Embed Size (px)
Citation preview

Chapitre 1
Perception du mouvement etmouvement propre
Nous avons choisi de consacrer ce premier chapitre a la perception du mouvement et au mou-vement propre pour deux raisons principales : d’une part parce que, comme nous l’avons apercuauparavant, la representation de l’espace dans le cerveau semble intimement liee a la perception dumouvement, et d’autre part parce que les travaux realises pour notre these portent sur des signauxde mouvement, les signaux neuronaux relatifs au mouvement de la tete.
La perception du mouvement propre, c’est-a-dire la perception du deplacement de l’observateurdans l’environnement (Warren 1990), repose sur un ensemble de signaux distincts, parmi lesquels lessignaux vestibulaires, les signaux de flux sensoriels optiques ou tactiles, les signaux proprioceptifsou kinesthesiques, ou encore les signaux de copie motrice efferente ou de decharges collaterales.Afin de cerner les bases de la perception du mouvement et du mouvement propre, nous debuteronspar la description du capteur specialise pour le mouvement, le capteur vestibulaire. Ensuite, nousconsacrerons deux parties a l’etude du flux optique et aux informations sur le mouvement que lecerveau peut en extraire. Puis nous verrons comment la perception du mouvement propre emergede la fusion de plusieurs informations sensorielles. Nous finirons par une description sommaire descaracteristiques generales du mouvement propre, et du couplage entre perception du mouvementet action.
1.1 Le systeme vestibulaire
- Le sens vestibulaire donne des informations sur le mouvement physique de la tete. Bien qu’actifen permanence, il possede la particularite de n’arriver a la conscience que lorsqu’il pose probleme.L’on prend generalement conscience de son existence en cas de mal des transports, d’intoxica-tion alcoolique aigue ou de vertiges (vertige de Meniere, vertige alternobarique declenche par undifferentiel de pression entre les deux oreilles moyennes, etc.).
Du point de vue des indices du mouvement propre, les indices vestibulaires (et les indicessomatosensoriels dependants de l’inertie) ont deux particularites par rapport aux autres indicessensoriels (Mergner & Becker 1990). D’abord, ils ne peuvent pas etre interrompus comme l’est parexemple le signal visuel lorsqu’on ferme les yeux. Ensuite, puisqu’ils mesurent l’acceleration dansl’espace physique, un message non nul signifie automatiquement un deplacement dans l’espace. Al’oppose, les autres signaux sensoriels comme la vision ou l’audition peuvent etre interrompus avolonte, et l’information qu’ils fournissent est ambigue, puisqu’elle est relative. Ainsi, en se basantsur la seule vision, le sujet ne peut savoir si c’est lui ou l’environnement qui bouge.
15

16 Perception du mouvement et mouvement propre
- La difficulte d’apprehender le sens vestibulaire est par ailleurs assez bien illustree par sonhistoire (Guerrier & Mounier-Kuhn 1980). Le labyrinthe est decouvert par d’Empedocle (535-475av J.C.), mais ce dernier le considere comme un organe de l’audition, affirmant que le son estcapte par cet organe puis ensuite regularise le long des canaux. On doit ensuite attendre GabrielFallope, au XVIe siecle, qui donne la premiere description anatomique complete de l’oreille interne,en distinguant ses deux cavites nommees labyrinthe et limacon. Ce n’est que bien plus tard encore,au XIXe siecle, qu’une premiere hypothese sur la fonction vestibulaire est avancee par Pierre-Jean-Marie Flourens, qui considere que le labyrinthe est l’organe peripherique dans lequel resideraientles forces moderatrices des mouvements. Le labyrinthe vestibulaire ne fait son entree en cliniqueque vingt ans plus tard, en 1861, avec le fameux rapport de Prosper Meniere qui decrit le premiersyndrome vertigineux en rapport avec l’oreille interne (maladie de Meniere). Enfin, c’est le physicienallemand Ernst Mach, qui en 1875, est le premier a decrire la geometrie des canaux semi-circulaires,notamment leur orientation respective ainsi que leur fonctionnement par couple (voir Henn 1997).
1.1.1 Le labyrinthe : un capteur de mouvement tridimensionnel
1.1.1.1 Anatomie et structure du capteur vestibulaire
Le capteur vestibulaire, ou labyrinthe membraneux, est situe dans l’oreille interne (Fig. 1.1A). Ilest compose de deux types de capteurs, les canaux semi-circulaires qui sont des capteurs de rotationde la tete, et les capteurs otolithiques ou maculaires qui percoivent les accelerations lineaires,soit sous forme de translation de la tete ou d’orientation de celle-ci par rapport a la gravite. Lescanaux semi-circulaires sont au nombre de trois, un canal anterieur, un canal posterieur et un canalhorizontal (Fig. 1.1A : bleu cyan, vert et rouge, respectivement). Ces canaux sont composes d’unepart du canal proprement-dit (hemi-tore) et d’un renflement encore appele ampoule, qui contientles cellules sensorielles. Les capteurs otolithiques sont au nombre de deux, l’utricule qui se trouvejuste a la base des canaux semi-circulaires, et le saccule qui se trouve en dessous de ce dernier (Fig.1.1A : en orange).
Les canaux semi-circulaires sont fixes par rapport a la tete, et sont orientes dans trois plans del’espace physique. Le canal horizontal est incline (d’environ 25◦ chez l’homme, voir Wilson & Melvill-Jones 1979, pp.15-19 ; environ 15◦ chez le singe, Reisine et al. 1988) par rapport au plan horizontalanatomique de la tete (plan de Horsley Clark ; Fig. 1.1B), ce qui entraıne qu’en conditions naturelles,les canaux horizontaux sont alignes avec l’horizontale definie par rapport a la terre. Les canauxverticaux anterieur et posterieur sont orientes diagonalement par rapport a la tete, suivant deuxlignes passant par deux reperes anatomiques (Fig. 1.1C). Plus generalement, l’organisation spatialedes canaux semi-circulaires suit trois grands principes : (1) une symetrie bilaterale (Fig. 1.1C), (2)une orthogonalite deux a deux des plans canalaires, et (3) un fonctionnement par couple excitateur-inhibiteur (push-pull ; Graf 1988). Les deux premieres proprietes ne sont toutefois verifiees quepartiellement, en particulier en raison de variabilite anatomique qui entraıne des deviations del’orthogonalite ainsi que de l’alignement des deux canaux d’une paire d’environ 10◦ (Wilson &Melvill-Jones 1979, pp.15-19 ; Reisine et al. 1988). De telles deviations pourraient toutefois etrecompensees par le fonctionnement en couple (Wilson & Melvill-Jones 1979).
Les capteurs otolithiques sont aussi orientes dans l’espace. Le capteur utriculaire est approxima-tivement aligne avec le plan des canaux semi-circulaires horizontaux, et le capteur sacculaire est dansun plan vertical perpendiculaire a celui-ci (Wilson & Melvill-Jones, pp. 20-22). Cette dispositiondes otolithes montre leur role complementaire, constituant ensemble un capteur gravito-inertiel.

Le systeme vestibulaire 17
Fig. 1.1 –

18 Perception du mouvement et mouvement propre
FIG. 1.1 – Localisation cranienne du systeme vestibulaire. A Coupe frontale schematique montrant lalocalisation du systeme vestibulaire dans l’oreille interne. Celui-ci est compose d’une part des canaux semi-circulaires, dont l’horizontal (en rouge), l’anterieur (en bleu cyan) et le posterieur (en vert), et d’autrepart des otolithes (saccule et utricule ; en orange). Les canaux semi-circulaires sont composes du canalproprement dit et d’un renflement situe au dessus de l’utricule, l’ampoule. B Vue laterale de la tete, etposition de l’appareil vestibulaire. Remarquons que le canal semi-circulaire horizontal (en rouge) n’est pasdans le plan horizontal du crane, mais est devie de celui-ci d’un angle d’environ 25◦. C Vue du dessus :les canaux semi-circulaires anterieurs (en bleu) et posterieurs (en vert) sont orientes a environ 45◦ du plansagittal, et approximativement perpendiculaires entre eux. L’orientation des canaux anterieurs est definiepar la ligne passant par le centre des structures vestibulo-cochleaires et le bord posterieur du foramenmagnum, et l’orientation des canaux posterieurs est definie par la ligne passant par le centre des structuresvestibulo-cochleaires et la selle turcique.
Plus generalement, l’ensemble des capteurs vestibulaires dote le cerveau d’informations sur lemouvement de la tete dans l’espace physique tridimensionnel. L’importance du capteur vestibulaireet de sa structure peuvent etre percues a travers sa phylogenie. Les premiers organismes fossilisespossedant un labyrinthe sont les Ostracodermes (familles des Agnathes, 350-400 millions d’annees).Ce labyrinthe primitif ne comprend que des canaux verticaux. Les descendants modernes des Gna-thostomes (organismes pourvus de machoire), les poissons cartilagineux et osseux (Chondrichtienset Osteichtiens) acquierent un nouvel element : les canaux horizontaux. Si le labyrinthe des pois-sons osseux est identique a celui des Mammiferes, celui des poissons cartilagineux ne possede pas departie commune entre les canaux anterieur et posterieur, laissant penser au developpement de deuxsolutions evolutives distinctes. Toutefois, remarquons que le capteur vestibulaire apparaıt relati-vement tot dans l’evolution des vertebres, et que sa structure tri-dimensionnelle, une fois acquise,reste inchangee. Le labyrinthe est donc une solution evolutive stable pour percevoir le mouvementdans l’espace tridimensionnel (Graf 1988).
1.1.1.2 Les canaux semi-circulaires sont des capteurs de rotation
Depuis la description de Mach en 1875, l’on sait que les canaux semi-circulaires sont adaptespour percevoir les rotations de la tete en trois dimensions. Chaque canal a ainsi un axe de ro-tation cephalique prefere, c’est-a-dire qu’une rotation autour de cet axe provoque une reponseneuronale maximale (Fig. 1.2A). L’axe prefere n’est, en general, pas aligne avec l’axe anatomiquedu canal, mais differe de celui-ci d’environ 10◦ en moyenne chez le primate (Reisine et al. 1988).Par ailleurs, les six canaux semi-circulaires fonctionnent en couple, definissant 3 axes de rota-tion majeurs. Les deux canaux horizontaux definissent l’axe vertical, les canaux anterieur-droitet posterieur-gauche definissent l’axe horizontal diagonal anterieur-droit posterieur-gauche, et lescanaux anterieur-gauche et posterieur-droit definissent le deuxieme axe horizontal diagonal.
Le fonctionnement des canaux semi-circulaires est base sur un mecanisme inertiel. Une trans-lation de la tete, donc de l’ensemble des canaux semi-circulaires, ne provoque aucun effet sur cesderniers. Par contre, dans le cas par exemple d’une rotation horizontale de la tete vers la droite, lecanal horizontal droit est active, le canal horizontal gauche inhibe (Fig. 1.2A). Le mouvement derotation de la tete provoque, par inertie du liquide endolymphatique a l’interieur du canal droit, unmouvement fluide vers la gauche, qui devie la cupule de l’ampoule du canal droit vers la gauche (Fig.1.2B). Cette deviation entraıne un mouvement des cils –des cellules ciliees de la base ampoulaire–qui sont solidaires de la cupule. L’inclinaison des cils vers le kinocil provoque a son tour l’ouverturede canaux ioniques transmembranaires, aboutissant a l’excitation des cellules ciliees qui sont encontact avec les cellules bipolaires du ganglion de Scarpa.

Le systeme vestibulaire 19
Fig. 1.2 – Transduction du message vestibulaire. A Representation schematique des canaux semi-circulaires.Les directions preferees de rotation de la tete sont representees pour les canaux horizontaux (en rouge),anterieurs (en bleu) et posterieurs (en vert). Les canaux semi-circulaires fonctionnent par paire. Ainsi, lorsd’une rotation horizontale de la tete vers la droite, le canal droit est active et le gauche inhibe. B Agran-dissement de l’ampoule du canal horizontal droit. Lors de la rotation de la tete vers la droite, le liquideendolymphatique, par inertie, deplace la cupule vers la gauche. Les cellules ciliees de la crete ampoulairesont activees par la deviation consequente de leurs cils apicaux (kinocils) qui entraıne l’ouverture de canauxioniques transmembranaires. C Generation du message vestibulaire. En l’absence de mouvement (phase 1),les cellules bipolaires reliees aux cellules ciliees dechargent toniquement. Lors d’une augmentation de la vi-tesse de rotation de la tete (acceleration positive, phase 2), les cils sont devies vers le kinocil (grand cil noir),la cellule ciliee activee (EPSP), la frequence de decharge de la cellule bipolaire augmente rapidement jusqu’auniveau de saturation qui est proportionnel a l’amplitude de l’acceleration. A la fin de l’echelon d’acceleration,la tete tourne a vitesse constante, la frequence de decharge de la cellule diminue jusqu’au niveau de dechargetonique de repos (phase 3). Lors de la deceleration de la tete (phase 4), les cils sont devies a l’oppose de ladirection preferee, la cellule ciliee inhibee (IPSP), la frequence de decharge de la cellule bipolaire descend endessous de la frequence de repos, pour revenir progressivement vers celle-ci apres la fin de la deceleration.Ainsi, le systeme vestibulaire ne distingue pas l’immobilite (phase 1) de la rotation a vitesseconstante (phase 3).

20 Perception du mouvement et mouvement propre
L’excitation des cellules ciliees aboutit a la generation d’un potentiel post-synaptique excita-teur (EPSP), puis a la generation de potentiels d’action nerveux par les cellules bipolaires (Fig.1.2C). A l’inverse, dans le canal horizontal gauche, les cils sont devies dans l’autre sens (a opposedu kinocil), les canaux ioniques ouverts provoquent une hyperpolarisation des cellules ciliees etune diminution de la decharge des cellules bipolaires : les deux canaux fonctionnement en coupleexcitation-inhibition.
De par sa nature inertielle, le capteur canalaire n’est sensible qu’a l’acceleration de la tete. Unmouvement a vitesse constante n’a pas d’effet inertiel. Ainsi, le capteur canalaire ne peut distinguerentre l’absence de mouvement et un mouvement a vitesse constante (Fig. 1.2C).
1.1.1.3 Les otholithes sont des capteurs d’acceleration lineaire
Les capteurs otolithiques sont aussi des capteurs inertiels, et leur fonctionnement intime presentede grandes similarites avec celui des canaux semi-circulaires. Ils se composent egalement de cellulesciliees incluses dans une lame basale maintenue par des cellules de soutien (Fig. 1.3A). Les cils descellules ciliees se trouvent, quant a eux, solidaires d’amas de cristaux particuliers appeles otoconies,et qui ne sont retenus que par les cils.
Fig. 1.3 – Transduction du message otholithique. A Les otolithes se composent d’une rangee de cellulessensitives ciliees soutenues par une lame basale qui contient les terminaisons afferentes du nerf sacculaire. Lescils de la partie apicale des cellules sensitives sont solidaires de cristaux appeles otoconies. Au repos, la teteverticale, aucune force n’est exercee sur les otoconies. Les cils ne sont pas devies, aucun message sensorieln’est produit. B Lorsque la tete est inclinee, la gravite exerce une force sur les otoconies, ce qui entraıne unedeviation des cils des cellules sensitives et la generation du message nerveux. Une acceleration lineaire de latete produit une force analogue sur les otoconies, et genere un message nerveux par le meme mecanisme. Lesotolithes mesurent ainsi les accelerations lineaires et l’orientation de la tete par rapport a la gravite.
Lorsque la tete est inclinee par rapport a la gravite (Fig. 1.3B), les otoconies ne sont plusorientes perpendiculairement a la gravite, et subissent une force gravitationnelle qui entraıne leur

Le systeme vestibulaire 21
deplacement. Ce deplacement provoque l’inclinaisons des cils des cellules ciliees et la generation dumessage nerveux. Ce type de reponse des otolithes a l’orientation de la tete constitue la fonctionotolithique statique.
De meme, lorsque la tete subit un mouvement de translation avec une composante d’accelera-tion, la resultante de la somme de l’acceleration cephalique et de la gravite n’est plus perpendiculaireaux otoconies. Ceux-ci subissent alors une force gravito-inertielle qui provoque l’inclinaison des cilset la generation du message nerveux. C’est la fonction otolithique dynamique (voir Benson 1990,p. 153). En resume, les capteurs otolithiques percoivent les acceleration lineaires, qu’elles soientprovoquees par des mouvements de translations de la tete, ou par une inclinaison de celle-ci parrapport a la gravite.
La nature inertielle des capteurs otolithiques contraint ceux-ci aux memes limitations que lescanaux semi-circulaires, a savoir l’impossibilite de distinguer l’immobilite d’un mouvement a vitesseconstante.
1.1.1.4 Aspects geometriques
Si l’organisation spatiale des canaux semi-circulaire respecte la condition d’orthogonalite mu-tuelle, on pourrait se demander pourquoi la solution diagonale pour les canaux verticaux a eteretenue au lieu, comme chez la pieuvre par exemple, d’avoir les canaux orientes dans les planscorporels principaux (i.e. horizontal, frontal et sagittal ; Graf 1988). Sans pour autant pretendreapporter une reponse a cette question, remarquons que les plans des canaux semi-circulaires sontremarquablement bien alignes avec ceux de certains de leurs effecteurs principaux, a savoir lesmuscles extra-oculaires (Fig. 1.4 ; voir aussi Wilson & Melvill-Jones 1979, p.255). De plus, il sem-blerait que le codage du monde visuel dans les neurones vestibulaires secondaires, ainsi que dansle systeme optique accessoire soit exprime selon les memes coordonnees (Graf 1988, Simpson et al.1988 : etudes chez le lapin). Ainsi, si la raison de l’orientation particuliere retenue par l’evolutionreste inexpliquee, l’alignement des axes des capteurs, de l’effecteur ainsi que de la representationvisuelle presente l’avantage certain de reduire les problemes de changement de coordonnees, placantde fait le systeme vestibulaire au centre de la representation de l’espace tridimensionnel.
1.1.2 Les signaux du capteur vestibulaire
1.1.2.1 Principales voies anatomiques vestibulaires
Bien qu’une description exhaustive des voies vestibulaires soit en dehors du champ de cetteetude, il paraıt neanmoins necessaire de parcourir rapidement les principales, en particulier celle quimene les signaux du capteur vestibulaire au cortex (Fig. 1.5). A partir des canaux semi-circulaires etdes otolithes, le signal vestibulaire passant par le ganglion de Scarpa arrive aux noyaux vestibulaires.Au sein de ceux-ci, les afferences vestibulaires sont distribuees dans des regions independantes pourune part, et se recouvrant d’autre part, permettant ainsi aux neurones vestibulaires secondaires(i.e. dont le corps cellulaire se trouve dans les noyaux vestibulaires) de transmettre de l’informationprovenant d’un ou plusieurs capteurs a la fois (pour une revue, voir Wilson & Melvill-Jones 1979,chap. 5 ; Fig. 1.5 a droite). Des noyaux vestibulaires, le signal est envoye au thalamus, dans le noyauxventro-posterieur (Buttner & Henn 1976) et dans la region peri-genouillee (Magnin & Fuchs 1977).Du thalamus, le message vestibulaire arrive au cortex, se repartissant directement dans differentesaires dites vestibulaires (Guldin & Grusser 1998 ; cf. chap 1.1.3).

22 Perception du mouvement et mouvement propre
Fig. 1.4 – Orientation 3D des plans des canaux semi-circulaires et des plans d’action des muscles extrao-culaires chez un animal aux yeux frontaux. Il existe une correspondance du plan du canal horizontal et decelui des recti horizontaux (en rouge), du plan du canal anterieur et de celui des recti verticaux (en bleu),et enfin du plan du canal posterieur et des muscles obliques (en vert). Remarquons neanmoins que cettecorrespondance n’est pas absolue : les paires de plans de meme couleur ne sont pas tout a fait paralleles.(d’apres Ezure & Graf 1984)
Par ailleurs, le premier relais central (noyaux vestibulaires) est etroitement lie, par projectionreciproque, au cervelet. Ce dernier aurait un role tres important dans l’elaboration et le controlede l’activite reflexe liee au systeme vestibulaire.
Retenons simplement pour la suite qu’il existe deux differences majeures avec la voie visuelleretino-corticale. La premiere est la presence des noyaux vestibulaires, relais dans le tronc cerebralentre le capteur vestibulaire et le thalamus, alors que l’information visuelle de la retine arrivedirectement au thalamus (noyau genouille lateral). La deuxieme est l’absence de cortex vestibulaireprimaire, qui, a l’instar du cortex visuel primaire, serait l’aire corticale a partir de laquelle seraittransmise l’information vers les autres aires du cortex. L’information vestibulaire thalamique estdistribuee directement dans les aires corticales vestibulaires.

Le systeme vestibulaire 23
Fig. 1.5 – Principales voies vestibulaires. Les afferences vestibulaires primaires provenant des canaux semi-circulaires (a,p,h) arrivent au premier relais central, en passant par le ganglion de Scarpa : les noyauxvestibulaires (VNs) (details a droite, points de couleur). Le canal horizontal projette sur les noyaux vesti-bulaires median (nVM) et lateral (nVL). Les canaux anterieur et posterieur, tous deux impliques dans laperception des rotations verticales et torsionnelles, projettent dans des regions voisines des noyaux vestibu-laires median (nVM) et anterieur (nVA). L’utricule projette sur le noyau median (nVM) et inferieur (nVI),et le saccule sur le noyaux inferieur (nVI). Des noyaux vestibulaires, les signaux vestibulaires rejoignentle faisceau longitudinal median du tronc cerebral, d’ou partent differentes afferences, notamment vers lamoelle (reflexe vestibulo-colique et coordination oculo-cephalique), vers les noyaux oculomoteurs (reflexe-vestibulo-oculaire), vers l’olive inferieure et le cervelet (vestibulocervelet : nodule, uvule, flocculus ; cortexcerebelleux ; coordination oculo-cephalique et modulation des reflexes vestibulaires), et enfin, vers le thala-mus (noyau ventro-posterieur et region peri-genouillee) d’ou partent les afferences vers les differentes regionsvestibulaires du cortex.

