Embed Size (px)
Citation preview

PLASTICIDAD FENOTÍPICA EN LA GERMINACIÓN DE Cephalocereus palmeri var. sartorianus (Rose) Krainz.,. (CACTACEAE) BAJO DOS CONDICIONES DE LUZ
TESIS QUE PRESENTA JOSÉ ANTONIO MIRANDA JÁCOME PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS
Xalapa, Veracruz, México 2008




DEDICATORIAS
A mis padres, por su amor, su compañía y por alentarme a seguir adelante
siempre.
A mis hermanos, por las batallas campales de la niñez (algo así de todos contra
todos y para todo); así mismo les agradezco haberme regalado buenos y malos
momentos que contribuyeron de manera significativa para mejorar mi historia de vida.
A mis sobrinos, por haberme demostrado que soy un poco más humano de lo
que pensaba.
A Rosa Evit por su apoyo incondicional y su grata compañía durante estos diez
placenteros años; mi reconocimiento y mi amor.
Y en especial a Paolo Miranda, por regalarme la felicidad inconmensurable de
estar junto a él, te amo hijo.
i

RECONOCIMIENTOS
Expreso mi agradecimiento sincero al Dr. Carlos Montaña, quien ha sido una
persona clave en mi realización profesional, primero como director de tesis de
licenciatura y ahora como director de la tesis de maestría, por su tiempo, apoyo,
estimulo y amistad, gracias infinitas.
Al Dr. Oscar Briones, le agradezco las sugerencias y la disponibilidad en todo
momento para discutir el manuscrito, de igual manera le agradezco la invaluable
amistad.
Al Dr. César Domínguez, por las observaciones, comentarios y sugerencias, ya
que gracias a ellas el trabajo cambió en algunos puntos radicalmente, mejorando el
diseño del marco conceptual del mismo.
Al Dr. Luis Eguiarte y la Dra. Maria Luisa Martínez (miembros del jurado) les
agradezco el cuidado de sus revisiones y las sugerencias al manuscrito de la tesis que
contribuyeron significativamente para mejorarlo, aunque si existen errores, la
responsabilidad es de un servidor.
A los Doctores Roger Guevara por la ayuda con algunos análisis estadísticos,
Fabiana Pezzani por la orientación en aspectos logísticos del trabajo de tesis y Joel
Flores por sus comentarios en algunos puntos del manuscrito de la tesis.
A mis compañeros del departamento de Biología Evolutiva: Hugo Piña, Yarenni
Perroni y Francisco Reyes, les agradezco haber hecho más placentera mi estancia en
el INECOL.
Agradezco a Alejandra Lepe y Miguel Munguía por acompañarme en las salidas
a campo. En especial a éste último le agradezco haberme mostrado sus resultados (aun
no publicados) sobre la polinización en C. palmeri var. sartorianus.
Así mismo agradezco a mis compañeros del posgrado: Sandra Amézquita, Vieyle
Cortés, Nadia Rivera y Pablo Berea su amistad y apoyo.
Agradezco al CONACyT el apoyo económico (No. 203971), que me otorgó para
el estudio de Posgrado (sep. 2005 - ago. 2007).
Agradezco al Instituto de Ecología A. C. las facilidades otorgadas para el
desarrollo de la tesis.
ii

RELACIÓN DE TABLAS
Tabla 1. Análisis de varianza del porcentaje de la cobertura vegetal registrado
para cada uno de los sitios. 24
Tabla 2. Tabla de contingencia aplicada a los resultados obtenidos de la
asociación de plántulas bajo cada una de las condiciones de luz
en los cinco sitios. 25
Tabla 3. Análisis de varianza del arcoseno de la germinación como una función
de sitios, familias genéticas y tratamiento de luz. 26
Tabla 4. Coeficiente de correlación intraclase (± error estándar) entre los
porcentajes de germinación obtenidos bajo dos calidades de luz en
cada uno de los sitios de estudio. 32
Tabla 5. Análisis de la varianza de la masa de semillas como una función de
sitios y familias genéticas. 34
RELACIÓN DE FOTOGRAFÍAS
Foto 1. Cephalocereus palmeri var. sartorianus (Rose) Krainz.,. (Fotos:
Antonio Miranda). 12
iii

RELACIÓN DE FIGURAS
Figura 1. Mapa de la República Mexicana donde se indica la ubicación de los
sitios de Cephalocereus palmeri var. sartorianus estudiados y su altitud. 15
Figura 2. Porcentajes de cobertura vegetal (± error estándar) de los cinco
sitios. TE= Tenampa; SI= San Ignacio; LR= Loma de Rogel; PA= Puente
Actopan; PN= Puente Nacional. Letras diferentes indican diferencias
significativas (Prueba de Tukey p<0.05). 24
Figura 3. Porcentajes de germinación (± error estándar) de los cinco sitios
bajo los dos tratamientos de luz. Luz blanca (barras claras), luz rica en
rojo-lejano (barras oscuras) TE= Tenampa; SI= San Ignacio; LR= Loma
de Rogel; PA= Puente Actopan; PN= Puente Nacional. Letras diferentes
indican diferencias significativas (Prueba de Tukey p<0.05). 27
Figura 4. Valor fenotípico inter-sitio expresado en porcentaje (± error
estándar) para la germinación de semillas en respuesta a los dos
tratamientos de luz. 28
Figura 5. Normas de reacción del porcentaje de germinación en respuesta a
los dos tratamientos de luz para las 15 familias de cada sitio. 29
Figura 6. Valor fenotípico expresado en porcentaje (± error estándar), para
la germinación de semillas en respuesta a los dos tratamientos de luz
para las 15 familias genéticas de cada sitio. 30
Figura 7. Relación entre los porcentajes de germinación obtenidos con dos
calidades de luz (luz blanca y rojo lejano) con semillas producidas en
15 familias genéticas de 5 sitios distintos. Para cada sitio se muestran
los puntos correspondientes a las 15 familias genéticas, y el valor del
iv

coeficiente de correlación intraclase (± error estándar). Valor crítico
(r 0.05, 2, 13 = 0.514). 33
Figura 8. Masa de semillas expresada en gramos (± error estándar) de los
cinco sitios. TE= Tenampa; SI= San Ignacio; LR= Loma de Rogel; PA=
Puente Actopan; PN= Puente Nacional. Letras diferentes indican diferencias
significativas (Prueba de Tukey p<0.05). El valor de la masa corresponde
al valor neto de las semillas multiplicado por 1000 para cada sitio. 35
Figura 9. Relaciones entre el porcentaje de germinación y la masa de
semillas obtenidos bajo dos tratamientos de luz en cada uno de los sitios
de estudio (○ familias genéticas sometidas al tratamiento de luz blanca;
● familias genéticas sometidas al tratamiento de luz rica en rojo-lejano).
Se muestran también los coeficientes de correlación (± error estándar)
calculados para cada tratamiento de luz. Valor crítico (r 0.05, 2, 13 = 0.514).
El valor de la masa corresponde al valor neto de las semillas multiplicado
por 1000 para cada familia genética de cada sitio. 36
Figura 10. Correlaciones entre el arcoseno de la germinación y las variables
ambientales (a= altitud; b= precipitación, c= temperatura y d= cobertura
vegetal). Se muestra la ecuación de la recta para la germinación contra
cada variable ambiental y el valor de p. TE = Tenampa; SI = San Ignacio;
LR = Loma de Rogel; PA = Puente Actopan; PN = Puente Nacional. 37
Figura 11. Relación entre la masa de semillas y las variables ambientales
(a= altitud; b= precipitación, c= temperatura y d= cobertura vegetal).
Se muestra la ecuación de la recta para el peso de semillas contra
cada variable ambiental y el valor de p. TE = Tenampa; SI = San Ignacio;
LR = Loma de Rogel; PA = Puente Actopan; PN = Puente Nacional. El
Valor de la masa corresponde al valor neto de las semillas multiplicado
por 1000 para cada familia genética de cada sitio. 39
v

INDICE
CONTENIDO Página
DEDICATORIA i
RECONOCIMIENTOS ii
RELACIÓN DE TABLAS iii
RELACIÓN DE FOTOGRAFIÁS iii
RELACIÓN DE FIGURAS iv
1. RESUMEN 1
2. INTRODUCCIÓN 2
2.1 Plasticidad fenotípica. 2
2.2 Germinación de cactáceas. 6
2.3 Germinación y masa de semillas. 9
2.4 Planteamiento del problema. 10
3. OBJETIVOS 11
3.1 Objetivo general. 11
3.2 Objetivos específicos. 12
4. MÉTODOS 12
4.1 La especie de estudio. 12
4.2 Los sitios de estudio. 14
4.3 Diseño experimental para medir la cobertura vegetal inter-sitios y la
asociación de plántulas de C. palmeri var. sartorianus en respuesta
a la luz. 17
4.4 Diseño experimental para medir la plasticidad fenotípica de la

germinación de C. palmeri var. sartorianus en respuesta a la luz. 18
4.5 Diseño experimental para estimar la variación intra e inter-sitio
en la masa de semillas de C. palmeri var. sartorianus. 20
4.6 Diseño experimental para evaluar el efecto ambiental sobre la
variación inter-sitio en la masa de semillas y la germinación de
C. palmeri var. sartorianus. 20
4.7 Análisis estadísticos. 21
5. RESULTADOS 23
5.1 Cobertura vegetal inter-sitio y asociación de plántulas en respuesta
a las condiciones lumínicas. 23
5.2 Germinación y calidad de la luz. 25
5.3 Correlaciones genéticas entre la respuesta de la germinación a las
distintas calidades de la luz. 31
5.4 Masa de semillas. 34
5.5 Variación de la germinación y la masa de semillas en función de las
variables ambientales. 37
6. DISCUSIÓN 40
6.1 Cobertura vegetal inter-sitio y asociación de plántulas. 40
6.2 Germinación y calidad de la luz. 42
6.2.1 Variación intra-sitio. 42
6.2.2 Variación inter-sitio. 43
6.2.3 Factor lumínico. 45
6.3 Correlaciones genéticas entre la respuesta de la germinación a las
distintas calidades de la luz. 47

6.4 Variación de la germinación en función de la masa de semillas. 49
6.5 Variación de la germinación y la masa de las semillas en función
de las variables ambientales. 51
6.6 Discusión general 53
7. REFERENCIAS BIBLIOGRÁFICAS 56

1. RESUMEN
Las condiciones ambientales extremas ocasionan que la germinación de semillas
de cactáceas sea un evento raro y esporádico. Muchas especies aumentan la
germinación cuando se establecen bajo la copa de los árboles de otras especies,
fenómeno conocido como nodricismo. El presente trabajo evaluó la respuesta
germinativa de semillas en cámara de germinación bajo condiciones de luz blanca
(hábitat abierto) y luz rica en rojo-lejano (hábitat cerrado), en 15 familias genéticas de
cinco sitios ubicados en un gradiente altitudinal en el que se distribuye la cactácea
columnar Cephalocereus palmeri var. sartorianus. El objetivo fue determinar si la
calidad de la luz es un factor importante en la promoción de la germinación y estudiar la
variabilidad de este efecto tanto a nivel inter como intra-sitio y su plasticidad fenotípica.
Se obtuvieron las normas de reacción para cada una de las familias por sitio. Se
observó que dicho rasgo posee una alta variación genética para la plasticidad a nivel
intra-sitio (77.20 % de la varianza explicada) y menor variación inter-sitios (r2= 5.8 %).
Las correlaciones genéticas mostraron que hay dependencia genética entre los estados
del carácter de acuerdo a la calidad de luz en la germinación para los sitios de San
Ignacio, Loma de Rogel, Puente Actopan y Puente Nacional. En contraste para
Tenampa existe independencia genética y, por lo tanto, existe la posibilidad de la
evolución de la norma de reacción. Se comprobó que existen diferencias en la masa de
semillas a nivel intra-sitio (r2= 50.5 %), e inter-sitio (r2= 19 %), pero éstas no afectaron a
la germinación entre sitios, ni entre tratamiento de luz. Por último, se demostró que la
germinación así como la masa de semillas se encuentran correlacionadas con variables
ambientales (temperatura, precipitación y altitud).
1

2. INTRODUCCIÓN
2.1 Plasticidad fenotípica.
El estudio de las respuestas fenotípicas al ambiente tiene una larga historia, ya
que se ha observado que individuos de la misma especie pueden variar
considerablemente en rasgos fenotípicos importantes debido a los diferentes ambientes
en que se desarrollan. Estas variaciones pueden ser originadas por un componente
genético o por un componente ambiental, siendo este último el que le dará a un
genotipo u organismo la oportunidad de expresar diferentes fenotipos en repuesta al
ambiente, lo que se le conoce como plasticidad fenotípica (Schmalhausen, 1949; Via,
1994).
Una manera efectiva para mantener la adecuación al ambiente es a través de la
plasticidad fenotípica (Bradshaw, 1965; Via & Lande, 1985; Schlichting, 1986; Sultan,
1987; Schlichting & Pigliucci, 1998; Wright & McConnaughay, 2002; Donohue, 2003), ya
que la plasticidad al ambiente promueve que individuos puedan hacer frente a la
heterogeneidad ambiental, dotándolos de cierta capacidad de responder frente a
condiciones bióticas o abióticas (McCullough & Shropshire, 1970; Hayes & Klein, 1974;
Evans & Cabin, 1995; Gutterman, 2000), involucrando así cambios en la fisiología,
morfología y desarrollo de los genotipos creciendo bajo diferentes ambientes (Pigliucci,
2001).
Las poblaciones de plantas pueden mostrar diferenciación en la plasticidad
fenotípica (Platenkamp, 1990; Sultan & Bazzaz, 1993; Oyama, 1994), puesto que los
patrones de variación ambiental pueden influenciar la divergencia en la plasticidad de
2

rasgos entre dichas poblaciones (Galloway, 1995; Weining, 2000). De hecho algunas
teorías predicen que mayores niveles de heterogeneidad ambiental pueden seleccionar
altas magnitudes de plasticidad fenotípica (Bradshaw & Hardwick, 1989), al existir
evidencia de que plantas que crecen bajo ambientes más cambiantes presentan niveles
mayores de respuesta fenotípica.
La plasticidad fenotípica se manifiesta por una interacción entre genotipo y
ambiente, cuya existencia se puede verificar mediante un Análisis de Varianza
(ANDEVA) que evalúe los efectos del genotipo y el ambiente. La significancia en esta
interacción, nos indica que existe el potencial para que dicha plasticidad pueda
evolucionar (Via & Lande, 1985).
Cuando se considera a la plasticidad fenotípica, es importante recordar que la
señal que dispara las diferencias fenotípicas es el ambiente y que esta habilidad o
plasticidad de responder a la señal ambiental está basado hasta que punto puede
evolucionar (Via, 1994). Para poder entender qué tanto la plasticidad puede
evolucionar, se debe determinar cómo se va a medir. La forma más utilizada es por
medio de la norma de reacción (Núñez-Farfán & Cordero, 1993). La norma de reacción
de un genotipo es su rango de respuestas fenotípicas a lo largo de un gradiente
ambiental (Woltereck, 1909; citado en Schlichting & Pigliucci, 1998). Normalmente se
presentan las normas de reacción en gráficas donde las respuestas fenotípicas se
miden en las ordenadas y los estados del gradiente ambiental se representan en el eje
de las abscisas.
La importancia de los estudios sobre la norma de reacción radica en mostrar la
variabilidad ya sea de caracteres de historia de vida, fisiológicos, morfológicos y de
comportamiento, dentro y entre poblaciones. Cualquier norma de reacción que no sea
3

paralela al eje ambiental (abscisas) indica por definición la presencia de plasticidad
(Lewontin, 1974). Estas diferencias poblacionales obedecen entre otras cosas a
factores ambientales y estas respuestas pueden o no tener origen plástico o genético.
Con ayuda de la genética cuantitativa se pueden analizar poblaciones para saber
si la variabilidad de los caracteres se encuentra asociada a la adecuación, además si
esta variabilidad tiene un componente genético o es producto del ambiente. La genética
cuantitativa pretende determinar qué fracción de la varianza fenotípica total (Vf) de un
carácter es de origen genético (Vg) y qué fracción de origen ambiental (Va), es decir, Vf
= Vg + Va (Núñez-Farfán & Cordero, 1993).
Las correlaciones genéticas entre caracteres son parámetros de interés para la
genética cuantitativa y estas muestran el grado de asociación entre dos caracteres
(Núñez-Farfán, 1991).
Dichas correlaciones genéticas, obtenidas del análisis fenotípico de los
individuos son fuentes importantes para el cambio evolutivo (Bradshaw, 1984) ya que
pueden ser usadas para predecir la respuesta potencial a la selección o para identificar
las restricciones a la evolución futura (Mitchell-Olds & Rutledge, 1986).
Una correlación genética significativa a través del ambiente, indica que los
estados de carácter (estos últimos tomados como la expresión de un carácter en un
ambiente determinado) no son independientes (Via, 1994). Por lo que cualquier
correlación genética a través de los ambientes con valor absoluto mayor a cero sugiere
una restricción genética en el ajuste independiente del fenotipo expresado dentro de
cada ambiente y por consiguiente una restricción en la evolución de una norma de
reacción adaptativa (Via, 1994).
4

