Embed Size (px)
DESCRIPTION
Citoesqueleto y polaridad.
Citation preview

Mas allá de la polaridad de polímeros: como el
citoesqueleto construye una célula polarizada.
Rong Li, Gregg Gundersen The Stowers Institute for Medical Research, 1000 East 50th Street, Kansas City, Missouri 64110, USA.
Department of Pathology & Cell Biology, Columbia University, 1217 Black Building, 630 West 168th Street, New York, New York 10032, USA.

Resumen
La polaridad celular depende de la organización asimétrica de los componentes celulares.
Los microtúbulos y la actina están bien capacitados para dar polaridad a la célula
Por su inherente polaridad estructural
Dinámica que les permite responder a las señales de polaridad. Actina: domina el proceso de pérdida de la simetría.
Microtúbulos: mantienen la estabilidad de la organización polar.
Comunicación continua entre las dos estructuras.

Objetivos de la revisión
¿Cómo los arreglos polarizados de actina y microtúbulos se generan o como contribuyen a la polaridad celular y el transporte?

Polaridad celular
Resulta de un eje vectorial que dirige la organización interna de
las células
Propiedades: -Acumulación asimétrica de
componentes móviles.
-organización polar del citoesqueleto a lo largo del eje
de polaridad.
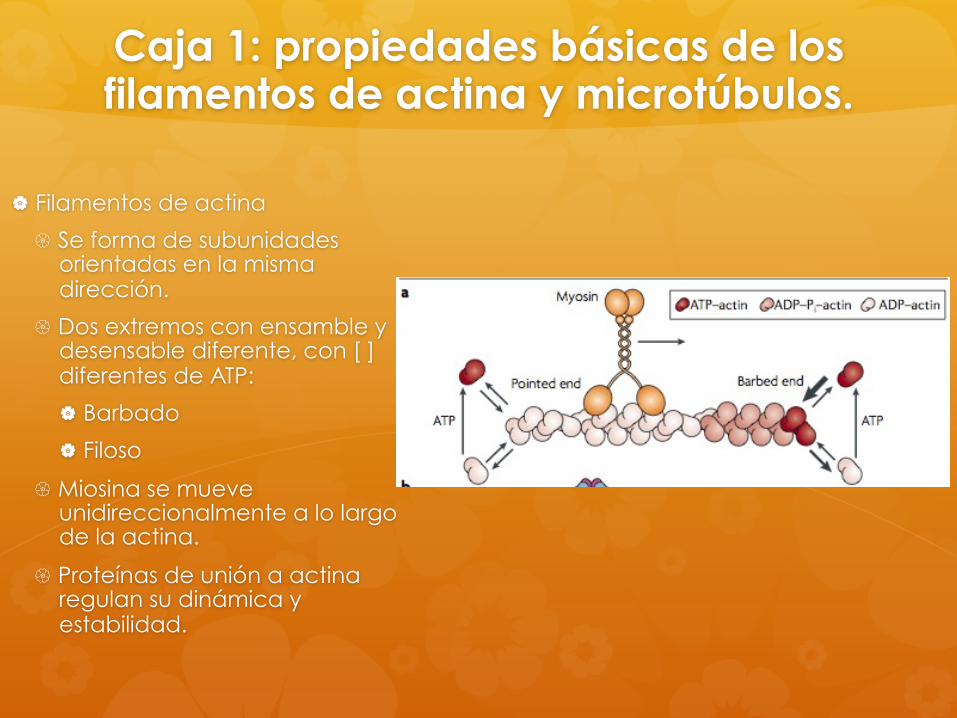
Caja 1: propiedades básicas de los filamentos de actina y microtúbulos.
Filamentos de actina
Se forma de subunidades orientadas en la misma dirección.
Dos extremos con ensamble y desensable diferente, con [ ] diferentes de ATP:
Barbado
Filoso
Miosina se mueve unidireccionalmente a lo largo de la actina.
Proteínas de unión a actina regulan su dinámica y estabilidad.
