Embed Size (px)
Citation preview

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 1/16
ORIGINAL ARTICLE
Transforming Growth Factor-A1 May Be a Key Mediator of the
Fibrogenic Properties of Neural Cells in LeprosyRafael B. Petito, PhD, Thaıs P. Amadeu, PhD, Bernardo M. O. Pascarelli, MSc,
Ma ´rcia R. Jardim, MD, PhD, Robson T. Vital, MD, Sergio L. Antunes, PhD,
and Euzenir N. Sarno, MD, PhD
AbstractFibrosis is the main cause of irreversible nerve damage in leprosy.
Phenotypic changes in Mycobacterium leprae (ML) Y infected Schwann
cells (SCs) have been suggested to mediate this process. We found that
SC line cultures stimulated with ML upregulated transforming growth
factor-A1 (TGF-A1), and that TGF-A1 or ML induced increased
numbers of >-smooth muscle actin (>-SMA) Y positive cells with
characteristic stress fibers. Mycobacterium leprae and TGF-A1 also
induced increased type I collagen and fibronectin mRNA and secretion
and augmented mRNA levels of SOX9 and ZEB1, which are involved
in the epithelial-mesenchymal transition. These effects could be
inhibited by the TGF-A1 type I receptor (ALK5) inhibitor, SB-431542.
In nerve biopsies from leprosy-infected patients with varying grades of
fibrosis (n = 11), type I and III collagen and fibronectin were found in
the endoneurium and perineurium, >-SMA Y positive cells filled the
fibrotic perineurium but not the endoneurium, and CD34-positive fi-
broblasts predominated in the endoneurium. Results of transcriptional
studies of 3 leprosy nerves and 5 controls were consistent with these
data, but >-SMA and other mRNA levels were not different from
those in the control samples. Our findings suggest that TGF-A1 mayorchestrate events, including reprogramming of the SC phenotype,
leading to transdifferentiation, connective tissue cell expansion, and
fibrogenesis in the evolution of leprosy nerve lesions during some
evolutionary stages.
Key Words: >-Smooth muscle actin, Fibrosis, Leprosy, Mycobac-
terium leprae, Peripheral neuropathy, Schwann cell, Transforming
growth factor-A1.
INTRODUCTION Nerve lesions in leprosy constitute one of the better
characterized processes in which chronic inflammation pro-
gresses to fibrosis that result in the deformities and permanent
paralyses that have stigmatized this disease for millennia.Although fibrosis is currently of great scientific interest inrelation to many diseases, nerve fibrosis has not gained its dueattention perhaps because it is an uncommon condition amongthe neurologic disorders and only rarely referred to in the fieldof leprosy.
Fibrosis is the widespread end stage of many inflamma-tory diseases, but because the pathways driving fibrosis areuncertain, it remains without effective treatment. Deposition of ECM components is a very common histopathologic charac-teristic in fibrotic disorders, including cirrhosis, renal and sys-temic sclerosis, pulmonary fibrosis (1 Y 4), hypertrophic scars,and keloids (5). Moreover, a variety of cell lineages seem tooriginate cells that overproduce collagen (4).
Nerve fibrosis has been described in 60% of the biop-sies taken from leprosy patients with peripheral neuropathy(6), yet the main cell involved in fibrogenic repair remainsunknown (7). Although long-term infection seems to be themain cause of nerve destruction in leprosy, progression tosevere fibrosis occurs even after effective bacterial clearance
by multidrug treatment has taken place, implying that thefibrogenic cells take command of later events. In addition,
posttreatment immunologic reactions may play an important role in the evolution of fibrosis (8).
The remodeling of ECM after nerve injury is an im- portant step in nerve regeneration, involving the effects of proteases and their inhibitors as well as de novo collagensyntheses, which is orchestrated mainly by transforminggrowth factor-A1 (TGF-A1) (9, 10). Our previous studysuggested that the maintenance of high metalloproteinaseexpression (MMP-2 and MMP-9), together with a tissueinhibitor of metalloproteinase 1, could lead the nerveenvironment toward fibrosis subsequent to Mycobacteriumleprae (ML) nerve infection (11).
Transforming growth factor-A, a critical regulator of ECM deposition, is a key growth factor in the development of tissue fibrosis (12). Many studies have attributed an important anti-immune/inflammatory role to TGF-A1 in ML infection(13, 14). Nonetheless, its role in determining the outcome of ML infection in the nerve requires further clarification. Wealso have reported induction of TGF-A receptor transcriptionin the ST88-14 Schwann cell (SC) line after in vitro stimula-tion with ML (15).
It is widely accepted that >-smooth muscle actin(>-SMA) Y posi tive cells (myofibroblasts) are the key cell
351 J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
J Neuropathol Exp NeurolCopyright Ó 2013 by the American Association of Neuropathologists, Inc.
Vol. 72, No. 4April 2013
pp. 351 Y 365
Leprosy Laboratory, Oswaldo Cruz Institute, FIOCRUZ (RBP, TPA, BMOP,MRJ, RTV, SLA, ENS); and Department of Neurology, FluminenseFederal University, Niteroi (RTV), Rio de Janeiro, Brazil.
Send correspondence and reprint requests to: Euzenir N. Sarno, MD, PhD,Leprosy Laboratory, Oswaldo Cruz Institute, Oswaldo Cruz Foundation,Av. Brasil 4365, Manguinhos, 21040-360 Rio de Janeiro, Brazil; E-mail:[email protected]
Rafael B. Petitoand Thaıs P. Amadeu contributedequallyto this work as co Y first authors.
The authors declare no financial or commercial conflicts of interest.This work was supported by CNPq, FIOCRUZ, CAPES, and FAPERJ.
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 2/16
mediators of fibrosis, acting as the primary collagen-producingcells in conjunction with a variety of proinflammatory activities(4). Whereas these cells are also present in normal tissue, theyare more frequently found in locations requiring mechanicalforces (16). It seems, however, that the mechanisms leading to>-SMA expression and persistence in various pathologic situ-ations are different (17). Among the cells capable of expressing>-SMA, CD34-positive fibroblasts, smooth muscle cells, and
pericytes are mutually distinguishable and may be involved inthe fibrotic processes (4, 18).
In chronic traumatic peripheral nerve lesions, perineu-rial cells can proliferate and form tumor-like lesions (i.e. peri-neuriomas); >-SMA immunoreactivity has been reported in60% of such lesions (19, 20). Epithelial cells can become a source of myofibroblasts through a process called epithelial-mesenchymal transition (EMT) (21). During morphogenesis,wound healing, fibrotic organ diseases, and tumor progression,some cells undergo EMT, acquiring increased migratory ca-
pacity and mesenchymal protein expression, which may beinduced by TGF-A1 (22 Y 25). In EMT (the most frequently
used study model of transdifferentiation), several transcriptionfactors have been described as E-cadherin repressors, amongwhich are the zinc figure E box Y binding (ZEB) transcriptionfactors (23). These proteins function downstream of the EMT-inducing signaling pathways activated by TGF-A1, fibroblast growth factor, and other molecules (25). Their increasedexpression results in rapid EMT. They also regulate prolifera-tion, susceptibility to apoptosis, and senescence (23). Zincfigure E box Y binding 1 has been demonstrated in S100-
positive cells in neurofibromas when EMT-related signalingin SCs seems to occur (24). Real et al (26) demonstrated theability of TGF-A1 Y cultivated quail SCs to transdifferentiate intomyofibroblasts, and Roh et al (27) found that the SOX9 geneexpression also increases in rat SCs transdifferentiated into
myofibroblasts by SOX10 downregulation. Thus, the high de-gree of plasticity of SCs suggests that they may have roles in
particular circumstances such as in repair.In the present study, we investigated the effects of ML
on SC TGF-A1 transcription and secretion, the effects of ex-ogenous TGF-A1 on >-SMA expression, morphology, andECM deposition in vitro and correlated the findings with thosein nerve biopsies of leprosy patients. Our data indicate that myofibroblasts may play a role in the fibrogenic evolution of leprosy neuritis.
MATERIALS AND METHODS
Nerve Biopsy Specimens Nerve biopsy specimens were collected at the Souza
Arau jo Ambulatory, Oswaldo Cruz Foundation, Rio de Janeiro,Brazil, for diagnostic purposes. Eleven patients (7 men and4 women) with a mean TSD age of 41.2 T 14.5 years wereselected. Four patients were diagnosed as having pure neuralleprosy based on criteria established in a previous publication
by our group (28) and others (29), 5 were determined to haverelapsed leprosy, and 2 had reactional neuritis. Histopathologicexamination of the biopsy specimens revealed an inflammatoryinfiltrate composed of mononuclear cells containing epithelioidgranulomas or foamy macrophages. The infiltrates were dis-
tributed within the surrounding epineurial and perineurial smallvessels, occupying a variable extension of the endoneurialcompartment. Endoneurial and/or perineurial fibrosis were
present in all the nerves, as evaluated by the extension of thegreen staining area in the Gomori trichrome. Inflammatory in-filtrate and fibrosis replaced the myelinated fibers, leavingsome with regenerated, degenerated, or remyelinated features.
