Embed Size (px)
Citation preview

ISSN 0026�8933, Molecular Biology, 2014, Vol. 48, No. 3, pp. 439–447. © Pleiades Publishing, Inc., 2014.Original Russian Text © D.G. Naumoff, A.A. Ivanova, S.N. Dedysh, 2014, published in Molekulyarnaya Biologiya, 2014, Vol. 48, No. 3, pp. 508–517.
439
INTRODUCTION
Glycoside hydrolases are a wide group of enzymesthat catalyze the hydrolysis of the O�glycosidic bond.Great variety of these enzymes is determined by bigdiversity of their natural substrates; currently, almost200 corresponding enzymatic activities are known(EC 3.2.1.1–EC 3.2.1.195). Based on the homology ofthe catalytic domains, glycoside hydrolases are classi�fied in 133 families (GH1–GH133) according to theCAZy database [1]. Glycoside hydrolase genes andtheir close homologues are found in genomes of themost living organisms; they compose approximately1% of all protein�coding genes. Several common fea�tures are typical for the gene evolution of glycosidehydrolases: they often undergo duplications, elimina�tions, and horizontal transfer [2]. As a result, thetopologies of their phylogenetic trees and ribosomalRNA gene trees are often significantly different.
Endo�β�xylanases (EC 3.2.1.8 and EC 3.2.1.32)are a widespread and industrially important group ofglycoside hydrolases [3–10]. They are responsible fordegradation of heteroxylans that are structurally aquite heterogeneous group of polysaccharides, whichcompose the basis of hemicelluloses in the higherplant cell wall. Currently known endo�β�xylanases areclassified in 13 glycoside hydrolase families in theCAZy database [1]. However, only two families of pro�teins (GH10 and GH11) have no other enzymaticactivities, except endo�β�xylanase. An exception issome GH10�enzymes of Ascomycota with the tomati�nase activity. This suggests that other uncharacterizedproteins of these two families have the same activity.The majority of biochemically characterized endo�β�
xylanases belong to these two families (about 500 pro�teins of various origin). The corresponding enzymesdigest their substrates with retaining the anomericconfiguration in the product [1]. The characteristicfeature of GH10 endo�β�xylanases is a broad substratespecificity [10].
The planctomycetes is a phylum of bacteria thathave unique morphological, ultrastructural, and bio�chemical features [11–14]. Due to difficulties inobtaining pure cultures of these bacteria, the number ofcharacterized Planctomycetes is quite limited. Cur�rently there are two orders in the phylum Plancto�mycetes: Planctomycetales, which includes 15 speciesof validly described heterotrophic Planctomycetes, and‘Candidatus Brocadiales’, represented by autotrophicanaerobic anammox (ANaerobic AMMonium OXida�tion) Planctomycetes [11]. The functional potential ofthese microorganisms is poorly characterized, despitetheir wide environmental distribution [14, 15]. Thereis no doubt that anammox Planctomycetes play animportant role in the global nitrogen cycle and thecleaning of wastewater [14, 16]. However, the func�tional capabilities of heterotrophic members of thisbacterial phylum are not so obvious.
Sphagnum peat bog is one of the habitats for Planc�tomycetes where they are an important microbialcommunity component, i.e., they comprise up to 13%of the total bacterial number [15, 17]. Planctomycetesare one of the dominant bacterial groups in decompos�ing Sphagnum [18]. Based on the available data, itcould be suggested that these microorganisms are animportant component of the hydrolytic microbialcommunities of peat bogs. Some peat�inhabitingmembers of the Planctomycetes were obtained in pure
Phylogeny of β�Xylanases from PlanctomycetesD. G. Naumoff, A. A. Ivanova, and S. N. Dedysh
Winogradsky Institute of Microbiology, Russian Academy of Sciences, Moscow, 117312 Russia;e�mail: [email protected]
Received November 15, 2013; in final form, December 3, 2013
Abstract—Here, we present the results of a computational analysis of a group of hypothetical GH10 endo�β�xylanases from the Planctomycetes, a bacterial phylum with poorly characterized functional capabilities.These proteins are encoded in all analyzed genomes of heterotrophic Planctomycetes and form a phylogenet�ically distinct and tight cluster. In addition, we determined nucleotide sequences for endo�β�xylanase genesfrom five strains of Isosphaera�Singulisphaera group of the Planctomycetes. The trees constructed for the 16SrRNA genes and the inferred amino acid sequences of endo�β�xylanases were highly congruent, thus suggest�ing the vertical transfer of endo�β�xylanase genes and their functional importance in Planctomycetes.
DOI: 10.1134/S0026893314030145
Keywords: Planctomycetes, Singulisphaera, Isosphaera, glycoside hydrolase, endo�glycosidase, β�xylanase,GH10 family, CAZy, hierarchical protein classification, protein evolution, protein phylogenetic tree, searchof homologues
BIOINFORMATICS
UDC 575.852'112:577.152.321:579.833.6

440
MOLECULAR BIOLOGY Vol. 48 No. 3 2014
NAUMOFF et al.
cultures and characterized as novel genera and species[19–24]. These microorganisms are capable of hydro�lyzing xylan [19–23], pectin [19–23], cellulose [23],and several heteropolysaccharides [19–23]. In thenucleotide sequence databases, there are genomes ofseveral Planctomycetes, including peat�inhabitingrepresentatives. This allows one to check whether theyhave genes that are responsible for the degradation ofpolymeric substrates. The present paper aims to searchfor endo�β�xylanase genes.
According to the CAZy database [1], glycosidehydrolase genes of 12 out of the 13 families, whichcontain endo�β�xylanases, are either presented onlyin some Planctomycetes genomes, or totally absent.The only exception is the GH10 family, which is notfound only in ‘Candidatus Kuenenia stuttgartiensis’and Pirellula staleyi DSM6068 [1]. We examined theposition of hypothetical (because their enzymaticactivity in vitro was not measured) endo�β�xylanasesfrom the Planctomycetes on the phylogenetic tree ofthe catalytic domains of the GH10 family of glycosidehydrolases.