Aquellos loci que introducen variación genética en los estados del carácter
expresados en cierto ambiente y que no están correlacionados con la variabilidad
genética en los estados del carácter expresados en otro ambiente (variación genética
no correlacionada) podrán facilitar el ajuste independiente del estado del carácter en
cada sitio y promocionar la aceleración en la evolución de una norma de reacción
adaptativa (Via, 1994).
Las correlaciones genéticas estiman el grado en el cual los fenotipos expresados
en dos ambientes tienen la misma base genética. Una correlación genética alta a través
de los ambientes implica que los mismos alelos o conjunto de alelos influencian los
estados del carácter de la misma manera en dos ambientes (Via & Lande, 1985).
Los estudios de genética de poblaciones en cactáceas son pocos, considerando
su importancia ecológica en regiones áridas y semi-áridas del hemisferio occidental, por
ejemplo en 2002 sólo existían estimaciones de diversidad genética (mediante el uso de
aloenzimas) para 10 especies (Hamrick et al., 2002).
Aunado al poco conocimiento que se tiene de la genética de poblaciones para
solo algunas especies de cactáceas, el estudio abordado bajo la genética cuantitativa
para dicha familia es sorprendentemente escaso, el cual puede resultar una poderosa
herramienta con la cual se podrían entender muchos de los procesos ecológicos de las
especies, producto de los componentes genéticos y de los ambientes en que se
desarrollan estas.
5

2.2 Germinación de cactáceas.
Factores como la temperatura, humedad y la luz (entre otros) fijan límites
latitudinales y altitudinales de distribución para muchas plantas, incluida la familia
Cactaceae (Godínez-Álvarez et al., 2003). A una escala más local, estas propiedades
juegan un papel fundamental, ya que la mayoría de los cactus muestran una
distribución espacial agregada, lo cual es un reflejo de la distribución aparchonada de
los recursos en los ambientes altamente heterogéneos que habitan.
Uno de los factores que explica su distribución agregada es la asociación de
plántulas de cactáceas con plantas adultas de otras especies (Shreve, 1931; Turner et
al., 1966; McAuliffe, 1984; Valiente-Banuet & Ezcurra, 1991; Sosa & Fleming, 2002;
Flores & Jurado, 2003). Esta asociación también ha sido llamado “síndrome de planta
nodriza” (Niering et al., 1963) o “interacción nodriza-protegida” (Cody, 1993).
El estudio de dicha asociación fue reportado por primera vez por Shreve (1910,
1931), quien sugirió que el establecimiento de plántulas en zonas áridas era un evento
raro y esporádico dado por los regimenes de altas temperaturas y baja humedad del
suelo. En respuesta a estas condiciones estresantes, el establecimiento de plántulas de
muchas especies de cactus aparecía con más frecuencia bajo las copas de plantas
adultas de otras especies, las cuales proveían de un microambiente menos estresante
(Ellner & Shmida, 1981). En esta interacción, las plántulas se benefician del
microambiente creado por la planta adulta, como el amortiguamiento de temperaturas
extremas, mayor disponibilidad de agua, menor evapotranspiración y mayor cantidad de
nutrientes en el suelo.
6

Flores y Jurado (2003) realizaron una compilación de trabajos respecto a la
interacción nodriza-protegida, encontrando dos patrones principales: que en los
ecosistemas áridos y semi-áridos es donde frecuentemente se reporta dicha interacción
y que las cactáceas es la familia con el mayor número de especies asociadas con
alguna nodriza.
En lo que respecta a la germinación, las cactáceas es una de las familias de
plantas perennes menos estudiadas (Rojas-Aréchiga et al., 1997) por lo que hay un
gran desconocimiento de la germinación para muchas especies, particularmente de los
requisitos lumínicos en la activación de la germinación para esta familia.
Estudios que han analizado la germinación de semillas de cactáceas en
condiciones de campo, consideran al agua y la temperatura como los factores más
importantes que limitan este proceso (Martínez-Holguín, 1983; Nobel, 1988; Rojas-
Aréchiga et al., 1997; Valiente-Banuet et al., 2002), mientras que la luz resulta ser un
factor que impacta de manera indirecta en la germinación de semillas, puesto que el
microambiente de sombra creado por el dosel sólo se visualiza como un buffer de
temperatura y humedad del suelo (Turner et al., 1966; Valiente-Banuet & Ezcurra,
1991), restando importancia a la luz en la germinación.
Aunque trabajos realizados en laboratorio muestran una tendencia contraria,
pues han demostrado que la exposición a determinada calidad de luz puede modificar
la respuesta germinativa en especies de cactáceas (ver Alcorn & Kurtz, 1959;
McDonough, 1964; Maiti et al., 1994; Rojas-Aréchiga et al., 1997; Zimmer & Buttner,
1982; Arias & Lemus, 1984), alterando dicho espectro lumínico a la germinación.
Según Rojas-Aréchiga et al. (1997), se podría hacer una clasificación de la
germinación de los cactus con respecto a la luz de acuerdo a su forma de vida: los
7

cactus de barril son fotoblásticos positivos (germinan sólo bajo condiciones de
irradiancia, Alcorn & Kurtz, 1959; McDonough, 1964; Rojas-Aréchiga et al., 2001);
mientras que los cactus columnares, pueden ser indiferentes a la luz (fotoblásticos
neutros, es decir, germinan bajo cualquier tratamiento lumínico, Rojas-Aréchiga et al.,
1997; Rojas Aréchiga & Vázquez-Yanes, 2000) o fotoblásticos positivos (Alcorn & Kurtz,
1959; McDonough, 1964; Rojas-Aréchiga et al., 2001).
El factor ambiental de interés para ser estudiado en el presente trabajo fue la luz,
pues ésta representa uno de los factores más heterogéneos, espacial y temporalmente
de entre los que afectan a las plantas (Gutterman, 1993; Rojas-Aréchiga et al., 1997;
Rojas-Aréchiga & Vázquez-Yanes, 2000).
Debido a su naturaleza energética, la luz representa un factor indispensable, por
lo que la acción individual o conjunta con otros factores de estrés ambiental pueden
poner a prueba la capacidad de resistencia y adaptación de las plantas a un medio
determinado (Manrique, 2003).
En este sentido, la copa de los árboles puede restringir la calidad y cantidad de la
luz que llega a las plantas que crecen bajo su sombra (i.e. disminución en la relación de
rojo: rojo lejano del espectro lumínico) (Rhodes & Stern, 1978; Gilbert et al., 2001). Las
semillas tienen fitocromos que les ayudan a distinguir la calidad e intensidad lumínica,
permitiéndole alterar la germinación (Roundy et al., 1992; Vazquez-Yanes et al., 1996;
Casal & Sanchez, 1998).
La variación en la germinación en respuesta a la luz puede ser considerada una
respuesta evolutiva de las plantas a la sombra producida por la vegetación (Smith,
1982), es decir, dependiendo de la composición espectral de la luz se estimulará o no la
respuesta germinativa de las semillas.
8

De acuerdo a ésta composición espectral, las especies se agrupan en patrones
de establecimiento como tolerantes a la luz, (aquellas especies pioneras beneficiadas
por los claros del dosel) y las intolerantes a la luz (especies de estadio sucesional
tardío), que necesitan de cobertura vegetal para evitar una serie de factores que
afectarían su desarrollo (i.e desecación). (Bazzaz, 1979; Bazzaz & Pickett, 1980;
Swaine & Whitmore, 1988).
Diversos trabajos han demostrado variación inter-específica de la germinación en
respuesta a la luz (Silvertown, 1980; Ballaré, 1994; Olff et al., 1994, Vázquez-Yanes &
Orozco-Segovia, 1994; Bazzaz, 1996; Flores et al., 2006). Sin embargo, poco se
conoce acerca de las diferencias intra-específicas en respuesta a este último factor,
puesto que sólo se han evaluado diferencias intra-específicas en la respuesta
germinativa con variables como la latitud, elevación, humedad y nutrientes en el suelo
(Baskin & Baskin, 1998).
2.3 Germinación y masa de semillas.
Además de la luz, otro factor que puede impactar de manera directa la
germinación es la masa de las semillas. Por lo general, en semillas grandes se obtienen
los mayores porcentajes de germinación o emergencia con respecto a semillas
pequeñas, debido principalmente a las reservas alimenticias dentro de la semilla (Grime
& Jeffrey, 1965; Baker, 1972; Silvertown, 1981).
Aunque diversos trabajos asumen que el tamaño o la masa de las semillas es
relativamente constante en especies de plantas que presentan un crecimiento
indeterminado (ver Harper et al., 1970), grandes variaciones en estos fenotipos han sido
9

reportados dentro de poblaciones (Harper & Obeid, 1967; Wulff, 1973; Waller, 1982;
Kidson & Westoby, 2000) y entre poblaciones (Baker, 1972; Schimpf, 1977) para una
misma especie. Además, estas variaciones pueden afectar significativamente la
germinación e incrementar dichas diferencias si se someten a diversas condiciones
ambientales (Wulff, 1986).
Si bien los mecanismos que originan y mantienen la variación en la masa de las
semillas en un gradiente geográfico son poco claros (Leishman et al., 2000), la variación
en la masa de las semillas con respecto a dicho gradiente pueden variar
sistemáticamente con la latitud y altitud, producto de las diferencias en la temperatura,
la radiación solar y la lluvia (Murray et al., 2004).
2.4 Planteamiento del problema.
Se eligió a Cephalocereus palmeri var. sartorianus (Rose) Krainz.,. como modelo
de estudio, ya que se distribuye a lo largo de todo el gradiente altitudinal de la selva
baja caducifolia del centro del estado de Veracruz. Dicho gradiente puede modificar por
un lado la respuesta germinativa en cada uno de los sitios donde se localiza la especie,
producto de la heterogeneidad ambiental (i.e. la calidad y cantidad de luz que llega a la
semilla puede variar en respuesta a las variaciones en la cobertura vegetal de cada
sitio) o por otro lado, esa amplia distribución puede dar paso a la existencia de
genotipos especializados a distintas calidades lumínicas para cada sitio, estimando así
la posible evolución de la norma de reacción.
10

Así mismo, la especie puede ser utilizada también como un modelo para
investigaciones que evalúen la germinación en cactáceas columnares y en las que sus
poblaciones se encuentren con alta heterogeneidad ambiental.
El estudio también evalúa la existencia de diferentes magnitudes en la plasticidad
fenotípica a nivel inter-sitios, que pueda determinar así una posible correlación entre los
sitios de acuerdo con su heterogeneidad ambiental y el nivel de plasticidad en la
germinación.
Además, el trabajo estimó la masa de semillas de cada familia genética para
determinar si los resultados de germinación se debían a la masa de las semillas (i.e.
efecto de la masa en las diferencias de la germinación de semillas en los diferentes
tratamientos lumínicos) o al atributo del genotipo para responder a las señales
ambientales. Así mismo, se evaluó si las variaciones en la masa de semillas inter-sitios
fueron producto del efecto ambiental sobre éstas.
La información de la especie en los requisitos ambientales para su germinación,
así como la información de genética cuantitativa en la germinación inter e intra-sitio
podría ayudar a entender mejor algunos aspectos ecológicos de la especie.
3. OBJETIVOS
3.1 Objetivo general.
Determinar si existe variación de la germinación en función de la calidad de la
luz, y en ese caso determinar si esa variabilidad se debe a plasticidad fenotípica.
11

3.2 Objetivos específicos.
Evaluar el efecto de la calidad de la luz en la germinación de C. palmeri var.
sartorianus.
Evaluar el efecto de la masa de las semillas en la germinación de C. palmeri var.
sartorianus.
Analizar la variabilidad fenotípica (variación inter e intra-sitio) en la germinación
de C. palmeri var. sartorianus, bajo dos condiciones de luz.
Estimar las correlaciones genéticas entre la respuesta de la germinación a las
distintas calidades de la luz.
4. MÉTODOS
4.1 La especie de estudio.
Se utilizó a Cephalocereus palmeri var. sartorianus (Rose) Krainz.,. una cactácea
columnar candelabriforme (Foto 1).
Foto 1. Cephalocereus palmeri var. sartorianus (Rose) Krainz.,. (Fotos: Antonio
Miranda).
12

Es una cactácea que mide de tres a cinco metros de altura con ramas casi
erectas. Sus ramas son de siete a diez cm de diámetro, color verde azulado. Posee
siete costillas de dos cm de alto con un surco a cada lado de las aréolas marcando los
podarios. Las aréolas son próximas y distantes entre sí a 1.5 cm, provistas de pelos
finos. Con frecuencia posee una espina central de un cm de color amarillo paja y
después grisácea y siete u ocho espinas radiales. Sus aréolas floríferas forman un
seudocefalio apical y lateral con numerosos pelos de cuatro a seis cm de longitud,
blancos. Las flores son campanuladas de seis a ocho cm de largo de color rosa sucio.
El fruto es globoso de cuatro a seis cm de diámetro, al principio verde, después de color
púrpura o rojizo. Las semillas tienen dos mm de longitud, son negras, brillantes y
finamente punteadas (la descripción de la especie se tomó con base en Bravo-Hollis,
1978).
En el Estado de Veracruz la especie se ha colectado en sitios como Huatusco,
Orizaba, Fortín, Puente Nacional, Las Vigas y Xalapa. En el Estado de Oaxaca se ha
colectado en Ixtlán de Juárez y en Tuxtepec.
Las flores de C. plameri var. sartorianus son hermafroditas y poseen rasgos
morfológicos y funcionales asociados a la polinización por murciélagos (ver Fleming, et
al., 1996; Valiente-Banuet, et al., 1996), entre las principales especies de murciélagos
que visitan las flores se encuentran Choeroniscus godmani, Glossophaga sp.,
Leptonyctetis nivalis y L. curasoae, ésta ultima puede realizar vuelos hasta de 30 km en
búsqueda de alimentos.
Según Munguía-Rosas, Sosa & Jácome (en preparación), en la parte central de
Veracruz el periodo de floración ocurre de abril-agosto y la fructificación y dispersión
sucede tres semanas posteriormente a cada uno de los tres o cuatro picos de floración;
13

siendo los murciélagos, lagartijas, hormigas y aves los principales animales encargados
de la dispersión.
Además, según los autores, la especie es xenógama y las flores poseen cierto
nivel de auto-compatibilidad, aunque no se sabe si las semillas producidas por la auto-
cruza experimentan depresión por endogamia.
4.2 Los sitios de estudio.
La variación en rasgos de historia de vida es común en especies que tienen una
distribución geográfica amplia (Roff, 1992; Stearns, 1992). Esta variación puede ser
creada por diferencias en la disponibilidad y calidad de los recursos entre los hábitats,
considerándose como una evidencia de estrategia adaptativa, producto de la
divergencia ambiental entre los sitios.
Debido a que los cambios ambientales pueden ser predecibles de acuerdo a la
elevación (Sears & Angilletta Jr., 2003), el gradiente altitudinal provee una oportunidad
para investigar la fuente de variación (i.e. la luz) en la germinación. Por lo que se
eligieron cinco sitios ubicados a lo largo de todo el gradiente altitudinal de la selva baja
caducifolia del estado de Veracruz en el que se distribuye C. palmeri var. sartorianus
(Figura 1).
14