24 Perception du mouvement et mouvement propre
1.1.2.2 Le message vestibulaire
Differentes methodes de test des fonctions vestibulaires se sont developpees, permettant d’etudierseparement les differents capteurs. Ainsi, la chaise tournante de Barany permet de stimuler uneseule paire de canaux semi-circulaires, les stimulations caloriques permettent de stimuler un seullabyrinthe, et enfin, des changements de position de la tete testent la reponse des otolithes.
La description detaillee des capteurs vestibulaires et de leur mecanisme inertiel a rapidementdonne lieu a un modele physique de la dynamique canal-cupule (Wilson & Melvill-Jones 1979, pp.41-76). Ce modele montre que le deplacement inertiel du liquide endolymphatique est proportionnela la (variation de) vitesse de la tete, avec en outre un effet de ”remise a zero” progressive dudeplacement cupulaire par l’elasticite de celle-ci. Ainsi, le modele canalo-cupulaire prevoit que dansune bande de frequences de stimulation moyenne, le capteur se comporte comme un transducteurde vitesse angulaire, deviant ensuite vers un ”accelerometre angulaire fuyant” aux basses frequences(Jones & Milsum 1965 ; Wilson & Melvill-Jones 1979).
–Afferences primaires
Canaux semi-circulairesLa premiere etude electrophysiologique des afferences vestibulaires primaires (Ross 1936, chez
la Grenouille) montre en effet que, dans une bande de frequence limitee, le systeme canalo-cupulaireintegre l’acceleration et produit un signal de vitesse angulaire de la tete. Chez le singe, la premiereetude detaillee decrit deux caracteristiques principales de ces afferences (Fernandez & Goldberg1971). La frequence de decharge spontanee est en moyenne tres elevee (91.3 ± 1.7 sp/s, allant de10 a 170 sp/s), ce qui permet un codage bi-directionnel (excitation-inhibition, cf. Fig. 1.2C). Ladeuxieme caracteristique concerne la regularite de la frequence de decharge de repos. Sont decritesdes unites regulieres, de une frequence elevee, qui correspondent probablement a des fibres de petitdiametre et qui sont en contact avec de nombreuses cellules ciliees et realisent ainsi une sorte demoyennage afferent. D’autre part, les unites irregulieres, dont la frequence de repos est plus faibleque les precedentes, qui correspondent a des fibres de gros diametre et qui ne sont en contactqu’avec un petit nombre de cellules ciliees (voir aussi Wilson & Melvill-Jones 1979, p.95).
La reponse des afferences canalaires primaires en fonction de la frequence de stimulation aete etudiee pour la premiere fois chez le primate par Fernandez et Goldberg (1971 ; Fig. 1.6A),pour des unites regulieres et irregulieres (symboles ouverts et symboles pleins, respectivement), al’aide d’une stimulation sinusoıdale. La courbe continue represente la reponse theorique d’apresle modele biophysique. Dans la bande de frequences moyennes (i.e. entre 0.1 et 1 Hz), le gain (Fig.1.6A, a gauche) est approximativement constant, la phase par rapport a la vitesse de la stimulation(Fig. 1.6A, a droite) est proche de zero : les neurones repondent a la vitesse angulaire de rotationde la tete. A basse frequence (< 0.1 Hz), les neurones se comportent comme des accelerometresfuyants (la phase tend vers 90◦ et le gain tend vers 0). On note toutefois deux deviations principalesdes reponses du modele biophysique : (1) les unites irregulieres sont en avance de phase meme dansles gammes de frequences moyennes et basses (phenomene d’adaptation des neurones), et (2) legain et la phase augmentent pour les hautes frequences (> 1 Hz), comme si les neurones etaientsensibles a la fois a la vitesse et a la position de la cupule.
OtolithesChez le primate, les neurones primaires sacculaires repondent preferentiellement a l’acceleration
lineaire verticale, et les neurones utriculaires a l’acceleration horizontale (Fernandez & Goldberg1976). La fonction otolithique statique est etudiee a l’aide de rotations lentes (10◦/s) qui permettentd’evaluer la reponse otolithique a l’orientation de la tete (Fig. 1.6B). Au cours d’un cycle de

Le systeme vestibulaire 25
rotation dans le plan sagittal, la frequence de decharge des neurones sacculaires varie continumentavec l’orientation de la tete par rapport a la gravite : ces neurones transmettent bien un signald’orientation (statique) de la tete.
La fonction otolithique dynamique (i.e. reponse a des accelerations de translation ou centri-fuges) est etudiee de maniere analogue a celle des canaux semi-circulaire, a l’aide de stimulationssinusoıdales. Pour les unites regulieres (Fig. 1.6C), le gain reste approximativement constant entre0 et 2 Hz, et la phase proche de 0 (± 10◦) entre 0 et 1 Hz : dans une large bande de frequences, lesneurones otolithiques (unites regulieres) se comportent comme de tres bons accelerometres lineaires(Fernandez & Goldberg 1976 ; voir aussi Wilson & Melvill-Jones 1979, p.118). Comme dans le casdes afferences canalaires, les reponses des unites otolithiques irregulieres sont beaucoup plus va-riables que celles des unites regulieres, et sont par consequent moins ”fiables” par rapport au signalphysique code (Wilson & Melvill-Jones 1979, p.121).
–Afferences secondaires (noyaux vestibulaires)
En provenance des canaux semi-circulairesL’activite dans les noyaux vestibulaires est relativement differente de celle de leurs entrees ves-
tibulaires (afferences primaires). Au contraire des neurones primaires qui repondent uniquement ala rotation angulaire ipsilaterale, divers types de reponses par rapport a la direction de mouvementsont decrits dans les noyaux vestibulaires (Duensing & Schaefer 1958). Le type I correspond auneurones qui repondent au mouvement ipsilateral. Le type II englobe ceux qui repondent au mou-vement controlateral, le type III ceux qui sont actives par les mouvements ipsi- et controlateral, etle type IV ceux qui sont inhibes par les mouvements des deux cotes. Les types I et II sont largementmajoritaires, avec une legere dominance du type I qui correspond aux reponses des afferences pri-maires. Par ailleurs, la frequence de decharge de repos est inferieure a celle des neurones primaires,et de ce fait certains neurones secondaires sont silencieux pendant une partie du cycle de stimula-tion sinusoıdale. Le gain (donc la sensibilite) des neurones secondaires est generalement plus eleveque celui des neurones primaires, les neurones vestibulaires secondaires transmettant toujours unsignal de vitesse angulaire de la tete (Wilson & Melvill-Jones 1979, p.152). Notons aussi que lesconstantes de temps (i.e. vitesse de diminution de la decharge neuronale, cf. Fig. 1.2C) des neuronesvestibulaires secondaires sont plus longues que celles des neurones afferents, atteignant la duree desconstantes de temps du nystagmus vestibulaire (mouvements oculaires reflexes, voir plus loin), ceteffet disparaissant sous anesthesie (Buettner et al. 1978).
En provenance des otolithesComme pour les neurones secondaires canalaires, on retrouve les differents types de reponses
par rapport a la direction des stimulations (les types α, β, γ, δ correspondent aux types I,II,III etIV respectivement ; Wilson & Melvill-Jones 1979, p. 161-2). Par ailleurs, apres cerebelectomie, onobserve une augmentation du rapport du nombre de reponses statiques sur le nombre de reponsesdynamiques trouvees, ce qui suggere un role du cervelet dans la distinction des reponses statique etdynamique, autrement dit, dans la distinction de l’acceleration gravitaire de l’acceleration lineairedue au mouvement (Wilson & Melvill-Jones 1979, p.166).
Codage des mouvements de la teteLes mouvements naturels de la tete se composent en general a la fois de translations et de
rotations : les canaux semi-circulaires et les otolithes sont donc stimules conjointement. S’appuyantsur la variation de phase et de gain des neurones vestibulaires canalaires et otolithiques en fonctionde la direction (3D) du mouvement de la tete, les travaux originaux de Holly et al. (1999) montrentqu’en theorie, ces variations permettraient un codage precis du vecteur de mouvement cephalique,au niveau des noyaux vestibulaires, codage base sur les differences de phase entre les neurones achaque mouvement.
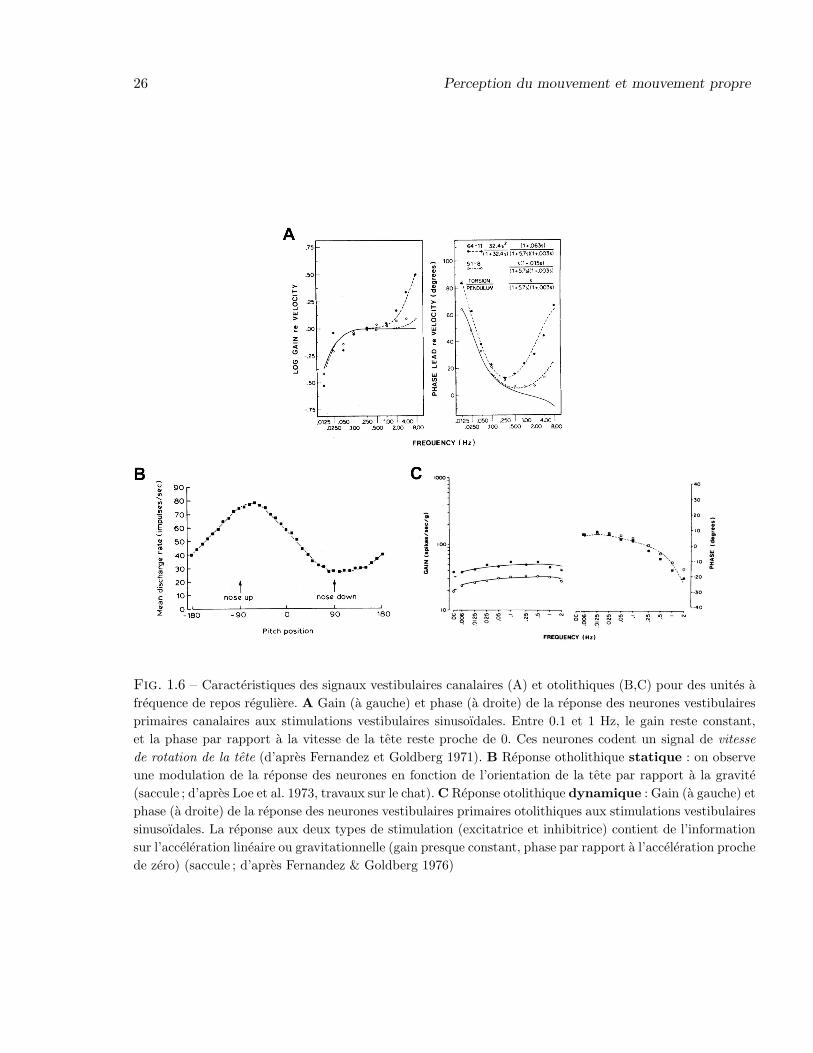
26 Perception du mouvement et mouvement propre
Fig. 1.6 – Caracteristiques des signaux vestibulaires canalaires (A) et otolithiques (B,C) pour des unites afrequence de repos reguliere. A Gain (a gauche) et phase (a droite) de la reponse des neurones vestibulairesprimaires canalaires aux stimulations vestibulaires sinusoıdales. Entre 0.1 et 1 Hz, le gain reste constant,et la phase par rapport a la vitesse de la tete reste proche de 0. Ces neurones codent un signal de vitessede rotation de la tete (d’apres Fernandez et Goldberg 1971). B Reponse otholithique statique : on observeune modulation de la reponse des neurones en fonction de l’orientation de la tete par rapport a la gravite(saccule ; d’apres Loe et al. 1973, travaux sur le chat). C Reponse otolithique dynamique : Gain (a gauche) etphase (a droite) de la reponse des neurones vestibulaires primaires otolithiques aux stimulations vestibulairessinusoıdales. La reponse aux deux types de stimulation (excitatrice et inhibitrice) contient de l’informationsur l’acceleration lineaire ou gravitationnelle (gain presque constant, phase par rapport a l’acceleration prochede zero) (saccule ; d’apres Fernandez & Goldberg 1976)

Le systeme vestibulaire 27
Autres signaux transmis par les neurones vestibulaires secondairesLa nature multimodale sur sens vestibulaire apparaıt tres tot, dans certains cas des les afferences
vestibulaires primaires. Ainsi, chez le poisson rouge (ou carassin dore, Goldfish), des reponses a delarges stimuli visuels (optocinetiques) ont ete decrites dans les neurones relies au canal horizontal(Klinke & Schmidt 1970). De telles reponses n’existent pas chez le primate (Keller 1976).
Dans les noyaux vestibulaires, des reponses visuelles ont ete mentionnees pour la premiere foispar Duensing et Schaefer (1958, chez le lapin). Dans la premiere etude detaillee chez le poissonrouge, Dichgans et al. (1973) ont montre que l’entree visuelle permet de lever l’ambiguıte vestibu-laire sur l’absence de mouvement ou la presence d’un mouvement a vitesse constante, ameliorantainsi significativement la fonction de ”speedometre” des neurones vestibulaires secondaires. Chezle singe, Henn et al. (1974) ont decrit des reponses visuelles dans les noyaux vestibulaires, reponsesqui, puisqu’elles sont absentes dans les neurones vestibulaires afferents, definissent une activitevestibulaire centrale distincte de l’activite vestibulaire peripherique (Keller 1976).
Plus generalement, les noyaux vestibulaires contiennent de nombreux types de neurones et dereponses differentes. Ainsi, en plus des reponses vestibulaires et visuelles, on trouve des reponses adiverses composantes du mouvement des yeux, notamment aux saccades oculaires (Miles 1974), al’acceleration angulaire (Fuchs & Kim 1975), a la poursuite oculaire (Keller & Kamath 1975) ouaux phases lentes du nystagmus optocinetique (Baker & Berthoz 1974). Parmi ces neurones lies aumouvement des yeux, certains repondent en phase avec la position et non pas avec la vitesse de latete (Keller & Kamath 1975).
Enfin, notons que les neurones otolithiques secondaires ont aussi des reponses visuelles, et qu’ilexiste une certaine convergence des afferences canalaires et otolithiques dans les noyaux vestibulaires(Wilson & Melvill-Jones, p.166, p. 176).
1.1.2.3 Activite reflexe : une fonction importante du systeme vestibulaire
Si une description exhaustive des activites reflexes impliquant le systeme vestibulaire est hors ducadre de ce travail, nous en verrons neanmoins le principe general, dans le cas particulier du reflexevestibulo-oculaire (VOR). De maniere generale, l’activation du labyrinthe provoque un ensemble dereflexes dans de nombreuses parties du corps, ces reflexes ayant pour objectif principal de stabiliserle regard dans l’espace. Pour ce faire, ils provoquent des mouvements compensatoires de la tetepar rapport au corps ainsi que des ajustements posturaux (reflexes vestibulo-coliques et vestibulo-spinaux du systeme vestibulo-spinal ; Wilson & Melvill-Jones 1979, chap.7), et des mouvementscompensatoires des yeux par rapport a la tete (reflexe vestibulo-oculaire ; Wilson & Melvill-Jones1979, chap.8). L’ensemble de ces reflexes prend toute son importance lors d’une activite aussicourante que la marche, laquelle serait litteralement impossible si le regard n’etait pas stabilise (cequi donnerait une vision similaire a celle d’un film tourne camera au poing).
Les caracteristiques geometriques des mouvements reflexes sont bien definies, et ont ete etudieesen premier lieu sur le VOR. Typiquement, lorsque la tete est tournee vers la droite, les yeux subissentune contre-rotation opposee de meme amplitude, en l’occurrence vers la gauche. Il en est de memepour les autres directions de rotation cephalique (Fig. 1.7). Lorsque la stimulation du labyrintheest prolongee, il apparaıt des mouvements oculaires rapides de repositionnement de l’oeil dansl’orbite, qui entrecoupent les mouvements compensatoires plus lents : cet ensemble de mouvementsoculaires, phases lentes et phases rapides, est appele nystagmus vestibulaire. Si l’on stimule lescanaux semi-circulaires individuellement, le nystagmus evoque (rotation des yeux) est dans le plandu canal stimule (Wilson & Melvill-Jones 1979, p.254).
Le circuit neuronal minimal sous-tendant le VOR comprend seulement trois neurones : leneurone vestibulaire afferent (corps cellulaire dans le ganglion de Scarpa), le neurone vestibulaire

28 Perception du mouvement et mouvement propre
secondaire (corps cellulaire dans le noyau vestibulaire et projetant directement sur le motoneu-rone), et le motoneurone extraoculaire (des noyaux oculomoteurs et abducens pour les mouvementhorizontaux)(voir aussi Fig. 1.7). Le premier neurone transmet un signal de vitesse de la tete, maisqui n’est pas maintenu a vitesse constante. Le second recoit le signal du premier, et aussi un signalvisuel qui est proportionnel a la vitesse du mouvement visuel, et qui, s’ajoutant au premier signal,permet le maintien de la reponse au mouvement de la tete meme a vitesse constante (entree op-tocinetique, Fig. 1.7). Le neurone vestibulaire secondaire est ainsi un speedometre fiable. Le signalde mouvement de la tete est ensuite transmis au motoneurone, a la fois directement et apres uneintegration pour indiquer la position finale de l’oeil (signal supplementaire de fixation). Le mouve-ment de l’oeil genere par ce circuit est identique mais de direction opposee a celui de la tete. Enpratique, le circuit tri-neuronal du VOR n’est pas isole. La formation reticulee du tronc cerebral,ainsi que le vestibulo-cervelet contribuent egalement au VOR.
Fig. 1.7 – Reflexe vestibulo-oculaire (VOR). Les entrees vestibulaire (V : θ′t,v, estimation de vitesse de la
tete) et optocinetique (Oc : θ′t,o, estimation de la vitesse de l’oeil dans l’orbite) sont combinees au niveau des
noyaux vestibulaires. Le signal de commande resultant (θ′o) est ensuite transmis d’une part aux motoneurones
extraoculaires (MN) directement et via un integrateur pour fournir la commande motrice de vitesse et deposition oculaires, et d’autre part θ′
o est reinjecte en copie efferente pour calibrer le signal optocinetiqueprovenant du glissement retinien (θ′
gr). Le mouvement physique de la tete (vitesse θt) est ainsi transformeen un mouvement physique oculaire (vitesse θo) d’amplitude identique et de direction opposee, suivi d’unefixation de l’oeil a la nouvelle position. (d’apres Robinson 1977)
L’influence de ce dernier a ete particulierement etudiee depuis les travaux theoriques fecondsde Ito (1972) qui propose que le cervelet agit comme un ”ordinateur” sur les reflexes vestibulaires,dans le sens ou, lorsque la stabilisation est inadaptee, le cervelet agit sur le signal d’erreur pourreadapter le reflexe. Il a ete demontre ensuite d’une part que le vestibulo-cervelet (en particulier lenodule) recoit des informations visuelles, et d’autre part que les cellules de Purkinje produisent uneinhibition selective (par rapport aux plans des canaux) sur la connexion labyrinthe–motoneuronesextraoculaires du circuit tri-neuronal (voir Wilson & Melvill-Jones 1979, pp.281-283, 302). En parti-culier, du fait de l’influence opposee des afferences visuelle et vestibulaire sur les cellules de Purkinje,ces dernieres contribuent largement a l’inhibition du VOR pour permettre la fixation d’un objetpendant le mouvement de la tete. La boucle de retroaction entre les noyaux vestibulaires et levestibulo-cervelet permet des adaptations rapides (i.e. passer du VOR a la fixation), par opposition

Le systeme vestibulaire 29
a la plasticite adaptative du VOR –consequente a une perturbation au long terme du rapport entreles afferentes visuelle et vestibulaire (port de prismes)– qui indique des changements plastiques etde longue duree du gain de VOR et qui depend de l’integrite du cortex vestibulaire cerebelleux(Wilson & Melvill-Jones 1979, p.310-316).
1.1.2.4 Activite vestibulaire, nystagmus et sensation de mouvement
Dans certains cas, il existe des dissociations entre l’activite vestibulaire et le comportement(nystagmus) ou la sensation de mouvement de la tete (Henn 1982). Ainsi, lorsqu’on augmente lavitesse d’un stimulus optocinetique, la vitesse des yeux et la sensation de mouvement continuent aaugmenter meme apres que les neurones vestibulaires aient atteint leur seuil de decharge maximale.Lors de la suppression du nystagmus, malgre l’absence de mouvements des yeux, l’activite dansles noyaux vestibulaire et la sensation de mouvement restent inchangees. Enfin, lors d’un conflitvisuo-vestibulaire ou le stimulus visuel tourne avec l’animal, malgre la presence d’une stimulationvestibulaire, l’activite dans les noyaux vestibulaires est inhibee mais elle reste inchangee dans lenerf vestibulaire. Pour de faibles accelerations, la sensation de mouvement est nulle. La presencede ces dissociations entre l’activite vestibulaire d’une part, et le comportement ou la sensationde mouvement de l’autre, montre un degre de complexite supplementaire du systeme vestibulaire,faisant probablement intervenir le cortex cerebral.
1.1.3 Le reseau vestibulaire cortical
1.1.3.1 De multiples aires corticales vestibulaires
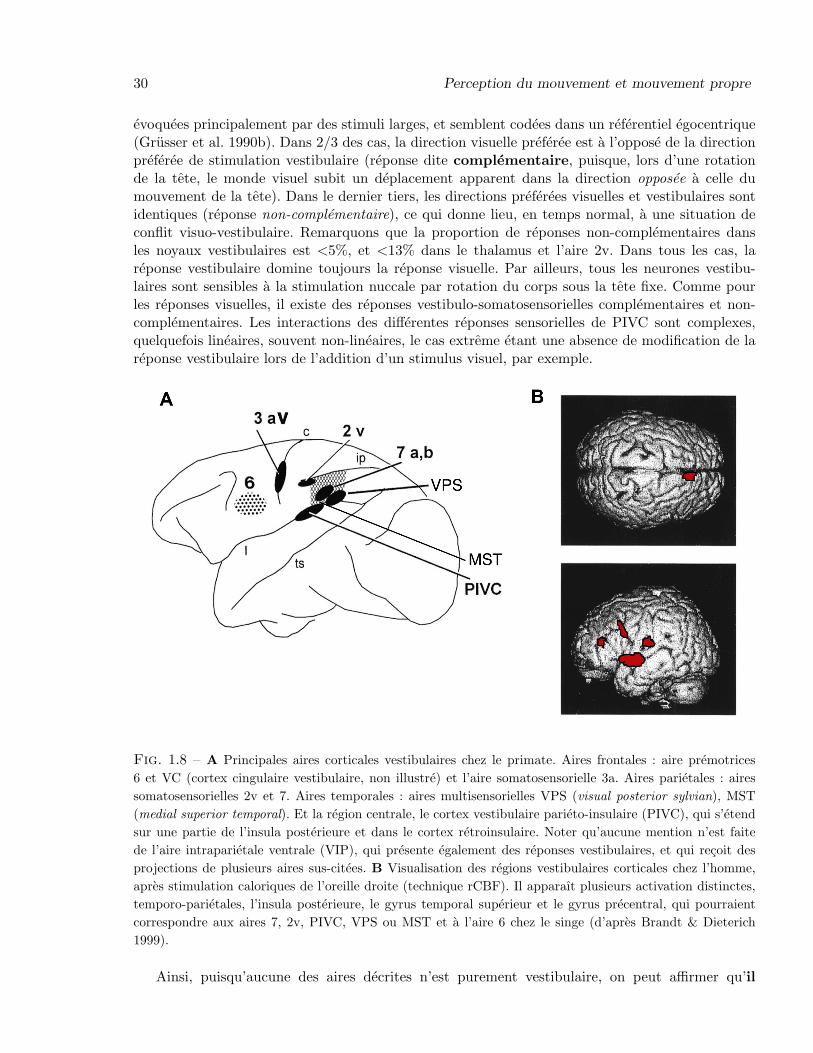
Ainsi que nous l’avons vu dans la section 1.1.2, apres un premier relais tronculaire et un se-cond thalamique, l’information vestibulaire arrive au cortex (Figure 1.5). La recherche de l’entreecorticale primaire, a l’instar du cortex visuel primaire, a donne chez le singe des resultats sur-prenants (comme auparavant chez le chat ; Sans et al. 1969). A l’aide de potentiels evoques puisd’electrophysiologie, une premiere aire (2v) est mise en evidence a la pointe anterieure du sillonintraparietal (Schwarz & Fredrickson 1971). Puis, dans l’aire somatosensorielle frontale 3a, desreponses vestibulaires sont decouvertes, et ensuite dans l’aire somatosensorielle 7a, et aussi dansune region du cortex insulaire, le cortex vestibulaire parieto-insulaire ou PIVC (Grusser et al. 1990b ;Fig. 1.8A ; voir aussi Guldin & Grusser 1998). Si toutes ces aires contiennent effectivement des neu-rones repondant a la stimulation vestibulaire, aucun de ces neurones ne repond exclusivement acette derniere.
Dans l’aire 2v, les neurones vestibulaires repondent aussi a la rotation des articulations desmembres superieurs, et 80% d’entre eux repondent egalement a de larges stimuli visuels (stimulioptocinetiques ; Buttner & Buettner 1978). Dans l’aire 3a, les neurones vestibulaires repondentaussi a des stimulations somatosensorielles (nuccales), ce qui est aussi le cas des neurones de l’aireparietale associative 7, et du PIVC. L’aire MST, quant a elle (Fig. 1.8A), fait partie de la voievisuelle dorsale et se trouve impliquee dans la perception du mouvement (cf. section 1.3.2). Enfin,l’aire visuelle post-sylvienne (VPS ; Guldin & Grusser 1998) possede, elle aussi, des neurones visuo-vestibulaires.
Le cortex vestibulaire parieto-insulaire a ete etudie en detail par Akbarian et al. (1988) etGrusser et al. (1990b). Situee au fond du sillon sylvien et s’etendant dans la partie retroinsulaire,c’est une aire corticale multisensorielle possedant en plus des reponses vestibulaires, des reponsesvisuelles (80% des neurones vestibulaires) et somatosensorielles (100% des neurones vestibulaires).A ce titre, elle ressemble particulierement a l’aire intraparietale ventrale (qui est l’objet principalde notre etude), sur laquelle elle se projette d’ailleurs (cf. section 2.5). Les reponses visuelles sont
30 Perception du mouvement et mouvement propre
evoquees principalement par des stimuli larges, et semblent codees dans un referentiel egocentrique(Grusser et al. 1990b). Dans 2/3 des cas, la direction visuelle preferee est a l’oppose de la directionpreferee de stimulation vestibulaire (reponse dite complementaire, puisque, lors d’une rotationde la tete, le monde visuel subit un deplacement apparent dans la direction opposee a celle dumouvement de la tete). Dans le dernier tiers, les directions preferees visuelles et vestibulaires sontidentiques (reponse non-complementaire), ce qui donne lieu, en temps normal, a une situation deconflit visuo-vestibulaire. Remarquons que la proportion de reponses non-complementaires dansles noyaux vestibulaires est <5%, et <13% dans le thalamus et l’aire 2v. Dans tous les cas, lareponse vestibulaire domine toujours la reponse visuelle. Par ailleurs, tous les neurones vestibu-laires sont sensibles a la stimulation nuccale par rotation du corps sous la tete fixe. Comme pourles reponses visuelles, il existe des reponses vestibulo-somatosensorielles complementaires et non-complementaires. Les interactions des differentes reponses sensorielles de PIVC sont complexes,quelquefois lineaires, souvent non-lineaires, le cas extreme etant une absence de modification de lareponse vestibulaire lors de l’addition d’un stimulus visuel, par exemple.
Fig. 1.8 – A Principales aires corticales vestibulaires chez le primate. Aires frontales : aire premotrices6 et VC (cortex cingulaire vestibulaire, non illustre) et l’aire somatosensorielle 3a. Aires parietales : airessomatosensorielles 2v et 7. Aires temporales : aires multisensorielles VPS (visual posterior sylvian), MST(medial superior temporal). Et la region centrale, le cortex vestibulaire parieto-insulaire (PIVC), qui s’etendsur une partie de l’insula posterieure et dans le cortex retroinsulaire. Noter qu’aucune mention n’est faitede l’aire intraparietale ventrale (VIP), qui presente egalement des reponses vestibulaires, et qui recoit desprojections de plusieurs aires sus-citees. B Visualisation des regions vestibulaires corticales chez l’homme,apres stimulation caloriques de l’oreille droite (technique rCBF). Il apparaıt plusieurs activation distinctes,temporo-parietales, l’insula posterieure, le gyrus temporal superieur et le gyrus precentral, qui pourraientcorrespondre aux aires 7, 2v, PIVC, VPS ou MST et a l’aire 6 chez le singe (d’apres Brandt & Dieterich1999).
Ainsi, puisqu’aucune des aires decrites n’est purement vestibulaire, on peut affirmer qu’il

