk ke dalam dinding sel dengan cara osmosis dan sel mengalami pemanjangan. Oleh karena itu, kalus yang remah mengadung banyak air karena belum mengalami lignifikasi dinding sel, antara kumpulan sel yang satu dengan yang lain relatif mudah untuk dipisahkan. Pada penelitian ini, kalus remah pada perlakuan
NAA dengan konsentrasi yang tinggi tumbuh akar, namun tidak mengalami pemanjangan, sedangkan pada perlakuan kombinasi NAA dan Kinetin yang tinggi tidak dapat berdiferensiasi menjadi tunas atau akar, karena zat pengatur tumbuh dengan konsentrasi tinggi ini cenderung menghambat diferensiasi kalus. Peristiwa pencoklatan ini adalah peristiwa alamiah, yang merupakan suatu proses perubahan adaptif bagian tanaman akibat adanya pengaruh seperti respon dari bekas perlukaan pada eksplan dan juga merupakan tahapan awal perubahan warna kalus menjadi putih kehijauan. Hal ini terbukti pada kalus yang berwarna coklat juga terdapat kalus remah yang berwarna putih dan massa kalus semakin bertambah dan juga ada yang tumbuh akar. Perubahan warna juga diduga karena adanya sintesis senyawa fenolik akibat adanya cekaman berupa pelukaan pada jaringan. Pernyataan tersebut diperkuat oleh Verpoorte et al. (1993), bahwa terjadinya pencokelatan pada jaringan adalah karena aksi polifenol oksidase yang disintesis akibat dari oksidasi jaringan ketika terluka. Selain itu warna coklat ini berarti terdapat proses degradasi klorofil karena tidak ada penambahan Kinetin dan konsentrasi Kinetin yang rendah, dimana Kinetin disini berperan dalam pembentukan klorofil, sehingga menyebabkan warna hijau tidak muncul. Hal ini sesuai dengan pendapat Santosa dan Nursandi (2002) bahwa kalus yang tidak hijau disebabkan oleh hilangnya polarisasi, sehingga terjadi proses dekomposisi klorofil.
4.2 Respon Organogenesis Kultur In Vitro
Nicotiana tabacum L. var Prancak 95 Proses organogenesis eksplan secara in vitro terjadi dengan dua cara yang berbeda yaitu secara langsung dan tidak langsung. Eksplan menunjukkan respon organogenesis secara tidak langsung apabila eksplan tumbuh melalui kalus, kemudian akan berdiferensiasi menjadi tunas dan akar. Eksplan menunjukkan respon secara organogenesis langsung apabila eksplan tumbuh langsung membentuk tunas dan akar, tanpa melalui
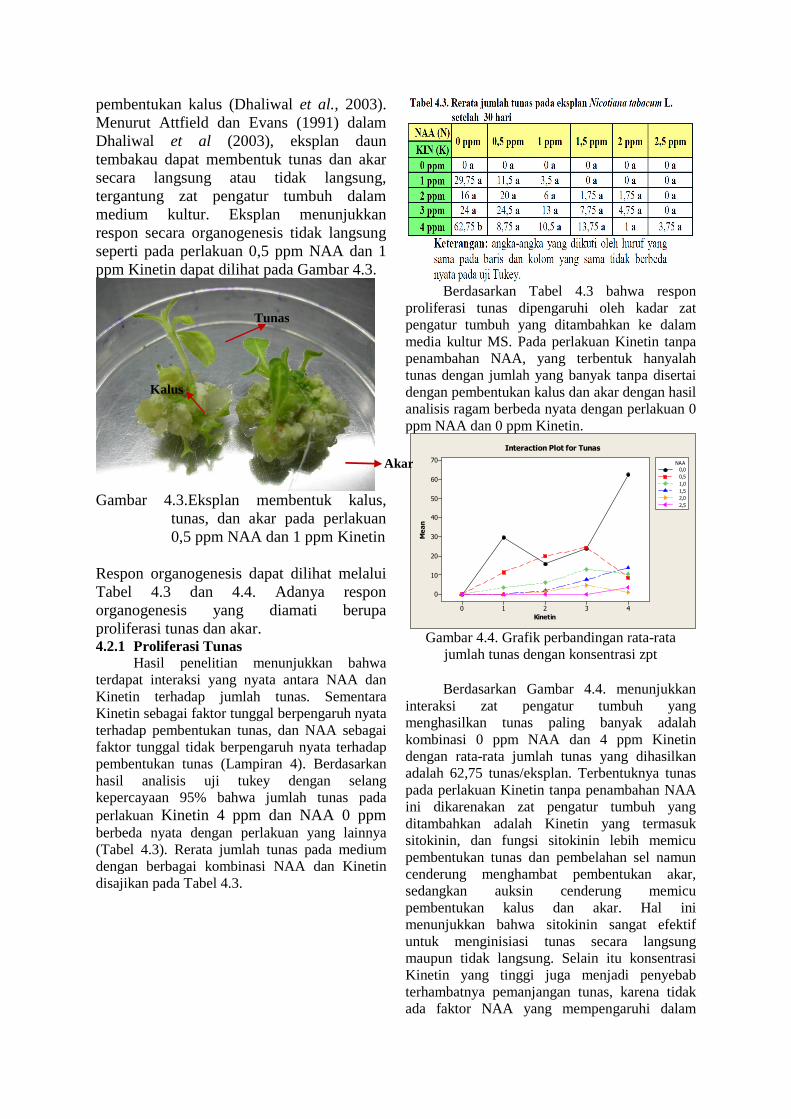
pembentukan kalus (Dhaliwal et al., 2003). Menurut Attfield dan Evans (1991) dalam Dhaliwal et al (2003), eksplan daun tembakau dapat membentuk tunas dan akar secara langsung atau tidak langsung, tergantung zat pengatur tumbuh dalam medium kultur. Eksplan menunjukkan respon secara organogenesis tidak langsung seperti pada perlakuan 0,5 ppm NAA dan 1 ppm Kinetin dapat dilihat pada Gambar 4.3.