#
#
#r#
CARDELCHICHICAXTLE
PUENTE NACIONAL
XALAPA
VERACRUZ
TAMARINDO
N
19°2
'00"
19°2'00"
19°1
7'30"
19°17'30"
19°3
3'00
" 19°33'00"
96°44'30"
96°44'30"
96°29'00"
96°29'00"
96°13'30"
96°13'30"
GOLFO DE MÉXICO
# Localidades
Carretera Xalapa-Veracruz
Límite Municipal
r Chichicaxtle
Simbología
10 0 10 20 Km
◙ TE (894 msnm)
◙ SI (867 msnm)
Simbología TE = Tenampa
Figura 1. Mapa de la República Mexicana donde se indica la ubicación de los sitios de
Cephalocereus palmeri var. sartorianus estudiados y su altitud.
Los cinco sitios son: TE= Tenampa, SI= San Ignacio, LR= Loma de Rogel, PA=
Puente Actopan y PN= Puente Nacional. La variación altitudinal de los cinco sitios
determina cambios en otras variables ambientales como son el tipo de suelo,
precipitación, temperatura, así como en la composición de la vegetación. Los cambios
en la vegetación se traducen en modificaciones en la altura y estructura de las copas de
los árboles, lo cual altera los niveles de luz recibida a nivel del suelo en cada sitio.
La descripción de cada sitio de acuerdo al gradiente es la siguiente: TE ubicada
a 894 msnm, cuenta con un suelo rocoso de origen volcánico de reciente formación, la
SI = San Ignacio LR = Loma de Rogel PA = Puente Actopan PN = Puente Nacional
◙ PA (259 msnm)
Xalapa ◙ LR (640 msnm)
◙ PN (87 msnm)
Veracruz
15

precipitación promedio anual es de 1741.1 mm, y una temperatura media de 17.11 ºC
(García, 2004). La vegetación arbórea y arbustiva es escasa, aquí se presenta la menor
cobertura vegetal con respecto a cada uno de los sitios (ver Figura 2) y las especies
dominantes son Lysiloma acapulcensis, Dodonacea viscosa, Quercus olioides,
Cnidoscolus acantifolius y Tonduzia longifolia. Los individuos de C. palmeri var.
sartorianus son de talla media; además de ser abundante, es la única cactácea
columnar presente en la zona.
En el sitio de SI (867 msnm) el estrato arbustivo presente es mayor con respecto
a la población de Tenampa (ver Figura 2), aunque la composición de especies es
parecida a la de TE. La vegetación es dominada principalmente por leguminosas de
baja altura como L. acapulcensis, L. microphylla, además de Q. olioides, D. viscosa, C.
acantifolius y T. longifolia. Los individuos de C. palmeri var. sartorianus presentan pocas
ramificaciones y al igual que en TE, es la única cactácea columnar presente en el
paisaje.
El sitio intermedio del gradiente es LR (640 msnm), los individuos de C. palmeri
var. sartorianus son de gran y mediana estatura, con ramificaciones hasta del tercer
orden; se pueden observar parches de vegetación que aumentan de manera
significativa la cobertura vegetal con respecto a los anteriores sitios (ver Figura 2), las
especies leñosas dominantes son T. longifolia, L. acapulcensis, D. viscosa, Acacia
cornigera, C. acantifolius, Comacladia engleriana y Plumeria rubra. En este sitio el suelo
es más desarrollado (se presenta todavía el suelo rocoso aunque en menor proporción
que en los sitios de TE y SI), la precipitación promedio anual en el sitio es de 1053.5
mm y la media de la temperatura anual es de 22.34 ºC (García, 2004). Además se
16

observa la incorporación de Neobuxbaumia euphorbioides (otra cactácea columnar) en
el paisaje.
Por último se encuentran los sitios de PA y PN (256 y 87 msnm respectivamente)
aquí los individuos de C. palmeri var. sartorianus son muy ramificados. La vegetación
arbórea y arbustiva para PA es dominada por Ceiba aescutifolia, Cochlospermum
vitifolium, C. engleriana, Esenbeckia berlandieri, Ipomoea arborescens, Leucaena
lueucocephalus y Randia laetevirens; mientras que para PN son: Tabebuia chrysantha,
Bursera simaruba, C. aescutifolia, Cordia dentata, Croton cortecianus, E. berlandieri y
Luehea candida; para ambos sitios se puede apreciar un porcentaje de cobertura mayor
con respecto al resto de los sitios (ver Figura 2). El suelo cuenta con mayor contenido
de materia orgánica, la precipitación promedio anual para PA es de 848.8 mm y la
temperatura promedio es de 24.69 ºC. (García, 2004); mientras que para PN la
precipitación promedio anual es de 1215.8 mm y la temperatura media es de 27.14 ºC
(García, 2004). Cabe destacar que en el último sitio se observó el mayor tamaño de
individuos y frutos con respecto al resto de los sitios. En PA y PN se integran al paisaje
Neobuxbaumia scoparia y Stenocereus griseus, (cactáceas columnares).
4.3 Diseño experimental para medir la cobertura vegetal inter-sitios y la asociación de
plántulas de C. palmeri var. sartorianus en respuesta a la luz.
En diciembre del 2007 se realizaron de manera aleatoria cinco transectos de 50
m para cada uno de los sitios (i.e. 5 transectos x 5 sitios). Se determinó el porcentaje de
cobertura vegetal inter-sitios registrando el inicio y fin de cada una de las copas de los
árboles que pasaban por dicho transecto.
17

Para determinar el patrón de asociación de plántulas de C. palmeri var.
sartorianus, todos los individuos sexualmente inmaduros (<90 cm de altura) que se
localizaron a 2.5 m de cada lado del transecto anteriormente descrito, fueron
registrados de acuerdo a su ubicación con respecto a las dos condiciones de luz (bajo
la copa de un árbol o si se encontraban sin cobertura del dosel). Lo anterior se realizó
con la finalidad de probar la hipótesis nula de que las plántulas no se asociaban con
alguna condición lumínica en particular en ningún sitio.
4.4 Diseño experimental para medir la plasticidad fenotípica de la germinación de C.
palmeri var. sartorianus en respuesta a la luz.
En junio del 2006 se colectó un fruto maduro por cada uno de 15 individuos
(familias genéticas) de cada sitio. A los frutos se les extrajeron las semillas y se les
retiró el resto del mucílago mediante lavado, posteriormente las semillas se pusieron a
secar a temperatura ambiente y se almacenaron en bolsas de papel.
Las semillas de cada planta fueron consideradas como una familia genética,
éstas tienen la mitad del parentesco de la planta madre, aunque se desconoce la
paternidad, pues pudieron haberse formado por autofecundación o fertilización cruzada.
Para evaluar el efecto lumínico se utilizaron dos niveles de luz: luz blanca, que
simuló las condiciones de una semilla en suelo descubierto (hábitat abierto) y luz rica en
rojo-lejano, que simuló las condiciones de una semilla bajo la copa de un árbol (hábitat
cerrado), puesto que la reducción de la proporción de rojo-lejano en la radiación
recibida a nivel de suelo funciona como un indicador de la presencia de un dosel de
plantas que debe ser atravesado por la luz incidente (Smith & Whitelam 1997).
18

Para simular las condiciones de luz rica en rojo-lejano dentro de la cámara de
germinación, se cubrieron las cajas petri con filtros Lee rojo y azul (número 071 y 182
respectivamente). De acuerdo con Valencia-Díaz & Montaña (2003), la proporción de
rojo/rojo-lejano obtenida con esos filtros es de 0.08 y la densidad de flujo de fotones en
el rango de 400 a 750 nm es de 0.51 μmol/m2/s mientras que para la luz blanca, la
proporción de rojo/rojo-lejano fue de 5.08 y la radiación fotosintéticamente activa de
32.79 μmol/m2/s.
Los experimentos de germinación se realizaron en cámara de germinación (Lab-
Line Biotronette, Melrose Park, IL, USA). Para esto se pusieron a germinar las semillas
de cada una de las 15 familias genéticas de cada sitio en cajas petri con agar al 2%
como sustrato. Además se agregó al agar Micostatin (Nistatina 500,000 U, 1g l-1) y
Cloramfenicol (2 ml l-1) para prevenir la contaminación por hongos y bacterias durante el
experimento.
La condición para la germinación fue un fotoperiodo de 12 horas luz a 25ºC y 12
horas oscuridad a 20ºC. La germinación (considerada una vez que la radícula fuese
visible) se registró diariamente por 30 días para la condición de luz blanca, mientras que
para el tratamiento de luz rica en rojo-lejano se registró la germinación hasta el término
del experimento (30 días posteriores a la siembra).
La plasticidad en la germinación se evaluó con el siguiente diseño: se pusieron a
germinar en cada uno de los tratamientos de luz (blanca y rica en rojo-lejano) las
semillas de las 15 familias (120 semillas por familia genética) de cada una de los cinco
sitios con tres repeticiones por tratamiento (i.e. 2 condiciones de luz x 5 sitios x 15
familias genéticas x 3 repeticiones = 450 unidades experimentales). La unidad
experimental fue de 20 semillas por caja petri.
19

4.5 Diseño experimental para estimar la variación intra e inter-sitio en la masa de
semillas de C. palmeri var. sartorianus.
Para evaluar la variación inter e intra-sitio en la masa de las semillas, se siguió
con la metodología empleada para estimar la plasticidad fenotípica en la germinación,
pues se respetó el origen (familia genética y sitio) de las semillas pesadas, el método de
colecta, el lavado y almacenado de semillas.
Posteriormente 50 grupos de 10 semillas, cada uno por familia genética de cada
sitio (i.e. 50 repeticiones de 10 semillas por cada una de las 15 familias de cada una de
los cinco sitios= 3750 unidades experimentales) fueron pesadas para registrar su masa
(g) en una balanza analítica Sartorius modelo 100.
El pesaje en grupos de a 10 semillas se realizó para disminuir el error de
medición que se habría registrado si hubiesen pesado las semillas de manera individual
ya que la masa aproximada para cada grupo fue de menos de 0.0012 mg.
Por último, cada uno de los pesajes se multiplicó por 1000 para aumentar así la
suma de cuadrados y observar las posibles diferencias entre los factores.
4.6 Diseño experimental para evaluar el efecto ambiental sobre la variación inter-sitio en
la masa de semillas y la germinación de C. palmeri var. sartorianus.
Debido a que las semillas se obtuvieron de diferentes sitios, los cuales difieren
entre sí en variables ambientales, se estimó el posible impacto que estas variables
pueden tener sobre la germinación y la masa de semillas a nivel inter-sitio.
20

En ambos casos (i.e. germinación y masa de semillas), se respetó el origen de
las semillas (sitio) y se tomaron en cuenta variables ambientales, como la precipitación,
temperatura, altitud y cobertura vegetal que se registraron para cada uno del los sitios.
4.7 Análisis estadísticos.
Se analizó la variación de los porcentajes de cobertura vegetal ínter-sitio, usando
un ANDEVA de una vía como una función de los sitios.
Para analizar la asociación de plántulas bajo alguna condición lumínica en
particular, se aplicó una prueba de heterogeneidad de chi cuadrada (X2) al total de
plántulas que se registraron en cada uno de los cinco transectos de cada sitio en las
dos condiciones de luz (i.e. 5 transectos x 5 sitios x 2 condiciones de luz); como los
valores fueron menores a la X2 de tablas (X20.05,4= 9.488) se pudieron agrupar las
réplicas de los transectos y se obtuvo un solo valor observado y esperado para cada
condición lumínica de cada uno de los sitios, al que finalmente se le practicó la prueba
de X2 (i.e. condición lumínica x sitio) (Zar, 1999).
Para evaluar la variación ínter-sitio de los porcentajes de germinación, se analizó
la variabilidad de la germinación como una función de los sitios, de las familias
genéticas (anidadas en los sitios con 15 niveles) y de las condiciones de luz (blanca y
rica en rojo-lejano) usando un ANDEVA para cada una de las variables de respuesta.
Para evaluar la variación intra-sitio se realizó, para cada sitio, un ANDEVA donde
se analizó la variabilidad de la germinación en función de la calidad de la luz (luz blanca
y rica en rojo-lejano) y de la familia genética. La interacción entre estas variables es la
interacción genotipo x ambiente necesaria para determinar si hay plasticidad fenotípica
21

(Via & Lande, 1985; Via, 1994). Posteriormente se construyeron los gráficos de las
norma de reacción de cada sitio.
Los porcentajes de germinación empleados en los ANDEVAS fueron
transformados al arcoseno para normalizar la distribución de los datos. Se usó un
ANDEVA anidado de dos vías implementado en una hoja de cálculo siguiendo el
procedimiento de Montgomery (1976).
Además, para facilitar las comparaciones entre las normas de reacción, se
obtuvo un valor fenotípico por familia genética, el cual fue calculado como un cociente
entre el porcentaje de geminación mayor registrado en un tratamiento lumínico y el
porcentaje de germinación menor registrado para la misma familia genética en el
tratamiento de luz complementario, es decir, dicho cociente mostró el porcentaje en la
plasticidad (media ± error estándar) producto de las diferencias entre la germinación
bajo las dos condiciones lumínicas para cada familia genética.
También se calculó el valor fenotípico inter-sitio como el cociente entre el
porcentaje mayor y el porcentaje menor de germinación de cada sitio, obtenido bajo los
dos tratamientos de luz.
Para estimar el grado de dependencia genética en respuesta a la variabilidad
ambiental (calidad de la luz), se realizaron las correlaciones genéticas (rG: Falconer,
1952; Via & Lande, 1985; Via, 1994) para cada sitio y para la variable de respuesta
(germinación), las cuales se calcularon mediante correlaciones, usando los promedios
registrados en la germinación de cada una de las familias genéticas bajo cada uno de
los tratamientos de luz.
22

Para evaluar la variación ínter e intra-sitio en la masa de semillas, se analizó la
variabilidad como una función de los sitios (cinco niveles), de las familias genéticas
(anidadas en los sitios con 15 niveles).
Para evaluar el efecto entre la masa de semillas y la germinación, se obtuvo un
coeficiente de correlación para cada sitio con la media aritmética de la germinación y la
masa de semillas para cada familia genética sometida en cada condición de luz (blanca
y rica en rojo-lejano).
Por último, para evaluar el efecto ambiental sobre la variación inter-sitio en la
masa de semillas y en la germinación, se realizaron correlaciones entre cada variable
ambiental (temperatura, precipitación y altitud) y cada una de las variables de respuesta
(germinación y masa de semillas). Además, en cada caso se calculó la ecuación de la
recta de mejor ajuste (Zar, 1999).
5. RESULTADOS
5.1 Cobertura vegetal inter-sitio y asociación de plántulas en respuesta a las
condiciones lumínicas.
El porcentaje de cobertura vegetal fue significativo para los sitios (92.5 % de la
varianza explicada, Tabla 1), apreciándose un aumento gradual en la cobertura vegetal
conforme disminuye el gradiente altitudinal (Figura 2).
23

Tabla 1. Análisis de varianza del porcentaje de la cobertura vegetal registrado para
cada uno de los sitios.
Fuente de variación S. C
varianza explicada gl C. M F
Valor crítico de F P
Sitio 9397.36 92.5 4 2349.34 61.65 2.86 ≤0.0001
Error 762.04 7.5 20 38.10
Total 10159.4 100 24
Fuente de variación: Sitio= Tenampa, San Ignacio, Loma de Rogel, Puente Actopan y
Puente Nacional. Transecto= 50 m; Cinco réplicas por sitio.
Cobertura vegetal entre los sitios
0
20
40
60
80
100
TE SI LR PA PN
sitios
Cob
ertu
ra v
eget
al (%
)
a b
b
c c
Figura 2. Porcentajes de cobertura vegetal (± error estándar) de los cinco sitios. TE=
Tenampa; SI= San Ignacio; LR= Loma de Rogel; PA= Puente Actopan; PN= Puente
Nacional. Letras diferentes indican diferencias significativas (Prueba de Tukey p<0.05).
24