Le systeme vestibulaire 31
n’existe pas de cortex vestibulaire primaire par analogie aux cortex visuel ou auditif pri-maires. A partir des deux observations suivantes, Brandt & Dieterich (1999) nous livrent unehypothese interessant sur cette absence de cortex vestibulaire primaire. D’une part, la stimulationnaturelle du systeme vestibulaire pendant le mouvement de la tete et la locomotion est toujoursmultisensorielle. D’autre part, par contraste avec les stimuli visuels et auditifs, les caracteristiquesphysiques de la stimulation vestibulaire sont definies completement par seulement deux parametres,la direction et l’amplitude de l’acceleration subie par la tete. Ainsi, il se pourrait tout simplementqu’une aire vestibulaire primaire ne soit pas necessaire.
1.1.3.2 Architecture du reseau vestibulaire cortical
Partant du constat qu’il existe plusieurs aires vestibulaires chez differentes especes de primates,Guldin & Grusser (1998) postulent l’existence d’un veritable reseau vestibulaire cortical, dont PIVCserait la region centrale, en particulier parce qu’elle possede largement plus de neurones vestibulairesque les autres aires. Si chacune des aires vestibulaires possede sa propre entree sensorielle thalamiqueet sa propre connexion corticofugale vers les noyaux vestibulaires, les travaux anatomiques deGuldin & Grusser devoilent nettement l’existence d’un reseau cortico-cortical (Figure 1.9).
Fig. 1.9 – Le reseau vestibulaire cortical. Seules sont representees les aires corticales presentant un marquagedense apres injection dans aux moins deux des trois aires vestibulaires representees dans une ellipse. Toutesles aires corticales definies comme etant vestibulaires a partir de la physiologie sont retrouvees (compareravec la figure 1.8). Remarquer le nombre important de connexions entre les differentes aires vestibulaires eten particulier les connexions reciproques, l’ensemble definissant un veritable reseau d’aires interagissant pourle traitement des informations vestibulaires dans le cortex. Remarquer cependant que seule PIVC recoit desafferences de toutes les aires du reseau. PIVC serait ainsi la region centrale du reseau vestibulaire cortical(d’apres Guldin & Grusser 1998)
Le role central de PIVC est confirme par l’anatomie : c’est la seule aire vestibulaire qui recoitdes entrees de toutes les autres aires vestibulaires corticales. Guldin & Grusser (1998) proposentque PIVC est l’aire ou l’information des differents centres corticaux est reunie en un concept[d’information sur] la tete dans l’espace.

32 Perception du mouvement et mouvement propre
Remarquons toutefois que l’aire intraparietale ventrale (VIP) ne figure pas dans le reseau vesti-bulaire decrit, alors que nous avons montre precedemment au laboratoire qu’elle possede de nom-breuses reponses vestibulaires (Graf et al. 1995 ; Bremmer et al. 1997), et qu’elle est connecteedirectement aux aires 2v, 3a, 7, MST et PIVC (Lewis & Van Essen 2000a,b ; cf. section 3).
1.1.3.3 Le cortex vestibulaire chez l’homme
Comme chez le singe, la stimulation vestibulaire (stimulation calorique, activation des canauxsemi-circulaires) provoque l’activation de multiples aires corticales chez l’homme (Fig. 1.8B). Brandt& Dieterich proposent que le cortex parieto-insulaire de l’homme est l’equivalent de PIVC chez lesinge. Deux differences semblent cependant exister : le cortex parieto-insulaire est active avec unedominance dans l’hemisphere cerebral droit, et il peut etre active par stimulation galvanique ce quisuggere une entree otolithique dans cette region chez l’homme. Or, aucune reponse a l’orientationstatique par rapport a la gravite n’a ete retrouvee dans PIVC chez le singe (Grusser et al. 1990b). Lalocalisation exacte de l’equivalent chez l’homme du PIVC est toujours sujet de discussion, puisqued’autres etudes semblent le placer dans la partie posterieure du sillon lateral (Lobel et al. 1998 ; deWaele et al. 2001).
L’existence d’entrees vestibulaires canalaires et otolithiques chez l’homme semble etre confirmeepar la pathologie. Lorsqu’il est lese, le cortex parieto-insulaire provoque deux ensembles symptoma-tiques. Le premier est une desorientation spatiale corticale statique, aboutissant a une inclinaisonde la verticale subjective et une latero-pulsion corporelle associee. Le second est une desorientationspatiale dynamique, avec mouvement apparent ou vertiges rotatoires (Brandt & Dieterich 1999).
1.1.4 Psychophysique et pathologie
1.1.4.1 Psychophysique vestibulaire
– Perception du mouvement angulaireLes resultats d’une experience de psychophysique ou les sujets devaient detecter une rotation
qui leur etait imposee montre que la courbe de gain de la detection en fonction de la frequence destimulation se superpose a celle obtenue par Fernandez et Goldberg (1971) sur les neurones vesti-bulaires secondaires chez le singe (Benson 1990). Entre 0.1 et 1 Hz, il apparaıt une predominance dela vitesse dans le mecanisme sensoriel, et en dessous de 0.1 Hz, une predominance de l’acceleration(cf. Fig. 1.6).
– Perception du mouvement lineaireTeste avec des mouvements lineaires ”aller-retour” sinusoıdes, le seuil de detection du mouve-
ment est plus faible dans le plan horizontal que pour les deplacements haut-bas (Benson 1990).Chez le singe, la difference de seuil vertical et horizontal est retrouvee qualitativement, mais elleest quantitativement plus importante : la perception des stimuli lineaires a basse frequence estdeterminee principalement par la dynamique de transduction des otolithes (cf. aussi Fig. 1.6).
– Perception de la verticale (posture)Des experiences a l’aide d’un siege orientable dans les plans sagittal et frontal montrent que
les estimations des sujets sains sont a 1◦ (± 2.2◦) de la verticale vraie. Avant d’attribuer cesperformances aux otolithes, deux reserves sont a emettre. D’une part, les sujets plonges dans unepiscine, prives ainsi de vision et d’information somatosensorielle, ont beaucoup de mal a retrouverla verticale (l’arc d’erreur devient ± 31◦). D’autre part, les patients labyrinthiques obtiennent des

Flux optique et autres informations sensorielles. 33
performances seulement tres faiblement inferieures aux sujets sains (Benson 1990). Ainsi, la visionet la proprioception jouent un role predominant dans la perception de la verticale.
– Les limites de la fonction vestibulaire qui causent des erreurs de perceptionL’existence d’un seuil, en dessous duquel aucun message vestibulaire n’est genere, implique que
de legers mouvements peuvent etre indetectables.Puisque le capteur vestibulaire est un capteur d’acceleration, un mouvement a vitesse constante
durant plus longtemps que la constante de temps ne genere plus de message nerveux (cf. Fig. 1.2),ce qui peut entraıner des sensations erronees de mouvement angulaire. Un mouvement de la teteautour d’un axe apres une rotation de longue duree autour d’un axe perpendiculaire a ce dernierproduit l’illusion d’un mouvement autour du troisieme axe orthogonal aux deux premiers.
Le systeme vestibulaire est incapable de distinguer l’influence de l’acceleration de celle de lagravite. Ainsi, lors d’une deceleration dans un avion, la resultante vectorielle de l’acceleration etde la gravite est dirigee vers l’arriere de l’appareil, donnant ainsi au pilote une fausse sensationd’ascension verticale : c’est l’illusion somatogravique.
Enfin, la maladie des transports resulte d’une incoherence entre les informations recues et cellesattendues par le cerveau, notamment les informations visuelles et vestibulaires.
1.1.4.2 Le sens vestibulaire et la representation de l’espace
De nombreuses preuves experimentales soulignent l’importance du sens vestibulaire dans larepresentation de l’espace. Nous ne citerons que quelques exemples.
– En ce qui concerne l’orientation spatiale et la mise a jour continue de la representationinterne de l’espace, l’importance du sens vestibulaire est demontree par les deficits de memoirespatiale consecutifs a un sejour prolonge en microgravite. La performance des sujets dans une tachede pointage vers une cible memorisee est degradee, a cause d’une erreur de jugement sur la positionde la cible et non sur la direction du bras(Watt 1997).
– Les pathologies du nystagmus vestibulaire, qui entraınent un nystagmus constant et irrepressible,provoquent egalement des desorientations spatiales dans la direction du nystagmus (Dieterich &Brandt 1999).
– Enfin, des lesions tout au long des voies vestibulaires, ainsi que des sejours prolonges enmicrogravite, provoquent, lors d’un mouvement de la tete (stimulation vestibulaire), une illusionspatiale particuliere. L’illusion de la piece penchee (room tilt illusion) correspond a l’impressiontransitoire d’etre a l’envers (la tete en bas), ou encore que la piece dans laquelle on se trouve esttournee de 90◦. Brandt (1999) propose que l’illusion de la piece penchee proviendrait d’un defautd’alignement transitoire des coordonnees spatiales 3D visuelles et vestibulaires. Ces disparites, d’uneamplitude de 90◦ ou 180◦ seraient en fait la consequence des ”essais” d’alignement des messageserrones (ou de resolution du conflit visuo-vestibulaire) au niveau du cortex. L’illusion disparaıtlorsque le systeme visuel impose sa reference au systeme vestibulaire, autrement dit, lorsque laverticale basee sur des informations visuelles empiriques (sens vertical des meubles ou de la porte...)est determinee comme la verticale unique.
1.2 Flux optique et autres informations sensorielles.
D’un point de vue pragmatique, si le systeme visuel sert a la locomotion, il doit representer lemouvement propre dans une forme appropriee pour agir dans l’environnement. De plus, la locomo-tion comporte un grand nombre de taches differentes qui a leur tour demandent des informations

34 Perception du mouvement et mouvement propre
visuelles differentes (specifiques). Par consequent, dans une certaine mesure, les informations per-tinentes dependent de la tache locomotrice a realiser. Ainsi, l’etude de la perception visuelle dumouvement propre a ete divisee en fonction des questions suivantes : comment savoir si on sedeplace, a partir du moment ou on se deplace ; vers ou va-t-on ; et enfin, une fois qu’on est en route,comment eviter les obstacles ? Ces differentes questions ont engendre des champs de recherchedistincts, a savoir : un champ de recherche theorique sur les indices disponibles qui caracterisentle mouvement propre par opposition, par exemple, ou mouvement exterieur au sujet ; un champconsacre a la perception de la direction de deplacement (heading) ; et enfin, un champ consacre auxaspects de prediction du temps de contact avec un obstacle (notamment la theorie τ ou time tocontact). Dans ce chapitre, qui comme le precedent est consacre a la description des informationssensorielles relatives au mouvement propre, le lecteur trouvera une presentation du flux optiquedans une premiere partie, et une revue tres succincte des autres informations sensorielles qui contri-buent a la perception du mouvement propre. Les autres questions relatives a l’information optiquerelative au mouvement meritent un chapitre a part entiere, et seront developpees en suivant.
1.2.1 Considerations theoriques et definition du flux optique
Avant de debuter, remarquons simplement avec Koenderink (1990) qu’en raison de l’absenced’effet inertiel en optique, il est impossible de mesurer directement l’acceleration optique commedans un systeme mecanique (tel le systeme vestibulaire). Par consequent, il faut extraire les pa-rametres optiques du mouvement par des procedes spatio-temporels a partir de l’image retiniennequi varie dans le temps.
1.2.1.1 Definition du flux optique
C’est Gibson (1966) qui le premier remarque que le champ de mouvement visuel contient desinformations pertinentes pour servir la locomotion. Il a notamment observe que l’ensemble des lignesreliant un objet dans l’environnement a un point de reference (centre de l’oeil de l’observateur) subitun mouvement d’expansion lorsque le point de reference se deplace vers l’avant. Seule la ligne quicoıncide avec la direction de mouvement reste inchangee. C’est l’ensemble des vecteurs associes achaque direction –qui decrivent le mouvement– qui est appele flux optique, et son centre invariantle foyer d’expansion (voir Figure 1.10).
Malheureusement, le flux optique est rarement aussi simple. Meme pour un sujet en translationpure, la retine etant fixee sur une serie de supports mobiles (dont l’oeil et la tete) qui tournentles uns par rapport aux autres, le flux optique resultant est rarement purement expansif. Ceciamene a distingue le flux optique (optical flow, optic array) qui ne depend que des mouvements detranslation du sujet, du flux retinien (retinal flow, optic flow) qui lui depend de la translation dusujet et des mouvements relatifs de rotation des differents segments du corps (Van den Berg 2000,Warren 1990). Du point de vue des formulations mathematiques du flux optique, il n’existe pasde consensus (Warren 1990). Ainsi, la description des differents formalismes depasse le cadre denotre etude (voir par exemple Koenderink 1990 et Zacharias, 1990). Les equations proposees parles auteurs dependent de la nature exacte du probleme qu’ils se posent. Par exemple, Koenderink(1990) propose un formalisme plus adapte a la recherche de la direction de deplacement (heading),et Zacharias (1990) une formule derivee adaptee a l’extraction de la forme a partir du flux optique.C’est pourquoi les deux formalismes different par leur expression de la rotation, le premier utilisantla rotation propre (egorotation) et le second la rotation induite par le mouvement (induced rotation).Ceci dit, les differents formalismes ont des caracteristiques communes. Le flux optique canonique estdefini comme tout champ de vecteurs dont les elements individuels changent leur position optique enfonction d’equations derivees sous les hypothese d’un environnement rigide et de trajet rectiligne de

Flux optique et autres informations sensorielles. 35
Fig. 1.10 – Representation d’un flux optique simulant un mouvement vers l’avant. Au centre (en rouge),le foyer d’expansion. Chaque vecteur de mouvement (lignes flechees, noires) est caracterise par sa ligne dedirection (ligne pointillee, en vert) le reliant au foyer d’expansion. Chacun de ces vecteurs represente lemouvement visuel provoque par le deplacement de l’observateur.
la lumiere (Warren 1990, p.15). Ces formalismes incorporent a la fois les mouvements de translationet de rotation du point de reference (observateur). Enfin, elles sont souvent suffisamment generalespour pouvoir etre exprimees independamment d’un systeme de coordonnees particulier (cf. parexemple Warren 1990).
1.2.1.2 les differents indices disponibles
- indices directement bases sur le flux optique
(i) Considerons les cas theoriques d’une translation et d’une rotation pures de l’observateur dansun environnement regulier et stable (Figure 1.11 ; Koenderink 1990). Dans le cas d’une translationpure (Figure 1.11A), la direction des lignes de champs indique celle du mouvement de translation.Les poles, situes sur l’horizon, correspondent aux lieux d’ou l’on vient et ou l’on va (Warren, 1990,p.16). Par contre, l’amplitude optique depend de la distance de l’observateur a l’objet qui generele flux optique. Pour une rotation pure (Figure 1.11B), l’axe defini par les deux poles est l’axedu mouvement de rotation, et l’amplitude de la rotation est directement determinee par celle desvecteurs de champ. Cette fois, l’amplitude ne depend pas de la configuration de l’environnement.Ainsi, les flux optiques consequents a une translation et une rotation ne donnent par les memesinformation sur le mouvement. La situation se complique encore dans la realite parce que le champvisuel ne couvre pas 360◦ (voir Figs. 1.11 C et D pour un mouvement de translation et de rotation,respectivement). Il s’agit alors d’extraire l’information sur le type de mouvement en cours a partir

36 Perception du mouvement et mouvement propre
Fig. 1.11 – A Representation du flux optique total du a un mouvement de translation pur. Les lignes dechamp sont des demi-cercles (ou meridiens) qui connectent les poles. B Flux optique du a un mouvement derotation pur. Les lignes de champ sont des cercles paralleles (ou latitudes). Les deux poles definissent l’axede la rotation. C Champ visuel restreint du flux optique de translation (A). Noter la difficulte de determinerla position des poles. Une petite incertitude sur les directions des vecteurs entraınerait de grandes variationsdans l’estimation des poles. D Champ visuel restreint du flux optique de rotation (B). Pour determiner laposition des poles, il faut estimer l’axe des centres de courbure de ces lignes de champ. Une petite incertitudesur ces direction peut meme rendre cette determination impossible. (d’apres Koenderink, 1990)
des lignes de champs visibles. On comprend alors l’importance des problemes poses par l’incertitudede mesure des directions, par exemple, qui peut rendre la tache d’extraction tres delicate. De plus,notons que les figures 1.11C et D sont assez ressemblantes. Ainsi, si le champ visuel est tres restreint,il peut devenir impossible de distinguer translation et rotation. Enfin, les mouvements naturelscomprennent en general translation et rotation, ajoutant un degre de complexite supplementaireau probleme d’extraction des parametres du mouvement propre a partir des informations du fluxoptique.
(ii) a propos de la distanceDans le systeme visuel, les seules mesures disponibles sont des mesures d’angle, ce qui pose
un certain nombre de difficultes pour mesurer les distances en profondeur (Koenderink 1990 ; cf.discussion de Wertheim (1990) sur le sujet). Cependant, dans la majorite des cas de la vie de tousles jours, l’environnement est compose d’ensembles d’objets distribues dans l’espace, et chaqueensemble possede son ordre propre de profondeurs relatives. L’information spatiale (de profondeur)est souvent partielle. De plus, elle n’est pas forcement necessaire en tant que telle : d’abord, ilest peu probable que l’on extraie la forme des objets a partir de la distance, et ensuite, dansde nombreuse situation, l’on peut s’affranchir d’estimer les distances et calculer directement desdescripteurs spatiaux d’ordre superieur (tilt, slant, curvature, ...). Par exemple, si un objet A

Flux optique et autres informations sensorielles. 37
occlut partiellement un objet B et que l’on peut identifier (la structure de) l’objet A a partir dela structure du recouvrement, de la dynamique de l’occlusion, ou d’autres indices visuels, il estfacile de determiner la direction du mouvement a realiser pour avoir une vue complete de l’objetcache. Aucune information de distance n’est necessaire, seules les relations radiales (radial orders)suffisent (Koenderink 1990).
Dans la mesure ou le flux optique est un champ de vecteurs de mouvement, d’autres pro-prietes plus globales que la direction et l’amplitude des vecteurs individuels peuvent apporter desinformations precieuses, par exemple les gradients, la divergence, et les trajets ou traces optiques(integration dans le temps) (Warren, 1990).
En dehors des informations vectorielles sur le mouvement, celles qui concernent la structurede l’environnement ont aussi leur importance (Koenderink, 1990). Trois ordres d’information sontdistingues. L’ordre zero est simplement la position des reperes. Le 1er ordre est la descriptiondes surfaces, le parametre principal etant l’orientation des unites de surfaces, qui generalementdependent de la position. Enfin, le 2e ordre decrit la courbure des unites de surface, pouvant etredecrite par un tenseur specifiant la rotation de la normale a l’unite de surface pour tout deplacementunitaire de l’observateur. De fait, cette categorie depend de la position et de l’unite de surface enquestion. Il est remarquable que l’observateur humain semble capable d’extraire les proprietes dezero, premier et second ordre independamment. Nous verrons des utilisations pratiques de cettecategorisation formelle a travers l’etude du mouvement relatif (section 1.3.1), de l’extraction de lastructure a partir du mouvement (section 2.1.1.2) et d’une approche theorique sur la metrique del’espace (2.7).
- indices indirects
Un certain nombre de ces indices indirects derive de l’etude de la perception de la vitessepropre (egospeed) et de l’altitude (Warren, 1990). La perception de la vitesse varie avec l’altitude,elle est maximale dans un vehicule tres bas sur la route, minimale dans un avion a haute altitude.Ainsi, on definit des variables telles que la variation de flux global qui est le quotient de lavitesse par l’altitude (v/h), la densite optique globale comme le quotient de l’altitude sur lataille des elements visibles au sol et la variation de texture optique definies comme le nombred’elements de reference depasses par unite de temps. L’influence des differentes variables peutetre illustree par des situations experimentales. Par exemple, lorsqu’un conducteur de vehicule sedeplace sur une route bordee de platanes a vitesse constante, si l’espacement entre les elementsde references (les arbres) diminue, le conducteur aura la sensation d’accelerer, et inversement sil’espacement augmente, il croira ralentir. Cet effet peut etre encore plus dramatique en vol, puisqueune diminution de densite optique globale (v apparente diminue)couplee aux effets de la variationde flux global (v/h) donnera l’illusion au pilote d’une augmentation de son altitude.
- flux non canoniques
La definition formelle du flux optique canonique (cf. 1.2.1.1) comprend l’hypothese d’un deplace-ment dans un environnement rigide. L’on parle de flux optique non canonique lorsque l’une aumoins des hypotheses initiales n’est par respectee. Ainsi, un deplacement dans un environnementnon rigide, ou donne lieu a un flux non canonique (Warren 1990). Un primate se deplacant dans unarbre aux branches secouees par le vent, un navigateur sur une mer agitee ou un passant dans unefoule en mouvement en sont quelques exemples. Le fait qu’il soit neanmoins possible dans ce genrede situation de percevoir son propre mouvement et de se deplacer sans difficulte illustre un degre decomplexite supplementaire dans la perception du mouvement propre, ce qui amene Warren (1990)