EXPERIMENTAL
Searching for Proteins in the Database
Amino acid sequences of the Planctomycetes pro�teins from the GH10 family of glycoside hydrolaseslisted in the CAZy database [1] were used to search forthe closest homologues in the GenPept database(“non�redundant protein sequences” section) by theblastp program. About 100 of the closest homologuesof each Planctomycetes protein were used for multiplealignments and subsequent phylogenetic analysis. Wegenerally did not use closely related proteins of strainsfrom the same species and species from the samegenus.
Possible Planctomycetes proteins among metage�nomic sequences were being searched by screeningthe GenPept database (“metagenomic proteins” sec�tion) by the blastp program, wherein sequence of oneof the Phycisphaera mikurensis protein (GenPept,BAM04356.1) was used as a query.
Analysis of Amino Acid Sequences
Multiple sequence alignment was produced manuallywith the BioEdit program (http://www.mbio.ncsu.edu/BioEdit/bioedit.html). Results of pairwise alignmentwith PSI�BLAST were taken into account.
After removing of the most variable regions, multiplesequence alignment was used to build the phylogenetictrees by the PROTPARS program (protein sequence par�simony method, MP) and the NEIGHBOR program(neighbor�joining method, NJ) in the PHYLIP package(http://evolution.gs.washington.edu/phylip.html). Theconfidence limits for each node were estimated by 100or 1000 bootstrap replicates. The TreeView Win32
program was used to obtain graphical images of trees(http://taxonomy.zoology.gla.ac.uk/rod/treeview.html).
Strains
We used strains of peat�inhabiting Planctomycetesof the Isosphaera�Singulisphaera group, namely, thepreviously described Singulisphaera acidiphilaMPL1015 [20] and ‘Candidatus Nostocoida acid�iphila’ OB1 [24], as well as three taxonomicallyuncharacterized strains of the Planctomycetes (Isos�phaera sp. PX4, S. acidiphila BW, and S. acidiphilaPX31), which were isolated from northern wetlands ofRussia and provided by Irina S. Kulichevskaya (Wino�gradsky Institute of Microbiology, Russian Academyof Sciences) as a courtesy.
Extraction of DNA and Polymerase Chain Reaction
The standard method was used for DNA extractionfrom the planctomycete cells [25]; this DNA was usedas a template in polymerase chain reaction. The PCRamplification of fragments of 16S rRNA genes (about1400 bp) was performed using universal bacterialprimers 9f and 1492r [26] by thermocycler PE Gene�Amp PCR System 9700 Perkin�Elmer (Applied Bio�systems, United States).
Specific primers were used for the PCR amplifi�cation of fragments (about 750 bp) of endo�β�xyla�nase genes: direct IsoF–5'�GCT(G/C)AAC�GAC(G/C)TGCG(C/T)AACCA�3' and reverse IsoR–5'�ATTCGGCCCA(G/A)GGGCG(G/C)TCGA�3'.These primers were designed based on nucleotidesequences of genes of the endo�β�xylanases fromGH10 family of the type strains of Isosphaera pallidaATCC43644 (GenBank, CP002353.1) and Sin�gulisphaera acidiphila DSM18658 (CP003364.1). Thefinal concentration of the primers in the reaction mixwas 0.3 mM. The temperature of primer annealing was64°С.
DNA Cloning and Sequencing
Amplicons of the 16S rRNA genes and endo�β�xylanase genes were cloned using the pGem�T EasyVector System II (Promega, United States) accordingto the manufacturer’s recommendations. Recombi�nant clones were checked for insert size by amplifica�tion of cloning fragments with vector�specific T7 andSP6 primers. Plasmid DNA was extracted using theWizard® Plus Minipreps DNA Purification System kit(Promega). Nucleotide sequences were determinedusing ABI 377A Perkin�Elmer sequencer (AppliedBiosystems).
Nucleotide Sequence Analysis
16S rRNA gene multiple nucleotide sequencealignment and building of the phylogenetic tree by the

MOLECULAR BIOLOGY Vol. 48 No. 3 2014
PHYLOGENY OF β�XYLANASES FROM PLANCTOMYCETES 441
Neighbor�Joining method was done using the ARBprogram package (http://www.arb�home.de). The con�fidence limits for each node were estimated by 1000bootstrap replicates (the PHYLIP program package).Nucleotide sequences of genes of GH10 family of gly�coside hydrolases from BW, MPL1015, OB1, PX31,and PX4 strains, as well as nucleotide sequences of 16SrRNA genes from BW, PX31, and PX4 strains, aredeposited in the GenBank database under accessionnumbers KF467521�KF467528, respectively.
RESULTS
Searching for Planctomycetes Genes That Encode Hypothetical Endo�β�Xylanases with Catalytic
Domains of the GH10 Family of Glycoside Hydrolases
When we began our research in the summer of2012, the domain of the GH10 family of glycosidehydrolases was found in six proteins encoded in thefour genomes of the following Planctomycetes: Isos�phaera pallida ATCC43644 (GenPept, ADV61272.1),Planctomyces brasiliensis DSM5305 (ADY58863.1),P. limnophilus DSM3776 (ADG69659.1) and Rhodop�irellula baltica SH1 (CAD76104.1, CAD78872.1, andCAD76526.1) according to the CAZy database [1]. Atpresent, additional four Phycisphaera mikurensisNBRC102666 proteins (BAM03104.1, BAM03106.1,BAM03107.1, and BAM04877.1) and one proteinfrom an uncultured Planctomycetales bacteriumHF0500_02G17 (ADI22410.1) are included into theCAZy database.