Neovascularity was detected within the perineurium and amidthe fibrotic endoneurium (Table).
All patients received the standard treatment describedin the Brazilian Ministry of Health guidelines for leprosy.
Normal sural nerves (n = 8) from healthy young male adults(20 Y 40 years of age) were obtained after nerve autograft surgeryto reconstruct the brachial plexus as a result of traumatic injury.These control specimens were considered to be normal becauseno peripheral neuropathy was found.
Nerve biopsy specimens were divided into 2 smallfragments. One was fixed in phosphate-buffered 4% parafor-maldehyde (PFA), processed, and embedded in paraffin for routine staining, and the other was preserved in liquid nitro-
gen until further use. Five-micrometer-thick paraffin sectionsstained with hematoxylin and eosin (for general evaluation),Gomori trichrome (for ECM evaluation) and Wade stains (toidentify acid-fast bacilli [AFB] Y loaded cells) were examinedvia a Nikon E400 light microscope (Japan). This examina-tion protocol revealed the morphologic status of each nerve(Table). Paraffin sections were also used in the immunohis-tochemical assays; cryosections of the frozen samples wereused in the immunofluorescence assays. This study was ap-
proved by the Ethics Committee on Human Research of theOswaldo Cruz Foundation (Protocol 538/09).
ML and ReagentsIrradiated armadillo-derived ML was generously donated
by Dr. P. Brennan (Department of Microbiology, ColoradoState University, Fort Collins, CO). The ML sample wasdetermined to be pure and without endotoxin. All stimuli usedfor in vitro cultures were shown to contain less than 0.1 U/mLendotoxin according to the limulus amebocyte lysate assay(Whittaker Bioproducts, Walkersville, MD). In a previous
publication, Marques et al (30) demonstrated that no armadillo protein was present in the dead ML sample used in this study.Human recombinant TGF-A1 was purchased from R&D Sys-tems (Minneapolis, MN), and the TGF-A1 type I receptor (ALK5) inhibitor SB-431542 was obtained from Tocris Bio-science (Bristol, UK).
SC CulturesThe ST88-14 tumor cell line generously donated by J. A.
Fletcher (Dana Farber Cancer Institute, Boston, MA) wasestablished from malignant schwannomas (neurofibrosarcomas)of patients with neurofibromatosis type I. The cells were grownin RPMI 1640 medium (Invitrogen, Carlsbad, CA) supple-mented with 100 U penicillin/mL, 100 Kg of streptomycin/mL,2 mmol/L L-glutamine, and 15% fetal calf serum (HyClone,Waltham, MA) in a humidified CO2 incubator at 37-C.
For the experimental assays, the attached ST88-14cells were released using trypsin/EDTA (0.25%, 1 mmol/L) for
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.352
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 3/16
1 minute. The cells were then washed in RPMI-FBS, suspendedin complete medium, and further cultured on 24-well plates(Falcon, Franklin Lakes, NJ) for 1 to 24 hours (1 Â 105 cells/ well) or 7 days (1 Â 104 cells/well); onto 6-well plates for 1 to6 hours (5 Â 105 cells/well); and onto 96-well plates for 7 days(4 Â 103 cells/well) for in vitro stimulation.
To evaluate the effects of blocking ALK5 responses,cultures were incubated with fresh medium containing10 Hmol/L SB-431542 or vehicle (dimethyl sulfoxide[DMSO]) for 1 hour before the addition of stimulus. TheTGF-A1 dosage and incubation times were determined in
previous kinetic and dose-response experiments. The time points tested were 1, 2, 4, and 7 days after adding TGF-A1 tothe cultures. Seven days were considered the optimal timeresponse to evaluate >-SMA and ECM expression. Different concentrations were also tested, and the optimal concentra-tion was 10 ng/mL.
RNA Isolation and cDNA SynthesisFive normal and 3 leprosy nerve fragments stored in
liquid N2 were recovered and homogenized using a PolitronPT-3000 in 1 mL of Trizol (Invitrogen). All samples were
processed simultaneously. Schwann cells were suspended in1 mL Trizol. Total RNA was extracted according to themanufacturer’s instructions. RNA purity was verified by theOD260/280 absorption ratio of 1.9 to 2.0, and cDNA wassynthesized via Superscript III first-strand reverse transcrip-tase polymerase chain reaction (RT-PCR) Kit (Invitrogen).
Semiquantitative RT-PCRA semiquantitative analysis of TGF-A1 (Tgfb1) mRNA
was determined in SC cultures as previously described (15)(Applied Biosystems, Foster City, CA). Sequences for Tgfb1
were: 5¶-CTACTACGCCAAGGAGGTCAC-3¶ and 5¶-TTGCTGAGGTATCGCCAGGAA-3¶. The primers were designedto avoid genomic DNA amplification. Polymerase chain re-action was performed as detailed previously (31), and thesamples were amplified in a DNA thermocycler 2400 (PerkinElmer Applied Biosystems). Polymerase chain reaction prod-ucts were subjected to electrophoresis on 1.7% agarose gelswhere specificity of the amplified bands was validated bytheir predicted size (A-actin, 661 bp; Tgfb1, 326 bp). Den-sitometer analysis was performed by scanning the gel im-ages (Video Documenting System, Amersham-Pharmacia,Piscataway, NJ), and densitometric values were obtained via ImageMaster software.
Real-time PCRSOX9 (Sox9), ZEB1 (Zeb1), type I collagen (Col1a2),
and fibronectin (Fn1) mRNA expression was determinedin SC cultures and human nerve tissues using a TaqMan real-time PCR kit; >Sma mRNA expression was determined via an SYBR-Green kit (Applied Biosystems) by the Illumina real-time PCR system (ECO, San Diego, CA). In nervetissues, TGF-A1 (Tgfb1) expression was also evaluatedusing a TaqMan real-time PCR kit. Polymerase chain reac-tion amplification was carried out by way of 100 ng cDNA T
A B L E .
C l i n i c a l a n d H i s t o p a t h
o l o g i c D a t a o f N e r v e B i o p s y S a m p l e s
P a t i e n t
S e x
A g e ,
y e a r s
C l i n i c a
l
D i a g n o s
i s
P e r i n e u r i a l
I n f l a m m a t i o n
E n d o n e u r i a l
I n f l a m m a t i o n
E p i t h e l i o i d
G r a
n u l o m a
P e r i n e u r i a l
F i b r o s i s
E n d o n e u r i a l
F i b r o s i s
S m a l l
M F
L o s s
L a r g e
M F
L o s s
A x o n
R e g e n e r a t i o n
A
x o n
R e m y e l i n a t i o n
N e o
B l o o d
V e s s e l s
1
M
7 5
P N L
+ + +
+ + +
+ + +
+ +
N
+ + +
+ + + +
N
N
+ / + +
2
F
3 9
P N L
N
+ +
N
N
N
+ + + +
+ + +
N
N
N
3
M
2 3
R L
+ +
+ + +
+ +
+ +
+ +
+ + + +
+ + + +
N
N
N
4
F
4 4
P N L
+ + +
+ + +
N
+ + +
+ + +
+ + + +
+ + + +
N
N
+ + +
5
M
3 5
P N L
+ +
+ +
+
+
+
+
+
+ +
+
+ + +
6
F
6 1
R L
+
+ +
+
+
+ + +
+ + + +
+ + + +
N
N
N
7
M
3 7
R N
+ + +
+ + +
+ +
+ +
+ +
+ +
+ +
+ +
+ +
+ + / + + +
8
M
3 3
R L
+ + +
+ + +
+ +
+ +
+ + +
+ + +
+ + + +
N
N
+ +
9
M
3 9
R L
N
+ +
N
+ +
+
+
+
+ +
+
+ +
+
1 0
M
3 3
R N
+
+ +
N
+
+
+ + +
+ + +
+
+
+ / + +
1 1
F
3 5
R L
+
+ +
N
+ + +
+ + +
+ + + +
+ + +
+
N
+ +
F ,
f e m a l e ; M ,
m a l e ; M F ,
m y e l i n a t e d fi b e r s ; N e o ,
p r e s e n c e o f n e w b l o o d v e s s e l s i n t h e p e r i n e u r i u m a n d / o r e n d o n e u r i u m ; P N L ,
p u r e n e u r a l l e p r o s y ; R L ,
r e l a p s e l e p r o s y ; R N ,
r e a c t i o n a l n e u r i t i s .