38 Perception du mouvement et mouvement propre
a s’interroger sur la notion information perceptible, et l’impossibilite de s’affranchir artificiellementou de contourner le lien perception/action.
1.2.1.3 Limites du systeme visuel
L’existence de limites du systeme visuel, et leur analyse, doivent etre considerees dans les hy-potheses sur les parametres (optiques) du flux optique supposes pouvoir etre extraits par le cerveau(comme la variation de texture optique, voir 1.2.1.2). Nakayama (1990) resume les principales li-mites.
- La resolution spatiale est variable : des experiences de perception du contraste (utilisant desgrilles dont le contraste varie dans l’espace selon une fonction sinusoıde) montrent qu’elle diminuelineairement du centre vers la peripherie de la retine. Le seuil y est alors 35 fois plus importantqu’au centre. Ce phenomene est d’ailleurs accentue par l’amplification visuelle corticale de la regioncentrale de la retine (c’est-a-dire le fait qu’environ autant de surface corticale est dediee a la retinecentrale qu’a la retine peripherique malgre une grande difference de surface entre ces deux regions).
- La resolution temporelle est variable : pour etre percus au centre de la retine, les stimuli visuelsdoivent etre au moins deux fois plus lents que les plus rapides pouvant etre percus en peripherie.
- L’etablissement de courbes spatio-temporelles montre qu’a chaque vitesse correspond unebande restreinte de frequences spatiales visibles : a faible vitesse, seules les frequences spatiales treselevees sont percues, et plus la vitesse augmente, plus les frequences spatiales visibles diminuent. Leparametre qui semble constant est la frequence temporelle (ftemp = fspa∗Vdepl). Ainsi, il existe doncune sorte de filtrage des frequences spatiales visibles en fonction de la vitesse de deplacement. Ceslimites (temporelles) sont probablement fixees tres tot, des le niveau des photorecepteurs (Nakayama1990).
Ces resultats sont potentiellement tres importants pour preciser le role des informations optiquesdans la perception du mouvement propre en particulier.
1.2.2 Les autres messages sensoriels influant sur la perception du mouvementpropre
Hormis les informations sensorielles vestibulaires (cf. 1.1.2, 1.4.1) et visuelles (cf. 1.2 ,1.3), ilexiste encore deux types de signaux principaux qui peuvent contribuer a la perception du mouve-ment propre, l’information somatosensorielle et les copies motrices efferentes (Barlow 1964).
L’information somatosensorielle se compose de deux elements. D’une part, les informationsproprioceptives (fuseaux neuro-musculaire, fibres α et recepteurs sensoriels tendineux) et ki-nesthesiques (arthrokinesie, recepteurs articulaires) –qui influent sur la sensation de mouvementpropre induite visuellement (vection, cf. 1.4.1, Mergner et al. 1993, 1997)–, et d’autre part les fluxsensoriels tactiles (recepteurs intra-dermiques) –qui peuvent aussi induire une sensation de mou-vement propre (Dichgans & Brandt 1978, cf. 1.4.1), et qui se retrouvent intimement lies aux fluxsensoriels visuels dans les neurones du cortex parietal (aire VIP, Duhamel et al. 1991, cf. 2.5).
Enfin, les informations motrices, en particulier oculomotrices et cephalomotrices, puisqu’ellescon- cernent directement le mouvement des capteurs visuels et vestibulaires par rapport au corpset dans l’espace, jouent aussi un role dans la perception du mouvement propre. Ces informations,telles que les copies efferentes des commandes motrices (Von Holst & Mittelstaedt 1950) oules decharges collaterales (copies des signaux des motoneurones, par exemple) participent parexemple a la decomposition du flux optique necessaire a la perception de la direction du deplacementde l’observateur (Crowell et al. 1998 ; cf. 1.3.3).

Flux optique et mouvement propre 39
1.3 Flux optique et mouvement propre
Cette partie est plus particulierement consacree a etudier comment le systeme nerveux extraitdu flux optique les informations sur le deplacement dans l’espace du sujet, ainsi que la direction dudeplacement en question.
1.3.1 Flux optique et perception du mouvement
Peut-on savoir, simplement a partir du flux optique, si l’on bouge ou non ? A priori, cettequestion peut sembler triviale, puisque dans la realite, l’information visuelle n’est jamais isolee.Cependant, du point de vue purement visuel, la reponse n’est pas aussi simple qu’il paraıt au depart,parce que le mouvement visuel peut etre provoque soit par le mouvement du sujet, soit par unmouvement externe au sujet, ou encore les deux a la fois. Ainsi, Duncker (1929) definit le mouvementrelatif au sujet (subject relative motion) comme tout mouvement en reference a l’observateur, quecelui-ci se percoive (ou soit) en mouvement ou non. Un autre mouvement important, parce qu’ildonne aussi des informations sur l’environnement, et qui est accessible par le flux optique, est lemouvement relatif entre objets (Duncker 1929 : object relative motion). Celui-ci peut etre illusoire,comme dans le cas ou les nuages en mouvement dans le ciel donnent l’impression que la lune sedeplace. Dans cet exemple, remarquons que l’existence de l’illusion illustre que, pour le systemevisuel, le mouvement d’un objet A par rapport a un objet B n’est pas equivalent au mouvementde l’objet B par rapport a l’objet A, mais depend de la position des yeux. L’objet poursuivi parle regard est automatiquement defini comme le point de reference, par rapport auquel se deplacele second objet. Changer le point de fixation change la reference, donc change tout le percept(Johansson 1978). Nous verrons au chapitre suivant (1.4) que ce constat reste valable entre unobjet et l’egocentre.
1.3.1.1 Comment extraire l’information sur les mouvements a partir du flux optique ?
Andersen (1990) propose trois etapes theorique pour la segregation du flux optique en deuxcomposantes relatives au mouvement des objets d’une part, et au mouvement propre d’autre part.La premiere etape consiste en l’extraction du champ des vecteurs 2D (flux optique 2D) a partirimages optiques successives. La deuxieme, c’est l’extraction des vitesses 3D a partir du flux 2D.La troisieme et derniere etape consiste a decomposer les vitesses 3D en mouvement propre etmouvement d’objet. La composante de mouvement propre est determinee comme etant le vecteur3D (constant) qui est applique a tout le flux optique. Une fois ce vecteur soustrait, il reste les vitesses3D dues au mouvement des objets. Un certain nombre d’indices particuliers peuvent faciliter unetelle decomposition, comme la profondeur relative, le mouvement commun (mouvement parallelede deux points), le mouvement proportionnel, le type de flux (radial, parallele,...) et la localisationretinienne.
En dehors des indices particuliers, notons qu’il existe aussi differentes contraintes pour l’extrac-tion des parametres 3D du mouvement propre et de la structure. En particulier, l’hypothese derigidite des objets est generalement largement satisfaite pendant le mouvement propre, alors quede nombreux objets se deforment quand ils bougent (Cornilleau-Perez & Droulez 1990). D’ou l’hy-pothese de Droulez (1989) que la metrique utilisee par le systeme visuel pour extraire l’information3D du flux optique est calibree essentiellement pendant le mouvement propre. Les parametres decette metrique sont ensuite ajustes pour minimiser la deformation optique de l’ensemble de l’image.
Parallelement, un autre ”indice” servant a extraire le mouvement du flux optique est celui de lalocalisation des vecteurs de mouvement sur la retine. Un certain nombre de resultats experimentauxsur la perception du mouvement ont donne lieu a la theorie bimodale de la perception. Selon cette

40 Perception du mouvement et mouvement propre
theorie, le mode focal correspondrait a perception du mouvement des objets qui serait basee surretine centrale. Le mode ambiant correspondrait au mouvement propre, et serait plus dependantde la retine peripherique (voir la revue de Dichgans & Brandt 1978). Mais Wolpert (1990) presenteun argument serieux contre la theorie bimodale.
Il teste l’hypothese que le l’information disponible dans les deux secteurs de la ”matrice optique”(optic array)(i.e. centre et peripherie) est utilisee differentiellement en fonction de la tache quel’observateur doit effectuer (p.116). Il realise deux experiences, en croisant les facteurs de ”regionretinienne” stimulee (la stimulation visuelle correspondant toujours a celle que voient la retinecentrale et la retine peripherique pendant pendant un mouvement normal) et de champ visuel.Places dans un simulateur de vol, les sujets doivent determiner s’ils perdent de l’altitude dans la 1re
experience, et s’ils acceleraient dans la deuxieme. Les principaux resultats sont superposables pourles deux taches. Les sujets font moins d’erreurs pour la vue laterale par rapport a la vue frontale,et quand la stimulation est localisee sur la retine centrale par rapport a la retine peripherique.Ces resultats vont a l’encontre de la theorie bimodale. C’est la structure de l’information optiquedisponible dans la matrice optique (flux optique obtenu sur retine peripherique en situation normale)laterale par opposition a celle dans la matrice optique frontale qui est critique, et non l’oppositionretine centrale–retine peripherique.
1.3.1.2 Mouvement relatif et strategies comportementales
En dehors des indices directement relies au mouvement, il existe aussi un certain nombred’”astuces” ou de strategies comportementales possibles pour contourner la difficulte relative al’extraction du mouvement relatif. En particulier, Koenderink (1990) fait remarquer que si l’ons’interesse au 1er ordre de description de l’environnement (surface, forme : voir section 1.2.1.2), ilimporte peu de connaıtre avec precision les mouvements de translation et de rotation (du sujet). Laseule composante du mouvement relatif qui a un effet sur la forme de l’image, c’est la composantede rotation autour de l’axe (vertical) perpendiculaire au regard (voir Figure 1.12).
Par ailleurs, pour resoudre des taches de mouvement relatif apparemment complexes, au lieu defaire une analyse complexe pour determiner la part du mouvement propre et celle du mouvement del’objet dans le flux optique, les sujets peuvent choisir des strategies comportementales tres simples.Par exemple, pour attraper une balle en etant eux-memes en mouvement, les sujets choisissent toutsimplement de maintenir constant l’angle (optique) entre la balle et la main (constant bearing anglestrategy) pendant toute la duree de l’approche (Lenoir et al. 1999). Cette strategie, basee unique-ment sur l’annulation de la vitesse angulaire optique relative entre la main et la balle est suffisanteet efficace pour l’interception. Une telle strategie permet de s’affranchir de calculs complexes sur leflux optique, ainsi que des calculs de predictions.
Pour finir, Berthelon et al. (1998) ont realise une experience avec des sujets sains independantset d’autres dependants du champ (field independant and field dependent ; Shoptaugh & Whitacker1984), qui se distinguent par leur capacite (ou non) a percevoir un objet independamment de soncontexte et a adopter une attitude analytique dans la resolution d’un probleme spatial. Les deuxgroupes ont ete testes sur leur capacite a analyser le mouvement specifique d’un autre vehiculependant une simulation d’arrivee a un carrefour routier. D’une part, pour les deux groupes, latache s’est trouvee facilitee par la presence d’un panneau routier (signe local). Par consequent,dans une situation complexe, les indices locaux (mouvement relatif) sont utilises. D’autre part,

Flux optique et mouvement propre 41
les sujets independants sont bien meilleurs que les sujets dependants, donc plus aptes a extrairel’information pertinente (de mouvement relatif) dans un environnement complexe. Ainsi, au-delades strategies comportementales, l’analyse du mouvement relatif peut dependre de styles perceptifs.
Fig. 1.12 – A Description qualitative du mouvement de translation pure de l’observateur passant a coted’un arbre. Observer le changement de l’orientation mutuelle entre l’objet et l’observateur : la portion visiblede l’objet (demi-ellipse grise puis rouge) varie en fonction de la position du sujet sur sa trajectoire. L’objet etl’observateur subissent un mouvement de rotation identique (ici dans le sens horaire, fleches) par rapport ala ligne qui connecte leurs centres (pointilles). Au moment du depassement, la distance objet-observateur estminimale, et la vitesse de rotation par rapport a la ligne connectrice est maximale. B Mouvement apparentde l’objet autour de l’observateur. L’amplitude de la rotation correspondant a la totalite du mouvement estde 180◦ (d’apres Koenderink, 1990).
1.3.2 Bases neurales de la perception du mouvement
Dans une reflexion theorique, Koenderink (1986) suggere qu’un systeme particulierement ap-proprie pour analyser le flux optique, et notamment en extraire l’information sur la forme 3Ddes objets, serait un systeme sensible aux changements relatifs dans le temps de l’orientations desdifferents details de l’image optique serait .
Chez les mammiferes, il existe trois voies afferentes des informations optiques dans le cerveau : lavoie geniculo-corticale, la voie du tectum (colliculus superieur), et le systeme optique accessoire. Des

42 Perception du mouvement et mouvement propre
neurones dont le champ recepteur (i.e. la partie de la retine qui, lorsqu’elle est stimulee, provoque unereponse du neurone) exhibe les proprietes necessaires en theorie, ont ete decrits extensivement dansle cortex des mammiferes (pour une revue, voir Bremmer et al. 2000), mais aussi dans le systemeoptique accessoire situe dans le tronc cerebral. Ce dernier systeme presente des proprietes tresinteressantes, en particulier celle d’exprimer le mouvement visuel dans une organisation geometriquecommune avec le systeme vestibulaire (Simpson et al. 1988 ; etude chez le lapin ; voir aussi Frost& Wylie 2000). Il existe en effet une similarite marquee entre les directions de mouvement visuelproduites par rotation autour des axes des trois canaux semi-circulaires et la direction prefereede la rotation visuelle des neurones du systeme optique accessoire. Cette coherence geometriquerend ce systeme pontique particulierement adapte a la perception des consequences visuelles dela rotation de la tete. Dans la suite, nous nous concentrerons plus particulierement sur les airescorticales impliquees dans le mouvement visuel, en exposant des donnees obtenues chez le primate.
1.3.2.1 Cortex temporal
Le flux de l’information visuelle dans le cortex est separee en deux voies principales, la voiedorsale qui analyse principalement la localisation et le mouvement visuels, et la voie ventrale quiest impliquee dans l’analyse de la nature et la reconnaissance des objets (Ungerleider & Mishkin1982). Pour l’analyse du mouvement, l’information visuelle transite depuis la retine, passe par lethalamus (noyau genouille lateral dorsal ; LGNd), arrive au cortex visuel primaire (V1, dans le lobeoccipital) qui projette directement dans l’aire MT situee sur la partie posterieure du sillon temporalsuperieur (Ungerleider & Desimone 1986 ; voir Fig. 1.13).
- aire MT
L’aire medio-temporale se distingue de l’aire visuelle primaire en particulier par la sensibilitedes neurones qui s’y trouvent au mouvement complexe. A l’instar des cellules de V1, les neuronesde l’aire MT repondent a la direction et a la vitesse d’un stimulus visuel, ainsi qu’a l’orientationstatique (Maunsell & Van Essen 1983 ; Albright 1984). Par contre, la reponse a l’orientation n’estpas maintenue dans le temps, comme pour les cellules de V1. Par ailleurs, lorsque sont presenteesdeux grilles en mouvement simultane, les neurones de V1 repondent au mouvement de l’une oude l’autre, alors que ceux de MT repondent a la direction de mouvement apparent, c’est-a-dire auvecteur resultant de la somme vectorielle des deux vecteurs de mouvement des grilles (Movshon etal. 1985). Dans le cas d’un mouvement transparent –compose de deux directions de mouvementopposees dans la meme zone du champ visuel–, les neurones de V1 continuent de repondre, alorsque les neurones de MT cessent de decharger lorsqu’un tel stimulus leur est presente, meme sicelui-ci contient un mouvement dans leur direction visuelle preferee (Snowden et al. 1991).

Flux optique et mouvement propre 43
Fig. 1.13 – Principales aires corticales sensibles au mouvement dans la voie corticale visuelle dorsale chezle primate. Sur la vue laterale de l’hemisphere cerebral, le sillon temporal superieur (STS) et le sillon intra-parietal (IP) sont deplies pour mettre en evidence les aires qu’ils contiennent. La representation schematiquesouligne le flux de l’information visuelle depuis la retine, ainsi que les connexions connues entre les differentesaires (connexion forte : ligne pleine ; connexion faible : ligne pointillee). LGN : noyau genicule lateral. V1 :cortex strie. V2 : aire visuelle secondaire. MT : aire medio-temporale. MST : aire medio-temporale superieure.VIP : aire intraparietale ventrale. PM : aire premotrice (d’apres Bremmer et al. 2000 )
Il existe une organisation anatomique topographique dans MT. Les neurones ayant une memedirection preferee de mouvement sont organises en colonnes (Maunsell & Van Essen 1983 ; Albright1984). Il en est de meme pour les vitesses preferees (Lagae et al. 1993). Enfin, les neurones voisinsont leurs champs recepteurs (retiniens) dans des regions voisines de la retine (Tanaka et al. 1993).
Des preuves de l’implication de MT dans la perception du mouvement ont ete amenees par desexperiences comportementales chez le primate. L’inactivation chimique de l’aire MT entraıne uneelevation du seuil de perception du mouvement visuel (Newsome & Pare 1988). De plus, des lesionsfocales de MT entraınent l’impossibilite d’analyser le mouvement visuel (dans la region retiniennecodee par MT) pour maintenir la poursuite oculaire (Newsome et al. 1985).
L’ensemble de ces resultats amene Wurtz et al. (1990) a proposer que l’aire medio-temporaleconcentre le mouvement visuel pour toute activite dependant de celui-ci, que ce soit la generationde mouvement ou bien la perception du mouvement.
- aire MST
Les neurones de l’aire medio-temporale superieure ont des champs recepteurs de plus grandetaille que ceux de MT (deja plus larges que ceux de V1 ; Desimone & Ungerleider 1986), et lagrande majorite de ces neurones repondent a la direction et a la vitesse d’un stimulus visuel (Saitoet al. 1986). L’aire MST est divisee en deux parties, sur la base des representations de la retine etdes proprietes electrophysiologiques. La partie latero-ventrale (MSTl) est plus specialisee dans lemaintien de la poursuite oculaire, alors que la partie dorsale (MSTd), repondant preferentiellement ades stimuli visuels de grande taille, est de ce fait plus susceptible d’etre impliquee dans la perceptiondu mouvement propre (Komatsu & Wurtz 1988 ; Wurtz et al. 1990). De plus, certains neurones sont

44 Perception du mouvement et mouvement propre
sensibles a la disparite binoculaire. Dans ce cas, on trouve une interaction entre la disparite et lasensibilite au mouvement. Par exemple, un neurone sensible au mouvement vers la droite pour unedisparite negative (i.e. en avant du point de fixation), sera sensible au mouvement vers la gaucheau-dela du point de fixation (Roy & Wurtz 1990). Or c’est exactement ce type de mouvement visueloppose en profondeur qui est genere lors d’une rotation de la tete, confirmant le role possible deMSTd dans la perception du mouvement propre.
La selectivite des neurones de MST relative a des stimuli correspondant a des flux optiquesgeneres pendant le mouvement propre a ete decrite initialement par Saito et al. en 1986. Unequantification plus systematique des reponses dans MST aux mouvements plans (dans le planfrontoparallele, correspondant a une translation du sujet, Fig. 1.14 A), radiaux (expansion oucontraction, correspondant a des mouvements de translation vers l’avant ou l’arriere, Fig. 1.14 B),ou circulaires (rotations horaires ou anti-horaires correspondant a une rotation de la tete dans leplan frontoparallele, Fig. 1.14 C) montre qu’il existe plusieurs categories de neurones (Duffy &Wurtz 1991a,b).
Fig. 1.14 – Composantes du flux optique. A Flux optique plan, correspondant successivement a un mou-vement propre simule de translation vers la droite, la gauche, le haut et le bas. B Flux optique radial,correspondant successivement a un mouvement propre vers l’arriere (contraction) et vers l’avant (expan-sion). C Flux optique circulaire, correspondant a une rotation de la tete du sujet vers l’epaule droite et versl’epaule gauche (d’apres Duffy & Wurtz 1991a ).
Certains repondent a une seule composante (plane, radiale ou circulaire), d’autres a deux com-posantes (reponses plano-radiales ou plano-circulaires) et d’autres encores aux trois composantes.Classer les neurones en fonction de leur type de selectivite et de l’intensite de leur reponse estimpossible : il existe un veritable continuum de reponses au flux optique dans MST. Les memesauteurs proposent que les reponses a plusieurs composantes seraient tout simplement le resultat de