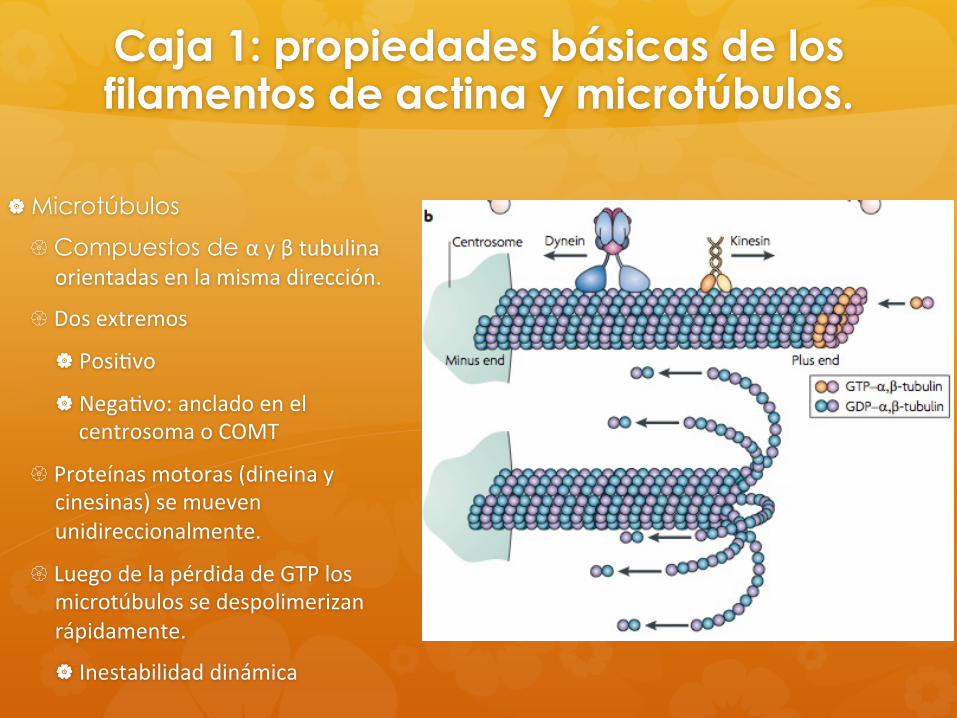
Caja 1: propiedades básicas de los filamentos de actina y microtúbulos.
Microtúbulos
Compuestos de α y β tubulina orientadas en la misma dirección.
Dos extremos
Posi8vo
Nega8vo: anclado en el centrosoma o COMT
Proteínas motoras (dineina y cinesinas) se mueven unidireccionalmente.
Luego de la pérdida de GTP los microtúbulos se despolimerizan rápidamente.
Inestabilidad dinámica
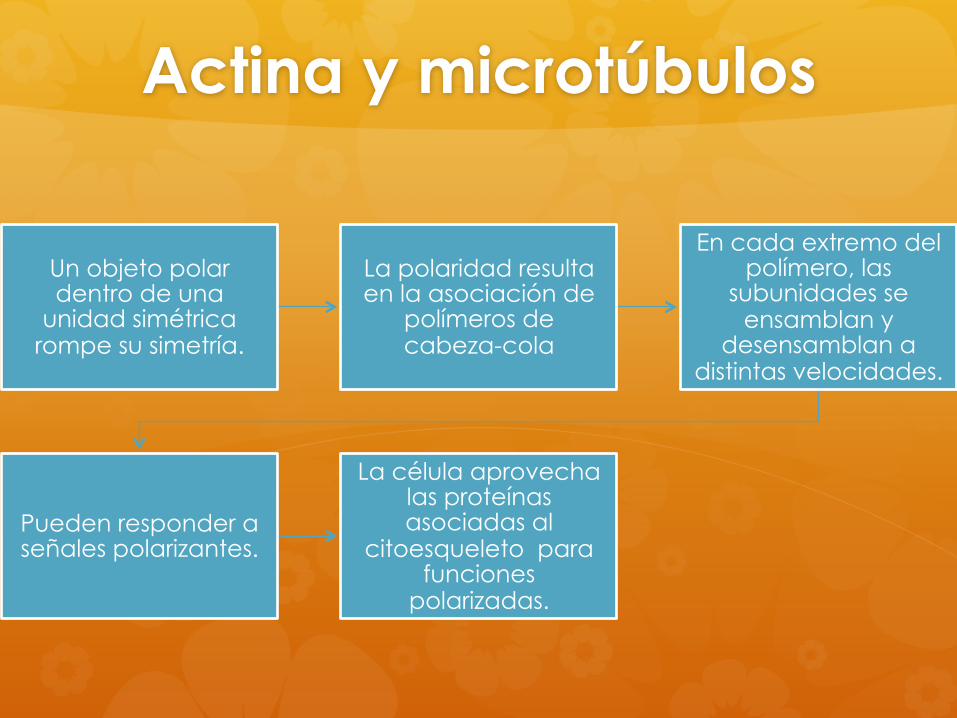
Actina y microtúbulos
Un objeto polar dentro de una
unidad simétrica rompe su simetría.
La polaridad resulta en la asociación de
polímeros de cabeza-cola
En cada extremo del polímero, las
subunidades se ensamblan y
desensamblan a distintas velocidades.
Pueden responder a señales polarizantes.
La célula aprovecha las proteínas asociadas al
citoesqueleto para funciones
polarizadas.

Los filamentos intermedios no se involucran en la polaridad de la célula.
Estructuras no polares
Sin embargo, se ha observado que proteínas no polares de la familia de septina (semejante a los filamentos intermedios) son importantes para la polaridad en diversas células.

Proteínas asociadas al citoesqueleto de importancia para la polaridad.