I n t e n s i t y : N ,
a b s e n t ; + ,
w e a k l y ; + + , m
o d e r a t e ; + + + ,
s t r o n g ; + + + + ,
v e r y s t r o n g .
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 353
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 4/16
added to triplicate tubes with 5 HL of the master mix and0.5 HL or 2 HL of the probe (FAM-MGB) containing for-ward and reverse primers for glyceraldehyde-3-phosphatedehydrogenase (Gapdh), Tgfb1, Sox9, Zeb1, Col1A2, or Fn1(Applied Biosystems). The primers used for >Sma wereforward: 5¶CCGACCGAATGCAGAAGG-3¶, reverse: 5 ¶-ACAGAGTATTTGCGCTCCGGA-3¶, target fragment length88 bp. Relative mRNA expression was analyzed during thePCR log phase by the comparative cycle threshold method (32).
None of the template controls for each gene showed any con-taminating DNA.
Flow Cytometry Transforming growth factor-A1 and >-SMA intracel-
lular staining was performed separately after cells wereharvested, washed, and fixed with 4% PFA. Cells werewashed in 1% bovine serum albumin (BSA) in PBS and
permeabilized in 0.1% saponin/PBS for 20 minutes at roomtemperature. Cells marked for TGF-A1 or >-SMA were sub-sequently washed in 0.1% saponin/PBS and incubated with
mouse monoclonal primary anti Y
TGF-A1 (1:50; R&D Sys-tems) or anti Y >-SMA (1:50; Dako, Carpinteria, CA) anti- bodies. The cells were then incubated with FITC conjugategoat anti-mouse antibody and resuspended in PBS beforeanalysis on an Accuri C6 flow cytometer (BD Biosciences).Positivity thresholds were set by way of irrelevant anti-mouseimmunoglobulin G (IgG) as negative controls. Data wereanalyzed by C Flow Plus software for Windows (BD Bio-sciences).
TGF-A1 ELISAsTransforming growth factor-A1 concentrations in cell-
free culture supernatants were determined using commer-cially specific ELISAs that had been processed according to
the manufacturer’s specifications (Promega, Charbonnieres,France) (15, 33).
Analysis of ECM Secretion by Indirect ELISATo determine the relative levels of type I collagen and
fibronectin in native matrices, a semiquantitative ELISA was set up according to Alves et al (34), as follows. Cells were grownin sterile ELISA 96-well plates for 7 days. After monolayer lysis with Triton X-100/ammonia solution, the whole plate was
blocked with 0.1% BSA/PBS-Ca 2+ for 2 hours and then incu- bated with primary anti Y type I collagen (1:5000) or anti-fibronectin (1:5000) antibodies diluted in PBS-Ca 2+/Tween for 1 hour at 37-C. After washing with 0.1% BSA/PBS-Ca 2+/ Tween 0.05%, horseradish peroxidase Y conjugated secondary
antibodies diluted in the same buffer were added for 1 hour.Colorimetric reaction was determined by citrate buffer 0.1 mol/L
pH 4.5 containing 0.002% H2O2 and o-phenylenediaminesubstrate 1 mg/mL and was stopped with H2SO4 3 mol/L.Absorbance was measured at 490 nm in an ELISA micro-
plate automatic reader (Spectra Max 190; Molecular Devices,Sunnyvale, CA).
Immunolabeling in SCsST88-14 SCs were cultured onto 24-well plates
containing glass coverslips covered with 4% silane (Sigma
Chemical Co., St. Louis, MO). The samples were washedwith PBS and fixed in cold methanol for >-SMA staining or with 4% PFA for type I collagen and fibronectin, blocked and
permeabilized in 0.5% Triton X-100/10% goat serum and10% fetal calf serum in PBS, for >-SMA, or only blocked in10% goat serum and 10% fetal calf serum in PBS for 1 hour at room temperature. The cells were then incubated with themouse anti
Y >-SMA (1:100; DAKO), rabbit anti
Y type I col-
lagen (1:200; Abcam, Cambridge, MA), or rabbit anti-fibronectin (1:200; Sigma) antibodies diluted in 1% BSA inPBS. After rinsing with PBS, Alexa 633 goat anti-mouse or Alexa 532 goat anti-rabbit IgG secondary antibody was addedand incubated for 1 hour at room temperature. Cellular nucleiwere stained with DAPI (Sigma), and coverslips weremounted with Permafluor (Thermo Scientific, San Jose, CA).Cells were analyzed by an Axio Observer Z1 Colibri micro-scope (Zeiss, Oberkochen, Germany), and images were
processed via AxioVision software (Zeiss).
Immunohistochemical and ImmunofluorescentStaining in Nerve Biopsy SpecimensTo characterize the distribution of >-SMA Y positive
cells, perineural cells and fibroblasts in the peripheral nerves, paraffin sections (5 Km) of normal and leprosy nerve tissueswere incubated overnight with mouse monoclonal antibodiesagainst human >-SMA (1:500; DAKO), glucose transporter 1(GLUT-1) (1:100; Thermo Scientific), and CD34 (1:50;DAKO), respectively. To detect CD34, antigen retrieval was
performed by way of proteolysis digestion for 30 minutes at 37-C. Before labeling, deparaffinized and rehydrated sectionswere incubated for 30 minutes in a 3% H2O2 methanol solu-tion to block endogenous peroxidase. After rinsing in PBS,the primary antibody was detected using the HiDef Detec-
tion HRP Mouse/Rabbit Polymer System (Cell Marque,Rocklin, CA). Afterward, sections were incubated withdiaminobenzidine (Cell Marque), counterstained with hema-toxylin, dehydrated, and mounted. The negative controls were
performed by omitting the primary antibody, and no labelingwas observed. The slides were examined on a Nikon E400light microscope (Japan), and the images were captured via a videomicroscopic system composed of a Nikon E400 micro-scope, CoolSNAP-Procf Color camera, and Infinity analysissoftware.
To identify the colocalization of GLUT-1 and >-SMAin nerve specimens and to characterize the distribution of theECM components, double immunofluorescences were per-formed in normal and leprosy nerve tissues. Cryosections were
fixed in acetone for 10 minutes, and the nonspecific bindingswere blocked with 10% goat serum and 10% fetal calf serum inPBS for 1 hour. Primary mouse monoclonal antibodies to detect >-SMA (1:50; DAKO), type I collagen (1:100; Abcam), typeIII collagen (1:500; Sigma), or fibronectin (1:50; Sigma) wereapplied overnight at 4-C, and the primary rabbit polyclonalantibody against GLUT-1 (1:50; Thermo Scientific) was ap-
plied for 2 hours. The slides were washed in PBS and stainedwith goat anti-mouse IgG (1:500, Alexa Fluor 532; Molecular Probes, Eugene, OR) for >-SMA, type I and III collagen or fibronectin, and with goat anti-rabbit IgG (1:500, Alexa Fluor
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.354
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 5/16
633; Molecular Probes) for GLUT-1 detection. Lastly, speci-mens were rewashed with PBS and mounted on Permafluor (Thermo Scientific). Nuclei were stained with DAPI; negativecontrols were performed by omitting the primary antibody, andno labeling was observed.
Quantification Analysis of CD34-Positive Cellsand Endoneurial FibrosisThe peripheral nerves were analyzed considering the
CD34-positive cells and connective tissue. The analysis was performed via a videomicroscopic system (Nikon E400 mi-croscope, Infinity capture color camera, and Image Pro soft-ware [Media Cybernetics, Rockville, MD]) for processingcaptured digital images, including a stereologic test systemcomposed of 36 test points (PT), as previously described (35,36): Vv = PP/PT (%) (PP represents the evaluated points that hit the structure). Five random fields were randomly analyzed
by blindly moving the stage of the microscope for each nervesection. Results are expressed as mean T SD.
Statistical AnalysisThe results were expressed as mean T SE or mean T SD.
Significant differences between 2 groups were determined by the Wilcoxon nonparametric test in which p e 0.05 wasconsidered significant. Friedman nonparametric test wasused to compare multiple groups using Dunn posttest inwhich p G 0.1 was considered significant. Analysis were
performed using Windows GraphPad Prism version 5.0(GraphPad Software, San Diego, CA). Quantitative analysisof CD34 and endoneural connective tissue was performed byan unpaired t -test with Welch correction using GraphPadInstat 3.1.
RESULTS
ML Induces TGF-A1 Expression in SCsPrevious work by our laboratory has shown that ML
(100:1) induced Tgfb1 upregulation and downregulation in thehuman SC lineage (ST88-14) after 1 and 3 hours, respectively(15). To elucidate the effect of ML on Tgfb1 expression, inthis study, we initially sought to evaluate if an ML concentra-tion of 50:1 would have the same effect on Tgfb1 expressionafter 1, 3, and 6 hours of interaction with ST88-14 SCs. Thus,a semiquantitative RT-PCR analysis was performed. After 1 hour of ML infection, the cultures showed significant Tgfb1upregulation in comparison with the unstimulated cells (p G0.05) (Fig. 1A). After 3 and 6 hours of ML exposure, Tgfb1
downregulation was detected in these same cultures in com- parison with unstimulated cells (p G 0.05) (Fig. 1A).