Flux optique et mouvement propre 45
la convergence sur le neurone en question de neurones repondant individuellement a chacune descomposantes.
Ces derniers resultats ont souleve la question de savoir si le cerveau analyse les flux optiquescomplexes a l’aide de tels neurones codant pour des mouvements particuliers, et qui formeraient unebase (au sens mathematique) de l’ensemble des mouvements. Pour prendre un exemple simple, unmouvement vers l’avant couple a une rotation de la tete vers la droite serait analyse par deux typesde neurones, le premier repondant exclusivement (ou presque) au mouvement d’expansion, le secondau mouvement plan. La recherche de categories de neurones pouvant servir de base dans l’espacespiral (qui comprend les mouvements d’expansion et les mouvements circulaires) s’est revelee etreun echec , puisqu’au lieu de categories, il existe en fait un continuum de reponses, avec tous lesintermediaires possibles (Fig. 1.15 ; Graziano et al. 1994 ; voir aussi Andersen et al. 2000).
Fig. 1.15 – Reponse d’un neurone dans l’espace spiral de stimulation visuelle A Reponse du neuroneexprimee en coordonnees polaires. L’axe des abscisses est oriente des rotations visuelles dans le sens tri-gonometrique (CCW) vers le sens des aiguilles d’une montre (CW). L’axe des ordonnees est oriente desmouvements visuels de contraction (correspondant a un deplacement du sujet vers l’arriere) vers les mouve-ments d’expansion (correspondant a un deplacement du sujet vers l’avant). Le neurone represente ne repondpas purement a une composante de l’un des axes, mais a une combinaison de contraction et de rotation dansle sens horaire. B Intensite de la reponse du neurone en fonction des divers types de mouvement spiral. Laselectivite du neurone est relativement large (d’apres Graziano et al. 1994 )
La reponse des neurones de MST a un type de mouvement particulier reste selective (a lacomposante du flux optique preferee) si l’on deplace le stimulus visuel sur la retine, et que parconsequent la direction du mouvement sur une region particuliere de la retine est modifiee (Grazianoet al. 1994 ; Duffy 2000). Par contre, l’intensite de la reponse varie en fonction de la positionretinienne du stimulus. En plus de cette relative invariance spatiale, les neurones de MST exhibentaussi une invariance par rapport a la forme des objets dont le mouvement constitue le flux optique(Geesaman & Andersen 1996). Ces deux proprietes d’invariance sont en accord avec l’hypotheseque MST analyse le flux optique pendant le mouvement propre.
Par ailleurs, le role de MST comme analyseur du flux optique se trouve renforce par une etudede Duffy et Wurtz (1997) qui ont montre que les neurones de MST sont modules par la distribution,non plus des directions des vecteurs de mouvements du flux optique, mais des vitesses. Ainsi, cesneurones seraient capables de differencier les flux optiques generes par les deux images de la figure1.16.
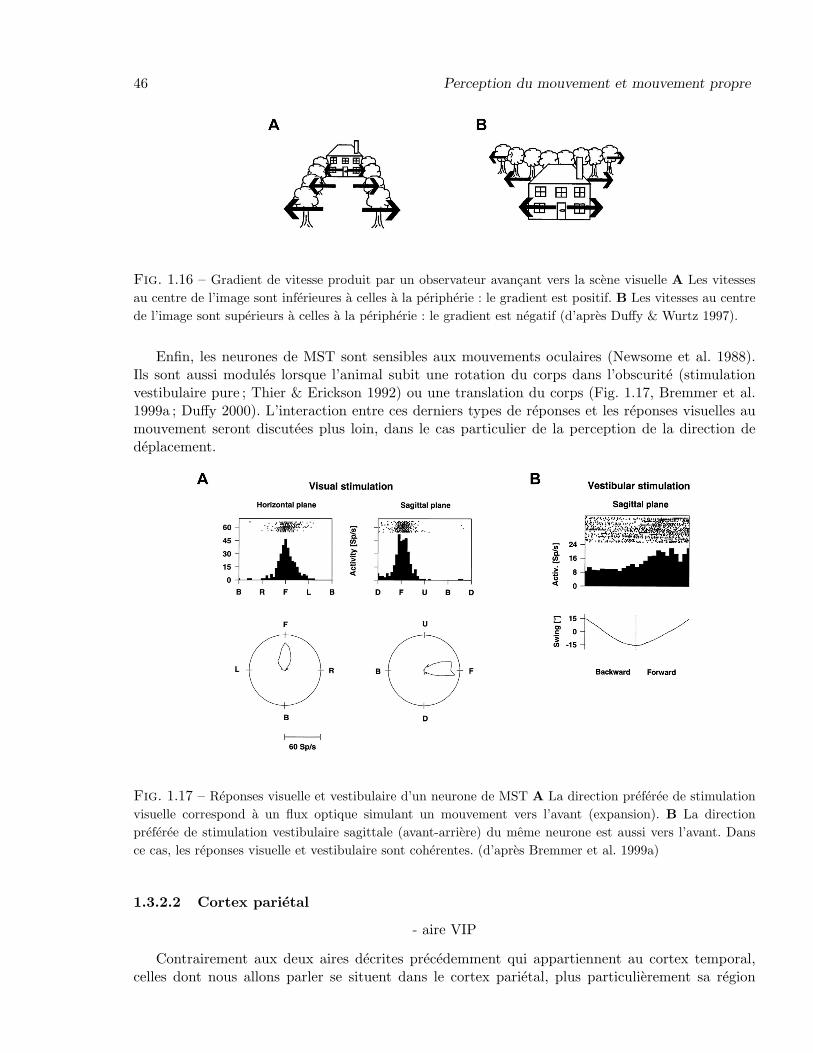
46 Perception du mouvement et mouvement propre
Fig. 1.16 – Gradient de vitesse produit par un observateur avancant vers la scene visuelle A Les vitessesau centre de l’image sont inferieures a celles a la peripherie : le gradient est positif. B Les vitesses au centrede l’image sont superieurs a celles a la peripherie : le gradient est negatif (d’apres Duffy & Wurtz 1997).
Enfin, les neurones de MST sont sensibles aux mouvements oculaires (Newsome et al. 1988).Ils sont aussi modules lorsque l’animal subit une rotation du corps dans l’obscurite (stimulationvestibulaire pure ; Thier & Erickson 1992) ou une translation du corps (Fig. 1.17, Bremmer et al.1999a ; Duffy 2000). L’interaction entre ces derniers types de reponses et les reponses visuelles aumouvement seront discutees plus loin, dans le cas particulier de la perception de la direction dedeplacement.
Fig. 1.17 – Reponses visuelle et vestibulaire d’un neurone de MST A La direction preferee de stimulationvisuelle correspond a un flux optique simulant un mouvement vers l’avant (expansion). B La directionpreferee de stimulation vestibulaire sagittale (avant-arriere) du meme neurone est aussi vers l’avant. Dansce cas, les reponses visuelle et vestibulaire sont coherentes. (d’apres Bremmer et al. 1999a)
1.3.2.2 Cortex parietal
- aire VIP
Contrairement aux deux aires decrites precedemment qui appartiennent au cortex temporal,celles dont nous allons parler se situent dans le cortex parietal, plus particulierement sa region

Flux optique et mouvement propre 47
posterieure. L’aire VIP (aire intraparietale ventrale) a ete definie au depart comme la zone deprojection de MT dans le cortex parietal (Maunsell & Van Essen 1983 ; Ungerleider & Desimone1986). De cette definition, on peut deduire que, a l’instar de ceux de MT, les neurones de VIPdevraient repondre a la direction et a la vitesse d’un stimulus visuel, ce qui a ete confirme (Duhamelet al. 1991 ; Colby et al. 1993). Ces reponses visuelles differencient VIP des aires intraparietalesadjacentes (cf. Chapitre 2.5 ; Colby & Duhamel 1991). Mais VIP recoit aussi des afferences de l’aireMSTd (Boussaoud et al. 1990). Par consequent, on retrouve dans environ 2/3 des neurones deVIP des reponses aux composantes du flux optique tres similaires a celles des neurones de MSTd(Bremmer et al. 1995 ; Schaafsma & Duysens 1996). Une comparaison precise des reponses au fluxoptique dans MSTd et VIP a ete realisee par Schaafsma et Duysens (1996). Si les neurones de VIPrepondent au flux optique plan, radial et circulaire ainsi qu’a des combinaisons de ces deux dernierescomposantes (mouvement spiral), il n’existe pas, en regle generale, de neurones repondant a uneseule composante. Par contre, pour les mouvements radiaux, l’expansion (simulant un mouvementpropre vers l’avant) est plus representee que la contraction, et parmi l’ensemble des composantesdu flux optique, les composantes radiales sont les plus representees (Bremmer et al. 1997).
Sur l’organisation spatiale des champs recepteurs, la reponse des neurones de VIP est globale-ment maintenue lorsque seule une partie du champ recepteur est stimulee, ou bien lorsque la tailledu stimulus est reduite (tout en gardant les directions de mouvement constantes ; Schaafsma & Duy-sens 1996). Ce dernier resultat semble compatible avec l’hypothese d’une organisation en champ devecteurs (vector field hypothesis), qui stipule que toutes les sous-parties du champ recepteur sontequivalentes. L’organisation spatiale des champs recepteurs de MSTd est, quant a elle, compatibleavec l’hypothese du champ de vecteurs pour certains neurones, et compatible avec l’hypothese demosaıque pour d’autres. Cette derniere propose que c’est l’ensemble des sous-parties du champrecepteur (dont chacune est specialisee pour percevoir une direction de mouvement particuliere)qui determine la reponse au stimulus global (Duffy & Wurtz 1991a,b).
Une autre question sur l’organisation spatiale des champs recepteurs est de savoir si les reponsesaux stimuli de flux optique sont determinees simplement par les caracteristiques du champ recepteur(localisation retinienne, forme et surface) et la direction preferee de mouvement dans le plan fron-toparallele, ou bien si ces reponses sont particulieres et ne peuvent pas etres deduites des autrestypes de reponses (Fig. 1.18 ; Bremmer et al. 1997 ; Bremmer et al. 2000). En realite, seul 50 % desneurones ont une reponse au flux optique pouvant etre deduite de la direction preferee du mouve-ment frontoparallele et des caracteristiques du champ recepteur (Fig. 1.18 A). Il est impossible deprevoir sur la meme base la reponse au flux optique de l’autre moitie des neurones enregistres (Fig.1.18 B). Ainsi, les reponses dans VIP aux stimuli optiques simulant des mouvements propres sontbien particulieres : elles ne peuvent pas etre expliquees simplement a partir des caracteristiquesspatiales du champ recepteur et de la sensibilite a la direction du mouvement visuel. Il en est dememe pour les neurones de MST (Lagae et al. 1994).
Par ailleurs, des reponses a la poursuite oculaire (mais aucune reponse aux saccades oculaires)et des reponses a la stimulation tactile ont ete documentees dans VIP (Duhamel et al. 1991). Deplus, des reponses purement vestibulaires ont ete decrites dans notre laboratoire (Bremmer et al.1995 ; Graf et al. 1996 ; Bremmer et al. 1997, 2000). Nous reviendrons sur ces deux derniers typesde reponses sensorielles, qui montrent d’interessantes proprietes spatiales pour les premieres (cf.Chapitre 2.5), et qui ont fait l’objet du travail experimental de notre these pour les secondes (cf.Chapitres 3 et 4).

48 Perception du mouvement et mouvement propre
Fig. 1.18 – Champ recepteur, direction preferee et reponse de neurones de VIP au flux optique. 1re ligne :Reponse a la direction du mouvement (graphique polaire) et cartographie du champ recepteur visuel. 2e
ligne : Direction des vecteurs de mouvement du flux optique surimposee sur le champ recepteur. 3e ligne :Histogramme de reponse du neurone au flux optique. A : Le neurone repond preferentiellement pour unmouvement allant vers la droite (en haut). D’apres les vecteurs de mouvement, on peut deduire que ce neuronerepondra preferentiellement a un mouvement d’expansion (au milieu). Le neurone repond effectivement al’expansion, et est inhibe par la contraction (en bas). Les reponses visuelles et au flux optique de ce neuronesont coherentes. B : La direction preferee de ce neurone est vers la droite (en haut). Il est delicat de prevoirla reponse du neurone d’apres les vecteurs de mouvement, mais on peut supposer que la reponse au fluxoptique circulaire sera faible (au milieu). Contrairement aux predictions, le neurone repond preferentiellementau mouvement de rotation anti-horaire (d’apres Bremmer et al. 2000).
Colby et al. (1993) offrent quelques elements de comparaison entre VIP et MT. Du point devue anatomique, la taille de VIP (5 ∗ 8 mm) environ est de l’ordre de celle de MT. Du point devue physiologique, les reponses de VIP au mouvement simple, et en particulier a la vitesse dumouvement, sont similaire a celles de MT, bien que VIP reponde a des vitesses globalement plusgrandes que les neurones de MT (voir aussi Bremmer et al. 1997). Enfin, au niveau des directionsdu mouvement frontoparallele, celles-ci sont representees uniformement dans VIP, alors qu’il existeune sur-representation de la direction vers le bas dans MT.
Si l’on compare maintenant les reponses au flux optique de VIP et MST, les differences entreles deux aires sont plus faibles et moins nombreuses que leurs ressemblances (Schaafsma & Duysens1996). Il en est de meme pour les reponses a la poursuite oculaire. En ce qui concerne la distanceen profondeur, Duhamel et al. (1991) rapportent qu’environ 20 % des neurones de VIP repondentpreferentiellement a des stimuli visuels presentes a moins de 20cm du visage. Certains neuronesde MST repondent en fonction de la disparite (Wurtz et al. 1990), c’est-a-dire a la distance dustimulus par rapport au point de fixation. Aucune etude systematique, comparant les deux aires,

Flux optique et mouvement propre 49
n’a ete realisee sur la distance en profondeur. Par contre, VIP differe nettement de MST, en ceque ses reponses sont modules par le comportement, ce qui est une caracteristique plus generaledu cortex parietal (Duhamel et al. 1991). VIP est aussi plus multimodale que MST, en particulier,aucune reponse a la stimulation tactile n’a ete trouvee dans MST, alors que la majorite des neuronesde VIP possedent de telles reponses (Duhamel et al. 1991,1998). Enfin, VIP differe de MST pourcertaines proprietes spatiales en relation avec la position des yeux (cf. le point suivant) et aussiparce qu’elle projette directement dans une region specifique du cortex premoteur, specifique dumouvement de la tete et du cou (Fig. 1.13 ; Rizzolatti et al. 1998).
1.3.2.3 Autres aires corticales
Les aires MST et VIP projettent toutes deux sur l’aire parietale 7A (Fig. 1.13). Ainsi, onretrouve dans 7A des reponses au flux optique, qui pour certaines sont plus intenses pour unedirection du flux (l’expansion est preferee a la contraction), et d’autres sont plus fortes pour unecomposante du flux optique (flux radial prefere au flux circulaire ; Sakata et al. 1985 ; Read & Siegel1997 ; Siegel & Read 1997). En outre, comme dans MST, les reponses au flux optique sont moduleespar la position du stimulus sur la retine, et par celle des yeux dans l’orbite (Bremmer et al. 2000 ;voir 1.3.3).
Par ailleurs, l’analyse de l’information visuelle n’est pas entierement separee entre les deuxgrandes voies corticales. L’aire STPa (aire supero-temporale polysensorielle anterieure) recoit desafferences des aires MST et 7A de la voie dorsale, et de l’aire TEO de la voie ventrale (Boussaoud etal. 1990 ; Distler et al. 1993). Des etudes electrophysiologiques (Heitanen & Perret 1996a,b) utilisantdes objets reels, ont montre que les neurones de STPa repondent uniquement au mouvement visuelprovoque par des objets. Ainsi, au contraire de VIP et MST, les neurones de STPa ne repondentpas au mouvement propre.
Chez l’homme, l’analyse du mouvement est aussi separee en deux voies corticales (etude PET,Zeki et al. 1991). L’equivalent de MT (V5) du primate est active par le mouvement visuel coherent,mais pas par le mouvement incoherent (Cheng et al. 1995). Par contre, les stimuli de flux optiquen’activent pas l’equivalent de MT, mais ils activent l’aire V3 dans l’hemisphere droit, ainsi que laregion occipito-temporale ventrale bilateralement. Ces resultats montrent une analyse specifique duflux optique dans les lobes parietaux chez l’homme (De Jong et al. 1994).
1.3.2.4 Voie dorsale et cortex vestibulaire
En plus du mouvement visuel, le mouvement propre induit aussi une sensation vestibulaire (voir1.4). L’aire VIP est connectee reciproquement aux aires MST et 7a, et recoit des entrees du PIVC.VIP est donc connectee directement au reseau cortical vestibulaire (Lewis & Van Essen 2000a,b ;cf. 1.1.3).
Des modulations de la reponse en fonction de la direction de stimulation vestibulaire ont etedecrites dans MST (Thier & Erickson 1992), et dans VIP (Graf et al. 1995 ; Bremmer et al. 1997,2000). Les neurones de VIP montrent un type d’interaction visuo-vestibulaire particulier puisque lesdirections preferees des stimulations visuelles et vestibulaires sont identiques (generant un conflitsensoriel). Seuls un tiers des neurones du cortex vestibulaire sont aussi ”non-complementaires” (cf.1.1.3). Cette interaction visuo-vestibulaire dans l’aire VIP a fait l’objet de la premiere partie denotre travail experimental (Chapitre 3).

50 Perception du mouvement et mouvement propre
1.3.3 Flux optique et direction de deplacement (vers ou va-t-on ?)
1.3.3.1 Perception de la direction du deplacement (ou heading)
Lors de la definition des composantes principales du flux optique, nous avons vu en particulierque le flux radial correspond a un mouvement propre vers l’avant (expansion) ou vers l’arriere(contraction ; Fig. 1.14B). Dans ce cas, il existe une singularite dans la distribution des vecteurs demouvement, qui est le point unique ou le vecteur de mouvement est nul, autrement dit ou la vitesselocale est nulle (comparer par exemple Fig. 1.14 A et B). C’est cette singularite du flux optique,encore appelee foyer d’expansion, qui determine la direction de deplacement de l’observateur. Aucuncadre de reference n’est necessaire pour evaluer la direction de deplacement. C’est l’ensemble desobjets visuels qui sert lui-meme de referentiel au sein duquel la direction de deplacement est specifieevisuellement. Par ailleurs, rappelons que le mouvement de translation a un effet sur le flux optiqueinversement proportionnel a la distance entre l’observateur et les objets qui generent le flux (cf.1.2.1.1). Par contre, les mouvements de rotation generent un flux optique uniforme sur la retine, dontl’amplitude est independante de la distance. La direction et l’amplitude du flux genere dependent enparticulier de l’axe de rotation de l’oeil et de sa vitesse de rotation. En consequence, les mouvementsdes yeux, parce qu’ils generent un flux optique qui s’ajoute au flux provenant du deplacement del’observateur, entraınent un deplacement du foyer d’expansion de la retine, voire sa disparition(Warren & Hannon 1990 ; Lappe & Hoffmann 2000). Ainsi, les mouvements des yeux font apparaıtreune ambiguıte en rompant la relation directe entre la position du foyer d’expansion sur la retineet la direction de deplacement, ce qui complique la determination de cette derniere a partir duflux optique (Fig. 1.19). Plus generalement, le flux optique sur la retine depend du deplacement dusujet, de la direction du regard par rapport a celle du deplacement, et de la rotation des yeux.
- temps d’apparition et precision de la perception de la direction de deplacement
La totalite des etudes psychophysiques sur la perception de la direction du mouvement propreutilisent des mouvements propres simules, realisees a l’aide stimuli visuels presentes aux sujetssur un ecran digital. Contrairement a la perception du mouvement propre induite visuellement(vection ; cf. 1.4.1) qui met plusieurs secondes a se mettre en place, la perception de la direction dedeplacement apparaıt en environ 400ms (Crowell et al. 1990 ; Hooge et al. 1999), indiquant que lesmecanismes sous-jacents sont differents pour les deux types de perception.
Warren et al. (1988) ont montre que la direction de mouvement propre par rapport a une ciblede reference peut etre determinee a 1-2◦ d’angle visuel de precision. Cette precision est considereecomme suffisante pour pouvoir eviter des obstacles dans les situations de la vie courante (Cuttinget al. 1992). Par ailleurs, la precision de jugement de la direction de deplacement varie peu enfonction du temps d’exposition au stimulus ou en fonction de la structure ou de la densite del’environnement (Warren et al. 1988).
Par contre, elle depend de la localisation du stimulus visuel sur la retine –elle diminue enfonction de l’excentricite–, et de la taille du stimulus optique. En particulier, si l’on offre au sujetune ”vision tunnel”, l’amplitude de l’erreur de jugement augmente tres fortement, surtout si lefoyer d’expansion est hors du champ de vision (Crowell & Banks 1993).
D’autre part, les sujets humains arrivent tres bien a differencier, sur la base du flux optique,une trajectoire courbe d’une trajectoire rectiligne (Turano & Wang 1994). Le seuil de detectionde la courbure dans la trajectoire simulee est plus de trois fois inferieur au seuil de discriminationde la direction de deplacement. Ceci suggere l’existence de mecanismes specialises pour detecterla deviation d’une trajectoire rectiligne, qui pourraient par exemple etre bases sur la perceptiond’asymetries de la distribution des vecteurs de mouvement du flux optique (Warren et al. 1991a).

Flux optique et mouvement propre 51
Enfin, les effets sur la perception de la direction de deplacement des objets visuels se deplacantindependamment sont tres reduits, ce qui suggere que le mouvement d’objet est largement supprimelors de l’analyse de la direction du mouvement propre (Van den Berg 2000, revue).
- le probleme du mouvement des yeux
La rotation de l’oeil provoque une modification voire la destruction du foyer d’expansion (Fig.1.19). Lorsque la position retinienne du foyer d’expansion est modifiee, si les sujets determinent ladirection de deplacement a partir de cette position, ils font une erreur systematique. D’ou l’ideed’etudier, toujours au moyen de mouvements simules, l’aptitude des sujets humains a decomposerle flux optique retinien pour en retirer la composante produite par la rotation des yeux. Ainsi,en particulier pour evaluer la contribution de signaux oculomoteurs extra retiniens, de nombreuxtravaux ont ete realises en comparant la perception de la direction de deplacement (ou l’aptitudea la decomposition du flux optique) avec soit des mouvements de poursuite oculaire, soit desmouvements de poursuite simules et l’oeil stationnaire (Warren & Hannon 1988, les premiers autiliser ce type de paradigme).
Les erreurs de jugement de la direction de deplacement sont toujours de faible amplitude (2-4◦)lorsque les sujets realisent le mouvement de poursuite, que le deplacement du regard soit provoquepar un mouvement des yeux ou de la tete (Warren & Hannon 1990, poursuite oculaire ; Crowellet al. 1998, mouvement de la tete). Lors de mouvements simules, si le flux simule un mouvementpar rapport au plan du sol (stimuli analogues a ceux de la Figure 1.19), on n’observe pas dedegradation de la precision de decomposition du flux optique (Warren & Hannon 1990). Par contre,la performance se degrade lorsque le stimulus visuel n’inclut pas d’indices de profondeur, ou lorsquela vitesse de rotation simulee du regard depasse 1.5◦/s. Cette vitesse seuil correspond aux vitessesmoyennes habituelles de poursuite lors d’un deplacement reel (marche a pied). La decompositiondu flux optique est donc possible meme avec un mouvement simule (Warren & Hannon 1988, 1990).Avec des rotations simulees du regard a des vitesses superieures, les resultats sont beaucoup plusvariables (voir aussi Banks et al. 1996). Cependant, la decomposition est plus robuste lorsque laperturbation du flux optique est provoquee par un mouvement actif que lorsqu’il est simule, ce quiimplique l’influence d’un signal extra-retinien (Van den Berg 1992).
Trois types (au moins) d’indices sensoriels contribuent au signal extra-retinien qui garantit laprecision de la decomposition du flux optique (Crowell et al. 1998). Un signal vestibulaire canalaire,un signal de proprioception nuccale et une copie efferente des mouvements des yeux et de la tete.Les signaux vestibulaire ou de proprioception nuccale seuls ne suffisent pas pour compenser. Maislorsqu’ils sont couples, dans une tache de poursuite passive oeil-tete ou lors de la stabilisation duregard sur le tronc en mouvement, ils suffisent a certains sujets pour compenser la deviation dufoyer d’expansion sur la retine due au mouvement (Crowell et al. 1998).
- strategies comportementales pour resoudre le probleme de la rotation
Pour evaluer la direction du deplacement seule, que la trajectoire simulee soit rectiligne oucourbe, c’est la direction et non la vitesse (amplitude) des vecteurs de mouvement qui importe(Warren et al. 1991b). En effet, aucune augmentation de l’erreur de jugement n’est constatee lorsqueles vitesses des stimuli optiques sont modifiees aleatoirement, alors qu’il apparaıt une serieusedegradation de la performance si ce sont les directions qui sont changees.