Gambar 4.3.Eksplan membentuk kalus,
tunas, dan akar pada perlakuan 0,5 ppm NAA dan 1 ppm Kinetin
Respon organogenesis dapat dilihat melalui Tabel 4.3 dan 4.4. Adanya respon organogenesis yang diamati berupa proliferasi tunas dan akar. 4.2.1 Proliferasi Tunas
Hasil penelitian menunjukkan bahwa terdapat interaksi yang nyata antara NAA dan Kinetin terhadap jumlah tunas. Sementara Kinetin sebagai faktor tunggal berpengaruh nyata terhadap pembentukan tunas, dan NAA sebagai faktor tunggal tidak berpengaruh nyata terhadap pembentukan tunas (Lampiran 4). Berdasarkan hasil analisis uji tukey dengan selang kepercayaan 95% bahwa jumlah tunas pada perlakuan Kinetin 4 ppm dan NAA 0 ppm berbeda nyata dengan perlakuan yang lainnya (Tabel 4.3). Rerata jumlah tunas pada medium dengan berbagai kombinasi NAA dan Kinetin disajikan pada Tabel 4.3.
Berdasarkan Tabel 4.3 bahwa respon
proliferasi tunas dipengaruhi oleh kadar zat pengatur tumbuh yang ditambahkan ke dalam media kultur MS. Pada perlakuan Kinetin tanpa penambahan NAA, yang terbentuk hanyalah tunas dengan jumlah yang banyak tanpa disertai dengan pembentukan kalus dan akar dengan hasil analisis ragam berbeda nyata dengan perlakuan 0 ppm NAA dan 0 ppm Kinetin.
Kinetin
Mean
43210
70
60
50
40
30
20
10
0
NAA
1,0
1,5
2,0
2,5
0,0
0,5
Interaction Plot for Tunas
Gambar 4.4. Grafik perbandingan rata-rata jumlah tunas dengan konsentrasi zpt
Berdasarkan Gambar 4.4. menunjukkan interaksi zat pengatur tumbuh yang menghasilkan tunas paling banyak adalah kombinasi 0 ppm NAA dan 4 ppm Kinetin dengan rata-rata jumlah tunas yang dihasilkan adalah 62,75 tunas/eksplan. Terbentuknya tunas pada perlakuan Kinetin tanpa penambahan NAA ini dikarenakan zat pengatur tumbuh yang ditambahkan adalah Kinetin yang termasuk sitokinin, dan fungsi sitokinin lebih memicu pembentukan tunas dan pembelahan sel namun cenderung menghambat pembentukan akar, sedangkan auksin cenderung memicu pembentukan kalus dan akar. Hal ini menunjukkan bahwa sitokinin sangat efektif untuk menginisiasi tunas secara langsung maupun tidak langsung. Selain itu konsentrasi Kinetin yang tinggi juga menjadi penyebab terhambatnya pemanjangan tunas, karena tidak ada faktor NAA yang mempengaruhi dalam
Kalus
Tunas
Akar

pemanjangan sel. Konsentrasi Kinetin 4 ppm yang ditambahkan disini merupakan konsentrasi optimal untuk memicu pembentukan tunas. Hal ini sesuai dengan penelitian Yunus (2007), bahwa jumlah tunas terbanyak tumbuh pada perlakuan Kinetin tunggal 4 ppm. Tunas yang terbentuk pada perlakuan Kinetin 4 ppm tanpa penambahan NAA dapat dilihat pada Gambar 4.5.
