-
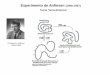
Membrana plasmtica
Bicapa lipdica Mezcla de
enzimas hidrolticas
Protenas de transporte glicosiladas
Degradacin lisosmica
- Son vesculas digestivas que contienen enzimas hidrolticas y
que sirven para la fagocitosis y procesos autofgicos.
- En condiciones de ayuno prolongado se activa la degradacin
selectiva de protenas por el lisosoma.
- La secuencia KFERQ es reconocida por la chaperona HSP70.
- La chaperona HSP70 dirige a la protena al interior del
lisosoma donde ser degradada.
- VIDEO: Funcin de los lisosomas
Lys-Phe-Glu-Arg-Gln (KFERQ)
-
Degradacin por la va Ubicuitina-Proteasoma (1)
Poli-ubicuitilacin
Degradacin proteosmica
Lisosoma
Formacin de autofagosomas
Protena nativa
Protena mal plegada
Agregados proteicos
Agregosoma
Toxicidad
Final de la vida til
- Las protenas mal plegadas pueden agregarse y tener efectos
txicos.
- La ubicuitilacin es el mecanismo principal de degradacin de
protenas mal plegadas o que es necesario eliminar.
- La ubicuitilacin de agregados proteicos determina su
destruccin por degradacin lisosmica
- La vida media de una protena est marcada por el aminocido que
est en su extremo amino terminal.
-
1) El extremo carboxlico terminal de la ubicuitina se activa por
adenilacin catalizada por la Enzima Activadora de Ubicuitina
(E1)
2) La adenilacin permite la incorporacin de la ubicuitina a un
grupo sulfhidrilo de la propia enzima E1
3) La Enzima Conjugante de Ubicuitina (E2) transfiere la
ubicuitina a otro grupo sulfhidrilo de ella
4) La enzima Ubicuitina ligasa (E3) cataliza la transferencia de
la ubicuitina desde la enzima E2 a la protena que va a ser
marcada.
5) Sucesivas reacciones de la enzima E3 permiten la incorporacin
de ms unidades de ubicuitina a la protena objetivo (target). La
enzima E3 es la responsable de la identificacin de las protenas que
se van a marcar.
Degradacin por la va ubicuitina-Proteasoma (2)
-
Degradacin por la va ubicuitina-Proteasoma (3)
Carboxilo terminal
ubicuitina
Enlaces Isopeptdicos
+ Lisina Carboxilo terminal
Enlace isopeptdico
Enlace Isopeptdico
- La ubicuitina es una protena pequea (76 aa), muy conservada,
que se utiliza para marcar las protenas que van a ser
degradadas.
- Las ubicuitinas se unen a la protena mediante un enlace amida
entre el grupo carboxlico terminal de la ubicuitina y un grupo
amino de una cadena lateral de la lisina 48.
- Es necesario acumular varias ubicuitinas consecutivas para que
la protena se degrade.
- La incorporacin de ubicuitinas en otras lisinas distintas de
la 48 tiene funciones reguladoras.
-
Protena para degradar
Corte
Descarga
Liberacin Reconocimiento
Apertura
Desplegado
Degradacin por la va Ubicuitina-Proteasoma (4)
a
b
b
a
Centro regulador
Ncleo cataltico
Protena
ubicuitinas
- El proteasoma (26S) es un complejo multiproteico con un Ncleo
Cataltico (20S) en el centro, formado por cuatro anillos con 7
subunidades (a o b) cada uno.
- Contiene un Centro Regulador en cada extremo responsable del
reconocimiento de la protena ubicuitilada, el desplegado de la
protena y la liberacin de las ubicuitinas.
- La ubicuitinas liberadas se reciclan.
- La incorporacin de ubicuitinas en otras lisinas distintas de
la 48 tiene funciones reguladoras.
-
Enzimas que facilitan el plegamiento de las protenas
Prolil-pptido Isomerasa
Protena-Disulfuro Isomerasa
Protenas de choque trmico (Chaperonas)
- La Prolil-Ptido Isomerasa corrige la isomerizacin a cis de los
enlaces peptdicos de la prolina
- La Protena-Disulfuro Isomerasa ayuda a encontrar los enlaces
disulfuro correctos para el plegamiento
- Las protenas de choque trmico mantienen a las protenas
desplegadas, pero protegidas, para evitar su plegamiento
incorrecto
-
Estructura de las Chaperoninas (Complejo GroEL-GroES)
- GroEL est formado por dos anillos (cis y trans) de 7
subunidades cada uno.
- Las subunidades trans tienen un sitio de unin para el ATP.
- Los dos anillos dejan un hueco diseado para el plegamiento de
la protena.
Anillo cis
Anillo trans
-
Mecanismo de accin de las Chaperoninas (Complejo
GroEL-GroES)
- Cuando el complejo contiene ATP, la protena desplegada se une
a la entrada del tubo GroEL.
- Se incorpora la subunidad GroES encerrando a la protena
(1).
- La hidrlisis de ATP provoca el plegamiento correcto de la
protena (2).
- La segunda mitad se carga con una protena desplegada y con ATP
(3).
- La entrada del ATP desprende la tapa GroES y provoca la salida
de la protena plegada, junto con los restos de ADP (4).
- El ciclo vuelve a empezar en la segunda mitad.