To investigate whether the effect observed in the presenceofMLon Tgfb1 expression was reflected in intracellular protein
production and secretion, TGF-A1 protein levels were mea-sured by flow cytometry and in supernatants by ELISA in ML-stimulated SCs, respectively. In contrast to unstimulated cells,the presence of ML increased intracellular TGF-A1 produc-tion after 1 and 6 hours (p G 0.05) (Fig. 1B). On the con-trary, however, downregulation of intracellular TGF-A1 wasobserved after 3 hours of ML exposure (p G 0.05) (Fig. 1B).
Intracellular TGF-A1 production remained higher after 24 hoursand 7 days of ML exposed versus unstimulated cells (p G 0.05)(Fig. 1B). The presence of ML also increased TGF-A1 secre-tion after 1 and 6 hours and 7 days versus unstimulated cells,
but without statistical difference (Fig. 1C). Similarly to the re-sults obtained after 3 hours, downregulation in TGF-A1 secre-tion was also observed at this time point (p G 0.05) (Fig. 1C).As a whole, these results demonstrate that ML induced TGF-A1 gene expression, intracellular production, and protein se-cretion in ST88-14 SCs.
FIGURE 1. Mycobacterium leprae (ML) induces increased trans- forming growth factor-A1 (TGF-A1) expression in Schwann cells(SCs). (A) Semiquantitative RT-PCR was performed to measureTgfb1 mRNA expression in ST88-14 SCs stimulated or not withdead ML (multiplicity of infection, 50:1) for 1, 3, and 6 hours.(B) Intracellular TGF-A1 production was evaluated by flow cy-tometry in ST88-14 SCs stimulated or not with dead ML (50:1) for 1, 3, 6, and 24 hours and 7 days of culture. (C) TGF-A1 levelswere verified in supernatants of ST88-14 SCs stimulated or notwith dead ML (50:1) for 1, 3, 6, and 24 hours and 7 days byELISA. Results represent mean T SE of 5 or more independentexperiments. NS, unstimulated cells. Wilcoxon nonparametrictest was used for statistical analysis in which * p G 0.05.
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 355
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 6/16
FIGURE 2. Transforming growth factor-A1 (TGF-A1) induces >-smooth muscle actin (>-SMA) expression in Schwann cell (SCs).ST88-14 SCs were incubated with recombinant human TGF-A1 (10 ng/mL) or Mycobacterium leprae (ML) (50:1) and preincubatedwith the TGF-A1 type I receptor (ALK5) inhibitor (SB-431542) (10 Kmol/L), followed by TGF-A1 or ML for 7 days. (A) Real-time RT-PCR for >Sma mRNA expression (n = 3). (B) Intracellular >-SMA expression detected by flow cytometry in ST88-14 SCs incubatedwith TGF-A1 or ML (n Q 4). NS, unstimulated cells. Friedman nonparametric test with Dunn posttest was used for statistical analysisin which * p G 0.1. (C) Representative immunofluorescence of >-SMA expression (green, Alexa Fluor 633) in ST88-14 SCs incubatedwith TGF-A1 or ML for 7 days. Results are representative of 5 independent experiments. Nuclei were labeled with DAPI. Imageswere visualized and obtained by a fluorescence microscope. Scale bar = 20 Km.
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.356
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 7/16
TGF-A1 Induces >-SMA Expression DuringSC-Myofibroblast Transdifferentiation
We next analyzed effects of TGF-A1 and ML on >-SMA(>Sma) mRNA expression and intracellular production on SCST88-14 cells after 7 days of culture. Polymerase chain reac-tion showed that TGF-A1 and ML induced increased >Sma
levels versus unstimulated cells (p G 0.1) (Fig. 2A). Flow
cytometry revealed that TGF-A1 and ML also induced higher intracellular >-SMA production in comparison with unstimu-lated cells (p G 0.1 for both) (Fig. 2B). When ML was addedto TGF-A1 in the culture, the same effect as TGF-A1 alone wasobserved (data not shown). Thus, these results strongly suggest that the presence of ML induces increased TGF-A1 levels. Toconfirm this observation, SCs were pretreated with SB-431542,an inhibitor of the TGF-A1 type I receptor (i.e. ALK5), andthen incubated with TGF-A1 or ML for 7 days. AlthoughSB-431542 inhibited the effect of TGF-A1 and ML on >Smaexpression, no statistical differences were observed. Data dem-onstrated that SB-431542 likewise inhibited the effect of TGF-A1 and ML on intracellular >-SMA production whencomparing this effect to their respective controls, althoughno statistical significance was observed (Fig. 2A, B).
The transdifferentiation accompanied by >-SMA ex- pression detected by flow cytometry analysis was also ob-served by immunofluorescence. In this analysis, some of the>-SMA Y positive SCs, stimulated with TGF-A1 or ML after 7 days, had stress fibers (Fig. 2C). The same results were
obtained when an experiment was performed with primaryhuman SCs prepared from nerve explants of adult humandonors (generously provided by Dr. Patrick Wood, Miller School of Medicine, Miami, FL) (37, 38) (data not shown).
TGF-A1 Induces an Increase in TranscriptionFactors Involved in SC-MyofibroblastTransdifferentiation
To examine Sox9 and Zeb1 mRNA expression, SCswere stimulated with TGF-A1 or ML for 1 hour because
preliminary results detected upregulation of TGF-A1 mRNAat this time point. Sox9 and Zeb1 levels were upregulated
by TGF-A1 versus unstimulated cells (p G 0.1 for both)(Fig. 3A, B). The presence of ML led to a significant up-regulation of Sox9 and Zeb1 levels versus the unstimulatedcells (p G 0.1 for both) (Fig. 3A, B). To confirm the effect of TGF-A1 on Sox9 and Zeb1 expression, SCs were pretreatedwith SB-431542 and then incubated with TGF-A1 or ML for 1 hour. SB-431542 significantly inhibited the effect of TGF-A1on Sox9 levels and ML on Zeb1 levels when compared withtheir controls without inhibitor (p G 0.1 for both) (Fig. 3B).
TGF-A1 Induces an Increase in Type I Collagenand Fibronectin by SCs
Mouse SCs increase nerve collagen deposition by ML
infection (39). Therefore, we evaluated effects of TGF-A1 andML on type I collagen and fibronectin gene expression andtheir secretion by SCs. Schwann cells were stimulated withTGF-A1 or ML for 7 days, after which a real-time RT-PCR analysis was performed. Results showed that TGF-A1 in-creased type I collagen (Col1A2) and fibronectin ( Fn1) mRNAlevels versus unstimulated cells (p G 0.1 for both) (Fig. 4A, B).
Mycobacterium leprae also significantly increased Col1A2 and Fn1 expression (p G 0.1 for both) (Fig. 4A, B). Pretreatment with SB-431542 significantly inhibited the expression of Col1A2 and Fn1 (p G 0.01 for both) (Fig. 4A, B).
FIGURE 3. Transforming growth factor-A1 (TGF-A1) inducesthe expression of Sox9 and Zeb1 mRNAs in Schwann cells(SCs). ST88-14 human SCs were incubated with recombinanthuman TGF-A1 (10 ng/mL) or Mycobacterium leprae (ML)(50:1) and preincubated with the TGF-A1 type I receptor (ALK5) inhibitor (SB-431542) (10 Kmol/L), followed by TGF-A1or ML for 7 days. (A, B) Real-time RT-PCR was performed todetect Sox9 (A) and Zeb1 (B) mRNA expression. Results arerepresented by mean T SE of 5 or more independent experi-ments. NS, unstimulated cells; DMSO, vehicle control. Fried-man nonparametric test with Dunn posttest was used for statistical analysis in which * p G 0.1.
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 357
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 8/16
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.358
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 9/16
Qualitative analysis of ECM as evaluated by indirect ELISA showed that TGF-A1 and ML increased the secretionof both type I collagen and fibronectin in opposition to theunstimulated cells (p G 0.1 for both) (Fig. 4C, D). To confirmthe effects of ML-induced TGF-A1 on type I collagen andfibronectin secretion, SCs were pretreated with SB-431542and then incubated with TGF-A1 or ML for 7 days. Thesefindings also demonstrated that SB-431542 inhibited theeffect of TGF-A1 and ML on both type I collagen (p G 0.1for both) (Fig. 4C) and fibronectin secretions (p G 0.1 for
both) (Fig. 4D), demonstrating that the effect observed in the presence of ML is mainly caused by TGF-A1 production.