Gambar 4.5. Tunas yang terbentuk pada
perlakuan 0 ppm NAA dan 4 ppm Kinetin
Sedangkan perlakuan dengan penambahan zat pengatur tumbuh NAA tanpa Kinetin tidak menghasilkan tunas, hanya kalus dan akar saja dengan hasil analisis ragam tidak berbeda nyata dengan perlakuan 0 ppm NAA dan 0 ppm Kinetin. Hal ini dikarenakan zat pengatur tumbuh yang ditambahkan adalah NAA. Auksin lebih berperan dalam inisiasi kalus dan akar. Tidak terbentuknya tunas dikarenakan tidak adanya Kinetin yang berperan dalam pembelahan sel serta diferensiasi sel yang lebih ke arah pembentukan tunas. Sehingga dapat diketahui bahwa sitokinin sangat penting dalam mengiduksi tunas eksplan daun tembakau Nicotiana tabacum L.
Adapun interaksi antara zat pengatur tumbuh NAA dan Kinetin dengan jumlah tunas dapat dilihat pada gambar 4.4. Adanya interaksi antara NAA dan Kinetin dapat mempengaruhi terbentuknya tunas dengan baik. Hal ini dibuktikan pada perlakuan interaksi tersebut banyak yang menghasilkan tunas. Hal ini dikarenakan terdapat pengaruh faktor NAA dan Kinetin sebagai zat pengatur tumbuh. Keseimbangan antara NAA dan Kinetin sangat penting dalam menginduksi tunas karena masing-masing zat pengatur tumbuh tersebut mempunyai peranan dalam menginduksi tunas. Skoog dan Miller (1950) dalam Kieber (2002) mengungkapkan bahwa dengan adanya auksin dan sitokinin dalam medium dapat menstimulasi sel-sel jaringan parenkim tembakau untuk
membelah. Pemanjangan sel, pembelahan sel, morfogenesis dan pengaturan pertumbuhan merupakan proses yang sangat penting dalam pembetukan kalus dan selanjutnya diikuti pembentukan tunas. Hal ini menunjukkan bahwa sitokinin (Kinetin) dan auksin (NAA) berperanan saling melengkapi dalam menginduksi tunas.
Tunas sebagian besar terbentuk pada bagian abaksial daun, sedangkan akar tumbuh pada bagian adaksial daun. Dhaliwal et al (2004) mengatakan bahwa pada eksplan daun tembakau primordia tunas tumbuh dari sel-sel mesofil palisade yang terletak di bagian adaksial daun, sedangkan akar tumbuh dari barisan sel parenkim dekat dengan pembuluh vaskuler. Skoog and Miller, (1957) dalam Kieber (2002) mengatakan sitokinin terlibat dalam berbagai aspek pada pertumbuhan dan perkembangan tanaman terutama pembentukan tunas. Proses perkembangan bakal tunas diawali dengan pembelahan sel secara periklinal di sisi lateral (periferal), agak di bawah daerah distal meristem pucuk. Pembelahan sel secara periklinal yang diikuti dengan pertumbuhan sel anak yang menyebabkan tonjolan yang membentuk primordial daun, sedangkan pembelahan sel secara antiklinal dapat meningkatkan luas permukaan primordia daun. Primordial daun ditopang oleh sel prokambium, selanjutnya prokambium tersebut akan menjadi tulang daun (Qosim, 2005).