No existe asociación de plántulas bajo el dosel para ningún sitio, puesto que las
plántulas muestreadas en cada uno de éstos presentaron una frecuencia igual bajo las
dos condiciones lumínicas (X2= 5.48 < X20.05,4= 9.48) (Tabla 2).
Tabla 2. Tabla de contingencia aplicada a los resultados obtenidos de la asociación de
plántulas bajo cada una de las condiciones de luz en los cinco sitios.
Sitio
Plántulas bajo el dosel
Plántulas en espacios abiertos
Total de plántulas
TE
9 9.81*
14 13.18*
23
SI
16 13.85*
11 13.14*
27
LR
22 20.40*
15 16.59*
37
PA
21 22.19*
5 3.80*
26
PN
19 22.17*
6 2.82*
25
∑
87 88.44*
51 49.55*
138
Niveles del tratamiento: Sitio= TE=Tenampa, SI=San Ignacio, LR=Loma de Rogel,
PA=Puente Actopan y PN=Puente Nacional; Condiciones de luz= bajo dosel y en
espacios abiertos; (*) Valores esperados; Total de plántulas= número total de plántulas
encontradas en cada uno de los sitios; Valor de X2 0.05, 4= 9.48
5.2 Germinación y calidad de la luz.
El factor más importante para la germinación fue la familia anidada en el sitio
(77.2% de la varianza explicada). También tuvieron efectos significativos la interacción
luz x familia y la interacción luz x sitio (5.8% y 2.0 % de la varianza explicada
respectivamente, Tabla 3).
25

Tabla 3. Análisis de varianza del arcoseno de la germinación como una función de
sitios, familias genéticas y tratamiento de luz.
Fuente de variación S. C
varianza explicada gl C. M F
Valor crítico de F P
Luz 188.03 0.13 1 188.03 1.56 3.98 0.2152
Sitio 7241.02 5 4 1810.2 1.12 2.5 0.3495
Familia (Sitio) 112143.84 77.20 70 1602.05 33.39 1.34 ≤0.0001
Luz * Sitio 2870.45 1.98 4 717.61 5.96 2.5 0.0003
Luz * Familia (Sitio) 8417.74 5.80 70 120.25 2.50 1.34 ≤0.0001
Error 14392.72 9.90 300 47.97
Total 145254 100
Fuente de variación: Sitio=Tenampa, San Ignacio, Loma de Rogel, Puente Actopan y
Puente Nacional; Familia (sitio)= 15 familias genéticas anidadas en sitio; Luz= luz
blanca y luz rica en rojo-lejano; Diseño factorial= 2 condiciones de luz x 5 sitios x 15
familias genéticas x 3 repeticiones por tratamiento= 450 unidades experimentales; La
unidad experimental fue de 20 semillas por caja petri.
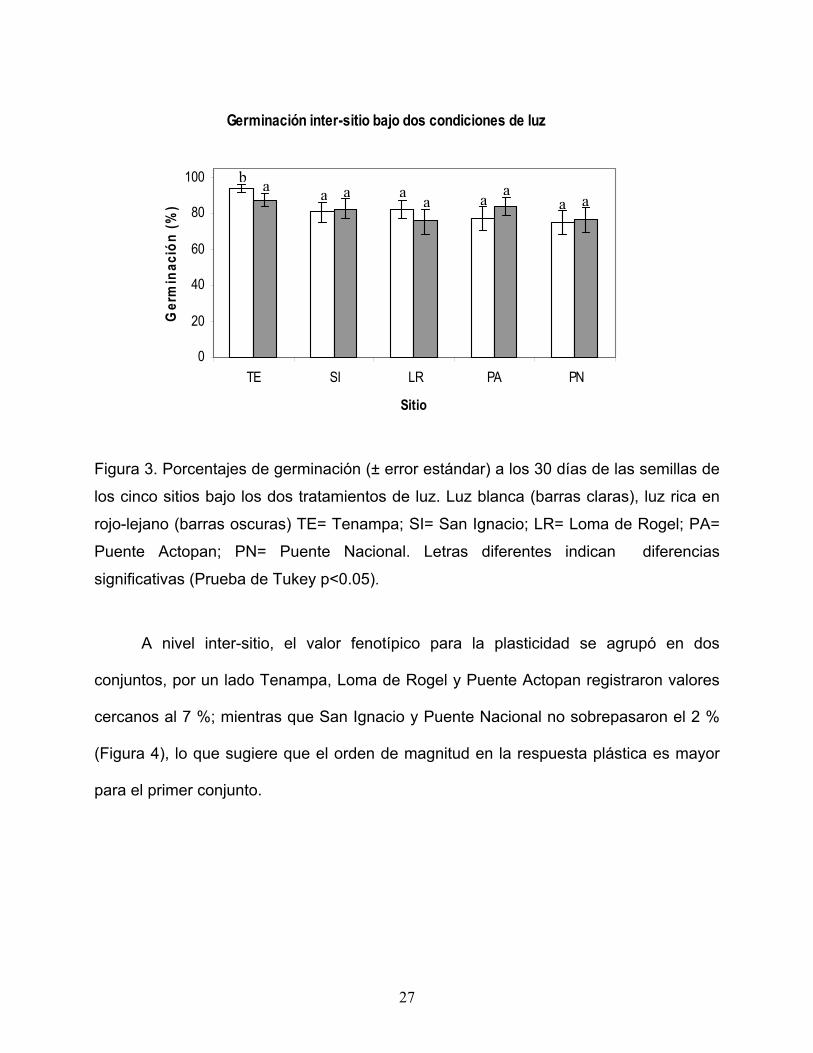
En la interacción tratamiento de luz * sitio se registró que para los sitios de San
Ignacio, Loma de Rogel, Puente Actopan y Puente Nacional, la germinación no difirió
significativamente entre los tratamientos lumínicos, mientras que la germinación para
Tenampa fue mayor en el tratamiento de luz blanca (Figura 3).
26
Germinación inter-sitio bajo dos condiciones de luz
0
20
40
60
80
100
TE SI LR PA PN
Sitio
Ger
min
ació
n (%
)b a a a aa
a aaa
Figura 3. Porcentajes de germinación (± error estándar) a los 30 días de las semillas de
los cinco sitios bajo los dos tratamientos de luz. Luz blanca (barras claras), luz rica en
rojo-lejano (barras oscuras) TE= Tenampa; SI= San Ignacio; LR= Loma de Rogel; PA=
Puente Actopan; PN= Puente Nacional. Letras diferentes indican diferencias
significativas (Prueba de Tukey p<0.05).
A nivel inter-sitio, el valor fenotípico para la plasticidad se agrupó en dos
conjuntos, por un lado Tenampa, Loma de Rogel y Puente Actopan registraron valores
cercanos al 7 %; mientras que San Ignacio y Puente Nacional no sobrepasaron el 2 %
(Figura 4), lo que sugiere que el orden de magnitud en la respuesta plástica es mayor
para el primer conjunto.
27

Plasticidad fenotípica inter-sitio
0
2
4
6
8
10
12
TE SI LR PA PN
Sitio
Valo
r fen
otíp
ico
(%)
a
b b
a a
Figura 4. Valor fenotípico inter-sitio expresado en porcentaje (± error estándar) para la
germinación de semillas en respuesta a los dos tratamientos de luz.
En la Figura 5 se muestran las normas de reacción (tratamientos de luz de las
familias genéticas de cada sitio). Las pendientes en las líneas (cada línea es una norma
de reacción) indican el grado de plasticidad fenotípica expresada para cada genotipo
(familia genética). Se observa una marcada variación en la plasticidad a nivel intra-sitio.
28

Tenampa
20
30
40
50
60
70
80
90
100
Luz blanca Rojo-lejano
Tratamiento de luz
Ger
min
ació
n (%
)San Ignacio
20
30
40
50
60
70
80
90
100
Luz blanca Rojo-lejano
Tratamiento de luz
Ger
min
ació
n (%
)
Loma de Rogel
20
30
40
50
60
70
80
90
100
Luz blanca Rojo-lejano
Tratamiento de luz
Ger
min
ació
n (%
)
Puente Actopan
20
30
40
50
60
70
80
90
100
Luz blanca Rojo-lejano
Tratamiento de luz
Ger
min
ació
n (%
)
Puente Nacional
20
30
40
50
60
70
80
90
100
Luz blanca Rojo-lejano
Tratamiento de luz
Ger
min
ació
n (%
)
Figura 5. Normas de reacción del porcentaje de germinación en respuesta a los dos
tratamientos de luz para las 15 familias de cada sitio.
29

Los valores fenotípicos muestran variación genética intra-sitio para la plasticidad
en todos los sitios del gradiente altitudinal (diferencias en la plasticidad fenotípica),
marcándose una mayor variación en San Ignacio, Loma de Rogel y Puente Actopan;
mientras que dicha variación es menor en los sitios de Puente Nacional y Tenampa,
localizadas al extremo sur y norte del gradiente respectivamente (Figura 6).
San Ignacio Plasticidad fenotípica
0
5
10
15
20
25
30
35
40
A B C D E F G H I J K L M N Ñ
familias genéticas
valo
r fen
otíp
ico
(%)
Tenampa Plasticidad fenotípica
0
5
10
15
20
25
30
35
40
A B C D E F G H I J K L M N Ñ
familias genéticas
valo
r fen
otíp
ico
(%)
Loma de Rogel Plasticidad fenotípica
0
5
10
15
20
25
30
35
40
A B C D E F G H I J K L M N Ñ
familias genéticas
valo
r fen
otíp
ico
(%)
Puente Actopan Plasticidad fenotípica
0
5
10
15
20
25
30
35
40
A B C D E F G H I J K L M N Ñ
familias genéticas
valo
r fen
otíp
ico
(%)
Puente Nacional Plasticidad fenotípica
0
5
10
15
20
25
30
35
40
A B C D E F G H I J K L M N Ñ
familias genéticas
valo
r fen
otíp
ico
(%)
Figura 6. Valor fenotípico expresado en porcentaje (± error estándar), para la
germinación de semillas en respuesta a los dos tratamientos de luz para las 15 familias
genéticas de cada sitio.
30

El ANDEVA aplicado a cada una de los sitios para evaluar la interacción luz *
familia genética, muestra que en todos los sitios la interacción resultó ser significativa,
Tenampa (F = 2.18, gl = 14, 89, p = 0.018), San Ignacio (F = 3.18, gl = 14, 89, p =
0.0008), Loma de Rogel (F = 2.67, gl = 14, 89, p = 0.004), Puente Actopan (F = 3.18, gl
= 14, 89, p = 0.0008) y Puente Nacional (F = 2.00, gl = 14, 89, p = 0.03) explican el
11.20 %, 5.25 %, 7.0 %, 6.12 % y 4.0 % de la varianza respectivamente. Estos
resultados indican que en todos los sitios hay plasticidad fenotípica en la respuesta de
la germinación a la calidad de luz.
5.3 Correlaciones genéticas entre la respuesta de la germinación a las distintas
calidades de la luz.
Los porcentajes de germinación bajo los dos tratamientos de luz estuvieron
correlacionados (es decir las correlaciones fueron distintas de 0) en los sitios de San
Ignacio, Loma de Rogel, Puente Actopan y Puente Nacional, confirmándose para esos
sitios una dependencia genética de los estados de carácter. Por el contrario, en el caso
del sitio de Tenampa dicha correlación no fue significativa (Tabla 4, Figura 7).
31

Tabla 4. Coeficiente de correlación intraclase (± error estándar) entre los porcentajes de
germinación obtenidos bajo dos calidades de luz en cada uno de los sitios de estudio.
Sitio
rG Germinación
error estándar rG Germinación
Tenampa 0.6928 0.1999
San Ignacio 0.8546 * 0.1439
Loma de Rogel 0.8795 * 0.1319
Puente Actopan 0.9062 * 0.1172
Puente Nacional 0.9308 * 0.1013
Niveles del tratamiento: Calidades de luz= luz blanca y luz rica en rojo-lejano; Valor
crítico (r 0.05, 2, 13 = 0.514); * Diferencias significativas.
32

rG San Ignacio
20
30
40
50
60
70
80
90
100
20 30 40 50 60 70 80 90 100
Luz blancaGerminación (%)
Roj
o-le
jano
Ger
min
ació
n (%
)
rG Tenampa
20
30
40
50
60
70
80
90
100
20 30 40 50 60 70 80 90 100
Luz blancaGerminación (%)
Roj
o-le
jano
Ger
min
ació
n (%
)
rG Loma de Rogel
20
30
40
50
60
70
80
90
100
20 30 40 50 60 70 80 90 100
Luz blancaGerminación (%)
Roj
o-le
jano
Ger
min
ació
n (%
)
rG Puente Actopan
20
30
40
50
60
70
80
90
100
20 30 40 50 60 70 80 90 100
Luz blancaGerminación (%)
Roj
o-le
jano
Ger
min
ació
n (%
)
rG Puente Nacional
20
30
40
50
60
70
80
90
100
20 30 40 50 60 70 80 90 100
Luz blancaGerminación (%)
Roj
o-le
jano
Ger
min
ació
n (%
)
rG= 0.6928 ± 0.19
rG= 0.8795* ± 0.13 rG= 0.9062* ± 0.11
rG= 0.9308* ± 0.10
rG= 0.8546* ± 0.14
Figura 7. Relación entre los porcentajes de germinación obtenidos con dos calidades de
luz (luz blanca y rojo lejano) con semillas producidas en 15 familias genéticas de 5 sitios
distintos. Para cada sitio se muestran los puntos correspondientes a las 15 familias
genéticas, y el valor del coeficiente de correlación intraclase (± error estándar). Valor
crítico (r 0.05, 2, 13 = 0.514); * Diferencias significativas.
33

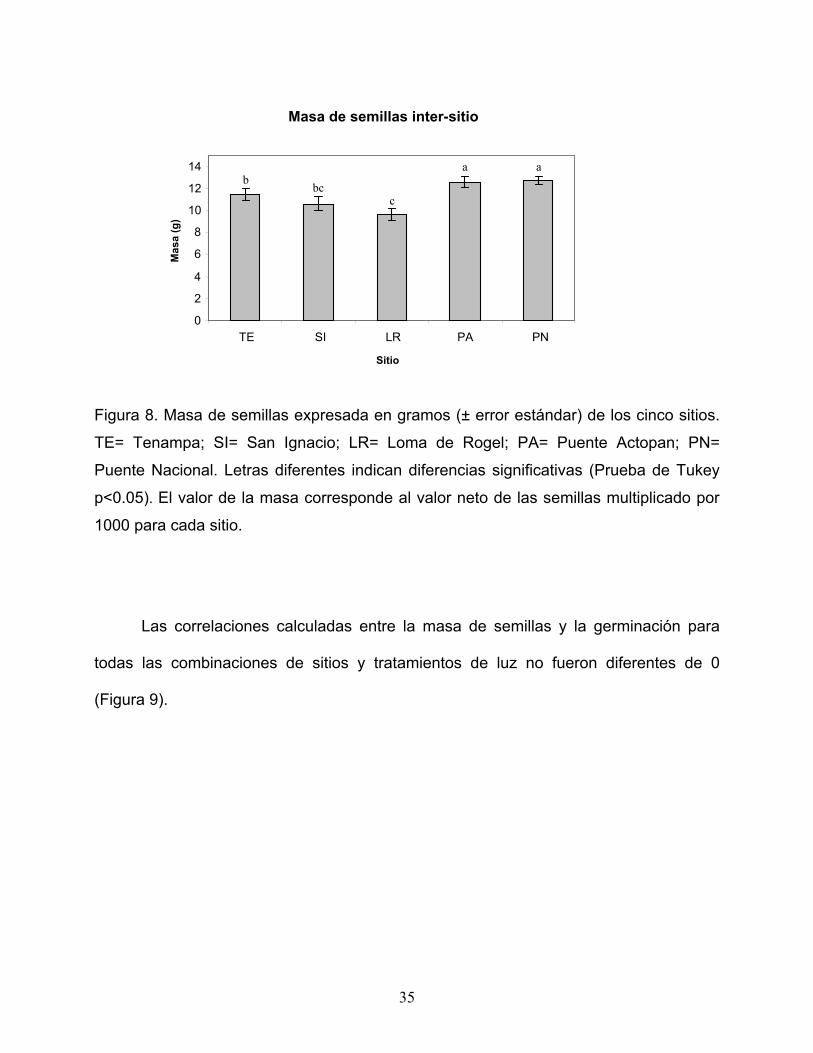
5.4 Masa de semillas.
El factor familia (anidado en sitio) explicó el 50.5 % de la varianza; mientras que el
factor sitio explicó el 19.05 % de la misma (Tabla 5). La mayor masa en las semillas se
registró en los sitios ubicados en la parte más baja del gradiente como Puente Nacional
y Puente Actopan (12.76 ± 0.38 g y 12.56 ± 0.53 g respectivamente); mientras que para
los sitios ubicados en la parte más alta Tenampa, San Ignacio y Loma de Rogel se
registraron las masas menores de semillas (11.45 ± 0.55 g, 10.57 ± 0.61 g y 9.62 ± 0.55
g respectivamente, Figura 8).
Tabla 5. Análisis de la varianza de la masa de semillas como una función de sitios y
familias genéticas.
Fuente de variación S. C.
Varianza explicada gl C. M. F
Valor crítico de F p
Sitio 5545.59 19.05 4 1386.39 6.59 2.97 ≤0.0001
Familia (Sitio) 14708.02 50.53 70 210.11 87.21 1.36 ≤0.0001
Error 8854.13 30.42 3675 2.41
Total 29107.75 100 3749
Fuente de variación: Sitio= Tenampa, San Ignacio, Loma de Rogel, Puente Actopan y
Puente Nacional; Familia (sitio)= 15 familias genéticas anidadas en sitio; Diseño
factorial= 50 repeticiones de 10 semillas por cada una de las 15 familias de cada una de
los cinco sitios= 3750 unidades experimentales)
34
Masa de semillas inter-sitio
0 2 4 6 8
10 12 14
TE SI LR PA PN
Sitio
Mas
a (g
) a a
cbc
b
Figura 8. Masa de semillas expresada en gramos (± error estándar) de los cinco sitios.
TE= Tenampa; SI= San Ignacio; LR= Loma de Rogel; PA= Puente Actopan; PN=
Puente Nacional. Letras diferentes indican diferencias significativas (Prueba de Tukey
p<0.05). El valor de la masa corresponde al valor neto de las semillas multiplicado por
1000 para cada sitio.
Las correlaciones calculadas entre la masa de semillas y la germinación para
todas las combinaciones de sitios y tratamientos de luz no fueron diferentes de 0
(Figura 9).
35