Immunofluorescence images confirmed changes in theamount and organization of type I collagen and fibronectinsecreted and deposited by SCs in contrast to the unstimulatedcells (Fig. 4E). Unstimulated cultures showed a few fibers of disorganized type I collagen and fibronectin, whereas in TGF-A1 Y and ML-stimulated SCs, an increase in the amount of type I collagen and fibronectin fibers was observed, althoughthey were generally more organized and at times had a parallel
orientation (Fig. 4E).
Distribution of ECM and Cellular Components inLeprosy Nerve Tissues
To determine the distribution of ECM components alongwith the presence of >-SMA Y positive cells and ‘‘fibroblast-like’’ cells in leprosy neural fibrosis, expression of GLUT-1(perineurial cell marker), CD34 (‘‘fibroblast-like’’ cell marker),and >-SMA (myofibroblast marker) in peripheral nerve speci-mens from control and leprosy patients were assessed. For this,all leprosy nerve specimens studied were AFB positive and the
patients had clinical and histologic evidence of neuropathy.The presence of AFB in the perineurium was detected in 5 of the 11 samples selected. In the perineurial and endoneurial
compartments of all 5 samples, a less fibrotic morphologicaspect was observed. The Table summarizes the histopatho-logic characteristics.
Leprosy is characterized as a chronic disease without cutaneous fibrosis, whereas the affected peripheral nerves havean excessive deposition of ECM components (39 Y 42). Our invitro data showed that the secretion of type I collagen and fi-
bronectin increased in ST88-14 SCs in the presence of ML. Toreveal the distribution of type I and III collagen and fibronectinin leprosy nerve tissues, double-immunofluorescence stainingwas performed. Immunofluorescent images showed expressionof GLUT-1, type I and III collagen, and fibronectin throughout
the epineurium, perineurium, and endoneurium in both controland leprosy nerves (Fig. 5A Y F). Leprosy nerve tissues hadhigher expression than controls of both type I and III collagenand fibronectin in both perineurium and endoneurium. In con-trast, in control nerves, higher levels of the abovementionedtend to occur more frequently in the endoneurial than the
perineurial area (Fig. 5).In normal nerves, >-SMA expression was only be
detected in the perineurium, in which some perineurial cellsshowed immunoreactivity (Fig. 6A). Smooth muscle cellssurrounding vessels also exhibited >-SMA immunoreactivityin controls and leprosy samples (Fig. 6A Y C). Leprosy nervetissues displayed >-SMA Y expressing cells in the thicker peri-neurium (asterisks) and blood vessel walls (arrows), whereasnone were observed in the endoneurium (Fig. 6B, C). In nor-mal nerves, only the perineurium showed immunoreactivity for GLUT-1 (Fig. 6F). However, in leprosy nerve tissues,
perineurial cells and some cells within the endoneurial area expressed GLUT-1 (asterisk and arrows) (Fig. 6G), suggesting
pericyte migration to the endoneurium, which could contribute
to neural fibrosis and/or microfasciculation (Fig. 6G). In normaland leprosy nerves, some perineurial cells express both >-SMAand GLUT-1 despite the absence of >-SMA Y positive cellsinside the endoneurium (Fig. 6H, I).
In leprosy nerve tissues, there were increased numbersof CD34-positive cells, principally in the endoneurial area,
but sometimes around microfascicles; this was not seen innormal nerve tissues (Fig. 6D, E). To correlate the presence of ‘‘fibroblast-like’’ cells (CD34-positive cells) and the deposi-tion of connective tissue in leprosy nerves, the volume densityof connective tissue and CD34-positive cells was also evalu-ated. The volume occupied by connective tissue (and thecorresponding neural fibrosis) was 96% higher in leprosynerve tissues than among the controls (65% T 17% vs 33% T
11%; p G 0.0001). The volume occupied by CD34-positivecells was at 76% significantly higher in leprosy nerves thanamong controls (37% T 12% vs 21% T 9%; p G 0.0001) (data not shown).
Expression of Some Transdifferentiation-RelatedGenes in Nerve Biopsy Specimens
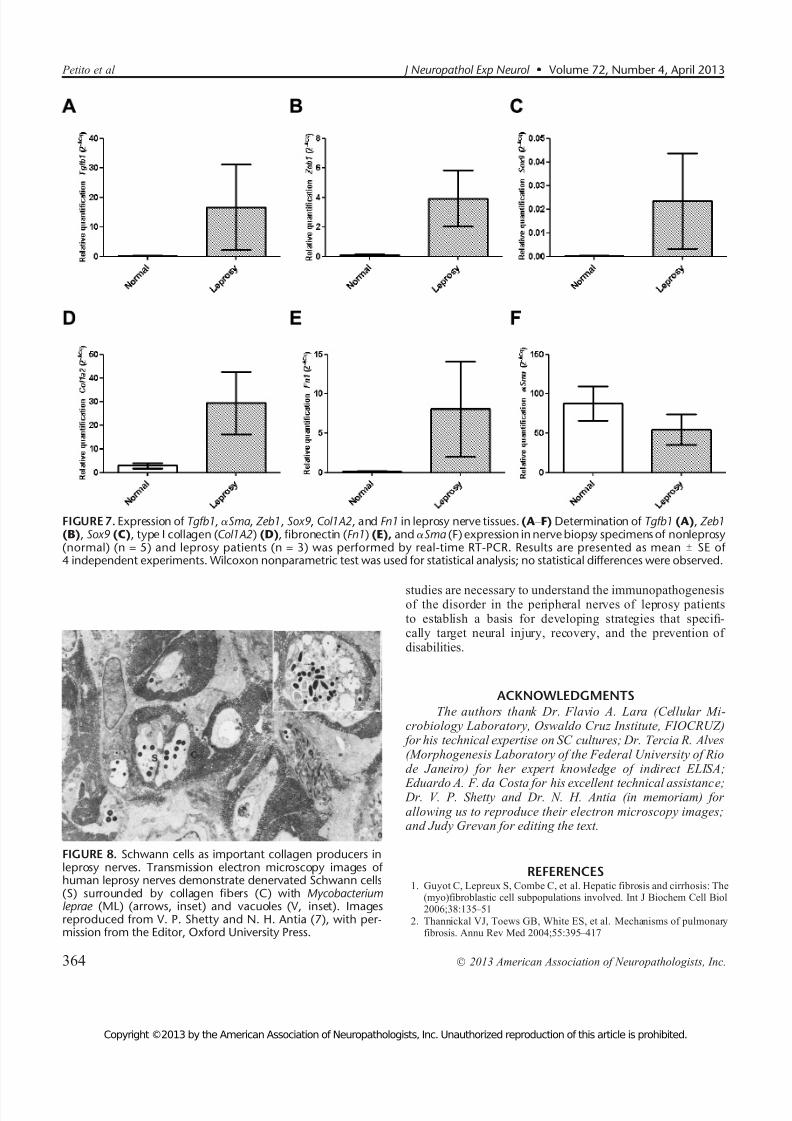
Because our in vitro results showed upregulation of Tgfb1, >Sma, Zeb1, Sox9, Col1a2, and Fn1 by TGF-A1 dur-ing SC-myofibroblastic transdifferentiation, fragments of nerve biopsies from 5 nonleprosy and 3 leprosy patients werealso processed and evaluated by real-time RT-PCR (Fig. 7).
FIGURE 4. Transforming growth factor-A1 (TGF-A1) induces type I collagen and fibronectin expression in human Schwann cells(SCs). ST88-14 SCs were incubated with recombinant human TGF-A1 (10 ng/mL) or Mycobacterium leprae (ML) (50:1) or preincubated with TGF-A1 type I receptor (ALK5) inhibitor SB-431542 (10 Kmol/L), followed by TGF-A1 or ML for 7 days. NS,unstimulated cells; DMSO, vehicle control. (A, B) Real-time RT-PCR was performed to measure the type I collagen ( Col1A2) (A)and fibronectin (Fn1) (B) mRNA expression. Results are represented by meanT SE of 4 or more independent experiments. Friedmannonparametric test with Dunn posttest was used for statistical analysis in which * p G 0.1. (C, D) Immobilized matrices wereinvestigated by indirect ELISA with anti Y type I collagen (C) and anti-fibronectin antibodies (D). Results are represented by mean T
SE of 5 independent experiments. Friedman nonparametric test with Dunn posttest was used for statistical analysis in which * p G
0.1. (E) Immunofluorescent images representative of type I collagen (top) and fibronectin (bottom) expression (red, Alexa Fluor 532) in ST88-14 SC cultures incubated with TGF-A1 or ML for 7 days. Results are representative of 5 independent experiments.Nuclei were labeled with DAPI. Scale bar = 50 Km.