4.2.2 Proliferasi Akar
Hasil analisis ragam menunjukkan bahwa terdapat interaksi yang nyata antara NAA dan Kinetin terhadap jumlah tunas dan jumlah akar. Sementara faktor NAA sebagai faktor tunggal berpengaruh nyata terhadap pembentukan akar (Lampiran 4). Berdasarkan hasil analisis uji Tukey dengan selang kepercayaan 95 % bahwa jumlah akar pada perlakuan 2,5 ppm NAA dan 0 ppm Kinetin berbeda nyata dengan perlakuan lainnya (Tabel 4.4). Rerata jumlah akar pada medium dengan berbagai kombinasi NAA dan Kinetin disajikan pada Tabel 4.4.
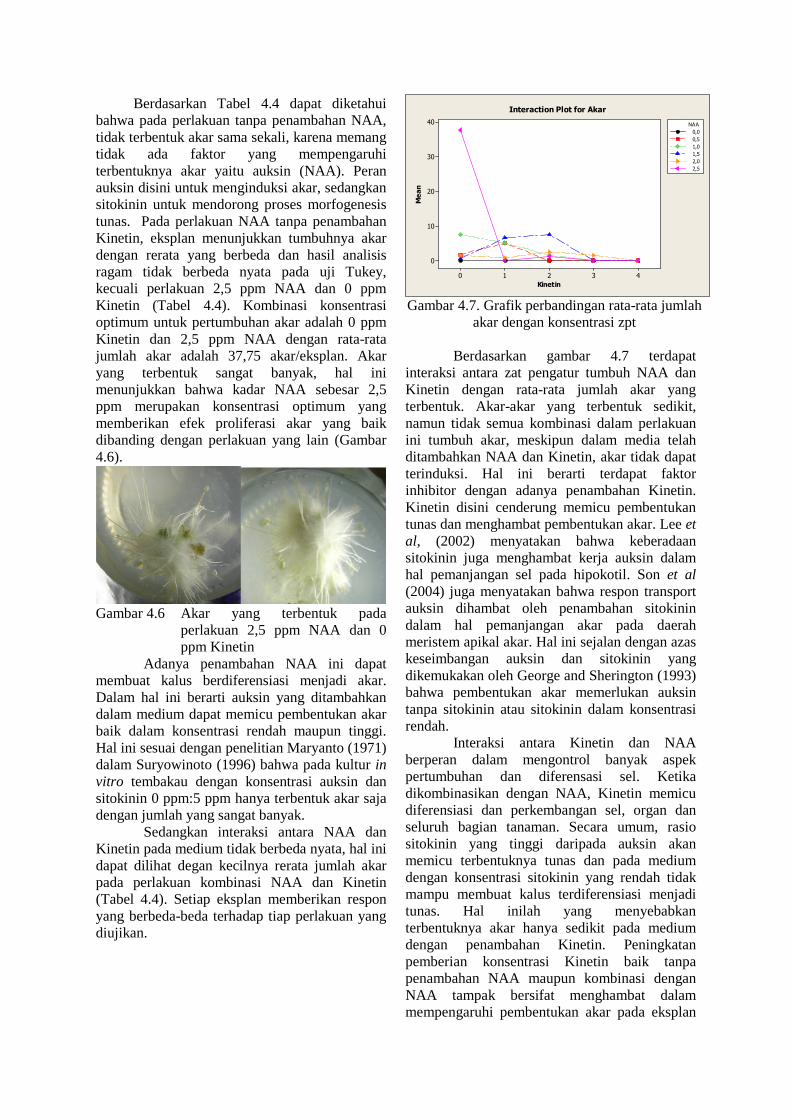
Berdasarkan Tabel 4.4 dapat diketahui bahwa pada perlakuan tanpa penambahan NAA, tidak terbentuk akar sama sekali, karena memang tidak ada faktor yang mempengaruhi terbentuknya akar yaitu auksin (NAA). Peran auksin disini untuk menginduksi akar, sedangkan sitokinin untuk mendorong proses morfogenesis tunas. Pada perlakuan NAA tanpa penambahan Kinetin, eksplan menunjukkan tumbuhnya akar dengan rerata yang berbeda dan hasil analisis ragam tidak berbeda nyata pada uji Tukey, kecuali perlakuan 2,5 ppm NAA dan 0 ppm Kinetin (Tabel 4.4). Kombinasi konsentrasi optimum untuk pertumbuhan akar adalah 0 ppm Kinetin dan 2,5 ppm NAA dengan rata-rata jumlah akar adalah 37,75 akar/eksplan. Akar yang terbentuk sangat banyak, hal ini menunjukkan bahwa kadar NAA sebesar 2,5 ppm merupakan konsentrasi optimum yang memberikan efek proliferasi akar yang baik dibanding dengan perlakuan yang lain (Gambar 4.6).