Proteínas motoras asociadas a actina
Miosina: movimiento unidireccional hacia el extremo barbado de actina (excepto miosina IV) por hidrólisis de ATP.
Miosina V: se mueve procesivamente a lo largo de los filamentos de actina (+ pasos consecutivos) por lo que es ideal para transportar por largas distancias.
Miosina II: baja procesividad, pero genera movimiento contractil deslizando los filamentos de actina.

Proteínas asociadas al citoesqueleto de importancia para la polaridad.
Proteínas motoras asociadas a filamentos intermedios
Cinesinas: se mueven en dirección al extremo positivo de los microtúbulos
Dineína: se mueve en dirección al extremo negativo en presencia del complejo de dinactina.
¿Cómo se desarrolla la polaridad celular?
Ruptura de la simetría
Mantenimiento de la polaridad
Polaridad celular
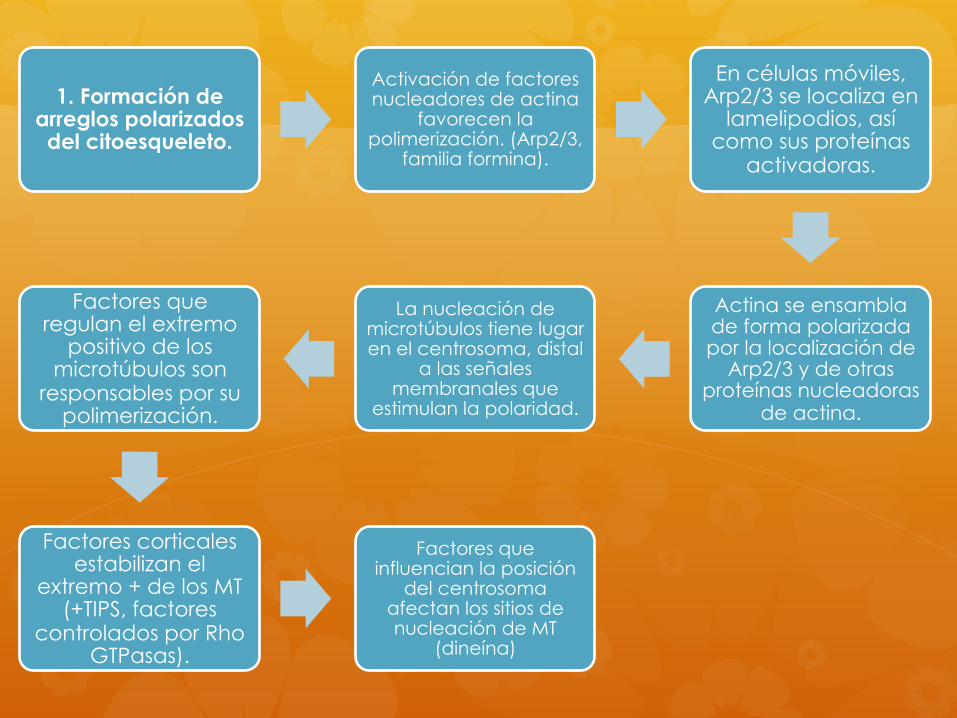
1. Formación de arreglos polarizados del citoesqueleto.
Activación de factores nucleadores de actina
favorecen la polimerización. (Arp2/3,
familia formina).
En células móviles, Arp2/3 se localiza en
lamelipodios, así como sus proteínas
activadoras.
Actina se ensambla de forma polarizada por la localización de
Arp2/3 y de otras proteínas nucleadoras
de actina.
La nucleación de microtúbulos tiene lugar en el centrosoma, distal
a las señales membranales que
estimulan la polaridad.
Factores que regulan el extremo
positivo de los microtúbulos son
responsables por su polimerización.
Factores corticales estabilizan el
extremo + de los MT (+TIPS, factores
controlados por Rho GTPasas).