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 359
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 10/16
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.360
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 11/16
Although the number of nerve samples at our disposal wasquite small, the results suggest that Tgfb1 (mean T SEM of normal vs leprosy: 0.1676 T 0.1579 vs 16.57 T 14.53), Zeb1(0.1201 T 0.02600 vs 3.924 T 1.895), Sox9 (0.0002170 T0.0001063 vs 0.02341 T 0.02032), Col1a2 (2.814 T 1.200 vs29.46 T 13.26), and Fn1 (0.1370 T 0.02561 vs 8.048 T 6.036)are upregulated in leprosy nerves that have fibrosis in compar-ison with what is typically found in normal nerves (Fig. 7A
Y E).
Although we observed >-SMA Y positive cells in leprosy nerves by immunohistochemistry and immunofluorescence assays, theexpression of >Sma in nonleprosy and leprosy specimens wasnot different (87.58 T 21.98 vs 54.41 T 19.52, respectively)(Fig. 7F).
DISCUSSIONThe most fascinating challenge in leprosy research lies
in elucidating the specific mechanisms by which bacterialinfection leads to nerve destruction. Nerve damage occursacross the entire spectrum of leprosy and may even progress
subsequent to clinically determined bacterial cure (43). Theaim of this study was to clarify some aspects of the complexinteractions among specific nerve cell components and MLthat may be critical to the development of nerve fibrosis. Anattempt was made to clarify the role of SC as an in vitro ML-induced profibrogenic cell. Nerve biopsy specimens were alsoexamined to gather enough information to support the con-tribution of different fibrogenic cells to nerve fibrosis in lep-rosy. In the present study, the presence of >-SMA Y positivecells and stress fibers in ST88-14 SC cultures stimulated withML and/or TGF-A1 strongly suggests the occurrence of SC-myofibroblastic transdifferentiation under TGF-A1 stimulusand the involvement of TGF-A1 in the production of ECMcomponents. Transcription factors ZEB1 and SOX9, which
are known to be enhanced in EMT, were also upregulatedunder both stimuli. Because no synergistic effect was shown,the minor effect of ML alone on ST88-14 transdifferentiationcompared with the exogenous effect obtained with TGF-A1was probably caused by the small amount of TGF-A1 pro-duced by SCs in response to ML. We previously demonstratedan increase of Tgfb1 and its receptors in ML-stimulated SCs(15). Confirming these results, our present investigationfound augmented Tgfb1 in ML-challenged SCs and alsodemonstrated that ML induced intracellular TGF-A1 and
protein secretion when stimulated with a lower bacterialconcentration. This raises the possibility that ML could am-
plify the TGF-A1 effect by inducing increased expression of its receptor even when there is a low bacterial level. Al-
though our results are also consistent with previous in vitrodata on quail SCs (26), caution is necessary when compar-ing these results because of differing experimental condi-
tions. Our data suggest that human SC transdifferentiationmay be one of the mechanisms involved in neural fibrosisin leprosy.
The role of SCs as collagen-producing cells has beeninferred by previous authors. Figure 8 shows denervated SCssurrounded by fibrillar collagen, suggesting that these cellsmay produce collagen (7). Ridley et al (44) implied that SCsmay play a role in collagen production in the leprosy nerve. Inanother study, the presence of collagen deposition in leprosynerve lesions was found to be very frequent (6). Perineurialfibrosis and endoneurial fibrosis were decisive findings indefining a leprosy diagnosis in histopathologic analyses of nerve biopsy samples, that is, multivariate analysis showedthat perineurial fibrosis could distinguish AFB-negative lep-rosy from nonleprosy nerves (45).
In our study, the vast majority of nerves had some degreeof fibrosis, but neither the presence nor the extent of fibrosiscorrelated with the number of lost nerve fibers or the amount of ML in the nerve. Among the 11 cases analyzed, 2 cases withfiber loss did not show any detectable endoneurial fibrosis. In
2 other cases, discrete fibrosis was seen in the absence of fiber loss when analyzed only by semithin sections. This suggeststhat the expansion of the collagen matrix may contribute
primarily to the progressive nerve fiber reduction observed at the end of leprosy neuropathy, principally at the lepromatousleprosy pole.
Both the absence of an adequate animal model capableof reproducing leprosy and the inability to grow ML in vitromake it almost impossible to construct a precise view of thishost-parasite interaction. The lengthy leprosy incubation pe-riod is also a deterrent to our ability to accompany the gradual
progression of the disease. What is left is the analysis of bi-opsy nerve samples taken at any one of the many stages of leprosy disease at a given time.
Several studies have extensively used the ST88-14 SCline as an in vitro model to understand ML-SC interaction toacquire applicable knowledge and deepen understanding of this disease (46, 47). Because the phenotype of this cell line isclose to that of nonmyelinating SCs (48), it has been used toanalyze ML entry and survival and its apparent escape fromthe bactericidal activity of macrophages (49).
It is known that TGF-A1 serves as an inhibitor of SC proliferation and myelination during development. On theother hand, TGF-A1 may promote the transition of SCs after
peripheral nerve injury to a proliferating nonmyelinating phenotype and thereby enhance the regenerative response(50). In addition to its role as an inducer of collagen and fi-
bronectin synthesis, it acts as a suppressor of inflammation
(51). Our data showing that ML can induce TGF-A1 produc-tion by SCs in the absence of inflammatory cells raise the pos-sibility that this cytokine (which was previously demonstrated
FIGURE 5. Distribution of type I and III collagen and fibronectin in leprosy nerve tissues. (A Y F) Double immunofluorescent imagesshow GLUT-1 Y positive cells (perineurial cell marker) and the expression of extracellular matrix components (type I and III collagenand fibronectin) in normal (left column) and leprosy nerve tissues (right column). GLUT-1 expression is observed in the perineuralarea (peri). The expression of type I collagen (A, B), type III collagen (C, D), and fibronectin (E, F) is mainly observed in theendoneurial area (endo). Type I and III collagen and fibronectin are in red (Alexa Fluor 532); GLUT-1 is in green (Alexa Fluor 633).Nuclei are labeled with DAPI. Results are representative of 2 normal and 6 leprosy cryostat samples. Scale bar = 100 Km.
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 361
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 12/16
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.362
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 13/16
in leprosy lesions [52]) could maintain a tolerant immune re-sponse in the nerve while inducing profibrogenic activity. Thisdynamic could explain the absence of acute neural symptomsin patients who have been seen to progress to complete nervefibrosis after long-term infection without having had any acuteneural symptoms.
There are several reports on TGF-A1 Y induced EMT indiverse cells by regulating the signaling of such moleculesand genetic programs as SOX9 and ZEB1 transcription fac-tors involved in scattering, migration, invasion, and control-ling the differentiation stages of SCs (23 Y 25). Roh et al (27)also reported on Sox10 downregulation and increased Sox9expression during rat SC-myofibroblast transdifferentiation.Our present results also suggest that TGF-A1 also contributesto human SC-myofibroblast transdifferentiation by regulatingthe expression of Sox9 and Zeb1 and the upregulation of their mRNA in leprosy nerve tissues, as has been described in other disease models.
Singh et al (41) showed that ML in vitro induces ex- pression of type I, III, and IV collagen in mouse SCs, but this
was not observed in another study on fibroblast cultures inmice (39). The authors concluded that collagen productionoccurs via increased deposition of these molecules by SCs.Our study has also demonstrated that TGF-A1 and ML induceincreased type I collagen and fibronectin mRNA, together with their secretion by SCs. We also previously showed that ML infection via SCs increased the production of MMP-2 andMMP-9, as well as their expression in leprosy nerve tissues,concluding that the imbalance between proteases and their respective inhibitors is an important mechanism of neural fi-
brosis and in support of the hypothesis that ML induces aninflammation-mediated remodeling of ECM (15).
Despite the convincing in vitro data revealing ECM pro-duction by SCs and SC transdifferentiation into >-SMA Y positive
cells, the main fibrogenic cell in vivo has not been proven. In thenerve biopsy specimens included in our study, >-SMA Y positivecells were only seen to have accumulated in layers of the peri-neurium and not in the endoneurium itself. The expansion or migration of constitutively >-SMA Y positive cells from the
perineurium could occur during the long process of ECM re-modeling and nerve destruction. That myofibroblasts derivedfrom transdifferentiated SCs participate in triggering a fibrogenicresponse in the early stages of infection cannot be ruled out inview of the fact that all the biopsy specimens under study weretaken from patients whose clinical neuropathy had alreadystalled. On the other hand, the numbers of other fibrogenic
cell Y like CD34-positive fibroblasts were found to be not onlysignificantly higher in fibrotic leprosy nerves versus normalspecimens but were augmented in fibrotic lesions within theendoneurium. Our results suggest that multiple sources of fibrogenic cells within the nerves are responding to ML infec-tion. Transdifferentiated SCs, >-SMA Y positive cells in the
perineurium, pericytes, and endoneurial fibroblasts would all becandidates for this role.