TENAMPA
0
20
40
60
80
100
6 8 10 12 14 16
Masa de semillas (g)
Ger
min
ació
n (%
)
SAN IGNACIO
0
20
40
60
80
100
6 8 10 12 14 16
Masa de semillas (g)
Ger
min
ació
n (%
)
LOMA DE ROGEL
0
20
40
60
80
100
6 8 10 12 14 16
Masa de semillas (g)
Ger
min
ació
n (%
)
PUENTE ACTOPAN
0
20
40
60
80
100
6 8 10 12 14 16
Masa de semillas (g)
Ger
min
ació
n (%
)
PUENTE NACIONAL
0
20
40
60
80
100
6 8 10 12 14 16
Masa de semillas (g)
Ger
min
ació
n (%
)
○ 0.5434 ± 0.233● 0.5104 ± 0.238
○ 0.2212 ± 0.270 ● 0.0501 ± 0.227
○ 0.3491 ± 0.259 ● 0.3926 ± 0.255
○ 0.4435 ± 0.248 ● 0.4612 ± 0.246
○ - 0.0993 ± 0.276 ● - 0.1510 ± 0.274
Figura 9. Relaciones entre el porcentaje de germinación y la masa de semillas
obtenidos bajo dos tratamientos de luz en cada uno de los sitios de estudio (○ familias
genéticas sometidas al tratamiento de luz blanca; ● familias genéticas sometidas al
tratamiento de luz rica en rojo-lejano). Se muestran también los coeficientes de
correlación (± error estándar) calculados para cada tratamiento de luz. Valor crítico (r
0.05, 2, 13 = 0.514). El valor de la masa corresponde al valor neto de las semillas
multiplicado por 1000 para familia genética de cada sitio.
36
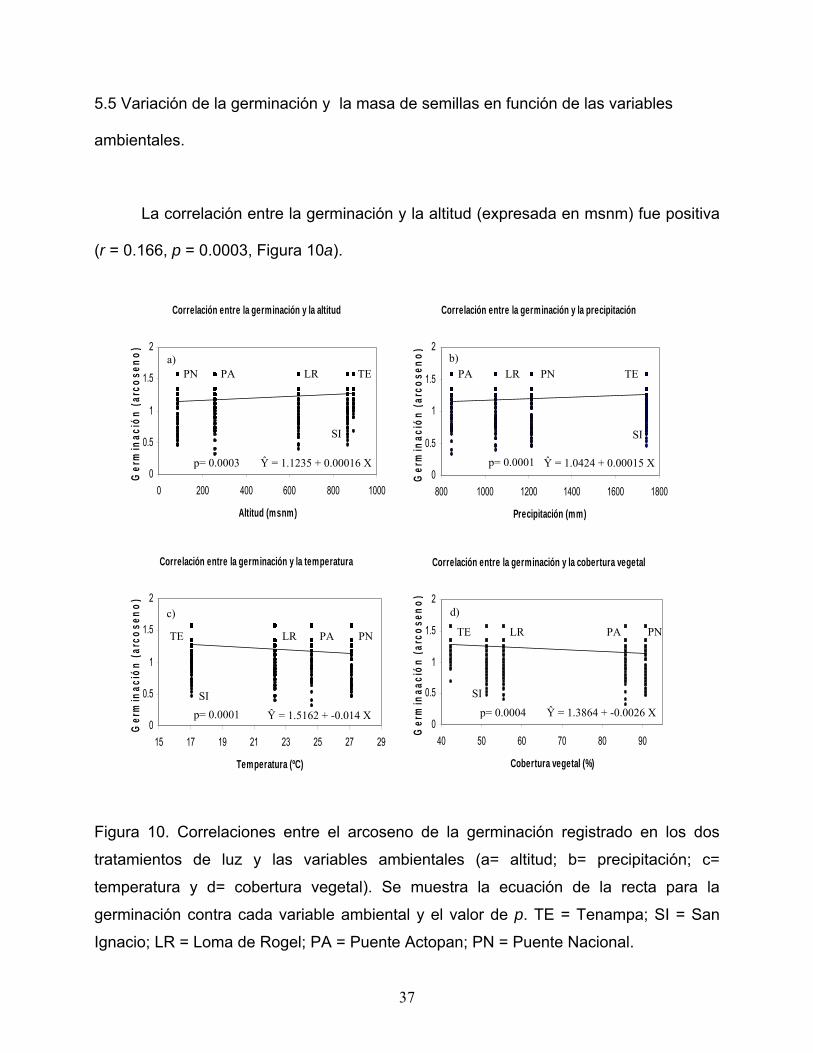
5.5 Variación de la germinación y la masa de semillas en función de las variables
ambientales.
La correlación entre la germinación y la altitud (expresada en msnm) fue positiva
(r = 0.166, p = 0.0003, Figura 10a).
Correlación entre la germinación y la precipitación
0
0.5
1
1.5
2
800 1000 1200 1400 1600 1800
Precipitación (mm)
Ger
min
ació
n (a
rcos
eno)
Correlación entre la germinación y la temperatura
0
0.5
1
1.5
2
15 17 19 21 23 25 27 29
Temperatura (ºC)
Ger
min
ació
n (a
rcos
eno)
Correlación entre la germinación y la altitud
0
0.5
1
1.5
2
0 200 400 600 800 1000
Altitud (msnm)
Ger
min
ació
n (a
rcos
eno) b) a)
PN PA LR TE PA PN LR TE
SI SI
p= 0.0001 p= 0.0003 Ŷ = 1.0424 + 0.00015 X Ŷ = 1.1235 + 0.00016 X
Figura 10. Correlaciones entre el arcoseno de la germinación registrado en los dos
tratamientos de luz y las variables ambientales (a= altitud; b= precipitación; c=
temperatura y d= cobertura vegetal). Se muestra la ecuación de la recta para la
germinación contra cada variable ambiental y el valor de p. TE = Tenampa; SI = San
Ignacio; LR = Loma de Rogel; PA = Puente Actopan; PN = Puente Nacional.
PA
SI
TE LR PN
Correlación entre la germinación y la cobertura vegetal
0
0.5
1
1.5
2
40 50 60 70 80 90
Cobertura vegetal (%)
Ger
min
aaci
ón (a
rcos
eno)
d) c) PN TE LR PA
SI
p= 0.0004 Ŷ = 1.3864 + -0.0026 X p= 0.0001 Ŷ = 1.5162 + -0.014 X
37

Una tendencia parecida mostró la correlación entre la germinación y la
precipitación (expresada en mm), con una r = 0.148 y p = 0.001 (figura 10b); mientras
que la correlación entre la germinación y la temperatura (ºC) muestra una tendencia
opuesta a las anteriores (r = -0.179; p = 0.0001) pues conforme aumenta la
temperatura, disminuye la germinación (Figura10c) y esta misma tendencia se presenta
en la correlación entre la germinación y la cobertura vegetal (r =.-0.165 y p = 0.0004,
Figura 10d).
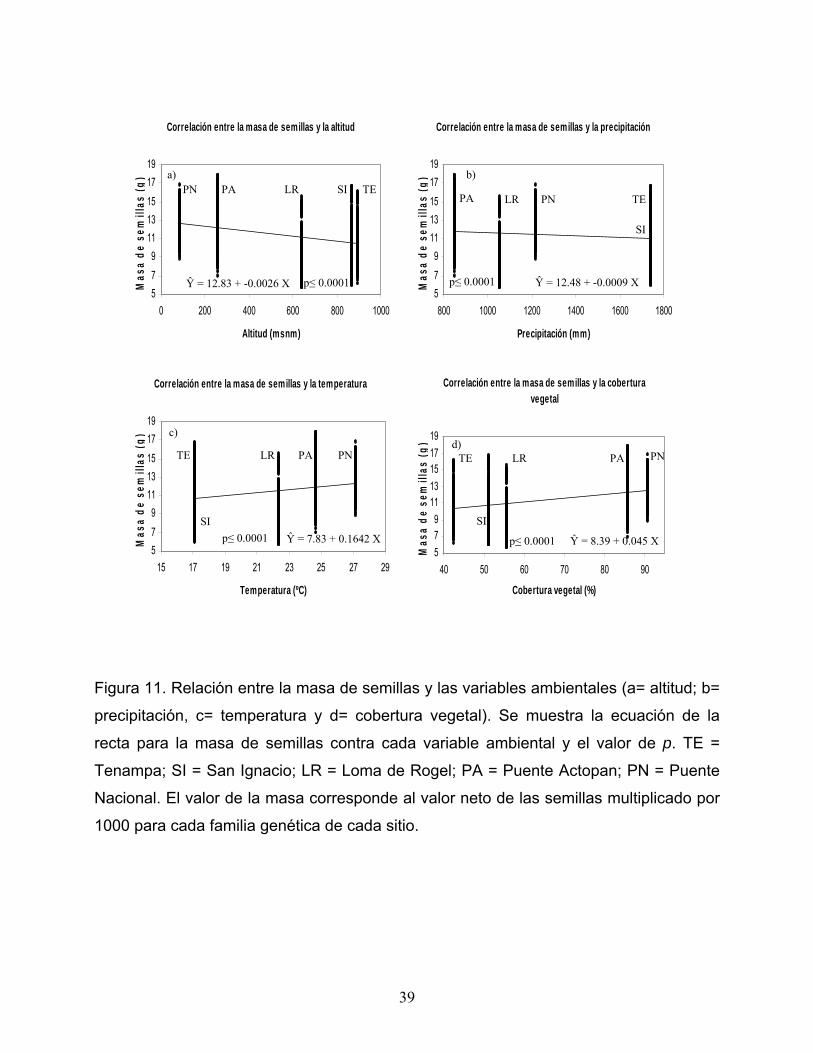
Todas las correlaciones realizadas para evaluar el efecto ambiental en la masa
de semillas resultaron también significativas.
La correlación entre la masa de semillas y la altitud, así como la correlación entre
la masa de semillas y la precipitación mostraron tendencias negativas (r = -0.349; p ≤
0.0001 y r = -0.123; p ≤ 0.0001 respectivamente), pues como se puede apreciar en las
figuras 11a y 11b, conforme se aumenta la altitud y la precipitación, disminuye la masa
de las semillas. Contrariamente, la correlación entre la masa de semillas y la
temperatura (r = 0.272 y p ≤ 0.0001, Figura 11c) y la correlación entre la masa de
semillas y la cobertura vegetal (r = 0.366; p ≤ 0.0001, Figura 11d) fueron negativas, es
decir, conforme aumenta la temperatura y la cobertura vegetal aumenta la masa de las
semillas.
38
Correlación entre la masa de semillas y la altitud
579
1113151719
0 200 400 600 800 1000
Altitud (msnm)
Mas
a de
sem
illas
(g)
Correlación entre la masa de semillas y la precipitación
579
1113151719
800 1000 1200 1400 1600 1800
Precipitación (mm)
Mas
a de
sem
illas
(g)
Correlación entre la masa de semillas y la temperatura
579
1113151719
15 17 19 21 23 25 27 29
Temperatura (ºC)
Mas
a de
sem
illas
(g)
b) a) PN PA LR TE SI
PA LR PN TE
SI
p≤ 0.0001 Ŷ = 12.48 + -0.0009 X p≤ 0.0001 Ŷ = 12.83 + -0.0026 X
Correlación entre la masa de semillas y la cobertura vegetal
579
1113151719
40 50 60 70 80 90
Cobertura vegetal (%)
Mas
a de
sem
illas
(g)
c)
Figura 11. Relación entre la masa de semillas y las variables ambientales (a= altitud; b=
precipitación, c= temperatura y d= cobertura vegetal). Se muestra la ecuación de la
recta para la masa de semillas contra cada variable ambiental y el valor de p. TE =
Tenampa; SI = San Ignacio; LR = Loma de Rogel; PA = Puente Actopan; PN = Puente
Nacional. El valor de la masa corresponde al valor neto de las semillas multiplicado por
1000 para cada familia genética de cada sitio.
TE
SI
LR PA PN
Ŷ = 7.83 + 0.1642 X
p≤ 0.0001
TE d)
PN LR PA
SI
p≤ 0.0001 Ŷ = 8.39 + 0.045 X
39

6. DISCUSIÓN
6.1 Cobertura vegetal inter-sitio y asociación de plántulas.
Nuestros resultados indicaron que existen diferencias en los porcentajes de
cobertura vegetal a nivel inter-sitios, siendo la heterogeneidad ambiental (i.e.
precipitación, temperatura y humedad) la que puede estar determinando la abundancia
y riqueza de especies leñosas para cada uno de los sitios de estudio (ver la descripción
de los sitios) y éstas influyendo a su vez en los porcentajes de cobertura vegetal para
cada de ellos.
Puesto que la heterogeneidad ambiental ha sido documentada en muchos casos
como uno de los factores que modelan a las comunidades (i.e. áridas y semi-áridas)
como resultado de las limitaciones impuestas por bajos niveles de humedad y altas
temperaturas (ver Shreve, 1931; Woodell, Money & Hill, 1969; Noy-Meir, 1973; Gerhardt
& Hytteborn, 1992), por lo que nuestros resultado pueden estar apoyados por este
principio.
Bajo el supuesto de que la cobertura vegetal actuaría como un sitio seguro, se
esperaba que el gradiente de cobertura entre los sitos pudiera determinar el grado de
asociación de plántulas bajo el dosel para cada uno de ellos, es decir, a mayor
cobertura vegetal mayor asociación de plántulas en ésta. Nuestros resultados difieren
con el supuesto ya que sin importar el sitio, la asociación de plántulas no mostró mayor
frecuencia bajo el dosel. Y es que se sabe que la facilitación es uno de los procesos
ecológicos más importantes para la germinación, la supervivencia y el crecimiento para
algunas plantas (Flores & Jurado, 2003).
40