Factores que influencian la posición
del centrosoma afectan los sitios de nucleación de MT
(dineína)
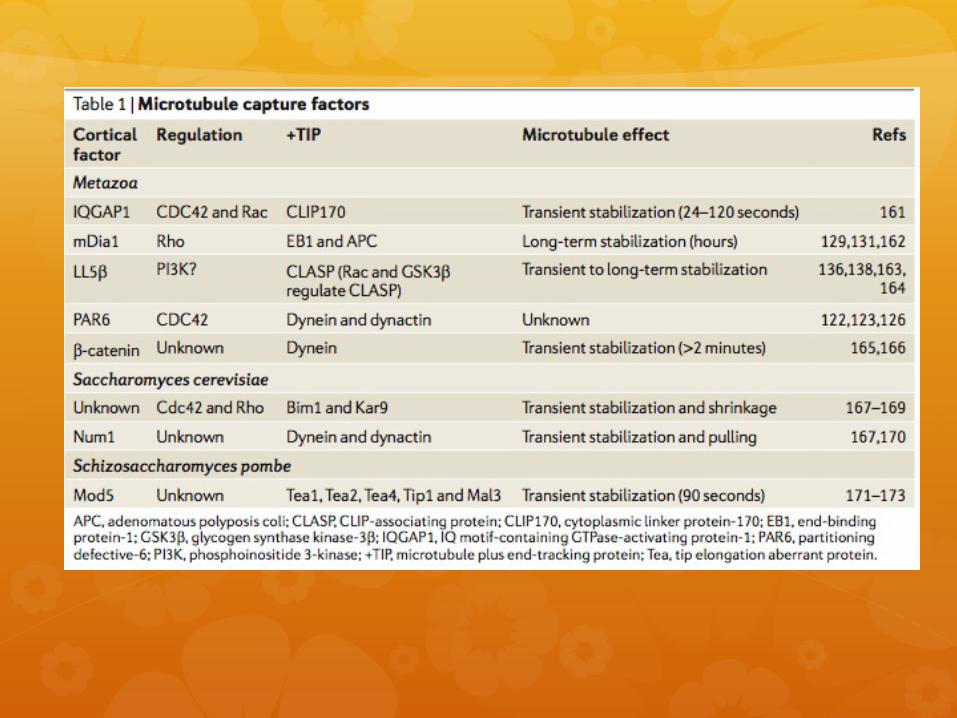
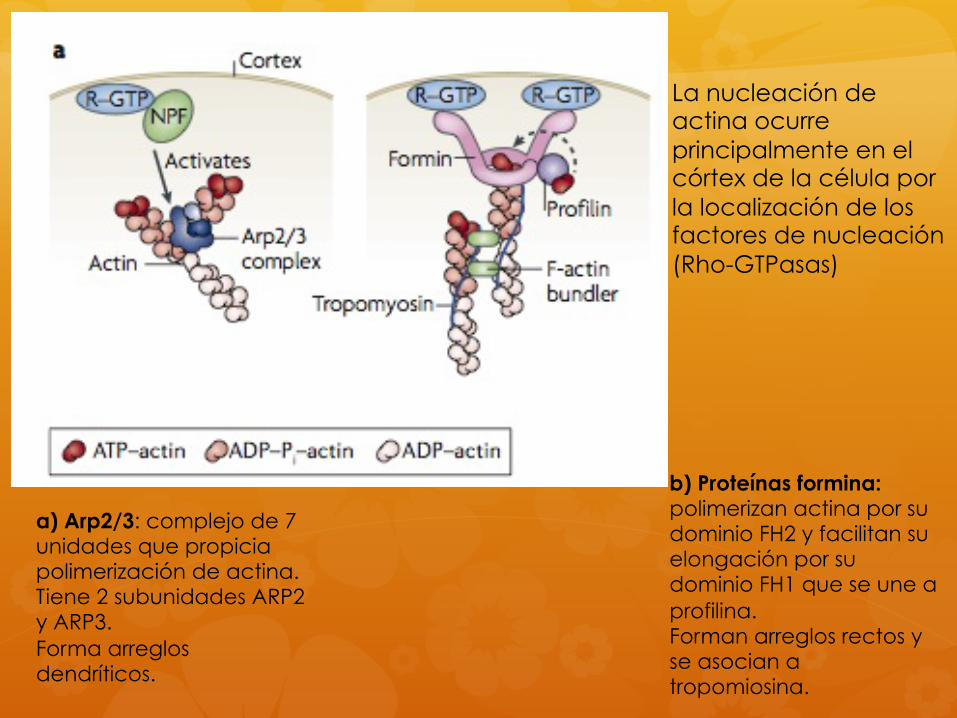
a) Arp2/3: complejo de 7 unidades que propicia polimerización de actina. Tiene 2 subunidades ARP2 y ARP3. Forma arreglos dendríticos.
b) Proteínas formina: polimerizan actina por su dominio FH2 y facilitan su elongación por su dominio FH1 que se une a profilina. Forman arreglos rectos y se asocian a tropomiosina.