52 Perception du mouvement et mouvement propre
Fig. 1.19 – Flux optique sur la retine, direction du regard et rotation des yeux. A La direction du regard(symbolisee par le cercle) est parallele a la direction de deplacement (DD ; croix). Le flux optique est un fluxd’expansion pur. Le foyer d’expansion est centre sur la retine. B L’observateur fixe un point sur l’horizon,decale a gauche par rapport a la DD. Ceci ne provoque pas de mouvement des yeux. Le foyer d’expansionindique toujours la DD, mais il n’est plus centre sur la retine. C Le regard de l’observateur est dirige surun point localise sur le sol, en avant et a gauche de la DD. Ceci provoque (i) un deplacement de l’imageretinienne (de l’horizon) vers le haut, et (ii) un mouvement des yeux pour maintenir le regard sur le pointfixe qui est en mouvement par rapport a l’observateur. Le flux optique se compose d’un flux radial du audeplacement, auquel s’ajoute un flux circulaire du a la rotation des yeux. La resultante est un flux spiral,dont la singularite est liee a la fixation du point sur le sol et n’est plus un indicateur de la DD. D Tout enavancant, l’observateur suit du regard un objet qui se deplace sur l’horizon (vers la gauche). L’effet de lapoursuite oculaire est un deplacement de l’image retinienne dans le sens oppose au mouvement des yeux (versla droite). Le flux optique resultant de cette combinaison d’un flux radial (deplacement de l’observateur) etd’un flux plan (rotation des yeux) donne l’impression que l’observateur se deplace sur une trajectoire courbe.Il n’y a plus de singularite du flux optique. (d’apres Lappe & Hoffmann 2000)

Flux optique et mouvement propre 53
Par contre, lorsqu’il s’agit de decomposer le flux optique, direction et vitesse sont toutes deuximportantes (Van den Berg, observations non publiees, Van den Berg 2000).
Van den Berg (2000) a compare les performances des sujets decomposant le flux optique pourdeterminer la direction de leur deplacement simule dans un certain nombre d’etudes, et il a montrequ’il existe des ”observateurs retiniens” –qui repondent a la position du foyer d’expansion sur laretine, et donc ne compensent pas l’effet de la rotation des yeux–, des ”observateurs de la directiondu deplacement” –qui eux compensent parfaitement–, ainsi que tous les intermediaires entre cesdeux comportements extremes.
Une strategie simple pour resoudre le probleme de decomposition est de fixer un point dansl’environnement, ce qui reduit la recherche de la direction du deplacement a l’intersection entre lemeridien de poursuite oculaire et l’horizon lorsque l’observateur se deplace parallelement au sol.D’autre part, fixer un point dans l’environnement fait dependre le decalage entre la direction dedeplacement et la direction du regard (H) de la vitesse de rotation de l’oeil (R) :
H = sin−1( RT/d)
pour un mouvement (de l’observateur) en translation (T ) et une distance de fixation (d)constantes (Van den Berg 2000). Une telle strategie peut effectivement etre utilisee par des su-jets humains (Banks et al. 1996).
Par ailleurs, le plan du sol fournit des informations sur la profondeur, independantes du fluxretinien, et qui peuvent egalement contribuer a la decomposition. Notons toutefois que si ces in-dices peuvent etre utilises, en particulier en situation experimentale, ils n’apportent pas d’avantagesignificatif lorsque le bruit visuel est faible (grand rapport signal sur bruit ; Van den Berg, 2000).Par contre, le fait que differents points alignes en profondeur ne restent pas sur une meme lignesauf s’ils sont alignes avec la direction de deplacement, constitue un indice tres fiable de la directiondu deplacement (Cutting et al. 1992).
- Perception de la direction de deplacement et mouvement propre
D’une part, le flux optique influe sur le mouvement propre, puisqu’il provoque des ajustementsposturaux (cf. 1.4.1). D’autre part, le fait de marcher introduit des oscillations horizontales etverticales qui s’ajoutent au flux optique, donc modifient celui-ci. Mais ces oscillations ne modifientpas la perception de la direction de deplacement (Cutting et al. 1992).
Par ailleurs, des sujets dont le champ visuel est devie lateralement par le port de prismes optiquesmarchent vers la cible visuelle en suivant une trajectoire courbe, alors que la strategie consistant amaintenir la cible au foyer d’expansion predit, malgre l’effet des primes, une trajectoire rectiligne(Rushton et al. 1998). Ce dernier resultat, ainsi que d’autres resultats experimentaux parallelesamenent Van den Berg (2000) a proposer que la direction de deplacement percue a travers l’analysedu flux optique n’est peut-etre simplement qu’un des elements de l’information visuelle utilisee pourguider nos actions dans l’environnement.
1.3.3.2 Bases physiologiques de la perception de la direction du deplacement
- aire MST
Les neurones de MST modulent leur reponse a un stimulus de flux optique en fonction de laposition de la singularite du flux (foyer d’expansion pour les stimuli radiaux). Plus de 90% des

54 Perception du mouvement et mouvement propre
neurones ont une preference pour la position de la singularite dans une certaine region de la retine,et dans toute la population de ces neurones, l’ensemble de la retine est representee (Duffy & Wurtz1995 ; Duffy 2000). Cependant, la selectivite a la position de la singularite est relativement large(modelisee par une sigmoıde 2D), la direction de deplacement ne peut donc pas etre deduite de lareponse d’un seul neurone mais a partir de l’ensemble de la population (Lappe et al. 1996 ; Lappe2000). Lappe et al. (1996) ont montre qu’un modele utilisant les moindres carres pouvait, a partirde la population de neurones de MST qu’ils ont enregistres, determiner la position (retinienne) dela singularite du flux optique (donc de la direction de deplacement) a 4.3◦ de precision.
De plus, les neurones de MST peuvent compenser la perturbation du flux optique provoqueepar un mouvement de poursuite oculaire (Bradley et al. 1996), et pourraient ainsi etre a la basede la decomposition du flux optique (voir section precedente). En effet, pendant la poursuite, onobserve un glissement des courbes de selectivite a la position de la singularite du flux optique sur laretine dans la direction du deplacement de la singularite, deplacement provoque par le mouvementdes yeux (Fig. 1.20).
Ainsi, lorsque la poursuite s’ajoute a un flux radial d’expansion, la courbe de selectivite estdecalee dans la direction du mouvement de poursuite (Fig. 1.20A), lorsque la poursuite s’ajoute aun flux radial de contraction, la courbe de selectivite est decalee dans la direction opposee a cellede la poursuite (Fig. 1.20B), et enfin lorsque cette derniere s’ajoute a un flux circulaire, la courbede selectivite est decalee selon un axe orthogonal a celui du mouvement oculaire (Fig. 1.20C). Parconsequent, la courbe de selectivite est toujours decalee de maniere a compenser les effets optiquesdu mouvement oculaire ; la selectivite de ces neurones a la position de la singularite du flux optiqueest exprimee en coordonnees allocentriques (de l’ecran) et non en coordonnees retinocentriques.Remarquons toutefois que seule une partie des neurones compensent de la sorte, et que l’amplitudede la compensation est toujours inferieure a celle du deplacement de la singularite provoque par lapoursuite. En outre, les mesures de compensation sont realises dans la direction preferee de reponsea la poursuite. Aucune donnee n’est disponible pour un neurone dans un ensemble plus large dedirection de mouvements oculaires.
Par ailleurs, dans les resultats de Bradley et al. (1996), les neurones compensent uniquementlorsque la deviation de la singularite est provoquee par des mouvements (de poursuite) actifs.Lorsque ceux-ci sont simplement simules, aucune compensation n’est observee. Cependant, Brem-mer et al. (1998) ont montre que, si les stimuli optiques simulent un deplacement au-dessus du solcombine avec un mouvement de poursuite (i.e. stimuli identiques a ceux de la Figure 1.19), certainsneurones de MST (ainsi que d’autres de VIP) peuvent compenser. Lorsque la vitesse du mouvementde poursuite realise par les animaux augmente, le deplacement de la courbe de selectivite augmenteparallelement, ce qui constitue un indice supplementaire indiquant que la compensation observeeest bien celle du mouvement des yeux (Shenoy et al. 1998).
Enfin, des microstimulations de MST peuvent affecter la perception de la direction de deplacement(Britten & Van Wezel 1998). L’ensemble de ces resultats conforte le role de MST dans la perceptiondu mouvement propre, et particulierement dans la determination de la direction de deplacement.Plus generalement, les neurones de MST pourraient etre impliques dans la perception de la stabilitede l’espace pendant le mouvement propre (Bradley et al. 1996).

Flux optique et mouvement propre 55
Fig. 1.20 – Deplacement de la singularite avec la rotation de l’oeil pour differentes composantes du fluxoptique. Dans chaque cas, le vecteur de mouvement oppose a celui genere par la poursuite est indique enrouge. A Flux optique radial d’expansion. Lors de la combinaison avec un mouvement de poursuite versla gauche, le foyer d’expansion est deplace aussi vers la gauche, dans la direction identique a celle de lapoursuite. B Flux optique radial de contraction. L’addition d’un mouvement de poursuite vers la gauchedeplace le foyer de contraction vers la droite, dans la direction opposee a celle de la poursuite. C Fluxoptique circulaire. L’addition d’un mouvement de poursuite vers la gauche deplace la singularite sur un axeorthogonal a celui de la poursuite, en l’occurrence vers le haut. (d’apres Bradley et al. 1996).

56 Perception du mouvement et mouvement propre
Shenoy et al. (1999) ont compare le deplacement de la courbe de selectivite a la position de lasingularite du flux optique pendant des mouvements de poursuite et de suppression du VOR (lesanimaux subissent une rotation de tout le corps et doivent maintenir la fixation sur une cible visuellesynchrone avec la rotation, ce qui provoque un mouvement de la tete dans l’espace, les yeux gardantune position orbitale constante). Leurs resultats indiquent que la compensation des neurones deMST est similaire dans les deux cas, c’est-a-dire que le mouvement du regard soit produit par unerotation de l’oeil ou par une rotation de la tete seule. Ces resultats constituent une preuve de plus del’implication de MST dans l’analyse du flux optique pour determiner la direction de deplacement.Par ailleurs, cette compensation basee sur des signaux vestibulaires est interessante, parce que lesexperiences de psychophysique montrent que les sujets humains ne peuvent pas compenser de lasorte (Crowell et al. 1998). Shenoy et al. (1999) proposent que la sous-population de neurones quicompensent a l’aide des signaux vestibulaires ne participe pas a la perception de la direction dedeplacement. Or, on pourrait aussi proposer que la perception de la direction de deplacement differequelque peu entre le macaque et l’homme. Il n’existe pas d’experiences de psychophysique chez lesinge dans la tache de suppression du VOR pour trancher.
- aire VIP
Nous insisterons surtout sur les differences des reponses de l’aire intraparietale ventrale aveccelles de MST. Comme dans MST, lorsqu’on varie la position de la singularite du flux optique, onobserve une modulation de l’intensite de la reponse des neurones de VIP (Bremmer et al. 2000), etl’ensemble des reponses de la population permet de determiner la position de la singularite sur laretine (Duhamel et al. 1997a).
Mais contrairement aux neurones de MST, de nombreux neurones de VIP codent l’informationvisuelle en coordonnees non retinocentriques explicitement (Duhamel et al. 1997b ; voir aussi2.6), ce qui permet en particulier de coder les autres informations sensorielles de ces neuronesdans le meme systeme de coordonnees, les coordonnees craniocentriques. A l’oppose, les champsrecepteurs des neurones de MST, de 7a et plus generalement des aires du cortex parietal posterieursont retinotopiques, avec simplement une modulation de l’intensite de decharge en fonction de laposition des yeux, ce qui est considere comme un codage non retinocentrique implicite (theoriedes gain fields, qui propose que la modulation de la reponse des neurones par la position de l’oeilagisse comme un gain, qui permettrait alors, au niveau de la population, de retrouver un codagecraniocentrique ; Andersen et al. 1985 ; Andersen & Zipser 1988 ; Andersen et al. 2000 ; Bremmer etal. 1997b ; Squatrito & Maioli 1996). Le codage non retinocentrique explicite de VIP place cette aireen premier lieu dans l’analyse du mouvement propre, de la direction du deplacement, et peut-etreegalement dans l’analyse du mouvement des objets.
1.4 Fusion multisensorielle et emergence de la perception du mou-vement propre
La perception du mouvement propre n’est pas dependante d’un seul sens, mais elle emergede l’interaction de plusieurs modalites sensorielles. Une des raisons de cette situation reside dansl’incompletude (ambiguıte) sensorielle, autrement dit, l’incapacite d’un seul sens a donner uneimage fiable du mouvement propre dans toutes les situations. Le systeme vestibulaire est ineffi-cace pendant un deplacement a vitesse constante, et le systeme visuel seul ne peut pas vraimentdeterminer l’origine (exterieure : mouvement d’objet, ou propre : mouvement du sujet) du mouve-ment qu’il percoit. L’emergence de la perception du mouvement propre sera revue a travers trois

Fusion multisensorielle et emergence de la perception du mouvement propre 57
problematiques. D’abord l’interaction visuo-vestibulaire, ensuite a travers l’influence de la proprio-ception, et enfin en considerant une approche plus theorique.
1.4.1 Le cas de la vection
Un des moyens d’apprehender, au sein du systeme nerveux, les mecanismes de fusion multisen-sorielle qui sous-tendent la perception du mouvement propre, est l’etude des illusions perceptives.Une des plus classiques dans le domaine est la vection, ou la sensation de mouvement propre in-duite visuellement (Dichgans & Brandt 1978 ; Mach 1875 ; von Helmholtz 1896). Chacun aura puexperimenter la vection lorsque, par exemple, un train voisin demarre et que l’on a l’impression,pendant un moment, que c’est celui dans lequel on se trouve qui se met en mouvement. Si le mou-vement induit par la vision est rotatoire on parlera de vection circulaire, si c’est un mouvement detranslation on parlera de vection lineaire (Dichgans & Brandt 1978), et s’il s’agit d’un mouvementsuivant une trajectoire courbe, on parlera de vection curvilineaire (Sauvan & Bonnet 1995). Enparticulier, Dichgans et Brandt (1978) partent de l’hypothese que l’illusion de mouvement proprede la vection provient du meme mecanisme physiologique que celui qui maintient la perception dumouvement reel du corps a vitesse constante. Un tel mecanisme de maintien de la perception dumouvement a vitesse constante est rendu necessaire en raison des deficiences du systeme vestibulairependant un tel mouvement (voir Fig.1.2C) : par sa nature de capteur d’acceleration, le systeme ves-tibulaire seul ne peut pas faire la difference entre l’immobilite et un mouvement a vitesse constante.A la base du mecanisme de maintien de la perception du mouvement, ces auteurs postulent que lesafferences visuelles et vestibulaires sont fusionnees tres tot, de telle sorte que lorsque les yeux sontouverts, la vision compense directement les deficiences du message vestibulaire.
L’induction d’une sensation de mouvement propre n’est pas l’appanage de la seule vision. Eneffet, il existe aussi une vection circulaire audiocinetique (von Stein 1910, in Dichgans & Brandt1978), ou provoquee par le mouvement passif de l’epaule ou vection arthrocinetique (Brandt etal. 1977), ou encore provoque par la stimulation de la surface du corps ou vection haptocinetique(Dichgans & Brandt 1978).
Le dispositif permettant de stimuler la sensation de vection est relativement simple. Pour la vec-tion circulaire, il est constitue par une chaise tournante disposee au centre d’un cylindre, lui-mememobile independamment de la chaise. Les parois du cylindres sont peintes de bandes verticales,noires et blanches en alternance, provoquant un stimulus optocinetique sur l’ensemble du champvisuel lorsque le cylindre est mis en mouvement. Dans ce cas, la rotation du cylindre engendreune sensation de mouvement propre apparent, de sens oppose a celui du cylindre, et qui est iden-tique a celle provoquee par une acceleration lente reelle suivie d’un mouvement a vitesse constante(Dichgans & Brandt 1978).
Les principales caracteristiques de la vection sont resumees sur la figure 1.21, dans le cas de lavection circulaire. La sensation de vection n’est pas immediate, mais elle apparaıt progressivement,au fur et a mesure que diminue la vitesse apparente du stimulus visuel. La vection a un seuil devitesse, entre 90 et 120◦/s, en deca duquel elle est proportionnelle a la vitesse du stimulus visuel.Enfin, la vection depend de la stimulation de la peripherie visuelle, en excentricite comme en profon-deur (Fig. 1.21, colonnes 4 et 6 ; Dichgans & Brandt 1978). Toutefois, l’opposition centre-peripherieest remise en question par Post (1989) qui montre qu’il s’agirait plutot d’une consequence desdifferences quantitatives d’aire stimulee dans les deux conditions. Par ailleurs, Howard et Heck-man (1989) ont montre qu’il est difficile de dissocier l’effet ”centre-peripherie” de l’effet ”premierplan-arriere plan” en montrant leur equivalence pour stimuler la vection.
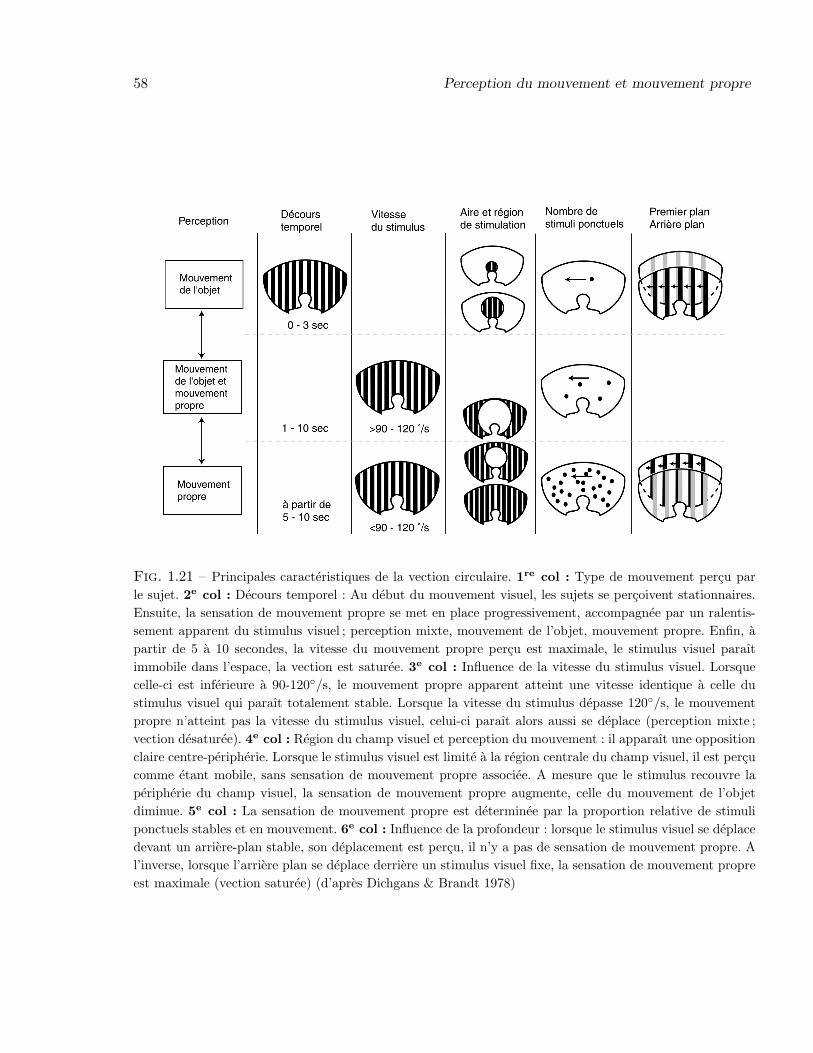
58 Perception du mouvement et mouvement propre
Fig. 1.21 – Principales caracteristiques de la vection circulaire. 1re col : Type de mouvement percu parle sujet. 2e col : Decours temporel : Au debut du mouvement visuel, les sujets se percoivent stationnaires.Ensuite, la sensation de mouvement propre se met en place progressivement, accompagnee par un ralentis-sement apparent du stimulus visuel ; perception mixte, mouvement de l’objet, mouvement propre. Enfin, apartir de 5 a 10 secondes, la vitesse du mouvement propre percu est maximale, le stimulus visuel paraıtimmobile dans l’espace, la vection est saturee. 3e col : Influence de la vitesse du stimulus visuel. Lorsquecelle-ci est inferieure a 90-120◦/s, le mouvement propre apparent atteint une vitesse identique a celle dustimulus visuel qui paraıt totalement stable. Lorsque la vitesse du stimulus depasse 120◦/s, le mouvementpropre n’atteint pas la vitesse du stimulus visuel, celui-ci paraıt alors aussi se deplace (perception mixte ;vection desaturee). 4e col : Region du champ visuel et perception du mouvement : il apparaıt une oppositionclaire centre-peripherie. Lorsque le stimulus visuel est limite a la region centrale du champ visuel, il est percucomme etant mobile, sans sensation de mouvement propre associee. A mesure que le stimulus recouvre laperipherie du champ visuel, la sensation de mouvement propre augmente, celle du mouvement de l’objetdiminue. 5e col : La sensation de mouvement propre est determinee par la proportion relative de stimuliponctuels stables et en mouvement. 6e col : Influence de la profondeur : lorsque le stimulus visuel se deplacedevant un arriere-plan stable, son deplacement est percu, il n’y a pas de sensation de mouvement propre. Al’inverse, lorsque l’arriere plan se deplace derriere un stimulus visuel fixe, la sensation de mouvement propreest maximale (vection saturee) (d’apres Dichgans & Brandt 1978)

Fusion multisensorielle et emergence de la perception du mouvement propre 59
Par ailleurs, puisque le phenomene de vection resulte d’une interaction visuo-vestibulaire, sonapparition est facilitee par une diminution du conflit visuo-vestibulaire potentiel. En particulier, lalatence de la vection est diminuee par une acceleration du sujet dans la direction du mouvementpropre percu (Dichgans & Brandt 1978). Pour la meme raison, la latence de la vection diminueavec l’augmentation du seuil de sensibilite vestibulaire des sujets –un sujet moins sensible aura unconflit visuo-vestibulaire moins aigu– (Lepecq et al. 1999). Enfin, la vection apparaıt plus tot etest plus forte chez des patients bilabyrinthectomises, que chez des patients hemilabyrinthectomises,eux-memes plus sensibles que les sujets sains (Jonhson et al. 1999).
La possibilite de provoquer la sensation d’un mouvement propre vers l’avant par stimulationvisuelle (vection lineaire ; Berthoz et al. 1975 ; in DB1978) prouve que l’activite des canaux semi-circulaire n’est pas la seul a pouvoir etre simulee, celle des otolithes peut l’etre aussi (Dichgans &Brandt 1978). Par ailleurs, il est egalement possible de provoquer des sensations de mouvementpropres dans les plans frontoparallele et sagittal (roll vection et pitch vection). Dans ce cas, lasensation de mouvement continu est accompagnee par celle d’un deplacement limite, probablementen raison de l’apparition d’un conflit entre la vision et les gravicepteurs somatosensoriels et otoli-thiques (Dichgans & Brandt 1978). D’autres aspects de la vection, en relation avec l’espace, serontdiscutes dans la section 2.1.1.1 qui traite de l’interaction mouvement-espace.
Dichgans et Brandt (1978) supposent que l’environnement est percu comme stable pendant lavection parce que l’information qui arrive au cortex visuel est neutralisee par un signal collateralprovenant du systeme vestibulaire. Un certain nombre de correlats neuronaux de la vection cir-culaire, et plus generalement de l’interaction visuo-vestibulaire dans le systeme nerveux, semblentleur donner raison.
Tout d’abord, il est connu depuis longtemps qu’une grande majorite de neurones vestibulairesenregistres chez le primate ont egalement des reponses aux stimuli optocinetiques, des les noyauxvestibulaires (Henn et al. 1974 ; in DB78), et aussi dans le flocculus cerebelleux (Waespe & Henn1977 ; Waespe et al. 1981), dans le thalamus vestibulaire (Buttner & Henn 1976), et dans les cor-tex vestibulaires (aire PIVC : Grusser et al. 1990 ; aire 2v : Buttner & Buettner 1978, Buttner &Henn 1981). Pendant la rotation sinusoıdale a basse frequence de l’animal a la lumiere, il apparaıtdans les noyaux vestibulaires, un renforcement de la reponse vestibulaire par rapport a celle obte-nue a l’obscurite, ainsi qu’une abolition de l’avance en phase caracteristique des reponses a bassefrequence (Henn et al. 1974). Tout se passe comme si la stimulation visuelle ”calait” la reponse ves-tibulaire aux parametres du mouvement. Il existe donc une certaine complementarite des reponsesneuronales visuelles et vestibulaires puisque, comme dans le cas de la vection au niveau percep-tif, le systeme visuel pallie les deficiences du systeme vestibulaire a basse frequence (Dichgans &Brandt 1978). Les voies anatomiques par lesquelles le signal visuel parvient au premier relais desafferences vestibulaires ne sont toujours pas clairement elucidees aujourd’hui. Ceci dit, la latencede 7 secondes pour les reponses visuelles dans les noyaux vestibulaires rapportee par Waespe etHenn (1977) laisse presager, par son importance, une voie multisynaptique.
Chez l’homme, des interactions visuo-vestibulaires ont ete documentees au niveau cortical.Contrairement a ce que l’on aurait pu penser au depart, la stimulation vestibulaire entraıne unediminution de l’activite dans le cortex visuel (Wenzel et al. 1996), et cette inhibition semble etrespecifique pour la direction du mouvement –elle n’est presente que lors de stimulations ou la di-rection (et le sens) des stimulations visuelle et vestibulaire sont identiques– (Loose et al. 1999).Par ailleurs, des patients ayant une hemianopsie homonyme ne percoivent pas la vection, ni nepresentent d’ajustements posturaux consequents a une stimulation visuelle etendue (Straube &Brandt 1987), ce qui souligne le role clef du cortex visuel dans la vection. A l’inverse de l’inhibitionvestibulaire sur le systeme visuel, il existe aussi une inhibition du cortex vestibulaire par la stimu-