Estudios en cactáceas han contribuido significativamente para poder entender
como opera la facilitación, mostrando que las plantas leñosas resultan ser en muchos
casos indispensables para el reclutamiento de nuevos individuos para esta familia
botánica producto de los ambientes estresantes que habitan (Godínez-Alvarez et al.,
2003; Bashan et al., 2002; López & Valdivia, 2007).
Así mismo, muchos trabajos han sugerido la importancia de micro-hábitats
favorables para que se lleven a cabo la germinación y el establecimiento de plántulas
(Turner et al., 1966; Steenbergh & Lowe, 1977; Franco & Nobel, 1989), esperándose
que la cobertura vegetal podría estar actuando como un sitio seguro para promocionar
ambos rasgos.
Lo anterior pudo deberse principalmente a factores biológicos, que van desde la
granivoría hasta el impacto que puede tener la depredación en plántulas. Munguía &
Sosa (2007) quienes trabajaron con esta especie, realizaron experimentos de
germinación y supervivencia de plántulas bajo tres micro-ambientes: espacios abiertos,
bajo la copa de árboles y dentro de cavidades rocosas, mostrando que la germinación y
supervivencia de plántulas disminuía drásticamente en todos los sitios cuando las
semillas y las plántulas no tenían protección, haciendo que las diferencias en la
germinación y supervivencia registradas para cada uno de los sitios sin protección
desaparecieran entre si, lo cual concuerda con nuestras observaciones en campo, por
lo que al parecer la condición lumínica no es un factor determinante para el
establecimiento de plántulas como resulta ser la depredación.
Por lo que al parecer C. palmeri var. sartorianus no es especialista hacia un
micro-ambiente en particular como ha sido reportada para otras cactáceas columnares
(Callaway, 1998), lo cual concuerda además con los resultados del presente trabajo
41

(germinación bajo los tratamientos lumínicos), sugiriendo así que la especie no requiere
de una nodriza para el éxito en la germinación y el establecimiento.
6.2 Germinación y calidad de la luz.
6.2.1 Variación intra-sitio.
Los resultados de germinación a nivel intra-sitio mostraron que existen
diferencias en los porcentajes de germinación en respuesta a la luz para las familias
genéticas de cada uno de los sitios, indicando que hay variación genética en el atributo
medido a este nivel para todos los sitios y que esta variabilidad es dada por plasticidad
fenotípica como se indica en la interacción luz * familia genética, realizado en el análisis
de varianza general y confirmado por el ANDEVA de una vía para cada sitio.
Lo anterior es congruente con diversos estudios de genética cuantitativa
realizados en varias especies de plantas que han demostrado la existencia de
variabilidad genética para muchos caracteres cuantitativos a nivel inter e intra-sitios
(Venable, 1984; Billington et al., 1988).
La gran variabilidad genética encontrada puede deberse a dos posibles causas,
la primera, que la acción de la selección natural no ha eliminado completamente la
varianza genética, lo que podría estar permitiendo una alta variabilidad, y la segunda
causa, es que esta variabilidad genética podría representar una adaptación a la
heterogeneidad ambiental en la cual estos genotipos se desarrollan.
Estas adaptaciones a dicha heterogeneidad sugieren por un lado la existencia de
genotipos especializados para confrontar el ambiente lumínico, y por el otro, a las
42

condiciones del micro-sitio en el que se encontraba la planta madre (la sombra de la
copa de los árboles o el hábitat abierto), por lo que dicha variación pudo haber sido
influenciada por el ambiente aumentando el efecto materno, el cual no se pudo estimar,
ya que para esto se tuvieron que haber realizado cruzas controladas.
La variación genética en las respuestas plásticas de diferentes rasgos ha sido
ampliamente observada para diversas especies de plantas no-perennes (Schlichting &
Pigliucci, 1998). Nuestros resultados muestran que gran parte de la varianza obtenida
está explicada por el factor familia genética (anidada en sitio) pese a ser estudiados
sólo 15 familias genéticas de cada sitio, lo cual sugiere que los sitios donde se localiza
C. palmeri var. sartorianus mantienen una alta variación genética que, aunada a la
plasticidad fenotípica observada en la germinación, provee a esta especie con un
sistema genético capaz de responder a los cambios ambientales.
Así pues, los resultados del presente trabajo aportan evidencia sobre la
diferenciación en la plasticidad fenotípica intra-sitio, dando soporte a la relación entre
heterogeneidad ambiental dentro de las poblaciones y la magnitud de las respuestas
plásticas.
6.2.2 Variación inter-sitio.
La diferenciación genética a nivel inter-sitio fue baja y en conjunto constituye un
acervo con poca variabilidad pese a las diferencias ambientales del gradiente
estudiado; dos posibles causas que pudieron explicar la discrepancia del presente
análisis son:
43

La primera, una causa estadística, ya que la misma variación intra-sitio pudo
haber minimizado las diferencias inter-sitio, puesto que las familias genéticas al
responder de diferente manera al estímulo ambiental, minimizaron las diferencias entre
sitios.
La segunda causa que pudo haber influenciado la poca variación genética a nivel
inter-sitio pudo haber dependido de aspectos de la biología de la especie, como pueden
ser: su patrón de distribución espacial, la forma primordial de reproducir descendencia
(síndrome reproductivo) y el tipo de polinizador (ver Ehrlich & Raven, 1969; Schemske
& Lande, 1985), ya que la especie al presentar poblaciones grandes, el poseer un
sistema reproductivo de autofecundación como de fecundación cruzada y una
dispersión de polen por murciélagos (ver Capitulo 4.1), se espera entonces que el flujo
genético entre sitios sea alto y que la diferenciación entre éstos sea el producto de los
anteriores factores.
Aunque diversos trabajos mencionan que las poblaciones de plantas pueden
mostrar diferenciación en la plasticidad fenotípica (Platenkamp, 1990; Sultan & Bazzaz,
1993; Oyama, 1994) e incluso sugieren que es más común la diferencia entre
poblaciones para una misma especie que la diferenciación intra-poblacional (Allard et
al., 1968; Bradshaw, 1984; Venable & Búrquez, 1989), puesto que los patrones de
variación ambiental pueden influenciar la divergencia en la plasticidad de rasgos entre
éstas poblaciones (Galloway, 1995; Weining, 2000); incluso algunas teorías predicen
que mayores niveles de heterogeneidad ambiental pueden seleccionar altas
magnitudes de plasticidad fenotípica (Bradshaw & Hardwick, 1989), al existir evidencia
de plantas que crecen bajo ambientes más cambiantes presentan niveles mayores de
respuesta fenotípica.
44

Sin embargo, nuestros resultado no mostraron las tendencias antes
mencionadas y probablemente se deba al rasgo que se estimó ya que los caracteres
relacionados directamente con los rasgos de historia de vida presentan poca variación
entre los sitios, por lo que frecuentemente presentan una variabilidad genética reducida
(Falconer, 1981; Venable, 1984; Mitchell-Olds & Rutledge, 1986).
En cuanto a la diferencia significativa encontrada para el sitio de Tenampa en la
interacción luz * sitio ésta podría deberse al gradiente altitudinal, el cual impone
características particulares para este sitio, por ello, los resultados de la germinación
bajo una condición lumínica u otra pudieron deberse a procesos de adaptación local;
pues la variación en los rasgos de historias de vida es común en especies de
distribución amplia (Roff, 1992; Stearns, 1992) y esta variación creada y mantenida por
las diferencias en la disponibilidad y calidad de recursos entre los hábitat, es lo que a
menudo se considera como una evidencia de estrategias adaptativas que se dan entre
ambientes distintos (Reznick, 1996; Blanckenhorn, 1998, 1999; Travis et al., 1999; Leips
et al., 2000; Sultan, 2001).
6.2.3 Factor lumínico.
Los resultados obtenidos del análisis del factor lumínico en la germinación de
semillas de C. palmeri var. sartorianus no mostraron diferencias en la germinación con
respecto a la calidad de la luz, confirmando los resultados aportados por Rojas-
Aréchiga et al. (1997); Rojas-Aréchiga & Vázquez-Yanes (2000), quienes proponen que
muchas especies de cactáceas (principalmente columnares) presentan un fotoblastismo
45

neutro, puesto que la germinación puede ser estimulada por diferentes calidades
lumínicas.
Aunque los resultados de la germinación con respecto al factor lumínico (luz
blanca y luz rica en rojo-lejano) no fueron diferentes entre los tratamientos de luz para la
especie, no se descarta que los requisitos lumínicos para la germinación pudieran estar
determinados por una combinación de factores como son la humedad en el suelo y la
temperatura.
En nuestro experimento, la calidad de la luz, el agua y la temperatura fueron
mantenidos constantes, por lo que las posibles interacciones entre estos efectos no
pudieron ser evaluadas, ya que se conoce que en condiciones naturales, la luz es un
recurso heterogéneo espacio-temporalmente que además interactúa con otros factores
que pueden afectar la germinación (i.e. temperatura, disponibilidad de agua, etc.) (Pons,
1992; Vázquez-Yanes & Orozco-Segovia, 1994).
Se sabe que el tiempo y lugar de la germinación de semillas condicionan de
manera importante el crecimiento subsiguiente y el desarrollo de las plantas, por lo que
las semillas tienen que “utilizar” ciertas señales ambientales para germinar cuando las
condiciones del medio resultan ser óptimas, activando así dicho proceso y completando
el ciclo de vida de las plantas (Casal & Sánchez, 1998).
Para el caso de las cactáceas, Shreve (1931) observó en condiciones naturales
que la germinación de semillas era un evento raro y esporádico, por lo que muchas
especies aumentaban su germinación cuando se establecían bajo la copa de los
árboles de otras especies (fenómeno conocido como nodricismo). Existen diverso
trabajo y compilaciones que han abordado la asociación nodriza-protegida, (Flores &
Jurado, 2003).
46

Un ejemplo que ha abordado el tema del nodricismo y que reviste importancia
para el presente trabajo, ya que estudiaron la germinación de semillas de C. palmeri
var. sartorianus en campo, es el que realizaron Munguía & Sosa (2007), quienes
registraron diferencias en la germinación de esta especie en campo bajo tres
tratamientos: bajo la copa del dosel, en cavidades rocosas y en áreas desprovistas de
cualquier tipo de sombra y encontraron que los dos primeros tratamientos registraron
los mayores porcentajes de germinación, argumentando que la sobra proporcionada en
estos sitios funcionaba como un buffer de temperatura y evaporación, activando así el
proceso de la germinación. Por lo que al parecer, la luz por si sola no afecta a la
germinación, pero el efecto conjunto con otros factores ambientales puede determinar el
éxito de dicho proceso.
6.3 Correlaciones genéticas entre la respuesta de la germinación a las distintas
calidades de la luz.
La limitada independencia genética de los estados del carácter (correlación
genética significativa) en la germinación registrada para los sitios de San Ignacio, Loma
de Rogel, Puente Actopan y Puente Nacional, sugiere que es poco probable que esas
normas de reacción puedan evolucionar (ver Via, 1994).
El significado de una correlación genética significativa entre los ambientes indica
que lo estados del carácter no son independientes. Por lo que cualquier correlación
genética entre ambientes cuyo valor absoluto sea mayor a cero sugiere una restricción
genética en el ajuste independiente del fenotipo expresado entre cada uno de los
ambientes y por consiguiente una restricción en la evolución de una norma de reacción
47

adaptativa, es decir, ésta correlación genética distinta de cero es el resultado de la
expresión de los mismos alelos o conjuntos de alelos que afectan los estados del
carácter en dos ambientes de la misma forma (i.e. están determinadas por una fracción
de genes en común).
En contraste, para Tenampa se observó una independencia genética, por lo que
existe la posibilidad de la evolución de la norma de reacción para la capacidad de
germinar bajo diferentes condiciones lumínicas (Via & Lande, 1985; Via, 1994).
La evolución de la norma de reacción implica que la plasticidad fenotípica puede
ser considerada como un mecanismo para que las poblaciones produzcan fenotipos
alternos que resulten apropiados en los diferentes ambientes en que se desarrollan
(Via, 1993).
Por lo que la carencia de correlación genética en los estados del carácter
expresados en dos ambientes facilita el ajuste independiente de los estados del
carácter para cada ambiente, acelerando la evolución de la norma de reacción;
sugiriéndose que los genotipos de Tenampa pueden producir fenotipos para cada uno
de los ambientes lumínicos, lo que les permite una mayor especialización con respecto
al resto de los sitios.
Como se pudo ver, las condiciones de luz predominantes para el sitio de
Tenampa contrastaban con el resto de los sitios, registrándose un mayor porcentaje de
áreas sin cobertura vegetal, lo cual pudo influenciar la plasticidad fenotípica registrada
en la germinación bajo los dos tratamientos de luz (i.e. mayor porcentaje de
germinación bajo el tratamiento de luz blanca), que aunado a la independencia de los
estados del carácter sugieren que la evolución de la norma de reacción puede ser el
resultado de la selección natural para que se puedan producir genotipos especializados
48

para germinar en espacios sin cobertura vegetal, resultando ser un ventaja con respecto
al resto de los sitios.
Es por todo lo anterior que las variaciones en las correlaciones genéticas entre
los caracteres deben ser tomadas en cuenta, ya que pueden cambiar de un sitio a otro
(Mitchell-Olds & Rutledge, 1986; Venable & Búrquez, 1990), lo que podría indicar que
las presiones de selección varían en intensidad o en dirección, fomentando la
diferenciación genética entre distintos sitios (Billington et al., 1988; Venable & Búrquez,
1990), como se pudo apreciar en el presente trabajo.
6.4 Variación de la germinación en función de la masa de semillas.
En nuestro trabajo se observaron diferencias entre la masa de las semillas a nivel
ínter-sitio, lo cual puede ser producto de que la masa está fuertemente influenciada por
las condiciones ambientales presentes para cada sitio, que se reflejan en las
condiciones ambientales en las cuales la planta madre es expuesta (Gutterman, 1980-
81) y que es un fenómeno recurrente para varias poblaciones (McWilliams et al., 1968;
Baker, 1972; Schimpf, 1977; Melzack & Watts, 1982).
Así mismo, los resultados de la masa de las semillas a nivel intra-sitio también
mostraron una alta variación, como la registrada en los trabajos de Harper & Obeid
(1967); Wulff (1973); Milberg et al. (1996); Kidson & Westoby (2000); Baloch et al.
(2001). Además, en algunos casos se menciona que la variación en el tamaño de
semillas puede desempeñar un factor importante en los procesos de germinación,
existiendo una relación positiva entre el tamaño de la semilla y el éxito en la
49

germinación (Jurado & Westoby, 1992; Milberg et al., 1996; Pereira de Souza & Válio,
2001).
En el presente trabajo las diferencias en la masa a nivel inter o intra-sitio no
afectaron de manera significativa la germinación. Lo anterior puede deberse a que este
es el rango de masa adecuada para asegurar la germinación producto de un balance
entre la madurez fisiológica y las reservas de la semilla. Ayala-Cordero et al. (2004)
demostraron que en Stenocereus benekei, pese a poseer una amplia variación intra-
sitio en el tamaño de las semillas, no se registraron diferencias en el éxito de la
germinación entre diferentes clases de tamaños.
Generalmente hay una relación entre el tamaño de las semillas y la cantidad de
luz que requieren para su germinación, pues semillas pequeñas requieren de mayor
cantidad de luz para germinar con respecto a semillas grandes (Grime et al., 1981;
Pons, 1992; Baskin & Baskin, 1998; Milberg et al., 2000; Maiti et al., 2003; y este mismo
patrón es sugerido para cactáceas por Rojas-Aréchiga & Batis (2001); aunque poco se
conoce acerca de las diferencias en los requisitos lumínicos entre semillas de distintos
tamaños para una misma especie.
En el presente trabajo se observó que no existen diferencias en la germinación
con respecto a la masa de las semillas con el tratamiento de luz, por lo que al parecer,
la masa de las semillas y las distintas condiciones lumínicas podrían estar
correlacionadas con la velocidad germinativa y no en el porcentaje final para esta
especie.
Puesto que los rasgos de germinación y de la masa de semillas son
considerados como íntimamente relacionados con la adecuación de las plantas (Waller,
1982; Mitchell-Olds & Rutledge, 1986; Mazer, 1987), se esperaría que presenten
50

variabilidades bajas o nulas; Sin embargo, diversos trabajos han demostrado para el
caso de la germinación, en especial en plantas con distribución latitudinal amplia, que
éstas pueden variar debido a la variación geográfica (clima, suelo, etc.), la cual
determina las condiciones ambientales de la pre y post-dispersión experimentadas por
la semilla (Donohue et al., 2005). Por lo que los factores climáticos son las causas
externas más comunes de variación ambiental que repercuten en la germinación y la
masa de semillas (Waller, 1982).
Otras fuentes de variación son los efectos maternos, pues influyen de manera
directa en el desarrollo de la semilla (Schaal, 1984; Schwaegerle & Levin, 1990), y
pueden ser una causa frecuente de diferenciación ambiental entre familias, ya que las
semillas están sujetas a dicho ambiente durante las primeras etapas de desarrollo,
influyendo en los valores fenotípicos de muchos caracteres métricos (i.e. masa de
semillas). Lo anterior ocasiona que los descendientes de una misma madre se parezcan
entre sí (disminución en la varianza intra-familiar) y exista una mayor diferencia entre
familias y poblaciones (aumentando las varianzas respectivas, producto de la ubicación
de la planta madre) (Falconer, 1981).
6.5 Variación de la germinación y la masa de las semillas en función de las variables
ambientales.
Nuestros resultados mostraron variabilidad en la germinación y en la masa de
semillas entre los sitios, lo cual concuerda con estudios que demuestran que dichos
rasgos pueden variar a nivel intra e inter-sitio para una misma especie (Janzen, 1977;
Cavers & Steel, 1984).
51