We screened the GenPept database using severaldomains of the GH10 family, which yielded 3672 pro�teins that contain domains of this family. Here, extraparalogues were found encoded in some of the abovePlanctomycetes genomes, including one in R. balticaSH1 (CAD71336.1) and two in P. mikurensisNBRC102666 (BAM03393.1 and BAM04356.1). More�over, one or several proteins of the family were found inseveral other Planctomycetes: Blastopirellula marinaDSM3645 (EAQ81247.1), Gemmata obscuriglobusUQM2246 (WP_010033755.1 and WP_010037050.1),Pirellula staleyi DSM6068 (ADB18958.1), Planctomycesmaris DSM8797 (EDL61831.1), R. baltica SH28(EKJ98661.1, EKJ99654.1, EKK00284.1, EKK01796.1,and EKK03334.1), R. baltica SWK14 (ELP30803.1,ELP32534.1, ELP33066.1, ELP33967.1, andELP34476.1), R. baltica WH47 (EGF24716.1,EGF29139.1, EGF29421.1, EGF29484.1, andEGF27587.1), ‘R. europaea’ 6C (EMB13996.1,EMB14634.1, EMB16514.1, EMB17801.1,EMB17802.1, and EMB18096.1), ‘R. europaea’SH398 (EMI25130.1, EMI25179.1, EMI25518.1,EMI26865.1, and EMI27338.1), ‘R. maiorica’ SM1(EMI16935.1 and EMI19362.1), ‘R. sallentina’ SM41(EMI51840.1, EMI52671.1, EMI53465.1, andEMI54157.1), Rhodopirellula sp. SWK7 (EMI42248.1,EMI44021.1, and EMI45012.1), Schlesneria paludicolaDSM18645 (WP_010583456.1), Singulisphaera acid�
iphila DSM18658 (AGA27804.1), and Zavarzinellaformosa DSM19928 (WP_020472577.1). However,amino acid sequences of an orthologous pair from‘R. europaea’ 6C and ‘R. europaea’ SH398 (EMB14634.1and EMI25518.1) were identical. Genes that encodeproteins of the GH10 family were not found in par�tially sequenced genomes of KSU�1 Planctomycetes(GenBank, NZ_BAFH00000000.1) and anammoxplanctomycete ‘Candidatus Kuenenia stuttgartiensis’(GenBank, CT030148.2 and CT573071.1–CT573074.1).
Among metagenomic proteins (with E�value ≤ 10–7),five non�full�length representatives were found thatprobably belong to Planctomycetes. Only one of them(GenPept, EBE27272.1) has a full domain of the GH10family. We used it in the multiple sequence alignment andfurther phylogenetic analysis. Another four proteins(EBL67503.1, EBQ98517.1, EBR14041.1, andECA07557.1) were excluded from subsequent analy�sis.
Comparative Analysis of Amino Acid Sequences of Planctomycetes Endo�β�Xylanases: Detection
of Two Subfamilies of GH10 Family
Pairwise comparison of amino acid sequences ofthe Planctomycetes proteins containing domain of theGH10 family of glycoside hydrolases allowed us tocombine them into two groups by degree of similarity.One of these groups contains one protein from eachorganism (as well as all five found metagenomic pro�teins). These proteins are the closest homologues toone another and could be considered as a distinct sub�family composed of the GH10 family of glycosidehydrolases. Studying this subfamily is the primary pur�pose of this research. The second group contains pro�teins from some Planctomycetes species only andusully is presented by several paralogues. The numberof paralogues could be different, even in strains of onespecies, as has been shown for Rhodopirellula balticaand ‘R. europaea’. Proteins of the Planctomycetesfrom this group are not always the closest homologuesfor each other. They could be considered to be mem�bers of another subfamily of the GH10 family of glyco�side hydrolases; this subfamily is more heterogeneousand contains numerous proteins from other organ�isms, including members from different phyla of bac�teria, as well as some Eukaryotes and Archaea. One ofthe Phycisphaera mikurensis proteins (BAM03393.1)most probably did not belong to either of these twogroups. It is one of the most divergent members of theGH10 family and apparently does not have any enzy�matic activity (data not shown).
The multiple alignment of 112 amino acidsequences was produced for phylogenetic analysis ofproteins from the first group. The alignment includednot only 20 Planctomycetes proteins but also theirclosest homologues from other living organisms.Results of the analysis confirmed the existence of adistinct subfamily that is formed by proteins of Planc�

442
MOLECULAR BIOLOGY Vol. 48 No. 3 2014
NAUMOFF et al.
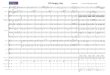
tomycetes. The cluster of Planctomycetes has 100 and99% support, respectively, on the trees that were builtby the maximum parsimony method (MP, Fig. 1) andneighbor�joining method (NJ, figure not shown). Aprotein (GenPept, EBE27272.1) from marine metage�nome falls into this cluster along with 20 proteins of thePlanctomycetes. The proteins of phyla of Cyanobacteria(ABW28526.1, ACK64415.1, ACU99283.1, andADN16869.1), Firmicutes (AGC67515.1, EEQ57087.1,EGN35770.1, and EMS72420.1), and Spirochaetes(AEF81843.1) are closest to proteins of this cluster,with 90 and 95% support on the MP and NJ trees,respectively. Three large, stable clusters (100% of sup�port) were detected on both MP and NJ trees; they areformed by proteins of bacteria (from Actinobacteriaand Firmicutes phyla), fungi, and plants, respectively.
Full�length amino acid sequences of some proteinsindicated in Fig. 1 were used to search for homologousproteins in the GenPept database (“non�redundantprotein sequences” section) by the blastp program.The results of counting the closest homologues num�ber that represent the same phylum of living organismsare shown in the table. Clusters of fungal and plantproteins (indicated in Fig. 1) correspond to numerousproteins, which are encoded in almost all genomes(with rare exceptions) of these organisms. However,the protein group that corresponds to the bacterialcluster is not so large and the genes that correspond toit are encoded only in a minority of sequencedgenomes of Actinobacteria and Firmicutes.