60 Perception du mouvement et mouvement propre
lation visuelle. Brandt et al. (1998) ont induit la vection circulaire frontoparallele chez des sujetssains pendant l’acquisition d’images PET, et ont pu montrer une baisse de l’activite cerebrale dansl’insula posterieure (l’equivalent chez l’homme du cortex vestibulaire PIVC chez le singe), ainsique dans le thalamus. L’ensemble de ces resultats a conduit Brandt et al. (1998) a postuler quel’interaction visuo-vestibulaire, en particulier dans le cas de la perception du mouvement propre,est basee sur une inhibition reciproque au niveau cortical. Un tel mecanisme presenterait l’avantagepouvoir reduire les conflits sensoriels, en permettant notamment de faire basculer la dominanced’un sens a l’autre en fonction des besoins comportementaux. Elle permettrait aussi, dans le casde la vection, d’assurer une certaine stabilite de cette derniere en evitant que de legeres perturba-tions vestibulaires (comme des mouvements de la tete pendant un trajet en voiture) ne viennentconstamment interferer avec la sensation de mouvement propre (Brandt et al. 1998 ; Brandt 1999).
Dans le cas de la vection lineaire, il est possible que les mecanismes mis en jeu soient differents.Une etude de magneto-encephalographie (Nishiike et al. 2001) montre que pendant l’accelerationinduite (par l’acceleration du stimulus visuel), il apparaıt une activation de la region parieto-temporale corticale correspondant au sulcus temporal superieur et a l’insula posterieure (aire MSTet cortex vestibulaire respectivement). Il reste neanmoins possible que la difference de ces resultatsavec ceux de Brandt et al. (1998) provienne de l’utilisation de stimuli accelerant et non a vitesseconstante.
1.4.2 Plasticite et interaction multisensorielle
Lorsque des informations sensorielles de nature differentes concourent a l’emergence d’une per-ception unique, ces informations doivent etre exprimees dans le meme systeme de coordonnees, ouen tous cas interagir de maniere efficace pour produire une perception stable.
En fonction de la tache comportementale, il est parfois possible, voire necessaire, de se basersur l’information provenant d’un sens plutot qu’un autre. Mergner & Becker (1990) ont etudie laplasticite de l’interaction visuo-vestibulaire, basee soit sur une reference visuelle (visual impression :mouvement d’objet) ou soit sur une reference vestibulaire (bodily feeling : mouvement propre), dansle cadre psychophysique de la vection circulaire. L’influence tres forte des instructions donneesau sujet (fixer ou non le stimulus visuel, par exemple) sur la perception eprouvee par la suite(l’induction de la vection circulaire) suggere que les deux signaux referents (vestibulaire et visuel)sont present a des niveaux cognitifs eleves, et que la dominance (en terme de signal referent) de l’unou de l’autre sens est modifiable par la volonte. Par contre, les patients vestibulaires ne peuvent passupprimer la vection (Mergner et al. 2000). Les auteurs proposent qu’en temps normal, la perceptiondu mouvement propre est definie en reference a l’environnement visuel considere a priori commeetant stable, et que si l’environnement paraıt bouger (mouvement d’objet), le signal (referent) visuelest supprime et la perception est alors basee sur l’information vestibulaire (Mergner et al. 2000). Leport de prismes met en evidence que ce type d’interactions visuo-vestibulaires est plastique (Kohler1956).
Par ailleurs, Mergner et Becker (1990) ont egalement cherche a preciser l’influence de differentssignaux sensoriels sur la perception du mouvement propre. Pour ce faire, ils ont mis en conflit lesdifferents indices sensoriels –indice retinien (objet visuel), indice oculomoteur (suppression ou nondes mouvements oculaires), indice vestibulaire et proprioception nuccale (mouvement tete et corpsou corps seul)– , et demande a leurs sujets d’evaluer leur deplacement dans l’espace. En comparantindividuellement les convergences sensorielles suivantes, retinienne-oculomotrice, retinienne-nuccaleet retinienne-canalaire, les auteurs ont montre qu’en fait, il existe une sommation lineaire des effetsdes differents messages sensoriels sur la perception du mouvement propre. Rappelons en passant

Fusion multisensorielle et emergence de la perception du mouvement propre 61
que Crowell et al. (1998) ont montre que des copies efferentes des mouvements des yeux et de latete, ainsi que des signaux vestibulaires et la proprioception nuccale, contribuent a la perceptionde la direction du deplacement de l’observateur a partir des informations du flux optique.
Plus recemment, Mergner et al. (1993) ont observe que les effets sur la perception du mouvementpropre de la proprioception liee aux membres inferieurs sont analogues a ceux de la proprioceptionnuccale (Heimbrandt et al. 1990). Plus precisement, lors d’une stimulation vestibulo-proprioceptivecombinee (par exemple deux rotations concomitantes de la tete et du tronc dans l’obscurite), la per-ception du deplacement de l’egocentre dans l’espace varie avec les deux informations afferentes. Parcontre, lorsque l’une d’elles est absente, comme dans le cas des patients vestibulaires, la perceptiondu mouvement propre est erronee. Ces patients considerent que leur tronc est stable dans l’espacedans tous les cas (Heimbrandt et al. 1990). Les auteurs proposent ainsi un modele plus general d’in-teraction visuo-proprioceptive, ou les differentes informations sensorielles se completeraient pouraboutir a la perception du mouvement propre (Mergner et al. 1993). Autrement dit, partant d’unsignal (vestibulaire) de mouvement de la tete dans l’espace, les differentes informations proprio-ceptives sont ensuite combinees pour calculer successivement la position (et le mouvement) desparties du corps dans l’espace. Mergner et al. (1997) rajoutent au modele un principe de migra-tion d’information du bas vers le haut, en proposant que le support corporel puisse interagir avec(voire corriger) le percept d’espace ”physique” base au depart sur la seule information vestibu-laire : ainsi, les afferences proprioceptives sont impliquees dans une veritable boucle de retroactionmultisensorielle qui aboutit a la perception du mouvement propre que nous ressentons.
1.4.3 Considerations theoriques : mouvement propre et mouvement d’objets
L’interaction visuo-vestibulaire qui preside a l’emergence de la perception du mouvement proprepose en meme temps la question de la perception du mouvement des objets. En effet, mouvementpropre et mouvement d’objet sont lies, parce que tous deux sont percus par le systeme visuel maisque sur la base de la vision seule, il est impossible de les distinguer.
Il existe deux theories opposees sur la perception du mouvement propre et du mouvement desobjets (Wertheim 1990). La theorie de la perception directe dit que, par defaut, l’environnement eststable. En consequence, le flux optique correspond uniquement au mouvement propre. A l’oppose,la theorie inferentielle repose sur l’existence d’une comparaison d’un signal extraretinien (copieefferente) au flux optique, qui serait a l’origine de la perception du mouvement propre. Or, cesdeux theories ont quelques difficultes avec l’apparition de la vection circulaire.
Un sujet place au centre d’un cylindre compose de bandes verticales que l’on fait tourner selonun axe vertical, percoit dans un premier temps le mouvement du cylindre seul, puis graduellement,apparaıt une sensation de tourner dans le sens contraire du mouvement du cylindre, le cylindresemblant ralentir puis s’arreter, le sujet atteignant alors la sensation de vitesse maximale (egale maisopposee a la vitesse du cylindre initialement percue). A ce moment, la vection circulaire est ditesaturee (Dichgans & Brandt 1978 ; cf. 1.4.1). La theorie de perception directe prevoit la sensation devection, puisque par defaut l’environnement est stable et que le flux optique genere par la rotation ducylindre ne contient pas d’invariant susceptible d’indiquer une rotation de l’environnement. Maisl’apparition graduelle de cette sensation, avec au depart une perception du seul mouvement ducylindre –qui suppose l’existence d’un invariant specifiant la rotation de l’environnement– pose unserieux probleme. A l’oppose, la theorie inferentielle explique pourquoi le mouvement du cylindreest percu. Le flux optique genere par le mouvement du cylindre est compare a un signal extra-retinien nul (ou n’indiquant pas de mouvement propre), d’ou la perception du mouvement del’environnement. Par contre, cette theorie n’explique par l’apparition de la vection circulaire.
Wertheim (1990) postule l’existence d’un signal de reference qui serait compose de plusieurs

62 Perception du mouvement et mouvement propre
elements, notamment une copie efferente des signaux des muscles extra-oculaires (von Holst &Mittelstaedt 1950), un signal vestibulaire (mouvement de la tete), et un signal de flux optique (ouoptocinetique), le tout serait envoye ensuite au comparateur –comparant le signal de reference ausignal visuel– dont la sortie specifierait le mouvement propre. La composante de flux optique dusignal de reference implique un principe d’autoreference, puisque le signal retinien participe a lafois au signal visuel et au signal de reference qui lui est compare. Ainsi, la vitesse percue d’un objetdepend de l’amplitude de la difference entre les signaux retinien et referent, moins le seuil (pluspetite difference perceptible : celle-ci augmente avec l’amplitude des signaux a comparer).
Le principe d’autoreference permet de resoudre le probleme theorique pose par la perception dumouvement des objets et de la stationarite dans l’installation et la saturation de la vection circulaire.Au debut du mouvement du cylindre autour de l’observateur, un signal retinien de mouvement del’objet (environnement) est genere immediatement et est compare au signal de reference qui estbien plus faible. S’ensuit une perception du mouvement du cylindre. Au cours de l’apparition dela vection, le signal de reference est graduellement amplifie par sa composante de flux optique. Lesignal de reference augmente parallelement a l’augmentation de la sensation de mouvement propre.La difference entre les signaux retinien et de reference diminue, et avec elle la perception de larotation du cylindre. Enfin, lorsque la vection est saturee, la sensation de mouvement propre estmaximale, le signal de reference atteint une amplitude comparable au signal retinien, le cylindresemble immobile.
Parmi les differentes predictions du modele propose, nous ne retiendrons que les plus impor-tantes dans le cadre de notre sujet. –La vitesse percue d’un objet depend de la difference entre lavitesse de cet objet et le seuil de detection de la vitesse dans la direction du mouvement. Ainsi,dans une tache de poursuite oculaire, la vitesse du stimulus est sous-estimee par les sujets parceque le seuil est augmente dans la direction du mouvement de poursuite. De plus, l’arret rapided’une poursuite oculaire a grande vitesse donne l’illusion que le stimulus poursuivi accelere. Cesphenomenes perceptifs sont, dans le modele, une consequence directe de la participation des mou-vements des yeux (copie efferente) au signal de reference, ce qui fait dependre le seuil de perceptionde la difference entre les signaux retinien et de reference de la vitesse de l’oeil. –L’influence de lacomposante vestibulaire du signal de reference a ete verifiee pendant le VOR, ou elle est dans le sensoppose a la composante de mouvement des yeux (phases lentes). Les sujets places sur une chaisetournante dans le noir, a l’interieur d’un cylindre peint de barres verticales, subissent une rotationsinusoıde d’axe vertical. Les murs du cylindres sont illumines brievement (400ms) au moment dupic de vitesse de la trajectoire des sujets. Ceux-ci, soit libres de bouger les yeux soit en train desupprimer leur VOR en fixant une diode solitaire de la chaise tournante, devaient estimer la vitessede leur propre mouvement. Conformement a la prediction du modele, la vitesse apparente augmentelors de la suppression du VOR, c’est-a-dire lorsque le signal de mouvement des yeux, de directionopposee a celle du mouvement de la tete, est supprime. –Les stimuli de grande taille (surround)sont frequemment percus comme etant stationnaires par rapports a des objets en mouvement pluspetits. Le modele de Wertheim propose une explication alternative a celle qui voudrait que, pardefinition, les objets plus grands servent de cadres de reference (theorie de perception directe). Cesstimuli larges generent une grande composante optocinetique du signal de reference, d’ou un seuilimportant de detection de leur mouvement, et par consequent une tendance accrue a les considerercomme stables. –Au cours du mouvement propre, le signal de reference est augmente par la com-posante vestibulaire, ce qui entraıne (dans le modele) une augmentation du seuil de perception dela difference entre les signaux retinien et de reference, d’ou une diminution de la perception dumouvement. Ceci explique un certain nombre d’observations psychophysiques comme par exemplel’augmentation du seuil de perception du mouvement d’un objet et la diminution de sa vitesseapparente lors du mouvement propre (cf. 1.5.2). Ainsi, Wertheim souligne que les composantes

Caracteristiques generales du mouvement propre et couplage perception-action 63
vestibulaire et optocinetique du signal de reference ont une fonction ”ecologique” importante : ellesinterfacent la perception du mouvement propre avec la perception du mouvement des objets ou dela stationarite.
En ce qui concerne les bases neurales possibles du signal de reference du modele, l’auteur indiqueles neurones enregistres dans les noyaux vestibulaires (Berthoz et al. 1981), dans le cervelet (Suzuki& Keller 1988) et dans le cortex vestibulaire (Buttner & Buettner 1978 ; Straube & Brandt 1987)qui repondent au deplacement physique de la retine dans l’espace independamment de sa cause(mouvement des yeux, de la tete, ou des deux), et qui, pour certains, ont aussi des reponses visuelles.Plus recemment, Shenoy et al. (1999) ont montre que les neurones de l’aire supero-temporale medialedorsale (MSTd) ajustent la structure spatiale de leurs champs recepteurs visuels en fonction desmouvements du regard, qu’ils soient provoques par un mouvement des yeux ou de la tete. Parailleurs, notons que le modele propose peut etre generalise a d’autres modalites sensorielles ou lemouvement d’un objet est percu et le capteur de mouvement se deplace dans l’espace, comme parexemple la somatosensibilite manuelle ou digitale.
1.5 Caracteristiques generales du mouvement propre et couplageperception-action
Lorsque l’on s’interesse aux mouvements produits par un sujet, une distinction s’impose d’embleeentre d’une part les mouvements actifs –realises directement par tout ou partie du corps du sujet–,et les mouvements passifs, subits par le sujet dans un vehicule ou un support mobile. Les mou-vements actifs sont eux-memes separes en mouvements actifs volontaires (locomotion, orientationdu regard) et en mouvements actifs involontaires ou reflexes (cf. 1.1.2.3 reflexe vestibulo-oculaire,ou plus generalement l’ensemble des reflexes qui visent a stabiliser le sujet –reflexes posturaux– oul’image du monde sur la retine –reflexes de stabilisation du regard). Par leur fonction, les mouve-ments reflexes interagissent avec les mouvements volontaires et sont fortement contraints par cesderniers. Les mouvements volontaires sont, quant a eux, gouvernes par les contraintes de but del’action, et par le temps de l’action (interaction, reponse a une attaque de congeneres, etc.). Dansce qui suit, nous exposerons tout d’abord un certain nombre de caracteristiques specifiques desmouvements actifs, notamment des mouvements des capteurs essentiels a la perception du mouve-ment propre (a savoir les capteurs visuels et vestibulaires). Ensuite nous verrons comment l’etudedu mouvement peut aussi renseigner sur le cadre spatial des mouvements des capteurs, et enfin,nous verrons que l’influence du mouvement sur la perception sensorielle n’est pas identique, suivantqu’il est genere activement ou subi passivement.
1.5.1 Le mouvement 3D : problemes specifiques
1.5.1.1 Description du mouvement et formulation mathematique
Tout mouvement tridimensionnel est decrit par six parametres, trois parametres de translation(en general selon les axes avant-arriere, droite-gauche, et haut-bas) et trois parametres de rotation(Fig. 1.22A). Rappelons que la structure du capteur vestibulaire indique qu’il percoit trois degresde translations (les otolithes) et trois degres de rotation (definis par l’orientation des canaux semi-circulaires, cf. 1.1.1). Les rotations posent un probleme specifique, du fait de la non-commutativitedu groupe des rotations 3D qui entraıne que le resultat final de la combinaison de plusieurs rotationsdepende de l’ordre sequentiel des mouvements (Tweed 1997 ; Fig. 1.22B).

64 Perception du mouvement et mouvement propre
Fig. 1.22 – A Les trois axes des rotations 3D. L’axe vertical (haut) est l’axe des rotations horizontales(droite-gauche). L’axe inter-aural (a droite) est l’axe des rotations de ”tangage” (mouvement haut-bas). Letroisieme axe, qui pointe vers l’avant de la figure, est celui des rotations de ”roulis” (mouvement epauledroite-epaule gauche). B Non-commutativite des rotations et VOR. A partir d’une position initiale, le sujeteffectue deux rotations successives, mais dont l’ordre est inverse. Dans le 1er cas (en haut), il realise d’abordune rotation d’un quart de tour vers la droite, puis une rotation d’un quart de tour vers le bas. En positionfinale, la tete est dirigee vers le bas. Dans le 2e cas, c’est d’abord un quart de tour vers le bas, puis un quartde tour vers la droite. En position finale, la tete est orientee horizontalement et dirigee vers le lecteur. Lesdeux positions finales de la tete sont donc tres differentes : le resultat final d’une serie de rotations dependde l’ordre des rotations effectuees. Parallelement, notons qu’un reflexe vestibulo-oculaire parfait, maintenantconstante la direction du regard au cours des deux series de rotations, aboutit a deux position des yeux(dans la tete) distinctes (regard dirige vers la gauche dans le 1er cas, et regard dirige vers le haut dans le 2e
d’apres Tweed 1997).

Caracteristiques generales du mouvement propre et couplage perception-action 65
Partant de la non-commutativite des rotations, Tweed (1997) se pose le probleme de l’integrateurneuronal pour le VOR. Depuis le modele de Robinson (1975), le VOR se concoit en partant dusignal vestibulaire de vitesse de la tete (cf. 1.1.2), multiplie par -1 –parce que pour etre compen-sateurs, les mouvements des yeux sont a l’oppose des mouvements de la tete– et ensuite integrepour aboutir a un signal de position des yeux qui est envoye aux motoneurones. Or, l’integrationmathematique est un operateur commutatif, donc le resultat d’une integration ne depend pas del’ordre sequentiel des mouvements integres, ce qui n’est pas le cas pour le VOR (Fig. 1.22B).
L’exemple precedent pose le probleme des representations mathematiques utilisees pour decrireune propriete donnee du cerveau, en l’occurrence le VOR. Manifestement, le modele de Robinson(1975) qui repose sur une integration de la vitesse angulaire de la tete ne decrit pas correctement leVOR. Tweed (1997) souligne l’interet de choisir des representations mathematiques dont certainesau moins des proprietes sont partagees par les representations dans le cerveau. Dans cette optique,les quaternions sont particulierement pertinents. –Ces objets mathematiques ont ete definis audepart comme sous ensemble de R4 ayant une structure de corps avec l’addition et la multiplicationmatricielle. Cette derniere n’etant pas commutative, le corps des quaternions est un corps noncommutatif. Par ailleurs, on peut aussi voir ce corps a quatre dimensions comme l’espace ”plat” leplus petit qui contient l’espace des rotations 3D –qui est un espace tridimensionnel ”courbe” (unerotation de 360◦ est identique a une rotation de 0◦). Par construction, les quaternions possedentdonc des proprietes tres interessantes pour l’etude des mouvements oculaires 3D. En particulier, dufait de la non commutativite de la multiplication des quaternions, la combinaison des rotations 3Ddevient une operation directe, tout comme la transformation de la vitesse angulaire en variationde position (Tweed 1997 ; Tweed et al. 1990)–. Ainsi, pour le VOR, Crawford & Vilis (1991) ontmontre que, si l’on decrit les mouvements des yeux avec des quaternions, le modele du VOR decritalors correctement le comportement biologique observe (Tweed 1997b).
1.5.1.2 Lois deduites de l’etude des mouvements des yeux
L’etude des mouvements des yeux reste encore actuellement tres marquee par les travaux dedeux precurseurs. Le premier, Frans Cornelis Donders, publie en 1848 un article dont la conclusionest connue aujourd’hui comme la loi de Donders : l’orientation tri-dimensionnelle de l’oeil pour unedirection donnee du regard est toujours la meme, quel que soit le chemin emprunte par l’oeil poury arriver (Henn 1997 ; Fig. 1.23A). Une consequence importante de cette loi est que l’ensemble despositions 3D de l’oeil est limite a une surface bi-dimensionnelle (Vilis 1997). Remarquons cependantque la loi de Donders pose un probleme, lie a la non-commutativite des rotations (Tweed & Vilis1990)). Si l’on imagine, comme sur la figure precedente (1.22 B), que l’oeil subisse une rotationhorizontale puis une rotation verticale, et que l’on inverse ensuite l’ordre de ces rotations a partird’une meme position initiale, la direction du regard obtenue en empruntant ces deux chemins seraidentique, mais la position 3D de l’oeil differente (en l’occurrence, les composantes horizontale etverticale seront identiques, mais la composante torsionnelle differente ; cf. Fig. 1.23 A). La solutionpour eviter cette violation de la loi de Donders est la loi issue des travaux du deuxieme precurseur,Listing. Johannes Benedict Listing publie en 1853 un traite d’optique physiologique ou il definitsa future loi : la tete droite et immobile, toutes les positions (3D) des yeux peuvent etre decritespar un axe de rotation unique a partir d’une position de reference, et tous ces axes font partied’un plan unique (Fig.1.23B ; Henn 1997 ; Tweed et al. 1990). De cette maniere, Listing offre unequantification de la loi de Donders, en specifiant la composante de torsion oculaire.