Cabe destacar que aunque ambos rasgos (germinación y masa) resultaron estar
correlacionados contra todas las variables ambientales, las pendientes de las
correlaciones para cada uno de ellos no fueron tan pronunciadas.
Lo anterior puede apoyarse en lo que varios autores han propuesto, ya que
mencionan que la germinación y el tamaño de las semillas (para el caso de estudio:
masa) son características estables en plantas (ver Harper, 1977); puesto que la planta
madre hace una distribución equitativa de los recursos en la progenie, ya sea por ser el
producto directo de la reproducción sexual, como por ser el principal medio para la
dispersión de la especie.
Por lo que ambos rasgos tenderían hacia un tipo de selección estabilizadora que
debería favorecer la constancia en la germinación y la masa de las semillas dentro de
una misma especie (Harper et al., 1970).
En el caso de los sitios estudiados se encontró una variabilidad moderada, esto
parece indicar que, aún cuando la germinación y la masa de las semillas están
determinados genéticamente, las diferencias en la disponibilidad de recursos del medio
ambiente o durante la ontogenia de la planta madre tienen un efecto importante sobre
ambos rasgos.
De forma natural C. palmeri var. sartorianus se distribuye desde los 87 hasta los
879 msnm, por lo que las diferencias en el gradiente altitudinal, nos permitieron
relacionar variables ambientales como la altitud, la temperatura, la precipitación y la
cobertura vegetal en las variaciones en la germinación y de la masa de las semillas
entre los sitios, demostrándose que estas variables ambientales pueden tener un papel
importante en la determinación de la germinación y la masa de las semillas.
52

6.6 Discusión general.
De acuerdo con los resultados de la asociación de plántulas en campo bajo los
ambientes lumínicos, al parecer C. palmeri var. sartorianus es no es especialista hacia
un micro-ambiente en particular como ha sido reportada para otras cactáceas
columnares, sugiriéndose que la especie no requiere de un ambiente de sombra
producida por árboles (i.e. nodriza) para aumentar la germinación y el establecimiento.
Lo anterior concuerda además con los resultados obtenidos de la germinación
de semillas sometidas bajo los tratamientos lumínicos en cámara de germinación, los
cuales muestran que para C. palmeri var. sartorianus el factor lumínico no fue
determinante en el éxito de la germinación de semillas, y que a su vez confirma los
resultados de Rojas-Aréchiga et al. (1997); Rojas Aréchiga & Vázquez-Yanes (2000),
quienes mencionan que las cactáceas columnares presentan un fotoblástismo neutro.
Por otro lado, aunque la luz no afecta de manera significativa la germinación de
la especie, sin embargo si constituye un agente selectivo (aun que no sea de
importancia demográfica), ya que existen respuestas diferenciales entre los genotipos
en respuesta al factor lumínico.
Lo anterior se demuestra en los resultados de germinación a nivel intra-sitio, ya
que mostraron la existencia de variación genética en el rasgo para cada uno de los
sitios estudiados, lo cual podría estar representando por un lado, la existencia de
genotipos especializados para confrontar el ambiente lumínico o una adaptación a la
heterogeneidad ambiental en la cual estos genotipos se desarrollan
53

Lo cual sugiere que los sitios donde se localiza C. palmeri var. sartorinus
mantienen una alta variación genética, pese a ser estudiados sólo 15 familias genéticas
de cada sitio.
Nuestros resultados muestran poca variabilidad genética a nivel inter-sitio que
pudo deberse a la biología de la especie, como puede ser: su patrón de distribución
espacial, el síndrome reproductivo y el tipo de polinizador. Por lo que se espera que el
flujo genético entre sitios es alto debido a la combinación de ambos factores, producto
de los “emigrantes” que han comunicado a los sitios en una escala de tiempo
considerable (por tratarse de una no-perenne) y de una manera recurrente,
presentándose así una alta combinación genética entre los sitios que disminuyó las
varianzas genéticas entre éstos.
Por otra parte, la dependencia genética de los estados de carácter en la
germinación para los sitios de San Ignacio, Loma de Rogel, Puente Actopan y Puente
Nacional, sugiere que es poco probable que esas normas de reacción puedan
evolucionar (ver Via, 1994). En contraste nuestros resultados indican que para
Tenampa existe la posibilidad de la evolución de la norma de reacción, debido a la
independencia genética, la cual puede facilitar el ajuste independiente de los estados
del carácter para cada ambiente, es decir, la expresión de genotipos apropiados para
confrontar cada uno de los ambientes lumínicos.
Así mismo se encontró variabilidad en la germinación y masa de semillas a nivel
inter-sitio, indicando que, aún cuando ambos rasgos están determinados
genéticamente, las diferencias en la disponibilidad de recursos del medio ambiente o
durante la ontogenia de la semilla en la planta madre tienen un efecto importante sobre
ambos rasgos (Janzen, 1977; Cavers & Steel, 1984)
54

Si bien las generalizaciones acerca de la distribución de la especie y la
plasticidad fenotípica basada solamente en la respuesta a la intensidad de la luz debe
ser considerada con precaución, en nuestros resultados se muestra que C. palmeri var.
sartorianus posee plasticidad fenotípica y que es ésta plasticidad la que posiblemente le
confiere a la especie oportunidades para distribuirse a lo largo de todo el gradiente
altitudinal en comparación con la distribución restringida que presentan las otras tres
cactáceas columnares de la selva baja caducifolia (N. scoparia, N. euphorbioides y S.
griseus). Lo anterior se apoya en la idea que diversos estudios han demostrado, pues
se ha observado que la plasticidad fenotípica tiende a estar asociada positivamente con
una distribución amplia de una especie (Cordell et al., 1998; Sultan et al., 1998; Bell &
Sultan, 1999; Valladares et al., 2000a).
55

7. REFERENCIAS BIBLIOGRÁFICAS
Alcorn, S.M., & Kurtz, E.B. 1959. Some factors affecting the germination of seed of the
saguaro cactus (Carnegiea gigantea). American Journal of Botany 46: 526-529.
Allard, R.W., Jain, S.K., & Workman, P.L. 1968. The genetics of inbreeding populations.
Advances in Genetics 14: 55-131.
Arias, I., & Lemus, L. 1984. Interaction of light, temperature and plant hormones in the
germination of seeds of Melocactus caesius Went (Cactaceae). Acta Científica
Venezolana 35: 151-155.
Ayala-Cordero, G., Terrazas, T., López-Mata, L., & Trejo, C. 2004. Variación en el
tamaño y peso de la semilla y su relación con la germinación en una población
de Stenocereus beneckei. Interciencia 29: 692-697.
Ballaré, C.L. 1994. Light gaps: sensing the light opportunities in highly dynamic canopy
environments. En: Caldwell MM, Pearcy RW, eds. Exploitation of environmental
heterogeneity by plants. San Diego: Academic Press, Inc, 73-110.
Baker, H.G. 1972. Seed weight in relation to environmental conditions in California.
Ecology 53: 997-1010.
Baloch, H.A., DiTomasso, A., Watson, A.K. 2001. Intrapopulation variation in Abutilon
theophrasti seed mass and its relationship to seed germinability. Seed Sci. Res.
11: 335-343.
Bashan, Y. Li, C.Y., Lebsky, V.K., Moreno, M., de Bashan, L.E. 2002. Primary
colonization of volcanic rocks by plants in arid Baja California, Mexico. Plant
Biology 4: 392-402.
56

Baskin, C.C., & Baskin, J.M. 1998. Seeds: ecology, biogeography and evolution of
dormancy and germination. Academic Press, San Diego, CA.
Bazzaz, F.A. 1979. The physiological ecology of plant sucession. Annu. Rev. Ecol.
Syst. 10: 351-371.
Bazzaz, F.A. 1996. Plants in changing environments. Cambridge: Cambridge University
Press.
Bazzaz, F.A. & Pickett, S.T.A. 1980. Physiological tropical succession: a comparative
review. Annu. Rev. Ecol. Syst. 11: 287-310.
Bell, D.L., & Sultan, S.E. 1999. Dynamic phenotypic plasticity for root growth in
Polygonum: a comparative study. American Journal of Botany 86: 807-819.
Billington, H.L., Mortimer, A.M., McNelly, T. 1988. Divergence and genetic structure in
adjacent grass populations. I. quantitative genetics. Evolution 42: 1267-1277.
Blanckenhorn, W.U. 1998. Adaptive phenotypic plasticity in growth, development, and
body size in the yellow dung fly. Evolution 52: 1394-1407.
Blanckenhorn, W.U. 1999. Different growth responses to temperature and resource
limitation in three fly species with similar life histories. Evolutionary Ecology 13:
395-409.
Bradshaw, A.D. 1965. Evolutionary signficance of phenotypic plasticity. Genetics
13:115-155.
Bradshaw, A.D. 1984. The importance of evolutionary ideas in ecology-and visce versa.
En: Shorrocks, B. (ed.). Evolutionay Ecology. Blackwell Scientific Publications,
Oxford. Pp. 1-25.
Bradshaw, A.D. & Hardwick, K. 1989. Evolution and stress-genotypic and phenotypic
components. Biol. J. Linn. Soc. 37: 137-155.
57

Bravo-Hollis, H. 1978. Las cactáceas de México, Vol. I. UNAM, México.
Callaway, R.M. 1998. Are positive interactions species-specific? Oikos 82: 202-207.
Casal, J.J. & Sanchez, R.A. 1998. Phytochrome and seed germination. Seed Science
Research 8: 317-329.
Cavers, P. & Steel, M. 1984. Patterns of change in seed weight over time on individual
plants. Amer. Nat. 124: 324-335.
Cody, M.L. 1993. Do cholla (Opuntia spp., Subgenus Cylindropuntia) use or need nurse
plants in the Mojave Desert? J. Arid Environm. 24: 139-154.
Cordell, S., Goldstein, G., Mueller-Dombois, D., Webb, D. & Vitousek, P.M. 1998.
Physiological and morphological variation in Metrosideros polymorpha, a
dominant Hawaiian tree species, along an altitudinal gradient: the role of
phenotypic plasticity. Oecologia 113: 188-196.
Donohue, K. 2003. Setting the stage: plasticity as habitat selection. Int. J. Plant Sci. 164:
S79-S92.
Donohue, K., Dorn, L., Griffith, C., Kim, E., Aguilera, A., Polisetty, C.R., & Schmitt, J.
2005. Environmental and genetic influences on the germination of Arabidopsis
thaliana in the field. Evolution 59: 740-757.
Ehrlich, P.R., & Raven, P.H. 1969. Differentation of populations. Science 165: 1228-
1232.
Ellner, S., & Shmida, A. 1981. Why are adaptations for long-range seed dispersal rare in
desert plants ? Oecologia 51: 133-144.
Evans, A.S., & Cabin, R.J. 1995. Can dormancy affect the evolution of post-germination
traits? The case of Lesquerella fendleri. Ecology 76: 344-356.
Falconer, D.S. 1952. The problem of environmental selection. Amer. Natur. 86: 293-298.
58

Falconer, D.S. 1981. Introduction to quantitative genetics. 2da. Ed., Longman, London.
Fleming, T.H., Tuttle, M.D. & Horner, M.A. 1996. Pollination biology and the relative
importance of nocturnal and diurnal pollinators in three species of Sonoran Desert
columnar cacti. The Southwestern Naturalist 41: 257-269.
Flores, J., & Jurado, E. 2003. Are nurse-protégé interactions more common among
plants from arid environments?. Journal of Vegetation Science 14: 911-916.
Flores, J., Jurado, E., & Arredondo, A. 2006. Effect of light on germination of seeds of
Cactaceae from the Chihuahuan Desert, Mexico. Seed Science Research 16:
149-155.
Franco, A.C. & Nobel, P.S. 1989. Effect of nurse plants on the microhabitat and growth
of cacti. J. Ecol. 77: 870-886.
Galloway, L.F. 1995. Response to natural environmental heterogeneity: maternal effects
and selection on life-history characters and plasticities in Mimulus guttatus.
Evolution 49: 1095-1107.
García, E. 2004. Modificaciones al sistema de clasificación climática de Köppen (para
adaptarlo a las condiciones de la República Mexicana). Serie Libros. Núm 6. 5 ta.
Edición. México, D.F. 90 pp.
Gerhardt, K. & Hytteborn, H. 1992. Natural dynamics and regenerations methods in
tropical dry forest, and introduction. J. Veg. Sci. 3: 361-364.
Gilbert, I.R., Jarvis, P.J., & Smith, H. 2001. Proximity signal and shade avoidance
differences between early and late succesion trees. Nature 411: 792-796.
Godínez-Álvarez, H., Valverde, T., & Ortega-Baes, P. 2003. Demographic Trends in the
Cactaceae. The Botanical Review 69: 173-203.
59

Grime, J.P., Mason, G., Curtis, A.V., Rodman, J., Band, S.R., Mowforth, M.A.G., Neal,
A.M., & Shaw, S. 1981. A comparative study of germination characteristics in a
local flora. J. Ecol. 69: 1017-1059.
Grime, J.P. & Jeffrey, D.W. 1965. Seedlings establishment in vertical gradients of
sunlight. Journal of Ecology 53: 621-642.
Gutterman, Y. 2000. Environmental factors and survival strategies of annual plant
species in the Negev Desert, Israel. Plant Species Biology 15: 113-125.
Gutterman, Y. 1993. Seed germination in desert plants (Adaptations of desert
organisms). Berlin, Springer-Verlag.
Gutterman, Y. 1980-81. Influences on seed germinability: phenotypic maternal effects
during seed maduration. Israel Journal of Botany 29: 105-117. Citado en: Wulff,
R. 1986. Seed size variation in Desmodium paniculatum. Journal of Ecology 74:
87-97.
Hamrick, J.L., Nason, J.D., Fleming, T.H., & Nassar, J.M. 2002. Genetic diversity in
columnar cacti. En: Fleming & Valiente-Banuet. 2002. Colomnar Cacti and their
mutualists. The University of Arizona Press. U.S.A. 371 pp.
Harper, J.L., & Obeid, M. 1967. Influence of seed size and depth of sowing on the
establishment and growth of varieties of fiber and oilseed flax. Crop Science 7:
527-532.
Harper, J., Lovell, P., Moore, K. 1970. The shapes and sizes of seeds. Ann. Rev. Ecol.
Syst. 1: 327-356.
Harper, J. 1977. Population biology of plants. Academic Press, New York, New York,
U.S.A.
60