The unexpected greater amount of >-SMA mRNA innormal samples versus leprosy nerves may be caused by manyfactors, including the size of the tissue specimen collected, theamount of vessels present in the samples, and the stage of leprosy nerve fibrosis. Although all these factors could in part explain the result, the stability of each phenotype in a verydynamic repairing environment may be questionable particu-larly because the end stage of the fibrotic process is an acel-lular collagen structure.
It is known that 3 local events are needed to generate >-SMA Y positive cells in vivo: TGF-A1 accumulation, the pres-ence of specialized ECM proteins, and intense extracellular
stress leading up to the mechanical properties of ECM.Although no mechanosensing has been reported in the pe-ripheral nerve structure, it is widely accepted that, in leprosy,the preferred localization of lesions in leprosy, that is, associ-ated with the superficial peripheral nerves, is the result of theconstant mechanical stress to which these nerves are subjected.Under healthy conditions, the multilayered perineurium mod-ulates the external stretching forces that could be extremelyharmful to the nerve, whereas in leprosy, the perineuriumseems to contribute to the increasing intraneural pressure andconsequently to edema and the inflammatory swelling of theendoneurium (53).
Bunge et al (54) suggested that perineurial cells may bemyofibroblast-like cells on account of their contractile cell-like
morphologic characteristics. In our study, >-SMA Y
positivecells were only found in the perineurium. On the other hand,myofibroblastic cells had previously only been detected innormal peripheral nerves and regenerated nerve trunks inrats (55). To our knowledge, this is the first description of >-SMA Y positive cells in normal human adult nerves. Nonethe-less, their precise role in peripheral nerve pathology remainselusive.
Together, our results suggest that ML-infected SCsundergo phenotypical changes and even death as a result of inflammatory mediators, leading them to secrete ECM that contributes to progressive nerve fiber loss and fibrosis. Future
FIGURE 6. Distribution of >-smooth muscle actin (>-SMA) Y
positive and CD34-positive cells in leprosy nerves. (A) In normal nerves,>-SMA expression is observed in the perineurium (asterisk). (B, C) In leprosy nerves, >-SMA is expressed in a thicker perineurium(asterisk and arrowheads) and the smooth muscle cells of blood vessels (arrow). (D) Normal nerve immunostained for CD34(arrows). (E) In leprosy nerves, a higher number of CD34-positive cells (arrows) is seen within the endoneurial area. The insetdemonstrates that perineurium is not immunostained for CD34 (asterisk). (F) In a normal nerve, GLUT-1 expression is distributed inthe perineurium (asterisk). (G) In leprosy nerve tissue, GLUT-1 expression is detected in perineurial cells (asterisks) and additionallyin many endoneurial cells (arrows). Photomicrographs are representative of 8 normal and 8 leprosy nerve tissues. (H, I) Double-immunofluorescent images showed that GLUT-1 Y positive cells are mostly >-SMA positive and are mainly distributed within theperineurium in both normal (H) and leprosy (I) nerves. In leprosy nerves, there is an increase of >-SMA Y positive cells within theperineurium (in detail); some cells demonstrate stress fibers. Alpha-SMA (red, Alexa Fluor 532) is double immunostained withGLUT-1 antibody (green, Alexa Fluor 633). Nuclei are labeled with DAPI. Double-positive cells are yellow. Data shown are repre-sentative of 2 normal and 6 leprosy cryostat nerve samples. Peri, perineurium; endo, endoneurium. Scale bar = 100 Km.
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 363
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.
7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 14/16
studies are necessary to understand the immunopathogenesisof the disorder in the peripheral nerves of leprosy patientsto establish a basis for developing strategies that specifi-cally target neural injury, recovery, and the prevention of disabilities.
ACKNOWLEDGMENTSThe authors thank Dr. Flavio A. Lara (Cellular Mi-
crobiology Laboratory, Oswaldo Cruz Institute, FIOCRUZ) for his technical expertise on SC cultures; Dr. Tercia R. Alves(Morphogenesis Laboratory of the Federal University of Riode Janeiro) for her expert knowledge of indirect ELISA;
Eduardo A. F. da Costa for his excellent technical assistance; Dr. V. P. Shetty and Dr. N. H. Antia (in memoriam) for allowing us to reproduce their electron microscopy images;and Judy Grevan for editing the text.
REFERENCES1. Guyot C, Lepreux S, Combe C, et al. Hepatic fibrosis and cirrhosis: The
(myo)fibroblastic cell subpopulations involved. Int J Biochem Cell Biol2006;38:135 Y 51
2. Thannickal VJ, Toews GB, White ES, et al. Mechanisms of pulmonaryfibrosis. Annu Rev Med 2004;55:395 Y 417
FIGURE 7. Expression of Tgfb1, >Sma , Zeb1, Sox9, Col1A2, and Fn1 in leprosy nerve tissues. (A Y F) Determination of Tgfb1 (A), Zeb1(B), Sox9 (C), type I collagen (Col1A2) (D), fibronectin (Fn1) (E), and >Sma (F) expression in nerve biopsy specimens of nonleprosy(normal) (n = 5) and leprosy patients (n = 3) was performed by real-time RT-PCR. Results are presented as mean T SE of 4 independent experiments. Wilcoxon nonparametric test was used for statistical analysis; no statistical differences were observed.
FIGURE 8. Schwann cells as important collagen producers inleprosy nerves. Transmission electron microscopy images of human leprosy nerves demonstrate denervated Schwann cells(S) surrounded by collagen fibers (C) with Mycobacteriumleprae (ML) (arrows, inset) and vacuoles (V, inset). Imagesreproduced from V. P. Shetty and N. H. Antia (7), with per-mission from the Editor, Oxford University Press.
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.364
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 15/16
3. Varga J, Abraham D. Systemic sclerosis: A prototypic multisystem fi- brotic disorder. J Clin Invest 2007;117:557 Y 67
4. Hinz B, Phan SH, Thannickal VJ, et al. Recent developments inmyofibroblast biology: Paradigms for connective tissue remodeling. AmJ Pathol 2012;180:1340 Y 55
5. Amadeu TP, Braune AS, Porto LC, et al. Fibrillin-1 and elastin are dif-ferentially expressed in hypertrophic scars and keloids. Wound Repair Regen 2004;12:169 Y 74
6. Chimelli L, Freitas M, Nascimento O. Value of nerve biopsy in the di-agnosis and follow-up of leprosy: The role of vascular lesions and use-fulness of nerve studies in the detection of persistent bacilli. J Neurol1997;244:318 Y 23
7. Shetty V. Pathology of nerve damage in leprosy. In: Antia N, Shetty V,eds. The Peripheral Nerve in Leprosy and Other Neuropathies. Calcutta,India: Oxford University Press, 1997:79 Y 137
8. Van Brakel WH, Khawas IB. Nerve damage in leprosy: An epidemio-logical and clinical study of 396 patients in west Nepal V Part 1. Defini-tions, methods and frequencies. Lepr Rev 1994;65:204 Y 21
9. Awatramani R, Shumas S, Kamholz J, et al. TGFbeta1 modulates the phenotype of Schwann cells at the transcriptional level. Mol Cell Neurosci 2002;19:307 Y 19
10. Shubayev VI, Angert M, Dolkas J, et al. TNFalpha-induced MMP-9 promotes macrophage recruitment into injured peripheral nerve. Mol Cell Neurosci 2006;31:407 Y 15
11. Oliveira AL, Antunes SL, Teles RM, et al. Schwann cells producingmatrix metalloproteinases under Mycobacterium leprae stimulation may
play a role in the outcome of leprous neuropathy. J Neuropathol Exp Neurol 2010;69:27 Y 39
12. Verrecchia F, Mauviel A. Transforming growth factor-beta signalingthrough the Smad pathway: Role in extracellular matrix gene expressionand regulation. J Invest Dermatol 2002;118:211 Y 15
13. Goulart IM, Mineo JR, Foss NT. Production of transforming growthfactor-beta 1 (TGF-beta1) by blood monocytes from patients with dif-ferent clinical forms of leprosy. Clin Exp Immunol 2000;122:330 Y 34
14. Kiszewski CA, Becerril E, Baquera J, et al. Expression of transforminggrowth factor-beta isoforms and their receptors in lepromatous and tu-
berculoid leprosy. Scand J Immunol 2003;57:279 Y 85
15. Oliveira RB, Sampaio EP, Aarestrup F, et al. Cytokines and Mycobac-terium leprae induce apoptosis in human Schwann cells. J NeuropatholExp Neurol 2005;64:882 Y 90