Gambar 4.6 Akar yang terbentuk pada perlakuan 2,5 ppm NAA dan 0 ppm Kinetin
Adanya penambahan NAA ini dapat membuat kalus berdiferensiasi menjadi akar. Dalam hal ini berarti auksin yang ditambahkan dalam medium dapat memicu pembentukan akar baik dalam konsentrasi rendah maupun tinggi. Hal ini sesuai dengan penelitian Maryanto (1971) dalam Suryowinoto (1996) bahwa pada kultur in vitro tembakau dengan konsentrasi auksin dan sitokinin 0 ppm:5 ppm hanya terbentuk akar saja dengan jumlah yang sangat banyak.
Sedangkan interaksi antara NAA dan Kinetin pada medium tidak berbeda nyata, hal ini dapat dilihat degan kecilnya rerata jumlah akar pada perlakuan kombinasi NAA dan Kinetin (Tabel 4.4). Setiap eksplan memberikan respon yang berbeda-beda terhadap tiap perlakuan yang diujikan.
Kinetin
Mean
43210
40
30
20
10
0
NAA
1,0
1,5
2,0
2,5
0,0
0,5
Interaction Plot for Akar
Gambar 4.7. Grafik perbandingan rata-rata jumlah akar dengan konsentrasi zpt
Berdasarkan gambar 4.7 terdapat
interaksi antara zat pengatur tumbuh NAA dan Kinetin dengan rata-rata jumlah akar yang terbentuk. Akar-akar yang terbentuk sedikit, namun tidak semua kombinasi dalam perlakuan ini tumbuh akar, meskipun dalam media telah ditambahkan NAA dan Kinetin, akar tidak dapat terinduksi. Hal ini berarti terdapat faktor inhibitor dengan adanya penambahan Kinetin. Kinetin disini cenderung memicu pembentukan tunas dan menghambat pembentukan akar. Lee et al, (2002) menyatakan bahwa keberadaan sitokinin juga menghambat kerja auksin dalam hal pemanjangan sel pada hipokotil. Son et al (2004) juga menyatakan bahwa respon transport auksin dihambat oleh penambahan sitokinin dalam hal pemanjangan akar pada daerah meristem apikal akar. Hal ini sejalan dengan azas keseimbangan auksin dan sitokinin yang dikemukakan oleh George and Sherington (1993) bahwa pembentukan akar memerlukan auksin tanpa sitokinin atau sitokinin dalam konsentrasi rendah.
Interaksi antara Kinetin dan NAA berperan dalam mengontrol banyak aspek pertumbuhan dan diferensasi sel. Ketika dikombinasikan dengan NAA, Kinetin memicu diferensiasi dan perkembangan sel, organ dan seluruh bagian tanaman. Secara umum, rasio sitokinin yang tinggi daripada auksin akan memicu terbentuknya tunas dan pada medium dengan konsentrasi sitokinin yang rendah tidak mampu membuat kalus terdiferensiasi menjadi tunas. Hal inilah yang menyebabkan terbentuknya akar hanya sedikit pada medium dengan penambahan Kinetin. Peningkatan pemberian konsentrasi Kinetin baik tanpa penambahan NAA maupun kombinasi dengan NAA tampak bersifat menghambat dalam mempengaruhi pembentukan akar pada eksplan

karena menghasilkan jumlah akar yang lebih sedikit. Menurut Sugiharto (2007) kadar sitokinin yang optimal untuk pertumbuhan tunas dapat menghambat pertumbuhan dan pembentukan akar. Pernyataan ini sesuai dengan hasil penelitian bahwa pada konsentrasi Kinetin 4 ppm yang dikombinasikan dengan berbagai macam konsentrasi NAA, tidak merespon pembentukan akar.
George and Sherington (1993) menyatakan bahwa auksin berpengaruh luas terhadap pertumbuhan, merangsang dan mempercepat pertumbuhan akar, serta meningkatkan kualitas dan kuantitas akar. Semakin cepatnya saat terbentuk akar pada media yang ditambahkan NAA menunjukkan bahwa auksin dapat mengaktifkan enzim-enzim yang berperan dalam pembuatan komponen sel sehingga begitu mulai terjadinya pembelahan sel maka NAA akan merangsang pembentukan sel-sel dengan cepat, sehingga dapat menginisiasi pembentukan akar (Wattimena, 1991).