Hayes, R.G., & Klein, W.H. 1974. Spectral quality influence of light during development
of Arabidopsis thaliana plants in regulating seed germination. Plant Cell Physiol.
15: 643-653.
Janzen, D. 1977. Variation in seed size within a crop of a Costa Rica Mucuna andreana
(Leguminosae). Amer. J. Bot. 64: 347-349.
Jurado, E. & Westoby, M. 1992. Germination biology of selected central Australian
plants. Aust. J. Ecol. 17: 341-348.
Kidson, R., & Westoby, M. 2000. Seed mass and seedling dimensions in relation to
seedling establishment. Oecologia 125: 11-17.
Leips, J., Travis, J., & Rodd, F.H. 2000. Genetic influences on experimental population
dynamics of the least killfish. Ecological Monographs 20: 289-309.
Leishman, M.R., Wright, I.J., Moles, A.T., & Westoby, M. 2000. The evolutionary ecology
of seed size. Seeds: the ecology of regeneration in plat communities, 2nd edn (Ed.
M. Fenner), CAB International, Wallingford, UK. Citado en: Murray, B.R., Brown,
A.H.D., Dickman, C.R. & Crowther, M.S. 2004. Geographical gradients in seed
mass in relation to climate. Journal of Biogeography 31: 379-388.
Lewontin, R.C. 1974. The genetic basis of evolutionary change. Columbia University
Press, New York. -ed. En español por Omega, Barcelona (1979).
López, R.P. & Valdivia, S. 2007. The importance of shrub cover for tour cactus species
differing in growth form in a Andean semi-desert. Journal of Vegetation Science
18: 263-270.
Maiti, R.K., Hernandez-Piñero, J.L., & Valdez-Marroquin, M. 1994. Seed ultrastructure
and germination of some species of Cactaceae. Phyton 55: 97-105.
61

Maiti, R.K., Singh, V.P., Baquie, A., Sánchez-Arreola, E., Wesche-Ebeling, P., Cuervo-
Parra, J.A., Perdome-Velázquez, H., & Lorenzo, J.L. 2003 Cactus: Biology,
propagation and conservation. Hisar, India, The Gaurav Society of Agricultural
Research Information Centre. Citado en: Flores, J., Jurado, E., & Arredondo, A.
2006. Effect of light on germination of seeds of Cactaceae from the Chihuahuan
Desert, Mexico. Seed Science Research 16: 149-155.
Manrique, E. 2003. Los pigmentos fotosintéticos, algo más que la captación de luz.
Ecosistemas 2003/1 (URL: http//www.aeet.org/ecosistemas /031/informe4.htm).
Martínez-Holguín, E. 1983. Germinación de semillas de Stenocereus griseus (Haw)
Buxbaum (Pitayo de mayo). Cactáceas y Suculentas Mexicanas 28: 51-57.
Mazer, S.J. 1987. The quantitative genetics of life history and fitness components in
Raphanus raphanistrum L. (Brassicaceae): ecological and evolutionary
consequences of seed-weight variation. American Naturalist 130: 891-914.
McCullough, J.M., & Shropshire, Jr. 1970. Physiological predetermination of germination
responses in Arabidopsis thaliana (L.) HEYNH. Plant Cell Physiol. 11: 139-148.
McDonough, W. 1964. Germination responses of Carnegia gigantea and Lemairocereus
thurberi. Ecology 45: 155-159.
McWilliams, E.L., Landers, R., & Mahlstede, J.P. 1968. Variation in seed weight and
germination in populations of Amaranthus retroftexus L. Ecology 49: 290-96.
Melzack, R.N., & Watts, D. 1982. Variation in sedd weigth, germination and seedling
vigour in the yew (Taxus baccata L.) in England. Journal of Biogeography 9: 55-
63.
Mitchell-Olds, T., & Rutledge, J.J. 1986. Quantitative genetics in natural populations: a
review. American Naturalist 127: 379-402.
62

Milberg, P., Andersson, L., & Thompson, K. 2000. Large-seeded species are less
dependent on light for germination than small-seeded ones. Seed Science
Research 10: 99-104.
Milberg, P., Anderson, L., Elfverson, C., & Regnér, S. 1996. Germination characteristics
of seeds differing in mass. Seed Sci. Res. 6: 191-197.
Montgomery, D.C. 1976. Design and Analysis of Experiments. John Wiley & Sons, New
York, NY.
Munguía-Rosas, Sosa & Jácome (en preparación)
Munguía-Rosas, M.A. & Sosa, V.J. 2007. Nurse plants vs. Nurse objects: Effects of
woody plants and rocky cavities on the recruitment of the Pilosocereus
leucocephalus Columnar cactus. Annals of Botany 1: 1-11.
Murray, B.R., Brown, A.H.D., Dickman, C.R. & Crowther, M.S. 2004. Geographical
gradients in seed mass in relation to climate. Journal of Biogeography 31: 379-
388.
Niering, W.A., Wittaker, R.H., & Lowe, C.H. 1963. The saguaro: a population in relation
to environment. Science 142: 15-23.
Nobel, P.S. 1988. Environmental Biology of Agaves and Cacti. New York: Cambridge
University Press.
Noy-Meir, I. 1973. Desert ecosystems: environment and producers. Annu. Rev. Syst. 4:
25-51.
Núñez-Farfán, J.S. 1991. Biología evolutiva de Datura stramonium L. en el centro de
México: Selección natural de la resistencia a los herbívoros, sistema de
cruzamiento y variación genética intra e interpoblacional. Tesis doctoral
(Ecología), Centro de Ecología-UACPyP, UNAM, México.
63

Núñez-Farfán, J.S., & Cordero, C. 1993. Tópicos de Biología Evolutiva. Centro de
Ecología, U.N.A.M. México.
Olff, H., Pegtel, D.M., Van Groenendael, J.M., & Bakker, J.P. 1994. Germination
strategies during grassland succession. Journal of Ecology 82: 69-77.
Oyama, K. 1994. Ecological amplitude and differentiation among populations of Arabis
serrata Fr. and Sav. (Brassicaceae). Int. J. Plant Sci. 155: 220-234.
Pereira de Souza, R., & Válio, I. F. M. 2001. Seed Size, Seed Germination, and
Seedling Survival of Brazilian Tropical Tree Species Differing in Successional
Status. Biotropica 33: 447-457.
Pigliucci, M. 2001. Phenotypic Plasticity: Beyond Nature and Nurture. John. Hopkins
University Press, Baltimore.
Platenkamp, G.A.J. 1990. Phenotypic plasticity and genetic differentiation in the
demography of the grass Anthoxanthum odoratum. J. Ecol. 78: 772-788.
Pons, T.L. 1992. Seed reponses to light. In: Fenner M, ed. The ecology of regeneration
in plants communities. Wallingford, UK: CAB International, 259-284.
Reznick, D. 1996. Life history evolution in guppies: a model system for the empirical
study of adaptation. Netherlands Journal of Zoology 46:172-190.
Rhodes, I., & Sterns, W.R. 1978. Competition for light. En: Wilson, J. R. (Ed.), Plant
relations in pastures. Commonwealth scientific and industrial researche
organization, Melbourne, 175-178.
Roff, D.A. 1992. The evolution of life histories. Chapman and Hall, New York, New York,
USA.
Rojas-Aréchiga, M., & Batis, A.I. 2001. Las semillas de cactáceas. . . ¿forman bancos
en el suelo? Cactáceas y Suculentas Mexicanas 46:76-82.
64

Rojas-Aréchiga, M., Casas, A., & Vázquez-Yanes, C. 2001. Seed germination of wild
and cultivated Stenocereus stellatus (Cactaceae) from the Tehuacán-Cuicatlán
Valley, Central México. Journal of Arid Environments 49: 279-287.
Rojas-Aréchiga, M., & Vázquez-Yanes, C. 2000. Cactus seed germination: a review.
Journal of Arid Environments 44: 85-104.
Rojas-Aréchiga, M., Orozco-Segovia, A., & Vázquez- Yanes, C. 1997. Effect of light on
germination of seven species of cacti from Zapotitlán Valley in Puebla, México.
Journal of Arid Environments 36: 571-578.
Roundy, B.A., Taylorson, R.B., & Sumrall, L.B. 1992. Germination responses of
Lehmann Lovegrass to light. Journal of Range Management 45: 81-84.
Schaal, B.A. 1984. Life-history variation, natural selection, and maternal effects in plant
population. In: Dirzo, R. & Sarukhán, J. (eds.). Perspectives on plant population
ecology. Sinauer, Sunderland, Mass. Pp. 188-206.
Schemske, D.W., & Lande, R. 1985. The evolution of self-fertilization and inbreeding
depression in plants. II. Empirical observations. Evolution 39: 41-52.
Schimpf, D.J. 1977. Seed weigth of Amaranthus retroflexus in relation to moinsture and
length of growing season. Ecology 58: 450-453.
Schlichting, C. 1986. The evolution of phenotypic plasticity. Annu. Rev. Ecol. Syst.
17:667-693.
Schlichting, C.D., & Pigliucci, M. 1998. Phenotypic evolution: a reaction norm
perspective. Sunderland: Sinauer Associates, Inc.
Schmalhausen, II. 1949. Factors of evolution. Chicago University Press. Chicago.
Schwaegerle, K.E., & Levin, D.A. 1990. Quantitative genetics of seed size variation in
Phlox. Evolutionary Ecology 4: 143-148.
65

Sears, M.W., & Angilletta, M.J. 2003.Life-history variation in the sagebrush lizard:
phenotypic plasticity or local adaptation?. Ecology 84:1624-1634.
Shreve, F. 1910. The rate of establishment of the giant cactus. Pl. World 10: 235-240.
Shreve, F. 1931. Physical conditions in sun and shade. Ecology 12: 96-104.
Silvertown, J.W. 1981. Seed size, lifespan and germination date as coadapted features
of plant life history. American Naturalist 118: 860-864.
Silvertown, J. 1980. Leaf-canopy-induced seed dormancy in grassland flora. New
Phytologist 85:109-118.
Smith, H. 1982. Light quality, photoperception and plant strategy. Annual Review of
Plant Physiology 33: 481-518.
Smith, H., & Whitelam, G.C. 1997. The shade avoidance syndrome: multiple responses
mediated by multiple phytochromes. Pl. Cell Environm. 20: 840-844.
Sosa, V.J., & Fleming, T.H. 2002. Why columnar cacti are associated to nurse plants?.
En: T. H. Fleming and A. Valiente-Banuet, editors. Evolution, ecology and
conservation of columnar cacti and their mutualists. University of Arizona Press.
306-323.
Steenbergh, W.F. & Lowe, C.H. 1977. Critical factors during the first years of life of
Saguaro (Cereus giganteus) at Saguaro National Monument, Arizona. Ecology
50: 823-834.
Sultan, S. E. 1987. Evolutionary implications of phenotypic plasticity in plants. Evol. Biol.
21: 127-178.
Sultan, S.E. and Bazzaz, F.A. 1993. Phenotypic plasticity in Polygonum persicaria. II.
Norms of reaction to soil moisture and the maintenance of genetic diversity.
Evolution 47: 1032-1049.
66

Stearns, S.C. 1992. The Evolution of Life Histories. Oxford University Press, New York.
Sultan, S.E., Wilczek, A.M., Bell, D.L., & Hand, G. 1998. Physiological response to
complex environments in annual Polygonum species of contrasting ecological
breadth. Oecologia 115: 564-578.
Sultan, S.E. 2001. Phenotypic plasticity for fitness components in Polygonom species of
contrasting ecological breadth. Ecology 82: 328-343.
Swaine, M.D. & Whitmore, T.C 1988. On the definition of ecological species group in
the tropical rain forest. Vegetatio 75: 81-86.
Travis, J., McManus, M.G., & Baer, C.F. 1999. Sources of variation in physiological
phenotypes and their evolutionary significance. American Zoologist 39: 422-433.
Turner, R.M., Alcorn, S.M., Olin, G., & Booth, J.A. 1966. The influence of shade, soil,
and water on saguaro seedling establishment. Bot. Gaz. 127: 95-102.
Valencia-Díaz, S., & Montaña, C. 2003. Effects of seed age, germination substrate,
gibberelic acid, light, and temperature on seed germination in Flourensia cernua
(Asteraceae), a Chihuahuan Desert shrub. Southwest. Nat. 48: 1-13.
Valiente-Banuet, A., & Ezcurra, E. 1991. Shade as a cause of the association between
the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana in the
Tehuacán Valley, Mexico. Journal of Ecology 79: 961-971.
Valiente-Banuet, A., Arizmendi, M.C., Roja-Martínez, A. & Domínguez-Canseco, I. 1996.
Ecological relationships between columnar cacto and nectar-feeding bats in
Mexico. Journal of Tropical Ecology 12: 103-119.
Valiente-Banuet, A., Arizmendi, M. C., Rojas-Martínez, A., Casas, A., Godínez-Álvarez,
H., Silva, C. & Dávila, P. 2002. Biotic interactions and population dynamics of
columnar cacti. En: Fleming, T. H. & Valiente-Banuet, A. (eds.), Columnar cacti
67

and their mutualists: Evolution, ecology, and conservation. University of Arizona
Press, Tucson.
Valladares, F., Martinez-Ferri, E., Balaguer, L., Perez-Corona, E., & Manrique, E. 2000a.
Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: a
conservative resource-use strategy? New Phytologist 148: 79-91.
Vázquez-Yanes, C., & Orozco-Segovia, A. 1994. Ecophysiology of seed germination in
the tropical humid forest of the World: a review. En: E. Medina, H.A. Mooney, and
C. Vázquez-Yanes (Eds.). Physiological ecology of plants of the wet tropics, pp.
37-50. Dr. W. Junk, The Hague, Netherlands.
Vazquez-Yanes, C., Rojas-Arechiga, M., Sanchez-Coronado, M.E., & Orozco-Segovia
A. 1996. Comparison of light-regulated seed germination in Ficus spp and
Cecropia obtusifolia: ecological implications. Tree Physiology 16: 871-875.
Venable, D.L. 1984. Using intraspecific variation to study the ecological significance and
evolution of plant life-histories. En Dirzo, R. & Sarukhán, J. (eds.). Perspectives
on plant population ecology. Sinauer, Sunderland, Mass. Pp. 166-187.
Venable, D.L., & Búrquez, M.A. 1989. Quantitative genetics of size, shape. Life-history,
and fruit characteristics of the seed-heteromorphic composite Heterosperma
pinnatum. I. Variation within and among populations. Evolution 43: 113-124.
Venable, D.L., & Búrquez, M.A. 1990. Quantitative genetics of size, shape. Life-history,
and fruit characteristics of the seed-heteromorphic composite Heterosperma
pinnatum. II. Correlation structure. Evolution 44: 1748-1763.
Via, S. 1993. Adaptative phenotypic plasticity: target or by-product of selection in a
variable environment?. American Naturalist 142: 352-365.
68

Via, S. 1994. The evolution of phenotypic plasticity: what do we really know?. En: Read
L, ed. Ecological genetics. Princeton: Princeton University Press, 35-57.
Via, S. & Lande, R. 1985. Genotype-environment interaction and the evolution of
phenotypic plasticity. Evolution 39: 505-522.
Waller, D.M. 1982. Factors influencing seed weight in Impatiens capensis
(Balsaminaceae). American Journal of Botany 69: 1470-1475.
Weinig, C. 2000. Plasticity versus canalization: population differences in the timing of
shade avoidance responses. Evolution 54: 441-451.
Woltereck, R. 1909. Weitere experimentelle Untersuchungen über Artveränderung,
speziell über das Wesen quantitätiver Artenunterschiede bei Daphniden. Verh.
D. Zool. Ges. 1909: 110-172. En: Schlichting & Pigliucci, 1998. Phenotypic
evolution. A reaction norm perspective (Sinauer, Sunderland, MA).
Woodell, S.R.J., Money, H.A. & Hill, A.J. 1969. The behaviour of Larrea divaricata
(creosote bush) in response to rainfall in California. J. Ecology 57: 37-44.
Wright, S.D., & McConnaughay, K.D.M. 2002. Interpreting phenotypic plasticity: the
importance of ontogeny. Plant Species Biology 17: 119-131.
Wulff, R. 1973. Intrapopulation variation in the germination of seeds in Hyptis
suaveolens. Ecology 54: 646-649.
Wulff, R. 1986. Seed size variation in Desmodium paniculatum. Journal of Ecology 74:
87-97.
Zar, J.H. 1999. Biostatistical analysis. Prentice Hall.
Zimmer, K., & Buttner, P. 1982. Ersatz des Lichtbedurfnisses bei der Keimung von
Kakteensamen durch Gibberellinsaure. Gartenbauwissenschaft 47: 97-101.
Citado en: Benítez-Rodríguez, J.L., Orozco-Segovia, A., & Rojas Aréchiga, M.
69

2004. Light effect on seed germination of four Mammillaria species from the
Tehuacán-Cuicatlán Valley, Central Mexico. The Southwestern Naturalist 49: 11-
17.
70





