The phylogenetic analysis of the GH10 family pro�teins (MP and NJ trees were built; figures are notshown) that belong to the second subfamily (v.s.) con�firmed that the Planctomycetes proteins do not form adistinct cluster. Their presence in only three genera ofthe Planctomycetes (Gemmata, Phycisphaera, andRhodopirellula) can be explained by multiple cases ofindependent horizontal gene transfers.
Amplification of Genes Encoding Endo�β�Xylanases from Planctomycetes
Distribution of the Planctomycetes proteins fromthe first group on the phylogenetic tree (Fig. 1) corre�sponds well to the taxonomic position of the corre�sponding organisms. This is unusual for either themajority of glycoside hydrolases [2] and for the GH10family proteins of other Prokaryota [27, 28]. We ampli�fied fragments of genes coding the GH10 family pro�
teins from some other strains of the Planctomycetesfor checking this regularity.
Some authors claimed [27, 28] that there are uni�versal primers allowing to amplify endo�β�xylanasegenes of the GH10 family from wide range of bacteria.However, our analysis showed that these primers hadno annealing sites in the corresponding genes of thePlanctomycetes (data not shown). Moreover, we werenot able to design a universal structure of primer for allknown GH10 family genes of the Planctomycetes (andeven for the first subfamily) because they had highvariability. Thus, we designed degenerate primersbased on nucleotide sequences of the correspondinggenes of Isosphaera pallida and Singulisphaera acid�iphila. Using these primers, fragments of genomesfrom five strains of Isosphaera�Singulisphaera group ofthe Planctomycetes were amplified and sequenced,and the sequences were used in phylogenetic analysis.All five proteins and proteins from I. pallida and S. aci�diphila fell into one cluster (Fig. 2b). Notably, the frag�ment of amino acid sequence of S. acidiphilaMPL1015 (AGT55542.1) determined by us was iden�tical to the previously known sequence of S. acidiphilaDSM18658 (AGA27804.1).
The 16S rRNA gene sequences of two of the fivestrains of the Planctomycetes that we experimentallystudied are known (GenBank, AM902525.1 andJQ067914.1). Sequences of three others were deter�mined in the present work. The phylogenetic tree wasbuild using the multiple alignment of 16S rRNA genenucleotide sequences of Planctomycetes (Fig. 2a).The topology of this tree correlates very well with thatof GH10 family proteins from the Planctomycetes(Fig. 2b).
DISCUSSION
Catalytic domains of the GH10 family proteinshave the (β/α)8�barrel structure, the most frequenttertiary structure among glycoside hydrolases. All, ornearly all, glycoside hydrolase families with this struc�ture have distant evolutionary relationships. Many ofthem with GH10 form GH�A clan at higher level ofhierarchy, the largest glycoside hydrolase clan by num�ber of grouped families [1, 2, 29].
Evolutionary histories of glycoside hydrolase genesand rRNA genes usually differ significantly [2]. Thefrequent gene duplications and eliminations, as well ashorizontal transfers, yield a mosaic structure to phylo�genetic trees of many families glycoside hydrolases.
Fig. 1. Structure of phylogenetic tree of GH10 family of glycoside hydrolases (112 proteins) built by maximum parsimonymethod. Statistical significance of the nodes was evaluated by bootstrap analysis; the number of supporting pseudoreplicas (outof 100) is indicated at each nodes. Protein clusters discussed in the text are marked. All proteins are signed by their accession num�bers according to the GenPept database as well as by the host phyla. Stable clusters of the higher plant and Ascomycota proteinsare marked by triangles. Bootstrap support for each cluster is shown inside a triangle, while the number of proteins in the clusterand the taxonomic affiliation of their host organisms is indicated on the right. Act is Actinobacteria, Bac is Bacteroidetes, Chl isCloroflexi, Cya is Cyanobacteria, Fir is Firmicutes, Met is marine metagenome, Pla is Planctomycetes, Spi is Spirochaetes, Theis Thermotogae, Thr is Thermobacullum terrenum, and Ver is Verrucomicrobia.

MOLECULAR BIOLOGY Vol. 48 No. 3 2014
PHYLOGENY OF β�XYLANASES FROM PLANCTOMYCETES 443
21 seq (Viridiplantae)ABG76966.1 (Fir)AEY67427.1 (Fir)
ADN02449.1 (Spi)ABX07436.1 (Chl)
EHA58720.1 (The)AAN16480.1 (The)
ADJ46284.1 (Act)ACZ42385.1 (Thr)
AEV81392.1 (Act)EDY63301.2 (Act)
ABX88978.1 (Act)BAG85014.1 (Act)AEY92972.1 (Act)
AEV98756.1 (Bac)EJG02102.1 (Bac)
CAZ98089.1 (Bac)ADB42365.1 (Bac)CCH53515.1 (Bac)WP_010663080.1 (Bac)
ADV47645.1 (Bac)EDY82726.1 (Ver)
13 seq (Ascomycota)EEG53601.1 (Fir)
CCH55999.1 (Bac)AEI51978.1 (Bac)
EHP35421.1 (Ver)AEW21052.1 (Bac)EEG22593.1 (Ver)
EHP32233.1 (Ver)EEG22123.1 (Ver)EIP99734.1 (Ver)CCH00389.1 (Bac)
ACB76939.1 (Ver)EDO60218.1 (Fir)EDO60219.1 (Fir)
CAQ00211.1 (Act)ACZ30464.1 (Act)AEG44163.1 (Act)ADQ47344.1 (Fir)ABP65852.1 (Fir)
ADQ41407.1 (Fir)ADL42025.1 (Fir)
EGD49414.1 (Fir)AEV66803.1 (Fir)
AEY64449.1 (Fir)AFK86467.1 (Fir)
ADL68523.1 (Fir)AEF17756.1 (Fir)ADQ57411.2 (Fir)
ABW28526.1 (Cya)ADN16869.1 (Cya)
ACU99283.1 (Cya)ACK64415.1 (Cya)
EGN35770.1 (Fir)AEF81843.1 (Spi)AGC67515.1 (Fir)EMS72420.1 (Fir)
EEQ57087.1 (Fir)ADI22410.1 (Pla)
BAM04356.1 (Pla)EBE27272.1 (Met)
WP_020472577.1 (Pla)ADV61272.1 (Pla)AGA27804.1 (Pla)
WP_010033755.1 (Pla)ADG69659.1 (Pla)
WP_010583456.1 (Pla)ADY58863.1 (Pla)EDL61831.1 (Pla)
EAQ81247.1 (Pla)ADB18958.1 (Pla)
EMI19362.1 (Pla)EMI53465.1 (Pla)EMI44021.1 (Pla)EMB14634.1 & EMI25518.1 (Pla)
CAD71336.1 (Pla)EKK00284.1 (Pla)
EGF24716.1 (Pla)ELP33066.1 (Pla)
100
93 27
89 4546
9567
5236
5098
91
38
6149
100
100
100
50
100
100
100
100
61
100
100
100
66
5585
77
94
96
64
34
100
100
100
100
100
100
100
98
45
4483
45
100
100
100
100100
100
100
44
90
73
42
53 9997
82
46 9738
68
6051
93
100
100100
100
10086
3555
Bacteroidetes &Verrucomicrobia
Actinobacteria& Firmicutes
Planctomycetes
Bacteroidetes &Verrucomicrobia
4329

444
MOLECULAR BIOLOGY Vol. 48 No. 3 2014
NAUMOFF et al.