La nucleación de actina ocurre principalmente en el córtex de la célula por la localización de los factores de nucleación (Rho-GTPasas)
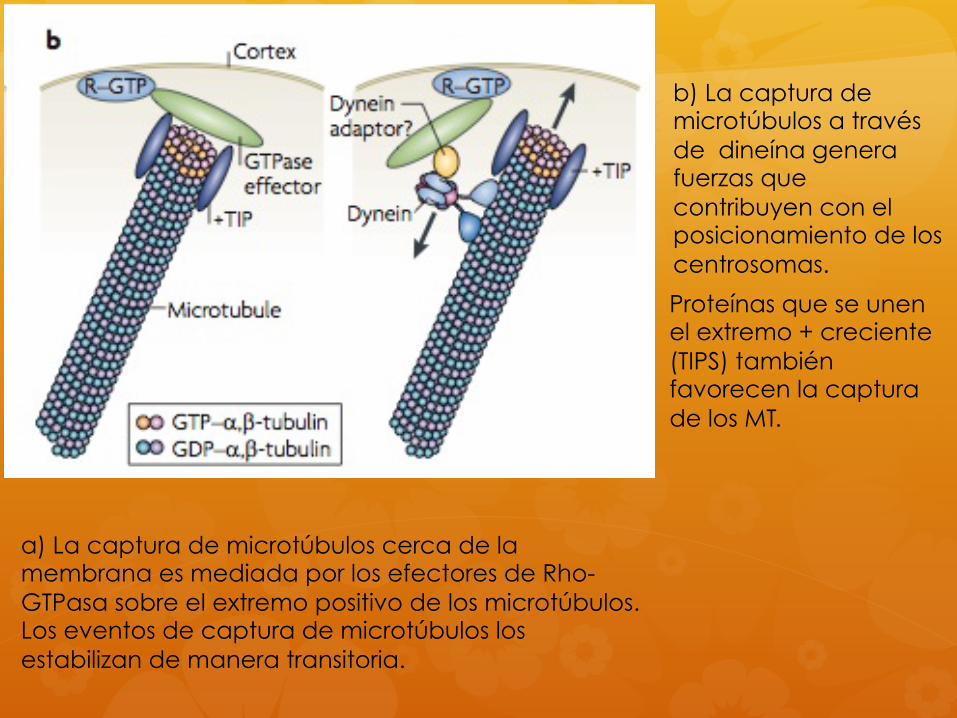
a) La captura de microtúbulos cerca de la membrana es mediada por los efectores de Rho-GTPasa sobre el extremo positivo de los microtúbulos. Los eventos de captura de microtúbulos los estabilizan de manera transitoria.
Proteínas que se unen el extremo + creciente (TIPS) también favorecen la captura de los MT.
b) La captura de microtúbulos a través de dineína genera fuerzas que contribuyen con el posicionamiento de los centrosomas.
2. Ruptura de la simetría celular.
Se ha demostrado que el
citoesqueleto no solo media las
funciones de as células
polarizadas.
Sino que contribuye
activamente en la pérdida de la
polaridad celular a través de proteínas
motoras y el ensable de estructuras.
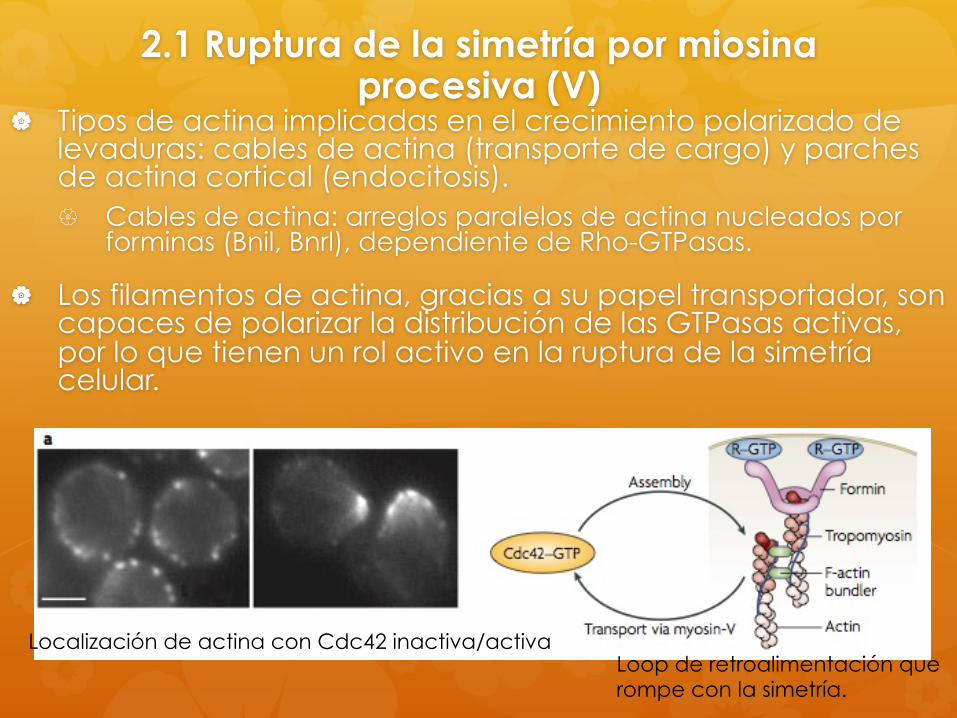
2.1 Ruptura de la simetría por miosina procesiva (V)
Tipos de actina implicadas en el crecimiento polarizado de levaduras: cables de actina (transporte de cargo) y parches de actina cortical (endocitosis). Cables de actina: arreglos paralelos de actina nucleados por
forminas (Bnil, Bnrl), dependiente de Rho-GTPasas.