IV KESIMPULAN Berdasar hasil penelitian dapat diperoleh kesimpulan sebagai berikut : 1. Penambahan zat pengatur tumbuh
Naphthalene Acetic Acid (NAA) dan Kinetin memberikan pengaruh terhadap jumlah tunas, jumlah akar pada kultur in vitro eksplan daun tembakau (Nicotiana tabacum L. var. Prancak 95).
2. Respon Jumlah Tunas terbaik ditunjukkan pada perlakuan dengan penambahan 4 ppm Kinetin 0 ppm NAA, dengan rata-rata sebesar 62,75 tunas/eksplan. Sedangkan respon jumlah akar terbaik ditunjukkan pada perlakuan dengan penambahan 2,5 ppm NAA 0 ppm Kinetin dengan rata-rata sebesar 37,75 tunas/eksplan.
3. Organogenesis pada eksplan terjadi secara langsung dan tidak langsung dan Callogenesis terjadi pada eksplan pada semua perlakuan kecuali perlakuan tanpa penambahan NAA.
DAFTAR PUSTAKA Abdullah, Ahmad dan Soedarmanto. 1982.
Budidaya Tembakau. CV Yasaguna. Jakarta.
Ali, G., F. Hadi, Z. Ali, M. Tariq, and M. A. Khan. 2007. Callus Induction and in vitro Complete Plant Regeneration of Different Cultivars ot Tobacco (Nicotiana tabacum L.) on Media of Different Hormonal
Concentration. Biotechnology. Vol 6(4): 561-566
Basuki, S, Suwarso, A. Herwati, dan S. Yulaikah. 1999. Biologi dan Morfologi Tembakau Madura. Balai Penelitian Tembakau dan Tanaman Serat. Malang
Dhaliwal, H. S., E. C. Yeung, and T. A. Thorpe. 2004. Tiba Inhibition of In vitro Organogenesis in Excised Tobacco Leaf Explants. In vitro cell. Dev. Biol. Plant 40:235-238. Plant Physiology Research Group, Departement of Biological Sciences, University of Calgary, Calgary, Alberta, T2N 1N4, Canada
Fitrianti, A. 2006. Efektivitas Asam 2,4-Diklorofenoksiasetat (2,4-D) dan Kinetin pada Medium MS dalam Induksi Kalus Sambiloto dengan Eksplan Potongan Daun. Skripsi. Biologi FMIPA UNS: Semarang
Fowke, L. C., P. J. Rennie, and F. Constable. 1983. Organelles Associated with the Plasma Membran of Tobacco Leaf Protoplast. Plant Cell Report 2(1983): 292-295
George, E. F., and Sherrington, Ph.D. 1993. Plant Propagation by Tissue Culture. Exegetic Limited. London.
Gunawan, L.W., 1995. Teknik kultur jaringan tumbuhan. Laboratorium Kultur Jaringan Tumbuhan, Pusat Antar Universitas (PAU), Institut Pertanian Bogor (IPB). Bogor.
Hanum, Chairani. 2008. Teknik Budidaya Tanaman Jilid 3. Direktorat Pembinaan Sekolah Menengah Kejuruan, Direktorat Jenderal Manajemen Pendidikan Dasar dan Menengah, Departemen Pendidikan Nasional. Jakarta
Hendaryono, Daisy.P.S dan Ari Wijayani. 1994. Teknik Kultur Jaringan (Pengenalan dan Petunjuk Perbanyakan Tanaman Secara Vegetatif-Modern). Penerbit Kanisius. Yogyakarta
Hidayat. 2007. Induksi Pertumbuhan Eksplan Endosperm Ulin dengan IAA dan Kinetine. Agritrop, 26 (4) : 147 – 152. Fakultas Pertanian Universitas Udayana Denpasar Bali - Indonesia
Imelda Maria, Wulansari Aida, Poerba Yuyu Suryasari. 2008. Regenerasi Tunas dari Kultur Tangkai Daun Iles-iles (Amorphophallus muelleri Blume). Biodiversitas Vol. 9, No. 3, Juli 2008, hal. 173-176

Judd. 2002. Plant Systematics. Sinauer Associates, Inc. Publisher. Sunder Land, Massachusetts U.S.A
Julia, H.L. 1990. Nicotiana tabacum L. Seccion Toxicologia Hospital de Clinicas "José de
San Martin" Cordoba 2351 Capital Federal 1120 Argentina. http://www.IPCS-INCHEM
Kieber, Joseph J., and Ingrid B D’Agostino. 2002. Molecular mechanisms of cytokinin action. Plant Biology 1999, 2:359–364. Department of Biological Sciences. Chicago
Lee, Dong Ju, Sung Soo Kim, Soung Soo Kim. 2002. The Regulation Of Korean Radish Cationic Peroxidase Promoter By a Low Ratio of Cytokinin To Auxin. Plant Science 162 (2002) 345–353.