Members of different high�level taxa are often neigh�bors, and proteins of closely related organisms (andeven paralogues of one organism) are spread through�out the tree. Examples of these families are GH13 [30,31], GH27 [32], GH36 [2, 33], GH78 [34, 35], GH97[36], GH106 [34, 35], and GH114 [37]. According tothe published data, the GH10 family could be added tothis list [27, 28]. However, no published tree of thisfamily has any proteins from the Planctomycetes. Ourdata show that the Planctomycetes proteins belong totwo different subfamilies. One of them (the second)has a typical mosaic structure; it has proteins of onlysome species of the Planctomycetes (usually as severalparalogues) that spread rather randomly on the tree(figure is not shown).
The first subfamily of the GH10 family is anunusual case. Genes that encode members of this sub�family are present strictly as single copies on all stud�ied genomes of the heterotrophic Planctomycetes.The phylogenetic tree of this subfamily is almost thesame as the tree that was built based on the 16S rRNAgene sequences (Fig. 2). These are arguments in favorof vertical gene transfer in this protein group and theirimportant functional role in all Planctomycetes.According to all known data about enzymatic activi�ties of proteins of the GH10 family of glycoside hydro�lases we could propose that they are endo�β�xylanasesand provide the Planctomycetes the ability of theeffective decomposition of hemicellulose from plantbiomass.
Closest homologues of some GH10 family proteins
Protein Organism Phylum Number of proteins
AGA27804.1 Singulisphaera acidiphila DSM 18658 Planctomycetes 19
EEQ57087.1 Clostridiales bacterium 1_7_47FAA Firmicutes 1
ACK64415.1 Cyanothece sp. PCC 8801 Cyanobacteria 4
EGN35770.1 Lachnospiraceae bacterium 3_1_57FAA_CT1 Firmicutes 1
AEF81843.1 Treponema azotonutricium ZAS�9 Spirochaetes 1
EMS72420.1 Clostridium termitidis CT1112 Firmicutes 2
EEG53601.1 Clostridium asparagiforme DSM 15981 Firmicutes 1
AEI51978.1 Runella slithyformis DSM 19594 Bacteroidetes 2
EHP35421.1 Opitutaceae bacterium TAV5 Verrucomicrobia 2
AEW21052.1 Tannerella forsythia ATCC 43037 Bacteroidetes 5
EIP99734.1 Opitutaceae bacterium TAV1 Verrucomicrobia 5
CCH00389.1 Fibrella aestuarina BUZ 2 Bacteroidetes 1
ACB76939.1 Opitutus terrae PB90�1 Verrucomicrobia 1
EDO60219.1 Clostridium leptum DSM 753 Firmicutes 21
AEG44163.1 Isoptericola variabilis 225 Actinobacteria 10
AEF17756.1 Thermoanaerobacterium xylanolyticum LX�11 Firmicutes 20
ABF94920.1 Oryza sativa Japonica Group Streptophyta 240
AEY67427.1 Clostridium sp. BNL1100 Firmicutes 3
ADN02449.1 Spirochaeta thermophila DSM 6192 Spirochaetes 2
ABX07436.1 Herpetosiphon aurantiacus DSM 785 Chloroflexi 1
EHA58720.1 Thermotoga maritima MSB8 Thermotogae 12
ADJ46284.1 Amycolatopsis mediterranei U32 Actinobacteria 7
AEY92972.1 Streptomyces hygroscopicus subsp. jinggangensis 5008 Actinobacteria 2
AEV98756.1 Niastella koreensis GR20�10 Bacteroidetes 10
CCH53515.1 Fibrisoma limi BUZ 3 Bacteroidetes 5
EDY82726.1 Verrucomicrobiae bacterium DG1235 Verrucomicrobia 1
CAA90075.1 Emericella nidulans ATCC 38163 Ascomycota 82
The accession number of proteins in the GenPept database (NCBI) is in the first column. Organism and phylum to which it belongs are inthe second and third columns. In the fourth column, there is a number of proteins from the same phylum that were found among the closesthomologues in the GenPept database (“non�redundant protein sequences” section) using full�sized amino acid sequences of the corre�sponding protein as the query (next in the list protein belongs to another phylum) by the blastp program. The database was screened on theJune 22, 2013.