Los filamentos de actina, gracias a su papel transportador, son capaces de polarizar la distribución de las GTPasas activas, por lo que tienen un rol activo en la ruptura de la simetría celular.
Localización de actina con Cdc42 inactiva/activa Loop de retroalimentación que rompe con la simetría.
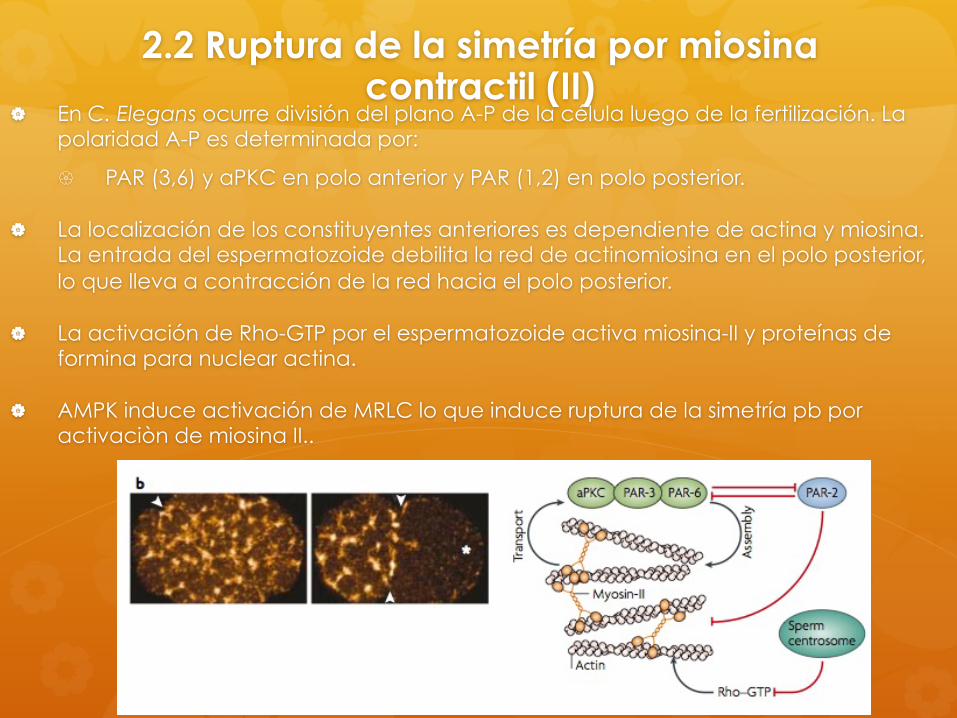
2.2 Ruptura de la simetría por miosina contractil (II)
En C. Elegans ocurre división del plano A-P de la célula luego de la fertilización. La polaridad A-P es determinada por:
PAR (3,6) y aPKC en polo anterior y PAR (1,2) en polo posterior.
La localización de los constituyentes anteriores es dependiente de actina y miosina. La entrada del espermatozoide debilita la red de actinomiosina en el polo posterior, lo que lleva a contracción de la red hacia el polo posterior.
La activación de Rho-GTP por el espermatozoide activa miosina-II y proteínas de formina para nuclear actina.
AMPK induce activación de MRLC lo que induce ruptura de la simetría pb por activaciòn de miosina II..
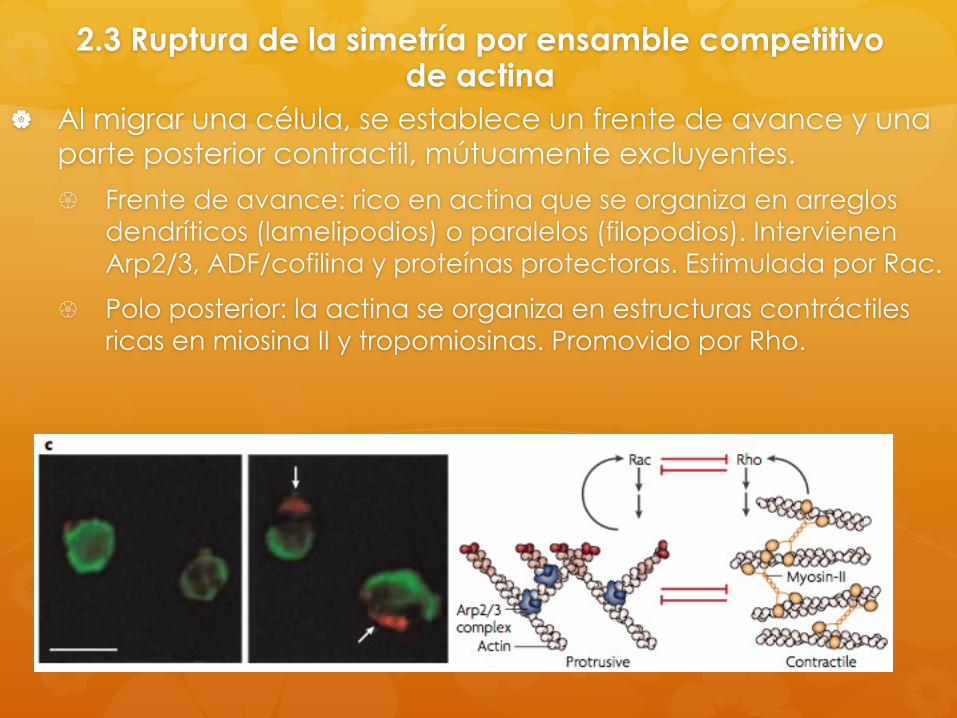
2.3 Ruptura de la simetría por ensamble competitivo de actina
Al migrar una célula, se establece un frente de avance y una parte posterior contractil, mútuamente excluyentes.