66 Perception du mouvement et mouvement propre
Fig. 1.23 – Les lois de l’oculomotricite. A Illustration des rotations oculaires et de la loi de Donders. Unmouvement des yeux est generalement constitue d’une composante de rotation horizontale (h), verticale (v)et torsionnelle (t). Selon la loi de Donders, si l’oeil est dans la position no.2 pour une certaine direction duregard, il sera toujours dans la position 2 lorsque la direction du regard sera identique. B La loi de Listing :pour toute direction du regard, la position (3D) de l’oeil est obtenue par une rotation unique a partir de laposition primaire (au centre). Les axes de rotation (en noir) correspondant a toutes les directions du regardsont contenus dans un plan, le plan de Listing (en bleu) (d’apres Tweed et al. 1990).
Des mesures experimentales effectuees dans les conditions indiquees par Listing qui ont eterealisees chez le singe (Haslwanter et al. 1992 ; Henn 1997) et chez l’homme (Suzuki et al. 1994),confirment que les position oculaires sont confinees dans une surface plane dont l’epaisseur –quirepresente la deviation par rapport au plan ideal– n’est que de 1◦ d’angle.
Plusieurs interpretations des lois de Donders et de Listing sont proposees par Hepp et al. (1997).Concernant la premiere, les auteurs notent que la loi de Donders ”simplifie” la vision, puisque dansle cas contraire, l’image d’un objet sur la fovea dependrait de la trajectoire de l’oeil, rendant sonidentification plus difficile. A propos de la loi de Listing, Hepp et al. (1997) proposent plusieursinterpretations. La premiere, l’interpretation visuelle, dit que si la loi de Donders s’applique pourun champ oculomoteur circulaire, la loi de Listing minimise la composante de torsion oculaire.L’interpretation motrice est la demonstration qu’un ensemble de positions oculaires reliees par dessaccades correspondant aux trajets les plus courts dans l’espace des rotations 3D (geodesiques) sanspour autant violer la loi de Donders, est un plan de Listing. Enfin, la troisieme, l’interpretationvisuomotrice est la suivante : si une cible est foveee par une saccade satisfaisant la loi de Listing,alors il y a une relation approximativement lineaire entre le vecteur saccadique –entendu commedifference entre les positions oculaires 3D initiale et finale– et l’erreur retinienne –entre la direc-tion de la cible et le centre de la retine–. La linearite de la relation entre le vecteur moteur etl’erreur visuelle reste valable pour des excentricites inferieures a 30◦, ce qui est le cas pour lamajorite des mouvements habituels. Cette propriete particuliere, engendree par la loi de Listing,servirait de base a la mise a jour de l’image visuelle au cours de la saccade oculaire, proprieteindispensable pour avoir une perception stable de l’espace environnant. (cf. 2.1.2). Pour les auteurscette interpretation visuomotrice, a caractere ”ecologique” est particulierement attrayante, et elleconstitue une justification suffisante pour la loi de Listing chez le singe et l’homme.
Par ailleurs, la loi de Listing, et par consequent celle de Donders, restent valables pour d’autres

Caracteristiques generales du mouvement propre et couplage perception-action 67
types de mouvements oculaires, avec toutefois quelques reserves. Pour les mouvements de poursuiteoculaire, les positions oculaires 3D pendant la poursuite sont contenues dans un plan chez le singe(Haslwanter et al. 1991) et chez l’homme (Tweed et al. 1992 ; Misslich 1997). Par contre, on observedes deviations par rapport a la loi de Listing lorsque la cible n’est plus ponctuelle, ou lors deschangement de sens dans une tache de poursuite d’une cible sur une trajectoire circulaire (Tweed etal. 1992). En ce qui concerne les mouvements de vergence, les plan de listing de chaque oeil tournenten fonction du degre de convergence : la loi de Listing au sens strict est donc violee (Nakayma 1983 ;Mikhael et al. 1995). Par contre, pour un degre de convergence donne, les positions oculaires suiventla loi de Listing, qui n’est donc violee que partiellement (Mok et al. 1992).
Un autre aspect du mouvement tridimensionnel est le probleme de la redondance : pour unedirection donnee du regard, une infinite de positions oculaires 3D est possible en theorie. Leslois de Donders et de Listing sont une solution a ce probleme de degres de liberte surnumeraires,puisqu’elles font correspondre une position oculaire 3D unique a chaque direction du regard. En fait,il semble que cette solution soit utilisee plus generalement par le systeme nerveux. En particulier,il a ete montre que les lois de Donders et de Listing s’appliquent aussi aux mouvements de latete (Straumann et al. 1991) et des membres (Straumann et al. 1991 ; Gielen et al. 1997). Pources derniers, le probleme de la redondance se pose aussi en raison de l’implication de plusieursarticulations independantes. Hormis la resolution du probleme de redondance qui assure ainsi unecertaine reproductibilite des mouvements, un autre interet ”ecologique” de ces lois est de faciliterla coordination des mouvements de differents segments corporels (oeil, tete, bras) en restreignantles vecteurs de rotation 3D a des plans voisins (Straumann et al. 1991). Le probleme du mouvementen 3D est ainsi ramene physiologiquement a un probleme 2D.
1.5.1.3 Mouvements des yeux et de la tete
Les capteurs essentiels au mouvement propre –l’oeil et le vestibule– etant localises dans la tete,il a paru necessaire de presenter quelques travaux relatifs aux mouvements 3D de la tete et leurinteraction avec les mouvements des yeux.
L’etude des mouvements de la tete realises pendant l’orientation vers une serie de cibles visuellesa montre que les positions 3D de la tete devient substantiellement de la loi de Listing (chez leprimate : Guitton & Crawford 1994). Tout se passe comme si, au lieu d’avoir des axes de rotationsindependants comme dans le cas des mouvements oculaires, la tete se comportait comme un cardande Fick (Fig. 1.24 ; Tweed et al. 1995). Ce comportement en cardan de Fick presente l’avantaged’induire des rotations verticales de la tete les plus courtes possibles, minimisant ainsi l’energienecessaire au maintien de la tete contre la gravite (Vilis 1997). Une autre raison possible est que cetarrangement permettrait d’eviter l’accumulation de torsion de la tete au cours de ses mouvements(Tweed et al. 1992). En fait, la tete suit la loi de Donders, mais moins strictement que l’oeil. Cetargument contredit l’hypothese de la commande motrice commune pour l’oeil et la tete formulee al’origine par Robinson et Zee (1981).
Crawford et al. (1997) proposent que le signal de commande existe, a la fois simultanementet de maniere intermittente, dans de multiples systemes de coordonnees motrices, en l’occurrenceles systemes de Listing et de Fick. Par ailleurs, il est possible que la difference observee entre lesmouvements des yeux et de la tete soit moins importante qu’elle n’y paraıt a priori. Une experienced’adaptation menee sur le primate portant un masque restreignant son champ visuel a la visionperifoveale a montre que lors de l’orientation vers des cibles visuelles, les mouvements de la tete serapprochaient de la loi de Listing (Crawford & Guitton 1995).

68 Perception du mouvement et mouvement propre
Fig. 1.24 – Cinematique des mouvements de la tete. Les mesures des positions angulaires 3D de la tetemontrent que celle-ci se comporte comme un cardan de Fick. Dans ce systeme, l’axe qui sous-tend les rotationsverticales (v) est situe dans un dispositif solidaire de l’axe des rotations horizontales (h). Par consequent,pour une position donnee de la tete, la rotation verticale est minimale (d’apres Tweed et al. 1995).
L’orientation statique a une influence sur le plan de Listing et la ”surface de Fick” (l’equivalentdu plan de Listing pour la tete). Lorsque la tete est inclinee dans le plan frontoparallele, l’oeil subitun mouvement de torsion dans le sens oppose (Haslwanter et al. 1992), ce qui amene le plan deListing a tourner par rapport a la tete. Parallelement, lorsque le corps est incline, la tete contre-balance, ce qui entraıne une rotation de la surface de Fick (Misslisch et al. 1994). Mais l’amplitudede ces mouvements de contre-rotation ne depassant pas 10% de l’amplitude du mouvement initial,Vilis (1997) conclut que le plan de Listing et la surface de Fick peuvent etre considerees comme fixespar rapport a la tete et au corps respectivement. D’une maniere generale, le systeme oculomoteurminimise l’excentricite radiale (torsionnelle), et le systeme cephalo-moteur minimise l’excentriciteverticale. Par ailleurs, si l’on mesure la position 3D des yeux dans l’espace, on s’apercoit qu’ellene se reduit pas exactement a une surface de Fick (Radau et al. 1994). En d’autres termes, lessujets ne choisissent pas des positions oculaires 3D identiques pour une direction du regard donneeatteinte par des mouvements combines de l’oeil et de la tete, mais des positions voisines : cecisemble exclure un role perceptif de la loi de Listing (Vilis 1997).
Le devenir des lois de l’oculomotricite pendant le mouvement de la tete a ete etudie sur lereflexe vestibulo-oculaire. Crawford et Vilis (1991) qui ont fait tourner des singes dans les troisplans (horizontal, sagittal et frontoparallele) et mesure les positions oculaires 3D a partir desquellesont ete calculees les vitesses angulaires 3D. Plusieurs systemes de coordonnees ont ete essayes pourexprimer leurs mesures. Les meilleurs resultats ayant ete obtenus avec le systeme de coordonneesde Listing, les auteurs suggerent que les coordonnees utilisees par le cerveau pour le controle duVOR et les saccades sont identiques. La vitesse des phases lentes du VOR est plus faible selonl’axe de torsion, ce qui entraıne une legere mais systematique deviation de celle-ci par rapport a lavitesse de la tete. Ainsi, l’oeil ne contre-balance pas exactement le mouvement de la tete pendantles phases lentes. Cependant, la vitesse des phases lentes reste suffisamment proche de celle de latete (qui reagit selon un cardan de Fick) pour devier significativement de la loi de Listing. Cettedeviation est ensuite compensee par les phases rapides, qui, en ramenant la torsion oculaire a zero,devient l’oeil du plan de Listing a l’oppose de la deviation engendree par les phases lentes. Ainsi,les phases rapides du VOR se comportent comme un operateur de Listing, dans le sens ou ellesminimisent la torsion oculaire (Misslisch 1997). Des resultats similaires ont ete obtenus sur le VOR

Caracteristiques generales du mouvement propre et couplage perception-action 69
chez l’homme (Tweed et al. 1994).
1.5.1.4 Vers un cadre de reference otholitique ?
Deux types d’influences du signal sensoriel otolithique sur la position oculaire 3D ont etedecrites. La premiere, l’influence otolithique statique a ete explicitee dans la section precedentea propos de la stabilite du plan de Listing (cf. Haslwanter et al. 1992 ; Crawford & Vilis 1991) :pour une orientation donnee de la tete dans le plan frontoparallele, une certaine quantite de tor-sion est ajoutee aux positions 3D de l’oeil au cours de la saccade et de la fixation. La deuxiemeinfluence otolithique (l’influence dynamique), est mise en evidence pendant des rotations lentes dusujet d’experience autour d’un axe legerement decale de la verticale. Ce dispositif experimentalpermet de faire varier continument la direction du vecteur gravitaire par rapport a la tete, enstimulant exclusivement les otolithes. Angelaki et Hess (1996) ont decrit chez l’animal un reflexeotolitho-oculaire d’inclinaison associe a une modulation des composantes verticale et torsionnellede la position oculaire 3D en phase avec la position de la tete par rapport a la gravite. Il existedonc une dependance des coordonnees oculomotrices (de Listing) de l’orientation de la tete dansl’espace 3D.
Ces derniers resultats ont fait naıtre l’idee que cette influence du systeme otolithique pour-rait etre a la base d’un veritable cadre de reference fixe par rapport a l’espace. Un paradigmeexperimental possible pour tester cette hypothese est le suivant. Apres avoir subi une rotationd’axe verticale a vitesse constante suffisamment longue pour que les canaux semi-circulaires nesoient plus stimules, le sujet est brusquement decelere puis incline dans le plan frontoparallele oule plan sagittal. On mesure alors la direction du nystagmus post-rotatoire. Si elle reste inchangeeapres l’inclinaison, le codage des signaux des canaux semi-circulaires est fixe par rapport a la tete.Dans le cas contraire, on postulera l’existence d’un systeme de coordonnees centrales inertielles.Angelaki et Hess (1994) ont utilise le paradigme precedent sur le singe rhesus, et ont montre qu’ilexiste une reorganisation spatiale du VOR dans les trois plans de l’espace : la direction du nystag-mus s’aligne parfaitement sur la gravite. Le mecanisme d’alignement differe toutefois en fonction dela direction du VOR initial. Pour le VOR horizontal, la compensation post-inclinaison est realiseepar une rotation du vecteur de vitesse des yeux, alors que pour les VOR vertical et torsionnel, lacompensation est obtenue par projection du vecteur de vitesse oculaire sur la direction de la gravite.Le clampage des canaux semi-circulaires n’a pas d’influence sur la compensation, ce qui prouve bienque ce mecanisme est otolithique (Angelaki & Hess 1995). Ainsi, chez le singe rhesus, le systemeotolithique est un systeme inertiel dont la fonction principale serait de transformer l’activite descanaux semi-circulaires en vitesse angulaire de la tete exprimee dans des coordonnees spatiales, uneinformation utilisee pour controler les mouvements de l’oeil, de la tete et du corps dans l’espace3D (Misslisch 1997). Par contre, chez l’homme, aucun alignement de la vitesse de l’oeil par rapporta la gravite n’a ete observe, que l’inclinaison de la tete aie ete generee passivement (Fetter et al.1996) ou activement (Fetter et al. 1992). Force est de conclure que chez l’homme, le codage del’information sur le mouvement de la tete est fixe par rapport a la tete (Misslisch 1997). L’utilisa-tion de referentiels differents chez le singe et l’homme conduit a penser qu’il existe des differencesinterspecifiques importantes en termes de perception spatiale et du mouvement propre. Autrementdit, l’etude de l’action permet d’inferer l’existence de differentes caracteristiques au niveau de laperception.
1.5.2 Mouvements propres passif et actif
Des lors qu’on s’interesse au mouvement, et plus particulierement au couplage perception-action,il convient de s’interroger avec Gibson (1966) pour savoir si l’observateur passif peut extraire la

70 Perception du mouvement et mouvement propre
meme information pendant la locomotion que l’observateur actif qui controle sa locomotion.Autrement dit, il s’agit d’evaluer si ces deux modes de deplacement ont une influence differente surla perception de l’environnement et des mouvements que l’on y realise. Intuitivement, l’engagementdans l’action semble avoir un effet perceptif au moins dans la situation de conflit sensoriel qu’estle mal des transports. Pendant un voyage sur une route montagne, le conducteur du vehicule, quicontrole le mouvement, n’est jamais malade. Par contre, si le conducteur susceptible cede le volant,il deviendra malade.
1.5.2.1 Resultats psychophysiques
–Sens du mouvement de son propre corpsEn ce qui concerne le sens du mouvement et du positionnement du corps (kinesthesie), Craske
et Crawshaw (1975) ont montre que les sujets plonges dans l’obscurite indiquent plus precisementla position de leur main apres un mouvement actif qu’apres le meme mouvement de leur bras realisepar l’experimentateur. Le mouvement du bras etant identique dans les deux cas, la difference deperception de la position de la main montre la contribution significative d’un signal que l’on peutqualifier de commande motrice ou encore de re-afference (von Holst 1954).
–Mouvements oculaires, cephaliques, et perception visuelle du mouvementLes mouvements des yeux et de la tete, parce qu’ils entraınent des deplacement des capteurs
retiniens dans l’espace, sont susceptibles d’interagir avec la perception visuelle du mouvement.Ainsi, s’il est vrai que les mouvements des yeux semblent peu influencer la sensation de mouvementpropre induite visuellement (Dichgans & Brandt 1978, cf. 1.4.1), la perception de la directionde deplacement du corps (heading) est nettement plus precise lorsque le flux optique complexe aanalyser est produit par des mouvements actifs du sujet (Crowell et al. 1998, cf. 1.3.3).
Par ailleurs, les mouvements actifs des yeux et de la tete influent sur la perception du mouvementdes objets. D’une part, les mouvements de poursuite oculaire elevent le seuil de perception dumouvement de l’arriere-plan sur lequel se deplace la cible poursuivie (Wertheim 1981). D’autrepart, les mouvements actifs de la tete elevent eux aussi le seuil de perception du mouvement d’unobjet. Cette elevation du seuil augmente avec la frequence du mouvement cephalique et atteint a1.5Hz trois fois le seuil mesure lorsque le sujet est immobile. Des mouvements passifs de la teteseule, obtenus par deplacement des sujets alors qu’ils fixent une cible visuelle solidaire de leurtete, augmentent aussi le seuil de perception du mouvement d’un objet (Probst & Wist 1982). Laconsequence de cette augmentation du seuil est la diminution de la vitesse apparente de l’objet enmouvement (Berthoz & Droulez 1982).
Enfin, dans la situation de deplacement passif-actif du corps qu’est la conduite de vehicule, laperception du mouvement du vehicule qui precede est egalement fortement diminuee par rapportau meme mouvement visuel observe par un sujet immobile (Probst et al. 1984). Cet ensemble deresultats semble pointer nettement une nouvelle variable de la perception du mouvement –qu’il soitexterne au sujet ou realise par celui-ci : le mouvement produit par le sujet observateur. Probst etal. (1984) suggerent que la diminution du seuil de perception et de la vitesse apparente des objetsenvironnants fait partie d’une hypothese plus generale que fait le cerveau, qui est que l’on se deplacedans un environnement majoritairement stable.
–Coordination oculo-manuelleLes premiers travaux sur la comparaison mouvement actif-passif ont ete realises dans le cadre de
l’apprentissage de taches visuo-manuelles spatiales, suite aux travaux precurseurs sur la necessitede signaux re-afferents ou stimulations provenant de l’action du sujet de von Holst (1954). Held &

Caracteristiques generales du mouvement propre et couplage perception-action 71
Hein (1958) ont utilise une tache de pointage manuel aveugle (les sujets ne pouvant pas voir leurmain pendant qu’ils pointent), avant et apres le port prolonge de prismes decalant l’image visuellevers la droite ou la gauche. Les sujets compensent la deviation due au prismes si et seulement s’ilsont la possibilite de bouger leur main eux-memes pendant qu’ils portent les prismes. Si par contre,l’experimentateur deplace la main des sujets, offrant a ceux-ci une stimulation somatosensorielleet visuelle coherente mais sans mouvement actif, aucune compensation n’apparaıt. Ainsi, l’appren-tissage d’une tache visuo-manuelle spatiale est contingente des signaux re-afferents ; le mouvementseul ne suffit pas, il doit etre actif. De meme, l’apprentissage au cours du developpement de compor-tements spatiaux guides par la vision ne peut se passer des informations fournies par le mouvementactif (Riesen 1958 chez le singe, Hein & Diamond 1983 chez le chat).
Mais l’influence du mouvement actif sur les performances motrices spatiales n’apparaıt pasuniquement a travers l’apprentissage. La poursuite d’une cible visuelle deplacee par le sujet estplus efficace que si le sujet poursuit tout en ayant la main deplacee passivement selon le mememouvement que celui de la cible (Vercher et al. 1996). De plus, les sujets deafferentes realisent desperformances temporelles identiques aux sujets sains. C’est donc le signal re-afferent de commandemotrice qui est implique dans les aspects temporels de la poursuite de la cible manipulee par lesujet, alors que le signal de proprioception motrice est implique dans les ajustements parametriquesde la poursuite (Vercher et al. 1996). Par ailleurs, le mouvement actif modifie aussi l’anticipation dela trajectoire visuelle d’un objet. Lorsque le sujet manipule lui-meme un objet qui se deplace, puisqui est cache tout en continuant a se deplacer en fonction du mouvement de la main, l’estimationde la position de l’objet est plus avancee (le long de la trajectoire) que sa position reelle, et surtoutplus avancee que l’estimation que donne le sujet a propos du meme deplacement de l’objet quecelui qu’il a realise precedemment, mais execute par l’ordinateur (Wexler & Klam 2001). Ainsi,pour un deplacement de l’objet identique, autrement dit pour une stimulation visuelle identique,l’engagement ou non dans une action motrice sur l’objet modifie la perception de la trajectoire del’objet.
–Deplacements de la tete et perception spatialeComme pour les taches spatiales de coordination oculo-manuelle, bouger soi-meme ou etre
deplace dans un environnement –tout en portant des prismes deplacant l’image horizontalement–ne revient pas au meme. Les sujets auxquels Held & Bossom (1961) ont demande d’indiquer ladirection ”droit devant”, apres une periode d’exposition a la modification des interactions visuo-motrices due au port de prismes, ne compensent le biais induit que lorsqu’ils se sont deplacesactivement dans l’environnement.
Plus recemment, Wexler et al. (2001) ont montre un effet immediat du deplacement actif dela tete sur la perception (visuelle) de la structure de l’environnement. Dans la condition active,les sujets provoquent leur propre deplacement. Dans la condition passive, ils recoivent la memestimulation visuelle que dans la condition active, mais ne bougent pas. En utilisant un objet virtuela l’aide duquel les indices spatiaux issus de la perspective et du mouvement sont mis en conflit,les auteurs ont pu montrer que, d’une part, l’information sur le mouvement propre fait partieintegrante de l’analyse visuelle de la structure 3D de l’environnement –l’effet du mouvement actifne se reduit pas seulement a la modification du flux optique qu’il entraıne–, et, d’autre part, que lessujets preferent la solution ou l’objet reste stationnaire dans l’espace (un referentiel allocentrique)meme si celui-ci se deforme. L’equivalence implicite dans les etudes consacrees a l’extraction de lastructure spatiale 3D a partir des informations de mouvement entre un sujet statique observantdes objets rigides subissant un mouvement et un sujet en mouvement actif observant des objetsstationnaires est donc erronee.
Dans une deuxieme serie d’experiences, Wexler (2002) a compare plus specifiquement mou-

72 Perception du mouvement et mouvement propre
vement actif et mouvement passif dans une tache spatial utilisant a nouveau un objet virtuelambigu. Il existe deux solutions possibles de mouvement pour ce stimulus ambigu, l’une dansun referentiel egocentrique et l’autre dans un referentiel allocentrique. Ces deux solutions corres-pondent a des axes de rotation de l’objet separes de 90◦. La tache du sujet etant d’indiquer l’axede rotation, le grand interet de ce paradigme est qu’il ne fait pas appel directement a un jugementspatial ego/allocentrique. Les resultats montrent que le mouvement actif influe sur la perceptionimmediate du mouvement et de l’espace 3D : les sujets ont tendance a utiliser plus frequemmentle referentiel allocentrique pendant le mouvement actif, alors que pendant un mouvement proprepassif equivalent, ils utilisent de preference le referentiel egocentrique. Ainsi, du point de vue de laperception de l’espace 3D, un mouvement propre actif du sujet n’est pas simplement l’equivalent dudeplacement du sujet dans l’espace. L’ensemble des resultats presentes semble indiquer que mou-vement et espace apparaissent reellement indissociables d’une part, et que d’autre part mouvementactif et deplacement passif ne sont pas equivalents pour le cerveau.
1.5.2.2 Resultats neurophysiologiques
Le nombre reduit de resultats indiquant le role du mouvement actif sur la perception contrasteavec la relative abondance de donnees psychophysiques.
–Perception du mouvementA propos du mouvement propre, et dans le cas particulier de la direction de deplacement, Brad-
ley et al. (1996) ont montre que les neurones de l’aire corticale MST ne compensent la perturbationdu flux optique due au mouvement de l’oeil que si le mouvement est execute par le singe. Le memeflux optique observe passivement ne permet pas de retrouver la direction de deplacement a partirdu flux optique. De plus, si l’animal subit une rotation pendant qu’il supprime son reflexe vestibulo-oculaire –ce qui provoque une perturbation du flux optique due au mouvement de la tete seule–,les neurones de MST peuvent compenser les effets de ce mouvement cephalique passif (Shenoy etal. 1999 ; cf. 1.3.3).
Par ailleurs, les neurones de MST ont une reponse plus faible lorsque le mouvement visuelsur la retine est provoque par un mouvement des yeux que lorsqu’il est totalement independantde l’animal (Erickson & Thier 1991), ce qui confirme l’influence plus generale des signaux extra-retiniens dus au mouvement sur l’analyse du mouvement dans le cortex temporal. Dans l’airetemporale polysensorielle (TPO, TGa), l’on trouve aussi des neurones sensibles au mouvementd’un objet. Par contre, ces neurones ne repondent pas lorsque c’est la main de l’animal qui esten mouvement, et ils repondent seulement faiblement si l’animal approche un objet nouveau danssa main afin de l’examiner (Heitanen & Perret 1993). Ainsi, les neurones des TPO sont sensiblesau mouvement des objets, preferentiellement si ce mouvement n’est pas provoque par l’animallui-meme.
–Neurones vestibulairesDeux etudes neurophysiologiques sur les reponses liees au mouvement de la tete dans les noyaux
vestibulaires ont compare ces reponses lors de mouvements cephaliques actifs et passifs, et ontmontre que les neurones reagissent differemment dans les deux cas. Robinson & Tomko (1987)ont compare les neurones vestibulaires secondaires chez le chat, lors de mouvements actifs del’animal et lors d’une repetition de ces mouvements par la table tournante. Environ un tiers des 35cellules enregistrees repondent plus intensement au mouvement actif qu’au mouvement passif. Ladifference de reponse dans les deux conditions pouvait etre expliquee par l’addition d’une entreesomatosensorielle nuccale au signal vestibulaire pur dans la condition active. Toujours dans les

Caracteristiques generales du mouvement propre et couplage perception-action 73
noyaux vestibulaires, mais chez le singe cette fois, tous les neurones repondent moins intensementau mouvement actif qu’au mouvement passif de la tete, la grande majorite (2/3) ne repondantpas du tout au mouvement actif (Mc Crea et al. 1999). L’hypothese avancee par les auteurs pourexpliquer cette difference est que le signal vestibulaire serait annule par la soustraction d’un signalde copie efferente motrice pendant le mouvement actif.
Si les resultats de ces deux etudes semblent a priori contradictoires, il n’en reste pas moins que lessignaux neuronaux relatifs au mouvement de la tete, des les premiers relais vestibulaires centraux,sont differents suivant que le mouvement est actif ou passif. Si le cortex parietal est implique dansla representation de l’espace, a la lumiere des resultats psychophysiques et neurophysiologiquesprecedents, il devient tres interessant de savoir si les signaux vestibulaires qui s’y trouvent sontaussi modifies par la nature du mouvement produit. La cinquieme etude de ce travail de these estconsacree a cette question.





