MOLECULAR BIOLOGY Vol. 48 No. 3 2014
PHYLOGENY OF β�XYLANASES FROM PLANCTOMYCETES 445
Fig
. 2. C
ompa
riso
n o
f Pla
nct
omyc
etes
ph
ylog
enet
ic t
rees
th
at w
ere
buil
t by
nei
ghbo
r�jo
inin
g m
eth
od b
ased
on
mul
tipl
e al
ign
men
ts o
f fra
gmen
ts o
f (a)
nuc
leot
ide
sequ
ence
s of
16S
rR
NA
gen
es a
nd
(b)
amin
o ac
id s
eque
nce
s of
th
e G
H10
pro
tein
s. S
tati
stic
al s
ign
ific
ance
of t
he
nod
es w
as e
valu
ated
usi
ng
the
boot
stra
p an
alys
is; t
he
num
ber
of s
uppo
rtin
gps
eudo
repl
icas
(ou
t of 1
000)
is in
dica
ted.
Th
e n
ames
of o
rgan
ism
s fo
r w
hic
h g
enes
the
GH
10 fa
mil
y pr
otei
ns
wer
e se
quen
ced
duri
ng
the
pres
ent w
ork
(see
text
) ar
e un
derl
ined
.
(a)
(b)
1000
988
531
987
998
802
580
963
1000
1000
1000
1000
1000
1000
970
627
594
980
991
965 51
085
9
992
100
1000
1000
1000
1000
1000
403 39
1
984
998
522
835
871
985
610
468
806
905
995
672
944
717
100
un
cult
ure
d P
lan
ctom
ycet
ales
bac
teri
um
HF
0500
_02
G17
Gem
mat
a ob
scur
iglo
bus
UQ
M 2
246
Zav
arzi
nella
form
osa
DS
M 1
9928
Isos
phae
ra p
alli
da A
TC
C 4
3644
Isos
phae
ra s
p. P
X4
‘Nos
toco
ida
acid
iphi
la’
OB
1
Sin
guli
spha
era
acid
iphi
la B
W
Sin
guli
spha
era
acid
iphi
la P
X31
Sch
lesn
eria
pal
udic
ola
DS
M 1
8645
Pla
ncto
myc
es m
aris
DS
M 8
797
‘Rho
dopi
rellu
la m
aior
ica’
SM
1
Rho
dopi
rellu
la s
p. S
WK
7
Phy
cisp
haer
a m
ikur
ensi
s N
BR
C 1
0266
6
Sin
guli
spha
era
acid
iphi
la M
PL
1015
Sin
guli
spha
era
acid
iphi
la D
SM
186
58
Pla
ncto
myc
es li
mno
phil
us D
SM
377
6
Pla
ncto
myc
es b
rasi
lien
sis
DS
M 5
305
Pir
ellu
la s
tale
yi D
SM
606
8
Bla
stop
irel
lula
mar
ina
DS
M 3
645
‘ Rho
dopi
rellu
la s
alle
ntin
a’ S
M41
‘Rho
dopi
rellu
la e
urop
aea’
SH
398
‘ Rho
dopi
rellu
la e
urop
aea’
6C
Rho
dopi
rellu
la b
alti
ca S
H 1
Rho
dopi
rellu
la b
alti
ca S
H28
Rho
dopi
rellu
la b
alti
ca S
WK
14
Rho
dopi
rellu
la b
alti
ca W
H47

446
MOLECULAR BIOLOGY Vol. 48 No. 3 2014
NAUMOFF et al.
An analysis of the phylogenetic tree (Fig. 1), thatwas built based on the sequences of Planctomycetesproteins from the first subfamily and the closest pro�teins of other organisms, shows that the proteins ofother bacterial phyla are spread on the tree rather cha�otically. This could indicate the multiplicity of dele�tions and horizontal transfers of genes encoding it dur�ing the evolution. For example, three clusters ofbranches were found that contain members ofBacteroidetes and Verrucomicrobia phyla at the sametime. The structure of clusters shows that there werethree or four horizontal transfers of endo�β�xylanasegenes between bacteria of these two taxa. The genesencoding proteins of this group (Fig. 1) are presentonly in a small number of known prokaryoticgenomes, it shows that their functions are optional inthe most bacterial groups.
Notably, the proteins from the phylum Proteobac�teria were absent from the phylogenetic tree (Fig. 1).We screened (data not shown) amino acid sequencedatabase by the blastp algorithm using protein of Sin�gulisphaera acidiphila DSM18658 (GenPept,AGA27804.1) as the query. This allowed us to find thenearest homologue from the Proteobacteria only withan E�value of 0.002 (from Rhizobium leguminosarum;WP_020051258.1; member of the GH10 family). Itshould be recalled that the Proteobacteria is the leaderamong prokaryotic phyla in both the number of validlydescribed species [38] and the number of thesequenced genomes [39]. Thus, it seems reasonable toexpect that they have members among all large groupsof bacterial proteins, which undergo horizontal trans�fer [34, 35]. Accordingly, the absence of proteobacte�rial proteins among the closest homologues of thePlanctomycetes proteins of this group could not beconsidered to be random. We previously showed thatthe Proteobacteria has no glycoside hydrolases ofsome other families totally or almost totally. Thus,these bacteria have no GH101 family that contains theendo�α�N�acetylgalactosaminidases [40, 41]. Only asmall number of species has GH78 and GH106 fami�lies composing by α�L�ramnosidases [34, 35]. Thesame phenomenon was found in other numerous bac�terial phyla, i.e., Firmicutes has no proteins of theGH114 family, which includes endo�α�1,4�polyga�lactosaminidases [37]. The explanation of this phe�nomenon has yet to be found.