Frente de avance: rico en actina que se organiza en arreglos dendríticos (lamelipodios) o paralelos (filopodios). Intervienen Arp2/3, ADF/cofilina y proteínas protectoras. Estimulada por Rac.
Polo posterior: la actina se organiza en estructuras contráctiles ricas en miosina II y tropomiosinas. Promovido por Rho.
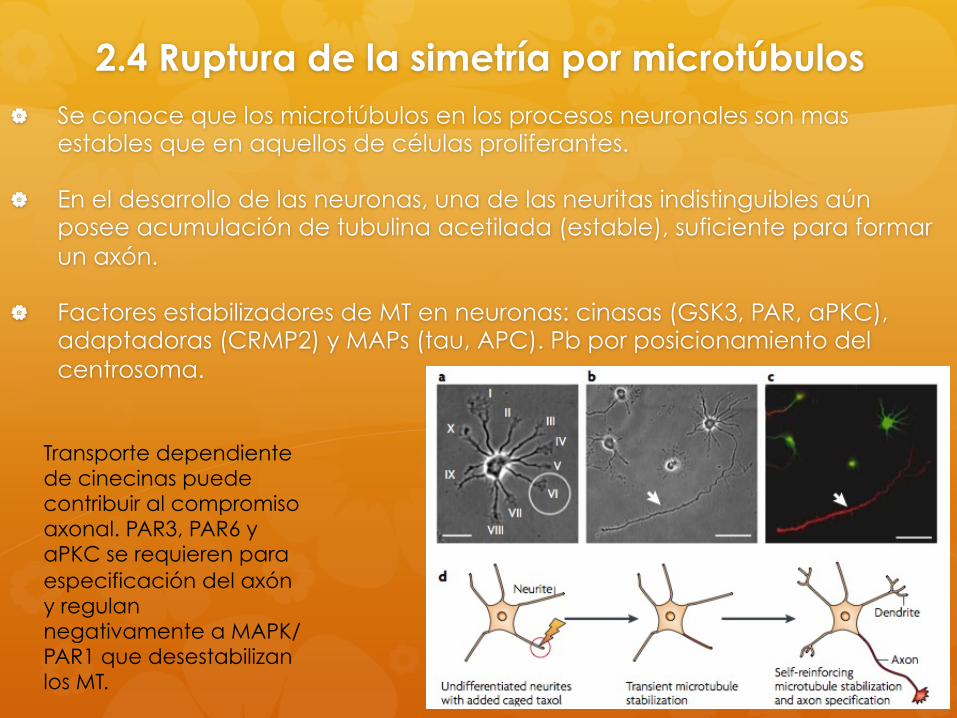
2.4 Ruptura de la simetría por microtúbulos Se conoce que los microtúbulos en los procesos neuronales son mas
estables que en aquellos de células proliferantes.
En el desarrollo de las neuronas, una de las neuritas indistinguibles aún posee acumulación de tubulina acetilada (estable), suficiente para formar un axón.
Factores estabilizadores de MT en neuronas: cinasas (GSK3, PAR, aPKC), adaptadoras (CRMP2) y MAPs (tau, APC). Pb por posicionamiento del centrosoma.
Transporte dependiente de cinecinas puede contribuir al compromiso axonal. PAR3, PAR6 y aPKC se requieren para especificación del axón y regulan negativamente a MAPK/PAR1 que desestabilizan los MT.
3. Mantenimiento de la polaridad celular
Una vez establecida, la polaridad se mantiene según el tipo celular.
La actina contribuye al mantenimiento de la
polaridad celular de forma dinámica por endocitosis.
Los MT refuerzan la polaridad inicial establecida
por el citoesqueleto de actina, en células grandes o que requieran de polaridad
por mucho tiempo.

3.1 Mantenimiento dinámico de la polaridad por endocitosis.
Las proteínas corticales polarizadas pueden difundir por la membrana o lejos de ella. Esto debe controlarse de manera dinámica. Por ej. el reciclaje de Cdc42.
El reciclaje endocítico ocurre en zonas ensambladas por Arp2/3 y miosina I.
Algunas proteínas endocitadas inducen endocitosis, regulando la dinámica de actina (Cdc42, Oskar).