16. Hinz B. Formation and function of the myofibroblast during tissue repair.
J Invest Dermatol 2007;127:526 Y
3717. Buttle DJ. Factors controlling matrix turnover in health and disease.
Biochem Soc Trans 2007;35:643 Y 46
18. Desmouliere A, Chaponnier C, Gabbiani G. Tissue repair, contraction,and the myofibroblast. Wound Repair Regen 2005;13:7 Y 12
19. Hornick JL, Fletcher CD. Intestinal perineuriomas: Clinicopathologicdefinition of a new anatomic subset in a series of 10 cases. Am J SurgPathol 2005;29:859 Y 65
20. Pina-Oviedo S, Ortiz-Hidalgo C. The normal and neoplastic perineurium:A review. Adv Anat Pathol 2008;15:147 Y 64
21. Radisky DC, Kenny PA, Bissell MJ. Fibrosis and cancer: Do myofibro- blasts come also from epithelial cells via EMT? J Cell Biochem 2007;101:830 Y 39
22. Willis BC, Liebler JM, Luby-Phelps K, et al. Induction of epithelial-mesenchymal transition in alveolar epithelial cells by transforminggrowth factor-beta1: Potential role in idiopathic pulmonary fibrosis. Am JPathol 2005;166:1321 Y 32
23. Lee K, Nelson CM. New insights into the regulation of epithelialmesenchymal transition and tissue fibrosis. Int Rev Cell Mol Biol 2012;294:171 Y 221
24. Arima Y, Hayashi H, Kamata K, et al. Decreased expression of neurofibromin contributes to epithelial-mesenchymal transition in neu-rofibromatosis type 1. Exp Dermatol 2010;19:136 Y 41
25. Shirakihara T, Saitoh M, Miyazono K. Differential regulation of epi-thelial and mesenchymal markers by deltaEF1 proteins in epithelial mes-enchymal transition induced by TGF-beta. Mol Biol Cell 2007;18:3533 Y 44
26. Real C, Glavieux-Pardanaud C, Vaigot P, et al. The instability of theneural crest phenotypes: Schwann cells can differentiate into myofibroblasts.Int J Dev Biol 2005;49:151 Y 59
27. Roh J, Cho EA, Seong I, et al. Downregulation of Sox10 with specificsmall interfering RNA promotes transdifferentiation of Schwannoma cells into myofibroblasts. Differentiation 2006;74:542 Y 51
28. Jardim MR, Antunes SL, Santos AR, et al. Criteria for diagnosis of pureneural leprosy. J Neurol 2003;250:806 Y 9
29. van Brakel WH, Khawas IB. Silent neuropathy in leprosy: An epidemi-ological description. Lepr Rev 1994;65:350 Y 60
30. Marques MA, Neves-Ferreira AG, da Silveira EK, et al. Deciphering the
proteomic profile of Mycobacterium leprae cell envelope. Proteomics2008;8:2477 Y 91
31. Moraes MO, Sarno EN, Almeida AS, et al. Cytokine mRNA expressionin leprosy: A possible role for interferon-gamma and interleukin-12 inreactions (RR and ENL). Scand J Immunol 1999;50:541 Y 49
32. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method.Methods 2001;25:402 Y 8
33. Waghabi MC, Coutinho CM, Soeiro MN, et al. Increased Trypanosomacruzi invasion and heart fibrosis associated with high transforminggrowth factor beta levels in mice deficient in alpha(2)-macroglobulin.Infect Immun 2002;70:5115 Y 23
34. Alves TR, da Fonseca AC, Nunes SS, et al. Tenascin-C in the extracellular matrix promotes the selection of highly proliferative and tubulogenesis-defective endothelial cells. Exp Cell Res 2011;317:2073 Y 85
35. Neves RH, Alencar AC, Aguila MB, et al. Hepatic stereology of Schis-tosomiasis mansoni Y infected mice fed a high-fat diet. Mem Inst OswaldoCruz 2006;101:253
Y 60
36. Mandarim-de-Lacerda CA. Stereological tools in biomedical research.An Acad Bras Cienc 2003;75:469 Y 86
37. Casella GT, Bunge RP, Wood PM. Improved method for harvestinghuman Schwann cells from mature peripheral nerve and expansion invitro. Glia 1996;17:327 Y 38
38. Tapinos N, Ohnishi M, Rambukkana A. ErbB2 receptor tyrosine kinasesignaling mediates early demyelination induced by leprosy bacilli. Nat Med 2006;12:961 Y 66
39. Singh N, Birdi TJ, Antia NH. Collagen profile in sciatic nerves of M. leprae Y inoculated mice correlates with in vitro collagen production by Schwann cells. Int J Lepr Other Mycobact Dis 1998;66:238 Y 40
40. Junqueira LC, Montes GS, Neto EA, et al. The collagen of permanentlydamaged nerves in human leprosy. Int J Lepr Other Mycobact Dis 1980;48:291 Y 97
41. Singh N, Birdi TJ, Chandrashekar S, et al. Schwann cell extracellular
matrix protein production is modulated by Mycobacterium leprae andmacrophage secretory products. J Neurol Sci 1997;151:13 Y 22
42. Antunes SL, Gallo ME, de Almeida SM, et al. Dermal extracellular ma-trix in cutaneous leprosy lesions. Int J Lepr Other Mycobact Dis 1999;67:24 Y 35
43. Wilder-Smith EP, Van Brakel WH. Nerve damage in leprosy and itsmanagement. Nat Clin Pract Neurol 2008;4:656 Y 63
44. Ridley MJ, Waters MF, Ridley DS. Events surrounding the recognition of Mycobacterium leprae in nerves. Int J Lepr Other Mycobact Dis 1987;55:99 Y 108
45. Antunes SL, Chimelli L, Jardim MR, et al. Histopathological examinationof nerve samples from pure neural leprosy patients: Obtaining maximuminformation to improve diagnostic efficiency. Mem Inst Oswaldo Cruz2012;107:246 Y 53
46. Teles RM, Krutzik SR, Ochoa MT, et al. Interleukin-4 regulates the ex- pression of CD209 and subsequent uptake of Mycobacterium leprae bySchwann cells in human leprosy. Infect Immun 2010;78:4634 Y 43
47. Mattos KA, Oliveira VG, D’Avila H, et al. TLR6-driven lipid droplets in Mycobacterium leprae Y infected Schwann cells: Immunoinflammatory platforms associated with bacterial persistence. J Immunol 2011;187:2548 Y 58
48. da Silva TP, da Silva AC, Baruque M da G, et al. Morphological andfunctional characterizations of Schwann cells stimulated with Mycobac-terium leprae. Mem Inst Oswaldo Cruz 2008;103:363 Y 69
49. Aung T, Kitajima S, Nomoto M, et al. Mycobacterium leprae in neuronsof the medulla oblongata and spinal cord in leprosy. J Neuropathol Exp
Neurol 2007;66:284 Y 94
50. Einheber S, Hannocks MJ, Metz CN, et al. Transforming growthfactor-beta 1 regulates axon/Schwann cell interactions. J Cell Biol 1995;129:443 Y 58
J Neuropathol Exp Neurol Volume 72, Number 4, April 2013 Fibrogenic Cells in Leprosy Neuropathy
Ó 2013 American Association of Neuropathologists, Inc. 365
Copyright ©2013 by the American Association of Neuropathologists, Inc. Unauthorized reproduction of this article is prohibited.

7/30/2019 Petito et al 2013.pdf
http://slidepdf.com/reader/full/petito-et-al-2013pdf 16/16
51. Kitani A, Fuss I, Nakamura K, et al. Transforming growth factor (TGF)-beta1 Y producing regulatory T cells induce Smad-mediated inter-leukin 10 secretion that facilitates coordinated immunoregulatory activityand amelioration of TGF-beta1 Y mediated fibrosis. J Exp Med 2003;198:1179 Y 88
52. Simoes Quaresma JA, de Almeida FA, de Souza Aarao TL, et al.Transforming growth factor A and apoptosis in leprosy skin lesions: Pos-sible relationship with the control of the tissue immune response in the
Mycobacterium leprae infection. Microbes Infect 2012;14:696 Y
701
53. Dastur DK, Manghani DK, Elavia JF. Therapeutic implications of morphological study of nerves from treated or untreated tuberculoidor lepromatous leprosy. Int J Lepr Other Mycobact Dis 1991;59:122 Y 24
54. Bunge MB, Wood PM, Tynan LB, et al. Perineurium originates fromfibroblasts: Demonstration in vitro with a retroviral marker. Science1989;243:229 Y 31
55. Chamberlain LJ, Yannas IV, Hsu HP, et al. Connective tissue response totubular implants for peripheral nerve regeneration: The role of myofibro-
blasts. J Comp Neurol 2000;417:415 Y
30
Petito et al J Neuropathol Exp Neurol Volume 72, Number 4, April 2013
Ó 2013 American Association of Neuropathologists, Inc.366





