It should be noted that in addition to the Plancto�mycetes cluster, the large stable clusters correspondingto fungi and plants were found on the phylogenetictree (Fig. 1). It seems that the endo�β�xylanases of thispart of the GH10 family are of vital importance. It islogical to assume that plants need them for synthesis ofa large number of hemicellulose but Fungi and Planc�tomycetes need them to hydrolyze it.
Thus, we established the similarity between thetopology of the Planctomycetes phylogenetic treesthat were built both based on amino acid sequences ofGH10 family of endo�β�xylanases and nucleotide
sequences of 16S rRNA genes. It suggests the verticalinheritance of the endo�β�xylanase genes of this bac�terial phylum that is unusual for glycoside hydrolasegenes.
ACKNOWLEDGMENTS
The work was supported by Russian Foundation forBasic Research (project no. 12�04�00480) and the“Molecular and Cell Biology” program of the Presid�ium of the Russian Academy of Sciences.
REFERENCES
1. Lombard V., Ramulu H.G., Drula E., Coutinho P.M.,Henrissat B. 2014. The carbohydrate�active enzymesdatabase (CAZy) in 2013. Nucleic Acids Res. 42, D490–D495. http://www.cazy.org/
2. Naumoff D.G. 2011. Hierarchical classification of gly�coside hydrolases. Biochemistry (Moscow). 76, 622–635.
3. Reilly P.J. 1981. Xylanases: Structure and function.Basic Life Sci. 18, 111–129.
4. Kulkarni N., Shendye A., Rao M. 1999. Molecular andbiotechnological aspects of xylanases. FEMS Microbiol.Rev. 23, 411–456.
5. Subramaniyan S., Prema P. 2000. Cellulase�free xyla�nases from Bacillus and other microorganisms. FEMSMicrobiol. Lett. 183, 1–7.
6. Beg Q.K., Kapoor M., Mahajan L., Hoondal G.S.2001. Microbial xylanases and their industrial applica�tions: A review. Appl. Microbiol. Biotechnol. 56, 326–338.
7. Subramaniyan S., Prema P. 2002. Biotechnology ofmicrobial xylanases: Enzymology, molecular biology,and application. Crit. Rev. Biotechnol. 22, 33–64.
8. Polizeli M.L., Rizzatti A.C., Monti R., Terenzi H.F.,Jorge J.A., Amorim D.S. 2005. Xylanases from fungi:Properties and industrial applications. Appl. Microbiol.Biotechnol. 67, 577–591.
9. Ahmed S., Riaz S., Jamil A. 2009. Molecular cloning offungal xylanases: An overview. Appl. Microbiol. Biotech�nol. 84, 19–35.
10. Pollet A., Delcour J.A., Courtin C.M. 2010. Structuraldeterminants of the substrate specificities of xylanasesfrom different glycoside hydrolase families. Crit. Rev.Biotechnol. 30, 176–191.
11. Ward N.L. 2010. Phylum XXV. Planctomycetes, Garrityand Holt 2001, vol. 4 of Bergey’s Manual of SystematicBacteriology. Eds. Krieg N.R., Staley J.T., Brown D.R.,et al. NY: Springer, pp. 879–925.
12. Fuerst J.A. 1995. The planctomycetes: Emerging mod�els for microbial ecology, evolution and cell biology.Microbiology. 141, 1493–1506.
13. Fuerst J.A. 2004. Planctomycetes: A phylum of emerg�ing interest for microbial evolution and ecology. WorldFed. Cult. Collect. Newsl. 38, 1–11.
14. Fuerst J.A., Sagulenko E. 2011. Beyond the bacterium:Planctomycetes challenge our concepts of microbialstructure and function. Nature Rev. Microbiol. 9, 403–413.

MOLECULAR BIOLOGY Vol. 48 No. 3 2014
PHYLOGENY OF β�XYLANASES FROM PLANCTOMYCETES 447
15. Ivanova A.O., Dedysh S.N. 2012. Abundance, diversity,and depth distribution of Planctomycetes in acidicnorthern wetlands. Front. Microbiol. 3, Art. 5.
16. Kuypers M.M.M., Sliekers A.O., Lavik G., Schmid M.,Jørgensen B.B., Kuenen J.G., Damsté J.S.S., Strous M.,Jetten M.S.M. 2003. Anaerobic ammonium oxidationby anammox bacteria in the Black Sea. Nature. 422,608–611.
17. Kulichevskaya I.S., Pankratov T.A., Dedysh S.N. 2006.Detection of representatives of the Planctomycetes inSphagnum peat bogs by molecular and cultivationapproaches. Microbiology (Moscow). 75, 329–335.
18. Kulichevskaya I.S., Belova S.E., Kevbrin V.V.,Dedysh S.N., Zavarzin G.A. 2007. Analysis of the bac�terial community developing in the course of Sphagnummoss decomposition. Microbiology (Moscow). 76,621–629.
19. Kulichevskaya I.S., Ivanova A.O., Belova S.E.,Baulina O.I., Bodelier P.L.E., Rijpstra W.I.C., Dam�sté J.S.S., Zavarzin G.A., Dedysh S.N. 2007. Schlesne�ria paludicola gen. nov., sp. nov., the first acidophilicmember of the order Planctomycetales, from Sphag�num�dominated boreal wetlands. Int. J. Syst. Evol.Microbiol. 57, 2680–2687.
20. Kulichevskaya I.S., Ivanova A.O., Baulina O.I., Bode�lier P.L.E., Damsté J.S.S., Dedysh S.N. 2008. Sin�gulisphaera acidiphila gen. nov., sp. nov., a non�fila�mentous, Isosphaera�like planctomycete from acidicnorthern wetlands. Int. J. Syst. Evol. Microbiol. 58,1186–1193.
21. Kulichevskaya I.S., Baulina O.I., Bodelier P.L.E., Rijp�stra W.I.C., Damsté G.S.S., Dedysh S.N. 2009. Zavar�zinella formosa gen. nov., sp. nov., a novel stalked, Gem�mata�like planctomycete from a Siberian peat bog. Int.J. Syst. Evol. Microbiol. 59, 357–364.