3.2 Microtúbulos en la polaridad de LT
Durante la presentación de antígenos a las células T, e crea una sinapsis inmunológica dependiente de actina. El centrosoma se posiciona cerca de la sinapsis gracias a la activación de CDC42.
Dineína interactúa con ADAP (proteína adaptadora promotora de adhesión y degranulación).
Dineína asociada a la sinapsis crea la fuerza para que los MT arrastren el centrosoma a la sinapsis para posicionar el aparato de Golgi a la sinapsis.
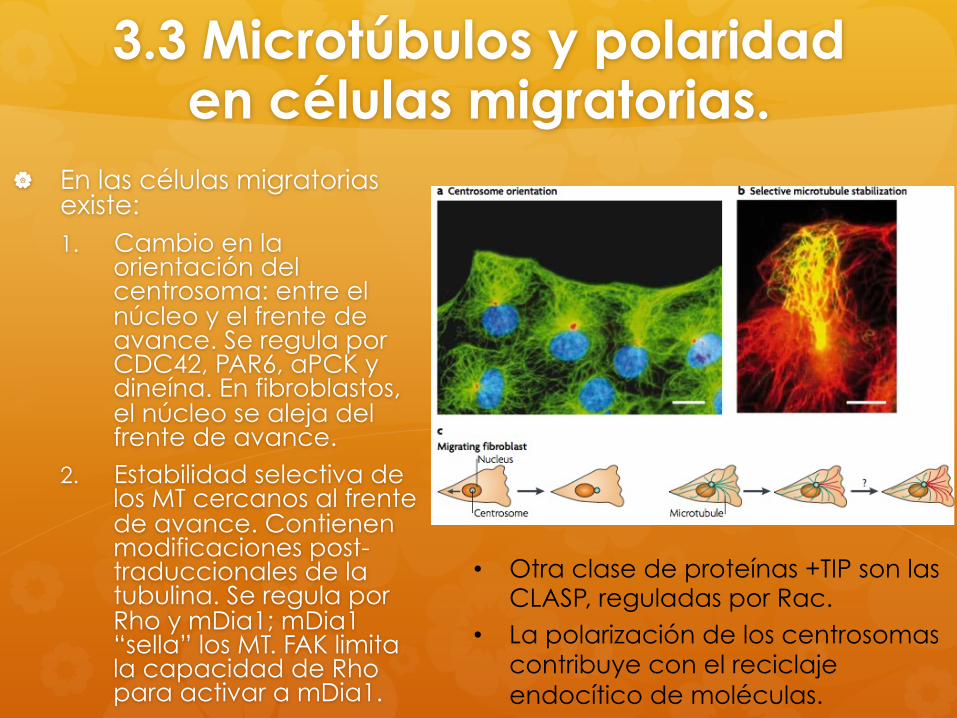
3.3 Microtúbulos y polaridad en células migratorias.
En las células migratorias existe: 1. Cambio en la
orientación del centrosoma: entre el núcleo y el frente de avance. Se regula por CDC42, PAR6, aPCK y dineína. En fibroblastos, el núcleo se aleja del frente de avance.
2. Estabilidad selectiva de los MT cercanos al frente de avance. Contienen modificaciones post-traduccionales de la tubulina. Se regula por Rho y mDia1; mDia1 “sella” los MT. FAK limita la capacidad de Rho para activar a mDia1.
• Otra clase de proteínas +TIP son las CLASP, reguladas por Rac.
• La polarización de los centrosomas contribuye con el reciclaje endocítico de moléculas.

3.4 Microtúbulos en el mantenimiento de la polaridad epitelial
Luego del establecimiento de la polaridad en células epiteliales, los MT sufren rearreglo de estilo radial centrosomal, a no centrosomal.
Epitelio columnar: arreglo paralelo al eje apical-basal, con el extremo positivo en el polo basal. Ecadherina de las uniones adherentes contribuye a la estabilidad de MT formando MT no centrosomales.

Comunicación entre filamentos de actina y microtúbulos
Debe existir comunicación bilateral entre los filamentos de actina y los MT para mantener la polaridad.
Disrrupción de MT por nocodazol o colchicina activa Rho en toda la célula, aumentando la contractibilidad por el citoesqueleto de actina.
Los focos de adhesión se componen de integrinas asociadas a fibras de estrés de actina que proveen de tracción para la migración celular. El desensamble es iniciado por los MT e involucra la endocitosis de integrinas.

Conclusiones
La polaridad celular dura desde minutos hasta días o años dependiendo del tipo celular.
La actina tiene un rol clave en la pérdida de la polaridad y permite una respuesta rápida a estímulos.
Los microtúbulos estabilizan la asimetría creada por actina. Pueden estar orientados por el centrosoma o por modificaciones post-traduccionales de los MT.





