Leupin, Ruth E., Leupin Marianne, Charles Ehret, Karl H. Erismann, And Witholt Bernard. 2000. Compact Callus Induction And Plant Regeneration Of A Non-Flowering Vetiver From Java. Plant Cell, Tissue And Organ Culture 62: 115–123. Switzerland.
Marlin. 2005. Regenerasi In Vitro Planlet Jahe Bebas Penyakit Layu Bakteri Pada Beberapa Taraf Konsentrasi 6-Benzil Amino Purine (BAP) dan 1 -Napthalene Acetic Acid (NAA). Jurnal-Jurnal Ilmu Pertanian Indonesia, volume 7 no-1: 8-14
Nugroho, A. 2004. Pedoman Pelaksanaan Teknik Kultur Jaringan. Penebar Swadaya. Jakarta.
Pant, B., and Manandhar, S. 2007. In Vitro Propagation Of Carrot (Daucus Carota) L. Scientific World, Vol. 5, No. 5, June. Central Department Of Botany, Tribhuvan University, Kirtipur, Kathmandu, Nepal.
Pauline, D. Kasi & Sumaryono. 2008. Perkembangan Kalus Embriogenik Sagu (Metroxylon sagu Rottb.) pada tiga sistem kultur in vitro. Menara Perkebunan, 76(1), 1-10. Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor.
Purwianingsih, Widi.Dra.M.Si, Kusdianti R, Dra.M.Si, Yuniarti Linda, S.Si. Anatomi Kalus Yang Berasal Dari Eksplan Daun Catharanthus Roseous (L). G. Don (Tapak Dara). Skripsi Yuniarti.
Qosim, Warid Ali, Poerwanto. R, Wattimena. G.A, Dan Witjaksono. 2005. Pembentukan Planlet Manggis Dari Kalus Nodular In Vitro. Zuriat, Vol. 16, No. 2. Bogor
Rachman, A. Machfudz Dan Heri Istiana. 1997. Teknik Budidaya Tembakau Madura. Balai Penelitian Tanaman Tembakau Dan Tanaman Serat. Malang.
Rahardja, P. C. 1995. Kultur Jaringan Teknik Perbanyakan Tanaman Secara Modern. Penebar Swadaya. Jakarta
Salisbury, F. B dan C.W. Ross. 1995. Fisiologi Tumbuhan. Jilid 3. Bandung: ITB.
Santosa, E. K. 2007. Pemanfaatan Daun Tembakau (Nicotiana Tabacum) Sebagai Pewarna Kain Sutera dengan Menggunakan Mordan Jeruk Nipis (Citrus Aurantifolia Swingle) Diterapkan Pada Lenan Rumah Tangga. Skripsi. Jurusan Teknologi Jasa dan Produksi Fakultas Teknik UNS. Semarang
Santoso, U. dan Nursandi, F. 2003. Kultur Jaringan Tanaman. Penerbit Universitas Muhammadiyah Malang. Malang.
Silva, J. A. T. 2005. Simple Multiplication and Effective Genetic Transformation (Four Methods) of in vitro-grown Tobacco by Stem Thin Cell Layers. Plant Science 169: 1046-1058
Smith dan Wood. 1992. Mollecular and Cell Biochemistry. Cell Biology. Chapman & Hall. London
Son, Ora, Hee-Yeon Choa, Kim Mi-Ran. 2004. Induction of a Homeodomain–Leucine Zipper Gene By Auxin Is Inhibited By Cytokinin In Arabidopsis Roots. Biochemical and Biophysical Research Communications 326 (2005) 203–209. Korea
Steenis, Van. 2008. Flora Untuk Sekolah Menengah di Indonesia. PT Pradnya Paramita : Jakarta
Sudarmadji. 2003. Penggunaan Benzil Amino Purine Pada Pertumbuhan Kalus Secara In Vitro. Buletin Teknik Pertanian Vol 8 No. 1 2003.