22. Kulichevskaya I.S., Detkova E.N., Bodelier P.L.E.,Rijpstra W.I.C., Damsté G.S.S., Dedysh S.N. 2012.Singulisphaera rosea sp. nov., a novel planctomycetefrom acidic Sphagnum peat, and emended descriptionof the genus Singulisphaera. Int. J. Syst. Evol. Microbiol.62, 118–123.
23. Kulichevskaya I.S., Serkebaeva Y.M., Kim Y., Rijp�stra W.I.C., Damsté G.S.S., Liesack W., Dedysh S.N.2012. Telmatocola sphagniphila gen. nov., sp. nov., anovel dendriform planctomycete from northern wet�lands. Front. Microbiol. 3, Art. 146.
24. Kulichevskaya I.S., Ivanova A.A., Belova S.E.,Dedysh S.N. 2012. A novel filamentous planctomyceteof the Isosphaera�Singulisphaera group isolated from aSphagnum peat bog. Microbiology (Moscow). 81, 446–452.
25. Marmur J. 1961. A procedure for the isolation of deox�yribonucleic acid from microorganisms. J. Mol. Biol. 3,208–218.
26. Weisburg W.G., Barns S.M., Pelletier D.A., Lane D.J.1991. 16S ribosomal DNA amplification for phyloge�netic study. J. Bacteriol. 173, 697–703.
27. Wang G., Wang Y., Yang P., Luo H., Huang H., Shi P.,Meng K., Yao B. 2010. Molecular detection and diver�sity of xylanase genes in alpine tundra soil. Appl. Micro�biol. Biotechnol. 87, 1383–1393.
28. Wang G., Meng K., Luo H., Wang Y., Huang H., Shi P.,Yang P., Zhang Z., Yao B. 2012. Phylogenetic diversityand environment�specific distributions of glycosyl
hydrolase family 10 xylanases in geographically distantsoils. PLoS ONE. 7, e43480.
29. Naumoff D.G. 2006. Development of a hierarchicalclassification of the TIM�barrel type glycoside hydro�lases. Proc. Fifth Int. Conf. Bioinformat. Genome Regul.Structure, Novosibirsk, Russia, July 16–22, 2006,vol. 1, pp. 294–298. http://www.bionet.nsc.ru/meeting/bgrs_proceedings/papers/2006/BGRS_2006_V1_067.pdf
30. Stam M.R., Danchin E.G., Rancurel C., Coutinho P.M.,Henrissat B. 2006. Dividing the large glycoside hydro�lase family 13 into subfamilies: towards improved func�tional annotations of α�amylase�related proteins. Pro�tein Eng. Des. Sel. 19, 555–562.
31. Gizatullina D.I., Naumoff D.G. 2009. Reclassificationof GH13 family of glycoside hydrolases. Proc. Int..Moscow Conf. Comput. Mol. Biol., July 20–23, 2009,pp. 249–250. http://mccmb.belozersky.msu.ru/2009/MCCMB09_Proceedings.pdf
32. Naumoff D.G. 2004. Phylogenetic analysis of α�galac�tosidases of the GH27 family. Mol. Biol. (Moscow). 38,388–399.
33. Naumoff D.G. 2004. The α�galactosidase superfamily:Sequence based classification of α�galactosidases andrelated glycosidases. Proc. Fourth Int. Conf. Bioinfor�mat. Genome Regul. Structure, Novosibirsk. Russia,July 25–30, 2004, vol. 1, pp. 315–318. http://www.bio�net.nsc.ru/meeting/bgrs_proceedings/papers/2004/BGRS_2004_V1_079.pdf
34. Naumoff D.G., Dedysh S.N. 2012. Lateral gene trans�fer between the Bacteroidetes and Acidobacteria: Thecase of α�L�rhamnosidases. FEBS Lett. 586, 3843–3851.
35. Naumoff D.G. 2013. Multiple lateral transfers and dupli�cations of genes as sources of diversity of α�L�rhamnosi�dases in Clostridium methylpentosum DSM5476. Micro�biology (Moscow). 82, 415–422.
36. Naumoff D.G. 2005. GH97 is a new family of glycosidehydrolases, which is related to the α�galactosidasesuperfamily. BMC Genomics. 6, Art. 112.
37. Naumoff D.G., Stepuschenko O.O. 2011. Endo�2α�1,4�polygalactosaminidases and their homologs: Struc�ture and evolution. Mol. Biol. (Moscow). 45, 647–657.
38. Yarza P., Ludwig W., Euzéby J., Amann R.,Schleifer K.�H., Glöckner F.O., Rosselló�Móra R.2010. Update of the all�species living tree project basedon 16S and 23S rRNA sequence analyses. Syst. Appl.Microbiol. 33, 291–299.
39. Pagani I., Liolios K., Jansson J., Chen I�M.A.,Smirnova T., Nosrat B., Markowitz V.M., Kyrpides N.C.2012. The Genomes OnLine Database (GOLD) v.4:Status of genomic and metagenomic projects and theirassociated metadata. Nucleic Acids Res. 40, D571–D579.
40. Naumoff D.G. 2010. GH101 family of glycosidehydrolases: Subfamily structure and evolutionary con�nections with other families. J. Bioinform. Comput. Biol.8, 437–451.
41. Naumoff D.G. 2013. Bioinformatic analysis of familyGH101 of glycoside hydrolases. FEBS J. 280 (S1), 540.
Translated by V. Kharcheva






















![Poe 2004 Phylogeny of Anoles[1]](https://img.pdfslide.tips/doc/110x75/577cdaad1a28ab9e78a63b09/poe-2004-phylogeny-of-anoles1.jpg)