Sugiharto Bowo, Rahayu Triastuti, Faatih Mukhiissul. 2007. Propagasi Tanaman Nilam (Pogostemon Cablin Benth.) Secara In Vitro Dengan Kombinasi Sitokinin Dan Auksin 2,4 D. MIPA, Vol. 17, No. 1, Januari 2007: 39 – 47. Jurusan Pendidikan Biologi FKIP UNS
Sunarjono, H. 2002. Budidaya Pisang Dengan Bibit Kultur Jaringan. Penebar Swadaya. Jakarta.
Suryowinoto, M. 1996. Pemuliaan Tanaman Secara In Vitro. Penerbit Kanisius.Yogyakarta.

Susilowati, E. Y. 2006. Identifikasi Nikotin dari Daun Tembakau Kering (Nicotiana tabacum) dan Uji Efektivitas Ekstrak Daun Tembakau sebagai Insektisida Penggerek Batang Padi (Scirpophaga innonata). Skripsi. Kimia FMIPA UNS. Semarang
Suwarso. 1991. Pemuliaan Tanaman Tembakau Virginia dan Tembakau asli dalam Prosiding Pemuliaan Tanaman I. PPTI Komda jatim. Malang.
Suwarso, A. Herwati, A. Rachman, dan Slamet. 1999. Pemuliaan Tembakau Madura. Balai Penelitian Tembakau dan Tanaman Serat: Malang
Suwarso, A. Herwati, dan A. S. Murdiyati. 2006. Varietas-varietas Baru Tembakau Madura. Balai Penelitian Tembakau dan Tanaman Serat: Malang
Suwarso, Anik Herwati. 2008. Varietas Unggul Tembakau Prancak 95. www.bpatp.litbang.deptan.go.id
Tohari. 1992. Tembakau dalam fisiologi tanaman budidaya tropik. Gajah Mada University Press p.747-836. Yogyakarta
Verpoorte R, Van der Heijden R, Schripsema J. 1993. Plant biotechnology for the production of alkaloids; present status and prospect. J Nat Prod 56:186-207.
Wang, H., M. Zhao, B. Yang, Y. Jiang, and G. Rao. 2008. Identification of Polyphenols in Tobacco Leaf and Their Antioxidant and Antimicrobial Activities. Food Chemistry 107: 1399–1406
Wattimena, G. A. 1992. Diktat Zat Pengatur Tumbuh Tanaman. Laboratorium Kultur Jaringan Tanaman PAU Bioteknologi IPB-Direktorat Jenderal Pendidikan Tinggi Departemen Pendidikan dan Kebudayaan. Bogor
Widiastoety, D. dan A.Santi. 1994. Pengaruh Air Kelapa terhadap Pembentukan Proticorm Like Bodies (PLBs) dari Anggrek Vanda dalam Medium Cair. Jurnal Hortikultura Volume 4 No. 2.
Yunus, Ahmad. 2007. Pengaruh IAA dan Kinetin terhadap Pertumbuhan Eksplan Bawang Merah (Allium ascalonicum L.) secara In Vitro. Jurnal Akta Agrosia Edisi Khusus No. 1: 53-58
Yusnita. 2003. Kultur Jaringan : Cara Memperbanyak Tanaman Secara Efisien. Jakarta. Agromedia Pustaka.
Zia, Muhammad, Riaz-ur-Rehman, and Chaudhary Muhammad Fayyaz. 2007.
Hormonal regulation for callogenesis and organgenesis of Artemisia absinthium L. African Journal of Biotechnology Vol. 6 (16), pp. 1874-1878. Plant Physiology Laboratory, Faculty of Biological Sciences, Quaid-i-Azam University, Islamabad, Pakistan.
Zulfiqar, Bushra, Akhtar Abbasi Nadeem, Ahmad Touqeer, and Ishfaq Ahmed Hafiz. 2009. Effect of explant sources and different Concentrations of plant growth regulators on in vitro shoot proliferation and rooting of avocado (persea americana mill.). Pak. J. Bot., 41(5): 2333-2346. Department of Horticulture, Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi, Pakistan.
Zulkarnain. 2009. Kultur Jaringan Tanaman. Bumi Aksara. Jakarta





























